Site structure of aquaculture farms in production zone 4
With focus on conditions in Nordfjord
In this report we model and discuss the connectivity of pelagic salmon lice and viruses among aquaculture-sites in production area 4 (PA4), with a focus on Nordfjord.
Our models indicate moderate to high concentrations of infectious salmon lice across almost the entire production area, except inner parts of the large fjords. We report high concentrations of infectious salmon lice in the areas of Sognesjøen, outer parts of Sognefjorden, Austefjord, Masfjorden, and Frøysjøen as well as in several bays and narrow straits along the coastal landscape in the production area.
A clustering-analysis on the connectivity of salmon lice among localities in the production area reveals 10 clusters of closely connected localities, with a similar distribution of clusters within fjords for lice and viruses. The current production strategy divided into season and even / odd-numbered years does not seem to affect the cluster-structure of the infection network in PO4 to a significant degree.
The ranking of dispersal factors in the production area shows a large geographical variation, with generally high exports of infectious lice from several localities in the areas of Sognesjøen, Austefjord, Masfjorden and Osterfjorden / Sørfjorden.
Specifically for the Nordfjord-Frøysjøen cluster, we observed low connectivity with localities in other clusters, with the exception of a few localities in Frøysjøen. The connections within the Nordfjord-Frøysjøen cluster were dominated by self-infection, especially on the virus side. Any coordinated de-lousing strategies taken within Nordfjord will therefore most likely have a good effect due to low external influence.
Sammendrag
I en bestilling fra Fiskeridirektoratet (Fdir) bes Havforskningsinstituttet (HI) å opparbeide et kunnskapsgrunnlag for en vurdering av lokalitetsstrukturen i Nordfjord. I et påfølgende møte mellom Fdir og HI den 16/11-20 ble det avtalt at HI skulle utføre en konnektivitetsanalyse mellom lokaliteter i produksjonsområde 4 (PO4) ut i fra dagens lokalitetsstruktur og lusenivåer–et såkalt «null-scenario».
Havforskningsinstituttet sine modeller indikerer moderate eller høye konsentrasjoner av infektive lakselus i stort sett hele PO4, unntatt indre deler av de store fjordene. Vi rapporterer høye konsentrasjoner av infektive lakselus i områdene Sognesjøen, ytre deler av Sognefjorden, Austefjord, Masfjorden, og Frøysjøen samt i flere viker og trange sund langs kystlandskapet i PO4.
Smittenettverket for lakselus i PO4 ser ut til å være delt opp i 10 klynger av tett sammenkoblede lokaliteter, med generelt lignende fordeling av klynger innad i fjorder for lus og virus. Nåværende utsett-strategi fordelt på sesong og partall/oddetalls-år ser ikke ut til å påvirke klyngestrukturen til smittenettverket i PO4 i nevneverdig grad.
Rangeringen av smittefaktorer i PO4 viser stor geografisk variasjon, med generelt høy eksport av smitte fra flere lokaliteter i områdene Sognesjøen, Austefjord, Masfjorden og Osterfjorden/Sørfjorden.
Spesifikt for klyngen Nordfjord-Frøysjøen observerte vi lav konnektivitet med lokaliteter i andre klynger, med unntak noen få lokaliteter i Frøysjøen. Koblingene innad i Nordfjord-Frøysjøen-klyngen var dominert av selv-smitte, og da særlig på virus-siden. Ethvert koordinert tiltak innad i Nordfjord vil derfor mest sannsynlig gi god effekt grunnet lav påvirkning utenfra.
1 - Bakgrunn og innfallsvinkel
Med bakgrunn i Havforskningsinstituttets (HI) og Veterinærinstituttets rapport “Endret lokalitetsstruktur i produksjonsområde 3” (Husebråten m. fl. 2020) har Fiskeridirektoratet (Fdir) etterspurt kunnskapsgrunnlag for tilsvarende analyser for produksjonsområde 4 (PO4).
Bestillingen etterspurte spesifikt det samme grunnlag som ble brukt i prosjektet med produksjonsområde 3, heretter kalt PO3 prosjektet – men grunnet knapp tidshorisont var Fdir åpen for justeringer i omfanget av data som skal leveres slik at grunnlaget ville bli tilstrekkelig for å svare opp mot prosjektet «Nordfjord 2020».
Det ble på møtet mellom Fdir og HI 16/11-20 besluttet at man i første omgang skulle simulere et «null-scenario». Dette vil si en konnektivitetsanalyse ut i fra dagens struktur og lusenivåer, og at at null-scenarioet skulle kjøres for i) lakselus ii) SAV/PD og iii) ILA.
I bestillingen ble det ytret ønske om at arbeidet skulle begrenses til området fra Frøysjøen i sør, til Sildagapet i nord. Forskningsgruppe Oseanografi og Klima ved HI hadde på dette tidspunktet allerede opparbeidet skript og data, samt etablert arbeidsflyt for å svare på bestillingen gjennom tidligere prosjekt «Lokalitetsstruktur PO3». I utgangspunktet ville alle avvik fra tidligere arbeidsflyt innebære merarbeid og der all eventuell skreddersøm inn mot det mindre området i prosjekt «Nordfjord 2020» vil redusere generaliteten av hva som potensielt kunne leveres for hele PO4 innenfor rammene av prosjektet. Det ble også sett på som høyst trolig at lokaliteter i området mellom Frøysjøen og Sildagapet ikke burde sees på i isolasjon, da det var forventet høyt smittepress fra oppstrøms lokaliteter, altså sørfra.
Det er derfor anbefalt fra HI sin side å ikke avgrense analysene til Nordfjord og området i dens umiddelbare nærhet, men heller utføre tilsvarende analyse som i PO3 for hele PO4, der et uttrekk fra dette datamaterialet så kan brukes som kunnskapsgrunnlag i «Nordfjord 2020» prosjektet.
En eventuell analyse av nye driftsformer, f.eks nye sonevise utsett, innføring av lukkede eller semi-lukkede anlegg, eller konsultasjoner med næringen og andre interessenter, slik det til dels ble gjort for PO3, ble vurdert. Men dette ville forsinke prosessen og ville ikke ha vært mulig å få til med tilfredsstillende kvalitet innenfor den korte tidsfristen til medio desember 2020. Denne rapporten er derfor begrenset til en analyse av nåsituasjonen, samt tre år tilbake i tid for å fange opp et tilfredsstillende spekter av dynamiske utfall i forhold til havstrømmene. Datamaterialet opparbeidet i forbindelse med denne rapporten er godt egnet for videre analyser av scenarier der lokalitetsstrukturen eller driftsform endres.
Basert på erfaringer og tilbakemeldinger fra næringen i etterkant av PO3-arbeidet er det i denne analysen brukt nauplieproduksjon i “sanntid”, dvs. modellert klekking av egg på hver lokalitet basert på lusetall, temperatur, og antall fisk som til enhver tid var rapportert til Mattilsynet (i motsetning til gjennomsnittlig nauplieproduksjon over tidsperioden, slik som i PO3-arbeidet). Der er også tatt høyde for at en lokalitet som i korte perioder står brakk (grunnet sonevise utsett eller andre problemer på lokaliteten) ikke mottar lus fra andre lokaliteter. Slik sett får man indirekte testet om dagens koordinerte utsett-strategi gir en observerbar effekt på topologien til smittenettverket i produksjonsområdet i sin helhet. Ulempen med denne innfallsvinkelen er at lokalitetens kvaliteter i mindre grad blir vurdert (altså, lokalitetens spredningspotensiale gitt geografisk posisjon i strømsystemene), til fordel for en mer “realistisk” gjenspeiling av driften på hver enkelt lokalitet.
I denne rapporten har kun lokaliteter med kommersiell lakseoppdrett i sjø blitt vurdert. Enkeltlokaliteter som har skiftet navn eller blitt flyttet i studieperioden er heller ikke med i analysene*.
*Merk at lokalitet Endal Ø 12236 ble flyttet og skiftet navn under studieperioden og er ikke med i analysen.
2 - Materialer og metoder
2.1 - Hav og spredningsmodell
I denne rapporten er det benyttet seg av en hydrodynamisk sirkulasjonsmodell som grunnstein for analyser av spredning. Denne havmodellen gir en beskrivelse av det fysiske miljøet (blant annet strømhastighet, temperatur og saltholdighet) for hele produksjonsområdet. Modellen inkluderer tidevann, vindpådrag, ferskvannsavrenning fra land og utvekslingsprosesser mellom fjord og havet utenfor. For detaljert beskrivelse av havmodellen se Albretsen m.fl. (2011).
For å simulere spredning av pelagiske lakselus og virus i det horisontale planet brukte vi Runge-Kutta 2. ordens metode basert på strømfeltene fra havmodellen som implementert i partikkelspredningsmodellen LADIM (Lagrangian Advection and DIffusion Model, Ådlandsvik 2020). Simuleringen ble kjørt for perioden 1/4-2017 til 30/9-2020. Her ble det brukt et tidssteg på to minutter og ved hvert tidssteg ble adveksjonen beregnet basert på strømmen hvor hver enkelt partikkel befant seg i med en medregnet tilfeldig horisontal diffusjon på opptil 0,2 m2/s.
Videre er det tilpasset en såkalt «individ-basert modell» (IBM) som forsøker å gjenskape en realistisk adferd til både lakselus og virus. Disse IBM-ene er basert på beste kunnskap om utslipp, utviklingshastighet, dødelighet, smittsomhet og eventuell adferd i forhold til lys og saltholdighet av lakselus.
2.2 - Parametrisering av lakselus-IBM
Basert på ukentlig innrapporterte antall kjønnsmoden hunn-lus per lokalitet modellerte vi nauplieproduksjon på lokalitetene som funksjon av antall fisk per lokalitet, gjennomsnittlig antall lus per fisk og observert temperatur på lokaliteten (Stien m.fl., 2005). Den ukentlige modellerte nauplieproduksjonen på hver lokalitet ble så interpolert til daglig produksjon per lokalitet.
Lakselusens adferd, dødelighet, individuell respons på miljøet og utvikling ble implementert som beskrevet i Sandvik m.fl. (2020). Kort oppsummert ble det implementert en døgnlig vertikal bevegelse som følge av lyspåvirkning, samt en gradvis vertikal unnvikelse for saltholdigheter lavere enn 31. Lusene opplevde en døgnlig dødelighet på 17%. Videre ble infeksjonssannsynligheten til lusen modellert som funksjon av alder (i døgngrader) og temperaturen i den vannmassen individet befant seg i (Skern m.fl. 2020).
2.3 - Parametrisering av virus-IBM
Det er antatt at mengden viruspartikler som blir utskilt fra fisk ved et eventuelt utbrudd av PD eller ILA er skalert med den totale biomassen som til enhver tid står på den smittede lokaliteten. For å utforske alle potensielle smitteveier (i motsetning til å kun modellere dokumenterte tilfeller) ble utslipp av virus i modellen skalert med den månedlige biomassen av laks på lokalitetene som rapportert til Fiskeridirektoratet, med en daglig temporær interpolasjon fra måned til måned. Dette er et kunstig scenario da disse virussykdommene per nå ikke er systemiske, men denne modelleringen vil gi nyttig kunnskap om hvordan smitteveien ville sett ut hvis sykdommen potensielt sprer seg til og fra alle lokaliteter hvis de en dag blir smittet. Videre, da virus i alle praktiske tilfeller kan sees på som passive partikler i vannmassene ble viruspartiklene satt opp til å bevege seg tilfeldig vertikalt i de øvre 20 meterne med opptil 0.001 m2/s, f.eks. som følge av turbulens i vannsøylen.
I litteraturen på virus bruker man halveringstid som mål på dødelighet, dvs. tiden det tar før halvparten av viruset er blitt inaktivt. Hovedprinsippet er at dødeligheten øker med økende temperatur, mens andre miljøfaktorer spiller mindre rolle. Basert på verdiene hentet fra Graham m.fl., 2007 finner vi en lineær sammenheng mellom halveringstid og temperatur for PD, men der variasjonen rundt regresjonslinjen er stor, noe som tilsier at det er stor usikkerhet knyttet til denne halveringstiden. Grunnet få publiserte verdier på halveringstiden til ILA og stor spredning mellom publikasjonene er det knyttet stor usikkerhet til hva halveringstiden til ILA er. I Huserbråten m.fl. (2020) ga simulering av spredning av ILA nær identiske resultater som PD i alle praktiske tilfeller. Grunnet usikkerhet knyttet til halveringstid og smittedose er det i denne rapporten ikke gjennomført en egen simulering for ILA. Det er derfor antatt at resultatene fra PD simuleringene vil gi en tilstrekkelig representasjon av spredningsbilde også for ILA. Regresjonslinjen for halveringstid for virus (altså, for PD der datagrunnlaget er mer solid) er gitt ved t1/2=a-b*T, der a = 8.33, b = -0.29 og T er temperaturen fra havmodellen på stedet der viruset til enhver tid innfinner seg.
2.4 - Nettverksanalyser
Antall lakselus eller virus sluppet ut fra lokalitet A som fortsatt var infektivt og ble registrert innen en radius på 240 m rundt lokalitet B ble så talt opp for alle lokaliteter i en ny N x N matrise (der hver rad og kolonne representerer hver lokalitet i produksjonsområdet) for hver dag av simuleringsperioden. Denne matrisen hvor hvert element viser smitte mellom to lokaliteter per dag, blir heretter referert til som «konnektivitetsmatrisen». Merk at hvis det ikke oppholdt seg fisk på lokalitet B ved det bestemte tids-steget ble det ikke registrert smitte fra lokalitet A til B. Den daglige smitten til hver lokalitet ble så integrert over hele simuleringsperioden og delt på antall dager det sto fisk på den smittede lokaliteten.
Én visuell måte å fremstille konnektivitetsmatrisen på er i såkalte nettverksplot. Her blir hver lokalitet fremstilt som en node i nettverket, og alle forbindelser denne noden har med andre noder i nettverket er representert med x-antall koblinger (markert med buede linjer)avhengig om lokalitet A ble smittet av lokalitet B. Tykkelsen på koblingene representerer da styrken av smitte mellom to lokaliteter. Grunnet mange svake koblinger mellom lokalitetene ble kun de sterkeste koblingene plottet i nettverksplottene.
For å objektivt evaluere klynger av lokaliteter som var sterkere koblet sammen enn andre, ble det foretatt en klyngeanalyse basert på konnektivitetsmatrisen (Rosvall & Bergstrøm 2008). Det betyr på generelt grunnlag at tiltak gjort innad i en klynge vil ha større effekt på lokalitetene i klyngen den tilhører enn på lokaliteter i andre klynger.
3 - Resultater og diskusjon
3.1 - Konsentrasjon av lus i PO4
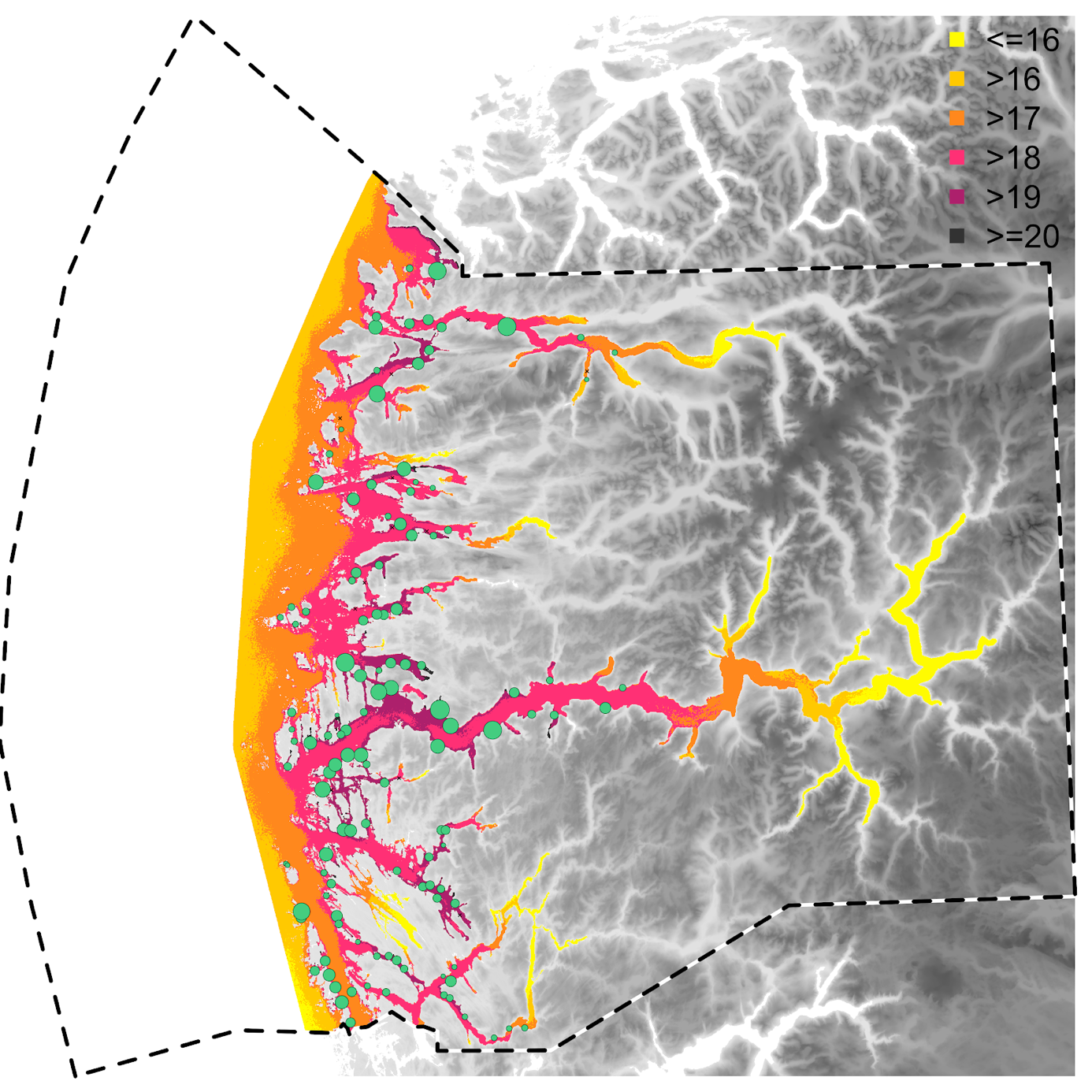
Ser man på konsentrasjonsfeltet av infektive lusestadier i PO4 er det store forskjeller mellom områder med høy og lav konsentrasjon integrert over studieperioden (Figur 1). Generelt kan man si at konsentrasjonen av infektive lusestadier over tid er størst i ytre deler av fjorder og i trange sund langs kystlandskapet, der også tettheten av lokaliteter er størst. Videre ser vi en moderat konsentrasjon lenger inn i fjordarmene, og en lav konsentrasjon innerst i fjordene. Denne fordelingen kan delvis forklares av at det ferskvannspåvirkede overflatelaget i hovedsak beveger seg ut av fjordene og at flere av fjordene er nasjonale laksefjorder som betyr at det generelt er lav tetthet av lokaliteter innerst i fjordene. Langs ytre kyst og på eksponerte områder vil igjen konsentrasjonen være moderat til lav, dette grunnet høy vannutskiftning og dermed høy fortynningseffekt, men også høy spredning nordover med kyststrømmen. Men merk at denne moderate konsentrasjonen i ytre kyst også kan være kunstig lav, tatt i betraktning at utslipp av frittlevende lusestadier i oppstrøms produksjonsområder (i all hovedsak PO3) ikke er inkludert i analysen.
Figur 1 . Konsentrasjonsfelt for lakselus i PO4, integrert over hele simuleringsperioden på 1250 dager (1/4-2017 til 30/9-2020) og kuttet mot grunnlinjen. Her er konsentrasjonen av lus klassifisert inn i seks nivåer (på log skala), der en økning til neste klasse representerer en tre-dobling. Størrelsen på grønne sirkler er skalert etter gjennomsnittlig produksjon av lusenauplier over dagene det har stått fisk på lokaliteten. Små, sorte kryss representerer lokaliteter som står oppført i akvakulturregisteret men som ikke har rapportert lusetall i studieperioden.
3.2 - Smittenettverket i PO4
3.2.1 - Lakselus
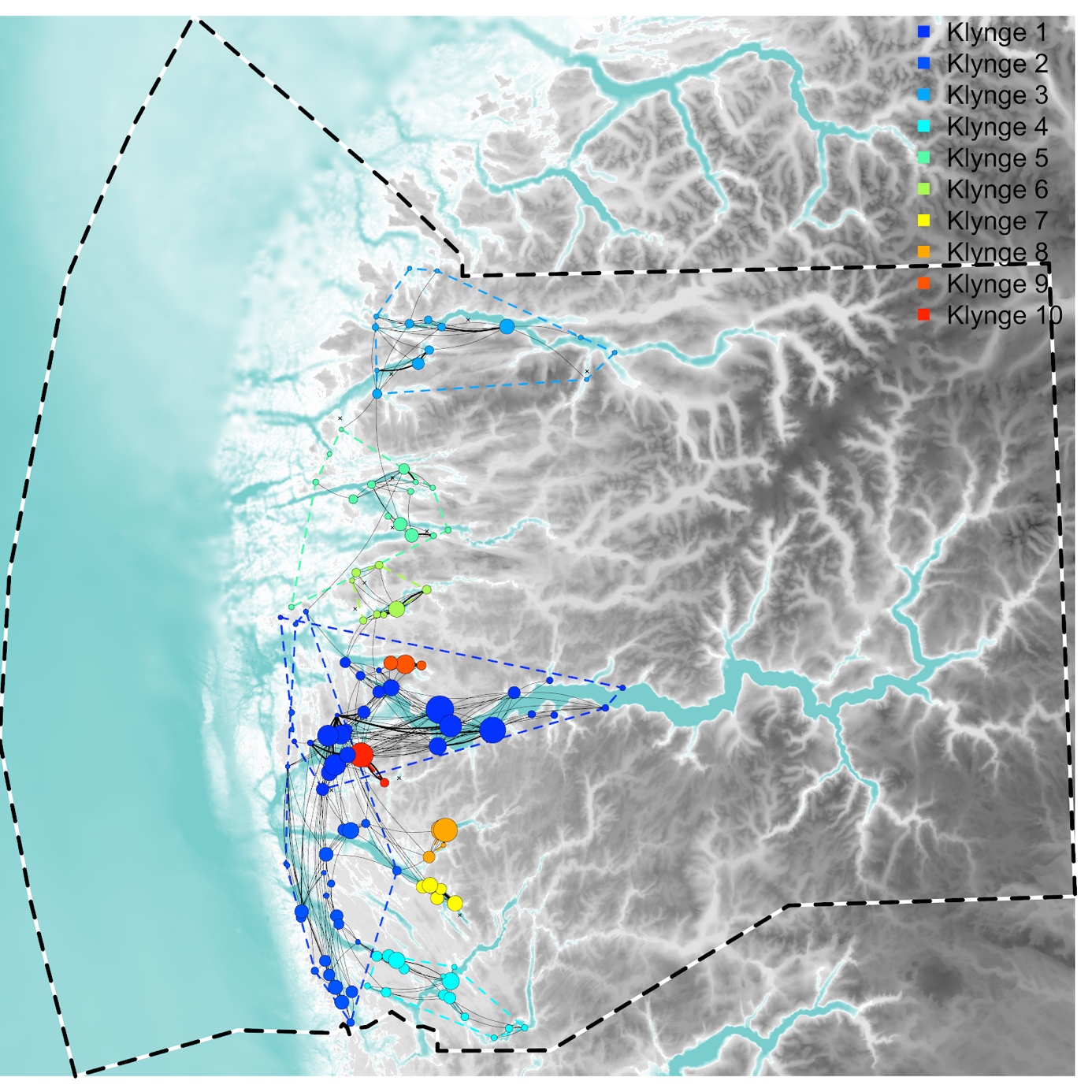
Basert på konnektivitetsmatrisen som oppsummerte lusesmitten mellom alle 122 lokaliteter som hadde rapportert lusetall i perioden 1/4-2017 til 30/9-2020 (av 134 som oppført i akvakulturregisteret under kommersiell lakseoppdrett av matfisk) trer det frem 10 klynger av sterkt sammenkoblede lokaliteter. Fra sør til nord omfavner klyngene lokaliteter fordelt over følgende fjorder og kyststrekninger (med referanse til klyngene i Figur 2): Osterfjorden, Sørfjorden, Herdlefjorden, og Radfjorden (K4); kyststrekningen fra Øygarden til Værlandet og halvveis inn i Austfjorden (K2); innerste del av Austfjord (K7); Masfjorden (K8); Gulafjorden (K10); Sognefjorden og Sognesjøen (K1); Ålfjorden (K9); Dalsfjorden og Stongfjorden (K6); Førdefjorden, Brufjorden, og Høydalsfjorden (K5); og Nordfjord og Frøysjøen (K3).
Det faktum at klyngene i nettverksanalysen i all hovedsak dannes “fjordvis” eller på tvers av kystlandskapet i ytre områder viser at dagens strategi med sonevise utsett i fordelt over sesonger (vår-høst) og år (partal og oddetalsår) ikke påvirker topologien til smittenettverket, og skaper ikke noen nevneverdig isolasjon mellom produksjonssykluser. Hvis en slik strategi hadde hatt en effekt ville vi sett et fragmentert nettverk av små klynger, som ville gjenspeilt de historiske koordinerte utsett-områdene.
Figur 2 . Smitte av lakselus mellom lokaliteter basert på strømmodell og lusetall fra perioden 1/4-2017 til 30/9-2020. Her representerer størrelsen på noder den relative eksporten av lus fra en lokalitet til alle andre og tykkelsen på koblinger den spesifikke eksporten fra en lokalitet til en annen (med klokken), der tykke linjer representerer de 100 sterkeste koblingene og tynne linjer de 500 sterkeste koblingene. Fargen på sirkler indikerer klyngetilhørighet med andre lokaliteter, der lokaliteter innad i klyngen er sterkere koblet med hverandre enn med andre klynger.
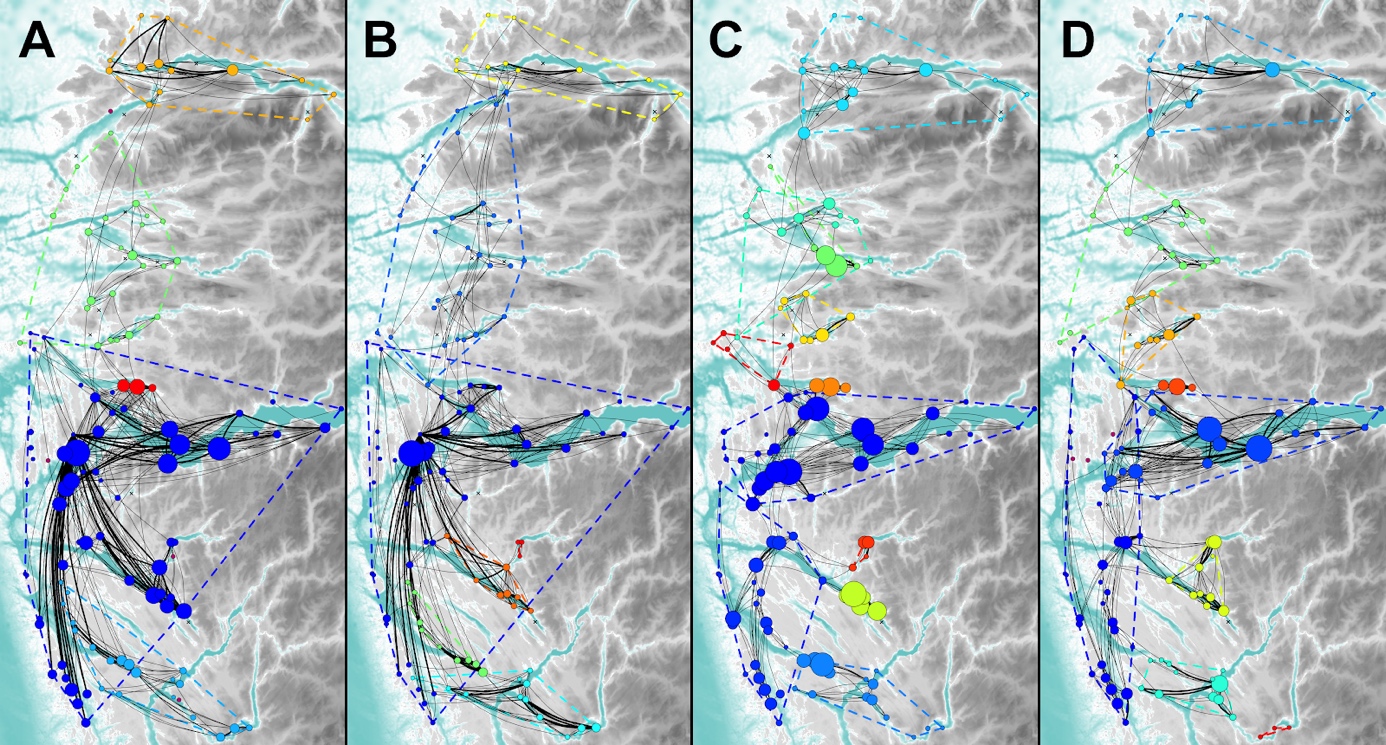
Ser vi på smittenettverket for lus fordelt over sesongene er det påfallende forskjeller på vinter-vår og sommer-høst (Figur 3). Her er den største forskjellen den store klyngen som omfatter lokaliteter på tvers av hele den søndre delen av PO4 (inkludert Sognefjorden) vinter-vår; men som fragmenteres i mindre klynger særlig på sommeren, men også på høsten. Den naturlige forklaringen på denne høye konnektiviteten på tvers av søndre deler av PO4 på vinteren og våren er at dette er tidsperioden hvor vinddrevne havstrømmer generelt er sterkest. Det kommer også klart frem fra analysene at klyngen som omfavner Nordfjord-Frøysjøen er mest stabil gjennom sesongene, da med unntak av noen lokaliteter som skifter klyngetilhørighet grunnet sin noe mer eksponerte plassering for import fra sør.
Figur 3 . Smittenettverk for lus delt opp i sesonger (antall klynger i parentes): A vinter (5 klynger); B vår (7 klynger); C sommer (11 klynger); og D høst (9 klynger). Lilla sirkler i panel A og D represente lokaliteter som ikke er koblet til nettverket, enten grunnet null koblinger til andre lokaliteter eller ingen drift i perioden. Ellers se forklaring Figur 2
3.2.2 - Virus
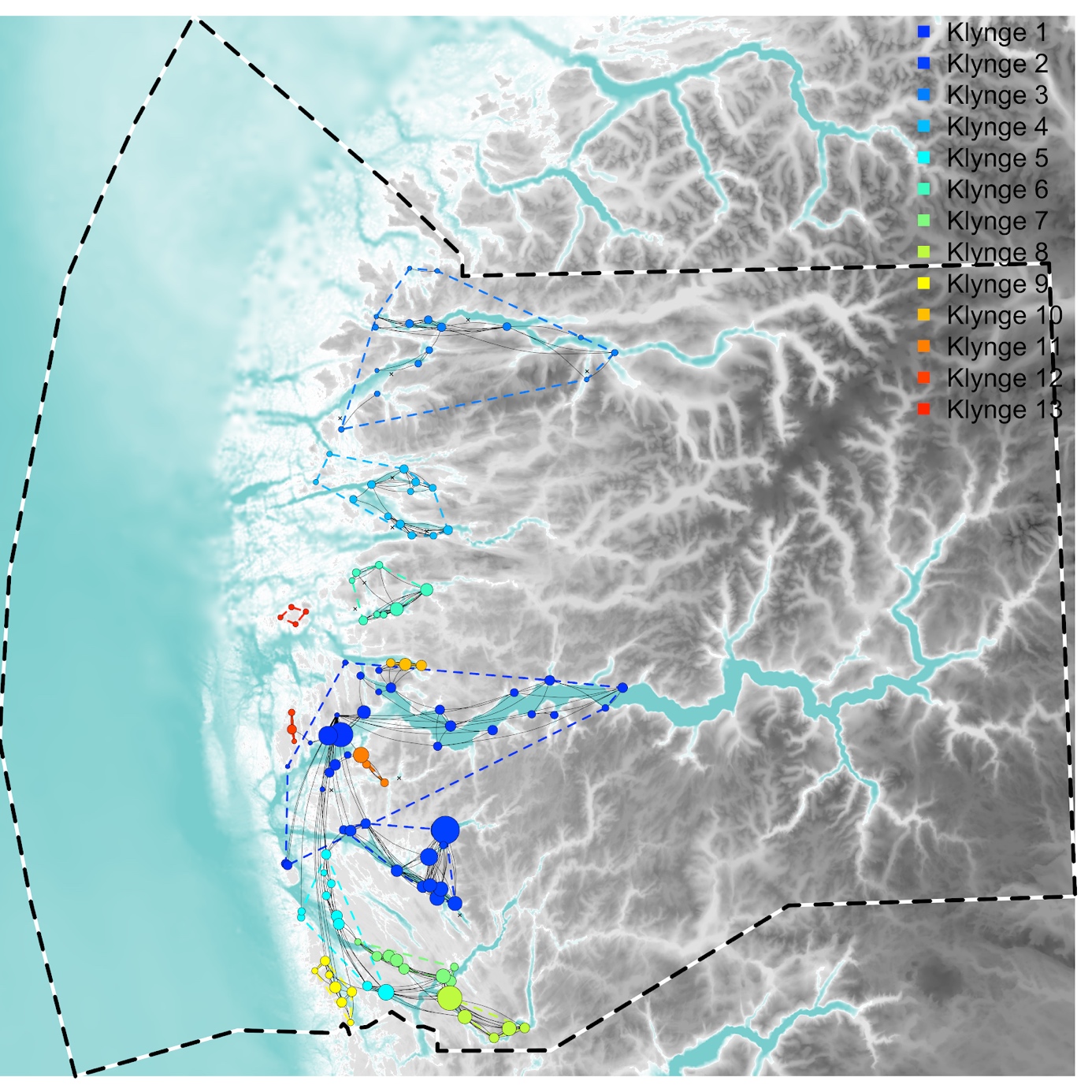
På samme måte som for lus danner virus-klyngene seg i høy grad fjordvis, men der den store klyngen som for lus inkluderte lokaliteter på tvers av kystlandskapet fra Øygarden til Værlandet brytes opp i flere mindre klynger (Figur 4). Den viktigste forklaringen på oppdelingen av denne store klyngen ute i kystlandskapet er den høyere dødeligheten (kortere halveringstid) hos virus som gir kortere transport av smitte. Som resultat forekommer størsteparten av virussmitten på de nære lokalitetene.
Figur 4 . Smitte av virus (ILA og PD) mellom lokaliteter basert på strømmodell og biomasse på lokaliteter fra perioden 1/4-2017 til 30/9-2020. For videre forklaring se Figur 2.
3.3 - Variasjon i smittefaktorer i PO4
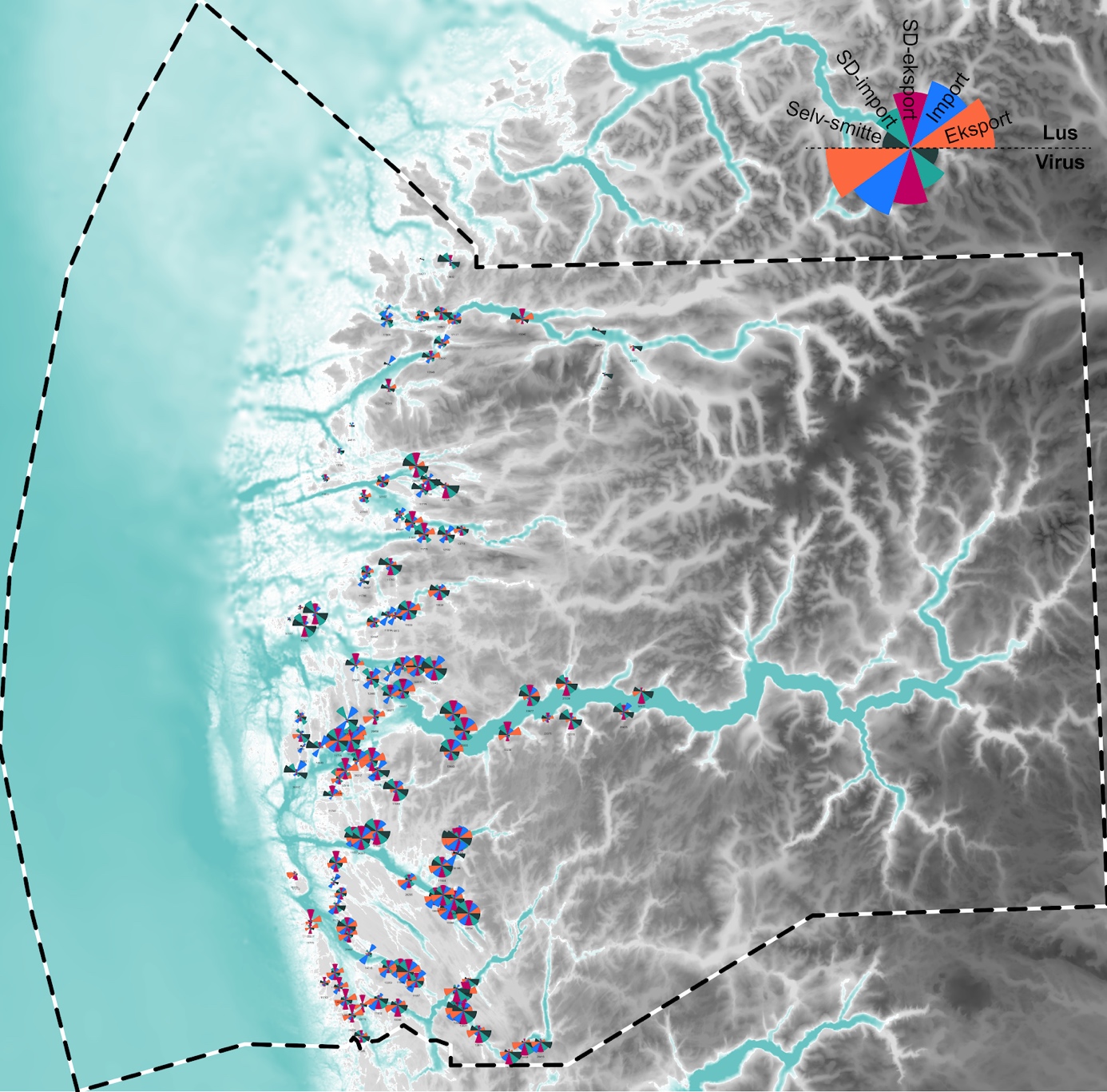
Konnektivitetsmatrisen lar seg analysere og presentere på flere måter enn nettverksplottene og klyngeanalyser presentert i foregående paragrafer. For eksempel kan man summere selv-smitten (diagonalen av matrisen), eller se på variasjon (her standardavvik) i import og eksport over tid. Ser man på disse variablene på tvers av PO4 vil man se en stor romlig variasjon, med høye rangeringer på tvers av alle nevnte faktorer for eksempel i ytre deler av Sognefjorden, Sognesjøen, Masfjorden og Austefjord; og generelt lave rangeringer på øyene utenfor Flora og i indre Nordfjord der f.eks selv-smitte ser ut til å være den viktigste faktoren (Figur 5). Det bør her noteres at da kyststrømmen som hovedregel transporterer lus nordover langs kysten, og analysen ikke inkluderer anlegg utenfor PO4, vil importen i sør undervurdert og likedan vil eksporten for anlegg i nord være underrepresentert.
Figur 5 . Rangering av smittefaktorer i PO4, der sektorer i sektordiagram representerer en rangering fra 1 til 256 (der 1 har størst sektor): (1) eksport til andre lokaliteter, (2) import fra andre lokaliteter, (3) standardavvik i eksport, (4) standardavvik i import, og (5) selv-smitte. Her okkuperer rangeringen av lusesmitte den øvre halvdelen av sektordiagrammet og virus den nedre.
3.4 - Smittepress i Nordfjord-Frøysjøen
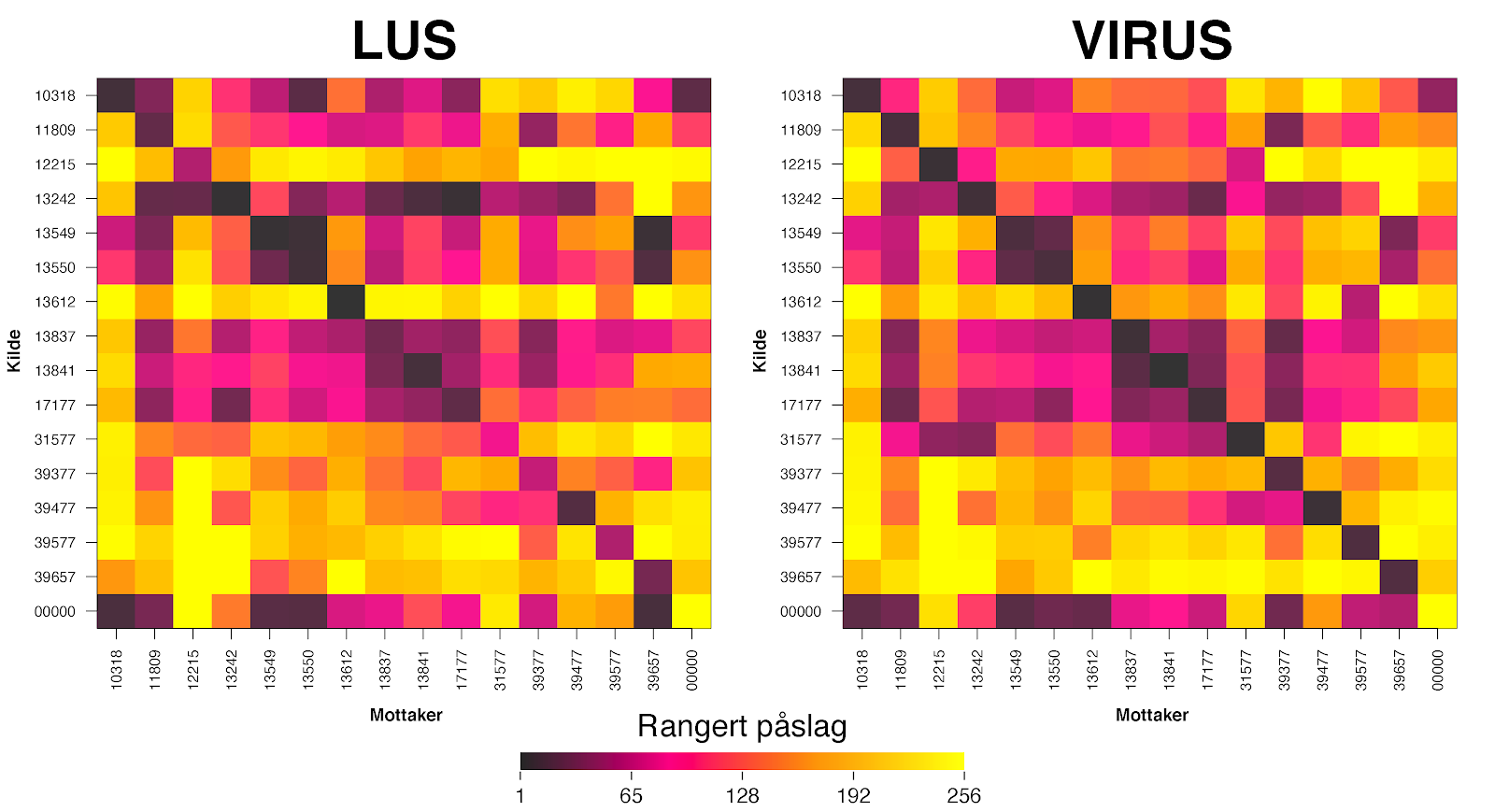
I klyngeanalysen kommer Nordfjord og Frøysjøen sammen ut som en egen klynge (K3, Figur 2). Dette mønsteret ser også ut til å være stabilt mellom sesongene (Figur 3). Lignende resultat så vi også for spredningen av virus (Figur 5). Videre, s er man på alle koblingene som kommer inn til Nordfjord-Frøysjøen fra andre klynger under ett, kommer ≈10% av lus og ≈1% av potensiell virussmitte fra lokaliteter utenfor klyngen. Det aller meste av denne importen fra andre klynger vil lande på lokalitetene i Frøysjøen (f.eks. 39657, 10318, 13550 og 13549 i Figur 6). Dette betyr at lokalitetene i Nordfjord er forholdsvis isolerte fra resten av produksjonsområdet, og at lokalitetene sammen kan oppnå god effekt av enhver koordinert innsats mot lus, og enda større effekt av lokale tiltak mot virus. Videre kan man se at diagonalene av konnektivitetsmatrisen (i.e., selv-smitte) bidrar sterkt i smittebildet, særlig da på virus-siden der alle de sterkeste koblingene består av selv-smitte. Her må det også legges til at en eventuell smitte fra lokaliteter i ytre Nordfjord og inn i PO5 ikke er modellert, der det antas at denne smitten til tider kan være moderat.
Figur 6 . Konnektivitet av lus og virus mellom lokaliteter i klyngen “Nordfjord-Frøysjøen” (K3 i Figur 2). Her er det rangerte påslaget (1 til 256, der 1 er størst påslag) fra en bestemt lokalitet representert med fargede kvadrater langs radene, til en bestemt lokalitet langs kolonnene, og kvadrater langs diagonalen er selv-smitte . Lokalitet “00000” representerer det totalet påslaget fra lokaliteter utenfor klyngen Nordfjord-Frøysjøen.
4 - Referanser
Albretsen J, Sperrevik AK, Sandvik AD, Asplin L (2011) NorKyst-800 Report No. 1 User Manual and technical descriptions. Fisken og Havet 2:1–48
Graham DA et al. (2013) Biophysical properties of the samonid alphaviruses: influence of temperature and pH on virus survival. Journal of Fish Diseases, 2007, 30, 553–543.
Huserbråten Mats, Bjørn Ådlandsvik, Øivind Bergh, Søren Grove, Ørjan Karlsen, Geir Lasse Taranger, Lars Qviller, Katherine Rose Dean, Britt Bang Jensen og Ingrid Askeland Johnsen (2020) Endret lokalitetsstruktur i produksjonsområde 3. Rapport fra Havforskningen 2020-12
Rosvall M, Bergstrom CT (2008) Maps of random walks on complex networks reveal community structure. Proc Natl Acad Sci U S A 105:1118–23
Skern R, Sissener NH, Sandvik AD, Meier S, Sævik PN, Skogen MD, Vågseth T, Dalvin S, Skern-Mauritzen M, Bui S (2020) Parasite development affect dispersal dynamics; infectivity, activity and energetic status in cohorts of salmon louse copepodids. Journal of Experimental Marine Biology and Ecology, 530-531:151429
Stien A, Bjørn PA, Heuch PA, Elston DA (2005) Population dynamics of salmon lice Lepeophtheirus salmonis on Atlantic salmon and sea trout. Mar Ecol Prog Ser 290:263–275
Ådlandsvik, B. (2020), LADiM documentation, https://ladim.readthedocs.io/