I denne risikovurderingen vurderer vi risiko for «Negative effekter på kysttorskbestander som følge av lakseoppdrett i åpne merder i sjø». Vi konkluderer med at risikoen er lav for området 67°N til og med Vest-Finnmark og moderat for området Jæren til 67°N. Vi påpeker at vurderingene er gjort på de samlede kysttorskbestandene og ikke nødvendigvis er gjeldene i hele de geografiske områdene som omfattes. Dette som følge av at lokale kysttorskbestander vil kunne være i bedre eller verre forfatning enn gjennomsnittet og at oppdrettsintensiteten vil variere.
Det mangler betydelig kunnskap om hvilket omfang eventuelle negative effekter av lakseoppdrett i åpne merder i sjø kan ha på kysttorskbestandene. Å øke kunnskapen om hvilke konsekvenser beiting på spillfôr og byttedyr forbundet med lakseoppdrett har på torskens reproduksjon, vurderes som særlig viktig. Dette tatt i betraktning, er det nødvendig å vite mer om hvorvidt kysttorsk tiltrekkes oppdrettsanlegg i en større sammenheng. Det vil si om tiltrekning varierer mellom geografiske områder og livsstadier. Identifisering av kunnskapshull anses som et viktig resultat av risikovurderingen, ettersom mindre usikkerhet knyttet til alvorlighetsgraden av endring i adferd og fyisologi, vil kunne endre risikobildet betraktelig.
Vurderingen er gjort for området Jæren til 67°N og for området 67°N til og med Vest-Finnmark. Det vil si at risikovurdering ikke omfatter området fra Svenskegrensen til Jæren eller Øst-Finnmark. Disse områdene er ekskludert ettersom produksjonen av laksefisk er betraktelig lavere her enn i de resterende delene av landet. Risikovurderingen som er gjort for området Jæren til 67°N omfatter forvaltningsområdet for kysttorsk fra 62°N – 67°N og forvaltningsområdet sør for 62°N. Dette gjøres fordi kysttorskbestandene ansees som svake i begge områdene, selv om tilstanden vurderes som spesielt svak i området sør for 62°N. Risikovurderingen som er gjort fra 67°N til og med Vest-Finnmark, omfatter til sammenligning kun kysttorsk i forvaltningsområdet nord for 67°N. Nord for 67°N har gytebiomassen økt de siste årene, og tilstanden til den samlede kysttorskbestanden vurderes som moderat ettersom fiskedødeligheten overskrider anbefalt nivå.
Risikovurderingen er basert på den nåværende tilstanden til kysttorsk, dagens produksjon av laksefisk i åpne merder og eksisterende kunnskap om de risikokildene og hendelsene vi anser som de viktigste for hvorvidt lakseoppdrett kan ha negative effekter på kysttorskbestandene. Om situasjonen, premissene eller kunnskapsgrunnlaget som ligger til grunn for risikovurderingen skulle endre seg i vesentlig grad, vil det være behov for en ny vurdering.
Lakseoppdrett har økt kraftig siden starten for 50 år siden og antas å utgjøre en av de største påvirkningene på det marine miljøet i kystsonen. Oppdrett av laks i åpne merder i sjø kan påvirke det lokale økosystemet, inkludert kysttorsken som lever der, først og fremst som en kilde til en rekke ulike utslipp. Tilstedeværelsen av anleggene kan føre til endringer i kysttorskens adferd og fysiologi, samt øke forekomsten av sykdom. Endringer i adferd kan forekomme ved at kysttorsk tiltrekkes eller frastøtes områder med lakseoppdrett. Eventuelle fysiologiske endringer vil kunne inntreffe som en konsekvens av beiting på spillfôr eller byttedyr som har spist spillfôr, samt ved eksponering for skadelige nivåer av fremmedstoffer. Sykdomsutbrudd blant laks i åpne merder kan eksponere kysttorsk for smitte, og gitt at den er mottakelig kan dette medføre infeksjoner og sykdomsutbrudd.
Eksisterende kunnskap viser at kysttorsken høyst sannsynlig tiltrekkes anleggene på grunn av økt mattilgang. Likevel er det stor variasjon mellom individer, og det finnes generelt lite kunnskap om hvorvidt kysttorsk oppholder seg ved oppdrettsanlegg på en større skala, samt hvorvidt den observerte tiltrekningen er representativ for ulike livsstadier eller om den varierer mellom ulike områder. Det at enkelte torsk tiltrekkes laksemerdene utelukker ikke muligheten for at andre individer kan frastøtes fra områder med lakseoppdrettsanlegg. For eksempel har fiskere observert at gytevandrende torsk har endret vandringsmønster og unngår gytefelt i oppdrettsintensive områder. Dette er ikke bekreftet av vitenskapelig studier, noe som kan skyldes at kysttorskens valg av gyteområder også påvirkes betydelig av andre faktorer. I lys av dette er det nødvendig med bedre kunnskap om i hvilken grad kysttorsk tiltrekkes og/eller frastøtes oppdrettsanlegg.
Av hendelsene som kan forårsake negative effekter på kysttorskbestandene ansees konsekvensene av de fysiologiske endringene som følge av beiting på spillfôr og byttedyr ved oppdrettsanleggene som spesielt alvorlige. Det finnes god kunnskap om at torsk som beiter på spillfôr og byttedyr forbundet med oppdrett vil få en endret fettsyresammensetning i vevet. Selv om ingenting foreløpig tyder på at en slik fysiologisk endring er negativ for torskens vekst og overlevelse, er det manglende kunnskap om hvorvidt dette påvirker reproduksjonen fordi enkelte marine fettsyrer er viktige for blant annet eggutvikling, befruktning og larveoverlevelse. Det er også manglende kunnskap om effektene av eksponering for fremmedstoffer fra spillfôr og nye impregneringsmidler.
Til tross for at kysttorsk kan samle seg rundt lakseanlegg og dermed overlappe i tid og rom med sykdomsutbrudd i laksemerdene, vurderes det som lite sannsynlig at dette vil føre til økt sykdomsforekomst blant kysttorsk og dermed bidra til risiko for negative effekter på kysttorskbestandene. Dette er fordi de aller fleste sykdomsutbruddene i lakseoppdrett er forårsaket av laksespesifikke agens som ikke vil smitte over på kysttorsk. Det er ikke gitt at det vil være de samme kjente agensene og sykdommene som vil være gjeldende i framtida. I tillegg må man også være forberedt på at nye ukjente agens eller nye varianter av kjente agens vil skape utfordringer.
Risikovurdering
Innledning
Bakgrunn
Norge eksporterte i 2025 i overkant av 1,4 millioner tonn oppdrettslaks (Salmo salar) og 86 903 tonn regnbueørret (Oncorhyncus mykiss) til en verdi på henholdsvis 124,67 og 7,42 milliarder kroner (tall fra Norges sjømatråd, januar 2026). Dette utgjorde 72,3 % av den samlede eksportverdien av sjømat på totalt 181,5 milliarder kroner. Lakseoppdrett, heretter definert som oppdrett av laks og regnbueørret, har økt kraftig siden starten for 50 år siden og antas å utgjøre en av de største påvirkningene på det marine miljøet i kystsonen.
Oppdrettsanlegg tiltrekker seg en rekke marine organismer, og da i hovedsak fisk som enten beiter på spillfôr fra anlegget, eller på andre organismer som lever nær anlegget. Langs norskekysten er det påvist at spesielt sei (Pollachius virens) samler seg rundt oppdrettsanlegg, men også andre arter, som torsk (Gadus morhua), kan forekomme i høye antall.
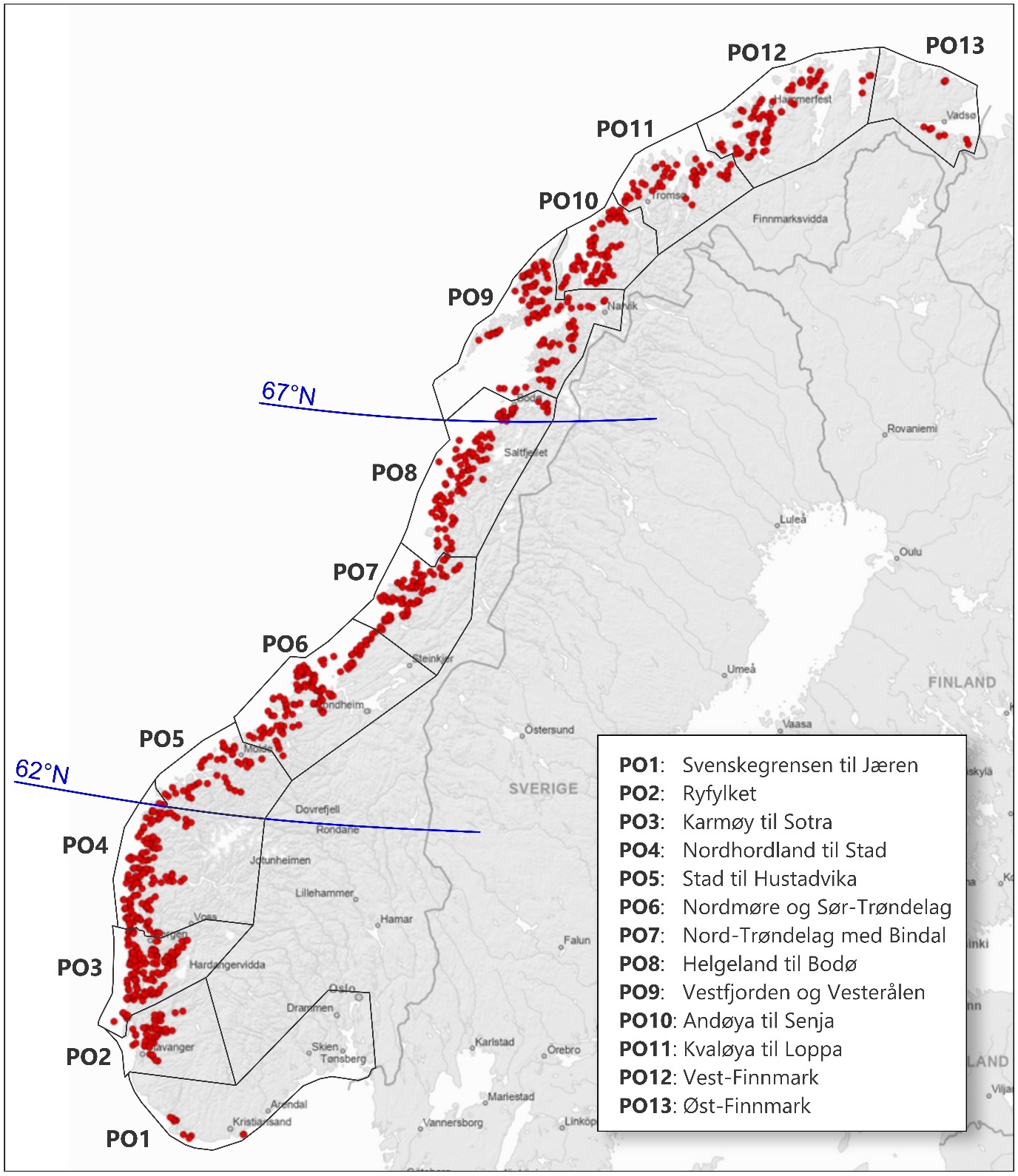
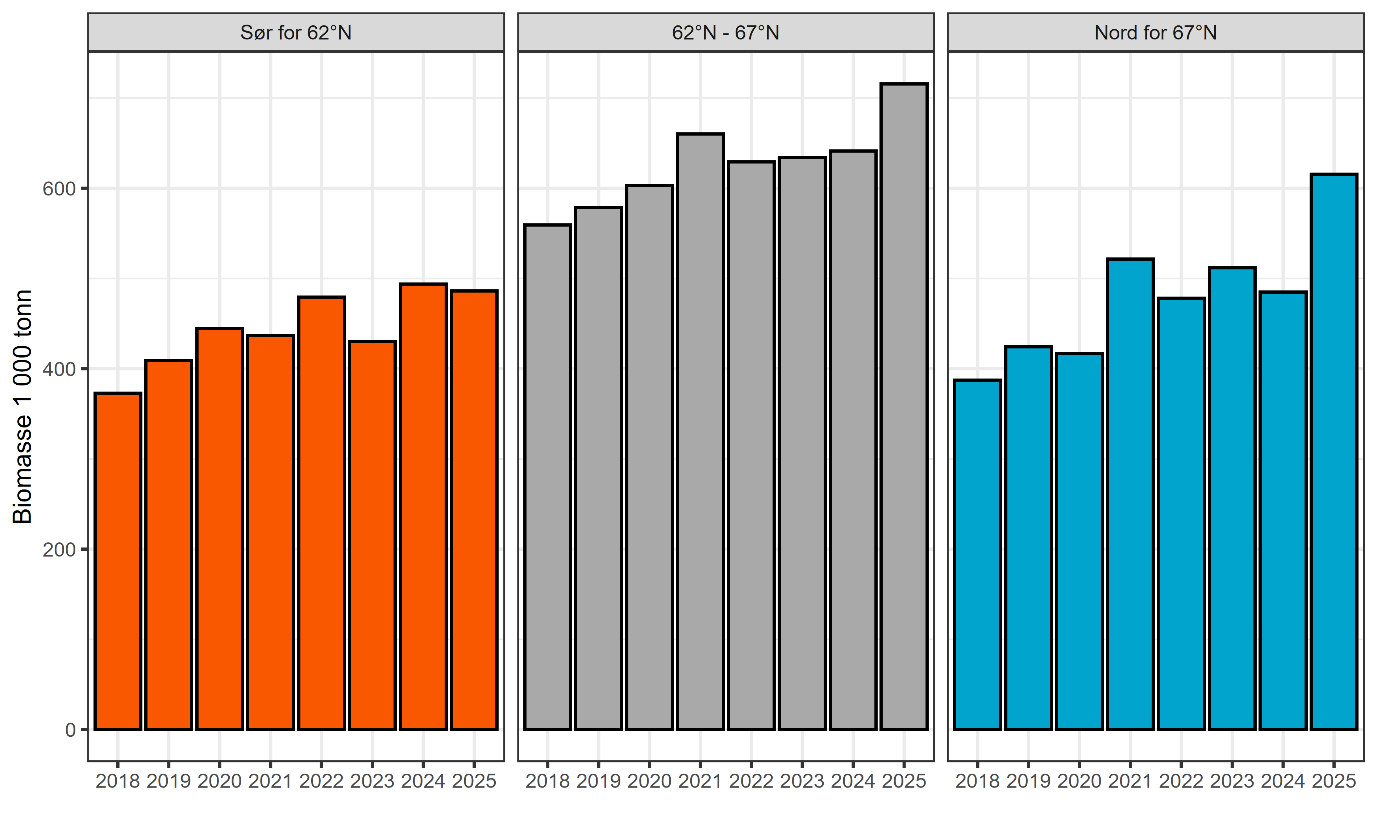
Bestandsstrukturen til torsk langs norskekysten er svært kompleks, og begrepet kysttorsk brukes ofte for å beskrive stedbundet torsk som kan skilles fra nordsjøtorsk og skrei. Frem til 2021 ble kysttorsk forvaltet i to geografiske områder; nord og sør for 62°N. Etter en metoderevisjon i 2021 ble det nordligste området delt i to ved 67°N og vi har nå tre forvaltningsområder; sør for 62°N (Skagerrak og Vestlandet), 62°N – 67°N (fra Sildegapet like sør for Stadlandet til Fugløyfjorden sørvest for Bodø) og nord for 67°N (nordlige del av Nordland inkludert Lofoten og Vesterålen samt Troms og Finnmark) (figur 1.1). En viktig bemerkning er at den samlede kysttorskbestanden i hvert forvaltningsområde består av en rekke lokale bestander. Dette kompliserer bestandsvurderingene ettersom varierende trender i rekruttering til lokale bestander kan forekomme innenfor de ulike forvaltningsområdene.
Kysttorskbestandene sør for 67°N har over tid gått kraftig ned. Det antas at fisketrykket har vært for høyt over lang tid, og at dette er en svært viktig medvirkende årsak til de observerte bestandsnedgangene, spesielt i de sørligste områdene. For å styrke bestandene av kysttorsk i Oslofjorden har det nylig blitt innført strenge tiltak fra regjeringen, og i 2020 kom Havforskningsinstituttet med forslag til forvaltningstiltak og ny gjenoppbyggingsplan for kysttorsk nord for 62°N der målet er vekst i bestanden. Nord for 67°N er den samlede bestanden i bedre forfatning, til tross for at fisketrykket overgår anbefalingene. Kysttorsk i dette området er likevel på et lavere nivå enn tidlig på 1990-tallet. Større bestander med god rekruttering, variert aldersstruktur og god genetisk status antas å være mer motstandsdyktige mot ytre påvirkninger som fiskeri, klimaendringer og oppdrett i kystsonen.
De tre forvaltningsområdene for kysttorsk sammenfaller relativt godt med den geografiske inndelingen av produksjonsområder (PO) for lakseoppdrett, hvor PO1 til PO4 (Svenskegrensen til Stad) tilsvarer forvaltningsområdet sør for 62°N, PO5 til PO8 (Stad til sør for Bodø) tilsvarer forvaltningsområdet fra 62°N – 67°N og PO9 til PO13 (Vestfjorden til Øst-Finnmark) tilsvarer forvaltningsområdet nord for 67°N (figur 1). Det ligger imidlertid 21 godkjente lokaliteter for oppdrett av laks og regnbueørret i PO8 (Helgeland til Bodø) som er nord for 67°N og tilsvarende to lokaliteter i PO4 (Nordhordland til Stad) som er i forvaltningsområdet 62°N – 67°N.
Figur 1.1. Produksjonsområder (PO) og grenser for forvaltningsområder for torsk (blå linjer). Røde sirkler viser 980 godkjente lokaliteter for oppdrett av laks og regnbueørret i sjø pr. 28. november 2025 (Kilde: Fiskeridirektoratets kart «Yggdrasil»).
Biomasse og tetthet av anlegg er to svært viktige forutsetninger for analysearbeidet. Det er betydelig variasjon i produksjon av laks og regnbueørret i åpne merder langs norskekysten. De mest oppdrettsintensive produksjonsområdene, basert på mengde fisk produsert per areal (tonn/km2), var i 2025 PO2 Ryfylke og PO3 Karmøy til Sotra, med henholdsvis 54,8 og 53,3 tonn/km2 (tabell 1.1). I andre enden av skalaen ligger PO1 Svenskegrense til Jæren og PO3 Øst-Finnmark med henholdsvis 2,4 og 3,7 tonn/km2 (tabell 1.1). Per i dag ansees produksjonen å være betydelig i alle produksjonsområder mellom Jæren og Øst-Finnmark. Likevel er det et tydelig skille i mengden som produseres per areal ved Helgelandskysten, og med unntak av PO10 (Andøya til Senja) har alle produksjonsområdene nord for Helgeland en betydeligere lavere produksjonstetthet enn landsgjennomsnittet på 24,8 tonn/km 2 (statistikk fra Fiskeridirektoratet, 20.01.2026).
Produksjon 2024
Produksjon 2025
Produksjonsområde
Areal (km2)
Mengde (tonn/år)
Mengde per areal (tonn/km2)
Mengde (tonn/år)
Mengde per areal (tonn/km2)
Område 1: Svenskegrensen til Jæren
3 864
22 823
5,9
9 133
2,4
Område 2: Ryfylke
1 977
88 705
44,9
108 402
54,8
Område 3: Karmøy til Sotra
3 558
186 714
52,5
189 749
53,3
Område 4: Nordhordland til Stad
5 584
195 380
35,0
178 869
32,0
Område 5: Stad til Hustadvika
3 950
80 602
20,4
120 356
30,5
Område 6: Nordmøre og Sør-Trøndelag
10 331
261 128
25,3
260 167
25,2
Område 7: Nord-Trøndelag med Bindal
5 182
120 370
23,2
148 855
28,7
Område 8: Helgeland til Bodø
12 766
178 817
14,0
186 231
14,6
Område 9: Vestfjorden og Vesterålen
16 115
156 704
9,7
186 544
11,6
Område 10: Andøya til Senja
4 640
114 392
24,7
159 351
34,3
Område 11: Kvaløy til Loppa
6 825
76 645
11,2
131 233
19,2
Område 12: Vest-Finnmark
10 683
125 204
11,7
124 537
11,7
Område 13: Øst-Finnmark
3 789
11 196
3,0
13 932
3,7
Tabell 1.1. Areal (sjøareal innenfor grunnlinjen, km2), produksjon (uttak slaktet fisk, tonn/år) og oppdrettsintensitet (tonn produsert fisk/km2) for hvert av de 13 produksjonsområdene i 2024 og 2025. (Statistikk fra Fiskeridirektoratet, 20.01.2026). Tallene gjelder for produksjon av atlantisk laks og regnbueørret.
Sett i lys av den dårlige tilstanden for kysttorskbestandene sør for 67°N og den høye produksjonen av laksefisk setter vi i denne analysen søkelyset på mulige negative effekter av lakseoppdrett på kysttorsk.
Mål og omfang
Målsetningen med denne analysen er å vurdere om lakseoppdrett utgjør en risiko for ytterligere svekkelse av kysttorskbestandene sør for 67°N og derved redusert mulighet for vellykket gjenoppbygging. Videre inkluderer målsetningen å vurdere risiko for svekkelse av kysttorsk bestanden nord for 67°N .
Kysttorskbestandene som inkluderes i denne analysen, er den samlede bestanden sør for 67°N og bestanden nord for 67°N. Kysttorsk sør for 67°N består av bestandene i to forvaltningsområder, 62°N – 67°N og sør for 62°N. Vi gjør denne inndeling på bakgrunn av at tilstanden til den samlede kysttorskbestanden nord for 67°N er bedre enn for de sørligere bestandene, samt at produksjonen av laks per areal samlet sett er lavere nord for 67°N. Likevel anerkjenner vi at tilstanden til kysttorsk er spesielt svakt sør for 62°N, hvor også produksjonstetteheten er høyest. For området sør for 67°N omfatter risikovurderingen ikke PO1 (Svenskegrensen til Jæren), mens for området nord for 67°N inkluderes ikke PO13 (Øst-Finnmark). Disse produksjonsområdene har svært liten produksjon av laks i åpne merder per dags dato, og det forventes en begrenset økning av produksjonen i tiden fremover. Det kan videre bemerkes at Oslofjorden med sin sårbare torskebestand, ligger i PO1, men at det ikke er fiskeoppdrett i denne fjorden. En risikovurdering her er derfor ikke relevant i denne sammenheng.
I likhet med Havforskingsinstituttets vurdering av «Negative effekter på kysttorskbestander som følge av torskeoppdrett i åpne merder i sjø», inkluderer heller ikke denne risikovurderingen nordsjøtorsk eller nordøstarktisk torsk (skrei). Dette fordi kysttorsken er mer stasjonær enn de havvandrende bestandene, noe som gjør den mer sårbar for lokal påvirkning.
Med åpne merder i sjø menes anlegg hvor det benyttes nøter for hold av fisken, og der det er fri gjennomstrømming av vann. Analysen omfatter kun negative effekter fra laksefiskoppdrett, ikke effekter av annen påvirkning på kysttorsken som for eksempel fiskeri (kommersielt fiske og turist- og fritidsfiske), klimaendringer eller andre ytre påvirkninger. Det antas at fisketrykket er den viktigste påvirkningsfaktoren for tilstanden til kysttorskbestandene, men en risikovurdering av samlet påvirkning som også inkluderer påvirkning fra akvakultur, finnes ikke per i dag. Det er derfor ikke mulig å vekte negative effekter fra akvakultur mot andre påvirkninger på kysttorsken.
Om situasjonen og premissene for årets risikovurdering skulle endre seg i vesentlig grad vil det være behov for å gjøre en ny vurdering.
Metodikk for risikovurdering
Metodikken som ligger til grunn for risikoresultatene presentert i denne rapporten, er utviklet av Havforskningsinstituttet og publisert i Andersen mfl. (2022). I dette kapittelet oppsummeres hovedpunktene i metodikken med søkelys på å forstå resultatene av risikovurderingen, hvordan de er fremkommet og hva de bygger på. For detaljer rundt grunnleggende elementer i metodikken og hvordan risikovurderingen gjennomføres, henviser vi til Andersen mfl. (2022).
I takt med forskningsfronten innen risikofaget finnes ingen objektive og korrekte risikotall som utgjør en form for fasit, det finnes kun usikkerhet knyttet til hva som ligger foran oss. Og det er nettopp denne usikkerheten fagekspertene sier noe om i denne rapporten. Målet er å skape risikoforståelse og risikoerkjennelse hos forvaltningen og andre interessenter som utgangspunkt for prioriteringer og beslutninger om veivalg og tiltak. Det er samtidig viktig at beslutningstakere og andre som skal benytte resultatene fra risikovurderingen, forstår at ufullstendig informasjon, utilstrekkelig kunnskap, hypoteser og antakelser er en del av, og i stor grad karakteriserer en slik analyse.
Vi har valgt å visualisere resultatet av risikovurderingene i form av grafiske hierarkiske strukturer (risikokart) som er ment å gi en hurtig og intuitiv forståelse for risikokilder, hendelser og mulige konsekvenser med tilhørende usikkerhet knyttet til næringens aktivitet. Det er imidlertid argumentasjonen som ligger til grunn for risikokartene, som skal skape tillit og eventuelt overbevise leseren om at risikovurderingen gir mening.
Fundamentale forhold knyttet til risikoanalysefaget som terminologi, definisjoner og metodisk tilnærming er forankret i Society for Risk Analysis, glossary (SRA, 2018); «Risk, Surprises and Black Swans – Fundamental Ideas and Concepts in Risk Assessment and Risk Management» (Aven, 2014) og NS 5814:2021 + AC:2023.
Risiko defineres i tråd med SRA (2018) som følger:
Med risiko menes konsekvenser (C) av aktiviteten med tilhørende usikkerhet (U)
Risikobegrepet introduserer dermed «konsekvenser» og «usikkerhet» (CU) som to sentrale aspekter som henger sammen. Usikkerheten knyttes til hva som kan skje, hvor ofte, samt til omfang og alvorlighetsgrad av konsekvensene. Vi kan måle denne usikkerheten ved hjelp av sannsynlighetsbetraktninger og forventningsverdier forankret i historiske data. Imidlertid vil en tilnærming der vi utelukkende bruker sannsynligheter som mål på usikkerhet, kunne føre til uheldige forenklinger og tap av viktige nyanser.
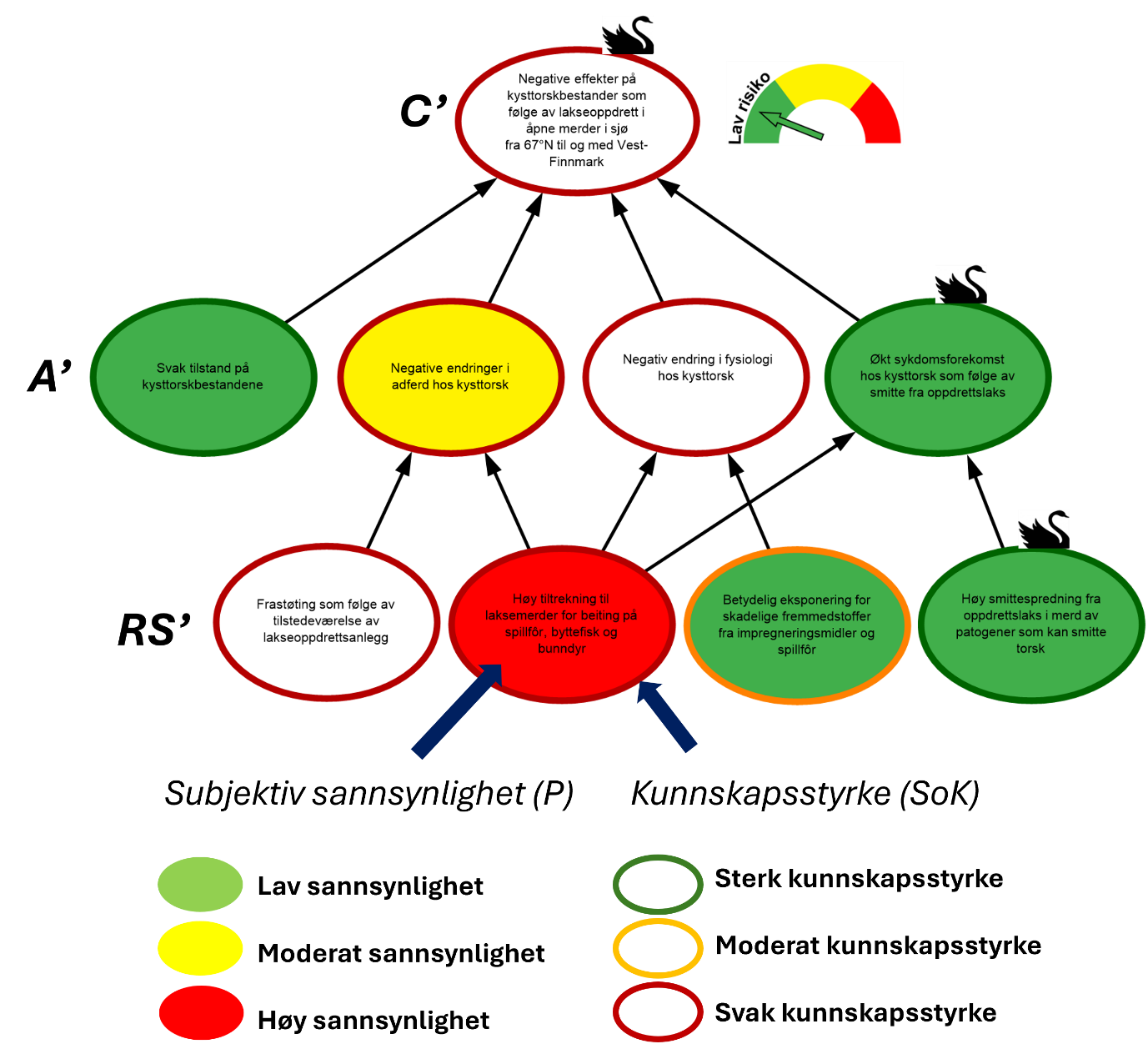
Med «usikkerhet» menes i denne risikovurderingen en kunnskapsbasert usikkerhet (epistemisk) som knyttes til det å ikke vite fremtidige konsekvenser. Risikovurderingene i denne rapporten måler og beskriver usikkerhet ved hjelp av subjektive sannsynligheter og kunnskapsstyrke knyttet til negative effekter på kysttorskbestander som følge av påvirkning fra lakseoppdrett. Vi måler og beskriver risiko ved (C’,Q,K) der C’ er spesifikke konsekvenser for kysttorskbestanden som følge av aktiviteter knyttet til lakseoppdrett, Q er et verktøy for å måle usikkerhet, og K er bakgrunnskunnskapen som vurderinger av C’ og Q bygger på. Vi velger å splitte opp de spesifikke konsekvensene C’, i risikokilder RS’, og hendelser A’ som kan føre til C’. Risiko beskrives da ved de fem variablene (RS’,A’,C’,Q,K). Vi måler usikkerheten Q ved hjelp av subjektive sannsynligheter P og vurderinger av kunnskapsstyrke SoK (Strength of Knowledge). Betraktninger om P og SoK hviler på bakgrunnskunnskapen K, og vi skriver Q (P,SoK,K ).
For denne aktuelle problemstillingen relateres usikkerheten eksempelvis til:
Status for kysttorskbestandene
Negative endringer i fysiologi og adferd hos kysttorsk
Sykdomsutbrudd i oppdrett og smittespredning til villfisk
Styrken til bakgrunnskunnskapen som danner grunnlag for vurderinger av RS’, A’, og C’ vurderes eksplisitt ved SoK, og gir innsikt i hvorvidt kunnskapen som danner grunnlaget for en bestemt konklusjon er sterk eller svak. Resultater som bygger på svak kunnskap, kan skjule deler av risikobildet gjennom eksempelvis feilaktige antakelser. Ved å inkludere en redegjørelse for kunnskapsstyrken i analysen, gis beslutningstakere mulighet til å ta stilling til hvorvidt kunnskapen på et område er tilstrekkelig som fundament for strategiske og operasjonelle veivalg. Dersom kunnskapen er svak vil det være naturlig å vurdere en risikostyringsstrategi basert på føre-var-prinsippet og diskutere tiltak som bidrar til å styrke kunnskapsgrunnlaget.
I arbeidet med denne rapporten har vi systematisert, strukturert og målt spesifikke risikokilder, hendelser og konsekvenser (RS’, A’, C’) knyttet til negative effekter på kysttorskbestander som følge av påvirkning fra lakseoppdrett i åpne merder. Det er lagt vekt på å inkludere all tilgjengelig kunnskap som utgangspunkt for å vurdere og formidle risiko. Kunnskapsgrunnlaget varierer mellom de ulike risikokildene, avhengig av om det er gjennomført mye vitenskapelige undersøkelser, om det eksisterer gode rapporteringssystemer og/eller overvåkningsprogrammer i tillegg til tidligere erfaringer i lignende situasjoner.
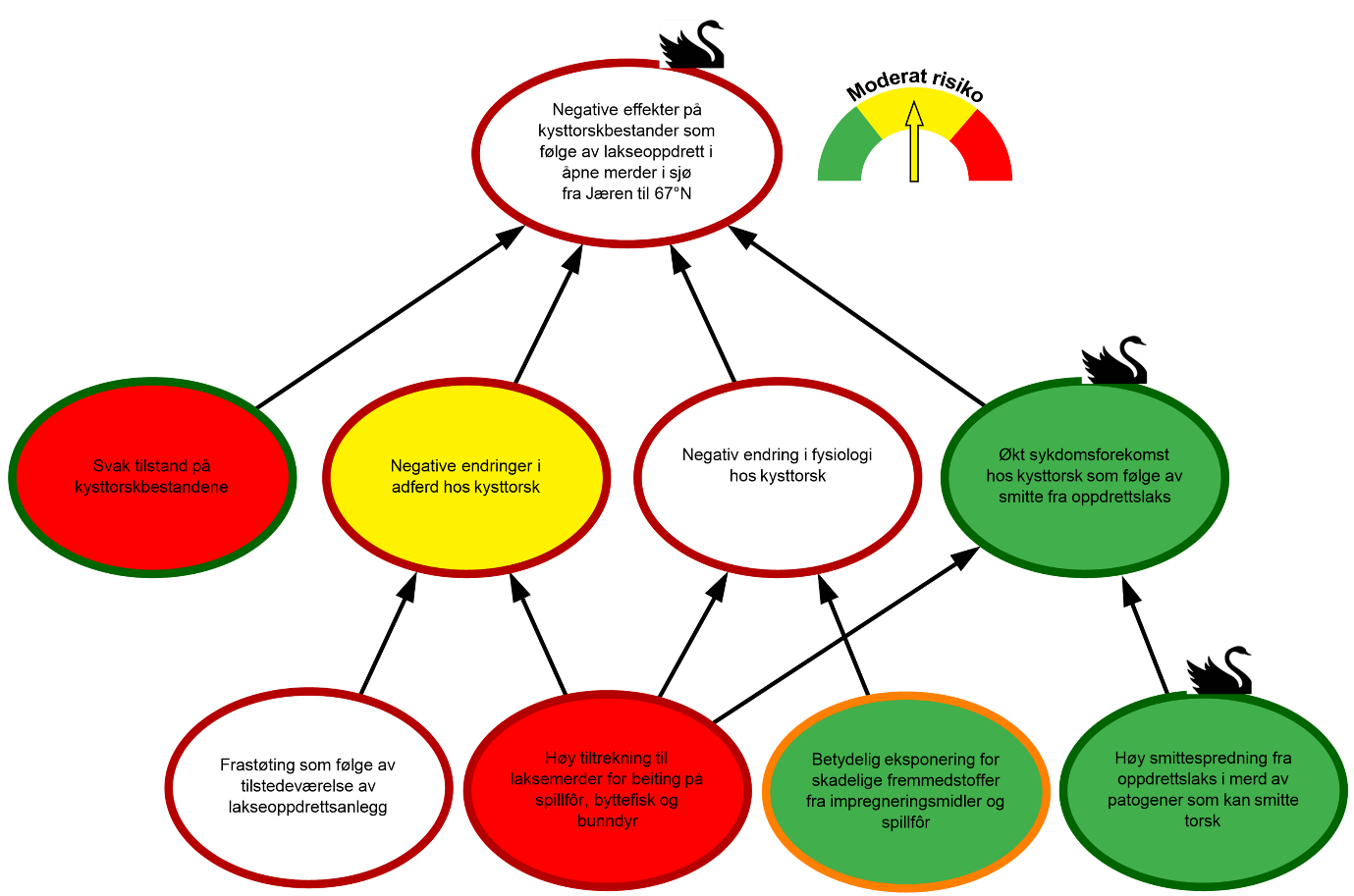
Bayesianske nettverk er valgt som et kvalitativt verktøy for å visualisere risiko, herunder; årsaks-sammenhenger mellom RS’, A’ og C’; sannsynligheter P knyttet til hvorvidt disse inntreffer, og styrken på kunnskapen (SoK) som disse vurderingene bygger på (figur 1.2). Hensikten er at de grafiske strukturene (kalt risikokart) og tilhørende argumentasjon skal bidra til hurtig og intuitiv risikoforståelse.
Figur 1.2. Eksempel på visualisering av risiko i et område. Risikokartet består av risikokilder (RS’), hendelser (A’) og konsekvenser (C’) på ulike kausale nivå med fargekoder som uttrykk for subjektiv sannsynlighet og kunnskapsstyrke.
I risikokartene benyttes fargekodene grønn, gul og rød på en node for å illustrere, lav, moderat og høy sannsynlighet. Fargen på sirkelen rundt noden illustrerer hvorvidt denne vurderingen bygger på sterk (grønn), moderat (oransje) eller svak (rød) kunnskap (SoK) (figur 1.2). Forskerne benytter all tilgjengelig kunnskap, samt refleksjoner rundt kunnskap vi eventuelt mangler, i arbeidet med å identifisere, analysere og fastsette sannsynligheter for at RS’, A’ og C’ inntreffer. Eksempelvis er sannsynligheten for «Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettslaks» vurdert som lav i figur 1.2, og grønn fargekode benyttes. Dette er basert på at sykdommene som rammer oppdrettslaks, generelt sett, i liten grad smitter over på torsk og kunnskapstyrken vurderes som sterk, noe som gir en grønn sirkel rundt noden. Denne hendelsen påvirkes av to underliggende risikokilder «Høy tiltrekning til laksemerder for beiting på spillfôr, byttefisk og bunndyr» og «Høy smittespredning fra oppdrettslaks i merd av patogener som kan smitte torsk», med henholdsvis høy og lav sannsynlighet. For å forstå hvordan hendelsen «Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettslaks» vurderes å ha lav sannsynlighet, må man lese forskernes argumentasjon. I dette tilfellet er forklaringen at de største sykdomsutfordringene hos laks i oppdrett forårsakes av agens som ikke kan smitte over på torsk. Da vi i tillegg har god oversikt over hvilke agens som forårsaker sykdom i laksenæringen, vurderes kunnskapsstyrken som sterk. Det totale smittepresset av agens som kan krysse artsbarrieren fra laks til torsk, vurderes derfor å være lavt. Dette vektes tyngre enn at torsk tiltrekkes laksemerder.
For både «Frastøting som følge av tilstedeværelse av lakseoppdrettsanlegg» og «Negativ endring i fysiologi» er kunnskapsstatus vurdert og funnet så svak at sannsynlighetsvurderinger ikke gjennomføres. Når kunnskapen er veldig svak mister sannsynlighetsvurderingen verdi og kan ikke tillegges vekt. Sannsynligheten kan være hva som helst, vi vet ikke, og hvorvidt en slik sannsynlighet oppgis, er ikke lenger viktig. Derimot er grunnen til at sannsynligheten ikke oppgis viktig da det tydeliggjør at betydelig kunnskap mangler. Dette visualiseres med hvite noder og rød ring (figur 1.2). Vurdering av sannsynlighet utelates også for konsekvensen «Negativ effekt på kysttorskbestander som følge av lakseoppdrett i åpne merder i sjø». Usikkerheten er naturlig høy grunnet manglende kunnskap, og dersom konsekvensene er alvorlige kan vi konkludere med høy risiko. Manglende kunnskap gir likevel ikke alltid grunn til en slik konklusjon. I slike tilfeller vil antagelser som legges til grunn og argumentasjonen som helhet forklare hvorfor vurderingen dreier mot lavere risiko.
Fra tid til annen forekommer det utbrudd av «nye», hittil ukjente sykdommer, eller sykdommer som er kjente, men nye for en gitt art eller i et område. Konsekvensene vil derfor variere, men i enkelte tilfeller kan de bli svært alvorlige for både norsk oppdrettsnæring og villfisk. Denne typen hendelser kan gi opphav til en mulig overraskelse med svært store konsekvenser og er visualisert med svarte svaner i risikokartet.
Vurderingen av konsekvensens alvorlighetsgrad, sannsynligheten for at den inntreffer og tilhørende vurdering av kunnskapsstyrke, ligger til grunn for konklusjoner om risiko. Manometeret i figur 1.2 uttrykker i dette tilfellet lav risiko for «Negative effekter på kysttorskbestander som følge av lakseoppdrett i åpne merder i sjø fra 67°N til og med Vest-Finnmark». Risikokartene er et nyttig redskap for å visualisere risikokilder og årsaks-sammenhenger, mens karakterisering og argumentasjon i teksten gir grunnlaget for å forstå hvordan måling av konsekvenser, sannsynligheter, kunnskapsstyrke og risiko er gjennomført.
Faktorer som medfører «Negative effekter på kysttorskbestander som følge av lakseoppdrett i åpne merder i sjø»
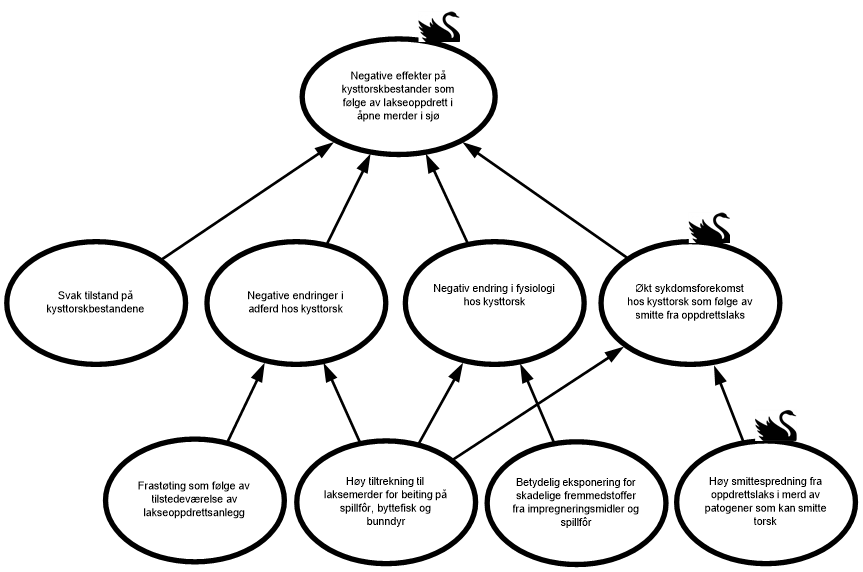
Produksjonen av laks i åpne merder i sjø vurderes som høy fra Jæren til Øst-Finnmark hvor alle produksjonsområdene har produksjonstetthet på over 11 tonn/km2. Med den høye produksjonen følger utslipp av ekskrementer, spillfôr, fremmedstoffer og smittsomme patogener. Dette kan føre til negative effekter på kysttorskbestandene og i verste fall svekke bestandene ytterligere og redusere mulighetene for en vellykket gjenoppbygging av bestandene. Faktorer som påvirker hvorvidt og i hvilken grad «Negative effekter på kysttorskbestander som følge av lakseoppdrett i åpne merder i sjø» inntreffer, er «Svak tilstand hos kysttorskbestandene», «Negative endringer i adferd hos kysttorsk», «Negative endringer i fysiologi hos kysttorsk» og «Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettslaks» (figur 1.3). Risikokilden «Svak tilstand hos kysttorskbestandene» er styrende for hvorvidt, eller i hvilken grad de andre risikofaktorene inntreffer og vil kunne bidra til en ytterligere svekkelse av kysttorskbestandene sør for 67°N og derved redusere mulighetene for en vellykket gjenoppbygging, samt føre til svekkede kysttorskbestander nord for 67°N.
Figur 1.3. Visualisering av risikokilder og hendelser som kan medføre «Negative effekter på kysttorskbestander som følge av lakseoppdrett i åpne merder i sjø».
«Svak tilstand hos kysttorskbestandene». En kysttorskbestand med dårlig rekruttering og lite eller ikke noe høstbart overskudd vurderes å ha en svak tilstand og dermed være mindre motstandsdyktig mot eventuelle negative effekter av lakseoppdrett.
Det er stor variasjon i hvor mye data som er tilgjengelig for kysttorsk i de tre forvaltningsområdene, og som følge av dette er det store forskjeller i nivået på bestandsvurderingene. For området sør for 62°N foreligger det per dags dato ingen kvantitativ bestandsvurdering, men det ble i 2025 utarbeidet en oppdatert statusrapport av Havforskningsinstituttet for kysttorsk i dette området. Til sammenligning gjøres det formelle bestandsvurderinger for kysttorsk 62°N – 67°N og for kysttorsk nord for 67°N av ICES. For området 62°N – 67°N gjøres det en bestandsvurdering med hensyn til relativ biomasse, men ikke absolutte estimater av biomasse. For området nord for 67°N estimeres biomasse både på gyte- og samlet bestand.
«Negativ endring i adferd hos kysttorsk» defineres som endring i kysttorskens naturlige adferd og vandringsmønster, og som vil kunne påvirke overlevelse, vekst og rekruttering. Hendelsen vil forekomme hvis kysttorsk enten unngår eller oppsøker områder med oppdrettsanlegg og avhenger av risikokildene «Frastøting som følge av tilstedeværelse av lakseoppdrettsanlegg» og «Høy tiltrekning til laksemerder for beiting på spillfôr, byttefisk og bunndyr».
«Negativ endring i fysiologi hos kysttorsk» kan oppstå hvis kysttorsk beiter på spillfôr fra lakseoppdrett og/eller organismer som har beitet på avfall fra anleggene. Dagens laksefôr inneholder en høy andel råstoff fra landbaserte planter, og næringssammensetningen er derfor svært forskjellig fra torskens naturlige føde. Det er knyttet bekymring til hvordan økt forekomst av ikke-marine fettsyrer og andre næringsstoffer vil kunne påvirke torskens reproduksjon, hovedsakelig i form av redusert kvalitet på gyteprodukter (egg, melke), økt deformasjon hos fiskelarver og redusert overlevelse for oppvoksende fisk.
Kysttorsken kan også eksponeres for miljøgifter og tungmetaller gjennom beiting på laksefôret og av impregneringsmiddel som lekker ut av nøtene. Eksponering for giftige fremmedstoffer i høye nok konsentrasjoner kan forårsake alvorlige, langsiktige effekter hos fisk, inkludert reproduksjonssvikt, immunforsvarssvekkelse, metabolismeforstyrrelser, organskade og atferdsendringer.
Hendelsen «Negativ endring i fysiologi hos kysttorsk» avhenger i hovedsak av de to risikokildene «Høy tiltrekning til laksemerder for beiting på spillfôr, byttefisk og bunndyr» og «Økt eksponering mot skadelige fremmedstoffer fra impregneringsmidler og spillfôr».
«Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettslaks» vil kunne forekomme dersom sykdomsutbrudd i åpne merder fører til forhøyet smittepress som spres til mottakelig villfisk gjennom horisontal smitte. Villfisk som oppholder seg i nærheten av et oppdrettsanlegg over tid, vil kunne utsettes for et høyere smittepress ved et pågående sykdomsutbrudd. Dette gjelder både torsk som oppsøker laksemerder for beiting, men også for stasjonær kysttorsk hvor oppdrettsanlegg overlapper med viktige oppvekstområder. Også feilernæring som følge av beiting på spillfôr, vil kunne påvirke den generelle helsestatusen og dermed mottakelighet for smitte.
Om mange nok individer blir kraftig svekket eller dør av sykdom som følge av smitte fra oppdrett, kan det i ytterste konsekvens føre til økt dødelighet ut over den naturlige dødeligheten i bestanden. Dette gjelder særlig for smittespredning av nye og endrede agens. Fra tid til annen forekommer det utbrudd av «nye», hittil ukjente sykdommer, eller sykdommer som er kjente, men nye for en gitt art eller i et område. Konsekvensene vil derfor variere, men i enkelte tilfeller kan de bli svært alvorlige for både oppdretts- og villfisk. Denne typen hendelser gir opphav til en mulig overraskelse med alvorlige konsekvenser og er visualisert med svarte svaner i risikokartet (figur 1.3).
Hendelsen «Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettslaks» påvirkes av risikokildene «Høy tiltrekning til laksemerder for beiting på spillfôr, byttefisk og bunndyr» og «Høy smittespredning fra oppdrettslaks i merd av patogener som kan smitte torsk».
«Frastøting som følge av tilstedeværelse av lakseoppdrettsanlegg» defineres som at kysttorsken unngår områder med lakseoppdrett. Dette kan forekomme hvis lakseanlegg er plassert på eller like ved gytefelt eller viktige oppvekstområder for torsk, og torsken endrer vandringsmønster og adferd og unngår disse områdene. For eksempel har fiskere lenge hevdet at gytevandrende torsk har endret vandringsmønster og unngår gytefelt i oppdrettsintensive områder. En slik endring av gyteområder kan føre til at torsken gyter i områder med mindre gunstig forhold for spredning av egg og larver.
«Høy tiltrekning til laksemerder for beiting på spillfôr, byttefisk og bunndyr». Oppdrettsanlegg tiltrekker fisk og andre marine organismer ved at de fysiske strukturene tilbyr ly og habitat, samt økt tilgjengelighet av mat i form av spillfôr og mulige byttedyr. Langs norskekysten er spesielt sei påvist å samle seg rundt oppdrettsanlegg, men også kysttorsk kan forekomme i store antall rundt åpne laksemerder. Adferdsundersøkelser gjort ved Smøla som er et (relativt) åpent kystområde med stor tetthet av lakseoppdrett, har vist at over halvparten av den lokale torskebestanden oppholdt seg ved anleggene, mens 9 % hadde en vedvarende tiltrekning og befant seg rundt laksemerdene i over en måned. Dette underbygges av en feltundersøkelse gjort i samme område der det ble vist at 19 % av 815 torsk fanget på ulike gyteplasser hadde en fettsyresammensetning i leveren ensbetydende med beiting på spillfôr og/eller byttedyr som hadde spist spillfôr. Det er foreløpig ikke gjort tilsvarende studier i andre områder langs kysten. Det er videre vist at aggregeringen av torsk nært oppdrettsanlegg kan variere mellom lokaliteter i samme område.
Tettheten av fisk som samler seg rundt et oppdrettsanlegg henger trolig sammen med fôrintensitet og mengden spillfôr.
På grunn av de antatt negative effektene på adferd og fysiologi hos kysttorsken, samt økt smittepress ved aggregering av villfisk, er det ikke ønskelig at en stor andel av torskebestanden tiltrekkes og lever ved oppdrettsanlegg over lengre tid.
«Betydelig eksponering mot skadelige fremmedstoffer fra impregneringsmidler og spillfôr». I denne analysen defineres fremmedstoffer som miljøgifter og tungmetaller fra bruken av impregneringsmidler og fra laksefôr. Avlusningsmidler vurderes ikke da bruken av disse har gått kraftig ned og har tilsynelatende svært begrenset effekt på villfisk ved anvendte doseringer.
Ved bruk av impregneringsmidler på nøtene vil både vannsøylen og bunnen under og rundt oppdrettsanleggene kunne påvirkes når midlene blør ut eller løsner fra nøtene, for eksempel ved høytrykksspyling. Inntil nylig var det bruken av kobber som ga den største bekymringen med hensyn til påvirkning på organismer som lever ved oppdrettsanleggene. Kobber er et naturlig tungmetall som forekommer i jordskorpen, i marine sediment og i sjøvann. Stoffet er en viktig faktor for enkelte enzymreaksjoner, men er giftig dersom konsentrasjonen av kobberforbindelser blir for høy. Kobber kan være giftig for ulike organismer i ulike utviklingsstadier. Det kan føre til redusert artsmangfold hvis konsentrasjonen i et gitt leveområde blir høyere enn artenes tålegrense, men kobber akkumuleres ikke oppover næringskjeden. Miljøkvalitetsmål for kobber i sediment er satt til 84 mg/kg tørt sediment. Kobberkonsentrasjoner under denne verdien antas å ikke ha noen giftig effekt på marine organismer. For kobber løst i vann er miljøkvalitetsmålet satt til 2,6 µg/l. Forbruket av kobber til impregnering i akvakulturnæringen var på 564 tonn i 2024.
I de senere årene har bruken av tralopyril som erstatningsstoff for kobber vært økende. Tralopyril er et ustabilt organisk pesticid med en halveringstid i sjø på 9 – 16 timer, men vi har manglende kunnskap om forekomst og giftighet til nedbrytningsproduktene og hvordan disse påvirker marine organismer. Forbruket av tralopyril til impregnering i akvakulturnæringen var på 108 tonn i 2024.
Antar vi at alle oppdrettsanlegg bruker impregneringsmidler vil eksponeringen være størst i områder med mye oppdrett og redusert vannutskiftning.
Skiftet fra marint til plantebasert fôr har ført til en betydelig endring i hvilke miljøgifter og tungmetaller som forekommer i laksefôret, fra persistente organiske miljøgifter (POP-er) og kvikksølv, til eksponering for pesticider som omfatter ugressmidler (herbicider) og insektmidler (insekticider) brukt i landbruket.
Mengde fôr som gis ved de ulike lokalitetene avhenger av antall og størrelsen på fisken (biomasse) som står i anlegget på et gitt tidspunkt. Ved å bruke et massebalansebudsjett som er basert på fôrsammensetning, er det beregnet at utslipp av fekalier utgjør 27,2 % av fôrmengden, mens spillfôr utgjør 5 %. Det totale fôrforbruket i områdene vi vurderer var i 2025 på 1 448 702 tonn sør for 67°N og 746 141 tonn nord for 67°N (tall fra Fiskeridirektoratet 20.01.2026). Dette gir en samlet mengde spillfôr på 72 435 tonn sør for 67 °N og 37 307 nord for 67°N. I 2025 var det henholdsvis 575 og 248 unike lokaliteter som meldte om uttak av fisk i de to områdene (tall fra Fiskeridirektoratet 20.01.2026). Tar man utgangspunkt i at alle lokaliteter har samme mengde spillfôr, utgjør det 126 tonn/lokalitet sør for 67°N og 150 tonn/lokalitet i området nord for 67°N i 2025. Prosentandelen spillfôr ved hver lokalitet vil variere avhengig av blant annet fôringsteknologi og appetitten til oppdrettsfisken. Videre vil vannutskiftningen ved lokalitetene påvirke hvor mye av spillfôret som er tilgjengelig for frittlevende organismer.
«Høy smittespredning fra oppdrettslaks i merd av patogener som kan smitte torsk». Det er per i dag høy produksjon av oppdrettslaks langs store deler av norskekysten og et høyt antall sykdomsutbrudd forårsaket av mange ulike agens. Generelt vil antallet anlegg og mengde fisk i sjø ha direkte innvirkning på utslipp av smittestoff fra oppdrett. Jo flere verter som frigir smittestoff, jo høyere blir smittepresset til omgivelsene og på mottakelige verter.
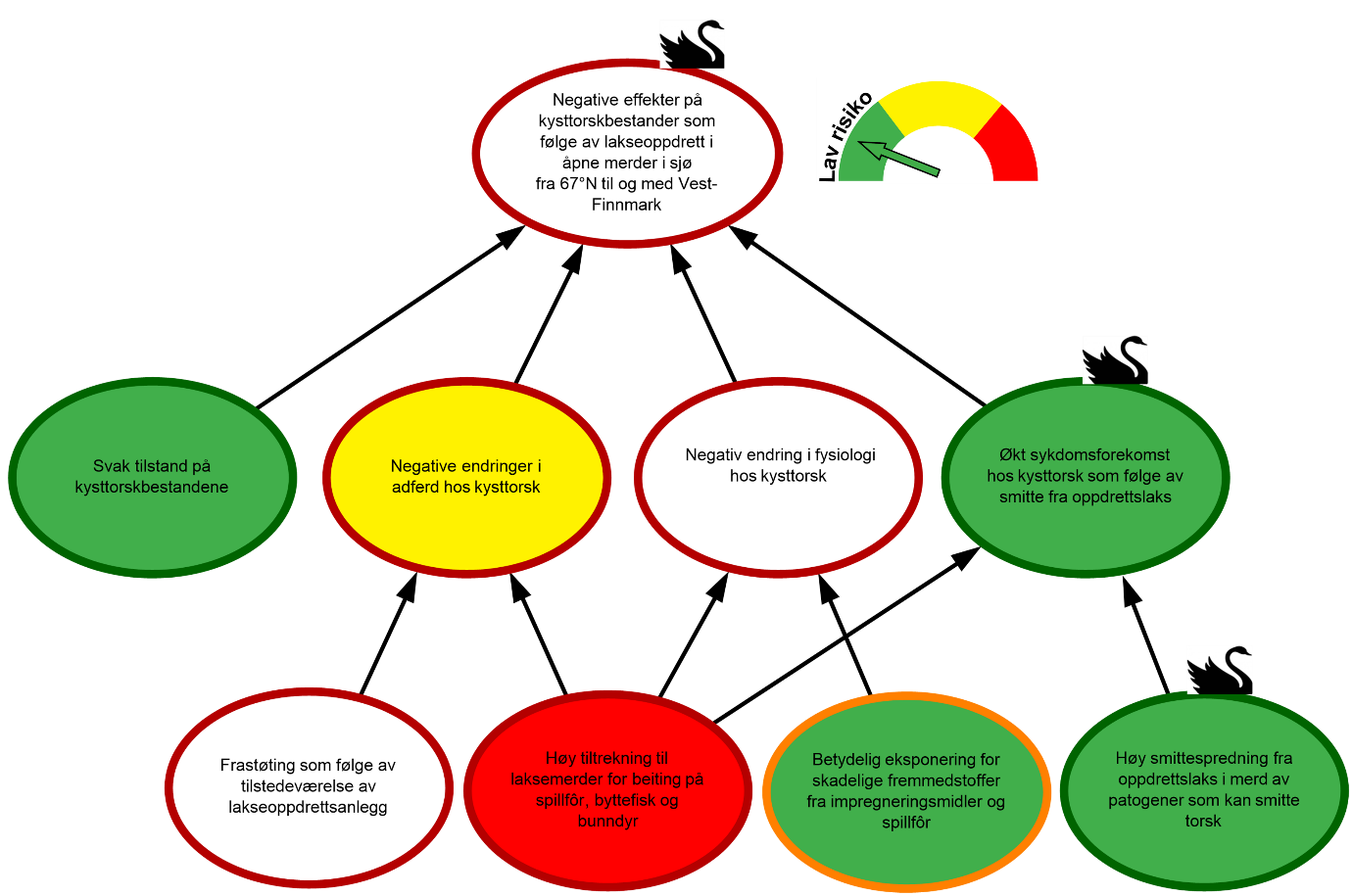
Risikovurdering «Negative effekter på kysttorskbestander som følge av lakseoppdrett i åpne merder i sjø fra 67°N til og med Vest-Finnmark»
Figur 1.4. Visualisering av risikobildet for negative effekter på kysttorskbestander som følge av lakseoppdrett i åpne merder i sjø fra 67°N til og med Vest-Finnmark.
«Svak tilstand på kysttorskbestandene» Nord for 67°N vurderes kysttorsk av ICES ved en aldersstrukturert bestandsmodell. Denne modellen viser en svak sammenheng mellom gytebestandsstørrelse og rekruttering, noe som kan tyde på lokal bestandsvariasjon innenfor forvaltningsområdet. Som følge av dette fastsettes det ikke en kritisk grense for gytebestanden.
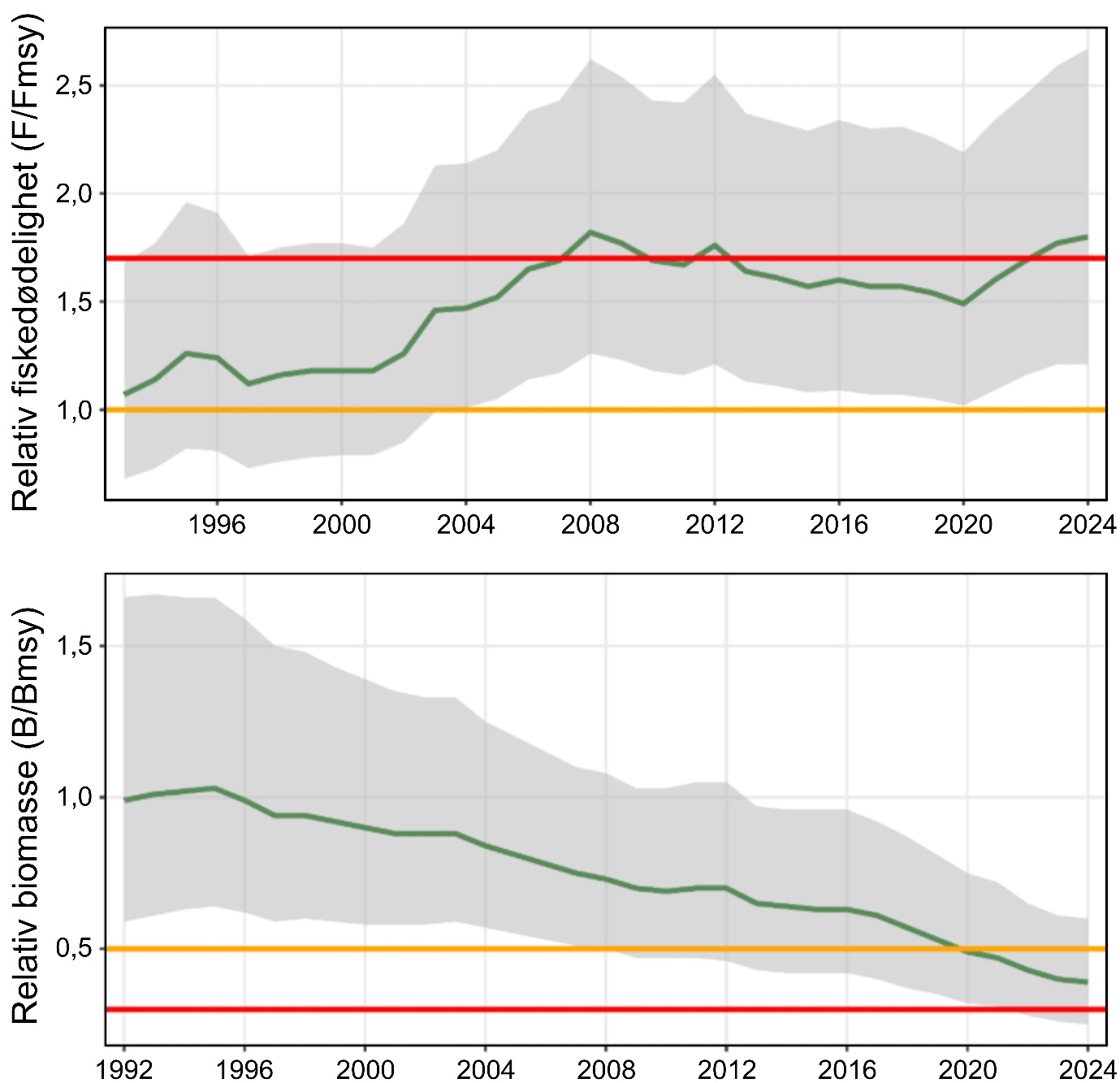
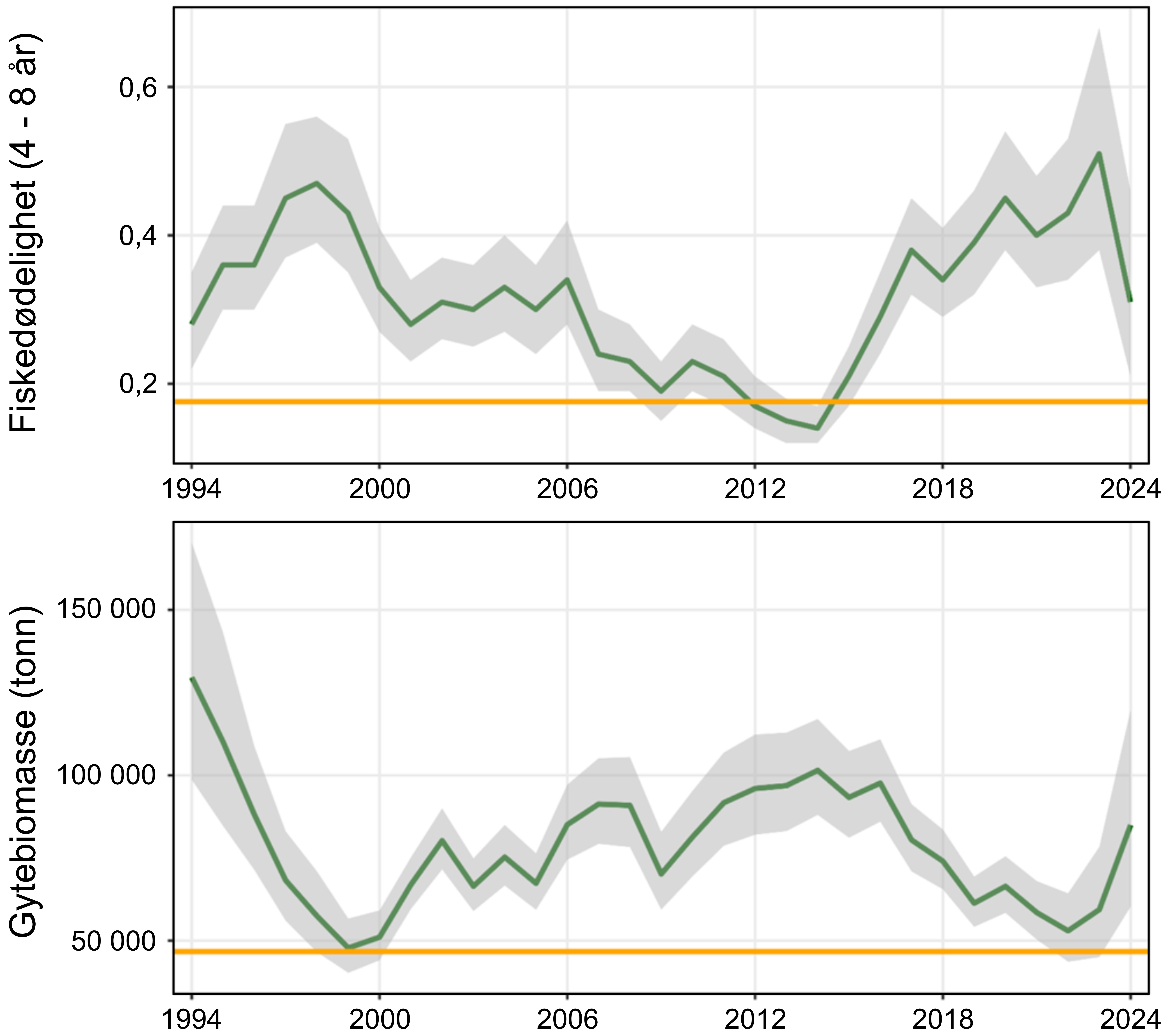
Siden midten av 1990-tallet har fiskedødeligheten for kysttorsk nord for 67°N for det meste vært over anbefalt nivå, og etter at det ble registrert spesielt høy dødelighet i 2023, returnerte den til et nivå som er bedre i samsvar med de siste års gjennomsnitt i 2024. Videre ble det i 2024 observert en 43 % økning i gytebestanden fra foregående år, sannsynligvis som følge av en sterk 2020-årsklasse samt økt vekst og tidligere kjønnsmodning for yngre fisk. Total bestandsbiomasse og biomassen på gytebestanden ble for 2024 estimert til henholdsvis 276 000 tonn (95 % konfidensintervall: 206 000 – 369 000 tonn) og 85 000 tonn (95 % konfidensintervall: 60 000 – 120 000 tonn). Disse er i relativt godt samsvar med bestandsestimatene siden tidlig 2000-tallet, men spesielt for gytebestanden er nåværende bestandsestimater betydelig lavere enn på starten av 1990-tallet. Samlet sett vurderes kysttorskbestandene nord for 67°N til å være i moderat forfatning fordi fiskedødeligheten overskrider anbefalt nivå. Likevel vurderes det som lav sannsynlighet for «Svak tilstand på kysttorskbestandene» siden bestanden har vist en positiv utvikling de siste årene. Kunnskapsstyrken vurderes som sterk gitt bestandsmodellen som brukes av ICES.
«Frastøting som følge av tilstedeværelse av lakseoppdrettsanlegg». Til tross for observasjoner fra fiskere om at gytevandrende torsk ungår områder med aktive oppdrettsanlegg, er det lite støtte for slik frastøting i den vitenskapelige litteraturen. Dette kan skyldes at årsak og virkning kan være adskilt over lang tid og at torskens valg av gyteområder også påvirkes av andre faktorer som for eksempel fiskepress og klimaendringer.
En laboratoriestudie har vist at torsk uten erfaring med oppdrett unngikk tanker med en liten andel vann fra både lakse- og torskeoppdrett. Dette kan tyde på en viss frastøting hos torsk fra områder med store fisketettheter i oppdrett. Til sammenligning viste en feltundersøkelse av kysttorsk på innsig ingen effekt av luktblokkering, da torsk satt ut i et område med oppdrettsanlegg ble værende på gytefeltet, mens torsk satt ut i ytre deler av fjorden forlot systemet. Denne responsen var uavhengig om luktesansen var intakt eller ikke. Disse undersøkelsene er utelukkende gjort på kjønnsmoden torsk, og det foreligger ingen kunnskap om hvorvidt yngre livsstadier unngår oppvekstområder i nærheten av oppdrettsanlegg.
Hvilket omfang eventuell frastøting har og hvilke mekanismer som er involvert i denne adferden, finnes det lite kunnskap om. Vi vurderer derfor kunnskapsstyrken til å være så svak at vi ikke gjør en sannsynlighetsvurdering av denne risikokilden. Dette visualiseres i risikokartet med en hvit node med rød ring. Vi velger likevel å ha med risikokilden i analysen siden det er rapportert observasjoner som kan tyde på at kysttorsken unngår områder med lakseoppdrett. Manglende kunnskap skaper usikkerhet, og det bør vurderes om det er behov for mer målrettet forskning rundt dette temaet for å øke kunnskapen og redusere usikkerheten.
«Høy tiltrekning til laksemerder for beiting på spillfôr, byttefisk og bunndyr». Adferdsundersøkelser gjort i et (relativt) åpent kystområde med stor tetthet av lakseoppdrett har vist at over halvparten av den lokale torskebestanden oppholdt seg ved anleggene, med høyere tiltrekning til anlegg hvor det var laks i merdene. Dette tyder på at tiltrekning styres av økt mattilgang og ikke av de fysiske strukturene. Sannsynligheten for «Høy tiltrekning til laksemerder for beiting på spillfôr, byttefisk og bunndyr» vurderes derfor å være høy.
Til tross for god kunnskap om at torsk kan tiltrekkes åpne laksemerder, er det usikkerhet knyttet til i hvilken grad torsk oppholder seg ved oppdrettsanlegg på en større skala, samt hvorvidt den observerte tiltrekningen er representativ for ulike livsstadier. Det er spesielt viktig å anerkjenne den manglende kunnskapen knyttet til hvorvidt torsk tiltrekkes laksemerder på lokalt bestandsnivå, ettersom kun et fåtall studier har estimert dette. Videre kan tiltrekningsgraden forventes å variere med hensyn til produksjonsintensitet, mengden kysttorsk, livsstadium, samt hvorvidt kysttorsken i området består av stasjonære eller vandrende individer. Kunnskapsstyrken vurderes derfor som svak.
«Negativ endring i adferd hos kysttorsk». Det vurderes å være høy sannsynlighet for høy tiltrekning av kysttorsk til lakseoppdrettsanlegg, mens det for faktoren frastøting ikke gjøres en sannsynlighetsvurdering siden det mangler grunnleggende kunnskap om risikokilden. Vi antar at tiltrekning har et større omfang enn frastøting.
Det er manglende kunnskap om hvorvidt tilstedeværelsen av oppdrettsanlegg fører til negativ endring i adferd hos kysttorsk, utover at de kan endre beitestrategi til en mer oppdrettsassosiert diett. Adferdsundersøkelser gjort i områder med mye oppdrett har ikke påvist gyting ved oppdrettsanleggene, og tilstedeværelsen av åpne laksemerder virket ikke til å hindre torsken i å oppholde seg på tradisjonelle gyteplasser under gyteperioden.
Sannsynligheten vurderes som moderat for «Negativ endring i adferd hos kysttorsk». Vurderingen hviler på svak kunnskap om i hvilket omfang kysttorsk tiltrekkes oppdrettsanleggene for å beite, og det mangler kunnskap om konsekvensene av adferdsendringen. Det mangler også kunnskap om hvorvidt den observerte tiltrekningen er representativ for ulike livsstadier og på bestandsnivå, og hvordan dette varierer mellom ulike områder. Kunnskapen rundt frastøting er svært begrenset. Totalt sett vurderes kunnskapsstyrken som svak. Det er behov for mer målrettet forskning rundt dette temaet for å øke kunnskapen og redusere usikkerheten.
«Betydelig eksponering for skadelige fremmedstoffer fra impregneringsmidler og spillfôr». Det finnes ikke data på hvilke anlegg som bruker impregneringsmidler, hvilke midler som brukes og frekvensen av høytrykksspyling. Det er dermed vanskelig å vurdere i hvilken grad kysttorsken blir eksponert for disse fremmedstoffene. Hvis vi antar at forbruket av impregneringsmidlene fordeles likt på alle anlegg langs kysten, blir de estimerte gjennomsnittlige utslippene lave. Miljøundersøkelsene (MOM C) underbygger dette med at de fleste anleggene nord for 67°N har lave verdier av kobber i sedimentene. Det er ingen overvåking av tralopyril som brukes som erstatningsstoff for kobber.
Omtrent 5% av fôrforbruket ender opp som spillfôr. I området nord for 67°N tilsvarer dette et årlig utslipp på 150 tonn/lokalitet. Hvor mye av dette spillfôret som spises av den oppdrettsassosierte kysttorsken vites ikke, men det antas at stimene med sei rundt anleggene spiser en relativt stor andel da undersøkelser har vist mye fôrpellets i magene til seien. Sei og andre organismer som spiser laksefôr kan likevel påvirke kysttorsk, ettersom torsk kan beite på disse.
Overgangen fra marint til plantebasert fôr har åpnet muligheten for at torsk som beiter på spillfôr vil bli eksponert for ugressmidler og insektmidler brukt i landbruket, som for eksempel glyfosat (Roundup-produkter), pirimiphos-methyl og chlorpyrifos (organofosfater). Siden vi ikke vet hvor mye spillfôr torsken faktisk spiser, er det vanskelig å si noe om graden av eksponering.
Med utgangspunkt at forbruket av impregneringsmidlene fordels likt mellom anlegg og at kysttorsken kun spiser en begrenset andel av spillfôret vurderes sannsynligheten for «Betydelig eksponering mot skadelige fremmedstoffer fra impregneringsmidler og spillfôr» som lav. Det bemerkes at i områder med mange oppdrettsanlegg som ligger tett, vil eksponeringen kunne være høyere. Det finnes data på fôrforbruk og forbruk av impregneringsstoffer for hele landet, men det er behov for data på lokalitetsnivå for å kunne gjøre bedre analyser av eksponering. Kunnskapsstyrken vurderes derfor som moderat.
«Negativ endring i fysiologi hos kysttorsk» forårsakes av «Høy tiltrekning til laksemerder for beiting på spillfôr, byttefisk og bunndyr» og «Betydelig eksponering for skadelige fremmedstoffer fra impregneringsmidler og spillfôr».
Bruk av kobber som antigroemiddel har tilsynelatende liten effekt på villfisk ettersom feltstudier har vist at hverken torsk eller sei fanget nært laksemerder har forhøyede nivåer av kobber i kroppen sammenlignet med fisk fra referansegrupper. Det er vist at tralopyril kan bli bioakkumulert til høye konsentrasjoner i blåskjell som vokser på tralopyril-impregnerte nøter, slik at det kan bli spredd i miljøet gjennom næringskjeden. Dette sannsynliggjør at også kysttorsk kan bli eksponert for tralopyril hvis den beiter på byttedyr som har fått i seg giften, men om dette gir negative endringer i fysiologien til torsken er det ikke gjort undersøkelser på.
I en nylig undersøkelse av pesticider i laksefilet ble ingen av de 43 påviste stoffene funnet over standard grenseverdi på 10 µg/kg, som regnes som et trygt nivå i mat. Selv om det ikke ble funnet glyfosat eller dens metabolitter i laksefilet, var dette stoffet den dominerende pesticiden i fôret (medianverdi 100 µg/kg). Dette tyder på at opptaket i laks er lavt, og at stoffet ikke akkumulerer opp i næringskjeden. Det kan likevel ikke utelukkes at pesticider funnet i fôret kan ha en påvirkning på fysiologien til fisken.
Torsk som beiter på spillfôr og byttedyr forbundet med oppdrett vil få en endret fettsyresammensetning i vevet. Det er lite som tyder på at en slik fysiologisk endring har en direkte negativ effekt på vekst eller overlevelse i voksne individer, og flere feltundersøkelser har vist at oppdrettsanlegg kan fungere som en effektiv energikilde for torsk. Dette kan isolert sett være positivt for bestanden, med resulterende økt individuell vekst og eggproduksjon (fekunditet).
Det er derimot knyttet bekymring til hvorvidt beiting på spillfôr og byttedyr forbundet med oppdrett kan redusere kvaliteten på gyteprodukter og overlevelsen til yngre livsstadier. Flere studier av diettkvalitet på stamfisk har vist at fekunditet og larveoverlevelse hos torsk avhenger av tilgang til enkelte langkjedede marine fettsyrer. Spesielt de essensielle fettsyrene ARA (arakidonsyre, 20:4n-6), EPA (eikosapentaensyre, 20:5n-3) og DHA (dokosaheksaensyre, 22:6n-3) er viktige for eggutvikling, befruktning, larveoverlevelse og vekst. I oppdrettsintensive områder er det påvist at 19 % av gytefisken fanget på gytefeltene beiter intensivt på spillfôr og har forhøyede nivåer av plantebaserte fettsyrer i lever og muskel, men også i rogn og melke. Gitt de observerte nivåene av terrestriske fettsyrer hos gytemoden torsk i oppdrettsintensive områder, samt fraværet av viktige langkjedede marine fettsyrer hos den samme gytefisken, anses muligheten for alvorlige negative konsekvenser for reproduksjonen til kysttorsk å være til stede. Usikkerhetene vurderes som høy ettersom det mangler konkrete eksperimentelle studier som kan bekrefte eller avkrefte dette.
Selv om det vurderes å være lav sannsynlighet for betydelig eksponering for skadelige fremmedstoffer, vektes påvisningen av endret fettsyresammensetning fra beiting på spillfôr da dette kan gi negative effekter på reproduksjonen. Det mangler grunnleggende kunnskap om hvor stor andel av kysttorskbestanden som tiltrekkes og beiter på spillfôr på en større skala og i hvilken grad dette varierer med intensiteten av lakseoppdrett. Det er også manglende kunnskap om eksponering av fremmedstoffer fra impregnering på lokalt nivå. Videre inneholder spillfôr landbruksbaserte ugress- og insektmidler som ikke er testet for helseeffekter på torsk. De langsiktige effektene beiting på laksefôr kan ha for kysttorskens reproduksjon er ikke studert. Dette er fremholdt som det viktigste kunnskapshullet for hvordan lakseoppdrett kan påvirke ville torskebestander. Vi vurderer derfor kunnskapsstyrken til å være så svak at vi ikke gjør en sannsynlighetsvurdering av denne risikokilden. Dette visualiseres i risikokartet med en hvit node med rød ring.
Manglende kunnskap skaper usikkerhet, og det er behov for mer målrettet forskning rundt dette temaet for å øke kunnskapen og redusere usikkerheten.
«Høy smittespredning fra oppdrettslaks i merd av patogener som kan smitte torsk». Med unntak av miljøbakterier (sårbakterier som Moritella viscosa, Tenacibaculum spp.) er de fleste av sykdomsutbruddene i lakseoppdrett forårsaket av laksespesifikke agens (f.eks. ILAV, SAV, HSMB, CMS) som ikke vil smitte torsk. Laks er vaksinert og dermed godt beskyttet mot sykdommer som kan smitte torsk, som for eksempel atypisk furunkulose og vibriose. Bruken av vaksiner holder antallet sykdomsutbrudd forårsaket av agens som kan smitte over på torsk, nede på et svært lavt nivå. Skottelus på villtorsk er i hovedsak en annen variant (genotype) enn den som vokser opp på laksefisk. Smitte mellom artene vil derfor forekomme i liten grad. Sannsynligheten for «Høy smittespredning fra oppdrettslaks i merd av patogener som kan smitte torsk» vurderes derfor som lav.
Selv om det ikke er fullstendig oversikt over annet enn de meldepliktige sykdommene (sykdommer på liste F), gir fiskehelserapporten en god oversikt over sykdomssituasjonen i norsk fiskeoppdrett og hvilke sykdommer som finnes i lakseoppdrett. Kunnskapsstyrken vurderes som sterk.
«Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettslaks». Det er vist at villtorsk kan samle seg rundt lakseanlegg for å beite på spillfôr og organismer forbundet med oppdrett. Overlapp i tid og rom mellom tilstedeværelse og sykdomsutbrudd vil kunne utsette villtorsk for økt smittepress. Det er derimot lav sannsynlighet for høy smittespredning av patogener som kan smitte videre fra oppdrettslaks til kysttorsk. På bakgrunn av dette vurderes sannsynligheten for økt sykdomsforekomst hos kysttorsk som lav. Fiskehelserapporten gir en god oversikt over sykdomssituasjonen i norsk fiskeoppdrett og hvilke sykdommer som finnes i lakseoppdrett. Kunnskapsstyrken vurderes som sterk.
«Negative effekter på kysttorskbestander som følge av lakseoppdrett i åpne merder i sjø fra 67°N til og med Vest-Finnmark». Det mangler betydelig kunnskap om endringer i adferd og fysiologi hos kysttorsken som følge av tiltrekning til lakseoppdrett og beiting på spillfôr. Dette bidrar til økt usikkerhet om hvorvidt og i hvilken grad lakseoppdrett i åpne merder i sjø kan ha negative effekter på den samlede kysttorskbestanden fra 67°N til og med Vest-Finnmark. Vi vurderer kunnskapsstyrken som for svak til å vurdere sannsynligheten for «Negative effekter på kysttorskbestander som følge av lakseoppdrett i åpne merder i sjø fra 67°N til Vest-Finnmark». Dette visualiseres i risikokartet med en hvit node for manglende grunnlag for sannsynlighetsvurdering og rød ring for svak kunnskap.
Basert på sterk kunnskap vurderes bestanden nord for 67°N å ikke være svak. Det vil imidlertid være lokal variasjon i bestandsstatus innenfor området. Kunnskapen om smitte mellom oppdrettslaks og kysttorsk vurderes også som sterk og sannsynligheten for «Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettslaks» vurderes som lav. Det mangler derimot kunnskap om i hvilken grad lakseoppdrett i åpne merder kan føre til negative endringer i adferd eller fysiologi hos kysttorsk. Kunnskapsstyrken vurderes derfor som svak for begge disse hendelsene. Vi legger spesielt vekt på manglende kunnskap om hvorvidt de observerte fysiologiske endringene, gjennom økt eksponering for terrestriske fettsyrer og fremmedstoffer via spillfôr, kan ha en negativ effekt på reproduksjonen. I tillegg vektlegges manglende kunnskap om hvor stor andel av bestandene som faktisk tiltrekkes lakseoppdrett, ettersom dette vil være en forutsetning for hvordan eventuelle negative effekter på reproduksjon påvirker bestandene.
Manglende kunnskap skaper usikkerhet og gitt de mulig alvorlige konsekvensene av beiting på spillfôr på kysttorskens reproduksjon, vil risikoen i utgangspunktet vurderes som høy. Basert på data om mengde spillfôr (37 000 tonn i 2025) og størrelsen på kysttorskbestanden (total bestandsbiomasse på 276 000 tonn og gytebestandsbiomasse på 85 000 tonn i 2024), antas det at den samlede produksjonen av oppdrettslaks i området ikke gir tilstrekkelige mengder spillfôr til å ha negative effekter på kysttorskbestanden. Videre antar vi at en stor andel av spillfôret aldri blir konsumert av kysttorsk ettersom det enten kan spres av vannstrømmen eller spises av andre organismer. I tillegg vurderes ikke kysttorskbestanden å være svak. Basert på dette konkluderer vi med lav risiko for «Negative effekter på kysttorskbestander som følge av lakseoppdrett i åpne merder i sjø fra 67°N til Vest-Finnmark».
Vi påpeker samtidig at tiltrekning av en ukjent andel av kysttorsken for beiting ved oppdrettsanleggene, med usikker påvirkning på reproduksjon, vil kunne øke risikoen i enkelte områder. Eksempelvis der hvor lakseoppdrett overlapper med leveområdet til små eller svake lokale bestander av kysttorsk, eller i spesielt produksjonsintensive områder. Videre, kan også mer isolerte lokale kysttorskbestander bestående av stasjonære kysttorsk være utsatt for høyere risiko, sammenlignet med bestander bestående av en større andel vandrende individer.
Risikovurdering «Negative effekter på kysttorskbestander som følge av lakseoppdrett i åpne merder i sjø fra Jæren til 67°N»
Figur 1.5. Visualisering av risikobildet for negative effekter på kysttorskbestander som følge av lakseoppdrett i åpne merder i sjø fra Jæren til 67°N.
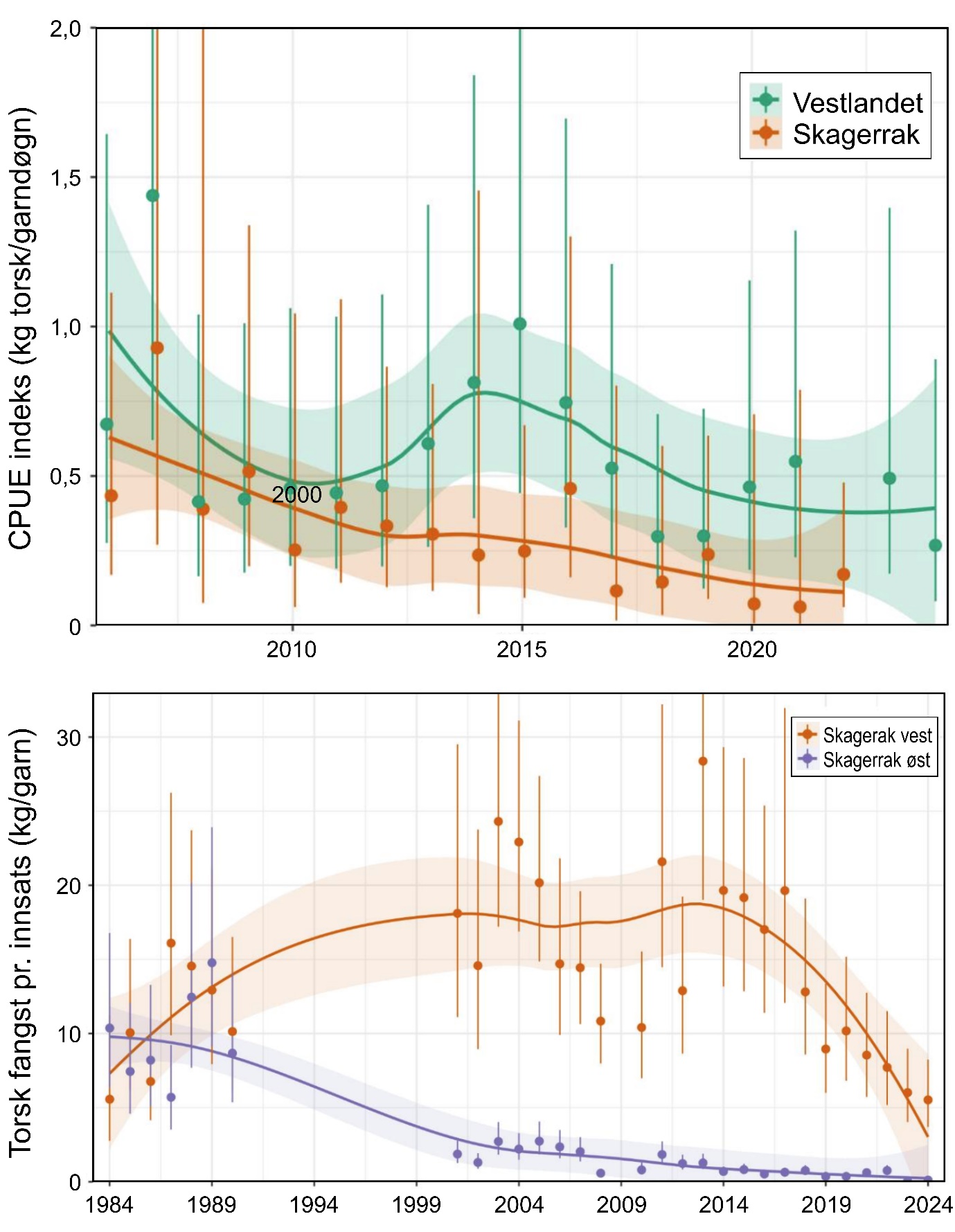
«Svak tilstand på kysttorskbestandene». Fra Jæren til 67°N forvaltes kysttorsk i to forvaltningsområder: sør for 62°N og mellom 62°N – 67°N. For området sør for 62°N foreligger det per dags dato ingen kvantitativ bestandsvurdering. Havforskningsinstituttet kom i 2025 med en oppdatert status for kysttorsk i dette området der alle tilgjengelig data tyder på at kysttorsken i dette forvaltningsområdet er i svært dårlig forfatning og biomassen antas i dag å være på historisk lave nivåer. Vekstforholdene (individvekst) begrenser ikke torskebestandene sør for 62°N. Av mulige påvirkningsfaktorer ansees fiskedødelighet som en viktig årsak til den observerte nedgangen, og da spesielt dødelighet fra turist- og fritidsfiske som har blitt beregnet å være høyere enn fra kommersielt fiskeri.
Vurdering av status gjort av ICES for kysttorskbestanden mellom 62°N og 67°N, konkluderer at fisketrykket er over kritisk nivå (rød status) og biomassen er mellom målnivå og kritisk lavt nivå (gul status). I bestandsvurderingen er det biomassen tilgjengelig for fiskeri som estimeres. Dette vil være et maksimalt estimat av gytebiomassen fordi modellen definerer høstbar biomasse som fisk over lengde 50 cm, som omtrent tilsvarer lengden der 50 % er kjønnsmodne. Den nåværende bestandsvurderingen støttes av kommersielle fangster og tetthetsindikatorer fra Havforskningsinstituttets overvåkningstokt som har vist en markant nedgang det siste tiåret. Det største problemet med den nåværende vurderingen av bestandsstatus for kysttorsk 62°N – 67°N er den høye usikkerheten rundt fangstene i turist- og fritidsfiske, da dette fisket sannsynligvis utgjør mer enn halvparten av de totale fangstene.
For kysttorsk sør for 67°N er det knyttet stor usikkerhet rundt de faktiske bestandsstørrelsene i begge forvaltningsområdene, ettersom en liten bestand med få individer, færre fiskeridata og lav robusthet kan være vanskelig å vurdere. Til tross for dette tilsier alle tilgjengelig datakilder at bestandene er på lave til svært lave nivåer, og følgelig vurderes sannsynlighet for «Svak tilstand på kysttorskbestandene» å være høy. Samlet sett vurderes kunnskapsgrunnlaget som sterkt, ettersom usikkerheten er knyttet til de faktiske bestandsstørrelsene og ikke bestandenes relative status og trender, eller hvorvidt de er redusert fra historiske nivåer.
«Frastøting som følge av tilstedeværelse av lakseoppdrettsanlegg». Til tross for observasjoner fra fiskere om at gytevandrende torsk ungår områder med aktive oppdrettsanlegg, er det lite støtte for slik frastøting i den vitenskapelige litteraturen. Dette kan skyldes at årsak og virkning kan være adskilt over lang tid og at torskens valg av gyteområder også påvirkes av andre faktorer som for eksempel fiskepress og klimaendringer.
En laboratoriestudie har vist at torsk uten erfaring med oppdrett unngikk tanker med en liten andel vann fra både lakse- og torskeoppdrett. Dette kan tyde på en viss frastøting hos torsk fra områder med store fisketettheter i oppdrett. Til sammenligning viste en feltundersøkelse av kysttorsk på innsig ingen effekt av luktblokkering, da torsk satt ut i et område med oppdrettsanlegg ble værende på gytefeltet, mens torsk satt ut i ytre deler av fjorden forlot systemet. Denne responsen var uavhengig om luktesansen var intakt eller ikke. Disse undersøkelsene er utelukkende gjort på kjønnsmoden torsk, og det foreligger ingen kunnskap om hvorvidt yngre livsstadier unngår oppvekstområder i nærheten av oppdrettsanlegg.
Hvilket omfang eventuell frastøting har og hvilke mekanismer som er involvert i denne adferden, finnes det lite kunnskap om. Vi vurderer derfor kunnskapsstyrken til å være så svak at vi ikke gjør en sannsynlighetsvurdering av denne risikokilden. Dette visualiseres i risikokartet med en hvit node med rød ring. Vi velger likevel å ha med risikokilden i analysen siden det er rapportert observasjoner som kan tyde på at kysttorsken unngår områder med lakseoppdrett. Manglende kunnskap skaper usikkerhet, og det bør vurderes om det er behov for mer målrettet forskning rundt dette temaet for å øke kunnskapen og redusere usikkerheten.
«Høy tiltrekning til laksemerder for beiting på spillfôr, byttefisk og bunndyr». Adferdsundersøkelser gjort i et (relativt) åpent kystområde med stor tetthet av lakseoppdrett har vist at over halvparten av den lokale torskebestanden oppholdt seg ved anleggene, med høyere tiltrekning til anlegg hvor det var laks i merdene. Dette tyder på at tiltrekning styres av økt mattilgang og ikke av de fysiske strukturene. Sannsynligheten for «Høy tiltrekning til laksemerder for beiting på spillfôr, byttefisk og bunndyr» vurderes derfor å være høy.
Til tross for god kunnskap om at torsk kan tiltrekkes åpne laksemerder, er det usikkerhet knyttet til i hvilken grad torsk oppholder seg ved oppdrettsanlegg på en større skala, samt hvorvidt den observerte tiltrekningen er representativ for ulike livsstadier. Det er spesielt viktig å anerkjenne den manglende kunnskapen knyttet til hvorvidt torsk tiltrekkes laksemerder på lokalt bestandsnivå, ettersom kun et fåtall studier har estimert dette. Videre kan tiltrekningsgraden forventes å variere med hensyn til produksjonsintensitet, mengden kysttorsk, livsstadium, samt hvorvidt kysttorsken i området består av stasjonære eller vandrende individer. Kunnskapsstyrken vurderes derfor som svak.
«Negativ endring i adferd hos kysttorsk». Det vurderes å være høy sannsynlighet for høy tiltrekning av kysttorsk til lakseoppdrettsanlegg, mens det for faktoren frastøting ikke gjøres en sannsynlighetsvurdering siden det mangler grunnleggende kunnskap om risikokilden. Vi antar at tiltrekning har et større omfang enn frastøting.
Det er manglende kunnskap om hvorvidt tilstedeværelsen av oppdrettsanlegg fører til negativ endring i adferd hos kysttorsk, utover at de kan endre beitestrategi til en mer oppdrettsassosiert diett. Adferdsundersøkelser gjort i områder med mye oppdrett har ikke påvist gyting ved oppdrettsanleggene, og tilstedeværelsen av åpne laksemerder virket ikke til å hindre torsken i å oppholde seg på tradisjonelle gyteplasser under gyteperioden.
Sannsynligheten vurderes som moderat for «Negativ endring i adferd hos kysttorsk». Vurderingen hviler på svak kunnskap om i hvilket omfang kysttorsk tiltrekkes oppdrettsanleggene for å beite, og det mangler kunnskap om konsekvensene av adferdsendringen. Det mangler også kunnskap om hvorvidt den observerte tiltrekningen er representativ for ulike livsstadier og på bestandsnivå, og hvordan dette varierer mellom ulike områder. Kunnskapen rundt frastøting er svært begrenset. Totalt sett vurderes kunnskapsstyrken som svak. Det er behov for mer målrettet forskning rundt dette temaet for å øke kunnskapen og redusere usikkerheten.
«Betydelig eksponering for skadelige fremmedstoffer fra impregneringsmidler og spillfôr». Det finnes ikke data på hvilke anlegg som bruker impregneringsmidler, hvilke midler som brukes og frekvensen av høytrykksspyling. Det er dermed vanskelig å vurdere i hvilken grad kysttorsken blir eksponert for disse fremmedstoffene. Hvis vi antar at forbruket av impregneringsmidlene fordeles likt på alle anlegg langs kysten, blir de estimerte gjennomsnittlige utslippene lave. Miljøundersøkelsene (MOM C) underbygger dette med at de fleste anleggene i området sør for 67°N har lave verdier av kobber i sedimentene. Unntaket er produksjonsområdene 2 og 3, Karmøy til Stadt der det finnes anlegg med nivå av kobber over grenseverdi. Det er ingen overvåking av tralopyril som brukes som erstatningsstoff for kobber.
Omtrent 5% av fôrforbruket ender opp som spillfôr. I området sør for 67°N tilsvarer dette et årlig utslipp på 126 tonn/lokalitet. Hvor mye av dette spillfôret som spises av den oppdrettsassosierte kysttorsken vites ikke, men det antas at stimene med sei rundt anleggene spiser en relativt stor andel da undersøkelser har vist mye fôrpellets i magene til seien. Sei og andre organismer som spiser laksefôr kan likevel påvirke kysttorsk, ettersom torsk kan beite på disse.
Overgangen fra marint til plantebasert fôr har åpnet muligheten for at torsk som beiter på spillfôr vil bli eksponert for ugressmidler og insektmidler brukt i landbruket, som for eksempel glyfosat (Roundup-produkter), pirimiphos-methyl og chlorpyrifos (organofosfater). Siden vi ikke vet hvor mye spillfôr torsken faktisk spiser, er det vanskelig å si noe om graden av eksponering.
Med utgangspunkt at forbruket av impregneringsmidlene fordels likt mellom anlegg og at kysttorsken kun spiser en begrenset andel av spillfôret vurderes sannsynligheten for «Betydelig eksponering mot skadelige fremmedstoffer fra impregneringsmidler og spillfôr» som lav. Det bemerkes at i områder med mange oppdrettsanlegg som ligger tett, vil eksponeringen kunne være høyere. Det finnes data på fôrforbruk og forbruk av impregneringsstoffer for hele landet, men det er behov for data på lokalitetsnivå for å kunne gjøre bedre analyser av eksponering. Kunnskapsstyrken vurderes derfor som moderat.
«Negativ endring i fysiologi hos kysttorsk» forårsakes av «Høy tiltrekning til laksemerder for beiting på spillfôr, byttefisk og bunndyr» og «Betydelig eksponering for skadelige fremmedstoffer fra impregneringsmidler og spillfôr».
Bruk av kobber som antigroemiddel har tilsynelatende liten effekt på villfisk ettersom feltstudier har vist at hverken torsk eller sei fanget nært laksemerder har forhøyede nivåer av kobber i kroppen sammenlignet med fisk fra referansegrupper. Det er vist at tralopyril kan bli bioakkumulert til høye konsentrasjoner i blåskjell som vokser på tralopyril-impregnerte nøter, slik at det kan bli spredd i miljøet gjennom næringskjeden. Dette sannsynliggjør at også kysttorsk kan bli eksponert for tralopyril hvis den beiter på byttedyr som har fått i seg giften, men om dette gir negative endringer i fysiologien til torsken er det ikke gjort undersøkelser på.
I en nylig undersøkelse av pesticider i laksefilet ble ingen av de 43 påviste stoffene funnet over standard grenseverdi på 10 µg/kg, som regnes som et trygt nivå i mat. Selv om det ikke ble funnet glyfosat eller dens metabolitter i laksefilet, var dette stoffet den dominerende pesticiden i fôret (medianverdi 100 µg/kg). Dette tyder på at opptaket i laks er lavt, og at stoffet ikke akkumulerer opp i næringskjeden. Det kan likevel ikke utelukkes at pesticider funnet i fôret kan ha en påvirkning på fysiologien til fisken.
Torsk som beiter på spillfôr og byttedyr forbundet med oppdrett vil få en endret fettsyresammensetning i vevet. Det er lite som tyder på at en slik fysiologisk endring har en direkte negativ effekt på vekst eller overlevelse i voksne individer, og flere feltundersøkelser har vist at oppdrettsanlegg kan fungere som en effektiv energikilde for torsk. Dette kan isolert sett være positivt for bestanden, med resulterende økt individuell vekst og eggproduksjon (fekunditet).
Det er derimot knyttet bekymring til hvorvidt beiting på spillfôr og byttedyr forbundet med oppdrett kan redusere kvaliteten på gyteprodukter og overlevelsen til yngre livsstadier . Flere studier av diettkvalitet på stamfisk har vist at fekunditet og larveoverlevelse hos torsk avhenger av tilgang til enkelte langkjedede marine fettsyrer. Spesielt de essensielle fettsyrene ARA (arakidonsyre, 20:4n-6), EPA (eikosapentaensyre, 20:5n-3) og DHA (dokosaheksaensyre, 22:6n-3) er viktige for eggutvikling, befruktning, larveoverlevelse og vekst. I oppdrettsintensive områder er det påvist at 19 % av gytefisken fanget på gytefeltene beiter intensivt på spillfôr og har forhøyede nivåer av plantebaserte fettsyrer i lever og muskel, men også i rogn og melke. Gitt de observerte nivåene av terrestriske fettsyrer hos gytemoden torsk i oppdrettsintensive områder, samt fraværet av viktige langkjedede marine fettsyrer hos den samme gytefisken, anses muligheten for alvorlige negative konsekvenser for reproduksjonen til kysttorsk å være til stede. Usikkerhetene vurderes som høy ettersom det mangler konkrete eksperimentelle studier som kan bekrefte eller avkrefte dette.
Selv om det vurderes å være lav sannsynlighet for betydelig eksponering for skadelige fremmedstoffer, vektes påvisningen av endret fettsyresammensetning fra beiting på spillfôr da dette kan gi negative effekter på reproduksjonen. Det mangler grunnleggende kunnskap om hvor stor andel av kysttorskbestanden som tiltrekkes og beiter på spillfôr på en større skala og i hvilken grad dette varierer med intensiteten av lakseoppdrett. Det er også manglende kunnskap om eksponering av fremmedstoffer fra impregnering på lokalt nivå. Videre inneholder spillfôr landbruksbaserte ugress- og insektmidler som ikke er testet for helseeffekter på torsk. De langsiktige effektene beiting på laksefôr kan ha for kysttorskens reproduksjon er ikke studert. Dette er fremholdt som det viktigste kunnskapshullet for hvordan lakseoppdrett kan påvirke ville torskebestander. Vi vurderer derfor kunnskapsstyrken til å være så svak at vi ikke gjør en sannsynlighetsvurdering av denne risikokilden. Dette visualiseres i risikokartet med en hvit node med rød ring.
Manglende kunnskap skaper usikkerhet, og det er behov for mer målrettet forskning rundt dette temaet for å øke kunnskapen og redusere usikkerheten.
«Høy smittespredning fra oppdrettslaks i merd av patogener som kan smitte torsk». Med unntak av miljøbakterier (sårbakterier som Moritella viscosa, Tenacibaculum spp.) er de fleste av sykdomsutbruddene i lakseoppdrett forårsaket av laksespesifikke agens (f.eks. ILAV, SAV, HSMB, CMS) som ikke vil smitte torsk. Laks er vaksinert og dermed godt beskyttet mot sykdommer som kan smitte torsk, som for eksempel atypisk furunkulose og vibriose. Bruken av vaksiner holder antallet sykdomsutbrudd forårsaket av agens som kan smitte over på torsk, nede på et svært lavt nivå. Skottelus på villtorsk er i hovedsak en annen variant (genotype) enn den som vokser opp på laksefisk. Smitte mellom artene vil derfor forekomme i liten grad. Sannsynligheten for «Høy smittespredning fra oppdrettslaks i merd av patogener som kan smitte torsk» vurderes derfor som lav.
Selv om det ikke er fullstendig oversikt over annet enn de meldepliktige sykdommene (sykdommer på liste F), gir fiskehelserapporten en god oversikt over sykdomssituasjonen i norsk fiskeoppdrett og hvilke sykdommer som finnes i lakseoppdrett. Kunnskapsstyrken vurderes som sterk.
«Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettslaks». Det er vist at villtorsk kan samle seg rundt lakseanlegg for å beite på spillfôr og organismer forbundet med oppdrett. Denne typen overlapp vil kunne utsette villtorsk for økt smittepress. Det er derimot lav sannsynlighet for høy smittespredning av patogener som kan smitte videre fra oppdrettslaks til kysttorsk. På bakgrunn av dette vurderes sannsynligheten for økt sykdomsforekomst hos kysttorsk som lav. Fiskehelserapporten gir en god oversikt over sykdomssituasjonen i norsk fiskeoppdrett og hvilke sykdommer som finnes i lakseoppdrett. Kunnskapsstyrken vurderes som sterk.
«Negative effekter på kysttorskbestander som følge av lakseoppdrett i åpne merder i sjø fra Jæren til 67°N». Det mangler betydelig kunnskap om endringer i adferd og fysiologi hos kysttorsken som følge av tiltrekning til lakseoppdrett og beiting på spillfôr. Dette bidrar til økt usikkerhet om hvorvidt og i hvilken grad lakseoppdrett i åpne merder i sjø kan ha negative effekter på den samlede kysttorskbestanden fra Jæren til 67°N. Vi vurderer kunnskapsstyrken som for svak til å vurdere sannsynligheten for «Negative effekter på kysttorskbestander som følge av lakseoppdrett i åpne merder i sjø fra Jæren til 67°N». Dette visualiseres i risikokartet med en hvit node for manglende grunnlag for sannsynlighetsvurdering og rød ring for svak kunnskap.
Det vurderes å være sterk kunnskap om at den overordnede tilstanden til kysttorskbestandene sør for 67°N er svak, selv om størrelsen på bestandene er svært usikker. Det vil imidlertid være lokal variasjon i bestandsstatus innenfor området, med generelt svakere bestander sør for 62°N. Kunnskapen om smitte mellom oppdrettslaks og kysttorsk vurderes også som sterk og sannsynligheten for «Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettslaks» vurderes som lav. Det mangler derimot kunnskap om i hvilken grad lakseoppdrett i åpne merder kan føre til negative endringer i adferd eller fysiologi hos kysttorsk. Kunnskapsstyrken vurderes derfor som svak for begge disse hendelsen. Vi legger spesielt vekt på manglende kunnskap om hvorvidt de observerte fysiologiske endringene, gjennom økt eksponering for terrestriske fettsyrer og fremmedstoffer via spillfôr, kan ha en negativ effekt på reproduksjonen. I tillegg vektlegges manglende kunnskap om hvor stor andel av bestandene som faktisk tiltrekkes lakseoppdrett, ettersom dette vil være en forutsetning for hvordan eventuelle negative effekter på reproduksjon påvirker bestandene.
Manglende kunnskap skaper usikkerhet og gitt de mulig alvorlige konsekvensene av beiting på spillfôr på kysttorskens reproduksjon, vil risikoen i utgangspunktet vurderes som høy. Kysttorskbestandene sør for 67°N er sårbare, men det finnes per i dag ikke pålitelige estimater av mengden kysttorsk i disse områdene. Videre er både produksjonen av oppdrettsfisk og mengden spillfôr (anslagsvis 72 000 tonn i 2025) betraktelig høyere fra Jæren til 67°N enn fra 67°N til og med Vest-Finnmark. Samtidig antar vi at en stor andel av spillfôret aldri blir konsumert av kysttorsk ettersom det enten kan spres av vannstrømmen eller spises av andre organismer. Vi vurderer risikoen som noe høyere sammenlignet med området nord for 67°N og konkluderer med moderat risiko for «Negative effekter på kysttorskbestander som følge av lakseoppdrett i åpne merder i sjø fra Jæren til 67°N».
Vi påpeker samtidig at tiltrekning av en ukjent andel av kysttorsken for beiting ved oppdrettsanleggene, med usikker påvirkning på reproduksjon, vil kunne øke risikoen i enkelte områder. Eksempelvis i områder hvor lakseoppdrett overlapper med leveområdet til små eller svake lokale bestander av kysttorsk, eller i spesielt produksjonsintensive områder. Videre, kan også mer isolerte lokale kysttorskbestander bestående av stasjonære kysttorsk være utsatt for høyere risiko, sammenlignet med bestander bestående av en større andel vandrende individer.
Ettersom både bestanden av kysttorsk er svakere og oppdrettsintensiteten er høyere fra Jæren til Stad enn mellom 62°N og 67°N, kan risiko for negative effekter på kysttorskbestandene være høyere i dette området. For fremtidige risikovurderinger bør det gjøres en egen vurdering i det sørligste forvaltningsområdet.
Kunnskapsstatus
Innledning
Denne kunnskapsstatusen er en oppdatert versjon av den som ble utgitt i 2025 (Strøm mfl. 2025), og presenterer en utvidet beskrivelse av faktorene knyttet til selve risikovurdering. Som et bakteppe både for kunnskapsstatusen og risikovurderingen er målsetningen om å ha kysttorskbestander (Gadus morhua) som kan gi et høstbart overskudd, samt motstandsdyktighet mot nåværende og forventede klimaendringer. Dette vil si kysttorskebestander med god rekruttering, variert aldersstruktur og god genetisk status.
Lakseoppdrett i Norge
Norge eksporterte i 2025 i overkant av 1,4 millioner tonn oppdrettslaks (Salmo salar) og 86 903 tonn regnbueørret (Oncorhyncus mykiss) til en verdi på henholdsvis 124,67 og 7,42 milliarder kroner (tall fra Norges sjømatråd, januar 2026). Dette utgjorde 72,3 % av den samlede eksportverdien av sjømat på totalt 181,5 milliarder kroner.
Per i dag reguleres den langsiktige produksjonen i lakseoppdrettsnæringen av trafikklyssystemet for lakseoppdrett som ble innført av Nærings- og fiskeridepartementet høsten 2017. I dette reguleringssystemet vurderes den årlige dødelighet på grunn av lakselus hos førstegangsvandrende (post-smolt) villaks i 13 forhåndsdefinerte geografiske produksjonsområder (PO) (figur 1.1) som grunnlag for videre produksjonsvolum. Det er betydelig variasjon i produksjon mellom produksjonsområdene og de mest oppdrettsintensive områdene, basert på mengde fisk produsert per areal (tonn/km2 ) var i 2025 PO2 Ryfylke og PO3 Karmøy til Sotra, med henholdsvis 54,8 og 53,3 tonn/km2 (tabell 1.1). I andre enden av skalaen ligger PO1 Svenskegrense til Jæren og PO3 Øst-Finnmark med henholdsvis 2,4 og 3,7 tonn/km2 (tabell 1.1). Per 1. desember 2025 er det 980 godkjente lokaliteter i sjø for kommersielt oppdrett laks, regnbueørret og ørret (figur 1.1).
Den geografiske inndelingen av produksjonsområder sammenfaller relativt godt med forvalningsinndelingen av kysttorsk, hvor oppdrett i PO1 til PO4 i stor grad overlapper med kysttorsk sør for 62°N, oppdrett i PO5 til PO8 hovedsakelig overlapper med kysttorsk fra 62°N – 67°N og oppdrett i PO9 til PO13 stort sett overlapper med kysttorsk nord for 67°N (figur 1.1). Det er likevel 21 godkjente lokaliteter for oppdrett av laks og regnbueørret i PO8 som er nord for 67°N og tilsvarende to lokaliteter i PO4 som er i forvaltningsområdet 62°N – 67°N.
Kysttorsk
Torsk finnes i indre og ytre strøk langs hele norskekysten, med betydelig variasjon i adferdsmønstre og livshistorietrekk både innad og mellom ulike områder (Strøm mfl. 2023; Svåsand mfl. 2025). Bestandsstrukturen til torsk er svært kompleks langs norskekysten og begrepet kysttorsk brukes ofte for å beskrive stedbunden torsk som kan skilles fra de havvandrende torskebestanden: nordøstarktisk torsk (skrei) og nordsjøtorsk. I beskrivelsen av torsk langs norskekysten er begrepet økotype ofte brukt i den vitenskapelige litteraturen (f.eks. Knudsen mfl. 2018, Barth mfl. 2019, Strøm mfl. 2023), og kan defineres som en gruppe individers genetiske tilpasninger til spesifikt miljøforhold (Clemens & Schreck 2021). Økotype begrepet brukes derfor til å skille lokal kysttorsk fra skrei (Strøm mfl. 2023), men også til å skille mellom genetisk ulike torskestammer som tradisjonelt sett kategoriseres som kysttorsk (Barth mfl. 2019).
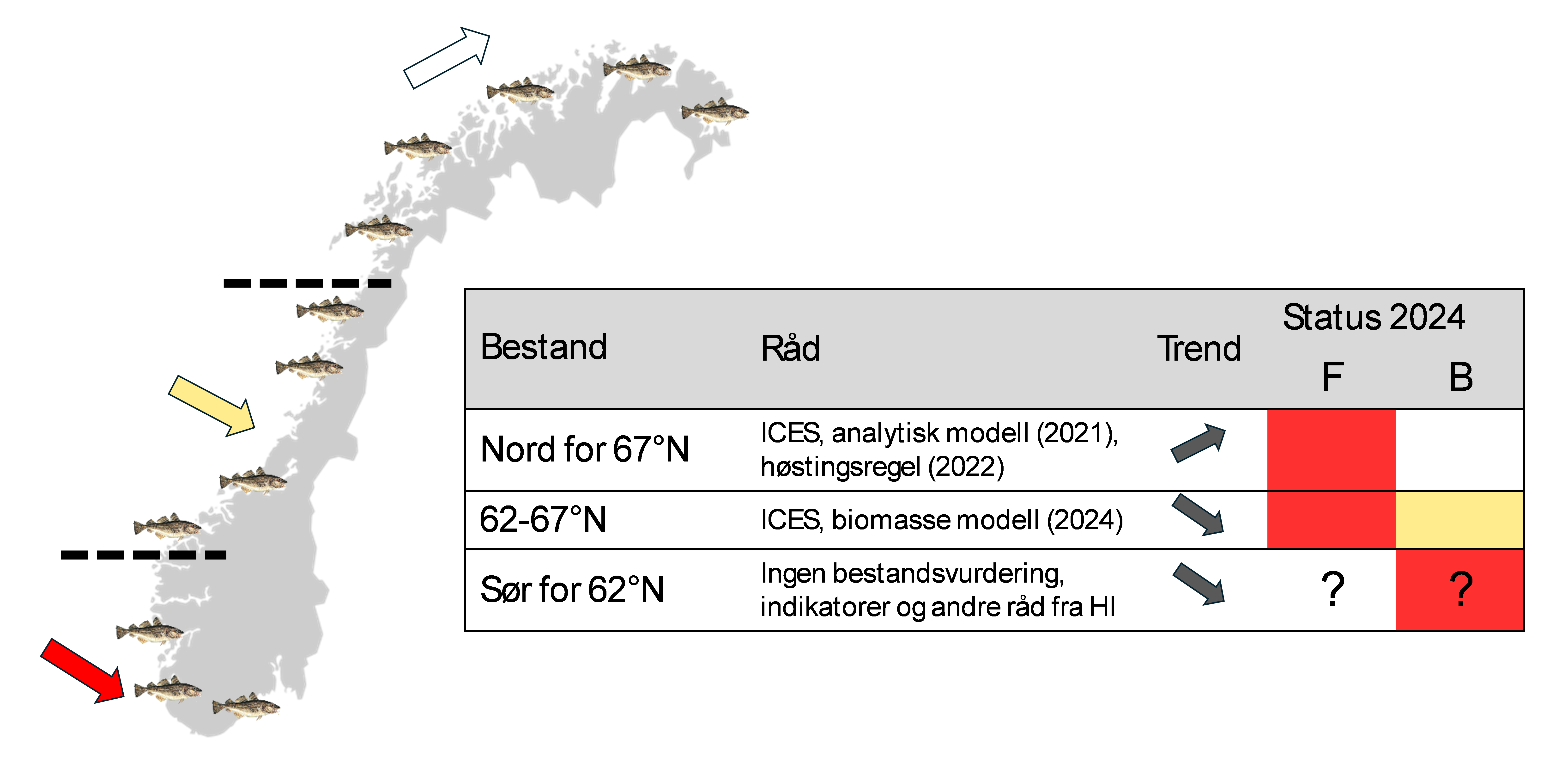
Frem til 2021 ble kysttorsk forvaltet i to geografiske områder: nord og sør for 62°N. Etter en metoderevisjon i 2021 ble det nordligste området delt i to ved 67°N (ICES 2021), og vi har nå tre forvaltningsområder (figur 2.1): sør for 62°N (Skagerrak og Vestlandet), 62°N – 67°N (fra Sildegapet like sør for Stadlandet til Fugløyfjorden sørvest for Bodø) og nord for 67°N (nordlige del av Nordland inkludert Lofoten og Vesterålen samt Troms og Finnmark). Kysttorsk i de tre områdene behandles som egne bestander med hensyn på overvåkning og bestandsvurdering (figur 2.1). De to nordligste bestandene har bestandsmodeller og kvoterådgivning gjennom ICES-systemet, mens det arbeides med å utvikle bestandsvurderinger for området sør for 62°N (Stock mfl. 2025).
Figur 2.1. Status for de tre kysttorsk bestandene med hensyn på fiskedødelighet (F) og biomasse (B) for 2025. Statuskategorier definert av ICES (ICES 2016). Grønn: biomasse over målverdi eller fiskedødelighet under målsetning. Gul: biomasse eller fiskedødelighet mellom målverdi og kritisk grense. Rød: biomasse under kritisk grense eller fiskedødelighet over kritisk nivå. Hvit: mål eller kritisk nivå ikke bestemt. Spørsmålstegn angir svakt datagrunnlag.
Kysttorsk fra nærliggende områder er mer genetisk lik enn kysttorsk som lever lengre fra hverandre, noe som tyder på isolering mellom ulike populasjoner langs kysten (Dahle mfl. 2018; Johansen mfl. 2020). Sør for 62°N er kysttorsken beslektet nordsjøtorsken. På Vestlandskysten beveger torsken seg mindre enn nordsjøtorsken og merkegjenfangst undersøkelser har vist at de aller fleste gjenfangstene er tatt mindre enn 10 km fra utsettingssted (Nedreaas mfl. 2008). Likevel er det en viss usikkerhet knyttet til om disse økotypene utgjør ulike bestander ettersom stedbunden torsk ofte er svært lik nordsjøtorsken genetisk (Stock mfl. 2025). Langs den norske delen av Skagerak er det påvist bred sameksistens mellom to genetisk adskilte økotyper: en nordsjølignende økotype, som er svært lik genetisk kjønnsmoden torsk fra Nordsjøen, og såkalt «fjordtorsk» som dominerer i beskyttede områder, vokser noe seinere, beveger seg mindre og er trolig tilpasset vann med lavere saltholdighet (Jorde mfl. 2018; Knudsen mfl. 2018; Barth mfl. 2019; Kristensen mfl. 2021).
I de to nordligste forvaltningsområdene (62°N – 67°N og nord for 67°N) sameksisterer kysttorsk med skrei. Denne sameksistens er størst under gyteperioden, men kan også forekomme andre tider på året, og det ble for eksempel i perioden 2021 – 2023 registrert 13 % skrei i torskefangsten nord for 67°N under Havforskningsinstituttets kysttokt i oktober-november (Johanna Fall pers. kom.). Genetisk innblanding med skrei øker nordover og har en viktig rolle for den genetiske variasjonen og bestandsstrukturen nord for 62°N (Breistein mfl. 2022). I flere fjorder i Troms og Finnmark er det påvist genetiske forskjeller mellom kysttorsk fanget i indre og ytre del av samme fjord, og mellom kysttorsk fanget i nærliggende fjordsystemer (Breistein mfl. 2022). Disse genetiske forskjellene er sterkt påvirket av ulik innblandingen av skrei og indikerer lokale kysttorskstammer (Breistein mfl. 2022). Videre har merke-gjenfangst data vist at avstand mellom gjenfangst- og utsettingssted hos torsk fanget langs norskekysten øker betydelig med breddegrad (Nedreaas mfl. 2008; Svåsand mfl. 2025). Dette tyder på at en mer vandrende kysttorsk lenger nord, noe som støttes av telemetriundersøkelser gjort i Finnmark som har vist at en betydelig andel av den umodne torsken forlater de lokale fjordsystemene (Strøm mfl. 2023).
Sør for 62°N gis det ikke et eget kvoteråd på kysttorsk, men Havforskningsinstituttet har gitt tekniske forvaltningsråd, slik som minstemål, maskevidde og verneområder for å redusere beskatningen (Nedreaas mfl. 2008; Aglen mfl. 2016; Moland mfl. 2021; Stock mfl. 2025). Fangsten av torsk innenfor 12 nautiske mil sør for 62°N trekkes fra i fangstregnskapet for torsk i Nordsjøen, og antas dermed å være utelukkende kysttorsk. Det fastsettes ikke egne fiskekvoter på de to bestandene av kysttorsk nord for 62°N, men kysttorskfangsten teller inn under den totale torskekvoten, som styres av størrelsen på skreibestanden. I årene 2022 – 2024 ble det i gjennomsnitt fanget 14 % kysttorsk i den totale norske torskefangsten nord for 67°N, mens i området 62°N – 67°N var den gjennomsnittlige andelen 87 % (ICES 2025a). En kysttorsk kan ikke uten videre skilles fra en skrei i felt, og det mangler tilstrekkelig infrastruktur og ressurser for å identifisere andelen kysttorsk i fangstene gjennom fangstsesongen. Kysttorskfangsten anslås derfor ved å analysere øresteiner (otolitter) fra fiskeriprøvetaking etter at fiskeåret er avsluttet. Det er også mulig å skille kysttorsk og skrei genetisk med få dagers svartid (Johansen mfl. 2018), og det foregår nå et prosjekt for å teste ut en slik «sanntidsovervåkning» i større skala. Kvoterådene for kysttorsk brukes allikevel som en rettesnor av forvaltningsmyndighetene for å vurdere behov for tekniske reguleringer som kan redusere fiskepresset på kysttorsken. Eksempel på reguleringer som har blitt innført de siste årene er økt minstemål for torsk innenfor 4 nautiske mil av grunnlinjen nord for 62°N, strengere begrensning på garnfisket innenfor fjordlinjene og havdeling der større fartøy må fiske lengre ut fra kysten (Regjeringen 2023a, Regjeringen 2023b, https://www.fiskeridir.no/Yrkesfiske/Nyheter/2024/styrker-vern-av-kysttorsken).
Kartlegging av gytefelt og oppvekstområder
Bærekraftig forvaltning av marine ressurser forutsetter ikke bare regulering av uttaket, men også beskyttelse av bestemte habitattyper og områder der artene lever. For mange fiskeriressurser befinner disse nøkkelområdene seg i kystnære strøk med til dels stor menneskelig påvirkning. Dette gjelder også for kysttorsk hvor menneskelige aktiviteter, som for eksempel etablering av akvakulturanlegg, vil kunne føre til fysiske og økologiske endringer av gytefelt og oppvekstområder som vil kunne påvirke både lokale bestander og større bestandskomplekser. Gjennom «Nasjonalt program for kartlegging av marine naturtyper» har Havforskningsinstituttet kartlagt gytefelt for kysttorsk langs hele norskekysten (Espeland mfl. 2013, figur 2.2). Denne kartleggingen består av vertikale håvtrekk gjennomført på forhåndsbestemte stasjoner, hvor mengden egg vurderes i sammenheng med simulering av strøm i det aktuelle området for å estimere tilbakeholdelse (retensjon) av egg. Produksjon, det vil si mengden egg, og retensjon gis så en verdi fra 0 til 3. Summen av disse verdiene gir en verdiindeks som klassifiserer den antatte viktigheten av et gytefelt, og hvor områder med en samlet verdiindeks på 1 eller retensjon lik 0, ikke regnes som gytefelt. Gytefelt med en verdiindeks på 6 klassifiseres som nasjonalt viktige siden disse både inneholder mye egg og har høy retensjon, mens gytefelt med verdiindeks på 5 klassifiseres som regionalt viktige. Gytefelt med en verdiindeks på 2 til 4 klassifiseres som lokalt viktige. Denne tilnærmingen gjør det mulig å identifisere små gytefelt med høy retensjon som biologisk viktige, noe som er svært relevant i en forvaltningssammenheng ettersom disse vil være mer sårbare for inngrep. Til sammenligning, kommer denne klassifiseringstilnærmingen til kort når det gjelder gytefelt som bidrar med store mengder egg over større områder. Dette gjelder spesielt for store fjordsystemer nord for 67°N hvor retensjon av egg ikke nødvendigvis bør vektes likt som produksjon ettersom disse kan bidra betydelig til den samlede rekrutteringen over større områder. I tillegg har Fiskeridirektoratet gjennomført intervjuundersøkelser for å kartlegge gyteområder. Denne kartleggingen inkluderer også områder som ikke dekkes av gytefeltkartleggingen gjort av Havforskingsinstituttet, i hovedsak en del ytre kystområder inkludert Lofoten og Vesterålen. I forvaltningen benyttes begge disse undersøkelsene (gytefelt og gyteområder) for å identifisere sjøarealer som kan være viktige for gyting.
Figur 2.2. Kartet viser områder (rød farge) som Havforskningsinstituttet har undersøkt med hensyn til kartlegging av gytefelt i «Nasjonalt program for kartlegging av marine naturtyper» siden 2007. For mer detaljert beskrivelse, se “Gytefelt torsk MB dekningskart” under Temalag “Kystnære fiskeridata” i Fiskeridirektoratets kartverk «Yggdrasil».
En viktig begrensning i denne kartleggingen er at de fleste gytefeltene kun er undersøkt en gang i en gytesesong, og bare et begrenset antall av gytefeltene er undersøkt i flere gytesesonger. To gytefelt i Vest-Finnmark (Repparfjorden og Revsbotn) har imidlertid vært gjenstand for en grundigere overvåkning med tre besøk i gytesesongene (april-mai) fra 2015 til 2025 (van der Meeren 2020, pers. kom.). Denne tidsserien viser at eggmengde kan variere betydelig både innad i og mellom gytesesonger (figur 2.3).
Figur 2.3. Variasjon i gyting gjennom gytesesong fra to fjorder i Vest-Finnmark fra 2015 til 2025. Antall egg pr. håvtrekk for egg i samme størrelse som torskeegg (1,2-1,65 mm diameter) er gitt som gjennomsnitt for 17 faste stasjoner i Repparfjorden og 15 faste stasjoner i Revsbotn. Data viser mengde yngre egg (stadium 1) og eldre egg (stadium 2 til 5).
Havforskningsinstituttets kartlegging av gytefelt ble primært gjennomført inne i fjorder og på indre deler av kysten. Som følge av dette er flere områder i ytre strøk, blant annet svært viktige gyteområder som Lofoten, Vesterålen og Senja, utelatt (figur 2.2). For disse områdene representer den intervjubaserte gytefeltkartleggingen gjort av Fiskeridirektoratet den best tilgengelige kunnskapen. Selv om det ofte er god overenstemmelse mellom informasjon gitt av fiskerne og data fra feltinnsamling av egg med tilhørende oseanografisk modellering, er det i enkelte tilfeller avvik mellom disse to metodene. For eksempel kan intervjubaserte gytefelt reflektere gode fiskeplasser for gytemoden fisk og ikke nødvendigvis områder hvor fisken faktisk gyter. Alternativt, kan intervjubasert undersøkelser gi historisk informasjon som er vanskelig å skaffe gjennom feltundersøkelser. Alle kartlagte gytefelt er tilgjengelig gjennom Fiskeridirektoratets kartløsning Yggdrasil under «Fiskeri» og temalaget “Kystnære fiskeridata”. Her viser kartlaget “Gytefelt Torsk MB” gytefelt kartlagt gjennom «Nasjonalt program for kartlegging av marine naturtyper», og kartlaget “Gyteområder torsk” viser gyteområder basert på Fiskeridirektoratets intervjuundersøkelser.
I motsetning til gytefelt, er oppvekstområder for kysttorsk i svært liten grad kartlagt. En av grunnen til dette er at torsk oppholder seg i et bredt spekter av habitater gjennom livet, og viktigheten av ulike områder varierer både med alder og sesong, også for de yngre livsstadiene (Dunlop mfl. 2022, Nickel mfl. 2025). Til gjengjeld har «Nasjonalt program for kartlegging av marine naturtyper» kartlagt forekomst av ålegressenger, tareskog og andre viktige naturtyper som kan fungere som oppvekstområder for kysttorsk. I tillegg gjøres undersøkelser av viktigheten til ulike habitatstyper som oppvekstområder for fiskeyngel på utvalgte steder gjennom prosjektet «Kartlegging av gytefelt og oppvekstområder for kommersielt viktige arter i kystsonen» (Espeland mfl. 2022). Resultater fra disse undersøkelsene, samt andre undersøkelser ledet av Havforskningsinstituttet, tyder på at flere ulike habitater som eksempelvis grunne sand- og grusområder med rødalger, og områder med brunalger også kan være viktige oppvekstområder for de yngste aldersgruppene (0+ og 1+) kysttorsk (Skaar 2021, Dunlop mfl. 2022). Disse naturtypene er i motsetning til ålegressenger og tareskog i liten grad kartfestet. For eldre ungfisk (≥ 2+) har en nytt studie med presis posisjonering av torsk ved bruk av akustisk telemetri vist en tydelig preferanse for skrånende og grunne bergområder (Gjelland mfl. i trykk), sannsynligvis relatert til forekomst av tareskog. Med tilsvarende metodikk er det også vist at størrelsen på torsken hjemmeområde øker med størrelse (Olsen mfl. i trykk).
Produksjonsområde 1 – 4, Svenskegrensen til Stad, forvaltningsområde sør for 62°N
Lakseoppdrett
Fra 2018 til 2024 har den totale årlige produksjonen av oppdrettslaks og regnbueørret i produksjonsområdene som overlapper med forvaltningsområdet for kysttorsk sør for 62°N økt med 30 % fra 373 000 – 486 000 tonn (figur 2.4). Dette tilsvarer en årlig vekst på 3,9 %. Produksjonen har vært relativt stabil siden 2019, da den har variert mellom 409 000 tonn i 2019 og 494 000 i toppåret 2024 (figur 2.4). Høyest produksjon av laks og regnbueørret forekommer i PO4 og PO3 (179 000 – 190 000 tonn), og høyest produksjonstetthet forekommer i PO3 og PO2, med henholdsvis 53 og 55 tonn/km2 (tabell 1.1). Til sammenligning er det lite lakseoppdrett i PO1, hvor tilnærmet all produksjonen på omtrent 9 000 tonn forekommer vest for Lindesnes.
Figur 2.4. Årlig produksjon av oppdrettslaks og regnbueørret for de ulike forvaltningsområdene for kysttorsk. Data er basert på tall fra Fiskeridirektoratet (20.01.2025).
Status kysttorsk sør for 62°N
Det finnes ikke en formell bestandsvurdering for kysttorsk sør for 62°N, men Havforskningsinstituttet har nylig publisert en rapport med oppdatert status for torsk i dette forvaltningsområdet (Stock mfl. 2025). Alle tilgjengelige data viser at bestanden er i svært dårlig forfatning. For eksempel viser fangst data fra referanseflåten en nedgang på 75 – 80 % i Skagerrak og på 50 % på Vestlandet siden 2006 (figur 2.5). Den negative utviklingen i Skagerrak tydeliggjøres ytterligere med data fra Havforskningsinstituttet garntokt, hvor det i Øst-Skagerrak ble observert en markant nedgang på 1990-tallet. Til sammenligning var torskebiomassen i Vest-Skagerrak relativ stabil før den gikk kraftig ned fra 2017 (figur 2.5). I dag er derfor biomassen av torsk antatt å være på historisk lave nivåer i begge områdene.
Figur 2.5. Fangst-per-enhet-innsats for garn for referanseflåten i perioden 2006 – 2024 (øverst) og fra garntokt i Skagerrak i periodene 1984 – 1990, 2001 – 2008 og 2010 – 2024 (nederst). Fargede felt viser 95 % konfidensintervall. Figur er er hentet fra Stock mfl. ( 2025 ).
Videre viser de kommersielle fangstene av torsk innenfor 12 nautiske mil av grunnlinjen en nedgang på ca. 90 % de siste 25 årene, sammenlignet med historiske fangster på 2000 – 4000 tonn fra 1920- tallet til rundt år 2000. Det er lite som tyder på at vekstforholdene (individvekst) begrenser torskebestandene sør for 62°N, ettersom vekt ved alder har økt i alle regioner for de fleste aldersgrupper. Merke-gjenfangst analyser har vist at fiskedødelighet (F) overgår naturlig dødelighet både i Skagerrak og på Vestlandet. Av mulige påvirkningsfaktorer (klimaendringer, vekst, rekruttering, naturlig dødelighet og fiskedødelighet) ansees fiskedødelighet og fangst av umoden fisk som sannsynligvis den viktigste årsaken til den observerte nedgangen i torskebestanden sør for 62°N (Stock mfl. 2025). Det vurderes som lite sannsynlig at nedgang i rekruttering er en viktig faktor hverken på Vestlandet eller i vest-Skagerrak, men det kan være det i øst-Skagerrak. Dårlig nylig rekruttering i øst-Skagerrak kan skyldes utarming av lokale gytere, mindre tilstrømning av larver fra Nordsjøen, miljøfaktorer eller en kombinasjon av disse faktorene.
Overlapp mellom oppdrettslokaliteter og gytefelt
For kystområdene i PO1 – PO4 er det en viss overlapp mellom viktige gytefelt for kysttorsk og oppdrettslokaliteter for laks og regnbueørret. Dette gjelder for Hornelsfjorden i Bremanger der oppdrettsanlegg er plassert i et nasjonalt viktig gytefelt, samt i Førdefjorden og fjordene rundt Osterøy, hvor det er en rekke oppdrettslokaliteter i regionalt viktige gytefelt.
Fra 2018 til 2025 har produksjonen av oppdrettslaks og regnbueørret i produksjonsområdene fra Stad til Bodø økt med 28 %, fra ca. 559 000 tonn til litt i overkant av 715 000 tonn (figur 2.4). Dette tilsvarer en årlig vekst på 3,6 %. Her er produksjonsvolumet desidert størst i PO6, som med sine 236 000 tonn representer det største volumet av alle produksjonsområdene (tabell 1.1). Produksjonstettheten av oppdrettslaks varierte i 2025 mellom 25 og 31 tonn/km2 for de tre produksjonsområdene sør for Helgeland (PO5 – PO7, tabell 1.1). I produksjonsområde 8 (Helgeland til Bodø) var mengden fisk produser per arealenhet betydelige lavere med 15 tonn/km2 (tabell 1.1).
Status kysttorsk 62°N – 67°N
Kysttorsk 62°N – 67°N er definert som kategori 2-bestand av ICES. Det betyr at bestandsvurderingen er godkjent med hensyn til relativ fisketrykk og biomasse, men ikke absolutte estimater av bestandens størrelse (B). Bestandsvurderingen gjøres ved en produksjonsmodell (Pedersen & Berg 2017). I en slik modell er relativ fiskedødelighet og den relative biomassen to sentrale elementer. Relativ fiskedødelighet gis ved å dele fiskedødelighet (F) på den fiskedødeligheten som gir det maksimale langtidsutbyttet (Fmsy), det vil si det fisket som er bærekraftig over tid. Ideelt sett bør dette forholdet (F/Fmsy) være mindre eller lik 1 og det anses som kritisk hvis den relative fiskedødeligheten overskrider 1,7. Tilsvarende er den relative biomassen av en bestand den faktiske biomassen (B) delt på biomassen som resulterer fra fiske ved Fmsy (Bmsy). Dette forholdet (B/Bmsy) bør være større eller lik 0,5 og 0,3 anses som kritisk nedre grense. Når bestanden når kritisk grense betyr det at gytebiomassen er så lav at bestandens rekruttering påvirkes negativt. Det er viktig å presisere at i denne type modeller refererer både relativ fiskedødelighet og biomasse til den fiskbare delen av bestanden. Det vil si biomassen av fisk med lengde over 50 cm, som omtrent tilsvarer lengden der 50 % er kjønnsmodne.
Bestandsmodellen inkluderer fangstdata fra kommersielt fiske, turist/fritidsfiske, samt biomasseindekser fra Havforskningsinstituttets kysttokt (akustikkdata og trålfangster) og garn-ruse tokt. Både kommersielle fangster og tetthetsindikatorer fra Havforskningsinstituttets overvåkningstokt har vist en tydelig nedgang det siste tiåret. Den relative fiskedødeligheten (F/Fmsy) økte fra midten av 1990-tallet og frem til 2008 hvor den flatet noe ut. For 2024 er denne estimert til å være 1,80 med 95 % konfidensintervall på 1,21 – 2,67 (figur 2.6), det vil si over bærekraftig nivå) (ICES 2025b). Dette gjenspeiler seg i den relative biomassen (B/Bmsy) som er mer enn halvert siden starten av 1990-tallet og for 2024 er estimert til 0,39 med 95% konfidensintervall på 0,25 – 0,60 (figur 2.6) (ICES 2025b). For 2025 forventes en total fangst på 6 834 tonn (ICES 2025a). Dette er over tre ganger høyere enn ICES sitt samlede råd for kommersielt-, turist- og fritidsfiske på 1 926 tonn for bærekraftig høsting for 2025 (ICES 2024) og omtrent 6 ganger høyere enn samlet råd for 2026 (ICES 2025b).
Det største problemet med denne bestandsvurderingen er den høye usikkerheten rundt fangstene i turist- og fritidsfiske ettersom dette fisket sannsynligvis utgjør mer enn halvparten (ca. 60 %) av de totale fangstene. Usikkerheten kan ikke kvantifiseres og er ikke inkludert i modellen. Fangstdata er kritisk for modellen ettersom det i stor grad bestemmer nivået på bestanden, mens trendene påvirkes av toktindeks. Når i tillegg sensitivitetstester av bestandsmodellen viser at absolutte estimater av bestandsstørrelse og fiskeridødelighet (F) er mye mer følsomme enn når disse utrykkes relativt Bmsy/Fmsy, og disse referansepunktene oppdateres hver gang modellen kjøres (se også ICES 2025a), er det åpenbart lav presisjon i bestandsestimatet for dette området.
For å kunne vurdere risiko fra lakseoppdrett for bestanden er del likevel behov for et anslag av biomassen. Fra modellen får vi et estimat på 30 000 tonn (95% konfidensintervall: 12 900 – 69 800) for 2024. Dette vil være et maksimalt estimat av gytebiomassen, da modellen definerer høstbar biomasse som fisk over lengde 50 cm, som omtrent motsvarer lengden der 50 % er kjønnsmodne. Historiske toktdata viser at omtrent 75 % av kysttorsken i de to bestandsområdene befinner seg nord for 67°N (Aglen mfl. 2020). I senere tid har bestandsutviklingen vært mer positiv for kysttorsken i dette området sammenlignet med området 62°N – 67°N der trenden er synkende, og andelen kysttorsk nord for 67°N har derfor økt (ICES 2025c). Ser vi på de totale fangstene (kommersielle + rekreasjonsfangster) i de to forvaltningsområdene så ligger området 62°N – 67°N på ca. 20 % av nord for 67°N. Gitt proporsjonalitet mellom bestandsstørrelse og fangster vil dette tilsi en gytebestandsstørrelse i dette forvaltningsområdet på i underkant av 20 000 tonn.
Figur 2.6. Relativ fiskedødelighet (F/ Fmsy ) og relativ biomasse (B/BMSY) fra bestandsvurderingen av kysttorsk 62°N – 67°N gjort i 2025 (ICES 2025b ). Mørkegrønne linjer viser estimater, grå områder viser 95 % konfidensintervall. Oransje linjer viser grenseverdier for bærekraftig høsting (F/ Fmsy = 1 og B/ Bmsy = 0,5), mens de røde viser kritiske grenser (F/ Fmsy = 1,7 og B/ Bmsy = 0,3).
Overlapp mellom oppdrettslokaliteter og gytefelt
For kystområdene inkludert i PO5 – PO8 forekommer det overlapp mellom viktige gytefelt for kysttorsk og lakseoppdrett. Dette gjelder for eksempel det nasjonalt viktige gytefeltet Nordfjorden – Beiarkjeften, hvor den samlede oppdrettskapasiteten i området er 36 000 tonn, inkludert 15 000 på selve gytefeltet (Fall mfl. 2024). I tillegg har områdene rundt Frøya og Hitra flere lokalt viktige gytefelt og totalt 43 oppdrettsanlegg (Fall mfl. 2024).
Produksjonsområde 9 – 13, Vestfjorden til Øst-Finnmark, forvaltningsområde nord for 67°N
Lakseoppdrett
Fra 2018 til 2024 har produksjonen av oppdrettslaks og regnbueørret nord for Bodø økt med 59 %, fra ca. 387 000 til omtrent 616 000 tonn (figur 2.4). Dette tilsvarer en årlig vekst på 8 %. Denne betydelige veksten påvirkes sterkt av en samlet produksjonsøkning på 27 % fra 2024 til 2025 (figur 2.4). I PO9 (Vestfjorden og Vesterålen) og PO12 (Vest-Finnmark) var produksjonstettheten litt under 12 tonn/ km2 i 2025 (tabell 1.1), mens den i PO11 (Kvaløy til Vest-Finnmark) var på 19 tonn/ km2 (tabell 1.1). Av produksjonsområdene nord for 67°N produseres det desidert mest fisk per arealenhet i PO10 (Andøya til Senja) med en produksjonstetthet på 34 tonn/ km2. I PO13 (Øst-Finnmark) ble til sammenligning kun produsert 3,7 tonn/km2 i 2025.
Status kysttorsk nord for 67°N
Datagrunnlaget for bestandsvurdering av kysttorsk nord for 67°N er bedre enn for kysttorsk i området 62°N – 67°N. Den samlede bestanden nord for 67°N er klassifisert som en kateogri-1 bestand av ICES, som gir sin bestandsvurdering og kvoteråd basert på en aldersstrukturert bestandsmodell (ICES 2021). Denne modellen har påvist en svak sammenheng mellom størrelse på gytebestanden og rekruttering, noe som kan tyde på varierende trender i rekruttering til lokale populasjoner innenfor bestandsområdet (ICES, 2022a). Som følge av dette er det ikke fastsatt en kritisk grense for biomasse i gytebestanden (Blim), et referansepunkt som normalt brukes som en del av høstingsregelen. En alternativ høstingsregel ble derfor foreslått og testet, der det ble beregnet en fiskedødelighet som var «føre-var» i hele spennet av observerte gytebestandsstørrelser og varierte lite mellom ulike antakelser om blant annet nivået på Blim (ICES, 2022b). Denne høstingsregelen, som ofte blir referert til som «Den norske forvaltningsplanen», ble vedtatt av Norge ved Nærings- og fiskeridepartementet og brukt for første gang i kvoterådet for 2023.
Siden midten av 1990-tallet har fiskedødeligheten (4 – 8 åringer) for kysttorsk nord for 67°N for det meste vært langt over anbefalt nivå. Etter en periode med lavere fiskedødelighet i 2011-2015, som følge av lavere fangster og gode årsklasser på slutten av 90-tallet, med påfølgende vekst i bestanden, har fiskedødeligheten igjen økt det siste tiåret (figur 2.7). Fiskedødelighet for yngre årsklasser (i.e., 3 – 6 åringer) var spesielt høy i 2023 før den i 2024 returnerte til et nivå i bedre samsvar med de siste års gjennomsnitt (ICES 2025c). Denne reduksjonen i dødelighet for ungfisk sammenfalt med en økning i minstemålet fra 44 til 55 cm, som tredde i kraft 1. januar 2024. En sterk 2020-årsklasse kombinert med økt vekst og sannsynlighet for kjønnsmodning hos yngre fisk (3 – 6 åringer) bidro til den relativt markante økningen i gytebiomasse fra 2023 til 2024 (ICES 2025a). De kommersielle fangstene av eldre fisk (7+) var mye lavere i 2024, noe som sannsynligvis har sammenheng med særdeles lav biomasse av disse aldersklassene i bestanden (ICES 2025a). For 2024 var gytebiomassen estimert til ca. 85 000 tonn (95 % konfidensintervall: 60 000 – 120 000 tonn), mens den totale biomassen var anslått å være ca. 276 000 tonn (95 % konfidensintervall: 206 000 – 369 000 tonn) (ICES 2025a). I 2024 ble totale fangsten estimert til ca. 36 000 tonn, hvorav rundt 80 % stammet fra kommersielt fiske (ICES 2025c). Dette tilsvarte en samlet reduksjon på 32 % sammenlignet med 2023 (ICES 2025c). Det er uvisst hvordan den reduserte torskekvoten, som reflekterer nedgangen i skreibestanden, vil slå ut på kysttorskfangstene i tid og rom. For 2026 har ICES anbefalt at fangsten av kysttorsk nord for 67°N ikke bør overskride 27 887 tonn (ICES 2025c).
Figur 2.7. Fiskedødelighet og gytebiomasse fra bestandsvurderingen av kysttorsk nord for 67°N (ICES 2025c). Mørkegrønne linjer viser estimater, grå områder viser 95 % konfidensintervall. Oransje linjer viser grenseverdi for fiskedødelighet (F) i Den norske forvaltningsplanen og grenseverdi for gytebiomasse (SSB).
Overlapp mellom oppdrettslokaliteter og gytefelt
En stor del av produksjonen nord for 67°N overlapper ikke med områder Havforskningsinstituttet har undersøkt i gytefeltkartleggingen for kysttorsk (figur 2.2). For disse områdene, som blant annet inkluderer Lofoten, Vesterålen og vestsiden av Senja, stammer tilgjengelig kunnskap om gyteområder fra intervjuer med fiskere. Til tross for at det er en viss usikkerhet knyttet til det samlede geografiske overlappet mellom lakseoppdrett og viktige gyteområder for kysttorsk nord for 67 °N, er det flere tilfeller hvor lakseoppdrett forekommer enten på, eller i nærheten av, svært viktige gytefelt. To eksempler på dette er Kvænangen og Revsbotn. I Kvænangen er det en samlet oppdrettskapasitet på 46 000 tonn, inkludert 24 000 tonn som er innenfor et regionalt viktig gytefelt til tross for at deler av fjorden er klassifisert som nasjonal laksefjord (Fall mfl. 2024). I Revsbotn er det to oppdrettsanlegg plassert innenfor et nasjonalt viktig gytefelt, med en samlet oppdrettskapasitet på 11 000 tonn (Fall mfl. 2024).
Risikokilder som kan gi negative effekter av lakseoppdrett på ville kysttorskbestander
I denne delen gir vi en beskrivelse av de ulike risikokildene og hendelsene knyttet til påvirkninger fra lakseoppdrett i åpne merder på ville kysttorskbestander. I den sammenhengen er kunnskap om naturlig vandring viktig ettersom store miljømessige endringer som tilstedeværelsen av oppdrettsanlegg kan påvirke denne. Vandringsadferden til torsk kan deles inn i fire hovedkategorier: 1) stasjonære individer som oppholder seg i samme begrensede område gjennom hele livet, 2) presise tilbakevandrere som vender tilbake til et spesifikt område flere ganger, 3) upresise tilbakevandrere som vender tilbake til et større geografisk område i løpet av livet og 4) spredere som tilsynelatende beveger seg bort, i et tilsynelatende tilfeldig mønster (Robichaud og Rose, 2004). Selv om mange kysttorskbestander kan klassifiseres som stasjonære (Knutsen mfl. 2018) er det er det stor variasjon i vandringsmønster, og i enkelte områder forlater en betydelig andel av den lokale bestanden fjordområdet (Strøm mfl. 2023). Mye tyder på at denne bestandsvariasjonen har en geografisk komponent styrt av oseanografiske forhold, med en større andel vandrende individer i mer åpne nordlige fjorder, hvor det kan forekomme genetisk adskilte bestander i indre og ytre deler av fjordsystemene (Breistein mfl. 2022). I tillegg til variasjon i vandringsmønster mellom bestander og individer, avhenger vandringsadferden også av fiskens livsstadium. Flere studier har vist at områdebruken til torsk øker med fiskestørrelse (Olsen mfl. 2023; i trykk; Strøm mfl. 2023; Nickel mfl. 2025), noe som kan forklares med at svømmeevne, metabolisme og fiskens posisjon i næringsnettet endres jo større den blir. I sum tilsier dette at vandringsadferd hos torsk styres av genetikk, miljømessige forhold, størrelse og fysiologisk status.
Endring i adferd hos kysttorsk
Tiltrekning til laksemerder
Oppdrettsanlegg fungerer som ansamlingsinnretninger for marine organismer (Sanchez-Jerez mfl. 2011, Uglem mfl. 2014, Callier mfl. 2018, Meier mfl. 2023), noen som ofte fører til høyere diversitet av dyreliv rundt disse, hovedsakelig drevet av økt forekomst av fisk (Barret mfl. 2019). Det er flere årsaker til hvorfor fisk og andre marine organismer oppholder seg rundt oppdrettsanlegg og der hvor enkelte arter og livsstadier tiltrekkes ved at de fysiske strukturene tilbyr ly og habitat (Sanchez-Jerez mfl. 2011), er tiltrekningen for andre drevet av økt mattilgang (Fernandez-Jover mfl. 2011). Videre kan også operasjonelle aktiviteter og regimer påvirke, og det er blant annet vist at bruk av kunstig belysning i lakseoppdrett kan tiltrekke seg både hvirvelløse dyr (evertebrater) og fisk i ulike livsstadier (McConnel mfl. 2010).
Langs norskekysten er det påvist at spesielt sei (Pollachius virens) samler seg rundt oppdrettsanlegg, men også andre arter, som torsk, kan forekomme i høye antall (Dempster mfl. 2009; 2010; Dunlop mfl. 2024; Keeley mfl. 2025). Telemetriundersøkelser gjort i oppdrettsintensive områder rundt Smøla, viste at rundt halvparten av torsken oppholdt seg ved anleggene (Skjæraasen mfl. 2021). Av torsken som var forbundet med oppdrett, var de fleste kun ved anleggene over en kort periode (< 1 uke), mens 16 % (dvs. 9 % av den undersøkte delen av bestanden) oppholdt seg i nærheten av laksemerden i mer enn 1 måned (Skjæraasen mfl. 2022). Dette tyder på vedvarende tiltrekning til lakseoppdrettsanlegg for en del av bestanden. Det ble videre observert at flere torsk befant seg i nærheten oppdrettsanlegg hvor det var laks (Skjæraasen mfl. 2022), noe som støtter forventningen om at økt mattilgang sannsynligvis er den viktigste faktoren for denne tiltrekningen hos torsk (Meier mfl. 2023). Likevel ble det ikke påvist at tiltrekningen til lakseoppdrettsanlegg endret torskens valg av gyteområder, da fisken holdt seg på de lokale gyteplassene i uker til måneder gjennom gyteperioden (Skjæraasen mfl. 2021).
Til tross for kunnskap om at enkelte torsk tiltrekkes lakseoppdrettsanlegg, og at dette kan omfatte en betydelig del av bestanden, er det fortsatt usikkert hvor sterkt torsk tiltrekkes lakseoppdrettsanlegg på en større skala. For eksempel har kameraundersøkelser vist at hvorvidt torsk aggregerer nært merdene kan variere mellom anlegg (Demster mfl. 2010). Videre kan det tenkes at tiltrekningen påvirkes av vandringsmønster på lokal skala, med ulik grad av tiltrekning for stasjonær og mer vandrende kysttorskbestander.
Frastøting fra områder med lakseoppdrett
Det at torsk kan tiltrekkes områder med lakseoppdrett i åpne merder, ekskluderer ikke muligheten for eventuell frastøting fra de samme områdene. I en telemetri undersøkelse gjort rundt Smøla ble det vist svært varierende grad av tiltrekning til lakseoppdrettsanlegg. Av den merkede torsken ble litt under halvparten ikke observert i nærheten av anleggene, mens 9 % opphold seg ved anleggene over lengre perioder (Skjæraasen mfl. 2022). I hvilken grad individuell variasjon i adferd er bestemmende for tiltrekking eller unnvikelse er uklart, men slike forskjeller i individuell adferd kan forventes å forekomme på bestandsnivå. Videre, kan det tenke seg at hvordan kysttorsken responderer på tilstedeværelse av lakseoppdrett avhenger av den lokale bestandens dominerende vandringsstrategi, og i flere områder langs norskekysten har fiskere lenge hevdet at vandrende kysttorsk unngår gytefelt i oppdrettsintensive fjorder. I en studie gjort av Maurstad mfl. (2007) ble det konkludert med at slike observasjoner må ansees som pålitelige, men at det trengs naturvitenskaplige studier for å konkludere med større sikkerhet.
Tilstedeværelsen av lakseoppdrettsanlegg kan påvirke torskens valg av gyteområder enten ved at vandrende individer ikke vender tilbake til påtenkte gytefelt eller ved at mer stasjonære individer frastøtes lokale gyteområder som følge av arealkonflikt med oppdrettsmerder. Begge tilfeller kan føre til at torsken gyter i et mindre gunstig område. Videre kan også endringer i vandringsmønster mellom gytefelt ha en negativ effekt på bestandsrektrutteringen. Det er vist at store hunntorsk benyttet seg av flere gytefelt i samme gyteperiode (Olsen mfl. 2023). Ettersom retensjonen og overlevelsen til egg ved gitte gytefelt vil kunne variere mellom år, vil det at de største individene med størst reproduktiv kapasitet benytter seg av flere gytefelt i samme gytesesong kunne være stabiliserende for bestanden.
For å undersøke muligheten for at kysttorsk unngår områder med lakseoppdrett har det blitt gjennomført en serie med laboratorieeksperimenter med hypotesen om at lukt spiller en viktig rolle i valg av område (Sæther mfl. 2007). Her ble det vist at torsk, antatt å være naiv til oppdrett, unngikk vanntanker med vann fra kar med oppdrettslaks. Denne adferden var ikke artsspesifikk ettersom liknende respons også ble observert ved eksponering for vann fra tanker med oppdrettstorsk. Videre ble det også vist en svakere frastøting for vill torsk fanget i nærheten av oppdrettsanlegg, samt en manglende adferdsrespons hos oppdrettstorsk eksponert for laks. Dette kan tyde på at frastøtende adferd fra områder med høye fisketettheter kan endres ved eksponering.
Som en videreføring av disse laboratorieeksperimentene ble adferden til gytevandrende kysttorsk i Finnmark undersøkt under naturlige forhold ved bruk av akustisk telemetri (Bjørn mfl. 2009). Her ble gyteklar kysttorsk fanget på innsig inn i en oppdrettsintensiv fjord, hvor halvparten av de merkede fiskene ble satt ut på fangststedet, mens resten ble transportert inn til det oppdrettsintensive området. I hver gruppe ble halvparten av fisken luktblokkert. Dette hadde ingen innvirkning på resultatene, da nesten all fisk satt ut i ytre del snudde og forlot fjorden innen en uke, mens majoriteten av torsk satt ut i det oppdrettsintensive området ble værende i ca. 3 uker (Bjørn mfl. 2009). Som følge av den manglende sammenhengen mellom luktesans og vandringsadferd, frembrakte studien ingen bevis på at gytevandrende kysttorsk unngår områder med oppdrett basert på lukt. Riktignok motstrider resultatene heller ikke fiskernes observasjoner om at torsken ikke lenger går inn til gytefeltet i fjorden.
I en telemetriundersøkelser gjennomført rundt Smøla ble det heller ikke påvist at torsk unngår gytefelt i oppdrettsintensive områder (Skjæraasen mfl. 2021, 2022). Her ble gytemoden kysttorsk fanget på gyteplassen og merket med akustiske sendere. Resultatene viste at flere torsk ble registrert ved gyteplassene enn ved oppdrettsanleggene, selv om de kunne tilbringe mye tid ved anleggene etter endt gyting (Skjæraasen mfl. 2021, 2022). Videre påvirket ikke nærheten til oppdrettsanleggene tilstedeværelsen av torsk ved gytefeltene (Skjæraasen mfl. 2021). Dette, kombinert med observasjoner av at gytingen foregikk på gytefeltene og ikke ved oppdrettsanleggene, tyder på at tilstedeværelsen av lakseoppdrett ikke hindrer torsken i å gyte på naturlige gyteplasser på lokal skala (Skjæraasen mfl. 2021). Dette burde ideelt sett blitt fulgt opp ved telemetristudier og merking av fisk fanget både ved gyteplasser og anlegg i samme område.
Endring i fysiologi hos kysttorsk
Torsk er en generalist som endrer diett gjennom livsløpet. Generelt sett består diettene til torsk under 20 cm på grunt vann i kystsonen hovedsakelig av ulike krepsdyr og små fisk (Arnott & Pihl 2000). Tilsvarende størrelse torsk hos de havgående bestandene spiser krill og andre små pelagiske byttedyr, men med større innslag av bunndyr og større byttefisk med økende størrelse (Link & Garrison 2002; Holt mfl. 2019). Videre varierer også torskens diett som en funksjon av byttedyrtetthet (Link & Garrison 2002). Denne fleksibiliteten kommer tydelig frem i diettundersøkelser i relasjon til nærhet til oppdrettsanlegg. Torsk tilknyttet laksemerder spiser mer spillfôr enn fisk fanget lengre bort fra anleggene (Dempster mfl. 2011, Fernandez-Jover mfl. 2011). Utnyttelsen av spillfôr kan variere med fiskestørrelse, og i en undersøkelse gjort av Sæther mfl. (2012) ble det påvist at for torsk større enn 60 cm var det færre individer med pellets i mageinnholdet (11 %) sammenlignet med mindre individer (32 %). Videre har en rekke studier vist at forekomsten av byttedyr i torskens diett endrer seg markant med hensyn på avstand til oppdrettsanlegg, enten i form av en økende andel fisk og flerbørstemark (Dempster mfl. 2011, Fernandez-Jover mfl. 2011) eller ved en endring i fiskearter og et større innslag av sediment-levende bunndyr (e.g., flerbørstemark, slangestjerner og muslinger) typisk forbundet med oppdrett (Keeley mfl. 2025).
Fôrsammensetning som brukes i norsk lakseoppdrett, har endret seg betraktelig de siste tiårene. I perioden 1990 til 2020 minket andelen marine ingredienser fra 90 til 25 %, mens andelen plantebaserte ingredienser økte fra ubetydelige nivåer til over 70 % (Aas mfl. 2022). Dette skyldes at fiskeoljer har blitt erstattet av rapsolje som inneholder svært høye nivåer av terrestriske fettsyrer som oljesyre (18:1n-9), linolsyre (18:2n-6) og α-linolensyre (18:3n-3), men mangler de lang-kjedede flerumettede fettsyrene som finnes i marine fiskeoljer (Bell mfl. 2003). Som følge av dette vil torsk som spiser spillfôr og byttedyr forbundet med oppdrett ha en diett bestående av en stor andel terrestriske omega-6-fettsyrer, samt en mindre andel omega-3-fettsyrer. Hos fisk reflekteres fettsyresammensetningen til føden i fiskens vev, og en rekke feltundersøkelser har vist at torsk fanget nært oppdrettsanlegg har høyere nivåer av terrestriske fettsyrer i muskel og lever enn fisk fanget lengre unna laskemerdene (Fernandez-Jover mfl. 2011, Meier mfl. 2023, Sanchez-Jerez mfl. 2024, Sanchez-Jerez mfl. 2025, Keeley mfl. 2025). I en feltundersøkelse gjort på gytende torsk rundt Smøla ble det også påvist en betydelig endring i fettsyresammensetningen hos gytebestanden. Her hadde 19 % av gytefisken fanget på gytefeltene en fettsyresammetning i leveren ensbetydende med beiting på spillfôr og/eller byttedyr som hadde spist spillfôr, mens de resterende individene hadde fettsyresammensetninger som reflekterte beiting på naturlige byttedyr (Meier mfl. 2023). Samme studie viste videre at de terrestriske fettsyrene ble overført til muskel og gonader, eksemplifisert med en 20 % økning av terrestriske fettsyrer i rognen til hunnfisk fra 7 til 27 % (Meier mfl. 2023). Slike resultater er entydige bevis på vedvarende inntak av byttedyr forbundet med oppdrett og/eller spillfôr, og støtter opp adferdsdata fra samme område som viste at en del av den kjønnsmodne kysttorsken oppholdt seg nært oppdrettsanlegg over lengre perioder (Skjæraasen mfl. 2022).
For torsk og andre marine fisker kan oppdrettsanlegg fungere som en effektiv energikilde, og i en studie gjort ved ni oppdrettslokaliteter langs norskekysten ble det dokumentert at torsk som beitet nært laksemerdene hadde bedre kondisjon og et vesentlig større energireservoar i form av leverstørrelse enn annen torsk (Dempster mfl. 2011, Fernandez-Jover mfl. 2011). Videre har undersøkelser gjort i laboratorier vist at torsk tolererer en plantebasert diett og kan opprettholde vekst så lenge fisken får nok marine omega-3 fettsyrer (Hansen og Hemre 2013). Til tross for dette representerer store mengder mat av avvikende kvalitet og fettsyresammensetning en risiko, ettersom det er svært lite kunnskap om hva langtidseffektene av en slik endring vil være på bestandsnivå (Bøhn mfl. 2024). Av spesiell interesse er hvordan overgangen fra en diett bestående av naturlige byttedyr med marine næringsstoffer til en diett dominert av terrestriske fettsyrer påvirker reproduksjon og rekruttering frem til nye voksne, kjønnsmodne individer, som igjen skal ha levedyktige avkom. Selv om økt mattilgang og kondisjon isolert sett er positivt for gonadevekst hos torsk (Wroblewski mfl. 1999, Skjæraasen mfl. 2010) har flere studier vist at både fekunditet og larveoverlevelse avhenger av tilgang til enkelte langkjedede marine fettsyrer (Pickova mfl. 1997, Salze mfl. 2005, Røjbek mfl. 2014, Norberg mfl. 2017). I en undersøkelse gjennomført av Barrett mfl. (2018), ble det påvist at kjønnsmoden torsk fanget i et område med høy tetthet av oppdrettsanlegg hadde egg som var 8 % mindre i diameter og larver som var 5 % kortere enn hos kjønnsmodne individer fanget i et område med mindre oppdrettsaktivitet. Det ble dog ikke påvist noen forskjell i klekkingssuksess mellom gruppene, og eventuelle kumulative effekter av lakseoppdrett på egg og larvestørrelse krever flere undersøkelser. I et pågående prosjekt ledet av Havforskningsinstituttet (MOTOR, Modningsfri torsk i merd, FHF, prosjektnr. 901815) undersøkes det blant annet hvorvidt forhøyede mengder terrestriske fettsyrer fra kommersielt torskefôr kan påvirke gonademodning, fekunditet og larveutvikling hos oppdrettstorsk. Foreløpige resultater viser at egg fra oppdrettstorsk som har spist torskefôr, har lavere befruktningsrate og lavere overlevelse i eggfasen sammenlignet med egg fra vill torsk. Forsøkene viste også at oppdrettstorsken fikk levedyktige avkom, og at larvene som overlevde, ikke hadde dårligere vekst enn larver fra vill torsk (Alix mfl. 2025). Det er behov for tilsvarende undersøkelser for å vurdere hvordan torskens reproduksjon påvirkes av laksefôr.
Eksponering mot skadelige fremmedstoffer
Miljøgifter og tungmetaller
Tidligere, da laksefôr hovedsakelig bestod av fiskeolje og fiskemel fra pelagisk fisk, kunne fôret inneholde relativt høye nivåer av persistente organiske miljøgifter (POP-er) og tungmetaller som kvikksølv (Hites mfl. 2004, Berntssen mfl. 2010). Felles for disse stoffene er at de oppkonsentreres i næringskjedene og rammer derfor predatorer høyere opp i disse. I en undersøkelse gjort i 2007 ble det påvist at torsk og sei som beitet under oppdrettsanlegg hadde henholdsvis 50 og 20 % høyere nivåer av organoklorid pesticider og bromerte flammehemmere i lever sammenlignet med fisk fra kontrollområder (Bustnes mfl. 2010). Som følge av dette konkluderte forfatterne at oppdrettsanlegg fungerte som punktkilde for enkelte fettløselige POP-er. Siden den gang har fôrsammensetningen endret seg betraktelig, og plantebasert fôr er vist å redusere forekomsten av POP-er i oppdrettslaks med 51–82 % (Berntssen mfl. 2010). Videre har nyere undersøkelser funnet høyere nivåer av POP-er i villaks enn i oppdrettslaks (Lundebye mfl. 2017), samt i sei fra områder ikke påvirket av oppdrett sammenlignet med sei tilknyttet oppdrettsanlegg (Pedersen 2013). Skiftet fra fiskebasert til plantebasert fôr har også redusert nivåene av uønskede tungmetaller i oppdrettslaksen betraktelig (Berntssen mfl. 2010), og en sammenligning gjort av villaks og oppdrettslaks har vist lavere nivåer av kvikksølv i sistnevnte (Lundebye mfl. 2017). Lignende resultater er også funnet for torsk og sei, der fisk fra kontrollområder generelt sett har hatt høyere nivåer av kvikksølv i muskel og lever sammenlignet med referansegrupper (Bustnes mfl. 2011, Arechavala-Lopez mfl. 2015, Fourdain mfl. 2022).
Et annet tungmetall som har fått en del fokus i risikovurderingen av lakseoppdrett, er kobber. Kobber er et naturlig tungmetall som forekommer i jordskorpen, i marine sediment og i sjøvann. Stoffet er en viktig faktor for enkelte enzymreaksjoner, men er giftig dersom konsentrasjonen av kobberforbindelser blir for høy. Kobber kan være giftig for ulike organismer i ulike utviklingsstadier. Det kan føre til redusert artsmangfold hvis konsentrasjonen i et gitt leveområde blir høyere enn artenes tålegrense, men kobber akkumuleres ikke oppover næringskjeden. Utslipp av kobber fra fiskeoppdrett kan forekomme via spillfȏr og ekskrementer, men de høyeste konsentrasjonene kommer fra impregnering av nøter der det brukes kobberstoff for å hindre begroing. Bruk av kobber som antigroemiddel har imidlertid gått kraftig ned de siste å rene og data fra 2024 viser en nedgang på 67 %, fra 1698 tonn i toppåret 2019 til 564 tonn i 2024 (tall fra Produktregisteret). Det er påvist forhøyede kobbernivåer i sedimentene rundt anlegg som ligger i områder med lite til moderat strøm og der det har vært oppdrett over lang tid (Grefsrud mfl. 2025). Miljøkvalitetsmål for kobber i sediment er satt til 84 mg/kg tørt sediment. Kobberkonsentrasjoner under denne verdien antas å ikke ha noen giftig effekt på marine organismer. Miljøkvalitetsmål for kobber i kystvann er på 2,6 µg/l, og målte konsentrasjoner har påvist nivå under dette (fra 0,5 - 1,9 µg/l). Studier på fisk fanget nært oppdrettsanlegg har vist at hverken torsk eller sei fanget under oppdrettsmerder hadde forhøyet nivåer av kobber i kroppen sammenlignet med fisk fra referansegruppene (Bustnes mfl. 2011, Fourdain mfl. 2022).
Sammenfallende med redusert bruk av kobber som antigroemiddlel, har forbruket av erstatningsstoffet tralopyril økt med 104 % fra 53 tonn i 2019 til 108 tonn i 2024 (tall fra Produktregisteret). Tralopyril er et ustabilt organisk pesticid med en halveringstid i sjø på ca. 9 – 16 timer, men vi har manglende kunnskap om forekomst og giftighet til nedbrytningsproduktene og hvordan disse påvirker marine organismer. Økt forbruk av tralopyril, samt bruken av andre erstatningsstoffer som sinkpyrithion og kobberpyrithion gjør at det er behov for å øke kunnskapen om hvilke effekter disse stoffene har på det marine miljøet, inkludert fisk som oppholder seg i områder med oppdrett.
Plantebaserte fôringredienser kan ha rester av sprøytemidler fra landbruk (Nácher-Mestre mfl. 2014). I dag er glyfosat (Roundup produktene) det mest brukte plantevernmiddelet både globalt og i Norge; en rolle virkestoffet har hatt siden midten av 1990-tallet da glyfosattolerante genmodifiserte planter, særlig soya, kom på markedet (Miyazaki mfl. 2019). I laksefôr er soya som hovedkilden for proteinkonsentrat, og i 2020 utgjorde soya 20,9 % av fôret (Aas mfl. 2022). Fôrundersøkelser gjort av Havforskningsinstituttet i 2022, 2023 og 2024 påviste glyfosat i henholdsvis 100, 88 og 93 % av fôrprøvene, med gjennomsnittskonsentrasjoner fra 70 til 100 µg/kg (Sele mfl. 2023, Lundebye mfl. 2024, 2025). Et nylig avsluttet prosjekt har utviklet screeningsmetoder for analyse av 200 ikke-klorerte-pesticider, inkludert også glyfosat og den viktigste metabolitten (AMPA) i laksefôr og laksefilet (Donald mfl. 2026). Her ble totalt 43 pesticider påvist, men ingen ble funnet med verdier over standard grenseverdi på 10 µg/kg i laksefilet, som regnes som et trygt nivå i mat. Selv om det ikke ble funnet glyfosat eller dens metabolitter i laksefilet, var det den dominerende pesticiden i fôret (medianverdi 100 µg/kg). Dette tyder på at opptaket av stoffet er lavt hos laks og at det ikke akkumulerer opp i næringskjeden. Likevel kan det ikke utelukkes at torsk som beiter på spillfôr kan bli eksponert for skadelige konsentrasjoner. Det finnes ingen kunnskap om hvorvidt glyfosat påvirker torsk via fôret, men det finnes det en rekke studier på at eksponering for glyfosat via vann kan påvirker adferd og fysiologi hos ulike fiskearter ved konsentrasjoner ned mot og under 100 µg/l (Lopes mfl. 2022). Som følge av dette er det behov for økt kunnskap om hvilke effekter glyfosat og glyfosatbaserte sprøytemidler kan ha på torsk. I tillegg til glyfosat er det også funnet en rekke andre sprøytemidler i laksefôr hvorav de vanligste er pirimiphos-methyl, chlorpyrifos og ulike former av DDT (Donald mfl. 2026).
Avlusningsmidler
Avlusningsmidlene som brukes av oppdrettsindustrien kan deles inn i to grupper: de som brukes til badebehandling og de som administreres til fisken via fôret. Siden toppåret i 2016, har det vært en årlig nedgang i bruk av legemidler mot lakselus. Denne reduksjonen er forårsaket av at oppdrettere har tatt i bruk ikke-medikamentelle metoder som varmt vann, ferskvann og mekanisk fjerning av lakselus.
I 2024 ble det brukt totalt 1 407 kg fôravlusningsmidler. Dette inkluderte 1 306 kg flubenzuroner og 101 kg emamektin-benzoat, bedre kjent ved legemiddelnavnet Slice. Flurbenzuroner hemmer syntesen av kitin og er derfor et effektive avlusningsmidler på alle stadier av lakselus som gjennomgår skallskifte, men stoffet er også dødelig for andre organismer som er avhengig av å produsere kitin i løpet av livssyklusen. I 2017 innførte Fiskeridirektoratet nye regler for å hindre uakseptabel bruk av kitinhemmere, hvor det blant annet ble presisert at slike legemidler ikke skal brukes innenfor 1000 m fra rekefelt. Undersøkelser av flurbenzuroner i fisk fanget nært oppdrettsanlegg har påvist generelt lave konsentrasjoner, med unntak av hos enkelte sei som sannsynligvis hadde spist spillfôr (Samuelsen mfl. 2015). Det foreligger lite kunnskap om hvorvidt flurbenzuroner påvirker torsk, men i et eksperiment gjort av Olsvik mfl. (2013) ble det påvist at selv om torsk kan ta opp stoffet, forsvinner det raskt fra kroppen og forårsaker kun små effekter på utrykket av enkelte gener som er aktive i avgiftningsprosesser.
Emamektin benzoat (emamektin), forårsaker hyperpolarisering av nerveceller og fører til død av alle utviklingsstadier hos lakselus. Stoffets er svært persistent i sediment og kan ha en halveringstid på over flere år under spesifikke miljøforhold (Samuelsen mfl. 2024). På grunn av denne lange halveringstid er det anbefalt og ikke utføre mer enn tre behandlinger i løpet av et år og ikke mer enn fem behandlinger i løpet av en to-årsperiode. Halveringstiden i vevsvæske, muskel og skinn hos torsk er beregnet til henholdsvis 180, 247 og 235 timer ved fôring på 9°C (Samuelsen 2010). Behandling av laks gitt emamektin via medisinert fôr med en dose på opptil 500 mg/kg kroppsvekt viste ingen dødelighet. Imidlertid ble det observert utvetydige tegn på giftighet, uttrykt ved blant annet fargeforandring, sløvhet og manglende appetitt, men kun ved den høyeste doseringen (Roy mfl. 2000).
Torsk som oppholder seg nært lakseoppdrett, kan i tillegg til å bli indirekte eksponert for skadelige kjemiske stoffer via spillfôr og beiting på oppdrettspåvirkede byttedyr, også utsettes for slike stoffer direkte via bruk av avlusningsmidler. Av badebehandlingene ble det brukt 731 tonn hydrogenperoksid i 2024, noe som representerer enn reduksjon på over 98 % sammenlignet med toppåret 2015 (Grefsrud mfl. 2026). Forbruket av azametifos og imidakloprid var i 2024 henholdsvis 795 kg og 1 587 tonn (Grefsrud mfl. 2026). Studier som har undersøkt effekten av hydrogenperoksid på fisk, er hovedsakelig gjort på laksefisk, ved konsentrasjoner brukt i behandling (Samuelsen mfl. 2024). Disse har vist at følsomheten øker ved temperatur, eksponeringstid og konsentrasjon (Samuelsen mfl. 2024). Det er lite sannsynlig at villfisk som oppholder seg i nærheten av oppdrettsanlegg vil bli eksponert for tilsvarende høye konsentrasjoner hydrogenperoksid sammenlignet med fisk under behandling. Dette bekreftes av feltundersøkelser gjort på Vestlandet, hvor vannprøver tatt innen 30 minutter fra behandling med hydrogenperoksid (1 330 mg/l) hovedsakelig innehold konstasjoner under 1 og 5 mg/l, avhengig av lokalitet og årstid (Samuelsen mfl. 2024). Undersøkelser gjort på egg fra torsk påviste 50 % dødelighet ved 343 mg/l noen som er langt over hva villfisk kan forventes å eksponeres for (Samuelsen mfl. 2024).
Azametifos er svært vannløselig og har dermed lav sannsynlighet for å binde seg til organisk materiale. Etter behandling vil stoffet hovedsakelig spres i miljøet via overflatestrømmer. Modelleringsstudier har vist at konsentrasjonen av azametifos rundt oppdrettsanlegg i løpet av 24 timer reduseres fra 100 µg/l til 0,2 µg/l i strømfattige områder og til 0,02 µg/l i områder med mye strøm (Samuelsen mfl. 2024). Dette er langt under nivåer som er påvist å ha en negativ effekt på fisk (Samuelsen mfl. 2024).
Sykdomsutbrudd og smittespredning i åpne laksemerder
I lakseoppdrett kan det forekomme sykdom forårsaket av både parasitter, virus og bakterier. Smittespredning vil påvirkes av mange ulike forhold, og vil variere med agens. Antallet og omfanget av utbrudd, dvs. mengden syk fisk som skiller ut smittestoff over tid, vil bestemme smittepresset på villfisk i omgivelsene. Agensets overlevelse i miljøet vil være viktig for spredningspotensialet, og vil påvirkes av miljøforhold som sollys (UV), temperatur, strøm og saltholdighet.
Mottakelige verter er en forutsetning for at smitte skal overføres fra oppdrettsfisk til villfisk. Smitte fra oppdrettslaks til villtorsk kan skje direkte, men også indirekte ved at andre mottagelige arter i miljøet overfører sykdom fra oppdrettslaks til torsk. Under et pågående sykdomsutbrudd ved en oppdrettslokalitet vil smitte spres til miljøet, og villfisk rundt oppdrettsanlegg vil kunne utsettes for økt smittepress. I dagens lakseoppdrett er det fiskelus, bendelorm, encellede gjelleparasitter, ulike virus og bakterielle sårinfeksjoner som dominerer. De fleste av disse agensene er verts-spesifikke, og har derfor liten evne til å smitte ubeslektede arter som torsk. Vi vil her omtale de agens og sykdommer som forekommer i lakseoppdrett med søkelys på de organismen som kan smitte villtorsk.
Parasitter
Fiskelus (Caligidae) utgjør en stor utfordring for den globale oppdrettsindustrien. Disse parasittiske krepsdyrene smitter direkte, enten ved planktoniske larver som spres i vannet eller ved at voksne lus bytter vertsindivid. I Norge finnes to vanlige arter fiskelus i åpne laksemerder: lakselus (Lepeophtheirus salmonis) og skottelus (Caligus elongatus). Lakselus forekommer bare på laksefisk, mens skottelus er lite artsspesifikk og kan forekomme hos over 80 marine fiskearter, inklusiv lakse- og torskefisk (Heuch mfl. 2007, Hemmingsen mfl. 2020). I motsetning til lakselus, som ikke forlater verten sin frivillig, kan skottelus skifte vertsindivid hyppig, samt leve fritt i vannmassene over lengre perioder. Påslag med voksne skottelus på oppdrettsfisk kan derfor være plutselige, og det er blitt spekulert om det har med innsig av villfisk å gjøre. Flere undersøkelser har identifisert rognkjeks (Cyclopterus lumpus) som foretrukken vert for skottelus, og disse kan ha langt over 100 lus på seg (e.g. Heuch mfl. 2007). Høye forekomster av skottelus er også registrert på torskeyngel (Neilson mfl. 1987), men når disse lusene blir voksne synes de å forlate fisken. Laboratorieeksperimenter har vist at større torsk ikke smittes med skotteluslarver (Karlsbakk mfl. 2009). Videre er chalimuslarver svært uvanlig på stor torsk (Heuch mfl. 2011) og det antas derfor at disse smittes av voksne skottelus fra vannmassene (Karlsbakk mfl. 2009).
Genetiske undersøkelser har identifisert to varianter av skottelus langs norskekysten, med forskjellig vertstilfang (Øines og Heuch 2007, Persson 2021). Skottelus av genotype 1 kan vokse opp på blant annet laksefisk og rognkjeks og bli værende på disse vertene som voksne lus. Genotype 2 skottelus kan derimot ikke utvikle seg på laks. Disse vokser opp på småfisk på grunt vann for så å bli frittsvømmende i vannet og gjøre et vertsskifte som voksne. Voksne genotype 2 er spesielt vanlig på villtorsk og annen torskefisk, men kan også smitte laks i oppdrett. Som følge av dette er det hovedsakelig genotype 1 skottelus som kan smitte fra laks til torsk. I tillegg kan det også tenkes at luselarver fra lus på oppdrettsfisk kan smitte torskeyngel, men ettersom torskeyngel virker lite påvirket av chalimuslarver og voksne lus vil forlate dem, antas dette å ha en begrenset effekt på villtorsk (Neilson mfl. 1987, Karlsbakk mfl. 2009, King 2023).
Av andre parasitter kan den encellede amøben Paramoeba perurans som forårsaker amøbegjellesykdom (AGD), gi store velferdsproblemer for laks i oppdrett. Amøben er ikke vertsspesifikk og kan infisere og forårsake sykdom også hos rensefisk (Haugland mfl. 2017). Amøben er ikke påvist hos torsk, selv om arten utvilsomt er eksponert. Andre gjelleproblemer på laks i oppdrett kan være knyttet til smitte med mikrosporidier og den ektoparasittiske encellede flagellaten Ichthyobodo salmonis. Disse parasittene er ikke påvist å infisere torsk.
Trichodina spp. er ciliater som angriper hud og gjeller og kan forårsake skader og dødelighet hos både torsk og laks i oppdrett (Khan 1991, 2004). Slike sykdomsutbrudd er knyttet til svekket fisk og dårlig vannkvalitet, og det er dermed lite aktuelt for villfisk. Videre er trolig Trichodina-artene delvis artsspesifikke, men ettersom dette ikke har blitt undersøkt nærmere kan man ikke utelukke mulige smitte fra oppdrettslaks til villtorsk.
Virussykdommer
De mest alvorlige virussykdommene i lakseoppdrett er forårsakes av virus som er spesifikke for laksefisk. Dette inkluderer blant annet salmonid alfavirus (SAV) som kan forårsake pankreas sykdom (PD), infeksiøs lakeanemi virus (ILAV) som kan forårsake infeksiøs lakseanemi og PRV1 som kan forårsake hjerte og skjelettmuskelbetennelse (HSMB). Ingen av disse virusene er påvist hos torsk eller annen torskefisk (Wiik-Nielsen mfl. 2012).
Oppdrettslaks plages av poxvirusinfeksjoner i gjellene, gjerne sammen med andre gjelleinfeksjoner. Smitten skjer primært i ferskvann, men kan også forekomme i sjø. Nylig er det påvist poxvirus også på oppdrettstorsk, men dette dreier seg om et eget, lite kjent virus (cod gill poxvirus, CGPV) (Gjessing mfl. 2024).
Et virus med lavere artsspesifisitet er infeksiøs pankreas nekrose virus (IPNV). Sykdommen infeksiøs pankreas nekrose (IPN) var i mange år en stor utfordring for oppdrettsnæringen inntil man startet å bruke en mer resistent laks. I 2023 ble viruset påvist på 43 lokaliteter med laks som representerer en dobling fra året før (Sommerset mfl. 2023). Viruset er sporadisk påvist i villtorsk, og har i et par tilfeller forårsaket sykdom hos yngel i oppdrett (Lorenzen mfl. 1995). Smitteforsøk har vist at torskeyngel er mottagelig for IPN fra laks (Jensen mfl. 2009, Urquhart mfl. 2009). IPNV er også vist å kunne overføres eksperimentelt fra IP-injisert torsk til uinfisert torsk via horisontal smitte (Weli mfl. 2025).
Et annet virus som kan smitte mellom marine fiskearter er viral hemoragisk septikemi virus (VHSV). Dette er et virus med bredt vertsspekter, med flere varianter (genotyper) i både fersk- og saltvann (Mohammadisefat mfl. 2023). VHS har forekommet i norsk oppdrett av regnbueørret, men hverken sykdommen eller viruset har vært påvist siden 2007 (Dale mfl. 2009), og i dag har Norge fristatus. Selv om det er påvist høy dødelighet hos juvenil torsk injisert med viruset, har flere eksperimentelle studier vist at torsk er svært lite mottagelig for VHSV (Snow mfl. 2000, 2005, Sandlund mfl. 2021).
Bakterielle sykdommer
Laksefisk i oppdrett er i dag godt beskyttet gjennom vaksinering mot bakterier som tidligere forårsaket alvorlige sykdommer. Det er derfor få utbrudd av vibriose (forårsaket av Vibrioanguillarum), kaldtvannsvibriose (Aliivibrio salmonicida) eller furunkulose (Aeromonassalmonicida subsp. salmonicida ) i oppdrett av laksefisk. Følgelig utøves det ikke noe relevant smittepress av disse bakteriene på villfisk. Det finnes varianter (serotyper) av V.anguillarum og underarter av Aeromonas salmonicida som er mer vanlige å finne hos torsk, men disse forekommer ikke hos laks i oppdrett. Det ansees derfor som lite sannsynlig at vibriose og furunkulose vil forekomme hos villtorsk som følge av smitte fra lakseoppdrett.
Sårproblematikk hos oppdrettsfisk kan være utfordrende til tider og skaper både velferdsutfordringer og dødelighet. Moritella viscosa, som forårsaker klassisk vintersår, og ulike varianter av Tenacibaculum spp. isoleres ofte fra laksefisk med sårskader. Fra oppdrettstorsk med sår er det blitt påvist både både M. viscosa (Colquhoun mfl. 2004) og Tenacibaculum spp. Trolig er det håndteringen av fisken som forårsaker sårene, ettersom bakteriene forekommer naturlig i sjøvann.
Pasteurellose er et problem for rognkjeks og laks i oppdrett. Det er to ulike varianter av bakterien, Pasteurella spp., som forårsaker sykdom hos de ulike artene (Gulla mfl. 2023). Bakteriene er ikke påvist hos torsk.
Yersiniose (Yersinia ruckeri) har vært en utfordring i lakseoppdrett i deler av Norge. Bakterien smitter hovedsakelig i ferskvann, men smitte i sjø kan forekomme, og på Island er det registrert utbrudd av yersiniose hos oppdrettstorsk (Gudmundsdottir mfl. 2014). På grunn av begrensede problemer i sjøfasen og fravær av påvisninger hos torsk ansees bakterien å utgjøre en liten fare for torsk i Norge.
Branchiomonas cysticola er en bakterie som er vanlig i forbindelse med gjellebetennelse og sykdommen epiteliocystis hos oppdrettslaks. Nylig er det påvist varianter av Branchiomonas i marin fisk, inklusivt i torsk med gjellesykdom (Gjessing mfl. 2024). Trolig er det snakk om egne typer av bakterien.
Referanser
Aas, T.S., Åsgård, T. & Ytrestøyl, T. (2022). Utilization of feed resources in the production of Atlantic salmon (Salmo salar) in Norway: An update for 2020. Aquaculture Reports 26, 101316. https://doi.org/10.1016/j.aqrep.2022.101316
Aglen, A., Nedreaas, K., Knutsen, J.A. & Huse, G. (2020). Kysttorsk nord for 62-grader nord. Vurdering av status og forslag til forvaltningstiltak og ny gjenoppbyggingsplan. Fisken og Havet nr. 2020-2. 64 s. https://www.hi.no/hi/nettrapporter/fisken-og-havet-2020-2
Alix, M., Yadav Ahir, S., Geitung, L., Perrichon, P., Fraser, T., Fjelldal. P.G., van der Meeren, T., Harboe, T., Utskot, S.O., Nola, V., Oppedal, F., Dempster, T. & Norberg, B. (2025). Reproductive biology and offspring development success in farmed cod are affected by nutrition, light and submergence. AQUA 2025, 22.-25. September, Valencia, Spania. In Aquaculture Europe 25 Abstracts, 50. https://eposters.blob.core.windows.net/eas-eposters/AE2025AbstractBook.pdf
Andersen, L.B., Grefsrud, E.S., Svåsand, T. & Sandlund, N. (2022). Risk understanding and risk acknowledgement: a new approach to environmental risk assessment in marine aquaculture. ICES Journal of Marine Science 79, 987-996. https://doi.org/10.1093/icesjms/fsac028
Arechavala-Lopez, P., Sæther, B.-S., Marhuenda-Egea, F., Sanchez-Jerez, P. & Uglem, I. (2015). Assessing the influence of salmon farming through total lipids, fatty acids, and trace elements in the liver and muscle of wild saithe Pollachius virens . Marine and Coastal Fisheries 7, 59-67. https://doi.org/10.1080/19425120.2014.1001540
Arnott, S.A. & Pihl, L. (2000). Selection of prey size and prey species by 1-group cod Gadus morhua: effects of satiation level and prey handling times. Marine Ecology Progress Series 198, 225-238. https://doi.org/10.3354/meps198225
Aven, T. (2014). Risk, Surprises and Black Swans: Fundamental Ideas and Concepts in Risk Assessment and risk management. Routledge, London. 276 s.
Barrett, L.T., Swearer, S.E., Harboe, T., Karlsen, Ø., Meier, S. & Dempster, T. (2018). Limited evidence for differential reproductive fitness of wild Atlantic cod in areas of high and low salmon farming density. Aquaculture Environment Interactions 10, 369-383. https://doi.org/10.3354/aei00275
Barrett, L.T., Swearer, S.E. & Dempster, T. (2019). Impacts of marine and freshwater aquaculture on wildlife: a global meta-analysis. Reviews in Aquaculture 11, 1022–1044. https://doi.org/10.1111/raq.12277
Barth, J.M.I., Villegas-Ríos, D., Freitas, C., Moland, E., Star, B., André, C., Knutsen, H., Bradbury, I., Dierking, J., Petereit, C., Righton, D., Metcalfe, J., Jakobsen, K.S., Olsen, E.M. & Jentoft S. (2019). Disentangling structural genomic and behavioural barriers in a sea of connectivity. Molecular Ecology 28, 1394-1411. https://doi.org/10.1111/mec.15010
Bell, J.G., McGhee, F., Campbell, P.J. & Sargent, J.R. (2003). Rapeseed oil as an alternative to marine fish oil in diets of post-smolt Atlantic salmon (Salmo salar): changes in flesh fatty acid composition and effectiveness of subsequent fish oil “wash out”. Aquaculture 218, 515–528. https://doi.org/10.1016/S0044-8486(02)00462-3
Berntssen, M.H.G., Olsvik, P.A., Torstensen, B.E., Julshamn, K., Midtun, T., Goksøyr, A., Johansen, J., Sigholt, T., Joerum, N., Jakobsen, J.-V., Lundebye, A.-K. & Lock E.-J. (2010). Reducing persistent organic pollutants while maintaining long chain omega-3 fatty acid in farmed Atlantic salmon using decontaminated fish oils for an entire production cycle. Chemosphere 81, 242-252. https://doi.org/10.1016/j.chemosphere.2010.06.031
Bjørn, P.A., Uglem, I., Kerwath, S., Sæther, B.S., Nilsen, R. (2009). Spatiotemporal distribution of Atlantic cod (Gadus morhua L.) with intact and blocked olfactory sense during the spawning season in a Norwegian fjord with intensive salmon farming. Aquaculture 286, 36-44. https://doi.org/10.1016/j.aquaculture.2008.08.038
Bustnes, J.O., Lie, E., Herzke, D., Dempster, T., Bjørn, P.A., Nygård, T. & Uglem, I. (2010). Salmon farms as a source of organohalogenated contaminants in wild fish. Environmental Science & Technology 44, 8736-8743. https://doi.org/10.1021/es102195d
Bustnes, J.O., Nygård, T., Dempster, T., Ciesielski, T., Jensen, B.M., Bjørn, P.A. & Uglem I. (2011). Do salmon farms increase the concentration of mercury and other elements in wild fish? Journal of Environmental Monitoring 13, 1687-1694. https://doi.org/10.1039/C1EM10083A
Bøhn, T., Strøm, J.F., Sanchez-Jerez, P., Keeley, N.B., Johansen, T., Gjelland, K.Ø., Sandlund, N., Sæther, B.-S., Sætra, I., Olsen, E.M., Skjæraasen, J.E., Meier, S., van der Meeren, T. & Bjørn, P.A. (2024). Ecological interactions between farmed Atlantic salmon and wild Atlantic cod populations in Norway: A review of risk sources and knowledge gaps. Reviews in Aquaculture 16, 1333-1350. https://doi.org/10.1111/raq.12899
Breistein, B., Dahle, G., Johansen, T., Besnier, F., Quintela, M., Jorde, P.E., Knutsen, H., Westgaard, J.-I., Nedreaas, K., Farestveit, E. & Glover, K.A. (2022). Geographic variation in gene flow from a genetically distinct migratory ecotype drives population genetic structure of coastal Atlantic cod (Gadus morhua L.). Evolutionary Applications 15, 1162-1176 https://onlinelibrary.wiley.com/doi/10.1111/eva.13422
Callier, M.D., Byron, C.J., Bengtson, D.A., Cranford, P.J., Cross, S.F., Focken, U., Jansen, H.M., Kamermans, P., Kiessling, A., Landry, T., O’Beirn, F., Petersson, E., Rheault, R.B., Strand, Ø., Sundell, K., Svåsand, T., Wikfors, G.H. & McKindsey, C.W. (2018). Attraction and repulsion of mobile wild organisms to finfish and shellfish aquaculture: a review. Reviews in Aquaculture 10, 924–949. https://doi.org/10.1111/raq.12208
Clemens, B.J. & Schreck, C.B. (2021). An assessment of terminology for intraspecific diversity in fishes, with a focus on “ecotypes” and “life histories”. Ecology and Evolution 11, 10772–10793. https://doi.org/10.1002/ece3.7884
Colquhoun, D.J., Hovland, H., Hellberg, H., Haug, T. & Nilsen, H. (2004). Moritella viscosa isolated from farmed Atlantic cod ( Gadus morhua ). Bulletin of the European Association of Fish Pathologists 24, 109-114. https://eafp.org/download/2004-Volume24/Issue%202/24_109.pdf
Dahle G., Quintela M., Johansen T., Westgaard J.I., Besnier F., Aglen A., Jorstad K.E. & Glover K.A. (2018). Analysis of coastal cod ( Gadus morhua L.) sampled on spawning sites reveals a genetic gradient throughout Norway's coastline. Bmc Genetics 19, 42. https://doi.org/10.1186/s12863-018-0625-8
Dale, O.B., Ørpetveit, I., Lyngstad, T.M., Kahns, S., Skall, H.F., Olesen, N.J. & Dannevig, B.H. (2009). Outbreak of viral haemorrhagic septicaemia (VHS) in seawater-farmed rainbow trout in Norway caused by VHS virus Genotype III. Disease of Aquatic Organisms 85, 93-103. https://doi.org/10.3354/dao02065
Dempster, T., Sanchez-Jerez, P., Uglem, I. & Bjørn, P.-A. (2010). Species-specific patterns of aggregation of wild fish around fish farms. Estuarine, Coastal and Shelf Science 86, 271–275. https://doi.org/10.1016/j.ecss.2009.11.007
Dempster, T., Sanchez-Jerez, P., Fernandez-Jover, D., Bayle-Sempere, J., Nilsen, R., Bjørn, P.-A. & Uglem, I. (2011). Proxy Measures of Fitness Suggest Coastal Fish Farms Can Act as Population Sources and Not Ecological Traps for Wild Gadoid Fish. PLOS ONE 6, e15646. https://doi.org/10.1371/journal.pone.0015646
Dempster, T., Uglem, I., Sanchez-Jerez, P., Fernandez-Jover, D., Bayle-Sempere, J., Nilsen, R. & Bjørn, P.A., (2009). Coastal salmon farms attract large and persistent aggregations of wild fish: An ecosystem effect. Marine Ecology Progress Series 385, 1–14. https://doi.org/10.3354/meps08050
Donald, C.E., Bernhard, A., Hauglid, M.F., Fürstenberg, A., Berntssen, M.H.G., Lundebye, A.-K. & Sele, V. (2026). Pesticide residues in aquaculture salmon filet and feed: Quantitative and qualitative screening profiles of legacy and current-use pesticides using GC-EI-MS/MS and LC-ESI-IMS-QToF. Journal of Chromatography B 1269, 124864. https://doi.org/10.1016/j.jchromb.2025.124864
Dunlop, K., Staby, A., van der Meeren, T., Keeley, N., Olsen, E.M., Bannister, R. & Skjæraasen, J.E. (2022). Habitat associations of juvenile Atlantic cod (Gadus morhua L.) and sympatric demersal fish communities within shallow inshore nursery grounds. Estuarine, Coastal and Shelf Science 279, 108111. https://doi.org/10.1016/j.ecss.2022.108111
Dunlop, K., Strammer, I. & Keeley, N. (2024). "Attraction and avoidance of wild demersal fish and crustaceans to open-net aquaculture pens resolved by baited and towed underwater camera surveys." Frontiers in Marine Science 11. https://doi.org/10.3389/fmars.2024.1400195
Espeland, S.H., Norderhaug, K.M., Thormar, J., Kleiven, P.J.N., Aanonsen, I.A.N., Saltskår, J., Jørgensen, T., Christensen, L., van der Meeren, T., Stiansen, S. & Humborstad O.-B. (2022). Metoder for kartlegging av oppvekstområder for kystnære bestander av fisk. Rapport fra Havforskningen Nr. 2022-43, 86 s. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-2022-43
Fernandez-Jover, D., Martinez-Rubio, L., Sanchez-Jerez, P., Bayle-Sempere, J.T., Lopez Jimenez, J. A., Martinez Lopez, F.J., Bjørn, P.-A., Uglem, I. & Dempster, T. (2011). Waste feed from coastal fish farms: A trophic subsidy with compositional side-effects for wild gadoids. Estuarine, Coastal and Shelf Science 91, 559-568. https://doi.org/10.1016/j.ecss.2010.12.009
Fourdain, L., Arechavala-Lopez, P., Uglem, I., Sæther, B.S. & Sanchez-Jerez, P. (2022). Spatiotemporal variations in trace element compositions in pollock populations under the influence of coastal Norwegian salmon farms. Marine and Coastal Fisheries 14, e10193. https://doi.org/10.1002/mcf2.10193
Gjelland , K.Ø., Strøm, J.F., Olsen, E.M., Baktoft, H., Skjæraasen, J.E., Johansen, T., Keeley, N.B.,Bygdnes, E.E., van der Meeren, T., Bjørn, P.A., Bøhn, T . (i trykk). Precise positioning of Atlantic cod reveals size-dependent preferences to substrate and bathymetric terrain. Marine Ecology Progress Series. https://doi.org/10.3354/meps15039
Gjessing, M.C., Tengs, T., Nilsen, H., Mohammad, S. & Weli, S.C. (2024). The first report of cod gill poxvirus in gills of Atlantic cod (Gadus morhua L.) suffering from cardiorespiratory disease. J. Fish Dis. e14078, https://doi.org/10.1111/jfd.14078
Grefsrud, E.S., Andersen, L.B., Grøsvik, B.E., Hansen, P.K., Husa, V., Samuelsen, O., Sandlund, N., Skern, R., Solberg, M.F. & Stien, L.H. (redaktører) (2025). Risikorapport norsk fiskeoppdrett 2026 - produksjonsdødelighet hos oppdrettsfisk og miljøeffekter av norsk fiskeoppdrett. Rapport fra havforskningen 2025-14.
Grefsrud, E.S., Andersen, L.B., Dunlop, K.M., Grøsvik, B.E., Hannisdal, R., Husa, V., Sandlund, N., Skern, R., Solberg, M.F. & Stien, L.H. (redaktører) (2026). Risikorapport norsk fiskeoppdrett 2026 - produksjonsdødelighet hos oppdrettsfisk og miljøeffekter av norsk fiskeoppdrett. Rapport fra havforskningen 2026.
Gudmundsdottir, B.K., Gudmundsdottir, S., Gudmundsdottir, S. & Magnadottir, B. (2014). Yersiniosis in Atlantic cod, Gadus morhua (L.), characterization of the infective strain and host reactions. Journal of Fish Diseases 37, 511-519. https://doi.org/10.1111/jfd.12139
Gulla, S., Colquhoun, D.J., Olsen, A.B., Spilsberg, B., Lagesen, K., Åkesson, C.P., Strøm, S., Manji, F., Birkbeck, T.H., Nilsen, H.K. (2023). Phylogeography and host specificity of Pasteurellaceae pathogenic to sea-farmed fish in the north-east Atlantic. Frontiers in Microbiology 14, 1236290. https://doi.org/10.3389/fmicb.2023.1236290
Hansen, A.-C. & Hemre, G.-I. (2013). Effects of replacing fish meal and oil with plant resources in on-growing diets for Atlantic cod Gadus morhua L. Aquaculture Nutrition 19, 641-650. https://doi.org/10.1111/anu.12078
Haugland, G.T., Olsen, A.-B., Rønneseth, A. & Andersen, L. (2017). Lumpfish (Cyclopterus lumpus L.) develop amoebic gill disease (AGD) after experimental challenge with Paramoeba perurans and can transfer amoebae to Atlantic salmon (Salmo salar L.). Aquaculture 478, 48-55. https://doi.org/10.1016/j.aquaculture.2016.04.001
Hemmingsen, W., MacKenzie, K., Sagerup, K., Remen, M., Bloch-Hansen, K. & Imsland, A.K.D. (2020). Caligus elongatus and other sea lice of the genus Caligus as parasites of farmed salmonids: A review. Aquaculture 522, 735160. https://doi.org/10.1016/j.aquaculture.2020.735160
Heuch, P.A., Øines, Ø., Knutsen, J.A. & Schram, T.A. (2007). Infection of wild fishes by the parasitic copepod Caligus elongatus on the south east coast of Norway. Disease of Aquatic Organisms 77, 149-158. https://doi.org/10.3354/dao01833
Heuch, P.A., Jansen, P.A., Hansen, H., Sterud, E., MacKenzie, K., Haugen, P. & Hemmingsen, H. (2011). Parasite faunas of farmed cod and adjacent wild cod populations. Aquaculture Environment Interactions 2, 1-13. https://doi.org/10.3354/aei00027
Hites, R.A., Foran, J.A., Carpenter, D.O., Hamilton, M.C., Knuth, B.A., Schwager, S.J. (2004). Global assessment of organic contaminants in farmed salmon. Science 303, 226-229. https://doi.org/10.1126/science.1091447
ICES (2022a). Norwegian request for evaluation of management of Norwegian coastal cod north of 67° N. I Report of the ICES Advisory Committee, 2022. ICES Advice 2022, sr.2022.08, 5 s. https://ices-library.figshare.com/ndownloader/files/35770520
ICES (2024). Cod (Gadus morhua) in Subarea 2 between 62°N and 67°N (Norwegian Sea), southern Norwegian coastal cod. In Report of the ICES Advisory Committee, 2024. ICES Advice 2024, cod.27.2.coastS. 7 s. https://doi.org/10.17895/ices.advice.25019210
ICES (2025a ICES (2025a). Cod (Gadus morhua) in Subarea 2 between 62°N and 67°N (Norwegian Sea), southern Norwegian coastal cod. In Report of the ICES Advisory Committee, 2025. ICES Advice 2025, cod.27.2.coastS. 7 s. https://doi.org/10.17895/ices.advice.28194458
ICES (2025c). Cod (Gadus morhua) in subareas 1 and 2 north of 67°N (Norwegian Sea and Barents Sea), northern Norwegian coastal cod. In Report of the ICES Advisory Committee, 2025. ICES Advice 2025, cod.27.1-2coastN, https://doi.org/10.17895/ices.advice.27202557
Johansen, T., Besnier, F., Quintela, M., Jorde, P.E., Glover, K.A. Westgaard, J.-I., Dahle, G., Lien, S. & Kent, M.P. (2020). Genomic analysis reveals neutral and adaptive patterns that challenge the current management regime for East Atlantic cod Gadus morhua L. Evolutionary Applications 13, 2673-2688. https://doi.org/10.1111/eva.13070
Johansen, T., Westgaard, J.-I., Seliussen, B.B., Nedreaas, K., Dahle, G., Glover, K.A., Kvalsund, R. & Aglen, A. (2018). “Real-time” genetic monitoring of a commercial fishery on the doorstep of an MPA reveals unique insights into the interaction between coastal and migratory forms of the Atlantic cod. ICES Journal of Marine Science 75, 1093-1104. https://doi.org/10.1093/icesjms/fsx224
Jorde, P.E., Kleiven, A.R., Sodeland, M., Olsen, E.M., Ferter, K., Jentoft, S., Knutsen, H. (2018). Who is fishin gon what stock: population-of-origin of individual cod (Gadus morhua) in commercial and recreational fisheries. ICES Journal of Marine Science 75, 2153-2163. https:// doi.org/10.1093/icesjms/fsy080
Karlsbakk E, Isaksen E, Hamre L (2009) Hva vet vi om parasitter og oppdrett av torsk? Fisken og havet 2009 – 2: 4 s.
Khan, R.A. (1991). Mortality in Atlantic salmon (Salmo salar) associated with trichodinid ciliates. Journal of Wildlife Diseases 27, 153-155. https://doi.org/10.7589/0090-3558-27.1.153
Khan, R.A. (2004). Disease outbreaks and mass mortality in cultured Atlantic cod, Gadus morhua L., associated with Trichodina murmanica (Ciliophora). Journal of Fish Diseases 27, 181-184. https://doi.org/10.1111/j.1365-2761.2004.00525.x
Keeley, N., Meier, S., Laroche, O., Steiner, K., Bøhn, T., Olsen, E., Johansen, T. & Bjørn, P.A. (2025). Connecting the dots: evidence of interactions between wild Atlantic cod Gadus morhua, the benthic environment and salmon farm discharges. Aquaculture Environment Interactions 17, 85-106. https://doi.org/10.3354/aei00496
King, A.M. (2023). The sea louse Caligus elongatus (Caligidae). Hosts susceptibility to the two genotypes. MSc oppgave i fiskehelse, Univ. Bergen. 48 s. https://hdl.handle.net/11250/2761777
Knutsen, H., Jorde, P.E., Hutchings, J.A., Hemmer-Hansen, J., Grønkjær, P., Jørgensen, K.-E.M., André, C., Sodeland, M., Albretsen, J. & Olsen, E.M. (2018). Stable coexistence of genetically divergent Atlantic cod ecotypes at multiple spatial scales. Evolutionary Applications 11, 1527–1539. https://doi.org/10.1111/eva.12640
Kristensen, M.L., Olsen, E.M., Moland, E., Knutsen, H., Grønkjær, P., Koed, A., Källo, K. & Aarestrup, K. (2021). Disparate movement behavior and feeding ecology in sympatric ecotypes of Atlantic cod. Ecology and Evolution 11, 11477–11490. https://doi.org/10.1002/ece3.7939
Link, J.S. & Garrison, L.P. (2002). Trophic ecology of Atlantic cod Gadus morhua on the northeast US continental shelf. Marine Ecology Progress Series 227, 109-123. https://doi.org/10.3354/meps227109
Lopes, A.R., Moraes, J.S. & de Martinez Gaspar Martins, C. (2022). Effects of the herbicide glyphosate on fish from embryos to adults: a review addressing behavior patterns and mechanisms behind them. Aquatic Toxicology 251, 106281. Https://doi.org/10.1016/j.aquatox.2022.106281
Lorenzen, E., Olesen, N.J., Strøm, A. & Evensen, Ø. (1995). Outbreaks of IPN in reared fry of Atlantic cod (Gadus morhua). In : European Association of Fish Pathologists, Seventh international conference "Diseases of Fish and Shellfish", 10.-15. Sept. 1995, Palma de Mallorca, Spain, s.38.
Lundebye, A.-K., Lock, E.J., Rasinger, J.D., Nøstbakken, O.J., Hannisdal, R., Karlsbakk, E., Wennevik, V., Madhun, A.S., Madsen, L., Graff, I.E. & Ørnsrud, R. (2017). Lower levels of Persistent Organic Pollutants, metals and the marine omega 3-fatty acid DHA in farmed compared to wild Atlantic salmon (Salmo salar). Environmental Research 155, 49-59. https://doi.org/10.1016/j.envres.2017.01.026
McConnell, A., Routledge, R., Connors, B.M. (2010). Effect of artificial light on marine invertebrate and fish abundance in an area of salmon farming. Marine Ecology Progress Series 419, 147-156. https://doi.org/10.3354/meps08822
Meier, S., van der Meeren, T., Skjæraasen, J.E., Bannister, R.J., Rasinger, J.D. & Karlsen, Ø. (2023). Terrestrial fatty acids from feed oil in feed for farmed salmonids are transferred to the liver, gonads, and muscle of wild Atlantic cod ( Gadus morhua ). ICES Journal of Marine Science 80, 1143-1154. https://doi.org/10.1093/icesjms/fsad051
Miyazaki, J., Bauer-Panskus, A., Bøhn, T., Reichenbecher, W. & Then, C. (2019). Insufficient risk assessment of herbicide-tolerant genetically engineered soybeans intended for import into the EU. Environmental Science Europe 31, 92. https://doi.org/10.1186/s12302-019-0274-1
Mohammadisefat, P., Zorriehzahra, M.J., Adel, M., Chamjangali, Z.A., Jabbari, M., Eftekhari, A., Farzipour, H. & Jazi, S.Y. (2023). Viral hemorrhagic septicemia virus (VHSV), past, present and future: a review. International Aquatic Research 15, 191-203. https://doi.org/10.22034/IAR.2023.1983457.1424
Moland, E., Synnes, A.-E., Naustvoll, L.-J., Brandt, C.F., Norderhaug, K.M., Thormar, J., Biuw, M., Jorde, P.E., Dahle, G., Jelmert, A., Bosgraaf, S., Olsen, E.M., Deininger, A. & Haga, A. (2021). Krafttak for kysttorsken - Kunnskap for stedstilpasset gjenoppbygging av bestander, naturtyper og økosystem i Færder- og Ytre Hvaler nasjonalparker. Rapport fra havforskningen 2021-2. 56 s. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-2021-2
Nácher-Mestre, J., Serrano, R., Portoles, T., Berntssen, M.H.G., Perez-Sanchez, J. & Hernández, F. (2014). Screening of pesticides and polycyclic aromatic hydrocarbons in feeds and fish tissues by gas chromatography coupled to high-resolution mass spectrometry using atmospheric pressure chemical ionization. Journal of Agricultural and Food Chemistry 62, 2165-2174. https://doi.org/10.1021/jf405366n
Nedreaas, K., Aglen, A., Gjøsæter, J., Jørstad, K., Knutsen, H., Smedstad, O., Svåsand, T. & Ågotnes, P. (2008). Kysttorskforvaltning på Vestlandet og langs Skagerrak-kysten. Vurdering av status for kysttorsk på strekningen svenskegrensen–Stad med forslag om forvaltningstiltak. Fisken og Havet, 5–2008. 106 s. https://www.hi.no/hi/nettrapporter/fisken-og-havet/74725/nr.5_2008
Neilson, J.D., Perry, R.I., Scott, J.S. & Valerio, P. (1987). Interactions of caligid ectoparasites and juvenile gadids on Georges Bank. Marine Ecology Progress Series 39, 221-232. https://www.jstor.org/stable/24825683
Nickel, A.K., Campana, S.E. & Ólafsdóttir, G.Á. (2025). Temperature and body size affect movement of juvenile Atlantic cod (Gadus morhua) and saithe (Pollachius virens) at nearshore nurseries. Fish Biology 106, 1554-1569. https://doi.org/10.1111/jfb.15850
Norberg, B., Kleppe, L., Andersson, E., Thorsen, A., Rosenlund, G. & Hamre, K. (2017). Effects of dietary arachidonic acid on the reproductive physiology of female Atlantic cod (Gadus morhua L.). General and Comparative Endocrinology 250, 21-35. https://doi.org/10.1016/j.ygcen.2017.05.020
Olsen, E.M., Strøm, J.F., Skjæraasen, J.E., van der Meeren, T., Jorde, P.E., Johansen, T., Gjelland, K.Ø., Karlsen, Ø., Bøhn, T., Bjørn, P.A. (i trykk). Intraspecific diversity in Atlantic cod (Gadus morhua) spatial ecology within subarctic fjords during the foraging season. Marine Ecology Progress Series. https://doi.org/10.3354/meps15087
Olsvik, P.A., Samuelsen, O.B., Erdal, A., Holmelid, B. & Lunestad, B.T. (2013). Toxicological assessment of the anti-salmon lice drug diflubenzuron on Atlantic cod Gadus morhua . Diseases of Aquatic Organisms 105, 27-43. https://doi.org/10.3354/dao02613
Pedersen, L.B. (2013). Levels of persistent organic pollutants (POPs) and physiological and biochemical changes in farm associated wild fish caught in Masfjorden. Masteroppgave, Universitetet i Bergen. 65 s.
Pedersen, M.W. & Berg, C.W. (2017). A stochastic surplus production model in continuous time. Fish and Fisheries 18, 226-243. https://doi.org/10.1111/faf.12174
Persson, H.L. (2021). The sea louse Caligus elongatus (Caligidae). Genetic variation and host use by its two genotypes. MSc thesis, Dept. Bioscienses, Univ. Bergen. 71 s.
Pickova, J., Dutta, P.C., Larsson, P.-O. & Kiessling, A. (1997). Early embryonic cleavage pattern, hatching success, and egg-lipid fatty acid composition: comparison between two cod (Gadus morhua) stocks. Canadian Journal of Fisheries and Aquatic Sciences 54, 2410-2416. https://doi.org/10.1139/f97-148
Robichaud, D. & Rose, G.A. (2004). Migratory behaviour and range in Atlantic cod: inference from a century of tagging. Fish and Fisheries 5, 185-214. https://doi.org/10.1111/j.1467-2679.2004.00141.x
Roy, W.J., Sutherland, I.H., Rodger, H.D.M. & Varma, K.J. (2000). Tolerance of Atlantic salmon, Salmo salar L., and rainbow trout, Oncorhynchus mykiss Walbaum, to emamectin benzoate, a new orally administered treatment for sea lice. Aquaculture 184, 19-29. https://doi.org/10.1016/S0044-8486(99)00307-5
Røjbek, M.C., Støttrup, J.G., Jacobsen, C., Tomkiewicz, J., Nielsen, A. & Trippel, E.A. (2014). Effects of dietary fatty acids on the production and quality of eggs and larvae of Atlantic cod (Gadus morhua L.). Aquaculture Nutrition 20, 654-666. https://doi.org/10.1111/anu.12124
Salze, G., Tocher, D.R., Roy, W.J. & Robertson, D.A. (2005). Egg quality determinants in cod (Gadus morhua L.): egg performance and lipids in eggs from farmed and wild broodstock. Aquaculture Research 36, 1488-1499. https://doi.org/10.1111/j.1365-2109.2005.01367.x
Samuelsen OB, Lunestad BT, Hannisdal R, Bannister R, Olsen S, Tjensvoll T, Farestveit E, Ervik A (2015). Distribution and persistence of the anti sea-lice drug teflubenzuron in wild fauna and sediments around a salmon farm, following a standard treatment. Science of the Total Environment 508: 115–121. https://doi.org/10.1016/j.scitotenv.2014.11.082
Sanchez-Jerez, P., Fernandez-Jover, D., Uglem, I., Arechavala-Lopez, P., Dempster, T., Bayle-Sempere, J., Pérez, C., Izquierdo-Gomez, D., Bjørn, P.-A. & Nilsen, R. (2011). Coastal Fish Farms as Fish Aggregation Devices (FADs), In Bortone, S.A., Pereira, B.F., Fabi, G. & Otake, S. (Eds.), Artificial Reefs in Fisheries Management. CRC Press, Boca Raton, FL, USA, 187-208.
Sanchez-Jerez, P., Atalah, J., Sætra, I.M., Bøhn, T., Saether, B.-S., Johansen, T., Keeley, N., van der Meeren, T. & Bjørn, P.A. (2024). Comparative study of fatty acid composition of muscles of Atlantic cod (Gadus morhua Linnaeus, 1758) with natural diet and feeding near salmon farms. Aquaculture Journal 4, 246-254. https://doi.org/10.3390/aquacj4040018
Sanchez-Jerez, P., Atalah, J., Sætra, I.M., Bøhn, T., Sæther, B.-S., Johansen, T., Keeley, N., van der Meeren, T. & Bjørn, P.A. (2025). Effects of salmon farming on Atlantic cod: insights from liver fatty acid composition in relationship with biological and genetic variables. Aquaculture Environment Interactions 17, 149-160. https://doi.org/10.3354/aei00501
Sandlund, N., Johansen, R., Fiksdal, I.U., Einen, A.C.B., Modahl, I., Gjerset, B. & Bergh, Ø. (2021). Susceptibility and pathology in juvenile Atlantic cod Gadus morhua to a marine viral haemorrhagic septicaemia virus isolated from diseased rainbow trout Oncorhynchus mykiss. Animals 11, 3523. https://doi.org/10.3390/ani11123523
Sele, V., Silva, M., Philip, A., Hamre, K., Skjærven, K., Espe, M., Liland, N., Lie, K., Berntssen, M. & Lundebye, A.-K. (2023). Program for overvåking av fiskefôr - Årsrapport for prøver innsamlet i 2022. Rapport fra havforskningen 2023-36. 38 s. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-2023-36
Skjæraasen, J.E., Meager, J.J., Karlsen, Ø., Mayer, I., Dahle, G., Rudolfsen, G. & Fernö, A. (2010). Mating competition between farmed and wild cod Gadus morhua . Marine Ecology Progress Series 412, 247-258. https://doi.org/10.3354/meps08670
Skjæraasen, J.E., Karlsen, Ø., Langangen, Ø., van der Meeren, T., Keeley, N.R., Myksvoll, M.S., Dahle, G., Moland, E., Nilsen, R., Elvik Schrøder, K.M., Bannister, R.J. & Olsen, E.M. (2021). Impact of salmon farming on Atlantic cod spatio-temporal reproductive dynamics. Aquaculture Environment Interactions 13: 399-412. https://doi.org/10.3354/aei00415
Skjæraasen, J.E., Karlsen, Ø., Langangen, Ø., Meier, S., Dunlop, K.M., van der Meeren, T., Keeley, N.B., Myksvoll, M.S., Dahle, G., Moland, E., Nilsen, R., Elvik, K.M.S., Bannister, R.J. & Olsen E.M. (2022). The attraction of cod Gadus morhua from local coastal spawning grounds to salmon farms. Aquaculture Environment Interactions 14, 229-242. https://doi.org/10.3354/aei00440
Skaar, I.C.A. (2021). Oppvekstområder for kysttorsk. Masteroppgave, Universitetet i Oslo. 77 s.
Snow, M., Cunningham, C.O. & Bricknell, I.R. (2000). Susceptibility of juvenile Atlantic cod Gadus morhua to viral haemorrhagic septicaemia virus isolated from wild-caught Atlantic cod. Diseases of Aquatic Organisms, 41, 225-229. https://doi.org/10.3354/dao041225
Snow, M., King, J. A., Garden, A. & Raynard, R.S. (2005). Experimental susceptibility of Atlantic cod, Gadus morhua (L.), and Atlantic halibut, Hippoglossus hippoglossus (L.), to different genotypes of viral haemorrhagic septicaemia virus. Journal of Fish Diseases 28, 737-742. https://doi.org/10.1111/j.1365-2761.2005.00682.x
Stock, B., Knutsen, H., Nedreaas, K., Otterå, H., Fall, J., Denechaud, C., Moland, E., Jorde, P.E. & Kleiven, A.R. (2025). Oppdatert status om torsk langs Vestlandet- og Skagerrakkysten: potensial for bestandsvurdering. Rapport fra Havforskningen 2025-42, 64 s. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-en-2025-42
Strøm, J.F., Bøhn, T., Skjæraasen, J.E., Gjelland, K.Ø. Karlsen, Ø., Johansen, T., Hanebrekke, T., Bjørn, P.A. & Olsen, E.M. (2023). Movement diversity and partial sympatry of coastal and Northeast Arctic cod ecotypes at high latitudes. Journal of Animal Ecology 92, 1966-1978. https://doi.org/10.1111/1365-2656.13989
Strøm, J.F., Berg, E., Bjørn, P.A., Bøhn, T., Espeland, S.H., Fall, J., Karlsbakk, E., Meier, S., Sandlund, N., Skjæraasen, J.E., Stock, B., van der Meeren, T., Grefsrud, E.S., (2025). Økologiske effekter av lakseoppdrett i åpne merder på ville kysttorskbestander – kunnskapsstatus. Rapport fra Havforskningen 2025-15, 36 s. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-2025-15
Svåsand, T., Michalsen, K., Fall, J., Bogstad, B., Johansen, T. & Glover, K.A. (2025). Analysis of long-term Atlantic cod tag-recapture data from Norway reveals insights into historical migration patterns and mortality rates of Norwegian coastal cod. ICES Journal of Marine Science 82, fsaf137. https://doi.org/10.1093/icesjms/fsaf137
Sæther, B.S., Bjørn, P.A. & Dale, T. (2007). Behavioural responses in wild cod ( Gadus morhua L.) exposed to fish holding water. Aquaculture 262, 260-267. https://doi.org/10.1016/j.aquaculture.2006.11.020
Sæther, B.-S., Løkkeborg, S., Humborstad, O.-B., Tobiassen, T., Hermansen, Ø. & Midling, K.Ø. (2012). Fangst og mellomlagring av villfisk ved oppdrettsanlegg. Nofima rapport nr. 8/2012. 53 s. https://nofima.brage.unit.no/nofima-xmlui/handle/11250/2504608
Uglem, I., Karlsen, Ø., Sanches-Jerez, P. & Sæther, B.S. (2014). Impacts of wild fishes attracted to open-cage salmonids in Norway. Aquacult. Environ. Interact. 6, 91-103. https://doi.org/10.3354/aei00112
Urquhart. K., Bowden, T.J., Buckett, B.E., Garcia, J., Fryer, R.J. & Ellis, A.E. (2009). Experimental study of the susceptibility of Atlatic cod, Gadus morhua (L.), to infection with an IPNV strain pathogenic for Atlatic salmon, Salmo salar L. Journal of Fish Diseases 32, 447-456. https://doi.org/10.1111/j.1365-2761.2009.01036.x
Vollset, K., Nilsen, F., Ellingsen, I., Karlsen, Ø., Paterson, R.A., Qviller, L., Skardhamar, J., Stige, L., Ugedal, O. & Lien V. (2023). Produksjonsområdebasert vurdering av lakselusindusert villfiskdødelighet i 2023. Rapport fra ekspertgruppe for vurdering av lusepåvirkning. 181 s.
Weli, S.C., Gjessing, M., Patel, S., Olsen, A.B. & Sandlund, N. (2025). Infectious pancreatic necrosis virus can be transmitted through cohabitation from Atlantic cod (Gadus morhua) to Atlantic cod, Atlantic salmon (Salmo salar), Atlantic halibut (Hippoglossus hippiglossus) and lumpfish (Cyclopterus lumpus). Journal of Fish Diseases 49, e70042. https://doi.org/10.1111/jfd.70042
Wiik-Nielsen, C.R., Løvoll , M., Sandlund, N., Faller, R., Wiik-Nielsen, J. & Bang Jensen, B. (2012). First detection of piscine reovirus (PRV) in marine fish species. Diseases of Aquatic Organisms 97, 255-258. https://doi.org/10.3354/dao02425
Wroblewski, J.S., Hiscock, H.W. & Bradbury, I.R. (1999). Fecundity of Atlantic cod (Gadus morhua) farmed for stock enhancement in Newfoundland bays. Aquaculture 171, 163-180. https://doi.org/10.1016/S0044-8486(98)00407-4