The Institute of Marine Research (IMR) is responsible for providing advice on the impact of anthropogenic sound and electromagnetic radiation on marine life. Such operations include scientific investigations, underwater explosions, seismic and electromagnetic surveys related to petroleum activity as well as offshore wind installations. This report intends to summarize available literature on how the above-mentioned sources impact marine life. Further, this knowledge is evaluated to show the reader the reasoning behind the advice given. This report includes one chapter on each of those sources (seismic, underwater explosions, electromagnetic investigations and offshore wind), again separated into subchapters of 1) review of scientific knowledge, 2) evaluation of knowledge and 3) advice. In addition, the report includes an overview of areas of vulnerable coral reefs in Norwegian waters. The reason for this is that many of the operations that produce sound involve placing various monitoring equipment on the sea bed, and these should not be placed in areas with corals. Hence spatial information about corals are relevant to include here. This report includes the advice from IMR for 2023 on the following topics: seismic surveys, electromagnetic surveys, underwater explosions and offshore wind farms. The advice for seismic and electromagnetics surveys are given as advisory maps; showing areas and periods to avoid for these activities. These maps are also part of the “meldesystem” of the Norwegian petroleum directorate.
Havforskningsinstituttets rådgivning for menneskeskapt støy i havet
— Kunnskapsgrunnlag, vurderinger og råd for 2023
Rapportserie:
Rapport fra havforskningen 2023-2
ISSN: 1893-4536
Publisert: 01.02.2023
Oppdatert: 24.02.2023
Prosjektnr: 14921
Forskningsgruppe(r):
Økosystemakustikk
Tema:
Seismikk,
Havvind,
Biologisk lyd
Program:
Nordsjøen
Research group leader(s):
Rolf Korneliussen (Økosystemakustikk)
Approved by:
Research Director(s):
Geir Huse
Program leader(s):
Henning Wehde
English summary
Sammendrag
Havforskningsinstituttet gir råd for alle operasjoner til havs som involverer bruk av lydkilder, samt elektromagnetiske kilder som kan påvirke marine økosystemer. Dette inkluderer vitenskapelige undersøkelser, utbygging og sprenging i sjø, seismiske og elektromagnetiske undersøkelser i forbindelse med petroleumsvirksomhet og utbygging og drift av vindkraftanlegg til havs. Denne rapporten er en sammenstilling av tilgjengelig kunnskap omkring hvordan de overnevnte typer av undersjøisk støy påvirker livet i havet. Videre evalueres denne kunnskapen i lys av hvilke konsekvenser dokumenterte effekter kan gi på økosystemet. Denne rapporten er inndelt med et kapittel for hver av de overnevnte støykildene (seismikk, elektromagnetiske undersøkelser, undersjøiske sprenginger og vindkraft til havs), som igjen er inndelt i delkapitler med 1) kunnskapsgrunnlag, 2) konsekvensvurdering og 3) råd. Rapporten inneholder konkrete råd fra Havforskningsinstituttet for 2023 for de ulike typene av støy. I tillegg inneholder rapporten en oversikt over hvor det finnes sårbare koraller i norske farvann. Årsaken til at dette er med er grunnet at mange av operasjonene til havs lager støy også innebærer å plassere ulike måleinstrumenter på bunn, og de bør ikke plasseres i områder med korall-forekomster. Derfor er det relevant å også ha med informasjon om romlig fordeling av koraller her. Rapporten inneholder konkrete råd fra Havforskningsinstituttet for 2023 for de ulike typene av støy og elektromagnetisk stråling. Råd for seismikk og elektromagnetiske undersøkelser gis i form av rådgivningskart, som viser perioder og områder som bør unngås. Disse kartene inngår også i Oljedirektoratets meldesystem.
Oppdatering 17.02.2023: I punktlisten i delkapittel 3.4.1, punkt 5, er ordet "frarådingsområdet" endret til "selve gyteområdet".
Oppdatering 24.02.2023: Forfatterlisten er oppdatert.
1 - Bakgrunn
Havforskningsinstituttet er rådgivende organ for alle operasjoner til havs hvor det brukes lydkilde som kan påvirke marine økosystemer. Herunder faller vitenskapelige undersøkelser, detonasjoner, utbygging til havs, seismiske og elektromagnetiske undersøkelser i forbindelse med petroleumsvirksomhet, samt utbygging og drift av vindkraftanlegg til havs. Havforskningsinstituttet er bedt om å dokumentere og sammenstille kunnskap om påvirkning, og basert på dette gi råd om slik påvirkning er bærekraftig eller kan føre til negative konsekvenser for økosystem og bestander (herunder dødelighet, skade og endret adferd hos marine dyr), mens Fiskeridirektoratet gir råd knyttet til konflikter med fiskeriaktivitet. Denne rapporten gjennomgår kunnskapsgrunnlag omkring effekter av de ulike støykildene på marint liv, samt en evaluering av alvorlighetsgrad og mulige konsekvenser som brukes som basis for de råd som gis. Havforskningsinstituttets råd for seismikk og elektromagnetiske undersøkelser blir gitt i form av rådgivningskart; kart som viser oversikt over hvilke områder som frarådes for disse aktivitetene i ulike perioder gjennom året. Disse kartene inngår i Oljedirektoratets meldesystem, slik at når selskapene melder inn eller planlegger en undersøkelse vil kunne se med en gang hvilke perioder det området de ønsker å undersøke vil medføre en fraråding fra Havforskningsinstituttet.
Kapittel 2 gir en introduksjon i mulige reguleringstiltak som har potensiale for å redusere påvirkningsgraden av støy på det marine økosystemet.
Mulige effekter av seismikk på marint liv og tilhørende råd for bruk av seismikk er beskrevet i kapittel 3, mens potensiell påvirkning av elektromagnetiske undersøkelser og råd for disse er behandlet i kapittel 4. De fleste råd fra Havforskningsinstituttet er i form av ulike områder som bør unngås til ulike tider på året. For hver 2-ukers periode gjennom året er det utarbeidet et kart over hvilke områder det rådes til å unngå å bruke seismikk eller elektromagnetiske undersøkelser. Disse kartene kalles Rådgivningskart, og aktuelle kart for 2023 er vist og forklart i Vedlegg I.
I forbindelse med oljeleting blir det i noen tilfeller også plassert utstyr på havbunnen, enten lyttekabler for seismikk eller i forbindelse med grunnundersøkelser uten seismikk. I slike tilfeller gir Havforskningsinstituttet også råd om det er særskilte hensyn som bør tas for å unngå skade på sensitiv bunnfauna, da i hovedsak koraller. Derfor inneholder også denne rapporten et avsnitt om korallrev og vernete områder på havbunnen (Vedlegg II).
I forbindelse med sprengningsarbeid til sjøs blir Havforskningsinstituttet også bedt om å levere råd om hvordan dette kan påvirke marine økosystem. Dette er gjennomgått i kapittel 5 med tilhørende råd. I forbindelse med konsesjonsrunder for utbygging av vindkraftanlegg til havs er Havforskningsinstituttet rådgivende organ. Her vurderes helhetlig påvirkning av en slik utbygging, herunder både støy og fysiske endringer av habitatet. Dette er helhetlig gjennomgått i en egen rapport (de Jong et al. 2020), mens den delen som omhandler støy fra vindkraftanlegg til havs med tilhørende råd er beskrevet i kapittel 6.
En annen lydkilde som ikke er behandlet i dette dokumentet er støy fra skipstrafikk. Dette utgjør en betydelig andel av den lavfrekvente, menneskeskapte støyen i havet, og denne ofte kontinuerlige støyen er påvist å påvirke adferd hos både fisk og sjøpattedyr. Denne typen støy blir imidlertid ikke regulert og forvaltet på tilsvarende måte som de overnevnte, og Havforskningsinstituttet har ikke noen rådgivende rolle. Av den grunn er skipstøy ikke behandlet som et eget tema i denne rapporten, men kunnskapen fra undersøkelser om denne typen lavfrekvent, kontinuerlig støy trekkes inn der den ansees som relevant. Havforskningsinstituttet har igangsatt aktivitet på å kartlegge og vurdere de ulike menneskelige påvirkninger fra de forskjellige sektorer (CoastRISK og BarentsRISK) hvor målsetningen er å lage et rammeverk (eller “risikokart”) for å identifisere den integrerte påvirkningen av de mest sentrale påvirkningsfaktorene, herunder støy, og utsatte områder og videre evaluere overvåkning og forvaltningstiltak for å redusere risiko. Her er også skipsstøy inkludert.
I denne rapporten behandles og rådgis ulike typer støy separat. Både på Havforskningsinstituttet og internasjonalt er det blitt økt fokus på den samlede belastningen av mange ulike typer menneskelig påvirkning på økosystemet de senere år, eks. Pirotta et al., 2022.
1.1 - Endringer fra 2022-2023
Rapporten blir oppdatert hvert år for å sikre at rådene våre tar hensyn til ny kunnskap. Her er en kort oppsummering om endringene i råd fra 2022 til 2023.
Rådene for alle de aktuelle støykildene er som før, med et unntak; for seismikk åpnes det opp for å komme nærmere gytefeltene enn 20/5 nautiske mil som har vært satt som buffersone. Dersom det skal gjøres et slikt unntak, pålegges seismikk selskapet å modellere i forkant at gytefeltet ikke eksponeres for lydeksponeringsnivå som overstiger 145 dB SEL over 10 sek. Dette skyldes ny forskning som viser at gytende fisk ikke skremmes bort fra gyteområdet sitt ved eksponering til seismikk på dette nivået.
Andre endringer er knyttet til oppdatering av rådgivningskartene. Disse er i stor grad knyttet til gytekart for fisk, da det i hovedsak frarådes å bruke seismikk i viktige gyteområder. Derfor gjennomføres hvert år et grundig arbeid for å oppdatere og forbedre eksisterende gytekart basert på ny kunnskap, samt vurdere om andre arter som tidligere ikke har vært inkludert i rådgivningskartene, bør inkluderes.
Prosjektet KINO II som har kartlagt gyteperioder for fisk i Nordsjøen har nå ferdigstilt sine resultater, og basert på dette har rådgivningsperiodene til bestandene Nordsjøtorsk og øyepål blitt endret. I tillegg har en ny art blitt inkludert; hvitting. Gytekart for kysttorsk er også oppdatert og disse endringene er gjort gjeldene i rådgivningskartene. Detaljer omkring dette finnes i Vedlegg 1.
Nytt for 2023 er også en tabell med hvilke arter som er med og ikke i frarådingskartene. Her er bestander av beinfisk og sjøpattedyr hvor HI har et bestandsansvar listet og vurdert basert på kommersiell viktighet, viktighet for økosystemet, sårbarhet (rødlistestatus) og mulighet til å negativt påvirkes av seismikk gjennom god hørsel, bruk av lyd eller dokumentert effekt av lydeksponering, og med tilhørende konklusjon om hvorvidt bestanden inkluderes i rådgivningskartene. Disse tabellene finnes i vedlegg 1.
1.2 - Referanser
de Jong, K., Steen, H., Forland, N. F., Wehde, H., Nyqvist, D., Palm, A.C.U., Nilssen, K.T., Albretsen, J., Falkenhaug, T., Biuw, M., Buhl- Mortensen, L. Sivle, L.D. (2020) Potensielle effekter av vindkraftanlegg på havmiljøet. Rapport fra Havforskningen 2020-42. ISSN:1893-4536. 42 pp.
Pirotta, E. Thomas, L., Costa, D.P., Hall, A.J., Harris, C.M., Harwood, J.,…& Tyack, P. (2022) Understanding the combined effect of multiple stressors: A new perspective on a longstanding challenge. Science of the Total Environment, 153322.
2 - Ulike typer av mulige reguleringstiltak
Det finnes mange måter å regulere og begrense støy i havet på. De vanligste er beskrevet i avsnittene under. Avbøtende tiltak er ofte en avveiing mellom å beskytte miljøet og å gjennomføre aktiviteten til havs. Hvilke tiltak som gir mest mulig utbytte av aktiviteten (eks. antall km med seismikk data) med minst mulig forstyrrelse av marint liv, burde vurderes i de ulike tilfeller og av ulike interessegrupper og myndigheter. Det er gjort forsøk på å veie økonomiske og miljøinteresser opp mot hverandre for å få et resultat som gir best mulig utnyttelse med minst mulig påvirkning, eks. med å beregne økonomisk utbytte/tap for seismikkdata for ulike scenarioer av områdebeskyttelse av hval i Arktis (Punt & Kaiser 2021) og ved å vekte påvirkning på naturen sammen med kostnader i tildeling av vindkraftkonsesjoner (
2.1 - Begrensninger av selve lydkilden
Det kan settes grenser for hvor kraftige lydkilder som kan brukes i undersøkelser. For seismiske kilder kan det være, for eksempel, begrensninger i maksimum tillatt kildenivå. Det kan også være tiltak for å erstatte lydkilden, for eksempel å bruke andre fundamenteringsteknikker enn pæling ved konstruksjoner.
2.2 - Verning av områder og perioder
Det kan settes restriksjoner for bruk av støykilder i spesifikke områder og perioder på året. Dette kan gjøres for å verne sensitive biologiske habitater, som for eksempel gyteområder for fisk eller viktige parings- eller beiteområder for sjøpattedyr.
2.3 - Ramp-up / Soft start
Dette innebærer at lydutsending starter på et lavt nivå og øker gradvis til operasjonell styrke. Det foregår typisk i 20–40 minutter. Hensikten er å sende en advarsel til dyr i området og gi dem tid til å søke bort fra kilden. Hovedformålet er å redusere fysisk skade ved å forhindre at dyr kommer nært nok til at dette kan skje.
Effekten av ramp-up er blitt testet eksperimentelt for arten knølhval, for både militære sonarer (Wensveen et al. 2017) og seismikk (Dunlop et al. 2016, Dunlop et al. 2020). Undersøkelsene viste en positiv, men noe begrenset effekt i den forstand at dyrene fikk en noe mindre total lyddose (SEL). Imidlertid ble det påpekt at ramp-up fungerer best for de artene som er mest følsomme for lyd fordi de oftere svømmer unna for å unngå eksponeringen. Basert på visuelle og akustiske observasjoner fra seismikkfartøy, konkluderte Stone (2015) med at ramp up var effektivt for sjøpattedyr, basert på at færre dyr ble observert ved bruk av ramp up, samt at flere dyr da ble observert å unnvike, hvilket er målet med denne prosedyren.
Modellstudier har vist at den risikoreduserende effekten av ramp-up avhenger av faktorer som dyrets responsterskel samt lydkildens og dyrets hastighet (von Benda-Beckmann et al. 2014).
For fisk er ramp-up blitt testet i en studie av Neo et al. (2016) som viste at dette ikke hadde noen effekt, med unntak av at det førte til mindre unnvikelse. Også i en studie med makrell som ble eksponert til gradvis økende nivå av seismikk, uteble respons til lyden (Sivle et al., 2017). Selv om begge disse studiene ble utført i oppdrettsmerder, noe som gir mindre mulighet for unnvikelse, kan resultatene tilsi at ramp-up kan føre til at fisk forblir i støyområdet i stedet for å forflytte seg på grunn av tilvenning til lyden.
Ramp up kan derfor virke som en relativt effektiv måte å redusere skade hos sjøpattedyr, da særlig sensitive arter. Ramp up kan muligens kan virke mot sin hensikt for fisk.
2.4 - Skadebegrensende (avbøtende) utstyr
Dette er tiltak som er beregnet å minimere fysisk skade på dyr i nærheten. Den vanligste typen er boblegardiner, som brukes både ved pæling og sprengninger under vann ( Verfuß 2014 , Grimsbø & Kvadsheim 2018). Tiltaket innebærer at man legger ut perforerte luftslanger rundt lydkilden. Luftboblene som stiger til overflaten, vil absorbere og spre tilbake deler av lydenergien fra aktiviteten. Det er også undersøkt om boblegardiner kan brukes for luftkanoner for å dempe de høyere frekvensene, over 200 Hz, som ikke er nyttige for seismikkundersøkelsen (Wehner & Landrø, 2020).
2.5 - Skadebegrensende (avbøtende) prosedyrer
Dette er prosedyrer for å forebygge at lyd skader eller forstyrrer dyr. En vanlig prosedyre er overvåkning av sjøpattedyr i en sone rundt fartøyet, som kan gjøres enten ved hjelp av sjøpattedyrobservatører (Marine Mammal Observer – MMO) eller passiv lytting etter vokalisering fra dyrene (Passive Acoustic Monitoring – PAM), som sørger for at det ikke befinner seg dyr i en viss radius fra lydkilden, som regel et seismikk-fartøy, enten ved oppstart eller under selve undersøkelsen. Disse tiltakene gjøres hovedsakelig for å hindre direkte skade på sjøpattedyr og begrenser seg derfor til en viss radius rundt fartøyet. Sikkerhetssoner kan ha flere nivåer, for eksempel total stopp av lydutsending hvis det befinner seg dyr tett ved kilden, redusert nivå om dyr på større avstander og kun registrering av observasjoner på lengre avstander.
Et annet eksempel er før-starts-undersøkelse, som innebærer typisk et transekt i oppstartsområdet i 30 minutter, hvor det speides etter sjøpattedyr. Dersom ingen dyr observeres, kan operasjonen starte.
2.6 - Begrensninger av eksponerte individer
Dette er reguleringskriterier som skal begrense hvor kraftig lyd dyr kan utsettes for. Det inkluderer blant annet å overvåke hvor mange dyr som har blitt eksponert for støy over et bestemt nivå. I en gitt undersøkelse er det kun lov å eksponere et gitt maksimum antall dyr for lydnivåer som overstiger dette nivået. Dette gjennomføres i praksis ved å telle dyr i det eksponerte området. Dette er hovedsakelig brukt for sjøpattedyr, hvor man har observatører (MMO) om bord som teller antall sjøpattedyr innenfor en radius hvor dette lydnivået antas å være oversteget.
2.7 - Bruk av avbøtende tiltak i Norge
Seismikkaktivitet er den av de menneskeskapte lydkildene som er mest utbredt, og som har den mest omfattende reguleringen i Norge. Tillatelse til å gjennomføre seismikkundersøkelser gis av Oljedirektoratet, men alle undersøkelser mottar råd både fra Havforskningsinstituttet og Fiskeridirektoratet.
I Norge har det mest brukte tiltaket vært verning av områder innenfor visse perioder (Sivle et al. 2021), og dette brukes både for seismikk, elektromagnetiske undersøkelser og sprenging i sjø. Seismikk har som følge av dette blitt frarådet i viktige gyteområder i gyteperioden samt i områder hvor det foregår viktig fiskeriaktivitet. I tillegg er det siden 2018 blitt lovpålagt å bruke ramp-up-prosedyrer ved oppstart av seismikkundersøkelser i enkelte områder. Norge er det eneste landet som etter vår kjennskap har regulering av seismikk knyttet til fisk og fiskerier.
Frem til 2018 hadde ikke Norge noen form for regulering av seismikk for sjøpattedyr. Men fra 2018 ble det lovpålagt å bruke ramp-up for å beskytte sjøpattedyr mot hørselsskade. Fra 2019 inkluderte Havforskningsinstituttet sjøpattedyr også i sine råd ved å fraråde seismikk i viktige beiteområder for bardehval.
For sprenging i sjø brukes også avbøtende utstyr, typisk boblegardin dersom dette er mulig.
2.8 - Referanser
Dunlop, R. A., McCauley, R. D., & Noad, M. J. (2020). Ships and air guns reduce social interactions in humpback whales at greater ranges than other behavioral impacts. Marine Pollution Bulletin, 154. doi:10.1016/j.marpolbul.2020.111072
Dunlop, R. A., Noad, M. J., McCauley, R. D., Kniest, E., Slade, R., Paton, D., & Cato, D. H. (2016). Response of humpback whales (Megaptera novaeangliae) to ramp-up of a small experimental air gun array. Marine Pollution Bulletin, 103(1-2), 72-83. doi:10.1016/j.marpolbul.2015.12.044
Grimsbø, E., Kvadsheim, P.H. (2018). Spregningsarbeider i sjø – effekter på marint liv og mulige tiltak.
Paper presented at the Fjellsprengningsdagen 2018.
Kavanagh, A. S., Nykanen, M., Hunt, W., Richardson, N., & Jessopp, M. J. (2019). Seismic surveys reduce cetacean sightings across a large marine ecosystem. Scientific Reports, 9. doi:10.1038/s41598-019-55500-4
Neo, Y. Y., Hubert, J., Bolle, L., Winter, H. V., ten Cate, C., & Slabbekoorn, H. (2016). Sound exposure changes European seabass behaviour in a large outdoor floating pen: Effects of temporal structure and a ramp-up procedure. Environmental Pollution 214, 26-34. doi:10.1016/j.envpol.2016.03.075
Punt, M. J., & Kaiser, B. A. (2021). Seismic shifts from regulations: Spatial trade-offs in marine mammals and the value of information from hydrocarbon seismic surveying. Environmental and Resource Economics, 80(3), 553-585.
Sivle, L.D., Forland, T.N., Hansen, R. R., Andersersson, M., Grimsbø, E., Linne, M. & Karlsen, H.E. (2017).
Behavioural effects of seismic dose escalation exposure on captive mackerel ( Scomber scombrus ). Rapport fra Havforskningen 34-2017. ISSN 1893-453.
Sivle, L.D., Vereide, E.H., deJong, K., Forland, T.N., Dalen, J. and Wehde, H. (2021). Effects of sound from seismic surveys on fish redroduction, the management case from Norway. Journal of Marine Science and Engeneering 9 (4): 436.
Stone, C. J. (2015a). Marine mammal observations during sources. Geophysical Research Letters, 31.
Verfuß (2014). Noise mitigation systems and low-noise installation technologies. In: Federal Maritime and Hydrographic Agency. In F. M. f. t. E. Nature Conservation and Nuclear Safety (Ed.), Ecological Research at the Offshore Windfarm alpha ventus. Wiesbaden: Springer Spektrum.
Von Benda-Beckmann, A. M., Wensveen, P. J., Kvadsheim, P. H., Lam, F. P. A., Miller, P. J. O., Tyack, P. L., & Ainslie, M. A. (2014). Modeling Effectiveness of Gradual Increases in Source Level to Mitigate Effects of Sonar on Marine Mammals. Conservation Biology, 28(1), 119-128. doi:10.1111/cobi.12162
Wehner, D., & Landrø, M. (2020). The impact of bubble curtains on seismic air-gun signatures and its high-frequency emission. Geophysics, 85, P1-P11. doi: https://doi.org/10.1190/geo2019-0451.1
Wensveen, P., Kvadsheim, P. Lam, F.P.A., von Benda-Beckman, A., Sivle., L., ….& Miller, P.J.O (2017). Lack of behavioural responses of humpback whales (Megaptera novaeangliae) indicate limited effectiveness of sonar mitigation. Journal of Experimental Biology, 220(22), 4150-4161. doi:10.1242/jeb.161232
3 - Seismikk
Seismiske undersøkelser, eller marine geofysiske undersøkelser, er den viktigste metoden for kartlegging og forståelse av geologien i havbunnen. Data om geologi og sediment-egenskapene nedover i havbunnen er avgjørende for å gjøre nye funn av olje og gass, men gjør det også mulig å få ut mer olje og gass fra allerede produserende felt. Derfor samles det inn seismiske data i ulike faser av utvinningsvirksomheten.
Enkelt beskrevet sendes lavfrekvente lydbølger fra en luftkanon ca. 5-15 m under havoverflaten ned i berggrunnen (Caldwell & Dragoset 2000, Prior et al. 2021). Kilden vil normalt generere kraftige, men korte lydsignaler med frekvensinnhold under 150 Hz ved hjelp av trykkluft som blir sluppet ut fra en luftkanon, eller fra en sammensetning av flere luftkanoner. Lydbølgene reflekteres mellom geologiske lag i berggrunnen, og ulike lag reflekteres forskjellig. De reflekterte lydbølgene blir fanget opp av mottakere (lyttekabler) som enten slepes etter seismikkskipet eller ligger på havbunnen. Rådataene blir så bearbeidet i datamodeller for å produsere bilder av berggrunnen som geologer tolker, slik at de kan få bedre kunnskaper om den geologiske strukturen, herunder olje- og gassforekomster.
Nedenfor gis det en kortfattet beskrivelse av ulike typer seismiske undersøkelser. Grundigere beskrivelser finnes i Gisiner (2016) og Birin & Maglić (2020).
3.1 - Ulike typer seismiske undersøkelser
3.1.1 - Borestedsundersøkelser
En borestedsundersøkelse (BSU, site survey) er en mindre seismisk undersøkelse som sammen med geoteknisk prøvetaking og topografisk kartlegging av bunnen utføres før en foretar prøveboringer. Luftkanonoppsettet er lite sammenlignet med de andre typene av seismiske undersøkelser, og det taues 1-4 lyttekabler med lengde på 100 til 1200 m. Det blir ofte brukt en liten (10 kubikktommer) kilde og en større som består av fire 40 kubikktommer (=160 kubikktommer) kilde som skyter annenhver gang omtrent hvert 5. sekund. Størrelsen på luftkanonen har betydning for frekvensinnholdet i signalet og dermed for oppløsningen til de seismiske dataene. Hovedformålet med undersøkelsen er å kartlegge de øverste delene av grunnen med tanke på eventuelle gasslommer som kan representere en sikkerhetsrisiko for prøveboringen.
3.1.2 - 2D-seismikk
Forskjellen på 2D, 3D, 4D og PRM -seismikk er lyttemetoden. Samme lydkilde, vanligvis flere luftkanon arrays med til sammen flere tusen kubikktommer volum, kan brukes ved disse metodene. Ved 2D-innsamlinger taues én lyttekabel. Lyttekabelen vil vanligvis være fra 4 til 12 km lang. Avstanden mellom kurslinjene kan være fra én til flere kilometer. Dette gir et todimensjonalt tverrsnitt av undergrunnen. En borestedsundersøkelse er ofte 2D-seismikk, men da med mindre volum på luftkanonoppsettet. Ellers blir 2D-innsamlinger brukt i letefasen og de dekker store områder. Kurslinjene kan være 100 – 1000 km lange.
3.1.3 - 3D-seismikk
Til forskjell fra 2D-undersøkelser sleper man her flere parallelle lyttekabler etter fartøyet, noe som gjør det mulig å avbilde undergrunnen i tre dimensjoner. Et seismikkfartøy som samler inn 3D-seismiske data, vil ha redusert manøvreringsevne som følge av det høye antallet lyttekabler (inntil 20 stk. med lengde 2 – 10 km, og avstand 50 – 150 m mellom hver kabel) etter fartøyet. 3D-seismikk blir brukt både i letefasen og i produksjonsfasen.
3.1.4 - 4D-seismikk
4D-seismikk er 3D-seismikk som gjentas i samme område med et visst tidsintervall, for eksempel en gang i året. Den fjerde dimensjonen er tid. Hovedmålsettingen med 4D-seismikk er å observere endringer i reservoaret over tid. Data fra slik seismikk gir informasjon om lokale trykkendringer i reservoaret og endringer i metning av henholdsvis olje, vann og gass i reservoaret. Denne informasjonen benyttes til å finne de beste posisjonene for nye brønner, slik at man kan optimalisere utvinningen fra feltet. 4D blir brukt i produksjonsfasen.
3.1.5 - Permanent reservoarovervåkning
Det er utviklet en type teknologi for 4D-seismikk der kabler eller noder med lyttesensorer er midlertidig plassert på, eller permanent gravd ned i havbunnen, og der det benyttes fartøy (kildefartøy) som kun sleper et luftkanonfelt under datainnsamling. Denne teknologien gir, som følge av at sensorene er fast posisjonert, seismikkdata med bedre kvalitet og bedre repeterbarhet enn det man får ved ordinær datainnsamling. I tillegg er denne metoden mer robust med hensyn til værforholdene. Siden det bare er luftkanonene som skal taues, tåles det mer bølger enn hva er tilfellet er når lyttekabler skal slepes. Slik teknologi betegnes som «Permanent Reservoir Monitoring – PRM». Det er økende bruk av slike metoder.
3.1.6 - Nye skytemønster og metoder
For ordinære seismiske undersøkelser er det mest vanlig å bruke to arrayer som blir avfyrt annenhver gang med avfyring hvert 10. sekund. De siste årene har lytte og signalbehandlingsmetodene blitt forbedret og dette har gjort det mulig å ta imot reflekterte signal kontinuerlig, og ved lavere signal til støy forhold. Dette har ført til nye metoder for avfyring der energien blir fordelt over hyppigere skudd med lavere støynivå. Dette blir gjort ved å dele luftkanon-kildene opp i mindre arrayer som blir avfyrt sekvensielt. Det kan for eksempel være tre arrayer med luftkanoner som blir avfyrt med henholdsvis 5.4 s, 5.4 s og 8.1 s mellom skuddene (Langhammer & Bennion, 2015). Eller seks eller flere luftkanonklynger som blir avfyrt hvert 1.8 sekund (Dhelie et al. 2018). Det totale antallet luftanoner er det samme som før, men det nye er at de blir gruppert i mindre grupper og avfyrt en gruppe av gangen med raskere skuddrate enn før. Den totale frigitte lydenergien over tid blir den samme som før. Det har også blitt mer vanlig å ha en egen båt som drar luftkanonene for å plassere de midt over lyttekablene som blir dradd av en annen båt, som for eksempel metoden kalt «TopSeis» (Vinje et al. 2017).
Enda mer kontinuerlige metoder er under utvikling. En metode kalt e-seismikk eller popkorn-seismikk går ut på å avfyre lufkanonene enkeltvis med svært kort tid (200 – 300 ms) mellom hver avfyring (Klüver og Høy, 2022, Hegna et al. 2018, Klüver et al. 2018, Abma & Ross, 2015) som resulterer i et tilnærmet kontinuerlig lydsignal med mye lavere spisstrykk enn for vanlig seismikk, men utsendt energi over tid er omtrent den samme som ved konvensjonell seismikk.
Det blir stadig strebet etter å få høyere signal-til-støy forhold for de laveste frekvensene, spesielt frekvenser lavere enn 7 Hz (Hegna og Parkes, 2011). En ny studie viser at en kan øke signalet for de lave frekvensene ved å plassere luftkanonen grunnere (Landrø og Amundsen, 2022). Dette er bare vist for en enkelt luftkanon.
3.1.7 - Andre lydkilder
En type kilde kalt «sparker» (“gnist”) er basert på en elektrisk utladning som lager en kort og kraftig akustisk puls. En elektrisk strøm går gjennom sjøvann mellom to eller flere elektroder. Den elektriske energien varmer opp sjøvannet ved elektrodene og resulterer i en boble av damp som utvider seg raskt og deretter kollapser. Dette lager lydpulsen. (English, 1991). Dette er en enkel og billig lydkilde som har mye svakere lyd enn luftkanoner og signalene når ikke like dypt ned i sedimentene. En sparker blir dermed bare brukt for kartlegging av grunne områder (mindre enn 1000 m), men har til gjengjeld høyere vertikal romlig oppløsning enn en får fra luftkanoner (Kluesner et al 2018, Wang et al. 2019). Det er vist at ved å kombinere en sparker med lyd i frekvensområdet 50- 500 Hz med luftkanoner kan en å få lyd med bredere frekvensbånd og dermed høyrere vertikal oppløsning (Yang et al. 2022).
En annen alternativ lydkilde til luftkanoner som kan brukes til seismiske undersøkelser er en marin vibrator (Laws et al. 2018). Dette er teknologi som har vært under utvikling siden 80-tallet og det har vært, og er fremdeles, flere parallelle utviklingsløp ulike steder (Pramik et al. 2015, Dellinger 2016, Feltham et al. 2017, Long & Tenghamn 2018, Oscarsson-Nagel et al. 2019). Til tross for det lange utviklingsforløpet er denne kilden ennå ikke tatt i bruk annet enn til uttesting av metoder. Den største utfordringen er å få nok utsendt energi ved lave frekvenser (under 16 Hz) og ulike metoder for å forbedre dette er i stadig utvikling (Morozov and Webb, 2022).
De fleste marine vibratorer er basert på et lukket gassfylt kammer der gassen blir utvidet og komprimert kontinuerlig slik at en membran som utgjør en eller alle veggene i kammeret vil vibrere frem og tilbake og dermed fungere som en høyttaler som setter vannet i bevegelse slik at det oppstår lydbølger. En kan kontrollere frekvensområdet og fasen, en fordel med det er at en bare sender ut det en har bruk for (i motsetning til en luftkanon som også lager mye lyd i frekvensområder som ikke blir brukt til datainnsamling).
En type marin vibrator, BASS (Gerez et al. 2020) er under utvikling i Norge. Denne kilden har et smalt frekvensbånd (typisk 0-150 Hz) og et typisk signal er et sveip på 10 sekunder som gjentas umiddelbart, slik at utsendingen blir kontinuerlig.
Fordelen med slike kontinuerlige lydsignaler er at de kan ha et lavere spisstrykk, og dermed mindre sannsynlig å medføre skade på hørselen til marine organismer. Frekvensbåndet på disse er smalere enn ved konvensjonell seismikk, men er innenfor frekvensområdet de fleste fisk kan høre. Imidlertid gir den kontinuerlige lyden lite rom for å lytte mellom signalene, og en bekymring er at slike derfor har større potensiale for maskering enn konvensjonell seismikk. Modelleringsarbeid viser likevel at fordi lydtrykket (SPL) er lavere, vil distansen som maskeringen skjer over være mindre; ~5 km for MV mot 10-50 km for luftkanoner (Matthews et al. 2020). Lignende resultat ble også funnet i Duncan et al. 2017.
Det er også testet ut å gjøre seismiske undersøkelser utenbruk av lydkilde, det vil si at seismikkfartøyet selv er lydkilden. Nyere forskning viser at det er mulig å bruke akustiske lydbølger generert av det seismiske fartøyet til å kartlegge hva som er under havoverflaten (Hegna 2021, Hegna 2022). Lydbølgene generert av fartøyet har mye lavere energi enn lydbølger generert fra tradisjonelle luftkanon arrayer, noe som begrenser hvor dypt under havbunnen man kan avbilde. Men disse lydbølgene dekker et bredt frekvensbånd. Reflektert fartøylyd fra havbunnen og lagene under kan registreres av hydrofonene i lyttekablene og brukes til å lage et bilde av havbunnen og de grunneste lagene under.
3.2 - Kunnskapsgrunnlag om effekter av seismikk på marine organismer
Her gjennomgår vi kunnskapsgrunnlaget for påvirkning av seismikk på fisk og sjøpattedyr.
3.2.1 - Direkte skade og dødelighet
Både fisk og sjøpattedyr har en velutviklet hørselssans, og lyd er svært viktig i mange økologiske funksjoner, som å finne mat, finne partner eller oppdage fiender. Fisk oppfatter partikkelbevegelsen i lyden ved hjelp av øresteiner (otolitter), som har høyere tetthet enn resten av fisken. Fisken, som har omtrent samme tetthet som vann, vil bevege seg med partikkelbevegelsen til lydbølgen i vannet, men den tyngre otolitten vil bevege seg tregere. Den relative bevegelsen mellom fisken og otolitten blir sanset av tynne hårceller (Popper & Hawkins, 2019). Sjøpattedyr har hørselsorganer som oppfatter lydtrykk, som hos mennesker. Både fisk og sjøpattedyr hører godt i frekvensspekteret til seismikk (ca. 10–200 Hz).
For begge grupper kan hørselen bli midlertidig svekket dersom lyden er kraftig nok. For sjøpattedyr er det også påvist kronisk hørselsskade, men det er uvisst om dette også er tilfelle for fisk. Kronisk hørselsskade, eller permanent terskelskifte (PTS), innebærer en uopprettelig svekkelse av hørselen, mens temporært terskelskifte (TTS) innebærer en midlertidig reduksjon. Slik redusert hørsel kan påvirke alle økologiske funksjoner og sammenhenger hvor lyd spiller en viktig rolle. Effektene på individer vil avhenge av skadeomfang og varighet og hvilket frekvensområde av hørselen som påvirkes.
Voksen fisk
Direkte skader fra kraftig lyd kan inkludere skade i svømmeblæren og leveren. Den plutselige endringen i trykket forårsaket av en impulsiv lyd kan i enkelte tilfeller føre til at svømmeblæren eksploderer inne i fisken (Halvorsen et al. 2012a,b). I en svært nøyaktig gjennomgang av hvordan forskjellige lydstyrker påfører skader, fant Halvorsen et al. (2012a, b) ut at selve lydstyrken er viktigere enn hvor lenge fisken er eksponert for lyden. Ved kortvarig eksponering kan imidlertid enkelte skader leges (Popper et al. 2016). I tillegg til direkte fysiske skader kan kraftig lyd påføre hørselstap som kan være midlertidig eller permanent. De fleste fisk hører bare lavfrekvent lyd (< 500 Hz), men noen arter, som sild og gullfisk, bruker svømmeblæren for å fange opp høyfrekvent lyd og viderefører den til øresteinene (otolittene). En direkte kanal gjør at svømmeblæren kan bli brukt for å videreføre lyd til otolittene. En slik kanal gjør fisk ekstra sårbare for hørselsskade (Halvorsen et al. 2013; Smith & Monroe 2016;). Det samme mønsteret er blitt vist også ved eksponering for seismikk (Popper et al. 2004, 2005).
70 artikler om påvirkning av seismikk på fisk og evertebrater som ble gjennomgått i Carroll et al. 2017 viste ingen resultat om dødelighet hos fisk som følge av seismikk. Nær luftkanonene kan det oppstå akustisk kavitasjon (Khodabandeloo & Landrø 2018) som kan føre til alvorlig skade eller død hos fisk (Cada et al. 2006). Kavitasjon skjer bare nær luftkanonene, så det vil uansett ikke ramme mange fisk.
Seismikkindusert hørselstap har vært påvist hos to arter av ferskvannsfisk etter kun fem skudd, men hørselen var tilbake til det normale etter 2–24 timer (Popper et al. 2005). Det er vist at flere skudd kan føre til mer langvarige skader på hårcellene i øret (McCauley et al. 2003), men denne studien trenger å repeteres for å verifiseres. For pæling, som også er en impulsiv lyd og derfor kan sammenlignes med seismikk, har grenseverdien for å unngå skader blitt satt til omtrent 179–181 dB re 1µPa²s SEL per slag, avhengig av antall slag (Halvorsen et al. 2011). Lydnivået fra et luftkanon array på 3590 in³ (59 liter) ble målt til 177 dB re 1µPa²s SEL ved avstand ca. 760 m fra kilden ved 758 m dyp og 63 m avstand i horisontal retning (Tashmukhambetov et al. 2008).
Disse studiene viser at det er mulig at fisk kan få hørselsskader fra kraftig lyd, men nivået som skal til, vil kreve at fisken befinner seg i umiddelbar nærhet til en luftkanon for å oppnå dette. I tillegg er alle de nevnte forsøkene gjort på fisk i tank eller akvarium, som dermed ikke har mulighet til å forflytte seg bort fra støykilden. Voksne fisk er mobile, og man kan anta at disse svømmer bort straks etter at lyden inntreffer, eller hvis lydkilden forflytter seg, før den kommer så nært at den induserer en hørselsskade. Direkte skade på voksen fisk anses derfor ikke som noe stort potensielt problem, og risikoen for populasjonseffekter på grunn av slike skader er minimal. Et mulig unntak kan være om fisken likevel blir i området grunnet store kostnader ved å flytte seg, og derfor blir værende til tross for at lydnivået kan påføre hørselsskade. Pena et al. (2013) viser at beitende sild ikke ble skremt vekk av en seismisk undersøkelse i nærheten. Det er også mulig at noen arter har en for stasjonær levemåte og dermed ikke vil forflytte seg fra området.
Fiskelarver og dyreplankton
Fisk i tidlige livsstadier (egg, larver og yngel) samt andre planktoniske organismer er potensielt utsatt for direkte skade fra seismikk da disse i ingen eller svært liten grad har mulighet til å forflytte seg bort fra luftkanonene. Det har vært utført et betydelig antall studier på egg og larver fra ulike arter (oppsummert i Dalen et al. 1996). Oppsummert framkommer det at dødelighet og skader i forbindelse med seismikkskyting kun forekommer i umiddelbar nærhet til luftkanonene. Høyeste dødelighetsrater og hyppigst forekommende skader ble funnet ut til ca. 1,5 m avstand. Lavere dødelighetsrater og mer sjeldent forekommende skader ble observert ut til ca. 5 m avstand.
Det er ikke gjort like mange studier på zooplankton, men en studie fra 2017 viser dødelighet hos flere arter av zooplankton på avstander ut til 1200 m (McCauley et al. 2017). Studien har svært få replikasjoner og unnlater også å undersøke andre mulige forklaringer på dødeligheten. En annen studie viste ingen dødelighet hos zooplankton 5 m fra en seismisk kilde (Fields et al., 2019). Et pågående prosjekt på Havforskningsinstituttet (ZoopSeis) har som målsetning å undersøke hvilke mekanismer og lydnivåer som forårsaker økt dødelighet hos ulike arter av dyreplankton.
Andre evertebrater
Evertebrater er den dyregruppen det har vært minst fokus på hvordan påvirkes av støy (Soto et al. 2016; Carroll et al. 2017), men de senere år har en rekke studier kommet til (Wale et al. 2021). Forskningen har konsentrert seg om kommersielle arter som krabbe og hummer (Kvadsheim et al. 2020; Wale et al. 2021). Men det er mange essensielle kunnskapshull; for det første er det fortsatt uklart hvordan evertebrater hører lyd. Det har vært antatt at de bruker statocystene (balanseorgan) til å fange opp vibrasjoner, men et nyere studie viser at krepsdyr muligens er sensitive til både partikkelakselerasjon og lydtrykk (Radford et al. 2022). Day et al 2022 har vist for juvenile hummer (Jasus edwardsii) at responsen som gjør at hummer snur seg tilbake hvis de havner på rygg ble ødelagt, og tiden mellom skallskifte økte etter eksponering for en seismisk undersøkelse på nært hold (< 500m), Juvenile som ble eksponert på 500 m avstand fikk tilbake responsen der de snur seg hvis de havner på rygg etter noen uker og tiden mellom skallskifte endret seg ikke. En krabbeart (Neohelice granulate) i Argentina som ble eksponert for lydsveip i frekvensbåndet 2.5 – 25 kHz viste økte konsentrasjoner av glukose og laktat, noe som kan indikere økt stressnivå som følge av lydeksponeringen (Filiciotto et al. 2016). Det har derimot blitt funnet minimal effekt på fangst av snøkrabbe i Canada (Morris et al. 2018) og overlevelse og stress hos en hummerart (Jasus edwardsii) i Australia (Fitzgibbon et al. 2017).
Mange evertebrater lever i bunnsedimenter og er svært viktig i økosystemet ved å mikse sedimentlagene som er nødvendig for å unngå opphoping av organisk materiale. Solan et al. (2016) studerte 3 arter av slike bunndyr; en musling (Ruditapes philippinarum), en sjøkreps (Nephoros norvegicus) og en slangestjerne (Amphiura filiformis) som ble eksponert til kontinuerlig eller impulsiv lyd over 7 dager. Ingen av disse lydene påvirket konsentrasjoner av fysiologiske stressfaktorer (i form av økt laktat og glukose). Imidlertid viste sjøkrepsen redusert aktivitet, mens muslingen reduserte sin overflateaktivitet og holdt seg lukket istedenfor å aktivt filtrere overflatelaget. Om denne typen reaksjoner vedvarer og omfanget er stort, vil det kunne medføre redusert miksing av substrat og videre mindre resirkulering av næringsstoffer i økosystemet. Et annet bunnlevende bløtdyr, kamskjell (Pecten fumatus og Mimachlamys asperrima) viste derimot ingen endring i dødelighet, muskelskade eller redusert kjøttkvalitet etter eksponering til seismikk i Australia (Przeslawski et al. 2018).
Sjøpattedyr
Midlertidig hørselstap er undersøkt i eksperimentelle studier på sel, delfiner og nise i fangenskap (Kastak et al. 1999; Finneran et al. 2003, 2010, 2015; Schlundt et al. 2000; Kastelein et al. 2012a, 2012b, 2013). Generelt viser disse studiene at det er lydenergien og varigheten av eksponeringen som er avgjørende for graden av hørselstap. For pulset lyd kan også enkeltpulsers egenskaper spille en stor rolle. Kortvarige lydpulser med rask stigetid og høy amplitude – egenskaper som er typiske for seismikk – har spesielt stor skadevirkning på øret hos pattedyr. Et nyere studie på seismikkindusert hørselstap hos nise indikerer imidlertid at det er mulig at noen arter kan begrense skade på egen hørsel (self mitigation) dersom lyden er forutsigbar (Kastelein et al. 2020).
Eksponeringskriterier for når ulike grupper av sjøpattedyr kan bli utsatt for hørselstap ble oppdatert i 2019 (Southall et al. 2019), og ytterligere for “veldig høyfrekvente hvaler” (hovedsakelig nise) og sel i 2022 (Tougaard et al. 2022). Terskelverdi for impulser som seismikk, oppgis både som spisstrykk (SPL dB re 1 µPa) og som total akkumulert energi over hele eksponeringen (innenfor 24 timer) (SEL dB re 1 µPa^2 ·s) (Tabell 1). Kriteriet tilsier at hørselskade vil oppstå dersom dyret utsettes for lyd som oppfyller ett av de to kriteriene (akkumulert energi eller spisstrykk). SEL-verdiene vektes i henhold til funksjoner som beskriver generiske hørselskurver for grupper av sjøpattedyr. Spisstrykk-verdien vektes ikke, fordi skade som følger av høye spisstrykkverdier ikke forventes å være særlig frekvensavhengig da de fører til mer direkte mekanisk påvirkning på hele det indre øret.
| Impulsive lydkilder | ||||
|---|---|---|---|---|
| Funksjonell hørselsgruppe |
TTS Terkselverdi |
PTS Terskelverdi |
||
|
SEL (Vektet) |
Spisstrykk SPL |
SEL (vektet) |
Spisstrykk SPL |
|
| Lavfrekvente hvaler | 168 | 213 | 183 | 219 |
| Høyfrekvente hvaler | 170 | 224 | 185 | 230 |
| Veldig høyfrekvente hvaler | 140 | 196 | 155 | 202 |
| Seler | 170 | 212 | 185 | 218 |
| Andre marine rovdyr | 188 | 226 | 203 | 232 |
Hos tannhvaler, som bruker ekkolokasjon i matsøk og til å orientere seg, vil nedsatt hørsel direkte svekke evnen til å finne mat, noe som vil få umiddelbare konsekvenser for overlevelsen. Nedsatt hørsel vil også redusere mulighetene til å kommunisere med andre individer. Mange sjøpattedyr lever i tette grupper, og nedsatt hørsel kan hemme flokkens kommunikasjonsevne og redusere den viktige mor–kalv-kontakten eller muligheten for å høre parrings-rop fra potensielle partnere.
Nise anses som den mest sensitive enkeltarten når det gjelder skadeeffekter. Det er også den arten som det finnes mest kunnskap om, både om hørsel og skadeeffekter (for eksempel Tougaard et al. 2015). Først og fremst har nise den laveste terskelverdien for midlertidig hørselstap som er målt for noen art. Nise er en av de mest tallrike artene av sjøpattedyr i norske farvann, men oppholder seg hovedsakelig i kystnære områder og ikke så mye til havs, hvor seismikkundersøkelser hovedsakelig foregår.
Som for fisk må også sjøpattedyr oppholde seg i nærheten av kilden for å risikere hørselstap, og man antar at dyrene vil svømme unna kilden dersom de har mulighet til det.
3.2.2 - Adferdsresponser
Studier av effekter av lyd på marine dyr viser at adferdsresponser er den vanligste reaksjonen (Duarte et al. 2021). Adferdsresponser er endringer i dyrenes oppførsel som følge av lydeksponeringen. Dette innebærer f.eks. unnvikelse av det eksponerte området, reduksjon eller opphold av viktige aktiviteter som beiting eller gyting, samt endring i hvordan dyrene selv lager lyd. For skader som følge av lyd, finnes det eksempler i litteraturen både for fisk og sjøpattedyr som sier noe om hvilke terskelverdier av lydnivåer skadene mest sannsynlig inntreffer ved (Southall et al. 2019; Popper et al. 2014; NOAA 2015), men tilsvarende terskelverdier for adferdsendringer er det svært lite kunnskap om. Slike terskelverdier for adferdsendringer vil sannsynligvis også ha langt større variasjon mellom arter enn det som er tilfelle for hørselsskader. Også innad i en art er det store variasjoner mellom ulike individer, mellom ulike livsstadier, ulike habitater og hvilken adferdsmodus dyret er i (for eksempel beiting, vandring, reproduksjon), samt dets fysiologiske tilstand (kondisjon) (de Robertis & Handegard 2013). Skader og hørselstap begrenser seg til nærområdet for lydkilden, mens adferdsresponser kan potensielt inntreffe overalt hvor dyrene hører lyden, og området hvor dette kan inntreffe, er derfor langt større, med tilsvarende mulighet for å påvirke mange flere individer (Slabbekoorn et al. 2010).
Fisk
Menneskeskapt lyd generelt kan påvirke fiskens forflytning (Engås et al. 1996), beiting; (Løkkeborg et al. 2012; Sabet et al. 2015), anti-predator adferd (Simpson et al. 2016), gyting (Sierra-Flores et al. 2015; de Jong et al. 2018a, 2020; Amorim et al 2022) og evne til å ta vare på avkom (Picciulin et al. 2010; Nedelec et al. 2017, 2022. Mange fiskearter bruker lyd til å kommunisere med artsfrender under gyting og beiting, og støypåvirkning vil kunne endre både kommunikasjon og adferd under reproduksjon (Picciulin et al. 2010; de Jong et al. 2018a, b). Med unntak av de to førstnevnte har disse studier imidlertid sett på effekter av mer kontinuerlig lyd som skipsstøy. Det er visst at lyd fra seismikk mulig kan bidra til maskering av kommunikasjon opptil 18 km fra kilden (Rogers et al. 2021).
Bunnfisk, som ulike arter av torskefisk og uer, kan reagere på seismikk med både horisontal og vertikal forflytning (Chapman and Hawkins, 1969, Pearson et al. 1992; Skalski et al. 1992; Engås et al. 1996, Paxton et al. 2017; Davidsen et al., 2019, van der Knaap et al., 2021), og i tillegg kan beitemotivasjonen reduseres (Skalski et al. 1992; Løkkeborg et al. 2012). Eksempelvis viste Engås et al. (1996) at mengde torsk og hyse målt med ekkolodd og ved fiskeri en nedgang på opptil 70% etter seismikk skyting sammenliknet med før seismikk-undersøkelsen startet, og Paxton et al. (2017) at mengde fisk på et tropisk rev filmet med video en nedgang på 78% etter en seismisk undersøkelse (Paxton et al. 2017).
Andre studier har rapportert om svært liten eller ingen reaksjon til seismikk hos bunnfisk. Meekaen et al. (2021) viste ingen endring i hverken sammensetning eller mengde av tropiske fisk i Australia hverken på kort (dager) eller lang (måneder) sikt som følge av seismikk-eksponering. Wardle et al. (2001) viste at det var få reaksjoner og ingen reduksjon i fiskemengde på et rev av svært stedbundne fisk i respons til lydeksponering fra luftkanoner i skotske farvann. Også andre studier viste at seismikk ikke påvirket artsfordelingen og fiskemengden på et rev og et bunnfisksamfunn (Miller & Cripps, 2013; McCauley et al. 2021).
I et eksponeringsforsøk med en fullskala seismikk-kilde i Nederland viste resultatene at torsk ikke forlot området under eksponeringen. Imidlertid ble det påvist noen mer subtile endringer i aktivitetsmønstre; torsken ble mer inaktiv under eksponering, samt at døgnmønsteret endret seg (van der Knaap et al., 2021). Endringer i aktivitetsnivå ble også påvist hos torsk i merd, men med stor individuell variasjon (Hubert et al., 2020). I tilsvarende forsøk på torsk og sei i merd i Norge, fant man noe redusert hjertefrekvens under eksponering til en luftkanon (Davidsen et al. 2019). Dette kan indikere en reaksjon tilsvarende en «startle respons». Imidlertid var alle reaksjoner kortvarige og minsket over tid, noe som tyder på en viss tilvenning til lyden (Davidsen et al., 2019). Et nytt studie i Norge med merket, frittsvømmende torsk i et gyteområde viste at gytende torsk ikke forlot gyteområdet sitt under eksponering til seismikk med lydnivå tilsvarende en undersøkelse et stykke unna (lydnivå opp til SEL på 145 dB re 1 µPa s-1) (McQueen et al. 2022).
I et studie med tobis, viste videoopptak en viss grad av irregulær svømming og noe alarmrespons under eksponering til seismikk, men det ble ikke påvist at den gravde seg ned i sanden, som var hypotesen i forkant av undersøkelsen (Hassel et al. 2004).
For pelagisk fisk er det dokumentert i studier av fisk i fangenskap at både makrell og andre arter som ikke finnes i norske farvann, reagerer på lavfrekvent lyd og seismikkpulser ved å øke svømmehastigheten og endre stimstrukturen til å gå tettere sammen (Sivle et al. 2016; Fewtrell & McCauley 2012). Endring i stimadferd ble også påvist hos en karuss som lever i Middelhavet (Sparus aurata), som responderte til seismikk ved å stime tettere (Mauro et al., 2020). Makrell viste imidlertid ikke slike umiddelbare reaksjoner ved en gradvis opptrapping av lydnivået til en ekte seismisk luftkanon (Sivle et al. 2017), noe som tyder på at måten lyden presenteres på, og eventuelt tilvenning, kan påvirke hvor sterk en respons er. I en studie under en reell seismikkundersøkelse studerte Slotte et al. (2004) horisontal og vertikal fordeling av sild og kolmule. Studien fant at kolmule forflyttet seg noe dypere i vannsøylen under seismikkeksponering, mens det ikke ble observert noen endring hos sild. Beitende sild viste ikke noen adferdsendring hverken ved horisontal eller vertikal forflytning eller endring i svømmehastighet og retning under seismikkeksponering (Pena et al. 2013). Det påpekes at dette kan skyldes den sterke motivasjonen til å beite, og at studien ble gjennomført etter at seismikkundersøkelsen hadde pågått en tid, slik at silda kan ha vent seg til lyden. Lignende tilvenning ble også vist i en akvariums-studie hvor stressresponsen hos havabbor forsvant etter noen uker med tilbakespilling av seismikk (Radford et al. 2016). Fordi den tilsynelatende tilvenningen kan være en følge av hørselsskade (Popper et al. 2009), kunne man ikke konkludere med at fisken ikke var påvirket av seismikk lenger. En studie av pelagisk fisk rundt et vindkraft-anlegg i Nordsjøen registrerte med ekkolodd færre, men tettere stimer under en seismikk-undersøkelse sammenliknet med før undersøkelsen startet, men samme typen endringer skjedde i kontroll området (Kok et al., 2021). Noen svakheter med dette studiet er imidlertid at det ikke oppgir hvilke arter av pelagisk fisk som er registret, samt at mangel på replikanter gjør det vanskelig å stadfeste om dette er en reell reaksjon til seismikken eller skyldes naturlig høy variasjon i stimdynamikk. Ut ifra de studier som er gjort på pelagisk fisk, kan det derfor tyde på at reaksjoner i form av endret stimstruktur og unnvikelse kan forventes, men at responsen sannsynligvis minsker over tid grunnet tilvenning.
Variasjonen i responser mellom studier og arter understreker viktigheten med å vurdere adferdseffekter i lys av forskjeller i habitat, levesett, art, adferdsmodus og hvordan lyden oppfattes og hva den forbindes med.
Kun et fåtall studier gir informasjon om omfanget av påvirkning i tid og rom. For torsk og hyse er det påvist unnvikelse ut til minimum 33 km (18 nautiske mil), med varighet på minimum fem dager etter avsluttet seismikkskyting (Engås et al. 1996). Dette resultatet støttes også av tidligere fangstforsøk (Soldal & Løkkeborg 1993). I studiet med torsk i Nederland var avstanden mellom fisk og seismikk kilden på det nærmeste 2,25 km (van der Knaap et al., 2021). Tobis ble eksponert for seismikk ved en avstand på 5 km fra luftkanonene (Hassel et al. 2004). Alle disse studiene brukte oppsett som er vanlige i seismiske undersøkelser.
Sjøpattedyr
Norske farvann er svært viktige som beite- og yngleområder for mange arter av sel og hval. Til tross for dette er det, med ett unntak av en studie på en spermhval (Madsen et al. 2002, resultater beskrevet lengre nede) ikke gjort noen norske studier på effekten av seismikk på adferd hos sjøpattedyr, men det er gjort utenlandske studier av enkelte av artene som finnes i norske farvann.
For sel viste en studie fra Alaska ingen endring i mengde sel i en sone på 250 m rundt et seismikkfartøy før og under aktiv skyting (Harris et al. 2001), men selene viste unnvikelse på avstander nærmere enn 150 m. En annen undersøkelse (Thompson et al. 1998) rapporterer at selartene havert og steinkobbe viste unnvikelse og nedsatt beiteaktivitet som respons på eksponering for seismikk.
Av tannhvalene er nise den arten som er best studert, og den har vist seg å reagere med unnvikelse når nivået oversteg 45 dB over høreterskelen (Tougaard et al. 2015). Reaksjoner ved relativt lave nivåer ble også vist av Thompson et al. (2013), men unnvikelse 5–10 km fra en seismisk kilde, og tilsvarende SPL og SEL på henholdsvis 165–172 dB re 1 µPa og 145–151 dB re 1 µPa2 s. Nisene returnerte imidlertid raskt tilbake etter eksponeringen var avsluttet og viste også tilvenning til lyden etter flere dagers eksponering. Pirotta et al. (2014) viste også at beiteadferden til nise gikk ned opptil 40 km unna en seismikkundersøkelse (SEL fra 130–165 dB re 1µPa2s). En nyere undersøkelse i Nordsjøen viste nedgang i akustisk deteksjon av niser på en rekkevidde av 8-12 km fra en fullskala seismisk undersøkelse, noe som indikerer enten at nisene forflytter seg unna, eller at de stopper å ekkolokalisere (Sarnocinska et al. 2020). Begge deler innebærer imidlertid en adferdsreaksjon, enten i form av unnvikelse, eller avsluttet beiteaktivitet/sosial adferd. Niser er imidlertid mest følsomme for høyere frekvenser, så seismikk-kilder med reduksjon av høye frekvenser er effektivt for å redusere adferdsresponser (Kastelein et al. 2019)
En annen tannhval som er studert, spermhval, viste ingen unnvikelse eller endring i dykkeadferd til fullskala seismiske undersøkelser hverken i Mexico (Miller et al. 2009) eller i Norge (Madsen et al. 2002).
Narhvalen, en art med et begrenset leveområde og vandringsrute, i tillegg til å være en dypdykkene art, ansees som særlig sårbar for menneskelig støy. Etter at Heide-Jørgensen (2013) viste at flere tilfeller av narhval som var blitt fanget i isen muligens skyltes seismikk, har denne arten vært fokus for flere studier som viser både fysiologiske og adferdsmessige responser. Forsøk med merkede dyr viste at narhvalen under eksponering til seismikk (opp til 152dB re 1µPa m) hadde en markant reduksjon i hjerteslagsfrekvens (baracardia) til tross for økt haleslagsfrekvens og aktivitet under dykk (Williams et al. 2022). Narhvalen viste også unnvikelse og økt svømmehastighet når den ble eksponert for seismikk, men også på skipet når det ikke skjøt seismikk (Heide-Jørgensen et al. 2021). Videre viste Teryo et al (2021) at narhvalen reagerte ved å avbryte beiteaktivitet under eksponering både av fartøy og seismikk på avstander på over 20 km, og ved lydnivåer som ikke overseg bakgrunnsstøyen. Dette tyder på at arten er særdeles følsom og kan skille ut lyd selv i bakgrunnsstøyen.
Bardehvaler bruker lavfrekvent lyd til å kommunisere over store avstander. Seismiske pulser inneholder mye energi i det samme frekvensbåndet, og seismikk vil derfor kunne maskere kommunikasjonen. Studier har vist at både knølhval (Cerchio et al. 2014), finnhval (Castellote et al. 2012), blåhval (Di Orio & Clark 2010) og grønlandshval (Blackwell et al. 2015) endrer vokaliseringen sin under seismiske undersøkelser. Grønlandshval reagerer for eksempel med å øke hyppigheten av vokaliseringen for å kompensere for økt støynivå, men bare dersom støyen er moderat (< 94 dB SEL re 1μPa2s) (Blackwell et al. 2015), mens de reduserer vokaliseringen dersom støyen øker (> 127 dB SEL re 1μPa2s), og blir helt stille ved nivåer over 160 dB SEL re 1μPa2s (Blackwell et al. 2015). En nyere studie viser også at grønlandshvalen kompenserer ved å øke styrken (Lombard effekt) på vokaliseringen sin med 10-15 dB når eksponert for svak (lengre borte) seismikkstøy, mens når støyen øker, har den ikke kapasitet til å øke sin styrke lengre (Thode et al. 2020). Finnhval kan endre både frekvens og båndbredde under seismikkeksponering (Castellote et al. 2012), slik at de vokaliserer på andre frekvenser enn seismikken bruker. Blåhvalen økte vokaliseringen signifikant under seismikkeksponering i et område hvor den hovedsakelig beiter og sosialiserer (Di Iorio & Clark, 2010), men det rapporteres at langdistansevokalisering hos den samme arten reduseres ved seismikkeksponering (Clark & Gagnon 2006). Grunnen kan være at det for dyr som kommuniserer på nært hold, kan være viktig å opprettholde kontinuerlig kontakt, mens det under langdistansesignalisering til mottakere som er langt unna, ikke betyr så mye med mindre avbrudd i kontakten. Dette viser at konteksten er av stor betydning, både for hvordan dyrene reagerer, og for hvordan de påvirkes (alvorlighetsgraden).
I Australia har en stor studie av hvordan knølhval reagerer på seismikk (BHRASS), ikke vist noen store responser (Dunlop et al. 2015, Dunlop et al. 2016), hverken i form av unnvikelse, svømmehastighet eller gruppestruktur i et vandringsområde. I en studie hvor de sammenlignet adferd under skyting med adferd under en kontroll hvor skipet var i nærheten uten å skyte seismikk, reagerte knølhval med 50 % sannsynlighet først ved lydnivåer på 150–155 dB re 1 μPa2·s og når de befant seg innenfor 2.5 km fra kilden (Dunlop et al. 2018). Under vandring mot parringsområdet, påviste Dunlop et al. (2020) at sannsynligheten for interaksjoner mellom ulike grupper, som er viktig adferd som leder opp til parring, ble sterk redusert som følge av tilstedeværelse av seismiske fartøy, og at mor-kalv par sinket farten og i noen tilfeller avventet å passere inntil fartøyet var borte. Dette var imidlertid tilfelle også når luftkanonen var avslått, noe som indikerer at selve fartøyet forstyrrer. Videre har Dunlop et al. (2021) brukt resultatene for å estimere mulige populasjonseffekter av seismikkundersøkelser på knølhval på vandring. Ved å bruke PCoD (Population Consecvences of Disturbance) modellen, beregnet de hvordan observerte unnvikelser kunne føre til forsinket vandring for mor-kalv par og mulige konsekvenser av dette på energitap; for voksne hunner til å produsere melk til kalvene, og for kalven til å overleve uten morsmelk i områder med forstyrrelse fra seismikk. Resultatene viste svært lav sannsynlighet for noen populasjonseffekt som følge av en seismisk undersøkelse, både for voksne hunner og for kalvene. Resultatene viser at den observerte unnvikelse ikke vil gi mer en maksimalt 24 t forsinkelse på vandringen, og det energetisk tapet (som følge av økt forbruk ved å vandre mer, samt tapt tid i beiteområde) vil være neglisjerbart. For diende kalver, vil selv et «worst-case-scenario», hvor en antar at mor og kalv blir værende i et eksponert område i over 48 t, og at dette fører til at kalven ikke får melk i denne perioden, vil dette sannsynligvis gi en relativt liten populasjonseffekt.
I motsetning til knølhval, har grønlandshval vist relativt kraftig unnvikelse fra seismiske kilder som er nærmere enn 8–10 km (Richardson et al. 1986; Robertson et al. 2013). I tillegg viser de endret dykkeadferd og respirasjonsrate ca. 8 km fra seismikk-kilden (Richardson et al. 1986; Robertson et al. 2016). Finnhval, en vanlig art i våre farvann, er også påvist å unnvike seismikk-kilden (Castellote et al. 2012). Imidlertid var dette under parring, som foregår i tropiske og subtropiske strøk, mens finnhvalen beiter i norske farvann. Det er derfor usikkert hvor overførbart dette resultatet er. Et nyere studie i Atlanterhavet viser at andel visuelle observasjoner av sjøpattedyr på aktive seismikkfartøy sammenliknet med kontrollfartøy var henholdsvis 88% og 53% færre under seismikk for henholdsvis bardehval og tannhval (Kavanagh et al. 2019), noe som kan indikere at sjøpattedyrene svømmer bort fra seismikkfartøyene.
Det er som nevnt ikke gjort studier av adferdsendringer som følge av seismikk på sjøpattedyr i Norge, utenom et studie på spermhval (Madsen et al. 2002), men det har vært gjort relativt omfattende undersøkelser på en annen kraftig, pulset lydkilde; militær sonar. Disse studiene har dokumentert effekter som unnvikelse av kilden, avbrutt beiteadferd og endret vokalisering (for eksempel Miller et al. 2012; Sivle et al. 2015; Harris et al. 2016). Slike sonarer opererer med høyere frekvenser enn luftkanoner (vanligvis 1–7 kHz), men begge typer signaler er godt hørbare for alle sjøpattedyr. Disse undersøkelsene viser også at det er stor variasjon mellom ulike arter av sjøpattedyr i hvor sterkt de reagerer, som ikke kan forklares ut i fra hørselsterskel eller taxonomi (se Kvadsheim et al. 2020 for utdypende forklaring).
Ofte brukes såkalt dose-respons studier for å vise ved hvilke lydterskler en gitt adferdsrespons er sannsynlig å inntreffe, som f.eks. av Miller et al. (2014) for spekkhogger. Imidertid er variasjonen ofte svært stor mellom individer ut ifra kontekst, leveområde, livshistorie at selv innad i en populasjon som responderer til et spesifikt signal, og Thomas og Tyack (2019) viste effekten kan underestimeres med så mye som 280 ganger for dose-respons funksjonen for spekkhogger. Slike enten-eller kriterier som er ment for å relatere eksponeringsnivåer for enkelt-hendelser og en adferdsrespons over en større taxonomisk gruppe og lydtype kan derfor medføre store feilkilder i å forutsi effekter, og er derfor ikke anbefalt å bruke uten en svært grundig evaluering (Southall et al. 2021).
Nyere studier tyder imidlertid på at responsterskel mer sannsynlig kan forklares ved «risk disturbance» hypotesen; at lyden oppfattes som en trussel som trigger anti-predator adferd, og at arter og populasjoner med høyt predasjonspress derfor er forventet å reagere sterkest (Miller et al. 2019).
3.2.3 - Kunnskapsgrunnlag for seismikk i nordområdene
I dag er kun den sørligste delen av Barentshavet åpen for kommersiell seismikk. Imidlertid foregår det til tider også noe seismikk i ikke-åpnede områder, enten i regi av Oljedirektoratet eller for ulike forskningsformål foretatt av forskningsinstitusjoner. Økosystemet i Barentshavet er sannsynligvis også det av våre havområder som er underlagt den største økningen i press fra ytre påvirkning som seismikk, skipstrafikk og fiskeri, og fra klimaendringer. Alle disse faktorene påvirker både leveforhold og utbredelse av dyrene som finnes her. Barentshavet er et grunt hav med store bankområder og er karakterisert av varme, atlantiske vannmasser i sør og kalde, polare vannmasser i nord som møtes langs den produktive polarfronten. Fordelingen av vannmassene har stor betydning for fordelingen av dyr på alle trofiske nivå i økosystemet. Det nordlige Barentshavet har et arktisk økosystem og er avhengig av isdannelse om vinteren. Sjøisdekket i dette området har vært rekordlavt de siste årene på grunn av økt innstrømming av atlantiske vannmasser og minsket innstrømming av sjøis fra nord (Lind et al. 2016). Disse to faktorene ligger til grunn for at Barentshavet er et av de områdene i verden som varmes opp raskest (Screen & Simmonds 2010; Comiso og Hall 2014; Lind et al. 2016). Redusert sjøisdekning forventes til å føre til økt skipstrafikk i omradet (Meld. St. 20 (2014–2015)). Dette kan medføre en økning i spesielt kontinuerlig støy, som kan påvirke fiskens reproduksjon og predatorunnvikelse (Simpson 2016, de Jong et al. 2018a, Blom et al. 2019, de Jong et al. 2020), men det har også blitt åpnet flere områder for seismikk i de siste årene.
I Klima- og miljødepartementets forvaltningsplan (Meld. St. 20 (2019-2020)) er flere områder i Barentshavet definert som særlig verdifulle og sårbare (SVO). I 2021 er det foretatt en harmonisering av SVO-vurderinger på tvers av havområdene og en helhetlig gjennomgang av alle SVO-ene for blant annet å synliggjøre årsak til verdi og sårbarhet (Eriksen et al. 2021). I denne prosessen ble det laget et forslag for SVO-områder i Barentshavet. Barentshavet er oppvekstområde for mange fiskearter, som sild, torsk og lodde. I tillegg til å være blant Norges viktigste fiskeriressurser er disse fiskeartene også en del av næringskjeden som fôrer verdens høyeste tettheter av sjøfugl. Store deler av havet nær norskekysten og rundt Bjørnøya er vernet (Meld. St. 20 (2019-2020)), mens området mellom disse per i dag er åpnet for kommersiell seismikk.
Barentshavet er også tilholdssted for mange arter av spesiell interesse for norsk og europeisk artsmangfold. Dette gjelder grønlandshval, klappmyss, hvithval, narhval, ringsel, vanlig uer, pigghå og hvalross som alle regnes som truede arter, og hvor 25 % eller mer av den europeiske bestanden finnes i Norge (såkalte ansvarsarter). I tillegg er det ulike arter av både fisk og sjøpattedyr som er listet som truet i Barentshavet: storskate, polartorsk, blålange, blåhval.
Særlig viktige arter av sjøpattedyr i arktiske områder
Noen arter av sjøpattedyr som kun finnes i Arktis, er ansett som særlig sårbare. Narhvalen, for eksempel, er regnet som ekstremt sensitiv for forstyrrelser og kan reagere på skipsstøy ved store forflytninger selv ved lave lydnivåer (94–105 dB re 1µPa) (Finley et al. 1990) og den har vist unnvikelse og økning av svømmehastighet under eksponering til seismikk (Heide Jørgensen et al. 2021). Ved tre anledninger har et stort antall narhval blitt fanget i isen grunnet sen utvandring fra sommerbeiteområde, sammenfallende i tid med seismikkundersøkelser og på en måte som er svært uvanlig. Heide-Jørgensen et al. (2013) knytter dette opp mot at narhvalen avventet migrasjonen bort fra området, da det ville ført den nærmere seismikkundersøkelsen, og dermed ble fanget da isen kom. Dette kan tyde på at seismikk kan forstyrre migrasjon hos narhvalen.
En annen arktisk art er grønlandshval, en av de største og desidert lengstlevende og saktevoksende dyrene som finnes. Denne ble i tidligere tider fangstet så kraftig at den nesten ble utryddet, og regnes i dag som truet. Den finnes i hele Arktis og deles i fem bestander, hvorav én av disse, Spitsbergen-bestanden, finnes i området mellom Øst-Grønland og østover til Karahavet, med hoved-konsentrasjon rundt Svalbard. Bestanden er svært lav, rundt 100 individer (Bjørge et al. 2010). Som nevnt i avsnitt 3.2.2 er grønlandshval påvist å unnvike seismikk, selv ved lave nivåer eller store avstander, samt endre dykk- og respirasjonsrate (Richardson et al. 1986; Robertson et al. 2013; Robertson et al. 2016). Grønlandshvalen er også en vokaliserende art, særlig i forbindelse med parringen i mars–april, hvor kommunikasjon er helt avgjørende for å lokalisere partnere og gjennomføre en suksessfull parring (Bjørge et al. 2010). Grønlandshvalen er derfor en art som bør anses som svært sårbar for seismikk.
Barentshavet er et viktig beiteområde for mange arter av hval, særlig på sommerstid, da mange bardehvaler kommer hit for å beite på de rike forekomstene av zooplankton og fisk. Det er særlig vågehval, knølhval og finnhval som er de mest tallrike (Bjørge et al. 2010, Øien, 2009). Disse kommer til Barentshavet om sommeren primært for å beite og har ofte migrert over lange distanser for å komme hit. En bardehval på rundt 40 tonn (tilsvarende en liten finnhval) må spise 600–1600 kg daglig for å få dekket energibehovet sitt (Bjørge et al. 2010). Bardehvalene er derfor helt avhengige av å være i områder med høye tettheter av byttedyr for å dekke energibehovet sitt.
Vågehval, finnhval og knølhval kan finnes over hele Barentshavet, men områdene vest for Spitsbergen skiller seg ut som høytetthetsområder hvor alle disse artene oppholder seg og beiter i perioden juni–september/oktober. På høsten er knølhval ofte også observert i store konsentrasjoner øst for Svalbard, hvor de oppholder seg før de starter migrasjon sørover mot parrings-/kalvingsområder langs ekvator. De senere år har også et stort antall knølhval blitt observert i fjorder langs kysten av Troms og Nordland i vinterstid, hvor de antagelig beiter på de store konsentrasjonene av overvintrende sild før de starter migrasjonen sørover.
I takt med økende sjøtemperatur og mindre isdekke har også flere hvalarter ekspandert lengre nord, og både våge-, fin- og blåhval er de senere årene hyppigere observert også nord og vest av Svaldbard (Solvang et al. 2021), og både våge-, fin og knølhval har nå de høyeste tetthetene i arktiske farvann nord for polarfronten om sommer og høsten (Ressler et al. 2015). Dette sammenfaller med en ekspansjon av mange dyreplankton arter, men også med konkurrerende predatorer som torsk, og mangel på korrelasjon mellom bytetetthet og hvalobservasjoner indikerer at tettheten av byttedyra her er for lav for suksessfull beiteaktivitet for de største artene finn-, blå- og knølhval, mens vågehvalen som krever lavere tettheter viser signifikante overlapp med fordelinger av zooplankton og 0-grupe fisk (Solvang et al. 2021). Dette viser at flere hvalarter oppholder seg i områder hvor det ikke er tilstrekkelige tettheter med byttedyr for effektiv beiting.
Det er uvisst hvordan seismikk påvirker beiteaktiviteten hos disse dyrene, men som beskrevet i avsnitt 3.2.2 har ulike arter av hval, blant annet knølhval, i området mellom Bjørnøya og Spitsbergen vist en signifikant reduksjon i beiteaktivitet ved eksponering for militær sonar. Da knølhvalen ikke regnes som en særlig sensitiv art, vil en kunne anta at arter som vågehval, som regnes som mer sensitive for forstyrrelser, kan komme til å reagere likt eller kraftigere. Det er derfor ikke urimelig å anta at både vågehval og finnhval vil kunne bli forstyrret eller avbrutt i beiteaktiviteten ved eksponering for seismikk.
Særlig viktige arter av fisk i arktiske områder
Nordøstatlantisk blåkveite er en selvstendig populasjon og en ansvarsart for Norge, det vil si at mer enn 25 % av populasjonen oppholder seg i norske farvann (Henriksen & Hilmo, 2015). Hoveddelen av voksen blåkveite er fordelt langs eggakanten mellom Fastlands-Norge og Svalbard gjennom hele året, og gytingen foregår også her. Hovedgytingen foregår på vanndybde rundt 500–800 m om høsten og vinteren på eggakanten mellom 70 og 75 °N. Dette er det eneste kjente gyteområdet i Norge for blåkveite og bør derfor tas spesielt hensyn til.
Kloskate er også en slik ansvarsart, og er den hyppigst forekommende arten av skater i Barentshavet og utgjør 96 % av antallet og omtrent 92 % av biomassen av skater fanget på vitenskapelige tokt og som bifangst (ICES 2018). Det er lite konkret kunnskap om viktige oppholdsområder.
Vanlig uer er en annen ansvarsart, som i tillegg er rødlistet som truet i Barentshavet (Artsdatabanken 2021). Arten lever på 100–500 meters dyp på kontinentalsokkelen, langs kysten og i noen fjorder. Utbredelsen går så langt nord som til Spitsbergen, men forekommer sjelden i fiskbare mengder nord for Tromsøflaket/Bjørnøya. Ueren legger ikke egg slik som de fleste andre fisk, men føder levende yngel. Det er ikke yngleområder i Barentshavet.
En art som synes til å være spesielt sensitive til endring i forekomst av sjøis er polartorsk (Huserbråten et al. 2019). Polartorsk bruker lyd under gyting, og endrer bevegelsesmønster som følge av lyd, noe som kan tyde på at lydforstyrrelse kan påvirke gytesuksess (Riera et al. 2018, Ivanova et al. 2020). Gyteområder har nylig blitt kartlagt, og er blitt inkludert i rådgivningskartene fra 2022.
I Barentshavet finnes det fem arter som er listet som truet på den globale rødlisten: storskate, steinkobbe, ål og håkjerring (Artsdatabanken, 2021). Men også for disse har man lite kunnskap om viktige oppholdsområder.
3.3 - Kunnskapsgrunnlag om lydutbredelse og effektsoner
3.3.1 - Biologiske forsøk med effektsoner
Når man forsker på effektsoner, avstander fra undersøkelsen der fisk blir påvirket, er storskalaforsøk med realistiske lydkilder den mest pålitelige metoden å innhente kunnskap. I slike forsøk må det gjøres observasjoner av fisk i et stort område før, under og eventuelt etter eksponering, og metodikken må være tilpasset å bestemme avstand, og helst lydnivå, for fisk som reagerer. Slike forsøk er fåtallig fordi de er svært kostbare å gjennomføre, og det finnes per i dag kun én slik studie som var spesifikt designet for å undersøke nettopp dette. Engås et al. (1996) brukte et 82-liters (5003 in³) luftkanonarray der det ble avfyrt skudd hvert 10. sekund i fem dager på Nordkappbanken i Barentshavet. Fiskeforekomster ble kartlagt ved hjelp av ekkolodd og fangst av torsk og hyse i et område på 40 x 40 nautiske mil (nmi) rundt seismikk-kilden. Resultatene viste stor nedgang i biomasse av fisk i en avstand på 20 nmi fra seismikkfartøyet under og etter eksponering. Fra dette forsøket kan en si at torsk og hyse kan reagere med unnvikelse i avstander ut til 20 nmi. Lydtrykket ble ikke målt, men teoretiske beregninger av lydnivået har vært utført (Handegard et al. 2013). Løkkeborg et al. (2012) gjorde et lignende forsøk i Vesterålen, der de foretok observasjoner av flere fiskearter rundt en ekte seismikkundersøkelse. Det ble også gjort målinger av lyden. I dette forsøket var reaksjonene hos fisken mer variable. En viktig forskjell mellom disse to studiene er at i Engås et al. (1996) var lydkilden innenfor et begrenset område på 3x10 nmi i sentrum av undersøkelsesområdet på 40x40 nmi, og lydeksponeringen varte i fem dager, mens i Løkkeborg et al. (2012) forflyttet lydkilden seg over et større område på 8x46 nmi løpet av 38 dager. Løkkeborg et al. (2012) var ikke designet for å finne noen lydterskel eller avstandsgrense for adferdsresponser, men så mer på fangsttall og mengde fisk generelt i området rundt en ekte seismikkundersøkelse før, under og etter.
Selv om vi ser at reaksjonen vist av Engås et al. (1996) skjer ut til en avstand på 20 nmi, kan det diskuteres om dette vil gjelde for alle områder. Det er derfor ønskelig å bruke modeller som kan finne ut ved hvilke avstander fra kilden lydnivået tilsvarer nivået som fisken opplevde ved 20 nmi fra kilden på Nordkappbanken. I Engås et al. (1996) ble det brukt en veldig stor lydkilde på 5000 kubikktommer. Denne er kraftigere enn de fleste av dagens kilder, men disse avfyres gjerne oftere. En mindre lydkilde som blir avfyrt oftere kan ha samme energi som en stor lydkilde i området nær kilden, men siden lyden er svakere vil den ikke bre seg like langt. Vi anser derfor at det er viktig å oppgi total lydenergi i en 10-sekundsperiode for å best sammenlikne mellom seismikk-kilder med ulike størrelse og tid mellom avfyringer.
3.3.2 - Modellering av lydforplanting
For å kunne beregne lydnivået ved en gitt avstand fra kilden er det viktig å ha god nok kunnskap om parameterne som må inngå i en slik modell (Farcas et al. 2016), som lydkilde, vanndyp, bunntopografi, egenskaper i bunn og vannkolonne (tetthet, lydhastighet og demping). Ulike modeller kan gi ulike resultater, og det er ikke alltid at modellen stemmer med aktuelle målinger. Ainslie et al. (2019) sammenlignet flere modelltyper og konkluderte med at de testede modellene hadde ganske godt samsvar (innenfor 3 dB) opp til 100 Hz, mens forskjellene økte for høyere frekvenser. Det som hadde størst betydning for lyden i fjernfeltet, var hvordan kildesignaturen, det vil si beskrivelsen av lyden ut fra kilden, var implementert i modellen. Denne må gjerne simuleres først i en separat modell, for så å brukes som input i en lydpropagasjonsmodell (Ainslie et al. 2016). Modeler for å beregne partikkelbevegelsen har også blitt sett på, og de viser at det er dårligere samsvar mellom ulike modeller for å beregne partikkelakselerasjon enn det er for lydtrykket. Dette er som forventet siden partikkelakselerasjonen modelleres ved å først modellere lydtrykket og deretter finne romlig gradient av dette. Det er størst usikkerhet rundt skarpe maksimum og minimum (Prior et al. 2019)
Direktiviteten til kilden vil spille en rolle for lydutbredelsen. De fleste luftkanonarray gir maksimal lyd nedover og mindre lyd horisontalt (Caldwell & Dragoset 2000). Kildens utstrålingsmønster er også forskjellig på langs og på tvers av linjeretningen (Heaney & Campbell 2019, Westerdal 2017). Dette kan påvirke resultatet og gi en lengre sikkerhetssone på tvers enn på langs i forhold til retningen på luftkanonarrayen. Lyden vil dessuten variere med dyp, så en må vurdere hvilket dyp man skal forholde seg til, eller om man skal bruke maksimalt lydtrykk uavhengig av dyp.
For å øke sjansene for at modellen gir et riktig resultat, må først modellen verifiseres ved å teste den mot et forenklet og kjent tilfelle. Det kan gjøres ved å sammenligne med en såkalt “benchmark model” eller referansemodell (Chin-bing et al. 1993). Deretter må modellen kalibreres for det aktuelle området og for lydkilden som skal modelleres. Kalibreringen gjøres ved å sammenligne modellen med målinger for aktuell lydkilde og område. Ved bruk av modeller og lydgrenser vil det bli nødvendig å måle lydnivået ved ulike avstander fra kilden og ulike dyp. Optimalt sett med hattmålinger både av lydtrykk og partikkelakselerasjonen i tre dimensjoner, men det er vanskelig i praksis. Noen få målinger av lydtrykk i ulike posisjoner er derimot realistisk. Det finnes gode modeller for lydpropagasjon, det som mangler er s kunnskap om parametere og kalibrering.
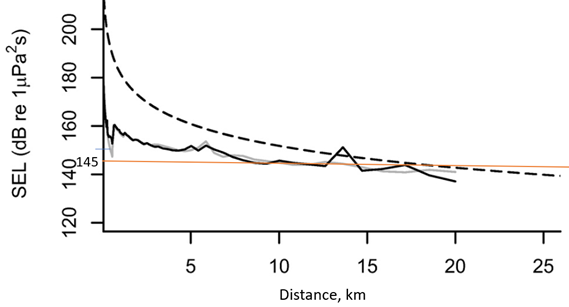
Vi mangler god kunnskap om hvilke lydtrykksnivåer som utløser fluktresponser hos fisk, men kan i påvente av dette bruke kunnskap om hvilke lydtrykksnivå som ikke utløser slike responser som terskler på sikkerhetssone, dvs en sone hvor det er usannsynlig at negativ påvirkning inntreffer. McQueen et al. (2022) viser at torsk som ble eksponert for lyd opptil 145 dB re 1 uPa2s ikke svømte vekk fra gyteområdet. Simuleringer av lydnivået fra kilden i Engås et al. (Handegard et al., 2013) viser at dette kan tilsvare en avstand mellom ca 8 – 18 km fra 5000 in3 kilden (Figur 2). For mindre kilder og et annet område/miljø vil trolig avstanden bli noe annet.. Dette eksempelet illustrerer at flere km fra kilden endrer ikke lyden seg så raskt med avstand lenger. En kan få store avstander som har omtrent samme lyd og det kan være utfordrende når en ønsker å vite ved hvilken avstand lyden er under ett gitt nivå.
3.4 - Konsekvensvurderinger av effekter av seismikk på marine organismer
Som vist i kunnskapsgrunnlaget i kapitlene ovenfor, kan både fisk og sjøpattedyr påvirkes av seismikk, enten ved direkte skade av hørsel eller endret adferd. Hvor alvorlig påvirkningen er avhenger av hvor stor andel av en bestand eller populasjon som eksponeres, hvor lenge forstyrrelsen varer, og hvilken type adferd som forstyrres. Enkelte biologisk viktige aktiviteter finner sted kun spesifikke tider på året, og på spesifikke steder, som for eksempel gyting hos fisk og beiting hos bardehval. Dersom disse aktivitetene forstyrres, er konsekvensen for en negativ effekt på populasjonen størst. I dette kapittelet er derfor mulige konsekvenser på slike biologisk signifikante livsfunksjoner vurdert for fisk og sjøpattedyr.
Konsekvensutredningen er gjort med bakgrunn i hvilke situasjoner forstyrrelsen og skade fra seismikk potensielt kan medføre negativ påvirkning på bestands- eller populasjonsnivå.
3.4.1 - Fisk
Biologisk viktige livsfunksjoner for fisk er beiting og reproduksjon (gyting). Konsekvenser for å forstyrre disse er gjennomgått i avsnittene under.
Konsekvenser for hørselskade
Basert på resultatene for hørselsskade på voksen fisk (se kap. 3.2.1) tyder dette på at fisk må være svært nær (<10 m) en luftkanon for å bli skadet. Frittsvømmende fisk antas også å svømme bort fra lydkilden før den kommer så nær at slik skade inntreffer. Eventuell skade vil derfor sannsynligvis hovedsakelig forekomme på fisk som befinner seg i umiddelbar nærhet til luftkanoen ved oppstart, da disse ikke har blitt «advart» og svømt bort før lydtrykket overstiger terskel for skade. Dette vil imidlertid gjelde et så lite antall fisk at en effekt på bestandsnivå vil være svært lite sannsynlig. Ramp up er et virkemiddel for å redusere slik skade ved oppstart, men svært lite kunnskap foreligger om dette for fisk. Faktisk viste Neo et al. (2016) at dette førte til mindre unnvikelse, riktignok for fisk i fangenskap. Fisk kan også regenerere skade på hørselsceller over tid (Smith & Monroe, 2016).
På bakgrunn av dette, har ikke Havforskningsinstituttet noen spesifikke råd for å begrense mulige fysiske skader på fisk ifm seismikk.
Konsekvenser for gyting og gytevandring
Suksessfull reproduksjon er helt essensielt for å opprettholde en bestand. For svært mange fiskearter foregår gyting ved at store deler av en bestand samles i store aggregeringer i samme område; slik at fisketettheten i dette området er mye større enn i de fleste andre perioder på året. En forstyrrelse i en slik aggregering vil derfor påvirke en større andel av bestanden enn utenom gytesesongen/gyteområdet.
Kunnskapsgrunnlaget som er gjennomgått i kap. 3.2 viser at i noen tilfeller under og etter seismikk-eksponering flytter fisk seg bort fra området, i andre tilfeller ikke. Torsk, f eks., forflyttet seg bort over store avstander på Nordkappbanken (Engås et al. 1996), mens en ikke så noen slik forflytning i en vindturbinpark i Nordsjøen (van der Knapp et al. 2021). Slike forskjeller kan muligens forklares ved hvor stedbunden fisken er, samt hvor stor konsekvensen er ved å forlate området.
Dersom fisk på gytefelt reagerer slik som beskrevet av Engås et al. (1996), kan en forvente at torskefisk forflytter seg bort fra området den oppholder seg i, og at denne forflytningen kan være i størrelsesorden 20 nmi (37 km) fra kilden. En slik forflytning bort fra gyteområdet kan føre til nedsatt gyting og økt yngeldødelighet grunnet viktigheten av gyting i tid og rom; eggene gytes på et tidspunkt og i et område der de kan drive med havstrømmene og klekkes i et område med rik tilgang på mat. Gytingen foregår derfor på en tid slik at klekking av egg sammenfaller med for eksempel våroppblomstring. Når fisken er på selve gytefeltet eller er på vandring mot dette, kan derfor forflytninger i størrelsesorden 20 nmi fra kilden få betydning for selve gytingen eller for fordelingen av egg på gytefeltet og dermed havner i et område som har endret strømningsmønstrer som kan fører til forandret overlevelse av larver/yngle. Eventuelle påførte forflytninger vil sannsynligvis også ha større betydning når gytebestanden er lav og gytefeltet er begrenset til et mindre, men optimalt område, enn når gytebestanden er høy eller gyteområdet stort. Den direkte effekten av en forstyrret eller forflyttet gyting vil i praksis ikke være mulig å måle på larve-nivå ettersom en ikke har noe direkte sammenligningsgrunnlag, og fordi de naturlige variasjonene i tallrikhet fra år til år kan være store. De påviste kraftige unnvikelsene er undersøkt for hovedsakelig torsk og hyse, men det vil være rimelig å anta at lignende reaksjoner kan inntreffe hos andre torskefisk som for eksempel sei, øyepål, hvitting og lysing. Arter som er avhengige av et helt spesielt bunnsubstrat for å gyte, som sild og lodde, eller som har en helt spesifikk bunntilknytning, som tobis, anses også som særlig sårbare, da en mulig forflytning på 20 nmi kan føre dem bort fra dette spesifikke bunnhabitatet og dermed forhindre gyting.
Under fiskens vandring til gyteområdene kan også forstyrrelser fra kraftig lyd potensielt gi avvik i det normale vandringsmønsteret, som igjen kan gi opphav til betydelige forstyrrelser i vandringsrutene. For de artene som har svært konsentrerte gytevandringer, det vil si der store deler av bestanden passerer små og smale områder like før de rekker gytefeltet, bør disse rutene skjermes for seismikk. Det gjelder hovedsakelig nordøstarktisk torsk (NEA-torsk), norsk vårgytende sild (NVG-sild) og lodde. Gytevandringen til lodde er med dagens kunnskap uforutsigbar og varierer mye fra år til år. Vandringsruten til lodde er vanskelig å kartfeste da den er svært dynamisk og varierer fra år til år. Siste delen av vandringsruten til lodde, når de nærmer seg gytefeltet, er mer konsekvent og har blitt beskrevet basert på Fall og Bogstad, (2019).
Havforskningsinstituttet fraråder på bakgrunn av dette seismikk i de konsentrerte delene av gyteområdene og gytevandringene for disse artene, samt i en buffersone rundt disse på 20 nmi. For borestedsundersøkelser som bruker mindre luftkanonoppsett er buffersonen satt til 5 nmi basert på lydtrykkmålinger fra typiske 3D kanonoppsett og BSU-liknende kanonoppsett presentert av Malme et al. (1986) og Pearson et al. (1987) i forhold til høre- og responsterskler (Popper et al. 2014) for bunnfisk. Det estimert at en ved en typisk borestedsundersøkelse kunne utløse skremmeatferd på ca. en fjerdedel av avstanden som ved en 3D-undersøkelse (Dalen, 2008). Disse grensene er basert på få undersøkelser, og det blir ikke tatt hensyn til at ulike luftkanonoppsett kan gi støy med ulik rekkevidde, og at lydpropagasjon varierer mellom områder, dyp og bunntype. En fullskala kilde i dag er ikke lenger den samme som i 1996. Nyere forskning på gytende torsk viser at i et gyteområde på kysten forflyttet torsken seg ikke vekk fra gyteområdet under eksponering til en nedskalert seismikk-kilde (McQueen et al. 2022). En kan derfor anta at nivåer lik eller lavere enn det torsken i dette studiet ble eksponert for (145 dB re 1 uPa2s SEL integrert over 10 sekund) lite sannsynlig medfører storskala forflytning.
Derfor anses det som akseptabelt at gyteområder kan eksponeres for lydnivåer opp til SEL≤145 dB re 1 uPa2s (integrert over 10 sekund). Som beskrevet over, og i kap 3.3, vil avstanden fra seismikk-kilden til hvor dette nivået inntreffer være høyst variabelt, og må derfor modelleres for den spesifikke kilden og område den skal brukes.
Havforskningsinstituttet vil derfor åpne opp for en mulighet til å unngå de lite fleksible og statiske grensene med 20 og 5 nmi som buffersone rundt gytefelt, og åpner opp for et alternativ hvor forutsetningen er at gytefelt ikke blir eksponert for lydnivåer som overstiger 145 dB re 1 uPa2s (SEL integrert over 10 sekund).
Havforskningsinstituttet foreslår derfor en prøveordning i 2023 hvor det kan åpnes for å få unntak fra regelen om 20 nmi buffersone rundt gytefeltene, angitt som frarådingsområder.
For å få et slik unntak må operatøren gå i dialog med Havforskningsinstituttet, og unntaket kan gis basert på akustisk modellering av lydforplantning og verifikasjonsmålinger utført av eller på vegne av operatøren. Lydeksponeringsnivået integrert over 10 sekunder i frarådingsområdet må dokumenteres før undersøkelsen ved hjelp av modellering til å være under 145dB re 1µPa2s. Den foreslåtte framgangsmåten består av modellering av lydforplantning fra den seismiske kilden som skal benyttes i undersøkelsene, verifisert med målinger under de seismiske undersøkelsene.
Følgende steg forventes:
- Det benyttes industristandard verktøy for å modellere kildesignatur (for eksempel Gundalf eller Nucleus). Kildesignaturen må kunne gjøres tilgjengelig for HI
- Verktøyet for modellering av lydforplantning må være dokumentert og egnet for å simulere lydforplantning av seismiske pulser og estimere SEL_10sek under vann. Offentlig tilgjengelige verktøy er å foretrekke.
- Modelleringen av lydforplantningen må være transparent slik at HI kan gjennomgå de ulike stegene og reprodusere resultatene.
- Innputdata til simuleringene må være relevante og dokumenterte, dette inkluderer lydhastighetsprofiler, bunndyp og sedimentsammensetning/elastoakustiske egenskapene til havbunnen.
- Resultatene må vise at maksimum vertikal SEL_10sek (dB re 1µPa2s) gjennom vannsøylen er mindre eller lik 145 dB re 1µPa2s i selve gyteområdet, uavhengig av hvor i den planlagte undersøkelsen kilden befinner seg for å kunne få et unntak. Dataene og beregningene må kunne gjøres tilgjengelig for HI. SEL_10sek må beregnes over frekvensbåndet ca. 1 Hz-1 kHz.
- Målinger av lydtrykk må utføres under det seismiske surveyet ved bruk av kalibrert(e) hydrofon(er) og dokumenteres. Beregning av SEL_10sek og sammenligning med resultater fra modellering må evalueres og beskrives. Data, kalibrering og beregninger må kunne gjøres tilgjengelig for HI.
Det er fremdeles viktig å få bedre klarhet i ved hvilken egenskap(er) av lyden og for hvilke terskler/varighet ved denne egenskapen(e) fisk begynner å reagere på. Videre hvilke reaksjoner som kan resultere i negative effekter for bestanden, og hvordan dette varierer mellom ulike arter. Å sette kriterier for lyd-eksponering av fisk er et komplisert tema som fortsatt har mange kunnskapshull, men det har vært mer fokus på miljøkonsekvenser fra lydpåvirkning av fisk internasjonalt de siste årene som førte til mer forskning på dette teamet (Hawkins et al. 2020).
Som nevnt ovenfor er lydgrensen på 145 dB re 1µPa2s basert på undersøkelser hvor det ikke ble observert at fisken forlot gyteområdet som følge av seismikkeksponering. Det er derfor uvisst ved hvilket lydnivå fisken faktisk starter å reagere. For å få bedre forståelse av dette, burde det gjennomføres følgeforskning, dvs. forskning ved å overvåke fisk før og under en reell seismikkundersøkelse, i et område i nærheten av et kjent gyteområde som vil medføre høyere lydtrykk enn denne grensen på et gytefelt. Selskaper som kan være interessert i å inngå samarbeid om en slik følgeforskning, bes kontakte Havforskningsinstituttet for nærmere dialog.
Konsekvenser for egg, larver og dyreplankton
Det er utført en verste-tilfelle-beregning av andelen larver i en bestand som kunne bli drept under en typisk seismisk 3D-undersøkelse som dekker et areal på 500 km². Det ble funnet ved et «worst case scenario» vil maksimalt 0,45 % av larvebestanden kunne bli drept av en undersøkelse. En har også beregnet en forventningsverdi som mer representerer det en i gjennomsnitt ville forvente ble drept i bestanden. Denne andelen er beregnet til om lag 0,3 ‰ av bestanden for hver 3D-undersøkelse. De daglige dødelighetsratene i løpet av undersøkelsen er beregnet til 0,18 ‰ per døgn i verste tilfelle og 0,012 ‰ per døgn i gjennomsnitt. Sett i relasjon til daglige naturlige dødelighetsrater på egg- og larvestadiet – 5–15 % per døgn, og senere 1–3 % per døgn fram til 0-gruppestadiet – er dette så lave dødelighetsrater at de kan regnes som ubetydelige i bestandssammenheng. (Sætre og Ona, 1996)
Havforskningsinstituttet fraråder derfor ikke seismikkaktivitet i områder med høye konsentrasjoner av egg og larver.
For dyreplankton anser vi inntil flere resultater foreligger at dødelighet kun inntreffer nærmere enn 5 m fra luftkanonen som mest relevante for norske forhold. Havforskningsinstituttet fraråder derfor ikke seismikkaktivitet i områder med høye konsentrasjoner av zooplankton.
Konsekvenser for beiting
I beiteperioden er de fleste fiskebestander fordelt over et mye større område enn under gyting, og en forstyrrelse vil derfor sannsynligvis ramme en mindre andel av bestanden.
Selve beiteadferden kan også forstyrres når fisk utsettes for seismikk (Løkkeborg et al. 2012) eller annen menneskeskapt støy (Purser & Radford 2011; Voellmy et al. 2014; Sabet et al. 2016). Dette medfører at energiinntaket reduseres, samtidig som energibruken gjerne øker som følge av økt svømmeaktivitet. Det er derimot usikkert hvor stor potensiell effekt dette kan ha på bestandsnivå, ettersom beitingen for, i motsetning til gyting, ofte foregår over svært store områder. En forflytning av beitende fisk på 20 nmi vil ikke nødvendigvis føre til nedsatt matinntak over lang tid eller for en så stor andel av bestanden at det vil gi konsekvenser på bestandsnivå. Vi kan anta at beitende fisk har større muligheter til å forflytte seg uten å miste beitemuligheter, derfor antas lokale forstyrrelser å ha mindre effekt. Store endringer i bakgrunnsstøy i store områder skal imidlertid unngås.
Havforskningsinstituttet fraråder derfor ikke seismikkaktivitet i beiteområder for fisk, men anbefaler at seismiske undersøkelser ikke skjer veldig nær hverandre i tid og rom, for å sikre at det finnes stillere områder å forflytte seg til for fiskene.
3.4.2 - Sjøpattedyr
Konsekvenser ved hørselsskade
Ut ifra de oppgitte terskelverdiene for permanent hørselsskade (NOAA 2015) vil skade som følge av enkeltpulser fra seismikk være begrenset til et område mellom 50 og 100 m dersom en antar et kildenivå på 220–240 dB re 1μPa·m og sfærisk propagasjonstap (20·logR)
Niser vil kunne risikere hørselstap dersom de utsettes for spisstrykk som overstiger 202 dB re 1µPa (Tabell 1), noe som vil inntreffe 450 m fra en seismikk-kilde med kildenivå 255 dB (re µPa m) eller akkumulert lyd (vektet SEL, 155 dB re 1µPa2 s) så nært som 30 m fra kilden. For en bardehval (lavfrevente hvaler, Tabell 1) vil spisstrykk kriteriet (219 dB re 1µPa) oppnås ved en avstand på ca 65m, mens SEL kriteriet (183 dB re 1µPa2 s) vil oppnås dersom dyret er nærmere enn 125m (for detaljer, se Kvadsheim et al. 2020).
Ved samme kriterier vil midlertidig terskelskifte (Tabell 1) kunne oppstå noen få hundre meter fra kilden, og dermed kun ramme dyr som befinner seg svært nær luftkanonene. Midlertidig terskelskifte er ofte brukt som et konservativt risikokriterium ved at det anbefales at dyrene ikke utsettes for nivåer som overstiger disse.
Risikoen for skade kan reduseres ved avbøtende tiltak som ramp-up eller soft-start, noe som innebærer en gradvis økning av lydnivået og gir dyr i nærheten av lydkilden mulighet for å svømme unna før nivået er høyt nok til å indusere skade (det forventes at sjøpattedyr unnviker lydkilder som nærmer seg nivåer for midlertidig terskelskifte). Modellstudier har vist at ramp-up reduserer risiko, men effekten avhenger av faktorer som dyrets responsterskel, lydkildens og dyrets hastighet (von Benda-Beckmann et al. 2014). Effekten av ramp-up har også vært undersøkt eksperimentalt på knølhval for både sonar (Wensveen et al. 2017) og seismikk (Dunlop et al. 2016). Disse studiene konkluderer med at ramp-up resulterte i lavere SEL, men at risikoreduksjonen var begrenset. Imidlertid har knølhval forholdsvis høy responsterskel sammenlignet med andre arter (Sivle at al. 2015), så dette er derfor ikke helt uventet. Dersom man skilte ut den mest responsive delen av bestanden, nemlig hunndyr med kalv, hadde ramp-up god effekt (Wensveen et al. 2017). Det er dermed forventet at ramp up vil ha en betydelig risikoreduserende effekt, men at denne varierer mellom ulike arter.
Ramp up kan derfor virke som en relativt effektiv måte å redusere skade hos sjøpattedyr, da særlig sensitive arter, og Havforskningsinstituttet tilråder derfor bruk av ramp up som avbøtende tiltak for å skjerme sjøpattedyr mot hørselskade.
Konsekvenser av endret vokalisering
Bardehvaler bruker lavfrekvent lyd til å kommunisere over store avstander. Seismiske pulser inneholder mye energi i det samme frekvensbåndet, og seismikk vil derfor kunne maskere slik kommunikasjon. Som beskrevet i kapittel 2.2.2 kan bardehvaler som finnhval, blåhval og grønlandshval endre kommunikasjonen sin enten ved å øke styrken eller lengden eller ved å endre frekvensen. Slike endringer kan ha økte metabolske kostnader, og Holt et al. (2015) viste at disse kostnadene var betydelige for delfiner ved endret vokalisering i form av økt lengde eller nivå. Alvorlighetsgraden av slike økte kostnader avhenger selvsagt av omfanget av den økte vokaliseringen, særlig dersom den sammenfaller med andre responser som kan øke den metabolske kostnaden ytterligere. Slike responser kan være økt overflateaktivitet eller dykkekostnader, eller responser som minker energinivået, som nedsatt beiteaktivitet. Det er derfor viktig å se på den kumulative effekten av den totale responsen for å vurdere forstyrrelsens alvorlighetsgrad (Lusseau & Bejder 2007). Dette har vi i dag for lite kunnskap om til å bruke som basis for råd. Grønnlandshvalen er en svært vokal hvalart, og Ahonen et al. (2017) rapporterer om deteksjon av syngende grønlandshval i Framstredet daglig mellom oktober og april. Denne populasjonen er sterkt utrydningstruet, men er beskyttet fra menneskelig aktivitet store deler av året grunnet isdekket gjør området ufremkommelig for skipsfart. Imidlertid er det forventet at isdekket gradvis vil minke grunnet global oppvarming, og at disse områdene blir mer attraktive for både oljeleting, turisme og annen skipsfart, noe som kan forstyrre den viktige vokaliseringen til grønlandshvalen (Ahonen et al. 2017).
Basert på overnevnte argumenter anbefaler Havforskningsinstituttet at dersom det gjennomføres seismikkundersøkelser i områder hvor grønlandshval kan påtreffes, bør det brukes dedikerte observatører slik at en unngår å skyte seismikk så lenge arten er i nærheten.
Konsekvenser av beiteforstyrrelser
Bardehvaler er såkalt «capital breeder», hvilket betyr at de bruker lagret energi for repoduksjon og overlevelse. Dette gjør dem i stand til å vandre lange distanser mellom reproduksjon og fødsel i tropiske strøk, og beiting i arktiske farvann. Særlig hunndyr er avhengig av enorme energilagre for å være istand å være gravid, samt å produsere melk til kalven som dier i 6-12 mnd etter fødselen, og i tillegg i denne perioden vandrer store avstander sammen med kalven til beiteområdet. Dette ekstreme energibehovet (faste under vandring og med diende kalv), medfører at kun store hunner har kapasitet til å føde og oppfostre kalver. En nedgang i næringstilgang kan redusere sannsynligheten for suksessfull reproduksjon (e.g. Lockyer, 1981). Den høye kostnaden med å die for bardehvaler medfører at lavere energireserver vil føre til mindre kalver med dårligere kondisjon (Christiansen et al. 2014). Finnhval, knølhval og vågehval er de mest tallrike artene av bardehval i norske farvann, og finnes hovedsakelig i Barentshavet om sommeren og høsten. Mesteparten av tiden disse tilbringer her, brukes til å beite eller vandre mellom klynger av byttedyr (Lydersen et al. 2020).
Det er økende mengder kunnskap som tilsier at de kraftigste responsene hos bardehval til lyd inntreffer når de beiter (eks. Goldbogen et al. 2013; Sivle et al. 2016; Kvadsheim et al. 2017; Southall et al. 2019; Boisseau et al. 2021).
Som beskrevet i avsnitt 3.2.2 er bare ett studium på seismikk på sjøpattedyr i våre farvann. Et svært omfatende studie i Australia har imidlertid funnet at effekten av seismikk på vandrende knølhval er liten, og at noen effekt på populasjonsnivå er høyst usannsynlig. Imidlertid bør en ta høyde for at a) studiet ble gjort på vandrende, ikke beitende knølhval som er i norske farvann, b) populasjonsmodellen antok forstyrrelser for bare en seismikkundersøkelse, c) knølhval er en art som er klassifisert som «lite responsiv» for lydforstyrrelser sammenliknet med f.eks vågehval og nebbhval.
Det finnes få studier av hvordan seismikk påvirker beiteaktiviteten hos sjøpattedyr, men for militære sonarer er det dokumentert avbrutt eller nedsatt beiteaktivitet for mange arter i norske farvann – både knølhval (Sivle et al. 2016), blåhval (Goldbogen et al. 2013; Friedlander et al. 2016), nebbhval (Miller et al. 2015), spermhval (Isojunno et al. 2016) og spekkhogger (Miller et al. 2012). Individer i beitemodus har høy sannsynlighet for å reagere, og som oftest gjør de det ved å avbryte beiteaktiviteten og svømme unna kilden (Goldbogen et al. 2013; Harris et al. 2015).
I mangel på kunnskap om hvordan seismikk påvirker beiteadferd hos sjøpattedyr, kan man anta at seismikk medfører de samme reaksjoner som vist for sonar. Særlig for arter som kun beiter intensivt i en sesong og er avhengige av tette konsentrasjoner av byttedyr kan slike reaksjoner være alvorlige. Om forstyrrelsen inntreffer i denne perioden, kan selv korte forstyrrelser føre til signifikant tap i energiinntaket (Goldbogen et al. 2013). Farmer et al. (2018) viser at avbrutt beiting over tid, for eksempel i et område med mye menneskelig aktivitet som seismikk og oljeutvinning, på sikt vil føre til reduserte fettreserver, noe som kan være kritisk ettersom disse brukes når pågående beiting opphører. Dersom dette skjer over tid, vil reservenene etter hvert bli stadig mindre, noe som resulterer i nedsatt evne til vellykket reproduksjon. Særlig er hunner som skal produsere melk, og kalver med lave fettreserver er utsatt. På sikt vil slike reduksjoner også kunne gi konsekvenser på populasjonsnivå da det går utover evnen til reproduksjon og reduserer sannsynligheten for at kalvene lever opp.
Adferdsstudier med sonar viser tydelige forskjeller mellom ulike arter. Noen skiller seg ut som særlig sensitive, som nebbhval, nise, vågehval og til dels spekkhogger (Harris et al. 2015; Sivle et al. 2015; Kvadsheim et al., 2017, Wensveen et al. 2019), mens arter som knølhval, spermhval og grindhval er mindre sensitive. Både nebbhval og vågehval er blant hvalfangere kjent for å være arter som er særdeles sky og vanskelige å tilnærme seg. Det er mye som tyder på at disse artene er særlig unnvikende og lett påvirkes av menneskeskapt lyd. Studer av nebbhval i områdene rundt Jan Mayen viser at disse reagerer særdeles kraftig på pulset lyd fra militær sonar ved å forlate området i opptil flere dager, avbryte all beiteaktivitet og anormale dykk som potensielt kan medføre fare for dykkersyke (Miller et al. 2015; Sivle et al. 2015). Derfor regnes nebbhvalen som særlig sårbar, og viktige beiteområder bør beskyttes for seismikkaktivitet, Imidlertid er konsentrerte beiteområder for nebbhval ikke godt nok kartlagt, og dagens kart over beiteområder viser svært store områder, og er derfor ikke inkludert i rådgivningskartene. Imidlertid pågår et arbeid med å bedre kartlegge disse, og vil derfor med stor sannsynlighet bli inkludert ved neste års gjennomgang, dvs i rådgivningskart for 2014.
Vågehval er en av de vanligste artene av sjøpattedyr i Norge, og har vist særdeles kraftige adferdsresponser til både sonar (1-3 kHz) (Sivle et al. 2015; Kvadsheim et al. 2017) og ADD (15 kHz) (Boisseau et al. 2021) i form av å svømme i rett linje bort fra lydkilden i høy hastighet, samt å endre dykkmønster. Boisseau et al. (2021) viser at en 15 minutters forstyrrelse kan forstyrre beiteadferd i opptil 45 minutter, og medføre et tap på rundt 5% av daglig matinntak. Dersom forstyrrelsen og responsen er langvarig/hyppig, vil dette over tid kunne medføre tapte energireserver som trenger til vandring og oppfostring av kalver.
Når en skal vurdere konsekvensene av en påvirkning, er det som regel effekten på en populasjon som er fokus. Såkalte PCoD (Population Consequenses of Disturbance) modeller brukes ofte til å predikere mulige slike effekter. For captital breeders, som de fleste bardehvalene er, viser slike modeller at redusert tid til å beite grunnet en forstyrrelse kan påvirke individets energibalanse og videre reproduksjon og overlevelse (Keen et al. 2021). Imidlertid varierer hvor stor denne effekten er veldig med tilgangen til mat, og hyppighet av forstyrrelser. For eksempel Pirotta et al. (2022) har modellert hvordan observert avbrutt beiting under eksponering til militær sonar hos blåhval i California påvirker overlevelse og reproduktiv suksess. Resultatene viste at selv om hvalene responderer til sonarlyd ved å avbryte beiteaktivitet, gav dette ikke konsekvenser for verken overlevelse eller reproduktiv suksess med et nivå av gjennomsnittlig 7.6 eksponeringer til sonar hvert år. Videre viser modellen at dersom mattilgangen er dårligere, vil negative effekter på reproduktiv suksess inntre ved lavere antall eksponeringer. Dette viser at bardehvaler kan tolerere en mengde støy uten at populasjonen påvirkes, men at dette er sårbart for svingninger i mattilgang. I år med dårlig mattilgang vil derfor toleransegrensen være lavere. Det er derfor viktig både å planlegge å legge lydforstyrrelse til perioder hvor beiteintensiteten er lavest, samt å følge med på mattilgang (eks overvåke biomasse av plankton, fisk etc) for å kunne begrense aktiviteten (støyen) dersom tilgangen på mat minker (Keen et al. 2021). Heller ikke agent-based modells (ABM) predikerte noen negativ påvirkning på populasjonsstørrelse hos niser eksponert for seismikk, men at når på sesongen forstyrrelsen finner sted er avgjørende; på våren når energilagrene er gode er sannsynligheten for en negativ effekt 7 ganger lavere enn på høsten når dyrene trenger å fylle opp lagrene for vinteren (Gallagher et al. 2021).
Basert på dette, fraråder Havforskningsinstituttet seismikkaktivitet i områder og perioder hvor det er høye tettheter av beitende bardehval.
3.4.3 - Konsekvensvurdering for nordområdene
Det er per i dag ikke åpnet for å gjøre ordinære seismikkundersøkelser i områdene nord for Bjørnøya. Derimot foregår det fra tid til annen undersøkelser i ikke-åpnede områder i regi av Oljedirektoratet eller i forskningsøyemed. I slike områder kan det forekomme arter som grønlandshval og narhval, som kan være særlig sårbare.
Grønlandshval er kjent for å være sensitiv til seismikk (Richardson et al. 1986; Robertson et al. 2013) og har rødlistestatus som «sterkt truet». Undersøkelser i områder nord for åpnede blokker bør unngås i områder med høy tetthet av grønlandshval og særlig i parringstiden (mars–april). Men siden arten er sjelden er høy tetthet vanskelig å predikere og kartfeste. Fraråding i hele området hvor de kan påtreffes året rundt vil hindre f.eks. viktige forskningstokt. Derfor anbefaler Havforskningsinstituttet at det brukes en dedikert observatør til å holde utkikk etter grønlandshval i denne artens oppholdsområder, og sikre at det ikke befinner seg grønlandshval i en sone i nærheten til seismikkfartøyet. Anbefalinger av hvor stor sonen rundt fartøyet som overvåkes burde være, varierer mellom landene som praktiserer dette (Weir & Dolman, 2007). I Storbritannia, Canada og Mexicogulfen brukes 500 m rundt luftkanonen som sikkerhetssone. Australia har 3000 m, mens for New Zealand varierer den fra 200 til 1500 m avhengig av art. I Russland er det en sikkerhetssone på 250 m for seler og 1000 m for hval. Bakgrunnen for disse sonene er som regel å unngå å utsette sjøpattedyr for lyd over en gitt terskel som kan gi hørselskade. F.eks. i Canada, er det argumentert at adferdsmessige og skadeeffekter kan inntreffe ved lydnivåer over henholdsvis 160 og 180 dB re 1 µPa. Ved å beregne lydtap kan man beregne ved hvilke avstander disse nivåene inntreffer. Imidlertid er jo dette svært varierende med bunntype, dyp, oseanografiske forhold og ikke minst kildestørrelse (se også kap. 3.3). I tillegg er det variasjon mellom arter ved hvilke nivåer både skade og adferdsreaksjoner inntreffer. Imidlertid må sonene være slik at det er praktisk gjennomførbart; mange ulike soner for ulike arter kan bli vanskelig for en person å ha kontroll på, og avstanden må relativt lett kunne bedømmes, samt ikke større enn at dyret relativt lett kan observeres i de fleste værforhold. For en veldig stor kilde (8600 in3) viser generelle beregninger at et nivå på 180 dB inntreffer ca 950 m fra kilden, mens en liten kilde på 250 in3 vil gi en sone på 50 m (Environment Australia, 2001). For grønlandshval, som dette er aktuelt for i Norge, er det ønskelig å forhindre både skade og endret adferd. På bakgrunn av dette anbefaler Havforskningsinstituttet å bruke en sone på 1000 m rundt luftkanonen. Dersom det observeres en grønlandshval nærmere enn dette, bør seismikken stanse, avventes i 30 min før den kan starte igjen med ramp up dersom hvalen ikke er i denne sonen. Områdene hvor det anbefales å bruke en slik dedikert observatør er kartfestet og vist i Vedlegg I.
I Nordområde er det også noen kjente områder med høye konsentrasjoner av beitende bardehval, hovedsakelig vågehval, knølhval og finnhval, som beskrevet i avsnitt 3.2.3. Havforskningsinstituttet gjennomfører årlige telletokt for hval i ulike områder, og områdene vest av Spitsbergen er vist å ha høye tettheter av vågehval, knølhval og finnhval, som beiter der i perioden juni–september/oktober. Av vågehval finner en særlig mange drektige hunner i dette området (pers. komm. Nils Øien, Havforskningsinstituttet). Knølhvalen beiter også i høye konsentrasjoner rundt Bjørnøya i denne perioden, samt øst av Spitsbergen på sensommeren og høsten (august–oktober), før den migrerer sørover.
Områdene som er beskrevet her, regnes som særdeles viktige beiteområder for bardehval, og aktivitet med seismikk frarådes i disse områdene og periodene. Dette er kartfestet og vist i Vedlegg I.
3.5 - Råd for seismikkundersøkelser i norske farvann
Basert på eksisterende kunnskapsgrunnlag og vurdering av konsekvenser for økosystemer og populasjoner gir Havforskningsinstituttet følgende råd for undersøkelser ved bruk av seismikk:
- Havforskningsinstituttet fraråder seismikkundersøkelser i viktige gytefelt for enkelte fiskebestander i gyteperioden, samt konsentrerte gytevandringsruter i perioder for slik innvandring. Hvilke bestander som inkluderes baseres på ulike kriterier viktigheten av bestanden og for risiko for at påvirkningen fører til populasjonseffekter. Vurderingen er beskrevet i detalj i Vedlegg 1 (delkapittel 7.1).
- Havforskningsinstituttet fraråder seismikk i en buffersone rundt gyteområdet og gytevandringsområdet på 20 nmi for ordinære undersøkelser og 5 nmi for borestedsundersøkelser. Alternativt må det dokumenteres at lydnivået ikke overstiger 145 dB re 1µPa2s SEL integrert over 10 sekund i gyte/gytevandrings området, og prosedyren som beskrevet over følges.
- Havforskningsinstituttet fraråder seismikkundersøkelser i områder og perioder hvor intensivt beite for bardehval forekommer.
- Kart for gyte-, gytevandrings- og beiteområder er slått sammen til rådgivningskart for 2 ukers bolker gjennom året. Rådgivningskart for 2023, samt beskrivelse av kartlag og hvordan disse kan lastes ned finnes i Vedlegg 1. Disse inngår også i meldesystemet til Oljedirektoratet.
- Havforskningsinstituttet fraråder seismikkundersøkelser i områder med høy tetthet av grønlandshval og råder til at det brukes en dedikert observatør ved seismikkundersøkelser i områder hvor denne kan påtreffes. Dersom en grønlandshval påtreffes innen en radius på 1000m tilråder Havforskningsinstituttet at seismikkskyting avventes i 30 minutter så lenge dyret ikke lenger er nærmere enn1000 m unna, og at ramp up brukes når seismikk starter opp igjen. Kart over områdene hvor dette er anbefalt finnes i Vedlegg 1.
3.7 - Referanser
Abma, R., & Ross, A. (2015). Practical aspects of the popcorn source method. In SEG Technical Program Expanded Abstracts 2015 (pp. 164-169). Society of Exploration Geophysicists.
Ainslie, M. A., Halvorsen, M. B., Dekeling, P. A., M., L. R., Duncan, A. J., Frankel, A. S., . . . Zeddies, D. G. (2016). Verification of airgun sound field models for environmental impacts assessment. Proceedings of Meetings on Acoustics., 27(1). doi: https://doi.org/10.1121/2.0000339
Ainslie, M. A., Laws, R. M., & Sertlek, H. Ö. (2019). International Airgun Modeling Workshop: Validation of Source Signature and Sound Propagation Models—Dublin (Ireland), July 16, 2016—Problem Description. IEEE Journal of Oceanic Engineering, 44(3), 565-574.
Ahonen, H., Stafford, K. M., de Steur, L., Lydersen, C., Wiig, Ø., & Kovacs, K. M. (2017). The underwater soundscape in western Fram Strait: Breeding ground of Spitsbergen's endangered bowhead whales. Marine pollution bulletin, 123(1-2), 97-112.
Amorim, M. C. P., Vieira, M., Meireles, G., Novais, S. C., Lemos, M. F., Modesto, T., ... & Fonseca, P. J. (2022). Boat noise impacts Lusitanian toadfish breeding males and reproductive outcome. Science of The Total Environment, 830, 154735.
Artsdatabanken (2021, 24. november). Norsk rødliste for arter 2021. https://www.artsdatabanken.no/lister/rodlisteforarter/2021
Birin, I., & Maglić, L. (2020). Analysis of Seismic Methods Used for Subsea Hydrocarbon Exploration. Pomorski zbornik, 58(1), 77-89. . doi:https://doi.org/10.18048/2020.58.05.
Bjørge A., Lydersen C., Skern-Mauritzen M., Wiig Ø. (2010). Sjøens pattedyr. Fisken og havet, særnummer 2-2010. (ISSN 0802 0620 ).
Blackwell, S. B., Nations, C. S., McDonald, T. L., Thode, A. M., Mathias, D., Kim, K. H., . . . Macrander, A. M. (2015). Effects of Airgun Sounds on Bowhead Whale Calling Rates: Evidence for Two Behavioral Thresholds. Plos One, 10(6). doi:10.1371/journal.pone.0125720
Blom, E.L., Kvarnemo, C., Dekhla I., Schöld S. Andersson, M.H., Svensson, O., Amorim, M.C.P. (2019). Continuous but not intermittent noise has a negative impact on mating success in a marine fish with paternal care. Scientific Reports, 9(5494). doi:https://doi.org/10.1038/s41598-019-41786-x.
Cada, G., Loar, J., Garrison, L., Fisher, R., & Neitzel, D. (2006). Efforts to reduce mortality to hydroelectric turbine-passed fish: Locating and quantifying damaging shear stresses. Environmental Management, 37(6), 898-906. doi:10.1007/s00267-005-0061-1
Caldwell, J., & Dragoset, W. (2000). A brief overview of seismic air-gun arrays. The leading edge, 19(8), 898-902
Carroll, A. G., Przeslawski, R., Duncan, A., Gunning, M., & Bruce, B. (2017). A critical review of the potential impacts of marine seismic surveys on fish & invertebrates. Marine Pollution Bulletin, 114(1), 9-24. doi:10.1016/j.marpolbul.2016.11.038
Castellote, M., Clark, C. W., & Lammers, M. O. (2012). Acoustic and behavioural changes by fin whales (Balaenoptera physalus) in response to shipping and airgun noise. Biological Conservation, 147(1), 115-122. doi:10.1016/j.biocon.2011.12.021
Cerchio, S., Strindberg, S., Collins, T., Bennett, C., & Rosenbaum, H. (2014). Seismic Surveys Negatively Affect Humpback Whale Singing Activity off Northern Angola. Plos One, 9(3). doi:10.1371/journal.pone.0086464
Chapman, C., Hawkins, A., 1969. The importance of sound in fish behaviour in relation to capture by trawls. FAO Fisheries and Aquaculture Report (FAO), 62(3):717-729.
Chin-Bing, S. A., King, D. B., Davis, J. A., & Evans, R. B. (1993). PE Workshop II. Proceedings of the Second Parabolic Equation Workshop. NAVAL RESEARCH LAB STENNIS SPACE CENTER MS.
Clark, C. W., & Gagnon, G. C. (2006). Considering the temporal and spatial scales of noise exposures from seismic surveys on baleen whales. Int. Whal. Comm. SC58/E9.
Comiso, J. C., & Hall, D. K. (2014). Climate trends in the Arctic as observed from space. . WIREs Climate Change, 5, 389-409.
Dalen, J., Ona, E., Soldal, A. V., & Sætre, R. (1996). Seismiske undersøkelser til havs: en vurdering av konsekvenser for fisk og fiskerier.
Dalen, J., Hovem, J.M., Karlsen, H.E., Kvadsheim, P.H., Løkkeborg, S., Mjelde, R., Pedersen, A., Skiftesvik A.B. 2008. Kunnskapstatus og forskningsbehov med hensyn til skremmeeffekter og skadevirkninger av seismiske lydbølger på fisk og sjøpattedyr. Rapport til Oljedirektoratet, Fiskeridirektoratet og Statens Forurensingstilsyn. ISBN 82-7257-661-9
Davidsen, J. G., Dong, H. F., Linne, M., Andersson, M. H., Piper, A., Prystay, T. S., . . . Hawkins, A. D. (2019). Effects of sound exposure from a seismic airgun on heart rate, acceleration and depth use in free-swimming Atlantic cod and saithe. Conservation Physiology, 7. doi:10.1093/conphys/coz020
Day, R. D., Fitzgibbon, Q. P., McCauley, R. D., Baker, K. B., & Semmens, J. M. (2022). The impact of seismic survey exposure on the righting reflex and moult cycle of Southern Rock Lobster (Jasus edwardsii) puerulus larvae and juveniles. In Environmental Pollution (Vol. 309, p. 119699). Elsevier BV. https://doi.org/10.1016/j.envpol.2022.119699
de Jong, K., Amorim, M. C. P., Fonseca, P. J., Fox, C. J., & Heubel, K. U. (2018). Noise can affect acoustic communication and subsequent spawning success in fish. Environmental Pollution, 237, 814-823. doi:10.1016/j.envpol.2017.11.003
de Jong, K., Amorim, M. C. P., Fonseca, P. J., & Heubel, K. U. (2018). Noise Affects Multimodal Communication During Courtship in a Marine Fish. Frontiers in Ecology and Evolution, 6. doi:10.3389/fevo.2018.00113
de Jong, K., Forland, T. N., Amorim, M. C. P., Rieucau, G., Slabbekoorn, H., & Sivle, L. D. (2020). Predicting the effects of anthropogenic noise on fish reproduction. Reviews in Fish Biology and Fisheries, 30(2), 245-268. doi:10.1007/s11160-020-09598-9
Dellinger, J., Ross, A., Meaux, D., Brenders, A., Gesoff, G., Etgen, J., ... & Harper, M. (2016). Wolfspar®, an “FWI-friendly” ultralow-frequency marine seismic source. In SEG Technical Program Expanded Abstracts 2016 (pp. 4891-4895). Society of Exploration Geophysicists.
De Robertis, A., & Handegard, N. O. (2013). Fish avoidance of research vessels and the efficacy of noise-reduced vessels: a review. Ices Journal of Marine Science, 70(1), 34-45. doi:10.1093/icesjms/fss155
Di Iorio, L., & Clark, C. W. (2010). Exposure to seismic survey alters blue whale acoustic communication. Biology Letters, 6(3), 334-335. doi:10.1098/rsbl.2009.0967
Dhelie, P. E., Danielsen, V., Lie, J. E., Branston, M., Campbell, R., & Ford, R. (2018, August). Improving seismic data by using smaller more compact seismic air-gun sources. In Marine Acquisition Workshop 2018 (pp. cp-560). European Association of Geoscientists & Engineers.
Duarte, C. M., Chapuis, L., Collin, S. P., Costa, D. P., Devassy, R. P., Eguiluz, V. M., Erbe, C., Gordon, T. A. C., Halpern, B. S., Harding, H. R., Havlik, M. N., Meekan, M., Merchant, N. D., Miksis-Olds, J. L., Parsons, M., Predragovic, M., Radford, A. N., Radford, C. A., Simpson, S. D., … Juanes, F. (2021). The soundscape of the Anthropocene ocean. In Science (Vol. 371, Issue 6529). American Association for the Advancement of Science (AAAS). https://doi.org/10.1126/science.aba4658
Duncan, A. J., Weilgart, L. S., Leaper, R., Jasny, M., & Livermore, S. (2017). A modelling comparison between received sound levels produced by a marine Vibroseis array and those from an airgun array for some typical seismic survey scenarios. In Marine Pollution Bulletin (Vol. 119, Issue 1, pp. 277–288). Elsevier BV. https://doi.org/10.1016/j.marpolbul.2017.04.001
Dunlop, R. A., McCauley, R. D., & Noad, M. J. (2020). Ships and air guns reduce social interactions in humpback whales at greater ranges than other behavioral impacts. Marine Pollution Bulletin, 154. doi:10.1016/j.marpolbul.2020.111072
Dunlop, R. A., Noad, M. J., McCauley, R. D., Kniest, E., Slade, R., Paton, D., & Cato, D. H. (2016). Response of humpback whales (Megaptera novaeangliae) to ramp-up of a small experimental air gun array. Marine Pollution Bulletin, 103(1-2), 72-83. doi:10.1016/j.marpolbul.2015.12.044
Dunlop, R. A., Noad, M. J., McCauley, R. D., Kniest, E., Paton, D., & Cato, D. H. (2015). The Behavioural Response of Humpback Whales (Megaptera novaeangliae) to a 20 Cubic Inch Air Gun. Aquatic Mammals, 41(4), 412-433. doi:10.1578/am.41.4.2015.412
Dunlop, R. A., Braithwaite, J., Mortensen, L. O., & Harris, C. M. (2021). Assessing Population-Level Effects of Anthropogenic Disturbance on a Marine Mammal Population. Frontiers in Marine Science, 8. doi:10.3389/fmars.2021.624981
Dunlop, R. A., Noad, M. J., McCauley, R. D., Kniest, E., Slade, R., Paton, D., & Cato, D. H. (2018). A behavioural dose-response model for migrating humpback whales and seismic air gun noise. Marine pollution bulletin, 133, 506-516.
English, J. M., Inkpen, S. T., & Guigne, J. Y. (1991). A New, High-Frequency, Broadband Seismic Source. In All Days. Offshore Technology Conference. OTC. https://doi.org/10.4043/6558-ms
Eriksen, E., van der Meeren, G. I., Nilsen, B. M., Quillfeldt, C. H. & Johnsen, H. (2021). Særlig verdifulle og sårbare områder (SVO) i norske havområder – Miljøverdi. Rapport fra havforskningen 2021-26.
Engås, A., Løkkeborg, S., Ona, E., & Soldal, A. V. (1996). Effects of seismic shooting on local abundance and catch rates of cod (Gadus morhua) and haddock (Melanogrammus aeglefinus). Canadian Journal of Fisheries and Aquatic Sciences, 53(10), 2238-2249. doi:10.1139/f96-177
Environment Australia (2001). Guidelines on the Application of the Environment Protection and Biodiversity Conservation Act to Interactions Between Offshore Seismic Operations and Larger Cetaceans.
Farcas, A., Thompson, P. M., & Merchant, N. D. (2016). Underwater noise modelling for environmental impact assessment. Environmental Impact Assessment Review, 57, 114-122. doi:10.1016/j.eiar.2015.11.012
Feltham A., Girard M., Jenkerson M., Nechayuk V., Griswold S., Henderson N. et al. 2017. The Marine Vibrator Joint Industry Project: four years on. Exploration Geophysics 49, 675–687
Fewtrell, J. L., & McCauley, R. D. (2012). Impact of air gun noise on the behaviour of marine fish and squid. Marine Pollution Bulletin, 64(5), 984-993. doi:10.1016/j.marpolbul.2012.02.009
Fields, D. M., Handegard, N. O., Dalen, J., Eichner, C., Malde, K., Karlsen, O., . . . Browman, H. I. (2019). Airgun blasts used in marine seismic surveys have limited effects on mortality, and no sublethal effects on behaviour or gene expression, in the copepod Calanus finmarchitcus. Ices Journal of Marine Science, 76(7), 2033-2044. doi:10.1093/icesjms/fsz126
Filiciotto, F., Vazzana, M., Celi, M., Maccarrone, V., Ceraulo, M., Buffa, G., ... & Buscaino, G. (2016). Underwater noise from boats: Measurement of its influence on the behaviour and biochemistry of the common prawn (Palaemon serratus, Pennant 1777). Journal of Experimental Marine Biology and Ecology, 478, 24-33.
Finley, K. J., Miller, G. W., Davis, R. A., & Greene, C. R. (1990). Reactions of belugas, Delphinapterus leucas, and narwhals, Monodon monoceros, to ice-breaking ships in the Canadian high arctic. . Canadian Journal of Fisheries and Aquatic Sciences, 224, 97-117.
Finneran, J. J. (2015). Noise-induced hearing loss in marine mammals: A review of temporary threshold shift studies from 1996 to 2015. Journal of the Acoustical Society of America, 138(3), 1702-1726. doi:10.1121/1.4927418
Finneran, J. J., Carder, D. A., Schlundt, C. E., & Dear, R. L. (2010). Temporary threshold shift in a bottlenose dolphin (Tursiops truncatus) exposed to intermittent tones. Journal of the Acoustical Society of America, 127(5), 3267-3272. doi:10.1121/1.3377052
Finneran, J. J., Dear, R., Carder, D. A., & Ridgway, S. H. (2003). Auditory and behavioral responses of California sea lions (Zalophus californianus) to single underwater impulses from an arc-gap transducer. Journal of the Acoustical Society of America, 114(3), 1667-1677. doi:10.1121/1.1598194
Finneran, J. J., & Schlundt, C. E. (2010). Frequency-dependent and longitudinal changes in noise-induced hearing loss in a bottlenose dolpHin (Tursiops truncatus) (L). Journal of the Acoustical Society of America, 128(2), 567-570. doi:10.1121/1.3458814
Fitzgibbon, Q. P., Day, R. D., McCauley, R. D., Simon, C. J., & Semmens, J. M. (2017). The impact of seismic air gun exposure on the haemolymph physiology and nutritional condition of spiny lobster, Jasus edwardsii. Marine Pollution Bulletin, 125(1-2), 146-156.
Gallagher, C. A., Grimm, V., Kyhn, L. A., Kinze, C. C., & Nabe-Nielsen, J. (2021). Movement and seasonal energetics mediate vulnerability to disturbance in marine mammal populations. The American Naturalist, 197(3), 296-311.
Gerez, D. et al. (2020). A practical marine-vibrator system optimized for geophysical applications. Conference paper. 82nd EAGE Conference & Exhibition 2020, Amsterdam, The Netherlands
Gisiner, R. C. (2016). Sound and marine seismic surveys. . Acosutics Today 12, 10-18.
Goldbogen, J. A., Southall, B. L., DeRuiter, S. L., Calambokidis, J., Friedlaender, A. S., Hazen, E. L., . . . Tyack, P. L. (2013). Blue whales respond to simulated mid-frequency military sonar. Proceedings of the Royal Society B-Biological Sciences, 280(1765). doi:10.1098/rspb.2013.0657
Halvorsen, M. B., Casper, M. B., Carlson, T. J., Woodley, C. M. & Popper, A. N. (2011). Assessment of barotrauma injury and cumulative sound exposure level in salmon after exposure to impulsive sound. In: The effects of noise on aquatic life: second international congress. Ed. Hawkins, A. A., New York, Springer Verlag
Halvorsen, M. B., Casper, B. M., Carlson, T. J., Woodley, C. M., & Popper, A. N. (2012a). Assessment of Barotrauma Injury and Cumulative Sound Exposure Level in Salmon After Exposure to Impulsive Sound. In A. N. Popper & A. Hawkins (Eds.), Effects of Noise on Aquatic Life (Vol. 730, pp. 235-237).
Halvorsen, M. B., Casper, B. M., Woodley, C. M., Carlson, T. J., & Popper, A. N. (2012b). Threshold for Onset of Injury in Chinook Salmon from Exposure to Impulsive Pile Driving Sounds. Plos One, 7(6). doi:10.1371/journal.pone.0038968
Halvorsen, M. B., Zeddies, D. G., Chicoine, D., & Popper, A. N. (2013). Effects of low-frequency naval sonar exposure on three species of fish. Journal of the Acoustical Society of America, 134(2), EL205-EL210. doi:10.1121/1.4812818
Handegard, N. O., Tronstad, T. V., & Hovem, J. M. (2013). Evaluating the effect of seismic surveys on fish - the efficacy of different exposure metrics to explain disturbance. Canadian Journal of Fisheries and Aquatic Sciences, 70(9), 1271-1277. doi:10.1139/cjfas-2012-0465
Harris, C. M., Sadykova, D., DeRuiter, S. L., Tyack, P. L., Miller, P. J. O., Kvadsheim, P. H., . . . Thomas, L. (2015). Dose response severity functions for acoustic disturbance in cetaceans using recurrent event survival analysis. Ecosphere, 6(11). doi:10.1890/es15-00242.1
Harris, R. E., Miller, G. W., & Richardson, W. J. (2001). Seal responses to airgun sounds during summer seismic surveys in the Alaskan Beaufort Sea. Marine Mammal Science, 17(4), 795-812. doi:10.1111/j.1748-7692.2001.tb01299.x
Hassel, A., Knutsen, T., Dalen, J., Skaar, K., Løkkeborg, S., Misund, O.A., Østensen, Ø., Fonn, M. and Haugland, E.K. Influence of seismic shooting on the lesser sandeel (Ammodytes marinus), ICES Journal of Marine Science, Volume 61, Issue 7, 2004, Pages 1165–1173,
Hawkins, A. D., Johnson, C., & Popper, A. N. (2020). How to set sound exposure criteria for fishes. Journal of the Acoustical Society of America, 147(3), 1762-1777. doi:10.1121/10.0000907
Heaney, K. D., & Campbell, R. L. (2019). Parabolic Equation Modeling of a Seismic Airgun Array. IEEE Journal of Oceanic Engineering, 44(3), 621-632. doi:10.1109/joe.2019.2912060
Hegna, S., & Parkes, G. (2011, September). The low frequency output of marine air-gun arrays. In 2011 SEG Annual Meeting. OnePetro.
Hegna S, Klüver T & Lima J (2018) Benefits of continuous source and receiver side wavefields, SEG Technical Program Expanded Abstracts 2018, pp. 41-45, Society of Exploration Geophysicists.
Hegna, S. (2021). Estimation of the acoustic wavefield generated by a seismic vessel from towed-streamer data. First International Meeting for Applied Geoscience & Energy Expanded Abstracts. First International Meeting for Applied Geoscience & Energy.
Hegna, S. (2022, June). The Acoustic Wavefield Generated by a Vessel Sailing on Top of a Streamer Spread, European Association of Geoscientists & Engineers, Conference Proceedings, 83rd EAGE Annual Conference & Exhibition, Volume 2022, p.1 - 5
Heide-Jørgensen, M. P., Hansen, R. G., Westdal, K., Reeves, R. R., & Mosbech, A. (2013). Narwhals and seismic exploration: Is seismic noise increasing the risk of ice entrapments? Biological Conservation, 158, 50-54. doi:10.1016/j.biocon.2012.08.005
Heide-Jørgensen, M. P., Blackwell, S. B., Tervo, O. M., Samson, A. L., Garde, E., Hansen, R. G., ... & Ditlevsen, S. (2021). Behavioral response study on seismic airgun and vessel exposures in narwhals. Frontiers in Marine Science, 665. https://doi.org/10.3389/fmars.2021.658173
Henriksen, S., & Hilmo, O. (2015). Norsk rødliste for arter 2015. http://www.artsdatabanken.no/Rodliste/EuropeiskPerspektiv
Hubert, J., Cambell, J. A., & Slabbekoorn, H. (2020). Effects of seismic air gun playbacks on swimming patterns and behavioural states of Atlantic cod in a net pen. Marine Pollution Bullentin, 160(111680).
Huserbråten, M. B. O., Eriksen, E., Gjosaeter, H., & Vikebo, F. (2019). Polar cod in jeopardy under the retreating Arctic sea ice. Communications Biology, 2. doi:10.1038/s42003-019-0649-2
ICES (2018). Report of the Working Group on Elasmobranch Fishes (WGEF), 19–28 June 2018, Lisbon, Portugal. ICES CM 2018/ACOM:16. 1306 pp.
Ivanova, S. V., Kessel, S. T., Espinoza, M., McLean, M. F., O'Neill, C., Landry, J., . . . Fisk, A. T. (2020). Shipping alters the movement and behavior of Arctic cod (Boreogadus saida), a keystone fish in Arctic marine ecosystems. Ecological Applications, 30(3). doi:10.1002/eap.2050
Kastak, D., Schusterman, R. J., Southall, B. L., & Reichmuth, C. J. (1999). Underwater temporary threshold shift induced by octave-band noise in three species of pinniped. Journal of the Acoustical Society of America, 106(2), 1142-1148. doi:10.1121/1.427122
Kastelein, R. A., Gransier, R., Hoek, L., & de Jong, C. A. F. (2012a). The hearing threshold of a harbor porpoise (Phocoena phocoena) for impulsive sounds (L). Journal of the Acoustical Society of America, 132(2), 607-610. doi:10.1121/1.4733552
Kastelein, R. A., Gransier, R., Hoek, L., Macleod, A., & Terhune, J. M. (2012b). Hearing threshold shifts and recovery in harbor seals (Phoca vitulina) after octave-band noise exposure at 4 kHz. Journal of the Acoustical Society of America, 132(4), 2745-2761. doi:10.1121/1.4747013
Kastelein, R. A., Gransier, R., & Hoek, L. (2013). Comparative temporary threshold shifts in a harbor porpoise and harbor seal, and severe sHift in a seal (L). Journal of the Acoustical Society of America, 134(1), 13-16. doi:10.1121/1.4808078
Kastelein, R. A., von Benda-Beckmann, S. M., Lam, F. P. A., Jansen, E., & de Jong, C. A. F. (2019). Effect of a Bubble Screen on the Behavioral Responses of Captive Harbor Porpoises (Phocoena phocoena) Exposed to Airgun Sounds. Aquatic Mammals, 45(6), 706-716. doi:10.1578/am.45.6.2019.706
Kastelein, R. A., Helder-Hoek, L., Cornelisse, S. A., von Benda-Beckmann, A. M., Lam, F. P. A., de Jong, C. A. F., & Ketten, D. R. (2020). Lack of reproducibility of temporary hearing threshold shifts in a harbor porpoise after exposure to repeated airgun sounds. Journal of the Acoustical Society of America, 148(2), 556-565. doi:10.1121/10.0001668
Kavanagh, A. S., Nykanen, M., Hunt, W., Richardson, N., & Jessopp, M. J. (2019). Seismic surveys reduce cetacean sightings across a large marine ecosystem. Scientific Reports, 9. doi:10.1038/s41598-019-55500-4
Khodabandeloo, B., & Landrø, M. (2018). Acoustically induced cavity cloud generated by air-gun arrays-Comparing video recordings and acoustic data to modeling. Journal of the Acoustical Society of America, 143(6), 3383-3393. doi:10.1121/1.5040490
Kluesner, J., Brothers, D., Hart, P., Miller, N., & Hatcher, G. (2018). Practical approaches to maximizing the resolution of sparker seismic reflection data. In Marine Geophysical Research (Vol. 40, Issue 3, pp. 279–301). Springer Science and Business Media LLC. https://doi.org/10.1007/s11001-018-9367-2
Klüver T, Hegna S & Lima J (2018) Processing of data with continuous source and receiver side wavefields: Real data examples. SEG Technical Program Expanded Abstracts 2018, pp. 4045- 4049, Society of Exploration Geophysicists
Klüver, T., & Høy, T. (2022, August). Efficient acquisition and imaging of high-resolution seismic data with the continuous wavefields method. In Second International Meeting for Applied Geoscience & Energy (pp. 36-40). Society of Exploration Geophysicists and American Association of Petroleum Geologists.
Kok, A.C., Bruil, L., Berges, B., Sakinan, S., Debusschere, E., Reubens, J., de Haan, D., Norro, A. and Slabbekoorn, H., 2021. An echosounder view on the potential effects of impulsive noise pollution on pelagic fish around windfarms in the North Sea. Environmental Pollution, p.118063.
Kvadsheim PH, DeRuiter S, Sivle LD, Goldbogen J, Hansen RR, Miller P, Lam FP, Calambokidis J, Friedlaender A, Visser F, Tyack P, Kleivane L & Southall, B (2017). Avoidance Responses of Minke Whales to 1-4 kHz Naval Sonar. Marine Pollution Bulletin (2017): http://dx.doi.org/10.1016/j.marpolbul.2017.05.037
Kvadsheim, P. H., Forland, T. N., de Jong, K., Nyqvist, D., Grimsbø, E., & Sivle, L. D. (2020). Effekter av støyforurensning på havmiljø – kunnskapsstatus og forvaltningsrådgiving. FFI-RAPPORT 20/01015. DOI: 978-82-464-3262-5
Langhammer, J., & Bennion, P. (2015, June). Triple-source simultaneous shooting (TS3), a future for higher density seismic?. In 77th EAGE Conference and Exhibition 2015 (Vol. 2015, No. 1, pp. 1-5). European Association of Geoscientists & Engineers.
Landrø, M., Amundsen, L. (2022). Improved Low Frequency Output from Seismic Air Guns by Firing at Shallow Depths? Conference Proceedings, Third EAGE Marine Acquisition Workshop, Volume 2022, p.1 - 4Langhammer, J., & Bennion, P. (2015, June). Triple-source simultaneous shooting (TS3), a future for higher density seismic?. In 77th EAGE Conference and Exhibition 2015 (Vol. 2015, No. 1, pp. 1-5). European Association of Geoscientists & Engineers
Laws, R. M., Halliday, D., Hopperstad, J. ‐F., Gerez, D., Supawala, M., Özbek, A., Murray, T., & Kragh, E. (2018). Marine vibrators: the new phase of seismic exploration. Geophysical Prospecting, 67(6), 1443–1471. https://doi.org/10.1111/1365-2478.12708
Lind, S., Ingvaldsen, R. B., & Furevik, T. (2016). Arctic layer salinity controls heat loss from deep Atlantic layer in seasonally ice-covered areas of the Barents Sea. Geophysical Research Letters, 43(10), 5233-5242. doi:10.1002/2016gl068421
Long, A., & Tenghamn, R. (2018). Marine vibrator concepts for modern seismic challenges. ASEG Extended Abstracts, 2018(1), 1-4.
Lydersen, C., Vacquié-Garcia, J., Heide-Jørgensen, M. P., Øien, N., Guinet, C. and Kovacs, K. M. 2020. Autumn movements of fin whales (Balaenoptera physalus) from Svalbard, Norway, revealed by satellite tracking. Sci. Rep. 10, 16966: 1-13. https://doi.org/10.1038/s41598-020-73996-z
Løkkeborg, S., Ona, E., Vold, A., & Salthaug, A. (2012). Sounds from seismic air guns: gear- and species-specific effects on catch rates and fish distribution. Canadian Journal of Fisheries and Aquatic Sciences, 69(8), 1278-1291. doi:10.1139/f2012-059
Matthews, M.-N. R., Ireland, D. S., Zeddies, D. G., Brune, R. H., & Pyć, C. D. (2020). A Modeling Comparison of the Potential Effects on Marine Mammals from Sounds Produced by Marine Vibroseis and Air Gun Seismic Sources. Journal of Marine Science and Engineering, 9(1), 12. https://doi.org/10.3390/jmse9010012
Malme, C.I., Smith , P.W. and Miles, P.R. 1986. Study of the Effects of Offshore Geophysical Acoustic Survey Operations On Important Commersial Fisheries in California. Technical Report No. 1, Report No. 6125. Contract No. MMS 14-12-0001-30273. Prepared by BBN Laboratories Inc., Cambridge, Mass., for Battelle, Ventura Office, CA, USA. 92 s.
Mauro, M., Pérez-Arjona, I., Perez, E. J. B., Ceraulo, M., Bou-Cabo, M., Benson, T., ... & Buscaino, G. (2020). The effect of low frequency noise on the behaviour of juvenile Sparus aurata. The Journal of the Acoustical Society of America, 147(6), 3795-3807.
McCauley, R. D., Day, R. D., Swadling, K. M., Fitzgibbon, Q. P., Watson, R. A., & Semmens, J. M. (2017). Widely used marine seismic survey air gun operations negatively impact zooplankton. Nature Ecology & Evolution, 1(7). doi:10.1038/s41559-017-0195
McCauley, R. D., Fewtrell, J., & Popper, A. N. (2003). High intensity anthropogenic sound damages fish ears. Journal of the Acoustical Society of America, 113(1), 638-642. doi:10.1121/1.1527962
McCauley, R. D., Meekan, M. G., & Parsons, M. J. (2021). Acoustic Pressure, Particle Motion, and Induced Ground Motion Signals from a Commercial Seismic Survey Array and Potential Implications for Environmental Monitoring. Journal of Marine Science and Engineering, 9(6), 571
McQueen, K., Meager, J. J., Nyqvist, D., Skjæraasen, J. E., Olsen, E. M., Karlsen, Ø., Kvadsheim, P. H., Handegard, N. O.,
Forland, T. N., & Sivle, L. D. (2022). Spawning Atlantic cod (Gadus morhua L.) exposed to noise from seismic airguns do not abandon their spawning site. In F. Juanes (Ed.), ICES Journal of Marine Science. Oxford University Press (OUP). https://doi.org/10.1093/icesjms/fsac203
Meld.St. 20 (2019-2020) Melding til Stortinget. Oppdatering av forvaltningsplanen for det marine miljø i Barentshavet og havområdene utenfor Lofoten.
Meekan, M.G., Speed, C.W., McCauley, R.D., Fisher, R., Birt, M.J., Currey-Randall, L.M., Semmens, J.M., Newman, S.J., Cure, K., Stowar, M., Vaughan, B., Parsons, M.J.G., 2021. A large-scale experiment finds no evidence that a seismic survey impacts a demersal fish fauna. PNAS, 118: doi.org/10.1073/pnas.2100869118
Miller, I., & Cripps, E. (2013). Three dimensional marine seismic survey has no measurable effect on species richness or abundance of a coral reef associated fish community. Marine Pollution Bulletin, 77(1-2), 63-70. doi:10.1016/j.marpolbul.2013.10.031
Miller, P. J. O., Kvadsheim, P. H., Lam, F. P. A., Wensveen, P. J., Antunes, R., Alves, A. C., . . . Sivle, L. D. (2012). The Severity of Behavioral Changes Observed During Experimental Exposures of Killer (Orcinus orca), Long-Finned Pilot (Globicephala melas), and Sperm (Physeter macrocephalus) Whales to Naval Sonar. Aquatic Mammals, 38(4), 362-401. doi:10.1578/am.38.4.2012.362
Miller, P. J. O., Johnson, M. P., Madsen, P. T., Biassoni, N., Quero, M., & Tyack, P. L. (2009). Using at-sea experiments to study the effects of airguns on the foraging behavior of sperm whales in the Gulf of Mexico. Deep-Sea Research Part I-Oceanographic Research Papers, 56(7), 1168-1181. doi:10.1016/j.dsr.2009.02.008
Miller, P. J., Antunes, R. N., Wensveen, P. J., Samarra, F. I., Catarina Alves, A., Tyack, P. L., ... & Thomas, L. (2014). Dose-response relationships for the onset of avoidance of sonar by free-ranging killer whales. The Journal of the Acoustical Society of America, 135, 975-993.
Miller, P. J. O., Kvadsheim, P. H., Lam, F. P. A., Tyack, P. L., Cure, C., DeRuiter, S. L., . . . Hooker, S. K. (2015). First indications that northern bottlenose whales are sensitive to behavioural disturbance from anthropogenic noise. Royal Society Open Science, 2(6). doi:10.1098/rsos.140484
Miller, P.J.O., Siegal, E., Isojuuno, S., Lam F.P.A, Kvadsheim, P. H., & Curé, C. (2019). The underwater soundscape of fear: responses of cetaceans to naval sonar and playback of predatory killer whale sounds. Paper presented at the World Marine Mammals Conference (https://www.wmmconference.org/), Barcelona, Spain.
Morris, C. J., Cote, D., Martin, B., & Kehler, D. (2018). Effects of 2D seismic on the snow crab fishery. Fisheries Research, 197, 67-77.
Morozov, A. K., & Webb, D. C. (2022). Experimental Marine Vibrator With a Helmholtz Bubble Resonator in the Frequency Range 5–16 Hz. IEEE Journal of Oceanic Engineering, 47(4), 984-994.
Nedelec, S. L., Radford, A. N., Gatenby, P., Davidson, I. K., Velasquez Jimenez, L., Travis, M., ... & Simpson, S. D. (2022). Limiting motorboat noise on coral reefs boosts fish reproductive success. Nature communications, 13(1), 1-9.
Nedelec, S. L., Radford, A. N., Pearl, L., Nedelec, B., McCormick, M. I., Meekan, M. G., & Simpson, S. D. (2017). Motorboat noise impacts parental behaviour and offspring survival in a reef fish. Proceedings of the Royal Society B: Biological Sciences, 284(1856), 20170143.
NOAA. (2015). National Oceanic and Atmospheric Administration draft guidance for assessing the effects of anthropogenic sound on marine mammal hearing: underwater acoustic threshold levels for onset of permanent and temporary threshold shifts. Draft Report, July 2015. . Retrieved from www.nmfs.noaa.gov/pr/acoustics/guidelines.htm
Oscarsson-Nagel, M. D. C., W. Söllner, Ø. Trætten, B. Armstrong, D. Nams, and P. Yeatman. "Marine vibrator source: Modular system with folded oscillating surface." In 81st EAGE Conference and Exhibition 2019, vol. 2019, no. 1, pp. 1-5. European Association of Geoscientists & Engineers, 2019
Paxton, A. B., Taylor, J. C., Nowacek, D. P., Dale, J., Cole, E., Voss, C. M., & Peterson, C. H. (2017). Seismic survey noise disrupted fish use of a temperate reef. Marine Policy, 78, 68-73. doi:10.1016/j.marpol.2016.12.017
Pearson, W.H, Skalski, J.R., and Malme, C.I. 1987. Effects of Sounds from a Geophysical Survey Device on Fishing Success. OCS Study MMS-86-0032. Prepared by BBN Laboratories Inc., Cambridge, Mass., og Battelle, Marine Research Laboratory, Washington, contract No. 14-12-0001-30273, to the Department of the Interior, Mineral Management Service, Pacific Outer Continental Shelf Region, Los Angeles, California. 293 s.
Pearson, W. H., Skalski, J. R., & Malme, C. I. (1992). EFFECTS OF SOUNDS FROM A GEOPHYSICAL SURVEY DEVICE ON BEHAVIOR OF CAPTIVE ROCKFISH (SEBASTES SPP). Canadian Journal of Fisheries and Aquatic Sciences, 49(7), 1343-1356. doi:10.1139/f92-150
Pena, H., Handegard, N. O., & Ona, E. (2013). Feeding herring schools do not react to seismic air gun surveys. Ices Journal of Marine Science, 70(6), 1174-1180. doi:10.1093/icesjms/fst079
Picciulin, M., Sebastianutto, L., Codarin, A., Farina, A., & Ferrero, E. A. (2010). In situ behavioural responses to boat noise exposure of Gobius cruentatus (Gmelin, 1789; fam. Gobiidae) and Chromis chromis (Linnaeus, 1758; fam. Pomacentridae) living in a Marine Protected Area. Journal of Experimental Marine Biology and Ecology, 386(1-2), 125-132. doi:10.1016/j.jembe.2010.02.012
Pirotta, E., Brookes, K. L., Graham, I. M., & Thompson, P. M. (2014). Variation in harbour porpoise activity in response to seismic survey noise. Biology Letters, 10(5). doi:10.1098/rsbl.2013.1090
Pirotta, E., Booth, C. G., Calambokidis, J., Costa, D. P., Fahlbusch, J. A., Friedlaender, A. S., ... & Southall, B. L. (2022). From individual responses to population effects: Integrating a decade of multidisciplinary research on blue whales and sonar. Animal Conservation.
Popper, A. N., Fewtrell, J., Smith, M. E., & McCauley, R. D. (2004). Anthropogenic sound: Effects on the behaviour and physiology of fishes. . Marine Technology Science J, 37, 35-40.
Popper, A. N., Gross, J. A., Carlson, T. J., Skalski, J., Young, J. V., Hawkins, A. D., & Zeddies, D. (2016). Effects of Exposure to the Sound from Seismic Airguns on Pallid Sturgeon and Paddlefish. Plos One, 11(8). doi:10.1371/journal.pone.0159486
Popper, A. N., & Hastings, M. C. (2009). The effects of anthropogenic sources of sound on fishes. Journal of Fish Biology, 75(3), 455-489. doi:10.1111/j.1095-8649.2009.02319.x
Popper, A. N., & Hawkins, A. D. (2019). An overview of fish bioacoustics and the impacts of anthropogenic sounds on fishes. Journal of Fish Biology, 94(5), 692-713. doi:10.1111/jfb.13948
Popper, A. N., Hawkins, A. D., Fay, R. R., Mann, D. A., Bartol, S., Carlson, T. J., . . . Tavolga, W. N. (2014). Sound Exposure Guidelines for Fishes and Sea Turtles: A Technical Report prepared by ANSI-Accredited Standards Committee S3/SC1 and registered with ANSI, . Retrieved from New York:
Popper, A. N., Smith, M. E., Cott, P. A., Hanna, B. W., MacGillivray, A. O., Austin, M. E., & Mann, D. A. (2005). Effects of exposure to seismic airgun use on hearing of three fish species. Journal of the Acoustical Society of America, 117(6), 3958-3971. doi:10.1121/1.1904386
Popper AN & Hawkins AD (2019). An overview of fish bioacoustics and the impacts of anthropogenic sounds on fishes. Journal of Fish Biology 94(5), 692-713.
Pramik, B., Bell, M. L., Grier, A., & Lindsay, A. (2015). Field testing the AquaVib: An alternate marine seismic source. In SEG Technical Program Expanded Abstracts 2015 (pp. 181-185). Society of Exploration Geophysicists.
Prior, M. K., Duncan, A. J., Sertlek, H. Ö., & Ainslie, M. A. (2019). Modeling acoustical pressure and particle acceleration close to marine seismic airguns and airgun arrays. IEEE Journal of Oceanic Engineering, 44(3), 611-620.
Prior, M. K., Ainslie, M. A., Halvorsen, M. B., Hartstra, I., Laws, R. M., MacGillivray, A., Müller, R., Robinson, S., & Wang, L. (2021). Characterization of the acoustic output of single marine-seismic airguns and clusters: The Svein Vaage dataset. In The Journal of the Acoustical Society of America (Vol. 150, Issue 5, pp. 3675–3692). Acoustical Society of America (ASA). https://doi.org/10.1121/10.0006751
Przeslawski, R., Brooke, B., Carroll, A. G., & Fellows, M. (2018). An integrated approach to assessing marine seismic impacts: lessons learnt from the Gippsland Marine Environmental Monitoring project. Ocean & Coastal Management, 160, 117-123.
Radford, C. A., Tay, K., & Goeritz, M. L. (2022). Comparative sound detection abilities of four decapod crustaceans. In Journal of Experimental Biology (Vol. 225, Issue 1). The Company of Biologists. https://doi.org/10.1242/jeb.243314
Radford, A. N., Lebre, L., Lecaillon, G., Nedelec, S. L., & Simpson, S. D. (2016). Repeated exposure reduces the response to impulsive noise in European seabass. Global Change Biology, 22(10), 3349-3360. doi:10.1111/gcb.13352
Richardson, W. J., Wursig, B., & Greene, C. R. (1986). REACTIONS OF BOWHEAD WHALES, BALAENA-MYSTICETUS, TO SEISMIC EXPLORATION IN THE CANADIAN BEAUFORT SEA. Journal of the Acoustical Society of America, 79(4), 1117-1128. doi:10.1121/1.393384
Robertson, F. C., Koski, W. R., Thomas, T. A., Richardson, W. J., Wursig, B., & Trites, A. W. (2013). Seismic operations have variable effects on dive-cycle behavior of bowhead whales in the Beaufort Sea. Endangered Species Research, 21(2), 143-160. doi:10.3354/esr00515
Robertson, F. C., Koski, W. R., & Trites, A. W. (2016). Behavioral responses affect distribution analyses of bowhead whales in the vicinity of seismic operations. Marine Ecology Progress Series, 549, 243-262. doi:10.3354/meps11665
Rogers, P., Debusschere, E., Haan, D. de, Martin, B., & Slabbekoorn, H. (2021). North Sea soundscapes from a fish perspective: Directional patterns in particle motion and masking potential from anthropogenic noise. Journal of the Acoustical Society of America, 150, 2174–2188.
Sabet, S. S., Wesdorp, K., Campbell, J., Snelderwaard, P., & Slabbekoorn, H. (2016). Behavioural responses to sound exposure in captivity by two fish species with different hearing ability. Animal Behaviour, 116, 1-11. doi:10.1016/j.anbehav.2016.03.027
Sarnocinska, J., Teilmann, J., Balle, J. D., van Beest, F. M., Delefosse, M., & Tougaard, J. (2020). Harbor Porpoise (Phocoena phocoena) Reaction to a 3D Seismic Airgun Survey in the North Sea. Frontiers in Marine Science, 6. doi:10.3389/fmars.2019.00824
Schlundt, C. E., Finneran, J. J., Carder, D. A., & Ridgway, S. H. (2000). Temporary sHift in masked hearing thresholds of bottlenose dolpHins, Tursiops truncatus, and wHite whales, Delphinapterus leucas, after exposure to intense tones. Journal of the Acoustical Society of America, 107(6), 3496-3508. doi:10.1121/1.429420
Screen, J. A., & Simmonds, I. (2010). Increasing fall-winter energy loss from the Arctic Ocean and its role in Arctic temperature amplification. Geophysical Research Letters, 37. doi:10.1029/2010gl044136
Sabet, S., Wesdorp, K., Campbell, J., Snelderwaard, P., & Slabbekoorn, H. (2015). Behavioural responses to sound exposure in captivity by two fish species with different hearing ability. Animal Behaviour, 116, 1-11. doi:10.1016/j.anbehav.2016.03.027
Sierra-Flores, R., Atack, T., Migaud, H., & Davie, A. (2015). Stress response to anthropogenic noise in Atlantic cod Gadus morhua L. Aquacultural Engineering, 67, 67-76. doi:10.1016/j.aquaeng.2015.06.003
Simpson, S. D., Radford, A. N., Nedelec, S. L., Ferrari, M. C. O., CHivers, D. P., McCormick, M. I., & Meekan, M. G. (2016). Anthropogenic noise increases fish mortality by predation. Nature Communications, 7. doi:10.1038/ncomms10544
Sivle, L. D., Forland, T. N., Hansen, R. R., Andersson, M., Grimsbø, E., Linne, M., & Karlsen, H. E. (2017). Behavioural effects of seismic dose escalation exposure on captive mackerel (Scomber scombrus). Rapport fra Havforskningen 34-2017. ISSN 1893-453.
Sivle, L. D., Hansen, R., Karlsen, H. E., & Handegard, N. O. (2016). Mackerel behaviour and seismic signals – a net pen study. Rapport fra Havforskningen 19-2016. ISSN 1893-4536.
Sivle, L. D., Kvadsheim, P. H., Cure, C., Isojunno, S., Wensveen, P. J., Lam, F. P. A., . . . Miller, P. J. O. (2015). Severity of Expert-Identified Behavioural Responses of Humpback Whale, Minke Whale, and Northern Bottlenose Whale to Naval Sonar. Aquatic Mammals, 41(4), 469-502. doi:10.1578/am.41.4.2015.469
Skalski, J. R., Pearson, W. H., & Malme, C. I. (1992). EFFECTS OF SOUNDS FROM A GEOPHYSICAL SURVEY DEVICE ON CATCH-PER-UNIT-EFFORT IN A HOOK-AND-LINE FISHERY FOR ROCKFISH (SEBASTES SPP). Canadian Journal of Fisheries and Aquatic Sciences, 49(7), 1357-1365. doi:10.1139/f92-151
Slabbekoorn, H., Bouton, N., van Opzeeland, I., Coers, A., ten Cate, C., & Popper, A. N. (2010). A noisy spring: the impact of globally rising underwater sound levels on fish. Trends in Ecology & Evolution, 25(7), 419-427. doi:10.1016/j.tree.2010.04.005
Slotte, A., Hansen, K., Dalen, J., & Ona, E. (2004). Acoustic mapping of pelagic fish distribution and abundance in relation to a seismic shooting area off the Norwegian west coast. Fisheries Research, 67(2), 143-150. doi:10.1016/j.fishres.2003.09.046
Smith, M. E., & Monroe, J. D. (2016). Causes and consequences of sensory hair cell damage and recovery in fishes. Fish hearing and bioacoustics, 393-417.
Soldal, A. V., & Løkkeborg, S. (1993). Seismisk aktivitet og fiskefangster. Analyse av innsamlede fangstdata. Fisken og Havet 4-1993. 44 p.
Soto, N. A. D. (2016). Peer-reviewed studies on the effects of anthropogenic noise on marine invertebrates: from scallop larvae to giant squid. The effects of noise on aquatic life II, 17-26.
Southall, B. L., Finneran, J. J., Rcichmuth, C., Nachtigall, P. E., Ketten, D. R., Bowles, A. E., . . . Tyack, P. L. (2019). Marine Mammal Noise Exposure Criteria: Updated Scientific Recommendations for Residual Hearing Effects. Aquatic Mammals, 45(2), 125-232. doi:10.1578/am.45.2.2019.125
Sætre, R.; Ona, E. Seismiske undersøkelser og skader på fiskeegg og -larver; en vurdering av mulige effekter på bestandsnivå. (Seismic investigations and injuries in fish eggs and larvae; an assessment of potential effects on stock level). Institute of Marine Research, Bergen, Norway, 1996, Fisken og Havet, no 8-1996. 25 p. (in Norwegian with English summary).
Tashmukhambetov, A. M., Ioup, G. E., Ioup, J. W., Sidorovskaia, N. A., & Newcomb, J. J. (2008). Three-dimensional seismic array characterization study: Experiment and modeling. Journal of the Acoustical Society of America, 123(6), 4094-4108. doi:10.1121/1.2902185
Thode, A. M., Blackwell, S. B., Conrad, A. S., Kim, K. H., Marques, T., Thomas, L., . . . Broker, K. (2020). Roaring and repetition: How bowhead whales adjust their call density and source level (Lombard effect) in the presence of natural and seismic airgun survey noise. Journal of the Acoustical Society of America, 147(3), 2061-2080. doi:10.1121/10.0000935
Thompson, D., Sjoberg, M., Bryant, M. E., Lovell, P., & Bjorge, A. (1998). Behavioural and physiological responses of harbour (Phoca vitulina) and grey (Halichoerus grypus) seals to seismic surveys. Report to European Commission of BROMMAD project. MAS2 C, 7940098
Thompson, P. M., Brookes, K. L., Graham, I. M., Barton, T. R., Needham, K., Bradbury, G., & Merchant, N. D. (2013). Short-term disturbance by a commercial two-dimensional seismic survey does not lead to long-term displacement of harbour porpoises. Proceedings of the Royal Society B-Biological Sciences, 280(1771). doi:10.1098/rspb.2013.2001
Tougaard, J., Beedholm, K., & Madsen, P. T. (2022). Thresholds for noise induced hearing loss in harbor porpoises and phocid seals. The Journal of the Acoustical Society of America, 151(6), 4252-4263.
Tougaard, J., Wright, A. J., & Madsen, P. T. (2015). Cetacean noise criteria revisited in the light of proposed exposure limits for harbour porpoises. Marine Pollution Bulletin, 90(1-2), 196-208. doi:10.1016/j.marpolbul.2014.10.051
van der Knaap, I., Reubens, J., Thomas, L., Ainslie, M.A., Winter, H.V., Hubert, J., Martin, B., Slabbekoorn, H., 2021. Effects of a seismic survey on movement of free-ranging Atlantic cod. Current Biology, 31(7):1555-62. https://doi.org/10.1016/j.cub.2021.01.050
Vinje, V., Lie, J. E., Danielsen, V., Dhelle, P. E., Silliqi, R., Nilsen, C., . . . Camerer, A. (2017). Shooting over the seismic spread. First Break, 35(6), 97-104.
Wang, X., Qi, B., Zhao, Q., Wang, W., & Yu, K. (2019). Feasibility of sparker source in marine seismic exploration: data analysis and processing. In Acta Geophysica (Vol. 67, Issue 5, pp. 1413–1418). Springer Science and Business Media LLC. https://doi.org/10.1007/s11600-019-00346-7
Wardle, C. S., Carter, T. J., Urquhart, G. G., Johnstone, A. D. F., Ziolkowski, A. M., Hampson, G., & Mackie, D. (2001). Effects of seismic air guns on marine fish. Continental Shelf Research, 21(8-10), 1005-1027. doi:10.1016/s0278-4343(00)00122-9
Weir, C.R. and Dolman, S.J. Comparative review of the regional marine mammal mitigation guidelines implemented during industrial seismic surveys and guidance towards a worldwide standard. Journal of International Wildlife Law and Policy, 10:1–27, 2007
Wensveen, P. J., Kvadsheim, P. H., Lam, F. P. A., von Benda-Beckmann, A. M., Sivle, L. D., Visser, F., . . . Miller, P. J. O. (2017). Lack of behavioural responses of humpback whales (Megaptera novaeangliae) indicate limited effectiveness of sonar mitigation. Journal of Experimental Biology, 220(22), 4150-4161. doi:10.1242/jeb.161232
Wale, M. A., Briers, R. A., & Diele, K. (2021). Marine invertebrate anthropogenic noise research – Trends in methods and future directions. In Marine Pollution Bulletin (Vol. 173, p. 112958). Elsevier BV. https://doi.org/10.1016/j.marpolbul.2021.112958
Wensveen P, Isojunno S, Hansen R, von Benda-Beckmann A, Kleivane L, van IJsselmuide S, Lam FP, Kvadsheim PH, DeRuiter S, Curé C, Narazaki T, Tyack P, Miller P (2019). Northern bottlenose whales in a pristine environment respond strongly to close and distant navy sonar signals. Proceedings of the Royal Society B 286:20182592.
Westerdahl, H. (2017). Lydutbredelse i havet. Presentert på konferansen «Fisk og Seismikk 2017». https://www.norskoljeoggass.no/miljo/prosjekter/fisk-og-seismikk/fisk-og-seismikk-2017/
Yang, Z., Wang, X., Hao, X., Qian, H., & Chen, X. (2022). Research on combined processing techniques of air gun and sparker source towed streamer seismic data. In Marine Geophysical Research (Vol. 43, Issue 2). Springer Science and Business Media LLC. https://doi.org/10.1007/s11001-022-09484-y
4 - Elektromagnetiske undersøkelser
4.1 - Hva er elektromagnetiske undersøkelser?
Elektromagnetiske undersøkelser (EMU) brukes, i likhet med seismiske undersøkelser, til å kartlegge petroleumsforekomster under havbunnen. En elektromagnetisk kilde taues vanligvis 30–50 m over bunnen eller 10 m under overflaten (Figur 2) med en hastighet på noen få meter per sekund, men kan også holdes stasjonært i ca. en time i bestemte posisjoner. Et rutenett av mottakere på havbunnen registrerer resistivitetsanomalier, som er en indikator for petroleumsforekomster (Figur 3). Mottakere er plassert på bunnen (1 m * 1 m * 0,15 m) med en avstand på 0,5–3 km. Frekvensen av det elektromagnetiske feltet er 0,05–10 Hz (Buchanan et al. 2006; EMGS 2011; Holten 2009; Mittet og Jensen 2018). Overflateundersøkelser kan gjennomføres i områder med dyp på maksimalt 500 m, mens man med bunnkilden kan utføre undersøkelser i områder med dyp ned til 3500 m. Vertikale undersøkelser utføres på dyp mellom 100 – 2100 m. (Buchanan et al. 2011, Ellingrud & Larsen 2019; Mittet 2016, Mittet & Jensen 2018).
4.2 - Kunnskapsgrunnlag om effekter av elektromagnetiske undersøkelser på marint liv
Mange marine dyr bruker elektriske og magnetiske signaler for å navigere, kommunisere, finne mat og unngå predatorer, men det mangler studier av hvordan elektromagnetiske undersøkelser kan påvirke disse dyrene. Det er imidlertid noen studier som har sett på hvordan organismer påvirkes av bestemte nivåer av elektriske eller magnetiske felt.
Effektene av elektromagnetiske undersøkelser på marint liv avhenger sannsynligvis av både feltstyrken, varighet av eksponering og dyrenes evne til å detektere slike felt. Som for seismikk kan effekter i teorien være både i form av skader og dødelighet eller adferdsendringer (Grimsbø et al. 2016).
Maksimalverdier for magnetiske og elektriske feltstyrker generert under elektromagnetiske undersøkelser er større enn magnetiske og elektriske felt som finnes naturlig i havet. De har også langt høyere nivåer enn det som har vært påvist å forårsake atferdsmessige og subtile fysiologiske effekter hos marine dyr (Nyqvist et al. 2020). Imidlertid dempes både elektriske og magnetiske felt raskt med avstand fra kilden. Dette medfører at høye nivåer kun finnes svært lokalt rundt kilden, og siden kilden som oftest konstant flyttes, blir stasjonære dyr eksponert bare i en kort periode.
4.2.1 - Naturlige elektriske og magnetisk felt og hvordan dyr bruker disse
Variasjon i magnetiske felt er vanlig i naturen, og jordklodens magnetfelt har en helling og en intensitet som begge varierer med geografisk plassering. Hellingen er 0° ved ekvator og 90° ved magnetpolene, mens intensiteten er rundt 60 000 nT ved polene, 40 000–50 000 nT ved midtbreddegrader og 30 000 nT ved ekvator. Dette resulterer i en gjennomsnittlig endring på 2–5 nT/km og 0,01°/km mellom ekvator og polene. Krystallstein og ikke-dipolkomponenter i jordens kjerne produserer også lokale anomalier og forårsaker gradienter på 10–100 nT/km (Kullnick 2000; Walker et al. 2003). I tillegg endrer det naturlige magnetiske feltet seg over tid, og historisk har det forekommet flere pol-reverseringer. I dag endres den totale feltintensiteten med en hastighet på 0–100 nT/år, avhengig av geografisk plassering.
Magnetfeltene varierer også med solstråling, og solstormer produserer med jevne mellomrom betydelig variasjoner (Klinowska 1986). I norske farvann vil dette variere fra noen få til hundrevis av mindre solgenererte magnetiske forstyrrelser (135–158 nT) årlig, med varighet på 0,5–2 timer. Slike forstyrrelser er for øvrig hyppigere og sterkere på høye breddegrader. Mer omfattende solstormer, i størrelsesorden > 1000 nT, med varighet over flere dager, inntreffer også en sjelden gang (5–10 ganger per 11-års-solar-syklus, Magnar Gullikstad Johnsen, UiT, personlig korrespondanse).
Naturlige elektriske felt blir indusert når saltvann beveger seg i det naturlige magnetiske feltet, og varierer med feltstyrke og strømhastighet. For eksempel måler elektriske felt i den engelske kanalen vanligvis 5–500 nV/cm (Kalmijn 1999). Fra Atlanterhavet, Golfstrømmen og Nordsjøen er tilsvarende elektriske feltstyrker på 350–500 nV/cm (Buchanan 2011). Under magnetstormer kan induktive elektriske felt nå styrker på 100 000 nV/cm (Kalmijn 1999). På tilsvarende måte blir også elektriske felt indusert når dyr, som også leder elektrisitet, svømmer i jordens magnetfelt (Kalmijn 1999).
En annen kilde til naturlige elektriske felt er levende organismer som genererer elektriske felt som følge av deres fysiologiske livsprosesser. Egenskapene til det genererte elektriske feltet varierer med art, posisjon og aktivitet og ligger typisk fra 2–100 μV/cm på svært nær avstand < 0,5 m (Haine et al. 2001). Noen fiskearter produserer også elektrisitet selv; for eksempel kan enkelte arter av skate produsere svake elektriske signaler for kommunikasjon, og elektriske rokker produserer relativt sterke elektriske felt som brukes under jakt (Bratton og Ayers 1987; Bray & Hixon 1978; Lowe et al. 1994).
Noen dyr, som bruker jordens magnetfelt for orientering, kalibrerer sitt interne kompass mot andre informasjons kilder (Cresci et al. 2019b, Goff et al. 1998, Muheim et al. 2006), dersom slik kalibrering forekommer relativt sjelden vil forstyrrelser i løpet av denne kalibreringstiden vil være spesielt uheldig. Migrerende sangfugler ( Chatarus sp. ) kalibrerer for eksempel sitt magnetiske kompass ved hjelp av solnedgangens retning eller tilhørende polariseringsmønster en gang om dagen (Cochran et al. 2004). Dyr kan også, som for eksempel tunfisk ( Thunnus maccoyii ), bruke magnetiske felt mer intensivt ved skumring og daggry, dette gir mindre forstyrrede «nattvinduer» for å få magnetisk informasjon med minimal påvirkning av naturlig magnetisk støy (Rodda 1984, Willis et al. 2009). Marine dyr som bruker slike kalibreringsvinduer, kan ende opp med å bevege seg i feil retning i en hel dag og bruke verdifull energi og tid, hvis de blir utsatt for et forvrengt magnetfelt i løpet av kalibreringen (Ferrari 2014, Vanselow et al. 2018). Dette vil forverre effekten av den elektromagnetiske forstyrrelsen utover eksponeringstidspunktet.
Noen marine dyr har utviklet særlige evner til å oppdage svake elektriske felt. Særlig kjent er bruskfisk (haier og skater), men også havmus, niøye, stjernetitterfisk, stør, maller og kvastfinnefisker er elektrofølsomme (Alves-Gomes 2001; Collin & Whitehead 2004; Walker 2001). I tillegg har laks og ål vist eksperimentelle responser på svake elektriske felt, og det er indisier på at rødspette potensielt orienterer seg i forhold til elektriske felt som genereres av havstrømmer (Metcalfe et al. 1993; Rommel Jr. & McCleave 1973a). Elektriske signaler har et bredere bruksområde for marine dyr enn magnetiske signaler, spesielt blant bruskfisk. Mens magnetiske signaler hovedsakelig brukes til orientering og i sosiale interaksjoner, brukes elektriske signaler til å oppdage bytte samt for å unngå predasjon, (Collin & Whitehead 2004). Elektriske felt har derfor potensial til å forstyrre et bredere adferdsspekter enn magnetiske felt.
Terskelverdier for elektriske felt i forhold til deteksjon, adferdsendring eller fysiologiske effekter hos marine dyr varierer mye, men bruskfisk er klart mest følsomme.
4.2.2 - Effekter av elektromagnetiske felt på marint liv
Mange forskjellige organismer kan oppfatte, bruke og reagere på magnetiske felt, fra bakterier (Blakemore & Frankel 1981) og protister (Bazylinski et al. 2000) til insekter, krepsdyr, fisk, havskilpadder, fugler og pattedyr (Wiltschko & Wiltschko 2005). Organismer kan oppfatte retning, intensitet og helling av det magnetiske feltet og kan bruke dette til å orientere seg eller navigere. Mange migrerende fisk orienterer seg ved hjelp av jordens magnetfelt.
Dyr som oppfatter og bruker naturlige endringer i elektriske og magnetiske felt, har også potensial for å bli påvirket av menneskeskapte endringer i disse, slik som under en elektromagnetisk undersøkelse, for eksempel ved at orienteringsevnen forstyrres. Imidlertid finnes det svært få studier på terskelverdier i forhold til deteksjon, adferdsendring eller fysiologiske effekter hos marine dyr.
Regnbueørrett som i embryostadiet blir utsatt for et spesifikt magnetfelt, statisk eller varierende, vil tiltrekkes av tilsvarende magnetfeltet også på larvestadiet (Jakubowska et al., 2021). A rter av ål og laks er mest undersøkt, men det er kjent at også tunfisk og larver av hyse og to korallfisker reagerer på endringer i magnetfelt (Bottesch et al. 2016; Cresci et al. 2019a; 2019b; 2020; Durif et al. 2013; Nishi et al. 2004; O'Connor and Muheim 2017; Putman et al. 2013; Walker et al. 2003; Walker 1984). Det er rapportert om sammenhenger mellom fangster av sild og nivå av geomagnetisk aktivitet. Silda migrerte fra grunne områder i Barentshavet til dypere vann i Norskehavet under større magnetiske stormer (Krylov et al. 2014). En annen studie har vist at bruskfiskarten liten skate ( Leucoraja erinacea ) økte svømmehastigheten sin og bevegde seg over et større område når den ble utsatt for elektromagnetiske felt fra en undervannskabel (Hutchison et al. 2018). Selv ved svært lave nivåer, 2–20 nT, ble amfipoder desorienterte (Tomanova & Vacha 2016).
Det er sannsynlig at mange dyr kan tilpasse seg variasjoner i magnetfeltet, enten ved å midlertidig orientere seg på andre måter eller ved å stanse aktiviteten sin (Freake et al. 2006; Rodda 1984). For eksempel er det blitt observert at ål som passerer et unaturlig magnetfelt grunnet en undervannskabel, endret svømmeretning, noe som resulterte i en mindre migrasjonsforsinkelse. Ålen fortsatte senere sin opprinnelige migrasjon (Öhman et al. 2007). Imidlertid kan selv små avvik i magnetfelt få fatale følger, som hvalstrandinger som har blitt assosiert med små avvikelser av magnetfeltet, som sannsynligvis forvirret hvalens navigasjon (Kirschvink et al. 1986). Påvirkninger av antropogent indusert magnetfeltet vil sannsynligvis også forekomme på bentiske krepsdyr, som reagerer på endringer av magnetfeltet i det geomagnetiske området og sterkere felt. Dette ble for eksempel rapportert hos en hummer art ( Panulirus argus) ( Boles and Lohmann, 2003; Ernst and Lohmann, 2016), men man fant ingen effekt på Europeisk hummer ( Homarus gammarus) (Taormina et al., 2020).
Bruskfisk og ål kan oppfatte elektriske signaler på over en kilometers avstand (Buchanan 2011; Peters et al. 2007). I teorien kan et oppfattet elektrisk felt midlertidig forstyrre beiting, orientering eller sosiale interaksjoner. For spadestør ( Polydon spathula ), en ferskvannsfisk som jakter plankton med hjelp av et elektriskeføleevne, resulterte menneskeskapt elektrisk støy i redusert predasjonsavstand ved 1000 nV/cm og umuliggjorde i prinsipp predasjon ved 50 000 nV/cm. I tillegg kan et elektrisk signal, avhengig av egenskaper og kontekst, påvirke fiskeadferden selv om det er svært svakt (Grimsbø et al. 2014; Kalmijn 1999).
Det er også en mulighet at de plutselige forandringene i elektriske (eller kanskje også magnetiske felt) under en EMU kan forårsake fluktresponser, stress eller adferdsendringer utover selve varigheten av eksponeringen, på en lignende måte som seismiske undersøkelser (Engås et al. 1996). For eksempel, selv om haier ikke frastøtes av den genererte elektriske feltstyrke, er det for eksempel ikke gitt at et gradvis økende elektrisk felt som beveger seg mot dem, ikke kan påvirke adferden deres. Et fluktuerende, bevegelig elektrisk felt fra en elektromagnetisk undersøkelse kan eksempelvis oppfattes som mer skremmende enn de relativt stabile elektriske feltene som genereres av havstrømmer.
Effekten fra elektromagnetiske felt tilsvarende styrken brukt i EMU på avstander fra 50 til 1000 m er de siste årene studert i en spesialkonstruert tank for ulike organismer. Disse viser at hyselarver reduserte svømmehastigheten sin med 30% under eksponering og reduksjonen vedvarte i minimum 5 timer etter en 15 minutters eksponering (Durif et al. 2022). Tilsvarende ble dokumentert for hyselarver eksponert til elektromagnetiske felt som genereres av undervannskabler (Cresci et al. 2022a). En annen studie i samme forsøksoppsett hvor hyseegg ble eksponert for feltstyrker fra elektromagnetiske undersøkelser tilsvarende avstander på 30 til 1000 m i 15 minutter viste at larvene økte hjertefrekvens samt brukte opp plommesekken mye raskere enn kontrollgruppen som ikke ble eksponert (Guillebon 2022).
Dette forsøksoppsettet er vanskelig å bruke på voksen fisk, men et pilotstudie med havsil (tobis) viste at disse ikke endret adferden sin under eller etter eksponering (Durif et al. 2022; Cresci et al. 2022b).
4.3 - Konsekvensvurdering av effekter av elektromagnetiske undersøkelser på marint liv
En omfattende miljøeffektrapport om elektromagnetiske undersøkelser konkluderer med at det er rimelig å anta at elektromagnetiske undersøkelser kan oppfattes av marine dyr og potensielt kan påvirke adferden deres. Rapporten konkluderer videre med at eventuelle effekter vil bli marginale i tid og rom og påvirke få individer i en populasjon (Buchanan 2011). Det er imidlertid store kunnskapshull på feltet, og konklusjonen i denne er basert på at effekten av påvirkningen opphører samtidig som påvirkningen opphører, noe som ikke nødvendigvis er tilfelle, da for eksempel hyselarver viste en vedvarende reduksjon i svømmehastighet i minst 5 timer etter endt eksponering til en EMU. Da de elektriske og magnetiske feltene begge dempes raskt, vil potensiell påvirkning være konsentrert til selve undersøkelsesområdet.. For de dyrene som befinner seg nettopp her, kan de imidlertid være av betydning. Konsekvensen av slike forstyrrelser vil sannsynligvis også avhenge av hvor ofte slike undersøkelser repeteres i samme område.
Fra tilgjengelige data synes bruskfisk å ha størst risiko for å bli påvirket av elektromagnetiske undersøkelser. I Norge finnes det 8 haiarter og 13 arter av skater. Pigghå er tradisjonelt den viktigste bruskfisken i fiskeriet, og den er listet som en truet art i Nordøst-Atlanteren. Selv om fiske etter pigghå er forbudt, tas den fortsatt som bifangst. Pigghå samles ofte i store aggregeringer, noe som gjør dem sårbare for lokale forstyrrelser dersom de overlapper med en slik aggregering. Eventuelle forstyrrelser fra en EMU vil da kunne påvirke en stor andel av bestanden. Gyteområdene og yngleplassene er lite kjent (Bakketeig et al. 2017). Mulige effekter av en EMU på pigghå er derfor særlig bekymringsverdig, men grunnet den lave kunnskapen om hvor og når aggregeringer er, er det vanskelig å fraråde spesifikke områder og perioder for å beskytte disse. Andre arter kan være fisk som sannsynligvis bruker magnetiske og elektriske felt til orientering under migrasjon; som for eksempel ål, laks eller hyse, eller andre fiskearter som bruker magnetiske felt til å orientere seg for eksempel mot gytefelt eller oppvekst/beiteområde. Sammenhenger mellom sildefangster og elektromagnetiske forstyrrelser, samt larvers orientering etter magnetfelt, viser at potensielt mange organismer kan påvirkes.
Inn- og utvandringsruter for ål og laks og parrings- og yngleplasser for pigghå og andre bruskfiskarter bør unngås i de periodene hvor dette foregår. Imidlertid er det stor usikkerhet både med hensyn til hvor innvandringsrutene for ål og laks er, når innvandringen finner sted, og hvor paringsområdene for pigghå og andre bruskfiskarter befinner seg. Det er derfor per i dag ikke mulig å framskaffe kart eller tidfeste aktiviteten særlig presist.
Siden forstyrrelser i elektriske, og kanskje særlig magnetiske, felt kan påvirke orienterings-evnen til ulike dyr, bør også viktige vandringsruter skjermes. Konsentrerte vandringer mot gytefeltene er en slik vandring som kan gi konsekvenser på bestandsnivå dersom den forstyrres på en slik måte at gytingen forringes. Om fisk ikke ankommer på gyteplassen, eller om ankomsten blir betydelig forsinket, kan dette få konsekvenser for gyteaktiviteten og rekrutteringen, se utdypende forklaring i avsnitt 3.4.1. Slike vandringsruter er godt kjent og kartfestet for artene torsk, sild og lodde.
Ny kunnskap viser at både egg og larver av hyse kan påvirkes betraktelig. Eggene påvirkes ved at hjerterytmen øker, og at plommesekken, som skal fungere som energireserve inntil larven klarer å spise selv, minker (Guillebon 2022). I tillegg viser studiene at større larver viser en betydelig og vedvarende reduksjon i svømmehastighet (Cresci et al. 2022b; Durif et al. 2022). I kombinasjon kan disse effektene være svært fatale; dersom plommesekken brukes opp før larven er i stand til å finne føde, eller er i et område med god fødetilgang vil den ikke overleve. Og dersom svømmehastigheten mot fødeområdet reduseres vil ankomsten skje senere. Det er derfor en betydelig økt risiko for at eksponerte egg og larver ikke vil overleve til voksen fisk. Imidlertid er den naturlige dødeligheten på disse livsstadiene svært høy, og effekten vil måtte ramme en relativt stor andel av egg, larver og yngel for å gi en effekt på populasjonsnivå (se avsnitt 3.4.1 om seismikk og effekter på fiskeyngel). Videre er det usikkert om de påviste effektene også gjelder andre fiskearter, eller om hyse er spesielt sensitiv. Det er planlagt både å gjøre en sensitivitets-analyse for å undersøke muligheten for populasjonseffekt, samt studier med andre arter for en bedre forståelse av omfanget. I påvente av denne kunnskapen beholdes derfor samme råd som for tidligere år, dvs. å fraråde konsentrerte vandringsruter, men dersom det er tilkommer ytterligere kunnskap som støtter oppunder effekter på tidlige livsstadier samt mulighet for påvirkning av en betydelig andel av populasjonen, er det mulig at rådet endres til å fraråde undersøkelser i områder og perioder med høye konsentrasjoner av egg og larver.
4.4 - Råd
Den viktigste forstyrrelsen av elektromagnetiske felt er sannsynligvis på fiskens orienteringsevne, med mulighet til å forstyrre vandringer. Grunnet mangel på kunnskap om høytetthetsområder for bruskfisk er disse ikke inkludert i noen kartfestet rådgivningskart til tross for at det kan antas å ha en påvirkning på disse. Det samme gjelder innvandringsruter for laks og ål. Rådet for elektromagnetiske undersøkelser er derfor følgende:
- Havforskningsinstituttet fraråder elektromagnetiske undersøkelser i områder og perioder med konsentrerte gytevandringer for fiskeartene torsk, lodde og sild. Kart for dette finnes i Vedlegg I.
4.5 - Referanser
Alves-Gomes, J. A. 2001. The evolution of electroreception and bioelectrogenesis in teleost fish: a phylogenetic perspective. Journal of Fish Biology, 58: 1489-1511.
Bakketeig, I., Hauge, M., and Kvamme, C. 2017. Havforskningsrapporten.
Bazylinski, D. A., Schlezinger, D. R., Howes, B. H., Frankel, R. B., and Epstein, S. S. 2000. Occurrence and distribution of diverse populations of magnetic protists in a chemically stratified coastal salt pond. Chemical Geology, 169: 319-328.
Blakemore, R., and Frankel, R. 1981. Magnetic Navigation in Bacteria. Scientific American, 245: 58-65.
Boles, L.C., Lohmann, K.J., 2003. True navigation and magnetic maps in spiny lobsters. Nature 421, 60–63. https://doi.org/10.1038/nature01226.
Bottesch, M., Gerlach, G., Halbach, M., Bally, A., Kingsford, M. J., and Mouritsen, H. 2016. A magnetic compass that might help coral reef fish larvae return to their natal reef. Current Biology, 26: R1266-R1267.
Bratton, B. O., and Ayers, J. L. 1987. Observations on the electric organ discharge of two skate species (Chondrichthyes: Rajidae) and its relationsHip to behaviour. Environmental Biology of Fishes, 20: 241-254.
Bray, R. N., and Hixon, M. A. 1978. Night-Shocker: Predatory Behavior of the Pacific Electric Ray. Science, 200: 333-334.
Buchanan, R. A., Fechhelm, R., Christian, J., Moulton, V. D., Mactavish, B., Canning, S., and Pitt, R. 2006. Orphan Basin Controlled Source Electromagnetic Survey Program Environmental Assessment. LGL Rep. SA899. 137 pp.
Buchanan, R., Fechhelm, R., Abgrall, P., & Lang, A. (2011). Environmental Impact Assessment of Electromagnetic Techniques Used for Oil & Gas Exploration & Production. LGL Rep. SA1084. by LGL Limited, St. John's, NL. In: Canada for International Association of Geophysical Contractors, Houston
Cochran, W. W., Mouritsen, H., and Wikelski, M. 2004. Migrating Songbirds Recalibrate Their Magnetic Compass Daily from Twilight Cues. Science, 304: 405-408.
Collin, S., and WHitehead, D. 2004. The functional roles of passive electroreception in non-electric fishes. Animal Biology, 54.
Cresci, A., Allan, B. J. M., Shema, S. D., Skiftesvik, A. B., and Browman, H. I. 2020. Orientation behavior and swimming speed of Atlantic herring larvae (Clupea harengus) in situ and in laboratory exposures to rotated artificial magnetic fields. Journal of Experimental Marine Biology and Ecology, 526: 151358.
Cresci, A., Durif, C. M., Paris, C. B., Shema, S. D., Skiftesvik, A. B., and Browman, H. I. 2019a. Glass eels (Anguilla anguilla) imprint the magnetic direction of tidal currents from their juvenile estuaries. Communications Biology, 2: 366.
Cresci, A., Paris, C. B., Foretich, M. A., Durif, C. M., Shema, S. D., O’Brien, C. E., Vikebø, F. B., et al. 2019b. Atlantic Haddock (Melanogrammus aeglefinus) Larvae Have a Magnetic Compass that Guides Their Orientation. iScience, 19: 1173-1178.
Cresci, A., Durif, C. M. F., Larsen, T., Bjelland, R., Skiftesvik, A. B., & Browman, H. I. (2022b). Magnetic fields produced by subsea high voltage DC cables reduce swimming activity of haddock larvae (Melanogrammus aeglefinus). PNAS Nexus. by subsea high voltage DC cables reduce swimming activity of haddock larvae (Melanogrammus aeglefinus). PNAS Nexus. https://doi.org/10.1093/pnasnexus/pgac175
Cresci, A., Perrichon, P., Durif, C. M. F., Sørhus, E., Johnsen, E., Bjelland, R., Larsen, T., Skiftesvik, A. B., & Browman, H. I. (2022). Magnetic fields generated by the DC cables of offshore wind farms have no effect on spatial distribution or swimming behavior of lesser sandeel larvae (Ammodytes marinus). Marine Environmental Research, 176, 105609. https://doi.org/https://doi.org/10.1016/j.marenvres.2022.105609
Durif, C. M. F., Browman, H. I., PHillips, J. B., Skiftesvik, A. B., Vøllestad, L. A., and Stockhausen, H. H. 2013. Magnetic Compass Orientation in the European Eel. PLOS ONE, 8: e59212.
Durif, C., Zhang, G., Cresci, A., Skiftesvik, A. B., Browman, H. I., Sivle, L. D., Nyqvist, D., Jensen, H. R., & Solheim, J. G. (2022). Effects of the electromagnetic field used in hydrocarbon surveys on marine organisms. Rapport fra havforskningen,2022-23, ISSN 1893-4536
Ernst, D.A., Lohmann, K.J., 2016. Effect of magnetic pulses on Caribbean spiny lobsters: implications for magnetoreception. J. Exp. Biol. jeb.136036-. https://doi.org/10.1242/jeb.136036.
Ferrari, T. E. 2014. Magnets, magnetic field fluctuations and geomagnetic disturbances impair the homing ability of honey bees (Apis mellifera). Journal of Apicultural Research, 53: 452-465.
Freake, Michael J., Muheim, R., and PHillips, John B. 2006. Magnetic Maps in Animals: A Theory Comes of Age? The Quarterly Review of Biology, 81: 327-347.
Goff, M., Salmon, M., and Lohmann, K. J. 1998. Hatchling sea turtles use surface waves to establish a magnetic compass direction. Animal Behaviour, 55: 69-77.
Grimsbø, E., Nortvedt, R., Hammer, E., and Roth, B. 2014. Preventing injuries and recovery for electrically stunned Atlantic salmon (Salmo salar) using High frequency spectrum combined with a thermal shock. Aquaculture, 434: 277-281.
Grimsbø, E., Nortvedt, R., Hjertaker, B. T., Hammer, E., and Roth, B. 2016. Optimal AC frequency range for electro-stunning of Atlantic salmon (Salmo salar). Aquaculture, 451: 283-288.
Guillebon, C. 2022. Effects of electromagnetic field on Atlantic haddock (Melanogrammus aeglefinus) early life stages. Sciences, Sorbonne Université Master 2022, 20 pp.
Holten, T., Flekkøy, E., Singer, B., Blixt, E., Hanssen, A., and Måløy, K. 2009. Vertical source, vertical receiver, electromagnetic technique for offshore hydrocarbon exploration. First Break, 27: 89-93.
Hutchison, Z., Sigray, P., He, H., Gill, A. B., King, J., and Gibson, C. 2018. Electromagnetic Field (EMF) Impacts on Elasmobranch (shark, rays, and skates) and American Lobster Movement and Migration from Direct Current Cables.
Jakubowska, M., Greszkiewicz, M., Fey, D.P., Otremba, Z., Urban-Malinga, B., Andrulewicz, E., Jakubowska, M., Greszkiewicz, M., Fey, D.P., Otremba, Z., Urban-Malinga, B., Andrulewicz, E., 2021. Effects of magnetic fields related to submarine power cables on the behaviour of larval rainbow trout (Oncorhynchus mykiss). Mar. Freshw. Res. 72, 1196–1207. https://doi.org/10.1071/MF20236
Kirschvink, J. L., Dizon, A. E., and Westphal, J. A. 1986. Evidence from Strandings for Geomagnetic Sensitivity in Cetaceans. Journal of Experimental Biology, 120: 1.
Krylov, V. V., Izyumov, Y. G., Izvekov, E. I., and NepomnyashcHikh, V. A. 2014. Magnetic fields and fish behavior. Biology Bulletin Reviews, 4: 222-231.
Kullnick, U. 2000. Influences of Electric and Magnetic Fields on Aquatic Ecosystems. In ICNIRP: Effects of Electromagnetic Fields on the Living Environment, pp. 113-132.
Lowe, C. G., Bray, R. N., and Nelson, D. R. 1994. Feeding and associated electrical behavior of the Pacific electric ray Torpedo californica in the field. Marine Biology, 120: 161-169.
Metcalfe, J., Schwartz, B., and Joaquim, S. 1993. The Cue-Familiarity Heuristic in Metacognition. Journal of experimental psychology. Learning, memory, and cognition, 19: 851-861.
Mittet, R. 2016. Presentation på faglig forum: Marine CSEM-electric and magnetic Field amplitudes.
Mittet, R., and Jensen, H. R. 2018. Marine CSEM - Electric and Magnetic Field Amplitudes.
Muheim, R., Moore, F. R., and PHillips, J. B. 2006. Calibration of magnetic and celestial compass cues in migratory birds - a review of cue-conflict experiments. Journal of Experimental Biology, 209: 2.
NisHi, T., Kawamura, G., and Matsumoto, K. 2004. Magnetic sense in the Japanese eel, <em>Anguilla japonica</em>, as determined by conditioning and electrocardiography. Journal of Experimental Biology, 207: 2965.
Nyqvist, D., Durif, C., Johnsen, M. G., De Jong, K., Forland, T. N., and Sivle, L. D. 2020. Electric and magnetic senses in marine animals, and potential behavioral effects of electromagnetic surveys. Marine Environmental Research, 155: 104888.
O`Connor, J., and Muheim, R. 2017. Pre-settlement coral-reef fish larvae respond to magnetic field changes during the day. The Journal of Experimental Biology, 220: 2874.
Peters, R. C., Eeuwes, L. B. M., and Bretschneider, F. 2007. On the electrodetection threshold of aquatic vertebrates with ampullary or mucous gland electroreceptor organs. Biological Reviews, 82: 361-373.
Putman, Nathan F., Lohmann, Kenneth J., Putman, Emily M., Quinn, Thomas P., Klimley, A. P., and Noakes, David L. G. 2013. Evidence for Geomagnetic Imprinting as a Homing Mechanism in Pacific Salmon. Current Biology, 23: 312-316.
Rodda, G. H. 1984. The orientation and navigation of juvenile alligators: evidence of magnetic sensitivity. Journal of Comparative Physiology A, 154: 649-658.
Rommel, S. A., Jr., and McCleave, J. D. 1973. Prediction of oceanic electric fields in relation to fish migration. ICES Journal of Marine Science, 35: 27-31.
Tomanova, K., and Vacha, M. 2016. The magnetic orientation of the Antarctic ampHipod Gondogeneia antarctica is cancelled by very weak radiofrequency fields. J Exp Biol, 219: 1717-1724.
Taormina, B., Poi, C. Di, Agnalt, A.-L., Carlier, A., Desroy, N., Helena Escobar-Lux, R., D’eu G, J.-F., Freytet, F., Durif, C.M.F., 2020. Impact of magnetic fields generated by AC/DC submarine power cables on the behavior of juvenile European lobster (Homarus gammarus). Aquat. Toxicol. 220, 105401. https://doi.org/10.1016/j.aquatox.2019.105401
Vanselow, K. H., Jacobsen, S., Hall, C., and Garthe, S. 2018. Solar storms may trigger sperm whale strandings: explanation approaches for multiple strandings in the North Sea in 2016. International Journal of Astrobiology, 17: 336-344.
Walker, M. M. 1984. Learned magnetic field discrimination in yellowfin tuna,Thunnus albacares. Journal of Comparative Physiology A, 155: 673-679.
Walker, M. M., Diebel, C. E., and Kirschvink, J. L. 2003. Detection and Use of the Earth’s Magnetic Field by Aquatic Vertebrates. In Sensory Processing in Aquatic Environments, pp. 53-74. Ed. by S. P. Collin, and N. J. Marshall. Springer New York, New York, NY.
Walker, T. 2001. Basslink project review of impacts of High voltage direct current sea cables and electrodes on chondrichthyan fauna and other marine life, Basslink Supporting Study No 29, Marine and Freshwater Resources Institute No 20, 68 pp.
Willis, J., PHillips, J., Muheim, R., Diego-Rasilla, F. J., and Hobday, A. J. 2009. Spike dives of juvenile southern bluefin tuna (Thunnus maccoyii): a navigational role? Behavioral Ecology and Sociobiology, 64: 57.
Wiltschko, W., and Wiltschko, R. 2005. Magnetic orientation and magnetoreception in birds and other animals. Journal of Comparative Physiology A, 191: 675-693.
Öhman, M., Sigray, P., and Westerberg, H. 2007. Offshore windmills and the effects of electromagnetic fields on fish. AMBIO A Journal of the Human Environment, 36: 630-633.
5 - Bruk av eksplosiver i sjø
5.1 - Bakgrunn
Havforskningsinstituttets rådgivning med hensyn til sprengningsarbeider har som hensikt å gi råd der målet er å begrense eller hindre skade på marint liv. Det er lite sannsynlig at lokale sprengninger i sjø har potensial til å påvirke fisk på populasjons- eller bestandsnivå (Govoni et al. 2008), og derfor peiler rådgivningen seg inn mot områder som lokale fiskebestander, gytefelt, akvakulturanlegg og annet marint liv av lokal karakter. Hvordan sprengninger kan påvirke marint liv, har vært gjennomgått i en rekke arbeider (Aune et al. 2018; Dalen 2009; Eliassen og Iversen 2004; Engås et al. 1989; Fauske 2007; Grimsbø og Kvadsheim 2018; Johnsen et al. 1994; Kjellsby 1993; Kjellsby og Kvalsvik 1997; Larsen et al. 1993; Soldal 1990; Trettenes 2006 , Dahl et al. 2020).
Rådgivningen denne rapporten omhandler gjelder i hovedsak sivile sprengningsarbeider, men vil også kunne gjelde demolering eller sprenging av militære etterlatenskaper i sjø. Havforskningsinstituttet er forøvrig involvert i det Europeiske programmet JPI Oceans ( http://www.jpi-oceans.eu/munitions-sea ) der man blant annet forsøker å finne løsninger på militære etterlatenskaper i sjø (Campana et al. 2016). Aktuelle sivile sprengningsarbeider Havforskningsinstituttet gir råd i forhold til kan være utbedring av skipsleder og andre arbeider der hele eller deler av sprengingen foregår i sjø eller har utslag i sjø. Et annet område det gis råd i forhold til er refraksjonsseismikk (Engås et al. 1989), der det brukes små sprengladninger for å frembringe et signal som brukes videre i grunnanalyser, ikke ulikt vanlig seismikk.
5.2 - Kunnskapsgrunnlag om effekter av eksplosiver på marint liv
5.2.1 - Direkte skade og dødelighet
I motsetning til en vanlig forbrenning, også kalt deflagrasjon, vil en detonasjon oppstå når forbrenningen er en sjokkbølge som forplanter seg med overlydsfart (raskere enn lydhastigheten til eksplosivet) i eksplosivet (Kiran 2016). Hastigheten sjokkbølgen forplanter seg i sprengstoffet med, kalles detonasjonshastighet og vil variere for de ulike sprengstoff-typene. Når eksplosjonen skjer, foregår det kjemiske reaksjoner som avgir mye energi og gass. Gassen som dannes, fortrenger mediet rundt så hurtig at det skaper en sjokkbølge. En sjokkbølge er en trykkbølge som gir en veldig brå endring i trykk, temperatur og tetthet i mediet, og som forplanter seg raskere enn lydhastigheten i mediet (Kjellsby 1993). Militære eksplosiver har generelt høyere detonasjonshastighet enn eksplosiver til sivilt bruk, for eksempel slike som blir brukt ved bergsprengning. For bergsprengning vil forøvrig detonasjonshastigheten kunne påvirkes av borehullsdiameter (Amundsen 1984; Higgins et al.1979; Petel et al. 2007) og trykk. I tillegg til selve trykkpulsen fra detonasjonen vil det være en betydelig gassutvikling, som også har en sprengningsvirkning. Eksplosivet sin sprengvirkning blir, for sivile sprengninger, ofte relatert til ANFO (Amonium nitrate fuel oil) sin sprengvirkning med et forholdstall. For militære eksplosiver blir gjerne TNT (Trinitrotoluen), som har en helt annen karakteristikk enn ANFO, brukt som referanse.
Ved bergsprengning er det trykkpulsen som først knuser berget (Fauske 2007), videre er det gassutviklingen som ytterligere river berget fra hverandre. Selv om mye av energien fra en sprengning går med til å knuse og rive berget fra hverandre, vil det i vann oppstå en kraftig trykkpuls samt påfølgende boblepuls (Cole 1948), se Figur 3.
Amplituden til trykkpulsen, ved tiden t1 i Figur 3, er bestemt av salve/landingsstørrelse og sprengstoffkarakteristikk samt av borehullets diameter (Amundsen 1984; Higgins et al. 1979; Petel et al. 2007). Det er trykkpulsene fra sprengningen som først og fremst forårsaker direkte skade på marint liv. Det er særlig luft- eller gassfylte organer, som svømmeblære og lunger som har høy risiko for å bli skadet (Gaspin 1975; Gaspin et al. 1976; Yelverton et al. 1975).Dette skjer ved at slike gasslommer komprimeres og ekspanderer og videre ved at vev strekkes og avrivninger oppstår med påfølgende blødninger (Ketten 1995; Yelverton et al. 1973). Trykkpulser med rask stigetid kan være mer skadelige enn pulser med høyere maksimalverdi, men lengre stigetid (Simenstad 1973).
Ved sprengningsarbeider i sjø kan man ofte observere en del skadet og død fisk flytende i overflaten nær sprengningsstedet; dette er fisk med svømmeblære. Skadet fisk uten svømmeblære vil derimot normalt synke og dermed ikke være synlig på overflaten, dette gjelder også fisk med svømmeblære der gassen unnslipper (Teleki & Chamberlain 1978). Trykkpulsen fra sprengingen vil også kunne slå i svime fisk (Grimsbø 2016; Lamboij et al. 2010) og naturlige reflekser vil da opphøre (Kestin et al. 2002). Det er også dokumentert at sprengningsarbeider har negativ innvirkning på yngel og egg (Faulkner et al. 2006), men modellering viser at dette gelder for korte avstander (McQueen et al. 2022) Sprengningsarbeider vil kunne ha et skadepotensiale knyttet til gyting og tidlige livsstadier (Faulkner et al. 2008; Krise 2001). Mulig skadepotensial kan også være knyttet til rystelser og vibrasjoner fra sprenginger som forplanter seg i grunnen (Dalen 2009: Toresen & Søvik 2012), selv om sammenhengene er noe uklare.
Det observeres ofte vann i svømmeblæren på fisk som har vært utsatt for sprengning (Soldal 1990); dette gjelder hovedsakelig arter med åpen svømmeblære, slik som laks. Dette antas å ha en sammenheng med at trykkpulsen fra sprengningen speiles i vannets overflate og skifter fase, samt genererer et kavitasjonsfelt (Fardal 2005; Gaspin & Price 1972). Når det befinner seg fisk der det oppstår undertrykk eller kavitasjon vil vann suges inn i den åpne svømmeblæren. En annen mulig årsak kan være boblepulsen i forbindelse med boblens kollaps. Vann som kommer inn i fiskens svømmeblære, fører til nedsatt almenntilstand og mulig økt dødelighet noe tid etter sprengningen er gjennomført. Det er ikke alltid sånn at faren for skader minker proporsjonalt med avstand. Et eksperiment med sardiner (Sardina pilcharus) ved ulike avstander (18-246 m) fra en detonasjon (4.66 kg TNT) viste at sardinene fikk skader i svømmeblære og nyrer. Skadene avtok som funksjon av avstand for de nærmeste 50 m, men etter det var det mer usystematisk fordeling av skader med avstand. Dette kan skyldes interferens med refleksjoner fra bunn (Dahl et al. 2020). Forsøket ble gjentatt med en annen art, stillehavsmakrell (Scomber japonicus). Skader på nyrer og svømmeblære og skader på hørselen ble funnet, men denne gangen var reduksjonen i skader ved økende avstand mer som forventet (Jenkins et al. 2022, Smith et al. 2022)
I tillegg til fisk kan dykkende fugl og sjøpattedyr som befinner seg i nærområdet til sprengningsarbeidet, skades. For sjøpattedyr vil det i hovedsak være snakk om hørselsskader (von Benda-Beckmann et al. 2015).
5.2.2 - Adferdseffekter
Når avstanden til en sprengning er stor, vil trykkpulsene som blir generert, ikke være et signifikant problem med tanke på direkte fysisk skade på marint liv, men de vil bidra til økt støynivå i flere kilometers radius (Kjellsby & Kvalsvik 1997; Koschinski 2011). Slik støy vil i likhet med seismikk inneholde lave frekvenser som er hørbare for fisk og adferdseffekter som dem beskrevet i kapittel 3.
5.2.3 - Effekter på fisk i oppdrettsanlegg
or Sprengningsarbeid i sjø finner ofte sted i kystnære områder hvor det kan finnes oppdrettsanlegg i nærheten. For fisk i oppdrettsanlegg, som ikke har mulighet til å svømme unna, kan både skader og adferdseffekter inntreffe.
I forbindelse med forsvarets ønske om etablering av permanent øvingsområde i Korsfjorden ble det gjennomført sprengningsforsøk med detonasjon av ladinger på 0.1- 2 kg TNT under vann. Detonasjonene ble gjort med og uten boblegardin (enkel og dobbel). Lydtrykk og fiskevelferd i nærliggende kommersielle laksemerder som lå i avstander 3.7 - 6 km avstand fra detonasjonene ble overvåket (Kvadsheim et al., 2022). Forsøkene viste en redusering av bredbånds spisstrykk med 12-20 dB re 1µPa og lydeksponeringsnivå (SEL) med 5-12 dB re 1 µPa 2 s når boblegardin ble benyttet. I disse forsøkene observerte man ingen signifikant forskjell i spisstrykk eller lydeksponeringsnivå ved bruk av enkel eller dobbel boblegardin. Fiskeadferd i merdene ble observert med ekkolodd og kamera før, under og etter detonasjon. Det ble observert kortvarig respons hos fisken ved de høyeste lydnivåene, >163dB re 1µPa spisstrykk i fiskens høreområde 20-500 Hz. Dette tilsvarer SEL nivåer på >147 dB re 1 µPa2·s, men denne responsen skiller seg ikke ut fra adferdsrespons som oppleves daglig i merdene grunnet andre årsaker. Det er verdt å merke seg at i fisken som ble tatt ut fra merden på anlegget nærmest detonasjonsstedet umiddelbart etter siste skudd, ble det funnet sirkulasjonsforstyrrelser og blødninger som er konsistent med barotraumer. I samme fisken så man samtidig forhøyede nivåer av stresshormonet kortisol i plasma, og kortisol- metabolitter i faeces. Dette kan skyldes håndtering av fisken, men man kan ikke utelukke subkliniske skader som følge av sprenginger. Derimot ble ikke observert signifikante endringer i forforbruk eller dødelighet på anlegget, og dermed har ikke sprengingene påvirket helsen generelt i populasjonen.
Selv om fisken ikke utsettes for direkte skade, kan den lavfrekvente støyen medføre økt stress. Langvarig stress som fisken ikke greier å tilpasse seg til kan medføre større dødelighet som følge av den ekstrabelastningen en sprengning representerer (Grimsbø & Kvadshein 2018; Iwama & Afonso 2006), og fisk som i utgangspunktet har redusert allmenntilstand for eksempel som følge av sykdom, er ekstra sårbare.
5.3 - Konsekvensvurdering av effekter ved bruk av eksplosiver i sjø og avbøtende tiltak for å redusere disse
Det er ofte vanskelig å si noe konkret på forhånd om hvor stor trykkbølgen fra en eksplosjon vil bli (Kjellsby & Kvalsvik 1997). Dermed er det vanskelig å predikere innenfor hvilke avstander det kan forventes skade i forbindelse med sprengningsarbeider – og følgelig er det ofte vanskelig å beregne på hvilke avstander det er nødvendig med tiltak. Det er heller ikke enkelt å sette noen generelle grenseverdier når det gjelder bergsprenging som gjelder under alle forhold, men i noen tilfeller er dette forsøkt (Dalen 2012; Hveding 2018). Kvadsheim et al. (2017) har oppsummert kunnskapsstatus og det fremkommer at faren for skade øker ved spisstrykksnivåer over 210 dB for fisk og 202 dB for sjøpattedyr. Det opereres for øvrig med en sikkerhetsavstand på 2 nmi til havbruksanlegg i forhold til eksplosivrydding under vann i Sjøforsvarets reglement (Svendsen 2012). Det finner også eksempler fra andre land på mer konkrete retningslinjer for sprengningsarbeider slik som Wrigth & Hopky (1998) og Department of Arts, Heritage and the Gaeltacht (2014).
Et tiltak som er mye brukt, er å sette av en fenghette eller en mindre ladning før hovedsalven for å skremme bort fisk og sjøpattedyr i nærområdet. Dette tilsvarer ramp-up for seismikk, men har ikke blitt dokumentert å ha noe entydig effekt i forbindelse med sprenging. Et slikt tiltak reduserer heller ikke de skadelige effektene fra en sprengning og er derfor ikke i seg selv tilstrekkelig for å unngå skade, selv om det antas å ha en gunstig effekt.
Ved å redusere størrelsen på salven/ladningen vil den skadelige trykkpulsens maksimaltrykk bli redusert. Dette kan oppnås ved å dele en salve opp i mindre delladninger som avfyres sekvensielt, selv om den totale ladningsstørrelsen fremdeles vil ha betydning for skadepotensialet. Man bør da etterstrebe en minst mulig ladningsstørrelse på hver delladning og gjerne ha ca. 20 millisekunds forsinkelse mellom hver ladning. Utover å redusere maksimaltrykket er hensikten med tidsforsinkelsen å plassere den ene delladningens trykkpuls t1 i foregående delladnings trykkminimum, altså mellom t1 og t2 til foregående ladning. Dermed vil delladningens trykkpuls helt eller delvis kunne bli kansellert av foregående delladnings undertrykk eller speilede trykkpuls.
Trykket t1 som oppstår som følge av detonasjonen, vil være avhengig ikke bare av sprengstoffmengden, men også av detonasjonshastigheten. Valg av riktig sprengstofftype (Meyer et al. 2002) og andre tiltak som reduserer detonasjonshastigheten, kan derfor forhindre skader på marint liv.
Ikke alle eksplosiver med moderat detonasjonshastighet, som for eksempel tradisjonell ANFO, kan brukes i vann eller vannfylte borehull da vannet endrer sprengstoffets sammensetning slik at detonasjon ikke oppnås. Sprengstoff som ikke detonerer ved initiering, kalles forsager og kan representere en betydelig forurensning av det marine miljøet, særlig med tanke på nitrogenforbindelser, og bør derfor unngås. Kjemisk forurensning vil også kunne oppstå etter en sprengning i form av restprodukter og udetonert sprengstoff.
Trykkbølgene i vannet som oppstår som følge av en sprengning, da særlig t1 (fig. 3), vil kunne speile seg i overflaten og endre fase. En slik speiling vil, som tidligere nevnt, skape en puls (Fardal 2005; Gaspin & Price 1972) som kan føre til langtidsskade på fisk. Slike uheldige speilinger i overflaten vil lettest oppstå når det er godt vær med blank sjø. Krusninger i overflaten og urolig sjø vil derimot bidra til å dempe trykkpulser og lyd fra sprengningen.
Et tiltak som er med på å redusere de voldsomme trykkbølgene fra sprengninger i vann, er bruk av såkalt boblegardin (Grimsbø & Kvadsheim 2018) (Figur 4). Boblegardin blir i andre deler av verden brukt nærmest som standard for å begrense skade på marint liv, men tiltaket er lite brukt i Norge.
Ved sprengning i vann må bruk av boblegardin, også kalt luftgardin, betraktes som det beste fysiske tiltaket for å begrense skadevirkningene på marint liv. En boblegardin lages ved at et perforert rør legges på bunnen og settes under trykk, slik at luftbobler stiger opp i vannsøylen og danner en sammenhengende vegg eller gardin av bobler (Domenico 1982a, 1982b; Croci et al. 2014; Grimsbø & Kvadsheim 2018; Johannessen et al. 2018; Keevin et al. 1997; Schmidtke 2010). Denne begrenser trykkpulsene fra sprengningen fra å forplante seg i sjøen ved at trykkpulsen dempes når den passerer luftboblene. Det er viktig at boblegardinen er en tett vegg av luftbobler som stiger mot overflaten og dermed i størst mulig grad hindrer trykkbølgen fra å passere.
Det kan være vanskelig å designe en boblegardin optimalt, slik at den gir tilsiktet effekt. Særlig utfordrende er det dersom bunnforholdene er vanskelige. Bruk av flere boblegardiner for å oppnå tilstrekkelig skjerming av sprengningsstedet kan da være lurt. Ved enkle bunnforhold – relativt flat og jevn bunn – kan boblegardinen lages ved å legge ut et perforert rør omkring sprengningsstedet. Kvadsheim et al. (2022) viste at det ikke var noen signifikant forskjell i lydutbredelsen, SEL eller SPL, ved bruk av dobbel eller enkel boblegardin.
Luftbobler kan også brukes til omrøring siden de fører til en oppadgående strøm i den omliggende vannsøylen, se Figur 5. Dette prinsippet er ofte brukt til omrøring av vannsjikt i mindre poller eller i ferskvann med stillestående vann mot bunnen som gir opphopning av hydrogensulfid (H2S). Brukt som boblegardin vil den oppadgående strømmen hindre vannet på den ene siden av gardinen i å blande seg med vannet på den andre siden, og i noen grad hindre giftige substanser og partikler fra å bli spredd. Dette utnyttes der man ønsker å hindre partikkelspredning og igjenslamming av områder (Laugesen 2006; Sharp et al. 2010), gjerne i forbindelse med anleggsarbeider.
Alternative løsninger basert på deflagrasjon av svartkrutt har også vært utprøvd for å etablere boblegardin i forbindelse med sprengningsarbeid i sjø (Samuelsen 1966).
5.4 - Råd
For å redusere mulige skadevirkninger som følge av selve sprengningen anbefales det at det benyttes boblegardin og oppdeling av salvene i mindre ladninger. Om mulig bør man sette av salvene når sjøforholdene i seg selv gir god demping, det vil si når det ikke er en speilblank overflate, men snarere litt vind og bølger. Ved sprengninger brukt i forbindelse med utfyllinger i sjø anbefales ikke at ladningen plasseres på fyllingsfot, men anbringes i rør i fyllingen. Det er svært uheldig med sprengninger i, eller som forplanter seg i, den frie vannsøylen. Derfor bør man være nøye med forladning i borehull. Man må gjerne også bruke fenghette for å skremme bort fisk og sjøpattedyr fra området umiddelbart før salven sprenges, men denne må da avfyres utenfor boblegardinen. Ikke minst bør man unngå å gjennomføre sprengninger nær gytefelt i tiden før, under og rett etter at gyting pågår, dette for å unngå å forstyrre gytingen og for å unngå mulig skade på egg og yngel. Når det gjelder sprengninger i nærheten av oppdrettsanlegg, er det en fordel å planlegge sprengningsarbeidet slik at det gjennomføres på et tidspunkt der anlegget er tømt for fisk eller ikke overstige lydnivåene Kvadsheim et al. (2022) observerte. For enkeltskudd betyr dette 169 dB re 1µPa spisstrykk og 152 dB re 1µPa 2 s SEL innenfor laks hørselsområde (20-500 Hz). For gjentatte sprenginger 160 dB re 1µPa spisstrykk og 145 dB re 1µPa 2 s SEL innenfor laks hørselsområde (20-500 Hz), men det må vurderes ut ifra hvor ofte og hvor mange eksponeringer fisken utsettes for.
Avhengig av prosjektets omfang bør det også vurderes å utføre biologisk overvåkning, da estimert lydtrykk ikke nødvendigvis er tilfredsstillende for å evaluere mulige negative adferdseffekter.
Havforskningsinstituttets råd for bruk sprengningsarbeider i sjø:
-
Sprengningsarbeider i sjø bør planlegges med tanke på marint liv på et tidlig tidspunkt i prosjekteringen. Her bør det gjøres lokale vurderinger blant annet for å ta hensyn til eventuelle gytefelt.
-
Havforskningsinstituttet tilrår å bruke boblegardin ved sprengningsarbeid i sjø.
-
Havforskningsinstituttet tilrår å dele opp salvene i mindre delladninger (sekvensiell sprengning).
-
Havforskningsinstituttet anbefaler å sette av salvene når sjøforholdene i seg selv gir god demping, altså ikke ved speilblank overflate.
-
Ved sprengning brukt i forbindelse med utfyllinger i sjø tilrår Havforskningsinstituttet at ladningen plasseres i rør i fyllingen, og ikke på fyllingsfot.
-
Man bør ta hensyn til forurensningsfare, i form av både partikler og kjemisk forurensning, fra sprengningsarbeider. Særlig gjelder dette i nærheten av oppdrettsanlegg for blåskjell.
-
Havforskningsinstituttet fraråder at sprengningsarbeid i sjø gjennomføres i gyteområder når fisken gyter samt i tiden rett etterpå, dette for å skjerme egg og yngel.
-
Havforskningsinstituttet tilrår at det utvises ekstra aktsomhet ved sprengningsarbeider i sjø dersom dette medfører irreversible endringer i strukturer og substrat i gyteområder. Dette rådet gjelder hele året.
I rådgivningsarbeidet opplever Havforskningsinstituttet ofte å komme inn i prosessen på et sent tidspunkt, når framdriftsplaner allerede er lagt. Dette gjør det vanskelig å foreta nødvendige justeringer for å ta hensyn til marint liv. Hensynet til marint liv bør derfor tas med i planleggingen på et tidlig tidspunkt.
-
Referanser
Amundsen, J. (1984). Emulsjonsslurry – erfaringer med borhulsdiameter 2 1/2" – 4". Fjellsprengingtekknikk, berg-mekanikk, geoteknikk, Norsk jord -og fjellteknisk forbund, p. 6.1- 6.18.
Aron, A.B., Slifko, J.P., Carter, A. (1948). Secondary Pressure Pulses Du to Gas Globe Oscillation in Underwater Explosions. I. Exprimental Data. The Jornal of the Acoustical Society of America, vol. 20, nr. 3, p. 271-276.
Aune, M., Clorenne, D. Billand, P., Bois, C., Folegot, T., Christensen, G.N., (2018). Vurdering av støy i forbindelse med undervannseksplosjoner i Båtsfjord, april 2018. Avaplan-Niva rapport no. 60306-01.
Campana, E. F., Amato, E., Bełdowski, J., Berglind, R., Boettcher, C., Angell-Hansen, K., Camerlenghi, A., Campostrini, P., Carniel, S., Grimsbø, E., Harms, J., Kvadsheim, H. P., Leffler, P., Marcinko, C., Moretti, P. F., Rogers, R. J., Rossland, H. K., Sprovieri, M., Sternheim, J. (2016): Research and innovation to address munitions in the sea by JPI Oceans Joint Programming Initiative - Healthy and Productive Seas and Oceans), NATO Research Workshop on “Sea Dumped Munitions and Environmental Risk”. Varna, Bulgaria. 11 to 13 Oct.
Cole, R. H. (1948). Underwater explosions. Princeton University Press, Princeton, NJ.
Croci, K., Arrigoni, M., Boyce, P., Gabillet, C., Grandjean, H., Grandjean, H., Jacques, N., Kerampran, S. (2014).Mitigation of underwater explosion effects by bubble curtains: experiments and modelling. 23rd MABS (Military Aspects of Blast and Shock), Oxford, UK, 7-12 September, United Kingdom.
Dahl, P. H., Keith Jenkins, A., Casper, B., Kotecki, S. E., Bowman, V., Boerger, C., Dall’Osto, D. R., Babina, M. A., & Popper, A. N. (2020). Physical effects of sound exposure from underwater explosions on Pacific sardines (Sardinops sagax). In The Journal of the Acoustical Society of America (Vol. 147, Issue 4, pp. 2383–2395). Acoustical Society of America (ASA). https://doi.org/10.1121/10.0001064
Dalen, J. (2009). Vurdering av påvirkning på torskelarver og -yngel i landbasert oppdrettsanlegg fra spregning på land og i sjøbunn og fra steinstøv i sjøvannet. Utredning for advokatfirmaet Schjødt DA, Oslo. Havforskningsinstituttet.
Dalen, J. (2012). Utredning for Statens Vegvesen, region vest tilknyttet prosjektet «Tunnel Fv7, Haukanesberget, Granvin» Havforskningsinstituttet.
Department of Arts, Heritage and the Gaeltacht (2014) Guidance to Manage the Risk to Marine Mammals from Man-made Sound Sources in Irish Waters . Dublin, Ireland, Department of Arts, Heritage and the Gaeltacht, 58pp. DOI: http://dx.doi.org/10.25607/OBP-176
Domenico, S.N. (1982a). Acoustic wave propagation in air-bubble curtains in water-Part I history and theory. Geophysics, VOL. 47, NO. 3, p.345-353.
Domenico, S. N. (1982b) Acoustic wave propagation in air-bubble curtains in water-Part II: Field experiment. Geophysics 47, 354-375.
Eliassen, R.A. & Iversen, M. (2004). Tålegrenser for fisk ved undersjøiske sprenginger, kasusstudie fra Kystverkets utbedringsarbeid i Lovund havn våren og sommeren 2004, Nordlandsforskning, NF-rapport nr 19/2004.
Engås, A., Olsen, S., Soldal, A. V. (1989). Undersøkelser av effekten på torsk i mær av refraksjonsseismiske sprenginger i Øygarden. Oppdragsrapport, Fangstseksjonen, Fiskeriteknologisk forskningsinstitutt, Institute of Fishery Technology research.
Fardal, R. (2005). Måling av lydtrykk fra undersjøiske mineeksplosjoner – Virkningerpå området for planlagt småbåthavn i Østhusvik. Forsvarets forskningsinstitutt, FFI/RAPPORT-05/03326.
Faulkner, S. G., Tonn, W. M., Welz, M., Schmitt, D. R. (2006). Effects of Explosives on Incubating Lake Trout Eggs in the Canadian Arctic. North American Journal of Fisheries Management, 26:4, p.833-842.
Faulkner, S. G., Welz, M., Tonn, W. M., Schmitt, D. R. (2008) Effects of Simulated Blasting on Mortality of Rainbow Trout Eggs. Transactions of the American Fisheries Society 137:1–12. DOI: 10.1577/T07-035.1
Fauske, A. (2007) Undervannssprenginger i nærheten av oppdrettsanlegg – begrensninger og krav til gjennomføring. Norsk Jord og Fjellteknisk Forbund, Fjellsprengningsdagen, Bergmekanikkdagen, Geoteknikkdagen, p.15.1 – 15.23.
Gaspin, J. B., Price, R. S. (1972). The inderpressure field from explosives in water as modified from cavitation. Naval ordnance laboratory, Wite Oak, Silver Spring, Maryland, USA
Gaspin, J. B. (1975). Experimental investigation of effects of underwater explosions on swimbladder fish, I: 1973 Chesapeake bay tests. Naval surface weapons center white oak laboratory silver spring, Maryland, NSWC/WOL/TR 75-58.
Gaspin, J. B., Wiley, M. L., Peters GB (1976). Experimental investigation of effects of underwater explosions on swimbladder fish, II: 1975 Chesapeake Bay tests. Naval surface weapons center white oak laboratory silver spring, Maryland, NSWC/WOL/TR 76-61.
Geers, T. L. & Hunter, K. S. 2002 An integrated wave-effects model for an underwater explosion bubble. The Journal of the Acoustical Society of America 111, 1584.
Govoni, J. J., West, M. A., Settle, L. R., Lynch, R. T., Greene, M. D. (2008). Effects of Underwater Explosions on Larval Fish: Implications for a Costal Engineering Project. Journal of Costal Research, Florida, 24, 2B, p.228-233.
Grimsbø, E. (2016). Measuring methods for fish welfare during slaughter based on electrical impedance, EEG, ECG and blood parameters. Dissertation for the degree of Philosophiae Doctor (PhD). University of Bergen.
Grimsbø, E., Kvadsheim, P. H. (2018). Sprengningsarbeider i sjø – effekter på marint liv og mulige tiltak (Blasting operations at sea – effects on marine life and possible actions). Fjellsprengningsteknikk Bergmekanikk/Geoteknikk 2018. Norsk Forening for Fjellsprengningsteknikk, Norsk Bergmekanikkgruppe og Norsk Geoteknisk Forening. Oslo 22-23 November 2018. ISBN: 978-82-8208-062-0. p. 33-51
Higgins, A. & Loiseu, J., Mi, X.C. (1979)Detonation velocity/diameter relation in gelled explosive with inert inclusions, American Institute of Physics, AIP Conference Proceedings.
Hveding, Ø.P. (2018) Sprengningsarbeid i Hagavika og mulig effekt på fisk. Oppdr.nr. 5164410, Dok.01, Ver.J02.
Iwama, G. K., Afonso, L. O. B. (2006) Stress in Fishes, Ed. Evans, D. H., Claiborne, J. B. The physiology of fishes, third edition, Taylor & Francis Group, FL. p.319-323.
Jenkins, A. K., Dahl, P. H., Kotecki, S. E., Bowman, V., Casper, B., Boerger, C., & Popper, A. N. (2022). Physical effects of sound exposure from underwater explosions on Pacific mackerel (Scomber japonicus): Effects on non-auditory tissues. In The Journal of the Acoustical Society of America (Vol. 151, Issue 6, pp. 3947–3956). Acoustical Society of America (ASA). https://doi.org/10.1121/10.0011587
Johannessen, T.C., Johnsen, A., Dullum, O., Bjerktveit, B. (2018). Trykkbølgedemping ved spregning under vann – småskader av boblegardin og luftfyldte materialer. FFI rapport 17/16837.
Johnsen, G.H., Kålås, S., Kambestad, A., (1994). Vurdering av skader på fisk ved undervannsspregninger i Raudbergsbukta i Lærdal, Erfaringer med spregningsarbeide høsten 1993 og våren 1994. Rådgivende biologer AS, Raport nr 139.
Keevin, T. M., Hempen, G. L., Schaeffer, D. J. (1997). Use of a bubble curtain to reduce fish mortality during explosive demolition of locks and dam 26. Mississippi river, International Society of Explosives Engineers, Proceedings of the twenty-third annual conference on explosives and blasting technique.Feb. 2-5, Las Vegas, Nevada U.S.A.
Kestin, S.C., Van de Vis, J. W., Robb, D. H. F. (2002). Protocol for assessing brain function in fish and the effectiveness of methods used to stun and kill them. The Veterinary Record 150, p.302-307.
Ketten, D. R. (1995). Estimates of blast injury and acoustic trauma zones for marine mammals from underwater explosions. In: Kastelein R, JA Thomas & PE Nachtigall (eds). Sensory Systems ofAquatic Mammals, De Spil Publishers, The Netherlands. ISBN 90-72743-05-9
Kiran, J. H. (2016). Undervannsdetonasjoner – en kort innføring. Forsvarets forskningsinstitutt, FFIrapport, 16/01500.
Kjellsby E (1993). Akustisk eksponering av fisk ved undervannssprenginger, Forsvarets forskningsinstitutt, FFI/rapport-93/2004.
Kjellsby, E., Kvalsvik, K. (1997). Begrensning av skade på marin fauna ved undervannssprenginger, Forsvarets Forskningsinstitutt, FFI/rapport-97/04847.
Koschinski, S. (2011). Underwater Noise Pollution From Munitions Clearance and Disposal, Possible Effects on Marina Vertebates and Its Mitigation. Marine Technology Society Journal, vol. 45, nr. 6, p. 80-88.
Krise, W. F. (2001) Sensitivity of Atlantic Salmon Eggs to Mechanical Shock during the First Six Hours after Fertilization .North American Journal of Aquaculture 63, 34-37.
Kvadsheim PH, Sivle LD, Hansen RR, Karlsen HE (2017). Effekter av menneskeskapt støy på Havmiljø - rapport til Miljødirektoratet om kunnskapsstatus. FFI-RAPPORT 2017/00075. http://www.ffi.no/no/Rapporter/17-00075.pdf
Kvadsheim, P. H., Søvik, A. A., Rokke, E., Kolstrup, M. L., Forland, T. N., Pedersen, G., Grimsbø, E., Dale, O. B., Cao, Y., Stene, A., Smedsrud, M. (2022) Risiko for skade på laksefisk i omkringliggende oppdrettsanlegg ved etablering av undervanns-skytefelt på Korsnes Fort. FFI-rapport 22/00814 ISBN 978-82-464-3401-8
Laugesen, J. (2006) Pilotprosjekt i Trondheim havn, Boblegardin mot spregning av muddermasser. Ref. 625 0512, DNV Consulting Rapport nr. 2006-025.
Larsen, T., Kjellsby, E., Olsen, S. (1993). Effekter av undervannssprengninger på fisk. Rapport fra senter for marine resurser nr.11, Havforskningsinstituttet.
McQueen, A. D., Wilkens, J. L., Katzenmeyer, A. W., Harrington, H. F., & Suedel, B. C. (2021). Assessing the Impacts of Dry Blasting on Fish Eggs in Adjacent Spawning Habitat. In Bulletin of Environmental Contamination and Toxicology (Vol. 108, Issue 2, pp. 190–195). Springer Science and Business Media LLC. https://doi.org/10.1007/s00128-021-03331-3
Meyer, M., Köhlner, J., Homburg, A. (2002). Explosives. Fifth Edition, Wiley-VCH Verlag GmbH & Co, KGaA.
Petel, O. E., Mack, D., Higgins, A. J., Turcotte, R., Chan, S. K. (2007). Minimum propagation diameter and thickness of high explosives. J. Loss Prev Process Ind 20(4-6): 578-583.
Samuelsen, E. (1966). Boblegardin fremstillt ved hjelp av kruttgassgenerator. Foredrag fra konferanse i Fjellspregningsteknikk, Norsk Forening for Fjellspregningsteknikk, Oslo.
Schmidtke, E. (2010). Schockwellendämpfung mit einem Luftblasenschleier zum Schutz der Meeressäuger. WTD 71, Forschungsbereich für Wasserschall und Geophysik, Berliner Stra β e 115, 24340 Eckernförde, Deutschland.
Simenstad, C. A. (1973). Biological effects of underground nuclear testing on marine organisms. I. Review of documented shock effects, discussion of mechanisms of damage, and prediction of amcHitka test effects. Conference of the environmental effects of explosives and explosions May 30-31 1973. Naval Ordnance Laboratory, Silver Springs, Maryland, USA.
Smith, M. E., Accomando, A. W., Bowman, V., Casper, B. M., Dahl, P. H., Jenkins, A. K., Kotecki, S., & Popper, A. N. (2022). Physical effects of sound exposure from underwater explosions on Pacific mackerel (Scomber japonicus): Effects on the inner ear. In The Journal of the Acoustical Society of America (Vol. 152, Issue 2, pp. 733–744). Acoustical Society of America (ASA). https://doi.org/10.1121/10.0012991
Soldal, A.V. (1990). Minespregning ved Helligvær, effekten på laks i merd. Oppdragsrapport, Fangsseksjonen, Fiskeriteknisk forskningsinstitutt.
Svendsen, P. (2012). Reglement for eksplosivrydding under vann - Fastsettes til bruk i Sjøforsvaret. KNM Tordenskjold 2012
Teleki, G. C., Chamberlain, A. J. (1978). Acute Effects of Underwater Construction Blasting on Fishes in Long Point Bay, Lake Erie. Journal of the Fisheries Research Board of Canada, 35(9), p.1191-1198.
Toresen, R., Søvik, G. (2012). Anmodning om faglig innspill i Engsbøsaken, Havforskningsinstituttet, Ref.2009/1183, arkiv.nr. 008, løpenr. 1444/2012.
Trettenes, V. (2006). Effekter av undervannssprenginger på torsk Gadus Morhua. Observasjoner fra sprengingene ved Nedre Stedje i Sogndal kommune 2005. Litteraturstudium over tidligere arbeid. Høgskulen i Sogn og Fjordane, N-Nr. 6/2006.
von Benda-Beckmann, A. M., Aarts, G., Sertlek, H. O., Lucke, K., Verboom, W. C., Kastelein, R. A., Ketten, D. R., van Bemmelen, R., Lam, F-P. A., Kirkwood, R. J., Ainslie, M. A. (2015). Assessing the Impact of Underwater Clearance of Unexploded Ordnance on Harbour Porpoises (Phocoena phocoena) in the Southern North Sea, Aquatic Mammals, 41, p.503-523
Yelverton, J. T., Richmond, D. R., Fletcher, E. R., Jones, R. K. (1973). Safe distances from underwater explosions for mammals and birds. Lovelace Foundation for Medical Education and Research, Albuquerque NM 87108, AD-766 952.
Yelverton, J. T., Richmond, D. R., Hicks, W., Saunders, K., Fletcher, E. R. (1975). The relationship between fish size and their response to underwater blast. Lovelace Foundation for Medical Education and Research, Albuquerque NM 87108, DNA 3677T.
6 - Vindkraft til havs
6.1 - Bakgrunn
Miljøvennlig produksjon av energi er blitt langt viktigere de senere år, og vindturbiner er en av de kraftproduserende metoder som etter hvert har fått stor, internasjonal utbredelse. Foreløpig plasseres de fleste vindturbiner på land, men det planlegges å plassere vindturbiner også i havområder. Havbaserte vindkraftanlegg er allerede i drift langs kysten av flere europeiske land, og i Norge har regjeringen nå åpnet to spesifikke havområder for vindkraftproduksjon: I juni 2020 ble det åpnet opp for at det kan søkes om konsesjon for bygging av havvind i områdene Utsira Nord og Sørlige Nordsjø II. (NVE 2010, 2012, https://www.nve.no/energiforsyning/kraftproduksjon/ vindkraft/vindkraft-til-havs/). Det ble tidligere foreslått flere områder for framtidig åpning, disse blir gjennomgått på nytt med mål om å tildele arealer tilsvarende 30 GW innen 2040 (NFD 2022) (Selv om slike vindkraftanlegg representerer en mer miljøvennlig produksjonsform for energi enn f.eks. kraftverk basert på fossile energikilder, er det samtidig viktig å være klar over at også vindkraftanlegg vil påvirke det marine miljø. Vindkraftanleggene vil legge beslag på sjøareal og påvirke strømforhold, samt produsere støy og elektromagnetisk stråling som vil kunne påvirke marine økosystemer og ressursutnyttelsen av disse. For at havbaserte vindkraftanlegg skal kunne produsere mest mulig miljøvennlig energi er det derfor viktig å utrede og redusere eventuelle utilsiktede konsekvenser denne type anlegg vil kunne påføre det marine miljø.
I forbindelse med etablering av nye vindkraftanlegg, har Havforskningsinstituttet en rådgivende rolle på lik linje som for seismikk og bruk av eksplosiver, ved at Norges Vassdrags- og Energidirektorat (NVE) ber om råd om mulige effekter på havmiljø i forkant av etablering av nye anlegg. I denne rapporten fokuserer vi på beskrivelsen av mulige effekter av støy fra havvindanlegg, men det minnes om at også andre påvirkningsområder blir introdusert av havvindaktiviteter som for eksempel forandringer av elektromagnetiske felt, strømningsmønstre i havsirkulasjonen eller lysfordelingen i vannsøylen.
6.2 - Kunnskapsgrunnlag om effekter støy fra vindkraftanlegg på marine organismer
Kunnskapsgrunnlaget er gjennomgått i detalj i en egen rapport (de Jong et al. 2020b) og derfor bare kort oppsummert her, samt at det er lagt til ny kunnskap som har tilkommet etter den rapporten ble publisert . Vindkraftanlegg kan påvirke mange ulike aspekter ved marine samfunn, men da denne rapporten omhandler effekter av støy, er det kun den delen av påvirkningen som er behandlet her.
Vindkraftanlegg til havs lager lyd som er hørbar for de fleste fisk og sjøpattedyr. Lydene kan deles opp i konstruksjonsstøy; altså lyd fra oppføringen av vindturbinene, og produksjonsstøy; lyd fra vindturbinene i drift.
6.2.1 - Konstruksjonsstøy
Lydkilder i denne perioden kan være pæling, boring, gravearbeid og økt båttrafikk (Thomsen et al, 2006).
For bunnmonterte vindturbiner blir det ofte brukt pæling der en hydraulisk hammer dunker ned påler i havbunnen. Impulsive lyder som dette beskrives gjerne ved spisstrykk (0-peak trykk) for å beskrive maksimaltrykket som er et mål på den høyeste positive eller laveste negative verdien i en definert periode, eller lydeksponeringsnivå (sound exposure level, SEL) for å beskrive energien i en lydpuls/hammerslag. Den samlede (kumulative) energien fra mange slag kan også beskrives ved å legge sammen energien til enkeltslag over en periode. Spisstrykket kan være omtrent 220 dB re 1 μPa 10 meter fra en påle med diameter på 0.75 cm og 200 dB re 1 uPa 300 meter fra en påle med diameter på 5 meter (Dahl et al. 2015). 1600 m fra pæling av en påle med diamenter på ca 2.5 m ble det målt spisstrykk på 190 dB re 1 μPa og SEL på 165 dB re 1μPa 2 s (Betke 2014). For pæling er spisstrykket typisk 20 - 25 dB høyere enn SEL.
Ved avstander på 1600 - 16 000 m fra pælingen er mesteparten av lyden i frekvensområdet rundt 50 -400 Hz, men lydnivået er høyere enn bakgrunnsstøyen fra under 20 Hz til ca 10 kHz (Betke 2014). Varigheten på lydpulsene øker med avstand. 600 m fra kilden kan varigheten være ca 0.1 sekund, men 17 km fra kilden kan varigheten være ca 0.6 sekund (Betke 2014). Med 1 sekund mellom slagene vil lyden kunne oppfattes som kontinuerlig støy ved store avstander.
I utbyggingsfasen kan lyd fra konstruksjonsarbeid som pæling skade hørsel og skremme vekk mobile dyr, men tiltak som boblegardiner kan redusere lydnivået betydelig. Forskning på effekter av lyd fra vindkraftanlegg på marine dyr har primært fokusert på utbyggingsfasen, men likevel vet man fortsatt lite om påviste effekter på enkeltindivider kan føre til effekter på populasjonsnivå.
Til konstruksjon av flytende turbiner brukes ikke pæling, men boring kan fortsatt være en del av konstruksjonsarbeid avhengig av hvilken ankermåte som blir brukt. Skipstrafikk vil øke i konstruksjonsfasen av alle vindkraftanlegg.
6.2.2 - Produksjonsstøy
Lydnivået av vindturbiner er mye lavere enn lydene under konstruksjonsfasen, men dette er kontinuerlig lyd som er tilstede så lenge vindturbinene er i drift, så tidsaspektet er derfor mye lengre enn for produksjonsstøyen. Lydnivået varierer og vil i stor grad påvirkes av vindstyrke (Pangerc et al. 2016).
Lyden som brer seg i luft fra rotasjonsbladene blir nesten fullstendig reflektert fra havoverflaten og får liten betydning under vann (Tougaard et al. 2009, Betke 2014). Under vann kommer støy fra vindturbiner hovedsakelig fra girkassen og fra generatoren som brer seg nedover tårnet og ut i vannet og i undergrunnen. Mekaniske deler som roterer resulterer ofte i ekstra høy lyd på smale frekvensbånd, der frekvensen er direkte knyttet til antall mekaniske svingninger/rotasjoner/vibrasjoner per sekund (toner). Vinkraftanlegg kan påvirke oseanografiske forhold som videre kan påvirke lydutbredelsen i vannet; lagdeling/skikt som påvirker lydutbredelse, temperatur som påvirker lydhastighet.
Det er flere eksempler på målt produksjonsstøy fra vindturbiner i litteraturen, men det er ofte målt i ulike avstander fra en eller flere vindturbiner. Nær vindturbinene (1-50 m) er det forhøyet bakgrunnsstøy over et bredt frekvensspekter fra 10 - 1000 Hz i tillegg til at det er enkelte frekvenser som skiller seg ut med høyere topper (toner). Frekvensen til tonene avhenger av vindturbintype og vindhastighet (Tougard et al. 2009, Pangerc et al. 2016, Cheesman 2016, Wahlberg og Westerberg 2005). Ved større avstander (>100 m) ser man gjerne bare tonene som skiller seg ut (Betke 2014, Thomsen et al, 2006).
På Hywind Demo, som er en av de første flytende turbiner i norske farvann, ble det i tillegg til det lave kontinuerlige lyd fra turbinen malt høye transiente lyder som kommer sannsynligvis fra strekk i ankerkjettingene (Martin et al. 2011). På Hywind Scotland ble det målt lavere transiente lyder, men i tillegg litt høyere kontinuerlige toner enn tidligere beskrevet for havvindanlegg (Burns et al. 2022; Wahlberg og Westerberg 2005). Generelt sett er det mye variasjon mellom anlegg i lydnivå (Tougaard et al. 2020; Støber & Thompsen 2021). Dette åpner opp for avbøtende tiltak som å bruke minst bråkete turbiner, og i muligens også å tilpasse lydnivåer og -karakteristikker til å unngå de frekvenser som er viktig for de mest sensitive arter i området som anlegget blir bygget i.
Flere rapporter har nylig beskrevet effekten og kunnskapshull om effekter av støy i utbyggingsfase (Thomsen et al. 2006; Copping & Hemery 2020; Farr et al. 2021). Det blir argumentert at fordi støy i utbyggingsfasen er mye høyere en støy i driftsfasen, er risikoen for påvirkning størst i den perioden. Imidlertid er utbyggingsfasen begrenset i tid (måneder), mens produksjonsfasen mye mer langvarig (tiår) og støyen her kontinuerlig. Dette påvirker særlig de dyrene som lever i eller nært selve området til vindkraftanlegget (Tougaard et al. 2009; Marmo et al. 2013). Mange dyr, fra krepsdyr til hval hører, bruker, og kan bli forstyrret av lyd (f.eks. Hawkins og Popper 2017; Popper & Hawkins 2019; Kvadsheim et al. 2020b). Støy kan dermed påvirke for eksempel predator-byttedyr interaksjoner (Simpson et al. 2016), reproduksjon (de Jong et al. 2020a) og hvordan evertebrater påvirker økosystemet (Solan et al. 2016). En nylig artikkel fant at to forskjellige fiskekor ble påvirket av både pæling og driftsstøy, med ett kor som økte både i intensitet og varighet under driftsfasen, mens andre endringer ikke var signifikante (Siddagangaiah et al. 2021). Det er imidlertid uklart om disse endringene oppsto fra endringer i fiskens kalleatferd, potensielt kompensert for høyere lydnivåer ved å synge lengre og høyere, eller endringer i mengden av fisk på grunn av tiltrekning til stedet.
Det har vært vanskelig å finne en måte å kvantifisere artenes eksponering for kontinuerlig lyd i havet, men det har blitt foreslått å bruke en kombinasjon av andelen av en populasjon som utsettes for økt lydnivå og hvor mye av tiden populasjonen blir utsatt (Merchant et al. 2017). Den metoden gir en målestokk for hvor utbredt lydpåvirkningen er, men den er fortsatt avhengig av at man setter en grenseverdi hvorunder støynivået er antatt akseptabelt. Fordelen med å bruke denne metoden er at man kan planlegge vindkraftanlegg på et sted hvor man unngår overlapp med viktige populasjoner.
6.3 - Konsekvensvurdering
Støy kan sammen med andre effekter av vindkraftanlegg (eks endringer i elektromagnetiske felt og strøm-mønster) føre til at habitatet endres, som potensielt kan gi negative effekter som nedsatt reproduksjon og/eller økt dødelighet. For å få oversikt over de reelle effektene av vindkraft til havs er det viktig å se effektene i sammenheng og trekke konklusjoner på populasjonsnivå.
For vindkraftkonsesjoner bør det derfor gjennomføres prosjektspesifikke evalueringer av mulige effekter på miljø og etableres overvåkingsprogram med relevante før- og etterundersøkelser for å skaffe et godt grunnlag om anleggenes påvirkning på marine organismer, både i anleggs- og driftsfasen. Kartlegging av bunntilknyttede organismesamfunn bør gjennomføres i de aktuelle områdene dersom slik informasjon mangler, og anleggene bør i minst mulig grad lokaliseres i områder med forekomster av viktige og sårbare ressurser og naturtyper, inkludert konsentrerte gyteområder og vandringsruter til økonomisk og økologisk viktige fiskebestander (de Jong et al. 2020b).
I utbyggingsfasen kan lyd fra konstruksjonsarbeid som pæling skade hørsel og føre til en skremmeeffekt for både fisk og sjøpattedyr, men tiltak som boblegardiner kan redusere lydnivået betydelig. For flytende vindturbiner kan det brukes sugeanker for å unngå boring. Det bør bruker forankringsmåter som ikke lager kraftig lyd ved bevegelse.
Kontinuerlig lavfrekvent støy fra turbinene i driftsfasen vil først og fremst påvirke dyrene inne i og nært vindkraftanleggene. Siden bunndyr etablerer seg i området og fisk ofte blir tiltrukket til anleggene, blir disse gruppene utsatt for produksjonsstøy over lengre tid. Kontinuerlig lavfrekvent støy kan påvirke atferd som, for eksempel, bunnmiksing, beiting, reproduksjon, antipredator adferd og kommunikasjon, men hvor stor disse effektene er sammenlignet med positive effekter av økt mattilgang og ly i vindkraftanlegg er ukjent. Det er også kunnskapshull omkring marine dyrs evne til å tilpasse seg støypåvirkninger over tid.
6.4 - Råd for støy fra vindkraft til havs
Kunnskapen om miljøeffekter av vindkraftanlegg til havs er fortsatt mangelfull, og derfor er det sannsynlig at rådene kommer til å endres de neste årene, etter hvert som mer kunnskap kommer til.
Da det er knyttet betydelig usikkerhet til omfanget av miljøeffekter fra vindkraftanlegg i norske farvann fraråder Havforskningsinstituttet foreløpig utbygging i områder som er spesielt viktige for ulike marine dyr. Dette omfatter gyteområder for fisk og kaste og beiteområder for sjøpattedyrarter. Utbygging bør også unngås i områder som er viktig for det marine økosystemet som for eksempel områdene rundt Lofoten, eller sårbare områder som korallrev.
Rådene som står oppgitt her er kun dem som er direkte knyttet til effekter av støy. Fullstendig liste over råd for utbygging av vindkraft til havs finnes i de Jong et al. 2020b.
-
Havforskningsinstituttet fraråder vindkraftutbygging i områder som er spesielt viktig for økosystemet og bestander som er viktig for bærekraftig bruk av ressurser, som gyteområder og gytevandringsruter for fisk, kasteområder for sel og beiteområder for sjøpattedyr. Hvilke arter som inkluderes, vurderes ut ifra hvilke som har en risiko for populasjonseffekt som følge av påvirkningen. Disse er de samme som for seismikk, som er gjennomgått i Vedlegg 1.
-
Havforskningsinstituttet anbefaler bruk av støydempende tiltak som boblegardin under utbygging, og at man unngår utbyggingsarbeid i gyteperioder for fisk, samt beite og kasteperioder for sjøpattedyr, i og nær områder dette gjelder.
-
Havforskningsinstituttet anbefaler bruk av materialer i fortøyning av flytende vindturbiner som lager minst mulig støy ved bevegelser av turbinene, f.eks unngå rykking og napping i forankring.
-
Havforskningsinstituttet anbefaler innsamling av data for å undersøke i hvilken grad, vindkraftanlegg endrer lydbildet. Innsamlete data bør være åpent tilgjengelig.
6.5 - Referanser
Betke K. 2014. Underwater construction and operational noise at alpha ventus. In: Federal Maritime and Hydrographic Agency, Federal Ministry for the Environment, Nature Conservation and Nuclear Safety (eds) Ecological Research at the Offshore Windfarm alpha ventus. Springer Spektrum, Wiesbaden.
Burns, R.D.J., S.B. Martin, M.A. Wood, C.C. Wilson, C.E. Lumsden, and F. Pace. 2022. Hywind Scotland Floating Offshore Wind Farm: Sound Source Characterisation of Operational Floating Turbines.Document 02521, Version 3.0 FINAL.Technical report by JASCO Applied Sciencesfor EquinorEnergy AS.
Cheesman S. 2016. Measurements of Operational Wind Turbine Noise in UK Waters. In The Effects of Noise on Aquatic Life II (pp. 153–160). Springer New York. https://doi.org/10.1007/978-1-4939-2981-8_18 .
Copping AE, Hemery LG. 2020. OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development Around the World. Report for Ocean Energy Systems (OES).
Dahl PH, de Jong CAF, Popper AN. 2015. The Underwater Sound Field from Impact Pile Driving and Its Potential Effects on Marine Life. Acoustics Today, 11, 18–25
de Jong, K., Forland, T. N., TN, Amorim, M. C. P., MCP, Rieucau, G., Slabbekoorn, H. and, Sivle, L. D. (2020a). LD. 2020. Predicting the effects of anthropogenic noise on fish reproduction from available data: the importance of types of noise and stages of reproduction. Reviews in Fish Biology and Fisheries 30:245-268. doi: 10.1007/s11160-020-09598-9.
de Jong, K., Steen, H., Forland, N. F., Wehde, H., Nyqvist, D., Palm, A.C.U., Nilssen, K.T., Albretsen, J., Falkenhaug, T., Biuw, M., Buhl- Mortensen, L. Sivle, L.D. (2020b) Potensielle effekter av havvindanlegg på havmiljøet. Rapport fra Havforskningen 2020-42. ISSN:1893-4536. 42 pp
Farr, H., Ruttenberg, B., Walter, R. K., Wang, Y.-H., & White, C. (2021). Potential environmental effects of deepwater floating offshore wind energy facilities. In Ocean & Coastal Management (Vol. 207, p. 105611). Elsevier BV. https://doi.org/10.1016/j.ocecoaman.2021.105611
Hawkins AD, Popper AN. 2017. A sound approach to assessing the impact of underwater noise on marine fishes and invertebrates. ICES Journal of Marine Science 74:635–651. DOI: 10.1093/icesjms/fsw205.
Kvadsheim PH, Forland TN, de Jong K, Nyqvist D, Grimsbo E, Sivle LD. 2020b. Effekter av støyforurensning på havmiljø - kunnskapsstatus og forvalningsrådgiving. FFI-Rapport 20/01015.
Merchant ND, Faulkner RC, Martinez R. 2017 . Marine Noise Budgets in Practice. Conservation Letters, 11, e12420. doi: 10.1111/conl.12420.
Marmo B, Roberts I, Buckingham MP, King S, Booth C 2013. Modelling of Operational Offshore Wind Turbines including noise transmission through various foundation types, Scottish Government Edingburgh
Martin, B, MacDonnell, J, Vallarta, J, Lumsden, E and Burns, R. 2011. HYWIND Acoustic Measurement Report: Ambient Levels and HYWIND Signature. Technical report for Statoil by JASCO Applied Sciences.
NFD (Nærings- og fiskeridepartementet). 2020. Veikart for grønt energiløft. https://www.regjeringen.no/contentassets/1c3d3319e6a946f2b57633c0c5fcc25b/veikart_skisse_uu_ja.pdf.
Pangerc T, Theobald PD, Wang LS, Robinson SP, Lepper PA. 2016. Measurement and characterisation of radiated underwater sound from a 3.6 MW monopile wind turbine. The Journal of the Acoustical Society of America, 140(4), 2913–2922. https://doi.org/10.1121/1.4964824
Popper AN, Hawkins AD. 2019. An overview of fish bioacoustics and the impacts of anthropogenic sounds on fishes. Journal of Fish Biology, 94, 692–713. doi: 10.1111/jfb.13948.
Simpson SD, Radford AN, Nedelec SL, Ferrari MC, Chivers DP, McCormick MI, Meekan MG. 2016. Anthropogenic noise increases fish mortality by predation. Nature Communications, 7, 10544. doi: 10.1038/ncomms10544.
Solan M, Hauton C, Godbold JA, Wood CL, Leighton TG, White P. 2016. Anthropogenic sources of underwater sound can modify how sedimentdwelling invertebrates mediate ecosystem properties. Scientific reports, 6, 20540.
Stöber, U., & Thomsen, F. (2021). How could operational underwater sound from future offshore wind turbines impact marine life? In The Journal of the Acoustical Society of America (Vol. 149, Issue 3, pp. 1791–1795). Acoustical Society of America (ASA). https://doi.org/10.1121/10.0003760
Thomsen F, Lüdemann K, Kafemann R, Piper W. 2006. Effects of offshore wind farm noise on marine mammals and fish, biola, Hamburg, Germany on behalf of COWRIE Ltd.
Tougaard J, Henriksen OD, Miller LA. 2009. Underwater noise from three types of offshore wind turbines: Estimation of impact zones for harbor porpoises and harbor seals. The Journal of the Acoustical Society of America, 125, 3766–3773. https://doi.org/10.1121/1.3117444.
Tougaard, J., Hermannsen, L., and Madsen, P. T. (2020). “ How loud is the underwater noise from operating offshore wind turbines?” J. Acoust. Soc. Am. 148(5), 2855–2893. https://doi.org/10.1121/10.0002453
Verfuß T. 2014. Noise mitigation systems and low-noise installation technologies. In: Federal Maritime and Hydrographic Agency, Federal Ministry for the Environment, Nature Conservation and Nuclear Safety (eds) Ecological Research at the Offshore Windfarm alpha ventus. Springer Spektrum, Wiesbaden. https://doi.org/10.1007/978-3-658-02462-8_16 .
Wahlberg M, Westerberg H. 2005. Hearing in fish and their reactions to sounds from offshore wind farms. Marine Ecology Progress Series, 288, 295–309. https://doi.org/10.3354/meps288295.
7 - Vedlegg I: Rådgivningskart for seismikk og elektromagnetiske undersøkelser
Havforskningsinstituttet fraråder seismikk-undersøkelser i og tett opptil gyteområder for en rekke viktige fiskebestander i gyteperioden og i viktige beiteområder for noe arter av bardehval i beiteperioden. Havforskningsinstituttet fraråder også elektromagnetiske undersøkelser (EMU) i gytevandringsområder i vandringsperiodene.
7.1 - Grunnlag for rådgivning for fisk
Kart for gyteområder, gytevandringer og beiteområder gjennomgås årlig sammen med bestandsansvarlige personer for hver av fiskebestandene. Om det i løpet av året er kommet inn relevant ny kunnskap, vil kartene oppdateres. I tillegg foregår en vurdering av hvilke bestander som skal inkluderes i rådgivningen, og det settes opp en liste over disse bestandene. Hvilke arter som blir inkludert vurderes etter følgende kriterier:
Kommersiell viktighet: Hovedsakelig gis råd for fiskebestander underlagt fiskeriforvaltning for kommersiell utnyttelse. Det er også for disse bestandene man har best grunnlag for å vite noe om gyteområde og gyteperiode, da det jevnlig, ofte flere ganger pr år, gjennomføres overvåkningstokt for disse.
Økosystemkomponent: Om arten innehar en nøkkelrolle i økosystemet, eks. om den er viktig næringsgrunnlag for en rekke arter av fisk og sjøfugl.
Bestandens tilstand: Basert på kategoriene fra rødlisten er alle bestander kategorisert som regionalt utdødd/kritisk truet, sårbar/nær truet eller livskraftig. Bestander som er kritisk eller nær truet bør en være særlig påpasselig med at ikke utsettes for negativ påvirkning.
Lydproduksjon og hørsel: Arter som selv hører godt og produserer lyd i de frekvenser som seismikken bruker, og særlig arter som bruker lyd ifm. gyting, regnes som ekstra sensitive for forstyrrelser av lyd fra seismikk.
Gyteområder- og perioder: I tillegg er første del av gyteperioden ansett som mer sårbar en siste del av gyteperioden. Dette skyldes at om fisken skremmes bort fra gyteplassen tidlig i sesongen, er det få fisk som har gytt. Senere i gytesesongen er en større andel av bestanden ferdig å gyte, og konsekvensene ved at fisken da forlater gyteplassen er mindre. For arter med definert tid for konsentrert gyteperiode og tid for «toppgyting» frarådes seismikk fra gytingen starter (bruker start for generell gyteperiode og ikke for konsentrert), og vi fraråder til og med «gytetoppen» eller den konsentrerte gyteperioden er over.
Gytevandring: Konsentrerte gytevandringer inn mot gytefeltene finnes for ulike bestander, men det er hovedsakelig for bestandene NEA torsk, NVG sild og lodde det finnes tilstrekkelig kunnskap om hvor og når og hvor denne vandringen finner sted.
Sensitivitet for elektromagnetisk stråling: (Gjelder kun for EMU). Arter med kjent sensitivitet for EM felt regnes som mer sårbare for forstyrrelser fra EM undersøkelser.
Tabell A1.1 viser alle fiskebestander som er vurdert, hvilke vurderinger som er gjort, og om arten er inkludert i rådgivningskartene eller ikke.
| Art | Vurderingsgrunnlag | Inkludert i fraråding | Kommentar |
|---|---|---|---|
| NVG sild (Clupea harengus) | høy kommersiell og økologisk viktighet, svært god hørsel, gyting konsentert i tid og rom | ja | |
| NEA torsk (Gadus morhua) | høy kommersiell og økologisk viktighet, god hørsel og lyd essensiellt for gyting. Viktigste gyteområder konsentrert i tid og rom. | ja | |
| Kysttorsk (Gadus morhua) | nøkkelart, god hørsel, bruker lyd under gyting, | ja | |
| Lodde (Mallotus villosus) | høy kommersiell og økologisk viktighet, kort gyteperiode | ja | |
| NEA hyse (Meanogrammus aeglefinus) | nøkkelart, god hørsel, bruker lyd under gyting, kommersiellt noe viktig | ja | |
| NEA sei (Pollachius virens) | Kommersiellt og økologisk viktig, god hørsel. | ja | |
| Polartorsk (Boreogadus saida) | nøkkelart, kritisk truet, kort gyteperiode | ja | |
| Nordsjøhyse (Meanogrammus aeglefinus) | Viktig økologisk, god hørsel, bruker lyd under gyting, kommersiellt noe viktig | ja | |
| Nordsjøtorsk (Gadus morhua) | høy kommersiell og økologisk viktighet, god hørsel og lyd essensiellt for gyting. Viktigste gyteområder konsentrert i tid og rom. | ja | |
| Nordsjøsild (Clupea harengus) | høy kommersiell og økologisk viktighet, svært god hørsel, konsentrerte gyteområder og perioder | ja | |
| Nordsjøsei (Pollachius virens) | Kommersiellt og økologisk viktig, god hørsel. | ja | |
| Blåkveite (Reinhardtius hippoglossides) | Livskraftig bestand, noe viktighet i økosystemet, med konsentrerte gytefelt og perioder | ja | |
| Brisling (Sprattus sprattus) | høy kommersiell og økologisk viktighet, mindre og konsentrerte gyteområder, svært god hørsel | ja | |
| Vanlig Uer (Sebastes norvegicus) | Nær truet bestand av noen kommersiell viktighet, påvist at seismikk kan føre til vertikal og horisontal unnvikelse | ja | |
| Blålange (Molva dipterygia) | Ikke veldig viktig kommersiellt eller økologisk, men en nær truet art, med konsentert gyteområde og periode | ja | |
| Hvitting (Merlangius merlangus) | Noe kommersiell viktighet, god hørsel, konsentrert gyteområde/periode | ja | |
| Øyepål (Trisopterus esmarkii) | Nøkkelart i økosystemet, god hørsel og gyteområde relativt konsentrert i tid og rom | ja | |
| Tobis (Ammodytus marinus) | høy kommersiell og økologisk viktighet, konsentrert gyteområder og periode | ja | |
| Laks (Salma salar) | Nær truet bestand, gyting foregår i ferskvann, men innvandringesruter kan være sårbare. Disse derimot ikke godt kartfestet. | nei | Inn- og utvandringsruter kan bli inkludert dersom disse blir bedre kartlagt |
| Lyr (Pollachius pollachius) | Livskraftig bestand, med god hørsel og lydproduksjon, men mangler kartlegging av gyteområder og kunnskap om gyteperiodens lengde | nei | kan bli inkludert dersom kartlagt konsentrerte gytefelt/perioder |
| Kveite (Hippoglossus hippoglossus) | Livskraftig bestand, men med relativt konsentert gyteområde og konsentert gytevandring, men dette ikke skikkelig kartlagt og uansett ikke i områder med vanlig seismikkaktivitet | nei | kan bli inkludert dersom kartlagt konsentrerte gytefelt/perioder |
| Ål (Anguilla anguilla) | Kritisk truet art, gyter ikke i norske farvann, men med konsentrerte gytevandringer opp i elver, men disse dårlig kartlagt | nei | kan bli inkludert, dersom får bedre kartlagt innvandringsruter |
| Makrell (Scomber scombrus) | gyter hovedsakelig utenfor norske farvann over en lang gyteperiode | nei | |
| Rødspette (Pleuronectes platessa) | Stort gyteområde, gyting over en lang periode, bruker ikke lyd og hørsel usikkert | nei | |
| Hestmakrell (Trachurus trachurus) | Livskraftig bestand med stort gyteområde og relativt lang gyteperiode. | nei | |
| Rognkjeks (Cyclopterus lumpus) | Livskraftig bestand med store gyteområder i områder utenfor vanlig seismikkaktivitet | nei | |
| Lysing (Merluccius merluccius) | Stort gyteområde, gyting over en lang periode, | nei | |
| Brosme (Brosme brosme) | Livskraftig bestand med stort gyteområde, liten kommersiell og økologisk betydning | nei | |
| Lange (Molva molva) | Livskraftig bestand med relativt stort gyteområde, men en relativt kort periode | nei | |
| Bergylt (Labrus begrylta) | Livskraftige bestand, med god hørsel og lydproduksjon i gytefasen, men store gyteområder (langs hele kysten) hvor seismikk ikke vanlig | nei | |
| Blåstål/rødnebb (Labrus mixus) | Livskraftige bestand, med god hørsel og lydproduksjon i gytefasen, men store gyteområder (langs hele kysten) hvor seismikk ikke vanlig | nei | |
| Bergnebb (Ctenolabrus ripestris) | Livskraftige bestand, med god hørsel og lydproduksjon i gytefasen, men store gyteområder (langs hele kysten) hvor seismikk ikke vanlig | nei | |
| Grøngylt (Symphodus melops) | Livskraftige bestand, med god hørsel og lydproduksjon i gytefasen, men store gyteområder (langs hele kysten) hvor seismikk ikke vanlig | nei | |
| Grasgylt (Centrolabrus exoletus) | Livskraftige bestand, med god hørsel og lydproduksjon i gytefasen, men store gyteområder (langs hele kysten) hvor seismikk ikke vanlig | nei | |
| Havnål (Entelurus aequoreus) | Livskraftig bestand med liten kommersiell og økologisk viktighet. | nei | |
| Vassild (Argentina silus) | av mindre økologisk betydning, stort gyteområde og en livskraftig bestand | nei | |
| Kolmule (Micromesistius poutassou) | Livskraftig bestand, med gyteområde utenfor norsk sone og derfor ikke aktuell for frarådingskart | nei | |
| Gråsteinbit (Anarchichas lupus) | Livskraftig bestand med antatt stort gyteområde og relativt lang gyteperiode. | nei | |
| Flekksteinbit (Anarhichas minor) | Livskraftig bestand med antatt stort gyteområde og relativt lang gyteperiode. | nei | |
| Blåsteinbit (Anarhichas denticalatus) | Livskraftig bestand med antatt stort gyteområde og relativt lang gyteperiode. | nei | |
| Makrellstørje (Thunnus thunnus) | Kommersiellt noe viktighet, men ikke nøkkelart i norske økosystemer, god hørsel, men gyteområder utenfor norsk sone. | nei |
7.2 - Grunnlag for rådgivning for hval
Tilsvarende som for fisk, gjennomgås også kunnskapsgrunnlaget for beiting hos bardehval sammen med ansvarlige fagpersoner årlig. Kart over viktige beiteområder oppdateres dersom ny kunnskap kommer til. På samme måte som for fisk vurderes det hvilke bestander som skal inkluderes i rådgivningen. Denne vurderingen tas på grunnlag av følgende:
Sesongbeiter:
Hvorvidt arten beiter hele året eller foretar intensivt beite kun deler av året. Noen hvalarter kommer til Barentshavet for å beite om sommeren på rike konsentrasjoner av zooplankton og fisk og fyller opp energilagrene til migrasjon til sørlige strøk hvor parring og kalving foregår, og beiter ikke under vandring og kalving/parring. Disse artene derfor særlig sårbare for å bli forstyrret i beiting, da selv små avbrekk kan gi konsekvenser. Andre hvalarter beiter kontinuerlig gjennom året, og er derfor mindre sårbare for forstyrrelser.
Bestandens tilstand: Basert på kategoriene fra rødlisten, er alle bestander kategorisert som regionalt utdødd/kritisk truet, sårbar/nær truet eller livskraftig. Særlig da bestander som er kritisk eller nær truet bør en være særlig påpasselig med at ikke utsettes på negativ påvirkning.
Økosystemkomponent: Om arten innehar en nøkkelrolle i økosystemet.
Lyd: Om arten aktivt selv bruker lyd i lave frekvenser eller om kjent negativ påvirkning av lavfrekvent lyd.
Hørsel: Om arten har god hørsel på lave frekvenser regnes den som mer sårbar for negativ påvirkning av lydforurensning på disse frekvensene.
Tabell A1.2 viser alle hvalbestander som er vurdert og hvilke vurderinger som er gjort, og hvilke som er inkludert i rådgivningskartene.
| Art | Vurderingsgrunnlag | Innkludert i fraråding | Kommentar |
|---|---|---|---|
| Knølhval (Megapthera novaeangliae) | Livskraftig bestand, sesongbeiter, hørsel og vokalisering i frekvensområde til seismikk, viktig i økosystemet | ja | |
| Finnhval (Balaenopthera physalus) | Livskraftig bestand, sesongbeiter, hørsel og vokalisering i frekvensområde til seismikk | ja | |
| Blåhval (Balaenopthera musculus) | Sterkt truet bestand, sesongbeiter, hørsel og vokalisering i frekvensområde til seismikk, | ja | |
| Vågehval (Balaenopthera acutorostrata) | Livskraftig bestand, ressurs for hvalfangst sesongbeiter, hørsel og vokalisering i frekvensområde til seismikk | ja | |
| Grønnlandshval (Baleana mysticetus) | Sterkt truet bestand, sesongbeiter, hørsel og vokalisering i frekvensområde til seismikk, sterk adferdsendring til seismikk | ja | |
| Nebbhval (Hyperoodon ampullatus) | Livskraftig bestand, ikke sesongbeiter, men vist ekstremt sensitivitet til lydforstyrrelse | nei | Bør inkluderes om får kartlagt bedre viktige beiteområder |
| spekkhogger (Orcinus orca) | Livskraftig bestand, ikke sesongbeiter, vokalisering, men på annet frekvensområde enn seismikk | nei | |
| Nise (Phocoena phocoena) | Livskraftig bestand, ikke sesongbeiter, vokalisering, men på annet frekvensområde enn seismikk | nei | |
| Kvitnos (Lagenorhynchus albirostris) | Livskraftig bestand, ikke sesongbeiter, vokalisering, men på annet frekvensområde enn seismikk | nei | |
| Hvithval (Delpnapterus leucas) | Sterkt truet, påvist negativ påvirket av seismikk, ikke sesongbeiter | nei | |
| Spermhval (Physeter macrosephalus) | Ikke sesongbeiter. Lydproduksjon i andre frekvensområder enn seismikk | nei |
7.2.1 - Beiteområder for bardehval
Kunnskap om beiteområder i Barentshavet kommer hovedsakelig fra årlige telletokt i regi av Havforskningsinstituttet, som viser utbredelsen av ulike arter i perioden juni-aug, i tillegg til det årlige økosystemtoktet i perioden aug.-okt. I tillegg er det gjennomført en del merking av hval med satelittmerker, som muliggjør sporing av dens bevegelse over tid. Forståelsen av utbredelse i tid og rom er ispedd observasjoner fra foto ID, rapporterte observasjoner fra fiskefartøy og andre vitenskapelige tokt uten dedikerte observatører, samt ved å kartlegge hvalturisme.
Knølhval. Denne arten har hatt en dramatisk økning i antall de siste fem år. I perioden juni-aug. finnes knølhvalen i store konsentrasjoner rundt Bjørnøya (Leonard og Øien 2020 a,b), mens på sensommeren (aug.-okt.) er den svært tallrik i området øst av Spitsbergen (Olgabassenget) (Mauritzen et al. 2011, WGIBAR 2018, 2020). I tillegg har det de siste 10 år blitt observert stadig mer hval som beiter også på vinterstid i norske farvann. Denne beitingen er knyttet til overvintrende sild i fjorder hovedsakelig i Troms. Denne er ikke kartlagt systematisk på samme måte som i Barentshavet, og vi har pr. i dag ikke godt nok kunnskapsgrunnlag til å kartfeste dette skikkelig i tid og rom. Derfor er disse områdene ikke inkludert i rådgivningskartene for 2023.
Finnhval. Beiter hovedsakelig vest av Spitsbergen, samt i området mellom Bjørnøya og Spitsbergen i perioden juni-aug (Leonard og Øien 2020a,b). Også finnhval finnes i stort antall på østsiden av Spitsbergen på sensommer og høst (aug-okt) (Mauritzen et al. 2011, WGIBAR 2020). En ny sammenstilling av merkedata for finnhval har identifisert enkelte områder sørvest av Spitsbergen, samt rundt Bjørnøya og Jan Mayen som viktige beiteområder for finnhval (Lydersen et al. 2020). Beiting i disse områdene foregår hovedsakelig frem til midten av september. Slike merkedata er særlig verdifulle da de i tillegg til å bare fortelle hvor hvalen er, kan brukes til å identifisere bevegelsesmønstre og hvilke type adferd den representerer, og derfor gir direkte informasjon om hvor hvalen faktisk beiter.
Vågehval har en større variasjon i høytetthetsområder, og har større variasjon i oppholdsområder fra år til år enn de to andre artene. De finnes hovedsakelig i de samme oppholdsområdene som de to overnevnte artene, men grunnet en mer opportunistisk diett ofte mer varierende (Skaug et al. 2014; Solvang et al. 2014). I 2020 var mer vågehval observert i østlige områder og mindre rundt Bjørnøya, muligens grunnet høyt beitepress rundt Bjørnøya fra store mengder knøl- og finnhval (Nils Øien, pers komm). Vågehvalen er en opportunistisk predator med en diett som inneholder både zooplankton og fisk (eks. Haug et al. 2002).
En sammenstilling av data for alle disse tre artene viser at de områdene hvor de er mest tallrike er rundt Svalbard og i nærheten av Jan Mayen (Hamilton et al. 2020).
7.3 - Rådgivningskart for seismikk og elektromagnetiske undersøkelser
7.3.1 - Endringer fra 2022 – 2023
Rådgivningskartene for 2023 har blitt oppdatert basert på ny kunnskap.
Flere av endringene som har blitt gjort har sammenheng med resultater fra KINOII prosjektet. En rask oppsummering av dette arbeidet er derfor gitt her. Egg og larve ble samlet inn ukentlig i en periode på 3 år fra 2017-2020 på fire posisjoner i Nordsjøen av forsyningsfartøy for oljeplattformer i samarbeid med Havforskningsinstituttet. Gyteperioden for mange arter ble kartlagt. Den relative mengden av egg og larver viste når det var mest intensiv gyting. Gyteperioden er dynamisk og kan endre seg fra år til år, og går ofte i svingninger, så et tre-års studie blir som et øyeblikksbilde, men likevel gir det nyttig kunnskap som vi har tatt med i rådgivningen i samråd med de bestandsansvarlige. I noen tilfeller bekrefter denne studien tidligere kunnskaper, i andre tilfeller tilføyer den ny kunnskap som fører til endringer av råd. Figur A1.1 viser innsamlingsområde og eksempel på resultat.
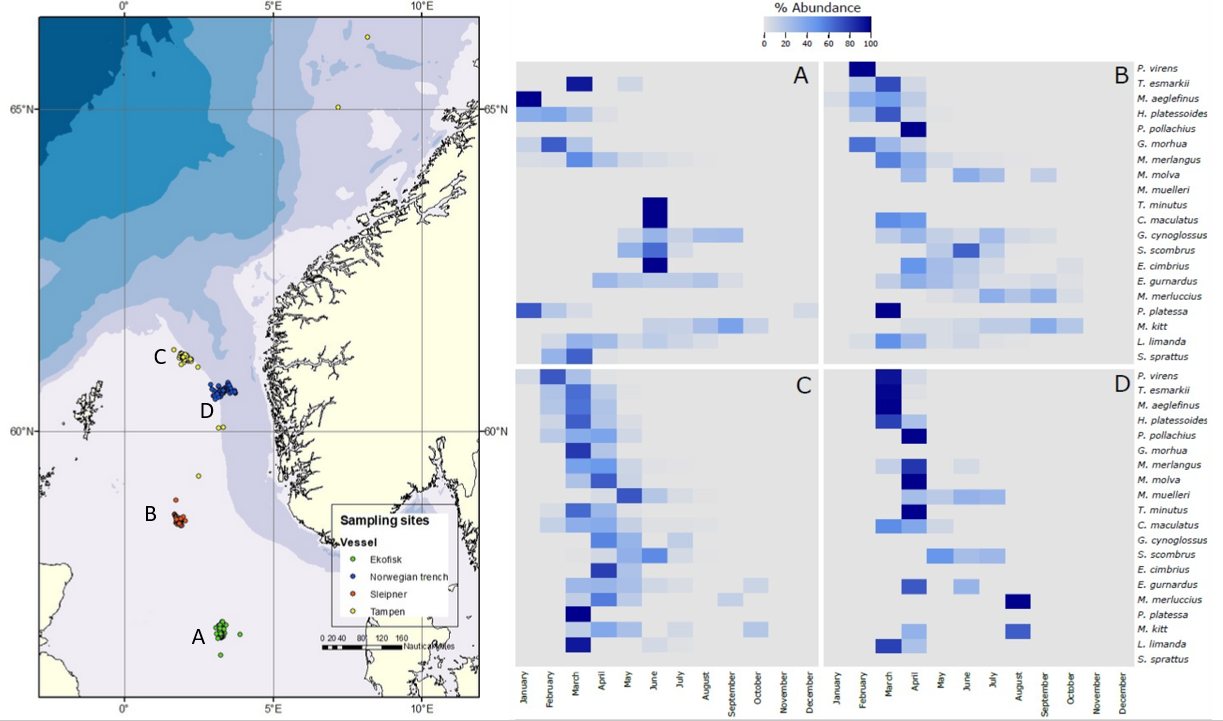
Her er en oversikt over endringene som har blitt gjort.
Ny art: Hvitting
Hvitting er en fisk i torskefamilien som gyter i Nordsjøen, som tidligere ikke har vært med hi rådgivningskartene grunnet stor usikkerhet knyttet til gyteperioden, og antatt lang gyteperiode. KINO II har gitt ny kunnskap om gyteperioden til hvitting (Krafft et al. 2021). Basert på dette blir tre av de konsentrerte gyteområdene til hvitting som ligger nær områdene for KINOII undersøkelsene tatt med i rådgivningskartene. Det nordligste området blir frarådet fra 1. mars – 30 april, og de to sørlige blir frarådet fra 1. mars – 31. mars basert på Krafft et al. 2021.
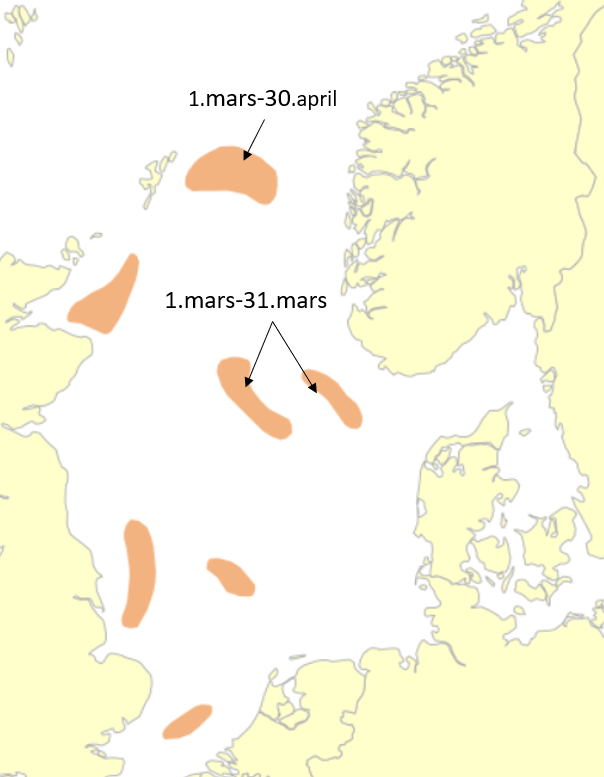
Oppdatert rådgivningsperiode: Nordsjøtorsk
Rådgivningskartet for nordsjøtorsk ble oppdatert i 2021 (Sivle et al. 2022). Ny kunnskap om gyteperioden fra KINOII gjør rådgivningsperioden endres. For nordsjøtorsk viste KINO II resultatene at gytingen starter tidligere i sør enn i nord, derfor har vi delt opp området i to perioder. 1. januar-31. mars i sør, og 1. februar til 31. mars i nord. Tidligere råd var 1. januar – 31. mars for alle områdene.
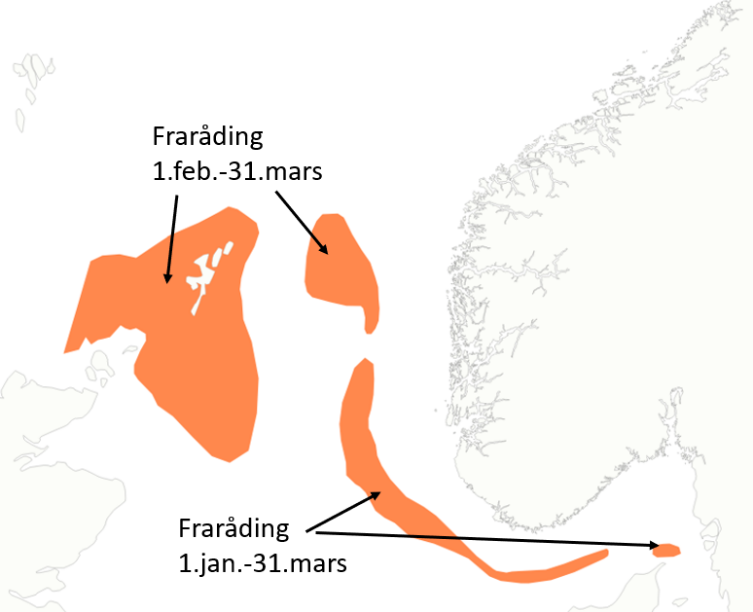
Oppdatert rådgivningsperiode: Øyepål
Basert på resultat fra KINOII prosjektet (Krafft et al. 2021) blir rådgivningsperioden for øyepål endret til 1.februar til 15. april. Tidligere periode var 1. jan til 30. april. Selve kartlaget er ikkje endret.
Oppdatert gytekart: Kysttorsk
Kartlegging av gytefelt i kystsonen er pågåande arbeid, hvor to årlige tokt dekker ulike områder i en rotasjon. Det tar 5-7 år å dekke hele Norgeskysten. Data om nye gytefelt blir levert til Fiskeridirektoratet og blir oppdatert på deres kartserver Yggdrasil. Havforskningsinstituttet har samkjørt sine kart for kysttorsk med disse. I løpet av 2022 har det kommet inn mer data som har blitt inkludert i rådgivningskartet for 2023. I områder som er definert som nasjonalt eller regionalt viktige og samtidig ligger ubeskyttet til langs kysten (ikke dekket av holmer og skjær) inngår i rådgivningskartene for seismikk.
Siden bare gytefeltene som ligger ubeskyttet i forhold til seismikk i havet er med i rådgivningskartene for kysttorsk må ikke dette kartlaget brukes i forbindelse med andre typer aktiviteter, som for eksempel sprengningsarbeid i fjorder.
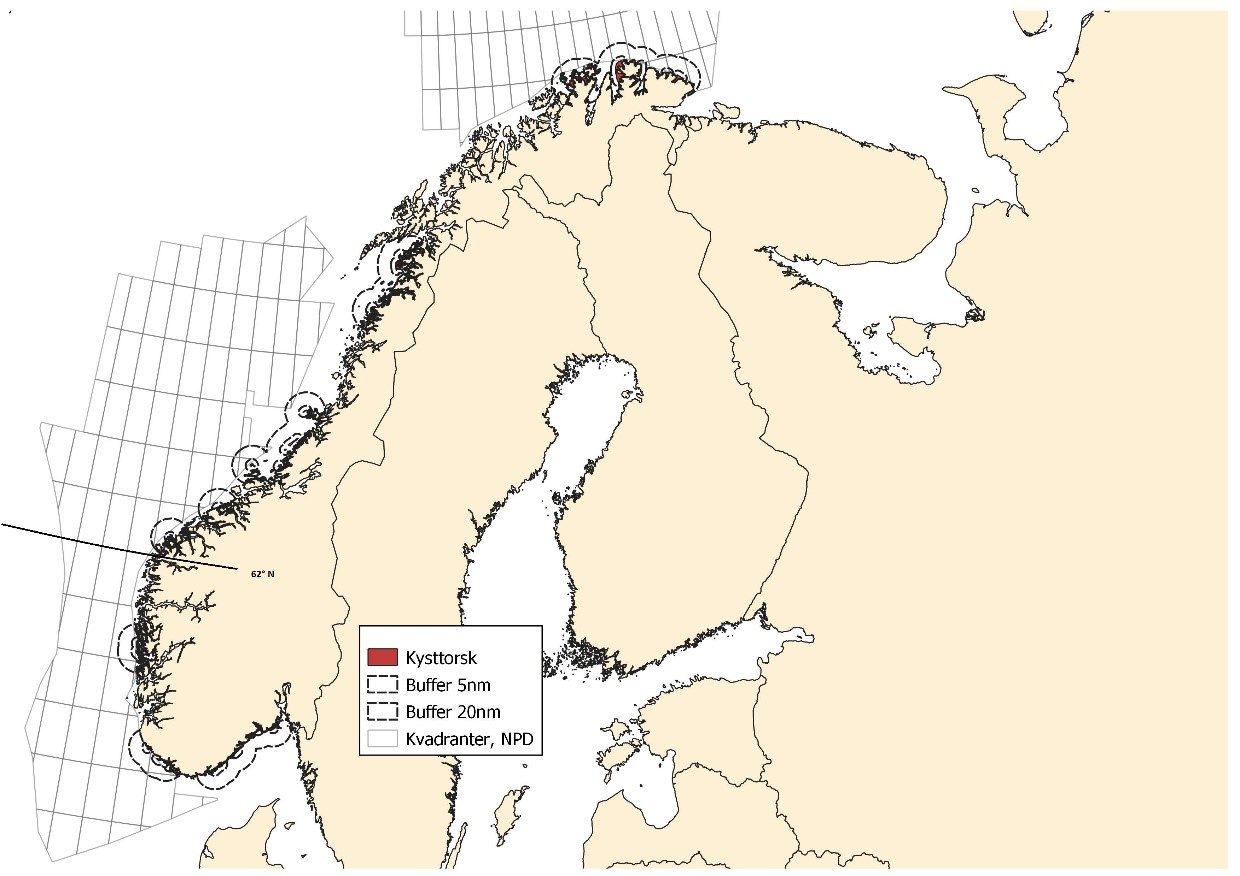
7.3.2 - Rådgivningskart
Siden 2018 har Havforskningsinstituttet benyttet rådgivningskart som viser for hvilke områder og perioder det er frarådet å drive med seismikk eller elektromagnetiske undersøkelser. Rådgivningskartene er sammenslått av ulike kartlag, som representerer et gyte- eller beiteområde. Rådgivningskartene er laget med en gyldighetsperiode på 2 uker, og kartlagene utgjør da gyte/beitekartene for de bestander som gyter/beiter i denne 2 ukers perioden. For eksempel består rådgivningskart for 1.-15. januar av gytekartene til alle bestander fra listen som gyter i perioden 1.-15. januar; (nordsjøtorsk, sei, kysttorsk sør, tobis, blåkveite og polartorsk), fisk som har gytevandring i denne perioden (NEA torsk) og hval som har intensivt beite i perioden (ingen) (Tabell A1.3). Rundt alle rådgivningskartene ligger også en buffersone på 20 og 5 nmi, som brukes for henholdsvis ordinære seismikkundersøkelser og borestedsundersøkelser.
Bestander og kartlag som rådgis for, og er inkludert i rådgivningskartene for 2023 er angitt i Tabell A1.3
| Seismikk/EMU | Art | Sårbar aktivietet | Periode for fraråding |
|---|---|---|---|
| seismikk | Nordsjøtorsk, vest | Gyting | 1. feb. - 31. mar. |
| Seismikk | Nordsjøtorsk, øst | Gyting | 1. jan. - 31. mar. |
| Seismikk | Nordsjøsei | Gyting | 1. jan. - 31. mar. |
| Seismikk | Øyepål | Gyting | 1.feb. - 15. apr. |
| Seismikk | Kysttorsk sør | Gyting | 1. jan. - 30. apr. |
| Seismikk | Polartorsk | Gyting | 1. nov. - 28. feb. |
| Seismikk | Tobis | Gyting | 1. des .-31. jan. |
| Seismikk | Blåkveite | Gyting | 1. des. -31. jan. |
| Seismikk | NVG Sild | Gytevandring, Trænabanken | 16. jan. - 31. jan |
| Seismikk | NVG Sild | Gytevandring langs kysten | 16. jan. - 31. mar. |
| Seismikk | NVG Sild | Gyting | 1. feb. - 31. mar. |
| Seismikk | Lodde | Gytevandring | 1. feb - 15. mar. |
| Seismikk | NEA sei | Gyting | 1. feb - 31. mar. |
| Seismikk | Nordsjøhyse | Gyting | 1. feb - 30. apr. |
| Seismikk | Kysttorsk nord | Gyting | 1. feb - 15. mai |
| Seismikk | Lodde | Gyting | 16.feb - 15. apr. |
| seismikk | Hvitting nord | Gyting | 1. mar. - 30. apr. |
| Seismikk | Hvitting sør | Gyting | 1.mar. - 31. mar |
| Seismikk | Vanlig Uer | Gyting | 1. mar. - 30. apr. |
| Seismikk | NEA hyse | Gyting | 1. mar. - 15. mai. |
| Seismikk | NEA Torsk | Gyting | 16.mar - 30. apr. |
| Seismikk | Brisling | Gyting | 1. mai - 30. jun. |
| Seismikk | Blålange | Gyting | 16. mai - 15. jun. |
| Seismikk | Hval | Beiting | 1. jul. - 31. okt. |
| Seismikk | NEA Torsk | Gytevandring | 1. jan. - 31. mar. |
| EMU | NEA Torsk | Gytevandring | 1. jan. - 31. mar. |
| EMU | NVG Sild | Gytevandring, Trænabanken | 16. jan -31. jan. |
| EMU | NVG Sild | Gytevandring langs kysten | 16. jan - 31. mar. |
| EMU | Lodde | Gytevandring | 1. feb. - 15. mar. |
Figurene A1.5-A1.7 viser alle rådgivningskartene for seismikk for 2023. Buffersonene på 5 og 20 nmi er inkludert. Rutenettet i figurene viser blokker som er åpne for oljeleting.
Figur A1.8 viser rådgivningskart for elektromagnetiske undersøkelser. Etter 31.mars er det ingen fraråding for denne typen undersøkelser.
Figur A1.9 viser kartlag for hvor det er tilrådet å ha en dedikert observatør for å sikre at det ikke skytes seismikk i nærheten av grønlandshval
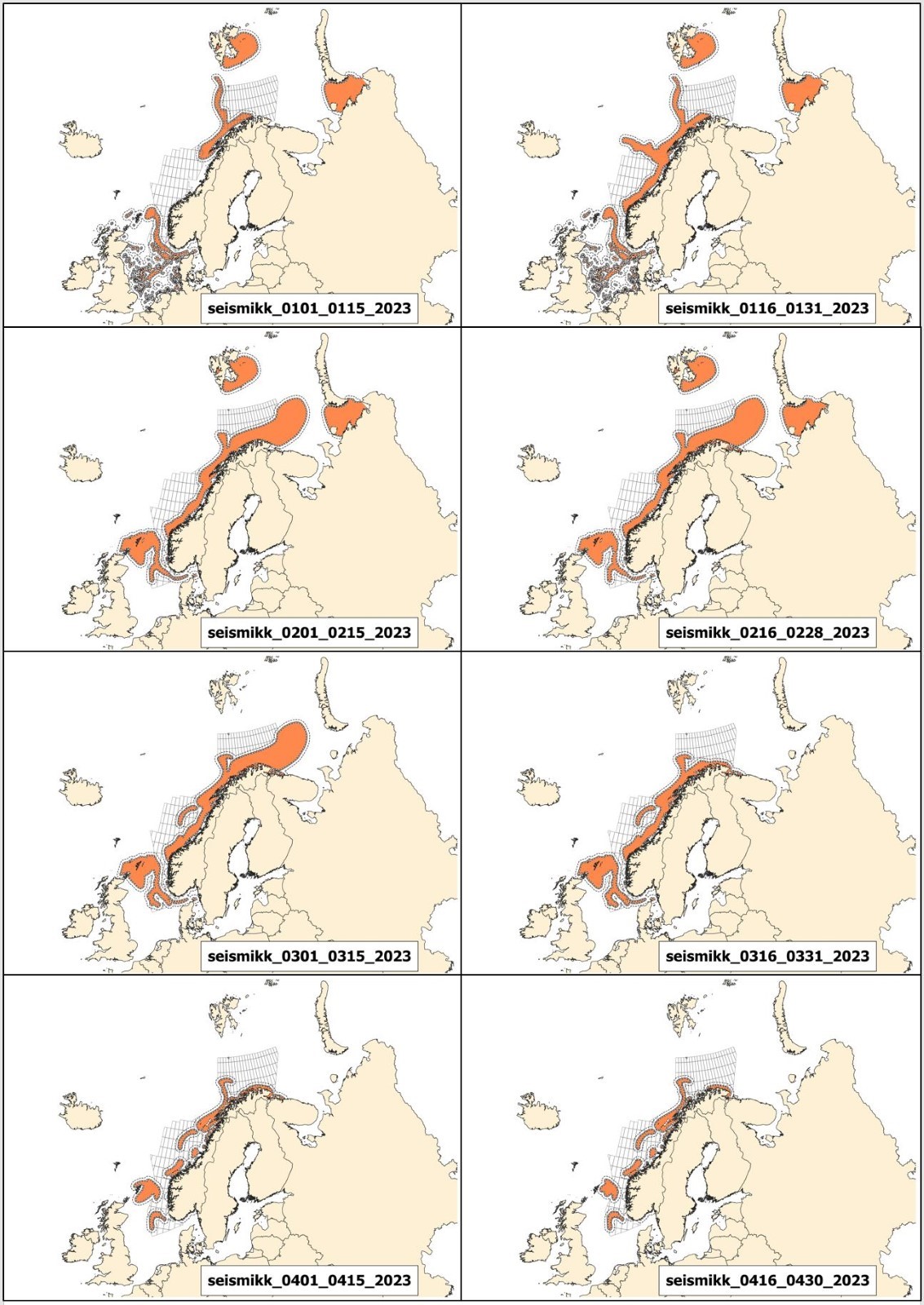
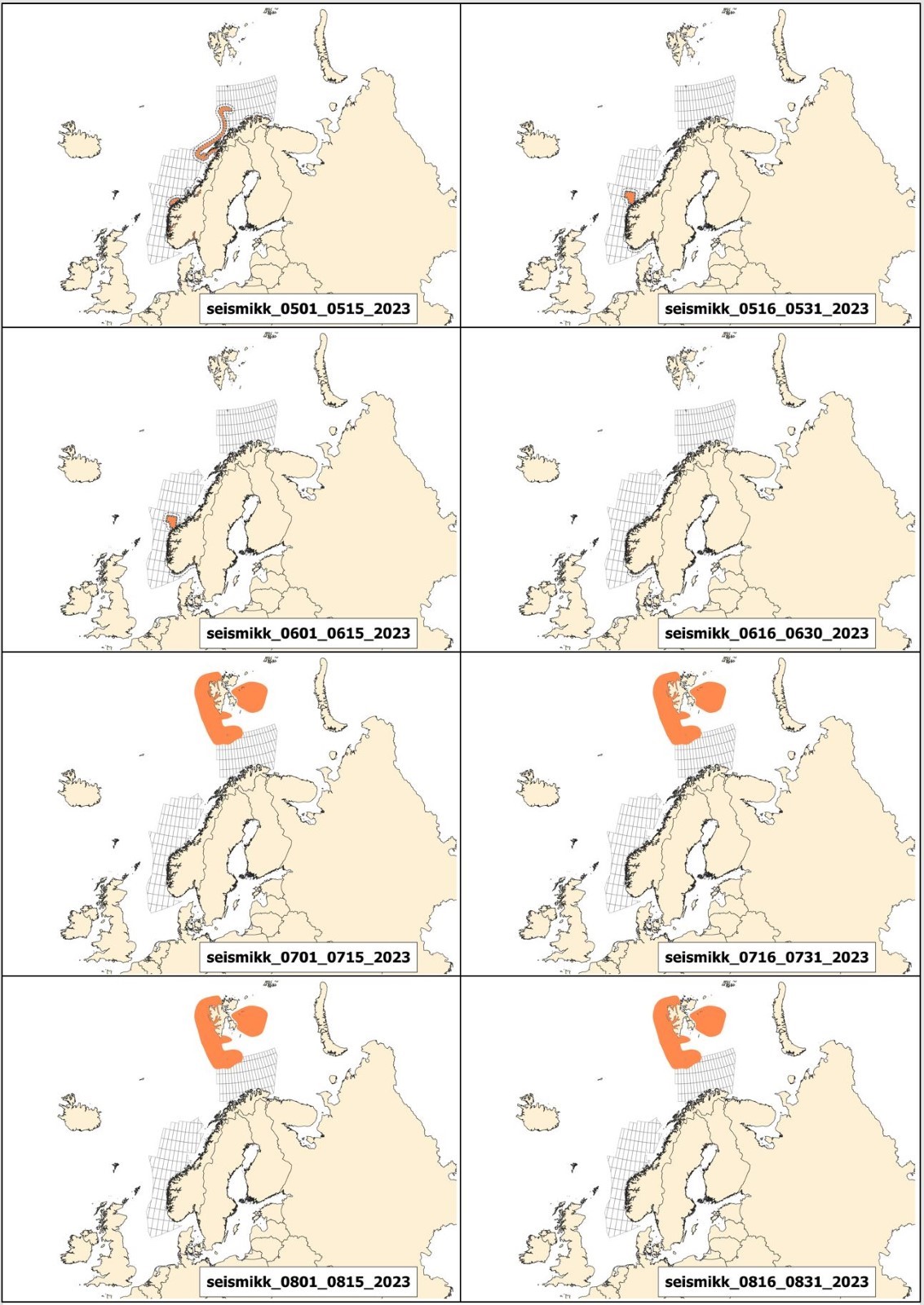
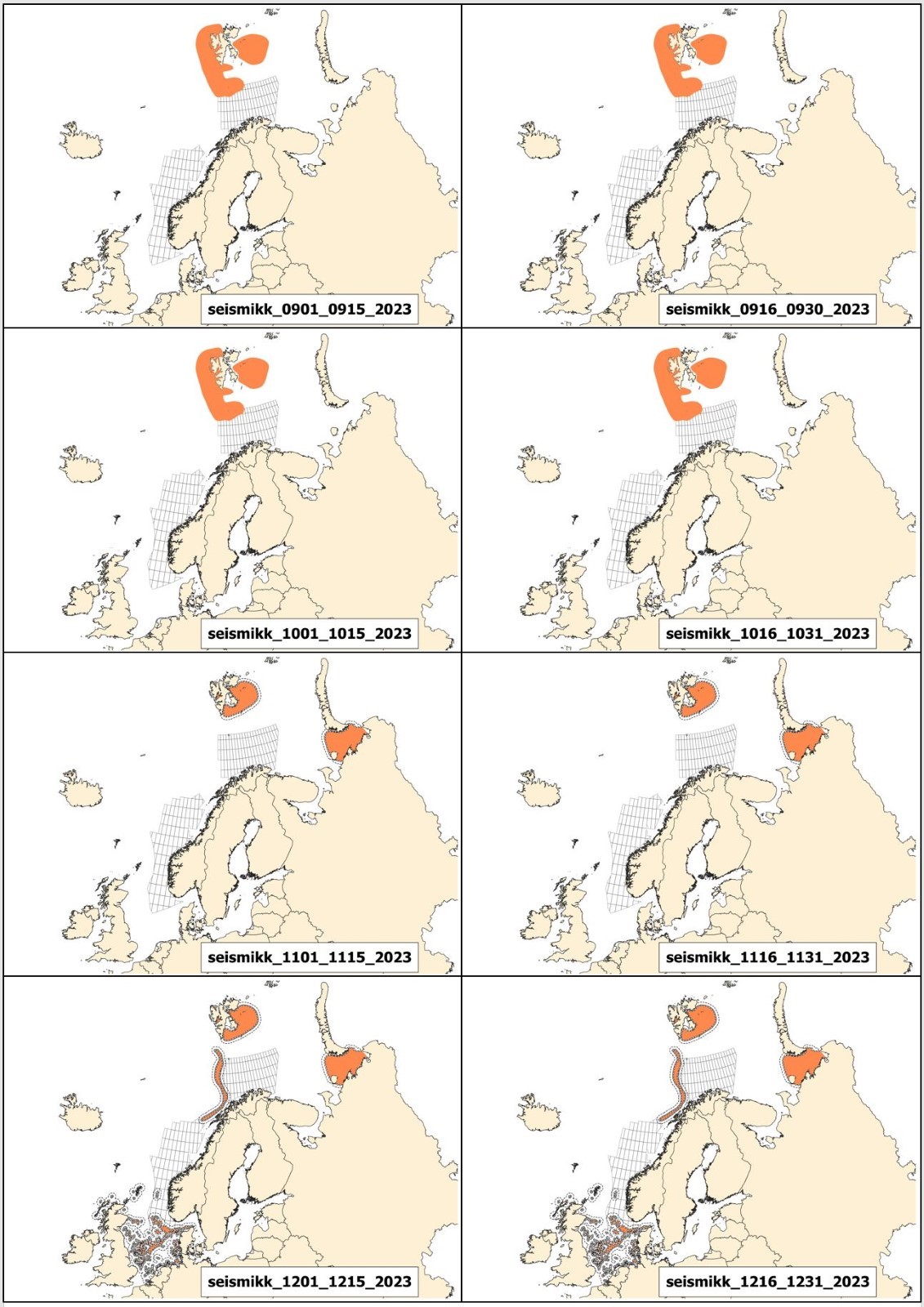
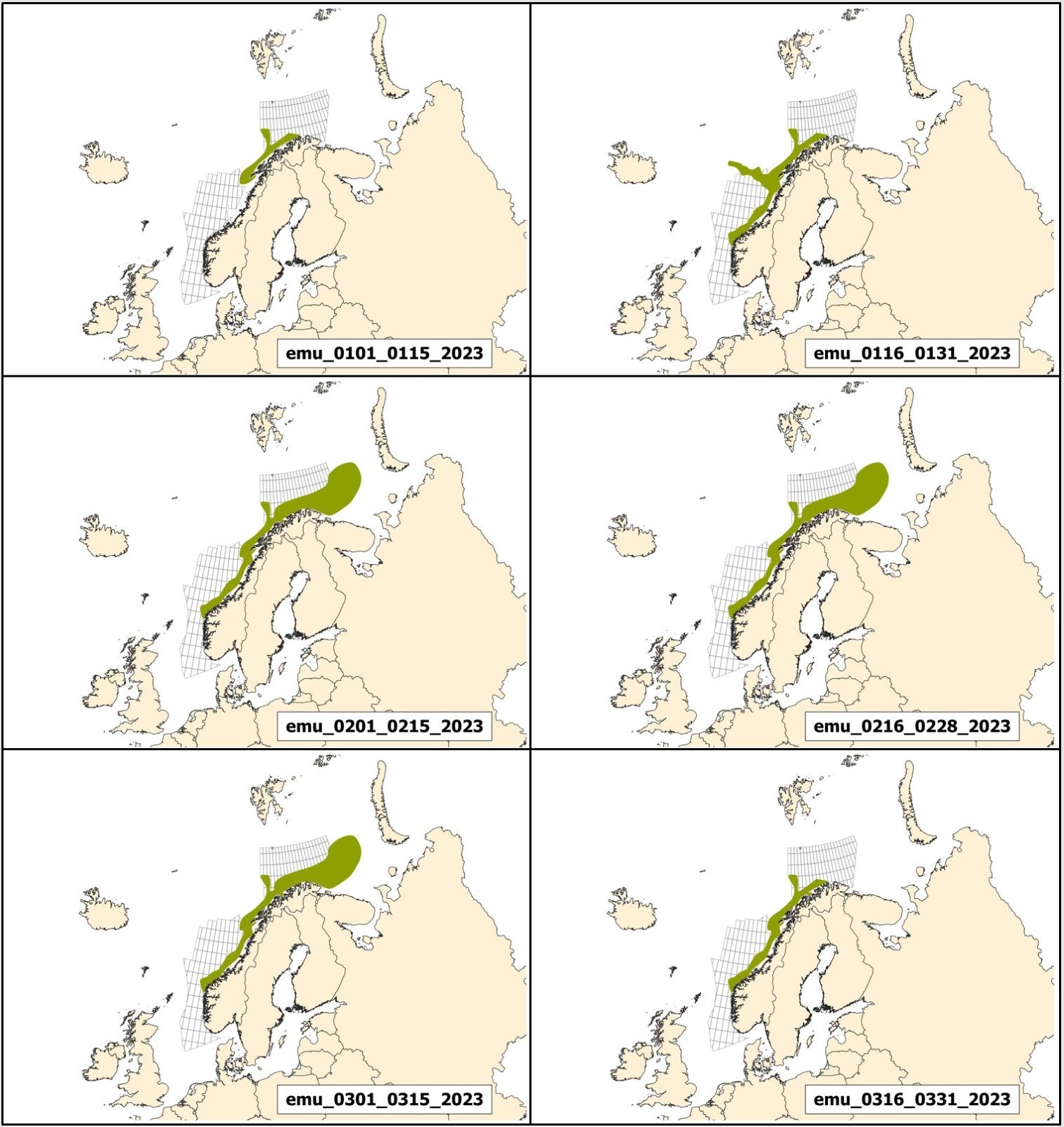
7.3.3 - Tilgang til rådgivningskart
Rådgivningskartene representerer Havforskningsinstituttet sine råd for undersøkelser til havs som bruker seismiske eller elektromagnetiske kilder. Kartene inngår i Oljedirektoratet sitt meldesystem der slike undersøkelser må registreres og saksbehandles. Seismikkselskaper og andre ønsker gjerne tilgang til digitale versjoner av disse kartene utenom meldesystemet, blant annet til bruk i planlegging av undersøkelser. Vi har derfor her gitt en beskrivelse av hvordan man kan få tilgang til disse
Her er informasjon om hvor og hvordan en kan se på eller laste ned rådgivningskartene.
Se på rådgivningskartene:
Kartene publiseres på HI sine kartdata-sider http://www.imr.no/geodata/geodataHI.html så snart som mulig, og ligger under menyvalget «Rådgivningskart 2023»
Se på og lagre rådgivningskartene som tjenester:
WMS-lenke: https://kart.hi.no/data/fraraadning/wms
WFS-lenke: https://kart.hi.no/data/fraraadning/wfs
Laste ned rådgivningskartene som shapefiler:
Det er mulig å laste ned alle kartene som en shapefil ved å følge denne lenken: https://kart.hi.no/data/ows?service=WFS&version=1.0.0&request=GetFeature&typeName=fraraadning%3Araadgivningskart_2023&maxFeatures=50000&outputFormat=SHAPE-ZIP
Shapefilen åpnes med et GIS-program (f.eks. ArcGIS eller QGIS).
Metadata-oppføring i Geonorge:
For teknisk informasjon om kartene, produktark, lisensinformasjon etc. kan man finne metadataoppføring i Geonorge sin kartkatalog her:
Datagrunnlaget for rådgivningsområdene (gytefelt og beiteområder med artsinformasjon) :
For å se på grunnlaget for områdene (gyte- og beiteområder) som er inkludert i rådgivningskartene for 2023 kan man bruke følgende lenke: https://kart.hi.no/data/fraraadning/wms?service=WMS&version=1.1.0&request=GetMap&layers=fraraadning%3Aomraader_inkludert_i_raadgivningskart_2023&bbox=-7.58931701699993%2C51.770196722%2C60.5965468019121%2C80.7907060000001&width=768&height=330&srs=EPSG%3A4326&styles=&format=application/openlayers
Man kan laste ned filen i shapefil-format her: https://kart.hi.no/data/ows?service=WFS&version=1.0.0&request=GetFeature&typeName=fraraadning%3Aomraader_inkludert_i_raadgivningskart_2023&maxFeatures=50000&outputFormat=SHAPE-ZIP
Eller bruke følgende WFS-lenke: https://kart.hi.no/data/fraraadning/omraader_inkludert_i_raadgivningskart_2023/wfs
7.4 - Referanser
Hamilton, CD., Lydersen, C., Aars, J., Biuw, M., Boltunov, A.N……….Kovacks, K.M. (2020). Marine mammal hotspots in the Greenland and barents Seas. Mar. Ecol. Prog. Ser. 659: 3-28 https://doi.org/10.3354/meps13584
Haug, T., Lindstrøm, U., & Nilssen, K. T. (2002). Variations in minke whale (Balaenoptera acutorostrata) diet and body condition in response to ecosystem changes in the Barents Sea. Sarsia: North Atlantic Marine Science, 87(6), 409-422.
Krafft, B. A., Mateos-Rivera, A., Mozfar, B., Skern-Mauritzen, R., Dahle, G., Sundby, S., ... & Wehde, H. (2021). Assessment of Fish Spawning in the North Sea; Final Project Report of KINO-2, 2017–2021. Rapport fra havforskningen.
Leonard, D. M. & Øien N. (2020a). Estimated abundances of cetacean species in the Northeast Atlantic from Norwegian Shipboard surveys conducted in 2002-2013. NAMMCO Scientific publications 11. https://doi.org/ 10.7557/3.4695
Leonard, D. M. & Øien N. (2020b). Estimated abundances of cetacean species in the Northeast Atlantic from Norwegian Shipboard surveys conducted in 2014-2018. NAMMCO Scientific publications11. https://doi.org/ 10.7557/3.4694
Lydersen, C., Vacquié-Garcia, J., Heide-Jørgensen, M. P., Øien, N., Guinet, C. and Kovacs, K. M. 2020. Autumn movements of fin whales (Balaenoptera physalus) from Svalbard, Norway, revealed by satellite tracking. Sci. Rep. 10, 16966: 1-13. https://doi.org/10.1038/s41598-020-73996-z
Mauritzen, M., Johannesen E., Bjørge, A. and Øien, N. 2011. Baleen whale distributions and prey associations in the Barents Sea. Mar. Eco Prog Ser Vol. 426: 289–30. doi: 10.3354/meps09027
Sivle, L.D.; Forland, T.N.; de Jong, K.; Nyqvist, D.; Grimsbø, E., Kutti., T and Wehde, H. Havforskningsinstituttets rådgivning for menneskeskapt lyd i havet – kunnskapsgrunnlag, vurderinger og råd for 2022. Report from Institute of Marine Research, Bergen, Norway, 2022-1, ISSN: 1893-4536 https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-2022-1
Skaug, H.J., Øien, N., Schweder, T., Bøthun, G. 2014. Abundance of minke whales (Balaenoptera acutorostrata) in the Northeast Atlantic: variability in time and space. Can.J.Fish.Aquat.Sci. 61:871-886. https://doi.org/10.1016/j.marpolbul.2015.12.044.
Solvang, H., Skaug, H.J., Øien, N. 2014. Abundance of minke whales in the Northeast Atlantic based on survey data collected over the period 2008-20013. Report.
WGIBAR 2020. Annex 4: The state and trends of the Barents Sea ecosystem in 2019 ICES SCIENTIFIC REPORTS 2:30.
8 - Vedlegg II: Korallrev og vernede områder - hvorfor det er viktig å beskytte revet mot utstyr som blir plassert på bunnen?
Den norske sokkelen, fra Stadt opp til Ingøydjupet vest for Nordkapp, er på mange måter karakterisert av sine utallige og ekstremt velutviklede korallrev, bygget av arten Lophelia pertusa . Lophelia er den vanligste revdannende dypvannskorallen i verden, men arten er særlig vanlig på norsk sokkel. Av alle registrerte forekomster av Lophelia globalt, så utgjør de norske 30%. Norge er derfor å regne som et kjerneområde for arten.
Korallrev fyller flere av de kriterier som definerer et sårbart økosystem. Korallrevene har en stor funksjonell signifikans ettersom de er levested for tusentalls andre arter og spiller en svært viktig rolle i karbonsyklusen i havet (Freiwald et al., 2012, Cathalot mfl 2015). Lophelia har lav motstandsdyktighet mot lokal endring i miljøforhold fordi den er fastsittende. I tillegg har arten en lav evne til å bygge opp ny bestand etter endt påvirkning. Den har uregelmessig rekrutering og langsom vekst (en korallkoloni vokser omtrent 1 cm per år), det vil derfor ta 100 til 1000-tals år å bygge opp nye populasjoner eller rev hvis de ødelegges (Mortensen & Rapp 1998, Rogers 1999, Roberts 2009, Brook & Järnegren 2013).
Revene er skjøre og mange rev har tidligere blitt ødelagt av bunntråling med tungt bunngear (Hall-Spencer et al., 2002, Fosså et.al., 2002). Korallrev bygget av Lophelia står derfor på Norsk Rødliste for naturtyper og er listet som et truet og minkende habitat av OSPAR kommisjonen. Allerede i 1999 etablerte Fiskeriforvaltningen tiltak for å beskytte revene mot ødeleggelse av bunnfiske. I 2004 hadde 9 korallrevområder fått særskilt beskyttelse mot bunntråling gjennom utøvelsesforskriftens § 66 og i 2016 ble dette utvidet til å gjelde 18 områder (Fosså et al., 2015). I tillegg står det i forskriften: For å beskytte korallrev mot ødeleggelser som følge av fiskeriaktivitet, og derigjennom bidra til en forsvarlig ressursforvaltning blant annet ved å sikre reproduksjons- og oppvekstområder for mange fiskeslag, må det utvises særlig aktsomhet ved fiske i nærheten av kjente forekomster av korallrev. Det er forbudt å ødelegge rev med hensikt.
Havforskningsinstituttet fraråder ikke seismikkundersøkelser i områdene med koraller, men for undersøkelser som skal plassere utstyr på bunnen i disse områdene råder vi til å bruke visuelt utstyr for å være sikker på at en ikke ødelegger koraller.
8.1 - Referanser
Brooke S, Järnegren J (2013) Reproductive periodicity of the scleractinian coral Lophelia pertusa from the Trondheim Fjord, Norway Journal of Marine Biology 160:139-153.
Cathalot C, Van Oevelen D, Cox T, Kutti T, Lavaleye M, Duineveld G, Meysman FJR (2015) Cold-water coral reefs: hotspots of benthic carbon cycling in the deep-sea. Frontiers in Marine Science 2:37. doi:10.3389/fmars.2015.00037
Fosså JH, Mortensen PB, Furevik D (2002) The deep-water coral Lophelia pertusa in Norwegian waters: distribution and fishery impact. Hydrobiologia 471:1-12.
Fosså JH, Kutti T, Buhl-Mortensen P, Skjoldal HR (2015) Vurdering av Norske korallrev. Rapport fra Havforskningen 8-2015, 64 pp.
Freiwald A, Beuck L, Wisshak M (2012) Korallenriffe im kalten Wasser des Nordatlantiks - Entstehung, Artenvielfalt und Gefährdung. In: Beck E (ed) Die Vielfalt des Lebens. Wiley-VCH, Weinheim, pp 89-96.
Hall-Spencer J, Allain V, Fosså JH (2002) Trawling damage to Northeast Atlantic ancient coral reefs. Proceedings of the Royal Society of London B 269: 507-511.
Mortensen PB, Rapp HT (1998) Oxygen and carbon isotope ratios related to growth line patterns in skeletons of Lophelia pertusa (L)(Anthozoa:Scleractinia): implications for determination of linear extension rates Sarsia 83:433-446.
Roberts, JM (2009) Cold-water corals: the biology and geology of deep-sea coral habitats, Cambridge University Press.
Rogers AD (1999) The biology of Lophelia pertusa (Linnaeus 1758) and other deep-water reef-forming corals and impacts from human activities. International Review of Hydrobiology 84:315-406. doi: 10.1002/iroh.199900032