Rapportserie:
Rapport fra havforskningen 2022-50ISSN: 1893-4536Publisert: 26.01.2023Prosjektnr: 15445Oppdragsgiver(e): FiskeridirektoratetReferanse: Fiskeridirektoratet, 18/8650 Approved by:
Research Director(s):
Geir Lasse Taranger
Program leader(s):
Terje Svåsand
In this report we have collected knowledge on use of copper as antifoulant in Norwegian aquaculture, on how much comes from feed spill and faeces, and compared levels with threshold values from feed. We show copper levels shown in the transition zone and remote zone and compare with environmental quality standards. We show levels from sediment traps after rinsing. Increased levels in the water column due to leakage from net pens are modelled and possible extra pulses due to rinsing are described.
The largest contribution of copper from aquaculture is copper(I)oxide (Cu2O) used as antifoulant on net pens. At sufficient high concentrations copper can damage sensitive species and give negative impact to the surrounding environment. In 2020, the last year we have knowledge on use, 1539 tonnes copper was registered to be used as antifoulants for aquaculture in Norway. This is the first year a decrease in such use has been registered.
Leakage experiments performed by the industry as part of documentation for approval have shown that approx. 28 % of copper content can be released to the environment if the net pens are not rinsed. If the net pens are rinsed, 20 % of original copper content may be lost during the first rinsing. Results from sediment traps show that a substantial fraction of the copper impregnation lost during rinsing can sediment to the seabed in the vicinity, but we lack data on how large fraction of copper is lost as particles and how much blead out as copper ions to the water column.
Model simulations where we assume that 28 % of copper has leaked to the water column, show that passive leaked copper can give a substantial contribution to the total copper concentration in a fjord system, of approx. 0.2-0.4 µg/l. In narrow fjords with less water exchange the contribution may be as high as 1 µg/l. Rinsing or high pressure rinsing of copper impregnated net pens may give higher pulses with higher concentrations. Environmental quality standards for copper in coastal water is 2,6 µg/l. We have measured concentration in fjords in Vestland from < 0.5 to 1.9 µg/l in 2021.
Monitoring of copper levels in sediment from the remote zone (> 1 km from nearest fish farm) in several fjords in Vestland showed a significant increase in copper levels in 4 of 13 stations in the period from 2018 to 2022.
Histologic analyses of blue mussels that grows on copper impregnated net pens shows high levels of copper in the soft tissue and that copper exposure that high copper concentration is related to tissue damage in digestive gland (tubular atrophy) and changed structure in gills. Such effects of copper exposure in blue mussels are now being tested in dose-response experiments.
Det large use of copper in Norwegian aquaculture combined with knowledge on passive leakage and extra discharges as pulses if the net pens are rinsed makes the Institute of Marine Research to recommend that measures are taken to reduce this impact.
Better information on which antifoulants used at different locations and praxis related to rinsing at different fish farms are needed to increase the strength of knowledge on impact of such discharge in the different production regions.
To reach the objective that discharge from aquaculture shall not impact the surrounding environment negatively, we recommend that the aquaculture industry increase the effort to develop solutions for antifouling that do not lead to discharge of toxic compound (copper or replacements) that can bioaccumulate in organisms.
Forord
Denne delrapporten er laget for å svare opp på bestilling fra Fiskeridirektoratet til Havforskningsinstituttet om kunnskapsstøtte på miljøeffekter av utslipp av kobber fra marine akvakulturanlegg og om miljøeffekter fra kobber på blåskjell (Fiskeridirektoratet, 18/8650). På grunn av at vi mangler data på hvor stor andel som blør ut som kobberioner i forhold til hvor stor andel som faller ned i anleggssonen som partikler etter spyling og har pågående dose-respons forsøk med blåskjell eksponert for kobber, vil vi følge opp resten av bestillingen etter hvert som disse resultatene er klare.
Sammendrag
Vi har sammenstilt kunnskap om bruk av kobber som grohemmende middel på nøter i norsk oppdrettsnæring med hvor mye som kommer fra fôrspill og fekalier, med grenseverdier fra fôr, kystvann og sediment. Vi viser også kobbernivå målt i sediment fra overgangssonen og i fjernsonen og fra sedimentfeller etter periode med spyling. Økte nivå på grunn av utblødning fra nøter er modellert og mulige ekstrapulser på grunn spyling er målt og beskrevet.
Det største bidraget av kobber fra fiskeoppdrett er kobber(I)oksid (Cu2O) brukt som grohemmende middel på nøter i oppdrettsnæringen. I høye nok konsentrasjoner kan kobber både føre til skade på følsomme arter og gi skadelige langtidsvirkninger i vannmiljøet. I 2020, som er siste året der vi har opplysninger om forbruk, ble det registrert 1539 tonn kobber til bruk som grohemmende midler i akvakultur. Dette er første året at det er registrert en nedgang i slik bruk.
Utlekkingsforsøk gjort av næringen som del av dokumentasjon for godkjenning har vist at ca 28 % av kobberinnholdet kan forsvinne etter endt levetid i sjøen dersom noten ikke blir spylt. Dersom noten blir spylt kan 20 % av opprinnelig kobbermengde forsvinne ved førstegangs spyling. Resultat fra sedimentfeller viser at en vesentlig andel av kobberimpregnering tapt ved spyling kan synke ned på sjøbunnen i anleggssonen, men vi mangler data på hvor stor andel kobber som faller ut som partikler og hvor mye som blør ut som kobberioner til vannsøylen.
Modellsimuleringer der vi har antatt at 28 % av kobberet lekker ut til vannsøylen, viser at passivt utlekket kobber kan gi et vesentlig bidrag til den totale kobberkonsentrasjonen i et fjordsystem, i størrelsesorden 0,2-0,4 µg/l. I trange fjorder med dårlig vannutveksling kan bidraget periodevis være opp til 1 µg/l. Spyling eller høytrykksspyling av kobberimpregnerte nøter vil kunne gi pulser med høyere konsentrasjoner. Miljøkvalitetsmål for kobber i kystvann er 2,6 µg/l. Vi har målt konsentrasjoner i fjorder i Vestland fra < 0,5 til 1,9 µg/l i 2021.
Overvåking av kobbernivå i sediment fra fjernsonen (> 1 km fra nærmeste oppdrettslokalitet) i flere fjorder i Vestland viser en signifikant økning i nivå på 4 av 13 stasjoner i perioden 2018-2022.
Histologiske analyser av blåskjell som vokser på kobberimpregnerte nøter viser høye nivå av kobber i mykvevet og sammenheng mellom kobbereksponering og vevsskader i fordøyelseskjertelen (atrofi eller innskrumping av tubuli) og endring av struktur i gjellene. Slike markører for kobbereksponering i blåskjell er under uttesting i dose-respons forsøk.
Det høye forbruket av kobber i norsk oppdrettsnæring kombinert med kunnskap om passiv utlekking og ekstra utslipp som pulser dersom nøtene blir spylt gjør at Havforskningsinstitutt anbefaler at det blir iverksatt tiltak for å redusere denne påvirkningen.
Mer informasjon om hvilke antibegroingsmidler som er i bruk på den enkelte lokaliteten og praksis i forhold til spyling på de enkelte anlegg vil være avgjørende for å øke kunnskapsstyrken om utslippsmengde i de enkelte produksjonsområdene.
For å nå målet om at utslipp fra oppdrett ikke skal påvirke miljøet rundt et oppdrettsanlegg negativt anbefaler vi at det blir satt i gang arbeid med å få til løsninger som ikke fører til spredning av giftige forbindelser som kobber eller erstatningsstoff som kan bli akkumulert i sediment eller i organismer.
1 - Innledning
Kobber er et naturlig forekommende tungmetall både i jordskorpen, i marine sediment og i sjøvann. Stoffet er en viktig faktor for enkelte enzymreaksjoner i levende organismer, men er giftig dersom konsentrasjonen av kobberforbindelser blir for høy. I tillegg til å være et essensielt metall tilsatt fôret til oppdrettsfisk, brukes kobber som antibegroingsmiddel på nøter. I det marine miljø kan kobber eksistere både som en-verdig Cu(I) og som to-verdig Cu(II) kobber. Kjemiske former og konsentrasjoner av uorganisk kobber i sjøvann blir påvirket av redoks-statusen til sjøvann og sediment.
Oppdrettsnøter som står i sjøen over lengre tid vil bli dekket av forskjellige organismer som alger og virvelløse dyr, der særlig sjøpung og blåskjell kan skape problemer. Begrodde nøter vil hindre vanngjennomstrømning og dette vil kunne redusere oksygentilførselen som fisken er avhengig av. Det vil også hindre at fekalier og fôrrester blir vasket ut av nøtene. I tillegg kan det føre til at rensefisk beiter på det som gror på nøtene istedenfor å beite på lakselus. For å unngå begroing impregneres nøtene med antibegroingsmiddel, som for eksempel kobber, i konsentrasjoner som er giftig for organismene. Over tid lekker det kobber ut i vannet som spres med vannstrømmen og en del faller av og synker ned under eller i nærsonen til anlegget avhengig av partikkelstørrelse, sedimentasjonshastighet og strømmønster. Dette betyr at sjøbunnen under og rundt fiskeoppdrettsanlegg kan inneholde høye konsentrasjoner da avsatt kobber kan akkumulere over tid.
Norske fiskeoppdrettsanlegg er pålagt å overvåke bunnen under anleggene og områdene rundt ved å følge Norsk Standard NS9410:2016. Overvåkingen er først og fremst for å dokumentere organisk påvirkning av bunnen, og består av en B-undersøkelse i anleggssonen og en C-undersøkelse i området hvor anlegget er lokalisert. Undersøkelsene utføres med jevne mellomrom og jo mer sedimentet er påvirket jo oftere skal en utføre målinger. Det blir ikke målt kobber i B-undersøkelsene, men i C-undersøkelsen er det pålagt å måle kobberkonsentrasjonen i sedimentet i anleggssonen (25-30 m fra anleggsrammen) og i overgangssonen (30-500 m) dersom det benyttes eller har vært benyttet kobber som antibegroingsmiddel, og disse verdiene er de vesentligste tall som inngår i kobberstatistikken. Resultatet av målingene i anleggssonen bestemmer om det er behov for ytterligere undersøkelser for å kartlegge kobberpåvirkningen.
Det er ikke ønskelig at organismer som lever i nærheten av et oppdrettsanlegg skal eksponeres for kobberkonsentrasjoner som over tid kan redusere normale fysiologiske prosesser som adferd, vekst, reproduksjonsevne eller helsetilstand. Bunnfaunaen utgjør en viktig økosystemtjeneste, og det er viktig å opprettholde denne for å kunne drive med langsiktig drift på lokaliteter. Dersom det blir målt for høye kobbernivå i sedimentene under anlegget vil dette medføre reduksjon i antall arter som kan leve der og lokaliteten vil bli stengt for videre oppdrettsaktivitet.
Når det skal vurderes risiko knyttet til påvirkning fra ulike utslippskilder er det viktig å kartlegge om påvirkningen er kort- eller langvarig, om den er reversibel eller irreversibel og om den kommer fra nedbrytbare stoffer eller stoffer som hoper seg opp i miljøet. Utslippene av kobber fra nøtene er i en slik størrelsesorden at man må forvente at det kan være påvirkning på marine organismer. Påvirkningen av kobber fra oppdrett er for det meste lokal og pågår i et begrenset geografisk område, men siden lokalitetene brukes over mange år vil påvirkningen kunne være langvarig (Grefsrud mfl. 2022 a og b).
Denne delrapporten er laget for å svare opp på bestilling fra Fiskeridirektoratet til Havforskningsinstituttet om kunnskapsstøtte på miljøeffekter av utslipp av kobber fra marine akvakulturanlegg og om miljøeffekter fra kobber på blåskjell (Fiskeridirektoratet, 18/8650).
2 - Potensial for spredning av kobber fra akvakultur-lokaliteter
Utslippsmengde fra et oppdrettsanlegg kommer fra fôrspill, fekalier og/eller kobber som faller av som store eller små partikler eller som lekker ut som kobberioner fra not-impregneringen.
2.1 - Utslipp fra fôr og fekalier
Det finnes en rekke fremmedstoffer som kommer ut i miljøet fra fiskeoppdrettsanlegg. Fiskefôr kan inneholde forskjellige miljøgifter som kommer fra fôringrediensene og disse kan tilføres miljøet både gjennom spillfôr og gjennom fiskens avføring (fekalier). Stoffene som blir tilført miljøet fra fiskefôr og fekalier kommer fra de ingredienser som fôret består av. Omtrent 70 % av fôringrediensene er i dag plantebasert og 30 % er basert på marine råstoff (Ytrestøyl mfl. 2015). Råstoffene brukt til fôrproduksjon inneholder blant annet halogenerte organiske forbindelser som PCB, dioksiner, furaner, klorerte pesticider, bromerte flammehemmere og tungmetallforbindelser som kvikksølv (Hg), arsen (As) og kadmium (Cd), kobber (Cu) og sink (Zn) (Tabell 1). Andre stoffer tilsettes fôret i små mengder og er nødvendige for at fisken kan ha god tilvekst. Dette inkluderer også kobber og sink, som derfor også kommer inn under kategorien mineraler når de blir tilsatt fôret.
Opptaket av kobber i fisken gjennom fôret er avhengig av konsentrasjonen i fiskefôret. Ved konsentrasjoner i fôret nær behovsgrensen (det oppdrettsfisken trenger for å opprettholde de fysiologiske funksjonene) er opptaket høyt, mens når konsentrasjonen er høyere enn behovsgrensen blir opptaket av kobber regulert eller redusert i tarmen til fisken. Som konsekvens gir dette et høyere utslipp av kobber gjennom fekalier. For kobber er retensjonen (tilbakeholdelsesgraden) i fisk 25% ved konsentrasjon på 5 mg/kg fôr, mens når konsentrasjon i fôret er 35 mg/kg er retensjonen kun 5,7 %. Ved en fôrkonsentrasjon på 10,9 mg/kg, som er gjennomsnittskonsentrasjonen i fiskefôr analysert i 2021, estimeres det at retensjon av kobber er 21%. Hele 79% av kobberet slippes dermed ut gjennom fekalier eller urin. Fôrforbruk per anlegg per år kan variere fra 700 tonn til 14000 tonn. Dersom et gjennomsnittlig oppdrettsanlegg bruker 2000 tonn fôr per år vil ca. 16 kg kobber gå gjennom fisken i anlegget (Tabell 1). Tabellen viser også beregnet utslipp av andre utvalgte miljøgifter fra fôret.
Tabell 1. Oversikt over utslipp av utvalgte stoffer gitt fôrforbruk per 1000 tonn og fôrspill på 0,5 eller 8 % basert på Sele et al. (2022) og retensjonsdata fra Havforskningsinstituttet.
Miljøgift
Øvre grenseverdi i fôr satt av EU kommisjonen
(EU 2002)
Gjennomsnitt fullfôr i 2021
Andel som går gjennom laksen
Utslipp gitt fôrforbruk på 1000 tonn gitt 0,5 % fôrspill
Utslipp gitt fôrforbruk på 1000 tonn gitt 8 % fôrspill
Hg
0,2 mg/kg
0,022 mg/kg
0,59
13 g
14 g
Cu
25 mg/kg
10,9 mg/kg
0,79
8,6 Kg
8,8 Kg
Cd
1,0 mg/kg
0,12 mg/kg
0,95
114 g
114 g
Zn
200 mg/kg
168 mg/kg
0,68
115 Kg
119 Kg
∑DDT
50 µg/kg
4,7 µg/kg
0,57
2,7 g
2,8 g
∑Klordan
20 µg/kg
1,1 µg/kg
0,67
0,7 g
0,8 g
HCB
10 µg/kg
1,0 µg/kg
0,57
0,6 g
0,6 g
PCB6
30 µg/kg
3,4 µg/kg
0,46
1,6 g
1,7 g
PBDE7
Ikke fastsatt
0,35 µg/kg
0,58
0,20 g
0,21 g
2.2 - Forbruk av kobber brukt som grohemmende middel i Norsk akvakultur
Mengdene av kobber fra fôrspill og fekalier er imidlertid langt mindre enn det som kommer fra kobber som impregneringsmiddel, som er det stoffet det er knyttet størst bekymring til.
Dikobberoksid (Cu2O) er den formen av kobber som det blir brukt mest av som grohemmende middel. I tillegg til dikobberoksid er disse godkjent som groehemmende middel (stoffgruppe 21): kobbertiocyanat, kobberpyrition, kobberflak (belagt med alifatisk syre).
Når spor av kobber fra Cu2O impregnert not går i løsning i sjøvannet blir det frigitt som Cu2+ ioner og er giftig for organismer som vokser på noten.
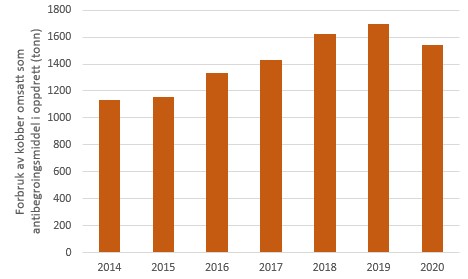
Det har vært en jevn økning i forbruk av kobber som grohemmende midler i norsk akvakultur siden 2003 (Skarbøvik et al., 2017). I 2014 ble det omsatt 1130 tonn kobber til bruk som grohemmende midler, mens tilsvarende forbruk i 2019 var 1698 tonn. Dette tilsvarer en økning på 50 % over denne perioden, og en gjennomsnittlig årlig økning på 9 %. I 2020 ble det for første gang registrert en nedgang i kobberforbruket til 1539 tonn (Figur 1) (Produktregisteret). Norge eksporterte 1391 tonn kobber som grohemmende midler i 2020, men dette er ikke inkludert i statistikken i Figur 1.
Figur 1. Omsetning av kobber til bruk som antibegroingsmiddel i norsk akvakultur i perioden 2014-2020. Kilde: Produktregisteret.
3 - Bakgrunnsnivå av kobber i kystvann og i marine sediment
3.1 - Kobber i kystvann
Løst kobber i sjøvann kan eksistere i flere kjemiske og fysiske former som partikler, ioner, og organiske og uorganiske komplekser. De kjemiske egenskapene til kobber i sjøvann er viktig for bioakkumulering og giftighet for marine organismer.
En litteraturgjennomgang av tilgjengelige data har satt bakgrunnsnivå i sjøvann til 0,1 µg/l (Sadiq 1992). For noen havområder er det rapportert om økende konsentrasjoner av kobber med økende dyp. Dette blir tilskrevet opptak av kobber i planteplankton eller biota i overflaten og regenerering av kobber i dypere vannmasser. For oksygenrikt og mindre oksygenrikt sjøvann er kobberklorid (CuCl) den mest vanlige formen, mens kobbersulfid (CuS) er mest vanlig i svært oksygenfattig sjøvann (Sadiq 1992).
Gjennomsnittsverdier på 0,140 µg/l kobber i den nordlige Nordsjøen og på 0,070 µg/l i Atlantisk vann er rapportert av Balls (1985). Miljødirektoratet har definert nivå mindre enn 0,300 µg/l som bakgrunnsnivå for kystvann i Norge (Tabell 2).
Russiske forskere har rapportert om gjennomsnittsnivå av kobber i Barentshavet til 1,267±0,103 µg/l og gjennomsnittsnivå i Arktisk vann til 0,985±0,086 µg/l (Novikov og Draganov, 2017).
Grenser for tilstandsklasser I–V for kobber i kystvann og sediment er vist i Tabell 2. PNEC (Predicted no effect concentration) for marine organismer og miljøkvalitetsstandard for kystvann er satt til 2,6 µg/l . Bakgrunnsverdier for kobber i kystvann og sediment er satt på grunnlag av verdier rapportert av OSPAR (2006). Nedre grenseverdier for Tilstandsklasse III Moderat og tilstandsklasse IV Dårlig er satt lik fordi forskjellen i konsentrasjoner som gir kronisk eller akutte effekter er relativt liten (Miljødirektoratet, 2014).
Tabell 2. Tilstandsklasser av kobber i kystvann og sediment (Miljødirektoratet, 2016).
I Bakgrunn
II God
III Moderat
IV Dårlig
V Svært dårlig
Bakgrunnsnivå
Ingen toksiske effekter
Kroniske effekter ved langtidseksponering
Akutte toksiske effekter
Omfattende akutt-toksiske effekter
Kobber i kystvann (µg/l)
< 0,3
0,3-2,6
<2,6
2,6-5,2
>5,2
Kobber i sediment (mg/kg) (tørr vekt)
< 20
20-84
<84
84-147
>147
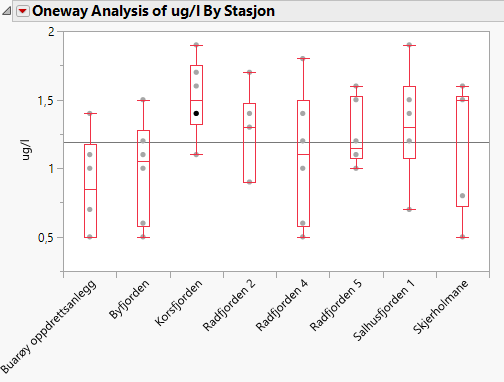
Målinger av kobber fra 3 forskjellige dyp fra 8 stasjoner i Vestland i 2021 gav gjennomsnittsnivå på 1,19±0,40 µg/l. Høyeste nivå ble målt i Korsfjorden til 1,52±0,28 µg/l kobber. Det var ikke forskjeller mellom dyp. Det var et spenn i målingene fra 0,5 µg/l (LOQ) til 1,9 µg/l (Figur 2). Overflatevann ved Os, Vestland er målt til 1,85±0,63 µg/ og ved Nordnes 0,24±0,01 µg/l. Vi finner relativ stor variasjon i målte nivåer og de høyeste målte verdiene er ikke så langt unna miljøkvalitetsstandarden for kystvann på 2,6 µg/l.
Figur 2. Boksplott av kobberinnhold på 3 forskjellige dyp fra 8 stasjoner i Vestland prøvetatt 20-21 januar 2021. Prøvene fra Buarøy oppdrettsanlegg og fra Skjerholmane ble prøvetatt 50 m fra oppdrettsanlegget. Dyp: Bunn + 5m, midt i vannsøylen og overflate – 5m.
3.2 - Kobber i marine sediment
Kobberkonsentrasjon i upåvirkete marine sediment er vanligvis under 10 mg/kg tørt sediment (Sadiq 1992). Kobber tilført marine sediment vil bli værende der i en eller annen kjemisk form. Det er derfor forventet at kjemiske nivå i marine sediment gradvis vil øke ved kontinuerlig tilførsel.
Den mest stabile formen for kobber er kobberferritt CuFe2O4 (s) i miljøer der redoks-statusen (pe + pH) er > 6 for oksygenrikt og for mindre oksygenrikt sjøvann. Den mest stabile formen i oksygenfattig sjøvann der redoks-statusen er < 6, er kobbersulfid Cu2S (s). Utfelling eller oppløsning kan påvirke konsentrasjonene i porevann og sjøvann (Sadiq 1992). Slik kan anaerobe og sulfidrike sediment i nærsonen binde kobber og redusere biologisk tilgjengelighet (Brooks og Mahnken 2003).
4 - Akkumulering av kobber i sediment over tid
4.1 - Økte konsentrasjoner av kobber i sediment i anleggsonen og innerste deler av overgangssonen
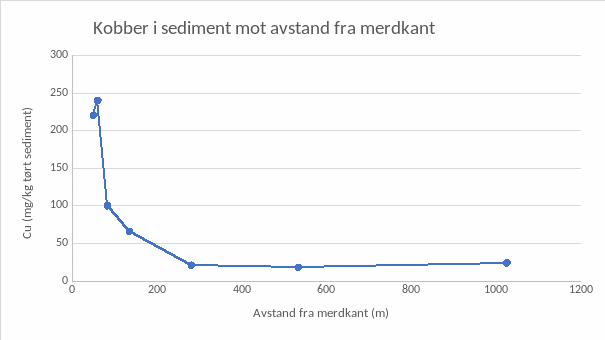
Målinger av kobbernivå i sediment i økende avstand ut fra merdkant viser en gradient med høyeste konsentrasjoner i anleggssonen (25 m fra merd) og avtagende nivå utover i overgangssonen (25-500 m) (Figur 3). Resultatene viser at kobberpartikler fra impregnert not synker ned under og rundt anlegget og at en for dette anlegget må ut i en avstand på ca 100 m fra anlegget for å komme under miljøkvalitetsstandard for kobber i sediment som er 84 mg/kg tørt sediment.
Figur 3. Transekt av kobber i sediment ut fra merdkant viser en gradient med høyeste konsentrasjoner i anleggssonen (25 m fra merd) og avtagende nivå utover i overgangssonen (25-500 m).
4.2 - Sedimentering av kobber rundt et anlegg etter periode med spyling
Analyser av hvor mye som sedimenterer til sedimentfeller rundt et anlegg som har blitt spylt er vist i Tabell 3. Sedimentfellene sto ute i 12 dager og på grunn av mye groe ble oppdrettsanlegget ble spylt to ganger i denne perioden. Dette kan forklare de relativt høye verdiene av sedimentert kobber over denne perioden. Resultatene viser også at spyling gir et høyt utslipp av partikler som sedimenterer. Nivå av kobber i sedimentprøve 50 m fra anlegget (158 m dyp) tatt samtidig med innhenting av sedimentfellene var 165±68 mg/kg tørt sediment. Anlegget bestod av 4 sirkelmerder.
Tabell 3. Sedimentering av kobber til sedimentfeller rundt et anlegg etter periode med spyling sammenlignet med nivå ved en referanse lokalitet.
Avstand til merd (m)
Dyp
Mengde tørt sediment etter 12 dager og to spylinger (g tørt sediment)
Kobberkonsentrasjon etter 12 dager og to spylinger (mg/kg tørt sediment)
65
40
0,48
160
66
40
0,29
200
74
40
0,19
370
74
140
0,20
140
Referanse
330
0,05±0,02
28±10
4.3 - Resultat fra C-undersøkelsene
For å vurdere påvirkning på sediment rundt et oppdrettsanlegg har vi brukt verdiene som er rapportert i C-undersøkelsene for anleggssonen og overgangssonen i forhold til tilstandsklassene som er satt av Miljødirektoratet. Miljødirektoratet har definert fem tilstandsklasser for kobber i marine sediment. Øvre grense for tilstandsklasse I er definert som bakgrunnsnivå, mens grensene for de andre tilstandsklassene er gjort etter vurderinger av giftighet til stoffet. Tilstandsklasse II-God: skal ikke føre til toksiske effekter, tilstandsklasse III-Moderat: kan gi kroniske effekter ved langtidseksponering, tilstandsklasse IV-Dårlig: kan gi akutte toksiske effekter ved korttidseksponering og tilstandsklasse V-Svært dårlig kan gi omfattende akutt-toksiske effekter. Når det gjelder giftighetsstudier for kobber er det ikke klare forskjeller på konsentrasjoner som gir kroniske effekter ved langtidseksponering og akutt giftighet ved korttidseksponering (Miljødirektoratet 2016). Den øvre grense for tilstandsklasse II- God grenser derfor både til tilstandsklasse III-Moderat og Tilstandsklasse IV-Dårlig (Tabell 1). Vi har derfor valgt å klassifisere tilstandsklasse I og II som God og tilstandsklasse III, IV og V som Dårlig i vurderingen vi har gjort av hvor stor prosentandel som har dårlig miljøtilstand for kobber i anleggssonen eller overgangssonen fra C-undersøkelsene fra de enkelte produksjonsområdene (Tabell 4). Produksjonsområde 1 og 2 (Svenskegrensen til Jæren og Ryfylke) er slått sammen for å få bedre tallgrunnlag.
Tabell 4. % lokaliteter som har dårlig miljøtilstand i anleggssonen eller i overgangssonen basert på C-undersøkelser i perioden 2017-2020, totalt 541 C-undersøkelser.
Produksjonsområde
Antall C-undersøkelser
% lokaliteter som har dårlig miljøtilstand av kobber i anleggssonen
% lokaliteter med dårlig miljøtilstand av kobber i overgangssonen
1. Svenskegrensen til Jæren og 2. Ryfylke
28
22
7
3. Karmøy til Sotra
58
53
17
4. Nordhordland til Stadt
63
41
4
5. Stadt til Hustadvika
30
15
0
6. Nordmøre og Sør-Trøndelag
84
11
6
7. Nord-Trøndelag med Bindal
49
21
4
8. Helgeland til Bodø
62
15
3
9. Vestfjorden og Vesterålen
62
2
0
10. Andøya til Senja
30
0
0
11. Kvaløy til Loppa
31
4
0
12. Vest-Finnmark
37
9
8
13. Øst-Finnmark
7
0
0
4.4 - Nivå i sediment i fjernsonen i fjorder i Vestland
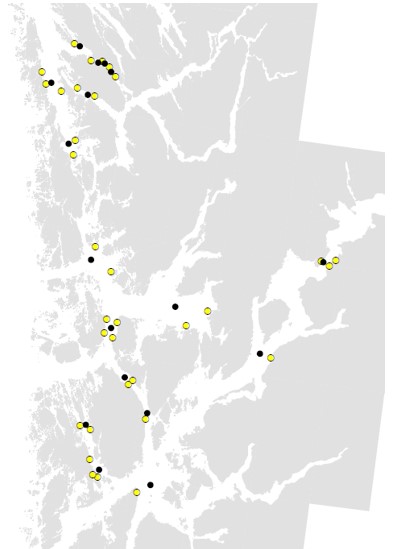
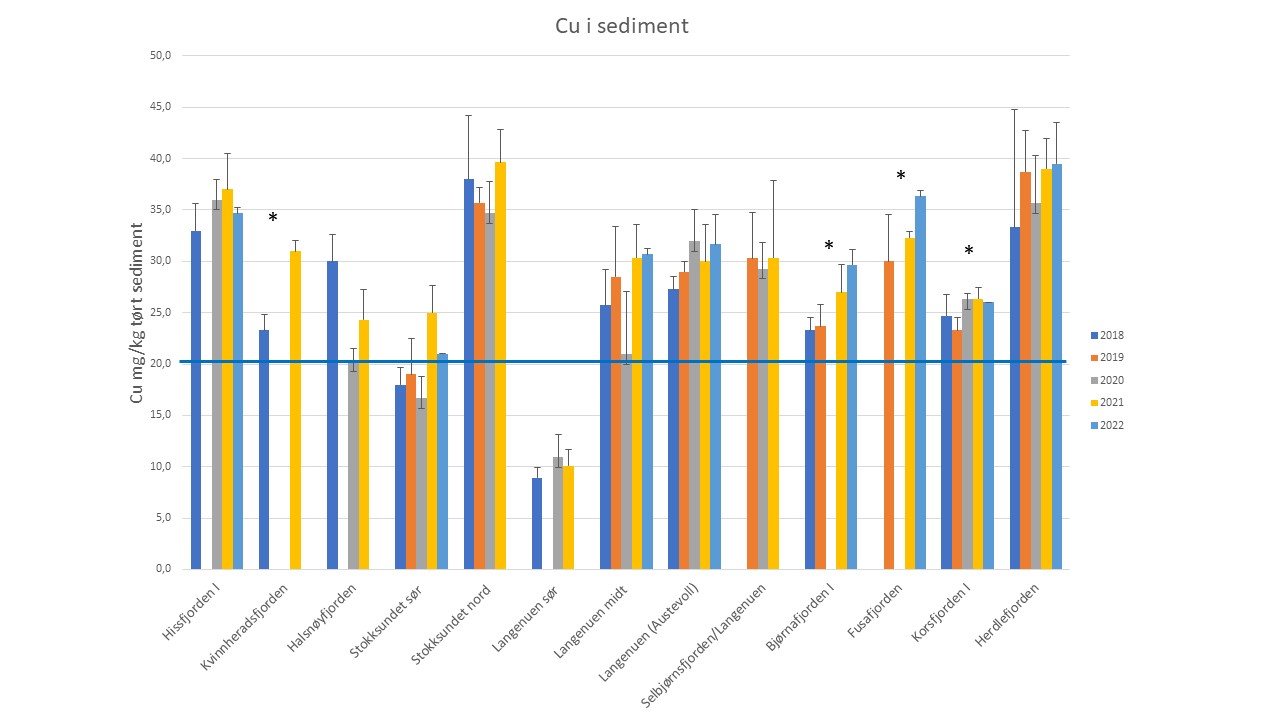
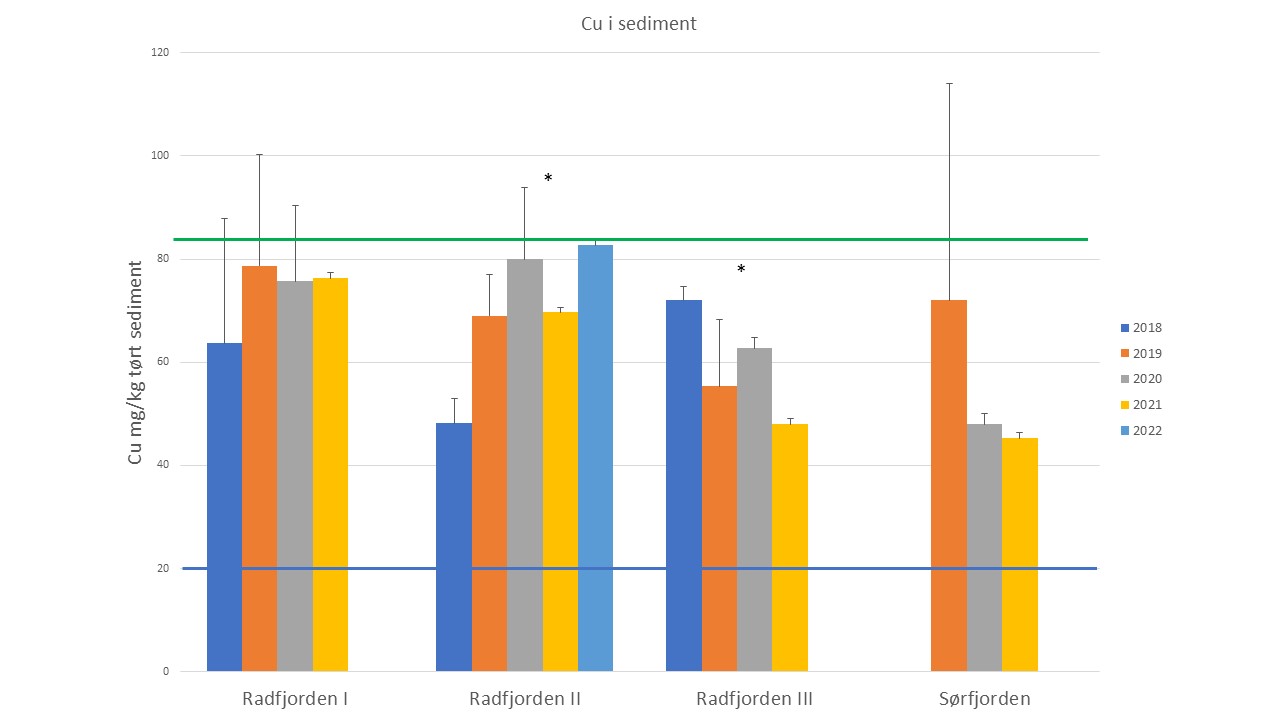
For å få bedre kunnskap om kobbernivå i fjernsonen endrer seg over tid startet Havforskningsinstituttet overvåking av miljøgifter i overflatesediment fra fjernsonen i flere fjorder i Vestland i 2018 (Figur 4, 5 og 6). Fem år (2018-2022) er en kort periode for å vise tidstrender, men foreløpige resultater for kobber i sediment viste at nivåene økte signifikant på 4 av 13 stasjoner i denne perioden (Kvinnheradsfjorden, Bjørnafjorden, Fusafjorden og Korsfjorden) (Figur 4). For de andre stasjonene var det stabile nivå eller svak økning og på stasjonen i Halsnøyfjorden var det en svak nedgang. De høyeste nivåene ble funnet Sørfjorden og i Radfjorden (Figur 6). På grunn av tidligere rapporter av høye nivå av kobber i sediment ved oppdrettsanlegg i Sørfjorden og i Radfjorden har Statsforvalteren i Vestland pålagt oppdrettsselskapene i disse to fjordsystemene å bruke kobberfrie antibegroingsmiddel. Stasjonen Radfjorden II viste en økning, mens stasjon Radfjord III viste en nedgang i denne perioden. Stasjonen i Sørfjorden viste også en nedgang (ikke signifikant) (Figur 6).
Figur 4. Overvåkingsstasjoner i sediment (svart sirkel) og oppdrettslokaliteter (gul sirkel). Overvåkingsstasjoner ligger i fjernsonen til oppdrettslokalitetene > 1000 m avstand.
Figur 5. Nivå av kobber i sediment i utvalgte fjorder i Vestland i perioden 2018-2021. Blå linje ved 20 mg/kg markerer grense mellom tilstandsklasse I (Bakgrunn) og Tilstandsklasse II (god). (*) Indikerer signifikant endring, p< 0,05.
Figur 6. Nivå av kobber i sediment fra tre stasjoner i Radfjorden og i Sørfjorden ved Osterøy i perioden 2018-2022. Blå linje ved 20 mg/kg markerer grense for tilstandsklasse 1 (Bakgrunn) og Tilstandsklasse II (god). Grønn linje ved 84 mg/kg markerer grense mellom Tilstandsklasse II (God) og Tilstandsklasse III Moderat og IV (Dårlig). (*) indikerer signifikant endring, p< 0,05.
5 - Giftighet av kobber
Kobber kan bli tatt opp (bioakkumulert) av marine organismer, men kobber blir ikke biomagnifisert til høyere nivå i næringskjeden. Bioakkumuleringsstudier viser at opptak av kobber til marine organismer hovedsakelig skjer som kobberion (Cu2+). Cu2+ er den giftigste formen etterfulgt av kobber-hydroksid kompleks (Cu-OH). Når kobber fra Cu2O og kobber thiocyanate lekker ut til oksygenholdig sjøvann vil den dominerende kobberformen være kobberionet Cu2+. Kobberion reduserer larvene sin evne til å feste seg på overflata av impregnert materiale ved at mikrolaget av vann og kobberion hemmer enzymaktivitet og bidrar til utfelling av enzym i cellene (EU, 2021).
5.1 - Ulike arters sårbarhet for kobber og kobberforbindelser
I høye nok konsentrasjoner kan kobber føre til skade på en rekke arter og gi redusert artsmangfold. Giftighetsstudier har vist at tidlige livsstadier av marine virvelløse dyr er mest følsomme for kobbereksponering, mens voksne stadier er mindre følsomme og til dels ganske robuste. Artene sin toleranse kan utrykkes som den konsentrasjonen av et stoff som ikke gir noen målbar effekt, såkalt PNEC ( Predicted no effect concentration) .
Eksponeringer for kobber i 2 døgn gav 50 % feilutvikling av embryo av stillehavsøsters (Crassostrea gigas) ved 5,3±0,5 μg/l og tilsvarende ved konsentrasjoner på 49±20 μg/l for larvestadie av krabben Cancer magister (Martin mfl. 1981). Kronisk eksponering av kobber på larvestadiet av den antarktiske pigghuden Sterechinus neumayeri har vist at halvparten av individene blir påvirket (EC50 verdi) ved 1,4 μg/l (King og Riddle 2001). Terskelverdier for konsentrasjoner som ikke gir effekt i en studie som inkluderte 24 arter (species sensitivity distribution) er blitt rapportert til 5,7 ug løst Cu/l (Foekema mfl. 2015). En annen gjennomgang av 65 marine arter har gitt tilsvarende terskelverdi på 5,6 ug løst Cu/l (Hall mfl. 1999).
PNEC for marine organismer er satt til 2,6 µg/l (Tabell 2). Denne verdien er basert på giftighetsstudier som gir HC5 verdi på 5,2 µg/l, med en sikkerhetsfaktor på 2 fordi en manglet mesokosmos studier eller feltdata (European Copper Institute, 2005).
En litteraturstudie om effekter av kobber på hoppekreps viste et stort spenn i konsentrasjoner som kan gi 50 % dødelighet (LC50). Nauplii stadiet var mest følsomt og pelagiske hoppekreps mer følsomme enn bunnlevende (Heuschele mfl. 2022).
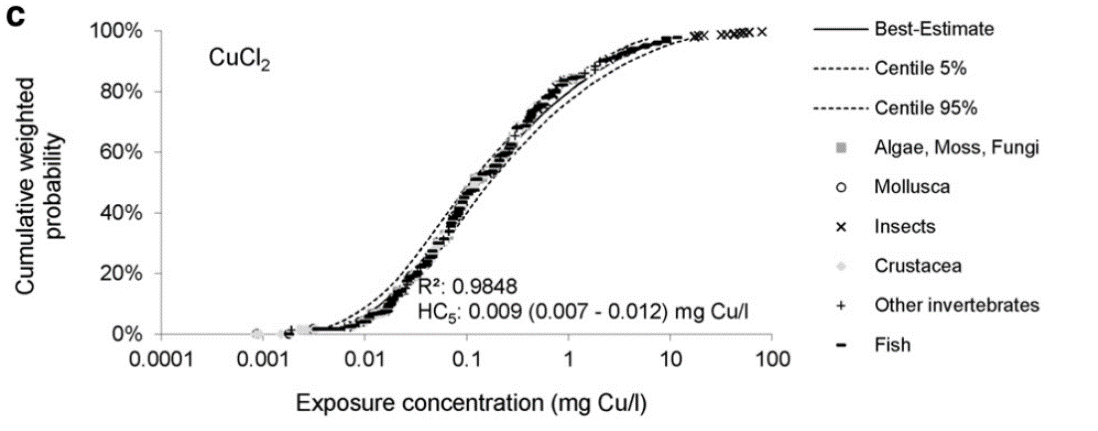
En studie som har sammenlignet giftighet av nanopartikler av CuO med CuCl2 salt i SSD kurver (artsfølsomhetsfordeling) viste at løst kobber var mer giftig enn nanopartikler (Adam mfl. 2015). HC5 for nanopartikler ble oppgitt til 150 ug/l kobber med konfidensintervall fra 50-470, mens HC5 for løst kobber ble oppgitt til 9 ug/l kobber med konfidensintervall på 7-12 µg Cu/l (Figur 7) (Adam mfl. 2015). HC5 står for det nivået der 95 % av artene skal være beskyttet. Krepsdyr var mest følsomme for kobbereksponering. Det antas at giftigheten både for kobber-nanopartikler og kobbersalt skyldes løste kobber-ion (Adam mfl. 2015).
Figur 7. SSD kurve for CuCl2 eksponering. Akutte toksisitetsdata (LC50/EC50) er tatt fra Ecotox database, 594 datapunkt (US EPA, Juli 2014). Figuren tatt fra Adam mfl. (2015).
5.2 - Giftighet og bioakkumulering i blåskjell (Mytilus spp.)
I tillegg til at Mytilus-arter er inkludert i SSD studiene nevnt over, er det blant annet vist i en eksponeringsstudie med tre mytilus arter (M. Edulis, M. galloprovincialis og M. trossulus) at 100 og 500 µg/l kobberklorid var dødelig for alle tre artene etter 21 dagers eksponering. Kontrollgruppene viste kobberklorid nivå mellom 1-2 mg/kg, mens skjell eksponert for 10 µg/l kobberklorid i 4 eller 21 dager gav en bioakkumulering til 6 mg/kg i M. galloprovincialis. M. edulis og M. trossulus ga bioakkumulering til 7-8 mg/kg Cu etter 4 dager og 17-18 mg/kg Cu etter 21 dager (Brooks mfl. 2015).
EC10 konsentrasjon for kobbereksponering av blåskjell larver (M. edulis) observert som påvirkning av larveutvikling er rapportert til 3,4 ug/l Cu (Deruytter et al., 2017). Kobbereksponering i en 48 timers embryo-larve test (M. trossulus) gav EC50 nivå på 6,9-9,6 ug/l (Nadella mfl. 2009).
Det er ikke gitt EQS-verdi for kobberinnhold i blåskjell (Miljødirektoratet, 2021). Kobbernivå i blåskjell er rapportert med gjennomsnittverdi på 0,99 mg/kg langs kysten av Norge i 2019 med et konsentrasjonsspenn på 0,72-1,5 mg /kg våtvekt (Duinker mfl. 2022). I overvåkingsprogrammet MILKYS viste de fleste stasjoner uendret tidstrend for kobbernivå i blåskjell langs norskekysten i 2019. En av 23 stasjoner gav økning og 3 av 23 stasjoner viste en nedgang (NIVA, 2020).
5.3 - Akkumulering i blåskjell tatt fra kobberimpregnert not
Blåskjell bioakkumulerer kobber over tid. Tabell 5 viser at blåskjell fra kobberimpregnerte nøter kan ha kobbernivå i innmat fra 2,9 til 86 mg/kg våtvekt. Laveste nivå fra not uten kobberimpregnering var 0,95 mg/kg. Flere av blåskjellprøvene som vokste på kobberimpregnert not inneholdt kobbernivå på tilsvarende nivå som rapportert fra Brooks mfl. (2015) der skjellene var eksponert for 10 µg/l kobberklorid.
Tabell 5. Bioakkumulering i blåskjell som vokser på kobberimpregnert not.
Sted
Dato
Cu i innmat (mg/kg våtvekt)
Størrelse på blåskjell (lengde, mm) Gjennom snitt ± stdav
Hardangerfjorden, kobberimpregnert not
Okt 2020
86
Austevoll, kobberimpregnert not
Okt 2020
2,9
Austevoll uten kobber (men med Tralopyril)
Okt 2020
0,95
Kobberprodusent: Blåskjell fra tau, ca 3 m fra ramme med kobberimpregnerte prøver
Mars 2021 Prøve nr 379
1,6
31±4
Kobberprodusent: Blåskjell fra ubehandla notlin, ca 3 m fra ramme med kobberimpregnerte prøver
Mars 2021 Prøve nr 380
1,7
28±4
Kobberprodusent: Blåskjell fra notlin 20 g/kg Cu2O og 25 g/kg tralopyril
Mars 2021 Prøve nr 381
10
31±3
Cu produsent: Blåskjell fra notlin 23 g/kg Cu2O
Mars 2021 Prøve nr 382
9,3
30±5
Cu produsent: Blåskjell fra notlin 26 g/kg Cu2O
Mars 2021 Prøve nr 383
8,2
28±9
Cu produsent: Blåskjell fra notlin 29 g/kg Cu2O
Mars 2021 Prøve nr 384
15
33±4
Cu produsent: Blåskjell fra notlin 28 g/kg Cu2O
Mars 2021 Prøve nr 385
21
28±8
Cu produsent: Blåskjell fra notlin 25 g/kg Cu2O og 1,5 g/kg Tralopyril
Mars 2021 Prøve nr 386
11
29±3
5.4 - Histopatologi i blåskjell som vokser på kobberimregnerte nøter
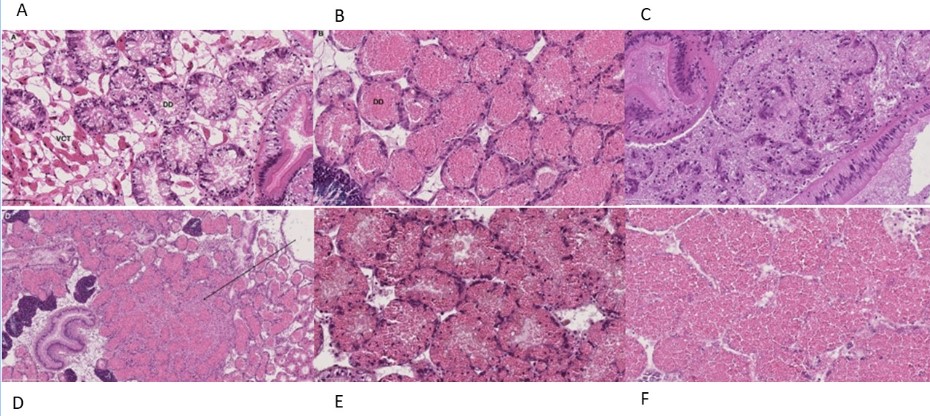
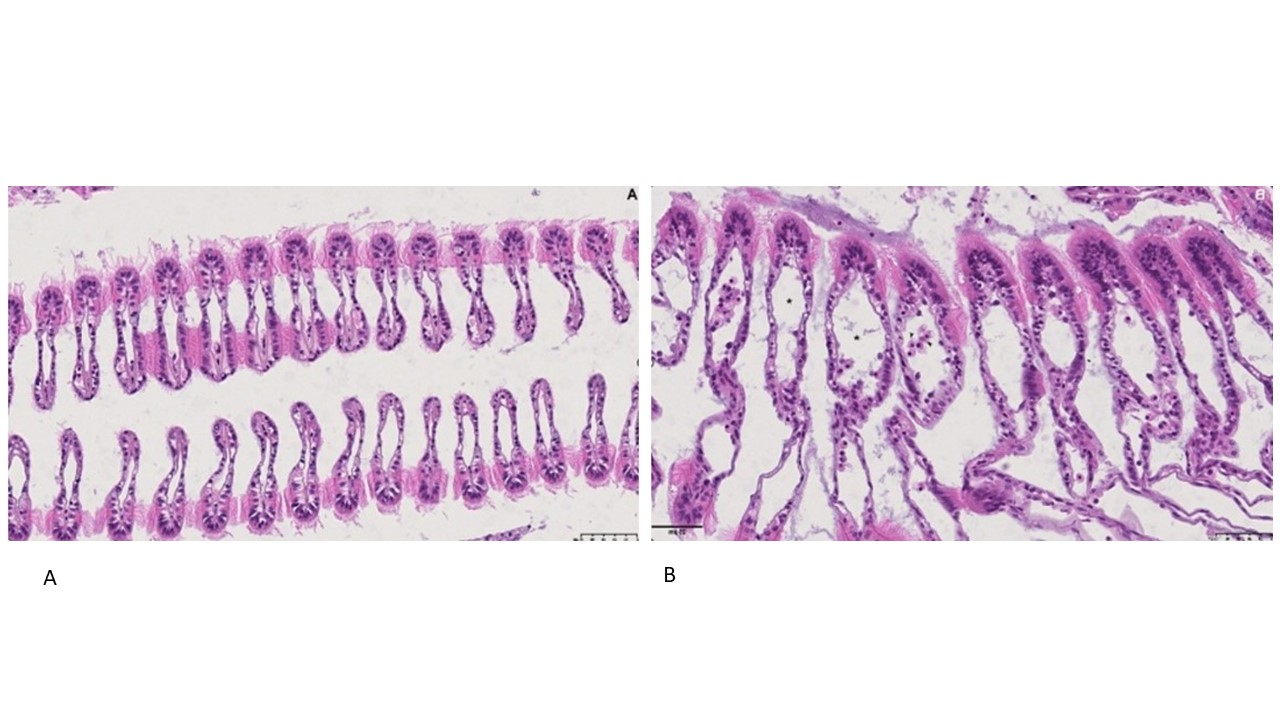
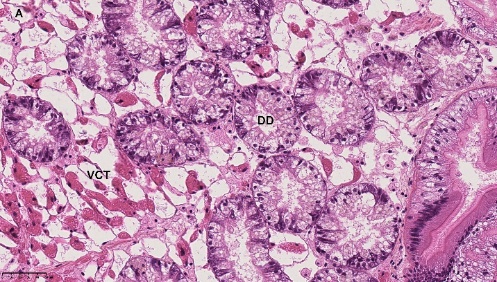
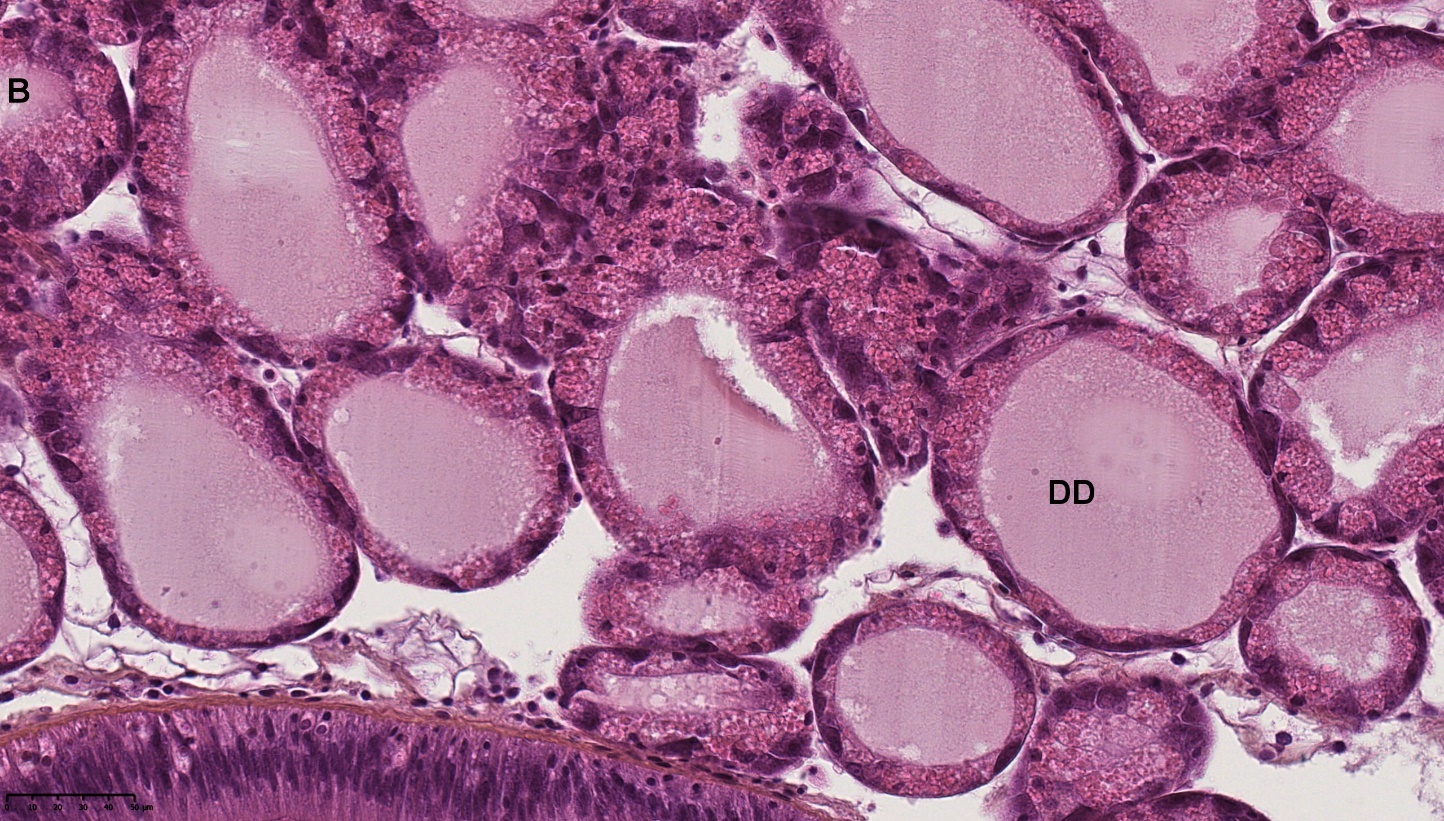
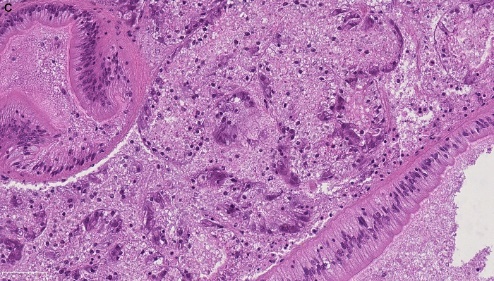
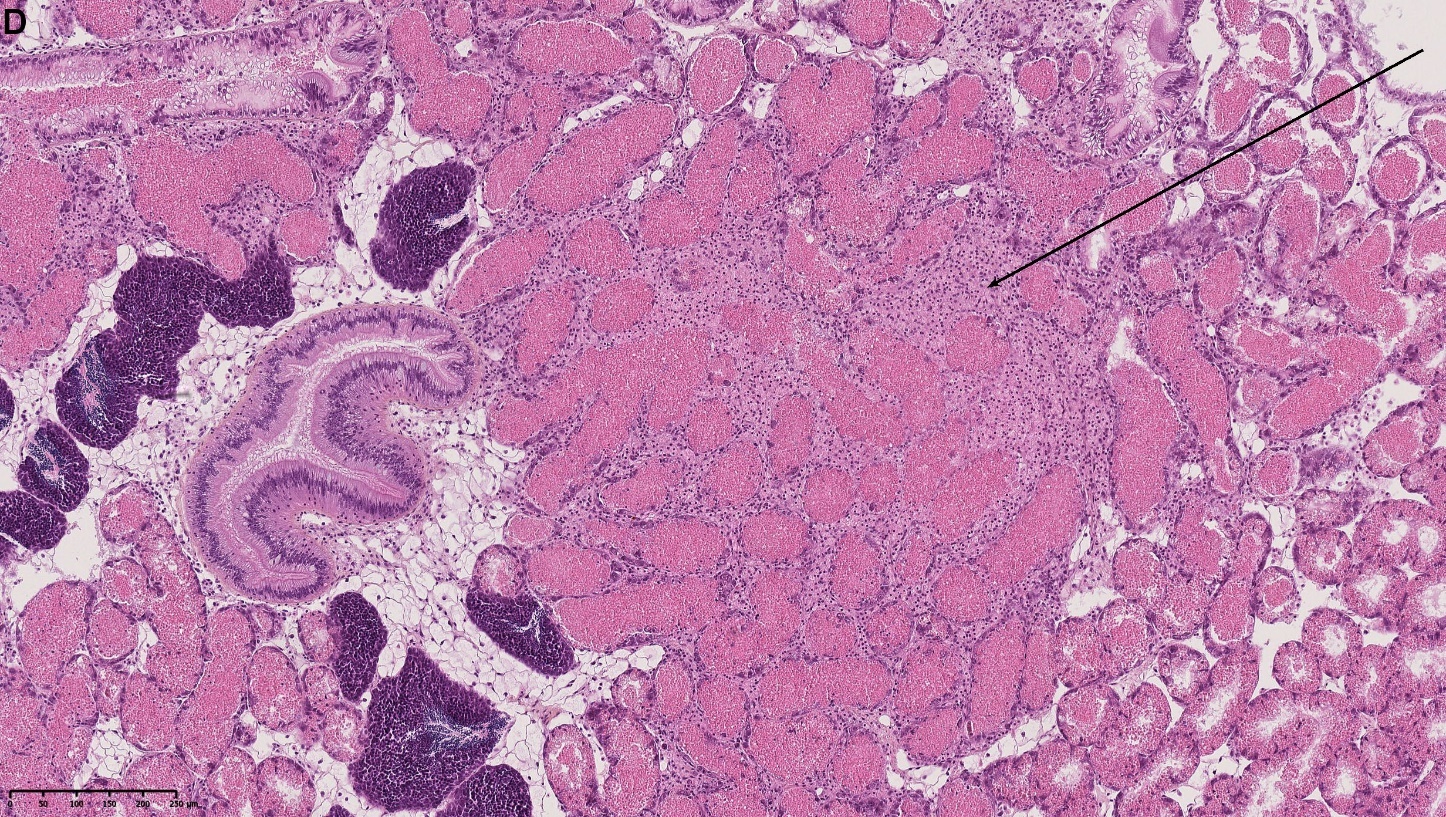
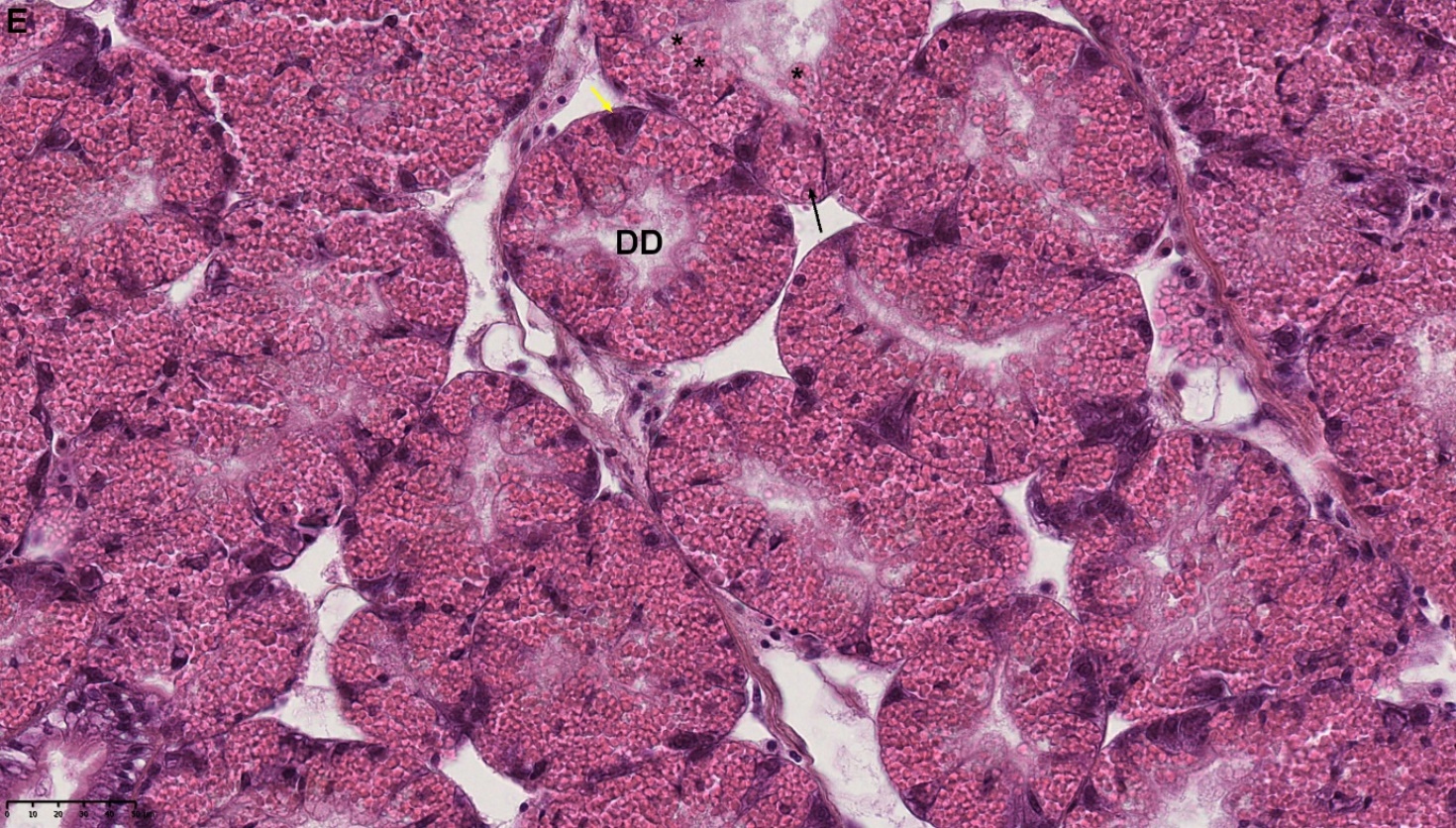
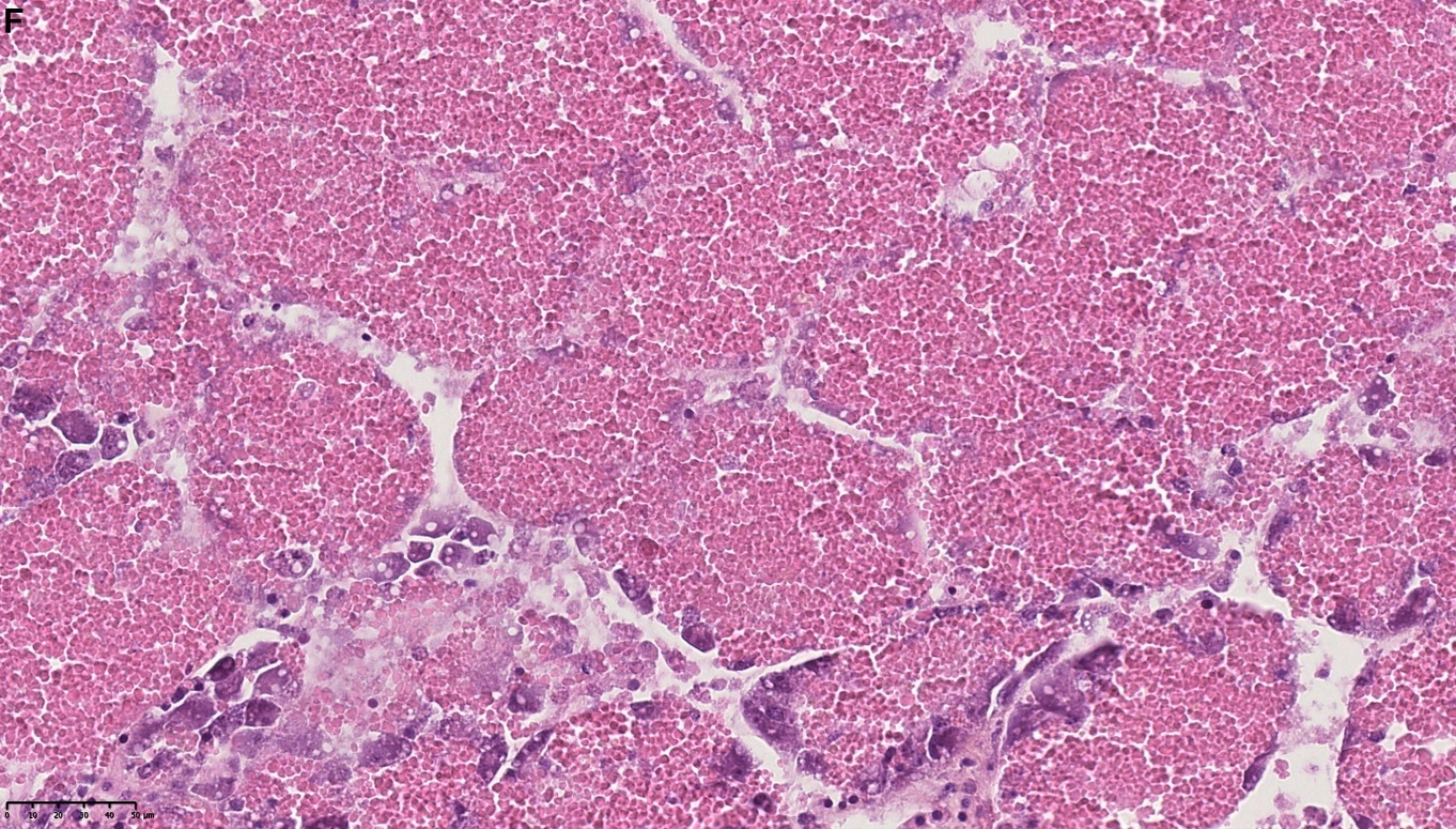
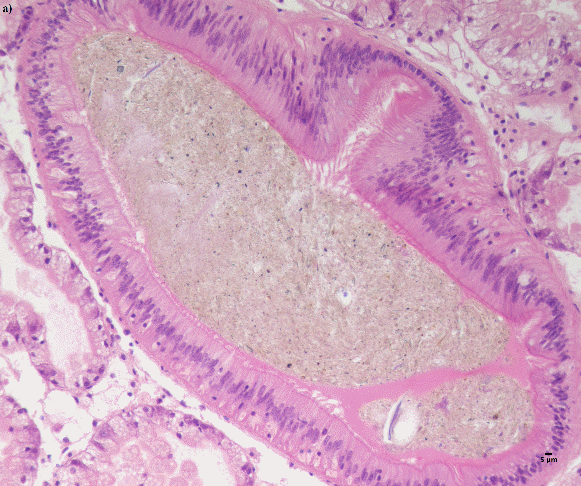
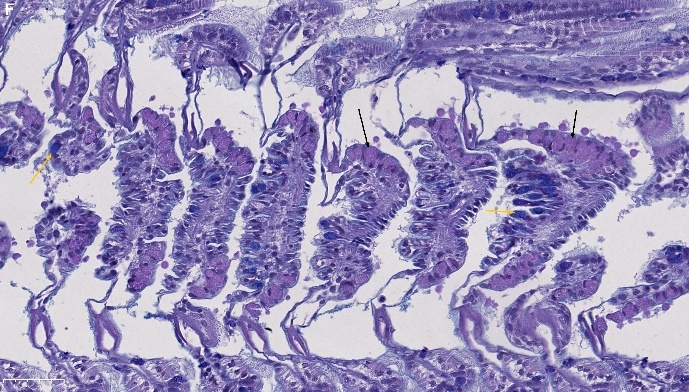
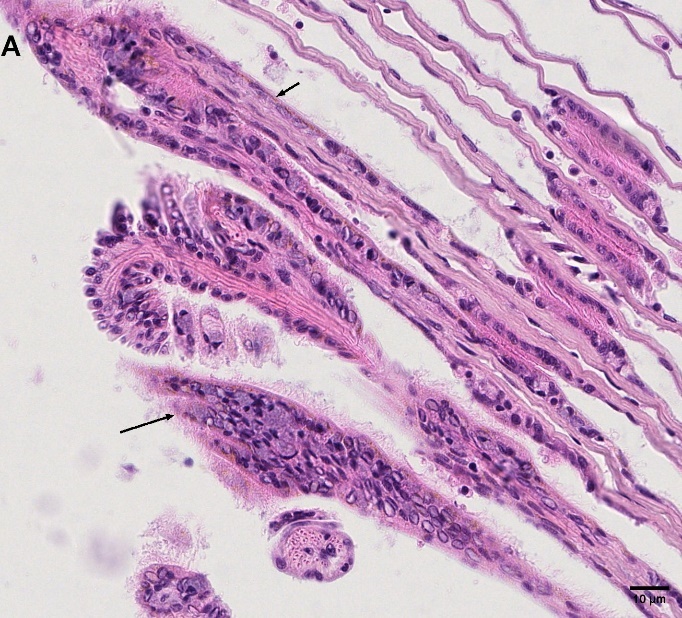
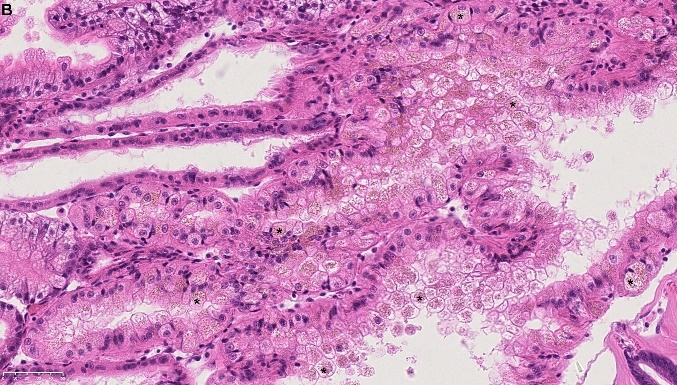
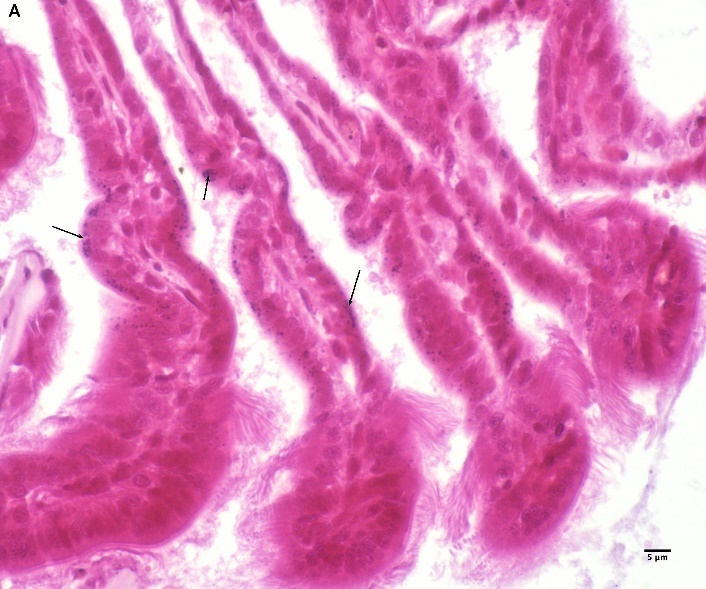
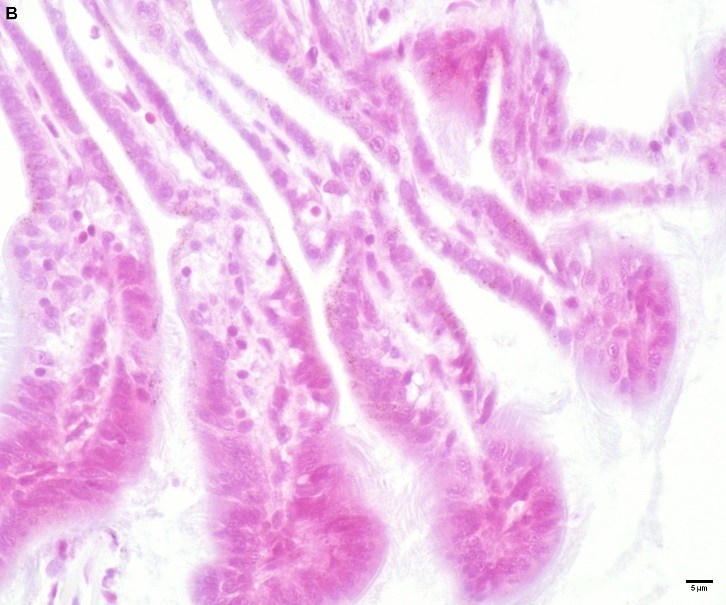
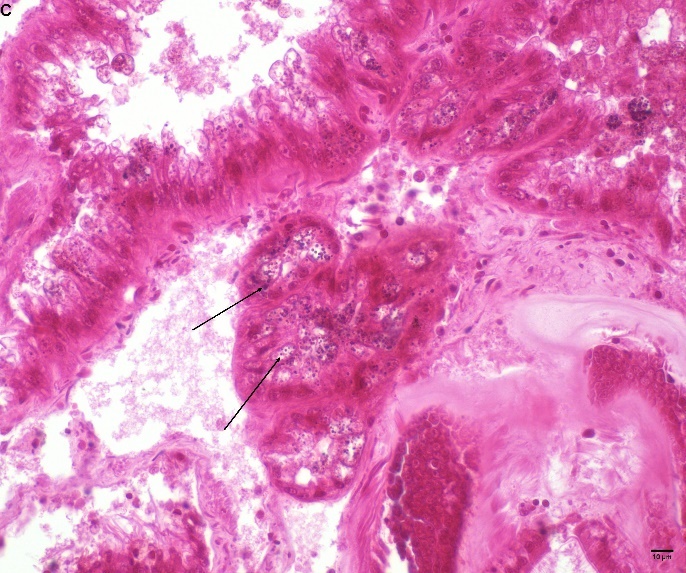
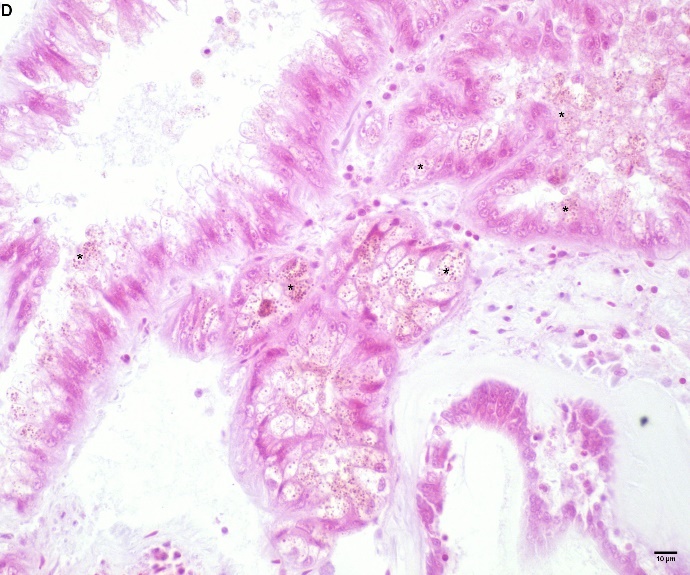
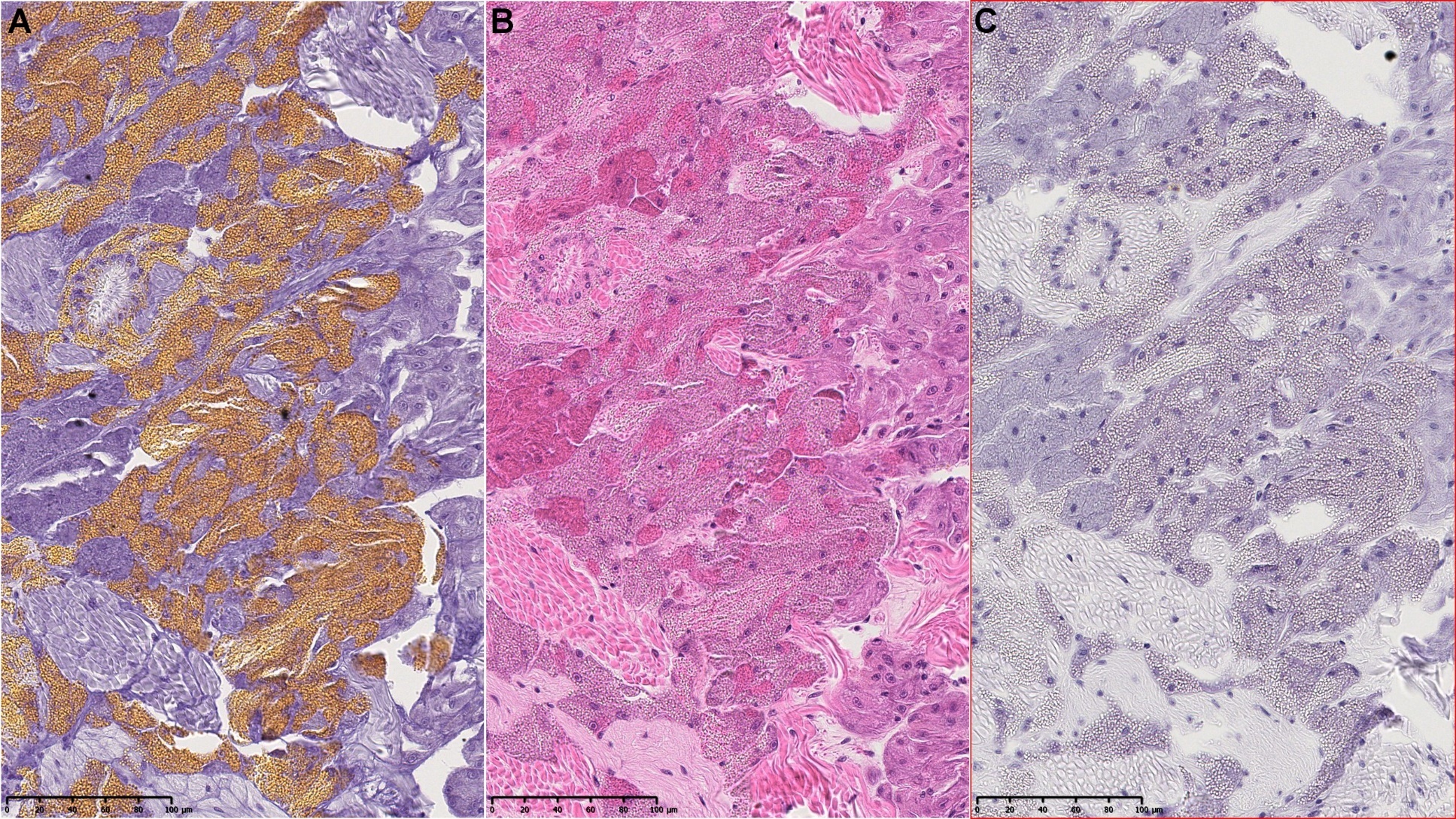
Blåskjell fra Kap 5.3 er blitt for undersøkt histologiske endringer i fordøyelseskjertel og gjelle. Det er sett på vevsendringer i blåskjell fra oppdrettsanlegg som bruker kobberimpregnerte nøter i Hardangerfjorden (n=15) og fra blåskjell som vokser på ulike kobber behandlete notprøver tilsendt fra en kobbeimpregneringsprodusent (n= 10 per gruppe, totalt 5 grupper). Metoden er beskrevet i Vedlegg 1. Vi observerte flere endringer i forøyelseskjertelen i blåskjell som vokste på kobberimpregnert merd i forhold til det som er normal histologi for fordøyelseskjertel i blåskjell. Disse endringene inkluderte høy forekomst av atrofi (innskrumping) av divertikler (utposninger) i fordøyelseskjertelen, i tillegg til nedbrytning av vev i fordøyelseskjerten, inflammasjon i bindevevet til fordøyelseskjertelen og lysosomale endringer i vesiklene i fordøyelsescellene og tegn på celledød i vevsstrukturen (Figur 8).
Figur 8. Endringer observert ved histologi i fordøyelseskjertel fra blåskjell som vokser på kobberimpregnert not. A) Normal fordøyelseskjertel, B) Atrofi (innskrumping) av divertikler (utposninger) i fordøyelseskjerten (digestive diverticula (DD)), C) Nedbrytning av vev i fordøyelseskjerten, 20x, D) Alvorlig inflammasjon i bindevevet (vesicular connective tissue (VCT)) til fordøyelseskjertelen (vist med pil), E) Lysosomale endringer i vesiklene i fordøyelsesceller i DD. F) Celledød i DD strukturen. Basalmembrane og basofile celler forsvinner.
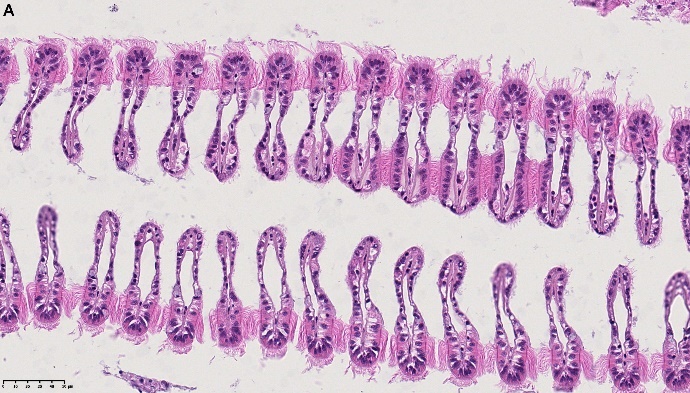
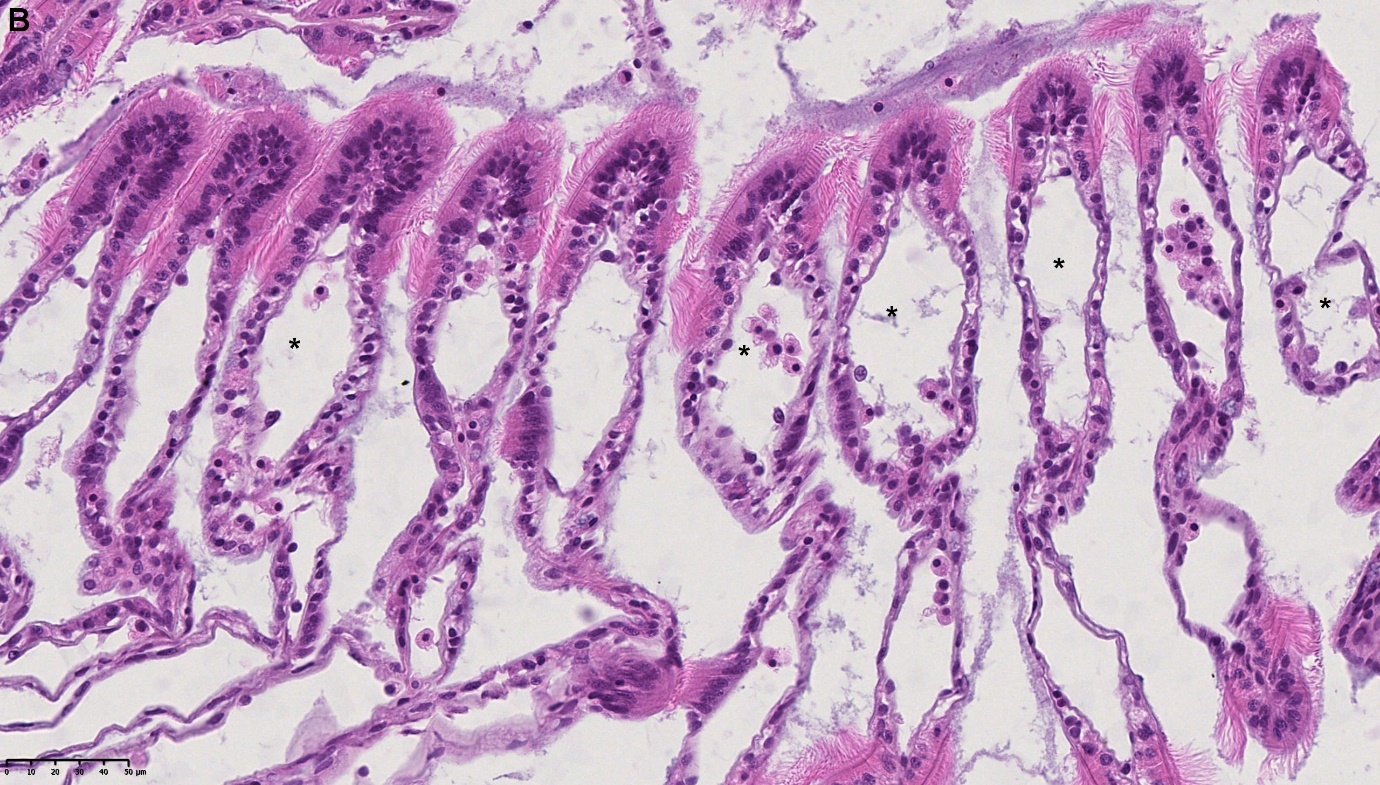
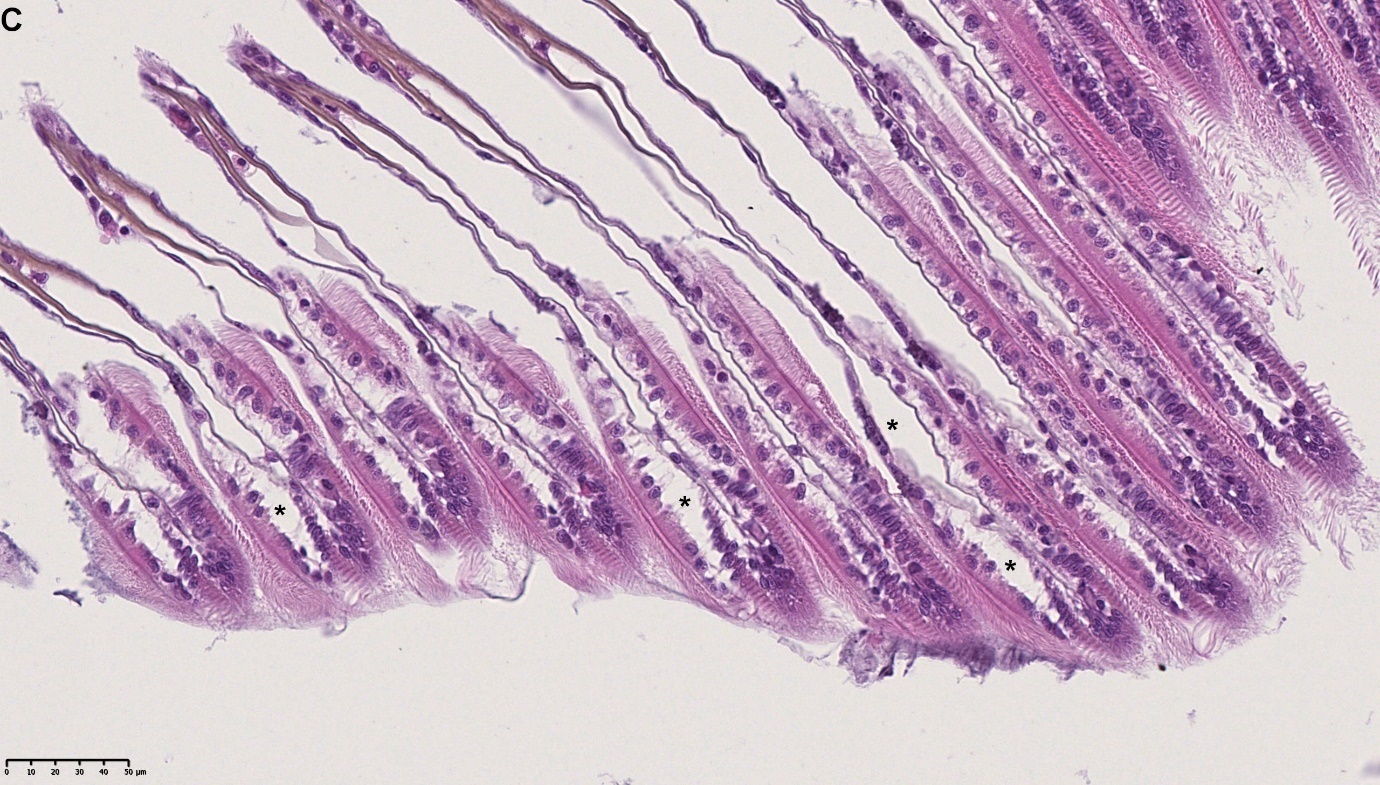
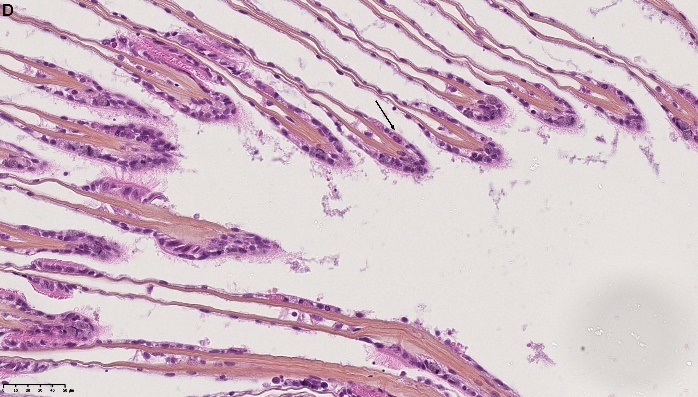
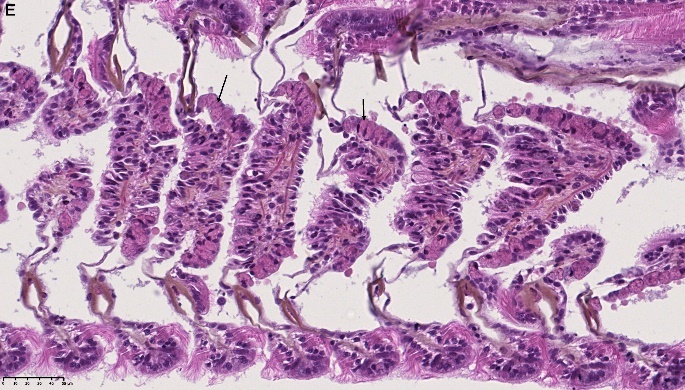
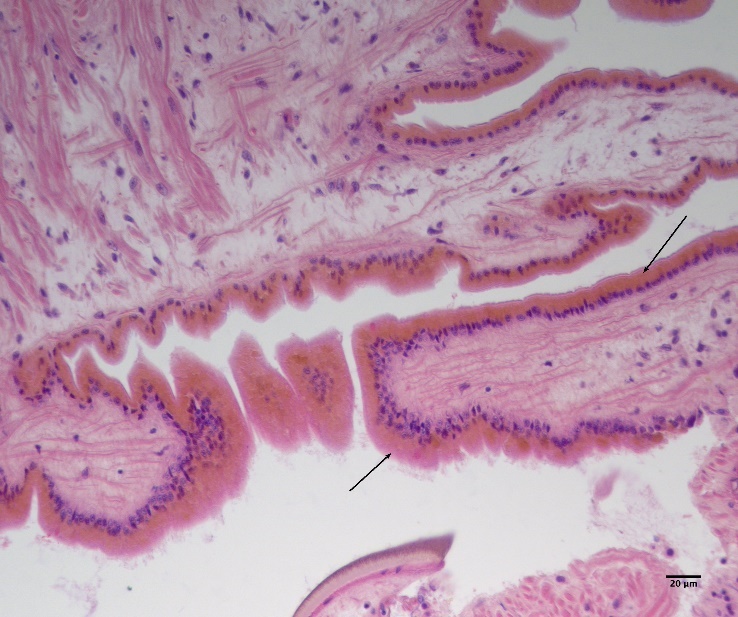
Det ble i tillegg observert endringer i gjellevevet som inkluderte forstørring av subepitelialt rom (Figur 9), men disse endringene var ikke så gjennomgående som endringene i fordøyelseskjertelen.
Figur 9. Endringer observert ved histologi i gjeller fra blåskjell som vokser på kobberimpregnert not. A) Normale gjeller. B) Forstørring av subepitelialt tom i blåskjell som er tatt fra kobbereksponert not med hemocytter (vist med pil) i det branchiale rommet (vist med *).
For å vurdere grader av atrofi i divertikler i fordøyelseskjertelen har vi brukt gradering som er vist i Tabell 6 og beskrevet i Ellis mfl. (1998) og beskrevet i Vedlegg 1. Denne klassifiseringen skiller grad av atrofi i fem klasser fra 0 (normal) og til svært stor grad av atrofi i divertiklene (4). Grad av histopatologiske endringer i blåskjell som vokser på kobberimpregnert not er vist i Tabellene 7-11. I blåskjell fra kobber-impregnert not fra Hardangerfjorden var 27 % normale og de resterende hadde ulik grad av atrofi, mens i skjellene fra både ubehandlet notlin og behandlet notlin hadde de fleste skjellene atrofi gradering fra 2-4 og vi såg klare avvik fra det som er forventet som normaltilstanden i blåskjell.
Resultatene viser at metoden er i stand til å fange opp endringer i skjell som vokser på kobberimpregnert not og som har ulik grad av kobberakkumulering i bløtdelene, men vi trenger å gjøre dose-respons og tid-respons forsøk for å validere om vi kan bruke disse metodene i feltovervåking av blåskjell og har pågående forsøk på dette.
Tabell 6. Gradering for atrofi (innskrumping) av divertikler i fordøyelseskjertelen.
Gradering
Beskrivelse
0
Normal veggtykkelse i flesteparten av tubuli (0% atrofi), få tubuli med litt atrofi.
1
Veggtykkelse: Mindre enn halvparten av tubuli har atrofi
2
Veggtykkelse i gjennomsnitt halvparten så tykk som normalt.
3
Mer enn halvparten av tubuli har atrofi. Noen vegger er ekstremt tynne.
4
Omtrent alle tubuli er atrofiert med ekstrem tynn vegg
Tabell 7. Histopatologisk skoring for atrofierte tubuli i fordøyelseskjertel i blåskjell fra Cu impregnert not fra Hardangerfjorden (n=15). Samleprøve av kobber i innmat ga kobber nivå på 86 mg/kg våtvekt).
Tabell 7. Histopatologisk skoring for atrofierte tubuli i fordøyelseskjertel i blåskjell fra Cu impregnert not fra Hardangerfjorden (n=15). Samleprøve av kobber i innmat ga kobber nivå på 86 mg/kg våtvekt).
Prøve ID
0
1
2
3
4
Prevalens (%)
27
20
7
13
33
Tabell 8. Histopatologisk skoring for atrofierte tubuli i fordøyelseskjertel i blåskjell fra tau, ca 3 m fra ramme (fra kobberprodusent) (n=10). Samleprøve av kobber i innmat ga kobbernivå på 1,6 mg/kg våtvekt).
Prøve ID
0
1
2
3
4
Prevalens (%)
50
10
40
Tabell 9. Histopatologisk skoring for atrofierte tubuli i fordøyelseskjertel i blåskjell fra kobberprodusent: Blåskjell fra ubehandlet notlin (fra kobberprodusent) (n=10). Samleprøve av kobber i innmat gav kobbernivå på 1,7 mg/kg våtvekt).
Prøve ID
0
1
2
3
4
Prevalens (%)
10
60
30
Tabell 10. Histopatologisk skoring for atrofierte tubuli i fordøyelseskjertel I blåskjell fra kobber produsent: Blåskjell fra ubehandla notlin (fra Cu produsent) (n=10). Samleprøve av Cu i innmat gav Cu nivå på 15 mg/kg våtvekt).
Prøve ID
0
1
2
3
4
Prevalens (%)
10
60
30
Tabell 11. Histopatologisk skoring for atrofierte tubuli i fordøyelseskjertel I blåskjell fra kobber produsent: Blåskjell fra notlin 29 g/kg Cu2O (fra Cu produsent) (n=10). Samleprøve av Cu i innmat gav Cu nivå på 21 mg/kg våtvekt).
Prøve ID
0
1
2
3
4
Prevalencs (%)
20
10
30
40
6 - Modellering av spredning av kobber brukt som antibegroingsmiddel
Spredning av kobber fra oppdrettsanlegg skjer via ulike prosesser:
Partikulært utslipp av kobber tilsatt i fôret, via forspill og fiskeavføring
En puls av partikulært kobber etter høytrykksspyling av nøter
En puls av vannløst kobber etter høytrykksspyling av nøter
Kontinuerlig utlekking av kobber til vannfasen fra impregnerte nøter
Kontinuerlig partikulært utslipp av kobber fra impregnerte nøter pga. erosjon
Vi har bare gjort detaljerte modellstudier av prosess 4 i denne rapporten.
6.1 - Fôrspill og fekalier
Fra Tabell 1 har vi at 1000 tonn fôr gir omtrent 9 kg kobber bundet til organiske partikler. Fôrforbruket i en typisk sirkelmerd med diameter 50 m (omkrets 157 m) er i størrelsesorden 500 tonn per år. Hovedtyngden av partiklene sedimenterer under anlegget, med mindre det er omfattende resuspensjon på stedet (Carvajalino-Fernández mfl. 2020). Dette gir en tilførsel på noen få gram kobber per m2 per år, dersom vi antar et verstefallsscenario der alt partikulært materiale faller rett ned og ikke blir resuspendert.
6.2 - Partikulært og løst kobber etter spyling
Mange anlegg har praktisert høytrykksspyling av impregnerte nøter for å hindre begroing. Store deler av impregneringsmaterialet vil også bli revet bort eller oppløst i denne prosessen. En av aktørene i bransjen (personlig kommunikasjon) estimerer at grovt regnet 20 % av opprinnelig kobbermengde kan forsvinne fra noten ved førstegangs spyling (varighet omtrent 2 timer). For en not med kobberinnhold 400 kg kobber (typisk verdi for sirkelmerd med 50 m diameter og 157 m omkrets) tilsvarer dette et enkeltutslipp på 80 kg kobber. Vi har ikke informasjon om hvor stor del av dette utslippet som er i form av partikulært og vannløst kobber, men resultat fra Kap 4.2 viser at spyling gir et høyt utslipp av partikler som sedimenterer.
Hvis vi antar at alt kobbertap under høytrykksspyling faller ned som partikulært materiale, tilsvarer dette et utslipp som er i størrelsesorden 20 ganger større enn et års kobberutslipp fra fôrtilsetning. Synkehastigheten til eroderte kobberpartikler er ikke kjent. Partikler med lav synkehastighet vil spres over et større område, mens tyngre partikler vil sedimenteres like i nærheten av merdområdet.
Hvis vi antar at kobberet løses fullstendig i vann, vil effekten være svært avhengig av strømforholdene på stedet. Kobbermengden må fortynnes til et volum på 1250m x 1250m x 20m for å komme ned i en konsentrasjon på 2,6 µg/L, som er miljøkvalitetsstandarden for kystvann. Vi har ikke gjort noen detaljert modellstudie på noen disse scenariene.
Et oppdrettsanlegg består gjerne av 4-10 slike merder. Spyling vil derfor kunne føre til utslipp av tilsvarende mange pulser av løst kobber gjennom en spylebehandling.
6.3 - Kontinuerlig utlekking av partikulært og vannløst kobber
En kobberimpregnert not vil avgi kobber selv uten spyling. I et feltforsøk av ble restinnholdet av kobber i 58 nøter målt etter endt levetid. Gjennomsnittlig restinnhold var 54 %, men for nøtene som ikke ble høytrykksspylt (7 av 58) var restinnholdet 72 %. Dette betyr at andel utlekket kobber uten høytrykksspyling var 28 %, mens nøter som ble høytrykksspylt mistet ytterligere 18 % av opprinnelig kobberinnhold (Ulriksen, 2020). Vi har ingen mulighet til å vurdere representativiteten i disse forsøkene, men de gir likevel en god indikasjon på hva som er en rimelig utlekkingsrate. Miljødirektoratet har estimert at 80-90 % av kobberet fra en kobberimpregnert not lekker ut av en nots levetid (180 dager i sjø) (Skarbøvik mfl. 2017; https://www.miljodirektoratet.no/ansvarsomrader/vann-hav-og-kyst/Akvakultur-fiskeoppdrett/).
Nøter som ikke blir høytrykksspylt, avgir både vannløst kobber og partikulært kobber som løsner fra noten (f.eks. i forbindelse med heving/senking av noten). Det antas at utlekking av vannløst kobber er dominerende.
6.4 - Modellert samlet utslipp av vannløst kobber for alle norske anlegg
Vi har utført simuleringer av kontinuerlig utlekking av vannløst kobber for alle anlegg langs norskekysten, med spesielt fokus på de oppdrettsintensive produksjonsområde 2 Ryfylke og 3 Karmøy til Sotra. I dette scenariet antar vi at alt kobber lekker ut som løste ioner i vannet. I virkeligheten vil mye av kobberet falle ned på bunnen som partikulært materiale og vil dermed få en mer lokal spredning. Simuleringen bør derfor betraktes som et verstefalls-estimat for spredning av løst kobber fra antibegroingsmidler. Bedre tall på andel som lekker som ioner i forhold til partikler som synker trengs for at denne modelleringen skal bli mer realistisk.
Havstrømmer er hentet fra modellsystemet NorKyst800, som dekker hele norskekysten med en horisontal oppløsning på 800 m og en vertikal oppløsning på 35 dybdelag (Albretsen mfl. 2011). Modellen er basert på ROMS (Regional Ocean Modeling System, Shchepetkin & McWilliams, 2005). Havmodellen drives av atmosfæriske drivkrefter fra AROME MetCoOp (Müller, 2017 ) og elveavrenning fra NVE (https://nve.no). Modellen har vist å gi gode resultater i sammenligning med direkte måledata (Dalsøren mfl. 2020). For simuleringer som bare omfatter et enkelt produksjonsområde har vi brukt NorFjords160, et modellsystem som er basert på NorKyst800, men med horisontal oppløsning på 160 m.
Spredning av kobber ble modellert med partikkelspredningsmodellen LADiM (https://github.com/bjornaa/ladim), med havstrømmer fra NorKyst800 og NorFjords160 som drivkrefter. Virtuelle partikler ble sluppet kontinuerlig fra alle oppdrettsanlegg i simuleringsområdet. Posisjonen til anleggene og maksimalt tillatt biomasse (MTB) ble hentet fra Akvakulturregisteret (https://www.fiskeridir.no/Akvakultur/Registre-og-skjema/akvakulturregisteret ).
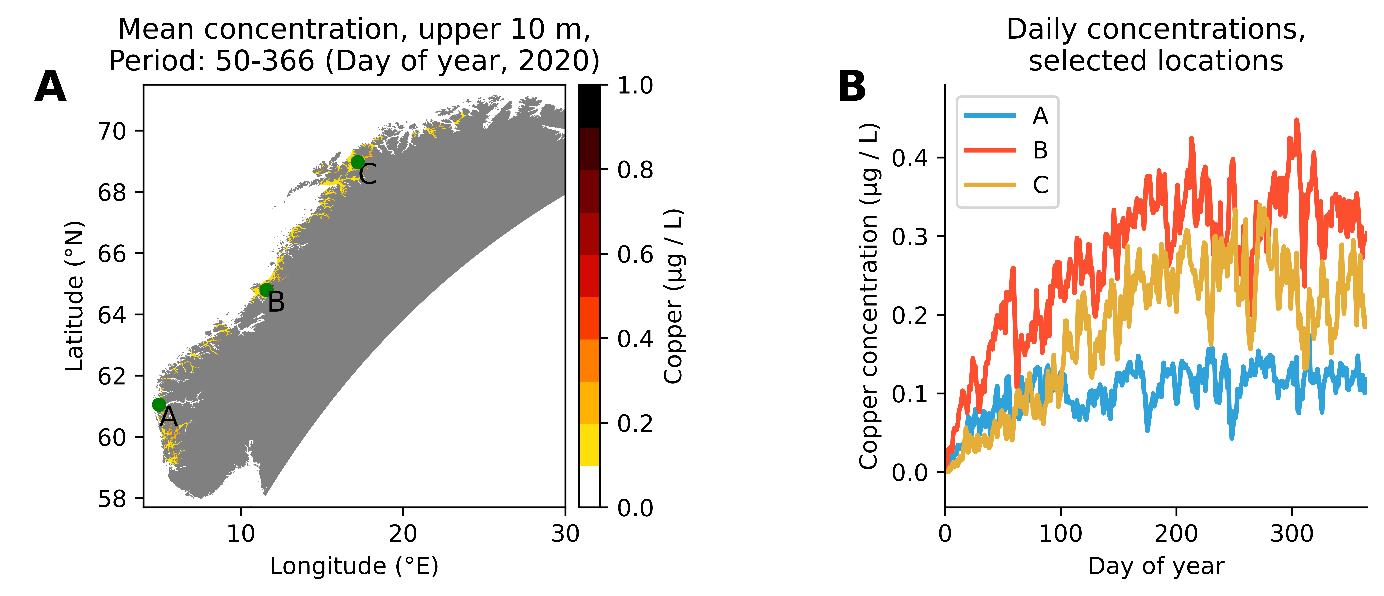
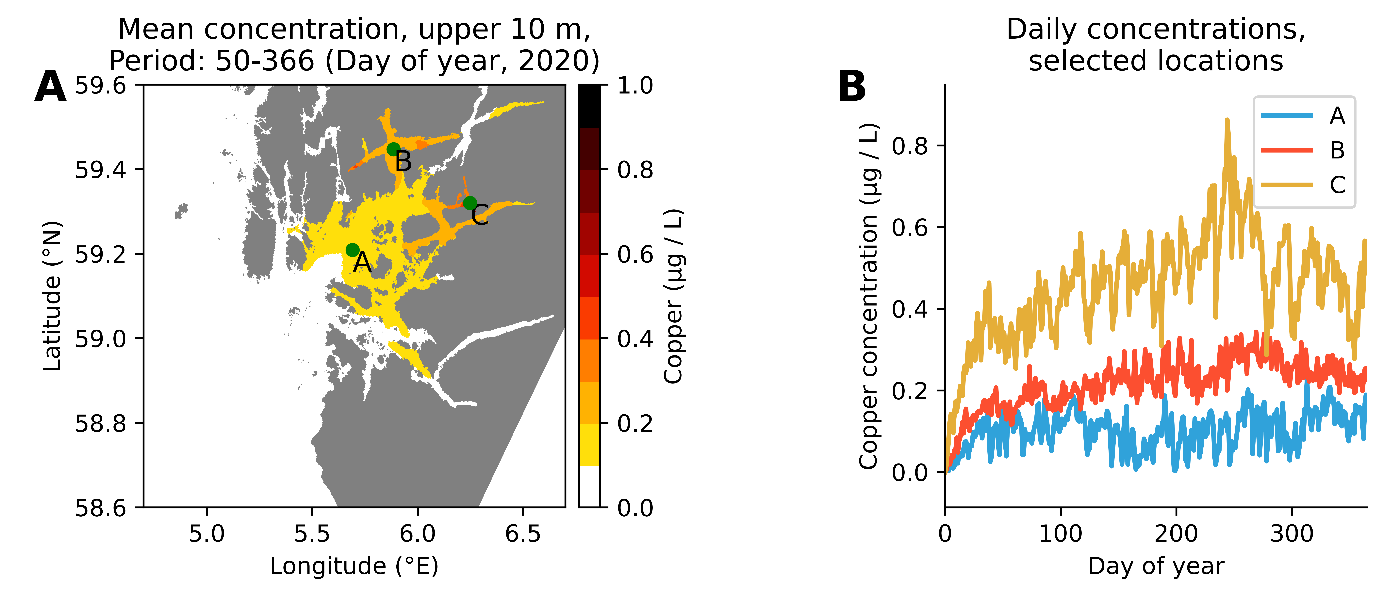
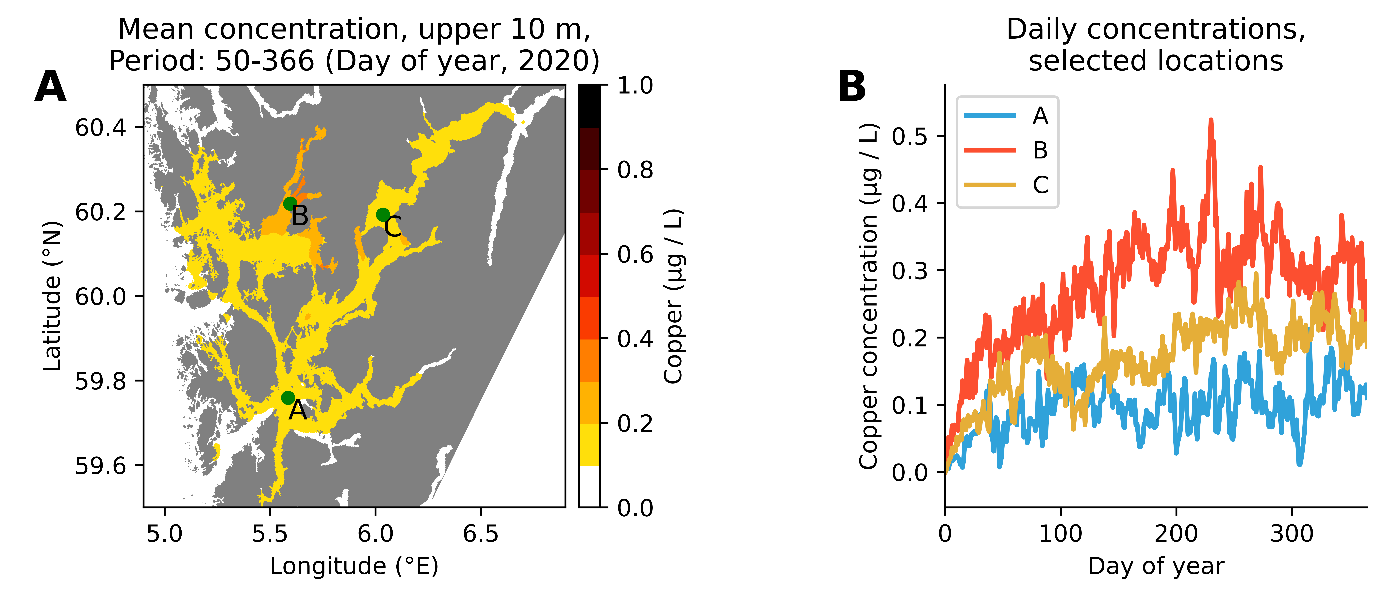
Vi har utført 3 sett med simuleringer: En for hele norskekysten (770 tonn kobber fordelt på 1251 anlegg), (Figur 10), en for produksjonsområde 2 Ryfylke (totalt 40 tonn kobber fordelt på 73 anlegg, Figur 11) og en for produksjonsområde 3 Karmøy til Sotra (totalt 81 tonn kobber fordelt på 172 anlegg, Figur 12). Simuleringene har vært kjørt fra 1. jan 2020 til 31. des 2020.
Forutsetninger for simuleringen:
Totalmengde kobberutslipp til vann er antatt å være 770 tonn for hele 2020. Dette er basert på tall fra Produktregisteret (Figur 1) og en utlekkingsrate på 50 % per år. Dette er et verstefalls-estimat, der vi antar at alt tap av kobber skjer som løste ioner i vann.
Mengde kobber per oppdrettsanlegg er vektet på maksimalt tillatt biomasse, slik at store anlegg antas å slippe ut mer kobber enn små anlegg. I realiteten er kobberutslipp proporsjonalt med anleggets totale notareal, men denne detaljerte informasjonen er ikke tilgjengelig. Maksimalt tillatt biomasse ble derfor valgt som en rimelig vektingsparameter.
I simuleringen antas det at alle anlegg bruker kobber til enhver tid. Vi mangler dessverre detaljert informasjon om hvor mange anlegg som bruker kobber. Det finnes fjorder der simuleringen viser betydelige kobberverdier, men som i praksis har mye lavere verdier fordi anleggene i området ikke benytter kobber.
Kobberpartiklene er sluppet i dybden 0-30 meter. Vi antar at en typisk not er sirkulær med konstant radius de øverste 15 meter, og deretter kjegleformet ned til 30 meters dybde.
Simuleringen viser at passivt utlekket kobber kan gi et vesentlig bidrag til den totale kobberkonsentrasjonen i et fjordsystem, i størrelsesorden 0,2-0,4 µg/l. I trange fjorder med dårlig vannutveksling kan bidraget periodevis være opp til 1 µg/l. Disse tallene er direkte proporsjonale til antatt utlekkingsrate, dvs. tallene ville vært halvparten så høye dersom totalmengden kobberutslipp til vann i 2020 var 385 tonn (25% utlekking) i stedet for 770 tonn. Dette vil for eksempel være tilfelle dersom vi antar at 25 % av opprinnelig kobbermengde lekker ut som løst kobber og 25 % faller ned som partikulært kobber og blir værende i det lokale sedimentet. En slik antagelse er ikke urimelig sett i lys av feltforsøket til Ulriksen (2020) som er beskrevet tidligere.
Simuleringen viser midlere konsentrasjoner på en grov skala (flere kilometer). Lokalt rundt et oppdrettsanlegg kan konsentrasjonen derfor være høyere enn simuleringen indikerer.
Simuleringen viser en viss akkumulering av konsentrasjon over tid, men etter 1-6 måneder (avhengig av lokale forhold) vil det dannes en likevekt mellom vannutskifting og tilført kobber, slik at konsentrasjonen ikke stiger ytterligere.
Figur 10. Simulert konsentrasjon av utlekket kobber fra oppdrettsnøter langs norskekysten. A: Midlere konsentrasjon i løpet av et år (ekskludert de første 50 dager), som viser høy fortynning utenfor grunnlinjen, men større konsentrasjon i oppdrettsintensive fjorder. B: Konsentrasjon av kobber dag for dag på tre utvalgte posisjoner (se kart til venstre), som viser at konsentrasjonen stiger inntil det dannes en likevekt mellom vannutskiftning og tilført kobber.
Figur 11. Simulert kobberkonsentrasjon i produksjonsområde 2 Ryfylke. A: Midlere konsentrasjon i løpet av et år (ekskludert de første 50 dager). B: Konsentrasjon av kobber dag for dag på tre utvalgte posisjoner (se kart til venstre).
Figur 12. Simulert kobberkonsentrasjon i produksjonsområde 3 Karmøy til Sotra. A: Midlere konsentrasjon i løpet av et år (ekskludert de første 50 dager). B: Konsentrasjon av kobber dag for dag på tre utvalgte posisjoner (se kart til venstre).
Modellsimuleringer viser at passivt utlekket kobber kan gi et vesentlig bidrag til den totale kobberkonsentrasjonen i et fjordsystem, i størrelsesorden 0,2-0,4 µg/l. I trange fjorder med dårlig vannutveksling kan bidraget periodevis være opp til 1 µg/l. Spyling eller høytrykksspyling av kobberimpregnerte nøter vil kunne gi pulser med høyere konsentrasjoner. Vi mangler data på hvor stor andel kobber som faller ut som partikler og som synker ned i anleggssonen og hvor mye som blør ut som kobberioner til vannsøylen. Det er likevel grunn til å anta at bidraget fra kobberimpregnerte merder kan gi et vesentlig bidrag til kobberkonsentrasjonen i vannsøylen i oppdrettsintensive fjorder. Miljøkvalitetsmål for kobber i kystvann er 2,6 µg/l. Vi har målt konsentrasjoner i fjorder i Vestland fra < 0,5 til 1,9 µg/l i 2021 (Figur 2).
7 - Erstatningsstoff
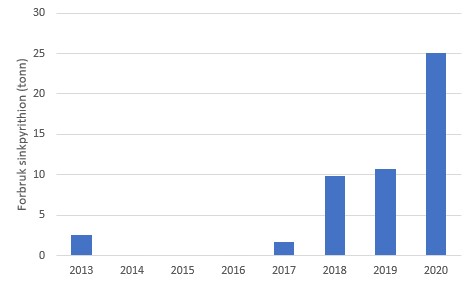
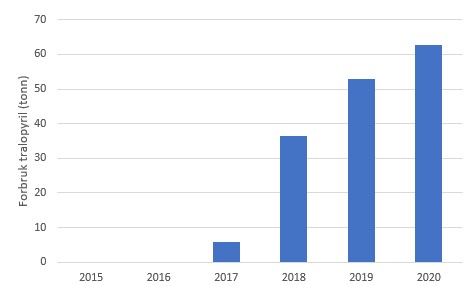
Etter at det i de siste årene har blitt mer fokus på miljøeffekter av kobber har bruken av erstatningsstoffene tralopyril og sinkpyrithion økt siden 2017 (Figur 13), gjerne i kombinasjon med økt hyppighet av spyling. Disse to erstatningsstoffene er effektive biocider og det er viktig at risikoen knyttet til bruk av disse stoffene blir vurdert slik at midlene som skal erstatte kobber ikke blir nye miljøutfordringer. Også her bør en dokumentere eventuell spredning av spyling.
Tralopyril er et bredspektret biocid brukt som antibgroingsmiddel i kobberfrie formuleringer. Tralopyril forstyrrer oksidativ fosforylering i mitokondrier (ECHA, 2014). Tralopyril ble godkjent til bruk i 2014 men med strenge krav til å redusere utslipp til miljøet (EU, 2014). EU kommisjonens gjennomføringsforordning skriver om godkjenning av tralopyril som et nytt aktivt stoff til bruk i biocidprodukter i produkttype 21, at «for godkjente produkter skal det fremgå av etiketter og eventuelle sikkerhetsdatablad at bruk, vedlikehold og reparasjon skal foregå innenfor et avgrenset område, helst på et hardt og ugjennomtrengelig underlag med oppsamling eller hvis underlaget er jord skal den være dekket av et ugjennomtrengelig materiale for å unngå spill og minske utslipp til miljøet. Eventuelt spill og avfall skal samles oppfor gjenbruk eller destruksjon.» Biocid-pt21 - regjeringen.no . Produktvurderingen skal ta særlig hensyn til eksponering, risiko og virkning knyttet til bruk som omfattes av en søknad om godkjenning, men som ikke er tatt i betraktning i risikovurderingen av det aktive stoffet på unionsplan 32014r1091.pdf (lovdata.no).
Miljøkvalitetsstandard for kobberpyrithione i kystvann er foreslått til 0,014 µg/l. Tilsvarende er ikke bestemt for tralopyril, men miljøkvalitetsstandard for ferskvann er foreslått til 0,002 µg/l (Martins mfl. 2018).
A
B
Figur 13. Forbruk av A: sinkpyrithion og B: tralopyril til bruk i akvakultur. Kilde: Produktregisteret.
8 - Oppsummering
Vi har sammenstilt potensial for spredning av kobber med grenseverdier i kystvann og sediment og med målte nivå i sjøvann og i sediment i anleggssonen, i overgangssonen og i fjernsonen. Økte nivå på grunn av utblødning fra nøter er modellert og mulige ekstrapulser på grunn av spyling er beskrevet.
Selv om vi har god kunnskap om konsentrasjoner av kobber under og i nærsonen til oppdrettsanleggene er det manglende kunnskap om hvor tilgjengelig ulike kobberformer er i sedimentet og det mangler også giftighetsdata for flere av artene som lever i sediment under oppdrettsanleggene.
Kobbertilførsler til marine vannressurser er noe som bør følges nøye med på. Per i dag finnes ingen overvåking av kobber i vannsøylen i tilknytning til områder med fiskeoppdrett. Overvåkning av økosystem i kystområder (Økokyst) måler innholdet av næringsstoffer (N, P, Si), oksygen, organisk karbon, karbon samt partikler (TSM) siktedyp, temperatur og saltholdighet i sine vannprøver. Det bør vurderes om også kobber skal inkluderes i denne overvåkingen, samt å utvide antall stasjoner som undersøkes langs kysten.
Spredning og fortynning er en viktig faktor for å forklare variasjonen av hvordan kobber akkumulerer på den enkelte oppdrettslokalitet. Det vil være behov for å utvikle bedre spredningsmodeller basert på kobberets egenskaper i de ulike fasene (gjennom fôrspill, fekalier, ved spyling (flak og større partikler) og løst i vann). Vi trenger mer data på hvor stor andel kobber som blør ut og hvor stor andel som synker ned i sedimentene i nærsonen og som spres til overgangssonen. Dette vil gi oss bedre grunnlag for modellering av spredning og påvirkning.
C-undersøkelsene inkluderer endringer i artssammensetning. Det er over tid samlet inn mye data om dette, men disse dataene har ikke vært systematisert og analysert. En samlet gjennomgang av dette materialet sett opp mot utslipp av kobber og andre påvirkningsfaktorer er planlagt og vil gi oss bedre data for fremtidige vurderinger av risiko knyttet til miljøeffekter av kobber. Det er likevel krevende å skille ulike påvirkninger ved endring i artssammensetning over tid, dvs å klare å skille mellom effekten av kobber i forhold til effekten av organisk materiale/ redusert oksygen/pH etc.
Mer informasjon om hvilke antibegroingsmidler som er i bruk på den enkelte lokaliteten og praksis i forhold til spyling på de enkelte anlegg være avgjørende for å øke kunnskapsstyrken om utslippsmengde i de enkelte produksjonsområdene og for å kunne gi gode råd til forvaltningsmyndigheter.
Modellert ekstrabelastning av kobber i kystsonen fra utblødning av kobberimpregnerte nøter gjør at en bør tilstrebe å redusere denne påvirkningen så mye som råd. Ekstra pulser av kobberkonsentrasjoner på grunn av spyling gir pulser kan gi konsentrasjoner som overstiger miljøkvalitetsstandard for sediment og kystvann. Vi mangler data for å gjøre modellsimuleringer av disse pulsene, men det høye forbruket av kobber i norsk oppdrettsnæring kombinert med kunnskap om passiv utlekking og ekstra utslipp som pulser dersom nøtene blir spylt gjør at Havforskningsinstitutt anbefaler at det blir iverksatt tiltak for å redusere denne påvirkningen.
I samtale med produsenter for antibegroingsmidler blir det opplyst at det ikke blir anbefalt å høytrykkspyle nøter som er impregnert med kobber. Dette bør kommuniseres klart i produktdatabladene.
For å nå målet om at utslipp fra oppdrett ikke skal påvirke miljøet rundt et oppdrettsanlegg negativt anbefaler vi at det blir satt i gang arbeid med å få til løsninger som ikke fører til spredning av forbindelser som kobber eller erstatningsstoff som er giftige og som kan bli akkumulert i sediment eller organismer. Alternativer kan være å unngå kobber eller andre biocider. Siden kobber fra fôrspill og fekalier også er en kilde til kobberutslipp fra oppdrettsanlegg, selv om dette utslippet ikke antas å kunne forklare de forhøyede verdiene av kobber nært oppdrettsanlegg, bør en gå gjennom hvor stor tilsats av kobber til fôret som er nødvendig og undersøke om kobber som blir tilsatt fôret også er i en form som gir godt opptak i laksen.
9 - Forkortelser
EC50 – Den konsentrasjonen som gir en effekt i 50 % av testorganismene.
HC5 – Den konsentrasjonen som påvirker 5 % av artene. Gjerne brukt i arts sensitivitets fordelinger (Species Sensitivity Distributions (SSD))
HC50 - Den konsentrasjonen som påvirker 50 % av artene
LC50 – Den konsentrasjonen som gir 50 % dødelighet
PNEC – Predicted No Effect Concentration – Den konsentrasjonen fra laboratorieforsøk som ikke gir signifikante endringer i forhold til kontrollgruppen
10 - Referanser
Adam N, Schmitt C, De Buyn L, Knapen D, Blust R. 2015. Aquatic cute species sensitivity distributions of ZnO and CuO nanoparticles. Science of the Total Environment 526: 233-242.
Albretsen, Jon, Ann Kristin Sperrevik, André Staalstrøm, Anne D. Sandvik, Frode Vikebø, and Lars Asplin. “NorKyst-800 Report No. 1: User Manual and Technical Descriptions.” Fisken Og Havet. Institute of Marine Research, 2011.
Balls PW. 1985. Trace metals in the Northern North Sea. Marine Pollution Bulletin. 16: 203:207.
Brooks SJ, Farmen E, Heierc LS, Blanco-Rayond E, Izagirre U. 2015. Differences in copper bioaccumulation and biological responses in three Mytilus species. Aquatic Toxicology 160: 1–12.
Brooks KM, Mahnken CVW (2003). Interactions of Atlantic salmon in the Pacific northwest environment III. Accumulation of zink and copper. Fisheries Research 62, 295-305.
Carvajalino-Fernández, M.A., P.N. Sævik, I.A. Johnsen, J. Albretsen, and N.B. Keeley. 2020. Simulating Particle Organic Matter Dispersal beneath Atlantic Salmon Fish Farms Using Different Resuspension Approaches. Marine Pollution Bulletin 161: 111685. https://doi.org/10.1016/j.marpolbul.2020.111685 .
Dalsøren, Stig B., Jon Albretsen, and Lars Asplin. “New Validation Method for Hydrodynamic Fjord Models Applied in the Hardangerfjord, Norway.” Estuarine, Coastal and Shelf Science 246 (November 5, 2020): 107028. https://doi.org/10.1016/j.ecss.2020.107028.
Deruytter D, Baert JM, Nevejean N, De Schamphelaere KAC and Janssen CR. 2017. Mixture toxicity in the marine environment: Model development and evidence for synergism at environmental concentrations. Environmental Toxicology and Chemistry, Vol. 36: 3471–3479.
Duinker A, Storesund J, Lunestad BT, Sanden M. 2022. Nasjonalt tilsynsprogram fro produksjon av skjell og andre bløtdyr- Prøver analysert I 2021 for kjemisk forurensende stoffer og mikroorganismer. Rapport fra Havforskningen Nr 2022-31. 22 s.
ECHA. 2014. Biocidal Products Commitee (BPC). Opinion on the application for approval on the active substance: Tralopyril. Product type: 21-ECHA/BPC/002(2014.
Ellis MS, Barber RD, Hillman RE, Kim Y and Powell Haskin EN. 1998. Histopathology analysis. NOAA Tech. Memo. NOS ORCA 130: 198-215.
EU. 2002. Directive 2002/32/EC of the European Parliament and of the Council of 7 May 2002 on undesirable substances in animal feed - Council statement. https://eur-lex.europa.eu/eli/dir/2002/32/oj/eng
EU. 2021. Risk assessment of a biocidal product (Family) for national authorisation applications. Aquanet 360 Biocidal Product Family: Product names; AquaNet LG 360, Aquanet HG360. AquaNet 360. Product type(s) 21 dicopperoxide and copper thiocyanate.Evaluating Competent Authority: Norway. Date 12th March 2021. Pp 383.
European Copper Institute. 2005. EU Risk Assessment - [Copper, Copper II sulphate pentahydrate, copper(I)oxide, copper(II)oxid, dicopper chloride trihydroxide] CAS [7440-50-8, 7758-98-7, 1317-39-1, 1317–38–0, 1332-65-6. PNEC derivation for copper in marine waters. ( https://echa.europa.eu/copper-voluntary-risk-assessment-reports).
Grefsrud ES, Andersen LB, Bjørn PA, Grøsvik BE, Hansen PK, Husa V, Karlsen Ø, Kvamme BO, Samuelsen O, Sandlund N, Stien LH, Solberg MF og Stien LH. 2022 a. Risikorapport norsk fiskeoppdrett 2022 – Risikovurdering. Fisken og Havet. ISSN: 1893-4536. Pp 235. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-2022-12
Grefsrud ES, Bjørn PA, Grøsvik BE, Hansen PK, Husa V, Karlsen Ø, Kvamme BO, Samuelsen O, Sandlund N, Stien LH, Solberg MF og Stien LH. 2022 b. Risikorapport norsk fiskeoppdrett 2022 – Kunnskapsstatus. Fisken og Havet. ISSN: 1893-4536. Pp 406. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-2022-13
Hall LW, Anderson RD. 1999. A Deterministic Ecological Risk Assessment for Copper in European Saltwater Environments. Marine Pollution Bulletin. 38(3): 207-281.
Heuschele J, Lode, T, Konestabo HS, Titelman J, Andersen T. 2022. Drivers of copper sensitivity in copepods: A meta-analysis of LC50s. Ecotoxicology and Environmental Safety 242, 113907. https://doi.org/10.1016/j.ecoenv.2022.113907
King CK, Riddle MJ (2001). Effects of metal contaminants on the development of the common Antarctic sea urchin Sterechinus neumayeri and comparisons of sensitivity with tropical and temperate echinoids. Marine Ecology Progress Series 215, 143-154.
Martin M, Osborn KE, Billig P, Glickstein N 1981. Toxicities of ten metals to Crassotrea gigas and Mytilus edulis embrys and Cancer magister larvae. Marine Pollution Bulletin 12, 305-308.
Martins SE, Fillmann G, Lillicrap A & Thomas KV. 2018. Review: ecotoxicity of organic and organo-metallic antifouling co-biocides and implications for environmental hazard and risk assessments in aquatic ecosystems, Biofouling, 34:1, 34-52, DOI: 10.1080/08927014.2017.1404036
Miljødirektoratet 2014. Kvalitetssikring av miljøkvalitetsstandarder. M-241. 197 s.
Miljødirektoratet 2016. Grenseverdier for klassifisering av vann, sediment og biota. M-608. 24 s.
Miljødirektoratet 2021. Proposed Environmental Quality Standards (EQSs) for blue mussel (Mytilus edulis). M-1939. 61 s.
Müller, Malte, Mariken Homleid, Karl-Ivar Ivarsson, Morten A. Ø Køltzow, Magnus Lindskog, Knut Helge Midtbø, Ulf Andrae, et al. “AROME-MetCoOp: A Nordic Convective-Scale Operational Weather Prediction Model.” Weather and Forecasting 32, no. 2 (April 1, 2017): 609–27. https://doi.org/10.1175/WAF-D-16-0099.1 .
Nadella SR, Fitzpatrick JL, Franklin N, Bucking C, Smith S, Wood CM. 2009. Toxicity of dissolved Cu, Zn, Ni and Cd to developing embryos of the blue mussel (Mytilus trossolus) and the protective effect of dissolved organic carbon. Comparative Biochemistry and Physiology, Part C 149: 340–348.
Novikov MA & Draganov DM. 2017. Spatial distribution of water mass pollution characteristics in the Barents Sea. Water quality and Protection: Environmental Aspects. 44: 768-772. DOI: 10.1134/S0097807817050086
OSPAR (2006). Agreement on Background Concentrations for Contaminants in Seawater, Biota and Sediment. OSPAR Agreement 2005-6 Revised by ASMO 2006 (ASMO 2006 Summary Record ASMO 06/12/01).
Sadiq M. Toxic metal chemistry in marine environments. Environmental Science and pollution; 1. 1992. Marcel Dekker Inc. New York. ISBN 0-8247-8647-5. Pp 390.
Sele V, Lundebye A-K, Berntssen M, Storesund J, Lie KK, Philip A, Nøstbakken OJ, Waagbø R, og Ørnsrud R. 2022. Program for overvåking av fiskefôr. Årsrapport for prøver innsamlet i 2021. Rapport fra Havforskningen Nr 22-2022. ISSN:1893-4536. pp 28.
Shchepetkin, Alexander F., and James C. McWilliams. “The Regional Oceanic Modeling System (ROMS): A Split-Explicit, Free-Surface, Topography-Following-Coordinate Oceanic Model.” Ocean Modelling 9, no. 4 (January 2005): 347–404. https://doi.org/10.1016/j.ocemod.2004.08.002 .
Skarbøvik E, Allan I, Sample JE, Greipsland I, Selvik JR, Schanke LB, Beldring S, Stålnacke P, Kaste Ø. 2017. Riverine inputs and direct discharges to Norwegian coastal waters–2016. NIVA report M ‐ 862–2017. Norwegian Institute for Water Research, Oslo, Norway.
Ytrestøyl T, Aas TS, Åsgard T (2015). Utilisation of feed resources in production of Atlantic salmon (Salmo salar) in Norway. Aquaculture 448, 365-374.
11 - Vedlegg 1. Report on the histopathology of blue mussel (Mytilus spp.) from copper polluted environment.
Materials and methods
Sampling, tissue processing and HES staining were performed according to the protocol described in the quality portal of the Institute of Marine Research (IMR) (MET.HIS.MET-11).
Sampling
Blue mussel samples were collected from copper treated net pen in Hardangerfjorden (n=15) and from a testing at Drøbak (n=10). Samples from Hardangerfjorden were dissected onboard whereas live blue mussel samples collected from Drøbak were transported to IMR for dissection. The samples from Drøbak were cultured in rafts impregnated with different concentrations of Cu2O; the samples contained 8 groups with each group containing 10 individuals (n=10) (Table 1). Dorsoventral section of the blue mussel that included the hepatopancreas, gills, mantle, muscle, and gonads were carefully incised and immediately fixed in seawater Davidson’s fixative containing 5% glacial acetic acid for 48 hrs.
Tissue processing
The samples were embedded in paraffin following dehydration in a progressive series of ethanol, cleared in xylene, and infiltrated in molten paraffin. Specimens of 3 µm thick sections were cut using a Rotary Microtome HM355S (Thermo Fisher Scientific) and stained with hematoxylin erythrosine saffron (HES) for routine histopathological assessment. Only samples with LIMS nr. 2021-379/390/384/385 & 386 were processed for histological analysis. One sample was stained with Alcian blue (AB) Periodic acid Schiff (PAS) to distinguish between acidic and neutral mucins and secretary cells of the gills.
Copper staining
Mallory hematein (SIGMA-ALDRICH, Product number: 51230-10G) and 5-(4-Dimethylaminobenzylidene)-rhodanine (MERCK, Product number 39090-10G) were used as special stains to demonstrate copper deposits in tissue sections. The staining protocol for Mallory hematein and 5-(4-Dimethylaminobenzylidene)-rhodanine were adopted from the US National Oceanographic and Atmospheric Administration (NOAA) Technical Memorandum NOS NCCOS 5 (Howard, 2004) and Abcam (Copper Stain Kit, ab150666) using the microwave method, respectively.
Mallory hematein : 0.5% Hematein stock solution was first prepared by dissolving in distilled water in dark room; then, hematein working solution was prepared by mixing 10-parts 0.5% hematein stock solution and 1-part dimethyl sulfoxide (DMSO) (Sigma-Aldrich, product number: D8418) prior to use. Tissue sections were deparaffinized, hydrated in distilled water and stained with hematein for 5 min. To turn positive reactions blue-black, sections were dipped 3X in ammonia water, rinsed in distilled water for 1 min. and counterstained in safranin for 2 min. The slides were washed in running tap water, de-stained in two changes of 100% ethanol for 1 min. each and cleared in two changes of xylene prior to mounting in histokitt (Howard, 2004).
Rhodanine : Rhodanine stock solution was prepared by dissolving 2 g rhodanine powder in 1000 ml absolute ethanol and kept at 4°C. For working solution, 4ml rhodanine stock solution was filtered and added into 46 ml acetate buffer (pH 8). The tissue sections were deparaffinized and hydrated in distilled water; the slides were, then, placed in prewarmed rhodanine working solution in a capped coplin jar. The coplin jar containing rhodanine working solution and tissue sections was loosely capped and heated in a microwave oven at full power until the solution was hot. The solution was cooled at room temperature; then the slides were checked for copper deposits by a microscope. Desired staining reaction was obtained by repeating the cooling and heating cycles. Excess stain was removed by rinsing the slides in two changes of acetate buffer for 1 min. Then, the specimens were stained in instant hematoxylin solution for 1 min, rinsed in three changes of acetate buffer solution for 1 min. each, dehydrated in three changes of 100% ethanol for 1 min. each and cleared in two changes of xylene prior to mounting in histokitt (Ahmad, Suhaimi et al. 2018).
NB. positive staining reactions were not confirmed with a positive test slide as known positive materials were lacking.
Analysis
Stained specimens were scanned at 40X objective using NanoZoomer S60 Digital slide scanner C13210-01 (Hamamatsu Phonotics K. K) and analyzed for histopathology by NDP.view2 software (Hamamatsu Phonotics K. K) and light microscopy (Leica DMRE).
The digestive gland (DG) and gills of all samples were analyzed for the presence of pathological changes, but semi-quantitative analysis with severity grades was not performed as there was lack of standardized histopathological scoring system for bivalves including mussels. Scoring of atrophied tubules for the DG was adopted from the method described by Ellis, B. et al (1998) (Table 2). The prevalence was given in percentage for the whole sample group. In addition, the kidney, mantle (& gills) and stomach were observed for the presence of lipofuscin aggregates, brown pigmentation, and debris, respectively.
Table 2. Histopathological scoring of atrophied tubules in the DG (Ellis et al., 1998).
Score
Description
0
Most tubules have normal wall thickness with few slightly atrophied
1
Less than one - half of wall thickness undergo atrophy with some normal tubules
2
Wall thickness averaging about one-half as thick as normal
3
More than one-half wall thickness atrophied, significant atrophy with extreme thinness
4
Extremely thin walls, almost all tubules atrophied
Results and discussion
Histopathology
The prevalence of atrophied tubules of the DG is presented in Table 2. The most obvious observations in most of the mussels from Drøbak were moderate to severe degeneration of the DG, atrophy of the digestive diverticula (DD), hemocyte infiltration in the vesicular connective tissue (VCT) surrounding the DD and high production of lysosomes in the DD (Figure 1). Over 60% of the samples (n=43) from Drøbak had moderate to severe atrophy of the digestive tubules (Table 4). There was no difference between groups.
Similarly, the DG of the mussels from Hardangerfjorden was also affected by degeneration and atrophy but with less severity and lower prevalence compared to the mussels from Drøbak. However, there was no extreme production of lysosomes in the hepatopancreas of the mussels from Hardangerfjorden. The lysosomes appeared to accumulate in the vesicles of the digestive cell and released into the lumen of the DD by breaking up the cell membrane (Figure 1E). Then, the basal membrane and pyramidal basophilic cells of the DD were completely dissolved leading to autolysis of the whole structure. It is not clear if the pathological changes in the DG of the mussels from Drøbak was associated with Cu – toxicity; no copper deposits were detected by hematein staining in both the gills and DG. Therefore, further investigation is required to understand the pathological processes, especially with respect to the lysosomal production. Lysosomal reactions to contaminant stressors are often used as markers for cell injury in different organisms including mollusks. Pollutants such as heavy metal ions can accumulate in the lysosomes causing physiological malfunction and destruction of the lysosomal membrane including DNA damage, lysosomal swelling, lipidosis and lipofuscinosis. The pollutants can trigger oxidative attack on protein machinery and cell constituents producing protein carbonyls and lipofuscin, which aggregate in the lysosomes. The generation of oxidative oxygen stress (ROS) aggravates oxidative damage, and the binding of lipofuscin with lysosomal hydrolases can limit the degradation of protein leading to failed autophagy (Moore et al., 2006).
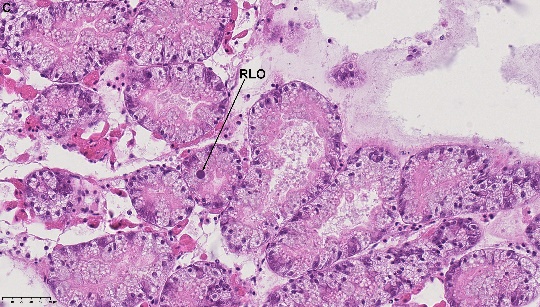
Significant number of mussels from both Drøbak – and Hardangerfjorden had brownish food debris in the gut; the food reflected producing silvery illumination when examined under darkfield polarized interference microscope (Leica DMRE) (Figure 2a &2b). It is not known if this food debris is indigestible material containing metals. Moreover, single rickettsia – like organism (RLO) was detected in the DG of two samples from Hardangerfjorden (Figure 2C).
Relatively, majority of the samples from Hardangerfjorden – and Drøbak had generally intact gills with minimal to mild hemocyte infiltration, loss of cilia and enlargement of subepithelial space (Figure 3). However, part of the gill structure in ca. 20% of the samples from Drøbak were moderately distorted. It was not clear if this distortion of gill structure is linked to pathology due to pollutants, infections, or processing such as fixation. The gills of the blue mussels are so fragile that they can easily be damaged during sampling, or processing. No bacteria or parasite was detected in any of the gills. Unlike in mussels from Hardangerfjorden, some of the gills of the mussels from Drøbak had high number of secretory cells and hyperplasia of epithelial cells, along the laterofrontal gill filaments (Figure 3E & 3F).
Mussels from Hardangerfjorden had deposition of brown pigmentation in the epithelial cells of the gills and accumulation of lipofuscin granules in the nephrocytes (Figure 4A & 4B). However, neither deposition of brown pigmentation in the gills nor aggregates of lipofuscin in the nephrocytes was observed in mussels from Drøbak. High lipofuscin accumulation is suggestive for oxidative stress; the pathological findings are often considered as general indices for cell injury due exposure to contaminants (Koehler et al., 2008).
Brown pigmentation was also observed in the epithelium along the margins of the mantle in the mussels from both sites (Figure 5). It is possible to observe brown pigmentation in the mantle from other wild samples. Therefore, it is not known whether the brown pigmentation of the mantles is specific to copper toxicity; further study is necessary to figure out if there is any association with copper toxicity. However, Culloty et al (2014) observed similar deposition of pigmented brown cells in the mantle and gills of Mytilus edulis in response to exposure to copper oxide (Cu2O) nanoparticles, and the pigmentation increased with elevated dosage (Hu et al., 2014). The deposition of pigmented brown cells may be attributed to buildup of granules affecting protein turnover, which impairs the function and structural integrity of the lysosomes (Lowe and Pipe 1994, Hu et al., 2014). Moreover, copper can interfere with enzyme activities and electron transport reaction while impairing membrane permeabilities and cell division (Davenport and Redpath 1984).
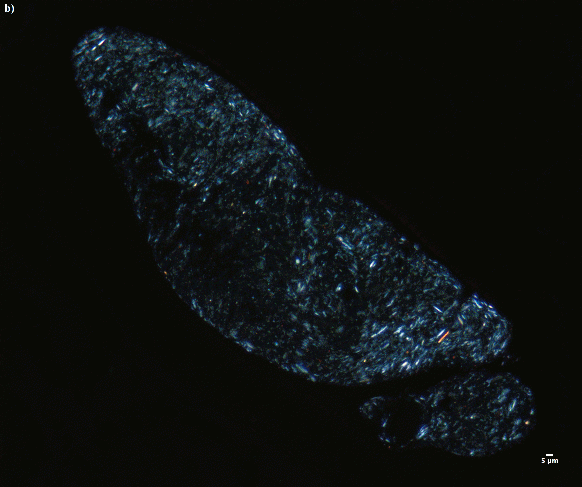
Staining with Mallory hematein exhibited the deposition of copper in the gills, as well as in the kidney, of the mussels from Hardangerfjorden (Figure 6); however, no positive reaction was evident in mussels from Drøbak. The copper deposits were blue-black in color as described in NOAA (Howard 2004). The deposits were localized in the nephrocyte lysosomes and in the cytoplasmic granular cells of postfrontal region of the gill and gill endothelial cells; however, the reaction was not confirmed with a positive test slide. Without hematein, the cytoplasmic granular and endothelial cells contained brown pigmented cells in parallel specimens stained with only safranin or HES. These brown pigmented cells might be lysosomes engaged in detoxifying excess copper (Everaarts 1990). Cajaraville et al. 1996 demonstrated the deposition and localization of copper deposits in the gills of Mytilus galloprovincialis using autometallography (Soto, Cajaraville et al. 1996).
Hematein is an oxidized form of hematoxylin; it chelates with metal ions, acting as mordants (Smith 2010). Therefore, hematein without a mordant can be used to demonstrate different minerals in tissue sections (Suvarna, Layton et al. 2018) from areas where there is heavy metal contamination. Tissues, specifically, the gills, kidney and DG including hemocytes and brown cells play vital role in the uptake, storage, and detoxification of metal pollutants such as copper from the environment (Janssen and Scholz 1979). These metals in tissue sections can act as mordants and react with hematein giving a specific color. For instance, as indicated in figure 6A & 6C, hematein copper complex gave blue-black color, which is specific for copper (Mallory and Parker Jr 1939).
Mussel samples from Hardangerfjorden and Drøbak were also stained with rhodanine using the microwave method, but without specific staining in the gills, kidney and the hepatopancreas, i.e. unlike hematein, there was no staining difference between the parallel sections stained with rhodanine and hemoxylin only. Mallory hematein gave consistent satisfactory result compared to rhodanine. However, some gland cells in the foot of mussels from Hardangerfjorden stained golden brown with rhodanine (Figure 7A). It is not known if this golden-brown color is specific to presence of copper in the mussel foot. No such staining was observed in mussels from Drøbak. Rhodanine is one of the most widely used special stains for determination of copper in humane patients with Wilson’s disease (Emanuele and Goodman 1998), a rare genetic disorder in which copper abnormally accumulates in the liver (Mak and Lam 2008). Rhodanine chelates with copper coupling proteins and results in light brown to reddish color in tissue sections stained with rhodanine (Emanuele and Goodman 1998, Kim, Zhang et al. 2020). No scientific research was possibly available explaining such copper coupling proteins are present in Mytilus edulis .
Recommendation
The results from this laboratory work throw light on how to proceed with further study regarding copper toxicity in blue mussels. However, it was not possible to deduce that the histopathological findings in this work is solely attributed to copper toxicity. Therefore, more work involving controlled research trial is required to make a possible inference.
Table 3. Histopathological scoring for atrophied tubules of the DG in mussels from Hardangerfjorden (n=15).
Sample ID
normal
Minimal
Mild
Moderate
Severe
0
1
2
3
4
BM1
4
BM2
0
BM3
1
BM4
2
BM5
BM6
4
BM7
1
BM8
1
BM9
0
BM10
4
BM11
3
BM12
4
BM13
3
BM14
4
BM15
0
Prevalence (%)
27
20
7
13
33
Table 4. Histopathological scoring for atrophied tubules of the DG in mussels from Drøbak.
Normal
Minimum
Mild
Moderate
Severe
Sample ID
0
1
2
3
4
Lims nr. 2021_379/1
4
Lims nr. 2021_379/2
3
Lims nr. 2021_379/3
2
Lims nr. 2021_379/4
4
Lims nr. 2021_379/5
2
Lims nr. 2021_379/6
4
Lims nr. 2021_379/7
2
Lims nr. 2021_379/8
2
Lims nr. 2021_379/9
4
Lims nr. 2021_379/10
2
Prevalence (%)
50
10
40
Normal
Minimum
Mild
Moderate
Severe
Sample ID
0
1
2
3
4
Lims nr. 2021_380/1
4
Lims nr. 2021_380/2
3
Lims nr. 2021_380/3
4
Lims nr. 2021_380/4
3
Lims nr. 2021_380/5
4
Lims nr. 2021_380/6
3
Lims nr. 2021_380/7
3
Lims nr. 2021_380/8
3
Lims nr. 2021_380/9
3
Lims nr. 2021_380/10
2
Prevalence (%)
10
60
40
Normal
Minimum
Mild
Moderate
Severe
Sample ID
0
1
2
3
4
Lims nr. 2021_384/1
1
Lims nr. 2021_384/2
2
Lims nr. 2021_384/3
3
Lims nr. 2021_384/4
2
Lims nr. 2021_384/5
3
Lims nr. 2021_384/6
1
Lims nr. 2021_384/7
3
Lims nr. 2021_384/8
1
Lims nr. 2021_384/9
3
Lims nr. 2021_384/10
4
Prevalence (%)
30
20
40
10
Continues to the next page.
Normal
Minimum
Mild
Moderate
Severe
Sample ID
0
1
2
3
4
Lims nr. 2021_385/3
4
Lims nr. 2021_385/6
4
Lims nr. 2021_385/9
1
Prevalence (%)
33
67
Normal
Minimum
Mild
Moderate
Severe
Sample ID
0
1
2
3
4
Lims nr. 2021_386/1
3
Lims nr. 2021_386/2
3
Lims nr. 2021_386/3
3
Lims nr. 2021_386/4
4
Lims nr. 2021_386/5
4
Lims nr. 2021_386/6
4
Lims nr. 2021_386/7
1
Lims nr. 2021_386/8
1
Lims nr. 2021_386/9
2
Lims nr. 2021_386/10
4
Prevalence (%)
20
10
30
40
Normal
Minimum
Mild
Moderate
Severe
0
1
2
3
4
Total prevalence (%, n=43)
0
14
21
33
33
Figure 1. A) Normal DG; VCT: Vesicular connective tissue; DD: Digestive diverticula, B) Atrophied DD. HES stained. NDP view 2, 40X.
Figure 1. C) Severely necrotized DG with distorted tissue architecture, 40X, D) Severe inflammation in the VCT of the digestive gland (black arrow), 10X. HES stained. NDP view2.
Figure 1. E) Normal tubules filled with lysosomes in the digestive cells; basophilic cells (yellow arrow); digestive cell (black arrow); the lysosomes appear to be released into the lumen in some tubules (Asterix), F) Autolysis of the entire structure of DD. Note the disappearance of the basal membrane and the basophilic cells. HES stained. NDP view 2, 40X.
Figure 2. a) Debris in the gut, b) The same specimen as in figure a) examined under darkfield polarized interference microscope (Leica), C) RLO in the digestive tubule. NDP view 2, 40x (HAMATSU Photonics)
Figure 3 . A) Normal gill. B) Enlargement of subepithelial space (*) with flow of hemocytes in the branchial space (black Asterix). NDP view 2, 40x (HAMATSU Photonics)
Figure 3. C) Detachment of the gill epithelium from the chitinous rod (black Asterix). D) Loss of cilia (black arrow). HES stained. NDP view 2, 40x (HAMATSU Photonics)
Figure 3. E) High number of secretory cells (black arrow) and hyperplasia of the epithelial cells, HES stained, F) Same specimen as in figure 3E stained with AB – PAS. The secretary cells are slightly PAS positive (black arrows), while the acidic mucins are AB positive (orange arrows). NDP view 2, 40X (HAMAMATSU Photonics).
Figure 4. Gill with brown pigmentation (black arrows), 40X (Leica DRME), B) Lipofuscin granules (Black Asterix) in the nephrocytes. HES stained. NDP view 2, 40X (HAMAMATSU Photonics).
Figure 5. a) Brown pigmentation (black arrow) in the epithelium along the margin of the mantle. 40x magnification, HES stained. NDP.view 2 (HAMAMATSU Photonics).
Figure 6. A) Blue – black precipitates (black arrow) in the gills stained with Mallory hematein for copper, B) Parallel sections of figure 6A stained with only safranin showing brown pigmented cells, where the metals are deposited, Leica DMRE, 60X.
Figure 6. C) Blue – black precipitates in the kidney stained with Mallory hematein for copper, D) parallel section of figure 6C stained with only safranin showing brown pigmented cells (Asterix)., Leica DMRE, 40x.
Figure 7. Three parallel sections of mussel foot from Hardangerfjorden stained with: A) rhodanine using the microwave method (golden brown color), B) hematoxylin and erythrosine, C) hematoxylin only. NDP view 2, 25x (HAMAMATSU Photonics).
References
Ahmad, M., et al. (2018). "Ternary copper (II) complex: NCI60 screening, toxicity studies, and evaluation of efficacy in xenograft models of nasopharyngeal carcinoma." PloS one13 (1): e0191295.
Davenport, J. and K. Redpath (1984). Copper and the Mussel Mytilus edulis. Toxins, Drugs, and Pollutants in Marine Animals , Springer : 176-189.
Ellis, M., et al. (1998). "Histopathology analysis." NOAA Tech. Memo. NOS ORCA130 : 198-215.
Emanuele, P. and Z. D. Goodman (1998). "A simple and rapid stain for copper in liver tissue." Annals of diagnostic pathology2 (2): 125-126.
Everaarts, J. (1990). "Uptake and release of cadmium in various organs of the common mussel, Mytilus edulis (L.)." Bulletin of environmental contamination and toxicology45 (4): 560-567.
Howard, D. W. (2004). Histological techniques for marine bivalve mollusks and crustaceans , NOAA, National Ocean Service, National Centers for Coastal Ocean Service ….
Hu, W., et al. (2014). "Toxicity of copper oxide nanoparticles in the blue mussel, Mytilus edulis: a redox proteomic investigation." Chemosphere108 : 289-299.
Janssen, H. and N. Scholz (1979). "Uptake and cellular distribution of cadmium in Mytilus edulis." Marine biology55 (2): 133-141.
Kim, P., et al. (2020). "Accurate measurement of copper overload in an experimental model of Wilson disease by laser ablation inductively coupled plasma mass spectrometry." Biomedicines8 (9): 356.
Koehler, A., et al. (2008). "Effects of nanoparticles in Mytilus edulis gills and hepatopancreas–a new threat to marine life?" Marine environmental research66 (1): 12-14.
Lowe, D. and R. Pipe (1994). "Contaminant induced lysosomal membrane damage in marine mussel digestive cells: an in vitro study." Aquatic toxicology30 (4): 357-365.
Mak, C. M. and C.-W. Lam (2008). "Diagnosis of Wilson's disease: a comprehensive review." Critical reviews in clinical laboratory sciences45 (3): 263-290.
Mallory, F. B. and F. Parker Jr (1939). "Fixing and staining methods for lead and copper in tissues." The American journal of pathology15 (5): 517.
Moore, M. N., et al. (2006). "Lysosomal and autophagic reactions as predictive indicators of environmental impact in aquatic animals." Autophagy2 (3): 217-220.
Smith, A. (2010). "Hematein chelates of unusual metal ions for tinctorial histochemistry." Biotechnic & Histochemistry85 (1): 43-54.
Soto, M., et al. (1996). "Tissue and cell distribution of copper, zinc and cadmium in the mussel, Mytilus galloprovincialis, determined by autometallography." Tissue and Cell28 (5): 557-568.
Suvarna, K. S., et al. (2018). Bancroft's theory and practice of histological techniques E-Book , Elsevier Health Sciences.