Knowledge for place-based ecosystem restoration in Færder- and Ytre Hvaler national parks
High fishing pressure over the last 100 years and technological development of the fisheries since the 1950s have played an important role in driving the decline of fish populations in the Oslo fjord and inner Skagerrak. Fisheries management not adapted to local conditions and local population dynamics caused serial depletion of local sub-stocks, where loss of older and larger individuals of top-predator species have resulted in a present-day fish assemblage dominated by smaller meso-predator species.
Coastal Atlantic cod in Skagerrak consist of two coexisting ecotypes. Young cod of both ecotypes were found throughout the study area in outer Oslo fjord, but very few individuals larger than the minimal legal size limit (40 cm) were observed by any method (≈ 5%). The 2016 year-class of cod was monitored throughout the project period. Annual apparent survival from 1- to 2-year old cod was estimated at 11 and 28% for the “North Sea”- and “fjord” ecotype (thus named based on previous molecular work), respectively. Findings indicate that both ecotypes recruit to coastal areas in outer Oslo fjord, and that both ecotypes should be considered important in efforts to rebuild populations in Skagerrak. A strong year-class of cod recruited to the study area in 2019.
Population genetics work indicate that sprat sampled in the Oslo fjord are separated from samples collected in Skagerrak and Kattegat. Revised management should assume that fjord sprat is stationary and self-recruiting, and thus vulnerable to harvesting. Rebuilding of fjord sprat and restoration of the species’ ecological function must be based on limiting fishing mortality within the fjord system.
Harbour seals and cormorants are native to Skagerrak and integral to an intact marine ecosystem. Both species are opportunistic predators with diets reflecting the available fish species assemblage. There is no data available to suggest that seals and cormorants are important drivers of the observed decline in fish populations compared to the effects of fisheries, natural mortality and reductions of suitable habitat. Harbour seals tagged with GPS-devices in outer Oslo fjord showed high mobility – a likely explanation for seals being more abundant during certain periods - in some areas - than suggested by IMR census counts (performed in August).
Parts of the Oslo fjord have poor environmental conditions, mainly due to effects of run-off, eutrophication and oxygen depletion. Essential habitat is damaged, disturbed or reduced due to chronic bottom-trawling, darkening of surface water and increased temperature. However, despite pending challenges that require action, there is little reason to believe that the outer Oslo fjord has entered a state where it is no longer suitable as habitat for cod and other depleted fish species. The best available science indicate that local population dynamics is the dominant pattern, suggesting that local management actions would be worthwhile.
Recommendations for management action build upon existing science and new data collected within the framework of the present project and the reports «Oppdatert kunnskapsstatus om kystnær torsk i Sør-Norge» (IMR) og «Forslag til helhetlig plan for Oslofjorden» (Mdir). The agriculture sector and cities/ municipalities must reduce their loadings to waterways in the catchment area draining into the Oslo fjord. Management of fisheries and life below water should shift to a place-based, adaptive strategy to ensure that harvest does not exceed productivity, and that essential ecosystem function is restored through rebuilding of broad age- and size structure in fish predator populations. Coastal development (coastal hardening) should be arrested, pollution should be eliminated, and existing polluted sediments should be rehabilitated according to best practice.
Forord
Ytre Oslofjord har en unik kystnatur av svært stor verdi for både fastboende og tilreisende. Færder- og Ytre Hvaler nasjonalparker ligger side om side ytterst i fjorden, og skal sikre naturverdiene for kommende generasjoner. Nasjonalparkene har utformet egne forvaltningsplaner og -mål, som gjenspeiler et ønske om å legge best mulig til rette for at fiskearter, fugler, planter og insekter skal fortsette å trives innenfor parkenes grenser. Generelle mål for naturverdiene er at de skal opprettholdes med god økologisk funksjon, og et høyt biologisk mangfold.
Det er viktig å sikre dette for kommende generasjoner, herunder flere fiskebestander som er sterkt redusert de siste tiårene. Et av forvaltningsmålene til nasjonalparkene er at bestanden av lokal kystnær torsk skal økes. Dette er bakgrunnen for prosjektet «Krafttak for kysttorsken», en felles satsning der følgende har vært med: Havforskningsinstituttet, Færder- og Ytre Hvaler nasjonalparker, Vestfold/Telemark- og Viken fylkeskommuner, Fiskeridirektoratet, Miljødirektoratet og representanter fra fiskernes organisasjoner.
Målet for prosjektet har vært:
å oppnå økt kunnskap om fiskesamfunnet og økosystemet gjennom ny forskningsaktivitet igangsatt av prosjektet og
peke på tiltak for om mulig å restaurere torskebestanden i fjorden.
Prosjektet har vært sektorovergripende med nært samarbeid mellom forskning og forvaltning. Bjørn Strandli har vært koordinator for den samlede aktiviteten i prosjektet og sekretariatet for styringsgruppen. I tillegg har følgende personer fra forvaltningen deltatt i arbeidet i sekretariatet og gitt verdifulle bidrag til prosjektet: Monika Olsen, nasjonalparkforvalter i Ytre Hvaler nasjonalpark; Atle Haga, fylkesbiolog i Viken fylkeskommune; Lars Wilhelm Solheim, seniorrådgiver i Vestfold og Telemark fylkeskommune.
Styringsgruppen vil takke alle som har deltatt i prosjektet for stor og verdifull innsats.
Havforskningsinstituttet står faglig ansvarlig for innholdet i sluttrapporten.
Styringsgruppe: Roar Jonstang, styreleder; Eivind Norman Borge (2017-19); Mona Vauger (2019-20); Elisabeth Rosendal (2017-18); Egil Postmyr (2018-20); Gunnstein Bakke; Andreas Lervik (2017-19); Anne Beathe K. Tvinnereim (2020); Aina Dahl (2017-19); Anne Lise Nilsson (2020); Thorbjørn Halvorsen; Olaf Thon (2017-18) og Kjetil Andersen (2019-20).
Prosjektledelse: Even Moland og Bjørn Strandli
24.mars 2021: Oppdaterte feil i referanselisten.
01.september 2021: Oppdatert tabell 3.3.6
Sammendrag
Høyt fiskepress gjennom de siste 100 år og teknologisk utvikling av fiskeriene siden 1950 er en viktig årsak til at flere fiskearter har hatt sterk tilbakegang i Oslofjorden og indre Skagerrak. Fravær av stedstilpasset fiskeriforvaltning har ført til gradvis utfisking av lokale bestander, der tap av eldre og større individer av toppredatorer har resultert i et fiskesamfunn som domineres av mindre arter. Fraværet av større individer av fiskespisende toppredatorer er tydelig.
Kystnær torsk i Skagerrak består av to sameksisterende genetiske varianter, kalt «økotyper». Unge torsk av begge økotyper ble funnet i hele studieområdet, men svært få individer større enn minstemålet (40 cm) ble observert (≈ 5%). 2016-årsklassen av torsk ble fulgt gjennom prosjektet. Overlevelse til torsk fra ett- til to-års alder ble beregnet til henholdsvis 11% og 28% for «Nordsjø»- og «fjord»-økotypen. Resultatene viser at begge økotyper rekrutterer til- og vokser opp i kystområdene i ytre Oslofjord, og at begge er viktige for restaurering av kystnær torsk i Skagerrak. En sterk årsklasse av torsk rekrutterte til studieområdet i 2019.
Genetisk kartlegging av populasjonsstruktur viser at brisling i Oslofjorden er genetisk forskjellig fra prøver hentet fra Skagerrak og Kattegat. En revidert forvaltning av brisling må gå ut fra at fjordbestanden er stedegen og derfor sårbar. Gjenoppbygging av denne bestanden og gjenoppretting av artens økologiske funksjon vil måtte baseres på begrensing av fiskedødelighet i selve fjordsystemet.
Både sel og skarv er naturlig hjemmehørende arter i et intakt marint økosystem. Artene er opportunister og spiser de fiskeartene det er mest av og som er lettest tilgjengelig. Sett i forhold til fiskeriene, annen naturlig dødelighet og reduksjon av leveområdene, har vi ikke data som tilsier at skarv og sel er viktige årsaker til torsk og andre fiskearters tilbakegang i Oslofjorden. Merkede steinkobber i ytre Oslofjord har høy mobilitet som er en sannsynlig forklaring på at det i perioder kan være flere steinkobber i noen områder enn hva Havforskningsinstituttets tellinger i hårfellingstida (august) viser.
Deler av Oslofjorden har til dels store miljøutfordringer med hensyn på avrenning, næringssaltbelastning og oksygenforhold i bunnvannet. Viktige habitater er svekket gjennom utbredt og langvarig bunntråling, endret lysgjennomtrenging ned i vannmassene og temperaturøkning. Samtidig er det ikke grunn til å hevde at det ikke lenger finnes livsgrunnlag for torsk og andre sterkt reduserte fiskebestander i ytre Oslofjord. Beste tilgjengelige kunnskap tilsier at lokal bestandsdynamikk er et fremtredende mønster.
Anbefalte tiltak bygger på evaluering av eksisterende data og ny kunnskap som har framkommet gjennom prosjektet, samt rapportene «Oppdatert kunnskapsstatus om kystnær torsk i Sør-Norge» (HI) og «Forslag til helhetlig plan for Oslofjorden» (Mdir). Landbruket og kommunene må redusere sine miljøskadelige utslipp til vassdragene på Østlandet og til Oslofjorden. Det er nødvendig med en mer aktiv stedtilpasset fiskeriforvaltning slik at beskatningen ikke overstiger produksjonsgrunnlaget, og at funksjoner i økosystemet gjenopprettes gjennom restaurering av bred alders- og størrelsessammensetning. Omfanget av tekniske inngrep og utbygginger må reduseres, utslipp av miljøgifter til fjorden må elimineres og eksisterende forekomster av miljøgifter i sjøbunnen bør saneres i henhold til beste tilgjengelig kunnskap.
1 - Oppdrag
Faglig innhold
Prosjektets oppdrag har vært å gi en oversikt over status for kystnær torsk i ytre Oslofjord, tilstanden for naturtyper som torsken er avhengig av, og økosystemet som den er en del av. Oppdraget omfattet også å gi råd om aktuelle tiltak for å restaurere kysttorsk og andre bestander av bunnfisk. Beskrivelsene fra prosjektet skulle bygge på beste tilgjengelige kunnskap, oppsummering av eksisterende data og data innhentet i forbindelse med gjennomføringen av prosjektet.
Forutsetningene i prosjektarbeidet har imidlertid endret seg vesentlig underveis. Dette skyldes at Stortinget, i et anmodningsvedtak datert 5. april 2018 ba regjeringen om å: «… legge fram en helhetlig plan for Oslofjorden – med mål om at fjorden skal oppnå god miljøtilstand, restaurere viktige naturverdier, fremme et aktivt friluftsliv og ivareta det biologiske mangfoldet i fjorden ». En innspillskonferanse ble arrangert sent i 2018 – der prosjektene «Krafttak for kysttorsken» og «Frisk Oslofjord» ble presentert. Tidlig i 2019 gav Klima- og miljødepartementet oppdraget til Miljødirektoratet om å lage forslag til en helhetlig plan for Oslofjorden. Denne ble lagt fram i desember 2019. «Forslag til helhetlig plan for Oslofjorden» (Miljødirektoratet 2019) er et stort arbeid utført i nært samarbeid med hele det norske fagmiljøet, både forskning og forvaltning, Havforskningsinstituttet inkludert. Med de ressursene som er tilgjengelig i Krafttak for kysttorsken, hadde det ikke vært mulig å lage en like grundig og helhetlig framstilling. Hensikten med å lage en særskilt analyse av miljøsituasjonen falt dermed bort.
I denne rapporten har vi derfor valgt å legge hovedvekt på beskrivelser som er relevante for fiskeriforvaltningen. I tillegg presenterer vi registeringer av miljøparametere i prosjektet med særlig relevans for vår vurdering av mulighetene for restaurering av økosystemet. Men for at leseren skal kunne danne seg et helhetsbilde av forholdene i fjorden har vi inkludert det brede og helhetlige kunnskapsgrunnlaget bak «Forslag til helhetlig plan for Oslofjorden» i vår oppsummering av hovedelementene som påvirker situasjonen i fjorden.
Sektorovergripende organisering
Initiativet til dette prosjektet ble tatt fra lokalsamfunnet knyttet til Færder- og Ytre Hvaler nasjonalparker i ytre Oslofjord. Prosjektet fikk en tydelig sektorovergripende og lokalt forankret organisering der følgende miljøer har vært involvert i prosjektet:
Færder- og Ytre Hvaler nasjonalparker
Havforskningsinstituttet
Fylkeskommunene på begge sider av fjorden
Fiskernes organisasjoner
Fiskeridirektoratet
Miljødirektoratet
Kommuner, fylkeskommuner, fylkesmenn, fiskeridirektoratet og miljødirektoratet sitter alle på virkemidler som er viktige i den marine forvaltningen. Ofte har virkemiddelbruken vært haltende fordi man de ikke har hatt et helhetlig kunnskapsgrunnlag og en felles forståelse rundt virkemiddelbruk.
Formålet med den sektorovergripende organisering har vært å skape forståelse for at samarbeid er nødvendig mellom de som leverer kunnskap, de forskjellige som påvirker fjorden og de som sitter med myndighet til å ta beslutninger om bruk av virkemidler. «Krafttak for kysttorsken» har hatt en sterk tro på viktigheten av samarbeid og forankring. Derfor kan også selve organisering av prosjektet ha stor overføringsverdi til andre prosjekter og til andre deler av kysten.
2 - Arter, naturtyper og økosystem
For å kunne overvåke og dokumentere effekter av forvaltningstiltak som marine bevaringsområder, eller andre fremtidige forvaltningsmessige- og/ eller miljøforbedrende tiltak, er det essensielt å samle inn tallmateriale som kan gi god informasjon om målarten/ -artenes tilstand. Kysttorsk er naturlig nok en målart i dette prosjektet, men i tråd med økosystemperspektivet i prosjektet har det vært nødvendig også å følge opp andre arter, samt utvikle metoder for å overvåke eventuelle endringer i det biologiske mangfoldet. Prosjektets datainnsamling støtter seg i hovedsak på en «før-etter-kontroll-tiltak» tilnærming (se Moland m.fl., 2013), med undersøkelser gjennomført før tiltak, med tanke på videreføring av sammenlignende datainnsamling i nær fremtid (etter tiltak).
2.1 - Fiskesamfunnet
I tillegg til eksisterende aktiviteter i ytre Oslofjord (overvåking av vannmiljø, strandnotserien, overvåking av bestandsutviklingen i bevaringsområder for hummer), ble ny datainnsamling gjennomført av Havforskningsinstituttet i tråd med målsetningene for «Krafttak for kysttorsken» (heretter: prosjektet). Flødevigens strandnotserie (Lekve m.fl. 1999, Barcelo m.fl. 2016) er utvidet til å inkludere 10 nye stasjoner i Færder nasjonalpark, og 10 nye stasjoner i Ytre Hvaler nasjonalpark i forbindelse med prosjektet (fra og med høsten 2017).
Datainnsamling for kombinert inventering av fiskesamfunnet og detaljert kartlegging av torskebestandene i begge nasjonalparker ble gjennomført i form av tokt i mai 2017-2019. Ruser ble benyttet på grunt vann (1–10 m) og fisketeiner/ havteiner ble brukt på dypere vann (10–30 m). Til sammen gjennomførte Havforskningsinstituttet henholdsvis 929 og 108 ruse- og teinedøgn i begge nasjonalparker og tilgrensende områder. Agnet stereovideo (se under) ble innført som ny observasjonsmetode for kartlegging av fiskesamfunnet, og et eget videotokt ble gjennomført i august 2017-2019. Det ble totalt registrert 9733 individuelle fisk fordelt på 37 arter fra 19 familier i prosjektperioden. Fiskesamfunnet på grunne habitater undersøkt med åleruser var dominert av et utvalg leppefisker (bergnebb og grønngylt) og små, stasjonære arter av ikke-kommersiell betydning. På dypere habitat undersøkt med teiner utgjorde flyndrefamilien 87% av totalfangst. Stereo-video registrerte i tillegg til bergnebb innslag av mobile kommersielle rovfisk (torsk, sei og hvitting). Arter med høyest relativ forekomst og gjennomgående høy tetthet var bergnebb og grønngylt for ruse, sandflyndre for teine og bergnebb og torsk for stereo-video. Temporale forskjeller i sammensetning av fiskesamfunnet i perioden 2017-2019 kunne ikke påvises. Rapport fra prosjektets inventering av fiskesamfunnet, basert på alle benyttede metoder, foreligger i form av en masteroppgave (Skjæveland 2020) og en doktorgradsavhandling (Synnes 2020) ved Universitetet i Agder.
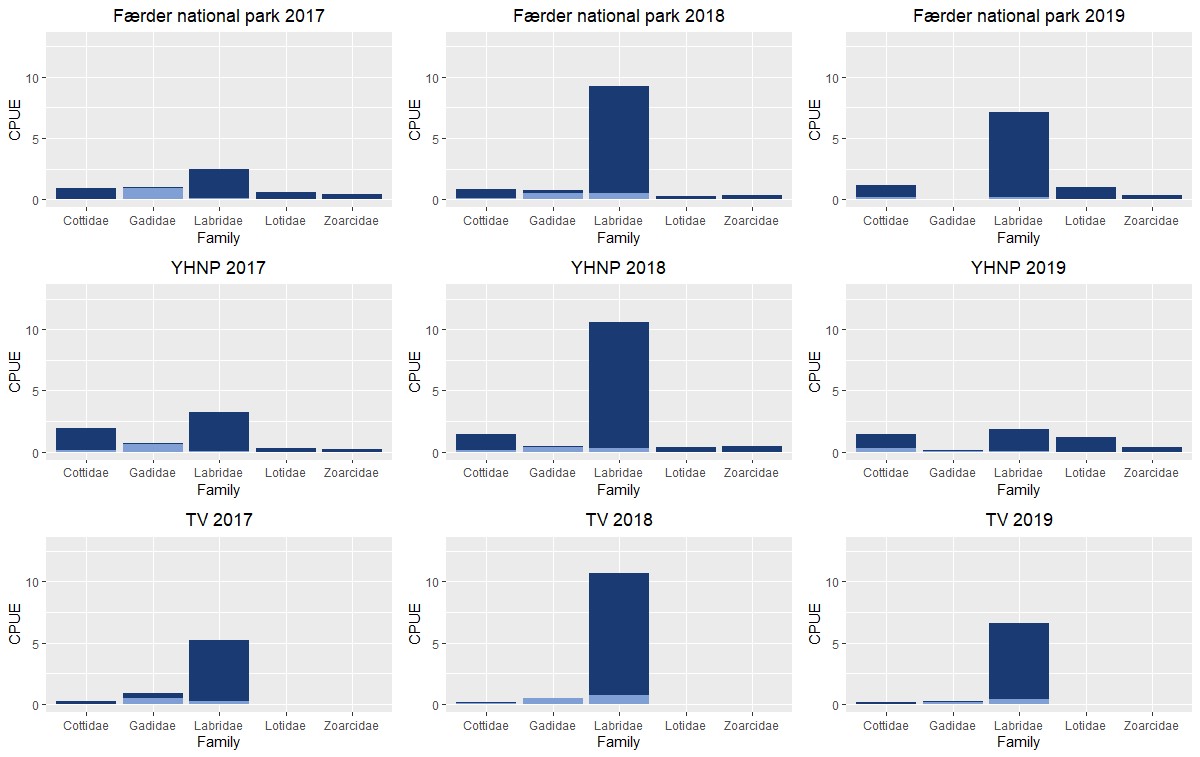
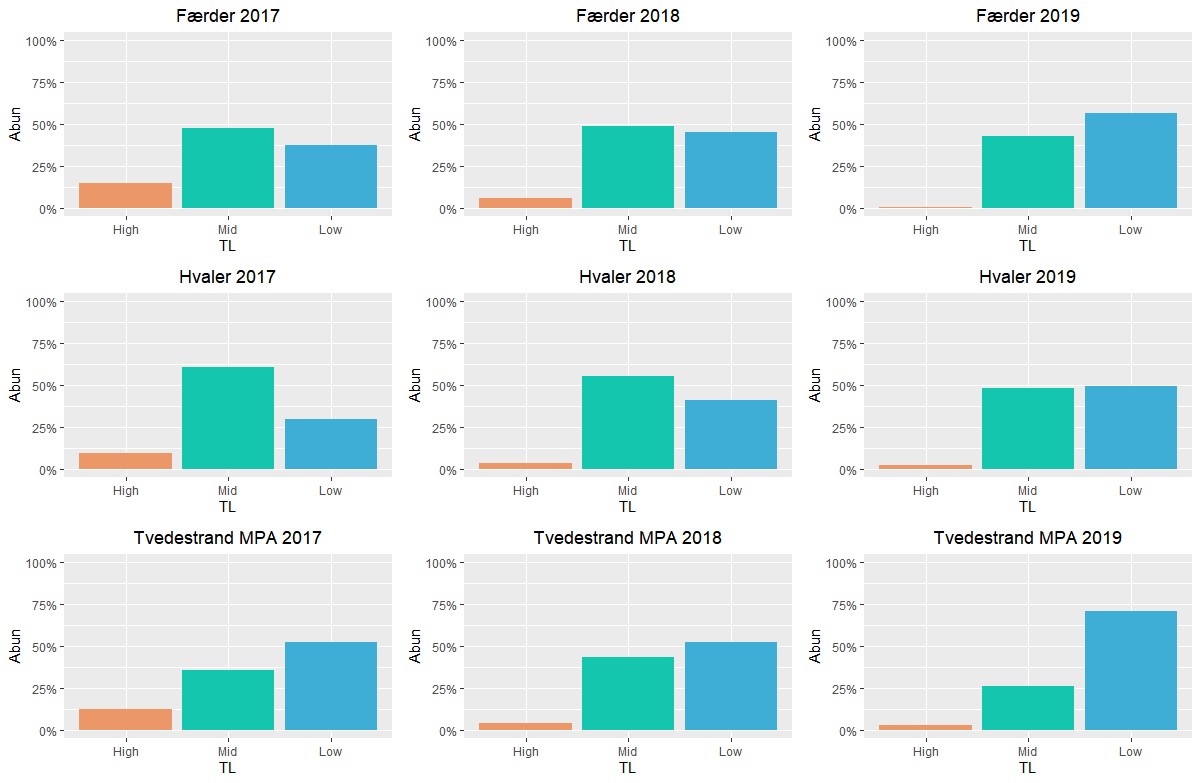
Fiskesamfunnet på grunt vann i ytre Oslofjord karakteriseres av en rekke mindre arter, med noen forskjeller mellom vest- og østsiden av fjorden. Hovedinntrykket harmonerer med lignende undersøkelser gjennomført i Kosterhavet nasjonalpark (Kraufvelin m.fl. 2017). Fraværet av større arter og fiskespisende toppredatorer er tydelig (se Fig. 2.1.1 og 2.1.2) og bekrefter inntrykket som beskrives av interessegrupper tilknyttet prosjektet (se tekst om torsk under). Et fiskesamfunn dominert av «mesopredatorer» (mesopredator = fisk som hovedsakelig beiter små evertebrater) kan medføre overbeiting på snegler og krepsdyr som igjen kontrollerer trådformede hurtigvoksende alger. Overfiske av rovfisk/ toppredatorer kan dermed gi inntrykk av et eutroft (overgjødslet) økosystem (Baden m.fl. 2010, 2012, Östmann m.fl. 2016). Vanlig ulke var svært tallrike i rusefangstene, særlig i Ytre Hvaler nasjonalpark og tilgrensende områder. Vanlig ulke er trolig en viktig predator på yngelstadiet til torsk og andre torskefisker (se Pedersen m.fl. 2020). Høy tetthet av denne arten i bunnslåings- og oppvekstområder for kystnær torsk er en sannsynlig kilde til en andel av naturlig dødelighet hos nylig bunnslåtte yngel av torskefisker i ytre Oslofjord, i tillegg til annen predasjon og svekkelse i perioder der næringsrike dyreplankton er mindre tilgjengelig som mat for pelagiske fiskelarver (se f.eks. Johannessen m.fl. 2012).
Figur 2.1.1 Fiskesamfunnet i ytre Oslofjord sammenlignet med tilsvarende undersøkelser i Tvedestrand (modifisert fra Synnes 2020). Fangst-per-enhet-innsasts (CPUE) av tallmessig dominerende fiskefamilier (ulker = Cottidae; torskefisker = Gadidae; leppefisk = Labridae; brosmefamilien = Lotidae; ålekvabbefamilien = Zoarchidae) samplet med ruser og fisketeiner (mai 2017, 2018 og 2019) i Færder- og Ytre Hvaler nasjonalparker og tilstøtende områder. Mørk blå = adulte stadier, lys blå = juvenile stadier. Høy temperatur i 2018 bidro til høy fangbarhet for leppefiskartene bergnebb (C. rupestris) og grønngylt (S. melops).
Figur 2.1.2 Fiskesamfunnet i ytre Oslofjord sammenlignet med tilsvarende undersøkelser i Tvedestrand (modifisert fra Synnes 2020). Tetthet (i %) av fiskearter samplet med ruser og fisketeiner (i mai 2017, 2018 og 2019) i Færder- og Ytre Hvaler nasjonalparker og tilstøtende områder, karakterisert i henhold til tre trofiske nivåer: «høyt» (= fiskespisende toppredatorer, oransje); «middels» (= beitere på større evertebrater, grønn); og «lav» (= beitere på mindre evertebrater, blå).
2.2 - Kystnær torsk i ytre Oslofjord
Kystnær torsk i indre Skagerrak og ytre Oslofjord består av lokale bestander tilhørende to økotyper (økotype = genetisk variant av art tilpasset lokale miljøforhold); en «fjord»- og en «Nordsjø»-økotype. De to typene er navngitt basert på tidligere populasjonsgenetiske funn som viser at «fjord»-økotypen forekommer hyppigere i indre områder av Skagerrak, mens «Nordsjø»-økotypen er mer vanlig i ytre områder. Nyere forskning viser at de to økotypene har stabil sameksistens i Skagerrak (Knutsen m.fl. 2018), og at ulik adferd og strukturelle forskjeller på genomet samvirker til å holde de to typene adskilt (Sodeland m.fl. 2016, Barth m.fl. 2019). Først og fremst har forskjellene – som stammer fra en hendelse der bestander av torsk var fysisk adskilt over tid – lagt grunnlag for at lokale tilpasninger oppstår og vedvarer. En bestandsstruktur preget av lokale bestander og lokale tilpasninger er sårbar for lokal utfisking og lokal utryddelse (se Cardinale m.fl. 2017).
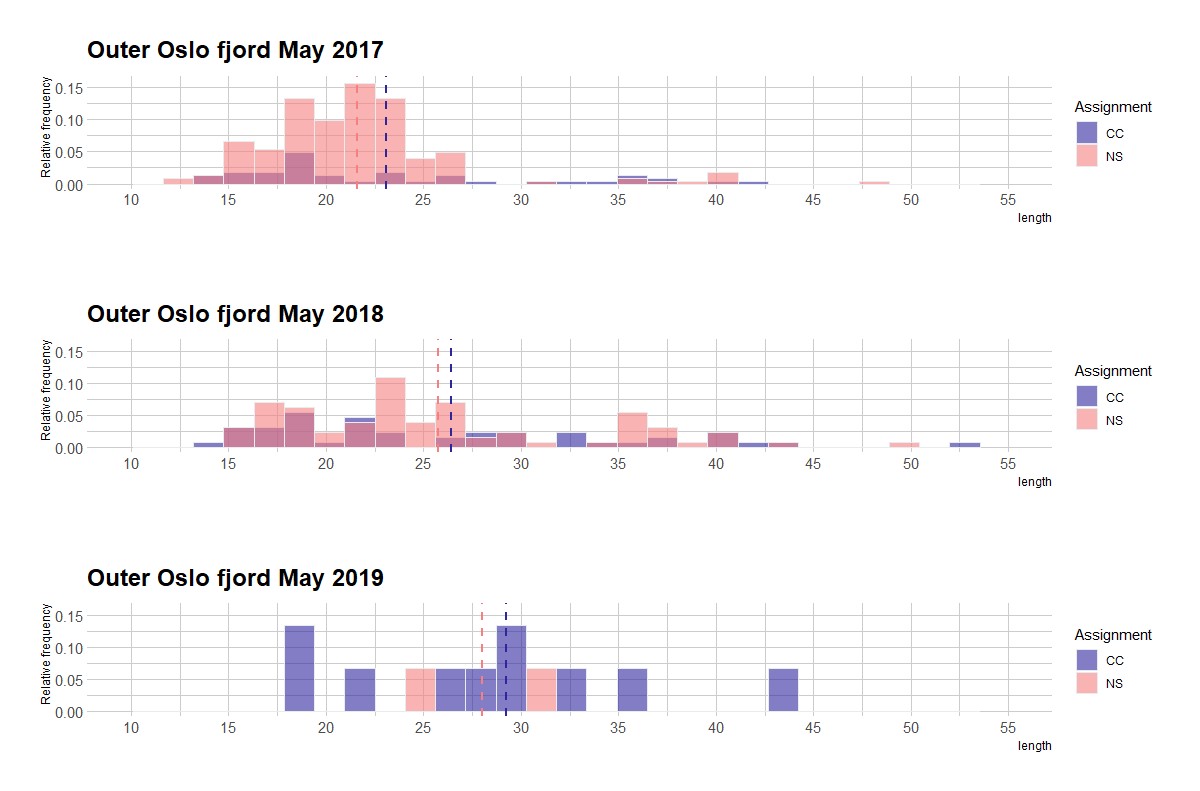
Figur 2.2.1 Lengdefordeling til torsk fanget i ruser og teiner under toktene gjennomført i mai 2017-2019. Blå = «fjord»-økotype», rosa = «Nordsjø»-økotype. Lilla = overlapp i lengdegruppe mellom de to økotypene. Svært få individer over minstemålet for torsk (>40 cm, eldre enn ca. 2 år) ble observert i alle redskapstyper i løpet av prosjektperioden (modifisert fra Synnes 2020).
Det ble funnet torsk av «fjord»- (N = 106) og «Nordsjø»- økotype (N = 262) på begge sider av fjorden i mai-toktene gjennomført i prosjektperioden (Fig. 2.2.1 og 2.2.2). I tillegg ble tallrike «0-gruppe»-torsk (årets rekrutter) funnet på strandnotstasjonene i ytre Oslofjord høsten 2017 og 2019, og i et fokusert prøvefiske i samarbeid med leppefisk-fiskere sommeren 2018 (Synnes 2020). Den geografiske fordelingen viser at det rekrutterer kystnær torsk til oppvekstområder på grunt vann i begge nasjonalparkene og tilstøtende områder. Begge økotyper ble altså funnet som fullt rekrutterte «1-gruppe»-, og som eldre torsk (>1 år) i hele prosjektperioden (2017-2019), med stor variabilitet mellom år. Svært få individer av torsk større enn minstemålet i fritidsfisket (40 cm) ble observert i fangstredskapene (N = 20, 5.4 % av totalt antall). Tilsynelatende årlig overlevelse (= reell overlevelse minus emigrasjon) for 2016-årsklassen av torsk (som ble fulgt gjennom prosjektet) fra 1- til 2-gruppe ble beregnet til henholdsvis 11% og 28% for «Nordsjø»- og «fjord»-økotypen (Synnes 2020). Prosjektets funn viser at begge økotyper rekrutterer til- og vokser opp i kystområdene i ytre Oslofjord, men at tilsynelatende årlig overlevelse er lav. Dette kan følge av at (1) fiskerier og/ eller andre predatorer fjerner begge økotyper effektivt, og/ eller (2) begge økotypene foretar vandringer knyttet til næringssøk, vekst og utvikling (ontogenetisk skifte i leveområde) vekk fra de undersøkte områdene, for eksempel til dypere leveområder. En kombinasjon av begge årsaker til svært lav «tilsynelatende overlevelse» er sannsynlig.
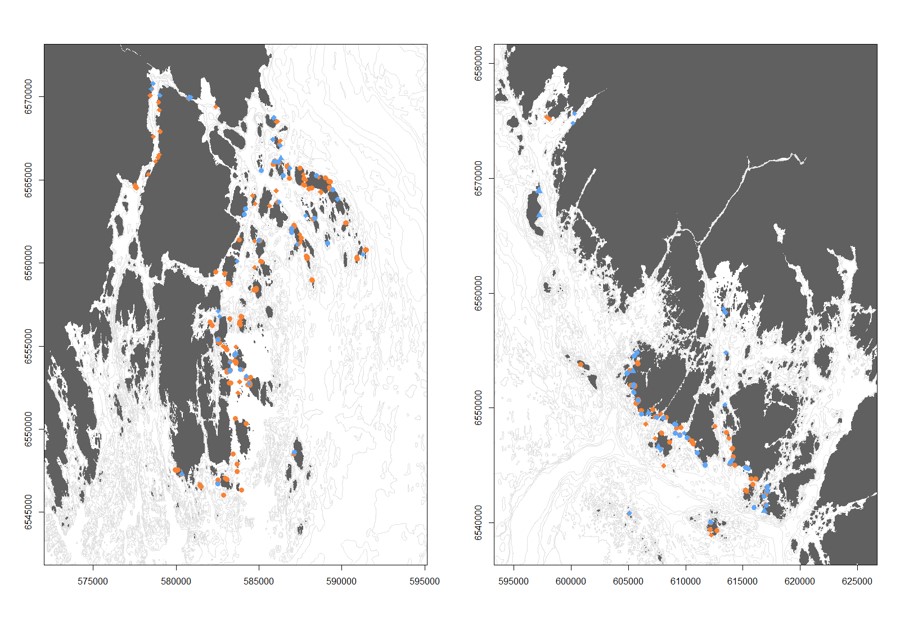
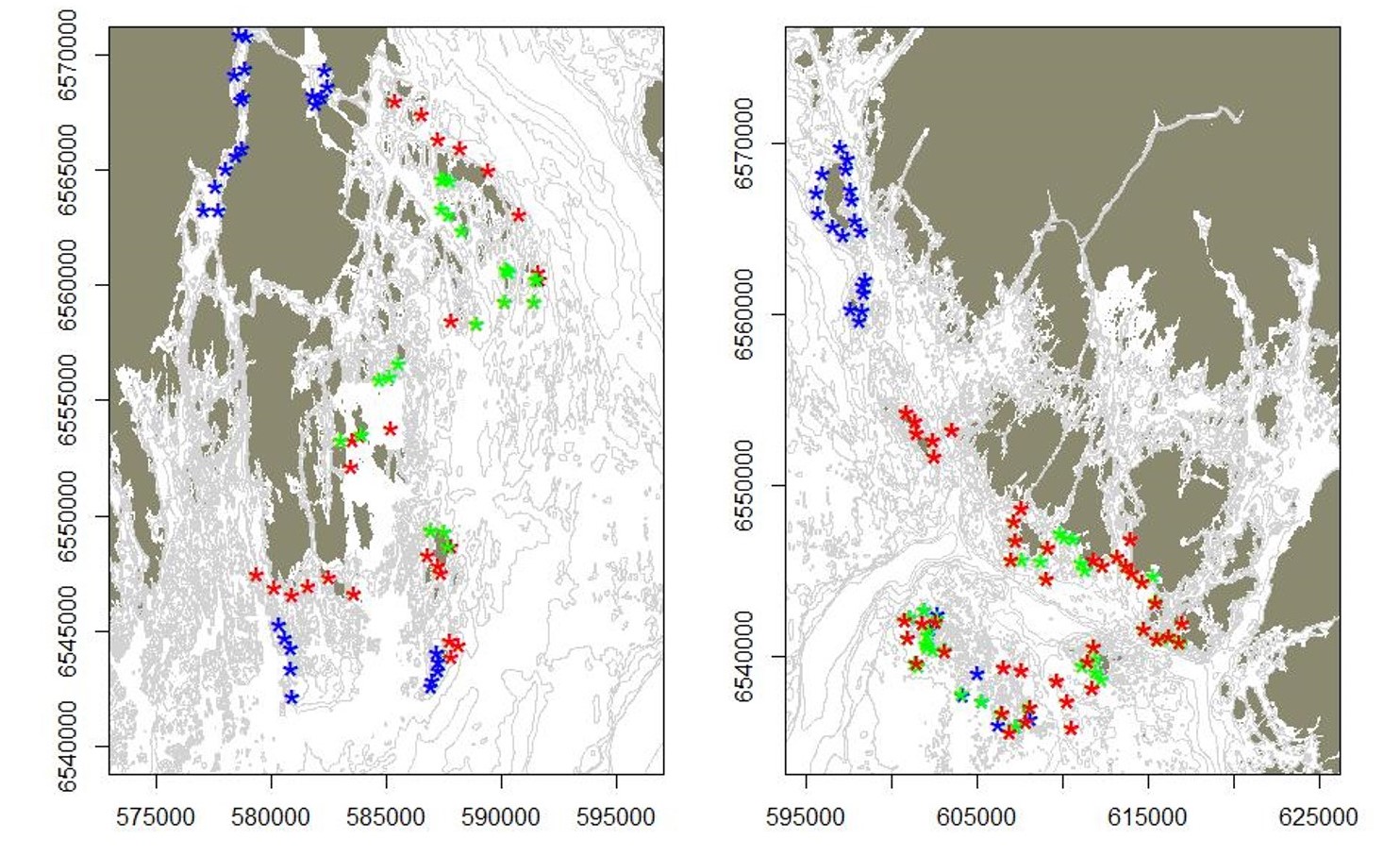
Figur 2.2.2 Økotype til torsk fanget i ruser og teiner i Færder- (venstre panel) og Ytre Hvaler (høyre panel) nasjonalparker under Havforskningsinstituttets tokt i mai 2017, 2018 og 2019. Røde symboler indikerer individer med økotype «Nordsjø», og blå symboler indikerer individer med økotype «fjord» basert på populasjonsgenetiske analyser.
Agnede stereovideorigger (Fig. 2.2.3) ble tatt i bruk i prosjektet under august-toktene 2017-2019 (se Fig. 2.2.4). Standard opptakstid er 60 minutter, der beregning av «fangst-per-enhet-innsats» gjøres basert på etablert analysemetodikk (Watson m.fl. 2005, Funk 2013). En fordel med stereo-metodikken er muligheten for lengdemåling av fisk fra bilder i opptakene (Langlois m.fl. 2012). Individstørrelsen for en art (eller størrelsesfordelingen til en bestand) kan dermed estimeres basert på et større antall individer enn hva som kan oppnås med fangstbasert metodikk.
Videoriggene har vist seg å være et godt alternativ til fangstbaserte metoder og agnet tiltrekker seg alle størrelsesgrupper av torsk (Fig. 2.2.3). Resultatene fra videoriggene bekrefter funnene i ruse- og teinefangster hvor svært få eldre torsk (> 1 år) er representert. Torsk over minstemålet på 40 cm var også fraværende i videostasjonene i 2019. Prosjektet har opprettet et omfattende stasjonsnett i ytre Oslofjord som representerer en basis/ grunnlinje for fremtidig overvåking.
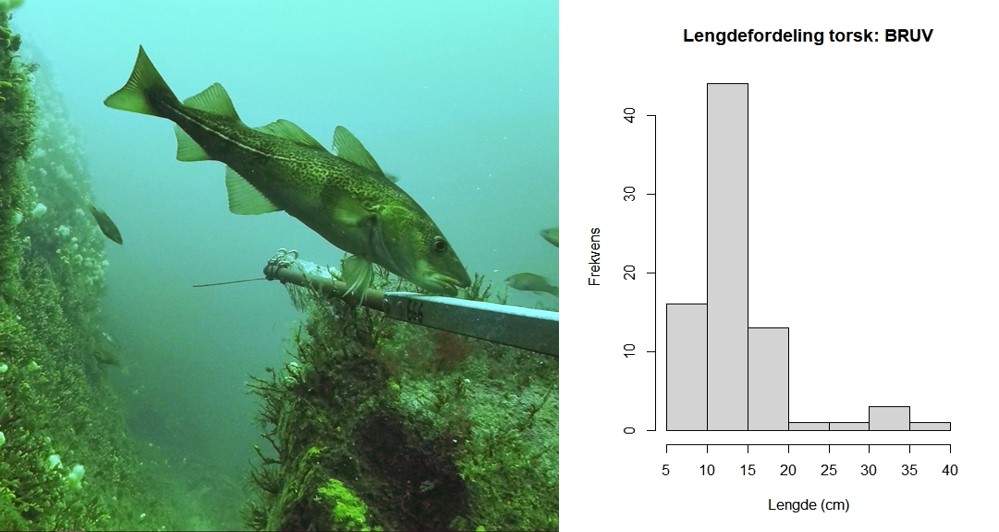
Figur 2.2.3 Venstre: eldre torsk (47 cm) ved Tjømebåen, august 2018 (22 m dyp). Høyre panel: lengdefordeling av torsk fra alle stereovideostasjoner i ytre Oslofjord august 2019. Fordelingen viser tydelig dominans av 2019-årsklassen («0-gruppe»), med individer fra 5-15 cm lengde. Lengdegrupper 15-30 cm antas å være 1-åringer (2018-årsklassen), lengdegrupper 30-40 cm antas å være 2-åringer (2017-årsklassen). Ingen torsk over minstemål i fritidsfisket (40 cm) ble observert med stereovideo i ytre Oslofjord i august 2019. Foto: E. Moland/ Havforskningsinstituttet.
Figur 2.2.4 Oversikt over stasjoner/ posisjoner for agnede stereovideo-rigger på vest- og østsiden av ytre Oslofjord under august-toktene i 2017 (røde symboler), 2018 (blå symboler) og 2019 (grønne symboler). Under toktet gjennomført i 2018 ble områder i Vestfjorden, Rauer og Misingene inkludert, samt at ytre områder (Tjømebåen, Tristeinene, Heia) ble prioritert. I 2019 ble repetisjon av tidligere besøkte stasjoner prioritert.
2.3 - Økosystemtilnærming
I tråd med begrunnelsen for Miljødirektoratets involvering i prosjektet, finansieringen fra Klima- og miljødepartementet, samt ambisjonen om å utvide prosjektet til et bredere og mer helhetlig økosystemperspektiv, ble det søkt om tilleggsfinansiering fra Regionalt forskningsfond (RFF) Oslofjordfondet i februar 2017. Ytterligere finansiering var nødvendig for å supplere aktiviteter i «Krafttak for kysttorsken», og for å kunne levere flere forskertimer i prosjektet. For å passe inn i utlysningen våren 2017 ble søknaden, og det omsøkte prosjektet, utformet som et «regionalt offentlig forskningsprosjekt» med Vestfold fylkeskommune som prosjekteier/ søker, med Østfold fylkeskommune, Færder- og Ytre Hvaler nasjonalparker som samarbeidspartnere, og med Havforskningsinstituttet som FoU-partner. RFF Oslofjordfondet bevilget i juni 2017 3 mill. kroner over 3 år til «Krafttak for kysttorsken». Hele søknadsbeløpet ble innvilget hvilket innebærer at ambisjonen om å jobbe bredere med økosystemet i ytre Oslofjord kunne realiseres. Oppstarten for denne nye delen av prosjektet ble satt til 1. januar 2018.
2.3.1 - Brisling
Brisling er en art som tidligere var grunnlag for et rikt fiskeri i Oslofjorden, med store fangster (≈2000 tonn årlig) gjennom 40-, 50- og 60-tallet. Det er sannsynlig at den betydelige historiske bestanden hadde en viktig funksjon i økosystemet, som omsetter av mindre dyreplankton til fiskebiomasse. De senere årene har kun mindre mengder brisling blitt høstet, hvor bare 139 tonn ble fanget av et enkelt fartøy i 2018. Dersom fjordbrisling rekrutterer fra lokale foreldre er det sannsynlig at overfiske er årsak til nedgangen og at sårbarheten til denne arten er blitt sterkt undervurdert.
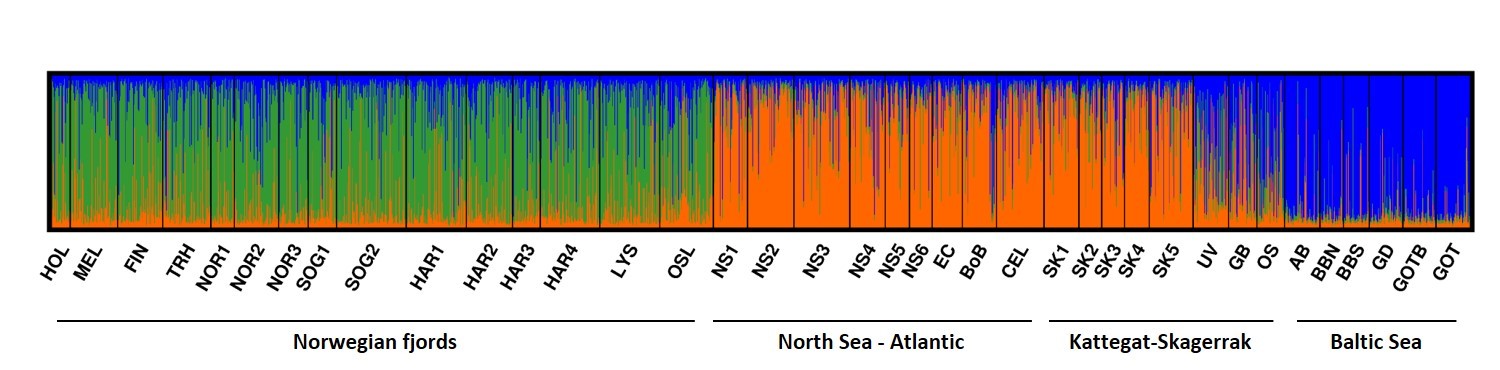
Havforskningsinstituttet har i prosjektperioden hatt et gjennombrudd i genetisk kartlegging av brisling og beskrivelse av populasjonsstruktur hos brisling (Quintela m.fl. 2020). Hovedfunnet i dette arbeidet, for Oslofjordens vedkommende, er at brisling funnet her er genetisk forskjellig fra brisling i Skagerrak og Kattegat (Fig. 2.3.1.1). En revidert forvaltning av brisling må gå utfra at fjordbestanden er stedegen og derfor sårbar. Gjenoppbygging av denne lokale bestanden vil måtte baseres på lavere uttak i fisket etter brisling og begrensing av fiskedødelighet i selve fjordsystemet.
Figur 2.3.1.1 Genetiske sammenligninger av brislingbestander viser at norske fjordbestander er delvis isolerte fra nabobestander i Nordsjøen, Atlanterhavet, Kattegat-Skagerrak og Østersjøen. Prøver fra Oslofjorden (OSL) tyder på større slektskap/ likhet med andre fjordbestander enn nabobestander i Skagerrak-Kattegat-området (kilde: Quintela m.fl. 2020).
2.3.2 - Kystsel
Figur 2.3.2.1 Steinkobbe (Phoca vitulina) trives i ytre Oslofjord. GPS-merking av sel ved Bolærne viste at dyrene bruker hvilesteder på begge sider av fjorden, med bevegelser fra Hurum til Väderöarna i Sverige. Foto: E. Moland/ Havforskningsinstituttet.
Sjøpattedyr er viktige komponenter i økosystemet. Fiskebestandene i ytre Oslofjord og østlige Skagerrak har, med noen få unntak, vist en negativ trend de siste tiårene. Det var nødvendig å inkludere steinkobbe i ytre Oslofjord fordi det er blitt en ganske vanlig oppfatning at arten er en betydelig konkurrent til mennesket om fiskeressursene, og en mulig årsak til torskebestandens tilbakegang i østlige deler av Skagerrak. Diettstudier utført i ytre Oslofjord på 90-tallet, og mer nylig i Agder og Telemark (Sørlie m.fl. 2020) viser imidlertid en svært sammensatt diett der selen spiser alle de fiskearter som er vanlig og lett tilgjengelig .
Steinkobbene trives i ytre Oslofjord, og dyr som observeres i områdene som inkluderer Færder- og Ytre Hvaler nasjonalparker er antatt å tilhøre ulike kolonier. Steinkobbe blir regnet for å være relativt stasjonære dyr. De er knyttet til områdene hvor de føder unger i siste halvdel av juni, dier i juli og har hårfelling i august. Resten av året sprer de seg noe mer ut for å finne føde, men de har likevel ikke regulære langdistansevandringer som flere andre arter sjøpattedyr.
Tellinger av steinkobbe kan tyde på at bestanden har økt i Vestfold fra fra1996-99 fram til i dag, men det er usikkerhet om bestandsstørrelsen fordi det kun ble gjennomført tellinger i deler av fylket 1996-99. I Vestfold ble det i 2014 og 2016 registrert henholdsvis 181 og 292 steinkobber i tellinger som dekket hele fylket. I Hvaler var det omkring 300 steinkobber i 1999-2000, mens det i 2001 ble det registrert 548 steinkobber. Det høye antallet i 2001 kan tyde på innvandring fra de større bestandene i svensk og dansk Skagerrak-Kattegat (ca. 25 000 dyr). I 2002 oppstod det en virusinfeksjon (PDV) hos steinkobbe i disse områdene, noe som førte til stor dødelighet i bestandene, også i norsk Skagerrak. Det tok 14-15 år før bestanden i Hvaler var tilbake på samme nivå (± 300 dyr) som rundt årtusenskiftet. Gjenvekst i de bestandene av steinkobbe som ble sterkt redusert av «selpesten» i 1988 og 2002, viser at steinkobbene under gode forhold kan ha en bestandsvekst på ca. 12% pr år. Informasjon om steinkobbers vandringsmønstre, diett og beiteområder er viktig for å evaluere påvirkning av sel på fiskebestandene i ytre Oslofjord, samt for å forklare selens rolle i økosystemet.
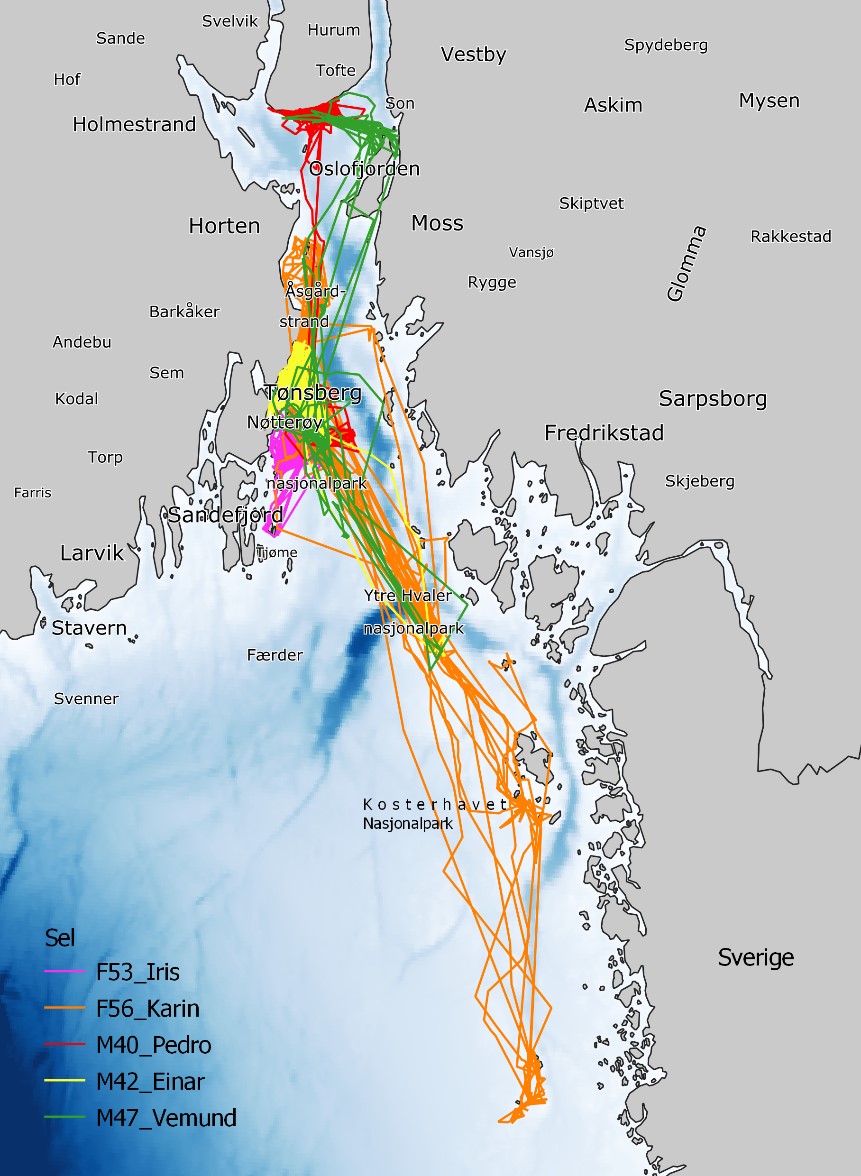
I november 2019 ble fem steinkobber fanget og merket ved Bolærne i Færder nasjonalpark. Disse dyrene besto av tre hanner og to hunner, som veide mellom 40 og 56 kg (Tabell 2.3.2.1). Merkene samlet GPS-posisjoner, dykkeprofiler og vanntemperatur mellom november 2019 og mars 2020. Foreløpige resultater viser at steinkobber i ytre Oslofjord er mer mobile enn tidligere antatt. Individer merket ved Bolærne har beveget seg hyppig mellom områder i Færder nasjonalpark, Ytre Hvaler nasjonalpark, samt Kosterhavet nasjonalpark og Väderöarna i Sverige (Fig. 2.3.2.2). Dette er svært relevant informasjon siden steinkobber forvaltes fylkesvis i Norge. Dessuten er steinkobbe i Norge og Sverige forvaltet som forskjellige bestander. Høy mobilitet hos steinkobber, som merkede individer viser, er en sannsynlig forklaring på at det i perioder kan gis et inntrykk av at det er flere steinkobber enn hva tellinger viser i farvannene i ytre Oslofjord.
Tabell 2.3.2.1 Oppsummering av data fra fem steinkobber merket ved Vestre Bolæren i november 2019. F = hunnkjønn; M = hankjønn.
Sel-ID
Kjønn
Vekt (kg)
Lengde (cm)
Merket
Siste posisjon
Antall dager
Antall GPS- posisjoner
Maks dybde (m)
F53_Iris
F
53
120
14.11.2019
29.03.2020
136
4918
86
F56_Karin
F
56
125
14.11.2019
21.03.2020
128
2553
284
M40_Pedro
M
40
113
14.11.2019
12.03.2020
119
5104
194
M42_Einar
M
42
116
14.11.2019
28.02.2020
106
2750
133
M47_Vemund
M
47
122
14.11.2019
26.12.2019
42
866
175
Figur 2.3.2.2 GPS-spor fra fem steinkobber merket ved Vestre Bolæren 14. november 2019. Steinkobbene sendte data mellom november 2019 og mars 2020.
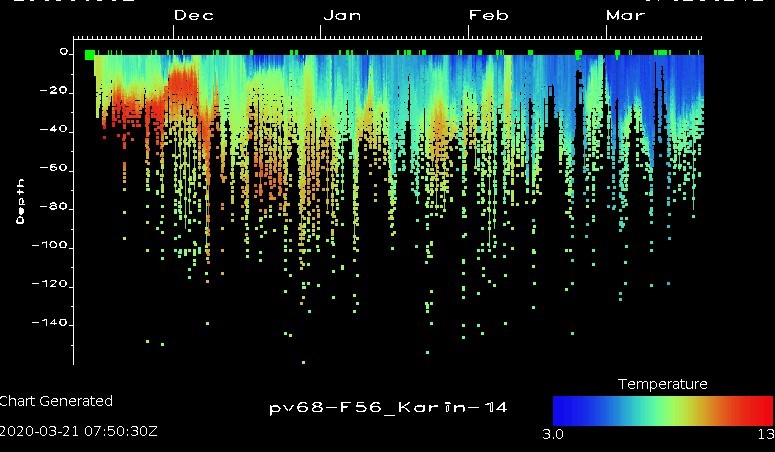
Data fra dybdemåleren i GPS-merkene viser at steinkobbene daglig dykker til 30 m og dypere (Fig. 2.3.2.3). Maksimal registrert dybde varierte mellom individer fra 86 m (F53_Iris) og 284 m (F56_Karin). Videre analyser skal undersøke hvilke områder og habitater er som er brukt når selene søker etter mat. Hvilepauser på land variert fra 11 minutter til 28 timer (gjennomsnitt 6 ± 0.5 timer). Varighet og tidspunkt av hvilepauser kan variere med miljøforhold (f.eks. vind, tidevann, vann- og lufttemperatur). Dette vil bli grundigere undersøkt i et masterprosjekt ved UiA.
Figur 2.3.2.3 Dybde- (m) og temperaturdata (°C) for steinkobben «F56_Karin» i løpet av 4 måneder etter merking.
Diettundersøkelser i ytre Oslofjord er gjennomført basert på analyser av selekskrement-prøver. Under feltarbeidet som ble gjennomført i forbindelse med fangst og merking ble 129 ekskrementprøver samlet fra holmer og skjær hvor steinkobber oppholder seg (77 fra Ytre Hvaler- og 52 fra Færder nasjonalpark). Flere prøver ble samlet høsten 2020. Prøvene ble frosset til senere identifisering av fiskeotolitter (øresteiner) i prøvene, samt genetiske analyser av byttedyr. Diettstudiet skal gjennomføres som en mastergradsoppgave ved Universitetet i Tromsø, i samarbeid med Universitet i Agder. Studiet er i gang og oppgaven forventes å bli levert i mai 2021.
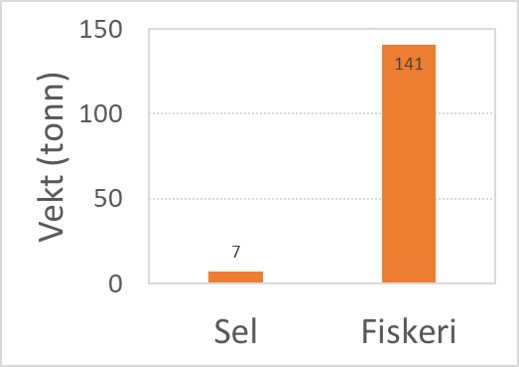
Et nylig publisert arbeid har undersøkt steinkobbens diett i tidligere Telemark- og Aust-Agder fylker. Resultater viser at torsk bare utgjør 2% av steinkobbens diett i området, mens de mest vanlige byttedyr var andre torskefisker som øyepål, sei, lyr og hyse, samt flatfisk (Sørlie m.fl. 2020). Videre ble det beregnet at steinkobbenes innhogg i torskebestanden utgjør ca. fem prosent sammenlignet med fangsten fra kommersielt fiskeri i indre Skagerrak (Fig. 2.3.2.4).
Figur 2.3.2.4 Årlig konsum av torsk fra steinkobber (5%) sammenlignet med årlig kommersielt fiskeri av torsk i samme område i tidligere Telemark- og Aust-Agder fylker 2016 (data fra Sørlie m.fl. 2020).
2.3.3 - Skarv
Figur 2.3.3.1 Den største kolonien av mellomskarv (Phalacrocorax carbo sinensis) i Norge har tilhold på Øra i Fredrikstad. I 2020 var hekkebestanden på 876 par. Foto: Per-Arne Johansen.
Så langt tilbake som vi har opplysninger, har storskarv (Phalacrocorax carbo) blitt observert vinterstid i Oslofjorden (Rasch 1838) og betydelige antall passerer våre områder i forbindelse med vår- og høsttrekk (Collett 1868). Denne torskarven tilhører en nordlig underart som hekker i Norge fra Boknafjorden til Finnmark. Også toppskarv, som hekker fra Rogaland til Finnmark, overvintrer i mindre antall i Oslofjorden.
Den andre underarten av storskarv (Phalacrocorax carbo sinensis), som også kalles mellomskarv (Fig. 2.3.3.1), har kommet sørfra og begynte å hekke i Norge første gang i Rogaland i 1996 og på Øra ved Fredrikstad i 1997 (Fredriksen og Johansen 1999). Senere har den bredt seg langs hele kysten mellom Østfold og Haugesund, med samlet 2500 hekkende par i 2012 (Lorentsen 2013). Etter dette har den norske hekkebestanden fortsatt å vokse og er i 2020 på i overkant av 3500 par ifølge Det nasjonale overvåkingsprogrammet for sjøfugl (Svein-Håkon Lorentsen pers. med.). Av disse finnes ca. 1130 par i Viken, 980 i Vestfold og Telemark, 1260 i Agder, og 200 i Rogaland. Kolonien på Øra er desidert størst på 876 par i 2020 (Per-Arne Johansen pers. med.).
Det er foretatt to undersøkelser av storskarvens fødevalg i den store hekkekolonien på Øra. Den første var i 2002 som hovedfagsoppgave ved Universitetet for Miljø- og Biovitenskap (UMB) (Skarprud 2003). Denne beskrev funn basert på 6716 otolitter (øresteiner) fra 240 gulpeboller, samt oppgulpet fisk. Grønngylt, svartkutling og bergnebb utgjorde 67.4% av næringsgrunnlaget. Andre arter som fantes i større antall var gressgylt, torsk, rødspette, brisling, skrubbe, vanlig ulke, ål og en rekke karpefisk. Totalt ble det funnet otolitter fra 26 fiskearter. Årlig totalt matkonsum for hekkekolonien ble anslått til 194-277 tonn.
I 2011 ble en ny masteroppgave gjennomført ved UMB på storskarvens sommerdiett på Øra, ved å undersøke i alt 6612 otolitter, fra 211 gulpeboller, samt oppgulpet fisk (Sørensen 2012). Minst 30 fiskearter var representert. Også denne gang utgjorde svarkutling, bergnebb, grønngylt og annen leppefisk over 50% av totalt antall fisk. Torsk utgjorde 8%, mens annen torskefisk herunder ubestemte arter utgjorde 30%. Totalkonsumet for kolonien i 2011 ble anslått til 205 – 293 tonn.
I 2017 ble en tilsvarende studie gjennomført i Aust-Agder, i Grimstad, Arendal og Tvedestrand, også denne gang som masteroppgave ved UMB (Gustavsen 2017). Her ble fødevalget påvist ved å analysere magesekken fra 84 skutte individer og oppgulpet fisk fra en stor hekkekoloni. Til sammen ble det funnet 354 fisk hvorav leppefiskarter og torsk utgjorde henholdsvis 35 og 8%. Majoriteten av fisk var mindre enn 100 gram.
Disse tre undersøkelsene samsvarer godt med et større svensk treårig arbeid på næringsvalget til storskarv ved Koster og samt to andre steder i hhv. Skagerrak (Hakefjorden) og Kattegatt (Bua) (Lunneryd og Alexandersson 2005). Totalt ble 59 fiskearter påvist. De viktigste artene var vanlig ulke (22-34%), flyndrefisk (10-36%), torsk (4-17%) og sei (0.8-13%). Tallmessig var dvergulke (Fig. 2.3.3.2) like hyppig i prøvene fra Koster. Dersom skarv medvirker til å kontrollere bestanden av ulker i ytre Oslofjord kan arten bidra til å promotere overlevelse til 0-gruppe torsk – ettersom ulkene er predatorer på juvenile torskefisker (se kapittel 2.1 Fiskesamfunnet og Pedersen m.fl. 2020).
Figur 2.3.3.2. Dvergulke (Taurulus bubalis) er tallrike i ytre Oslofjord og en viktig art i dietten til skarv ved Koster (jfr. Lunneryd og Alexandersson 2005). Foto: E. Moland/ Havforskningsinstituttet.
Lorentsen m.fl. (2021) oppsummerte fødevalget til storskarv i Norge og konkluderte med at arten er en opportunist som tar de fiskeartene som er lettest tilgjengelig nær hekkekoloniene. Majoriteten bestod av fisk opp til 100-150 mm lengde, dvs. for en stor del unge individer. Studien konkluderer derfor med at storskarv lokalt vil kunne påvirke rekruttering av voksen fisk, men påpeker at betydningen av dødelighet representert ved skarv sett forhold til andre kilder til naturlig dødelighet ikke er kjent.
Både sel og skarv er naturlig hjemmehørende arter i et intakt marint økosystem, og de hører hjemme i en frisk Oslofjord. Artene er opportunister og spiser de fiskeartene det er mest av og som er lettest tilgjengelig. Sett i forhold til både fiske, naturlig dødelighet og reduksjon av leveområdene, har vi ingen data som tilsier at skarv og sel er viktige årsaker til torsk og andre fiskearters tilbakegang. I sentrale deler av Østersjøen er en stor bestand av selarten havert (Halichoerus grypus) vurdert til å være en mindre viktig påvirker enn fiskerier og miljøvariasjon (Costalago m.fl. 2019). I moderne forvaltning hvor en økosystemtilnærming står sentralt vil en inndeling i «nyttige arter» som skal beskyttes, og «unyttige/ skadelige arter» som bør bekjempes virke utdatert. Den historiske innsatsen for å begrense Skandinaviske selbestander som skadedyr (i perioden 1889-1927) er senere blitt vurdert som lite nyttig fra et samfunnsøkonomisk perspektiv (Olsen m.fl. 2018). En litteraturgjennomgang av Lennox m.fl. (2018) fant få eksempler på at reduksjon av predatorer («culling») faktisk førte til bestandsøkning hos ønsket art. Likevel vil det være hensiktsmessig å overvåke bestander av sel og skarv da forvaltningen ønsker god informasjon om bestandsutviklingen, også som indikator for økosystemets helse. Det gjennomføres tellinger av steinkobbe under hårfellingsperoden (august) langs hele norskekysten, inkludert ytre Oslofjord. Det telles årlig i delområder av kysten, som ender opp i et totalt estimat for hele norskekysten (bortsett fra Svalbard) hvert femte år. Det gis jaktkvoter i alle områder hvor bestanden er tilstrekkelig stor, avhengig om bestanden er over målnivåene for hvert fylke. Samtidig er det verdt å påpeke at verdien av naturopplevelser knyttet til sel og skarv i frilufts- og turismesammenheng er viktig og trolig økende i framtiden.
2.3.4 - Habitater og interaksjoner
Ålegressenger, tang- og taresamfunn er oppvekstområder for kysttorsk og andre kystfisk, men habitatenes struktur og funksjon varierer og avhenger av miljøforholdene. I ytre Oslofjord finnes ålegrasenger i grunne bløtbunnsområder. Sukkertare dominerer middels eksponert kyst på hardbunn. Stortare med vekslende innslag av fingertare dominerer ytre kyst og tilstanden for stortare har vært god. I tidevannsonen vokser tangsamfunn i hele eksponeringsgradienten.
I «Krafttak for kysttorsken»-prosjektet og andre prosjekter Havforskningsinstituttet har gjennomført med aktiviteter i Bolærne-området har forekomst og tilstand for disse naturtypene vært undersøkt. Det har også vært behov for å utvikle metoder for å kvantifisere juvenil fisk. Både små teiner, videorigger, droner og akustikk (bunnstående ekkolodd) har vært testet for å kartlegge habitater som er oppvekstområder for fisk, deriblant kysttorsk. Habitater i området fra Tønsberg til Bolærne, som representer en gradient i bølgeeksponering og avrenning fra land har blitt overvåket 2018-2020. Undersøkelsene har vært gjort på stasjoner som ligger i transekter på stasjoner på bløt- og hardbunn fra indre kyst ved Tønsberg til ytre deler av Bolærne (Fig 2.3.4.1).
Figur 2.3.4.1 Stasjonskart for 10 Undervannsdrone-transekter ved Bolærne på dybder mellom 1-10 m i ulike habitat (hardbunn, ålegress, tangsamfunn, sandbunn og mudderbunn). Transektlengder fra 50-150 m.
Tilstanden for ålegressenger
Nedre voksegrense for ålegras kan gi informasjon om vannkvaliteten (lysforholdene) og endringer i disse. I 2018 ble nedre voksegrense fysisk avmerket med pinner i seks ålegressenger. Dette for å kunne følge år til år variasjon i dybdeutbredelse. Slike variasjoner kan oppstå ved f.eks. endrede lysforhold som følge av eutrofiering eller avrenning av partikler fra land. I 2019 ble 5 av stasjonene gjenbesøkt - to ytre, og 3 indre (som er mest påvirket av slike endringer). 17 av 25 pinner ble gjenfunnet (Fig. 2.3.4.2), og det ble ikke funnet noen endring i nedre utbredelse av ålegresset (usikkerhet på ± 5cm).
Figur 2.3.4.2 Markeringspinner for nedre voksegrense for ålegras, ett år etter markering. Foto: J. Thormar/ Havforskningsinstituttet.
I 9 ålegressenger har 4-16 ruter på 50 × 50 cm blitt avfotografert per eng. Tetthet av eng og påvekstalger ble klassifisert på en 4-trinns skala («enkeltskudd» til «heldekkende eng», og «lite» til «dominerende» mengde påvekstalger). Påvekstalger er hurtigvoksende alger som kan overgro og «kvele» ålegraset. Mengden av påvekstalger kan brukes som indikator på helsetilstanden til ålegraset (se Fig. 2.3.4.3).
Figur 2.3.4.3 Tetthet og mengde påvekstalger på ålegras i 2019. Størrelse på grønne sirkler angir tetthetsklasse fra enkeltskudd til heldekkende, og tykkelse på den røde ringen er klassifisering av påvekstalger fra lite til dominerende.
Tilstanden for sukkertare
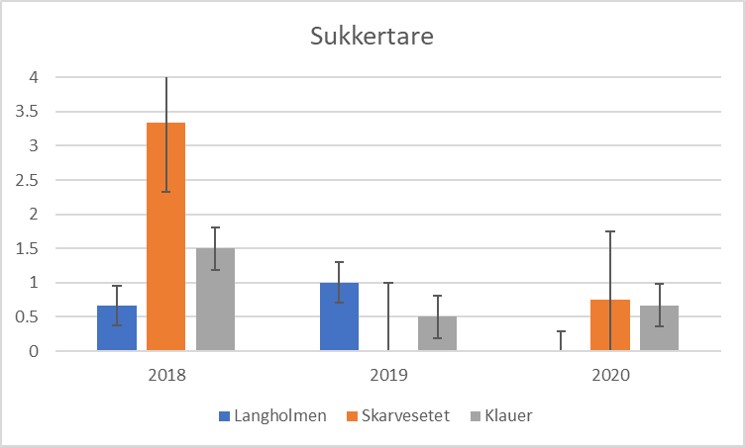
Tilstanden for sukkertare har vært varierende og i perioder svært dårlig i hele Skagerrak siden slutten av 1990-tallet. I 2018 viste Havforskningsinstituttet sammen med amerikanske kollegaer at hetebølger slår ut sukkertare og hurtigvoksende trådalger tar over bunnen (Filbee-Dexter m.fl. 2020). Også i ytre Oslofjord ble omfattende taredød registrert på tre stasjoner i Bolærne (Figur 2.3.4.4). En markant nedgang i antall sukkertare per 0.25 m2 ruter ble registrert i august 2018 sammenlignet med august 2019 og 2020. Endringen fra store flerårige tarer som representerer tredimensjonalt habitat med stor produksjon, til flate tepper av trådformede alger som samler sediment er betydelig, og teppene hindrer taren i å rekruttere. Algeteppene kan derfor hindre taren i å komme tilbake i lang tid etter at hetebølgen er over.
Figur 2.3.4.4 Foreløpige resultater. Antall sukkertare per 0.25 m2 ruter på Langholmen, Skarvesetet og Klauer i Bolærne, august 2018-2020.
Tilstanden for stortare
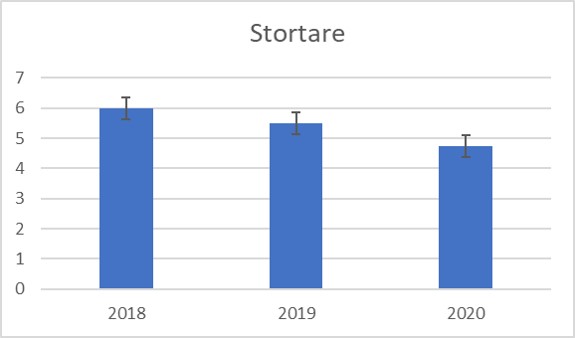
Nedre voksegrense for stortare kan si noe om vannkvaliteten (lysforholdene) og endringer i disse. Nedre voksegrense i ytre Oslofjord er redusert fra 25 m dyp i 1950 til 12-15 m i dag (se kapittel 3.4). Stortare står i tette skoger på ytre, bølgeeksponerte lokaliteter ved Bolærne. Tilstanden på ytre kyst har vært relativt uforandret i over tid i ytre Oslofjord, inkludert årene Krafttak for kysttorsken har blitt gjennomført. Det ble vurdert god tilstand for stortare på utsiden (østsiden) av Bolærne (Fig. 2.3.4.5).
Figur 2.3.4.5. Foreløpige resultater. Antall stortare per 0.25 m2 ruter på Langholmen, Skarvesetet og Klauer i Bolærne, august 2018-2020.
Transektundersøkelser og telling av fisk
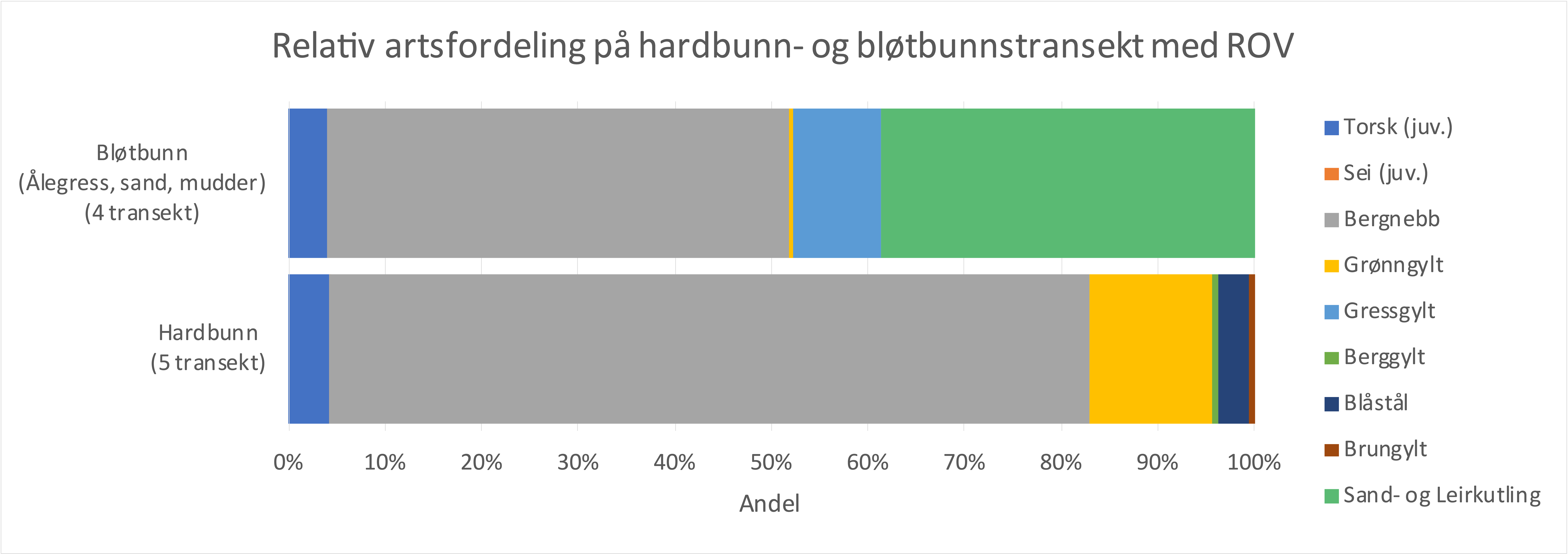
Fisk ble registrert langs dronetransektene og knyttet til bunntype. Foreløpig analyse av de 10 transektene viser relativt flere leppefisk på eksponert hardbunn med tang eller blandede makroalger, mens bløtbunntransektene i ålegress, sand og mudderbunn har større andel av gressgylt og kutling. Den relative fordelingen av juvenil torsk var relativ lav og lik på hard- og bløtbunn (Fig. 2.3.4.6).
Figur 2.3.4.6 Relativ artsfordeling av fisk på hard- og bløtbunn.
Bunnstående/ oppadrettet ekkolodd for registrering av fisk
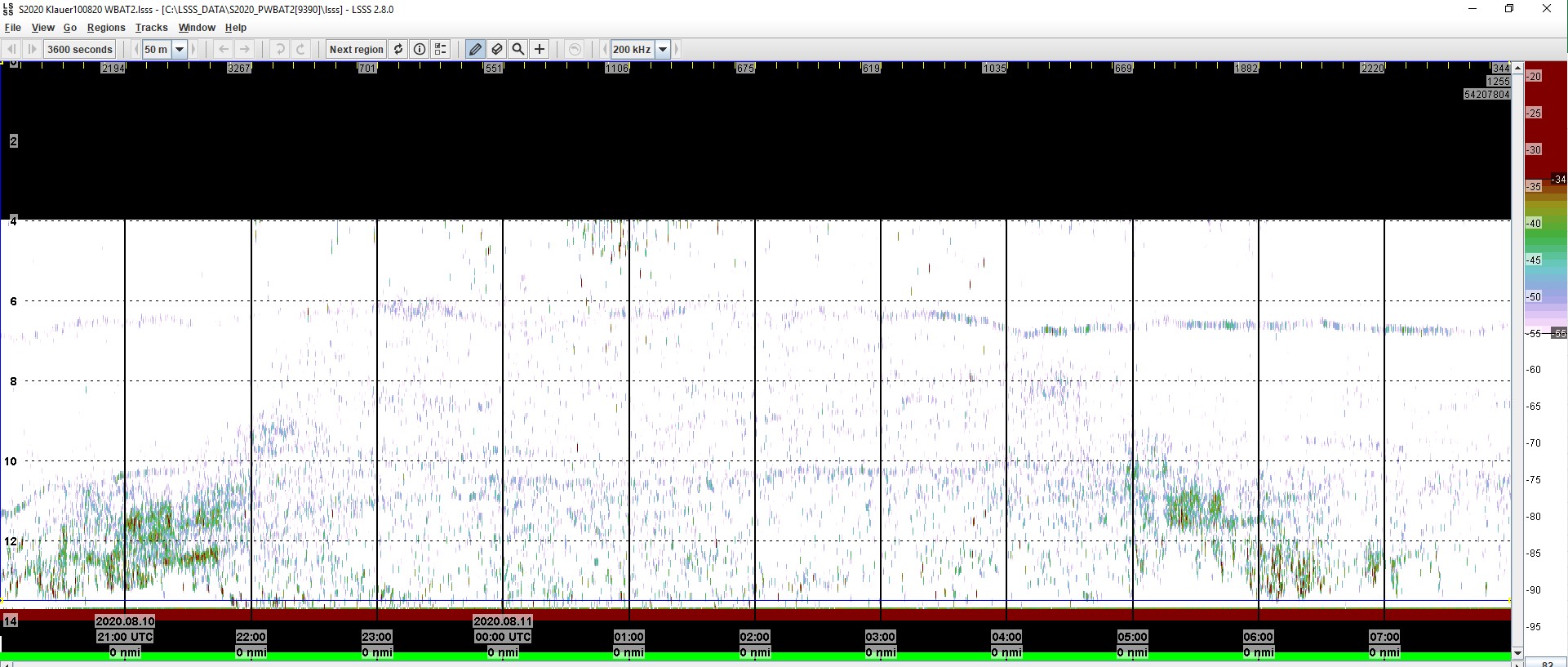
Visuelle undersøkelser som video er vanskelig å bruke om natten og på dyp med lite lys. Vi testet bunnstående ekkolodd på 8 stasjoner i Bolærne i 2020 for å se på aktivitet for fisk og andre dyr gjennom døgnets mørke timer. Ekkoloddene ble plassert ved nedre grense for tareskogen (eller trådalger). Ekkogrammene kan brukes til å beregne biomasse og telle antall fisk gjennom døgnet. Det ble registrert stor aktivitet om natten og spesielt i timene før og etter soloppgang og -nedgang (Fig. 2.3.4.7).
Figur 2.3.4.7 Ekkolodd-bilde (ekkogram) fra Klauer 10-11 august 2020 fra bunnstående ekkolodd (WBAT) på 10 m dyp. Ekkogrammet viser stor aktivitet rundt solnedgang (21:15) og soloppgang (05:30). Om natten er det yrende liv over tareskogen.
2.3.5 - Fremmede arter
Koordinert med andre pågående prosjekter ble det i 2017-2019 gjennomført inventeringer av stillehavsøsters i ytre Oslofjord. Antatt «gunstige» områder /habitater inne i nasjonalparkområdet (Færder nasjonalpark), i kjente lokaliteter for stillehavsøsters, og i nærliggende områder (influensområder) hvor arten kan spres via egg/larvestadier. Det er tatt prøver for genetiske analyser av stillehavsøsters.
Det ble gjennomført «Rapid Coastal Surveys» (RCS) for potensielle fremmede arter i «hot spot» områder (områder med kjent høy risiko for tilførsel av fremmede arter), i marinaer og havner i influensområdet rundt nasjonalparkene supplert med strandinventering i områder nær marinaer.
Konkrete funn 2019 - alger
Japansk sjølyng (Dasysiphonia japonica) ble funnet i havnen ved Tjøme og i marina ved Årøy. Den fremmede rødalgen Gracilaria vermicophylla ble funnet ved Husøy Båtforening ved Tønsberg. Sporadiske funn av denne algen ble gjort innimellom tepper av bleiktuste (Spermatochnus sp.).
Evertebrater
Stillehavsøsters ble funnet på en rekke lokaliteter både utenfor og inne i nasjonalparkområdene, bl.a. ved Bolærne og Ildverket. Ingen observasjoner ble gjort i ytre deler av skjærgården (Østre Bolærne og ut mot Fulehuk). I en foreløpig genetisk analyse ved hjelp av ddRAD, viser 455 SNP’er (Single Nukleotid Polymorfismer) svært liten genetisk struktur i norske populasjoner i hele området fra Svenskegrensa til Hordaland. Det er rimelig å anta at stillehavsøsters i hele Ytre Oslofjorden har etablert seg som følge av tilførsel av larver som transporteres med strømmer i Skagerrak og Kattegat.
Et nytt funn av ny art for området ble gjort for amerikansk knivskjell Ensis leei (tidligere Ensis directus) med funn i Jensesundet, Bolærne: N 59.217069°, E 10.540524°. Amerikansk knivskjell forekommer i store tettheter langs kanalkysten fra Nederland og Belgia og sørover, men foretrekker sandbunn med høy energi, og ser ikke ut til å konkurrere med hjemlige knivskjellarter om plass.
3 - Fiskeri- og miljøpåvirkninger
3.1 - Konsekvenser av samlet beskatningstrykk i ytre Oslofjord
Tallmaterialet samlet i prosjektet for å vurdere status for kystnær torsk i ytre Oslofjord (oppsummert i kapittel 2.2) supplerer allerede tilgjengelig informasjonen som ble lagt til grunn for anbefalte tiltak i HI-rapporten «Oppdatert kunnskapsstatus om kystnær torsk i Sør-Norge» (Aglen m.fl. 2016). I ytre Oslofjord finnes torsk av «fjord-» og «Nordsjø»-økotype i alle undersøkte områder, og «Nordsjø»-økotypen ser generelt ut til å være i flertall (71% av alle fullt rekrutterte individer observert i prosjektet, Synnes 2020). I tilgrensende kystområder lenger sør (Risør til Kristiansand) – hvor fritidsfisket og yrkesfisket overlapper tilsvarende, er det påvist større andel «fjord»-økotype i fangster i fritidsfisket enn fangster i yrkesfisket (Kleiven m.fl. 2016, Jorde m.fl. 2018). I lys av oppdatert kunnskap om stabil sameksistens mellom de to økotypene i Skagerrak (jfr. Knutsen m.fl. 2018, Barth m.fl. 2019) bør størrelsen på innslaget av de to økotypene i de ulike fiskeriene tillegges mindre vekt. Begge kan ansees som viktige for gjenoppbygging av kystnær torsk og den observerte variasjonen bør bevares. Det er i denne sammenheng relevant å nevne at ICES vurderer all torsk innenfor 12 nautiske mil langs Norskekysten som adskilt fra nordsjøtorsken.
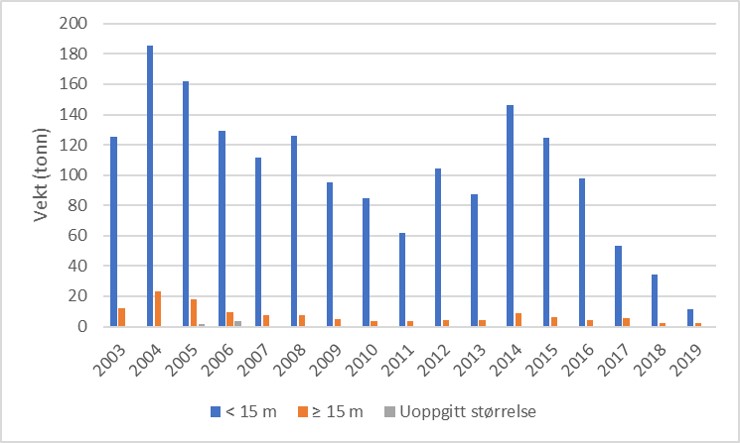
I områdene øst for 9 grader øst (Telemark-Østfold) viser Havforskningsinstituttets strandnot- og trollgarnserier minkende forekomster av torsk, og beregninger av totaldødelighet er foruroligende høye. Dette representerer i hovedsak de indre fjord- og kyststrøk. Rapporterte landinger fra det kystnære yrkesfisket med bunntrål, som forekommer på dypere vann (>60 m), viser også en negativ trend i de aller siste årene selv om de minste fartøyene (<15 m) har betydelig bifangst av torsk i perioder der torsk er tilgjengelig i området (Fig. 3.1.1).
Figur 3.1.1 Fangst av torsk som bifangst i trål for fartøyer over og under 15 m lengde i fylkene rundt Oslofjorden, 2003-2019 (kilde: Fiskeridirektoratet).
Det nyutviklede fiskeriet etter leppefisk indikerer perioder med relativt gode forekomster av «0-» og «1-gruppe» av torsk i deler av dette området, det samme rapporteres av krepsefiskere som benytter teiner på større dyp. Det er sannsynlig at denne torsken ved høyere alder bidrar til bifangst av torsk i bunntrålfisket etter reker på dypere vann (jfr. Aglen m.fl. 2016). For bunntrålfisket etter reke i Gullmarfjorden (Sverige) viste prøvetagning foretatt av Havfiskelaboratoriet i Lyskil mellom 2011 og 2014 en gjennomsnittlig fangst av småtorsk (yngel) på 33.8 individer per tråltime (Cardinale m.fl. 2017). Trålflåten i Gullmarfjorden kan fiske et maksimum antall dager i året (80 dager), og med en gjennomsnittlig daglig innsats på 11.3 tråltimer beregnet forskerne at 62% av all torskeyngel produsert av lokale gytere står i fare for å bli fanget i reketrål.
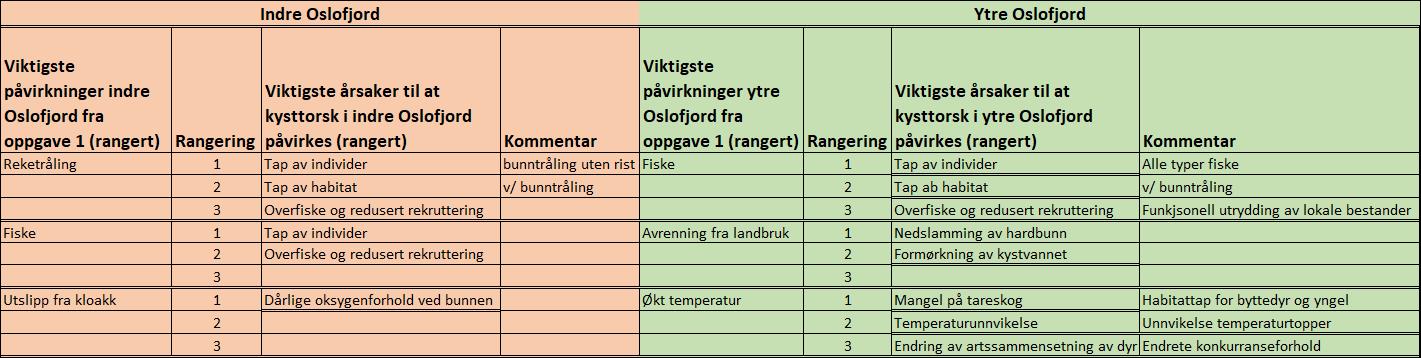
I forbindelse med innrapportering av kunnskapsgrunnlaget til «Forslag til helhetlig plan for Oslofjorden» (Miljødirektoratet 2019) rangerte Havforskningsinstituttet fiske (alle typer) som viktigste påvirkning på torsk i både indre- og ytre Oslofjord (Tabell 2). Med sterkt reduserte bestander vil alle typer fiske, inkludert fritidsfiske, bidra til å forverre situasjonen (Kleiven m.fl. 2016, 2019). Fiskeridirektoratet innførte i juni 2019 forbud mot landing av torsk fra Svenskegrensen til og med Telemark, med fiskeforbud fra 1. januar til 30. april i 14 definerte gytefelt på Skagerakkysten og i Oslofjorden, som et av flere foreslåtte tiltak i Aglen m.fl. (2016). Det er nå behov for ytterligere, varige stedsbaserte tiltak for å lykkes med restaurering av bunnfiskbestander i både indre- og ytre Oslofjord.
Tabell 3.1.1 Havforskningsinstituttets rangering av viktigste årsaker til at kysttorsk påvirkes i indre- og ytre Oslofjord (kilde: «Forslag til helhetlig plan for Oslofjorden», Miljødirektoratet 2019).
Stor arealutbredelse av fiskerier og vedvarende høyt fiskepress vurderes som en av hovedårsakene til svekkede fiskebestander og endringer i det marine miljøet (Halpern m.fl. 2008, Nellemann m.fl. 2008, IPBES 2019). Bevaringsvitenskapen har bidratt med empirisk kunnskap som gir håp for gjenoppbygging av marine bestander forutsatt en fortsatt vekst i strengt marint vern (Duarte m.fl. 2020). Fredningsområder for hummer (der bare krokfiske er tillatt) har bidratt til å utvikle bevaringsvitenskap for lokale forhold, og utgjør nå et veldokumentert og utbredt forvaltningsverktøy i Skagerrak, ikke minst i Oslofjorden. Fredningsområder for hummer har også vist positive effekter for torsk i form av økt gjennomsnittsstørrelse og tetthet (Moland m.fl. 2013), oppnådd som effekt av redusert fiskedødelighet (Fernandez-Chacon m.fl. 2015). Positive effekter av slike tiltak er også vist for leppefisk (Halvorsen m.fl. 2017). For å oppnå positive økosystemeffekter vil noe areal måtte omdisponeres fra fiskeri- til bevaringsformål. Konvensjonen om biologisk mangfold (CBD) forplikter partsnasjonene til å sette av 10% av kyst- og havområder for effektivt varig vern innen 2020. Det er foreslått å utvide dette målet til 30% innen 2030. Aglen m.fl. (2016) anbefalte opprettelse av tre referanseområder med null-fiske i Skagerrak for å sammenligne og vurdere effekten av øvrige forvaltningstiltak og høstingsregler. Anbefalt størrelse på slike referanseområder ble i rapporten oppgitt til minimum 5 km2. Dette er for små arealer til å oppnå en langsiktig bevaringseffekt på kystnær torsk i studieområdet. Betydelig større områder designet som transekt fra land til største dyp i Oslofjorden vil ha større sannsynlighet for å lykkes med å demonstrere en positiv effekt på torsk og andre langlivete og storvokste arter av bunnfisk. Detaljert forsøksfiske gjennomført i dette prosjektet har bekreftet at det haster å innføre treffsikre tiltak med potensial til å gjenoppbygge bred alders- og størrelsessammensetning i lokale bestander av kysttnær torsk i Oslofjorden/ indre Skagerrak. Hvorvidt det er mulig å kombinere bunntråling og gjenoppbygging av bunnfiskbestander – i samme økosystem – er usikkert. Derimot vet vi fra Øresund at et område med langvarig bunntrålforbud (siden 1932) var i stand til å bevare bred alders- og størrelsessammensetning hos torsk, i samme tidsrom som torskebestander kollapset i tilgrensende områder (Lindegren m.fl. 2010; Sundelöf m.fl. 2013).
3.2 - Økosystempåvirkning fra bunntrål i Oslofjorden
Figur 3.2.1 Bunntråling etter dyphavsreke (Pandalus borealis) er utbredt i ytre Oslofjord. De minste trålerne har ikke hatt krav om sporingsutstyr (VMS). Det er derfor ikke mulig å beregne den totale innsatsen og områdebruken til denne delen av flåten. Foto: E. Moland/ Havforskningsinstituttet.
Av historiske fiskeripolitiske årsaker er fiskeri med bunntrål tillatt i alle områder dypere enn 60 m fra Jærens rev til Svenskegrensen (Jærens rev til Nord-Trøndelag: dypere enn 100 m, nord for Nord-Trøndelag: dypere enn 170 m), også innenfor grunnlinjen. Dette har medført at hele Oslofjorden er sterkt påvirket av bunntråling. Blant de ulike fiskeriene er det sannsynlig at bunntråling er den viktigste påvirkningsfaktoren på fjordøkosystemet gjennom de siste 100 årene. Skagerrak er blant de mest intenst trålte kystområder i verden, målt i antall timer trålt per kvadratkilometer (Kroodsma m.fl. 2018). Bunntråling ble introdusert til Oslofjorden (indre Oslofjord/ Bunnefjorden) ca. år 1900 etter anbefaling og instruksjon fra Johan Hjort. Et rekefiskeri basert på tråling ble etablert fra ca. 1910 med fangster fra 20 til 100 tonn årlig (Broch m.fl. 1932). Fisket var i starten svært godt og man erkjente at man høstet fra en jomfruelig/ akkumulert bestand. Ruud (1968) beskrev hvordan noen fiskearter (håkjerring, svarthå, kveite) forsvant helt fra indre Oslofjord i tidsrommet mellom grundige undersøkelser gjennomført i 1897 og årene 1963-1967. En rekke arter som tidligere var nokså vanlige på dypt vann indre Oslofjord (kloskate, havmus, sølvtorsk, uer, steinbit, m.fl.) ble svært sjeldne i samme periode. I tilfellet håkjerring – som ble ansett som skadedyr, tilskrives ikke forsvinningen bunntråling, men heller utbetaling av premie for landing av haiarten, administrert av «Foreningen til fremme av fiskeriet i Oslofjorden innenfor Drøbak». Premie ble utbetalt for siste gang i 1920 for de to siste kjente landingene av håkjerring (fra Bunnefjorden). Begge fiskene ble rapportert til å være 3.5 m lange, med vekt 200 kg.
I østlige Skagerrak og Kattegat er effekten av bunntråling på økosystemet knyttet til havbunnen vurdert ved å sammenligne artsinventeringer gjennomført i 1921-1938 og ≈70 år senere i 2004-2009. I perioden der bunntråling ble etablert, modernisert og intensivert er artsmangfoldet halvert, og tap av arter og individer har ført til funksjonell homogenisering (tap av mangfold av funksjoner) (Obst m.fl. 2018). For ytre Oslofjord rapporterte Walday m.fl. (2019) i NIVA-rapporten «Overvåking av Ytre Oslofjord i 2014-2018» at bunnområdene i de åpne, ytre deler viser en tilsvarende negativ utvikling ved at bløtbunnsfaunaen har blitt mer fattig, både med hensyn til antall individer og antall arter. På den mest trålpåvirkede stasjonen fant man reduksjon i andelen dyptgravende dyr, og økning i andelen dyr som lever i sedimentoverflaten, som er tegn på at faunasammensetningen er påvirket av tråling. Restaurering av bunnfauna etter langvarig og dyptpenetrerende trålpåvirkning kan ta opptil 10 år (Rijnsdorp m.fl. 2020). I tillegg til negativ påvirkning av økosystemet knyttet til sedimentene medfører bunntråling resuspendering av sedimentpartikler. I Gullmarfjorden er konsentrasjoner opp til 4.8 mg/l målt i forbindelse med reketråling (Wikström m.fl. 2016), som er høyere enn torskens toleransegrense på 3 mg/l (Westerberg m.fl. 1996). Det er grunn til å anta at utstrakt hyppig bunntråling i ytre Oslofjord medfører resuspendering på tilsvarende nivå.
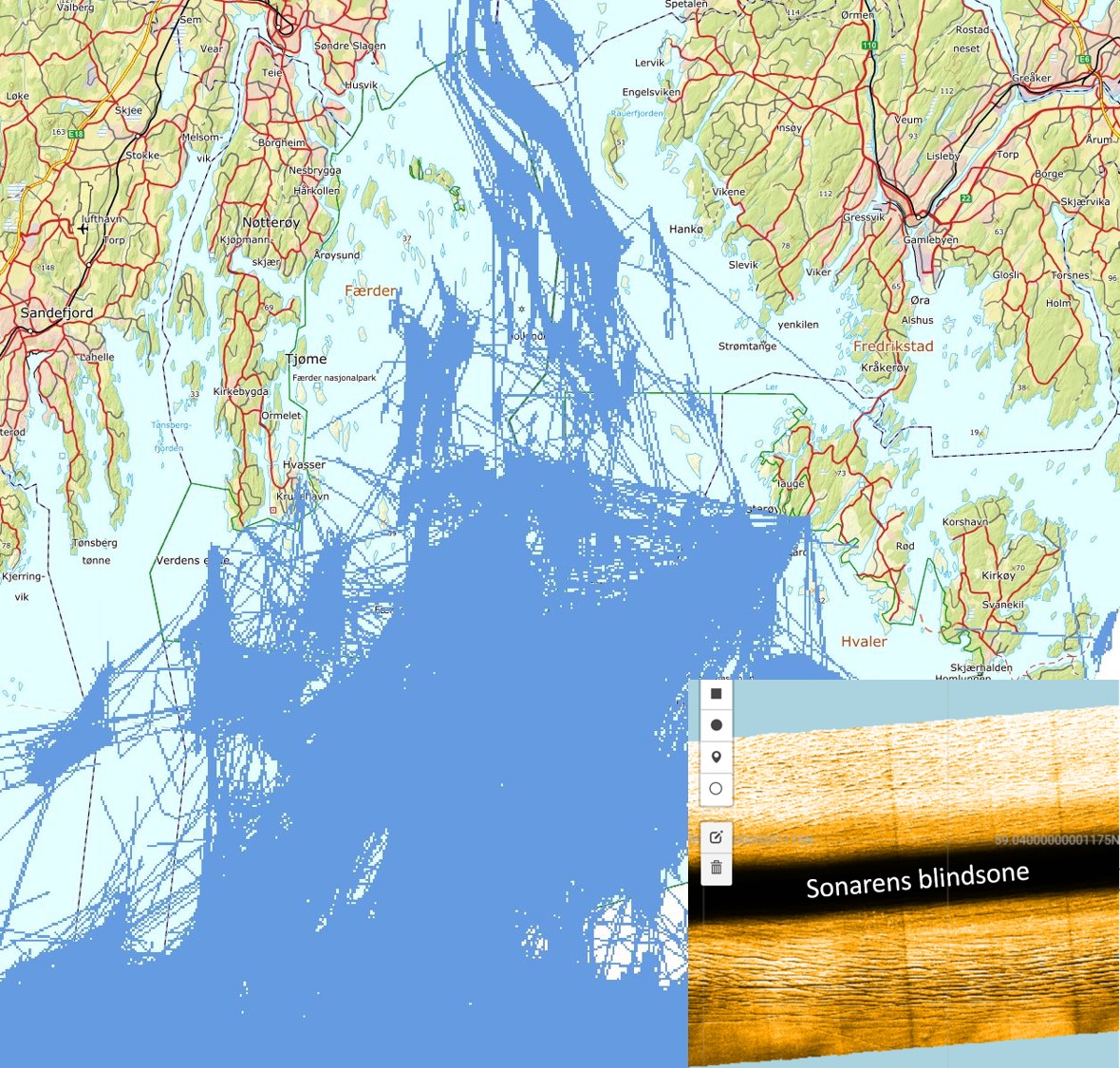
Torskefangstene tatt som bifangst i reketrål av fartøyer med tilhørighet i fylkene rundt Oslofjorden viser et betydelig uttak av torsk i fartøygruppen <15 m lengde (Fig. 3.1.1) med totale årlige landinger på over 120 tonn frem til 2014 og 2015, men med en betydelig reduksjon i perioden 2016-2019. Sporingsdata finnes ikke fordi utstyret som kreves (AIS/ VMS) ikke er påbudt for denne fartøygruppen. Sporingsdata innhentet fra fartøy >15 m indikerer svært høy utnyttelse av tillatte dyp og egnete deler av fjordbunnen i ytre Oslofjord (Fig. 3.2.2). Et pilotprosjekt med kartlegging av bunnen ved bruk av sonar fra autonom undervannsfarkost (AUV) ble gjennomført i søsterprosjektet «Frisk Oslofjord». En visuell vurdering av sonarbildene fra seilingsruten til AUV’en «Hugin» viste nær 100% dekningsgrad av trålspor på bunnen dypere enn 60 meter (E. Moland, A. Hestnes, pers. obs.).
Figur 3.2.2 VMS-data («slepestrek») for fartøy som fisker med bunntrål (fartøy over 15 m lengde) i ytre Oslofjord i perioden 2011–2019 (kilde: Fiskeridirektoratet). Nede til høyre: sonarbilde av trålspor på havbunnen i ytre Oslofjord tatt med Kongsberg maritimes autonome undervannsfarkost Hugin under tokt i regi av søsterprosjektet «Frisk Oslofjord» (kilde: Kongsberg maritime). «Glatting» av VMS-data vil være nødvendig for å benytte disse til vurderinger av grad av belastning fra denne flåtegruppen i perioden.
3.3 - Vannkvalitet og miljø: historiske data og nye registreringer
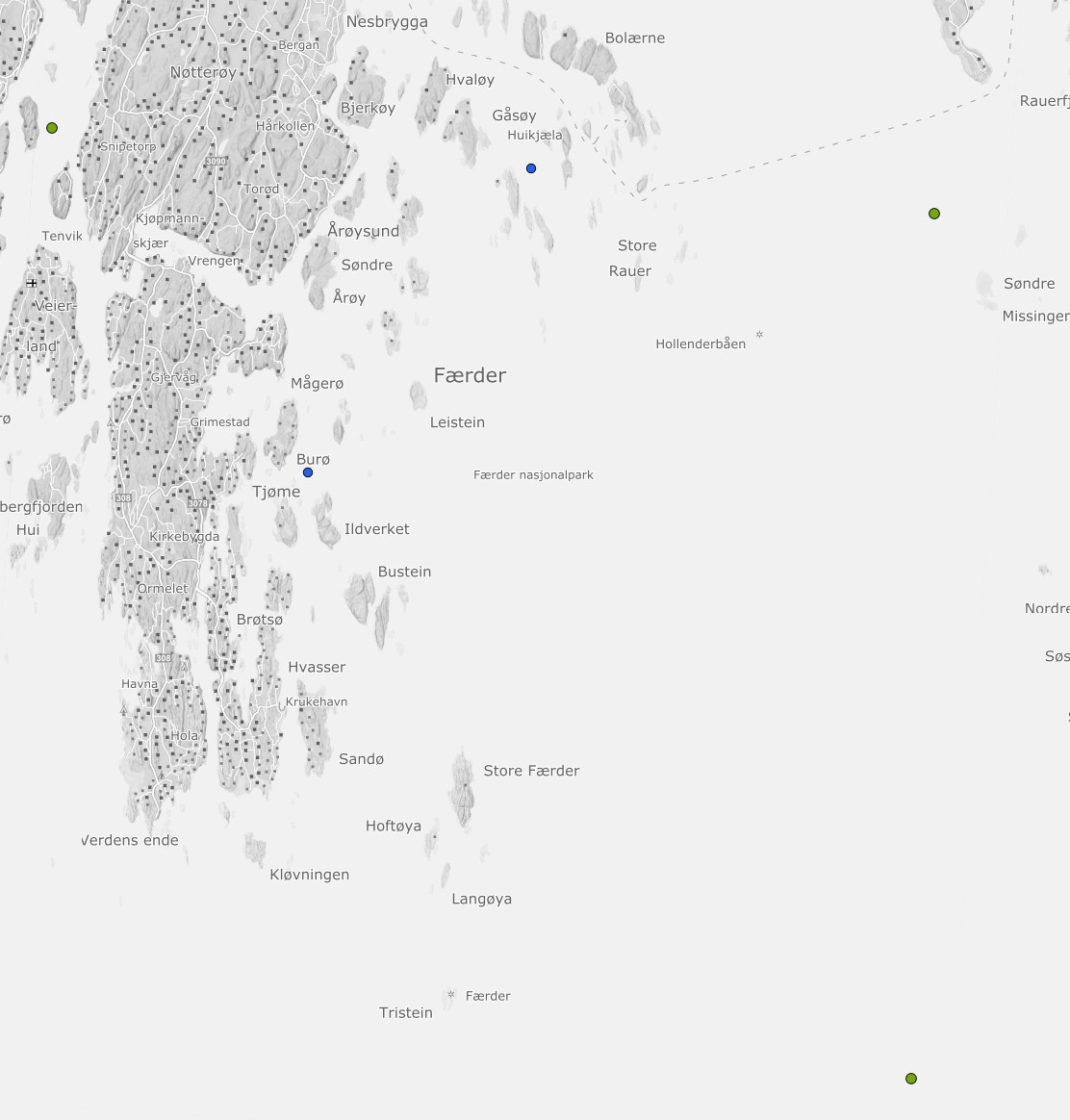
I prosjektet er det foretatt analyser av eksisterende datagrunnlag i ytre Oslofjord basert på data innhentet av Havforskningsinstituttet og Fagrådet for Ytre Oslofjord. I tillegg er det gjennomført prøvetakning ved 2 stasjoner i Færder nasjonalpark. Stasjonene OF 1 (Torbjørnskjær) og OF 2 (Missingen) har vært rutinemessig overvåket fra 1995. Stasjonen i Vestfjorden ved Tønsberg er inkludert for å beskrive forholdene i ett beskyttet fjordsystem i studieområdet. Stasjonen Kongsholmen og Bolærne (Fig. 3.3.1, blå prikker) er inkludert for å beskrive de moderat eksponerte områdene i Færder Nasjonalpark. Ved alle stasjoner er det innhentet vannprøver for kjemiske analyser (næringssalter), oksygen og planteplankton (klorofyll a). I tillegg er det innhentet fysiske data, samt målinger av temperatur og saltholdighet.
Figur 3.3.1 Undersøkt område i ytre Oslofjord. Grønne punkt indikere lokaliteter med tidsserier (Vestfjorden, Missingen, Torbjørnskjær), blå punkter (Huikjæla, Burø) er nye lokaliteter opprettet i forbindelse med prosjektet.
Næringssalter
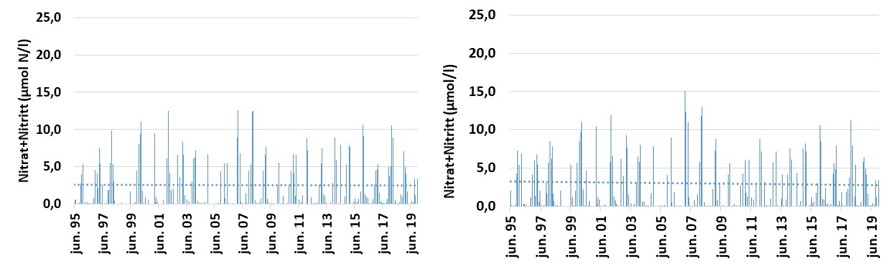
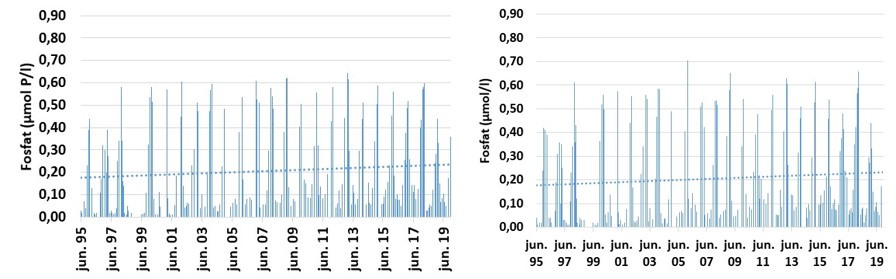
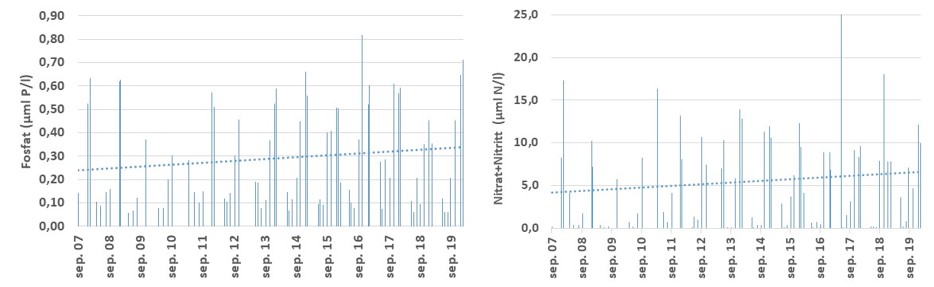
Det er først og fremst konsentrasjonen av fosfat og nitrogen (Nitrat+Nitritt) som er inkludert i tilstandsvurderinger og beskrivelse av eutrofieringssituasjonen. I dette materialet er det i tillegg inkludert totalt nitrogen og fosfat. For nitrogenforbindelsen Nitrat+Nitritt, har konsentrasjonen vært svakt avtakende fra et i utgangspunktet høyt nivå i perioden 1995-2019 ved Missingen og Torbjørnskjær i ytre Oslofjord (Fig. 3.3.2). Dataene indikerer at eutrofieringssituasjon i de ytre, åpne områdene har bedret seg over tid. For stasjonene Bolærne og Kongsholmen er dataserien for korte til å si noe om utviklingen. Dataene som foreligger, indikerer at Nitrat+Nitritt-konsentrasjon ikke avviker mye fra stasjon OF1 og OF2. I dette arbeidet er det også inkludert data fra stasjonen «Vestfjorden» (Tønsberg) hvor det i perioden 2007-2019 er observert økende Nitrat+Nitritt-konsentrasjon i overflatelaget (Fig. 3.3.4). Forskjellen mellom stasjon «Vestfjorden» og de ytre stasjonene kan forklares med økt tilførsel av nitrogenforbindelser i avrenning fra land. Samtidig med at nitrogenkonsentrasjon har vist en svak avtakende konsentrasjon i de områdene som ligger mot Oslofjorden, er det økende mengder med fosfat ved Torbjørnskjær (OF1) og Missingen (OF2) (Fig. 3.3.3). Det er fortsatt noe uvisst hva som er årsaken til den økende fosfatkonsentrasjon i dette området, men man har sett tilsvarende trender i ved flere lokaliteter i Oslofjorden (Aure m.fl. 2014) og ved «Vestfjorden» (Fig. 3.3.4). I utenforliggende områder, f.eks. Langesundsbukta, er det ikke observert en like tydelig økning, mens det i åpne Skagerrak har vært registret en reduksjon i fosfat i overflatelaget i den samme perioden. Generelt er det god miljøtilstand basert på kjemiske parametere i de åpne delene av ytre Oslofjord basert på samlet vurdering for perioden 2017-2019. De mer beskyttede sidefjordene er mer utsatt for avrenning og har redusert sirkulasjon med utenforliggende områder og viser dårlig tilstand. For stasjonen Torbjørnskjær og Missingen har man tidligere registrert forhøyede konsentrasjoner av Nitrat+Nitritt i vinterperioden. Med den observerte reduksjon i nitrogen har forholdene bedret seg de senere årene, med forbehold om at disse årene var preget av lite nedbør i sommerhalvåret. For prosjektperioden er forholdene «god» ved stasjonene. For stasjonen Bolærne og Kongsholmen er tilstanden «god». For stasjonen Vestfjorden er derimot tilstanden «moderat». For denne lokaliteten er det Nitrat+Nitritt og totalt fosfat som er de utslagsgivende. I tillegg er det redusert tilstand i bunnvannet på grunn av lave oksygenkonsentrasjoner (se under).
Figur 3.3.2 Nitrat+Nitritt konsentrasjon (µmol/l) ved stasjonene Torbjørnskjær (venstre) og Missingen (Høyre) i overflatevannet (0-10 m) i perioden 1995-2019. Prikket linje gir trendlinje over perioden.
Figur 3.3.3 Fosfat konsentrasjon (µmol/l) ved stasjonene Torbjørnskjær (venstre) og Missingen (Høyre) i overflatevannet (0-10 m) i perioden 1995-2019. Prikket linje gir trendlinje over perioden.
Figur 3.3.4. Næringssaltkonsentrasjon (µmol/l) ved stasjonen «Vestfjorden», Fosfat (venstre) og Nitrat+Nitritt (høyre) i overflatelaget i perioden 2007-2019.
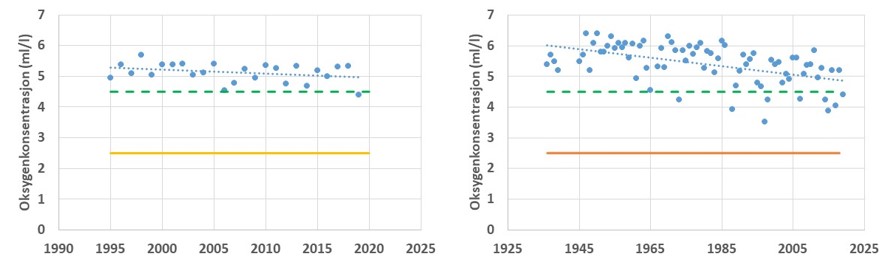
Oksygenforhold
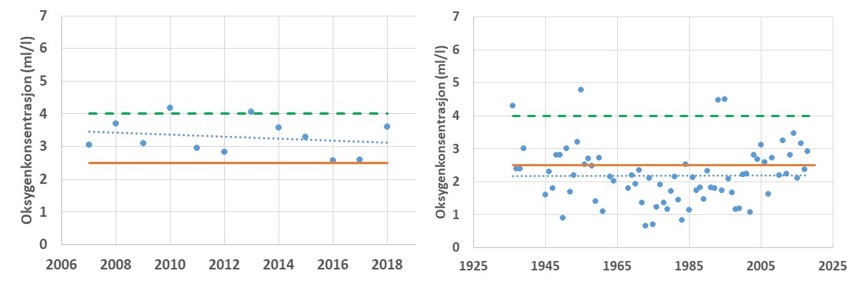
Oksygenkonsentrasjonen i overflatevannet påvirkes av biologisk produksjon i vannlagene. I dypvannet vil oksygenkonsentrasjon påvirkes av organisk belastning og nedbryting av materialet. Stor tilførsel av organisk materiale - fra vannsøyleproduksjon eller tilførsel via avrenning fra land - vil kunne resultere i reduserte oksygenkonsentrasjoner. Hvor lav konsentrasjonen blir i bunnvannet vil også avhenge av bunnvannets fornyelse. Bunndyrenes tilstedeværelse, sammensetning og mengde vil påvirkes av oksygenkonsentrasjonen. I nye undersøkelser har man funnet at dersom oksygenkonsentrasjonen faller under et gitt nivå (ca. 2.5 ml/l oksygen) vil torsk unngå i å oppholde seg i slike vannmasser. Lave oksygenkonsentrasjoner i bunnvannet vil dermed føre til at torskens habitat reduseres og bunndyr i slike områder vil være utilgjengelig føde. Ved stasjonen Torbjørnskjær og Missingen er forholdene generlt sett gode (Fig. 3.3.5). Oksygenkonsentrasjon holder seg relativt høy ved disse stasjonene, da de er i tett kontakt med et åpent Skagerrak og viser jevnlig utskiftning i bunnvannet. Ved Missingen er det registrert en reduksjon i oksygenkonsentrasjonen over tid, noe som indikerer en økt organisk belastning i dette området. Samme mønster er registret ved Torbjørnskjær, men reduksjonen er betydelig mindre. Ved begge disse stasjonene er oksygenkonsentrasjonen i bunnvannet såpass høy at torsk vil kunne oppholde seg i disse dypene også ved de laveste målte nivåene. Tilsvarende observeres i data fra moderat eksponerte områder Bolærne-Kongsholmen (Fig. 3.3.7), selv om tidsserien fra dette området er betydelig kortere og dermed danner grunnlag for mer usikre vurderinger.
Figur 3.3.5 Oksygenkonsentrasjonen i bunnvannet ved stasjonene Torbjørnskjær (venstre, 1995-2019) og Missingen (Høyre, 1936-2019). Prikket blå linje gir trendlinje over perioden, grønn linje gir konsentrasjon for «god» tilstand. Gul og oransje linje gir nedre tålegrense for torsk.
For andre områder i Oslofjorden er ikke forholdene like gode. I Vestfjorden på innsiden av Tjøme viser målinger i perioden 2007-2018 at oksygenkonsentrasjonen er redusert og har i perioder vært helt nede på grensen for hva torsk vil oppholde seg i (Fig. 3.3.6). I andre deler av Oslofjorden er forholdene for torsk dårlige, som for eksempel ved Steilene i indre Oslofjord eller i de indre delen av Hvaler. Data fra de to sistnevnte områdene viser at de mer beskyttede fjordområdene med lav utskiftning av dypvann er sårbare for tilførsel av organisk materiale og vil kunne oppleve perioder der torskefisk ikke vil kunne utnytte disse bunnområdene.
Figur 3.3.6 Oksygenkonsentrasjonen i bunnvannet ved stasjonene Vestfjorden, Tønsberg (venstre, 2007-2018) og Steilene i indre Oslofjord (Høyre, 1936-2018). Prikket blå linje gir trendlinje over perioden, grønn linje gir konsentrasjon for «god» tilstand. Oransje linje gir nedre tålegrense for torsk.
Figur 3.3.7 Oksygenkonsentrasjon ved stasjonene «Kongsholmen» og «Bolærne» i perioden 2017-2019. Grønn linje gir konsentrasjon for «god» tilstand og Oransje linje gir nedre tålegrense for torsk.
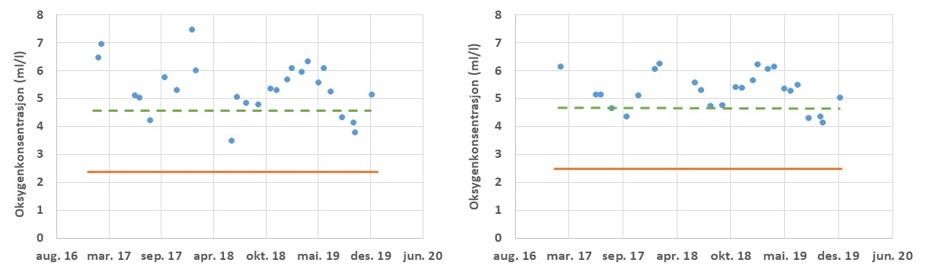
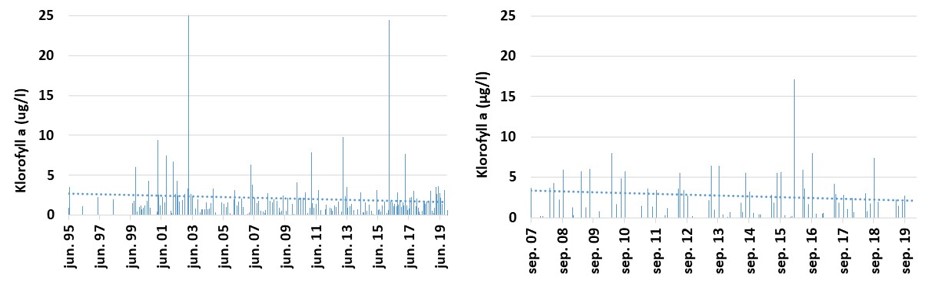
Planteplankton biomasse (klorofyll a)
Planteplankton er encellede planter som svever fritt i vannmassene. Planteplanktonet er avhengig av næringssalter (nitrogen, fosfat og silikat) for å kunne vokse, samt at de fysiske forholdene må ligge til rette for biomasse økning. Planteplankton respondere raskt på endringer i miljøet, som for eksempel tilførsel av næringssalter via avrenning eller innblanding av dypereliggende vannlag med høyere konsentrasjoner av næringssalter. Planteplanktonet gjennomgår en mer eller mindre fast suksesjon gjennom året, som starter med en våroppblomstring i mars-april og avsluttes i oktober-november på grunn av endringer i det fysiske miljøet. I sommerperioden er normalt sett biomassen lav, men i fjordområder vil man kunne ha sommeroppblomstringer i etterkant av avrenningsperioder. Stasjonene «Torbjørnskjær» og «Vestfjorden» viser begge en avtakende trend i mengden klorofyll a (Fig. 3.3.8). Tilsvarende trender er registrert ved flere stasjoner i Oslofjorden. Også i de utenforliggende områdene registreres det en negativ trend i planteplanktonbiomassen (Frigstad m.fl. 2018, Havforskningsinstituttet upubliserte data). Reduksjon i planteplanktonproduksjon kan knyttes til reduksjon i nitrogen konsentrasjonen i en rekke områder. For «Vestfjorden» er det derimot både en reduksjon i klorofyll a og en økning i nitrogenkonsentrasjon. Rent teoretisk skulle en økning i næringssalter stimulere til økt planteplanktonproduksjon. Manglede respons i planteplanktonet kan skyldes endringer i andre faktorer som påvirker planteplanktonvekst. I fjorder med stor elvetilførsel kan endringer i lysforholdene medfører redusert planteplanktonvekst selv under gode næringssaltbetingelser (se Frigstad m.fl. 2020).
Figur 3.3.8 Klorofyll a konsentrasjonen (µg/l) ved lokalitetene Torbjørnskjær OF1 (1995-2019) og ved stasjonen «Vestfjorden» ved Tønsberg (2007-2019).
3.4 - Formørking av kystvannet
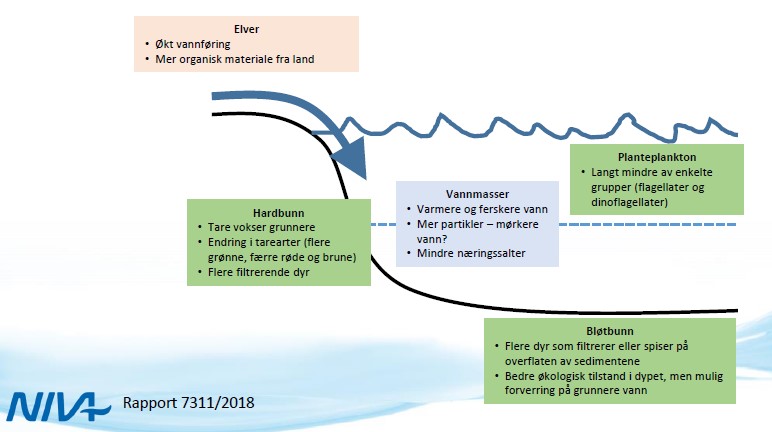
Klimaendringer vil påvirke marine økosystemer gjennom endret og økt belastning. Nordsjøen og Skagerrak betegnes allerede som et endret økosystem som følge av forhøyet gjennomsnittstemperatur (Arneberg m.fl. 2018). I tillegg viser klimaframskrivinger økt nedbørsmengde og -intensitet, og endret avrenningsmønster (f.eks. flere flomperioder/ -episoder). Med endret avrenningsmønster vil tilførsel av næringssalter endres. Man vil også få endret tilførsel av humus (CDOM) og organisk materiale (ter-OM) og -partikler (erosjonspartikler) fra landsiden. Fjord og kystområder der større vassdrag ender vil være mest utsatt (Deininger m.fl. 2020, Frigstad m.fl. 2020). Tidligere studier har vist at det har vært en økning i tilførselen av organisk materiale med terrestrisk opprinnelse til kystområdene (Aksnes m.fl. 2009, Frigstad m.fl. 2013, Deininger m.fl. 2020). En økning i tilførsel av organisk materiale vil resultere i økte DOC (løst organisk materiale, da spesielt den fargede komponenten CDOM), som igjen vil påvirke lyssvekkingen i vannsøylen. Endringer i lysforholdene vil kunne påvirke planteplanktonproduksjon ved at lysmengde tilgjengelig for vekst nedover i vannsøylen reduseres. Lyssvekkelse vil også kunne ha negativ betydning for visuelle predatorer som fisk, og vil kunne stimulere ikke-visuelle predatorer (maneter) og bakterieproduksjon (Thingstad m.fl. 2008, Aksnes m.fl. 2009, Deininger og Frigstad 2019).
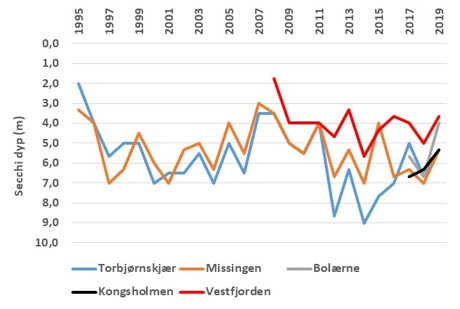
Målinger av CDOM i Oslofjorden viser at det er stor variasjon mellom lokaliteter og sesonger (Fagerli m.fl. 2020). Data fra «Økokyst-Skagerrak» viser at det er en tett kobling mellom saltholdighet og CDOM, som indikere at for dette området er opphavet fra landbasert kilder som transporteres ut i kystvannet ved avrenning. Målinger av lys (PAR-sensor) eller bruk av Secchi-skive gir ikke et direkte mål for CDOM, men gir informasjon omkring lysforholdene der ulike forhold påvirker svekkingen av lys med dypet. Viktige faktorer i lyssvekking vil være planteplanktonproduksjon (klorofyll), organisk materiale og fysiske forhold. Secchi-dypet vil variere innen året og mellom årene, men det generelle mønsteret er at stasjoner lokalisert i de åpne eksponerte områdene har større Secchi-dyp enn stasjoner som ligger inne i elvepåvirkede områder (Fig. 3.4.1). Ved bruk av lyssensorer (PAR) er det mulig å estimere svekkingen av lys mot dypet. For stasjonen OF1 Torbjørnskjær er det foretatt systematisk måling av PAR i perioden 2017-2019 (Fig. 3.4.2). I 2017 var det relativt klart vann med lite lyssvekking store deler av året, med unntak av høstperioden august-oktober. I 2019 var det derimot høy lyssvekking i mai-juli. I begge tilfellene sammenfaller perioden med lavere saltholdighet i overflatelaget som indikere stor avrenning og sannsynligvis stor tilførsel av organisk materiale. Se også kapittel 3.6 Samlet vurdering av miljøtilstand, basert på hovedmomenter fra «Forslag til helhetlig plan for Oslofjorden».
Figur 3.4.1 Siktdyp (m) basert på Secchi-skive-målinger for perioden 1995-2019, for Missingen og Torbjørnskjær, Vestfjorden (Tønsberg) 2008-2019, samt Kongsholmen og Bolærne i perioden 2017-2018. Kun data fra sommerperioden benyttet (juni-august).
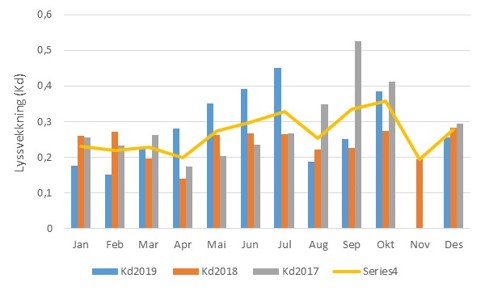
Figur 3.4.2 Lyssvekking ved stasjon OF 1 Torbjørnskjær i perioden 2017-2019. Svekkingskoeffisienten (Kd) er basert på data fra overflaten ned til 1% lysdyp. Gul linje gir gjennomsnittlig lyssvekking for perioden.
3.5 - Miljøgifter
Norskekysten viser hovedsakelig nedadgående trender for forekomst av undersøkte miljøgifter selv om kostholdsråd for fisk og skalldyr stadig er et faktum i flere fjorder. Indre Oslofjord er et område med forhøyede miljøgiftkonsentrasjoner som gir grunnlag for bekymring og ikke minst behov for overvåking, nærmere undersøkelser og tiltak (se Green m.fl. 2015). Laboratorieforsøk viser negative konsekvenser av miljøgifter for vannlevende organismer, men det finnes få feltbaserte studier som viser at miljøgifter har hatt bestandsregulerende effekter på marine fiskearter. Ono m.fl. (2019) brukte Havforskningsinstituttets strandnotserie for å undersøke i hvilken grad miljøgiftene kvikksølv, kadmium og hexaklorbensen – overvåket gjennom konsentrasjoner i blåskjell (se f.eks. Schøyen m.fl. 2017) – har hatt effekt på rekruttering av torsk i Skagerrak i perioden mellom 1980-2015. Arbeidet viste at kvikksølv kan ha bidratt til redusert rekruttering av torsk i noen fjorder i Skagerrak, med forbehold om andre kilder til variasjon i data. For Oslofjorden fant forfatterne av studien at rekruttering av torsk aldri falt under 50% av middelverdien i perioden, selv i årene med høyeste målte verdier av kvikksølv i blåskjell. Studien konkluderte at reduksjon av miljøgifter alene ikke vil kunne bidra til gjenoppbygging av lokale torskebestander, fordi dette mest sannsynlig avhenger av fiskerireguleringer og habitatrestaurering.
3.6 - Samlet vurdering av miljøtilstand, basert på hovedmomenter fra «Forslag til helhetlig plan for Oslofjorden»
Oppdraget i prosjektbeskrivelsen fra 2017 omfatter en vurdering av årsakene til at kysttorsken har gått sterkt tilbake. Disse vurderingene skal bygge på eksisterende kunnskap og data framskaffet gjennom prosjektet. Samfunnets fokus på Oslofjorden er stort og i prosjektperioden ble det satt i gang et omfattende arbeid med en helhetlig plan for Oslofjorden, ledet av Miljødirektoratet. Forslag til planen ble lagt fram i desember 2019. Dette planarbeidet involverte det aller meste av norske fagmiljøer, innen forskning og forvaltning. Miljøaspektene i Oslofjorden er godt dekket opp i dette dokumentet.
Helhetlig plan er nå under sluttbehandling i Klima- og miljødepartementet og det er varslet at Klima- og miljødepartementets anbefaling av tiltak vil foreligge i løpet av 2020. I og med at dette omfattende arbeidet nå er gjennomført og vil bli lagt til grunn for miljøtiltak i hele Oslofjordregionen, har prosjektet funnet det lite hensiktsmessig å gi en ny gjennomgang av alle fjordens miljøaspekter. Dette kapittelet er derfor en oppsummering av hovedelementer fra «Forslag til helhetlig plan for Oslofjorden».
Oslofjorden har gjennom mange tiår vært påvirket fra ulike aktiviteter på landsiden. De største menneskeskapte påvirkningene er tilførsler av organisk materiale, næringssalter og jordpartikler. De menneskeskapte kildene er jordbruk, kommunal kloakk og noe industri. Kildene er i stor grad lokale (Østlandet, se Fig. 3.6.2), men også langtransport helt fra Østersjøen. Algevekst og jordpartikler gir svekket lystilgang til naturen under vann og tilslamming av havbunnen. Tareskogenes utbredelse er redusert og en rekke områder har fått mer eutrof undervannsvegetasjon, som delvis også overgror tang, tare og ålegras .
Kystovervåkingsprogrammets målinger viste at vannkvaliteten i vårt kystvann ble bedre med reduksjon i kontinentale utslipp, men samtidig som langtransporterte tilførsler med kyststrømmen ble mindre, fikk lokale tilførsler større betydning for lokal vannkvalitet. Ulike tekniske inngrep bidrar også til å svekke fiskens leveområder, særlig i gruntvannsområder, med havner, utfylling, mudring, sprengning, moringer osv. Forringelse av leveområder (habitatforringelse), er en betydelig trussel mot biologisk mangfold. For eksempel trenger fisk ulike steder og typer av leveområder i ulike deler av sitt livsløp: oppvekstområder for yngel, lek- og gyteplasser, skjulesteder og områder for næringssøk.
Klimaendringer har også her en avgjørende betydning ved at de kommer på toppen av annen menneskelig aktivitet og inngrep, og medfører økte og/ eller uventede effekter. En samlet analyse av klima- og overvåkingsdata fra Skagerrakkysten viste at antall flomepisoder økte fra 1990 og utover, antall episoder med frysing/ tining av jorda gjennom vinteren økte betydelig, samt økt nedbør i vinterhalvåret. Alle tre faktorer medfører økt transport av jord- og leirpartikler, næringssalter, mm. fra land til kystvannet (Frigstad m.fl. 2018, Fig. 3.6.1). Grumsete vann sammen med temperaturøkning ble framholdt som mest sannsynlig forklaring på den kraftige tilbakegangen i sukkertare langs kysten av Skagerrak (se også Filbee-Dexter m.fl. 2020).
Figur 3.6.1 Oppsummering av funn fra overvåkingsprogrammer i rapporten «Fra land til hav: Endringer i Skagerrak de siste 30 år» (Frigstad m.fl. 2018).
Oslofjorden er Norges mest urbane og folketette kystlinje. Befolkningsøkning og fortetning forventes, med konsekvenser for utslipp og overflatevann. Fylkene rundt Oslofjorden har landets største landbruksarealer (gjennomsnittlig 17%, mot de øvrige fylkers gjennomsnitt på 3.4%). I tillegg medfører Sør-Norges topografi at vann fra «hele» Østlandet drenerer ut i Oslofjorden (Fig. 3.6.2).
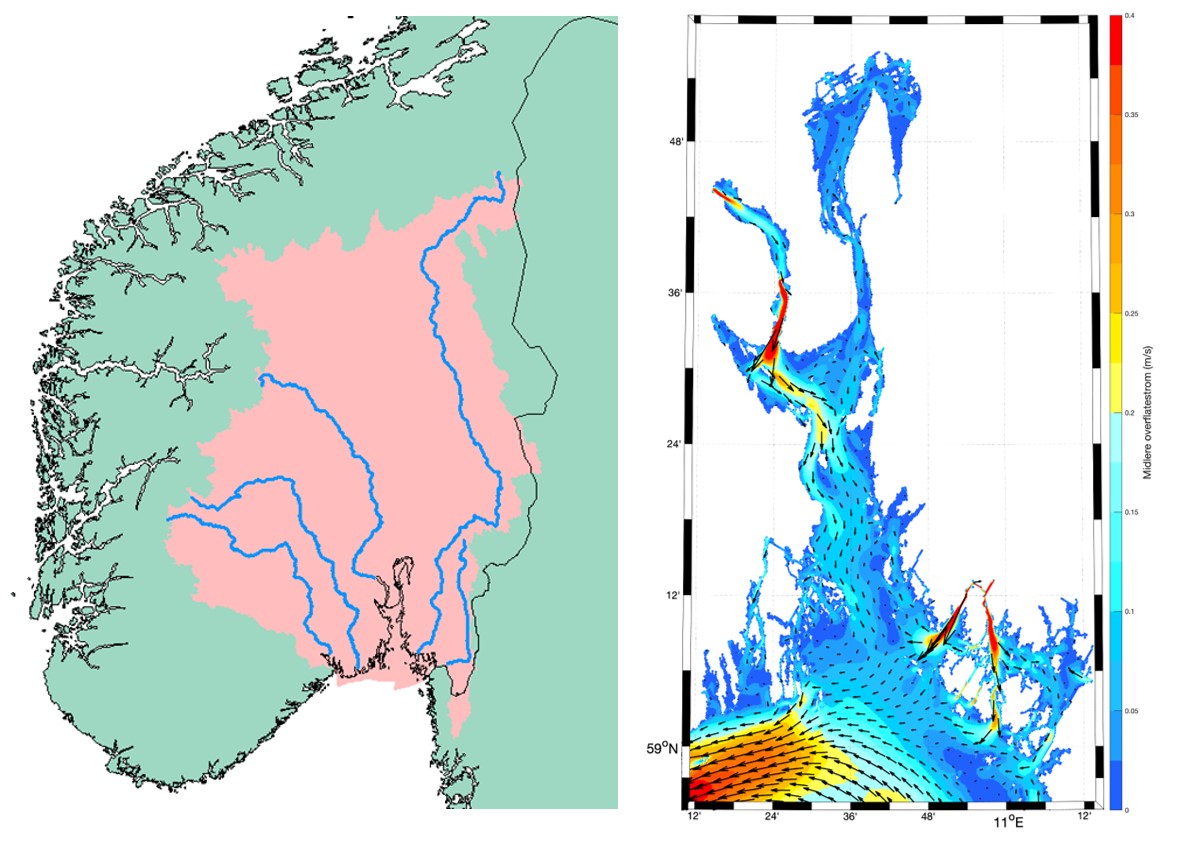
Figur 3.6.2 Venstre: kart over nedbørsfelt (Regine) som drenerer til Oslofjorden. De store elvene (fra vest mot øst) er Skiensvassdraget, Numedalslågen, Drammenselva, Glomma og Haldenvassdraget (kilde: NVE-Atlas https://atlas.nve.no). Høyre: gjennomsnittlig overflatestrøm i Oslofjorden og ytre Oslofjord (J. Albretsen/ Havforskningsinstituttet).
3.7 - Er det håp for fiskebestandene i Oslofjorden? Samlet vurdering
Det er godt dokumentert at menneskelig påvirkning er årsaken til fiskebestandenes tilbakegang i verdens kyst- og havområder, og i Oslofjorden. En eksakt vektfordeling mellom ulike hovedfaktorer for Oslofjordens vedkommende er en komplisert øvelse, og de ulike faktorenes innslag og relative viktighet ville måtte plasseres langs en tidslinje. Det er likevel overveiende sannsynlig at endringene skyldes en kombinasjon av mange års overbeskatning fra fiskeriene, påvirkning av fjordbunnen og bunndyrsfaunaen som følge av bunntråling, og en negativ utvikling av miljøtilstanden i fjorden der også klimaendringene spiller inn (for en uttømmende liste, se Tabell 6. s. 106-107 i «Forslag til helhetlig plan for Oslofjorden»). I forbindelse med innrapportering av kunnskapsgrunnlaget til «Forslag til helhetlig plan for Oslofjorden» rangerte Havforskningsinstituttet fiske, avrenning fra landbruk og økt temperatur som viktigste påvirkninger på «miljøelementet kysttorsk» i ytre Oslofjord (se kapittel 3.1).
Klimaendringer har trolig medført økosystemendringer i Nordsjøen og Skagerrak (Johannessen m.fl. 2012, Arneberg m.fl. 2018). Samtidig er grunnlaget svakt for å hevde at det ikke lenger finnes livsgrunnlag for torsk og andre sterkt reduserte fiskebestander i Oslofjorden. Dette prosjektet har vist at ytre Oslofjord fremdeles huser et mangfoldig fiskesamfunn, og at individer fra de mest reduserte fiskeartene fremdeles forekommer i hele området. Beste tilgjengelige kunnskap tilsier at lokal bestandsdynamikk er et fremtredende mønster, både for torsk og brisling. Tilsvarende genetisk informasjon er underveis for en rekke marine arter. Dette krever lokale tiltak som monner. Torsk og andre bunnfiskers mulighet til å fylle en rolle i fjordens økosystem vil i overskuelig framtid avhenge av hvilke tiltak som innføres innenfor fiskeri- og miljøforvaltningen. Torskebestanden i Øresund – der bunntråling har vært forbudt siden 1932 – har vist at en lokal torskebestand med bred alders- og størrelsessammensetning kan bestå gjennom perioder preget av ugunstig miljø og dårlig rekruttering (Lindegren m.fl. 2010, Sundelöf m.fl. 2013).
4 - Tiltak for gjenoppbygging og restaurering
Basert på oppdatert materiale presentert i denne rapporten og henvisninger til støttelitteratur legges følgende kunnskap til grunn for anbefalinger: Høyt fiskepress gjennom de siste 100 år og teknologisk utvikling av fiskeriene siden 1950 er en sentral årsak til flere fiskearters sterke tilbakegang i Oslofjorden/ indre Skagerrak. Mangel på lokal fiskeriforvaltning har ført til gradvis utfisking av lokale bestander, der tap av eldre og større individer av toppredatorer har resultert i et fiskesamfunn som domineres av mindre arter («meso-predatorer»).
Utbygginger, utfyllinger, tekniske inngrep og miljøgifter har over tid forringet fiskens leveområder. Samtidig har tilførsel av næringssalter og organisk materiale resultert i økt algevekst og økt oksygenforbruk i utsatte vannmasser. Økte nivåer av næringssalter har bidratt til økt tilgroing av påvekstalger i ålegrasenger og tareskog. Tilførsel av jord- og leirpartikler har bidratt til formørking av kystvannet og svekket lystilgangen til naturen under vann. Dette har hatt konsekvenser for dybdeutbredelsen av tareskog som er blitt sterkt redusert i løpet av de senere tiårene. Denne trenden forsterkes nå av klimaendringene (milde vintre, økt nedbørintensitet, økt avrenning).
Disse utfordringene krever oppfølging: Landbruket og kommunene må redusere sine miljøskadelige utslipp til vassdragene på Østlandet og til Oslofjorden. Det er nødvendig med en mer aktiv fiskeriforvaltning slik at beskatningen ikke overstiger produksjonsgrunnlaget, og at den økologiske balansen i fiskesamfunnet får mulighet til å reetableres. Omfanget av tekniske inngrep som utbygginger må reduseres, utslipp av miljøgifter til fjorden må elimineres og eksiterende forekomster av miljøgifter i sjøbunnen bør saneres i henhold til beste tilgjengelig kunnskap.
Klimaendringene vil bidra til å marginalisere kaldtvannsarter, og beskatningstrykk må justeres i tråd med endringene slik de manifesteres i miljøet. Særlig robuste områder, såkalte «miljørefugier» eller «klimarefugier» - er områder med mulighet for å bevare gode og stabile miljøforhold inn i en usikker fremtid. Slike områder må gis prioritet i plassering av null-fiskeområder og bevaringsområder for kystnær torsk og andre sterkt reduserte arter og bestander.
Anbefalte tiltak (under) bygger på kunnskapsgrunnlaget som har framkommet gjennom prosjektet, «Oppdatert kunnskapsstatus om kystnær torsk i Sør-Norge» (Aglen m.fl. 2016) og «Forslag til helhetlig plan for Oslofjorden» (Miljødirektoratet 2019). Med den tilgjengelige kunnskapen som er innhentet i prosjektet så er det sannsynlig at disse tiltakene vil kunne bremse den negative påvirkningen av Oslofjorden og gi grunnlag for et friskere økosystem . Noen tiltak er naturbaserte løsninger som kan iverksettes umiddelbart, mens andre krever mer planlegging og utredning. Noen av tiltakene krever i mindre grad økonomi, mens andre er mer kostbare og avhengig av både politisk mot og vilje.
4.1 - Fiskeriforvaltning
Regulering av høsting/ fiskerier:
Etablering av et nettverk av nullfiskeområder, særlig viktige gyte- og oppvekstområder
Redusere omfanget av bunntråling i Oslofjorden gjennom revisjon av dybdegrense (i dag: dypere enn 60 m øst for Jærens rev, Jærens rev til Nord-Trøndelag 100 m, nord for Nord-Trøndelag: 170 m)
Etablering av større trålfrie soner i Oslofjorden og indre Skagerrak
Krav om sorteringsrist uten oppsamlingspose der reketråling forekommer i fremtiden, for i størst mulig grad å beskytte store individer av bunnfisk
Fortsatt forbud mot bunngarn
Innføre maksimumsmål for fangst av viktige arter rovfisk/ storvokste og langlivete toppredatorer
Innføre forbud eller ytterligere restriksjoner mot lysfiske etter brisling i Oslofjorden
Innføre generelt forbud mot ikke-nedbrytbare fiskeredskaper slik at tapte fiskeredskaper ikke fortsetter å fiske
Innføring av et registreringssystem for fiskere i sjøområder etter samme mønster som i ferskvannsfiske og hummerfiske
Anbefalinger gitt i «Oppdatert kunnskapsstatus om kystnær torsk i Sør-Norge» (Aglen m.fl. 2016):
Ekstraordinære tiltak Telemark-Østfold:
1. Generelt forbud mot fiske av torsk innenfor definerte fjordlinjer*
a. Forbud mot garnfiske innenfor definerte fjordlinjer
*Rapporten anbefalte nedsetting av en egen arbeidsgruppe for å definere fjordlinjer fra Stad til svenskegrensen.
Framtidig dokumentering av effekter av reguleringene
Det er en kontinuerlig utfordring å evaluere effekter av reguleringstiltak, da de ofte er vanskelig å skille fra naturlige svingninger. For å forstå bedre hvilken effekt fiske har på torskebestanden og kystøkosystemene, og videre for å analysere effekter av forvaltningstiltak, anbefales det å etablere et utvalg av representative referanseområder langs kysten. Innenfor referanseområdene vil det være forbud mot høsting, slik at fiskedødeligheten er lik null. Det anbefales at det opprettes tre referanseområder på Vestlandet og tre i Skagerrak, da det vil være variasjoner i hvordan ulike områder responderer på høstingsforbud (Moland m.fl. 2013). For at referanseområdene skal kunne beskytte ulike livsfaser uten for store lekkasjeeffekter, bør områdene være av en viss størrelse. Som et minimum bør hvert område dekke rundt 5 km2**.På denne måten gjøres arbeidet i et adaptivt rammeverk der man er i stand til å evaluere utviklingen underveis og kan justere tiltak for å nå målsettingen om restaurering av levedyktige bestander med bred alders- og størrelsessammensetning.
**Denne rapporten konkluderer med at referanseområder (nullfiskeområder for effektstudier) må være større enn dette og designet som transekt fra land til største dyp (se Kapittel 3.1 - Konsekvenser av samlet beskatningstrykk i ytre Oslofjord).
4.2 - Miljøtiltak
Påvirkninger fra landsiden:
Redusere tilførsler av organisk materiale og næringssalter fra jordbruksarealene i Oslofjordens nedbørsfelt på Østlandet
Ambisjonsnivået i vanndirektivarbeidet må løftes betraktelig. Det må tas i bruk forskrifter for jordbruksområdene under marin grense, da det har gått for sakte med den frivillige innsatsen.
Forbud mot høstpløying under marin grense, økt bruk av grønne belter og fangdammer langs vassdragene er blant de viktige tiltakene.
Redusere tilførsel av næringssalter fra kommunale avløp i fjordens nedbørsfelt på Østlandet
Investeringer i infrastrukturen for kloakkhåndtering må trappes opp over hele Østlandet.
Det er viktig å sette i gang pilotprosjekter for «Utslippsfrie boliger». Teknologien er kjent og utprøvd. «Frisk Oslofjord» har sendt inn brev om dette til KLD våren 2020.
Redusere tilførsel av miljøgifter fra avløpsnettet. Strengere krav til oppsamling av spesialavfall og spyling av båter under vannlinjen i båthavnene langs fjorden
Det har lenge vært på trappene å innføre strengere krav til håndtering av spesialavfall i båthavnene, særlig bunnstoff. Dette er ikke systematisk iverksatt og må forseres gjennom krav til lokale havner
Redusere tekniske inngrep i det marine miljø generelt og gruntvannsområdene spesielt:
Kystkommunene må gjennom operative planer vektlegge det marine miljøet sterkere. Operative planer må innføres med strengere regler for gjennomføring av tekniske inngrep i marine miljøer:
Nye brygge- og havneområder
Mudring og sprengning
Deponering av masser
Moringer og moringhavner
4.3 - Andre restaureringstiltak
Andre restaureringstiltak:
Restaurere sedimenter i båthavner med stor forekomst av miljøgifter
Dette representerer store kostnader og Staten må trolig gi bistand til kommunene, eks. Sandefjord kommune
Prøve ut teknikker for reetablering av tareskog
Eksempler på dette er «grønn grus» og kunstig utplassering av andre voksemedier for tare
Prøve ut restaurering av ålegrasenger
De viktigste tiltakene vil være reduksjon av næringssalter og restaurering av rovfiskbestandene. Andre aktuelle tiltak vil være utplanting av ålegress.
Det må ryddes opp i tilfeldig og ikke omsøkte tekniske inngrep, bl.a. moringer
Havneloven, plan- og bygningsloven og grunneiers rettigheter i grunntvannsområder må brukes mer aktivt som virkemidler
Sjøørreten har gått sterkt tilbake de seneste årene. Det må innføres flere tiltak for å kunne restaurere denne viktige ressursen:
Styrke arbeidet med å restaurere vannkvalitet og gyteforhold i sjøørretførende bekker: vegetasjonsbelter og fangdammer langs vassdrag og bekker, fjerne oppgangshindre, utlegging av gytegrus osv.
Det må innføres begrensninger på fisket i sjø, særlig i perioden mars – mai.
Registrering og opprydding av tapte fiskeredskaper. Det ligger store mengder av tapt fiskeredskap på fjordbunnen.
Det pågår flere frivillige prosjekter og Fiskeridirektoratet har satt i gang et prosjekt, som del av Frisk Oslofjord. Et helhetlig prosjekt gjennomført i Raet nasjonalpark har høstet nyttige erfaringer med tiltak for begrensning og opprydding av tapte redskaper.
5 - Referanser
Aglen, A., Nedreaas. K., Moland, E., Knutsen, H., Kleiven, A.R., Johannessen, T., Wehde, H., Jørgensen, T., Espeland, S.H., Olsen, E.M., Knutsen, J.A. 2016. Oppdatert kunnskapsstatus om kystnær torsk i Sør-Norge. Fisken og havet, særnr. 4-2016. Havforskningsinstituttet, Bergen.
Aksnes D.L., Dupont, N., Staby, A., Fiksen, Ø., Kaartvedt, S. Are, J. 2009. Coastal water darkening and implications for mesopelagic regime shifts in Norwegian fjords. Marine Ecology Progress Series 387:39-49
Arneberg, P., van der Meeren, G.I. og Frantzen, S. (red.). 2018. Status for miljøet og ytre påvirkning i Nordsjøen og Skagerrak – rapport fra Overvåkingsgruppen 2018. Fisken og Havet, særnummer 3-2018, Havforskningsinstituttet.
Aure, J., Danielssen, D.S, Naustvoll, L.J. 2014. Miljøundersøkelser i norske fjorder: Ytre Oslofjord 1937-2011. Fisken og Havet nr. 5. 2014
Baden, S., Boström, C., Tobiasson, S., Arponen, H., & Moksnes, P. O. 2010. Relative importance of trophic interactions and nutrient enrichment in seagrass ecosystems: A broad‐scale field experiment in the Baltic−Skagerrak area. Limnology and Oceanography 55:1435-1448
Baden, S., Emanuelsson, A., Pihl, L., Svensson, C. J., & Åberg, P. 2012. Shift in seagrass food web structure over decades is linked to overfishing. Marine Ecology Progress Series 451: 61-73
Barceló, C., Ciannelli, L., Olsen, E. M., Johannessen, T., & Knutsen, H. (2016). Eight decades of sampling reveal a contemporary novel fish assemblage in coastal nursery habitats. Global change biology 22:1155-1167
Barth, J. M., Villegas‐Ríos, D., Freitas, C., Moland, E., Star, B., André, C., Knutsen, H., Bradbury, I., Dierking, J., Petereit, C., Righton, D., Metcalfe, J., Jakobsen, K. S., Olsen, E. M., & Jentoft, S. (2019). Disentangling structural genomic and behavioural barriers in a sea of connectivity. Molecular ecology 28:1394-1411
Broch, H., Huitfeld-Kaas, H., Ruud, J. 1932. Beretning 1932. Foreningen til fremme av fiskeriet i Oslofjorden innenfor Drøbak. Oslo 1933, A.W. Brøggers Boktrykkeri, 24 p.
Cardinale, M., Svenson, A., Hjelm, J. 2017. The "easy restriction" syndrome drive local fish stocks to extinction: The case of the management of Swedish ocastal populations. Marine Policy 83:179-183
Costalago, D., Bauer, B., Tomczak, M.T., Lundström, K., Winder, M. 2019. The necessity of a holistic approach when managing marine mammal–fisheries interactions: Environment and fisheries impact are stronger than seal predation. Ambio 48:552–564. doi:10.1007/s13280-018-1131-y
Collett, R. 1868. Zoologisk botaniske Observationer fra Hvaløerne. Nyt Magazin for Naturvidenskaberne 15:1-83
Deininger, A., Frigstad, H. 2019. Reevaluating the role of organic matter sources for coastal eutrophication, oligotrophication, and ecosystem health. Frontiers in Marine Science 6:210 doi:10.3389/fmars.2019.00210
Deininger, A., Kaste, Ø., Frigstad, H. et al. 2020. Organic nitrogen steadily increasing in Norwegian rivers draining to the Skagerrak coast. Scientific Reports 10:18451 doi:10.1038/s41598-020-75532-5
Duarte, C.M., Agusti, S., Barbier, E., et al. 2020. Rebuilding marine life. Nature 580:39-51 doi:10.1038/s41586-020-2146-7
Eriksson, B. K., Ljunggren, L., Sandström, A., Johansson, G., Mattila, J., Rubach, A., ... & Snickars, M. 2009. Declines in predatory fish promote bloom‐forming macroalgae. Ecological Applications 19:1975-1988
Fagerli, C.W., Trannum, H.C., Staalstrøm, A., Eikrem, W., Sørensen, K., Marty, S., Frigstad, H., Gitmark, J. 2019. ØKOKYST – DP Skagerrak. Årsrapport 2019. ØKOKYST – DP Skagerrak. 2019 report. Miljødirektoratet M-1603-2020
Fenberg, P.B., Caselle, J., Claudet, J., Clemence, M., Gaines, S., García-Charton, J.A., Gonçalves, E.J., Grorud-Colvert, K., Guidetti, P., Jenkins, S., Jones, P.J.S., Lester, S., McAllen, R., Moland, E., Planes, S., Sørensen, T.K. 2012. The science of European marine reserves: status, efficacy, and future needs. Marine Policy 36:1012-1021
Fernandez-Chacon, A., Moland, E., Espeland, S.H., Kleiven, A.R., Olsen, E.M. 2015. Demographic effects of partial versus full protection: inference from an empirical before-after control-impact study. Journal of Applied Ecology 52:1206-1215 doi:10.1111/1365-2664
Filbee-Dexter, K., Wernberg, T., Grace, S.P. J. Thormar, S. Fredriksen, C. N. Narvaez, C. J. Feehan & K. M. Norderhaug. 2020. Marine heatwaves and the collapse of marginal North Atlantic kelp forests. Scientific Reports 10:13388. doi:10.1038/s41598-020-70273-x
Fredriksen, Å., Johansen, P.-J. 1999. Storskarven Phalacrocorax carbo sinensis i Øra naturreservat 1999. Natur i Østfold 18:168-170
Frigstad, H., Andersen, T., Hessen, D.O., Jeansson, E., Skogen, M., Naustvoll, L.J., Miles, M.W., Johannessen, T., Bellerby, R.G.J. 2013. Long-term trends in carbon, nutrients and stoichiometry in Norwegian coastal waters: Evidence of a regime shift. Progress in Oceanography 111:113-124
Frigstad, H., Andersen G.S., Trannum, H.C., Naustvoll, L.J., Kaste, Ø., Hjermann, D.Ø. 2018. Synthesis of climate relevant results from selected monitoring programs in the coastal zone. Part 2: Quantitative analyses, NIVA rapport. ISBN 978-82-577-7046-4. No 7311 (54 sider)
Frigstad, H., Kaste, Ø., Deininger, A., Kvalsund, K., Christensen, G., Bellerby, R.G.J., Sørensen, K., Norli, M., King, A. 2020. Influence of riverine input on Norwegian coastal systems. Frontiers in Marine Science 7:332 doi:10.3389/fmars.2020.00332
Funk, B.M. 2013. BRUVs vs. Teiner: Hvilken metode er best egnet til å overvåke effektene av bevaring på torsk (Gadus morhua) i Norge? Universitet i Agder, Bacheloroppgave, 32 pp.
Green, N.W., Schøyen, M., Øxnevad, S., Ruus, A., Allan, I., Hjermann, D., Severinsen, G., Høgåsen, T., Beylich, B., Håvardstun, J., Lund, E., Tveiten, L., Bæk, K. 2015. Contaminants in coastal waters of Norway 2014, NIVA-rapport 6917.
Gustavsen, F. 2017. Great cormorant Phalacrocorax carbo diet related to the supply of marine shallow-living fish species in southeastern Norway; opportunistic or selective foraging? Master’s Thesis Norges miljø- og biovitenskapelige universitet, 33 s.
Halpern, B.S., Walbridge, S., Selkoe, K.A., et al. 2008. A global map of human impact on marine ecosystems. Science 319, 948–952
Halvorsen, K.T., Larsen, T., Sørdalen, T.K., Vøllestad, L.A., Knutsen, H., Olsen, E.M. 2017. Impact of harvesting cleaner fish for salmonid aquaculture assessed from replicated coastal marine protected areas, Marine Biology Research 13:359-369
Haug, L.M. H. 2013. Vurdering av BRUVS som metode for studier på artsdiversitet og torskebestand (Gadus morhua), Universitet i Agder, Bachloroppgave, 41 pp.
IPBES 2019. Global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. E. S. Brondizio, J. Settele, S. Díaz, and H. T. Ngo (editors). IPBES secretariat, Bonn, Germany.
Johannessen, T., Dahl, E., Falkenhaug, T., Nastvoll, L. J. 2012. Concurrent recruitment failure in gadoids and changes in the plankton community along the Norwegian Skagerrak coast after 2002. ICES Journal of Marine Science, 69:795-801.
Kleiven, A.R., Nordahl, J-H., Moland, E., Espeland, S.H., Knutsen, H., Olsen. E.M. 2016. Harvest pressure on coastal Atlantic cod (Gadus morhua) from recreational fishing relative to commercial fishing assessed from tag-recovery data. PLoS ONE e0149595. doi:10.1371/journal.pone.0149595
Kleiven, A. R., Moland, E., and Sumaila, U. R. 2019. No fear of bankruptcy: the innate self-subsidizing forces in recreational fishing. ICES Journal of Marine Science 77:2304-2307 doi:10.1093/icesjms/fsz128.
Knutsen, H., Jorde, P. E., Hutchings, J. A., Hemmer‐Hansen, J., Grønkjær, P., Jørgensen, K. E. M., André, A., Sodeland, M., Albretsen, J., & Olsen, E. M. 2018. Stable coexistence of genetically divergent Atlantic cod ecotypes at multiple spatial scales. Evolutionary Applications 11:1527-1539
Kraufvelin, P., Svensson, F., Fredriksson, R., Bergström, L., Karlsson, M., Wennhage, H., Wikström, A., Bergström, U. 2017. Inventering och modellering av fisk-och kräftdjurssamhällen i Kosterhavets nationalpark. Länsstyrelsen Västra Götaland, Göteborg. Pp: 29-41; 65-75
Kroodsma, D.A., Mayorga, J., Hochberg, T., et al. 2018. Tracking the gobal footprint of fisheries. Science 359: 904–908
Pedersen, T., Strand, H.K., Christie, H., Moy, F. 2020. Predation mortality from ambush and cruising predators on newly-settled 0-group gadoids. Journal of Experimental Marine Biology and Ecology 529:151396
Langlois TJ, Fitzpatrick BR, Fairclough DV, Wakefield CB, Hesp SA, et al. 2012. Similarities between Line Fishing and Baited Stereo-Video Estimations of Length-Frequency: Novel Application of Kernel Density Estimates. PLoS ONE 7:e45973 doi:10.1371/journal.pone.0045973.
Lekve, K., Stenseth, N. C., Gjøsæter, J., Fromentin, J. M., & Gray, J. S. 1999. Spatio-temporal patterns in diversity of a fish assemblage along the Norwegian Skagerrak coast. Marine Ecology Progress Series 178:17-27
Lennox, R.J., Gallagher, A.J., Ritchie, E.G., Cooke, S.J. 2018. Evaluating the efficacy of predator removal in a conflict-prone world. Biological Conservation 224:227-289
Lindegren, M., Diekmann, R., Möllmann, C. 2010. Regime shifts, resilience and recovery of a cod stock. Marine Ecology Progress Series 402: 239-253 doi: 10.3354/meps08454
Lorentsen, S.-H. 2013. Status of the breeding population of the P. c. sinensis subspecies of Great Cormorants in Norway in 2012. Page 75- 78 in IUCN/Wetlands International Cormorant Research Group Report: National reports from the 2012 breeding census of great cormorants Phalacrocorax carbo in parts of the Western palearctic. Technical report nr 22 from Danish Centre for Environment and Energy.
Lorentsen, S.-H., Anker-Nilssen,T., Barett, R.T., Systad, G.H. 2021. Population status, breeding biology and diet of Norwegian Great Cormorants. Ardea, i trykk.
Lunneryd, S. G., Alexandersson, K. 2005. Födoanalyser av storskarv, Phalacrocorax carbo i Kattegatt- Skagerrak. Finfo (Fiskeridirektoratet informerar), rapport 2005:11, 21 s.
Miljødirektoratet. 2019. Forslag til helhetlig plan for Oslofjorden. M-1550 2019, 167 s.
Miljøverndepartementet. 2010. Internasjonale mål vedtatt på FNs tiende partsmøte under konvensjonen om biologisk mangfold i Nagoya, Japan, 2010. (UNEP/CBD/COP/DEC/X/2)
Moksnes, P. O., Gullström, M., Tryman, K., & Baden, S. 2008. Trophic cascades in a temperate seagrass community. Oikos 1175:763-777
Moland, E., Olsen, E.M., Knutsen, H., Garrigou, P., Espeland, S.H., Kleiven, A.R., André, C. Knutsen, J.A. 2013. Lobster and cod benefit from small scale northern marine protected areas: inference from an empirical before-after control-impact study. Proceedings of the Royal Society B Biological Sciences 280:20122679
Nellemann, C., Hain, S., Alder, J. (Eds). 2008. In Dead Water – Merging of climate change with pollution, over-harvest, and infestations in the world’s fishing grounds. United Nations Environment Programme, GRID-Arendal, ISBN: 978-82-7701-048-9, 64 s.
Obst, M., Vicario, S., Lundin, K., Berggren, M., Karlsson, A., Haines, R., Williams, A., Goble, C., Mathew, C., Güntsch, A. 2018. Marine long-term biodiversity assessment suggests loss of rare species in the Skagerrak and Kattegat region. Marine Biodiversity 48:2165-2176. doi:10.1007/s12526-017-0749-5
Olsen, M.T., Galatius, A., Härkönen, T. 2018. The history and effects of seal−fishery conflicts in Denmark. Marine Ecology Progress Series 595:233-243
Ono K, Knutsen H, Olsen EM, Ruus A, Hjermann DØ, Chr. Stenseth N. 2019 Possible adverse impact of contaminants on Atlantic cod population dynamics in coastal ecosystems. Proceedings of the Royal Society B: Biological Sciences 286:20191167 doi:10.1098/rspb.2019.1167
Pedersen, T., Strand, H.K., Christie, H., Moy, F. 2020. Predation mortality from ambush and cruising predators on newly-settled 0-group gadoids. Journal of Experimental Marine Biology and Ecology 529:151369
Quintela, M., Kvamme, C., Bekkevold, D., Nash, R.D.M., Jansson, E., Sørvik, A.G., Taggart, J.B., Skaala, Ø., Dahle, G., Glover, K.A. 2020. Genetic analysis redraws the management boundaries for the European sprat. Evolutionary Applications 19:1906-1922
Rasch, H. 1838. Fortegnelse og Bemerkninger over de i Norge forekommende Fugle. Nyt Magazin for Naturvidenskaberne 1:357-398
Rijnsdorp, A. D., Hiddink, J. G., van Denderen, P. D., Hintzen, N. T., Eigaard, O. R., Valanko, S., Bastardie, F., Bolam, S. G., Boulcott, P., Egekvist, J., Garcia, C., van Hoey, G., Jonsson, P., Laffargue, P., Nielsen, J. R., Piet, G. J., Sköld, M., and van Kooten, T. 2020. Different bottom trawl fisheries have a differential impact on the status of the North Sea seafloor habitats. ICES Journal of Marine Science 77:1772-1786
Ruud, J. 1968. Changes since the turn of the century in the fish fauna and the fisheries of the Oslofjord. Helgoländer wiss. Meeresunters. 17:510-517
Schøyen M, Allan IJ, Ruus A, Håvardstun J, Hjermann D, Beyer J. 2017 Comparison of caged and native blue mussels (Mytilus edulis spp.) for environmental monitoring of PAH, PCB and trace metals. Marine Environmental Research 130: 221-232 doi:10.1016/j.marenvres.2017.07.025
Skarprud. M. 2003: Sommerføden til storskarven Phalacrocorax carbo i Øra naturreservat, Fredrikstad. Cand. scient hovedoppgave, Norges landbrukshøgskole, Ås. 43 s.
Skjæveland, S. B. 2020. Kvalitativ og kvantitativ inventering av fiskesamfunnet i Ytre Oslofjord – en beskrivelse av diversitet og generelle trekk ved fiskesamfunnet i Færder- og Ytre Hvaler nasjonalparker for perioden 2017-2019. MSc - Masteroppgave ved Universitet i Agder
Sodeland, M., Jorde, P. E., Lien, S., Jentoft, S., Berg, P. R., Grove, H., Kent, M. P., Arnyasi, M., Olsen, E. M., & Knutsen, H. (2016) “Islands of Divergence” in the Atlantic cod genome represent polymorphic chromosomal rearrangements. Genome biology and evolution 8:1012-1022
Sundelöf, A., Wennhage, H., and Svedäng, H. 2013. A red herring from the Öresund (ICES40G2): the apparent recovery of the Large Fish Indicator (LFI) in the North Sea hides a non-trawled area. ICES Journal of Marine Science 70:1081-1084
Synnes, A.E.W. 2020. Seascape ecology of Atlantic cod (Gadus morhua) in coastal Skagerrak: population structure, connectivity, and role in fish assemblage. PhD - Doktorgradavhandling, Universitetet i Agder
Sørensen, A. 2012: Sommerdiett hos storskarv (Phalacrocorax carbo sinensis) i Øra naturreservat 15 år etter kolonietablering. Masteroppgave, Universitetet for Miljø- og Biovitenskap, 33 s.
Sørlie, M., Nilssen, K.T., Bjørge, A., & Freitas, C. 2020. Diet composition and biomass consumption of harbour seals in Telemark and Aust-Agder, Norwegian Skagerrak, Marine Biology Research 16:299-310 doi:10.1080/17451000.2020.1751205
Thingstad, T. F., Bellerby, R.G.J., Bratbak, G., Borsheim, K.Y., Egge, J.K., Heldal, H., Larsen, A., Neill, C., Nejstgaard, J., Norland, S., Sandaa, R.A., Skjoldal, E.F., Tanaka, T., Thyrhaug, R., Topper. B. 2008. Counterintuitive carbon-to-nutrient coupling in an Arctic pelagic ecosystem. Nature 455:387-U337
Walday, M., Borgersen, G., Beylich, B., Eikrem, W., Gitmark, J., Naustvoll, L.J., Selvik, J.R. 2019. Overvåking av Ytre Oslofjord i 2014-2018. NIVA rapport l.nr. 7423-2019. 106 s.
Watson DL, Harvey ES, Anderson MJ, Kendrick GA. 2005. A comparison of temperate reef fish assemblages recorded by three underwater stereo-video techniques. Marine Biology 148:415-425 doi:10.1007/s00227-005-0090-6.
Westerberg, H., Rännbäek, P., Frimansson, H. 1996. Effects of suspended sediments on cod egg and larvae and on the behaviour of adult herring and cod, ICES CM 1E (1996)
Wikström, A., Linders, T., Sköld, M., Nilsson, P., Almén, A. 2016. Bottentrålning och resuspension av sediment, Länsstyrelsen i Västra Götalands län, rapport no. 36, 2016.
Östman, Ö., Eklöf, J., Eriksson, B. K., Olsson, J., Moksnes, P. O., & Bergström, U. 2016. Top‐down control as important as nutrient enrichment for eutrophication effects in North Atlantic coastal ecosystems. Journal of Applied Ecology 53:1138-1147