Særlig verdifulle og sårbare områder (SVO) i norske havområder - Miljøverdi
—
En gjennomgang av miljøverdier og grenser i eksisterende SVO og forslag til nye områder
Author(s):
74 medforfattere - se kapittel 15
Editor(s):
Elena Eriksen (HI), Gro I. van der Meeren (HI), Bente M. Nilsen (HI), Cecilie H. von Quillfeldt (NP) and Hanne Johnsen (NP)
Particularly valuable and vulnerable areas (SVO) in Norwegian seas - Environmental values
Assessment of environmental values and borders of present SVOs and proposals for new areas
Forord
Det er utviklet forvaltningsplaner for alle norske havområder, det vil si Barentshavet og havområdene utenfor Lofoten (St.meld. nr. 8 (2005-2006), oppdatert Meld. St. 10 (2010-2011)), Norskehavet (St.meld. nr. 37 (2008-2009), oppdatert Meld. St. 35 (2016-2017)) og Nordsjøen og Skagerrak (Meld. St. 37. (2012-2013)). I alle planene er det identifisert særlig verdifulle og sårbare områder (SVO) på grunnlag av ulike fagutredninger. I april 2020 la regjeringen fram en stortingsmelding som oppdaterer forvaltningsplanene for alle havområdene (Meld. St. 20 (2019-2020)). Det faglige grunnlaget for meldingen viste at det var behov for harmonisering av SVO-vurderinger på tvers av havområdene og en helhetlig gjennomgang av alle SVO-ene for blant annet å synliggjøre årsak til verdi og sårbarhet. I stortingsmeldingen står det at en slik gjennomgang av miljøverdi og sårbarhet for alle SVO-ene skal sluttføres i løpet av 2021. Samtidig skal det utredes om områder med undersjøiske fjell oppfyller kriteriene for særlig verdifulle og sårbare områder.
På oppdrag fra Faglig forum har Havforskningsinstituttet ledet en ekspertgruppe som har gjennomført første del av denne oppgaven, dvs. miljøverdivurderingene. Tidsplanen er nå justert, men hele gjennomgangen, inkludert andre og tredje del av denne oppgaven som omfatter sårbarhetsvurdering og beskrivelse av næringsaktiviteter innenfor de særlig verdifulle områdene, skal være gjennomført i løpet av våren 2022. Rapportene fra dette arbeidet vil inngå som underlagsrapporter til det faglige grunnlaget for stortingsmeldingen om helhetlige forvaltningsplaner for de norske havområdene i 2024. Det er Faglig forum som skal utarbeide det faglige grunnlaget og identifisere de særlig verdifulle og sårbare områdene.
Arbeidet med miljøverdivurderingene som er presentert i denne rapporten har vært gjennomført av en tverrfaglig ekspertgruppe med fageksperter fra Havforskningsinstituttet, Norsk Polarinstitutt, Norsk institutt for naturforskning, Norges geologiske undersøkelse, Akvaplan-niva, og universitetene i Bergen, Oslo og Tromsø.
Dette er en faktarapport som avgrenser, beskriver og begrunner hvordan et utvalg av områder er funnet å ha miljøverdier som møter kriteriene for særlig verdifulle områder. Etter ønske fra Faglig forum ble EBSA-kriteriene1 i Konvensjonen om biologisk mangfold (CBD) lagt til grunn for miljøvurderingene.
Oppdatering 02.07.2021. Endring i rekkefølge av redaktører: Elena Eriksen (HI), Gro I. van der Meeren (HI), Bente M. Nilsen (HI), Cecilie von Quillfedt (NP) og Hanne Johnsen (NP)
Change in listed editors: Elena Eriksen (HI), Gro I. van der Meeren (HI), Bente M. Nilsen (HI), Cecilie von Quillfedt (NP) and Hanne Johnsen (NP)
1 - Sammendrag
I forvaltningsplanene for alle de norske havområdene, dvs. Barentshavet og havområdene utenfor Lofoten, Norskehavet og Nordsjøen/Skagerrak, er det identifisert særlig verdifulle og sårbare områder (SVO). SVO er områder som har vesentlig betydning for det biologiske mangfoldet og den biologiske produksjonen i havområdet, ofte også utenfor områdene selv. SVO-er gir ikke direkte virkninger i form av begrensninger for næringsaktivitet, men signaliserer viktigheten av å vise særlig aktsomhet i disse områdene.
Det faglige grunnlaget i 2019 for stortingsmeldingen om helhetlige forvaltningsplaner for de norske havområdene (Meld. St. 20 (2019-2020), viste at det var behov for en helhetlig gjennomgang av alle SVO-ene i de norske havområdene. I stortingsmeldingen står det at en slik gjennomgang av miljøverdi og sårbarhet for alle SVO-ene skal sluttføres i løpet av 2021. Samtidig skal det utredes om områder med undersjøiske fjell oppfyller kriteriene for særlig verdifulle og sårbare områder.
På oppdrag fra Faglig forum har en tverrfaglig ekspertgruppe med deltagere fra åtte forskningsinstitusjoner , ledet av Havforskningsinstituttet, gjennomført første del av denne oppgaven, dvs. miljøverdivurderingene. Resultatene fra dette arbeidet presenteres i denne rapporten. Tidsplanen er nå justert, men hele gjennomgangen, inkludert andre og tredje del av denne oppgaven som omfatter sårbarhetsvurdering og beskrivelse av næringsaktivitetene innenfor de særlig verdifulle områdene, skal være gjennomført i løpet av våren 2022.
Siden denne første rapporten kun vurderer miljøverdiene, ikke sårbarhet, har vi valgt å bruke betegnelsen foreslått SVO gjennom hele rapporten, for nye og endrete områder som foreløpig ikke har vært gjenstand for sårbarhetsvurderinger. Tidligere arbeid med SVO-er har vist at det er forekomst av miljøverdier som er avgjørende for avgrensning av områdene, ikke sårbarhet. Dessuten blir det da også lettere i forhold til omtale av eksisterende SVO-er.
Med utgangspunkt i tidligere miljøverdivurderinger og ny kunnskap, har ekspertgruppen utført miljøverdivurderinger basert på kriterier definert i FNs Konvensjon for biologisk mangfold (CBD) for å vurdere økologiske eller biologiske viktige områder (Ecologically or Biologically Significant marine Areas in need of protection, in open ocean waters and deep sea habitats, EBSA). På dette grunnlaget kom ekspertgruppen fram til et forslag med et revidert utvalg av SVO-er.
Den foreliggende rapporten er en fagrapport, der de foreslåtte SVO-ene er basert på en omfattende kunnskapssammenstilling om miljøverdiene, og hvor EBSA-kriteriene er brukt i verdivurderingene som ligger til grunn for avgrensning av områdene.
Ofte er det abiotiske forhold som topografi, strøm, vannmiljø eller bunnforhold som er avgjørende for økologiske prosesser og som gjør områder særlig viktig for enkelte økosystemkomponenter, men også for økosystemer som helhet. Eksempler på dette er bankområder, oppvellingsområder, retensjonsområder og isdekke.
Det er viktige områder for gyting/fødsel/hekking, beiting, overvintring og myting/hårfelling, samt viktige områder for truete eller sårbare arter og naturtyper som særlig har vært avgjørende for lokalisering av foreslåtte SVO-er i forvaltningsplanområdene. Slike områder er viktige for økosystemenes struktur og funksjon og reflekterer ofte områder med stor betydning for produksjon og biologisk mangfold, ofte også utenfor områdene selv.
I den foreliggende vurderingen ble alle de sju EBSA-kriteriene (Unikhet/Sjeldenhet, Livshistorisk viktige områder, Viktighet for truede eller nedadgående arter og/eller habitater, Sårbarhet, skjørhet, følsomhet, eller lav restitusjonsevne, Viktighet for biologisk produktivitet, Viktighet for biologisk mangfold og Naturlighet) vurdert. Sju miljøverdigrupper er vurdert i forhold til dette kriteriesettet: Isbiota, Plankton (plante- og dyreplankton), Fisk, Mesopelagisk fauna, Bunnsamfunn, Sjøpattedyr og Sjøfugl. Fysiske og kjemiske faktorer som virker inn på de økologiske sammenhengene og påvirker miljøverdiene er beskrevet. Økologiske prosesser innen og mellom gruppene er påpekt.
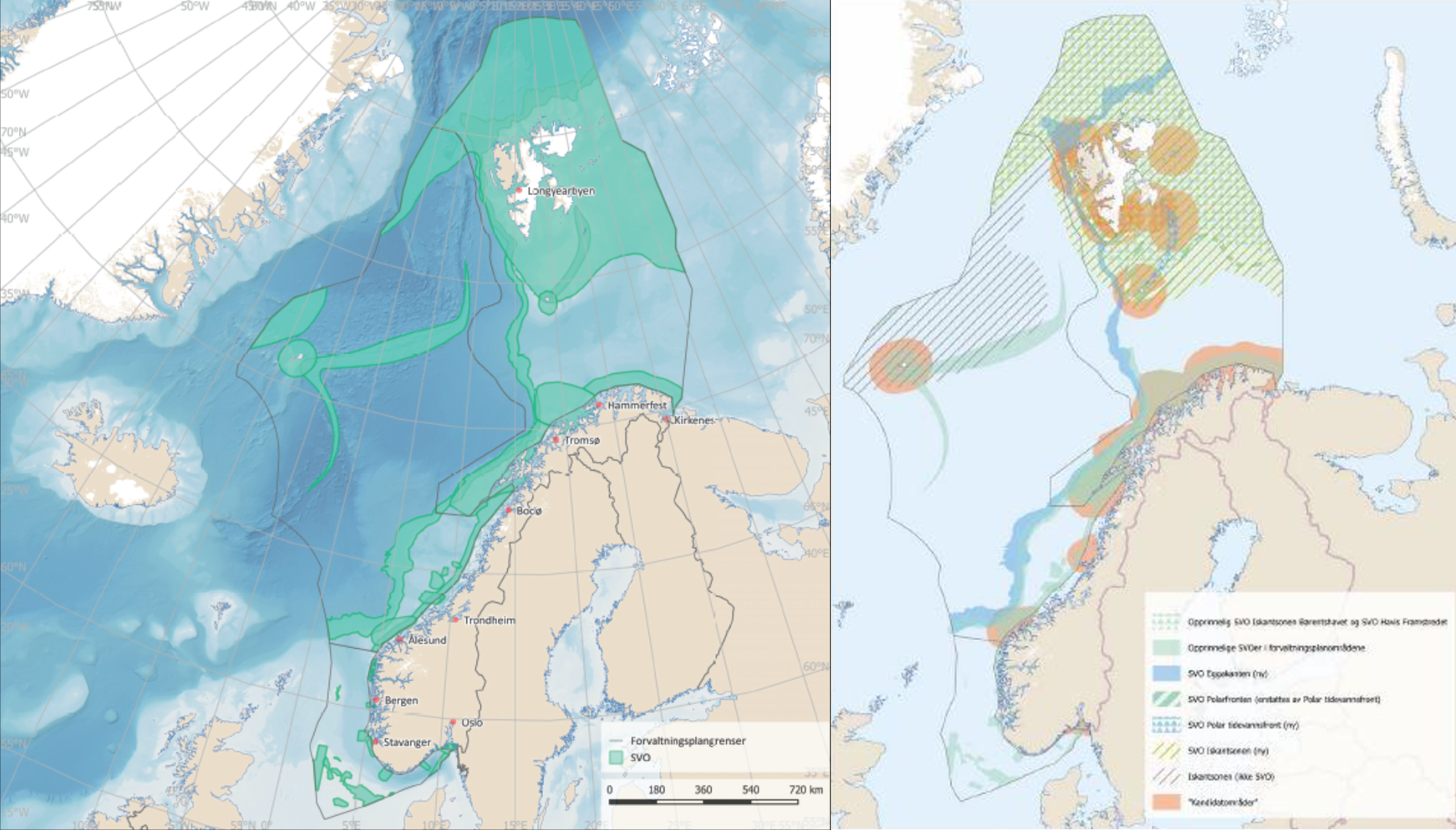
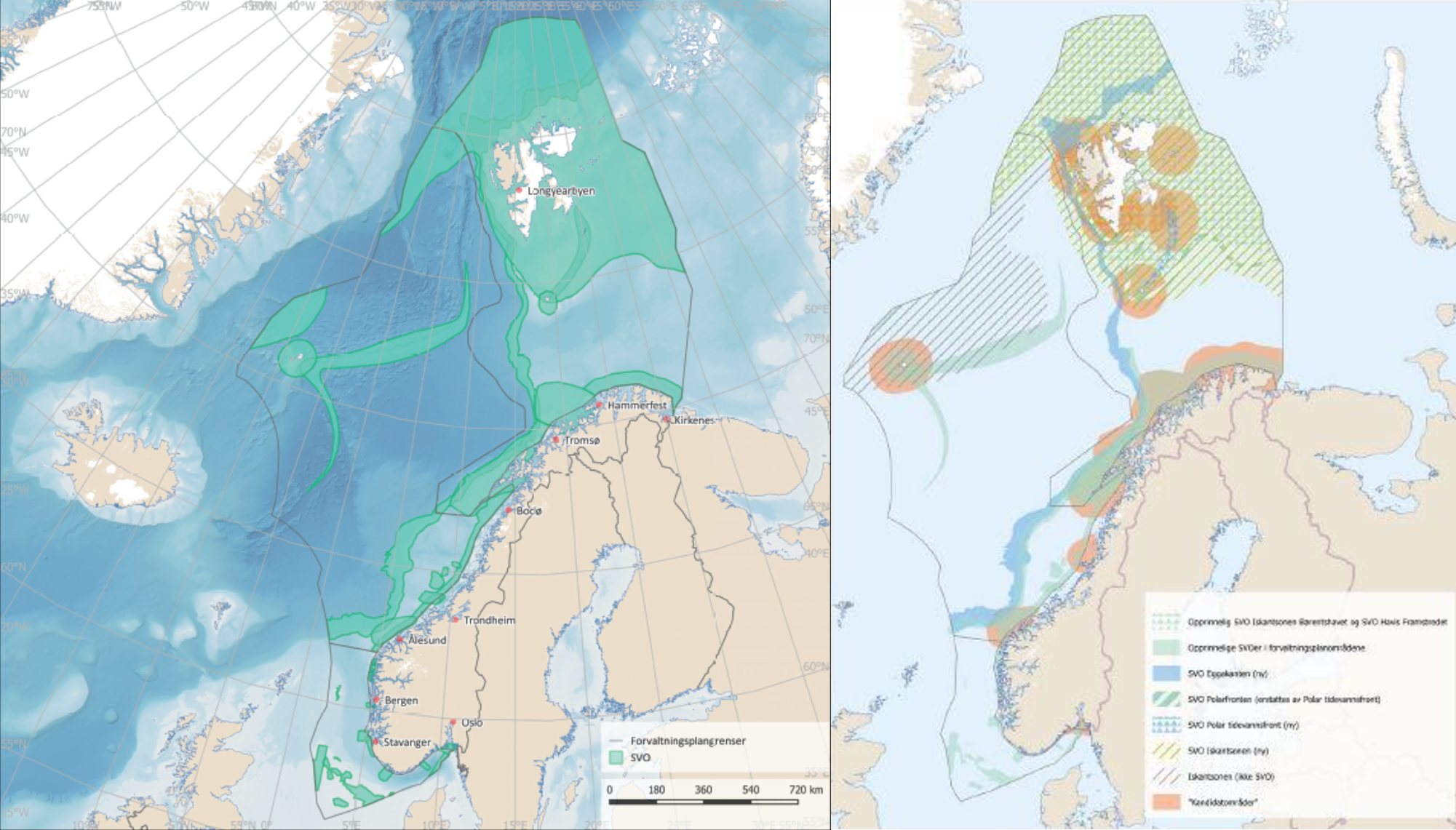
Den tverrfaglige ekspertgruppen mener at SVO-ene bør reflektere samspill mellom prosessene og miljøet som danner grunnlaget for miljøverdiene i området. Det er derfor foreslått at SVO inkluderer ikke bare et gytefelt, en sjøfuglkoloni eller utvalgte koraller, men i tillegg også reflekterer økologisk samspill i et større område. I forhold til tidligere SVO-er (figur 1.1) er det derfor foreslått færre, men større arealer for hvert SVO (figur 1.2). Selv om de foreslåtte SVO-ene har blitt større, varierer miljøverdiene innenfor områdene i tid og rom, dvs. ulike miljøverdier slår ut i ulike sesonger og i ulike deler av områdene. Denne tilnærmingen førte til størst endring i Nordsjøen/Skagerrak og til dels i Norskehavet, mens Barentshavet allerede hadde flere større områder. Dette vil også legge til rette for en bedre vurdering av status og utvikling over tid i de foreslåtte SVO-ene, da mindre SVO-er sjelden har hatt stedsspesifikk overvåking.
Figur 1.1. Øverst) De eksisterende SVO-er (Meld. St. 20 (2019-2020)). Nederst) Kart over kandidatområder (oransje felt) fra SVO-utredningen (Faglig forum for norske havområder 2019 b).
Norske havområder er blant verdens mest produktive og alle havområdene er i seg selv verdifulle. Det er derfor en utfordring å peke ut hvor de særlig verdifulle områdene ligger. Tidligere vurderinger og valgte SVO-er er gode veiledere (Olsen og von Quillfeldt 2003; Ottersen og Auran 2006, Ottersen mfl. 2010, von Quillfeldt 2018) sammen med ny kunnskap, blant annet fra rapporten om viktige områder for sjøfugl (Systad mfl. 2019) og notatet om avgrensning av SVO Havområdene rundt Svalbard (NP 2019).
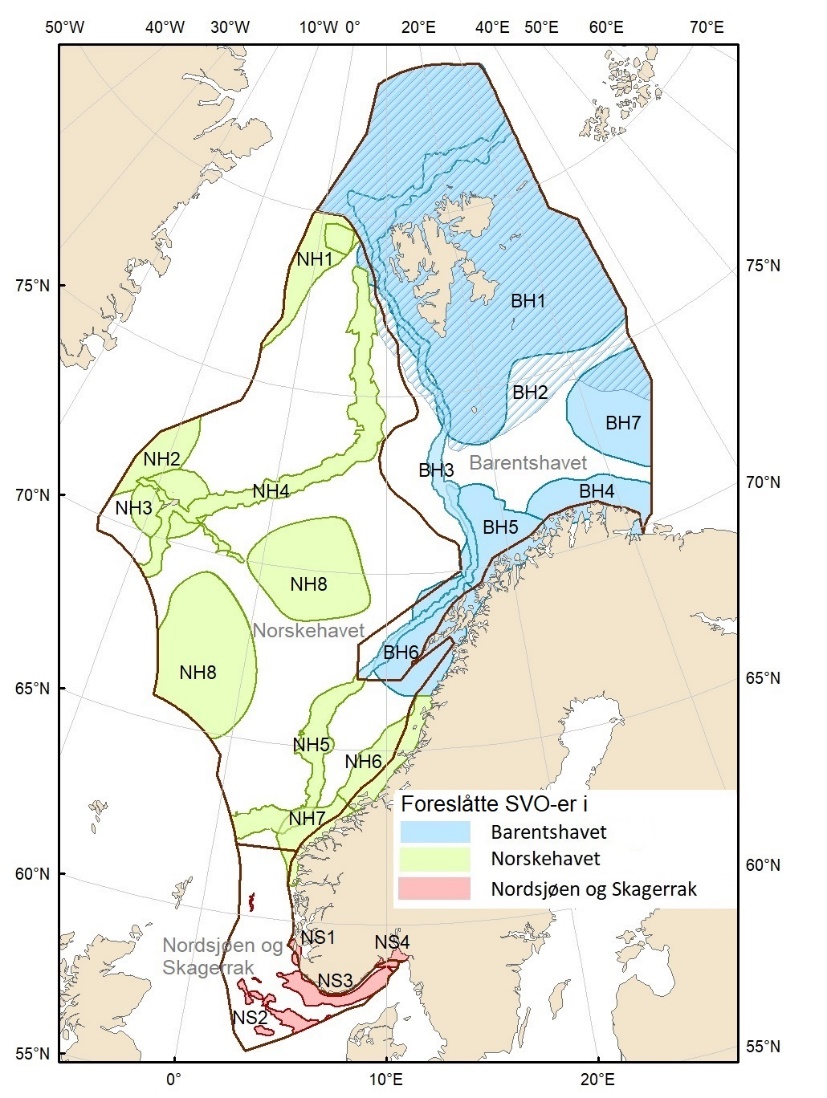
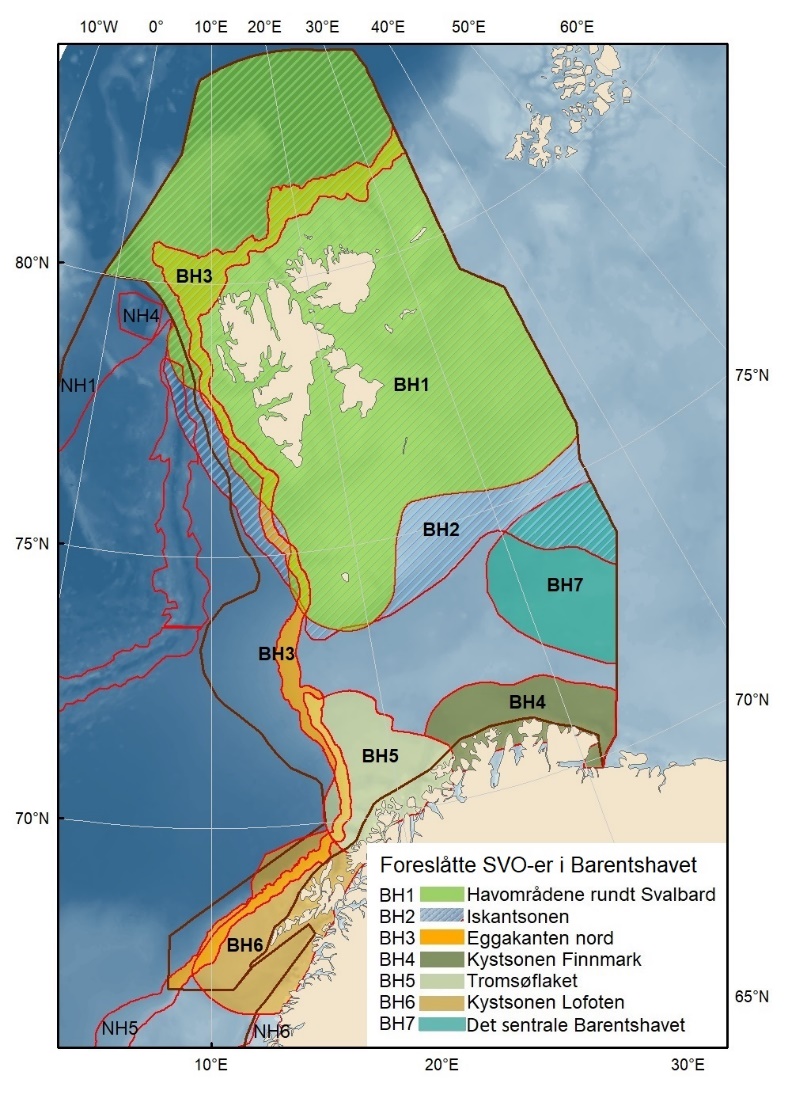
Den foreliggende rapporten beskriver to eksisterende SVO-er, 13 forslag til endrete SVO-er og fire forslag til nye SVO-er, totalt 19 områder fordelt på Barentshavet (sju), Norskehavet (åtte) og Nordsjøen/Skagerrak (fire).
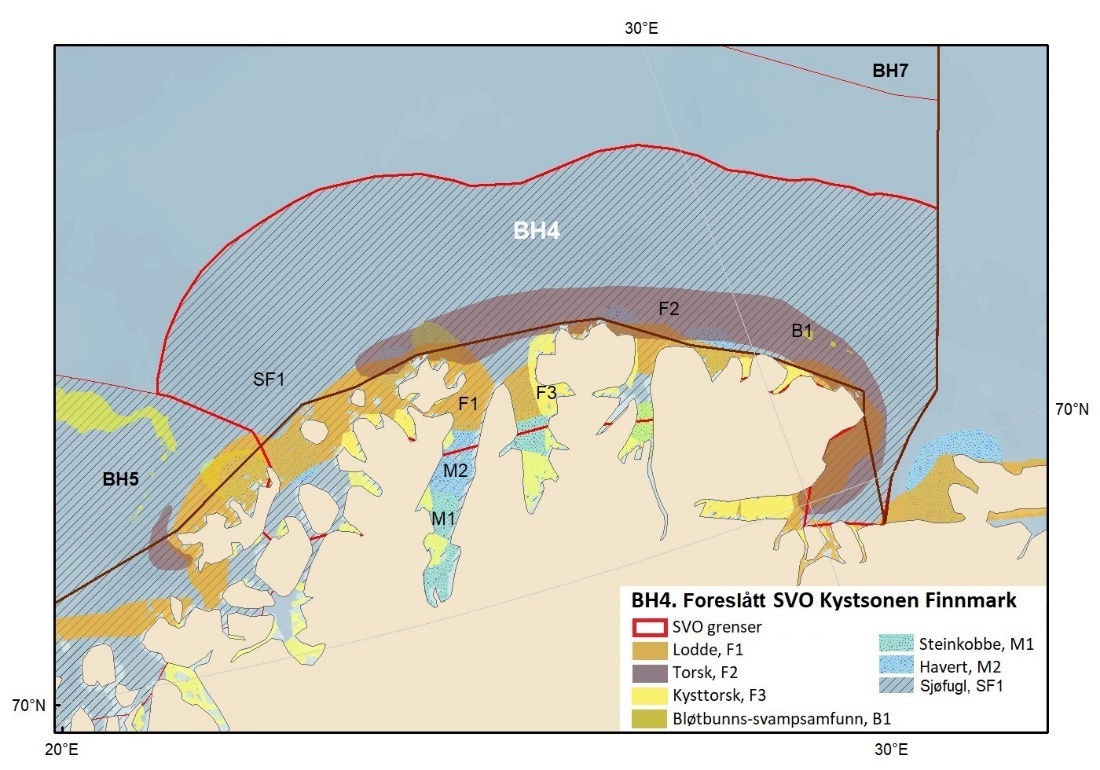
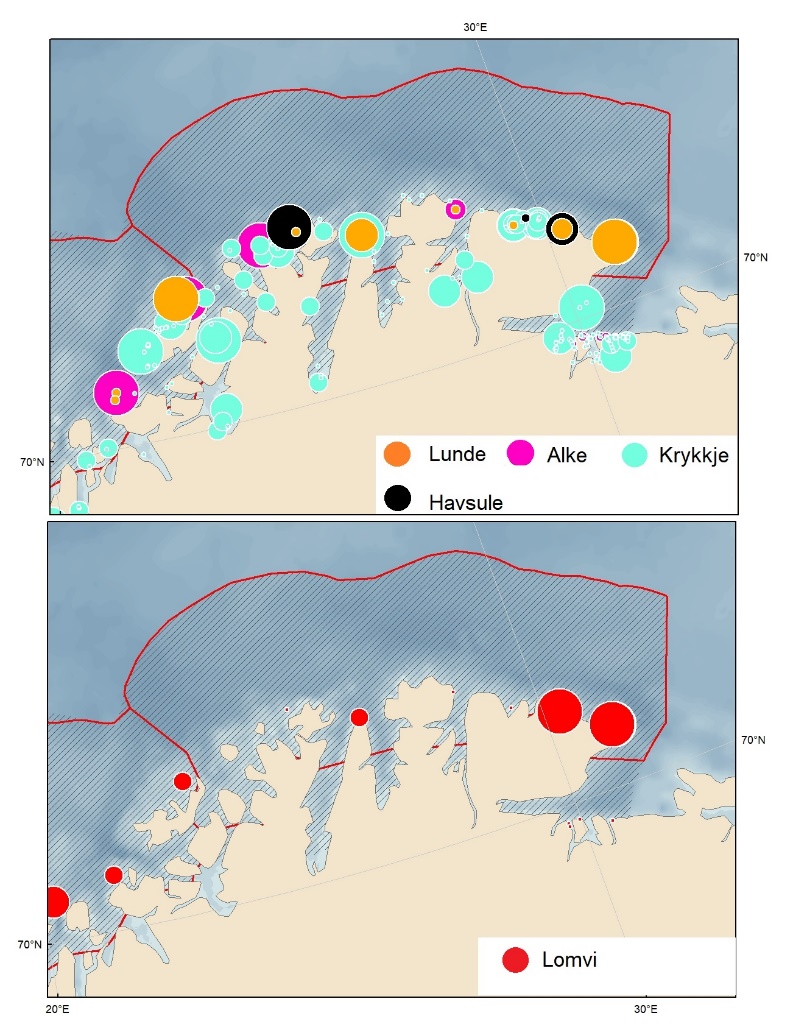
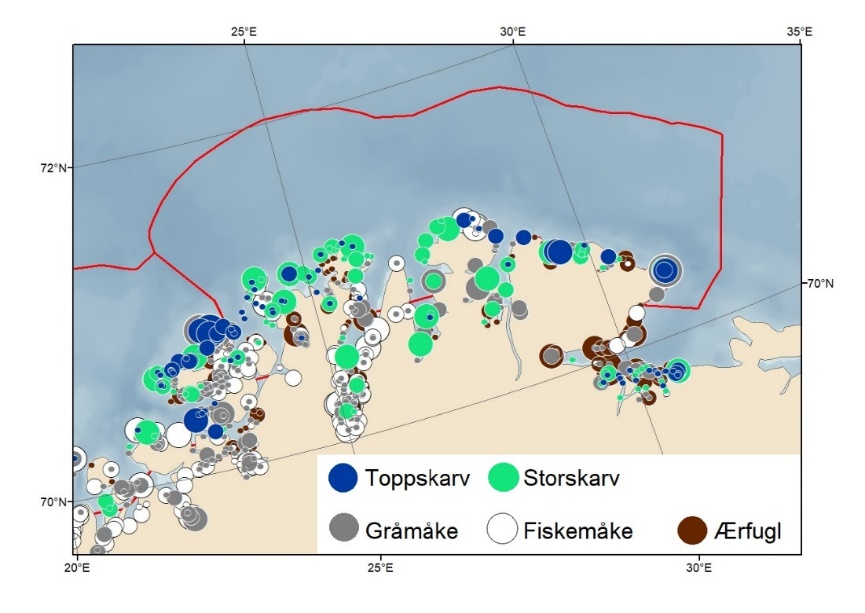
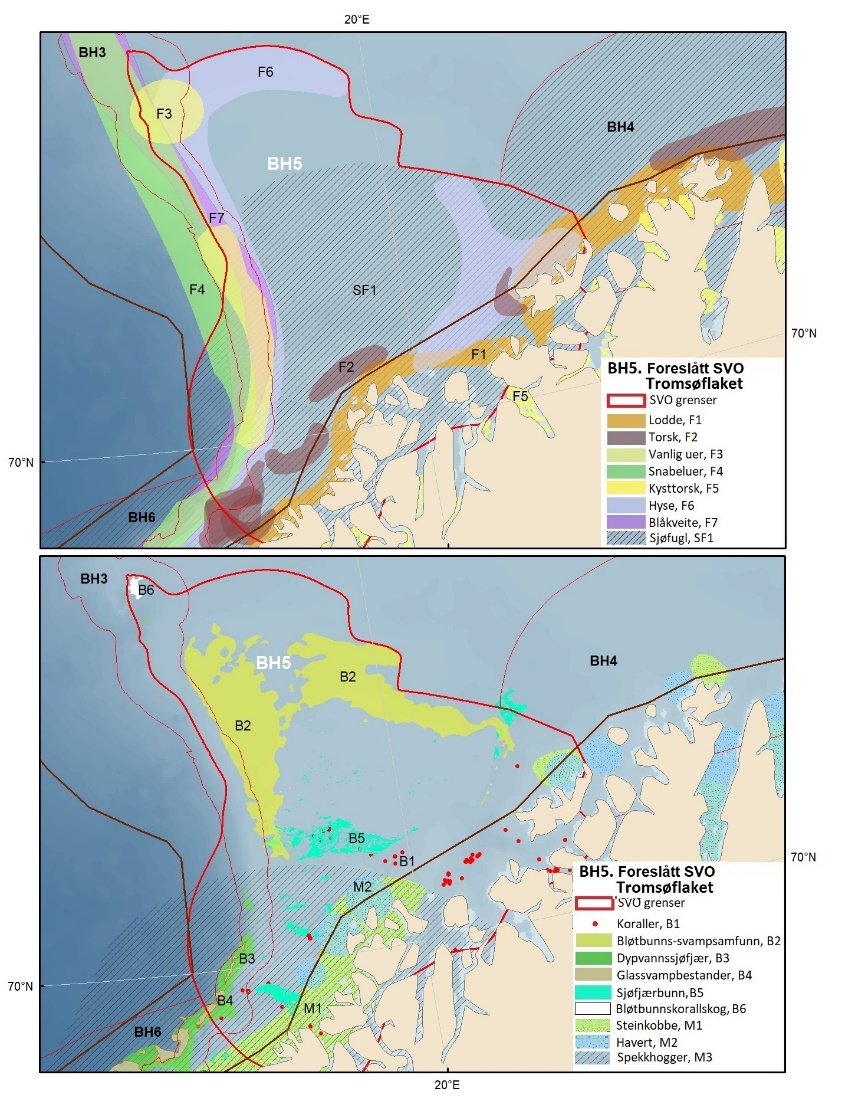
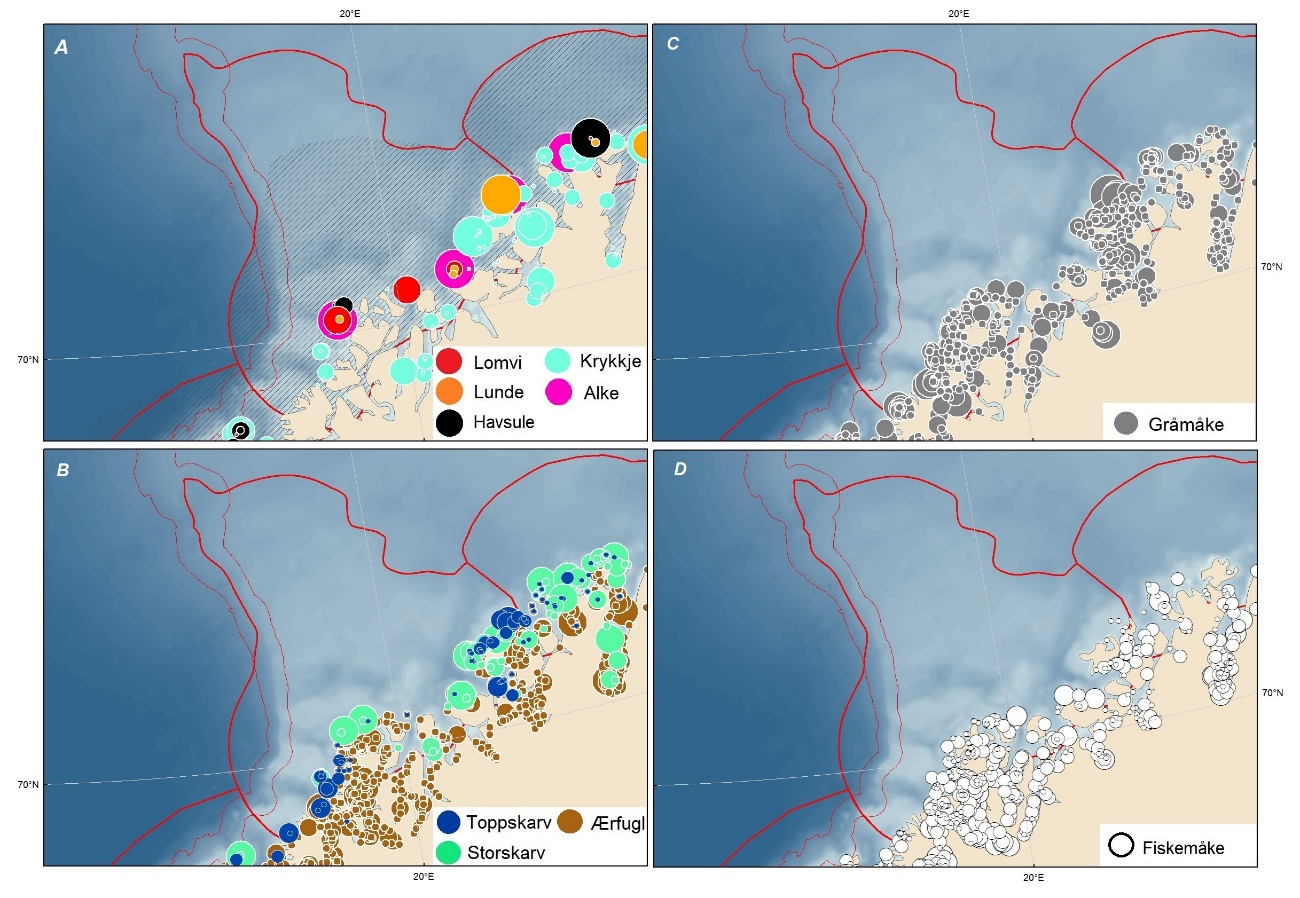
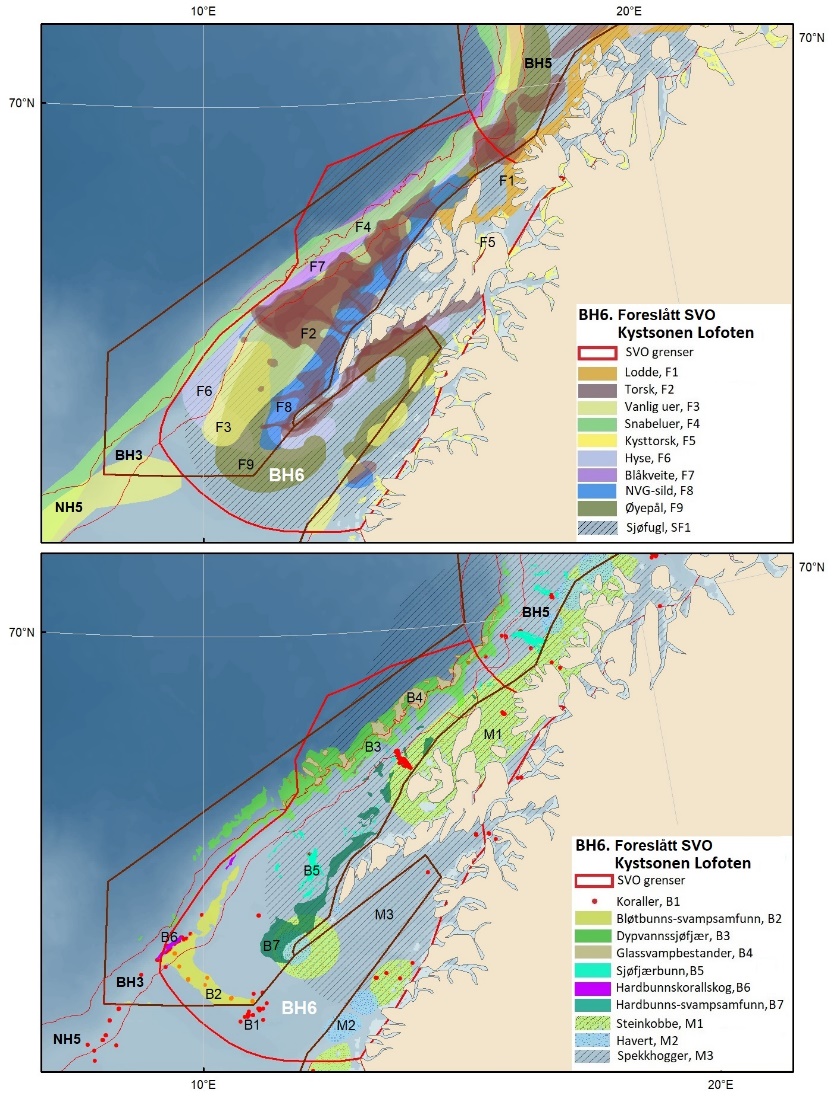
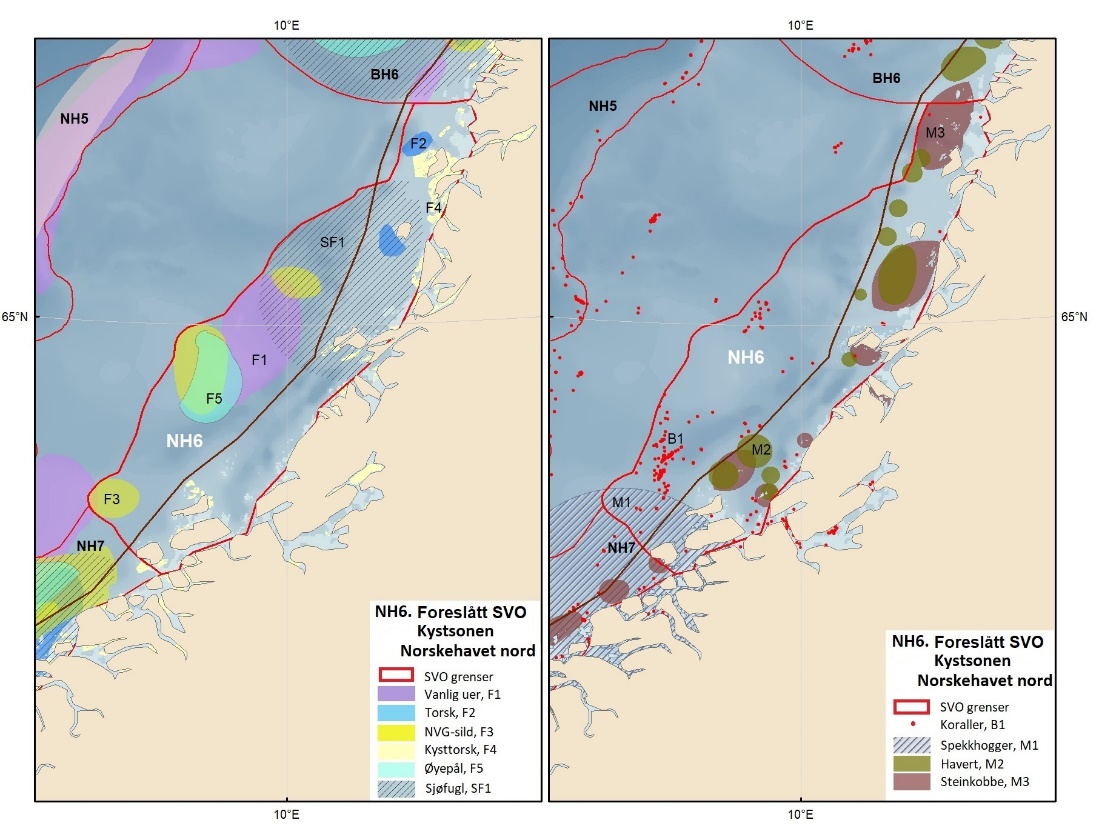
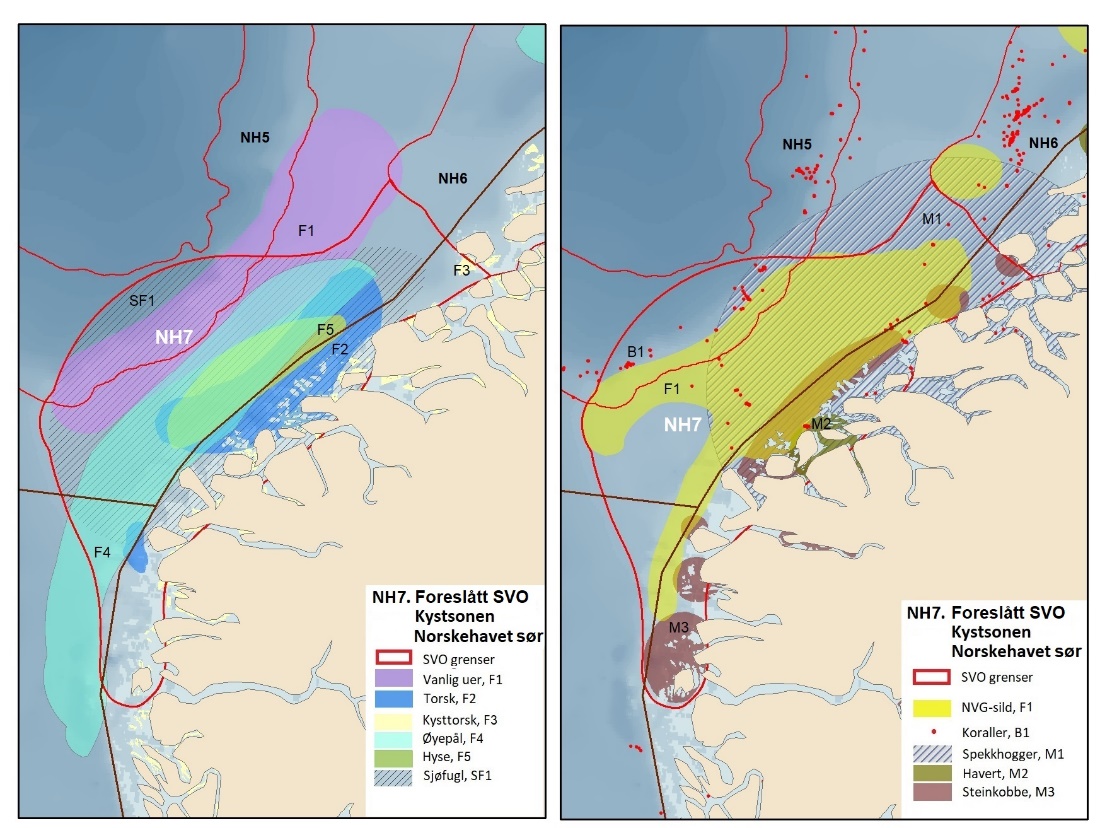
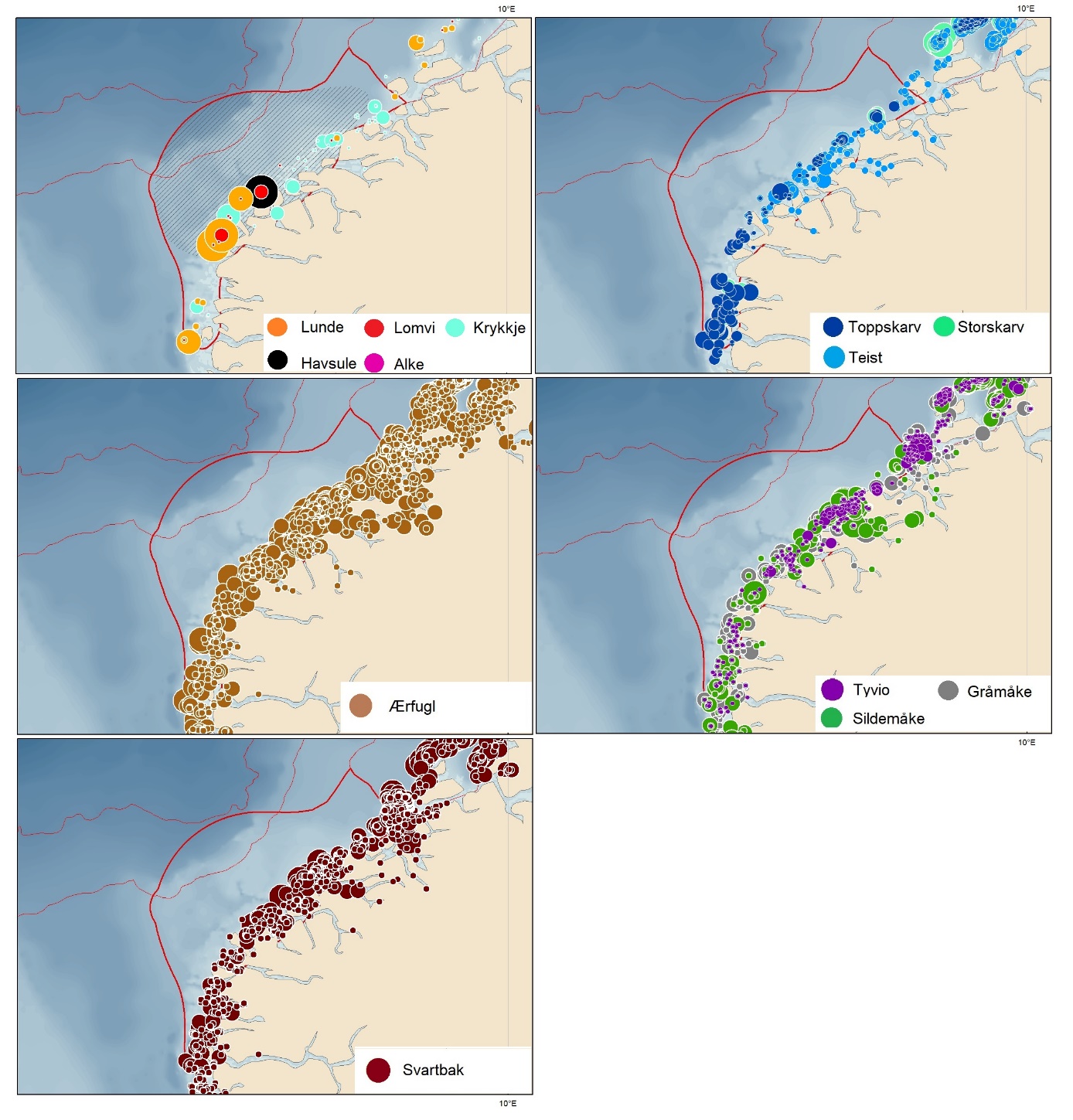
Foreslåtte SVO-er i Barentshavet er vist i figur 1.2 og tabell 1.1. Et foreslått n ytt SVO, Det sentrale Barentshavet, er særlig viktig som endepunkt for svømmetrekket etter hekkesesongen for lomvi og polarlomvi. To foreslått utvidete SVO-er, Iskantsonen og Havområdene rundt Svalbard omfatter alle miljøverdigruppene i større eller mindre arealer innenfor områdene, men med ulik betydning avhengig av årstid. I foreslått utvidet SVO Havområdene rundt Svalbard er det identifisert delområder med særlig høy verdi, og hele området er noe utvidet for å få med et av de viktigste beiteområdene for lodde. Foreslått utvidet SVO Iskantsonen dekker utvidetlsen til maksimal isutbredelse for å fange opp den delen av oppblomstringen i iskantsonen som strekker seg sør for iskanten og som legger grunnlag for at også dette er et viktig beiteområde. Tre foreslåtte SVO-er, Eggakanten nord, Havområdene rundt Svalbard og Iskantsonen, har til dels store overlapp, men dette er begrunnet med ulike miljøverdier og biologiske prosesser som slår ulikt ut for områdene der de overlapper. Foreslått utvidet SVO Tromsøflaket har fått grensene utvidet mot nord for å dekke rike bunnsamfunn og de viktige transportbanene for plankton, som fiskelarver og krepsdyr, og inn i fjordene som er beiteområder for fisk, sjøfugl, sel og vinterbeite for hval. Foreslått utvidet SVO Kystsonen Finnmark dekker det eksisterende SVO Kystnære områder fra Tromsøflaket til grensen mot Russland. De kystnære foreslåtte SVO-ene er alle utvidet med sjøfuglenes beiteområder både et stykke inn i fjordene, men også ut mot havet. Foreslått endret SVO Kystsonen Lofoten dekker størstedelen av det eksisterende SVO Havområdene utenfor Lofoten til Tromsøflaket. I tillegg er Vestfjorden, som nå er inkludert i det eksisterende SVO Kystsonen Norskehavet, inkludert siden miljøverdiene i Vestfjorden passer naturlig bedre sammen med miljøverdiene i dette området.
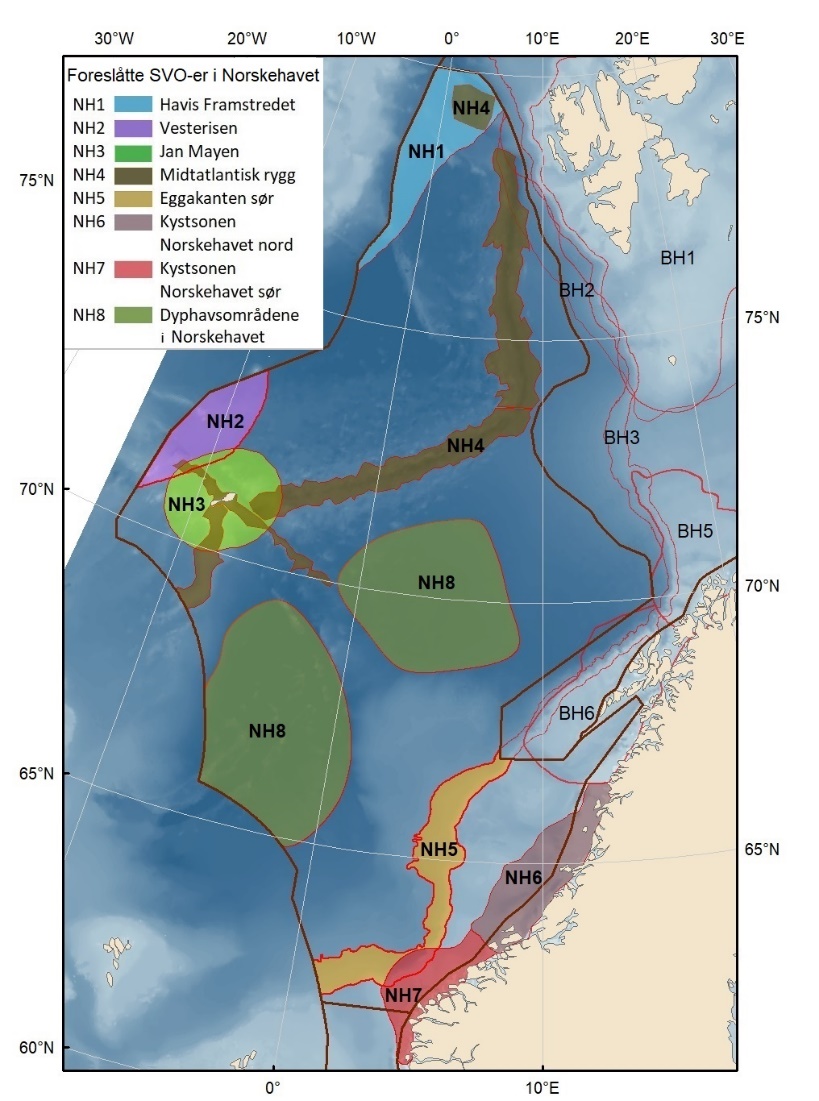
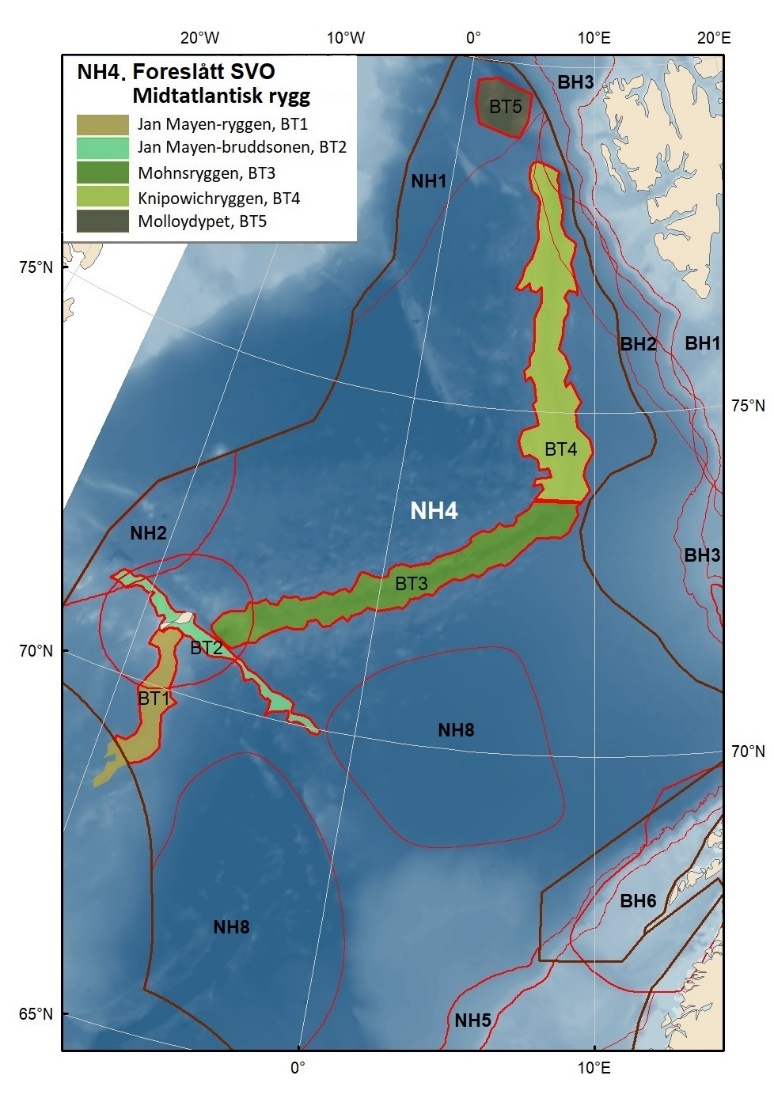
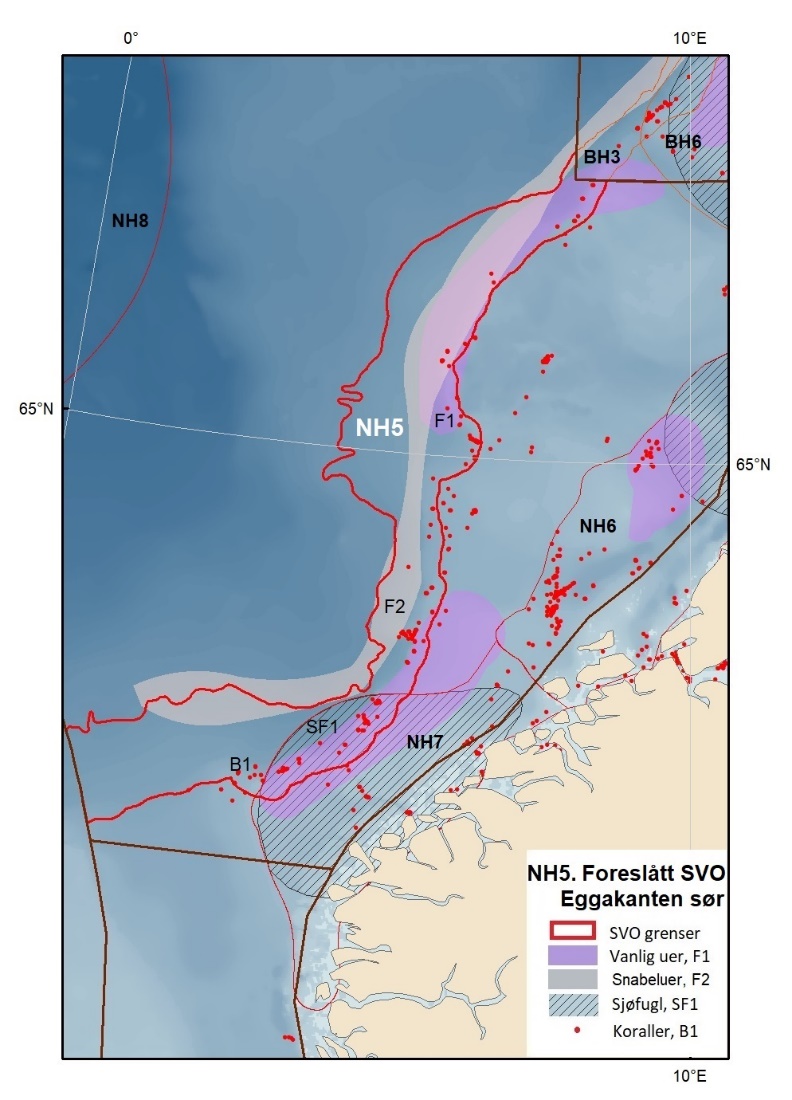
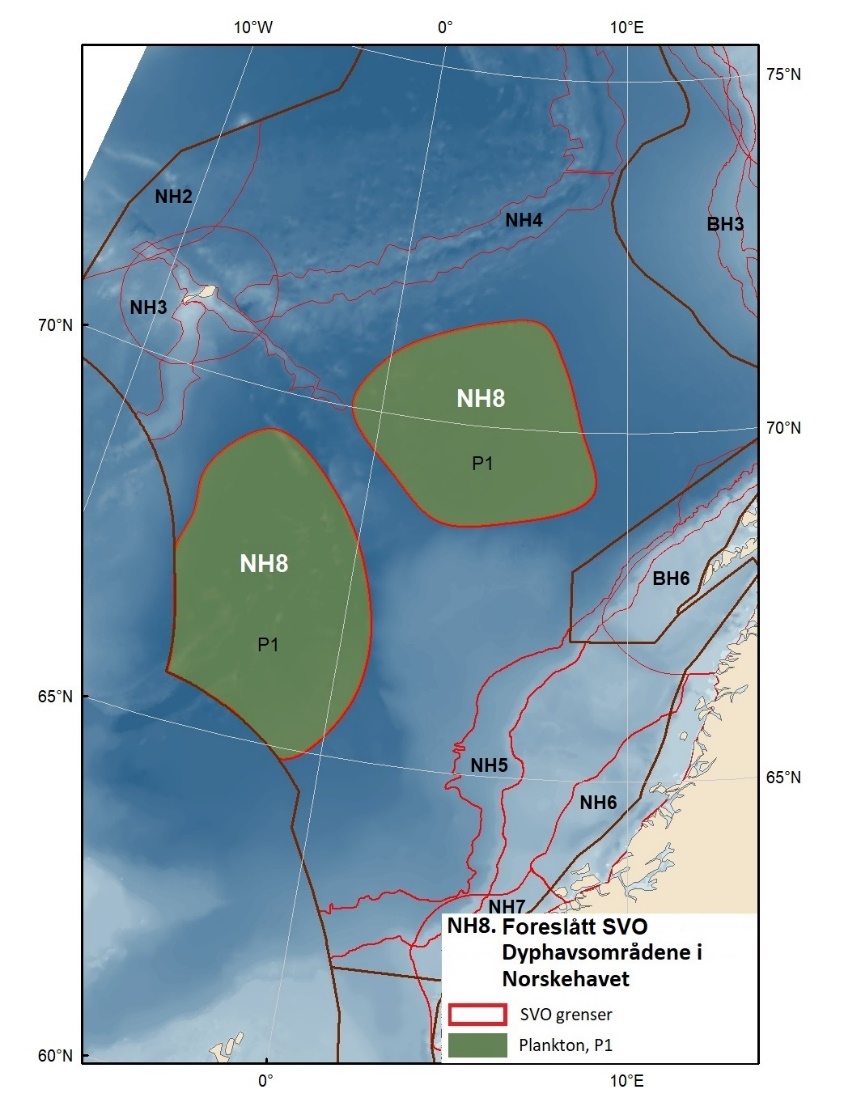
Foreslåtte SVO-er i Norskehavet er vist i figur 1.2 og tabell 1.1. I tillegg til de eksisterende SVO Havis Framstredet og SVO Vesterisen som er forslått uendret, er mange små kystnære SVO-er samlet i to større forslag til kystnære SVO-er, Kystsonen Norskehavet nord og Kystsonen Norskehavet sør, somder begge omfatter kystnære gyteområder av stor betydning og sjøfuglkolonier som er sterkt tilknyttet beiting på fisk. Foreslått endret SVO Jan Mayen er dekker bedre dekke viktige beiteområder for sjøfugl. Høy produktivitet var et viktig argument for det eksisterende SVO Den arktiske front, men siden dette ikke er godt nok undersøkt dokumentert, blir SVO Den arktiske front ikke lenger vurdert som et selvstendig SVO og foreslås fjernet som eget SVO. To nye områder, Midtatlantisk rygg og Dyphavsområdene i Norskehavet er forslått agt til nye SVO-er, til tross for at SVO Den arktiske front til en stor grad har overlapp med det nye forslaget SVO Midtatlantisk rygg. I likhet med foreslått endret SVO Eggakanten sør, er det likevel de dyptlevende livsformene som er særlig verdifulle her.
Den midtatlantiske ryggen er svært produktiv, har mye endemisk og unik fauna, og er gyte- og oppvekstområde for langsomtvoksende fisk . Bunnsamfunn nær og på Den midtatlantiske rygg er svært sårbare og har liten eller ingen restitueringsevne. Dyphavsområdene er overvintringshabitat for Calanus spp. og særdeles viktige for å opprettholde populasjonene. Disse planktonreservoarene er også essensielle for produktivitet og reproduksjon av fisk og sjøfugl langs Eggakanten og i kystsonens SVO-er i både Norskehavet og Barentshavet.
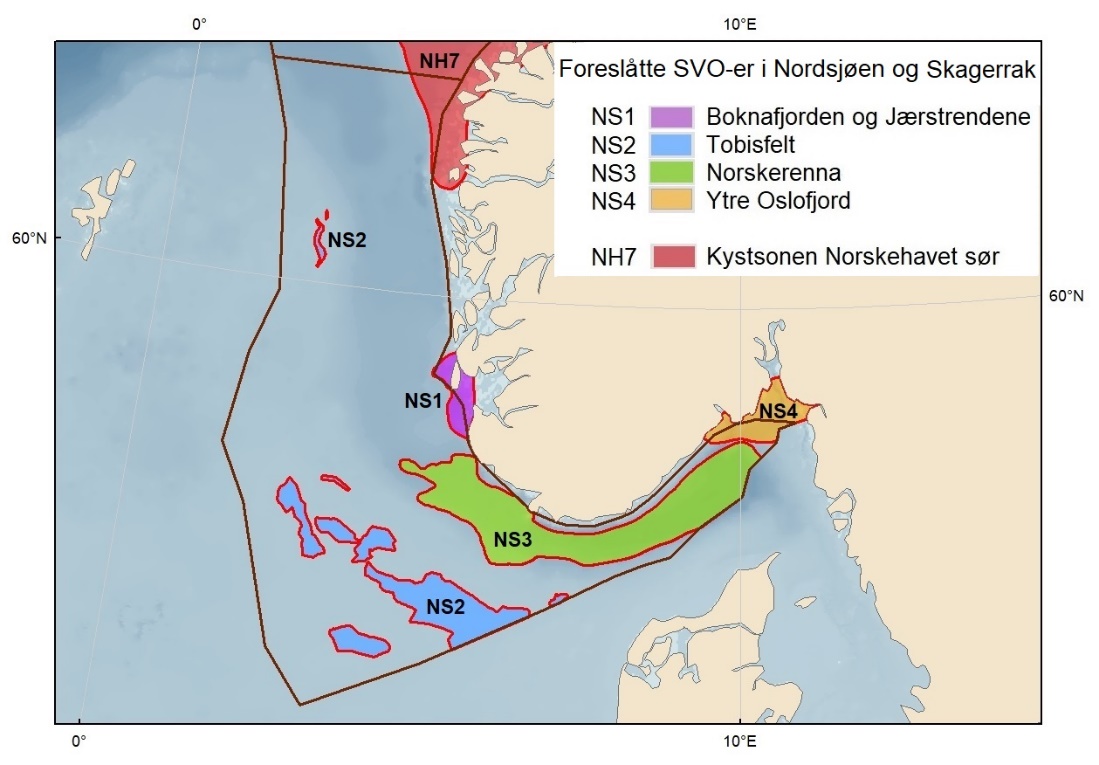
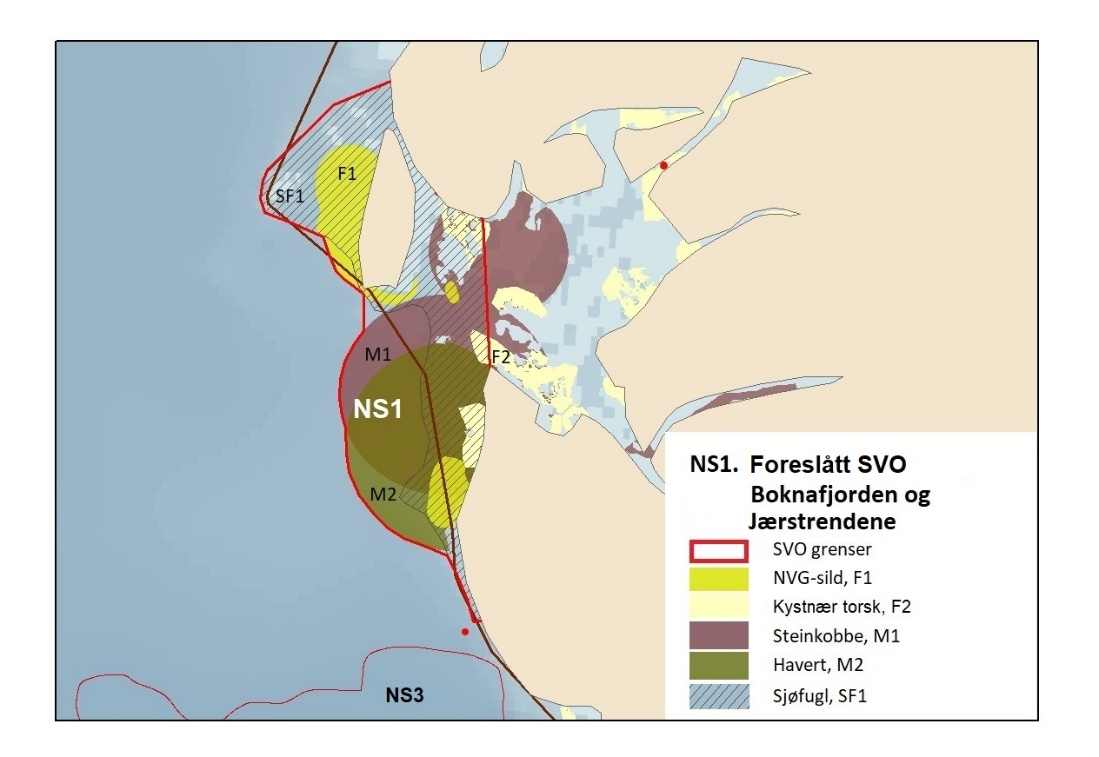
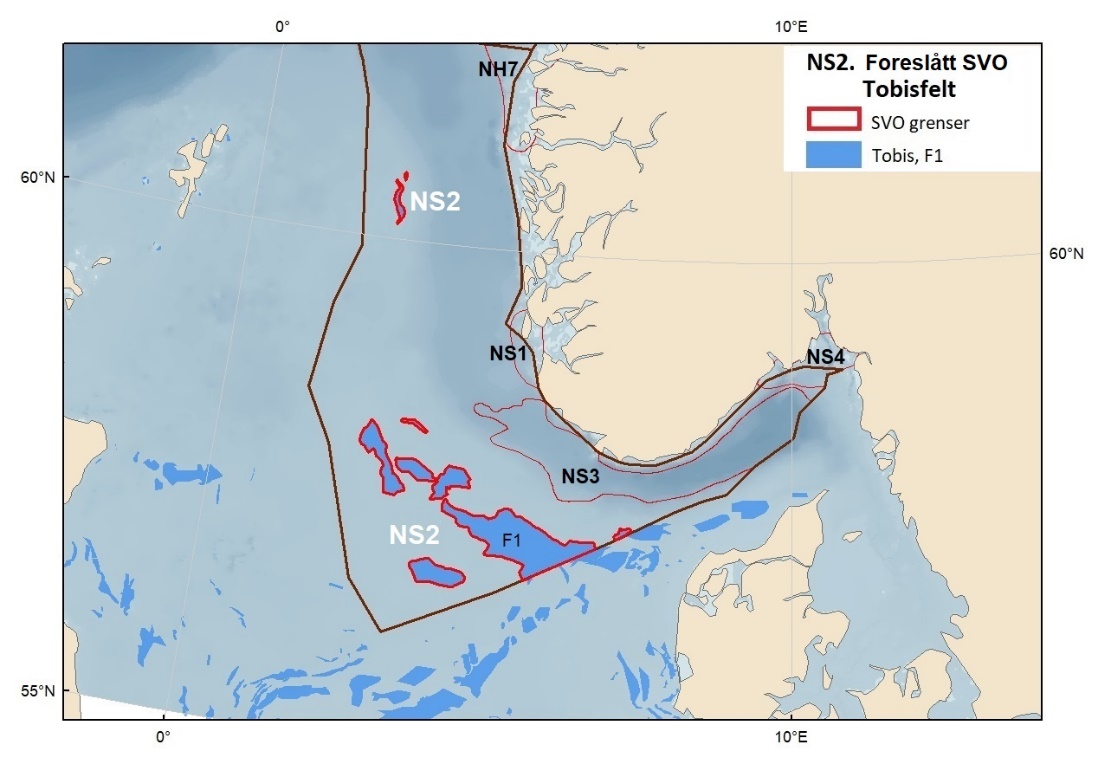
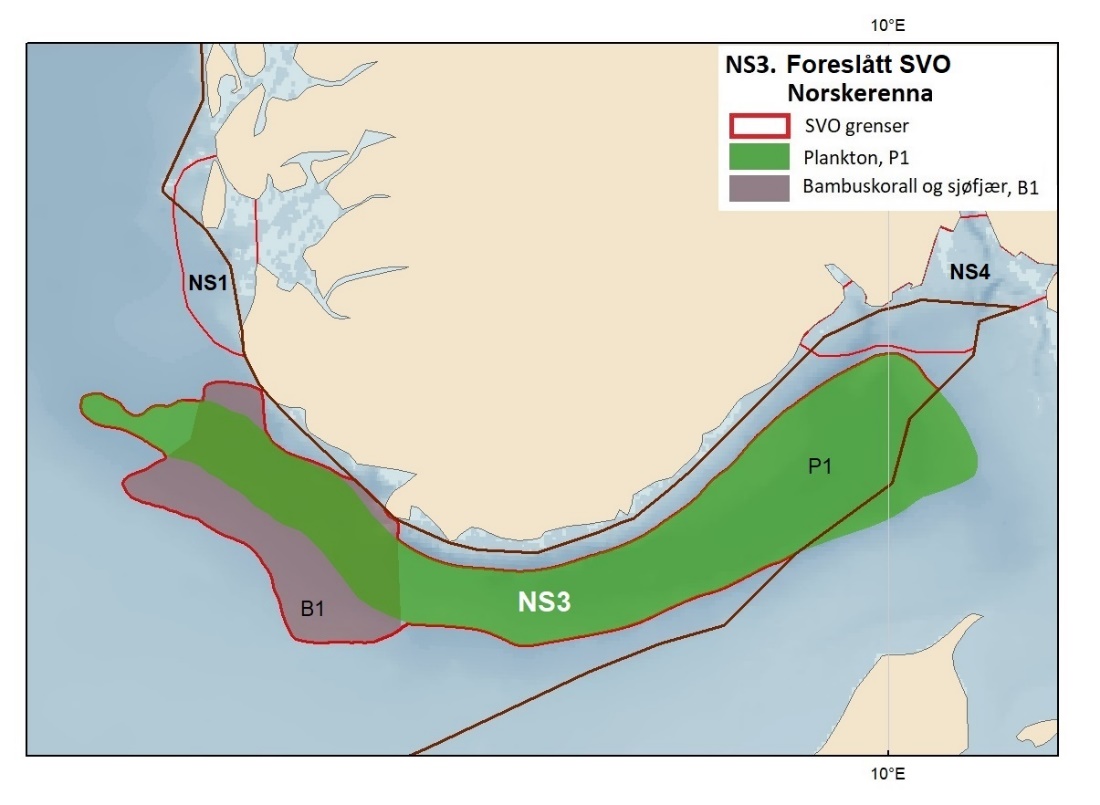
Foreslåtte SVO-er i Nordsjøen og Skagerrak er vist i figur 1.2 og tabell 1.1. Det er størst endringer i dette forvaltningsplanområdet, der antallet SVO-er er foreslått redusert til fire. To foreslått endringer i SVO-er er kystnære, Boknafjorden og Jærstrendene og Ytre Oslofjord, der ett område dekker for Skagerrak og ett for kystnært område ved Nordsjøen. Foreslått endret SVO Ytre Oslofjord er særlig viktig om vinteren som beiteområde for sjøfugl fra hele Nordsjøområdet. Området har helt spesielle fysiske/kjemiske og klimatiske forhold, og Oslofjorden har høyere planteplanktonproduksjon enn andre fjordsystemer. Foreslått endret Boknafjorden, inkludert Karmøyflaket og inn i Boknafjorden, har både historisk viktige gytefelt for norsk vårgytende sild (NVG-sild, videre omtalt som sild), større selkolonier, inkl. den sørligste kjente ynglekolonien av havert, og er et viktig overvintringsområde for kystnære sjøfugl. Begge områdene er antatt å ha et særlig høyt biologisk mangfold siden områdene er så varierte og omfatter overgangen mellom fjord, kyst og hav. Foreslått endret SVO Tobisfelt dekker alle tobisfeltene i Norsk Økonomisk Sone. Foreslått nytt SVO Norskerenna omfatter Nordsjøens eneste dype område, med sin særegne fauna og sitt reservoar av plankton og mesopelagisk fisk. I nord er det eksisterende SVO Bremanger til Ytre Sula anbefalt innlemmet i foreslått SVO Kystsonen Norskehavet sør. Eksisterende SVO Makrellfelt er anbefalt fjernet da det er vist at det ikke er en egen makrellstamme i Nordsjøen, og eksisterende SVO Skagerrak er inkludert i foreslått nytt SVO Norskerenna. Eksisterende SVO Transekt Skagerrak og andre kystnære SVO-er er delvis inkludert i samme område eller fjernet siden de ikke vil bidra med ytterligere verdier utover de som allerede er omfattet av de to kystnære foreslåtte SVO-ene som gjenstår.
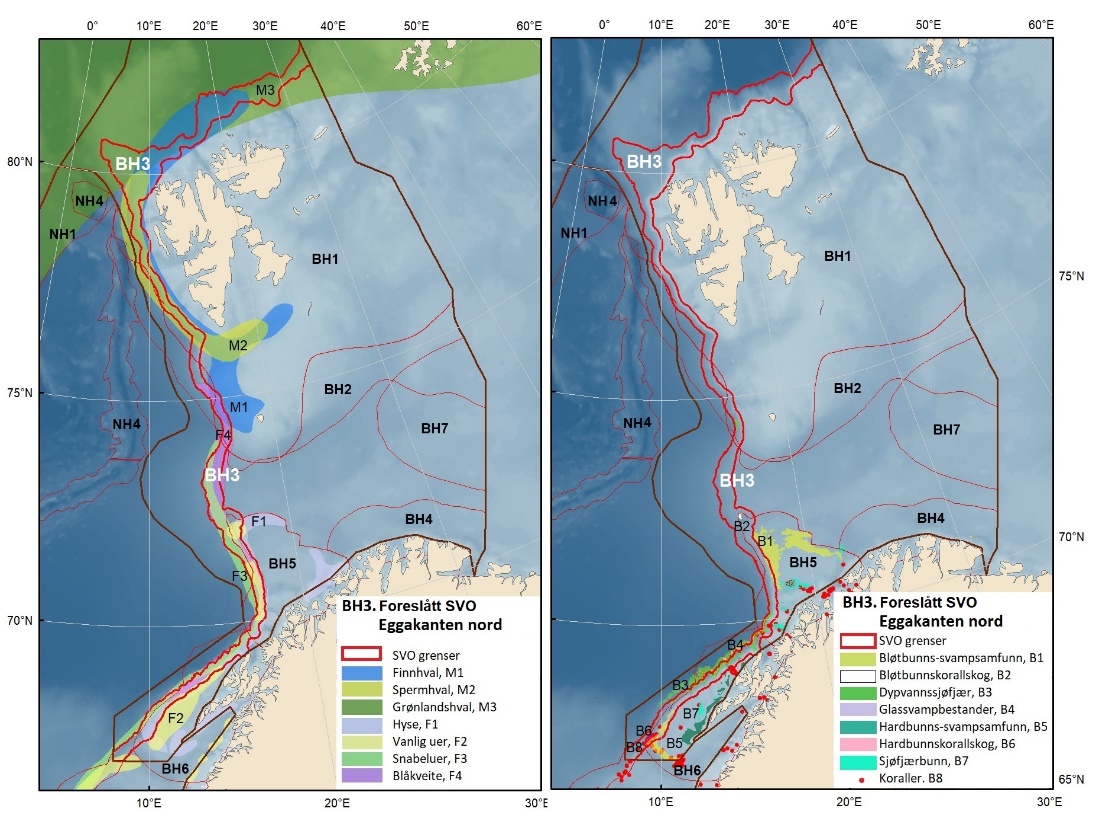
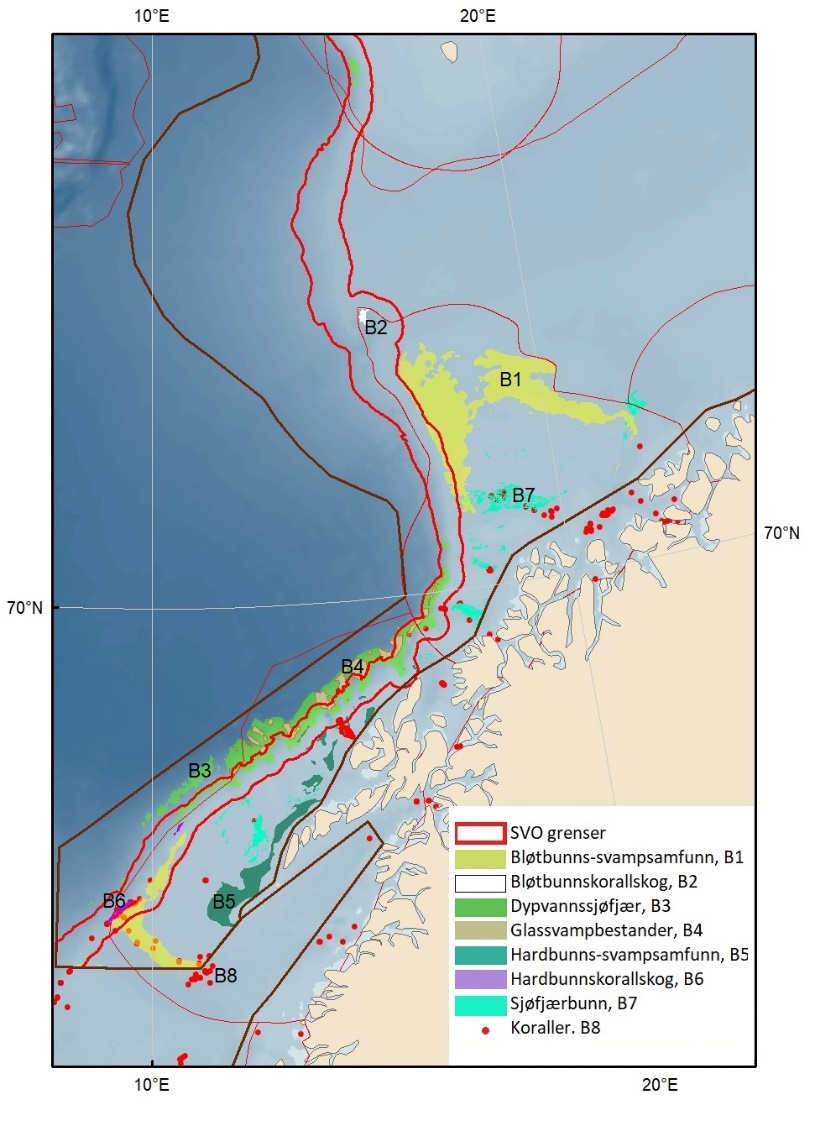
Figur 1.2. Kart over forslag til særlig verdifulle og sårbare områder (SVO) presentert i denne rapporten. Sju SVO-er ligger i Barentshavet (BH1–-BH7), åtte i Norskehavet (NH1–-NH8) og fire i Nordsjøen og Skagerrak (NS1–-NS4). Foreslått endret SVO Iskantsonen (skravert) overlapper med foreslått endrett SVO Havområdene rundt Svalbard (BH1), deler av foreslått endret SVO Eggakanten nord (BH3) og deler av foreslått nytt SVO Det sentrale Barentshavet (BH7). Grensene for forvaltningsplanområdene er markert med brune linjer.
Forslag SVO-kode Miljøverdi-rapport 2021
SVO-kode Meld. St.2019-2020
SVO-navn Meld. St.2019-2020
Forslag SVO-navn Miljøverdirapport2021
Forslag til endringer
Barentshavet
BH3
Polar tidevannsfront
Havområdene rundt Svalbard
Inkludert i Havområdene rundt Svalbard
BH1
BH4
Havområdene rundt Svalbard
Havområdene rundt Svalbard
Utvidet med sommerbeite for lodde
BH2
BH6
Iskantsonen
Iskantsonen
Utvidet til maksimal isutbredelse
BH3
BH7
Eggakanten
Eggakanten nord
Starter i sør ved grensen til forvaltningsplan-området for Norskehavet, ellers uendret
BH4
BH1
Kystsonen Finnmark
Kystsonen Finnmark
Utvidet nordover med Kandidatområdene og et stykke inn i fjordene for å dekke beiteområder for sjøfugl, sel og hval
BH5
BH2
Tromsøflaket
Tromsøflaket
Utvidet sør til Senja, nord til Sørøya og litt inn i fjordene, inkl. Lopphavet
BH6
BH5
Lofoten til Tromsøflaket
Kystsonen Lofoten
Endret til å omfatte Vestfjorden og Røst til Senja. Utvidet vestover med Kandidatområdene
BH7
Det sentrale Barentshavet
Nytt område
Norskehavet
NH1
NH11
Havis Framstredet
Havis Framstredet
Uendret
NH2
NH12
Vesterisen
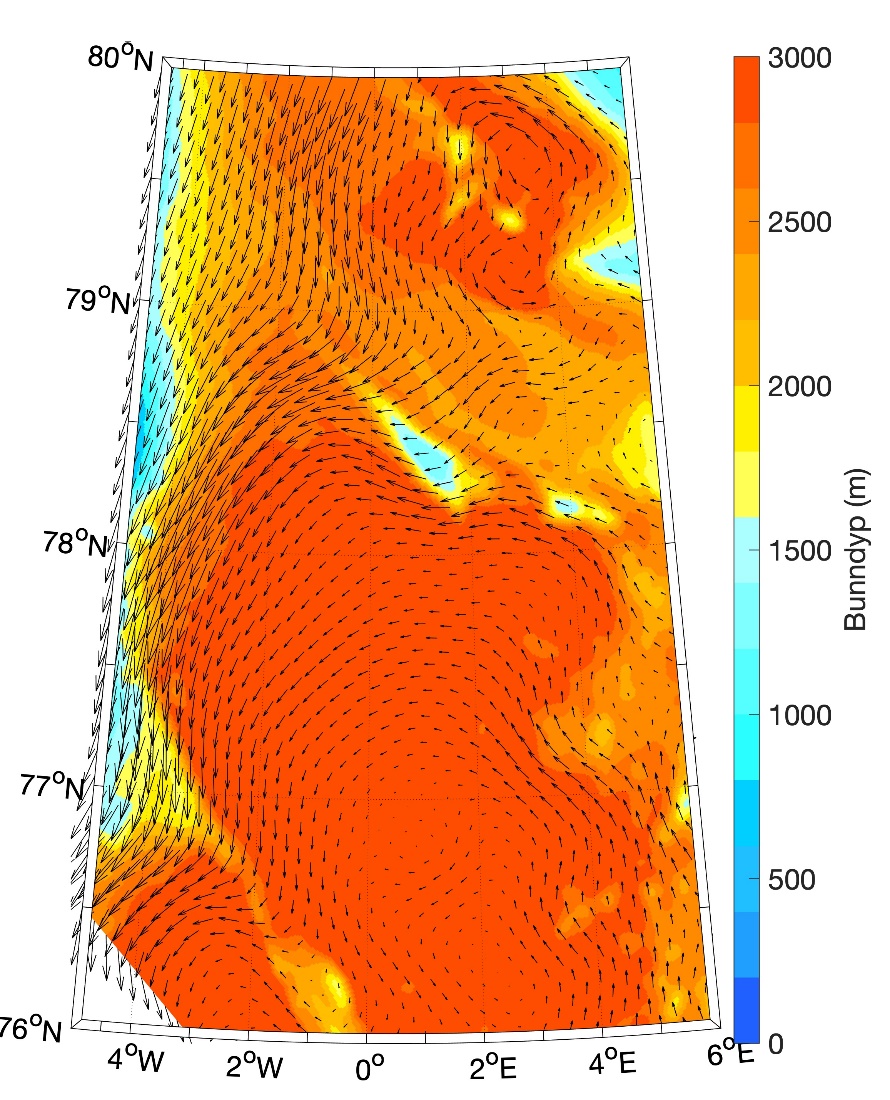
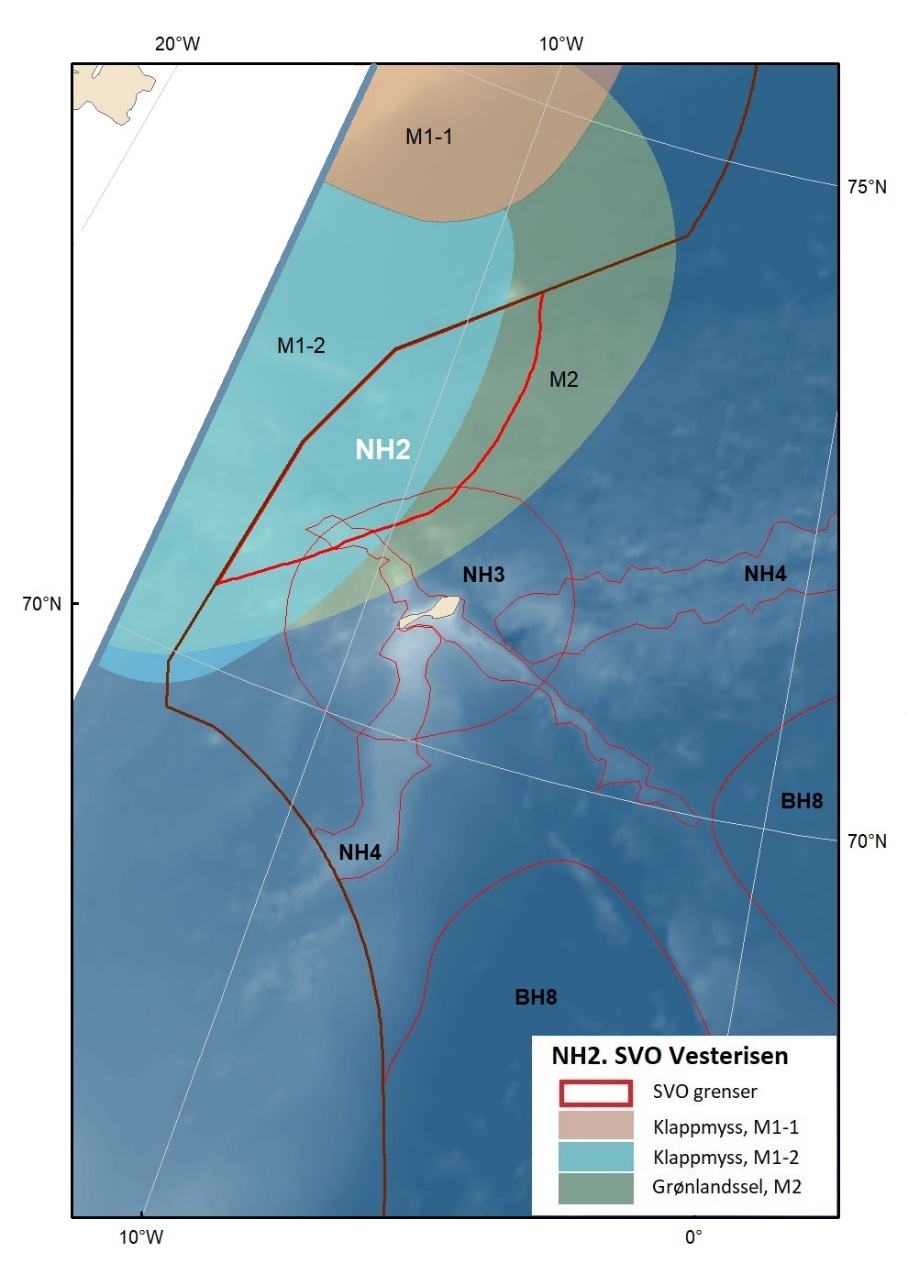
Vesterisen
Uendret
NH3
NH8
Jan Mayen
Jan Mayen
Utvidet med Kandidatområdet
NH4
NH6
Den arktiske front
Midtatlantisk rygg
Nytt område. Overlapper delvis med SVO Den arktiske front som ikke lenger anses som et SVO
NH5
NH7
Eggakanten, sør
Eggakanten, sør
Starter i nord ved grensen til forvaltningsplan-området for Barentshavet, ellers uendret
NH6
NH1
Iverryggen
Kystsonen Norskehavet nord
Inkludert i Kystsonen Norskehavet nord
NH6
NH2
Haltenbanken
Kystsonen Norskehavet nord
Inkludert i Kystsonen Norskehavet nord
NH6
NH3
Sklinnabanken
Kystsonen Norskehavet nord
Inkludert i Kystsonen Norskehavet nord
NH6
NH5
Froan med Sularevet
Kystsonen Norskehavet nord
Inkludert i Kystsonen Norskehavet nord
NH6/NH7
NH9
Kystsonen Norskehavet
Kystsonene Norskehavet sør/ nord
Inkludert i Kystsonen Norskehavet sør/nord
NH6
NH10
Remman
Kystsonen Norskehavet nord
Inkludert i Kystsonen Norskehavet nord
NH7
NH4
Mørebankene
Kystsonen Norskehavet sør
Inkludert i Kystsonen Norskehavet sør. Utvidet nordover slik at området ligger inntil Kystsonen Norskehavet nord.
NH8
Dyphavsområdene i Norskehavet
Nytt område
NH6
Den arktiske front
Utelatt, opprinnelig begrunnelse med forhøyet produksjon er ikke tilstrekkelig dokumentert
Nordsjøen og Skagerrak
NH7
NS7
Bremanger til Ytre Sula
Kystsonen Norskehavet sør
Inkludert i Kystsonen Norskehavet sør
NS1
NS4
Karmøyfeltet
Boknafjorden og Jærstrendene
Inkludert i Boknafjorden og Jærstrendene
NS1
NS8
Boknafjorden og Jærstrendene
Boknafjorden og Jærstrendene
Utvidet med Karmøyfeltet, og litt inn i fjorden
NS2
NS11
Vikingbanken
Tobisfelt
Inkludert i Tobisfelt
NS2
NS5
Tobisfelt sør
Tobisfelt
Inkludert i Tobisfelt
NS3
NS2
Transekt Skagerrak
Norskerenna
Delvis inkludert i Norskerenna
NS3
NS3
Listastrendene og Siragrunnen
Norskerenna
Siragrunnen inkludert i Norskerenna
NS3
NS10
Skagerrak
Norskerenna
Inkludert i Norskerenna
NS4
NS6
Ytre Oslofjord
Ytre Oslofjord
Utvidet med Kandidatområdet for bedre å dekke vinterbeite for sjøfugl
NS9
Korsfjorden
Utelatt, fyller kriteriene, men kun for kystmiljøverdier
NS1
Gytefelt makrell
Utelatt, fyller ikke kriteriene
Tabell 1.1. Oversikt over SVO-navn og -koder fra Meld. St. 20 (2019-2020) og foreliggende rapport. Forslag til endringer er kort forklart .
Knølhval. Foto: Kjell-Arne Fagerheim, HI
2 - Abstract
In the Norwegian Management plans for the Barents Sea, Norwegian Sea and North Sea and Skagerrak, particularly valuable and vulnerable areas (in Norwegian “ Særlig Verdifulle og Sårbare Områder” SVOs) have been identified. SVOs are sea areas of significant importance for biological diversity and production within, and often also outside, the area. SVOs do not directly impose restrictions on business activity; but signal the importance of exercising particular caution in these areas. While preparing the knowledge base for the updated report to the Norwegian parliament on the Management plans for the Norwegian Sea areas in 2019–2020, it became clear that a common and harmonised valuation of all the SVOs in the Norwegian sea areas was needed. In Report to the Storting (white paper) St. 20 (2019–2020) the Norwegian government stated that by the end of 2021 a review of environmental value and vulnerability of all SVOs in Norwegian sea areas should be completed. At the same time, it should be investigated if areas with deep sea mountain ranges, fulfil the criteria for particularly valuable and vulnerable areas.
Commissioned by the management forum for Norwegians sea areas, the Institute of Marine Research has led a national, cross-topic expert group, with experts from eight research institutes, including three universities to perform the first part of this task, i.e. the environmental valuations . Participating research institutions were, beside the Institute of Marine Research, the Norwegian Polar Institute, Norwegian Institute for Nature Research, Geological Survey of Norway, Akvaplan-niva, as well as the Universities of Bergen, Oslo and Tromsø.
The time frame has now been modified, but the complete review, including the second and third part of this task which encompass vulnerability assessment and description of the human activities within the particularly valuable areas, is to be completed by spring 2022.
Even if this first report presents only results related to environmental valuations, not vulnerability assessments, we have chosen to use the term SVO throughout this report, including new areas that have never been the subject of vulnerability assessments. Previous determinations of SVOs indicate that environmental value is decisive in delimiting these areas, not vulnerability; this also applies relative to existing SVOs. Based on previous environmental value assessments and new knowledge, expert groups from nine research institutions representing numerous disciplines have conducted environmental value assessments using criteria defined in the UN Convention on Biological Diversity (CBD) to assess ecologically and biologically significant areas (EBSAs). This enabled the expert group to suggest a revised selection of SVOs.
This present report is a scientific report, where the suggested SVOs are based on a comprehensive compilation of knowledge about the environmental values, using the EBSA-criteria for the environmental value assessment that forms the basis for the delineation of the SVOs.
It is often found that abiotic conditions such as topography, currents, mixing within the water column, or bottom conditions, are crucial for ecological processes, making certain areas particularly important for individual ecosystem components and for ecosystems as a whole. This can be exemplified by fishing grounds, upwelling areas, retention areas and ice cover.
Important areas for spawning/birth/breeding, feeding, overwintering, molting/shedding, and essential habitats for endangered or vulnerable species have been particularly decisive for delineating SVOs in the management plans. Such areas are essential to ecosystem structure and function and are often important for biological production and diversity, often also outside the SVO itself.
In the present assessment, all seven EBSA criteria (uniqueness/rarity, importance for life history stages of species, importance for threatened, endangered or declining species and/or habitats, vulnerability, fragility, sensitivity, or slow recovery, importance for biological productivity, importance for biological diversity, and naturalness) have been implemented. Relative to these criteria, seven environmental value groups have been considered: ice biota, plankton (phytoplankton and zooplankton), fish, mesopelagic fauna, benthic fauna, marine mammals, and sea birds. Physical and chemical factors which affect ecological conditions impacting these groups are described. Ecological interactions within and between these groups are discussed.
The cross-topic expert group agrees that SVOs should reflect interactions between the processes and the environment that forms the basis for the environmental values within the SVO. For this reason, the SVOs should encompass not only a spawning ground, a seabird colony, or selected coral reefs, but also reflect ecological interactions within a larger area. Compared to previous SVOs (Fig. 1.1), it is therefore proposed fewer, but larger areas for each SVO (Fig. 1.2). But even if the SVOs have become larger, the environmental values within the SVOs vary in time and space, i.e. different environmental values are of importance in different seasons and in different parts of the SVOs. This approach led to the greatest SVO increases in the North Sea / Skagerrak and the Norwegian Sea, whereas the Barents Sea already had several large areas. This will facilitate better assessment of status and development over time, as smaller SVOs have rarely had site-specific monitoring.
Figure 1.1. Top) Existing SVOs (Meld. St. 20 (2019-2020): Bottom) Map of candidate areas (orange areas) from the SVO Report (Management forum for Norwegian sea areas 2019 b).
Norwegian sea areas are among the most productive in the world and all are intrinsically valuable. It is, therefore, a challenge to delineate particularly valuable areas. Previous assessments and selections of SVOs offer guidance (Olsen and von Quillfeldt 2003, Ottersen and Auran 2006, Ottersen et al. 2010, von Quillfeldt 2018) together with new information, including reports on important areas for seabirds (Systad et al. 2019) and areas around Svalbard (NP 2019).
The present report describes 19 existing, expanded, or new SVOs partitioned amongst the Barents Sea (seven), the Norwegian Sea (eight), and the North Sea / Skagerrak (four).
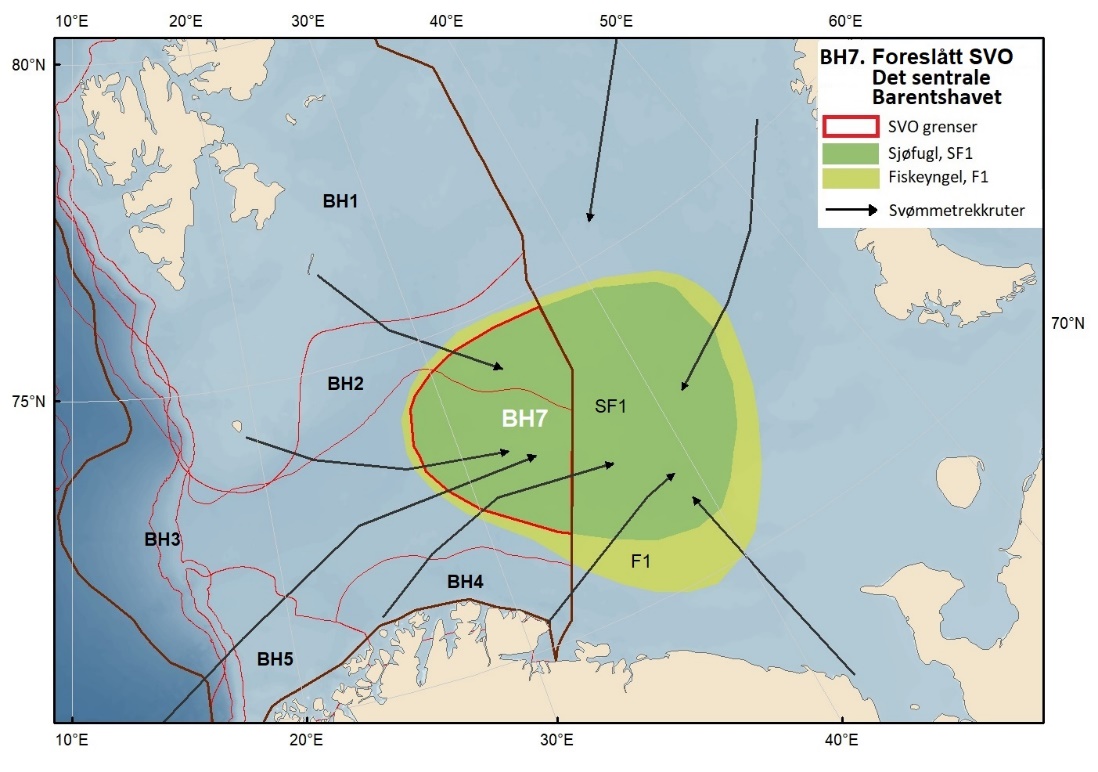
Proposed SVOs in the Barents Sea are shown in Fig. 1.2 and Table 1.1. A new SVO, The central Barents Sea, is particularly important as a destination for seabird – common murre/common guillemots (Uria aalge) and Brünnich’s guillemot/thick-billed murre (Uria lomvia) – swimming migrations after the breeding season. SVOs delimiting the marginal ice zone and the areas around Svalbard include all the environmentally valuable criteria in larger or smaller areas within the SVOs, but with varying significance depending on the season. In the SVO Sea areas around Svalbard, sub-areas with particularly high value have been identified, and the entire SVO has been expanded to include one of the most important grazing areas for capelin (Mallotus villosus) .
SVO Marginal ice zone has been extended to maximum ice extent to capture the part of the spring bloom extending south of the ice edge to establish an important feeding area. Three SVOs —Eggakanten north, The sea areas around Svalbard, and the Marginal ice zone – have some spatial overlap; this is justified, however, by different environmental values that have different effects in areas of overlap. SVO Tromsøflaket has had its borders expanded northward to include rich benthic communities and important transport zones for plankton, such as fish larvae and crustaceans, into the fjord grazing areas for fish, seabirds, seals, and whales. SVO Coastal zone in Finnmark includes the existing SVO Coastal areas from Tromsøflaket to the border with Russia. Coastal SVOs have all been expanded with the seabirds' grazing areas some distance both into the fjords, and out to sea. SVO Tromsøflaket and the SVO Coastal zone in Finnmark simultaneously represent spawning grounds for capelin and other fish species. The SVO Coastal zone of Lofoten includes most of the existing SVO Areas off Lofoten to Tromsøflaket; also included in this SVO is Vestfjorden, which previously was included in the SVO Coastal zone of Norwegian Sea.
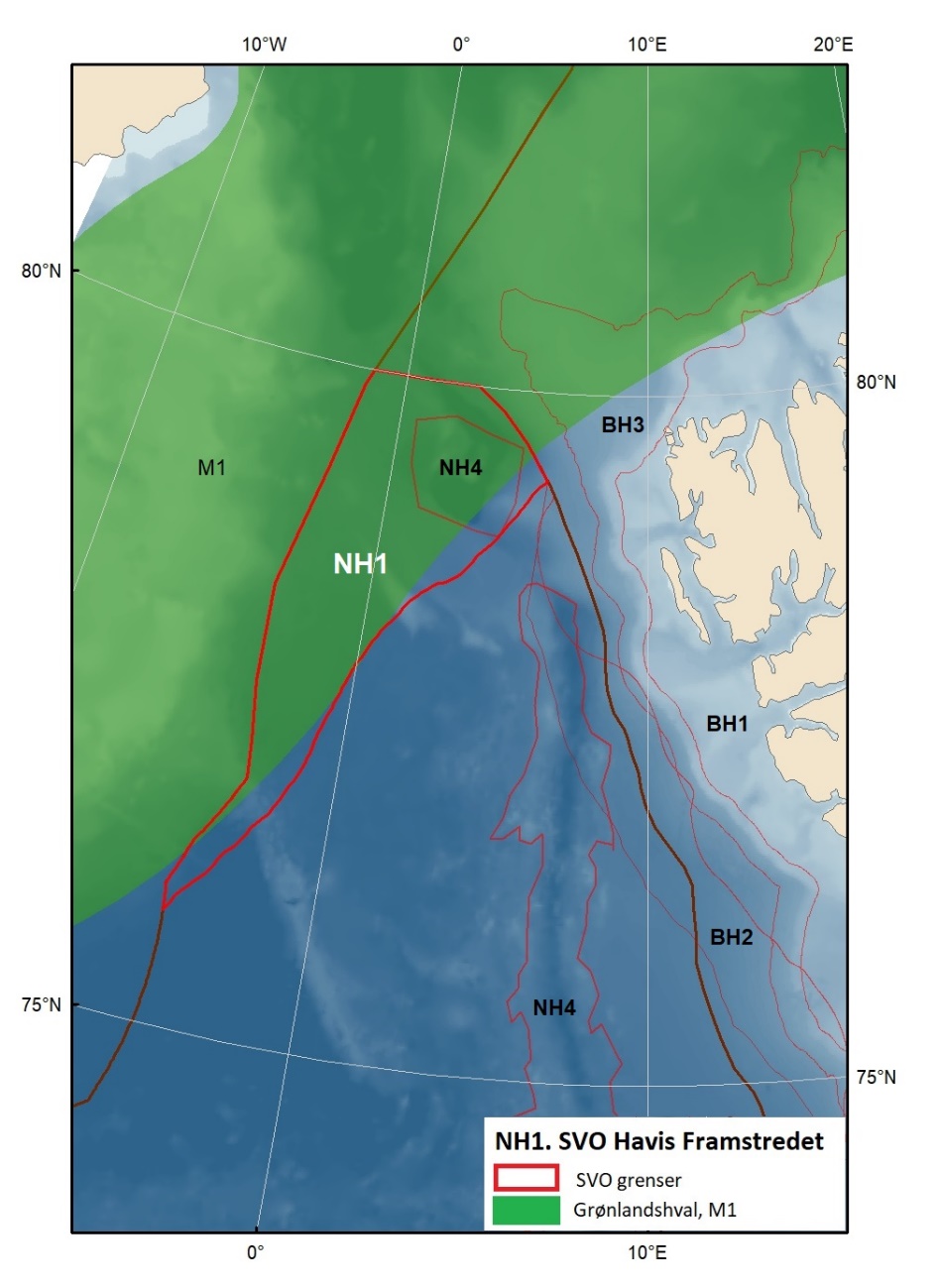
Proposed SVOs in the Norwegian Sea are shown in Fig. 1.2 and Table 1.1. In addition to the unchanged SVOs – SVO Havis Framstredet and SVO Vesterisen – many small coastal SVOs are grouped into two larger coastal SVOs: Coastal zone Norwegian Sea north and Coastal zone Norwegian sea south; both include coastal spawning areas of great importance and seabird colonies strongly associated with fish foraging. SVO Jan Mayen has been expanded to better cover important feeding areas for seabirds. SVO Mid-Atlantic Ridge and the SVO Deep-sea areas in the Norwegian Sea are proposed new SVOs. Similar to SVO Eggakanten south, deep-living life forms are particularly valuable here. The Mid-Atlantic Ridge has an abundance of endemic and unique fauna, is very productive, and is a spawning and rearing area for slow-growing fish species. Benthic communities near and on the Mid-Atlantic Ridge are very vulnerable and have limited or no capacity to recover. Deep-sea areas are overwintering habitats for Calanus spp. and are particularly important for maintaining the populations. These plankton resources are also essential for the production and productivity of fish and seabirds along the Eggakanten and in coastal zone SVOs in both the Norwegian Sea and the Barents Sea.
Proposed SVOs in the North Sea and Skagerrak are shown in Fig. 1.2. and Table 1.1. The largest changes were made in this area, where the number of SVOs has been reduced to four (Fig. 1.2, Table 1.1). Two are coastal SVOs, SVO Boknafjorden and Jærstrendene and SVO Ytre Oslofjord; one in Skagerrak and one in the North Sea. SVO Ytre Oslofjord is an especially important winter-feeding area for seabirds from the entire North Sea area. Boknafjorden, including Karmøyflaket and areas within the fjord, have historically important spawning grounds for Norwegian spring-spawning herring (Clupea harengus / henceforth referred to as herring), larger seal colonies, including the southernmost known breeding colony of grey seal (Halichoerus grypus) , and are important wintering areas for coastal seabirds. Both SVOs are believed to have a particularly high biological diversity since these areas are so varied and include transition zones between fjord, coast, and sea. SVO Ytre Oslofjord has special physical, chemical, and climatic conditions and Oslofjord has significantly higher phytoplankton production than other fjord systems. SVO Tobisfelt includes all sandeel grounds in the Norwegian Economic Zone. The new SVO Norwegian trench covers the North Sea's only deep-water area, with its distinctive fauna including plankton and mesopelagic fish populations. In the north, the existing SVO Bremanger to Ytre Sula has been incorporated into the SVO Coastal zone Norwegian Sea south. Makrellfelt SVO has been removed, as it has been determined that the North Sea mackerel stock is not separate. SVO Skagerrak is included in the new SVO Norwegian trench, while SVO Transekt Skagerrak and other coastal SVOs are partly included in the new SVO Norwegian trench or have been removed since they will not contribute additional value beyond that already covered by the two described coastal SVOs remaining.
Figure 1.2. Map of particularly valuable and vulnerable areas (SVOs) presented in this report. Seven SVOs are located in the Barents Sea (BH1-BH7), eight in the Norwegian Sea (NH1-NH8) and four in the North Sea and Skagerrak (NS1-NS4). The SVO Marginal ice zone (shaded) overlaps with the SVO Sea areas around Svalbard (BH1), parts of SVO Eggakanten north (BH3), and parts of SVO The central Barents Sea (BH7). The boundaries of the management plan areas are outlined in brown.
SVO-code
Miljøverdi-rapport 2021
SVO-code Meld. St.
2019-2020
SVO-name
Meld. St.
2019-2020
SVO-name Environmental values report
2021
Revisions and changes
The Barents Sea
BH3
The Polar Tidal Front
The Sea Areas Surrounding Svalbard
Included in The Sea Areas Surrounding Svalbard
BH1
BH4
The Sea Areas Surrounding Svalbard
The Sea Areas Surrounding Svalbard
Extended to cover the summer feeing groud og capelin
BH2
BH6
The Marginal Ice Zone
The Marginal Ice Zone
Extended to cover the statistical long-term maximal ice coverage in April
BH3
BH7
Eggakanten
Eggakanten North
Southern border at the southern border of the Barents Sea management area and northern border of Eggakanten South. Otherwise no changes.
BH4
BH1
Coastal zone Finnmark
Coastal Zone Finnmark
Extended to the north and to some extent into the fjords to better cover the feeding area of seabirds and sea mammals.
BH5
BH2
Tromsøflaket
Tromsøflaket
Extended south to Senja, north to Sørøya and to some extent into the fjords, incl. Lopphavet
BH6
BH5
Lofoten to Tromsøflaket
Coatsal Zone Lofoten
Changed to cover the Vestfjorden the south and from Røst to Senja west and north, to better cover breeding seabird feeding area, among ither reasons.
BH7
The Central Barents Sea
New area
The Norwegian Sea
NH1
NH11
Sea Ice Framstredet
Sea ice Framstredet
No changes
NH2
NH12
The West Ice
The West Ice
No changes
NH3
NH8
Jan Mayen
Jan Mayen
Extended to better cover seabird
NH4
NH6
The Arctic Front
The Mid-Atlantic Ridge
New area, including parts of the present SVO The Arctic Front, which is suggested removed from the list of dedicated SVOs.
NH5
NH7
Eggakanten
Eggakanten South
Northern border at the southern border of the Barents Sea management area and southern border of Eggakanten North. Otherwise no changes.
NH6
NH1
Iverryggen
Coastal Zone Norwegian Sea North
Included in the Coastal Zone Norwegian Sea North
NH6
NH2
Haltenbanken
Coastal Zone Norwegian Sea North
Included in the Coastal Zone Norwegian Sea North
NH6
NH3
Sklinnabanken
Coastal Zone Norwegian Sea North
Included in the Coastal Zone Norwegian Sea North
NH6
NH5
Froan with the Sula Reef
Coastal Zone Norwegian Sea North
Included in the Coastal Zone Norwegian Sea North
NH6/NH7
NH9
Coastal Zone, The Norwegian Sea
Coastal Zones, The Norwegian Sea North/ South
Included in the Coastal Zone Norwegian Sea North/South
NH6
NH10
Remman
Coastal Zone Norwegian Sea North
Included in the Coastal Zone Norwegian Sea North
NH7
NH4
Mørebankene
Coastal Zone Norwegian Sea South
Included in Coastal Zone Norwegian Sea South. Extended northwards to line up with the Coastal Zone Norwegian Sea North and southwards to include Bremanger to Sula.
NH8
Deep sea basins of the Norwegian Sea
New area.
NH6
The Arctic Front
Suggested removed from the list of dedicated SVOs.due to lack of sufficient documentation of the expected cause for environmental values.
The North Sea and Skagerak
NH7
NS7
Bremanger to Ytre Sula
Coastal Zone Norwegian Sea South
Included in the Coastal Zone Norwegian Sea South.
NS1
NS4
Karmøy field
The Boknafjord and Jæren Beaches
Included in The Boknafjord and Jæren Beaches
NS1
NS8
The Boknafjord and Jæren Beaches
The Boknafjord and Jæren beaches
Extended with the Karmøy Field and to some extent into the fjord .
NS2
NS11
The Viking bank
The Sandeeel Fields
Included in the exteded Sandeel Fields
NS2
NS5
Sandeel Fields South
The Sandeeel Fields
Extended with the Viking Bank
NS3
NS2
Transect Skagerrak
The Norwegian Trench
Partly included in The Norwegian Trench
NS3
NS3
Listas beaches and Siragrunnen
The Norwegian Trench
Siragrunnen is included in the Norwegian Trench
NS3
NS10
Skagerrak
The Norwegian Trench
Included in the Norwegian Trench
NS4
NS6
Outer Oslofjord
Outer Oslofjord
Extended to better cover the winter feeding areas of sea bird.
NS9
The Kors fjord
Removed, due to lack of interactive environmental values to the open sea, although filling several of the EBSA criterias for particular valuable coastal area.
NS1
Mackerel Spawning Field
Removed, does not fill the criterias.
Table 1.1. Overview of SVO names and codes from Meld. St. 20 (2019-2020) and the present report (Miljøverdirapport 2021). Proposed changes are briefly explained.
3 - Innledning
Det er utviklet forvaltningsplaner for alle norske havområder, det vil si Barentshavet og havområdene utenfor Lofoten (St.meld. nr. 8 (2005-2006), oppdatert Meld. St. 10 (2010-2011)), Norskehavet (St.meld. nr. 37 (2008-2009), oppdatert Meld. St. 35 (2016-2017)) og Nordsjøen og Skagerrak (Meld. St. 37 (2012-2013). I alle planene er det identifisert særlig verdifulle og sårbare områder (SVO) på grunnlag av ulike faglige utredninger (se tabell 1.1). I Meld. St. 20 (2014-2015) vises det til at behovet for å oppdatere avgrensingen av SVO-ene Iskant, Polarfront og Havområdene rundt Svalbard vil bli vurdert i forbindelse med revidering av forvaltningsplanen i 2020. Videre er det pekt på at det som en del av det faglige grunnlaget for denne revideringen også vil bli foretatt en gjennomgang av definisjonen som ligger til grunn for beregningen av iskanten. I det faglige grunnlaget i 2019 for stortingsmeldingen om helhetlige forvaltningsplaner for de norske havområdene (Meld. St. 20 (2019-2020) ble dette fulgt opp (Faglig forum for norske havområder 2019). Det faglige grunnlaget viste også at det var behov for en helhetlig gjennomgang av alle SVO-ene. I stortingsmeldingen står det at en slik gjennomgang av miljøverdi og sårbarhet for alle SVO-ene skal sluttføres i løpet av 2021. Samtidig skal det utredes om områder med undersjøiske fjell oppfyller kriteriene for særlig verdifulle og sårbare områder. I Meld. St. 29 (2020-2021) legges det vekt på at SVO-ene skal være sentrale arealer i videre arbeid med å utrede marint vern.
På oppdrag fra Faglig forum har en ekspertgruppe, ledet av Havforskningsinstituttet, gjennomført første del av denne oppgaven, dvs. miljøverdivurderingene. Planen er nå justert, men hele gjennomgangen, inkl. sårbarhet og aktivitet innenfor de verdifulle områdene skal være gjennomført i løpet av våren 2022.
Betegnelsen SVO gir ikke direkte virkninger i form av begrensninger for næringsaktivitet, men signaliserer viktigheten av å vise særlig aktsomhet i disse områdene. For å beskytte verdifulle og sårbare økosystemkomponenter og prosesser (miljøverdier) kan det, for eksempel med hjemmel i gjeldende regelverk, stilles særlige og konkrete krav til aktivitet som utøves.
Denne rapporten er en del av et større arbeid med SVO-er i norske havområder. Den tar kun for seg miljøverdivurderingene. Disse er gjennomført for isbiota, plankton (planteplankton og dyreplankton), fisk, mesopelagisk fauna (bl.a. fisk, krepsdyr og blekksprut), bunnsamfunn (bentos), sjøpattedyr og sjøfugl.
I denne delrapporten er det valgt å bruke betegnelsen foreslått SVO gjennom hele rapporten, for nye og endrete områder som foreløpig ikke har vært gjenstand for sårbarhetsvurderinger. Tidligere arbeid med SVO-er har vist at det er forekomster av miljøverdier som er avgjørende for avgrensning av områdene, mer enn sårbarhet. Dessuten blir det da lettere i forhold til omtale av eksisterende SVO-er.
SVO er områder som har særlig betydning for det biologiske mangfoldet og den biologiske produksjonen i havområdet, også utenfor områdene selv. Alle havområdene er i seg selv verdifulle og utvelgelsen av SVO-er er vanskelig. Med utgangspunkt blant annet i tidligere SVO-utredninger (Olsen og von Quillfeldt 2003, Ottersen og Auran 2007, Ottersen mfl. 2010, Faglig forum for norske havområder 2019 a), et notat om avgrensning av SVO Havområdene rundt Svalbard (NP 2019), en rapport med viktige områder for sjøfugl (Systad mfl. 2019), samt en rapport om tilleggsverdier i kandidatområdene (Faglig forum for norske havområder 2019 b), er alle SVO-er og kandidatområder i de norske havområdene vurdert igjen. I tillegg er forvaltningsplanområdene revurdert i sin helhet for bl.a. å identifisere eventuelle nye forslag til SVO-er.
Områdene er identifisert ved hjelp av forhåndsdefinerte kriterier, hvor betydning for biologisk mangfold og biologisk produksjon er blant de viktigste. Slike viktige områder er ofte der det er spesielle topografiske eller oseanografiske forhold. Eksempler på miljøverdier i særlig verdifulle og sårbare områder er viktige leve- eller gyteområder for fisk, viktige leveområder for sjøfugl og sjøpattedyr og korallforekomster. For å kunne beskrive en helhetlig miljøverdi av hvert område, må verdiene ses samlet i forhold til alle kriteriene. I tillegg skal sårbarheten i forhold til menneskelig aktivitet vurderes i en egen rapport, basert på miljøverdiene som er trukket fram i denne verdivurderingen.
Faglig forum har utarbeidet en plan for arbeidet med særlig verdifulle og sårbare områder (Plan for gjennomgang av Særlig verdifulle og sårbare områder 2020-2022 (2).pdf). Det er også utarbeidet et metodedokument for arbeidet med miljøverdivurderinger (Særlig verdifulle og sårbare områder – metodedokument for miljøverdivurderinger (5).pdf). Faglig forum har pekt på et behov for en ensartet vurdering av miljøverdiene i de eksisterende SVO-ene, tilgrensende kandidatområder og i eventuelle nye SVO-er. Derfor har kandidatområdene som ligger i tilknytning til eksisterende SVO-er ikke blitt vurdert enkeltvis, men sett som en del av de respektive SVO-ene. Dersom vurderingene viste at eksisterende SVO-er ikke fyller kriteriene, ble disse områdene foreslått fjernet (tabell 1).
CBD (Konvensjonen om biologisk mangfold) ble lagt til grunn for vurderingene, noe som også vil gjøre dette arbeidet relevant i internasjonal sammenheng (https://www.cbd.int/ebsa/about). For å gjøre det mer tydelig i tabellen hvilke miljøverdier som slår ut på kriteriene, er det valgt å vise dem hver for seg og ikke gjøre en samlet verdivurdering som EBSA-prosedyren tilsier. Det må i tilfelle gjøres i etterkant skal verdivurderingen bli helt sammenlignbar med øvrige EBSA-områder under CBD. Den valgte løsningen bygger likevel på at verdivurderingene i henhold til EBSA-kriteriene skal være harmonisert mellom og innen fagekspertgrupper for alle miljøverdiene.
Figur 3.1 – Stortareskog. Foto: Espen Bierud, HI
4 - Metodebeskrivelse
I 2020-2021 er alle eksisterende SVO-er, samt kandidatområder vurdert av fagekspertgrupper, hentet fra åtte ulike forskningsmiljøer, med en faglig bredde fra kjemisk og fysisk oseanografi til spisskompetanse på biologiske arter og økologi (Kap. 15). I tillegg til fagekspertgruppene, har en referansegruppe med seks eksperter gått igjennom fagekspertgruppenes vurderinger og begrunnelser. I tillegg er flere kart blitt tegnet og diskutert med referansegruppen for kart, for å legge til rette for at kartene skal bli tilgjengelige også i Arealverktøyet.
Arbeidet startet våren 2020, men på grunn av smittevernrestriksjoner og tilpasning av nye samarbeidsformer, kom selve rapportarbeidet først i gang for fullt høsten 2020. Det ble gjennomført en rekke nettmøter, der EBSA-vurderingen ble forklart og hvert SVO ble diskutert på tvers av fagekspertgruppene. Hver fagekspertgruppe hadde dessuten interne diskusjoner og jobbet sammen fram begrunnelsene for vurderingene i rapporten.
I januar 2021 ble det gjennomført en tredagers digital workshop, med alle ekspertene for å gå gjennom miljøverdivurderingene for alle kriteriene, samt valg av miljøverdier i alle SVO-ene. Målet var å harmonisere vurderingene på tvers av fag, miljøverdi og område. Faglig forum var invitert som observatører, for å kunne gi innspill på tekst og begreper som måtte gjøres lettere forståelig for lesere uten fagbakgrunn.
Et siste arbeidsgruppemøte ble avholdt tidlig i februar, for å fastslå områdegrensene og legge en plan for eventuelle mangler i tekst, figurer og EBSA-tabellene. I mars ble en samlet rapport for alle de tre forvaltningsplanområdene sendt til referansegruppa for kvalitetssjekk og til Faglig forum for å sikre at språk og begrunnelser var forståelige.
Endelig rapport ble oversendt til Faglig forum 28. april 2021.
4.1 - EBSA Miljøverdivurdering, metodebeskrivelse
EBSA omfatter sju kriterier, som hver for seg kvalifiserer til betegnelsen som EBSA. Det er i tillegg fire graderinger av relevansen av den faglige begrunnelsen.
De sju kriteriene er:
Unikhet/Sjeldenhet (Uniqueness or Rarity)
Livshistorisk viktige områder (Special importance for life history stages of species)
Viktighet for truede eller nedadgående arter og/eller habitater (Importance for threatened, endangered or declining species and/or habitats)
Sårbarhet, skjørhet, følsomhet eller lav restitusjonsevne (Vulnerability, Fragility, Sensitivity, or Slow recovery)
Viktighet for biologisk produktivitet (Biological Productivity)
Viktighet for biologisk mangfold (Biological Diversity)
Naturlighet (Naturalness)
Hva som må legges til grunn for hvert kriterium er gitt i tabell 4.1.
Metoden er internasjonalt brukt for å gi en samlet verdi (kryss) for hvert kriterium, fulgt av faglige begrunnelser hentet fra publisert dokumentasjon. I begrunnelsen skal det fremkomme hvordan ulike miljøverdier har bidratt til relevansen for hvert kriterium. I denne rapporten har vi imidlertid valgt å synliggjøre i tabellen hvilke miljøverdigrupper som slår ut for hvert kriterium fremfor kun å vise samlet verdi angitt som et kryss. Vår tilnærming gjør det lettere å raskt se hva som slår ut med hvilken relevans for hvert kriterium, noe som i enkelte tilfeller kan være mer hensiktsmessig for praktiske formål i for eksempel forvaltningen. Når det gjelder krav til dokumentert begrunnelse har vi fulgt samme prosedyre som i CBD.
Kriterium
Beskrivelse
Unikhet/sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, populasjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem; og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Sårbarhet, skjørhet, følsomhet eller lav restitusjonsevne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet.
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter eller har et høyere genetisk mangfold.
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
Tabell 4.1. EBSA-kriteriene med beskrivelser av kriteriekravene (Kilde CBD).
Gradering av relevans:
«Ikke tilstrekkelig kunnskap»: svært sannsynlig at en miljøverdi har betydning, men ikke publisert kunnskap
«Lav relevans»: Kriteriet oppfylles, men verdien finnes også utenfor området
«Middels relevans»: Kriteriet oppfylles og dette er et sentralt område for denne verdien
«Høy relevans»: Kriteriet oppfylles og dette området er helt sentralt for denne verdien
Makrell. Foto: E. Svensen
5 - Pågående og framtidige endringer
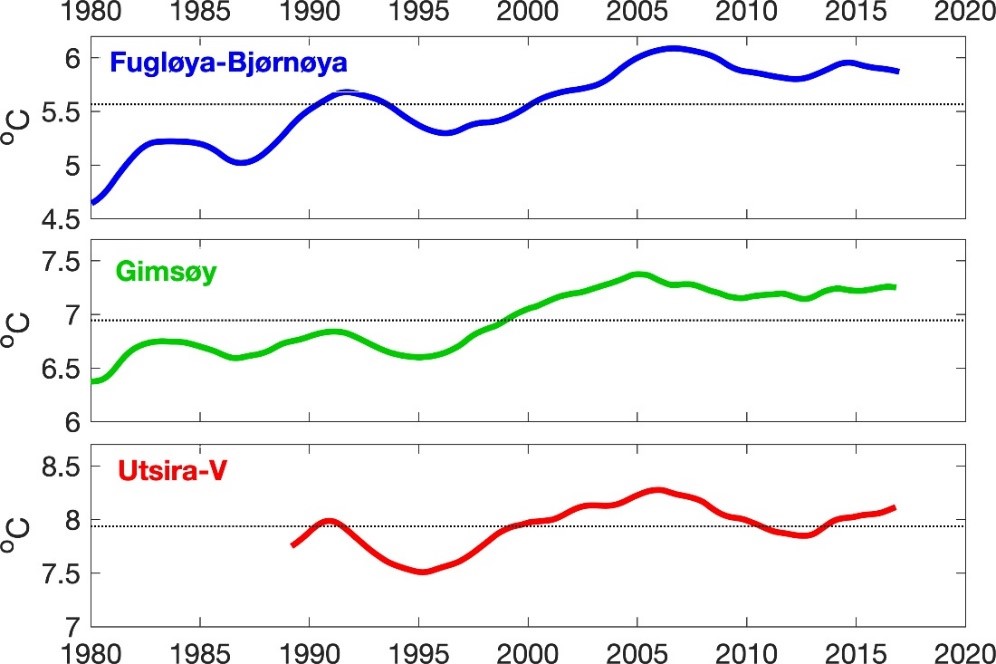
I store trekk varierer temperatur i de norske havområdene i takt (figur 5.1). Temperaturvariasjonene er i stor grad styrt av variasjoner i atlanterhavsvannet lenger sør, men også modifisert av lokalt varmetap og blanding på vei nordover. Havforskningsinstituttets overvåkning viser at over en 40-års periode steg temperaturen i atlanterhavsvannet i Nordsjøen, Norskehavet og Barentshavet frem til ca. 2006, og var på det tidspunktet rekordhøye (González-Pola mfl. 2019) (figur 5.1). Siden har temperaturen i Nordsjøen og Norskehavet stabilisert seg og også gått noe ned, mens temperaturøkningen i Barentshavet nådde ny topp rundt 2015-2016 før den gikk litt ned frem mot 2020. Til tross for temperaturnedgangene har havområdene vært varmere enn langtidssnittet i perioden 1982-2010. Den midlertidige nedgangen i temperatur skyldes naturlige variasjoner i klimasystemet, og prognosene fra International Panel for Climate Change (IPCC, 2019) viser at på sikt vil økningen i havtemperatur vil fortsette, og dessuten at vi må vente økt forsuring, i likhet med den globale utviklingen for boreale og polare havområder. IPCC-rapporten trekker fram påvirkninger på økosystemene og biologiske prosesser, i global skala. I det følgende er overordnede framtidsutsikter for effekter av klimaendringer som gjelder alle de tre norske havområdene kort beskrevet for de syv miljøverdigruppene isbiota, plankton, fisk, mesopelagisk fauna, bunnsamfunn, sjøpattedyr og sjøfugl. Deretter blir prognoser for havforsuringsprosessen i de norske havområdene presentert.
Figur 5.1. Tidsserier av temperatur i atlanterhavsvannet i Nordsjøen (Utsira-V-snittet), Norskehavet (Gimsøysnittet) og Barentshavet (Fugløya-Bjørnøya-snittet).
5.1 - Endringer som følge av økt havtemperatur
Økning i temperatur fører til en rekke økologiske endringer, som konkurransen mellom ulike arter, forskyving i tilgang på mat til tidlige livsstadier og forflytninger. Biogeografiske endringer er observert i Barentshavet og Nordsjøen og kan i nær fremtid forekomme i sørlige Norskehavet dersom økningen i havtemperaturen fortsetter. Dette vil kunne ha konsekvenser for produksjonen av dyreplankton. Arter med høyere optimal temperatur vil kunne bli mer tallrike og sammen med en betydelig reduksjon i isdekket, har dette allerede ført til en nordlig ekspansjon av boreale arter i alle norske havområder og en reduksjon av arktiske arter i Barentshavet. Denne utviklingen er ventet å fortsette i framtiden (Edwards mfl. 2016, Fossheim mfl. 2015, ICES 2020).
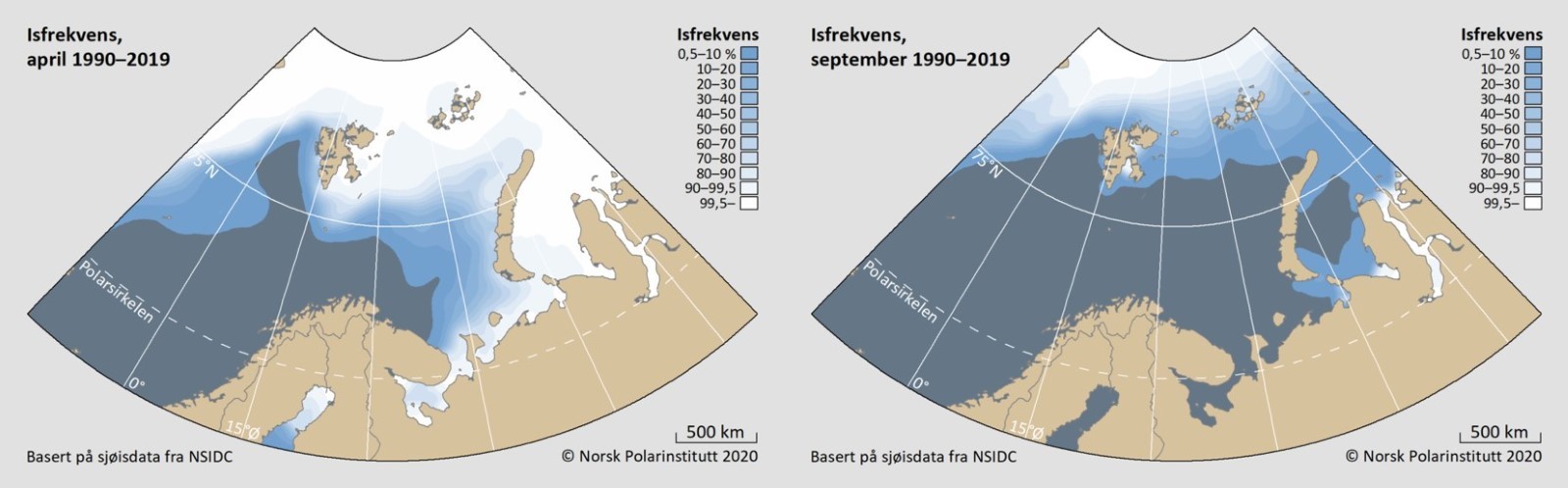
Isutbredelsen er allerede betydelig redusert, og perioden med isdekke er kortere. Framtidsscenarioer antyder at overflatetemperaturene i dette området vil øke med 1-1,5 °C frem mot 2060-2069 (Hanssen-Bauer mfl. 2018). Isbiota og andre arter med hele livssyklusen i havis, står i fare for å forsvinne eller bli sterkt redusert selv om det finnes mekanismer som bidrar til å holde isfauna i vannmassene (Berge mfl. 2012, Kunisch mfl. 2020).
Endring i dyreplanktonsamfunn kan gi betydelig endring i næringsinnhold og fettsyreprofiler, som igjen påvirker overlevelse og vekst hos fiskelarver (Deschutter mfl. 2019). I likhet med andre havområder har klimaendringer i norske havområder ført til at flere planktonarter forflytter seg i takt med et varmere hav. Dette er vist både i Nordsjøen (Beaugrand mfl. 2002, O’Brien 2013, Edwards mfl. 2016) og i Barentshavet (Aarflot mfl. 2018, Stige mfl. 2019, Eriksen mfl. 2017). For Norskehavet er det imidlertid ikke ventet vesentlige endringer i dyreplanktonbiomasse eller -produksjon i nær fremtid siden dette havområdet er sterkt påvirket av atlanterhavsvann og polare vannmasser, deriblant smeltevann fra Grønlandsbreene (ICES 2020, Kjesbu mfl. innsendt).
I et varmere hav utvider flere fiskearter sitt leveområde nordover (Shackell mfl. 2012, Engelhard mfl. 2013). Dette gjelder i alle tre havområder. Beiteområdene for planktonspisende fisk utvider seg i takt med at planktonarter forflytter seg. Oppvarmingen kan føre til endring i reproduksjonssuksess, fordi de tidlige livsstadiene ikke lenger opptrer samtidig som føden er tilgjengelig, og forskyvning av utbredelsen til ulike arter vil avhenge av at fisken finner nye gyteplasser lenger nord. I nord får de arktiske artene større konkurranse, mens de trolig ikke har muligheten for å trekke videre nordover ut i polhavsbassenget (Fossheim mfl. 2015, Eriksen mfl. 2017).
Dyphavene og miljøverdiene der reagerer senere på havoppvarmingen og det er ikke forventet særlige endringer på grunn av klimaeffekter (Sutton mfl. 2017).
Derimot er det sett en forskyving nordover av mengde bunndyr og varmekjære arter av bunnsamfunn, særlig i Barentshavet (Jørgensen mfl. 2015, Manushin mfl. 2021), men også i kystnære farvann langs fastlandskysten brer mer varmekjære arter seg nordover (Brattegard 2011).
Hekkesuksessen til sjøfuglene blir negativt påvirket om klimaendringer fører til at fiskearter endrer preferanser for gytefelt og trekker vekk, eller at de gyter usynkront med fuglenes hekking (Systad mfl. 2019). Det kan også tenkes at tilgjengeligheten av fiskelarver øker i noen områder som følge av endrede gyteområder og strømforhold. Nedgangen i sjøfuglbestandene som allerede har foregått over lengre tid (Barrett mfl. 2006, Fauchald mfl. 2015), er forventet å fortsette.
For sjøpattedyr er det mest i nord at oppvarmingen har vært negativ for en rekke særlig sensitive arktiske sel- og hvalarter (Laidre mfl. 2008, Hamilton mfl. 2017, 2018 og 2019a,b). I Nordsjøen og Skagerrak er det ikke noe klart trendmønster (Nilssen og Bjørge 2019).
5.2 - Endringer i pH-nivået
Siden den industrielle revolusjonen har pH nivået i våre havområder avtatt med rundt 0,1 enheter (Jiang mfl. 2019). Denne havforsuringen er drevet av havets opptak av menneskeskapt CO2 og gjør at pH reduseres. Surere vann kan gi problemer for ulike typer marint liv fordi metningsgraden for de ulike kalkmineralene (aragonitt og kalsitt) som brukes i skall og korallrev synker; dette kan være problematisk for, for eksempel kalkflagellater og koraller. pH og metningsgrad for kalkmineraler er nøkkelindikatorer for havforsuring.
I norske havområder finner vi lavest pH i det varme atlanterhavsvannet, mens den laveste metningsgraden for kalkmineraler forekommer i kaldere vann, altså i arktiske vannmasser mot nord og vest (Fransner mfl. 2020). Metningsgraden avtar dessuten nedover i vannsøylen, og i norske havområder finner vi i dag metningshorisonten for aragonitt på rundt 2000 m, dvs. at alt vann dypere enn dette er undermettet med dette kalkmineralet. Kalsitt er mindre løselig enn aragonitt, og hele vannsøylen er overmettet.
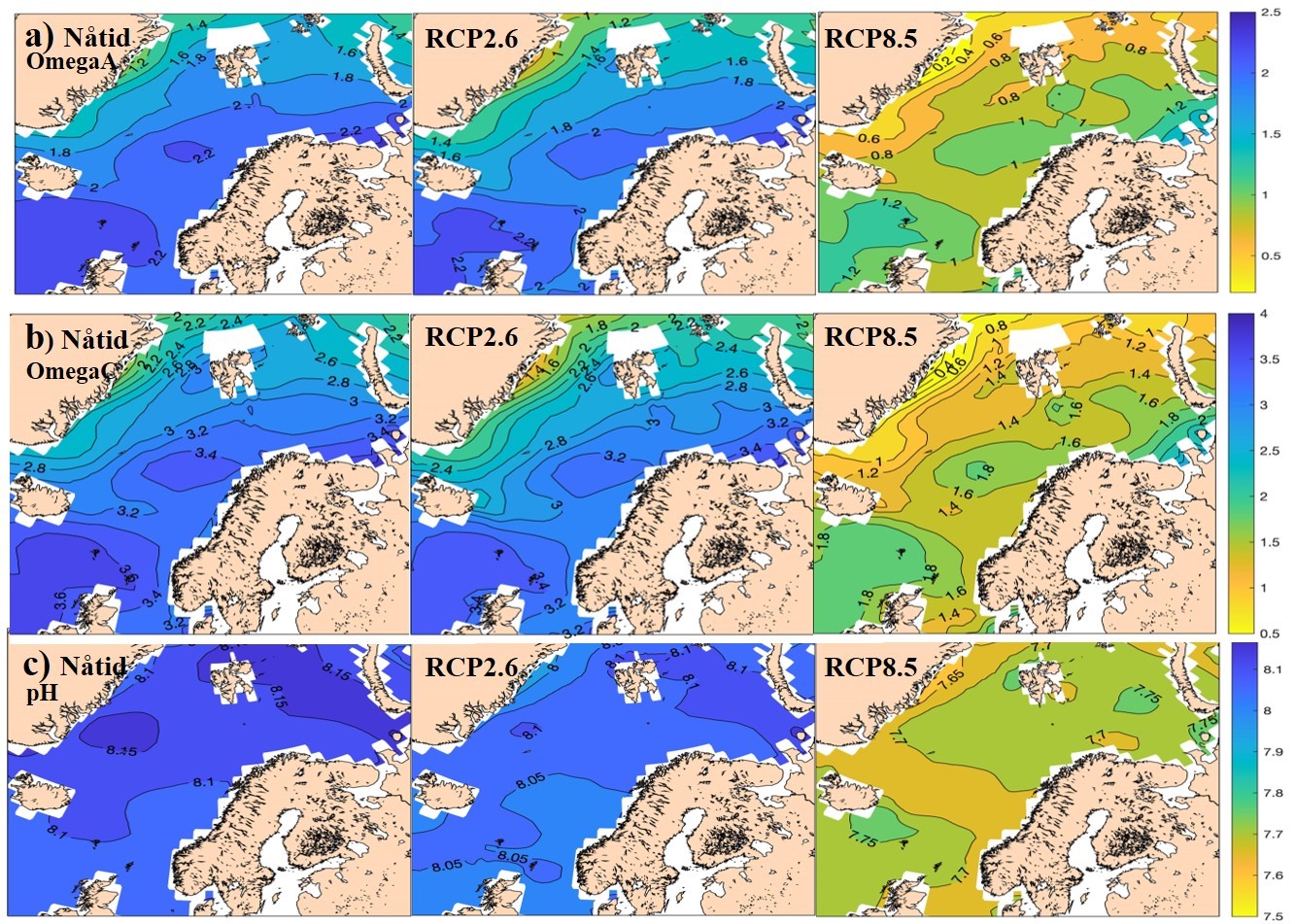
Måleserier siden 1990-tallet viser en signifikant nedgang i pH i nesten alle områder der vi har tilstrekkelig med data til å undersøke trendene i Nordsjøen og Norskehavet (Fransner mfl. 2020). Det eneste unntaket er sørvestre Barentshavet, der pH avtar, men trenden ikke er signifikant. I bunnvannet i de nordlige delene av Barentshavet er det imidlertid observert havforsuring og lav aragonittmetning (lavere enn 1,2; Chierici and Fransson 2018), og dette kan ha alvorlige konsekvenser for kalsifiserende marine organismer som for eksempel vingesnegl (Manno mfl. 2017, Niemi mfl. 2021). Trendene i pH ligger på rundt 0,02 – 0,03 enheter per år i overflatevannet, reduseres gradvis nedover i dypet, og er typisk ikke signifikante på større dyp enn 1000-2000 m. Simuleringer med den norske jordsystemmodellen viser at dersom utslippene fortsetter å øke i samme takt som nå (IPCC scenario RCP 8.5) vil pH ligge på rundt 7,7 i overflatevannet ved slutten av århundret. Dersom vi klarer å redusere utslippene i pakt med 2-graders målet (IPCC scenario RCP 2.6) vil pH være omtrent som i dag.
Metningsgraden for aragonitt er typisk i overkant av to i atlanterhavsvannet (dvs. 2x metnings-konsentrasjonen), og rett under to i arktiske vannmasser. Alt dette er lavt i forhold til resten av verdenshavene, der graden av metning er rundt tre-fire. I overflatevannet synker denne metningsgraden nå med typisk 0,01 til 0,02 enheter per år (Fransner mfl., 2020). Dersom utslippene forsetter å øke i samme takt som frem til nå (IPCC scenario RCP 8.5) vil store deler av våre havområder være undermettet med aragonitt før utgangen av dette århundret (Frasner mfl. 2020, Lauvset mfl. 2015), dette kan gi problemer for kaldtvannskoraller og vingesnegl. For kalsitt er ikke situasjonen like kritisk. Selv om trenden er omtrent den samme som for aragonitt, er dette mineralet mye mindre løselig. Vi forventer ikke undermetning ved slutten av århundret, selv under de mest pessimistiske (RCP 8.5) utslippsscenariene (Fransner mfl. 2020, Lauvset mfl. 2015).
Figur 5.2. Modellert nivå på (a) metningsgrad for aragonitt, (b) metningsgrad for kalsitt og (c) pH for nåtid (middel over årene 2010-2020) og for slutten av dette århundret (middel over perioden 2090-2100) for to ulike IPCC -scenarier, RCP2.6 og RCP8.5. Kartene er laget ved å ta utgangspunkt i klimatologiske kart for disse dataene (Lauvset mfl. 2016) som er sentrert på 2002, og legge på trender simulert med den norske jordsystemmodellen (NorESM), som beskrevet i Fransner mfl. (2020). Kystområder dekkes ikke, og de andre hvite flekkene er områder som er blanket ut i klimatologien pga. for store usikkerheter.
6 - Kunnskapsbehov
Det ble i rapporten Særlige verdifulle og sårbare områder: Faggrunnlag for revisjon og oppdatering av forvaltningsplanene for norske havområder (von Quillfeldt 2019), vist til hvor viktig langsiktig overvåking er, både av fysisk miljø og økologiske komponenter i havområdene generelt, men også spesielt for de beskrevne miljøverdiene som ligger til grunn for at et område kvalifiserer som SVO. Det vises her til de forskjellige behovene og utfordringene som er knyttet til bunnen, frie vannmasser, klima- og miljøpåvirkninger og på de ulike miljøverdiene som har spesielle behov på ulike tider av året, i livssyklusen og i dynamiske økosystem. Kartleggingen i regi av Mareano har vist seg verdifull, men mye gjenstår å kartlegge, i alle de norske havområdene. De nyetablerte, faste observasjonsplattformene Lofoten-Vesterålen Cabled Observatory (LoVe, et samarbeid mellom Havforskningsinstituttet, Equinor og Metas (Rogne og Johnsen 2019)) er utvidet med ytterligere plattformer, som vil kunne gi ny kunnskap om bunnsamfunn i Lofoten-Vesterålen-området, fra grunt vann og etter hvert ned til 2500 m dyp. Disse observasjonsplattformene vil kunne levere viktig og ny kunnskap døgnet rundt, fra endringer i atferd pga. forstyrrelser til naturlige endringer knyttet til fysiske parametere, samt innsikt i dyphavsøkologi og økologisk samspill generelt som tidligere ikke har vært mulig å fange opp.
Utvikling av ny observasjonsteknologi, inkludert autonome fartøy (for eksempel seildroner, lytte- og måleredskap, som trådløst sender informasjon, vil kunne gi svært mye ny informasjon. Behov for kvalifiserte forskere til å bearbeide og analysere all informasjonen vil være viktig.
Mye av overvåkingen har historisk vært knyttet til kunnskap som er viktig for forvaltning av havet, ikke minst fiskeriene, men også olje- og energisektoren. Dette er viktig kunnskapsinnhenting, der ytterligere overvåking må knyttes til videre næringsutvikling i havet, som mineralutvinning og vindenergi. Dette blir fulgt opp videre i neste del av SVO-gjennomgangen der sårbarhetsvurderingen av de miljøverdiene som er trukket fram i denne foreliggende rapporten, skal gjennomføres. Betydningen av fortsatt klima- og forsuringsovervåking blir ikke behandlet her, men dette er grunnleggende kunnskap i arbeidet for å oppnå økt økologisk og artsspesifikk innsikt.
I tillegg til å fortsette de verdifulle, lange tidsseriene, er det behov for særlig overvåking direkte på de ulike miljøverdiene SVO-ene omfatter, knyttet til både brede økologiske prosesser og de enkelte miljøverdiene. Noen særlig utvalgte kunnskapsbehov, men ikke en uttømmende liste, er gitt under:
6.1 - Økologiske prosesser
Det er behov for mer kunnskap om bentisk-pelagiske koblinger. Det er begrenset kunnskap om den økologiske rollen og dermed verdien, av ikke-kommersielle arter, særlig bunnlevende små fiskearter og virvelløse dyr. Dette gjelder både for sokkelhavene Barentshavet og til dels Nordsjøen og i dyphavet Norskehavet. I dyphavet vil dette også gjelde koblingen mellom bunndyr, det mesopelagiske sjiktet mellom bunnen og 200 m dyp og artene over 200 m, i lyssonen opp mot overflaten.
6.2 - Isbiota
Isbiota inngår i verdivurderingen i forslag til utvidet SVO Iskantsonen, forslag til utvidet SVO Havområdene rundt Svalbard og eksisterende SVO Havis Framstredet. Samlet utgjør dette ulike isforhold, dvs. fastis – drivis; årsis – flerårsis; hvor lenge det er is i området; driftsmønster m.m., men kunnskapsbehov for isbiota i disse tre områdene er sammenlignbare.
Det er behov for mer kunnskap om:
Primærproduksjon og biodiversitet i ulike sesonger og hvordan dette endres med endrete isforhold?
Hvordan biologiske prosesser påvirkes av at isen dannes senere og smelter tidligere enn før.
I hvilken grad det relative forholdet mellom is- og pelagisk produksjon endres med endrete isforhold.
Relevans organisk produksjon i isen har for næringsnettet, dvs. fra dyreplankton til sjøfugl, sel og hval, samt eksport til bunnsamfunn.
Særlig for Framstredet: Det forventes mindre eksport av flerårsis gjennom Framstredet; vil dette føre til tap av endemiske arter?
Det bør etableres et overvåkingsprogram hvor man følger fordeling av produksjon i is, vannsøylen og på havbunnen gjennom målinger på utvalgte kystnære lokaliteter med noen års mellomrom.
6.3 - Plankton
Produksjonen av plante- og dyreplankton er et mål på den totale produksjonskapasiteten til økosystemet. Artssammensetningen, samt produksjonen av enkelt-arter/grupper innenfor et område, er viktig for å kunne kvantifisere trofiske interaksjoner. Det fullstendige artsmangfoldet av planteplankton og dyreplankton, fra mikroorganismer til maneter, er derimot ikke beskrevet for de foreslåtte SVO-ene og de fleste områdene mangler in situ produksjonsestimater.
Det er behov for:
Bedre kunnskap om plante- og dyreplanktonets artssammensetning gjennom hele sesongen, og eventuelle mellomårlig endringer og langtidstrender.
Bedre forståelse av hvilke faktorer som driver endringer i fordeling, mengde og fenologi hos sentrale arter/grupper av dyreplankton.
Kunnskap om rekrutteringsmekanismene til økologisk viktige dyreplanktonarter, særlig Calanus spp.
Bedre produksjonsestimater for plante- og dyreplankton arter/grupper på storskala nivå, og i lokale områder som er særlig viktige for høyere trofisk nivå, som f.eks. sentrale områder for fiskelarver og -yngel.
Etablere årsakssammenhenger mellom planktondynamikk og fordeling og mengde av bestander på høyere trofisk nivå.
Bedre forståelse av bentisk-pelagiske koblinger.
6.4 - Fisk
Fisk er en relativt godt beskrevet miljøverdi innenfor de foreslåtte SVO-ene, selv om det er en hovedvekt på kommersielle arter. Det er behov for mer kunnskap om ikke-kommersielle arter, særlig de mindre, bunnlevende artene da disse kan ha en viktig funksjon i økosystemet som bentisk-pelagisk kobling i næringsnettet. For en del ikke-kommersielle arter finnes nyttige data fra tidsserier av forskningstokt gjennomført med finmasket forskningstrål (Wenneck 2005), håv-redskap og akustiske målinger. Grundig analyse av slike data vil kunne gi mer kunnskap.
6.5 - Mesopelagisk fauna
I dypbassengene i Norskehavet er det behov for mer kunnskap om mesopelagisk fisk for å få en bedre innsikt i arter, mengde, livshistorie og økologiske sammenhenger. Pelagiske samfunn i den mesopelagiske sonen (200 til 1000 m) er i begrenset grad undersøkt. Sammenhenger innenfor dette samfunnet og de gjensidige påvirkningene mellom det mesopelagiske økosystemet og økosystemene i de øvre vannmasser er ikke godt nok undersøkt (St. John mfl. 2016).
6.6 - Bunnsamfunn
Bunnsamfunnet i forslag til utvidet SVO Iskantsonen bør følges opp for å få mer kunnskap om den bentisk-pelagiske koblingen (forhold mellom dyr på bunnen og livsformer oppe i vannsøylen) og hvordan dette arktiske bunnsamfunnet påvirkes av klimaendringene.
Det er betydelige kunnskapsbehov når det gjelder dyreliv og økologi for sjøbunnen generelt, men særlig når det gjelder dyphavet i Norskehavet og den midtatlantiske rygg (Mareano.no, van der Meeren mfl. 2021).
6.7 - Sjøpattedyr
Alle norske havområder har marine pattedyrbestander, men kunnskapen om deres antall, utbredelse og økologi er mangelfull for mange av disse.
Det er behov for mer kunnskap om:
Bestandsstørrelse og trender, særlig for de arktisk endemiske artene hvor informasjon er enten ikke-eksisterende eller fra flere tiår siden.
Utbredelse for en rekke arter, som spermhval, blåhval, finnhval, kvitskjeving m.m.
Hvordan de ulike marine pattedyrartene responderer på pågående endringer i deres habitater – vi trenger å studere endringer i utbredelse (satellittsporing, akustikk) og responser på endringer i byttedyrtilgang (diettstudier – med bruk av minimalt invasive metoder for rødlistearter).
Er dagens høsting av ikke-kommersielle arter bærekraftig (når nok data om bestandsforhold blir tilgjengelig).
Potensielle effekter av endret støyregime under vann som følge av økt skipstrafikk, olje og gassutvikling, seismikk, på ulike marine pattedyr.
Konkurranse og predasjonsforhold som følge av at andre marine pattedyr og andre taxa (f.eks. ulike haiarter) endrer utbredelsesområdet mot nord.
Risikovurdering i forhold til økt sykdomseksponering som følge av at boreale arter kommer mer og mer inn i arktiske områder og endringer i eksponeringsgrad til ulike forurensningsstoffer (inklusiv mikroplast).
Kumulative effekter av alle disse potensielle påvirkningsfaktorene – som bør være et hovedfokusområde for fremtidige studier på marine pattedyr i norske havområder.
Nasjonale overvåkningsprogrammer for marine pattedyr må utvides for at vi skal kunne fremskaffe data for en god forvaltning av disse bestandene.
6.8 - Sjøfugl
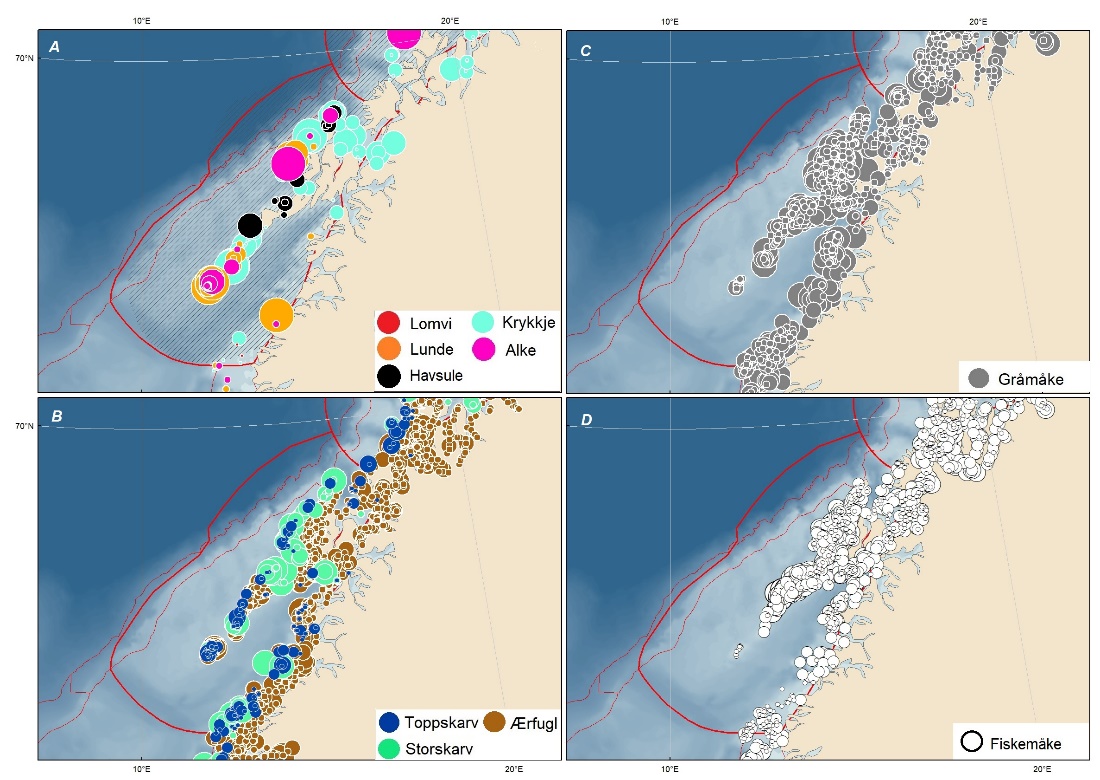
Sjøfugl overvåkes og kartlegges i norske havområder gjennom forskningsprogrammet SEAPOP og prosjektet SEATRACK som er tilknyttet dette. Disse prosjektene har vært helt essensielle for oppbyggingen av den kunnskapen vi har om sjøfugl nå, men må fortsettes slik at vi kan følge de store endringene i systemene som vi har sett utvikle seg.
Vi mangler kunnskap rundt følgende temaer:
Kartlegging av utbredelse må gjennomføres jevnlig, og er hittil gjort i et tiårsperspektiv.
Nøkkellokalitetene må gjennomgås i forhold til representativitet.
Forekomster av nye arter som ikke er dekket av pågående forskning må følges opp – flere arter er kommet til og har fått større betydning i norske havområder.
Ikke-hekkende sjøfuglbestander i norske havområder må kartlegges bedre. Nåværende kartlegging i åpent hav er todelt – den tradisjonelle metoden har vært båttransekter, og dette arbeidet har primært dekket havområdene langs norskekysten. SEATRACK basert på loggere på utvalgte sjøfuglbestander har tatt over mye av denne kartleggingen. Data for ungfugl som ikke har startet hekking, og bestander med annen hekkeutbredelse enn i norske kolonier er eksempler på manglende kunnskap.
Forandringer i næringsforhold på grunn av endringer i fiskebestandene og gyteområder bør ha et særlig fokus.
Kunnskap om prosesser som driver endringene i sjøfuglbestandene må bedres på generell basis. Dette gjelder hvilke effekter som har betydning, kumulative effekter, kaskadeeffekter med betydning for sjøfuglsamfunnene og økosystemene i sin helhet.
Flere av de samme problemstillingene som er oppgitt for sjøpattedyr gjelder også for sjøfugl.
7 - Leserveiledning til SVO-kapitlene
I de påfølgende kapitler blir hvert forvaltningsplanområde presentert for seg i kapittel 8–10. Hvert av disse kapitlene innledes med en oversikt i kart og tabell over alle de eksisterende og foreslåtte SVO-ene som beskrives i det aktuelle forvaltningsplanområdet. Dette er fulgt av en kort generell innledning som beskriver hele forvaltningsplanområdet geografisk og topografisk, med hensyn til fysisk og kjemisk oseanografi der strømmer, vannmasser og vannkvaliteter av betydning for økosystemene blir omtalt. Den generelle innledningen inneholder også en beskrivelse av dyre- og planteplankton, for hele forvaltningsplanområdet. Plankton styres bl.a. av lys, vind, vannmasser og de store havstrømmene i så stor grad at det ikke er mulig å forstå systemet bare ved å beskrive plankton innenfor hvert enkelt SVO. Pågående og framtidige endringer som følge av klimaendringer og havforsuring beskrives også på overordnet nivå for hvert forvaltningsplanområde, før den generelle beskrivelsen av hvert havområde avsluttes med en kort oversikt over løpende kartleggings- og overvåkningsaktivitet innenfor området.
Etter den overordnede beskrivelsen av hvert havområde, blir forslag til nye og endrete SVO-er samt eksisterende SVO-er som foreslås videreført, presentert område for område. Presentasjonen av hvert enkelt av disse områdene innledes med et sammendrag som beskriver den geografiske lokaliseringen og de miljøverdiene som er særlig relevante for området.
Videre følger et kart der miljøverdiene er vist med romlig fordeling, etterfulgt av tabellen med gradering av miljøverdiene i henhold til EBSA-kriteriene, med kort begrunnelse for hvert kriterium. Ikke alle kriteriene er nødvendigvis relevante for et område eller en miljøverdigruppe, og der vil begrunnelsesfeltet for kriteriet være blankt. Der det forekommer flere miljøverdilag i kartet enn de som er beskrevet i tabellen, er det på grunn av direkte eller indirekte økologiske sammenhenger med betydning for miljøverdiene i tabellen. Slike sammenhenger er nærmere beskrevet i etterfølgende underkapitler.
For hvert område er det så en mer detaljert beskrivelse av lokalisering og avgrensing av området, samt eventuelle endringer i forhold til eksisterende SVO-er beskrevet i Meld. St. 29 (2020-2021), f.eks. endring i avgrensing, navn eller utvidelser på grunn av kandidatområder for sjøfugl. Der det er overlapp mellom SVO-er, er dette også påpekt. Ved overlapp mellom SVO-er, er det ulike miljøverdier og biologiske prosesser som er vektlagt i de overlappende SVO-ene.
Etter kapitlet om lokalisering, kommer underkapitler som utdyper områdets topografi, inkl. fysiske og kjemiske forhold, samt alle miljøverdiene og sammenhengen mellom de ulike miljøverdigruppene. Dersom miljøverdiene er til stede kun i en begrenset del av året eller kun i deler av området, blir dette beskrevet her. Spesialkart for enkelte miljøverdier er lagt til i miljøverdibeskrivelsene, der oversiktskartene ikke har tilstrekkelig detaljert informasjon. Isbiota og mesopelagisk fauna er bare omtalt i relevante områder.
For noen områder der det finnes mer stedsspesifikke endrings- og fremtidsvurderinger, er dette også beskrevet. Dette gjelder de foreslåtte SVO-ene Havområdene rundt Svalbard, Iskantsonen og Midtaltantisk rygg samt eksisterende SVO Havis Framstredet.
Etter alle vurderingene, presenteres en referanseliste for hvert hovedkapittel (Kap. 11–14) og liste med ekspertene som har vært involvert, enten i fagekspertgrupper eller som referansepersoner (Kap. 15).
Helt til slutt er det lagt ved en oversikt over marine verneområder og andre effektive arealbaserte bevaringstiltak som berører hele eller deler av ett eller flere SVO-er i norske havområder (Vedlegg 1).
8 - Resultater for Barentshavet
8.1 - Oversikt over særlig verdifulle områder i Barentshavet
Det er beskrevet sju særlig verdifulle områder, BH1–BH7, i Barentshavet (figur 8.1, tabell 8.1). Lokalisering, beskrivelser og miljøverdivurderingene for hvert av disse er beskrevet i kapitlene 8.3. til 8.9.
Det er gjennom vurderingene foretatt nye avgrensninger av eksisterende SVO-er, bl.a. gjennom utvidelser av Kandidatområder og noen forskyvninger mellom eksisterende SVO-er. I tillegg er et nytt område (BH7) identifisert. Avgrensningene omtales nærmere under hvert område.
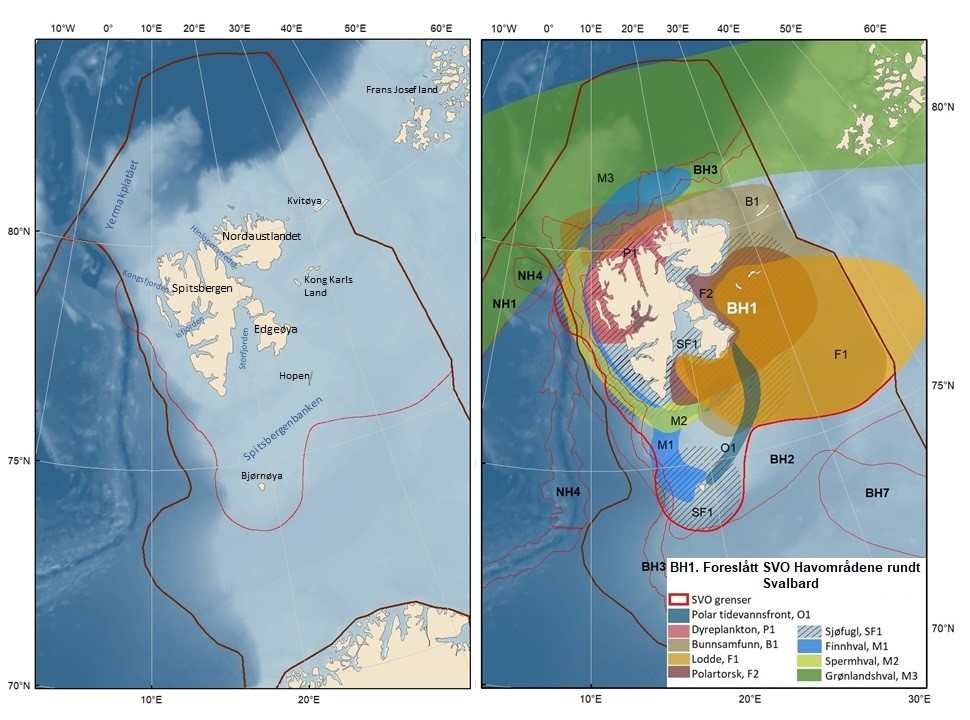
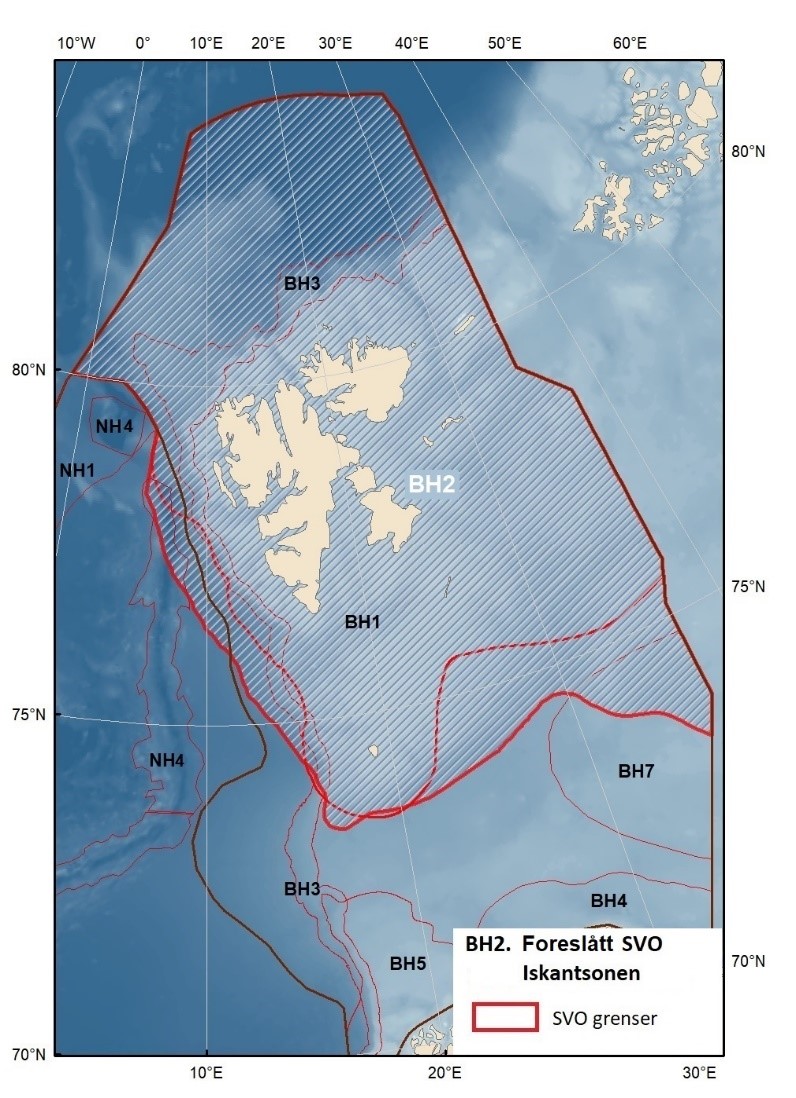
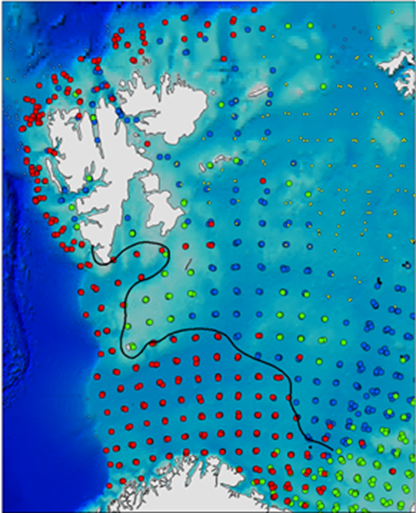
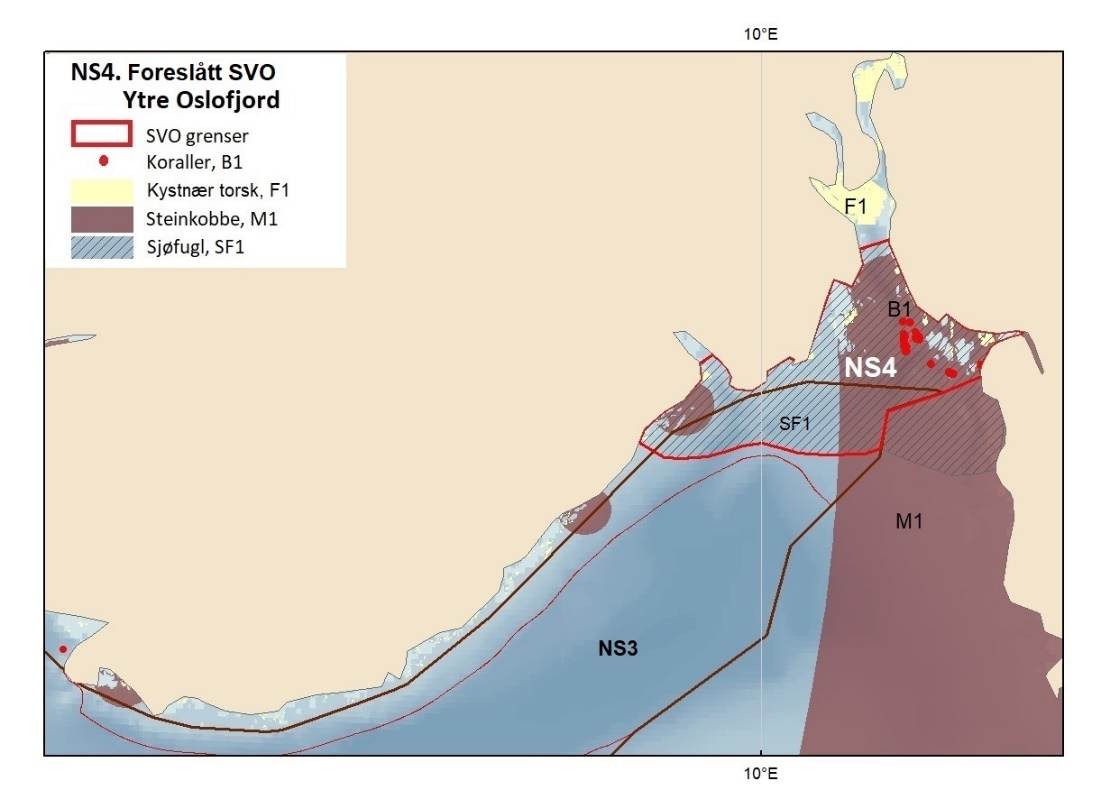
Figur 8.1. Forslag til særlig verdifulle og sårbare områder (SVO) i Barentshavet. To forslag til endrede SVO-er ligger i det sentrale og nordlige Barentshavet (BH1-Havområdene rundt Svalbard og BH2-Iskantsonen), ett foreslått endret SVO ligger langs kontinentalskråningen vest i Barentshavet (BH3-Eggakanten nord), tre foreslått endrete SVO-er ligger ved kysten av Nord-Norge (BH6-Kystsonen Lofoten, BH5-Tromsøflaket og BH4-Kystsonen Finnmark) og ett foreslått nytt SVO ligger sentralt i Barentshavet (BH7-Sentrale Barentshavet). Merk at BH1 og BH2 i stor grad overlapper. Grensene for forvaltningsplanområdet er vist med brune linjer.
Forslag SVO-kode Miljøverdi-rapport 2021
SVO-kode Meld. St. 2019-2020
SVO-navn Meld. St.2019-2020
Forslag SVO-navn Miljøverdi-rapport 2021
Forslag til endringer
BH1
BH3
Polar tidevannsfront
Havområdene rundt Svalbard
Inkludert i Havområdene rundt Svalbard
BH1
BH4
Havområdene rundt Svalbard
Havområdene rundt Svalbard
Utvidet med sommerbeite for lodde
BH2
BH6
Iskantsonen
Iskantsonen
Utvidet til maksimal isutbredelse
BH3
BH7
Eggakanten
Eggakanten nord
Starter i sør ved grensen til forvaltningsplanområdet for Norskehavet, ellers uendret
BH4
BH1
Kystsonen Finnmark
Kystsonen Finnmark
Utvidet nordover med Kandidatområdene, og et stykke inn i fjordene for å dekke beiteområder for sjøfugl, sel og hval
BH5
BH2
Tromsøflaket
Tromsøflaket
Utvidet sør til Senja, nord til Sørøya og litt inn i fjordene, inkl. Lopphavet
BH6
BH5
Lofoten til Tromsøflaket
Kystsonen Lofoten
Endret til å omfatte Vestfjorden og Røst til Senja. Utvidet vestover med Kandidatområdene
BH7
Det sentrale Barentshavet
Nytt område
Tabell 8.1. Samlet oversikt over koder for foreslåtte SVO-er i foreliggende rapport, sammenlignet med koder og navn på eksisterende SVO-er i Barentshavet (fra Meld. St. 20 (2019-2020)), navneforslag på de foreslåtte SVO-ene og kort beskrivelse av forslag til endringer i grensesetting.
Ismåke. Foto: Elvar H. Hallferdsson, HI
8.2 - Generell beskrivelse av økosystem Barentshavet
Utstrekning
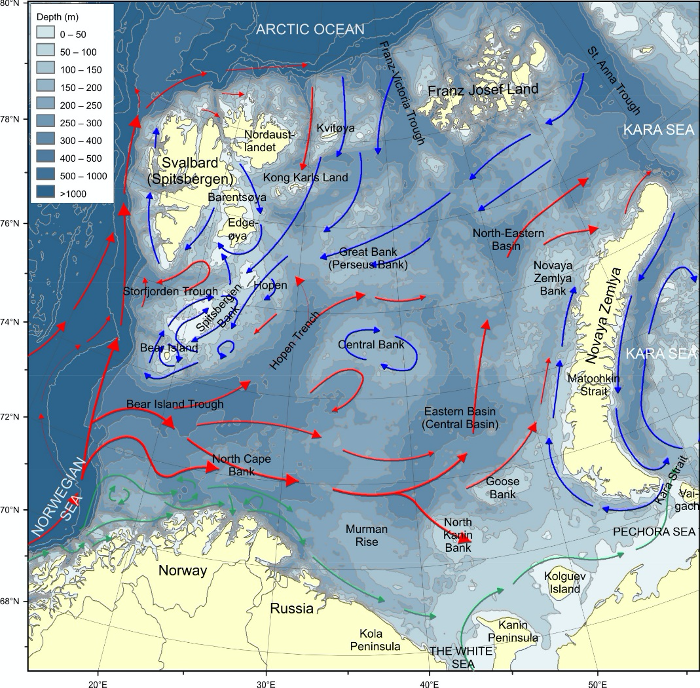
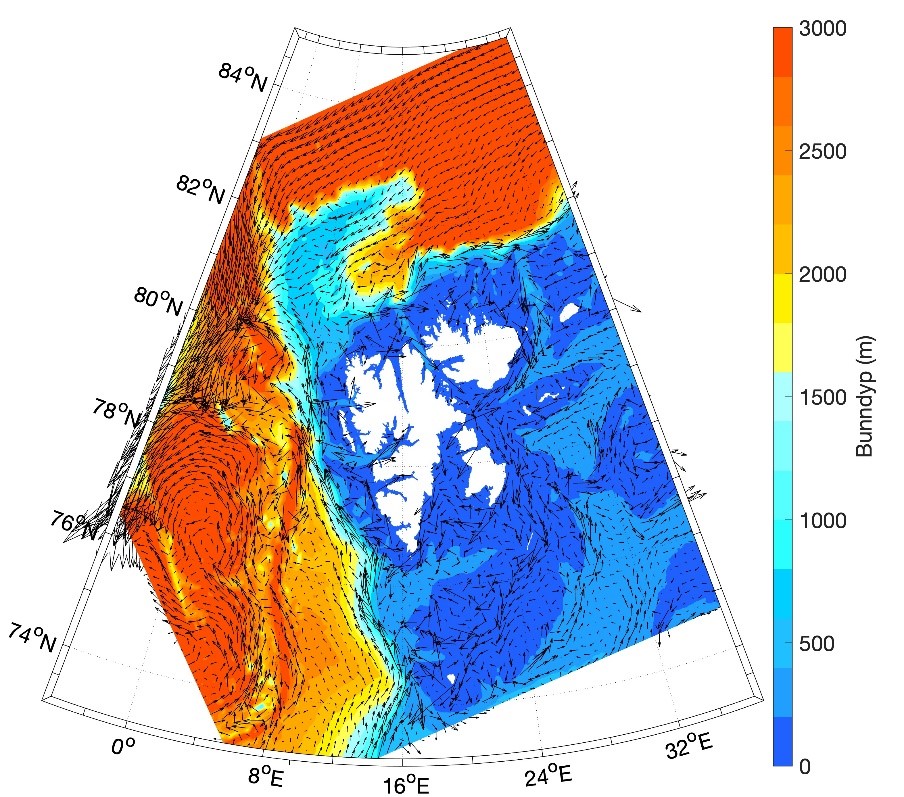
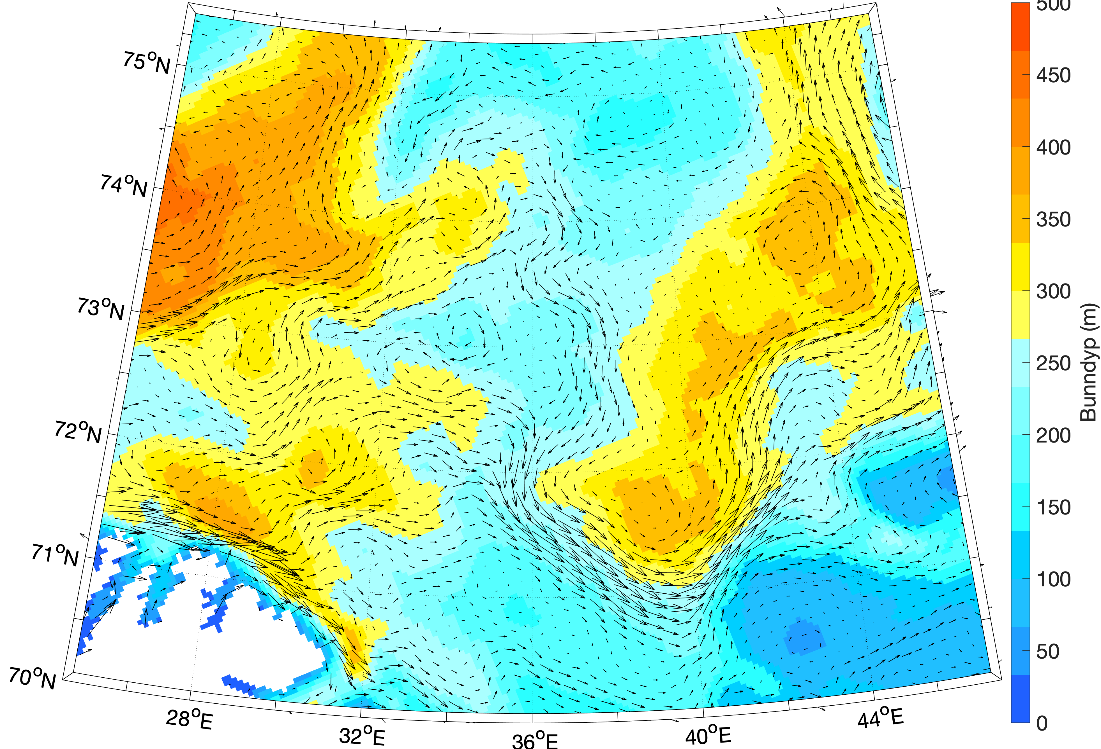
Barentshavet er et stort sokkelhav som ligger nordøst for Nord-Atlanteren. Det grenser til det dypere Norskehavet i vest og Polhavet i nord. I øst er grensen mot det grunne Karahavet og Novaja Semlja, og den norske og russiske kysten i sør. Dybdekonturen på 500 m langs kontinentalskråningen brukes til å avgrense Barentshavet mot vest (fra Norskehavet) og nord (fra Polhavet). Barentshavet dekker et område på omtrent 1,6 millioner km2 (Carmack mfl., 2006), har en gjennomsnittlig dybde på ca. 230 m, og en maksimal dybde på rundt 500 m ved den vestlige enden av Bjørnøyrenna (figur 8.2.1).
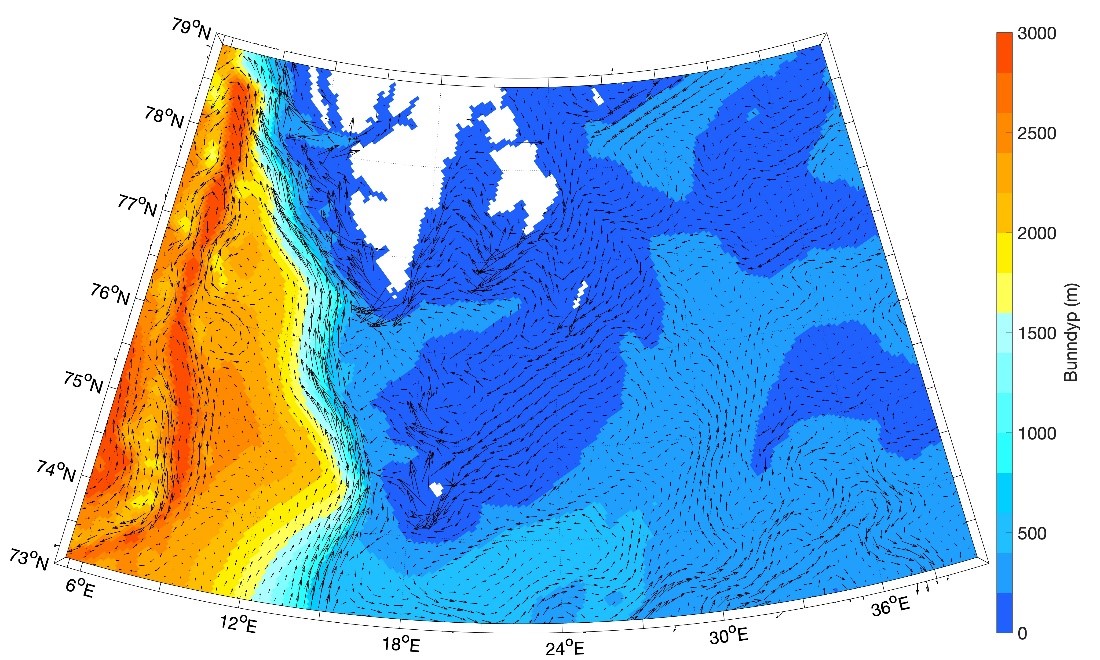
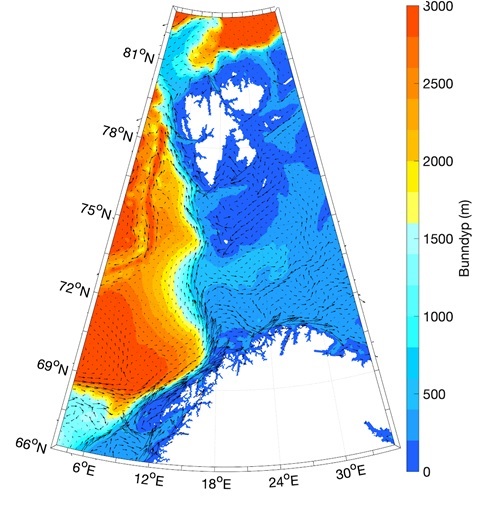
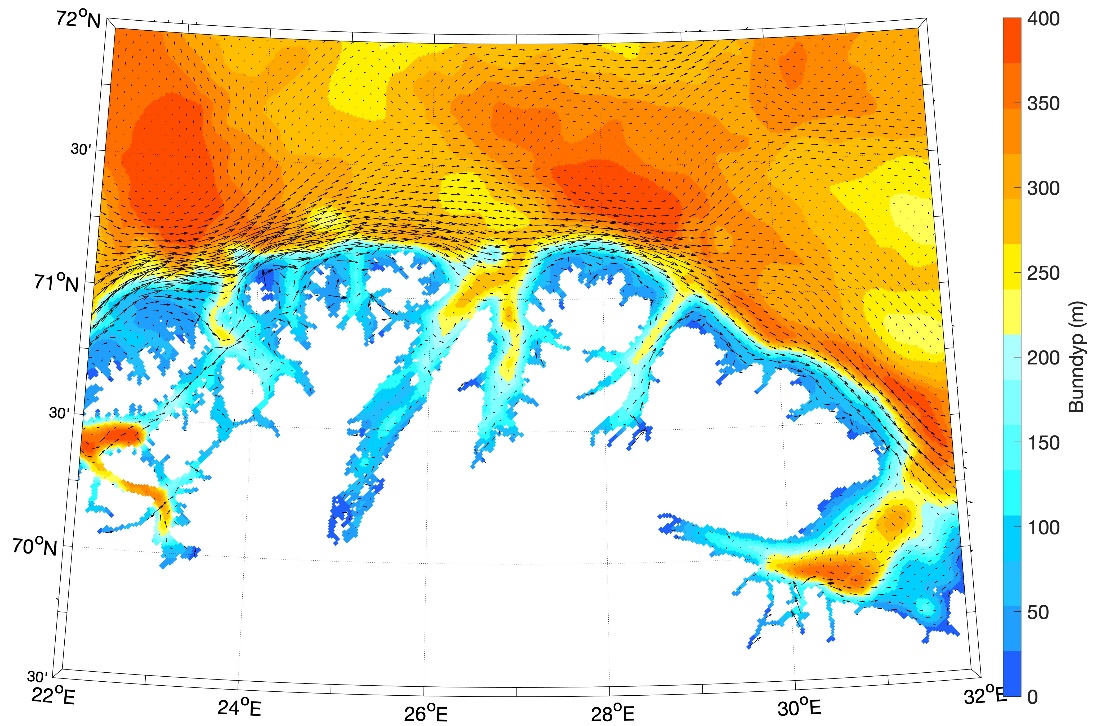
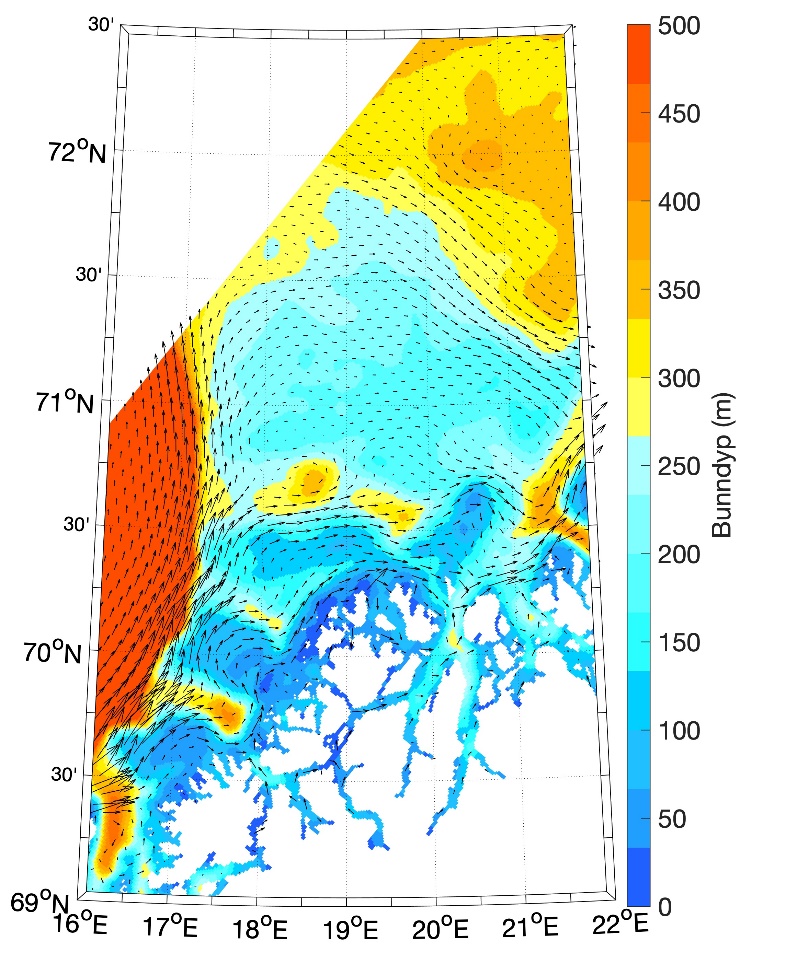
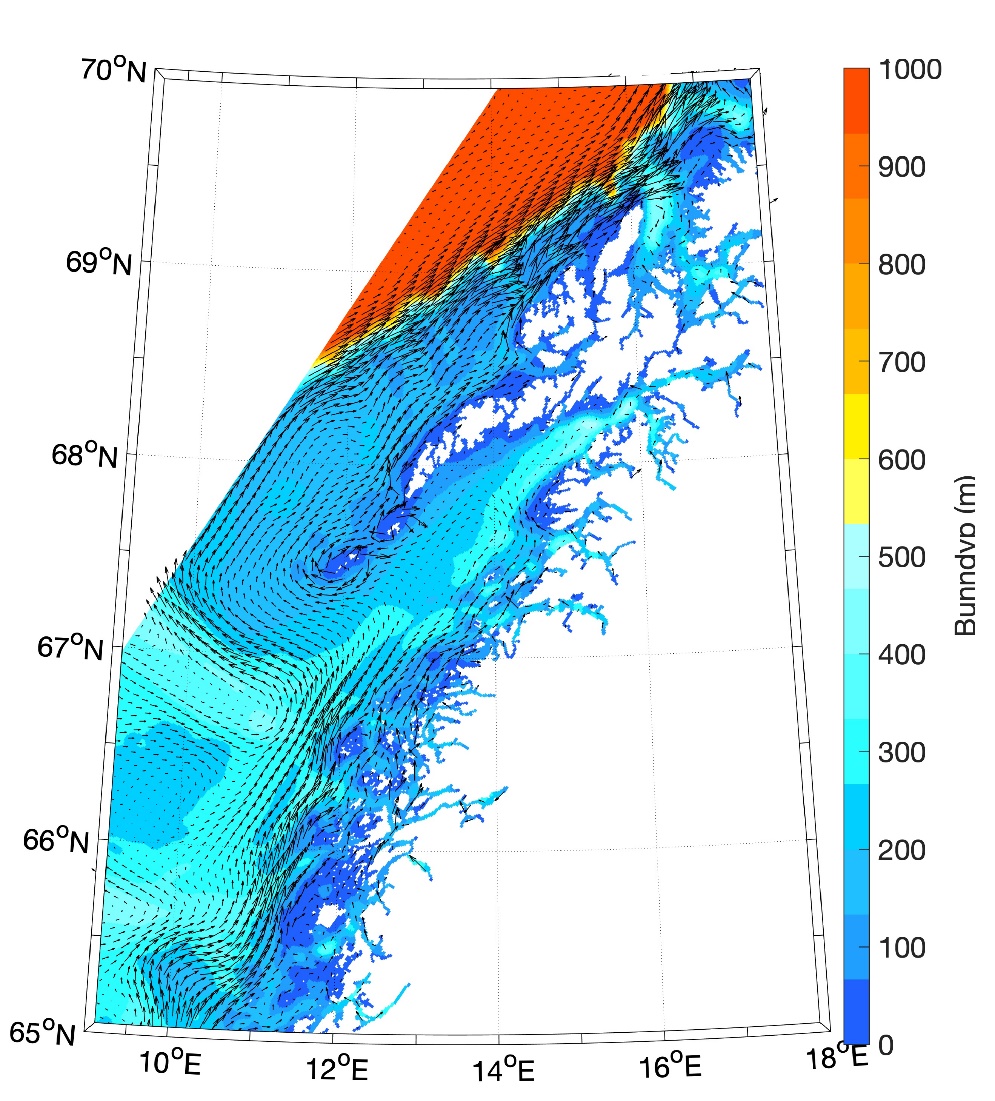
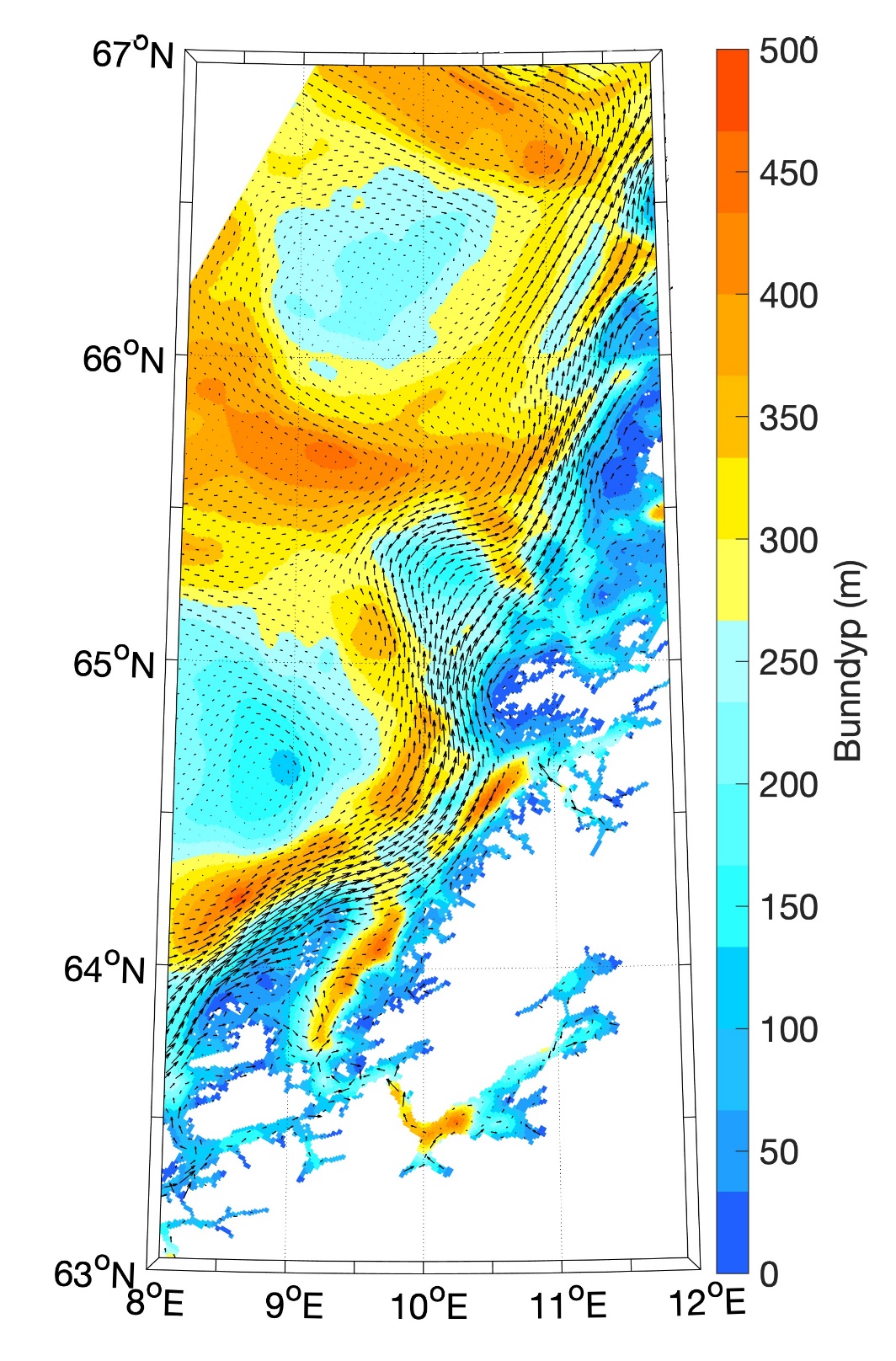
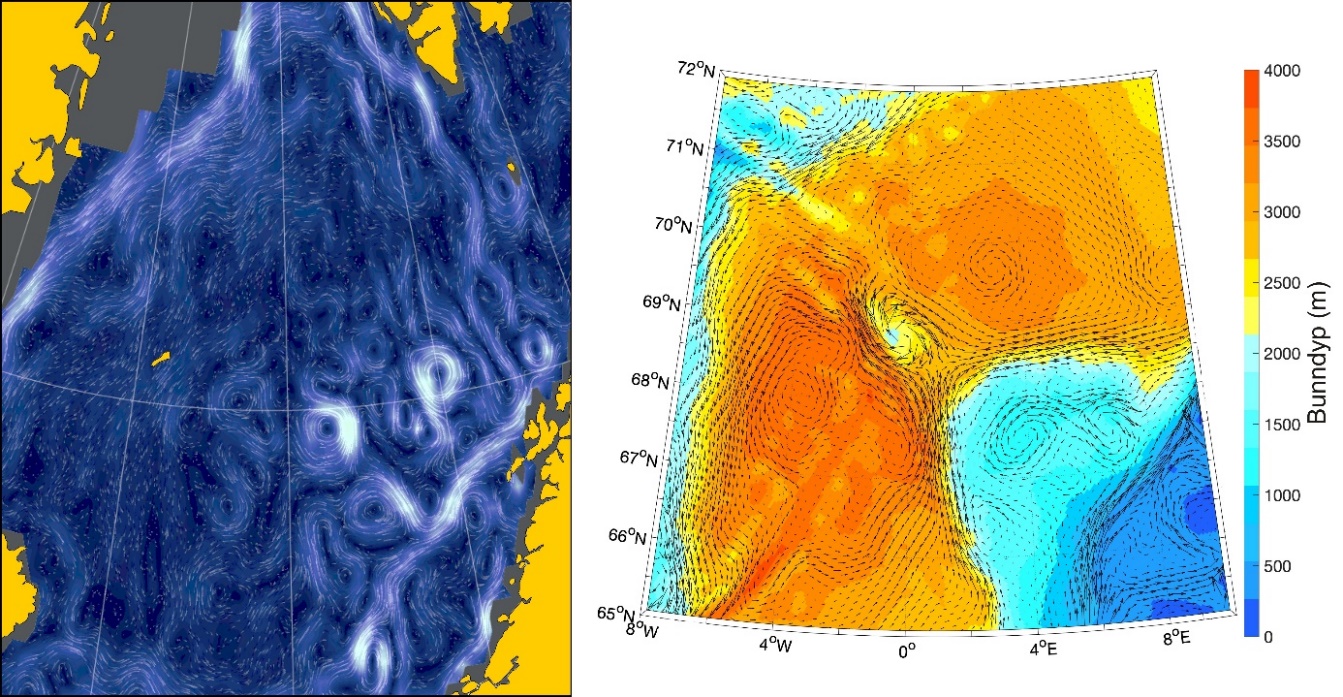
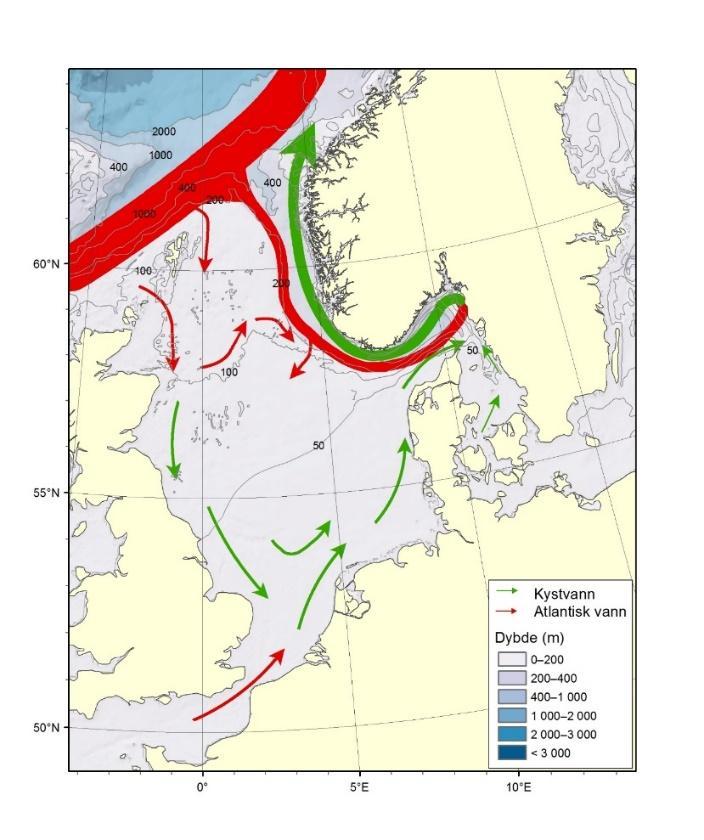
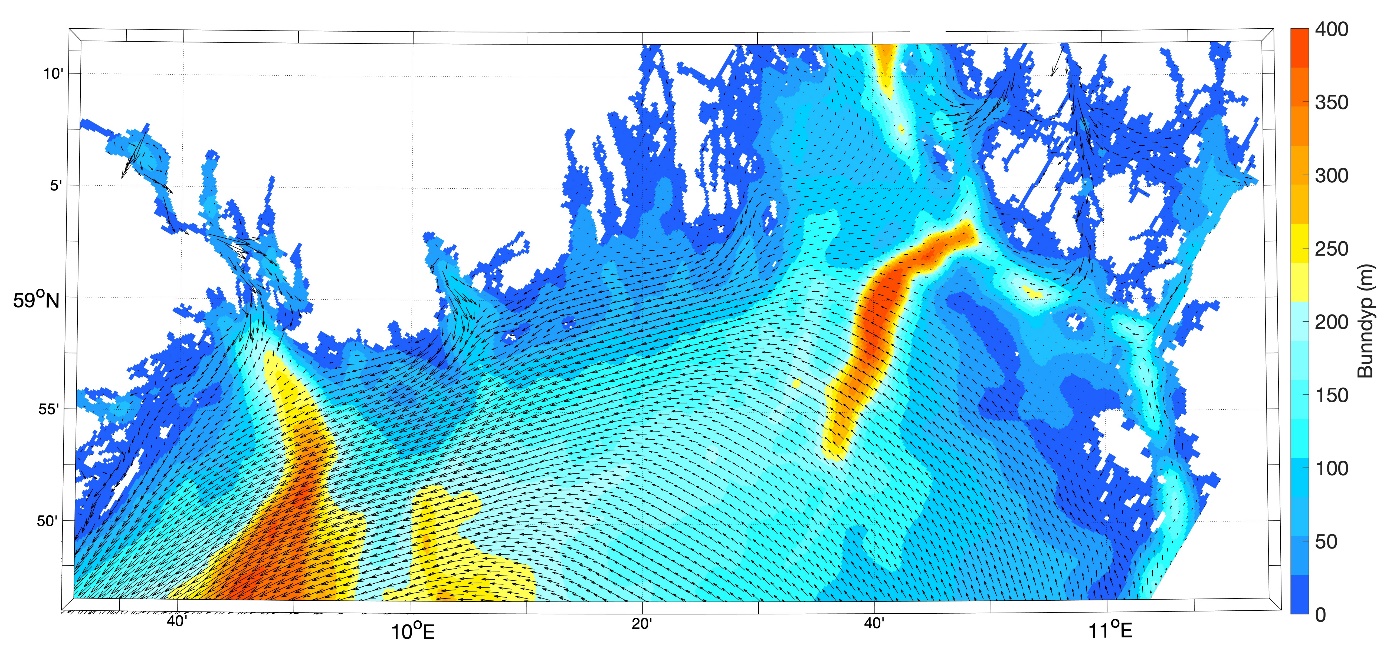
Figur 8.2.1. Kart over Barentshavet med bunntopografi og viktigste havstrømmer (atlanterhavsvann - rødt, arktisk vann - blått, kystvann - grønt).
Havstrømmer og vannmasser
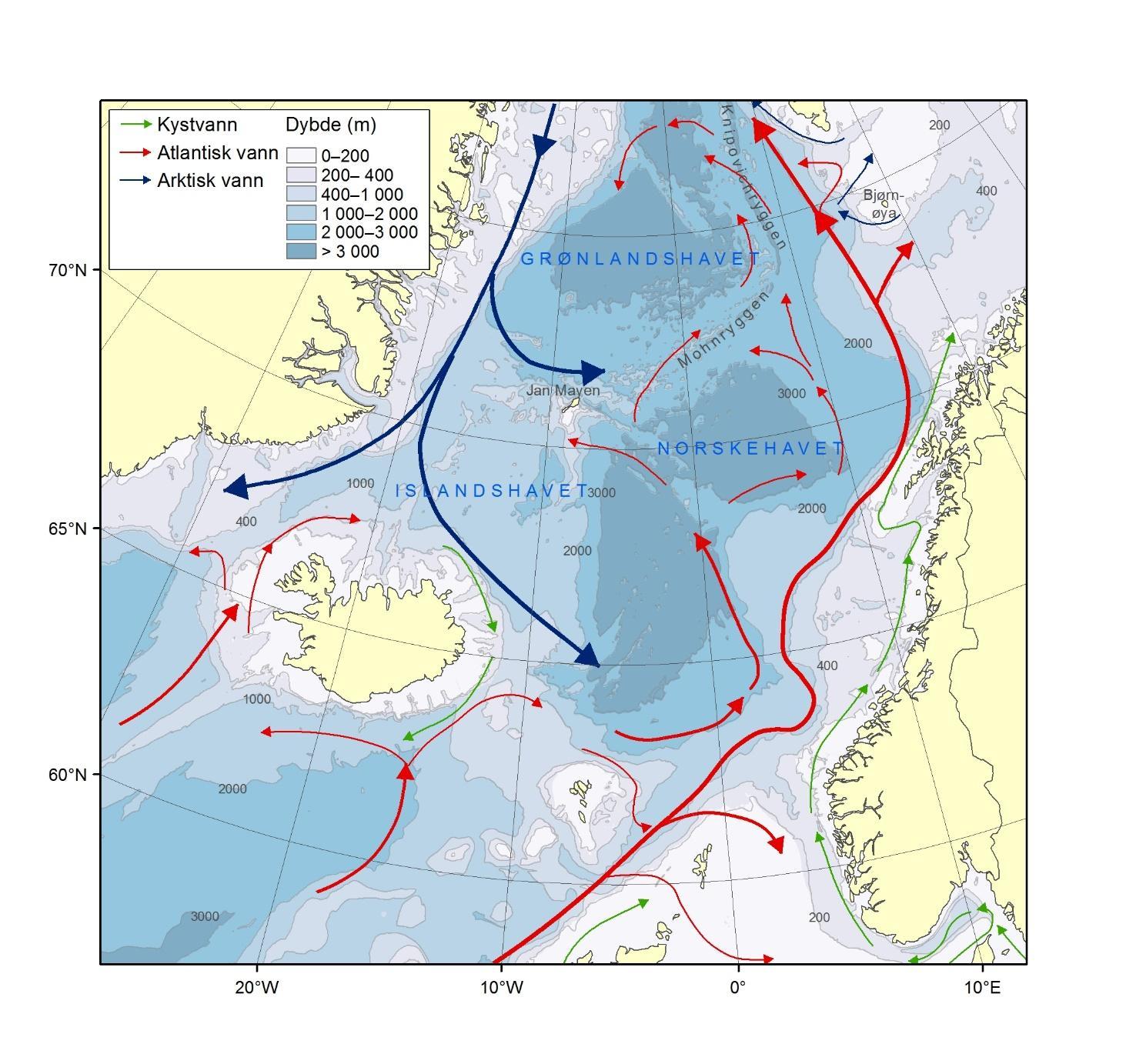
Topografien er preget av renner og bassenger, adskilt av grunne bankområder. De tre største bankene er Sentralbanken, Storbanken og Spitsbergenbanken. Varmt og salt atlanterhavsvann strømmer inn fra sørvest og fyller de sørlige og sentrale områdene, mens kaldt og ferskt arktisk vann dominerer i det nordlige Barentshavet (Loeng 1991). Storskala atmosfæriske trykksystemer påvirker volumfluksen av atlanterhavsvann, som igjen påvirker oseanografiske forhold både i Barentshavet og i Polhavet (Ingvaldsen mfl. 2004). Der atlanterhavsvannet møter kaldere arktiske vannmasser skapes en oseanografisk front (polarfronten). Nord for polarfronten er havet lagdelt med arktisk vann over atlanterhavsvann, og det arktiske vannet beskytter isdekket fra varmen i dypet (Loeng 1991, Lind mfl. 2018). Variasjoner i næringssaltkonsentrasjoner og karbonkjemi observeres både i nordlig og sørlig del av Barentshavet. I nordlig del skyldes dette variasjoner i isdekke og det kalde og ferskere arktiske vannet, mens i sør skyldes det variasjoner i det varmere og saltere atlanterhavsvannet. Barentshavet er et område hvor det tas opp mer atmosfærisk CO2 i havet enn det slippes ut, og dette er hovedsakelig drevet av biologisk CO2-opptak (Fransson mfl. 2001, Lauvset mfl. 2013). Det er ikke store havdyp i dette området, så mesopelagisk fauna er ikke vurdert.
Klimaendringer
I Barentshavet har havtemperaturene økt jevnt siden tidlig på 1980-tallet fram til 2016. I samme periode har Barentshavet hatt minkende isdekke (Årthun mfl. 2012, Onarheim mfl. 2018) og store økologiske endringer, særlig i de nordligste delene, hvor lagdelingen har blitt svekket og det har vært en svært rask oppvarming i de øvre 100 m (Lind mfl. 2018). Minket isdekke, oppvarming, økt ferskvann i overflaten og endringer i primærproduksjon gir endringer i karbonsyklus og hav-atmosfære CO2-utveksling (Chierici og Fransson 2018). På grunn av oppvarming og redusert fiskepress har særlig torskebestanden vokst og utvidet sitt utbredelsesområde i Barentshavet (Kjesbu mfl. 2014). Arktiske arter har samtidig fått et mer begrenset leveområde (Fossheim mfl. 2015). Etter 2016 og til 2020 har imidlertid temperaturen gått litt nedover, og samtidig har isdekket økt noe igjen, men temperaturen ligger fortsatt over langtidsmiddelet (González-Pola mfl. 2019, ICES 2020 a-under forberedelse).
Kort om økosystem og ressurser
Generelt vil det være større produksjon og mer stabil, høy biomasse i områder med kompleks topografi. For eksempel gir eggakanten mulighet for oppstrømning av næringssalter fra dypvannet som gir grunnlag for økt biomasse av planteplankton over sokkelen (Naustvold mfl. 2020). Også frontsystemer vil kunne gi grunnlag for økt biomasse og produksjon, da blanding av ulike vannmasser kan føre til innblanding av næringssalter til overflaten. For områder der fronter og topografiske forhold sammenfaller, kan man få særlig stor produksjon og biomasse av planteplankton (Lien mfl. 2018, Vikebø mfl. 2019). I slike områder vil også planteplanktonproduksjon være mer stabilt høy, sammenlignet med åpne havområder. Inne ved kysten vil man også ha tilførsel av næringssalter fra land, sammen med ferskvann, som vil resultere i tidlig og stor planteplanktonproduksjon i perioder av året (eks våroppblomstringen). I mange av de foreslåtte SVO-ene vil kyststrømmen gå gjennom og bringe med seg planteplankton og næringssalter. Kystnære områder vil samtidig kunne bringe planteplankton ut i havområdene. Det fysiske og kjemiske miljøet legger til rette for planteplanktonproduksjon i iskantsonen, der issmelting fører til vertikal stabilitet, vinterkonsentrasjoner av næringssalter i overflaten og forbedrede lysforhold (von Quillfeldt mfl. 2018). Den konsentrerte og intense produksjonen i iskantsonen har stor betydning for sekundærproduksjon og bunnlevende organismer i disse områdene (Reigstad mfl. 2011). I tillegg er det primærproduksjonen i isen som starter tidligere enn i vannmassene, som bidrar til å forlenge den produktive sesongen i områder med havis (Hegseth 1998, Leu mfl. 2015).
Det er fire nøkkelarter i Barentshavet: den pelagiske stimfisken lodde (Gjøsæter 1998, Orlova mfl. 2005, 2011, Gjøsæter mfl. 2009), polartorsk som beiter på ny planktonproduksjon om våren og sommeren (Hop og Gjøsæter 2013), nordøstarktisk torsk (Hylen mfl. 2008, Kjesbu mfl. 2014, Fossheim mfl. 2015, ICES 2020 a) og ungsild (Dragesund mfl. 1980, Sætre mfl. 2002, Sundby mfl. 2013). Lodde og polartorsk er nøkkelarter i Barentshavets økosystem hele året. Sildelarvene som kommer drivende fra kystbankene lenger sør er viktig næringskilde for fisk og andre organismer som beiter på eggene i perioden etter gyting. Ungsilda beiter hovedsakelig på raudåte og den kan være en stor predator på loddelarver, noe som førte til kollaps i loddebestanden i 1982–83, 1989 og 1997–1999 (Krysov og Røttingen 2011). Ungsilda er selv en viktig matressurs for rovfisk som torsk, sei og annen bunnfisk i tillegg til hval. Store flokker av spekkhoggere følger silda på dens vandringer.
De rike forekomstene av krepsdyr, fisk og fiskelarver gir grunnlag for en rik sjøfuglfauna i Barentshavet (Anker-Nilssen mfl. 2015, Fauchald mfl. 2015). Sjøfuglsamfunnet er satt sammen av arktiske og subarktiske arter. Mange av bestandene er i kraftig nedgang som en følge av endret næringsgrunnlag, særlig i de sørlige områdene av Barentshavet (Anker-Nilssen mfl. 2020, Fauchald mfl. 2015). Vi ser også at arter som tidligere hadde en mer sørlig utbredelse nå i økende grad får en mer nordlig forflytning av hekkebestandene, for eksempel havsule, lunde og storjo (Fauchald mfl. 2015).
De rike forekomstene av krepsdyr, fisk og fiskelarver gir grunnlag for en rik sjøfuglfauna i Barentshavet (Anker-Nilssen mfl. 2015, Fauchald mfl. 2015). Sjøfuglsamfunnet er satt sammen av arktiske og subarktiske arter. Mange av bestandene er i kraftig nedgang som en følge av endret næringsgrunnlag, særlig i de sørlige områdene av Barentshavet (Anker-Nilssen mfl. 2020, Fauchald mfl. 2015). Vi ser også at arter som tidligere hadde en mer sørlig utbredelse nå i økende grad får en mer nordlig forflytning av hekkebestandene, for eksempel havsule, lunde og storjo (Fauchald mfl. 2015).
Også sjøpattedyr utnytter de rike forekomstene av blant annet dyreplankton og fisk i dette havområdet(Laidre mfl. 2008, Hamilton mfl. 2017, 2018 og 2019 a,b).
Plankton i forvaltningsområdet
Barentshavet er et produktivt område som gir gode levevilkår for noen av verdens største bunnfiskbestander som torsk og hyse samt pelagiske arter som lodde, for sjøfugl og for arktiske og andre sjøpattedyr (Hunt mfl. 2013). Det rike og varierte planktonsamfunnet i Barentshavet opprettholder disse øvre trofiske nivåene (ICES 2019). Viktige arter av kopepoder (Calanus finmarchicus og Calanus glacialis) og krill (Thysanoessa inermis) beiter i hovedsak på planteplankton (Dalsgaard mfl. 2003, Falk-Petersen mfl. 2007), og muliggjør en effektiv trofisk overføring av energi fra planteplankton til høyere trofiske nivåer i økosystemet.
Sammensetningen og biomassen av dyreplankton i Barentshavet og rundt Svalbard er påvirket av de atlantiske og arktiske havstrømmene. Atlanterhavsstrømmen og den norske kyststrømmen transporterer en høy biomasse av borealt/atlantisk dyreplankton, for de meste C. finmarchicus inn i Barentshavet og delvis via en forgrening av atlanterhavsstrømmen som fortsetter nordover langs vest og nord av Svalbard og inn i Polhavet. En slik stadig påfylling av plankton gjør at arter kan utvide sine leveområder, og medvirker dermed til å fremme regionalt biologisk mangfold og produksjon (Dalpadado mfl. 2012). Sørgående arktiske vannmasser med typisk arktiske arter som C. glacialis dominerer i det nordlige Barentshavet øst for Svalbard. Calanus-artene er de viktigste mht. variasjon i biomassen av mesozooplankton i Barentshavet, og utgjør omtrent 80 prosent av den totale biomassen (Aarflot mfl. 2018).
Planteplankton som element har en økosystemfunksjon, som basis for marine næringskjeder. Planteplankton er til stede i alle våre havområder og vil gjennomgå markante endringer gjennom året både når det gjelder biomasse og sammensetning. Planteplankton vil også vise stor variasjon mellom årene. Artene vil være til stede over store områder (større enn SVO), har høy vekstrate i perioder og vil til enhver tid utnytte de fysiske og kjemiske forholdene. Arter som er knyttet til is er unike i forhold til åpent isfritt hav. På grunn av planteplanktonets store variasjon i tid og rom og deres frittflytende pelagiske levesett, er det ofte vanskelig å benytte i en utredning omkring miljøverdier inne i enkeltområder.
Atlanterhavsvannet transporterer ikke bare varme og plankton, men utgjør også et storstilt transportsystem for fiskelarver for de aller fleste av våre kommersielle fiskebestander: sild på kystbanker utenfor Vest- og Midt-Norge, torsk i hovedsak i Lofoten og Vesterålen, og lodde utenfor Troms, Finnmark og Kola. Larvene driver med strømmene fra gytefeltene til gunstige oppvekstområder for yngelen i Barentshavet.
8.2.1 - Pågående og framtidige endringer i Barentshavet
De overgripende klimaendringene og havforsuring er forklart i kapittel 5. Her trekkes det fram hvordan økosystem og miljøverdiene i Barentshavet blir og kan bli direkte eller indirekte påvirket av temperaturendringer og havforsuring.
Temperaturfallet siden 2016 er en forbigående nedgang, og klimamodeller viser at oppvarmingen og nedgangen i isdekke i norske havområder vil fortsette (IPCC 2019, Årthun mfl. 2019), og dessuten at vi må vente økt havforsuring.
Økning i temperatur og reduksjoner i isdekke er betydelig og har allerede ført til en nordlig ekspansjon av boreale arter i norske havområder og reduksjon av arktiske arter i Barentshavet. Dette er ventet å fortsette i fremtiden (Fossheim mfl. 2015). Barentshavet er den delen av Arktis som opplever de raskeste klimaendringene og har fått redusert tilførsel av is og ferskvann og svekket lagdeling i den nordlige arktiske delen (Lind mfl. 2018). Endringer i temperaturen vil også kunne endre konkurransen mellom ulike arter, og arter med høyere optimal temperatur vil kunne bli mer tallrike.
Den vedvarende forhøyete temperaturen skyldes i stor grad mindre varmetap fra hav til atmosfære (Skagseth mfl. 2020). Påvirkninger på økosystemene, fra organismenivå, biologiske prosesser og i økologisk sammenheng, har fokus på de seks miljøverdigruppene isbiota, plankton, bunnsamfunn, fisk, sjøpattedyr og sjøfugl.
Fremtidige endringer i temperatur, isdekke, nedbør, ferskvannsavrenning og vind vil kunne føre til endringer i primær- og sekundærproduksjon (plante- og dyreplankton) ved at næringssalttilgang og sesongdynamikk vil kunne endres (von Quillfeldt mfl. 2018).
Større tilførsel av ferskvann kan gi sterkere lagdeling og dermed mindre innblanding av næringsrikt vann fra underliggende vannmasser. Dette kan igjen føre til reduksjon i næringssalter i overflaten og dermed lavere i planteplanktonproduksjon og biomasse, som vil føre til redusert næring og energi videre i næringsnettverket.
Endringer i lagdelingen vil kunne ha konsekvenser for artssammensetningen. Sterk lagdeling pga. økt tilførsel av ferskvann kan være et konkurransefortrinn for små flagellater og fureflagellater (Li mfl. 2009), mens økt grad av miksing vil stimulere kiselalger. Dyp miksing favoriserer flagellater (Sakshaug 2004).
Sterkere vind og en økning i antall og/eller intensitet i stormer kan ha en positiv effekt på planktonproduksjonen gjennom svakere lagdeling og dermed økt innblanding av næringssalter.
Økt ferskvannstilførsel vil bidra til økt havforsuring og begrense kalkdannelse og gi økt kalkoppløsing på skall og skjelett i øvre vannmasser. Dessuten kan miksing av vannmasser bli så kraftig at den fører algene ned til dyp der fotosyntese ikke lenger forekommer.
Isbiota
Isbiota og andre arter med hele livssyklusen i havis, står i fare for å forsvinne eller bli sterkt redusert selv om det finnes mekanismer som bidrar til å holde isfauna i vannmassene (Berge mfl. 2012, Kunisch mfl. 2020). Se for øvrig foreslått SVO Havområder rundt Svalbard (Kap. 8.3.6) og foreslått SVO Iskantsonen (Kap. 8.4.6).
Plankton
Reduksjon av is i Barentshavet vil føre til høyere primærproduksjon fordi mer lys når ned i havet, men vil også føre til endret artssammensetning av planteplankton og dyreplankton (Dalpadado mfl. 2020). I likhet med andre havområder har klimaendringer ført til at flere arter forflytter seg i takt med et varmere hav. Stadig mer isfrie forhold vil kunne fremme «borealisering» av planktonsamfunnet i Barentshavet, og en del endringer ser vi allerede. Dette åpner for nye beiteområder for boreale arter, som tidligere har unngått rene polare leveområder. For organismer som er avhengige av produksjon i isdekte områder, eller lever i kaldere vannmasser, kan derimot borealiseringen av planktonsamfunnet ha negative konsekvenser. Vi ser at det er nedgang av typisk kaldtlevende arter som kopepoden Calanus glacialis og amfipoden Themisto libellula i varme år i Barentshavet (Aarflot mfl. 2018, Stige mfl. 2019). På den annen side er det rapportert en dobling av krill-biomasse i Barentshavet fra 1990- til 2010-tallet (Eriksen mfl. 2017). Isbaserte SVO-er i Norskehavet, som SVO Havis Framstredet (Kap. 9.3) og SVO Vesterisen (Kap. 9.4) har samme framtidsutsikter som de foreslått endrete SVO-ene Havområdet rundt Svalbard og Iskantsonen.
Fisk
Mens gytefelter vil forandre seg lite, kan vanntemperatur, transport og matforsyning bli påvirket av klimaendringer, enten av større eller hyppigere uoverensstemmelse mellom vårblomstring og gytetid eller av en endret atlanterhavsstrøm og derfor raskere eller tregere transport (Solemdal mfl. 1997, Hylen mfl. 2008, Dragesund mfl. 2008, Sundby og Nakken 2008, Sundby mfl. 2013).
Lodde (Mallotus villosus) og polartorsk (Boreogadus saida) beiter på ny planktonproduksjon om våren og sommeren (Gjøsæter 1998, Orlova mfl. 2005, 2011 Gjøsæter mfl. 2009, Hop og Gjøsæter 2013). Nordøstarktisk torsk (torsk, Gadus morhua) beiter både på plankton, når den er liten og reker og fisk, særlig lodde, når den blir større (Hylen mfl. 2008, Kjesbu mfl. 2014, Fossheim mfl. 2015, ICES 2020 a,b, Townhill mfl. 2020).
Polartorsk, som lever i arktiske vannmasser og er knyttet til isen, er avhengig av arktisk dyreplankton som en viktig matkilde. Nedgang i arktisk dyreplankton og ismengde vil mest sannsynlig påvirke fisk som polartorsk (Huserbråten mfl. 2019, Dalpadado mfl. 2020), og høyere trofiske nivåer, samt påvirke biodiversiteten i det arktiske økosystemet. Ytterligere oppvarming kan føre til at den allerede reduserte bestanden av polartorsk i Barentshavet vil fortsette å minke (Eriksen mfl. 2015, 2019, 2020, Huserbråten mfl. 2019, Gjøsæter mfl. 2020, Kortsch mfl. 2015). Klimatiske endringer innen fiskesamfunn er ytterligere diskutert, i forhold til ekspansjonsmuligheter og tilgjengelig habitat (Hollowed mfl. 2013a, b, Fossheim mfl. 2015, Haug mfl. 2017, Hollowed mfl. 2018). I et varmere hav med mindre is i Arktis, utvider flere arter sitt leveområde nordover (Fossheim mfl. 2015, Eriksen mfl. 2017). Forskyvning av utbredelsen til ulike arter vil avhenge av at fisken finner nye gyteplasser lenger nord og nye løsninger for geografisk lukking av sin livssyklus. Lodda forskyver seg øst, vest og nordover i Barentshavet i forhold til vekslinger mellom kalde og varme år (Gjøsæter 1998, Ingvaldsen og Gjøsæter 2013). Det er imidlertid sannsynlig at lodda ikke har særlig mer rekkevidde å hente på sin beitevandring nordover i Barentshavet ved dagens plassering av gytefeltene utenfor Troms, Finnmark og Kola (Loeng 2006). Torsken har imidlertid stor vandringsrekkevidde, slik at den fortsatt kan ha sine gytefelter langs kysten av Nord-Norge selv om den går lenger nord for å beite.
Bunnsamfunn
Det er vist en betydelig forskyvning av boreale bunndyrarter og endring i bunndyrsamfunn i den arktiske delen av Barentshavet bare de siste 20 årene, med en påfølgende nedgang i biomasse av epibenthos etter 2014 (Zakharov og Jørgensen 2017, Weslawski mfl. 2018, Zakharov mfl. 2020). Endringer i biodiversitet, særlig økning av boreale arter og tilbaketrekking av arktiske arter er registrert i Svalbard-området (Jørgensen mfl. 2019). Dette er delvis relatert til økning i makroalger (Beuchel mfl. 2006, Weslawski mfl. 2010). Det er i mindre grad forstått hvordan dette kan påvirke det arktiske økosystemet.
Relaterte endringer i næringsnettstrukturen er tenkelig, for eksempel påvirket av 1) økte forekomster (Kortsch mfl. 2012, Al-Hababeh mfl. 2020) og tilførsel av karbon fra makroalger (Renaud mfl. 2015) og 2) påvirkning av isdekke på næringsnettets egenskaper (Kortsch mfl. 2018).
Sjøfugl
Endringer i planktonsamfunnene vil gjenspeiles i sammensetningen av dietten og påvirke den generelle økologien for eksempel til spesialiserte sjøfugl. Alkekongen (Alle alle) behøver tilgang til store energirike planktonorganismer (C. glacialis) for bedre overlevelse av ungene (Weslawski mfl.1999, Pedersen og Falk 2001).
Sjøpattedyr
Klimaendringer har generelt negativ påvirkning på beiteområdene og yngleplassene til en rekke arktiske sjøpattedyr. Dette omtales kort i beskrivelsen for hver art under miljøbeskrivelsene i de aktuelle SVO-ene. De som i særlig grad har vist seg følsomme for dette er beskrevet her.
På samme måte som for andre arktiske hvalarter er klimaendringene en trussel for hvithvalene (på Svalbard. Spekkhoggere (Orcinus orca) er en trussel for hvithval (Delphinapterus leucas) i områder med lite sjøis. I tillegg blir områdene foran isbreer hvor de hovedsakelig jakter, mindre og mindre ettersom isbreene smelter og trekker seg tilbake. Hvithvalen viser imidlertid en del plastisitet her og tilbringer nå mer tid ute i fjordene vekk fra brefronter på vestsiden av Svalbard og jakter da antakelig på atlantiske fiskearter (Hamilton mfl. 2019 a).
Narhval (Monodon monoceros) er vurdert å være en av de arktiske marine pattedyrartene som er mest sensitive i forhold til klimaendringer på grunn av deres spesialiserte dyptdykkende furasjeringsvaner, isavhengighet, begrensede utbredelsesområde og sensitivitet i forhold til undervannsstøy (Laidre mfl. 2008).
Etter de drastiske endringene i isforholdene som startet i 2006, har atferden til ringselene (Pusa hispida) på Svalbard endret seg en god del. Dette gjelder både for de som vandrer ut og de som forblir kystnært. De bruker mye mer tid til dykking og hviler mindre og alt tyder på de jobber hardere for å finne seg mat (Hamilton mfl. 2017, 2018 og 2019 a,b).
8.2.2 - Løpende kartleggings- og overvåkingsaktivitet
Et utvalg av løpende kartleggingsprogrammer og faste overvåkingsprogrammer, tokt og oseanografiske snitt utføres av Havforskningsinstituttet, alene og i samarbeid med andre nasjoner og forskningspartnere. Norsk Polarinstitutts overvåkningsaktiviteter bidrar med kunnskap til økosystemovervåking blant annet til MOSJ (Miljøovervåking Svalbard og Jan Mayen), helhetlig nasjonal havforvaltning, miljøsamarbeidet med Russland, AMAPs (Arctic Monitoring and Assessment Programmes) og CBMP (Circumpolar Biodiversity Monitoring Program). Samlet henter begge institutt inn data for følgende komponenter av de fysiske, kjemiske og biologiske systemene: oseanografi (vanntemperatur, saltholdighet, strømforhold og vannkjemi), havnivå, havistykkelse og -utbredelse, isbiota, planteplankton og dyreplankton, fisk, bunnsamfunn, marine pattedyr (observasjonsprogram og akustisk overvåkning av hvaler og støy (AURAL), sjøfugl (inkludert data som er en del av SEAPOP) og isbjørn. Dataene benyttes også internasjonalt i rapportering til OSPAR og ICES-gruppene WGIBAR og WGICA.
Kysttokt nord for 62 breddegrad, Barentshavet og Norskehavet
BH4-BH6
X
Årlig
Skreigytetoktet i Lofoten og Vesterålen
BH6
X
Årlig
Sildelarvetokt langs kysten
BH4-BH6
X
Årlig
Egga nord
BH3
X
Hvert 2. år
Bestandsundersøkelse rognkjeks
BH4-BH6
X
Årlig
Hvaltelling
BH1-BH4
X
Årlig, rullerende
Økologisk overvåking vågehval og grønlandssel
BH1-BH7
X
Hvert 2. år
Oseanografiske snitt
Gimsøy, nordvest (forlenget)
BH3, BH6
X
4x årlig
Fugløya-Bjørnøya
BH1-BH2, BH5
X
6x årlig
Bjørnøya-Sørkapp
BH1-BH2
X
4x årlig
Vardø nord; forlenget
BH1-BH2, BH4, BH7
X
Årlig
Sørkapp
BH1
X
Årlig
Hinlopen
BH1-BH2
X
Årlig
Faste stasjoner
LoVe bunnobservatorier
BH6
X
X
X
X
X
X
Kontinuerlig
Norsk Polarinstitutt
Oseanografisk. inkl. havisovervåking
BH1-BH2
Regulært
Kongsfjord-overvåkningen
BH1
X
X
X
X
Regulært
Hvalross
BH1-BH2
X
Hvert 5. år
Isbjørn
BH1-BH2
X
Årlig
Artssammensetning av dyreplankton i Kongsfjorden
BH1
X
Årlig
Artssammensetning av planteplankton i Kongsfjorden
BH1
X
Regulært 10-12x årlig
Tabell 8.2. Tabellen viser et utvalg av løpende kartleggings- og overvåkingsprogram, tokt og oseanografiske snitt, der foreslåtte SVO-er i Barentshavet helt eller delvis inngår i kartleggingen/overvåkingen. Foreslåtte SVO-er og miljøverdier som kartlegges/overvåkes er oppført i tabellen.
FF Helmer Hanssen. Foto: G.I. van der Meeren, HI
8.3 - Forslag til endret SVO Havområdene rundt Svalbard (BH1)
8.3.1 - Sammendrag
Områdebeskrivelse og strukturer
Forslag til endret SVO Havområdene rundt Svalbard (BH1) består av en rekke store og små øyer, fra Bjørnøya i sør til Rossøya i nord. Området omfatter både åpent hav, kyst og fjorder og har følgelig en kompleks og varierende bunntopografi, strømforhold og hydrografi, inkludert varierende isforhold gjennom året og mellom år. Havområdene rundt Svalbard har både arktiske og atlantiske vannmasser. Atlanterhavsvannet dominerer i områdene vest og nord av Svalbard, mens kaldere og ferskere arktisk vann dominerer øst for Svalbard og i noen fjorder. Det arktiske vannet strømmer sørover på østsiden av Svalbard og nordover som en kyststrøm også vest av Spitsbergenkysten innenfor atlanterhavsstrømmen. Ferske og kalde vannmasser okkuperer vanligvis øvre vannlag inne i fjordene, mens atlanterhavsvann strømmer inn i dypet. Vestspitsbergenstrømmen holder det meste av havområdene vest og nord av Svalbard isfrie hele året, men disse områdene påvirkes også av tilførsel av havis utenfra og fra fjordene. Både deler av havområdene rundt Svalbard og enkelte av fjordene kan ha havis i deler av året. Havisens beskaffenhet og betydning beskrives i større detalj for foreslått SVO Iskantsonen.
Den polare tidevannsfronten, som i Meld. St. 20 (2019-2020) er oppgitt som eget SVO, er foreslått lagt inn under dette SVO-et. Den ligger på den grunne Spitsbergenbanken, som er dominert av relativt sterk strøm langs kanten og sterke tidevannsstrømmer. De grunneste delene av Spitsbergenbanken er stort sett vertikalt gjennomblandet på grunn av tidevann og vind. Dette fører til at det i sommerhalvåret settes opp en såkalt «aktiv» tidevannsfront med sterk vertikalblanding rundt banken på 50-100 m dyp.
Miljøverdier
Havområdene rundt Svalbard overlapper med store deler av foreslått SVO Iskantsonen. Begge SVO-ene deler derfor miljøverdier. Produksjonsforhold, forekomst av arter, sårbarhet for ulike typer av påvirkning og hvordan dette varierer gjennom året og mellom år har betydning for i hvilken grad iskantsonen er verdifull og sårbar. Imidlertid vil hele iskantsonen alltid være viktig for flere arter og/eller biologiske prosesser uavhengig av årstid.
Havområdene rundt Svalbard er et forslag til sammensatt SVO med mange miljøverdier som opptrer lokalt og sesongmessig i deler av området. Svalbard har et rikt fugle- og dyreliv som i hovedsak er konsentrert langs kysten og knyttet til det marine miljøet og drivisen. Det meste av dyrelivet og deler av plantelivet på Svalbard er direkte eller indirekte avhengig av næring fra havet.
Området har flere unike biotoper som er sammensatt av alle miljøverdiene. Innenfor området finnes viktige produksjonsområder for plante- og dyreplankton, sentrale gytefelt og det er gode beiteområder for pelagisk beitende sjøfugl som hekker på Svalbard samt beite- og reproduksjonsområder for marine pattedyr med tilhold i Svalbard-området gjennom hele året.
Isbiota forekommer både i havis og fjordis i dette området. Isbiota utgjør en rekke ulike samfunnstyper og består av et høyt antall arter fra en rekke ulike artsgrupper og har en unik egenverdi i seg selv både mht. samfunnstyper og artssammensetning, samt betydning for produksjon som kanaliseres videre i næringsnettet. I tillegg til fjorder, kan særlig områder påvirket av atlanterhavsvann ha relativt høy dyreplanktonbiomasse, sammenlignet med mer åpne havområder. For fisk finnes viktige gyteområder for polartorsk på østsiden, oppvekstområder for polartorsk, uer og torsk, og beiteområder hovedsakelig for nøkkelarten lodde (særlig på østsiden av Svalbard), men også for den arktiske polartorsken og de boreo-arktiske toppredatorene torsk og hyse. Det er flere polynjaer (områder med åpent vann, omgitt av is) i området, blant annet i Storfjorden. Disse har forhøyet produksjon og er derfor områder hvor sjøfugl og sjøpattedyr samles, også vinterstid. Brefronter rundt hele Svalbard er viktige beiteområder, blant annet for hvithval og ringsel. Arter som isbjørn, hvalross, storkobbe og ringsel er nært knyttet til isen i fjordene og drivisen som leveområde. På vestkysten av Spitsbergen finnes også verdens nordligste bestand av steinkobbe. Havområdene rundt Svalbard er viktige for både arktiske endemiske og sommergjestende marine pattedyr og området er et av de mest artsrike i Arktis for denne dyregruppen. Alle de nordøstatlantiske arktiske endemiske marine pattedyrene finnes her og mange av disse er blitt sterkt overbeskattet. Disse bestandene som er i ferd med å ta seg opp igjen, er av spesiell bekymring siden de er is-assosierte og forventet å bli negativt rammet av global oppvarming. Flere millioner sjøfugl hekker på Svalbard, særlig på øygruppens sørlige og vestlige deler, som er eksponert mot de produktive delene av Barentshavet. Man antar at ca. 45 prosent av totalbestanden av polarlomvi finnes i Storfjord-området.
Området er også viktig for truede, sårbare og skjøre arter innenfor isbiota, sjøfugler som alkefugler og ismåke, og bunnsamfunn i form av sent restituerende, saktevoksende og skjør megabentos som sjøfjær og svamper. På Yermakplatået er det unike, uberørte bunndyrsamfunn, men også områder nord og nordøst av Svalbard har rike, skjøre og mangfoldige bunndyrsamfunn. Kalkalger som vokser løstliggende på bunnen (rhodolitter) bidrar til økt biologisk mangfold og har en viktig økologisk funksjon på sokkelområdene ved Svalbard. Det er også betydelige rekeforekomster nær kysten og inne i fjorder og sund. Det er også flere rødlistede naturtyper i området, f.eks. polar havis (kritisk truet-CR) og kaldvannsbasseng (sterkt truet-EN).
Samlet sett er Spitsbergenbanken et svært spesielt område med mange ulike og særegne verdier og prosesser. På grunn av gunstige fysiske forhold er den årlige primærproduksjonen på Spitsbergenbanken kanskje blant de høyeste i hele Barentshavet. Det har avgjørende betydning for økosystemet i dette området. Området har betydning for beiting og oppvekst for flere fiskeslag. Dette er også et viktig næringsområde for de store sjøfuglkoloniene i områdene rundt. Spitsbergenbanken er nøkkelområde for overvintring av praktærfugl, muligens for hele Svalbard- og Øst-Grønland- populasjonen. Store deler av primærproduksjonen når bunnen, og dette gjenspeiles i høy bentisk biomasse. En ny ubeskrevet biotop for Barentshavet ble i 2017 registrert på Spitsbergenbanken med massivt forekommende skjellsandbunn med øyer av stein med brunpølse (Cucmaria frondosa), buskformet bryozo (mosdyr) og hydroider.
Alker ved Svalbard. Foto: H. Strøm, NP
Figur 8.3.1. Kart over forslag til endret SVO Havområdene rundt Svalbard (BH1). Venstre kart viser navn på ulike geografiske områder i og rundt området. Høyre kart viser grensen for det foreslåtte SVO-et med tykk rød strek, mens grenser for overlappende og nærliggende foreslåtte SVO-er er vist med tynn rød strek. Høyre kart viser også utbredelsen av SVO Polar tidevannsfront, samt utbredelsen av noen viktige miljøverdier med betydning for avgrensningen av det foreslåtte SVO-et. Bunnsamfunn, sommerbeiteområde for lodde og beiteområder for sjøfugl og hval har vært avgjørende for avgrensingen. For sjøpattedyr er det kun noen arter som er markert på kartet, men området har også stor verdi for flere andre hval- og selarter. I tillegg er forhøyet produksjon av dyreplankton og gytefelt for polartorsk markert, som viktige komponenter i næringsgrunnlaget til lodde, sjøfugl og sjøpattedyr.
8.3.2 - Vurdering i forhold til EBSA-kriteriene – Forslag til endret SVO Havområdene rundt Svalbard (BH1)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til endret SVO Havområdene rundt Svalbard (BH1)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet /sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Dyreplankton Plante-plankton
Fisk
Isbiota Bunnsamfunn Sjøpattedyr Sjøfugl
Issamfunn har en unik egenverdi i seg selv både mht. samfunnstyper (Syvertsen 1991, Bluhm mfl. 2017a) og artssammensetning, samt betydning for produksjon som kanaliseres videre i næringsnettet (Søreide mfl. 2010, 2013, Leu mfl. 2011, Kohlbach mfl. 2016, 2017, Wang mfl. 2016). Betydningen øker med breddegrad (Wassmann mfl. 2006). Noen arter forekommer bare i havis og er derfor også relevante miljøverdier for foreslått SVO Iskantsonen (Bluhm mfl. 2017b, Hop mfl. 2020). Svalbard har også fjorder med fastis og tilhørende isbiota (Gulliksen 1984, Weslawski mfl. 1993, Bluhm mfl. 2018, Marquardt mfl. 2018, Pitusi 2019, Leu mfl. 2020).
Enkeltområder, særlig Spitsbergenbanken kan ha spesielt høy primærproduksjon sammenlignet med øvrige områder i Arktis. Den er estimert til over 300 g C m-2 år-1 (Sakshaug 2009). Det kan også være forhøyet produksjon under fjordis nær brefronter pga. lokalt gode lysforhold, sterkere lagdeling og næringstilførsel som følge av oppstrømming (Vonnahme mfl. 2021). Det er strøm og topografi ved polarfronten på Spitsbergenbanken som fører til høy produksjon av plante- og dyreplankton her (Lien mfl. 2018) og i Hinlopen (ICES 2016, 2017, Knutsen mfl. 2017).
Områdets varierte bunnforhold og fjordene gir i tillegg til strømmene viktige beite, leve- og gyteområder for fiskebestander i hav og for egne fjordbestander (Madsen mfl. 2016). De grunne bankene øst for Svalbard er de viktigste beiteområdene for lodde (Gjøsæter 1999, Orlova mfl. 2010, Carscadden mfl. 2013, Fall mfl. 2018, Aarflot mfl. 2020). Det gjelder også arter som dvergkjeks og svartkjeks, som er knyttet til grunne områder og flere arter av arktiske ålebrosmer som finnes i dypt og kaldt vann (Wienerroither mfl. 2011, Bergstad mfl. 2017, Johannesen mfl. 2017, Mecklenburg mfl. 2018).
I den nordvestre delen av området, på Yermakplatået finnes det eneste habitatet i Barentshavet med stor langtidslevende arktisk megabenthosfauna som sjøfjæren Umbellula encrinus (Jørgensen mfl. 2020).
I et norsk perspektiv er Svalbard-området det viktigste (og i noen tilfeller eneste) habitatet for mange arter av arktisk endemiske marine pattedyr (Kovacs og Lydersen 2006, Hamilton mfl. 2021).
For flere av sjøfuglartene som hekker på Svalbard, utgjør bestanden her mer enn 25 prosent av europeisk bestand, f.eks. havhest, alkekonge, polarmåke og polarlomvi (Strøm mfl. 2020).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Bunnsamfunn
Dyreplankton
Isbiota
Fisk Sjøpattedyr Sjøfugl
Havis er svært viktig for arter med hele livssyklusen i isen (Lønne og Gulliksen 1991, Arndt og Swadling 2006, Bluhm mfl. 2017b, Hop mfl. 2020), men mesteparten av isen i iskantsonen og i fjordene på Svalbard er årsis og har derfor arter med bare deler av livssyklus i isen (Barber mfl. 2015, Pitusi 2019). Flere av disse kan også overleve i vannmasser (Berge mfl. 2012) eller på havbunnen (Poltermann 1998, Marquardt mfl. 2018).
Det er lokale populasjoner av den arktiske arten Calanus glacialis i enkelte fjorder som påvirkes lite av de større havstrømmene, for eksempel Billefjorden (Arnkværn mfl. 2004) og Rijpfjorden (Søreide mfl. 2010), hvor arten drar nytte av fjordis (landfast is) til å gjennomføre livssyklusen i løpet av en kort sommer (Søreide mfl. 2010, Leu mfl. 2011). C. glacialis lever også på sokkelen i Arktis generelt, men har forhøyet biomasse i disse fjordene.
Området øst av Svalbard er hovedgyteområdet og oppvekstområdet til den truede polartorsken innenfor norsk sone (Hop og Gjøsæter 2013, Eriksen mfl. 2019, Huserbråten mfl. 2019). Området er også det viktigste for både umoden og modnende lodde i beitefasen (Gjøsæter 1999, Orlova mfl. 2010, Carscadden mfl. 2013, Fall mfl. 2018, Aarflot mfl. 2020). Dette er hovedutbredelsesområdet for arktiske fisk i norske farvann (Wienerroither mfl. 2011, Mecklenburg mfl. 2018).
Dette området er livshistorisk viktig for bunnsamfunn, særlig den nordøstlige delen av området, som er den med mest arktiske bunndyrarter i nåtiden (Vassilenko og Petriashov 2009, Weslawski mfl. 2018) og sannsynligvis i framtiden.
Havområdene rundt Svalbard utgjør antakelig hele utbredelsesområdet for Svalbards hvithvaler de siste tiårene (Vacquiè-Garcia mfl. 2019). Området omfatter de viktigste sommerbeiteområdene for de vandrende bardehvalartene blåhval, finnhval og knølhval (Leonard og Øien 2020). Denne regionen er også Norges viktigste yngleområde for ringsel, storkobber og isbjørn og det viktigste furasjeringshabitatet for de fleste arktiske selarter enten på helårsbasis (voksne ringsel og storkobber) eller sesongmessig (unge ringsel og hvalross), og en betydelig del av Svalbards isbjørnbestand (rundt 300 dyr) oppholder seg på øygruppen året rundt (Laidre mfl. 2015, CAFF 2017, Hamilton mfl. 2021).
En høy andel av de arktiske sjøfuglartene hekker på Svalbard, med en rekke svært store kolonier, både på Spitsbergen og på øyene rundt (Bjørnøya, Hopen, Hinlopenstredet (Systad mfl. 2019). Hekkebestandene i øygruppen omfatter en rekke arter av arktisk og subarktisk opprinnelse, dominert antallsmessig av polarlomvi og alkekonge mens også mer sørlige arter forekommer i økende grad. Bjørnøya huser blant annet den største lomvikolonien i norske områder. I tillegg finnes arter som polarmåke, sabinemåke, rosenmåke og ismåke (Strøm mfl. 2020).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Bunnsamfunn
Dyreplankton
Isbiota Fisk Sjøpattedyr Sjøfugl
Isbiota har havis som habitat (Boetius mfl. 2015, Gradinger 2020). Både type og mengde havis i Barentshavet og fjordene på Svalbard er gjenstand for raske endringer som følge av klimaendringer (von Quillfeldt mfl. 2018, Dahlke mfl. 2020) og påvirker dermed forekomst og overlevelse av arter samt biologiske prosesser i isen (Bluhm mfl. 2017b, von Quillfeldt mfl. 2018, Tedesco mfl. 2019, Hop mfl. 2020).
Enkelte pelagiske dyreplanktonarter, som Calanus glacialis, er også påvirket av sjøis. For denne arten er oppblomstringen av isalger under isen før den pelagiske oppblomstring av planteplankton viktig for å kunne fullføre en generasjon i de korte somrene ved høy breddegrad (Søreide mfl. 2010, Leu mfl. 2011, Daase mfl. 2013, Ershova mfl. 2021). Nedgang i en nordlig C. glacialis-populasjon har følgelig blitt observert i år med lite is (men også i år med ekstremt mye is, noe som reduserer total primærproduksjon og dermed mattilgang) (Ershova mfl. 2021), og C. glacialis-populasjonsstørrelsen sørvest av Svalbard viser en negativ sammenheng med sjøtemperatur og positiv sammenheng med isutbredelse året før (Kwasniewski mfl. 2012). Men på større områder og lengre tidsserier har ikke C. glacialis vist en nedgang i bestandsstørrelse enda (Hop mfl. 2019, ICES 2020 a).
Områdene rundt Svalbard er et svært viktig habitat for polartorsk (Boitsov mfl. 2013, Hop og Gjøsæter 2013, Eriksen mfl. 2019, Gjøsæter mfl. 2020) og andre arktiske fiskearter, flere av disse har en nedadgående trend (Johannesen mfl. 2017).
I hvilken grad dette området er særlig viktig for bunndyrsamfunnene er ikke direkte dokumentert, men artssammensetningen i den nordlige og nordøstlige delen har en betydelig del av saktevoksende og skjøre arter (Jørgensen mfl. 2020) og arter som forekommer i kaldt vann (Jørgensen mfl. submitted).
Områdene rundt Svalbard er et svært viktig habitat for mange truede arktisk endemiske marine pattedyr som alle er is-assosierte og derfor sårbare for de pågående klimaendringene (Kovacs mfl. 2011, Hamilton mfl. 2021).
På Svalbard (som også inkluderer Bjørnøya) er det observert bestandsreduksjoner hos sjøfugl der polarlomvi har hatt en nedgang på 25-50 prosent siden slutten av 1990-tallet (Arneberg og Jelmert 2017). Hvis denne raten ikke endrer seg, har polarlomvi på Svalbard en høy sannsynlighet for å gå mot kvasi-ekstinksjon (så lavt nivå at de muligens ikke er i stand til å komme seg) innen de neste 50 årene.
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Fisk
Isbiota Bunnsamfunn Sjøpattedyr Sjøfugl
Isbiota, særlig arter i flerårsis vil være utsatt ved klimaendringer. Redusert isutbredelse, sen isdannelse/tidlig smelting og mindre fjordis (Dahlke mfl. 2020) gir kortere produktiv sesong for isbiota (Wassmann og Reigstad 2011, Hop mfl. 2013, Barber mfl. 2015). Havis som habitat er også følsomt i forhold til oljesøl fra skipstrafikk (Rusten 2014).
Området inneholder arktiske arter inkludert polartorsk som er sårbare for klimaendringer, inkludert reduksjon i havis (Bergstad mfl. 2017, Fossheim mfl. 2015, Huserbråten mfl. 2019).
Vest og øst for Svalbard: store oppreiste, skjøre megabentiske arter som kan ha lav restitusjonsevne og som bidrar til den høyeste biodiversiteten i, og biomassen av, megafauna i Barentshavet (Jørgensen mfl. 2015a, b, Jørgensen mfl. 2019). Restitusjon av hardbunnsbentossamfunnet kan ta 10-20 år i Svalbard-området (Keck Al-Habebeh mfl. 2020).
Kystnære isdekte områder er viktige yngle- og furasjeringshabitater for arktisk endemiske marine pattedyr. Det blir mindre både av årsis og også breis som følge av global oppvarming, og dette er en trussel for disse svært viktige områdene for denne dyregruppen (Lydersen mfl. 2014, Meredith mfl. 2019). Det samme gjelder for sjøfugl som er avhengige av arktisk planktonfauna knyttet til iskantsonen, særlig arter som alkekonge og krykkje (Amélineau mfl. 2019).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Sjøfugl
Planteplankton Dyreplankton Bunnsamfunn
Isbiota Fisk
Primærproduksjonen i isen starter tidligere enn i vannmassene noe som bidrar til å forlenge den produktive sesongen i et område (Hegseth 1998, Leu mfl. 2011, 2015). Isalger bidrar relativt sett mer til den totale primærproduksjonen jo lenger nord man kommer (Wassmann mfl. 2006, Wassmann og Reigstad 2011). Primærproduksjonen i isen er viktig for pelagiske og bentiske næringsnett både i fastis og drivis (Kohlbach mfl. 2019).
Innenfor det foreslåtte SVO-et er det flere områder (f.eks. Spisbergenbanken og Hinlopen) med høyere primærproduksjon i vannmassene på grunn av topografiske forhold som fører til vertikal gjennomblanding (Lien mfl. 2018). Det kan også være forhøyet produksjon under fjordis nær brefronter pga. lokalt gode lysforhold, sterkere lagdeling og næringstilførsel som følge av oppstrømming (Vonnahme mfl. 2021). Området overlapper med SVO Iskantsonen som også har forhøyet produksjon (von Quillfeldt mfl. 2018).
Det er høy sekundærproduksjon av dyreplankton i fjordene rundt Svalbard, og dette er høyere enn i Framstredet og nord for 83°N (Hop mfl. 2019 a, Hop mfl. 2019b). Strøm og topografi ved polarfronten på Spitsbergenbanken fører også til høy produksjon av dyreplankton her (Lien mfl. 2018) og i Hinlopen (ICES 2016, 2017, Knutsen mfl. 2017).
Området overlapper med polarfront/iskantsone som er viktig beiteområde for mange fiskeslag i sommerhalvåret – se foreslått SVO Iskantsonen. Dette er et viktig oppvekstområde for polartorsk i Nordvest-Barentshavet, hvor 0-gruppe polartorsk beiter på plankton gjennom sommeren og høsten (Boitsov mfl. 2013, Eriksen mfl. 2015, 2019, Gjøsæter mfl. 2020). Områdene øst for Svalbard er viktige beiteområder for lodde og korrelert med høy vekst (Gjøsæter 1999, Berg mfl. innsendt). Dette gjelder også torsk som her beiter på lodde og polartorsk i sommerhalvåret (Johannesen mfl. 2012, Fall mfl. 2021).
Megabentosfauna er kjent for å tilføre bunnen økt habitatvariasjon og leveområder. Vest for Svalbard gir den høye biomassen av store svamper bidrag til den årlige produksjonen av ny biomasse, og produksjon:biomass ratio er særlig høy nord og nordvest for Svalbard i dette SVO-et (Degen mfl. 2016).
Sjøfugl har en viktig funksjon i å bringe biologisk materiale fra marine til terrestre miljøer, både i form av gjødslingseffekter og som byttedyr for terrestre arter. For eksempel fjellrev, men også isbjørn, spiser sjøfugl og egg i koloniene (Prop mfl. 2015).
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Planteplankton Dyreplankton
Isbiota Fisk Bunnsamfunn Sjøpattedyr Sjøfugl
Isbiota utgjør en rekke ulike samfunnstyper og består av et høyt antall arter fra en rekke ulike artsgrupper (Syvertsen 1991, Horner mfl. 1992, Gradinger mfl. 2010, Poulin mfl. 2011, Bluhm mfl. 2011, Bluhm mfl. 2017a, b og 2018, Gradinger 2020, Hop mfl. 2020).
Planteplanktonsamfunnene består av både tilførte og lokale arter (von Quillfeldt 1996, Hasle og Heimdal 1998, Hegseth mfl. 2019). Stor variasjon i topografiske og oseanografiske forhold som påvirker stabilitet, lys og næringsforhold, favoriserer også arter på ulikt vis, noe som samlet sett fører til mange ulike samfunn (Heimdal 1989, von Quillfeldt 1996, Poulin mfl. 2011, Assmy mfl. 2017, Lovejoy mfl. 2017, Johnsen mfl. 2018, Vonnahme mfl. 2021). Det finnes også arter som ikke forkommer i farvann lenger sør, f.eks. Fossula arctica, en art som kan forekomme tidlig i våroppblomstringen (Hasle mfl. 1996).
Sammenlignet med Framstredet og Barentshavet, har Svalbardfjorder som er sterkt påvirket av å være kystnære, en høy andel meroplankton (planktoniske larvestadier av bentiske arter (Gluchowska mfl. 2017). Området inneholder både arktiske og atlantiske vannmasser og dermed flere dyreplanktonsamfunn (Blachowiak-Samolyk mfl. 2008).
Området har stor variasjon i dybdeforhold og oseanografiske forhold, og har derfor en rik fiskefauna (Bergstad mfl. 2017). Både yngel og voksen polartorsk og lodde danner hotspots som er attraktive beiteområder for andre fisk, sjøfugl og sjøpattedyr (Eriksen mfl. 2017, Gjøsæter mfl. 2020).
Områdene vest, nord og øst for Svalbard har den største biodiversiteten i Barentshavet av megafauna (Jørgensen mfl. 2019). Vest-, nord- og østsiden har forskjellige bunnsamfunn som er tilpasset de forskjellige miljøene. Samfunnet øst for Svalbard har lavere temperaturpreferanse enn samfunnet på vestsiden. Rhodolittene bidrar til økt biologisk mangfold og har en viktig økologisk funksjon på sokkelområdene ved Svalbard (Teichert 2014).
Områdene rundt Svalbard er et nøkkelområde for mange arktisk endemiske marine pattedyr og sjøfugl i Barentsregionen (Laidre mfl. 2015, CAFF 2017, Hamilton mfl. 2021).
Sjøfuglbestandene som hekker i øygruppen, omfatter en rekke arter av arktisk og subarktisk opprinnelse. De arktiske artene domineres antallsmessig av polarlomvi og alkekonge, men også mer sørlige arter forekommer i økende grad. Bjørnøya huser blant annet den største lomvikolonien i norske områder, og storjo er blitt vanlig flere steder. I tillegg finnes arter som polarmåke, sabinemåke, rosenmåke og ismåke (Strøm mfl. 2020).
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
Dyreplankton
Sjøpattedyr Sjøfugl
Isbiota
Fisk Bunnsamfunn
Klimaendringer har endret isdekke, temperatur, lagdeling og avrenning i Svalbardfjorder og havområdene rundt (Førland mfl. 2011, Onarheim mfl. 2014, Lind mfl. 2018, Pavlova mfl. 2019, Tverberg mfl. 2019, Skogseth mfl. 2020), og det er økende skipsfart rundt øygruppen. Med unntak av de pågående klimaendringene (Tedesco mfl. 2019), er isbiota lite direkte påvirket av menneskelig aktivitet som skipstrafikk, fiskeri, forurensning, introduserte arter m.m. (Eamer mfl. 2013).
Selv om enkelte arter viser økende trender (Calanus finmarchicus) eller nedgang (Pseudocalanus sp.) (Hop mfl. 2019, Carstensen mfl. 2019) i takt med klimaendringer, så er dyreplankton i havområdene rundt Svalbard likevel relativt lite påvirket av menneskelig aktivitet.
Deler av området har vært isdekket og/eller har så lave forekomster av kommersielt viktige arter at det ikke har vært fisket, f.eks. på Yermakplatået. (Bergstad mfl. 2017, Huserbråten mfl. 2019, Gjøsæter mfl. 2020, Jørgensen mfl. 2020).
Selv om tidligere tiders fangstaktivitet hadde stor innvirkning på mange arktisk endemiske marine pattedyr i Svalbardområdet, så gjør lovgivning og isdekke at det er en begrenset effekt av menneskelige aktivitet i marine områder på Svalbard slik at enkelte bestander er på vei opp igjen i disse områdene (f.eks. isbjørn - Aars mfl. 2017; hvalross – Kovacs mfl. 2014).
Øygruppen er også et refugium for sjøfugl med mindre menneskelig påvirkning enn på fastlandet, og mangler flere av predatorene på fastlandet (Strøm mfl. 2020).
8.3.3 - Lokalisering
Forslag til avgrensing av SVO Havområdene rundt Svalbard i Meld. St. 20 (2019-2020) er kun basert på en vurdering av miljøverdiene i området. Sårbarhet har aldri vært vurdert for dette området. Området strekker seg fra 100 km sør for Bjørnøya til nordlig grense av forvaltningsplanen for Barentshavet (figur 8.3.1). Kandidatområdene hvor beitearealene for pelagisk beitende sjøfugl strekker seg 100 km ut fra de viktigste hekkekoloniene på øygruppen var en del av den opprinnelige begrunnelsen for avgrensing av området i Meld. St. 20 (2019-2020). I sør er deler av området utvidet noe i forhold til forslag til avgrensning i stortingsmeldingen. Dermed inkluderes også viktige beiteområder for lodde.
Forslag til endret SVO Havområdene rundt Svalbard overlapper med foreslått SVO Iskantsonen og nordlige deler av foreslått SVO Eggakanten nord (figur 8.3.1). Området dekker også SVO Polar tidevannsfront som fagekspertgruppen nå anbefaler å inkludere i foreslått SVO Havområdene rundt Svalbard på lik linje med andre viktige delområder i dette SVO-et, bl.a. Spitsbergenbanken hvor denne fronten er lokalisert.
De marine verneområdene på Svalbard og flere av de «nye fiskeområdene» som er definert i reglene som regulerer fiskeriene med bunnredskap i det nordlige Barentshavet og områdene rundt Svalbard, vedtatt i 2019, inngår også i dette området.
8.3.4 - Introduksjon til området
Havområdene rundt Svalbard har en varierende bunntopografi, strømforhold og hydrografi, og har svært varierende isforhold. Isutbredelsen, iskantens struktur og form og iskonsentrasjonen i disse havområdene er delvis kontrollert og stedvis sterkt påvirket av land, vind, havbunnens topografi og havstrømmer (Walczowski mfl. 2012, 2017). Isdekke og iskonsentrasjon i området har stor sesongmessig (figur 8.3.2) og mellomårlig variasjon, hvilket kan ha betydning for arter som er avhengig av isen som habitat (von Quillfeldt mfl. 2018).
Figur 8.3.2. Figuren viser isfrekvens i april (venstre) og september (høyre) for perioden 1990–2019. Isfrekvensen er prosentandelen av dager det forekommer havis i et gitt område innenfor en nærmere angitt periode (Kilde: Norsk Polarinstitutt).
Havområdene rundt Svalbard har både arktiske og atlantiske vannmasser. Det er lite is på vestsiden fordi Vestspitsbergenstrømmen bringer varmt og salt atlanterhavsvann nordover (figur 8.3.3). Atlanterhavsvannet dominerer i områdene vest og nord av Svalbard, mens kaldere og ferskere arktisk vann dominerer øst for Svalbard (Helland-Hansen og Nansen, 1909, Loeng, 1991). Det arktiske vannet strømmer sørover på østsiden av Svalbard og nordover som en kyststrøm også vest av Spitsbergen, nærmere kysten enn atlanterhavsstrømmen (Svendsen mfl. 2002, Fig. 8.3.3). Ferske og kalde vannmasser okkuperer vanligvis øvre vannlag inne i fjordene, mens atlanterhavsvann strømmer inn i dypet (Nilsen mfl. 2016). Det siste tiåret har atlanterhavsvann strømmet inn også i øvre del av vannsøylen i noen av fjordene, i fravær av ferskere og lettere vannmasser sin tilstedeværelse og da har disse fjordene vært isfrie hele vinteren (Skogseth mfl. 2020).
Figur 8.3.3. Gjennomsnittlig overflatestrøm (piler) og bunndyp (farger) i forslag til endret SVO Havområdene rundt Svalbard og omkringliggende områder.
Vestspitsbergenstrømmen holder det meste av havområdene vest og nord av Svalbard isfrie hele året, men disse områdene påvirkes også av tilførsel av havis fra Polhavet, lokal isdannelse i fjordene og tilførsel av havis rundt Sørkapp og nordover nært land sammen med kalde, ferske vannmasser i kyststrømmen. Havisen rundt Svalbard er typisk satt sammen av førsteårsis og ulike unge isklasser. Flerårsis kan opptre, transportert inn fra Polhavet, særlig nord og øst for Svalbard (f.eks. Kwok mfl. 2005, 2008, Onarheim mfl. 2014). Fastis i fjordene på vestkysten av Spitsbergen (f.eks. Kongsfjorden og van Mijenfjorden) blir typisk ikke like tykk som fastisen i fjorder som er mindre påvirket av atlanterhavsvann (f.eks. Rijpfjorden og Storfjorden), se Gerland mfl. (2008, deres figur 3). Overflatesnø (snø oppå havisen) og isdannelse og smelting er viktige for utviklingen av havisdekket, og værforhold som lufttemperatur, nedbør og vind spiller en viktig rolle i denne prosessen. I tillegg er havisoverflatens topografi viktig for is og snødannelse (se f.eks. Nicolaus mfl. 2003, Cheng mfl. 2008, 2014, Wang mfl. 2015).
Det kjemiske miljøet i havet rundt Svalbard og i dets fjorder er særlig påvirket av fluktuasjoner i det varme, salte atlanterhavsvannet fra sørvest, med høy uorganisk karbon/CO2 og alkalinitet, og det kalde, ferske arktiske vannet fra nordøst med lavt innhold av uorganisk karbon/CO2 og alkalinitet (Fransson mfl. 2016, Chierici og Fransson 2018, Chierici mfl. 2019). Isbiota og marint liv i fjordene ved Svalbard vil i særlig grad påvirkes av de fysiske og kjemiske forholdene i fjordene som forårsakes av tilførsel av ferskvann fra breissmelting ved kysten og i Svalbardfjordene (med isbreer) (Fransson mfl. 2015, 2016, 2020, Hopwood mfl. 2020). Med økt tilførsel av ferskvann øker potensialet for CO2-opptak og aragonittmetningen (ΩAr) minker (Fransson mfl. 2015, 2016, Ericson mfl. 2019, Chierici mfl. 2019). I fjorder på Vest-Spitsbergen (Tempelfjorden, Kongsfjorden) er lave aragonitt (ΩAr)-verdier (<1,4) observert, hvilket kan gi alvorlige konsekvenser for kalsifiserende marine organismer som vingesnegl (Fransson mfl. 2015, 2016, Hopwood mfl. 2020). På den annen side kan smeltevannet fra isbreer bidra med kjemiske stoffer som for eksempel jern, som er viktige for primærproduksjonen, og dermed CO2-opptak (Hopwood mfl. 2020). Høy biologisk produksjon er observert nær brefronter i Svalbardfjorder, eksempelvis i Kongsfjorden (Halbach mfl. 2019, Hopwood mfl. 2020). I tillegg er havet rundt Svalbard i deler av året påvirket av havisdannelse og havissmelting, hvilket bidrar til endringer i de kjemiske forhold i overflaten. Prosesser i isen kan bidra med endringer i både alkalinitet og CO2, avhengig av om det er isdannelse eller issmelting, og i havet rundt Svalbard (i det arktiske vannet) bidrar isprosessene til at CO2-innholdet i overflaten er relativt lavt (lavere enn atmosfæriske verdier, ca. 410 µatm) og havet kan dermed ta opp mer CO2 fra atmosfæren (Fransson mfl. 2017, 2020, Chierici mfl. 2019). Stor primærproduksjon er hovedgrunnen til at Barentshavet tar opp CO2 fra atmosfæren (Fransson mfl. 2001, Lauvset mfl. 2013). Modellresultater antyder endringer i pH i overflaten i havet rundt Svalbard og varierer fra 0 til -0,006 per år i perioden 1998-2016, med den største endringen i Storfjorden (Becker mfl. 2020) der det er et stort opptak av CO2 på grunn av dypvannsdannelse (Anderson mfl. 2004). Sør for Svalbard (ved 72,8°N) viser observasjoner (2012-2018) at konsentrasjonen av uorganisk løst karbon ved 300-400 m dyp er knyttet til mellomårlige variasjoner i polarfronten (Chierici mfl. 2017, Jones mfl. 2019). Fra 2016 til 2018, minket aragonittmetningen (ΩAr) fra 1,8 til 1,4 (Jones mfl. 2019). I området øst for Svalbard, i de nordlige delene av Barentshavet, vises små endringer i karbonkjemi og havforsuring fra 2012 til 2018 på 250-300 m dyp. I denne perioden økte pH med 0,006 per år, men uten tydelig trend. Aragonittmetning varierte i perioden med middelverdi på 1,3 (Jones mfl. 2019). Dette området er påvirket av økt kaldt og ferskt arktisk vann, hvilket også bidrar til stor variasjon i karbonkjemien (Chierici mfl. 2017). I de øvre 50 m i overflaten er aragonittmetningen ca. 1,5-1,6 i de nordlige delene av Barentshavet og 1,7-2,0 i de sørlige delene (Lauvset mfl. 2016). Høy dynamisk omrøring og høy primærproduksjon bidrar til at disse områdene har høyt karbonopptak fra atmosfæren og sannsynligvis hurtig CO2-utveksling (Lauvset mfl. 2013). På høsten, i forbindelse med vertikal omrøring, transporteres karbon og næringssalter til overflaten, og dette gir stor sesongvariasjon i kjemiske parametere som næringssalter, pH og kalkmetning. I de øvre 50 m i overflaten er aragonittmetningen 1,8-2,0 og pH ca. 8,10 (Lauvset mfl. 2016), med forventet minking med økt CO2-opptak.
Den polare tidevannsfronten, som ligger på Spitsbergenbanken, inngår nå som en del av dette SVO-et (figur 8.3.1). Spitsbergenbanken er grunn (40-100 meter dyp) og karakterisert av relativt sterk strøm langs kanten (Loeng 1991, Lien mfl. 2018) og sterke tidevannsstrømmer (Gjevik mfl. 1994). De delene av banken som er grunnere enn 50-60 meter er stort sett vertikalt gjennomblandet på grunn av tidevann og vind (Fer og Drinkwater 2014). Dette fører til at det i sommerhalvåret settes opp en såkalt «aktiv» tidevannsfront med sterk vertikalblanding rundt banken på omtrent 50-100 m bunndyp (Fer og Drinkwater 2014). Slike fronter er ofte assosiert med økt primærproduksjon (Le Fèvre 1987), fordi næringssalter tilføres den eufotiske sonen gjennom økt vertikalblanding eller oppstrømming (f.eks. Allen mfl. 2005), men dette er i liten grad tilfelle her, bortsett fra en kort, intens våroppblomstring (Lien mfl. 2018). Spitsbergenbankvannet fører også med seg høy CO2 og lav alkalinitet som gir lav pH og aragonittmetning (Fransson mfl. 2015). «Aktive» fronter har også ofte høye konsentrasjoner av dyreplankton og fiskelarver (Franks 1992, Munk mfl. 2009) og kan være “hotspots” for det marine livet (Belkin mfl. 2009). Den polare tidevannsfronten bidrar dermed til å gjøre deler av Spitsbergenbanken til en "hotspot" med tanke på biologisk produksjon (Lien mfl. 2018). Ettersom Spitsbergenbanken er svært grunn, har den store variasjoner i sjøtemperatur (fra under –1 °C til over 5 °C) mellom sommer og vinter (Bergstad mfl. 2017), og det er tilsvarende store forskjeller mellom år (Årthun mfl. 2011). I tillegg er det store variasjoner i den kjemiske tilstanden mellom sesonger (Lauvset mfl. 2013) og mellom år (Becker mfl. 2020), med stort opptak av atmosfærisk CO2.
Som også beskrevet for foreslått SVO Iskantsonen, er det for isdekket på tidevannsfronten ved Spitsbergenbanken stor sesongvariasjon, med dannelse av førsteårsis om vinteren og isfrie somre (Ådlandsvik og Loeng 1991, Gammelsrød mfl. 2009, Vinje 1998 og 2001). Siden Spitsbergenbanken er lokalisert i den sørvestre delen av Barentshavet, kan hele eller deler av området være isfritt året rundt i varme år (særlig siste tiår) på grunn av innstrømning av varmt atlanterhavsvann (f.eks. Helland-Hansen og Nansen 1909, Loeng 1991). Det er også vist at det er en trend mot forsinket isdannelse på vinteren (Stroeve og Notz 2018), der én sannsynlig årsak er at det er stor oppvarming av overflatevann om sommeren (Lind mfl. 2018). I kalde år er området isdekt med maksimum i mars-april. Den polare tidevannsfront er også påvirket av innstrømning av is fra nordlige Barentshavet, som stammer fra Polhavet/Karahavet, og flerårsis kan opptre enkelte år (King mfl. 2017, Lind mfl. 2018).
8.3.5 - Beskrivelse av miljøverdiene
Isbiota
Da store deler av det foreslått endrete SVO Havområdene rundt Svalbard også omfatter det foreslått endrete SVO Iskantsonen, vil isbiota slik den er beskrevet for foreslått SVO Iskantsonen også gjelde for foreslått SVO Havområdene rundt Svalbard. I tillegg kommer isbiota i fjordis hvorav noe er fastis. Det er relativt få studier av isbiota i fjordene på Svalbard, og kunnskap er hovedsakelig basert på nyere studier i van Mijenfjord og Hornsund, noe fra Billefjorden (f.eks. Vonnahme mfl. 2021) og Kongsfjorden og Rijpfjorden (Søreide 2019), hvorav mye ennå ikke er publisert.
Hvor lenge en fjord er islagt spiller inn på utvikling av issamfunn over tid, men det er uansett snakk om sesongis som koloniseres hvert år (Weslawski mfl. 1993). Tilgjengelig lys, istykkelse og avstand til land har betydning for forekomst av arter (Bluhm mfl. 2018).
Issamfunn i fastis i kystnære områder skiller seg fra issamfunn i drivisen ved at det ofte er høy algebiomasse (Leu mfl. 2015) og stor forekomst av ismeiofauna, ofte med en stor andel av meroplankton (dvs. larver og unge stadier av bentiske dyrearter, f.eks. flerbørstemark) (Gradinger mfl. 2009, Bluhm mfl. 2018, Pitusi 2019, Gradinger og Bluhm 2020). Noen ganger skiller ikke sammensetningen av mikroalger og flercellede arter i fjordis seg fra den i underliggende vannmasser (Weslawski mfl. 1993), men det er mer typisk at det kan være en større relativ forekomst av f.eks. pennate kiselalger og da gjerne mer typiske isassosierte arter enn i vannmassene (Leu mfl. 2010 og 2020, van Leeuwe mfl. 2018). Isfauna som er avhengig av havis gjennom hele livssyklusen og som gjerne forekommer i flerårsis, observeres hovedsakelig i fjordissamfunn de årene hvor drivis kommer inn i fjordene (Weslawski og Adamski 1987), men er ellers sjelden under isen (Gulliksen 1984).
Mengde isfauna varierer med sesong og kan relateres til forekomst av isalger (Bluhm mfl. 2018). Forekomst av enkelte arter som f.eks. Sympagohydra tuuli i fjordis på vestsiden av Svalbard, styrker teorien om at for enkelte arter av isfauna er det en viktig kobling mellom is og havbunn i områder hvor det ikke er is hele året (Marquardt mfl. 2018). Sammensetning av isfauna reflekteres også i dietten til viktige beitere som polartorsk (Weslawski mfl. 1993).
Plankton
Det foreligger en del studier av planteplanktonbiomasse, produksjon og sammensetning fra farvannet rundt Svalbard. Mange av disse studiene er fra de kystnære områdene, bl.a. Kongsfjorden (Hop mfl. 2002, Hodal mfl. 2012, Hegseth mfl. 2019). For den vestlige delen vil innstrømmende atlantiske vannmasser være viktig for produksjonen på den vestlige sokkelen (Hegseth mfl. 2019, Vernet mfl. 2019). I de nordlige og østlige delene vil det være store forskjeller i planteplanktonproduksjon og biomasse, da den pelagiske produksjonen forekommer i både isfylte og isfrie områder, avhengig av årstid. Studier i den østlige delen viser derfor stor variasjon (Hegseth 1998, Owrid mfl. 2000), der planteplanktonproduksjonen er høyere i de kystnære områdene påvirket av smeltevann og/eller oppstrømning, som f.eks. Kongsfjorden. Også feltstudier (Hasle og Heimdal 1998, Owrid mfl. 2000) og modelleringer (Vernet mfl. 2019) tyder på høy produksjon vest i det foreslåtte SVO-et. Undersøkelser i nordlig del av Svalbard tyder på at også områder som f.eks. Hinlopen er viktige områder for planteplanktonproduksjon (SI_ARCTIC, HI upubliserte data). Tidevannsfronten ved Spitsbergenbanken er på grunn av sine fysiske særtrekk også et område hvor planteplankton-oppblomstringen er mer intens enn i omkringliggende områder, inkludert øvrige deler av polarfrontområdet, men også mer kortvarig (Lien mfl. 2018). Primærproduksjonen på Spitsbergenbanken er estimert til over 300 g C m-2 år-1, noe som er høyt sammenlignet med andre deler av Arktis (Sakshaug 2009), men det kan være store variasjoner fra et år til et annet. Senere studier har også vist forhøyet produksjon under fjordis nær brefronter pga. lokalt gode lysforhold, sterkere lagdeling og næringstilførsel som følge av oppstrømming (Vonnahme mfl. 2021).
Svalbardområdets planteplanktonsamfunn består av arter med ulik opprinnelse, både tilførte og lokale (von Quillfeldt 1996, Hasle og Heimdal 1998, Hegseth mfl. 2019). Stor variasjon i topografiske og oseanografiske forhold (fjord/åpent hav, grunne banker/dyphav, arktiske/atlantiske vannmasser, havis i deler av området m.m.) påvirker stabilitet, lys og næringsforhold, alle faktorer som favoriserer forskjellige planteplanktonarter på ulikt vis. Hele området sett under ett kan derfor ha et relativt høyt antall arter (Heimdal 1989, von Quillfeldt 1996, Poulin mfl. 2011, Assmy mfl. 2017, Lovejoy mfl. 2017, Johnsen mfl. 2018, Vonnahme mfl. 2021). Det finnes også arter som ikke forkommer i farvann lenger sør, f.eks. Fossula arctica, en art som kan forekomme tidlig i våroppblomstringen (Hasle mfl. 1996).
Dyreplanktonbiomasse rundt Svalbard styres av forholdene mellom atlantiske og arktiske strømmer (Blachowiak-Samolyk mfl. 2008, Wassmann mfl. 2015, Gluchowska mfl. 2017, Basedow mfl. 2018, Hop mfl. 2019), og prosesser relatert til fjordøkologi og sjøis (McGovern mfl. 2020, Leu mfl. 2011, Trudnowska mfl. 2014). Atlanterhavsstrømmen transporterer høy biomasse av atlantiske arter, som Calanus finmarchicus, nordover langs vestkysten av Svalbard, inn i noen av de åpne fjordene på vestkysten og øst- og nordover nord for Svalbard. Sørgående arktiske vannmasser, med typiske arter som Calanus glacialis dominerer på østsiden av Svalbard og i sør opp til Hornsund. Arktiske arter som C. glacialis er større og mer energirike enn atlantiske arter som C. finmarchicus, og derfor har utbredelsen av atlantiske og arktiske vannmasser stor betydning for mengden og tilgjengeligheten av mat for høyere trofiske nivåer som beiter på dem. Men samtidig har C. finmarchicus høyere årlig 2. produksjon enn C. glacialis. Selv om artene har ulikt lipidinnhold, som er viktig for arter som sjøfuglen alkekonge (Alle alle), som fanger individuelle hoppekreps, er det ikke gitt at beiting på større mengder C. finmarchicus vil være et betydelig problem (Karnovsky mfl. 2003, Kwasniewski mfl. 2012).
Ettersom dette er et stort område, finnes det flere dyreplanktonbiomassemålinger i området. Tidsserier av dyreplankton eksisterer fra vestkysten: Kongsfjordentransekt mot Framstredet (1996-nå) (Hop mfl. 2019), Isfjorden (2011-nå; Isfjorden Marine Observatory Svalbard), i nord: Rijpfjorden på Nordaustlandet 2010–2014 (Hop mfl. 2019b), rundt øygruppen og i Barentshavet (ICES 2017, 2018, 2019, 2020 a) og i Framstredet (Carstensen mfl. 2019), mens data fra fjorder ved østkysten er det færre av.
Ishavsåte. Foto: Allison Bailey, NP
Biomasse av dyreplankton i Svalbardfjordene er ofte høyere enn i det åpne havet rundt. For eksempel viser Hop mfl. (2019) at dyreplanktonbiomasse målt i en tyveårig tidsserie i Kongsfjorden er opp til fem ganger så høy som i østlige Framstredet og dobbelt så høy som på sokkelen utenfor fjorden. Biomasse i en høyarktisk fjord, Rijpfjorden, på Nordaustlandet viser samme trend, med høyere biomasse inne i fjorden enn både i atlantiske vannmasser på kontinentalsokkelen og i arktiske vannmasser nord for sokkelen (Daase og Eiane 2007, Hop mfl. 2019b). Nötig mfl. (2015) viser også at dyreplanktontetthet er høyere i den atlantiske Vestspitsbergenstrømmen enn lenger vest i blandede og polare vannmasser i Framstredet. I en oversikt over dyreplanktonbiomasse i Barentshavet de siste tre tiår fant Dalpadado mfl. (2020) at havområdet på vest- og nordkysten av Svalbard var ett av tre områder med høyest dyreplanktonbiomasse. I flere områder rundt Svalbard (foruten fjordene) er dyreplanktonbiomassen ofte relativt høy nord for øygruppen (over 10 g tørrvekt/m2) og i Storfjordrenna sør for øygruppen, mens i arktiske vannmasser sørøst for Svalbard kan biomassen være mye lavere (under 3 g tørrvekt/m2) (ICES 2017, 2018, 2019, 2020 a).
Hop mfl. (2019) viser at det er en økning i den atlantiske hoppekrepsarten C. finmarchicus, men ingen endring i den arktiske slektningen, C. glacialis i Kongsfjorden. Mindre hoppekreps som Oithona similis og Pseudocalanus arter har vist en nedgang fra 25-50 prosent fra 1996-2006 til 2006-2016 i Kongsfjorden (Hop mfl. 2019) og i Vestspitsbergenstrømmen (Carstensen mfl. 2019).
Hinlopenstredet og områdene nord for dette er et meget dynamisk område, med en blanding av ulike vannmasser, noe som gir gode betingelser for mange arter av dyreplankton. Området er påvirket av atlantisk innstrømning og har varierende grad av isdekning. De atlantiske vannmassene tilfører næringsstoffer, biomasse og varme (Menze mfl. 2020). Gjennomsnittlig dyreplanktonbiomasse i Hinlopenområdet er ca. 20 g tørrvekt/m2 (Knutsen mfl. 2017), mye høyere enn i Barentshavet (6-8 g tørrvekt/m2; Dalpadado mfl. 2020). Atlantiske og arktiske vannmasser, og blanding av disse, gir forutsetninger for de tre vanligste Calanus-artene, den arktiske boreale C. finmarchicus, den arktiske neritiske C. glacialis, og arktiske C. hyperboreus (Daase mfl. 2008). Alle utviklingsstadier av C. finmarchicus og C. glacialis blir observert i Hinlopenområdet (Walkusz mfl. 2003, Søreide mfl. 2008), hvilket indikerer at disse kopepodene kan reprodusere i disse omgivelsene. C. finmarchicus blir observert i varierende grad i hele vannsøylen, C. glacialis i de øverste 200 m og C. hyperboreus under 200 m dyp (Daase mfl. 2008). I tillegg bidrar små kopepoder som Oncaea borealis, Oithona similis og Pseudocalanus spp. mye til antallet av dyreplanktonorganismer. I tillegg til kopepoder, dominerer krill og amfipoder i de øverste 50 meter i Hinlopenområdet (Knutsen mfl. 2017). I de dypere områdene (250-600 m) nord for Hinlopenstredet viser akustiske registreringer og trålfangster et relativt svakt lag bestående av mesopelagisk fisk og større dyreplankton (Knutsen mfl. 2017). Akustiske undersøkelser viser også stor tetthet av krill i Hinlopenområdet (Ressler mfl. 2015).
Spitsbergenbanken er dominert av sterke tidevannsstrømmer og sterk vertikal blanding (Lien mfl. 2018), noe som gir grunnlag for en meget høy primærproduksjon over banken, der karbonmengden er beregnet til 300 g C/m2 per år (Sakshaug mfl. 2009), og gode levemuligheter for dyreplankton (Orlova mfl. 2005, Dalpadado mfl. 2020), fisk (Loeng og Drinkwater 2007) og hval (Skern-Mauritzen mfl. 2011).
Spitsbergenbanken er påvirket både av atlantiske og arktiske vannmasser, dette gjenspeiles i sammensetningen av dyreplankton i området, hvor den arktiske C. glacialis og atlantiske C. finmarchicus dominerer blant hoppekrepsene. Isdekket over banken og issmeltingen varierer mellom år, avhengig av temperaturen, dette påvirker starten på primærproduksjonen, den påfølgende sekundærproduksjonen og artssammensetningen (Orlova mfl. 2005, Dalpadado mfl. 2020). I det varme året 1983 ble det funnet 3 mg C. glacialis/m3 i august, et år da iskanten trakk seg tidlig tilbake og årets produksjon kunne starte tidlig. I det kalde året 1987 med sen produksjonsstart ble det observert 89 mg C. glacialis/m3 i august (Orlova mfl. 2005).
Undersøkelser av Spitsbergenbanken og nærliggende områder (1989-2017) viste i likhet med flere banker, lave dyreplanktonbiomasser (4-6 g tørrvekt/m2) i Barentshavet, i forhold i de fleste andre områdene hvor biomassene var over 10 g tørrvekt/m2 (Dalpadado mfl. (2020). Den til enhver tid eksisterende biomassen av dyreplankton over banken vil nødvendigvis ikke gjenspeile den høye produksjonen. Bestanden av dyreplankton over grunne bankområder vil lett kunne reduseres pga. beiting fra visuelle predatorer, siden planktonet ikke kan unnslippe ved å oppsøke dypere, mørke vannmasser (Aarflot mfl. 2019). Dyreplanktonet over Spitsbergenbanken blir spist av fisk, særlig lodde, men også hval (Skern-Mauritzen mfl. 2011, ICES 2019). Lodda på næringsvandring kan muligens spise alt av dyreplankton i de øvre opplyste vannmassene i løpet av få dager (Hassel mfl. 1991). Undersøkelser av ernæringen hos lodde på banken i august-september viste ikke bare en meget høy grad av magefylling, men også at mageinnholdet var dominert av hoppekreps (C. finmarchicus, C. glacialis, C. hyperboreus og Metridia longa) og krill (Thysanoessa inermis). På samme tid var næringsopptaket hos lodde i dypere områder mindre intens og besto kun av krill. Krillen på banken ble samtidig spist av torsk (Michalsen mfl. 2007). Det mangler kunnskap om endringer i artssammensetning, mengde og biomasse i dyreplankton gjennom året i dette viktige området.
Fisk
Polartorsk (Boreogadus saida) som er nøkkelart i det nordlige Barentshavet, er tilknyttet isfylte farvann og lever i de frie vannmassene, men er oftest fordelt ned mot bunnen, gjerne i svært tette konsentrasjoner (Hop og Gjøsæter 2013). Den lever av planktonorganismer og når den blir større, beiter den også på fiskeyngel. Området inneholder viktige gyteområder for polartorsk (figur 8.3.1), og beiteområder for stedegne og vandrende fiskearter. Polartorsk gyter om vinteren i fjorder og øst for Svalbard, mens egg og larver driver med klokken rundt Svalbard om våren og sommeren (Eriksen mfl. 2019).
For de kommersielt viktige fiskeartene er havområdene rundt Svalbard først og fremst et beiteområde, og i noen grad også et oppvekstområde, med for eksempel god fødetilgang i lodde (Mallotus villosus) og reker (Jakobsen og Ozhigin 2011, Eriksen mfl. 2017). Akustiske observasjoner fra dette området viser at tilbakespredning fra fisk er mye høyere enn på sokkelen nord fra Svalbard (Menze mfl. 2020). Lodda er en viktig brikke i økosystemet. Den omsetter store mengder planktonføde og er selv føde for mange arter, både fisk, sel, hval og sjøfugl, i tillegg til andre byttedyr, som dypvannsreker (Pandalus borealis). Lodda sitt sommerbeiteområde (figur 8.3.1.) er et viktig område for moden lodde hvor den beiter på Calanus og krill og danner matgrunnlag for både fisk, fugl og sjøpattedyr om sommeren (Gjøsæter 1998, Gjøsæter mfl. 2009 og 2011). Utbredelsen av sommerbeitet varierer mellom kalde og varme år og er nær knyttet til bestandsstørrelsen (når bestanden blir stor, utvides grensene mot øst og nord, Loeng og Ottersen 2011). Torsken (Gadus morhua) er særlig avhengig av lodde for å vokse godt, og av det totale årlige konsumet, som i senere år har vært beregnet til mellom 5 og 6 millioner tonn, kan lodda utgjøre omtrent halvparten. Torsken følger etter lodde på beitevandring og området (F1, figur 8.3.1) er et viktig beiteområde for torsk i løpet av sommer/høst (Loeng og Ottersen 2011, Townhill 2020). Hyse (Melanogrammus aeglefinus) og blåkveite (Reinhardtius hippoglossoides) beiter i området om sommeren og høsten. I tillegg finnes det en god del ikke-kommersielle arter (for eksempel ringbuker, enkelte ålebrosmer og ulker) som har hele livssyklusen i dette området (Wienerroither mfl. 2011, Bergstad mfl. 2017).
Det foreslått endrete SVO Havområdene rundt Svalbard er et yngelområde for flere kommersielle og økologisk viktige fiskearter, blant annet snabeluer (Sebastes mentella) og den sterkt truede (EN) vanlig uer (Sebastes norvegicus) (Drevetnyak og Nedreaas 2009, Jakobsen og Ozhigin 2011, Albert og Vollen 2015, Eriksen mfl. 2020). Blåkveitas hovedgyting foregår på dypt vann (500–800 m) om høsten og vinteren på eggakanten mellom 70 og 75°N (Albert mfl. 2001), og omtales for foreslått SVO Eggakanten nord. Snabeluer føder levende yngel i mars–april, mens vanlig uer føder levende yngel i april–mai langs eggakanten lenger sør, og atlanterhavsstrømmen sørger for at larver og yngel av disse artene blir transportert nordover og inn i Barentshavet hvor de beiter på plankton. Egg og larver driver med strømmen avhengig av hvor gytingen foregår. De siste ti årene er hoveddelen av blåkveiteegg og -larver blitt ført nordover langs Svalbard og østover mot Frans Josef land, inn i oppvekstområdet i det nordlige Barentshavet. Området er et også yngelområde for torsk, hyse, sild og lodde (Eriksen mfl. 2020). Disse artene gyter langs norskekysten fra Mørebankene (sild) og Lofoten (torsk, hyse) til Finnmarkskysten (lodde). Egg og larver driver med strømmen avhengig av styrken og retningen på strømmen. Området vest av Svalbard har noe mindre betydning for larvetransporten enn områdene sør og øst for Svalbard og ellers inne i Barentshavet (ICES 2020 a). Fiskeyngel (torsk, hyse, sild) blir spist av for eksempel torsk, grønlandssel og vågehval.
Med hensyn til biodiversitet er det i tillegg et mangfold av ikke-kommersielle fiskearter innenfor området (Wenneck mfl. 2005).
Bunnsamfunn
Dypvannsreke er utbredt i sentrale deler av Barentshavet og rundt Svalbard (NAFO, ICES 2020 a). Reken i Barentshavet og ved Svalbard utgjør en egen genetisk bestand, helt ulik rekene langs norskekysten (Jorde mfl. 2015). Bestanden er i god forfatning og har siden 1990-tallet fluktuert uten trend (NAFO, ICES 2020 a). Rekene ved Svalbard vokser langsomt, og kjønnsskifte fra hann til hunn skjer ved en alder av 6-8 år (Hansen og Aschan 2000). Utbredelsen av bestanden i Barentshavet har de siste årene flyttet seg mot nordøst, og det har blitt mindre reke på de tradisjonelle fiskefeltene i vest. Landingene har økt de siste tre årene, både fordi Russland igjen har begynt å fiske reker og fordi tredjeland lander mer enn tidligere (NAFO, ICES 2020 a). Tredjeland har bare mulighet til å fiske reker i Svalbardsonen og i Smutthullet. Se også omtalen av dypvannsreker under foreslått SVO Det sentrale Barentshavet (Kap. 8.9.5).
Kalkalger som vokser løstliggende på bunnen (dvs. rhodolitter, mergel-bunner), i fjorder og kiler har status utilstrekkelig data (DD) i rødlisten (Arneberg og Jelmert, 2017). Ny kunnskap om rhodolittene har vist at denne samfunnstypen bidrar til økt biologisk mangfold og har en viktig økologisk funksjon på sokkelområdene ved Svalbard. Det finnes to typer, en kompakt og en hul som følge av borende muslinger (Teichert 2014). Det er særlig den siste varianten som bidrar til økt biologisk mangfold ved at den etter hvert huser et rikt dyreliv, i tillegg til et bredt spekter av bentiske mikroorganismer. Rhodolitter er også viktig for lagring av karbon. Tredjeland har bare mulighet til å fiske reker i Svalbardsonen og i Smutthullet. Kalkalger som vokser løstliggende på bunnen (dvs. rhodolitter, mergel-bunner), i fjorder og kiler har status utilstrekkelig data (DD) i rødlisten (Arneberg og Jelmert, 2017). Ny kunnskap om rhodolittene har vist at denne samfunnstypen bidrar til økt biologisk mangfold og har en viktig økologisk funksjon på sokkelområdene ved Svalbard. Det finnes to typer, en kompakt og en hul som følge av borende muslinger (Teichert 2014). Det er særlig den siste varianten som bidrar til økt biologisk mangfold ved at den etter hvert huser et rikt dyreliv, i tillegg til et bredt spekter av bentiske mikroorganismer. Rhodolitter er også viktig for lagring av karbon.
På det arktiske Yermakplatået finnes et sjeldent og unikt, gammelt (opptil 70 år), bunnsamfunn med både dyphavsarter og arktiske arter. Denne miksen av arter er en konsekvens av møtet mellom dyphavet og arktiske forhold. Nede i skråningen utenfor Norges vestkyst finnes en rekke arter som vi gjenfinner på Yermkplatået (for eksempel Umbellula encrinus, se Mareano.no). Deler av den varme Atlanterhavsstrømmen som går langs skråningen utenfor Norges vestkyst blir presset over Yermakplatået (Menze mfl. 2019). Dessuten ligger Yermakplatået under is mesteparten av året (Onarheim mfl. 2014) og fungerer slik som habitat for arktiske arter. Dette spesielle miljøet gjør at vi finner dyphavs arter grunnere på Yermakplatået (300-500 m) sammenliknet med dypere på kontinentalskråningen langs vestkysten (se Mareano.no). Bunndyrsamfunnet på Yermakplatået består bl.a. av sjøfjæren Umbellula encrinus (står oppreist opp til 2,5 m over havbunnen og blir 70 år gamle; de Moura Neves mfl. 2018), den arktiske reka Bythocaris sp, de største amfipodene i Barentshavet Eurythenes gryllus og Amathillopsis spinigera, og den bunndyrspisende hvite blekkspruten Cirroteuthis moelleri som kan bli 150 cm lang (Jørgensen mfl. in prep). C. moelleri, med relativ ukjent biologi, blir mest sannsynlig gammel, har langsom vekst og lav fekunditet (https://www.iucnredlist.org/). Vi vet lite om disse store og gamle arktiske artene.
I de relativt kalde (Helland-Hansen og Nansen, 1909, Loeng, 1991) vannmassene på østsiden av Svalbard finnes den høyeste megafauna biodiversitet i Barentshavet (figur 8.3.1). Dette kan delvis forklares med møtet mellom to vannmasser (atlantisk og arktisk, Loeng 1991) som gir et miljø egnet for både arktiske og atlantiske (boreale) arter, men også det sesongpåvirkede miljøet som veksler mellom isdekke og isfritt og som dermed gir anledning både for arter tilpasset til å plukke opp partikler fra havbunnen etter for eksempel isalger som faller fra havisen, og for arter tilpasset å filtrere Atlanterhavsstrømmen som passerer området for partikler (Lind mfl. 2012). Til sammen utgjør disse strukturelle bunnhabitater som øker biodiversiteten (Jørgensen mfl. 2019). Arter som sjøliljer (Heliometra glacialis), blomkålskoraller (Nephtheidae, strekker seg opp til 25 cm over havbunnen) og medusahoder Gorgonocephalus spp. (opptil 40 cm) karakteriserer dette komplekse samfunnet, og disse artene kan ha en levetid på flere tiår. Slangestjerner kan dekke havbunnen som et teppe, og dette bidrar til å gi biomasseakkumuleringer som øker områdets produksjon (Degen mfl. 2016). På de store bløtbunnsslettene i Hopendypet registrerte Mareano sjøfjær, mens Økotoktet registrerte store mengder av reker. Området utgjør overgangen fra kaldt bunnvann til varmere atlanterhavsvann. Dette reflekteres i en overgang fra arktiske pigghuder som dominerer samfunnene nord for Polarfronten til mer boreale svampedominerte bunnsamfunn sør for Polarfronten (Jørgensen mfl. 2015).
I atlantisk vann i overgangen mellom kontinentalsokkelen og skråningen vest for Svalbard oppstår det sterkere strømmer (Menze mfl. 2019) som ser ut til å være gunstig for svampen Geodia (svampespikelbunn – se Mareano.no). Disse svampe-aggregeringene finnes på sokkelkanten langs vestkysten av Norge, forbi vestkysten av Svalbard og videre østover langs sokkelkanten nord for Karahavet (Jørgensen mfl. 2015 a, b) og viser dermed en affinitet for atlantisk (strømrikt) vann (Bluhm mfl. 2020). Geodia-svampene er blandet med andre typer av høye og komplekse svamper (Axinella, Phakellia, Haliclona) (Jørgensen mfl. 2015 a, b) som bidrar til å oppnå de høyeste artsrikdommene blant megafauna (Jørgensen mfl. 2019).
På den grunne Spitsbergbanken er det relativt sterk strøm langs kanten (Loeng 1991, Lien mfl. 2018) og sterke tidevannsstrømmer (Gjevik mfl. 1994). Filtrerende organismer får derfor en fordel av å kunne filtrerer fødepartikler ut av denne vannstrømmen. Det finnes store aggregeringer av sjøpølsen Cucumaria frondosa (Jørgensen mfl. 2015 a, b) som bruker tentakler til å vifte til seg partikler. Cucumaria frondosa står sammen med buskformet bryozoa (mosdyr) og hydroider på skjellgrus mellom 30 og 50 m dyp (Lien mfl. 2018, Kedra mfl. 2013). I dette området er dyrene konsentrert til øyer av stein hvor de kan holde seg fast i den sterke strømmen. Tilsvarende miljø er kjent fra trange sund på kysten, men langt til havs er slike forhold uvanlig. Dette miljøet og dyresamfunnet utgjør en særegen biotop, “brunpølsebunn”, som tidligere ikke har vært beskrevet offshore i Barentshavet. Utenom Spitsbergenbanken, Sentralbanken og på banker i sørøstlige Barentshavet (russisk område) har vi grunn til å tro at «Brunpølsebunn» utgjør et særegent habitat på banker og grunne områder i Barentshavet (Jørgensen pers medd.).
Sjøpattedyr
Dette høyproduktive området rundt Svalbard er også attraktivt habitat for et betydelige antall hval. God korrelasjon mellom finnhval (Balaenoptera physalus), knølhval (Megaptera novaeangliae), vågehval (Balaenoptera acutorostrata) og store mengder krill er observert i områder øst fra Hinlopenstredet (Ressler mfl. 2015). Andre undersøkelser viser også store mengder av finnhval og blåhval (Balaenoptera musculus) i dette området (Ingvaldsen mfl. 2017). Her følger presentasjoner av marine pattedyr i området.
Hvithval (Delphinapterus leucas) i den norske sektoren av Barentsregionen finnes generelt bare i farvannene rundt Svalbard hvor de okkuperer en unik nisje blant hvalene ved at de konsentrerer seg svært nær kysten (Lydersen mfl. 2001, Storrie mfl. 2018, Vacquie-Garcia mfl. 2018). De tilbringer mesteparten av tiden foran brefronter eller på vandring mellom slike i periodene hvor det ikke er sjøis i fjordene. Sporingsstudier viser at de generelt tilbringer mer tid på østkysten om høsten og vinteren og mer tid på vestkysten om sommeren (Vacquie-Garcia mfl. 2018). Sent på høsten blir de mange steder presset ut fra kysten på grunn av at isen legger seg, og de finnes da i drivisen ofte i områder med mer enn 90 prosent isdekke (Lydersen mfl. 2002, Vacquie-Garcia mfl. 2018). De fleste observasjonene av mødre med nyfødte kalver er gjort nord i Storfjorden, som bør betraktes som et yngleområde for arten på Svalbard. Den første tellingen av denne bestanden ble utført i 2018 og resulterte i et estimat på 549 dyr (95 % konfidensintervall 436-723) (Vacquie-Garcia mfl. 2020), noe som gjør svalbardbestanden til en av de minste av verdens 20 hvithvalbestander (Hobbs mfl. 2019). Hvithvalene på Svalbard er genetisk forskjellige fra hvithvalene på Vest-Grønland (O’Corry-Crowe mfl. 2010), mens deres genetiske slektskap med hvithvalene på Frans Josef land er ukjent. De er følsomme for klimaendringer som fører til mindre is og endrete beiteforhold (se Kap. 8.2.2).
Grønlandshval (Balaena mysticetus). I et historisk perspektiv var grønlandshvalene vanlig i fjordene på Svalbard, men disse ble raskt fangstet bort etter at Svalbard ble oppdaget. I dag kommer en og annen grønlandshval inn i disse områdene om somrene, særlig i år hvor iskanten i nord ligger ned mot øygruppen, og de betraktes som sjeldne i åpne farvann (Storrie mfl. 2018). Økt skipstrafikk og generell undervannsstøy antas å være en trussel for disse svært vokale hvalene (Ahonen mfl. 2017).
Narhval (Monodon monoceros). Som for grønlandshval er narhval sjelden å se i kystnære farvann rundt Svalbard (Storrie mfl. 2018). I motsetning til grønlandshval har aldri narhval vært vanlig nær Svalbard, da ville de vært mer dominerende i gamle jaktstatistikker (Gjertz 1991). Det finnes registreringer fra mange fjorder over hele øygruppen hvor da de fleste er sett i fjordene på Nordaustlandet (Storrie mfl. 2018). Den skjulte atferden i isen og lite innsats for å studere disse hvalene kan lett gjøre at vi undervurderer antallet narhval i kystnære strøk på Svalbard. Det eneste sporingsstudiet som er utført på øygruppen var på tre unge individer som ble fanget i Wahlenbergfjorden og disse svømte kystnært og tilbrakte perioder dykkende over en dyp kløft nordøst for Nordaustlandet (Lydersen mfl. 2007). Narhval er vurdert å være en av de arktiske marine pattedyrartene som er mest sensitive i forhold til klimaendringer (se Kap. 8.2.2).
Hvalross. Foto: Kit Kovacs og Christian Lydersen, NP
Bardehval. Området rundt Svalbard er kjerneområdet for sommerbeitet til de vandrende bardehvalartene (Skern-Mauritzen mfl. 2011). Disse omfatter artene vågehval (Balaenoptera acutorostrata), seihval (Balaenoptera borealis), finnhval (Balaenoptera physalus), blåhval (Balaenoptera musculus) og knølhval (Megaptera novaeangliae). Vågehval er vidt utbredt i hele Nord-Atlanteren; seihval er sporadisk forekommende. Blåhval har særlig tilhold vest og nord for Spitsbergen, knølhval i området Bjørnøya-Hopen (Spitsbergenbanken) og videre nordover mot Olgastredet, og finnhval er særlig knyttet til kontinentalskråningen mot Norskehavet/Grønlandshavet, men har i de senere årene hatt en vidstrakt utbredelse vest av Spitsbergen og helt til iskanten.
Knølhval og vågehval ble observert i loddeområdene under økosystemtoktene som tyder på at det er et viktig beiteområde (Klepikofsky og Øien, 2017).
Kvitnos (Lagenorhynchus albirostris). Kvitnos er en tallrik stedegen art som er vidt utbredt i det sørlige Barentshavet, og sokkelområdet vest av Spitsbergen er artens nordligste beiteområde (Fall og Skern-Mauritzen 2014).
Spermhval (Physeter macrocephalus) har sin sentrale utbredelse i Norskehavet utenfor sokkelkanten vest av Nord-Norge (Bleik-djupet er en ‘hotspot’) (Øien 2009), men har i de seinere årene opptrådt langs sokkelkanten lenger nordover med de nordligste observasjonene nord for Spitsbergen. Den observeres også sporadisk inne i Barentshavet selv om den i utgangspunktet er en dyphavsart.
Hvalross (Odobenus rosmarus). Historisk sett var hvalross svært tallrike på Svalbard, men de ble fangstet slik at de nesten ble utryddet. Antakelig er bestanden gradvis bygd opp med basis i dyr fra Frans Josef land som også er en del av denne bestandens utbredelsesområde. Hvalrossene fra Svalbard overvintrer inne i drivisen og bruker land som hvilested om somrene når sjøisen over furasjeringsområdene blir borte. I løpet av de siste tiårene har bestanden tatt seg kraftig opp, de tar igjen gamle liggeplasser som ikke har vært brukt på lang tid og bestandsstrukturen er i ferd med å normaliseres (Lydersen mfl. 2008, Kovacs mfl. 2014, www.mosj.no/no/fauna/hav/hvalross-bestand.html). De fleste hunnene med kalver finnes på østsiden av Svalbard, hovedsakelig nord på Nordaustlandet og øst for denne øya. Hvalrossene på Svalbard synes å reagere lite på turisttrafikk som til nå ikke er påvist å ha noen negativ betydning (Øren mfl. 2018). Den største trusselen til denne bestanden i forhold til klimaendringer er redusert tilgjengelig vinter (yngle) habitat og endringer i bunndyrfaunaen de ernærer seg fra.
Ringsel (Pusa hispida). Fjordene på Svalbard er det viktigste ynglehabitatet for ringselene (Lydersen 1998). De fleste voksne dyrene oppholder seg i områder inne i fjordene hvor vinterisen legger seg nær brefronter og disse individene holder seg her også resten av året (Freitas mfl. 2009, Hamilton mfl. 2017). Om våren finnes dominerende kjønnsmodne dyr innerst i fjordene der isen er mest stabil, mens unge individer og gamle hanner presses ut mot iskanten hvor forholdene er mer ustabile (Krafft mfl. 2006). Etter hårfellingsperioden vandrer de fleste unge dyrene opp til drivisen nord for Svalbard hvor de jakter gjennom sommeren (Freitas mfl. 2008, Hamilton mfl. 2015b) mens de voksne individene forblir nær kysten (Hamilton mfl. 2017). Etter de drastiske endringene i isforholdene som startet i 2006, har atferden til ringselene på Svalbard endret seg en god del (Kap. 8.2.2). Ringselene viser noen grad av atferdsmessig plastisitet med å legge seg opp og hvile på land enkelte steder om sommeren i stedet for alltid på is (Lydersen mfl. 2017). Bestandsstørrelse på Svalbard har ikke blitt estimert siden 2002/2003 da man fant rundt 10 000 dyr på isen under hårfellingsperioden i fjordene på Spitsbergen (Krafft mfl. 2006). Isforholdene de siste årene har vært så dårlige at flytellinger i denne perioden ikke er noen vits i å utføre. Man antar at antallet ringsel i Barentsområdet er på vei nedover, gitt de store endringene som har skjedd i kastehabitat (lite is og ofte ikke snø til huler som fører til stor ungedødelighet) og furasjeringshabitat (Hamilton mfl. 2019b). Tendensen disse selene har til å oppholde seg i små arktiske refugier (brefronter) gjør at de er svært sårbare i forhold til klimaendringer (Kovacs mfl. 2011 og 2012, Gilg mfl. 2012).
Storkobbene (Erignathus barbatus) yngler langs kysten og inne i fjordene på Svalbard, i tillegg til ute i drivisen i Barentsregionen (Kovacs 2018). Storkobbene inne i fjordene på Svalbard er svært stedfaste, noe som gjelder for alle aldersgrupper, bortsett fra en spredning av årsunger i en periode etter at de er avvendt (Hammill mfl. 1994, Gjertz mfl. 2000, Hamilton mfl. 2019b og 2019c). De har i tillegg et bemerkelsesverdig individuelt furasjeringsmønster hvor noen spiser foran brefronter, andre inne i fjordene og andre igjen foretar kortere turer ut på sokkelen offshore for så å returnere inn i fjordene igjen for å hvile seg på is (ofte bre-is) (Hamilton mfl. 2018). Vi vet lite om økologien til storkobbene som oppholder seg ute i drivisen i Barentsregionen, bortsett fra at de voksne hannenes typiske sanger i parringstiden høres ute i isen i Framstredet og også nord for Svalbard nær kontinentalskråningen (Moore mfl. 2012, Ahonen mfl. 2017, Martínez Llobet mfl. innsendt), noe som indikerer at yngling foregår i disse områdene. Det antas at tettheten av storkobber rundt Svalbard er relativt høy sammenliknet med andre steder i Arktis, siden hvalrossene her nesten ble utryddet, noe som reduserer potensiell konkurranse om bentiske byttedyr (Weslawski mfl. 2000). Det finnes ingen estimater for størrelsen på bestanden av storkobber i dette området. De forekommer generelt i lav tetthet og tilbringer mye av tiden i vannet (opptil 95 prosent) ofte i områder med mye is, noe som gjør det utfordrende å utføre tellinger på denne arten. Etter at sjøisforholdene i fjordene på Svalbard endret seg kraftig i 2006 har storkobbene skiftet fra å bruke sjøis til biter av breis som flyter rundt i fjordene, for å kaste på og også for diing av ungene (Kovacs mfl. 2020b). De har aldri blitt påvist å kaste på land, og breene som produserer disse «nye» kasteplattformene smelter og trekker seg etter hvert inn på land, så bruk av disse er ikke en langvarig fremtidig løsning. Selv om denne arten synes å være mer tilpasningsdyktig enn ringsel, så er fortsatt global oppvarming en alvorlig trussel for storkobbene og den vil sannsynligvis redusere artens utbredelsesområde i denne regionen (Kovacs mfl. 2011 og 2012).
Steinkobbene (Phoca vitulina) på Svalbard er en egen genetisk bestand og er mest beslektet med den lille truete bestanden av arten som finnes på Grønland (Andersen mfl. 2011). Hoved-utbredelsesområdet er langs vestkysten av Svalbard og de jakter lokalt, hovedsakelig i atlantiske vannmasser nær kontinentalskråningen og i de senere år også mer og mer inne i fjordene (Blanchet mfl. 2014 og 2015). Som for andre steinkobbebestander foretrekker de å hvile på land og ikke på is, men bruker is som hvileplattform om vinteren når liggeplassene på land er iset ned (Hamilton mfl. 2014). Mesteparten av ynglingen foregår på vestsiden av Prins Karls Forland (Reder mfl. 2003). Steinkobbene på Svalbard er på den nasjonale rødlisten fordi bestanden er så liten (ca. 2000 dyr, Merkel mfl. 2013) og isolert. Steinkobbene synes å respondere positivt på de pågående reduksjonene i isdekke og økning av atlantiske byttedyr lenger nord i Barentshavet, så de øker i antall og utvider sitt utbredelsesområde og vil antakelig måtte betraktes som «klimaendrings-vinnere».
Isbjørn (Ursus maritimus). Opp mot rundt 300 isbjørn lever i Svalbardområdet året rundt (Aars mfl. 2017). Isbjørn som det meste av året lever i iskantsonen, kan også oppholde seg i Svalbardområdet i perioder; en andel av binnene fra disse områdene kommer til Svalbard på høsten for å gå i hi (og føde unger). Isbjørn på Svalbard tar mest ringsel, og mye ringselunger, på våren (Iversen mfl. 2013). Seinere utover sommeren er storkobber minst like viktige byttedyr som ringsel (Derocher mfl. 2002). Mødre med små unger jakter foran brefronter tidlig på våren, mens andre isbjørn i større grad bruker områder lengre fra land (Freitas mfl. 2012). I senere år har brefrontene blitt mindre viktige jaktområder for isbjørn utpå sommeren, dette fordi det er mindre sjøis foran disse brefrontene enn det var for en del år tilbake (Hamilton mfl. 2017). Siden en stor del av isbjørnbestanden følger iskanten, som nå oftest er lenger nord enn før, er sjøisområdene rundt øyene benyttet av en stadig lavere andel av bestanden (Lone mfl. 2018). På Svalbard er isbjørn lokale innen ulike områder på øygruppa både over tid (Lone mfl. 2013) og over generasjoner (Zeyl mfl. 2009).
Ismåkeprosjektet. Foto: H. Strøm, NP
Sjøfugl
Havområdene rundt Svalbard er svært viktige for sjøfugl. For flere av artene som hekker på Svalbard, utgjør bestanden her mer enn 25 prosent av europeisk bestand, f.eks. havhest (Fulmarus glacialis), alkekonge (Alle alle), polarmåke (Larus hyperboreus) og polarlomvi (Uria lomvia) (Henriksen og Hilmo 2015). Hekkebestandene i øygruppen omfatter en rekke arter av arktisk og subarktisk opprinnelse. De arktiske artene domineres antallsmessig av polarlomvi og alkekonge. I tillegg finnes arter som polarmåke, sabinemåke (Xema sabini), rosenmåke (Rhodostethia rosea) og ismåke (Pagophila eburnea) (Strøm mfl. 2020), alle relativt fåtallige. Store deler av storjobestanden i Norge hekker på Svalbard, med den største kolonien på Bjørnøya. Lomvi har en mer sørlig utbredelse i Nord-Atlanteren enn polarlomvi, men den største lomvikolonien i Norge finnes også på Bjørnøya. Etter en kraftig tilbakegang i koloniene på fastlandet, hekker nå størstedelen av krykkjene (Rissa tridactyla) i Norge på Svalbard. Havhest er også tallrik på de arktiske øyene, og svært fåtallige på fastlandet.
De største sjøfuglkoloniene på Svalbard ligger på Bjørnøya, Hopen, i Storfjorden og på vestkysten av Spitsbergen. Sjøfuglkoloniene på Svalbard domineres av polarlomvi, krykkje, havhest, polarmåke, og mindre forekomster av lunde (Fratercula arctica) og teist (Cepphus grylle). I tillegg finnes meget store kolonier av alkekonge, særlig i Hornsund-området og på Nordvest-Spitsbergen. Lomvikolonien på Bjørnøya (>100 000 par i 2020) er den største i Norge. Alkekongen er trolig Svalbards mest tallrike fugleart, og den er den eneste av alkefuglene som primært lever av dyreplankton. Flere av de kolonihekkende artene (bl.a. alkefuglene) har stor aksjonsradius i hekkeperioden, og store deler av havområdet innenfor en radius av 100 km fra kolonien må regnes som svært viktige næringsområder. Det største fuglefjellet på Spitsbergen er Koval’skijfjella/Stellingfjellet i Storfjorden, der det primært hekker polarlomvi. I Hornsund finnes store hekkekolonier for alkekonge. På vestkysten er det ellers særlig Bellsund med koloniene på Midterhuken og i Ingeborgfjellet, samt Fuglehuken på Prins Karls Forland som peker seg ut som særlig viktige hekkeområder for artene.
Krykkjebestandene på Bjørnøya og Spitsbergen har holdt seg stabile eller vist en svak positiv endring, selv om dietten deres har endret seg på noen lokaliteter (Arneberg og Jelmert 2017, Vihtakari mfl. 2017).
Nedgangen i hekkebestanden til polarlomvi i området har fortsatt siden 2009 (Arneberg og Jelmert 2017). På Svalbard (som også inkluderer Bjørnøya) er det observert bestandsreduksjoner på 25-50 prosent siden slutten av 1990-tallet. Hvis denne raten ikke endrer seg, har polarlomvi på Svalbard en høy sannsynlighet for å gå mot kvasi-ekstinksjon (så lavt nivå at de muligens ikke er i stand til å komme seg) innen de neste 50 årene.
Endring av sjøis og arktisk planktonfauna tilknyttet isen vil kunne påvirke sjøfugl som er avhengige av arktisk planktonfauna knyttet til iskantsonen, særlig arter som alkekonge og krykkje (Amélineau mfl. 2019).
Figur 8.3.4. Figuren viser hekkekolonier for krykkje (turkis), polarlomvi (lysegul) og havhest (lysegrønn) i foreslått SVO Havområdene rundt Svalbard. Grensene for det foreslåtte SVO-et er markert med rød strek.
Hekkebestanden til lomvi på Bjørnøya har vokst siden 2009 (Arneberg og Jelmert 2017). Havhest (antagelig stabil) og alkekonge (muligens i nedgang) er også tallrike arter på Svalbard (S. Descamps, pers. medd.). Studier fra Frans Josef land tyder også på at brefronter blir brukt i økende grad av alkekonge som en respons på mindre sjøis (Grémillet mfl. 2015). Havsule har spredd seg nord til Bjørnøya og i 2011 ble to reir observert der (Anker-Nilssen mfl. 2020). I dag er det 75 reir i kolonien (H. Strøm, pers. med.). Ringgås hekker ikke i samme grad som tidligere på Tusenøyane grunnet predasjon fra blant annet fjellrev. Foreløpig er det imidlertid ikke grunnlag for å avskrive dette som et hekkeområde for ringgås.
I 2015 ble det startet årlig overvåking i en av de største polarlomvi- og krykkjekoloniene på Svalbard (Alkefjellet i Hinlopenstredet), som har ca. 70 000 par polarlomvi og minimum 5000 par krykkjer (Anker-Nilssen mfl. 2020). Samlet sett antas flere hundre tusen sjøfugl å hekke i dette området, noe som gir dette området høy verdi. Det ble også etablert overvåkingsfelt for havhest og polarmåke på samme lokalitet. I tillegg har noen av polarlomviene, krykkjene og havhestene fått påsatt lysloggere for å kartlegge vinterområdene. Til tross for kort tidsserie er det indikasjoner på at polarlomviene fra Alkefjellet har ganske ulik vinterutbredelse sammenlignet med polarlomvi fra Vest-Svalbard og Bjørnøya, dvs. har tilhold i Barentshavet hele året. Dette kan muligens også forklare hvorfor det kan se ut som om polarlomvi ikke minker på samme måte i øst som i vest (og på Bjørnøya) hvor fuglene har andre overvintringsområder.
Spitsbergenbanken og Hopenbanken mellom Bjørnøya og Hopen er et område som i økende grad er isfritt også vinterstid, noe som endrer områdets betydning for overvintrende sjøfugl. Både havelle, ærfugl og praktærfugl dykker ned til bunnen og beiter på organismer der, siden bankene er nokså grunne. Området vurderes som særlig verdifullt siden det er et nøkkelområde for overvintring av praktærfugl, muligens for hele Svalbard- og Øst-Grønland-populasjonen (H. Strøm upubl.).
8.3.6 - Pågående og framtidige endringer
Siden dette området allerede har vist seg følsomt og allerede påvirket av klimaendringene, legges det til ytterligere informasjon om dette her. Framtidsscenarioer antyder at overflatetemperaturene i dette området vil øke med 1-1,5 °C frem mot 2060-2069 (Hanssen-Bauer mfl. 2018). Det forventes økt havforsuring på grunn av fortsatt opptak av atmosfærisk CO2 i perioden 2000-2065 (Skogen mfl. 2014). Økt ferskvann fra breissmelting gir økt havforsuring og kan gi kritiske nivåer av aragonittmetning for kalsifiserende marine organismer (Fransson mfl. 2015 og 2016). Modellresultater forventer en undermetning av aragonitt i overflaten i år 2100 og minket pH ned til 7,7 (RCP8.5 scenario; Lauvset mfl. 2016, Fransner mfl. 2020), hvilket kan gi alvorlige konsekvenser på marine organismer.
Flere fjorder på vestsiden av Svalbard som tidligere var dekket av fjordis, har i liten grad vært isdekket siden 2006, med unntak av enkelte terskelfjorder som i noen grad er batymetrisk beskyttet fra vind og bølgeaktiviteter, f.eks. Billefjorden og van Mijenfjorden (Gerland og Hall 2006). Med den pågående oppvarmingen er det sannsynlig at både meteorologiske og kryosfæriske forhold på østsiden av Svalbard vil ligne de som allerede observeres i fjorder på vestsiden (hvor endringene også vil akselerere), dvs. mindre typisk arktisk klima noe som vil påvirke forekomst av arter (mengde og type) og biologiske prosesser (Dahlke mfl. 2020). I tillegg vil prosesser som i dag påvirkes av brefronter som ender i fjordene, endres når brefrontene trekker seg tilbake og blir liggende på land (Vonnahme mfl. 2021). Se ellers SVO Iskantsonen.
Ytterliger isreduksjon i området vil ha dramatiske konsekvenser for polartorskens gyting ved Svalbard og dermed bestandsutvikling (Eriksen mfl. 2019, Husebråten mfl. 2019, Gjøsæter mfl. 2020). Klimaendringer kommer til å påvirke flere faktorer som er viktig for produksjon av dyreplankton rundt Svalbard, inkludert redusert utbredelse og endret timing av landfast is og sjøis (Søreide mfl. 2010, Leu mfl. 2011), økt tilførsel av smeltevann fra breer og permafrost i fjorder (McGovern mfl. 2020, Hopwood mfl. 2020). Endringer i dyreplanktonsammensetningen som allerede har blitt påvist, kommer sannsynligvis til å fortsette (dvs. flere atlantiske arter) og etter hvert inneholde færre arktiske arter (Hop mfl. 2019 a, ICES2020 a, Carstensen mfl. 2019). Dette kan påvirke beiteforholdene for høyere trofiske nivåer (Kwasniewski mfl. 2012). Byttedyrene til de marine pattedyrene vil sannsynligvis bli utbredt på en mindre forutsigbar romlig måte og ha mindre lipidinnhold enn de tradisjonelle arktiske byttedyrene. Dette gjør at det må brukes mer energi for å finne maten og at denne har mindre energiinnhold. Det samme gjelder for sjøfugl. Det forventes en økning og innvandring av mer sørlige sjøfuglarter på Svalbard, en prosess som allerede er i gang. Storjo er etablert på øygruppen, havsule hekker nå på Bjørnøya og kan forventes å etablere seg også lenger nord (www.seapop.no). Lunde og lomvi finnes på Svalbard allerede, og kan øke i antall dersom klimaendringene gjør dette området bedre egnet for disse artene. Nordlige, arktiske arter kan ha større problemer med endringene. Sjøfugl som er avhengige av arktisk planktonfauna knyttet til iskantsonen, særlig arter som alkekonge og krykkje (se bl.a. Amélineau mfl. 2019, von Quillfeldt mfl. 2018) er utsatte dersom isutbredelsen minker både når det gjelder fastis og brefronter.
Fysiske og biotiske endringer i det marine miljøet vil påvirke de marine pattedyrbestandene direkte eller indirekte og antakelig kumulativt med økte menneskelige aktiviteter (skipstrafikk, turisme, industriell virksomhet, fiskeri etc.) i områder som tidligere var beskyttet på grunn av et utstrakt isdekke (f.eks. Pizzolato mfl. 2016, Dawson mfl. 2018). I tillegg, ettersom klimaet blir varmere, er arktiske marine pattedyr mer utsatt for miljøgifter og effekter av disse (Sonne mfl. 2017) samt økt risiko for eksponering for nye sykdommer (Van Wormer mfl. 2019).
8.4 - Forslag til endret SVO Iskantsonen (BH2)
8.4.1 - Sammendrag
Områdebeskrivelse og strukturer
Forslag til endret SVO Iskantsonen (BH2) er et relativt stort areal, som er valgt for å representere sonen som defineres av vinter- og sommerutbredelsen av isdekket hav. Dette er en overgangssone mellom isfritt og isdekket hav, hvor iskonsentrasjonen er mellom 15 og 80 prosent. Den faktiske iskantsonen er svært dynamisk og beveger seg fra Bjørnøya i sør til nord for Spitsbergen, avhengig av årstid, med mest is øst for Spitsbergen. Foreslått endret SVO Iskantsonen er som en statisk geografisk sone, ikke alltid identisk med den faktiske iskantsonen. I tillegg til sesongmessige variasjoner, forekommer mer kortvarige variasjoner, for eksempel forårsaket av vindretning og -styrke. Maksimal sørlig utbredelse av iskantsonen i Barentshavet styres i stor grad av polarfrontens beliggenhet, mens avsmelting om sommeren avhenger av blant annet lufttemperatur, temperaturen på underliggende vannmasser, mengde av snø på havisen og vind.
Miljøverdier
Produksjonsforhold, forekomst av arter, sårbarhet for ulike typer av påvirkning og hvordan dette varierer gjennom året og mellom år har betydning for i hvilken grad iskantsonen er miljømessig verdifull og sårbar. Imidlertid vil hele iskantsonen alltid være viktig for flere arter og/eller biologiske prosesser uavhengig av årstid.
I likhet med foreslått endret SVO Havområdene rundt Svalbard (Kap. 8.3), SVO Havis Framstredet (Kap. 9.3) og SVO Vesterisen (Kap. 9.4) gjør tilstedeværelse av havis dette til et unikt leveområde. Isalger er tilpasset lite lys og primærproduksjonen i isen starter tidligere enn i vannmassene, noe som bidrar til å forlenge den produktive sesongen i området. Det er et stort artsmangfold i området, inkludert bunnsamfunn med stor biodiversitet av megafauna. Det fysiske og kjemiske miljøet ligger til rette for planteplanktonproduksjon i iskantsonen, der issmelting fører til vertikal stabilitet og bedre lysforhold. Planteplanktonoppblomstringen vil følge isen når den trekker seg nordover. Primærproduksjonen i iskantsonen følges av dyreplankton, fisk, sjøfugl og sjøpattedyr som beiter seg nordover, i tillegg til at en stor del av produksjonen sedimenterer ut av vannsøylen og kommer bunnlevende organismer til gode. For de kommersielt viktige fiskeartene i Barentshavet er iskantsonen først og fremst et beiteområde, og i noen grad også et oppvekstområde. Med unntak av to pelagiske arter (polartorsk og istorsk (Arctogadus glacialis), er de fleste fiskeartene i iskantsonen i Barentshavet sterkt tilknyttet havbunnen. Av sjøfuglene kan særlig polarlomvi og alkekonge forekomme i store konsentrasjoner i iskantsonen og der det er råker om våren, men også teist og ismåke er vanlig. I tillegg observeres havhest, polarmåke og krykkje i iskantsonen gjennom hele året. Flere selarter bruker isen som kaste-, hårfellings- og hvileområde, men betydningen av iskantsonen varier mellom arter og gjennom året. Grønlandshval, hvithval og narhval er de eneste hvalartene som er tilpasset til å være i områder med is hele året. Spitsbergenbestanden av grønlandshval utgjør en egen genetisk enhet forskjellig fra andre bestander. I tillegg beiter andre bardehvaler (blåhval, finnhval, knølhval og vågehval) og tannhvaler (spekkhogger) langs iskanten i sommermånedene.
Det er mange både sårbare, skjøre, sent restituerende og rødlistede fisk-, sjøfugl- og sjøpattedyrarter i området. Naturtypen polar havis er kritisk truet (CR). Dette unike området er dessuten regnet som naturlig i henhold til EBSA-kriteriene, dersom man ser bort fra klimaendringene.
Figur 8.4.1. Kart over forslag til endret SVO Iskantsonen (BH2, skravert område). De foreslåtte grensene for SVO-et er vist med tykk rød strek, mens grensene for de overlappende foreslåtte SVO-ene Havområdene rundt Svalbard (BH1), Eggakanten nord (BH3), Det sentrale Barentshavet (BH7) og Midtatlantisk rygg (NH4), samt andre nærliggende foreslåtte SVO-er, er vist med tynn rød strek. Sørlig grense for området er anbefalt satt ved maksimal isutbredelse i april (0,5 prosent isfrekvens) basert på en tidsserie med satellittobservasjon av isutbredelse for 30-årsperioden 1990-2019.
8.4.2 - Vurdering i forhold til EBSA-kriteriene – Forslag til endret SVO Iskantsonen (BH2)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til endret
SVO Iskantsonen (BH2)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet /sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Issamfunn har en egenverdi i seg selv både mht samfunnstyper (Syvertsen 1991, Bluhm mfl. 2017a) og artssammensetning, samt betydning for produksjon som kanaliseres videre i næringsnettet (Søreide mfl. 2010, 2013, Leu mfl. 2011, Kohlbach mfl. 2016, 2017, Wang mfl. 2016). Betydningen øker med breddegrad (Wassmann mfl. 2006). Noen arter forekommer bare i havis (Bluhm mfl. 2017, Hop mfl. 2020).
Iskantsonen er et viktig leveområde for isamfipoder (f.eks Gammarus wilkitzkii G, Apherusa glacialis) og hoppekrepsen Calanus glacialis. (Arndt and Pavlova 2005, Kohlbach mfl. 2018). Høye konsentrasjoner av egg av C. glacialis er funnet under isen i mai, noe som tyder på at det er viktig gyteområde knyttet til våroppblomstring (Werner og Hirche 2001).
Iskantsonen, som har høy plante- og dyreplanktonproduksjon, utgjør leve- og beiteområdet for store økonomisk og økologisk viktige fiskearter, sjøpattedyr, og sjøfugl (Dolgov mfl. 2011, Hunt mfl. 2013, Fauchald mfl. 2015, ICES 2019, Husebråten mfl. 2019, Stige mfl. 2019, Dalpadado mfl. 2020). Polartorsk og lodde som er nøkkelarter i det nordlige Barentshavet er tilknyttet isfylte farvann (Ajiad mfl. 2011, Boitsov mfl. 2016, Eriksen mfl. 2015, 2019, Huserbråten mfl. 2019, Gjøsæter mfl. 2020, Gjøsæter 1995).
Integrert over tid (antakelig mange tiår) har det etablert seg et tydelig skille mellom et borealt svampedominert samfunn som utnytter partikler som drifter inn i Barentshavet med Atlanterhavsvannet, og et mer arktisk pigghud-dominert samfunn som utnytter organisk materiale som har landet på havbunnen fra den overliggende is, eller driftet inn i området (Jørgensen mfl. 2015, Zakharov mfl. 2020). Oppe på bankene utnytter filtrerende organismer både den tettere kontakten til isdekkets algeproduksjon, men også vannstrømmen. Dette er et unikt iskantsonefenomen som kan endre seg med endrede isforhold og bunntemperaturer. Ingen sesongmessige /tidsmessige variasjoner.
Bløtbunnsamfunnene i iskant-/polarfrontområdene i vestlige del av Barentshavet har vesentlig høyere sekundærproduksjon enn områdene sør og nord for frontområdene, noe som er knyttet til den forholdsvis høye pelagiske primærproduksjonen i disse områdene og til tett bentisk pelagisk kobling på bankene (bl.a. Spitsbergenbanken, Hopenbanken) (Carroll mfl. 2008, Cochrane mfl. 2012, Mareano data under utarbeidelse).
Iskantsonen er det viktigste habitatet for flere truede arter av arktisk endemiske marine pattedyr som isbjørn (Lone mfl. 2018a, grønlandshval (Kovacs mfl. 2020a) og narhval (Vacquie-Garcia mfl. 2017 b).
Iskantsonen er også det viktigste beitehabitatet til ismåke (Gilg mfl. 2010).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Bunnsamfunn
Isbiota
Dyreplankton Fisk Sjøpattedyr Sjøfugl
Svært viktig for arter med hele livssyklusen i isen (Lønne og Gulliksen 1991, Arndt og Swadling 2006, Bluhm mfl. 2017b, Hop mfl. 2020), men mesteparten av isen i iskantsonen er årsis og har derfor arter med bare deler av livssyklus i isen (Barber mfl. 2015). Flere av disse kan også overleve i vannmasser (Berge mfl. 2012) eller på havbunnen (Poltermann 1998).
Iskantsonen er veldig viktig for organismer som lever i og under isen og i tillegg de som er avhengige av iskantproduksjonen for reproduksjonen f.eks. Calanus glacialis (Søreide mfl. 2010, Leu mfl. 2011).
Området er et viktig oppvekstområde for polartorsk (Ajiad mfl. 2011, Boitsov mfl. 2016, Eriksen mfl. 2015, 2019, Huserbråten mfl. 2019, Gjøsæter mfl. 2020), og også for andre populasjoner av både kommersielle og ikke-kommersielle fisk (Eriksen mfl. 2020). Området er viktig som overvintringsområde og i oppstarten av beitinga for unglodde (Gjøsæter 1998, Fall mfl. 2018).
Det kan se ut som det arktiske pigghudsamfunnet kan være knyttet til iskantsonen (Jørgensen mfl. 2015, Zakharov mfl. 2020). Dette gjelder særlig på bankene.
Alle arktisk endemiske marine pattedyr er sterk tilknyttet iskantsonen (Hamilton 2021), og alle (bortsett fra hvithval) bruker den nordlige iskanten i Barentsregionen for furasjering og andre deler av sin livshistorie. Flere arktisk endemiske marine pattedyr yngler i iskantsoner i nordre Barentsregionen; som hvalross (Freitas mfl. 2009, Lowther mfl. 2015) ringsel (Wiig mfl. 1999) og storkobber (Llobet mfl. innsendt).
Iskantsonen er også det viktigste beitehabitatet til ismåke (Gilg mfl. 2010, Amélineau mfl. 2019).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Fisk Bunnsamfunn
Isbiota Dyreplankton Sjøpattedyr Sjøfugl
Isbiota har havis som habitat (Boetius mfl. 2015, Gradinger 2020). Både type og mengde havis i Barentshavet er gjenstand for raske endringer som følge av klimaendringer (von Quillfeldt mfl. 2018) og påvirker dermed forekomst og overlevelse av arter samt biologiske prosesser i isen (Bluhm mfl. 2017b, von Quillfeldt mfl. 2018, Tedesco mfl. 2019, Hop mfl. 2020).
Ikke umiddelbart, men hvis isen forsvinner helt, vil det påvirke artssammensetning, leveområder for is-assosierte pelagiske organismer som f.eks. amfipoder, og de øvrige trofiske nivåer (Arndt og Pavlova 2005, Kohlbach mfl. 2018, Huserbråten mfl. 2019, ICES 2019).
Polartorsk er tilpasset is og arktiske vannmasser og har vist en generell nedadgående trend i Barentshavet de siste 15 år (Ajiad mfl. 2011, Boitsov mfl. 2016, Eriksen mfl. 2015, Gjøsæter mfl. 2020).
I løpet av år 2005 til 2017 har grensen mellom de boreale og arktiske artene av bunnsamfunn flyttet seg 150-200 nm. Oppe på bankene har grensen flyttet seg 150 nm (Zahkarov mfl. 2020).
Iskantsonen er et svært viktig habitat året rundt for flere arktisk endemiske marine pattedyr som blant annet isbjørn (Lone mfl. 2018), grønlandshval (Kovacs mfl. 2020) og narhval (Vacquie-Garcia mfl. 2017b).
Ismåke, krykkje, havhest, alkekonge, teist og polarlomvi beiter alle nær og i iskantsonen om sommeren, flere av disse artene gjennom hele året (bl.a. Amélineau mfl. 2019, von Quillfeldt mfl. 2018).
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Dyreplankton Fisk
Isbiota Bunnsamfunn Sjøpattedyr Sjøfugl
Isbiota, særlig arter i flerårsis vil være utsatt ved klimaendringer. Redusert isutbredelse, sen isdannelse/tidlig smelting gir kortere produktiv sesong for isbiota (Wassmann og Reigstad 2011, Hop mfl. 2013, Barber mfl. 2015). Havis som habitat er også følsomt i forhold til oljesøl fra skipstrafikk (Rusten 2014). Oppvarming av havet kan medføre at isen forsvinner nesten helt. Dette vil ha stor påvirkning på alle organismer som lever i iskantsonen (Kelly 2016).
Området inneholder arktiske fiskearter inkludert polartorsk som er sårbare for klimaendringer, inkludert reduksjon i havis (Bergstad mfl. 2017, Boitsov mfl. 2016, Eriksen mfl. 2015, Huserbråten mfl. 2019).
Overlappet i utstrekning med foreslått SVO Havområdene rundt Svalbard gjør at samme begrunnelse og referanser for miljøverdier av skjøre og saktevoksende bunndyr gjelder også i foreslått SVO Iskantsonen.
Alle arktisk endemiske marine pattedyr er sterk tilknyttet iskantsonen (Hamilton 2021), og alle (bortsett fra hvithval) bruker den nordlige iskanten i Barentsregionen for furasjering og andre deler av sin livshistorie. Dette habitatet i dette området er svært negativt påvirket av pågående klimaendringer (Laidre mfl. 2015).
Sjøfugl som bruker iskantsonen er utsatte for endringer i isutbredelse, særlig i hekkesesongen da de kan erfare økt avstand til optimale beiteområder. Dette gjelder flere rødlistede arter, som for eksempel ismåke, krykkje og alkekonge (bl.a. Gilg mfl. 2010, Amélineau mfl. 2019).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Bunnsamfunn
Isbiota Planteplankton Dyreplankton Fisk
Primærproduksjonen i isen starter tidligere enn i vannmassene noe som bidrar til å forlenge den produktive sesongen i et område (Hegseth 1998, Leu mfl. 2015). Isalger bidrar relativt sett mer til den totale primærproduksjonen jo lenger nord man kommer (Wassmann mfl. 2006, Wassmann og Reigstad 2011). Primærproduksjonen i isen er viktig for pelagiske og bentiske næringsnett (Kohlbach mfl. 2019).
Det fysiske og kjemiske miljøet ligger til rette for forhøyet planteplanktonproduksjon i iskantsonen (von Quillfeldt mfl. 2018).
Produksjonen av dyreplankton er generelt høyest fra våren til sommeren og er knyttet til den sesongmessige planteplanktonproduksjonen i disse områdene (Dalpadado mfl. 2020). Selv om planteplanktonproduksjonen utgjør mye mindre i iskantsonen sammenlignet med det totale Barentshavet (Hegseth 1998, Wassmann mfl. 2006, Hunt mfl. 2013) er det relativt viktig når det gjelder sesongmessig fenologi og reproduksjon av bl.a. arktiske kopepoder (Søreide mfl. 2010, Leu mfl. 2011).
Høye biomasser av 0-gruppe fisk av torsk, hyse, lodde og uer, som er viktig mat for både fisk, sjøfugl og sjøpattedyr i området på sommeren-høsten, sørger for høy produktivitet i området (Eriksen mfl. 2012, Eriksen mfl. 2017, ICES 2017, 2018, 2019, 2020). Mesteparten av populasjonen av lodde som ikke er gyteklar overvintrer nær iskanten og følger isen nordover ved smelting om våren (Gjøsæter 1998).
Bunndyrene utnytter nedfall fra algeproduksjonen under og nær isen (Carroll og Carroll 2003, Ambrose mfl. 2005)
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Fisk Sjøfugl Planteplankton
Isbiota Dyreplankton Bunnsamfunn Sjøpattedyr
Isbiota utgjør en rekke ulike samfunnstyper og består av et høyt antall arter fra en rekke ulike artsgrupper (Syvertsen 1991, Horner mfl. 1992, Gradinger mfl. 2010, Poulin mfl. 2011, Bluhm mfl. 2011, Bluhm mfl. 2017a og b, Hop mfl. 2020).
I tillegg til isalger bidrar pelagiske planteplanktonarter til en høy diversitet (von Quillfeldt mfl. 2018).
Dyreplanktonet bidrar ytterligere til næring for øvrige trofiske nivåer i dette økosystemet (ICES 2019, Dalpadado mfl. 2020).
Polartorsk er en viktig matressurs for både fisk, sjøfugl og sjøpattedyr og sørger derfor for mangfold i området (Boitsov mfl. 2016, Eriksen mfl. 2015, Korsch mfl. 2015).
Områdene vest, nord og øst for Svalbard har den største biodiversiteten i Barentshavet av megafauna (Jørgensen mfl. 2019). Vest-, nord- og østsiden har forskjellige bunnsamfunn som er tilpasset de forskjellige miljøene, og som samlet bidrar til det biologiske mangfoldet i områder som kan ha havis.
Iskantsonen i Barentsregionen er en svært artsrik region for marine pattedyr i en sirkumpolar arktisk sammenheng (CAFF 2017). I tillegg utgjør alle de endemiske hvalene, samt isbjørn og hvalross, i dette området egne unike genetiske bestander som er svært viktige å bevare i forhold til global biodiversitet (hvalross: Andersen mfl. 1998, hvithval: O’Corry-Crowe mfl. 2010, narhval: Louis mfl. 2020, grønlandshval: Bachmann mfl. 2020, isbjørn: Zehl mfl. 2009).
Iskantsonen er svært viktig for en rekke sjøfuglarter av arktisk og subarktisk opprinnelse. De arktiske artene domineres antallsmessig av polarlomvi og alkekonge. I tillegg finnes arter som polarmåke, sabinemåke, rosenmåke og ismåke (Strøm mfl. 2020), alle relativt fåtallige. Store deler av storjobestanden i Norge hekker på Svalbard, med den største kolonien på Bjørnøya. Lomvi har en mer sørlig utbredelse i Nord-Atlanteren enn polarlomvi, men den største lomvikolonien i Norge finnes også på Bjørnøya. Også krykkje er nå vanligere som hekkefugl her enn på fastlandet.
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
Isbiota Dyreplankton Fisk
Iskantsonen er i så liten grad påvirket av menneskelig aktivitet at den kan regnes som naturlig i henhold til EBSA-kriteriene. Med unntak av de pågående klimaendringer, så er isbiota lite påvirket av menneskelig aktivitet som skipstrafikk, fiskeri eller forurensning, introduserte arter m.m. (Eamer mfl. 2013). Lite menneskelig påvirkning på dyreplankton foreløpig, unntatt menneskeskapte klimaendringer (Dalpadado mfl. 2020). Nordlig del av Barentshavet er ansett som viktig refugie for f.eks. grønlandshval, narhval, hvalross og isbjørn (Kovacs mfl. 2011).
8.4.3 - Lokalisering
Iskantsonen er en overgangssone mellom isfritt og isdekket hav, hvor iskonsentrasjonen er mellom 15 og 80 prosent. Iskantsonen omfatter området mellom ca. 74°N – 83°N.
Avgrensningen av iskantsonen som et særlig verdifullt og sårbart område er i forvaltningsplanen for Barentshavet–Lofoten satt der det forekommer havis 15 prosent av dagene i april (15 prosent isfrekvens), basert på satellittobservasjoner av isutbredelse fra 30-årsperioden 1988–2017. Denne grensen kan oppdateres med nye isdata ved senere oppdateringer av forvaltningsplanene (Meld. St. 20 (2019-2020)). Fagekspertgruppen anbefaler at sørlig grense for dette SVO-et endres til maksimal isutbredelse, dvs. yttergrensen for det største aggregerte arealet med isfrekvens større enn 0,5 prosent, for å fange opp en større andel av de biologiske prosessene som er knyttet til dette området.
Forslag til endret SVO Iskantsonen overlapper med SVO Polar tidevannsfront, foreslått SVO Havområdene rundt Svalbard og deler av de foreslåtte SVO-ene Eggakanten nord, Det sentrale Barentshavet og Midtlantisk rygg (Fig. 8.4.1).
Kandidatområdene i forslag til endret SVO Havområdene utenfor Svalbard hvor beitearealene for pelagisk beitende sjøfugl strekker seg 100 km ut fra de viktigste hekkekoloniene på øygruppen, vil også inngå i forslag til endret SVO Iskantsonen. Det samme gjelder deler av de marine verneområdene på Svalbard og «nye fiskeområder» som er definert i de nye reglene som regulerer fiskeriene med bunnredskap i det nordlige Barentshavet og områdene rundt Svalbard, vedtatt i 2019.
8.4.4 - Introduksjon til området
Iskantsonen er svært dynamisk og beveger seg fra Bjørnøya og Sentralbanken i sør til nord for Spitsbergen, avhengig av årstid, med mest is øst for Spitsbergen (von Quillfeldt mfl. 2018). Maksimal sørlig utbredelse av iskantsonen i Barentshavet styres i stor grad av polarfrontens beliggenhet, mens avsmelting om sommeren avhenger av blant annet solinnstråling, lufttemperatur, temperaturen på underliggende vannmasser, mengde av snø på havisen og vind. I tillegg til sesongmessige variasjoner, forekommer mer kortvarige variasjoner, for eksempel forårsaket av vind. Iskantsonen er også preget av sterke gradienter i isegenskaper, og store variasjoner i isflakstørrelse og form (Wadhams 1986, Zhang mfl. 2015). En av de mest markante konsekvensene av klimaendringer i Arktis er redusert isutbredelse (både i tid og rom) og redusert is-tykkelse (Polyakov mfl. 2017, Kwok 2018, Onarheim mfl. 2018, Årthun mfl. 2019, Docquier mfl. 2020, Spreen mfl. 2020). Tilførsel av havis fra Karahavet og Polhavet er viktig for isdekket i Barentshavet, og gir dessuten ferskvann når den smelter som opprettholder lagdelingen og et arktisk havklima nord i Barentshavet (Lind mfl. 2018). Ferskvann fra issmelting bidrar generelt til økt havforsuring (Fransson mfl. 2009, Chierici og Fransson 2009) med økt opptak av atmosfærisk CO2 i ferskere vann. Isprosessene bidrar til at det arktiske vannet har et økt potensiale for opptak av CO2 fra atmosfæren, hvilket øker havforsuring (Fransson mfl. 2017). Barentshavregionen er det området i Arktis hvor isreduksjonen har vært mest dramatisk de senere årene (Rodrigues 2008 og 2009, Smedsrud mfl. 2013, Onarheim mfl. 2018), særlig om vinteren, men samme trend observeres også i Framstredet (Hansen mfl. 2013, 2014 og 2015, Spreen mfl. 2020). Trenden i begge områdene viser at isen trekker seg nordover over tid, både for sommer- og vintersesongen, men det er store mellomårlige variasjoner (Pavlova mfl. 2014, Onarheim og Årthun 2017, Stroeve og Notz 2018).
Figur 8.4.2. Gjennomsnittlig overflatestrøm (piler) og bunndyp (farger) i sørlig del av forslag til endret SVO Iskantsonen og omkringliggende områder.
Isen i hele den polare iskantsonen kan være påvirket av smelteprosesser. Årsis er den mest vanlige istypen i Barentshavets iskantsone (Kvingedal mfl. 2005, Hansen mfl. 2014 og 2015, King mfl. 2017). Isen er generelt av mer regional opprinnelse enn isen i Framstredet og med lavere driftshastighet kan isen være utsatt for frysing og smelting over lengre tid (Vinje 1998 og 2001, Vinje og Kvambekk 1991). Generelt er også den isdekte delen av Barentshavet langt grunnere enn Framstredet, og særlig bankområdene har tilknytning til havisen.
I iskantsonen trekker isen seg i nordlig eller sørlig retning avhengig av sesongen. Dette området har dypvannsrenner (Franz Victoria) og bassenger (Olgadypet), separert med grunnere bankområder (Spitsbergen-, Stor- og Sentralbankene).
Figur 8.4.3. Figuren viser isfrekvens i april (venstre) og september (høyre) for perioden 1990–2019. Isfrekvensen er prosentandelen av dager det forekommer havis i et gitt område innenfor en nærmere angitt periode (Kilde: Norsk Polarinstitutt).
Snødekket på havisen er også relevant for fysiske og biologiske prosesser. Snøen kan isolere og redusere isveksten om vinteren og våren, den har høy albedo, og den kan danne grunnlag for smeltedammer i sommerhalvåret. Med hensyn til økosystemet har den blant annet betydning for demping av solstråling, og for sel som lager snøhuler. Generelt viser isen i Framstredet større snøtykkelse enn isen i Barentshavet. Det kan være flere faktorer som bidra til dette bildet, både regionale forskjeller i nedbør, og fordeling av istyper med mer flerårsis i Framstredet som kan samle mer snø om høsten enn årsis som dannes først i løpet av vinter og vår.
Smelting av havisen om våren/sommeren fører til at det dannes et overflatelag som er ferskere enn det arktiske vannet. Ferskere vann tar generelt opp mer CO2 fra atmosfære og bidrar til havforsuring (Fransson mfl. 2009, Chierici og Fransson 2009). Når isdekket forsvinner, blir det mer åpent hav med økt potensial for direkte CO2-opptak fra atmosfæren (Fransson mfl. 2009 og 2015). Aragonittmetning i de øvre 50 m i overflaten i perioden 2010-2020 viser verdier på 1,7-2,0 og pH på 8,10-8,15 (Lauvset mfl. 2016), med forventet minking med økt CO2-opptak. Tykkelsen av smeltevannslaget og graden av stabilitet mellom dette og vannet under avhenger blant annet av istykkelse og -konsentrasjon, smeltehastighet og hvor mye vinddrevet blanding som forekommer underveis i smeltesesongen. Hvis det øvre vannlaget er stabilt og med gunstige lysforhold og innhold av næringsstoffer, vil det resultere i primærproduksjon, som vil ta opp CO2.
Vi ser betydelig reduksjon i ismengden i dette området siden 2000-tallet samtidig med en nedgang i mengden av arktisk vann i området (Lind mfl. 2018, ICES 2019). Med mindre is har områdene med åpent vann økt betydelig og det samme har varigheten av de isfrie områdene. Avtagende havis har for noen organismer gitt habitatgevinst (mer åpent vannområde) mens for andre kan det forventes dårlige vilkår på grunn tap av habitat (mindre leveområde med is) i økosystemet. Mer åpent vann over et lengre tidsrom om sommeren har gitt økt planteplanktonproduksjon (PP) pga. mer inntrenging av lys i vannet. I Barentshavet har klorofyllmengden som gir grunnlaget for PP økt med 62 prosent i perioden 1998-2018 (Lewis mfl. 2020). I iskantsonen har også klorofyllmengden økt, men ikke så mye som i den nordøstlige delen av Barentshavet i den isfrie perioden (Dalpadado mfl. 2020).
8.4.5 - Beskrivelse av miljøverdiene
Isbiota
Figur 8.4.4. I og under isen er det ulike mikrohabitater med arter fra en rekke forskjellige artsgrupper. Modifisert fra Bluhm mfl. (2017a). Ill.: M. Daase. Kilde: von Quillfeldt mfl. (2018).
Issamfunn forekommer på under- og oversiden av isen og inne i isen (Horner mfl. 1992, Bluhm mfl. 2017a). Mer enn 2000 arter fra en rekke artsgrupper (f.eks. virus, bakterier, archaea, mikroalger, sopp, ciliater, kraveflagellater, amøber, foraminiferer; flercellete hjuldyr, nematoder, hoppekreps, amfipoder m.m.) har tilknytning til havis i hele eller deler av sin livssyklus (Bluhm mfl. 2017a og b, Hassett og Gradinger 2016, Ehrlich mfl. 2020, Gradinger 2020). Isens egenskaper (alder, tykkelse, lysforhold, drift m.m.) er bestemmende for type samfunn og utvikling av disse (Syvertsen 1991, Gradinger mfl. 2010, Fernandez-Mendez mfl. 2018, Hop mfl. 2020). Issamfunn utvikler seg over tid både med hensyn til struktur og funksjon, det vil si økt biodiversitet og kompleksitet når det gjelder interaksjoner mellom arter, biomasse og resirkuleringskapasitet (Poulin mfl. 2011, Bluhm mfl. 2017a, Hop mfl. 2020). Årsis har derfor ofte enklere samfunn enn flerårsis og består av arter som koloniserer isen hvert år på nytt (Ehrlich mfl. 2020). Selv for isbakterier er mangfold og mulig funksjon tydelig forskjellig mellom års- og flerårsis (Hatam mfl. 2016). Det kan også være variasjoner mellom år innenfor et område (Bluhm mfl. 2017 b).
Isalger (mikroalger som lever i tilknytning til isen) utgjør en konsentrert, næringsrik matkilde for både dyr som lever i isen og pelagiske beitere (Søreide mfl. 2010, Leu mfl. 2011, Wang mfl. 2015, Kohlbach mfl. 2016). Isalger er blitt anslått å utgjøre 16-22 prosent av den totale primærproduksjonen i iskantsonen i Barentshavet (Hegseth 1998) og opp til 50 prosent i det sentrale Polhavet (Gosselin mfl. 1997, Fernandez-Mendez mfl. 2015). Endrete isforhold vil imidlertid påvirke det relative forholdet mellom isalge- og planteplanktonproduksjon. Isalger er tilpasset lite lys, og primærproduksjonen i isen starter tidligere enn i vannmassene noe som bidrar til å forlenge den produktive sesongen i området (Hegseth 1998, Leu mfl. 2011, 2015). I tillegg kan en del sedimentere til havbunnen og utnyttes av bunndyr (Carroll og Carroll 2003, Ambrose mfl. 2005, McMahon mfl. 2006, Moran mfl. 2012, Rybakova mfl. 2019). Stor biomasse av bunndyr nordøst for Svalbard blir forklart med fødetilgang av isalger (Søreide mfl. 2013) og arter som livnærer seg ved å filtrere små partikler, større levende byttedyr eller nedfall fra havis (von Quillfeldt mfl. 2018). Den vertikale fluksen av isalger, andre organismegrupper og partikler til havbunnen henger sammen med havdyp, prosesser i vannmassene, havstrømmer m.m. (Olli mfl. 2002, Wiedmann mfl. 2020).
Isalger bidrar relativt sett mer til den totale primærproduksjonen jo lenger nord man kommer og i områder med flerårsis sammenlignet med områder med yngre is hvor en større andel av primærproduksjonen også foregår i vannmassene (Wassmann mfl. 2006, Wassmann og Reigstad 2011). Årsis kan imidlertid ha betydelige mengder med isalger (Szymanski og Gradinger 2016), men gjerne andre arter enn i flerårsis (Hop mfl. 2020). Allikevel er det også i årsis regionale forskjeller, blant annet økende andel arter som kan være viktige i planteplanktonoppblomstringen jo nærmere iskanten man kommer (Syvertsen 1991, Hop mfl. 2020).
Mens noen isfaunaarter (samlebetegnelse for ulike dyregrupper med tilknytning til isen) tilbringer kun deler av livssyklus i isen, har flere arter av amfipoder tilknytning til havis gjennom hele livssyklusen. Vanlige endemiske arter i isen er Gammarus wilkitzkii, Apherusa glacialis, Onisimus nanseni og O. glacialis. Den karnivore arten G. wilkitzkii er vesentlig større enn alle de andre artene, men den herbivore arten A. glacialis kan ofte være mest tallrik (Hop mfl. 2000, Gradinger og Bluhm 2004, Werner og Auel 2005). Selv om amfipodene deler samme habitat, okkuperer de ulike trofiske nisjer og har ulik diett (Scott mfl. 2001, Kohlbach mfl. 2016, Brown mfl. 2017). Disse amfipodene har en nøkkelrolle når det gjelder energioverføring, hovedsakelig til unge polartorsk, Boreogadus saida (Hop og Gjøsæter 2013, Kohlbach mfl. 2017), den mest tallrike fisken i Arktis. Havtemperatur og isdekke er viktige styrende faktorer for utbredelse i Barentshavet (Huserbråten mfl. 2019). Polartorsk er helt essensiell for mange arter av marine pattedyr og sjøfugl (Bluhm og Gradinger 2008).
Effekter av klimaendringer på havis og underliggende vannmasser er store og skjer med en hastighet som gjør det krevende å beskrive pågående endringer i isbiota i iskantsonen. I tillegg er det også regionale og temporære variasjoner (Sakshaug mfl. 2009, Reigstad mfl. 2011). Mindre flerårsis og mer årsis reduserer kompleksiteten i issamfunn, noe som allerede har vært observert i flere arktiske områder (Melnikov mfl. 2002, Hop mfl. 2020). Endringer i sjøis og snøkarakteristikk vil føre til endringer i isalgeproduktiviteten (Tedesco mfl. 2019). Isfylte områder med økt påvirkning av varmt atlanterhavsvann fører til økt smelting av isens underside og dette gjenspeiles for eksempel i Barentshavet ved redusert biomasse av isalger i den dypeste delen av isen (Barber mfl. 2015). Redusert isalgebiomasse forklarer også nedgangen av de isamfipodene som har isalger som fødepreferanse i Nansenbassenget nord for Svalbard (Hop mfl. 2013, Barber mfl. 2015). I tillegg er noen artsgrupper fraværende (Acoela, Platyhelminthes og Nematoda) og andre sterkt redusert (hjuldyr og isamfipoder) i isen nord for Svalbard de siste tiårene pga. endret kobling mellom is produsert på grunne sokkelområder og pakkisen (Ehrlich mfl. 2020). Med tynnere og derfor mer mobil is vil det løstsittende isalgesamfunnet på undersiden av isen lettere løsne på grunn av isens bevegelse og/eller fordi isen driver inn over varmere atlanterhavsvann (Assmy mfl. 2013). Dette kan være en av årsakene til at dette samfunnet ikke observeres så ofte vest i Barentshavet som tidligere (von Quillfeldt pers. medd.). Dette underissamfunnet utgjør mest biomasse sammenlignet med andre typer isalgesamfunn. Lenger sesong med åpent vann har også ført til at en mindre andel av primærproduksjonen kommer fra isalger sammenlignet med tidligere (Wassmann og Reigstad 2011, Barber mfl. 2015) samtidig som den pelagiske produksjonen er trukket lenger nordover (Renaut mfl. 2018). Tynnere is og mer råker fører også til tidlige planteplanktonblomstringer under isen (Assmy mfl. 2017). På den annen side vil tynnere is og relativt sett mer snø som tynger isen ned føre til at sjøvann trenger inn i snølaget på isens overflate noe som resulterer i en egen samfunnstype (infiltrasjonssamfunn) som tidligere sjeldent ble observert i Arktis (McMinn og Hegseth 2004, Fernández-Méndez mfl. 2018).
Plankton
Iskantsonen er overgangen mellom isdekket hav og isfritt hav, der råker og smeltedammer på isen har betydning for planteplanktonproduksjonen (Assmy mfl. 2017). Iskantsonen er dynamisk og vil bevege seg nordover etter som isen smelter (von Quillfeldt mfl. 2018). Det fysiske og kjemiske miljøet ligger til rette for planteplanktonproduksjon i iskantsonen, der issmelting fører til vertikal stabilitet, vinterkonsentrasjoner av næringssalter i overflaten og forbedrede lysforhold (Sakshaug mfl. 2009, von Quillfeldt mfl. 2018). Planteplanktonoppblomstringen vil følge isen ettersom den trekker seg nordover, men er mindre markant i de nordlige delene (Wassmann mfl. 2006). Modellering av planteplanktonproduksjon indikerer at størstedelen av planteplanktonproduksjonen skjer i iskantsonen og reduseres betydelig når iskonsentrasjonen er mer enn 80 prosent (von Quillfeldt mfl. 2018). Det er stor romlig variasjon i mengde planteplanktonproduksjon, i tillegg til variasjon mellom år. Det har også vært antydet at den totale primærproduksjonen i Barentshavet er opp mot 30 prosent høyere i år med lite is sammenlignet med år med mye is (Sakshaug mfl. 2009). På grunn av den store mellomårlige variasjonen i forekomst av havis vil også produksjonsforholdene i den delen av Barentshavet hvor iskantsonen kan befinne seg variere mer sammenlignet med den sørlige delen av Barentshavet som ikke har is (Reigstad mfl. 2011).
Starttidspunkt for oppblomstringen i iskantsonen avhenger av isens maksimale sørlige utbredelse om vinteren (enten den ligger sør for polarfronten eller ikke), men starter vanligvis i midten av april i sør og så sent som september i de nordligste delene. Dette er 6-8 uker tidligere enn i isfrie områder lenger sør hvor det stabile overflatelaget er resultat av oppvarming, og ikke issmelting (Sakshaug mfl. 2009, Assmy mfl. 2017). Den estimerte årlige produksjonen vil imidlertid være høyere i de isfrie delene av Barentshavet enn i isdekkete områder, men den konsentrerte og intense produksjonen i iskantsonen har stor betydning for sekundærproduksjon og bunnlevende organismene i disse områdene (Reigstad mfl. 2011).
Planktonisk isalge Nitzschia frigida. Foto: P. Assmy, NP
Figur 8.4.5. Stabilt næringsrikt overflatelag og tilstrekkelig lys fremmer en planteplanktonoppblomstring i iskantsonen. Ill.: M Daase. Kilde: von Quillfeldt mfl. (2018).
Atlanterhavsvann som blir transportert fra Norskehavet inn i Barentshavet og til vest- og nordsiden av Svalbard tilfører varme og bringer med seg plankton og næringssalter som er viktig for å opprettholde dyreplanktonbestander i iskantsonen (Dalpadado mfl. 2012).
Biomassen av dyreplankton om høsten, perioden når overvåkning i iskantsonen forgår, har vært relativt stabil (7 g tørrvekt per m2) i perioden 1989-2017 (Dalpadado mfl. 2020). Høyest biomasse er observert i det nordlige Svalbardområdet og Hopendjupet (ca. 23 g tørrvekt per m2) og lavest i Sentralbank- og Franz-Victoria-områdene (<2 g tørrvekt per m2). I de mer sørlige atlantiske vannmassene utgjøre raudåte Calanus finmarchicus den største biomassen, mens i den arktiske delen er C. glacialis dominerende (Søreide mfl. 2010, Aarflot mfl. 2018). Mange arktiske sekundærprodusenter har tilpasset sin livssyklus til den korte, men intense algeoppblomstringen om våren, dvs. de er avhengig av en god kobling ("match") i tid og rom med algeoppblomstringen (Søreide mfl. 2010). Alle de tre vanligste Calanus-artene er herbivore («planteetende»), men C. glacialis er særskilt tilpasset et liv i isfylte farvann ved at den får energi til reproduksjon via isalger, mens avkommet primært ernærer seg av pelagisk planteplankton. Hos de litt større dyreplanktonartene ser vi mer markerte endringer. Flere studier viser at krillbestanden har økt, særlig mer sørlige krillarter som Meganyctiphanes norvegica og Thysanoessa inermis (Orlova mfl. 2015, Eriksen mfl. 2016, Stige mfl. 2019). Disse inntar i større grad de nordlige delene av isfrie områder i Barentshavet. Som følge av mindre arktisk vann i de varme periodene, ser vi en nedgang i den arktiske amfipoden, Themisto libellula (ICES 2019, Stige mfl. 2019). Alle dyreplanktonartene som er nevnt her er viktig føde for lodde, polartorsk, torsk, sjøpattedyr og sjøfugl (Dolgov mfl. 2011, Hunt mfl. 2013, Fauchald mfl. 2015, ICES 2019). Den observerte nedgangen i bestanden av polartorsk i de siste årene kan skyldes reduksjon i havis (Husebråten mfl. 2019), og i tillegg en reduksjon av viktige byttedyrarter som T. libellula (Stige mfl. 2019, Dalpadado mfl. 2020).
Fisk
Polartorsk (Boreogadus saida) er i det nordlige Barentshavet tilknyttet isfylte farvann. Denne nøkkelarten lever i de frie vannmassene, men er oftest fordelt ned mot bunnen, gjerne i svært tette konsentrasjoner (Hop og Gjøsæter 2013). Den lever av planktonorganismer og når den blir større beiter den også på fiskeyngel. Lodde (Mallotus villosus) er en annen viktig art som beiter på produksjonen i iskantsonen. Bestandene av polartorsk og lodde har svingt betydelig i de siste 40 årene, og i siste tiåret har de vært på et lavt nivå (ICES 2020 a,b). Med unntak av tre pelagiske arter (lodde, polartorsk og istorsk (Arctogadus glacialis), er de fleste fiskeartene i iskantsonen i Barentshavet sterkt tilknyttet havbunnen (Christiansen 2012).
For de kommersielt viktige fiskeartene i Barentshavet er iskantsonen først og fremst et beiteområde, og i noen grad også et oppvekstområde (von Quillfeldt mfl., 2018, se også foreslått SVO Havområdene rundt Svalbard, Kap. 8.3.5). Torsk (Gadus morhua) kan for eksempel beite langt inn i iskantsonen om sommeren og høsten. Særlig unglodde og deler av torskebestanden vil dessuten ha tilhold i iskantsonen om vinteren. I tillegg finnes det en god del ikke-kommersielle arter (for eksempel ringbuker, enkelte ålebrosmer og ulker) som har hele livssyklusen i dette området. Von Quillfeldt mfl. (2018) gjorde en verdisetting av iskantsonen delt i gyte-, beite- og overvintringsområder for fisk. Iskantsonen ble også i forrige vurdering gitt en høy verdi for arktiske arter som polartorsk og andre ikke-kommersielle arter. For lodde, torsk, blåkveite (Reinhardtius hippoglossoides) og gapeflyndre (Hippoglossoides platessoides) ble området gitt middels verdi og for hyse (Melanogrammus aeglefinus) og snabeluer (Sebastes mentella) lav verdi siden disse artene også kan beite over større deler av Barentshavet, men det er vist at lodden får lavere fettinnhold når isen har trukket seg langt nord. Det er negativt for overvintring og gyting (Orlova mfl. 2013). Bankområdene er særlig viktige som beiteområder for fisk. Sammenlignet med fiskesamfunn i atlantiske vannmasser i Barentshavet har de arktiske fiskesamfunnene i iskantsonen også annerledes næringsnettstruktur, med mer spesialisert diett og mindre kannibalisme (Kortsch mfl. 2015).
Bunnsamfunn
Alger utgjør en konsentrert, næringsrik matkilde for både dyr som lever i isen og pelagiske beitere (Søreide mfl. 2010, Leu mfl. 2011) og i tillegg kan en del sedimentere til havbunnen og utnyttes av bunndyr (Carroll og Carroll 2003, Ambrose mfl. 2005). Den relative betydningen av isalger for dyreplankton i forhold til for bunndyr avhenger blant annet av om isalgene frigjøres gradvis eller mesteparten på en gang. Dersom iskantsonen befinner seg over dype havområder vil dessuten mindre alger nå havbunnen enn på sokkelområder, selv om isalger har vært observert ned til 4400 meter (Boetius mfl. 2013). Også vertikal fluks av andre organismer og partikler til havbunnen henger sammen med havdyp, prosesser i vannmassene, havstrømmer m.m. (Fig. 8.4.2). I dype havområder vil imidlertid denne produksjonen uansett være utilgjengelig for høyere trofiske nivå. Stor biomasse av bunndyr nord-øst for Svalbard blir forklart med fødetilgang av isalger (Søreide mfl. 2013) og arter som livnærer seg ved å filtrere små partikler, større levende byttedyr eller nedfall fra havis. Det er også høyere megabentisk sekundærproduksjon i de delene av Barentshavet som har sesongmessig isdekke, sammenlignet med isfrie områder, noe som sannsynligvis skyldes tettere pelagisk-bentisk kobling og lokale miljøforhold som blant annet innvirker på omrøring av vannmasser (Degen mfl. 2016).
Endrete isforhold påvirker den bentisk-pelagiske koblingen i iskantsonen, noe som kan få integrerte effekter i hele økosystemet. En reduksjon av havis kan ha en dramatisk effekt på timing, omfang og romlig fordeling av istilknyttet og pelagisk primærproduksjon, og som et resultat av dette, tilgangen av denne matkilden til de pelagiske og bentiske konsumentene (Carroll og Carroll 2003, Hansen mfl. 2003, Grebmeier mfl. 2006, Wassmann mfl. 2006). Endringer i biomasse som følge av redusert havis kan for eksempel ha en mulig konsekvens for megabenthossamfunnet, men det er ikke mulig å si noe om sammenhengen mellom havis og biomasse gjennom året i Barentshavet fordi overvåkningsperioden frem til i dag er for kort. Videre vil dette kunne føre til endret mattilbud for marine pattedyr, for eksempel storkobbe og hvalross, og for arktiske fiskearter som har spesialisert seg på beiting på bunndyr, slik som knurrulker og ringbuker.
Bunndyrene i Barentshavet viser et tydelig biogeografisk mønster, der artene har tydelige preferanser når det kommer til dybde, temperatur, saltholdighet, sedimentets beskaffenhet, strømforhold og graden av isdekke, dvs. antall dager per år som et gitt område er dekket av havis (Cochrane mfl. 2012, Carroll og Ambrose 2012, Jørgensen mfl. 2015a).
Der det er en forholdsvis sterk kobling mellom de fysiske og biologiske prosessene i de frie vannmasser og på bunnen er det dominans av arter som er aktive oppe på havbunnen, mens det i dypere områder med mindre strøm og tykkere sediment er flere gravende arter (Cochrane mfl. 2012). Iskantsonen og de nordlige delene av Barentshavet er generelt kjennetegnet av et høyt antall arter. Basert på store bunndyr (megabenthos) kan iskantsonen i Barentshavet grovt sett deles inn i to store biogeografiske regioner: nordvest og nordøst (Jørgensen mfl. 2015a).
Den nordvestre delen av iskantsonen, som kjennetegnes av relativt grunt vann, lave temperaturer, lav saltholdighet og relativt høy grad av isdekke (området er stort sett dekket av is om vinteren) har høyest artsdiversitet av store bunndyr.
Den nordøstre delen av iskantsonen, som kjennetegnes av relativt dypt vann, lave temperaturer, høy saltholdighet og høy grad av isdekke har høyest antall av store bunndyr. Det er også registrert biomasse «hotspots», særlig på dypere vann vest for Svalbard samt oppå og langs sidene på Spitsbergenbanken og Sentralbanken i det sentrale Barentshavet. Hopendypet har relativt mindre biomasse, men derimot et høyt antall små individer. Høyt antall arter, stor biomasse og mange individer samtidig som følge av særlig komplekse biologiske habitater, finner man på bankene og på sidene av bankene i Barentshavet, samt skråningen vest for Svalbard (Jørgensen mfl. 2015a).
Megabentossamfunnene i iskanten kan skilles mellom de mer boreale svampedominerte i sør (se røde stasjoner i figur 8.4.6) og de som er dominert av arktiske og subarktiske slangestjerner, sjøstjerner og sjøpølser lenge nord (blå stasjoner i figur 8.4.6), på ca. 250-300 m dyp i Barentshavet. De grunnere bankene (ca. 100 m) er dominert av filtrerende sjøpølser (Cucumaria frondosa, brunpølse) og haneskjell (Chlamys islandica). De grunne megabentossamfunnene har tilpasset seg et levesett som effektivt kan fange eller filtrere en mulig direkte og indirekte økt produksjon av føde fra havisen, mens dypere samfunn har en høyere andel av detrivorer (dvs. dyr som plukker opp døde eller levende matpartikler fra havbunnen) og predatorer.
Skillet mellom det svampe-dominerte og pigghud-dominerte områdene følger delvis den bentiske polarfronten (arktisk bunnvann T< 0 °C møter atlantisk bunnvann T> 3 °C). Dette skillet illustrerer langtidstilpasningen (min. 7 år) til miljøet da megafauna bruker tid på å «flytte på seg» som en respons på isdekke eller bunntemperaturer. «Bentosfronten» er derfor langt mer stabil enn den faktiske iskant-fronten eller variasjoner i bunntemperatur.
Figur 8.4.6. Bunndyrsamfunnene i Barentshavet delt opp i farger etter hvilke arter som dominerer. De røde er svampedominert, mens de blå er pigghud-dominert og de grønne er dominert av fauna hovedsakelig funnet på de grunne bankene (modifisert etter Jørgensen mfl. 2015).
Snøkrabben (Chionectes opilio) er en nyankommen art i den sørlige delen av iskantsonen som setter sitt preg på det opprinnelige bunndyrsamfunnet (Hjelset 2014, Frainer mfl. 2017).
Sjøpattedyr
Alle de arktiske endemiske (stedegne) sel- og hvalartene i Barentsregionen er is-assosierte og de fleste finnes i iskantsonen enten sesongmessig eller året rundt (Hamilton mfl. 2021). Isen gir beskyttelse mot dårlige værforhold (som er særlig viktige for unge dyr) og mot predatorer som finnes i åpne farvann, i første rekke spekkhoggere. Alle sel og hvalartene her, både de stedfaste og de som bare kommer hit om sommeren, drar nytte av den høye produksjonen stimulert av oppstrømning ved iskanten og næringsfrigjøring når isen smelter om sommeren som skaper et forutsigbart furasjeringsområde. Iskantens geografiske plassering varier både sesongmessig og mellom år, men de marine pattedyrene er svært mobile og forholder seg uten problemer til denne variasjonen. Isen er også en hvile- og hårfellingsplattform for ulike selarter. Fordi det er høye tettheter med ulike selarter i iskantsonen, er denne også et viktig jaktområde for en stor del av isbjørnbestanden i Barentsområdet. Mange av bestandene av marine pattedyr som finnes i iskantsonen ble overbeskattet i tidligere tider og noen er fortsatt truet. I tillegg vil alle de arktiske isavhengige marine pattedyrene bli negativt påvirket av reduksjonen i isdekket som skjer i dag som følge av global oppvarming. Utbredelsene er allerede endret og endringer i atferd, diett og annet er allerede påvist (se under for effekter på individuelle arter). Hvis/når iskanten flytter seg så den ligger over det dype Nordishavet i stedet for over grunnere sokkelområder i Barentshavet, vil antakelig marine pattedyr som finnes i iskanten og dypere inne i isen bli svært negativt påvirket.
Spitsbergenbestanden av grønlandshval (Balaena mysticetus) er klassifisert som kritisk truet (CR) både på den norske rødlisten og av IUCN (Cooke og Reeves 2018). Akustisk overvåkning i drivisen i Framstredet mellom Svalbard og Grønland har vist at arten er til stede her året rundt med avansert sangaktivitet døgnet rundt i vintermånedene, noe som tyder på at dette området er et forplantningsområde for denne bestanden (Stafford mfl. 2012, Ahonen mfl. 2017). I en flytelling i isen nord for Svalbard i 2015 ble det gjort 15 observasjoner av grønlandshval som førte til et estimat på 27 dyr innenfor et 52 919 km2 stort område, noe som gjør at man antar at hele Spitsbergenbestanden består av noen få hundre dyr (Vacquié-Garcia mfl. 2017b). På tross av lav tetthet har sporingsstudier av disse grønlandshvalene vist at de oppholder seg over hele det historiske utbredelsesområdet i iskantsonen fra Øst-Grønland videre nord for Svalbard og østover til Frans Josef land (Kovacs mfl. 2020 a). I motsetning til andre og større bestander av grønlandshval lengre vest, så vandrer ikke dyrene fra Svalbardbestanden i en klassisk forstand. De sprer seg heller fra vinterområdene nord i utbredelsesområdet om våren og returnerer til disse om høsten igjen. De holder seg stort sett hele tiden i iskanten eller inne i isen, med et vandringsmønster motsatt andre bestander (vandrer sør om sommeren og nord om vinteren) (Lydersen mfl. 2012, Kovacs mfl. 2020 a). Spitsbergenbestanden av grønlandshval utgjør ellers en egen genetisk enhet forskjellig fra andre bestander og må derfor forvaltes/beskyttes i forhold til dette (Bachmann mfl. 2020). De pågående økningene i havtemperaturen samtidig med reduksjon i isutbredelsen kan bli kritiske for grønlandshvalene både på grunn av tap av habitat og også termisk stress. I tillegg vil mindre is kunne gjøre at vi får en økning i predasjon fra spekkhoggere noe som allerede er et alvorlig problem for grønlandshvalbestanden i Okhotskhavet (Shpak og Paramonov 2018). Eksponering for sykdommer, endringer i næringsnettet som kan føre til mindre tilgjengelig mat (særlig fettrike kopepoder) og økt menneskelig aktivitet i Barentsregionen med tilhørende økning i undervannsstøy er også bekymringsverdig. Hvis grønlandshvalene i Spitsbergen-bestanden opprettholder sin sterke preferanse for kalde, islagte områder vil deres utbredelse flyttes mot nord til offshore, dype områder hvor de vil måtte ta til takke med pelagiske zooplanktonarter som nesten helt sikkert vil bremse potensiell vekst og da føre til en reduksjon i denne bestanden.
Narhvalene (Monodon monoceros) i Barentsregionen har fått svært lite oppmerksomhet. De få studiene som har vært gjort viser at deres foretrukne habitat er inne i drivisen i Framstredet og den nordlige Barentsregionen. Data fra en flytelling (Vacquié-Garcia mfl. 2017b) og passiv akustisk overvåkning (Ahonen mfl. 2019) bekrefter at narhvalene i Barentsregionen, i motsetning til andre narhvalbestander, oppholder seg i drivisen hele året og foretrekker områder med opptil 90 prosent isdekke. Genetiske studier viser liten genetisk diversitet mellom narhval fra ulike områder i deres utbredelsesområde, men at narhval fra Svalbard og Øst-Grønland skiller seg fra andre bestander og også at Svalbard-hvalene er noe forskjellig fra de på Øst-Grønland (Louis mfl. 2020). Bestanden av narhval i Barentsregionen er på den nasjonale Rødlisten og klassifiseringen er akkurat blitt oppdatert til sterkt truet-EN (fra Datamangel-DD) basert på en flytelling fra 2015. Her ble narhval sett ved 11 anledninger med totalt 58 individer innenfor et område på 52 919 km2, noe som førte til et estimat på 837 (CV = 0.501) narhval i dette området. Narhval foretrekker kalde arktiske vannmasser (0,6–1,5 °C; Heide-Jørgensen mfl. 2020) og studier av effekter av tidligere tiders klimaendringer på fylogeografi og demografi tyder på at de responderte kraftig på temperaturendringer og endringer i isutbredelse og vil antakelig gjøre det igjen som følge av dagens endringer i disse parameterne (Louis mfl. 2020). Andre utfordringer for narhvalene som følge av redusert isdekke er økt predasjon fra spekkhoggere noe som allerede er dokumentert i Arktisk Canada (Lefort mfl. 2020). Narhval er vurdert til å være en av de arktiske marine pattedyrartene som er mest utsatt for effekter av pågående klimaendringer (Laidre mfl. 2008, Reeves mfl. 2014).
Hvalrossene (Odobenus rosmarus) i det nordlige Barentshavet består av en enkelt bestand (Andersen mfl. 1998, Andersen mfl. 2017) som tilbringer vintrene inne i drivisen og somrene rundt Svalbard og Frans Josef land. Her hviler de seg på land på de mange øyene i disse områdene når sjøisen over sokkelområdene har flyttet seg lenger nord (Hamilton mfl. 2015a). Det yngler og parer seg om vinteren inne i isen mellom disse øygruppene opptil 600 km nord for iskanten i iskonsentrasjoner som ofte er over 90 prosent (Freitas mfl. 2009, Lowther mfl. 2015). Denne hvalrossbestanden er klassifisert som sårbar (VU) på den norske rødlisten hovedsakelig på grunn av relativt få hunndyr og dårligere isforhold (Kovacs mfl. 2011). Størrelsen på hele bestanden er ikke kjent fordi data ikke er tilgjengelige fra Frans Josef land området, men for Svalbard er det siste estimatet fra 2018 på 5503 (95 % konfidensintervall 5031-6036) individer som da var det andre overvåkningsintervallet med økende trend (Lydersen mfl. 2008, Kovacs mfl. 2014, www.mosj.no/no/fauna/hav/hvalross-bestand.html).
Ringsel (Pusa hispida). Iskantsonen er kjent som et viktig beiteområde for ringsel (Wathne mfl. 2000) og en ukjent fraksjon av Barentshavbestanden yngler ute i drivisen (Wiig mfl. 1999). Vi vet lite om ynglesuksess ute i drivisen, og det beste yngleområde for ringsel i dette området er antatt å være den mer stabile landfaste isen (Lydersen 1998). Unge ringsel fra Svalbard sammen med enkelte voksne individer foretar vandringer opp til iskantssonen sent på sommeren og tidlig høst (Freitas mfl. 2008a). Hovedbyttedyrene ved iskanten er Themisto libellula og polartorsk (Boreogadus saida). De selekterer områder med rundt 50 prosent isdekke og unngår områder med for høye konsentrasjoner (80-100 prosent; Lone mfl. 2019). Disse vandringene til iskanten har blitt stadig lengre ettersom sommerisen har trukket seg lenger nordover og dette har endret atferden til disse dyrene (Hamilton mfl. 2015b, Lone mfl. 2019). I senere tid må ringselene arbeide hardere for å finne mat i dette området – de dykker mer av tiden, dykker mindre rett under isen (for is-assosierte byttedyr), de hviler mindre og svømmer lenger og har færre perioder med mye dykking innenfor et begrenset område som indikerer at de finner færre steder hvor maten står konsentrert (Hamilton mfl. 2015b). Klimaendringene er en alvorlig trussel for ringselene over hele deres sirkumpolare utbredelsesområde på grunn av effekter på sjøis med tilhørende endringer i næringsnettet og endring i snøforhold som trengs for å grave ut kastehuler som er essensielle for ungeoverlevelse.
Storkobber (Erignathus barbatus) har blitt lite studert i iskantsonen. Man vet at de yngler i drivisen i Barentshavet og hannenes svært typiske undervannsang i parringstiden høres hele tiden på akustiske lyttebøyer i Framstredet og i drivisen nord for Svalbard (Moore mfl. 2012, Ahonen mfl. 2017, Martínez Llobet mfl. submitted ms). Som for de andre is-assosierte marine pattedyrene så antar man at klimaendringer representerer en alvorlig trussel også for storkobber (Kovacs mfl. 2011).
Grønlandssel (Pagophilus groenlandicus) og klappmyss (Cystophora cristata) bruker drivisiskanten som hvileområder gjennom beitesesongen når de veksler mellom hvile og matsøk (CAFF 2017). Fordelingen av hvalobservasjoner på norske hvaltelletokt indikerer at iskantsonen er viktig sommerbeiteområde for finn- og knølhval (Leonard og Øien 2020).
Isbjørn (Ursus marinus). Den største andelen av isbjørnene i Barentshavområdet benytter iskantsonen mye av året, i et belte omtrent 100 km nordover fra iskanten (Aars mfl. 2009). Siden iskantsonen har flyttet seg lengre nordover de siste tiårene, har isbjørnene fulgt etter. Isbjørnen foretrekker ikke for tykk sjøis med åpne råker hvor det er lettest å jakte sel, og hvor det er høy produktivitet (Durner mfl. 2009, Lone mfl. 2018a). De selartene som finnes i iskantsonen, vil være de viktigste byttedyrene for isbjørnbestanden. Det at iskantsonen mye av året er lengre nord enn tidligere har også ført til at tidligere viktige yngleområder (hiområder) er blitt mindre egnet enn før. Drektige binner må ha sjøis mellom jaktområdene og hiområdene i perioden på høsten for å kunne komme seg til hiområdene, helst før midten av november. Hopen (Derocher mfl. 2011) og Kongsøya (Aars 2013) har i senere år ofte manglet sjøis på høsten, og det har trolig skjedd et skifte mot at flere isbjørn nå går i hi på Frans Josef land, men gode data fra Russisk Arktis mangler for å dokumentere dette. Alternative hiområder på Svalbard for binner som vandrer fra iskanten er Nordaustlandet, Kvitøya og i noen grad Hinlopen og Edgeøya eller Barentsøya (Andersen mfl. 2012). Vi finner langt høyere tettheter av isbjørn langs iskanten i østlige områder av Barentshavet hvor havområdene er grunne enn i de mer vestlige områdene som ligger over mye dypere vann. En ytterligere forskyvning av iskanten nordover vil således kunne forventes å påvirke bestanden negativt på sikt. Tap av sjøishabitat de senere tiår har for Barentshavbestanden vært mye raskere enn for noen av de andre 18 isbjørnbestandene i Arktis (Stern og Laidre 2016), og forventes også å være raskere i dette området i kommende tiår enn hva som er predikert for isbjørnens øvrige leveområder (Durner mfl. 2009).
Sjøfugl
Iskantsonen kan bli brukt av krykkje (Rissa tridactyla), polarlomvi (Uria lomvia) og lomvi (Uria aalge), alkekonge (Alle alle) og ismåke (Pagophila eburnea) fra hele forvaltningsplanområdet (Arneberg og Jelmert, 2017, S. Descamps pers. medd.). Det betyr at det kan være fugl der fra områder med nedgang i hekkebestanden (krykkje: fastlandskysten, lomvi og polarlomvi: vestlige og nordlige deler av fastlandskysten), eller med stabil eller voksende hekkebestand (krykkje: Bjørnøya og Spitsbergen, lomvi: østlige deler av fastlandskysten og Bjørnøya). Foreløpige resultater, blant annet fremkommet ved bruk av lysloggere, har vist at særlig de nordlige sjøfuglbestandene på Spitsbergen (og Frans Josef land og Jan Mayen) forekommer i iskantsonen i større grad eller lengre perioder av året, enn de sørlige populasjonene (von Quillfeldt mfl. 2018). Med unntak av ismåke er det fortsatt store kunnskapsmangler med hensyn til betydning av iskantsonen for ulike arter av sjøfugl. I en verdisetting av iskantsonen som livshistorisk viktig område antas imidlertid iskantsonen å ha høy verdi gjennom hele året for ismåke, for polarlomvi middels om vår og høst og lav om sommer og vinter, alkekonge middels hele året unntatt vinter (lav), for krykkje middels om høsten ellers lav, for havhest og polarmåke lav gjennom hele året (von Quillfeldt mfl. 2018). Sjøfugl som bruker iskantsonen er utsatt for endringer i isutbredelse, særlig i hekkesesongen da de kan erfare økt avstand til optimale beiteområder. Dette gjelder flere rødlistede arter, som for eksempel ismåke, krykkje og alkekonge (bl.a. Gilg mfl. 2010).
Særlig polarlomvi og alkekonge kan forekomme i store konsentrasjoner i iskantsonen og inne i råker om våren, men også teist og ismåke er vanlig (Lønne og Gabrielsen 1992, Hunt mfl. 1996, Gilg mfl. 2010, Fort mfl. 2013). I tillegg observeres havhest, polarmåke og krykkje i iskantsonen gjennom hele året, men den relative betydningen av iskantsonen som furasjeringsområde er dårlig kjent for disse artene. Ismåken er den eneste arten som er direkte avhengig av iskantsonen gjennom hele sitt livsløp, men kan trekke over store områder om våren og høsten, selv om de normalt ikke forlater iskantsonen under trekkperiodene.
Det er først gjennom de siste års raske utvikling av ulike typer sporingsteknikker (lysloggere, små satellittsendere) at det har blitt mulig å få kunnskap om de ulike artenes bruk av iskantsonen. Bruken av disse teknikkene er likevel kun i startfasen og få studier er ennå publisert. Det forventes at bare innen noen få år vil kunnskapstilfanget være vesentlig bedre. Verdisettingen i dette dokumentet er derfor basert på en subjektiv vurdering basert på tilgjengelig kunnskap per i dag. Området som er vurdert begrenser seg til den norske delen av Barentshavet, samt iskantsonen i Grønlandshavet.
8.4.6 - Pågående og framtidige endringer for isbiota og habitatforhold
Iskantsonen er allerede markant påvirket av den globale oppvarmingen. Den har trukket seg nordover og sesongen med isdekke er blitt betydelig kortere. Volumet av isen har også blitt redusert med betraktelig mindre andel av flerårsis (Onarheim mfl. 2018, Docquier mfl. 2020). Det er stor sannsynlighet at denne trenden vil fortsette i fremtiden (IPCC, 2019). Framtidsscenarioer antyder at overflatetemperaturene i dette området vil øke med 1-1,5 °C frem mot 2060-2069 (Hanssen-Bauer mfl. 2018). Det forventes økt havforsuring på grunn av fortsatt opptak av atmosfærisk CO2 i perioden 2000-2065 (Skogen mfl. 2014). Modellresultater forventer også en undermetning av aragonitt i overflaten i år 2100 og minket pH ned til 7,7 (RCP8.5 scenario; Lauvset mfl. 2016, Fransner mfl. 2020), hvilket kan gi alvorlige konsekvenser for marine organismer.
Andel årsis og ung is vil fortsette å øke sammenlignet med flerårsis og resultere i mindre komplekse isbiotasamfunn (Olsen mfl. 2017). Videre må rekruttering av issamfunn i større grad måtte skje fra vannsøylen eller havbunnen. Hvor isen dannes og hvilke arter som er til stede (og som kan overleve i isen) har derfor stor betydning (von Quillfeldt mfl. 2018).
Endringer i tidspunkt for isdannelse (og inkorporering av organismer) vil sannsynligvis føre til større innslag av atlantiske arter i isen (Hop mfl. 2020) og endringer i balansen mellom is- og pelagisk produksjon (Wassmann og Reigstad 2011).
Arter med hele livssyklusen i havis, står i fare for å forsvinne eller bli sterkt redusert selv om det finnes mekanismer som bidrar til å holde isfauna i vannmassene (Berge mfl. 2012, Kunisch mfl. 2020) eller i bentiske samfunn i grunne områder (Poltermann 1998, Weslawski mfl. 2010).
Den tette koblingen mellom havis og det pelagiske næringsnettet gjør at endringer i havisdekket og isalgeproduksjon vil ha store konsekvenser for funksjonen til næringsnett og karbondynamikk i det pelagiske systemet (Kohlbach mfl. 2016, Flores mfl. 2019, Kohlbach mfl. 2021).
Tidligere issmelting og mer gjennomsiktig isdekke (flere smeltedammer og råker) vil redusere det relative bidraget fra isalger til den totale primærproduksjonen ytterligere og har potensiale til å øke frekvensen og intensiteten av planteplanktonblomstringen under isen (Ardyna mfl. 2020).
Endringer i biomasse som følge av redusert havis kan for eksempel være en mulig konsekvens for megabenthossamfunnet, men det er ikke mulig å si noe om sammenhengen mellom havis og biomasse gjennom året i Barentshavet, fordi overvåkningsperioden frem til i dag er for kort. Videre vil dette kunne føre til endret mattilbud for marine pattedyr, for eksempel storkobbe og hvalross, og for arktiske fiskearter som har spesialisert seg på beiting på bunndyr, slik som knurrulker og ringbuker (von Quillfeldt mfl. 2018). Sjøfugl som er avhengige av arktisk planktonfauna knyttet til iskantsonen, særlig arter som alkekonge og krykkje (se bl.a. Amélineau mfl. 2019, von Quillfeldt mfl. 2018) er utsatte dersom isutbredelsen minsker både når det gjelder fastis og brefronter. For mer om marine pattedyr og sjøfugl, se Kap. 8.3.6. Forslag til endret SVO Havområdene rundt Svalbard.
Polartorsk. Foto: E. Svensen
8.5 - Forslag til endret SVO Eggakanten nord (BH3)
8.5.1 - Sammendrag
Områdebeskrivelse og strukturer
Forslag til endret SVO Eggakanten nord (BH3) er en fortsettelse av forslag til endret SVO Eggakanten sør (NH5) og strekker seg fra grensen til forvaltningsplanområdet for Norskehavet og nordover til isdekket hav vest og nord av Svalbard, inkludert Yermakplatået. Hele lengden av Eggakanten har sammenlignbare oseanografiske prosesser, men det er stor variasjon i miljøforhold i bratte deler av skråningen. Eggakanten har en topografi som er karakterisert av sterk, nordgående atlanterhavsstrøm.
Sjøtemperaturene varierer i stor grad med varmetransporten i atlanterhavsstrømmen utenfor norskekysten og møtet med det kalde, ferske arktiske vannet fra nord. Vestspitsbergenstrømmen bringer varmt atlanterhavsvann langs sokkelskråningen vest og nord av Spitsbergen og holder området her isfritt store deler av året, men havis dekker vanligvis deler av området nord for Svalbard (sørøstlig del av Yermakplatået) og videre nordøstover.
Miljøverdier
Generelt er det forhøyet biologisk produksjon og stort biologisk mangfold langs eggakanten. Området inneholder flere sårbare naturtyper som bl.a. dyphavsjøfjær, grisehalekorallskog, Lophelia-rev, hardbunnskorallskog, og kaldtvanns-svampsamfunn. Den eneste kjente utbredelsen av grisehale-korallskog i Norge er begrenset til et lite område, nordvest for Tromsøflaket. Flere Lophelia-rev forekommer i sørlige del av området. Røstrevet, det største kjente dypvannsrevet av Desmophyllum pertusum (tidligere kalt Lophelia pertusa), ligger øverst i raskanten ved eggakanten i et område hvor det har gått et stort undersjøisk ras. Særlig i bratte deler av skråningen, hvor miljøforhold forandrer seg over korte distanser, kan biologisk mangfold være høyt i små områder. Vertikalblandingsprosesser som øker lokal primærproduksjon og adveksjon av næringsrik og høybiomasse atlanterhavsvann nordover er to grunner til at området er svært viktig for produktiviteten i området og øvrige områder i Barentshavet, rundt Svalbard, og i Polhavet. I den nordlige delen finnes flere arktiske ålebrosmer som bare finnes i dette området innenfor norske farvann. Området inneholder de viktigste gyte- og yngleområdene for mange kommersielt og økologisk viktige fiskearter som gyter eller yngler på kontinentalskråningen, inkludert torsk, hyse, sild, snabeluer, vanlig uer (sterkt truet-EN), samt blåkveite som har sitt eneste gyteområde i denne nordlige del av Eggakanten. Egg, larver og yngel av de ulike fiskeartene driver nordover i kjernen av atlanterhavsstrømmen og finner føde i strømmen på grunn av forhøyet planktonbiomasse. For sjøfugl er dette et viktig sommerbeiteområde, særlig for de pelagisk beitende artene i hekketiden vest av Spitsbergen, ved Bjørnøya og ved Norskekysten. Eggakanten er også et viktig beiteområde for hval som spiser mye dyreplankton, f.eks. finnhval og blåhval.
Figur 8.5.1. Kart over forslag til endret SVO Eggakanten nord (BH3). Grensene for det foreslåtte SVO-et er vist med tykk rød strek, mens grenser for overlappende og nærliggende foreslåtte SVO-er er vist med tynnere rød strek. Grensene for forvaltningsplanområdene er markert med brun linje. Kartet viser utbredelsen av utvalgte miljøverdier med betydning for området. Miljøverdier knyttet til fisk (ynglefelt for vanlig uer og snabeluer, og gytefelt for de øvrige artene) og sjøpattedyr er markert i venstre kart og miljøverdier knyttet til bunnsamfunn er markert i høyre kart.
Svamp og korall på Eggakanten nord. Foto: Mareano / HI
8.5.2 - Vurdering i forhold til EBSA-kriteriene – Forslag til endret SVO Eggakanten nord (BH3)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til endret
SVO Eggakanten nord (BH3)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet /sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Dyreplankton Sjøfugl
Fisk Bunnsamfunn
Den sterke, topografiskstyrte havstrømmen er unik i det europeiske Arktis og er svært viktig for produktiviteten i området og øvrige områder i Barentshavet, rundt Svalbard, og Polhavet (Wassmann mfl. 2019, Vernet mfl. 2019, Popova mfl. 2013). En rekke arter og grupper av plankton har eggakanten inkludert i sine utbredelsesområder (Torgersen mfl. 1997, Dalpadado mfl. 1998).
Området er svært viktig gyteområde for flere fiskearter som er særlig knyttet til dette området (Drevetnyak og Nedreaas 2009, Albert og Vollen 2015). Blåkveite gyter bare i nordlige del av Eggakanten (Nizovtsev 1989, Albert 2003) og en av de viktigste yngleområder for vanlig uer ligger utenfor Vesterålen (Mukhina mfl. 1992). Også hyse har sitt hovedgyteområde på kontinentalskråningen ved Lofoten (Bergstad mfl. 1987, Solemdal mfl. 1989). I den nordlige delen er det kalde vannmasser der det er dypt - her finnes flere arktiske ålebrosmer som bare finnes i dette området innenfor norske farvann (Bergstad mfl. 2017, Johannesen mfl. 2017).
Bestanden av grisehalekorall (Radicipes gracilis) er den eneste sikre observasjonen av en bløtbunnskorallskog dannet av denne arten i Norge mens den strømrike Eggakanten fører til et særlig rikt variert svampsamfunn (Jørgensen mfl. 2015 a, b, 2019).
For sjøfugl er dette et viktig beiteområde, særlig for de pelagisk beitende artene i hekketiden vest av Spitsbergen, ved Bjørnøya og ved Norskekysten (Systad mfl. 2019).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Sjøpattedyr
Fisk Sjøfugl
Området er svært viktig gyteområde for flere fiskearter som er særlig knyttet til dette området (Drevetnyak og Nedreaas 2009, Albert og Vollen 2015). Blåkveite gyter bare i nordlige del av Eggakanten (Nizovtsev 1989, Albert 2003) og en av de viktigste yngleområder for vanlig uer ligger utenfor Vesterålen (Mukhina mfl. 1992). Også hyse har sitt hovedgyteområde på kontinentalskråningen ved Lofoten (Bergstad mfl. 1987, Solemdal mfl. 1989). I den nordlige delen er det kalde vannmasser der det er dypt - her finnes flere arktiske ålebrosmer som bare finnes i dette området innenfor norske farvann (Bergstad mfl. 2017, Johannesen mfl. 2017).
Romlig fordeling av observasjoner på norske hvaltelletokt indikerer at eggakanten kan være viktig habitat for spermhval og finnhval (Leonard og Øien 2020, Skern-Mauritzen mfl. 2009; se også foreslått SVO Havområdene rundt Svalbard). Eggakanten og den øvre sokkelskråningen er også kjent for å være viktig beitehabitat for klappmyss som sannsynligvis beiter på blåkveite og andre dyptlevende pelagiske arter (Folkow og Blix 1999), selv om dette kan ha endret seg noe i senere tid (Vacquie-Garcia mfl. 2017 a).
For sjøfugl er dette et viktig beiteområde, særlig for de pelagisk beitende artene og særlig der Eggakanten ligger nært kolonien (Systad mfl. 2019).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Bunnsamfunn
Fisk
Sjøfugl
Eggakanten er delvis yngleområde for vanlig uer, som er sterkt truet (EN) (ICES 2020).
Bunnsamfunn langs Eggakanten har mye svamp og andre sentvoksende, men sårbare arter (Mareano.no).
Flere av sjøfuglartene som bruker eggakanten er sårbare og rødlistet (Systad mfl. 2018).
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Fisk
Sjøfugl Bunnsamfunn
Eggakanten er et viktig område for mange fiskearter, inklusive truede arter som vanlig uer (ICES 2020) og har noen bunnhabitater som kan bli skadet av fiske med bunntrål og aktivitet av oljeindustrien (Sundby mfl. 2013).
Mange dyphavsarter, som uer eller koraller, vokser veldig sakte og trenger derfor mye tid for restitusjon (Jakobsen og Ozhigin 2011). Siden Eggakanten er et stort område, er det noe bufret mot menneskeskapte forstyrrelser.
Sjøfuglene beiter på fiskelarver og dyreplankton i disse områdene, særlig i hekketiden, og er derfor ekstra sårbare for påvirkninger her (Systad mfl. 2018).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Planteplankton
Dyreplankton Fisk
Vertikalblandingsprosesser som øker lokal primærproduksjon og adveksjon av næringsrik og høybiomasse atlanterhavsvann nordover er to grunner til at området er svært viktig for produktiviteten i området og øvrige områder i Barentshavet, rundt Svalbard, og i Polhavet (Wassmann mfl. 2019, Vernet mfl. 2019, Popova mfl. 2013).
Det ville ikke har vært like mye Calanus finmarchicus i Barentshavet eller rundt Svalbard uten strømmene ved Eggakanten (Vestspitsbergenstrømmen) som fører høy biomasse fra Norskehavet nordover (Wassmann mfl. 2019, Carstensen mfl. 2019, Gluchowska mfl. 2017). Dette er et viktig byttedyr for mye av det pelagiske næringsnettet rundt Svalbard.
Området er svært viktig som gyteområde for flere fiskearter (Drevetnyak og Nedreaas 2009, Albert og Vollen 2015). Blåkveite gyter bare i nordlige del av Eggakanten (Nizovtsev 1989, Albert 2003) og en av de viktigste yngleområder for vanlig uer ligger utenfor Vesterålen (Mukhina mfl. 1992).
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Dyreplankton
Sjøpattedyr
Fisk Bunnsamfunn
Det foreligger ikke informasjon som tilsier at Eggakanten har høyere artsmangfold av dyreplankton, men arter som har sokkelen med kystvann eller oseaniske områder med atlantisk vann som sine habitater, vil kunne «møtes» ved eggakanten og ha dette området som habitat.
Området er svært viktig som gyteområde for flere fiskearter (Drevetnyak og Nedreaas 2009, Albert og Vollen 2015).
Den rike variasjonen av svamper og annen sentvoksende og sårbar megabenhosfauna utgjør i seg selv en rik biodiversitet (Jørgensen mfl. 2015, a, b, 2019). Generelt fører strøm, dybdeforhold og topografi til svært høyt lokalt biologisk mangfold. Fordi samfunn på eggakanten er styrt av dybde og derfor er avhengig av hvor bratt kontinentalskråningen er, kan biologisk mangfold bli veldig høyt over en kort distanse (Faglig forum for norske havområder 2019).
I tillegg er det beiteområde for sjøpattedyr, som klappmyss og om sommeren også sjøfugl (Skern-Mauritzen mfl. 2009, Systad mfl. 2019, Leonard og Øien 2020).
Samlet er det et høyt biologisk mangfold.
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
8.5.3 - Lokalisering
Avgrensing av SVO Eggakanten ble gjort i Meld. St. 20 (2019-2020), og er ikke endret i forhold til dette. Området er imidlertid nå foreslått endret ved å dele inn i foreslått SVO Eggakanten nord som beskrives her, og foreslått SVO Eggakanten sør som beskrives under Norskehavet. Eggakanten strekker seg fra Stad til nord av Svalbard, og foreslått SVO Eggakanten nord starter ved grensen til forvaltningsplanområdet for Norskehavet. Området omfatter hele sokkelskråningen og et stykke inn på sokkelen. Det er ulik bredde på området, avhengig av hvor bratt skråningen er (figur 8.5.1), og for Yermakplatået (se figur 8.3.1) omfatter avgrensningen av eggakanten omtrent halve platået, med dyp ned til ca. 800 meter. Man kunne i tillegg brukt et minimumsbunndyp, for å sikre at skråningen rundt hele platået ble tatt med, men den delen av området som har høyest biologisk aktivitet er med slik det nå er avgrenset.
Området overlapper delvis med de foreslåtte SVO-ene Kystsonen Lofoten, Tromsøflaket, Iskantsonen og Havområdene rundt Svalbard.
Eggakanten går inn i Forlandet nasjonalpark helt i vest, og inn i nordvesthjørnet av Nordvest-Spitsbergen nasjonalpark.
8.5.4 - Introduksjon til området
Foreslått SVO Eggakanten nord er sokkelskråningen som går opp fra Lofotenbassenget og 10 km inn på sokkelen. Det er ulik bredde på området, avhengig av hvor bratt skråningen er. Atlanterhavsstrømmen fø lger sokkelskråningen nordover. Sokkelskråningen har en sterk styrende virkning på strømmen (Orvik og Niiler 2002), og det resulterer i en sterk, ensrettet og smal atlanterhavstrøm langs foreslått SVO Eggakanten nord med strømhastigheter på 30-100 cm per sek (Anderson mfl. 2011) og relativt høye sjøtemperaturer i forhold til breddegradene (Mork og Skagseth 2010). Atlanterhavsstrømmen transporterer med seg næringssalter, plankton og fiskeyngel (Rey 2012, Dalpadado mfl. 2012, Sundby mfl. 2013).
Foreslått SVO Eggakanten nord har stor variasjon i miljøforhold i bratte deler av skråningen. For Y ermakplatået (nordvest av Spitsbergen) omfatter avgrensningen av eggakanten omtrent halve platået, med dyp ned til ca. 800 meter. Y ermakplatået er flatere enn resten av foreslått SVO Eggakanten nord, og har dermed både svakere strøm og mindre romlig variasjon enn Eggakanten for øvrig.
Sjøtemperaturene i foreslått SVO Eggakanten nord varierer i stor grad med varmetransporten i Atlanterhavsstrømmen utenfor Norskekysten. T emperaturen har hatt en positiv trend gjennom de siste 40 årene (González-Pola mfl. 2019). Den kjemiske variasjonen avhenger i stor grad av innstrømming atlanterhavsvann i sør, som transporterer næringsstoffer og høyt innhold av CO2 , og møtet med det kalde, ferske arktiske vannet fra nord med lavt innhold av CO2, karbon og næringsstoffer. I perioden 2012-2019 var variasjon i pH, CO2 og aragonittmetning drevet av fluktuasjoner i innstrømmingen av det arktiske og atlantiske vannet (Jones mfl. 2020). pH i overflaten er generelt lavere i sør enn i nord, og pH er høyere om sommeren enn om vinteren. Det er ikke noen tydelig trend i observasjonene for årene 2010-2018 (Jones mfl. 2020), og ikke heller i et lengre modellperspektiv er det noen tydelig trend (Fransner mfl. 2020). Når det gjelder kalkmetning av aragonitt (ΩAr), er det høyere ΩAr i de sørlige delene enn i de nordlige, med en tydelig undermetning av ΩAr (ΩAr <1) i 2013 både sommer og vinter (Jones mfl. 2020).
Foreslått SVO Eggakanten overlapper delvis med de foreslått endrete SVO-ene Iskantsonen og Havområdene rundt Svalbard i de nordlige og nordøstlige delene og i områder vest av Svalbard. Beskrivelsen av havis i disse delene av foreslått SVO Eggakanten nord reflekterer derfor havisen i disse overlappende SVO-ene. Flerårsis fra Polhavet kan opptre særlig nord og nordøst for Svalbard (Smedsrud mfl. 2013, Onarheim mfl. 2014, 2017, Hop mfl. 2019). I foreslått SVO Eggakanten nord er det observert drivende havis fra Polhavet langs vestkysten av Svalbard, området nord for Svalbard (sørøstlige deler av Yermak-platået) og videre nordøstover. Vestspitsbergenstrømmen (fortsettelsen på atlanterhavsstrømmen) bringer varmt atlanterhavsvann langs vestkysten og sokkelen nord for Spitsbergen og holder ellers området her isfritt store deler av året, og helt isfritt noen år (se f.eks. Svendsen mfl. 2002). Om vinteren er isen langs vestkysten av Spitsbergen typisk førsteårsis og ulike typer unge isklasser.
Lengst sør har Eggakanten stor betydning for strømmene som transporter egg, larver, yngel og annet plankton inn mot de foreslåtte SVO-ene Kystsonen Lofoten og Tromsøflaket og videre ut i Barentshavet (Sundby 2013).
Figur 8.5.2. Gjennomsnittlig overflatestrøm (piler) og bunndyp (farger) i forslag til endret SVO Eggakanten nord og omkringliggende områder.
8.5.5 - Beskrivelse av miljøverdiene
Plankton
Den sterke strømmen langs eggakanten fører til vertikale blandingsprosesser som øker konsentrasjoner av næringssalter og dermed primærproduksjon. Strømmene gjør også at området er en svært viktig transport vei for næringssalter, planteplankton (Vernet mfl. 2019) og dyreplankton (Wassmann mfl. 2019, Gluchowska mfl. 2017, Basedow mfl. 2018) fra Norskehavet og opp til vest-Spitsbergen, inn i Framstredet og videre til Polhavet nordøst for Svalbard. Planteplankton hotspots har lenge blitt påvist i forbindelse med eggakanten, for eksempel nord for Andøya (Slagstad mfl. 1999), og en modell har nylig vist at adveksjon av planteplankton nordover langs eggakanten fra Norskehavet til Svalbard er en viktig del av primærproduksjonen i Barentshavet og rundt Svalbard, og fører til en dobling av primærproduksjonen vest for Svalbard kontra Framstredet (Vernet mfl. 2019). Den atlantiske hoppekrepsarten Calanus finmarchicus (raudåte) dominerer dyreplanktonbiomassen i vannet som transporteres nordover fra Norskehavet til Svalbard og Polhavet og modeller anslår at hele 1,5 millioner tonn karbon C. finmarchicus forlater sokkelen ved Nord-Norge og transporteres nordover hvert år (Wassmann mfl. 2019). Den topografiske styringen av havstrømmene gjør at det er høyere konsentrasjoner av C. finmarchicus ved eggakanten utenfor nord-Norge enn i havområdene mot vest (Edvardsen mfl. 2006). I de atlantiske vannmassene som strømmer nordover ved eggakanten er mengden av dyreplankton dermed høy, noe som er viktig for marine dyr lengre nord. Det at denne biomassen fornyes kontinuerlig via nye innstrømmende vannmasser gjør at beitende dyr har tilgang til opptil 12 ganger så mye dyreplankton i løpet av et år i områdene rundt eggakanten nordvest for Svalbard enn områder uten like mye adveksjon og gjør at områdene er svært viktig for beitende fisk, sjøfugl, og marine pattedyr (Wassmann mfl. 2019). Denne nordgående havstrømmen ved eggakanten er en unik transportvei til Polhavet for næringssalter og biologisk biomasse på den atlantiske siden. Sammen med en lignende nordgående havstrøm på Stillehavssiden av Polhavet, utgjør disse tilførsler hele 20 prosent av produktiviteten i Polhavet (Popova mfl. 2013). Generelt vil høyere breddegrader føre til reduksjon i planteplanktonbiomassen (Vernet mfl. 2019) og dyreplanktonbiomassen (Wassmann mfl. 2019, Carstensen mfl. 2019) , mens i Vestspitsbergenstømmen skjer denne reduksjonen lengre nord og øst enn i andre områder . Dette støtter oppunder at produktiviteten i innstrømmende vannmasser er høyere enn for øvrig ved høye breddegrader.
Klimaendringer har økt den atlantiske innstrømningen til Framstredet og Polhavet (Muilwijk mfl. 2018), og det er påvist en økende tetthet av atlantiske raudåte (C. finmarchicus) i Vestspitsbergen-strømmen (Hop mfl. 2019, Carstensen mfl. 2019) og en utvidelse av atlantiske planteplanktonarter nordover (Oziel mfl. 2020).
Fisk
Eggakanten er et viktig gyte- eller yngleområde for flere kommersielt og økologisk viktige fiskearter, blant annet vanlig uer (Sebastes norvegicus) (F2), snabeluer (Sebastes mentella) (F3), blåkveite (Reinhardtius hippoglossoides) (F4) og hyse (F1), se figur 8.5.1 (Drevetnyak og Nedreaas 2009, Albert og Vollen 2015). Blåkveita er en flatfisk og har et aktivt levesett med migrasjoner både vertikalt og horisontalt, men forekommer sjelden i vann varmere enn ca. 4 °C (Vollen og Albert 2008). Mer enn 40 byttedyrgrupper er funnet i magen til blåkveite, men fisk dominerer (hovedsakelig lodde og polartorsk) i tillegg til blekksprut, reker og avfall fra fiskebåter. Når fisken blir større, forsvinner de minste byttedyrgruppene (reker og lodde) og andelen torsk (Gadus morhu a), hyse (Melanogrammus aeglefinus) og fiskeavfall øker (Dolgov and Smirnov 2001, Hovde mfl. 2002, Vollen mfl. 2004).
Blåkveita er en langlivet art som kun tåler lav beskatning. Det viktigste gyteområdet (F4, figur 8.5.1) er lokalisert til midtre del av foreslått SVO Eggakanten nord sør for Bjørnøya. Den nordøstarktiske blåkveitas hovedgyting foregår på dypt vann (500–800 m) om høsten og vinteren på eggakanten mellom 70°N og 75°N (Albert mfl. 2001). Egg og larver driver med strømmen avhengig av hvor gytingen foregår. De siste ti årene er hoveddelen av egg og larver blitt ført nordover langs Svalbard og østover mot Frans Josef land, inn i oppvekstområdet i det nordlige Barentshavet. Bestanden gikk dramatisk ned i 1970-årene, og kvotereguleringer ble innført i 1977. I 1992 ble fiske etter blåkveite forbudt, med unntak av forskningsfiske og et begrenset norsk kystfiske. Forbudet ble opphevet i 2009. Den fangstbare mengden blåkveite (fisk >45 cm), økte først, men har gått noe tilbake etter 2014 (ICES 2015, ICES 2020 a).
Snabelueren lever langs kontinentalskråningen mot Norskehavet fra 400 til 600 meters dyp nordover til Andøya (Cadrin mfl. 2010). I Barentshavet er det vist “gytevandring” av hunnfisk mot eggakanten (figur 8.5.1). Her finnes det lite snabeluer mindre enn 28–30 cm. Snabeluer yngler i mars–april, og den sterkt truede (EN) vanlig ueren yngler i april-juni med hovedtyngde i mai. Parringen foregår om høsten langs eggakanten . Yngleområdet strekker seg langs eggakanten fra Shetland til Tromsøflaket. Kyststrømmen og atlanterhavsstrømmen sørger for at larver og yngel av disse arter transporteres nordover og inn i Barentshavet mens de blir forsynt med plankton.
Mesopelagisk fauna
Eggakanten og områdene omkring har større mengde av de mesopelagiske fiskene nordlig lysprikkfisk (Benthosema glaciale), laksesild (Maurolicus muelleri) o g liten laksetobis (Arctozenus risso) sammenliknet med mer vestlige deler av Norskehavet (Dalpadado mfl. 1998, Salvanes 2004).
Der finnes potensielt gyte/oppvekstområde for isskate (Amblyraja hyperborea) basert på funn av store eggforekomster (Lynghammar 2014, Forsberg 2018).
Det er stor sesongmessig variasjon i mesopelagiske samfunn nord fra Svalbard med manet (Cyanea), polartorsk, laksetobis (Arctozenus risso, Paralepis coregonoides), og lysprikkfisk (Myctophidae arter) dominerende om sommeren sammen med veldig stor mengde amfipoder (T. libellula) og atlantiske krillarter. Om vinteren dominerer krill, snabeluer og Cyanea. Biomassen er 15–20x høyere om sommeren (Geoffrey 2019).
Bunnsamfunn
Dette området inneholder flere sårbare naturtyper: Dypvannssjøfjær (Umbellula encrinus), grisehalekorallskog (Radicipes gracilis), Lophelia -rev, hardbunnskorallskog, og kaldtvanns-svampsamfunn (dominert av glassvamp) (figur 8.5.3) .
Figur 8.5.3. Kart med ulike bunnsamfunn i den sørlige delen av foreslått SVO Eggakanten nord. Grensene for det foreslåtte SVO-et er vist med tykk rød strek, mens grenser for overlappende og nærliggende foreslåtte SVO-er er vist med tynnere rød strek. Grensene for forvaltningsplanområdene er markert med brun linje.
Den kjente utbredelsen av grisehalekorallskog i Norge er begrenset til et lite område, Bjørnøyaraset, nordvest for Tromsøflaket. Denne naturtypen er rødlistet. Lophelia -revene i området forekommer i den sørlige delen, nord til ytre del av renna kalt Malangsdjupet. Lenger mot sør er deler av rev-området i Hola (utenfor Vesterålen) også innenfor dette SVO-et, men de aller største forekomstene av Lophelia-rev finner vi på sokkelkanten, sørvest av Røstbanken i korallrevområdet kalt Røstrevet. Dette korallrevområdet er om lag 45 km langt og er vernet mot fiske med aktivt bunnredskap (Mareano.no).
Overlappet med de foreslåtte SVO-ene Iskantsonen og Havområdene rundt Svalbard fører til at beskrivelsene av megabentosfauna som er beskrevet der, også er dekkende for SVO Eggakanten nord, med rik variasjon av svamper (Jørgensen mfl. 2015 a,b, 2019).
Sjøpattedyr
Romlig fordeling av observasjoner på norske hvaltelletokt indikerer at eggakanten kan være viktig habitat for spermhval (Physeter macrocephalus) og finnhval (Balaenoptera physalus) (Leonard og Øien 2020, Skern-Mauritzen mfl. 2009; se også forslag til endret SVO Havområdene rundt Svalbard, figur 8.3.1). Eggakanten og den øvre sokkelskråningen er også kjent for å være viktig beitehabitat for klappmyss (Cystophora cristata) som sannsynligvis beiter på blåkveite og andre dyptlevende pelagiske arter (Folkow og Blix 1999), selv om dette kan ha endret seg noe i senere tid da nyere studier viser at klappmyss beiter mer over sokkelområdene mellom Øst-Grønland og Norge (Vacquie-Garcia mfl. 2017 a).
Sjøfugl
Foreslått SVO Eggakanten nord overlapper med mange av områdene som allerede er nevnt, og med områder som nevnes senere. For sjøfuglene er det foreslåtte SVO Eggakanten nord generelt viktig i hekkesesongen (Christensen-Dalsgaard mfl. 2018), særlig for pelagisk beitende arter som alkefugl og krykkje (Rissa tridactyla), men for hekkebestandene er disse områdene allerede dekket inn gjennom utvidelse av de andre områdene. Sjøfuglkoloniene legges gjerne i områder med stabil og forutsigbar næringstilgang i form av fiskelarver og annet som driver forbi, og de store sjøfuglkoloniene er knyttet opp mot slike områder (Sandvik mfl. 2016). Områder som dekker slike funksjoner i hekkesesongen omfatter eggakanten vest for Bjørnøya, langs vestkysten av Svalbard og langs Norskekysten (Systad mfl. 2019). Utenom hekkesesongen brukes hele eggakanten av en rekke pelagiske beitende arter, f.eks. lunde (Fratercula arctica) og krykkje (https://seatrack.seapop.no).
8.6 - Forslag til endret SVO Kystsonen Finnmark (BH4)
8.6.1 - Sammendrag
Områdebeskrivelse og strukturer
Forslag til endret SVO Kystsonen Finnmark (BH4) ligger nord for Finnmarkskysten ved kanten av kontinentalsokkelen sørvest i Barentshavet og strekker seg fra Tromsøflaket til grensen mot Russland og 100 km ut i havet. Kyststrømmen som følger skråningen utenfor kysten østover, er den sentrale transportåren i området. Der hvor skråningen er bratt ligger kyststrømmen tett mot kysten. Kyststrømmen påvirkes av grunne banker omgitt av områder med større dyp og det oppstår stasjonære virvler som øker oppholdstiden i noen områder. Området omfatter en rekke fjorder med unike miljøforhold der de munner ut mot Barentshavet.
Miljøverdier
Området er et av de viktigste hekkeområdene for sjøfugl på fastlandet, med høye andeler av en rekke norske bestander som beiter på fiskelarver og –yngel som passerer gjennom området. Området er også et viktig overvintringsområde for havdykkender, lommer og måker fra andre områder i Arktis. Stellerand er den mest sjeldne dykkand i verden, og hele 5–10 prosent av hele verdensbestanden overvintrer i Varangerfjorden. Området er også myteområde for norske og russiske bestander av ærfugl, praktærfugl og andre havdykkender. Området er viktig for lunde (sårbar-VU), lomvi (kritisk truet-CR), alke (sterkt truet-EN) og krykkje (EN), samt de store måkene som nå er i nedgang, uten at dette er reflektert i rødlisten.
Området er hovedgyteområde for lodde, som er en nøkkelart i økosystemet. Lodde gyter fra begynnelsen av mars, ved å legge eggene på bunnen. Larvene driver i de øverste 50 m med kyststrømmen østover og etter hvert nordover ut i Barentshavet sammen med store mengder plankton, egg og larver fra andre fiskearter (f.eks. sild, torsk, hyse, sei, uer) i løpet av vår/tidlig sommer.
Området har flere viktige yngleområder for havert og viktige habitater for steinkobbe. I de grunne områdene, og nær kysten, finnes det kaldtvannskoraller.
Porsangerfjorden har polare forhold og økosystem innerst i fjorden, med arter som ellers bare finnes lenger nord i Barentshavet, inkludert polartorsk.
Lodde. Foto: Maria Tennngen, HI
Figur 8.6.1. Kart over forslag til endret SVO Kystsonen Finnmark (BH4). De foreslåtte grensene for SVO-et er vist med tykk rød strek, mens grenser for nærliggende foreslåtte SVO-er er vist med tynnere rød strek. Grensene for forvaltningsplanområdet er markert med brun linje. Kartet viser utbredelsen av utvalgte miljøverdier som er av betydning for området. Kandidatområdene for pelagisk beitende sjøfugl er skravert, og utbredelsen av miljøverdier knyttet til fisk (gytefelt), bunnsamfunn og sjøpattedyr er vist med fargefelt.
Ung krykkje. Foto: Erik Olsen, HI
8.6.2 - Vurdering i forhold til EBSA-kriteriene – Forslag til endret SVO Kystsonen Finnmark (BH4)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til endret
SVO Kystsonen Finnmark (BH4)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet /sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Fisk
Sjøfugl
Strømrike lokaliteter mellom 25-75 m med sand/grussubstrat er viktige for loddegyting i mars-april (Gjøsæter 1998, Gjøsæter mfl. 2011, Sundby mfl. 2013).
Et av de viktigste hekkeområdene for sjøfugl på fastlandet, med høye andeler av en rekke norske bestander (Systad mfl. 2019), som beiter på fiskelarver og -yngel som passerer gjennom området (Sandvik mfl. 2016).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Fisk Sjøpattedyr Sjøfugl
Området er et viktig gytefelt for lodde (Gjøsæter 1998, Gjøsæter mfl. 2011, Sundby mfl. 2013, Sundby mfl. 2013) og en viktig transportvei for fiskelarver (sild, torsk, hyse, sei, uer og lodde) (Gjøsæter 1998, Dragesund mfl. 2008, Castano-Primo mfl. 2014, Sundby mfl. 2013).
Områdene utenfor Sørøya, Gjesvær, ytre Porsangerfjord og ytre Laksefjord og særlig Kongsfjorden er viktige yngleområder for havert (Nilssen og Bjørge 2019). Områdene rundt Sørøya, indre fjordområder i Porsangerfjord, Tanafjord og Kongsfjord er viktige habitater for steinkobbe (Nilssen og Bjørge 2019). Varangerfjorden er viktig habitat for niser (NAMMCO og IMR 2019).
Gjesværstappan er nå den største lundekolonien i Norge med mer enn 300 000 par hekkende lundefugl her. Rundt 80-100 000 hekker på Hjelmsøya like ved, og noen mindre kolonier i Kongsfjorden, Syltefjord og Hornøya (Systad mfl. 2019). Området er også et viktig overvintringsområde for havdykkender, lommer og måker fra andre områder i Arktis (Systad mfl. 2019).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Sjøfugl
Området er viktig for lunde (sårbar-VU), lomvi (kritisk truet-CR), alke (sterkt truet-EN) og krykkje (EN), samt de store måkene som nå er i nedgang, uten at dette er reflektert i rødlisten (Henriksen og Hilmo 2015).
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Sjøfugl
Langtlevende sjøfugl har lav restitusjonsevne og er utsatt for en rekke påvirkningsfaktorer, inkludert oljesøl, næringssvikt og klimaendringer. Sjøfuglene har opplevd en omfattende hekkesvikt samt langvarig bestandsnedgang, dette gjelder særlig pelagisk tilknyttede arter som lomvi og krykkje. Lundefuglene i dette området, samt lomvi på Hornøya, og den delen av lomvibestanden som hekker i steinur på Hjelmsøy, er unntak som bekrefter regelen (Systad mfl. 2019, Christensen-Dalsgaard 2020).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Plante-plankton
Dyreplankton Fisk
Høy produksjon av dyreplankton i området vil være viktig mat for fisk (Dalpadado mfl. 2014, ICES 2017, 2018, 2019, 2020 a). Fjordområdene vil ha høy produksjon av planteplankton, særlig på våren og sommeren (ØKOKYST).
Området er viktig for larver og yngel for flere av de store fiskeressursene i Barentshavet (Bergstad mfl. 1987, Solemdal mfl. 1997, Hylen mfl. 2008, Dragesund mfl. 2008, Sundby and Nakken, 2008, Sundby mfl. 2013, Eriksen mfl. 2011, 2012). Store deler av populasjonen av modnende lodde migrerer normalt inn til dette området for å gyte og dø, og gjør energi tilgjengelig for organismer på høyere trofisk nivå (Bogetveit mfl. 2008, Mikkelsen og Pedersen, 2017).
Sjøfugl har betydning for transport mellom hav og land, både i form av gjødslingseffekter og som næring for predatorer på land (Zmudczyńska-Skarbek mfl. 2017, Prop mfl. 2015).
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Bunnsamfunn
Fisk
Sjøfugl
Lodde er viktig mat for mange sjøfugl-, fiske- og sjøpattedyrarter og bidrar til biologisk biomangfold i området (Solemdal mfl. 1997, Hylen mfl. 2008, Dragesund mfl. 2008, Sundby and Nakken, 2008, Sundby mfl. 2013, Eriksen mfl. 2011, 2012, Systad mfl. 2019).
I de grunne områdene, og nær kysten, finnes det kaldtvannskoraller (Mareano.no).
Området har høy diversitet når det gjelder sjøfugl gjennom hele året (Systad mfl. 2019).
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
8.6.3 - Lokalisering
Forslag til endret SVO Kystsonen Finnmark dekker det eksisterende SVO Kystsonen Finnmark, sist oppdatert i Meld. St. 20 (2019-2020), men grensen for SVO-et er foreslått utvidet ved at kandidatområdene for sjøfugl som strekker seg 100 km ut fra de viktigste hekkekoloniene langs kysten er inkludert, og områder innenfor grunnlinjen er innlemmet av samme grunn.
Området ligger nord for Finnmarkskysten ved kanten av kontinentalsokkelen og strekker seg fra Tromsøflaket til grensen mot Russland (figur 8.6.1).
Forslag til endret SVO Kystsonen Finnmark dekker også delvis to områder som er foreslått som marine verneområder, indre del av Porsangerfjorden og Transekt fra Tanafjorden.
8.6.4 - Introduksjon til området
Kyststrømmen som følger skråningen utenfor k ysten østover er den sentrale transportåren i området (Fig. 8.6.2). Der hvor skråningen er bratt ligger Kyststrømmen tett mot kysten (Sandvik mfl. 2016). Kyststrømmen påvirkes av grunne banker omgitt av områder med større dyp og det oppstår stasjonære virvler som øker oppholdstiden i noen områder (Sandvik mfl. 2016). De lange og stedvis dype fjordene har i tillegg ulike former for vannkvaliteter i form av sjiktninger med ulik saltholdighet og temperatursoner, som gir opphav til svært spesielle levemiljøer.
Den sesongmessige variasjonen i karbon og næringssalter er hovedsakelig drevet av store variasjoner i vannmasser og saltholdighet. Dette er et resultat av svingninger i forholdet mellom ferskvannstilførsel fra den va rme norske kyststrømmen som har lav karbon/CO2 og alkalinitet og dessuten høy pH i de øvre 50 m, elvevann fra land nær kystsonen og tilførsel av det salte, varme atlanterhavsvannet med høy karbon/CO2 og alkalinitet (Chierici mfl. 2017). Havområdet tar opp atmosfærisk CO2, og det største opptaket skjer i mai og september på grunn av biologisk CO2-opptak (Lauvset mfl. 2013). En minking av pH i overflaten med 0,001 per år er beregnet i perioden 1998-2016 (Becker mfl. 2020). Dypere ned i vannsøylen, under 50 m, er pH og aragonittmetning lavere enn i overflaten (Chierici mfl. 2017). Korallrevene har samme karbonatkjemiske tilstand (aragonittmetning ca. 1,7 og DIC lavere enn 2170 µmol/kg) som bunnvannet som er funnet på andre steder i Atlanteren (Järnegren og Kutti 2014, Jones mfl. 2020).
Figur 8.6.2. Gjennomsnittlig overflatestrøm (piler) og bunndyp (farger) i forslag til endret SVO Kystsonen Finnmark og omkringliggende områder.
8.6.5 - Beskrivelse av miljøverdiene
Plankton
De kystnære områdene av det sørlige Barentshavet (sør for 72°N) inneholder både høy biomasse og høy modellert sekundærproduksjon av Calanus finmarchicus sammenlignet med Barentshavet generelt (i 1998 og 2006 over 7 g tørrvekt per m2 biomasse og opp mot 60 g tørrvekt per m2 år produksjon enkelte plasser ved kysten sammenlignet med 0-40 g tørrvekt per m2 perår ellers i Barentshavet) (Dalpadado mfl. 2014). Det er få studier som sammenligner kystområdene med utenforliggende havområder. Data sammenstilt av Dalpadado mfl. (2014) indikerer at det er høyere planteplanktonbiomasse i de kystnære områdene. I Oziel mfl. (2017), er det derimot estimater som tyder på at planteplankton-biomassen kan være like høy eller høyere i det åpne Barentshavet. Innenfor fjorden og mellom fjordene vil det være forskjell i produksjon (Walday mfl. 2020), der fjorder med avrenning har noe høyere produksjon.
Nyere biomassemålinger støtter opp under at dette er et viktig område i forhold til dyreplankton- biomasse, med relativt høye verdier som er tilsvarende flere andre steder i Barentshavet (ICES 2017, 2018, 2019, 2020 a). I vurderinger av hvor mye dyreplankton som er tilgjengelig for høyere trofiske nivåer som fisk, så er det både sekundærproduksjon (hvor mye biomasse som er produsert før det blir beitet på) og hvor mye planktonbiomasse som blir tilført med kyststrømmen, som har betydning. I dette området er havstrømmene relativt sterke (Vernet mfl. 2019). Dermed kan tilførselen av dyreplankton være høyere her enn i atlantershavsstrømmen lengre ute.
Dyreplankton i området domineres av C. finmarchicus, som transporteres nord- og østover hver sommer fra store overvintringspopulasjoner i Norskehavet og forbi foreslått SVO Kystsonen Finnmark (Tande 1989, Halvorsen mfl. 2003, Aarflot mfl. 2018). Bestandsstørrelsen av C. finmarchicus øker i Barentshavet (Aarflot mfl. 2018, ICES 2020 a). De fleste dyreplanktonartene som opptrer i det foreslåtte SVO-et finnes også utenfor området. Det som advekteres forbi har derimot god primærproduksjon i området og nærliggende områder å forsyne seg av (Vernet mfl. 2019), og dermed gode vekstvilkår. Det er ingen informasjon om andre dyreplanktonarter enn C. finmarchicus. Utover Havforskningsinstituttet sine årlige tokt i Barentshavet, er det generelt mindre forskning i dette området enn ellers i Norskehavet, Lofoten til Tromsøflaket, og rundt Svalbard.
Fisk
Lodde (Mallotus villosus) er en pelagisk stimfisk som lever hele sitt korte liv i Barentshavet (Gjøsæter 1998, Orlova mfl. 2005, 2011). Den modnende delen av bestanden, som består av 3–5 år gammel fisk vandrer mot kysten for å gyte, som oftest i begynnelsen av mars. Lodda gyter langs kysten av Troms, Finnmark og Kola, på ganske grunt vann (20 til 90 m dyp), på bunnsubstrater av sand, singel og grus, og derfor er foreslått SVO Kystsonen Finnmark et viktig gytefelt (F1, se figur 8.6.1) for lodde (Gjøsæter 1998, Gjøsæter mfl. 2009). Det foregår også en del gyting seinere på året, såkalt sommergyting, da helst i Øst-Finnmark og lengre øst på russisk side av grensa (F1). Lodda er hovedsakelig engangsgyter, dvs. at det meste av lodda dør like etter gyting (Gjøsæter 1998, Gjøsæter mfl. 2009, 2011). De klebrige eggene er festet til bunnsubstratet og klekkes etter 3 til 4 uker. Larvene stiger opp i det øvre laget (0-50 m) og driver med s trømmen østover og sprer seg etter hvert nordover ut i Barentshavet. Lodde er en nøkkelart i Barentshavets økosystem hele året. Gytelodde er viktig ved kysten som mat for både fisk og sjøpattedyr om vinteren (Gjøsæter 1998, Gjøsæter mfl. 2009, 2011, Eriksen mfl. 2020) . På grunn av det korte livsløpet og enkelte år med svikt i rekrutteringen, fluktuerer loddebestanden voldsomt. Det har være observert fire bestandskollapser siden 1971, der loddelarver og -yngel er beitet ned av ungsild (Clupea harengus) (Huse og Toresen 2000, Hallfredsson og Pedersen 2009, Gjøsæter mfl. 2016). Fiskeriet etter lodde er stengt i kollapsperiodene. I perioder med høyere bestand har man tillatt et fiskeri hvor kvotene settes med hensyn til matbehovet til torsk (Gadus morhua) (Gjøsæter mfl. 2009). I de siste 15 årene har en hatt en stor torskebestand, og 360 000 tonn er det høyeste kvantum lodde fisket pr år. Det ble ikke tilrådd fiske etter lodde i Barentshavet i 2019-2021 pga. lav bestand (ICES 2020 b). SVO Kystsonen Finnmark er også et svært viktig område for transporten av yngel (sild, torsk, hyse (Melanogrammus aeglefinus), delvis sei (Pollachius virens)) og larver (lodde, torsk, sild, hyse og delvis sei) fra gytefelt til Barentshavet om våren (Olsen mfl. 2010). I hovedsak foregår driften i de øverste 50 m av sjøen, men kan strekke seg ned mot 100 m.
Fjorder (Kvænangen, Porsanger- og Tanafjorden) er mangfoldige og både voksen og yngre torsk, (inkludert kysttorsk som er en nøkkelart), hyse, sei, vanlig uer (Sebastes norvegicus), lodde og sild er til stede hele året (Søvik mfl. 2020). Yngre stadier av fisk er et godt matgrunnlag for sjøfugl i området (se sjøfugl beskrivelser nede).
Bunnsamfunn
I de grunne områdene og nær kysten, finnes kaldtvannskoraller (Mareano.no).
I 2018-2019 ble det gjennomført bunntråltokttokt med vekt på dypvannsreke (Pandalus borealis) i Kvænangen, Tana- og Porsangerfjorden (Søvik mfl. 2020). Hovedformålet var å kartlegge fjordøkosystemene i Tana- og Porsangerfjorden som har vært stengt for bunntråling siden tidlig på 1970-tallet. Rekebestandene i Kvænangen, ytre Porsangerfjorden og Tanafjorden er i stor grad sammenlignbare til tross for at de to Finnmarksfjordene har vært u-trålte i nærmere 50 år, mens Kvænangen hele tiden har hatt et aktivt rekefiske (Båtevik 2020, Søvik mfl. 2020). Rekene i indre Porsangerfjorden skiller seg ut med en svært høy tetthet og en avvikende bestandsstruktur. Genetiske studier viser at reker i Barentshavet og fra kyststrekningen Trøndelag til Troms (inkludert Kvænangen) utgjør to forskjellige biologiske populasjoner, mens fjordene i Finnmark inneholder en blanding av reker fra kysten og fra Barentshavet (Hansen 2020). Stort sett alle kommersielle rekefelt langs norskekysten har blitt kartlagt (fiskeridir.no). Ut ifra det rekefiskere forteller, kan det se ut som om de forskjellige livsstadiene delvis befinner seg på ulike felt og dyp. Noen felt ser ut til å være viktige på ulike tider av året, mens andre felt ser ut til å være viktige året rundt. Dette inntrykket bekreftes av studier av andre rekebestander (Shumway mfl. 1985). Se også omtalen av dypvannsreke under foreslått SVO Tromsøflaket (Kap. 8.7.5).
Rød kongekrabbe (Paralithodes camtschaticus) er etablert i området siden siste del av forrige århundre (Hjelset 2014, ICES 2020 a). Den har satt preg på bunnsamfunnet på permanent basis (Falk-Pedersen mfl. 2011).
Sjøpattedyr
Produksjonen av kystselen havert (Halichoerus grypus) har vært stabil på om lag 200 unger årlig (Nilssen og Bjørge 2019). Områdene utenfor Sørøya, Gjesvær, ytre Porsangerfjord og ytre Laksefjord og særlig Kongsfjorden er viktige yngleområder for havert. Haverten danner kolonier, særlig i forbindelse med ungekasting (fødsel) og parring (september–desember) og hårfelling (februar–april). I resten av året er de spredt langs store deler av kysten.
Steinkobbe (Phoca vitulina) , vår mest tallrike kystsel, er en liten selart, samles på land i forbindelse med forplantning (juni-juli) og hårfelling (august), og de legger seg helst opp på litt beskyttede lokaliteter i skjærgården (skjær og sandbanker som tørrlegges ved fjære sjø) (Nilssen og Bjørge 2019). I yngle- og hårfellingstiden fra midten av juni til begynnelsen av september er steinkobbene svært stedbundne og knyttet til stedet der de ble født. Fra andre del av september til tidlig juni, er det næringssøk som har høyeste prioritet. I denne perioden vil de oppsøke steder med god tilgang på mat, ofte mange titalls kilometer fra stedet der de ble født.
Områdene rundt Sørøya, indre fjordområder i Porsangerfjord, Tanafjord og Kongsfjord er viktige habitater for steinkobbe og bestanden teller i underkant av 1850 dyr (Nilssen og Bjørge 2019).
Sjøfugl
Kyststrekningen fra Tromsøflaket til Russegrensen huser en svært stor andel av norske sjøfugl.
Gjennom overvåkingsprogrammene Seapop og SEATRACK er det nå ny kunnskap til stede om utstrekningen av beiteområdet rundt sjøfuglkoloniene i hekketiden (Systad mfl. 2019). Sjøfugl beiter ofte ut til 100 km fra kolonien. Dominerende arter er lunde (Fratercula arctica), lomvi (Uria aalge), alke (Alca torda) og krykkje (Rissa tridactyla) av de pelagisk beitende artene, og ærfugl (Somateria mollissima), toppskarv (Phalacrocorax aristotelis) og storskarv (Phalacrocorax carbo) av de bunnbeitende artene. De største fuglefjellene omfatter Gjesværstappan med over 300 000 par lunde, forekomster av lomvi, alke og den største havsulekolonien (Morus bassanus) i Nordnorge, Hjelmsøy med nærmere 80-100 000 par lunde, 15 000 par lomvi samt forekomster av alke, krykkje, havsvale (Hydrobates pelagicus), storjo (Stercorarius skua) med flere, Hornøya med den største lomvikolonien på fastlandet, Syltefjordstauran og Ekkerøy, begge de siste store krykkjekolonier. Overflatebeitende arter som gråmåke (Larus argentatus), svartbak (Larus marinus) og tyvjo er relativt tallrike i dette området. Teist (Cepphus grylle) har sitt viktigste hekkeområde i Norge i dette området (Systad mfl. 2019).
I området er det en positiv bestandsutvikling for arter som har negativ bestandsutvikling lengre sør (Systad mfl. 2019). Unntaket er krykkje, som har hatt en svært dårlig utvikling, og flere kolonier er redusert kraftig eller blitt borte.
Varangerfjorden og kysten av Varangerhalvøya ut til Hamningberg er et viktig overvintringsområde for bl.a. stellerand (Polysticta stelleri), ærfugl (russiske bestander), praktærfugl (Somateria spectabilis) og havelle (Clangula hyemalis) og myteområde for norske og russiske bestander av ærfugl, praktærfugl og andre havdykkender. Stelleranda er den mest sjeldne dykkand i verden, og 5–10 prosent av verdensbestanden overvintrer i dette området.
Figur 8.6.3. Kart viser fordeling av hekkende sjøfugl (i farger) med pelagiske beiteområder (skravert felt) i foreslått SVO Kystsonen Finnmark og nærliggende områder. De anbefalte grensene for SVO-et er markert med rød strek. Området er utvidet med tidligere foreslåtte kandidatområder i tråd med kunnskap om hvor store beiteområder de pelagiske sjøfuglene bruker (Systad mfl. 2019).
Figur 8.6.4. Kart viser fordeling av hekkende sjøfugl (i farger) med kystnære beiteområder i foreslått SVO Kystsonen Finnmark og nærliggende områder. De anbefalte grensene for SVO-et er markert med rød strek. Beiteområdene til disse artene er mer kystnære enn utstrekningen for SVO-et (Systad mfl. 2019).
Også i dette området har sjøfuglene opplevd en omfattende hekkesvikt samt langvarig bestandsnedgang, dette gjelder særlig pelagisk tilknyttede arter som lomvi og krykkje. Lundefuglene i dette området, samt lomvi på Hornøya, og den delen av lomvibestanden som hekker i steinur på Hjelmsøy, er unntak som bekrefter regelen.
Flere svært store krykkjekolonier er kraftig redusert eller forsvunnet siden 2009. Dette gjelder f.eks. Hjelmsøya, Gjesværstappan, Sværholt, Omgang og Syltefjordstauren . Disse koloniene utgjorde i 2009 majoriteten av hekkebestanden i Norge, men er nå redusert til en brøkdel. Fra årtusenskiftet til 2013 ble krykkjebestanden redusert med mer enn 80 prosent i dette området og er nå mindre enn 10 prosent av bestanden på 70-tallet (Systad mfl. 2019).
Lomvi har nærmest sluttet å hekke på åpne hyller i de vestlige delene av dette området (Hjelmsøya, Gjesværstappan), tilsvarende utviklingen lenger sør, og polarlomvi som kun hekker åpent, er på veg til å forsvinne som hekkefugl. Andelen lomvi som hekker skjult i steinur, øker imidlertid, men den samlede hekkebestanden er fremdeles lav sammenlignet med tidlig på 1980-tallet. I de østlige delene (Hornøya) har bestanden vokst, og der finnes det ennå en mindre bestand med hekkende polarlomvi. Imidlertid opplever krykkje-, lomvi og polarlomvibestandene på Hornøya nå en betydelig forstyrrelse fra havørn og kråkefugler, noe som har ført til manglende reproduksjon de siste to årene (Christensen-Dalsgaard 2020).
Vi har ikke god oversikt over de store lundekoloniene på Sørfugløy og Nordfugløy i Troms, og heller ikke på Loppa, men kartleggingen utført av SEAPOP (https://seapop.no/) viser at disse koloniene ikke har hatt samme nedgang for denne arten som lenger sør. Det samme gjelder for Gjesvær og Hjelmsøy. Kolonien på Hornøya er imidlertid i vekst (Anker-Nilssen mfl. 2020).
Både lomvi og krykkje som hekker langs fastlandskysten, er sårbare for forstyrrelser fra en økende havørnbestand. I noen lomvikolonier observerer man at fuglene i økende grad hekker inne i steinurer der de er mer beskyttet for denne forstyrrelsen (Arneberg og Jelmert 2017).
Den observerte tilbakegangen i hekkebestandene av krykkje kan muligens være relatert til næringsforholdene. Man har sett at voksenoverlevelse hos krykkjer på Hornøya er knyttet opp mot tilgangen på lodde i hekketiden, og til en nakensnegl i overvintringsområdene ved Grand Banks utenfor Newfoundland (Reiertsen mfl. 2014).
De viktigste fuglekoloniene er lokalisert ved områder med naturlig opphoping av larver. Samlet sett er området det mest verdifulle området for larver sett under ett for torsk, sild, hyse og lodde, noe som har sammenheng med oseanografiske forhold (Sandvik mfl. 2016). Hvis klimaendringer endrer disse forholdene i så stor grad at det påvirker driften og retensjonsområdene for torskelarver, vil dette kunne få store konsekvenser for sjøfuglbestandene, ikke bare i Barentshavet, men langs hele kysten. Den viktigste faktoren i dette er imidlertid hvor larvene driver fra og timingen av larvedriften forbi koloniene.
Blomkålkorall og torsk. Foto: Jan Helge Fosså, HI
8.7 - Forslag til endret SVO Tromsøflaket (BH5)
8.7.1 - Sammendrag
Områdebeskrivelse og strukturer
Forslag til endret SVO Tromsøflaket (BH5) er et bankområde ved kanten av kontinentalsokkelen helt sørvest i Barentshavet. Her sprer kyststrømmen fra sør seg i to grener, én tett mot land og én som følger topografien rundt Tromsøflaket . Dette fører til forlenget oppholdstid for vannmassene (retensjonsområde) på Tromsøflaket. Variabiliteten i kyststrømmen er i stor grad knyttet til variasjoner i vind, og strømmen er sterkere om vinteren enn om sommeren. Om våren og sommeren, når vannmassene i området er lagdelt, har vinden sterk påvirkning på overflatesirkulasjonen. Dette påvirker utvekslingen mellom sokkel og hav, og vindretningen er avgjørende for driftsbaner og oppholdstid av vannet. Tidevannsstrømmene bidrar også til utvekslingen mellom sokkel og dyphav i dette området. Noe atlanterhavsvann kommer også inn i nordkant av Tromsøflaket, men mesteparten går på nordsiden.
Miljøverdier
Forslag til endret SVO Tromsøflaket er et særlig viktig transportområde/gjennomstrømmingsområde for plankton, egg og larver på vei videre øst og nord i Barentshavet, og oppholdstiden i dette området er lang på grunn av strømsystemet (retensjonsvirvel). Nordenden av Tromsøflaket er et viktig gyteområde for flekksteinbit. Lodde gyter inn til kysten i mars-april der eggene klistrer seg på bunnen i 3-4 uker. Loddelarver klekkes, stiger mot overflaten, og transporteres med strømmen. Fiskelarver (sild, torsk, hyse, sei, uer, lodde) i store mengder passerer gjennom området i de øverste 50 m i løpet av vår-tidlig sommer, og finner godt beite på banken. Store deler av en årsklasse av torsk og hyse passerer forbi her hver sommer. Yngleområdet for vanlig uer strekker seg langs eggakanten og nordover og østover inn i Barentshavet, langs hele det foreslåtte SVO Tromsøflaket. I fjordene gyter kysttorsken i beskyttede områder. Gytefelt for kysttorsk er delvis inne i det foreslåtte SVO-et, men siden indre grensesetting er basert hovedsakelig på de miljøverdiene som er knyttet til forvaltningsplanområdet utenfor, er gytefelt for kysttorsk ikke fullstendig integrert i de kystnære foreslåtte SVO-ene.
På Tromsøflaket er det rike bunnsamfunn, inkludert verdens nordligste kaldtvannskorallrev («Korallen» utenfor Sørøya), og omfattende bløtbunns-svampsamfunn. I Lopphavet er det dypvannsrenner og en stor korallfauna som er oppvekstområde for flere fiskeslag.
Opphoping av planktoniske organismer fører til særlig gode næringsforhold for en rekke hekkende og overvintrende sjøfugl, hvorav flere er rødlistet. Det er hekkekolonier av nasjonal betydning for lunde, lomvi og historisk også krykkje. Sørøya er en av de største lundekoloniene i landet. Området har noen av de største koloniene av toppskarv i Norge, og er også et viktig overvintringsområde for havdykkender, lommer og måker fra andre områder i Arktis.
Hval, særlig spekkhogger og knølhval, følger om vinteren sild som går inn i fjordene for å gyte. Kystsel beiter i området hele året.
Figur 8.7.1. Kart over forslag til endret SVO Tromsøflaket (BH5). De foreslåtte grensene for SVO-et er vist med tykk rød strek, mens grensene for det overlappende og de inntilliggende foreslåtte SVO-ene Eggakanten nord (BH3), Kystsonen Finnmark (BH4) og Kystsonen Lofoten (BH6) er vist med tynnere rød strek. Grensene for forvaltningsplanområdet er markert med brun linje. Kartet viser utbredelsen av utvalgte miljøverdier av betydning for området. Miljøverdier knyttet til fisk (ynglefelt for vanlig uer og snabeluer, og gytefelt for de øvrige artene) og sjøfugl er markert i øverste kart og miljøverdier knyttet til bunnsamfunn og sjøpattedyr (blant annet vinterbeiteområde for spekkhogger) er markert i nederste kart.
8.7.2 - Vurdering i forhold til EBSA-kriteriene – Forslag til endret SVO Tromsøflaket (BH5)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til endret
SVO Tromsøflaket (BH5)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet /sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Dyreplankton
Fisk Sjøfugl
Tromsøflaket er et retensjonsområde med generelt høye dyreplanktonkonsentrasjoner (Vikebø mfl. 2005, Broms 2017). Calanus finmarchicus, som bidrar mest til totalbiomassen i det vestlige Barentshavet, har unge stadier i mai (sammenheng med våroppblomstringen), og når høye sesongmessige verdier i juni-august da voksne bidrar mest til biomassen (Aarflot mfl. 2018).
Viktig transportvei for fiskelarver (sild, torsk, hyse, sei, uer og lodde) (Gjøsæter 1998, Vikebø mfl. 2005, Dragesund mfl. 2008, Castano-Primo mfl. 2014).
Lundebestanden i området utgjør ca. ¼ av bestanden i Norge. Området er også svært viktig for hekkebestandene av teist i Norge, og har solide bestander av gråmåke, svartbak, toppskarv og storskarv. Nordfugløy og Sørfugløy er to av de fem største lundekoloniene i Norge, området rundt nordenden av Sørøya er et av de viktigste hekkeområdene for toppskarv i Norge, og teistbestanden her utgjør en vesentlig del av bestanden totalt sett. Området er også et viktig overvintringsområde for havdykkender, lommer og måker fra andre områder i Arktis (Systad mfl. 2019).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Sjøpattedyr
Dyreplankton
Fisk Sjøfugl
Viktig næringsområde for fiskelarver/yngel, særlig torsk og sild (Bjørke og Sundby 1987).
Området er et viktig gytefelt for lodde (Bergstad mfl. 1987, Hylen mfl. 2008, Jakobsen and Ozhigin, 2011, Sundby mfl. 2013) og en viktig transportvei for fiskelarver (sild, torsk, hyse, sei, uer og lodde) (Gjøsæter 1998, Vikebø mfl. 2005, Dragesund mfl. 2008, Castano-Primo mfl. 2014).
Hekkekolonier av nasjonal betydning for lunde, lomvi, og historisk utnytter også krykkje banken som beiteområde (Anker-Nilssen mfl. 2015).
Hval, særlig spekkhogger og knølhval, følger om vinteren sild som går inn i fjordene for å gyte (Similä mfl. 1995), og det er selkolonier som beiter i området (Nilssen og Bjørge 2019).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Fisk
Sjøfugl
Vanlig uer har gytefelt der det foreslåtte SVO-et overlapper med foreslått SVO Eggakanten nord, mens kysttorsk har sine viktigste gytefelt inne i Lopphavet og ut i de kystnære områdene i det foreslåtte SVO-et (Olsen mfl. 2010, Husa mfl. 2020).
Svært viktig hekkeområde for truede sjøfuglarter, som alke (sterkt truet-EN), lunde (sårbar-VU), lomvi (kritisk truet-CR) og krykkje (EN) der det ser ut til at disse artene får fram unger i noen kolonier (Anker-Nilssen mfl. 2015). To av de fem største lundekoloniene i Norge finnes her, Sør-Fugløy (> 100 000) og Nordfugløy (>200 000 par) (Fauchald mfl. 2015).
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Bunnsamfunn
Sjøfugl
Fisk
Viktig transportvei for fiskelarver (sild, torsk, hyse, sei, uer og lodde) (Gjøsæter 1998, Dragesund mfl. 2008, Vikebø mfl. 2005, Castano-Primo mfl. 2014). Egg av hyse er sårbare for oljepåvirkning. (Søhus mfl. 2016).
Mareano har dokumentert rike svampsamfunn på mudder- og grusbunn med ulik artssammensetning på Tromsøflaket. Områdene har forholdsvis lite hardbunn. Svampsamfunnene bidrar til forholdsvis store forekomster av trollkreps, reke, rødpølse, anemoner, sjøstjerner (Paraniomorpha) og kråkeboller (Echinus). Den rike epifaunasamfunnene gir grunnlag for bl.a. steinbitfiskerier i området.
Bestandene av sjøfugl som lunde, alke, lomvi og krykkje er langtlevende og har lang restitusjonstid. Høye tettheter av toppskarv i området ved lille Kamøy ved nordenden av Sørøya er blant de største koloniene i Norge (Anker-Nilssen mfl. 2015).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Planteplankton Sjøfugl
Dyreplankton
Fisk
Særlig fjordområdene anses som områder med høy planteplanktonproduksjon (Degerlund og Eilertsen 2010).
Deler av området inneholder relativt høye konsentrasjoner av C. finmarchicus (Broms 2017, Aarflot mfl. 2018).
Området har betydning som larve- og yngelområde for flere av de viktige fiskeressursene i Barentshavet (Bergstad mfl. 1987, Solemdal mfl. 1997, Hylen mfl. 2008, Dragesund mfl. 2008, Sundby og Nakken, 2008, Sundby mfl. 2013, Eriksen mfl. 2011, 2012). Loddegyting om våren sørger for energitransport fra hav til kyst og økt lokal produksjon (Bogetveit mfl. 2008, Mikkelsen og Pedersen, 2017).
Sjøfugl har betydning for transport mellom hav og land, både i form av gjødslingseffekter (Anderson og Polis 1999) og som næring for predatorer på land (Prop mfl. 2015).
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Bunnsamfunn Sjøpattedyr
Fisk Sjøfugl
Det er et mangfold av fiskearter, som gyter eller som transporteres som egg (sild, torsk, hyse, delvis sei) og larver (lodde, torsk, sild, hyse, uer og delvis sei) fra gytefelt lenger sør og til Barentshavet om våren (Olsen mfl. 2010). Den rike epifaunasamfunnene gir grunnlag for bl.a. steinbitfiskerier i området.
Mareano har dokumentert rike svampsamfunn på mudder- og grusbunn med ulik artssammensetning på Tromsøflaket. Svampsamfunnene bidrar til forholdsvis store forekomster av trollkreps, reke, rødpølse, anemoner, sjøstjerner (Paraniomorpha) og kråkeboller (Echinus). Kaldtvannskorallrev (Desmophylllum pertusum, tidligere kalt Lophelia pertusa), er registrert på 24 lokaliteter i Tromsøflaket/Lopphavet-området.
Området har høy diversitet når det gjelder sjøfugl gjennom hele året, og bestandene er store (Systad mfl. 2019).
Det er en stabil ungeproduksjon hos kystselen (Nilssen og Bjørge 2019).
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
8.7.3 - Lokalisering
Forslag til endret SVO Tromsøflaket dekker det eksisterende SVO Tromsøflaket, sist oppdatert i Meld. St. 20 (2019-2020), men grensene for SVO-et er anbefalt utvidet mot nord for å dekke rike bunnsamfunn, og inn mot kysten for å bedre dekke sjøfuglenes beiteområder og viktige gytefelt for fisk (figur 8.7.1). I tillegg er området anbefalt utvidet sørover for å inkludere et område som er tatt ut av det foreslåtte SVO Kystsonen Lofoten, siden miljøverdiene i dette området naturlig hørte bedre hjemme i det foreslåtte SVO Tromsøflaket . Området som er utvidet korresponderer med beiteområdene for sjøfugl, primært lundefugl, men også alke og lomvi, som hekker på Sørfugløya (Systad mfl. 2019).
Tromsøflaket er et bankområde ved kanten av kontinentalsokkelen, rett nord for kysten fra Tromsø til Hammerfest. Området inkluderer også Lopphavet som er et 70 kilometer bredt åpent havstykke mellom Fugløya og Arnøya i Troms og Sørøya i Finnmark.
Tromsøflaket som særlig verdifullt og sårbart område overlapper delvis med foreslått SVO Eggakanten nord (figur 8.7.1).
Tromsøflaket dekker med sin nye indre grense, også tre marine verneområder, Rystraumen, Rossfjordstraumen og Ytre Karlsøy, i tillegg til Lopphavet som er foreslått som marint verneområde.
8.7.4 - Introduksjon til området
Tromsøflaket er et bankom råde ved kanten av kontinentalsokkelen helt sørvest i Barentshavet. Hovedkomponentene av havsirkulasjonen er kyststrømmen som brer seg rundt Tromsøflaket før den strømmer videre østover. Kyststrømmen har i dette området to grener, én tett mot land og én som følger topografien rundt Tromsøflaket (Skagseth mfl. 2011). Dette fører til forlenget oppholdstid for vannmassene på Tromsøflaket. Variabiliteten i kyststrømmen er i stor grad knyttet til variasjoner i vind, og strømmen er sterkere om vinteren enn om sommeren (Skagseth mfl. 2011). Om våren og sommeren, når vannmassene området er lagdelt, har vinden sterk påvirkning på overflatesirkulasjonen. Dette påvirker utvekslingen mellom sokkel og hav, og vindretningen er avgjørende for driftsbaner og oppholdstid for organismer i strømmen (Skardhamar og Svendsen 2005). Tidevannsstrømmene bidrar også til utvekslingen mellom sokkel og dyphav i dette området, og kan være viktig for bentiske dyr og prosesser (Skardhamar mfl. 2015). Noe atlanterhavsvann kommer også inn i nordkant av Tromsøflaket, men mesteparten går på nordsi den av dette bankområdet. Strømhastighetene i kyststrømmen (og atlanterhavsstrømmen) går betydelig ned når vannet kommer inn i Barentshavet (Sundby 2013).
Den sesongmessige variasjonen i karbon og næringssalter er hovedsakelig drevet av variasjoner i vannmasser og saltholdighet. Dette er et resultat av svingninger i forholdet mellom ferskvannstilførsel fra den varme norske kyststrømmen som har lav karbon/CO2 og alkalinitet, og dessuten høy pH i de øvre 50 m, elvevann fra land nær k ystsonen og tilførsel av det salte, varme atlanterhavsvannet med høy karbon/CO2 og alkalinitet (Chierici mfl. 2017). De øvre 50 m i overflaten viser en aragonittmetning på ca. 2,1 og pH på 8,05-8,10 (Lauvset mfl. 2016) med en forventet minking med økt CO2-opptak. Dypere ned i vannsøylen (under 50 m) er pH og aragonittmetning lavere enn i overflaten. En minking av pH med 0,001 per år er beregnet i perioden 1998-2016 (Becker mfl. 2020).
Figur 8.7.2. Gjennomsnittlig overflatestrøm (piler) og bunndyp (farger) i forslag til endret SVO Tromsøflaket og omkringliggende områder.
8.7.5 - Beskrivelse av miljøverdiene
Plankton
Når det gjelder planteplankton foreligger det data fra enkelte fjordområder som indikerer høy produksjon innen området. Planteplanktonproduksjonen i de kystnære områdene er i stor grad styrt av lokale hydrografiske og topografiske forhold. Planteplanktonet viser en sesongmessig dynamikk, med lav produksjon på vinteren og høy produksjon og biomasse på våren og sommeren (Velvin mfl. 2020). For de ytre sokkelområdene foreligger det lite systematisk sammenstilte data som belyser primærproduksjon. Satellittinformasjon indikerer imidlertid at de kystnære områdene vil ha høyere planteplanktonbiomasse enn de åpne havområdene (Johnsen mfl. 2009). Det foreligger dessuten flere studier fra fjordområdene i Troms (f.eks. Hegseth mfl. 1995, Archer mfl. 2000, Degerlund og Eilertsen 2010) som viser høy planteplanktonproduksjon i forbindelse med våroppblomstringen.
Dette området er lokalisert ved inngangen til Barentshavet, der plante- og dyreplankton blir transportert med kyststrømmen fra Norskehavet inn i Barentshavet (Vikebø mfl. 2005). Tromsøflaket er et retensjonsområde for fiskelarver og yngel og et viktig beiteområde for disse (Bjørke og Sundby 1987, Vikebø mfl. 2005). Raudåten Calanus finmarchicus dominerer dyreplanktonet og utgjør den største delen av dietten til ung torsk (Sundby 2000). Dyreplanktonbiomassen i dette området utgjør ca. 5-10 g tørrvekt per m2 (Broms 2017). I området nær Fugløya ved Tromsøflaket er tettheten av C. finmarchicus høy (ca. 30 000 no. m2, årlig gjennomsnitt i perioden 1995-2019), og 300 ganger større enn tettheten av C. glacialis og C. hyperboreus (Dalpadado mfl. 2012; Havforskningsinstituttet database). Relativt høye dyreplanktontettheter (> 1500 no. m3) ble observert i Nordvestbank-området i juni 2000 (Zhou mfl. 2009). Tilstedeværelsen av høye konsentrasjoner av dyreplankton, f.eks. flere kopepodestadier (Aarflot mfl. 2018, Havforskningsinstituttet database), er viktig som mat for larver og yngel av torsk.
Fisk
Lodde (Mallotus villosus) i Barentshavet gyter langs kysten av Troms, Finnmark og Kola på ganske grunt vann i mars-april, 2 0 til 90 m dyp, på bunnsubstrater av sand, singel og grus (Gjøsæter 1998, Gjøsæter mfl. 2009). De klebrige eggene er klistret til bunnsubstratet og klekkes etter 3 til 4 uker. Larvene stiger opp i det øvre laget (0-50 m) og driver med strømme r østover og nordover og ut i Barentshavet. Foreslått SVO Tromsøflaket er et viktig område for loddelarver sent på våren- tidlig sommeren og for ungsild (Clupea harengus) som beiter på loddelarver (Huse og Toresen 2000, Hallfredsson og Pedersen 2009, Gjøsæter mfl. 2016). Det er ikke sett endringer i gytetid eller sted selv om sjøtemperaturen har steget (Carcadden mfl. 2013a, b). Lodde er en nøkkelart i Barentshavets økosystem hele året og er viktig ved kysten når gytelodde vandrer til gytefeltet om vinteren og danner matgrunnlag for både fisk og sjøpattedyr om vinteren (Gjøsæter 1998, Gjøsæter mfl. 2009, 2011) .
Foreslått SVO Tromsøflaket er et svært viktig område for transport av egg (sild, torsk (Gadus morhua), hyse (Melanogrammus aeglefinus) , delvis sei (Pollachius virens)) og larver (lodde, torsk, sild, hyse, uer og delvis sei) fra gytefelt til Barentshavet om våren (Olsen mfl. 2010). I hovedsak foregår denne driften i de øverste 50 m av sjøen, men kan strekke seg ned mot 100 m. I tillegg er det et viktig beiteområde året rundt for lodda, siden dyreplankton oppkonsenteres i området, og både dyreplankton og lodde er bytte for annen fisk (Gjøsæter 1998, Gjøsæter mfl. 2009, 2011) .
Nordøstarktisk torsk (skrei) og kysttorsk er samme art, men har litt ulik genetisk sammensetning (Husa mfl. 2020). Skreien gyter eggene i de frie vannmasser i mars–april (F2, se figur 8.7.1). Både egg og larver driver med strømmen inn i Barentshavet, og yngelen bunnslår seg der sent på høsten. I fjordene (F5, figur 8.7.1) gyter kysttorsken i beskyttede områder, ofte innerst i fjordarmene, i poller, våger eller bukter, hvor gytingen foregår typisk på 20–60 meters dyp. På kysten benyttes havbukter, men også mer beskyttede lokaliteter blant øyer, holmer og sund. I verneområdet er det store gyteområder for skrei på kontinentalsokkelen vest og nord for Andøya. Det er også gyteområder for kysttorsk i Bunkefjorden og Leikvika på Senja. Informasjon om disse gyteområdene bygger på intervju med fiskere (Husa mfl. 2020).
Vanlig uer (Sebastes norvegicus) er vurdert som sterkt truet (EN) på grunn av en sterk nedgang i bestanden langs norskekysten. Bestanden er på et historisk lavmål og regnes for å ha redusert rekrutteringskapasitet på grunn av sterkt redusert gytebestand. Vanlig uer lever på 100-500 meters dyp på kontinentalsokkelen, blant annet langs Tromsøflaket og noen steder inne i fjordene. Uer parrer seg om høsten og føder levende yngel i vår, vanlig uer i april-juni (F3, figur 8.7.1) og snabeluer i mars–april langs Eggakanten (F4, figur 8.7.1). Atlanterhavsstrømmen sørger for at larver og yngel av disse arter blir transportert nordover og inn i Barentshavet mens de blir forsynt med plankton. Yngleområdene for vanlig uer og snabeluer strekker seg langs eggakanten og nordover og østover inn i Barentshavet, langs hele det foreslåtte SVO Tromsøflaket.
Bunnsamfunn
Mareano har også her kartlagt bentos med video og innsamlingsred sk aper . Observasjoner fra videopptak er sammen med resultater fra den oseanografiske modellen Norkyst 800, sedimentkart og topografi brukt til å modellere utbredelsen av sårbare habitater. Antallet observerte taxa av bunndyr gir en god indikasjon på biomangfoldet på havbunnen.
Kaldtvannskorallrev (Desmophylllum pertusum, tidligere kalt Lophelia pertusa), er registrert på 24 lokaliteter i Tromsøflaket/Lopphavet-området. Denne arten er vurdert som nær truet (NT) i Norsk rødliste for arter 2015. Sjøtre (Paragorgia arborea), som er en viktig art i hardbunnskorallskog i dette området, er vurdert som nær truet (NT, Husa mfl. 2020). Det samme er den solitære kjøttkorallen Anthomastus grandiflorus som er registrert ved ett tilfelle i bomtrål på sokkelen utenfor Troms..
Naturtypene korallrev, hardbunnskorallskog og svampspikelbunn er vurdert som nær truet (NT). Svampspikelbunn er en naturtype som dannes av store kålrabisvamper (Geodia spp.), der bunnen består av et tykt porøst lag med sedimenterte svampspikler. Både koraller og svamp er særlig utsatt for fiske med bunnredskaper og ekstra sedimentering (utslipp).
Området har verdens nordligste korallrev («Korallen»), nord for Sørøya. Dette revet ble vernet i 1999.
Dypvannsreken (Pandalus borealis) finnes på rekefelt i alle fjordene i Troms. De kartlagte rekefeltene viser kommersielle rekefelt, og arten bruker sannsynligvis en større del av bunnen (Zimmermann mfl. 2019). Dypvannsreken er heller ikke begrenset til bunnen, men gjennomfører vertikalvandringer gjennom døgnet for å beite på plankton. Det pelagiske larvestadiet bidrar til utveksling av individer mellom geografisk adskilte felt. Rekene lever i flere år, og vekst og utvikling er avhengig av bl.a. temperatur (Rasmussen 1953, Hopkins and Nilssen 1990, Båtevik 2020). De forskjellige livsstadiene kan til dels befinne seg på ulike felt og dyp (Hopkins and Nilssen 1990). Noen felt ser ut til å være viktige på ulike tider av året, mens andre felt ser ut til å være viktige året rundt. Dette inntrykket bekreftes av studier av andre rekebestander (Shumway mfl. 1985). Dypvannsreken langs norskekysten har frem til nylig vært lite studert sammenlignet med bestandene i Barentshavet og i Skagerrak og Norskerenna. Tidligere fantes det et stort rekefiske i fjordene i Nord-Norge, og fremdeles er rekefisket viktig lokalt. Se også omtalen av dypvannsreke under foreslått SVO Kystsonen Finnmark (Kap. 8.6.5).
Sjøpattedyr
Steinkobben (Phoca vitulina), vår mest tallrike kystsel, er en liten selart. I yngle- og hårfellingstiden fra midten av juni til begynnelsen av september er steinkobbene svært stedbundne og knyttet til stedet der de ble født (Bjørge og Øien 1999). Fra andre del av september til tidlig juni, er det næringssøk som har høyeste prioritet. I denne perioden vil de oppsøke steder med god tilgang på mat, ofte mange titalls kilometer fra stedet der de ble født, men de er likevel knyttet til faste steder (Nilssen og Bjørge 2019).
Spekkhoggere (Orcinus orca)er sosiale dyr som opptrer i familiegrupper av varierende størrelse, vanligvis om lag 5-15 individer som vinteren de kan opptre i store ansamlinger i forbindelse med overvintringen av sild (Similä mfl. 1995). Spekkhoggeren i kystnære farvann er først og fremst knyttet til sildeforekomstene i overvintringsperioden: For tida vil dette i særlig grad gjelde foreslått SVO Tromsøflaket, og historisk for foreslått SVO Kystsonen Lofoten og foreslått SVO Kystsonen Norskehavet sør (Jourdain og Vongraven 2017).
Springere (Lagenorhynchus spp.) trolig mest kvitnos (Lagenorhynchus albirostris), indikerer at Tromsøflaket kan være viktig beitehabitat for disse (Leonard og Øien 2020).
Sjøfugl
Sørfugløya og Nordfugløya huser to av de fem største lundekoloniene (Fratercula arctica) i Norge. To krykkjekolonier (Rissa tridactyla) har en stabil bestand, Andotten og Seiland, i motsetning til de fleste andre norske krykkjekolonier (https://seapop.no/). Kunnskap fra Seapop-programmet viser at sjøfuglene beiter ut til 100 km fra hekkekoloniene (Systad mfl. 2018). Krykkjene er borte fra Sørfugløy, Nordfugløy (Fugløykalven), Loppa og Galten. Sørfugløya, Nordfugløya samt øya Loppa har store og viktige sjøfuglkolonier i området med tilsvarende verdier som i Lofoten-Vesterålen, både for bentisk beitende arter som ærfugl (Somateria mollissima) og toppskarv (Phalacrocorax aristotelis), og for pelagiske arter som lunde, alke (Alca torda) og lomvi (Uria aalge). Totalestimater av koloniene i området indikerer at disse lundekoloniene ikke har gått tilbake på samme måte som de lenger sør. Viktigheten av disse koloniene øker relativt sett ettersom den nasjonale andelen av disse øker. Status for disse bestandene er imidlertid ikke godt kjent, da det ikke utføres overvåkning i dette området. Lundekoloniene i området utgjør en økende andel av den nasjonale bestanden. Dette fører til at negativ påvirkning i dette området får større betydning nasjonalt, men siden disse koloniene virker å produsere unger, er de mer robuste enn kolonier lenger sør der ungeproduksjonen er minimal (bl.a. Anker-Nilssen mfl. 2015).
Figur 8.7.3. Kart viser fordeling av hekkende sjøfugl (i farger) i foreslått SVO Tromsøflaket og nærliggende områder. A) Hekkende sjøfugl med pelagiske beiteområder (skravert felt). B), C) og D) Hekkende sjøfugl med kystnære beiteområder. Beiteområdene til disse artene er mer kystnære enn utstrekningen for området (Systad mfl. 2018). De anbefalte grensene for SVO Tromsøflaket og inntilliggende og overlappende foreslåtte SVO-er er vist med røde streker.
8.8 - Forslag til endret SVO Kystsonen Lofoten (BH6)
8.8.1 - Sammendrag
Områdebeskrivelse og strukturer
Forslag til endret SVO Kystsonen Lofoten (BH6) er karakterisert av en smal kontinentalsokkel avgrenset av en svært bratt kontinentalskråning. Sokkelen har flere markante grunnere banker og dypere raviner som Røstbanken, Sveinsgrunnen, Malangsgrunnen, Bleiksdjupet og Andfjorden. Hovedkomponentene i havsirkulasjonen er kyststrømmen over kontinentalsokkelen og den sterke, ensrettede og smale atlanterhavsstrømmen langs sokkelskråningen. Kyststrømmen påvirker, og påvirkes av, vannmasser i skjærgård og fjorder, bestemt av topografiske forhold som terskler og bassengdyp.
Miljøverdier
Den smale kontinentalsokkelen og dermed smal og sterk kyststrøm fører til konsentrasjon av dyre-plankton (og andre organismegrupper som transporteres med strømmen), Dette er bl.a. en svært viktig passasje for sildelarver og torskeegg og -larver og til dels viktig for hyseegg og hyselarver om våren.
Vestfjorden, mellom Lofoten og Salten i Nordland fylke har tradisjonelt vært et hovedgyteområde for nordøstarktisk torsk og hyse på senvinteren og utover våren. Det foreslåtte SVO-et omfatter noen av de viktigste yngleområdene for vanlig uer (sterkt truet-EN) på Vesterålsbankene i tillegg til en rekke andre fiskearter. Området er et viktig overvintringsområde for norsk vårgytende sild, som er viktig byttedyr for spekkhoggere, selv om den i perioder konsentrerer seg i andre fjordsystem lenger nord. Lofotenområdet er et viktig beiteområde for brugde (sterkt truet). Området har stor variasjon av marine naturtyper og landskaper som inkluderer et av verdens største kaldtvannskorallrevkomplekser (Røstrevet og Hola), Andfjorden med sterkt truet bambuskorallskog og sjøfjærbunn, det store Steinaværrevet og Bleiksdjupet, en av Europas største undervannsraviner.
Området innehar en stor andel sjøfugl nasjonalt sett, d e fleste av sjøfuglartene i Norge hekker her og det er et av de viktigste beiteområdene for sjøfugl i Norge, både vinterstid og i hekkesesongen Området er viktig overvintringsområde for kystnære arter som ærfugl og skarv. En stor andel av verdensbestanden av gulnebblom bruker området under trekk og overvintring. I økende grad brukes området som rasteplass under vårtrekket for arktiske gjess. Lomvi, lunde og krykkje som hekker i de store sjøfuglkoloniene i området, er alle truede og sårbare sjøfuglarter.
Områder rundt Øksnes og Andøya er viktige habitater for steinkobbe.
For ulike arter er det variasjoner gjennom året for når området er særlig viktig, men på grunn av det rike artsmangfoldet, er det til enhver tid viktig for en eller flere grupper av fisk, sjøfugl, bunndyr og sjøpattedyr.
Figur 8.8.1. Kart over forslag til endret SVO Kystsonen Lofoten (BH6). De foreslåtte grensene for SVO-et er vist med tykk rød strek, mens grensene for det overlappende foreslåtte SVO Eggakanten nord (BH3) og det inntilliggende foreslåtte SVO Tromsøflaket (BH5) er vist med tynnere rød strek. Grensene for forvaltningsplanområdene er markert med brun linje. Kartene viser utbredelsen av utvalgte miljøverdier av betydning for området. Miljøverdier knyttet til fisk (ynglefelt for vanlig uer og snabeluer, og gytefelt for de øvrige artene) og sjøfugl er markert i øverste kart og miljøverdier knyttet til bunnsamfunn og sjøpattedyr (blant annet vinterbeiteområde for spekkhogger) er markert i nederste kart.
8.8.2 - Vurdering i forhold til EBSA-kriteriene – Forslag til endret SVO Kystsonen Lofoten (BH6)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til endret
SVO Kystsonen Lofoten (BH6)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet /sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Dyreplankton Bunnsamfunn Sjøfugl
Fisk
Viktig område på grunn av en rik forekomst av raudåte som er mat for torskelarver (Ellertsen mfl. 1989, Hernandez 2019, Zhou mfl. 2009). Retensjonen i de grunne områdene er viktig årsak for oppkonsentrasjonen av plankton (Torgersen og Huse 2005).
Området er et svært viktig gytefelt for torsk og hyse (Bergstad mfl. 1987, Solemdal mfl. 1997, Hylen mfl. 2008, Sundby og Nakken 2008, Jakobsen og Ozhigin 2011, Castano-Primo mfl. 2014). Området er en viktig passasje for fiskeegg og larver (sild, torsk, hyse og uer (Dragesund mfl. 2008, Vikebø mfl. 2005, Skagseth mfl. 2015, Jakobsen og Ozhigin, 2011). Beiteområde for sjelden brugde (Junge mfl. under forberedelse). Det er også gytefelt for kysttorsk i området som kan dominere inne i Vestfjorden under gytetiden (Nordeide og Båmstedt 2020).
Unike korallrevsområder (Røstrevet, Hola og i Andfjorden), forekomster av bambuskorall i Andfjorden og en av Europas største undervannsraviner med særegne korall- og svampsamfunn i Bleiksdjupet (Mareano.no).
Området innehar en stor andel sjøfugl nasjonalt sett (Fauchald mfl. 2015).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Dyreplankton
Fisk Bunnsamfunn Sjøfugl
Vestfjorden er et viktig overvintringsområde for raudåte for å opprettholde en lokal høy populasjon (Espinasse mfl. 2016). Området er et svært viktig gytefelt for torsk og hyse (Bergstad mfl. 1987, Hylen mfl. 2008, Jakobsen og Ozhigin 2011) og en viktig transportvei for fiskeegg og larver (sild, torsk, uer og delvis hyse) (Dragesund mfl. 2008, Vikebø mfl. 2005, Skagseth mfl. 2015, Castano-Primo mfl. 2014).
Det har gjennom historisk tid vært et viktig overvintringsområde for sild, og selv om det nå er mer av den lenger nord, må SVO- området regnes som et viktig for overvintrende sild (Sætra mfl. 2002, Olsen mfl. 2010).
Strøm, topografi og vannkvaliteten bidrar til korallvekst, som bambuskorall i Andfjorden (Husa mfl. 2020).
Området er fremdeles et av de viktigste beiteområdene for sjøfugl i landet, både vinterstid og i hekkesesongen (Systad mfl. 2019).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Fisk Sjøfugl
Vanlig uer er klassifisert som en sterkt truet art (EN) (Henriksen og Hilmo 2015). Bestanden er lav og fortsetter å minke. De senere årene er det satt i gang tiltak for å bedre situasjonen (Bakketeig mfl. 2015). Brugde er klassifisert som sterkt truet (EN) i Norsk rødliste og Lofoten-området er et viktig beiteområde for brugde (Junge mfl. under forberedelse).
Lundebestanden (sårbar-VU) i Lofoten-Vesterålen har gått kraftig tilbake de siste 40 årene, Også flere av de andre sjøfuglartene har gått kraftig ned, krykkjebestanden (sterkt truet-EN) er redusert til mindre enn 20 prosent i perioden 1984-2013 (Anker-Nilssen mfl. 2015).
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Fisk Bunnsamfunn Sjøfugl
Området er gyteplass også for den sterkt truede vanlige ueren (EN) (Husa mfl. 2020) og det er identifisert som et viktig beiteområde for brugde (observasjonsdata fra https://havfunn.dugnadforhavet.no og Junge mfl. in prep).
Kaldtvannskorallrev, svampsamfunn (Svampspikelbunn og svamphage), sjøfjærbunn, korallskog er skjøre og svært gamle (Mareano.no). Syv ulike sårbare naturtyper karakterisert av fastsittende megafauna er blitt modellert basert på Mareanos observasjoner.
Sjøfuglbestandene i området er langtlevende og har lang restitusjonstid. De produserer dårlig, og er utsatte for flere påvirkninger. Bestandstrendene er negative over flere tiår for de fleste sjøfuglartene (bl.a. Anker-Nilssen mfl. 2015).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Sjøfugl Planteplankton
Dyreplankton Bunnsamfunn
Fisk
Kystnære områder har høyere planteplanktonproduksjon (Huseby 2002), knyttet til våroppblomstringen (Vikebø mfl. 2019).
Det er en god tilgang på raudåte om våren, hvorav store deler har opphav fra sentrale Norskehavet. Den blir stående lenge i retensjonsstrømmer i området (Sundby mfl. 2013). Vestfjorden har også en overvintringsbestand av raudåte og denne lokale overvintringen kan være viktig for å opprettholde høye konsentrasjoner av raudåte på sokkelen tidlig i torskelarvenes beitesesong (Espinasse mfl. 2016).
Høy produksjon av fiskelarver og egg (ichthyoplankton) i mars-mai, og deres næringsorganismer, larver av raudåte (Ellertsen mfl. 1989, Toresen mfl. 2019). Området har betydning på egg-, larve- og yngelstadiet for flere av de viktige fiskeressursene i Norskehavet og Barentshavet (Jakobsen og Ozhigin 2011, Sundby mfl. 2013).
Kaldtvannskorallrev og svampsamfunn (Svampspikelbunn og svamphage) danner habitat som utnyttes av fisk og bunndyr Mareano.no).
Sjøfugl er en viktig faktor for å bringe biologisk materiale fra hav til land (Zmudczyńska-Skarbek mfl. 2017) også i dette området.
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Fisk Bunnsamfunn Sjøfugl
Området inneholder store mengder av egg-, larve- og yngel av mange fiskearter, som er viktig mat for mange sjøfugl -, fiske- og sjøpattedyrarter og bidrar til biologisk biomangfold i området (Solemdal mfl. 1997, Hylen mfl. 2008, Dragesund mfl. 2008, Sundby and Nakken, 2008, Sundby mfl. 2013, Systad mfl. 2019).
Kaldtvannskorallrev, svampsamfunn (Svampspikelbunn og svamphage), samt høyt biomangfold observert med video danner komplekse leveområder for fisk og bunndyr (Mareano.no). Mareano har dokumentert rike benthosforekomster som inkluderer et av verdens største kaldtvannskorallrevkomplekser (Røstrevet og Hola). Til sammen er det registrert mer enn 400 individuelle korallrev. er det et særlig høyt mangfold mellom Vestfjorden og Troms, på grunn av overlappende temporære og arktiske arter i dette overgangsområdet mellom to klimasoner (Brattegard og Holte 1997, Brattegard 2011).
De fleste av sjøfuglartene i Norge hekker i dette området (Systad mfl. 2019).
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
Dyreplankton
Bunnsamfunn
Lite menneskelig påvirkning på dyreplankton foreløpig, unntatt menneskeskapte klimaendringer (Dalpadado mfl.2020).
Intakte kaldtvannskorallrev, omfattende svampområder og korallskog befinner seg i området (Mareano.no).
8.8.3 - Lokalisering
Forslag til endret SVO Kystsonen Lofoten dekker størstedelen av det eksisterende SVO Lofoten til Tromsøflaket, sist oppdatert i Meld. St. 20 (2019-2020), men er anbefalt utvidet slik at kandidatområdene for sjøfugl er innlemmet, og i tillegg er Vestfjorden, som nå er inkludert i SVO Kystsonen Norskehavet, foreslått inkludert i området (figur 8.8.1). Nordlig grense for SVO-et er dessuten justert ved at det nordligste området er tatt ut og i stedet inkludert i forslag til endret SVO Tromsøflaket, da miljøverdiene i dette området naturlig hørte bedre sammen med miljøverdiene der. Avgrensingen er endret primært på grunn av beiteområdene til sjøfugl i de to områdene (Systad mfl. 2019). Fagekspertgruppen foreslår derfor at navnet endres til SVO Kystsonen Lofoten.
Forslag til endret SVO Kystsonen Lofoten er lokalisert rundt Lofoten, Vestfjorden og nordover til Troms, og området overlapper delvis med foreslått SVO Eggakanten nord (figur 8.8.1).
Området dekker også et marint verneområde, Karlsøyfjorden nord for Bodø, samt to områder som er foreslått som marine verneområder, Røstrevet og Andøyatransektet.
8.8.4 - Introduksjon til området
Havområdene utenfor Lofoten er karakterisert av en smal kontinentalsokkel avgrenset av en svært bratt kontinentalskråning. Sokkelen har flere markante grunnere banker og dypere raviner som Røstbanken, Sveinsgrunnen, Malangsgrunnen, Bleiksdjupet og Andfjorden. Vestfjorden ligger mellom Lofoten og Salten i Nordland fylke.
De to ho vedkomponentene i havsirkulasjonen er kyststrømmen over kontinentalsokkelen og atlanterhavsstrømmen som følger sokkelskråningen. Sokkelskråningen har en sterk styrende virkning på strømmene (Orvik og Niiler 2002), og det resulterer i en sterk, ensrettet og smal atlanterhavstrøm med strømhastigheter på 30–100 cm per sek (Anderson mfl. 2011). Kyststrømmen på innsiden av atlanterhavsstrømmen brer seg ut over det meste av sokkelens bredde med hastigheter på opptil 35 cm per sek (Sundby mfl. 2013). Kyststrømmen påvirker, og påvirkes av, vannmasser i skjærgård og fjorder, avhengig av topografiske forhold som terskler og bassengdyp. Kjennetegn på kyststrømmen er først og fremst relativ lav saltholdighet. Kyststrømmen er viktig for transport nordover av fiskeegg og larver (Vikebø mfl. 2005, Skagseth mfl. 2015). Temperaturen i kystsonen økte (~0,8 °C) fra 1960-tallet til 2000-tallet, blant annet som følge av varmere atlanterhavsvann (Albretsen mfl. 2012; https://www.hi.no/hi/temasider/hav-og-kyst/klimaet-i-havet/klimastatus/kysten).
Mindre grener av atlanterhavsvann skjærer inn mot øst over sokkelen i de dypere ravinene, og også de grunne bankene kan generere permanente og stabile strømsystemer. Også mindre bankstrukturer ned mot størrelser på 5–10 km i diameter kan ha en slik styrende virkning på strømmen (Sundby 2013).
Figur 8.8.2. Gjennomsnittlig overflatestrøm (piler) og bunndyp (farger) i forslag til endret SVO Kystsonen Lofoten og omkringliggende områder.
8.8.5 - Beskrivelse av miljøverdiene
Plankton
Planteplanktonet viser stor variasjon innenfor området, der de kystnære områdene har høyere planteplanktonproduksjon (Huseby 2002), knyttet til våroppblomstringen (Vikebø mfl. 2019) enn lenger ute fra kysten. Topografiske forhold sikrer oppstrømning av næringssalter som stimulerer stabil og høy planteplanktonproduksjon gjennom sesongen. For det foreslåtte SVO Kystsonen Lofoten er det mange likheter med forhold omtalt i de foreslåtte SVO-ene Tromsøflaket og Eggakanten nord. Ved Skrova pågår det fast overvåkning av blant annet planteplankton, som viser den generelle sesongmessige variasjonen i området (Naustvoll mfl. 2020). Det foreligger betydelig mindre informasjon fra dette området sammenlignet med fjordområdene i Troms.
Store deler av dyreplanktonet i området har opphav i sørlige og sentrale deler av Norskehavet og driver med kyststrømmen nordover. Raudåta, Calanus finmarchicus, er en viktig komponent i området og den mest tallrike dyreplanktonorganismen som utgjør til tider opp mot 80–98 prosent av organismene (Hernandez 2019). Vesterålen og områdene lengre nord er viktig særlig for torskelarver i deres første næringsopptak da larvene beiter på de tidligste stadiene av raudåte nauplier (Ellertsen mfl. 1989). Disse naupliene kan dominere i planktonet i mai (Zhou 2009). Biomassene i Lofoten–Vesterålen i mai er ganske stabile både mellom år og områder og varierer mellom 5–15 g tørrvekt per m2, i begrensede områder kan det forekomme høye verdier opp til 30 g tørrvekt per m2 (Broms 2015, Broms 2016). Til sammenlikning er langtidsgjennomsnittet for hele Norskehavet i mai ca. 9 g tørrvekt per m2. Et enkelt år er det observert fra 500 til over 3000 organismer per m3, det høyeste antallet i et begrenset område utenfor Andøya (Zhou mfl. 2009).
Vestfjorden har høy biomasse av dyreplankton (ca. 16–18 g tørrvekt per m 2) på våren sammenli g net med omkringliggende områder (ICES 202 0a og 2020 b , Wiborg 1954). Basert på storskala overvåkning gjelder dette særlig de siste årene, men også tidligere er det rapportert høye biomasser i ytre deler av Vestfjorden. Lavere biomasser er tidligere registrert i indre deler av fjorden (Wiborg 1954), noe som støttes av nye satellittbilder fra våren (Basedow mfl. 2019). Det kan være mellomårlige variasjoner. C. finmarchicus finnes i høyt antall på våren (mai), sensommeren (august) og så sent som i oktober, men det er store sesongvariasjoner i antallet (Strand mfl. 2020). Vestfjorden har også et høyt antall overvintrende C. finmarchicus , og denne lokale overvintringen kan være viktig for å opprettholde C. finmarchicus populasjonen på sokkelen tidlig i torskelarvenes beitesesong (Espinasse mfl. 2016). Det er registrert en rekke arter og grupper i området (data fra Continuous Plankton Recorder, Strand mfl. in progress), men det er ikke gjort studier av artsdiversiteten langs kysten som kan si om Vestfjorden har høyere diversitet enn andre områder. Det er lokal produksjon av Calanus hyperboreus i fjorden (Sømme 1934), en arktisk art som har hovedutbredelsen i Grønlandshavet, men som også finnes i dype norske fjorder.
Fisk
Lofoten og kystområdene i sør (Vestfjorden) og i nord (Vesterålen) i det foreslåtte SVO Kystsonen Lofoten inneholder de viktigste gytefeltene for flere fiskearter. Lofoten-området og det smale sokkelhavet nord for Lofoten er svært viktige områder for egg (torsk (Gadus morhua), hyse (Melanogrammus aeglefinus) , delvis sei (Pollachius virens)) og larver (torsk, sild (Clupea harengus), hyse og delvis sei). Siden dette er et område med gytefelt for uvanlig mange fiskearter, blir det presentert en rekke av dem her.
Nordøstarktisk torsk (torsk) gyter i mars–april og eggene blir gytt i frie vannmasser (F2, figur 8.8.1). Området inneholder store konsentrasjoner av egg og larver. Fra gytefeltene driver eggene med strømmen inn i Barentshavet og langs kysten av Vest-Spitsbergen, og yngelen bunnslår seg der sent på høsten (Hylen mfl. 2008). Gytebestanden av torsk er i dag på et høyt nivå. Både totalbestanden og gytebestanden har vokst siden 2006 og nådde toppen i 2013. Mesteparten av bestanden finner man i Barentshavet, på den varme siden av Polarfronten (til ca. 76°N og 50°Ø). I varme år går utbredelsen lenger nord og øst (Hylen mfl. 2008, Kjesbu mfl. 2014, Fossheim mfl. 2015, ICES 2020 a,b). Torsken er en altetende fisk og en av de viktigste rovfiskene i Barentshavet (Eriksen mfl. 2020). Det er også gytefelt for kysttorsk i området, som kan dominere inne i Vestfjorden under gytetiden (Nordeide og Båmstedt 2020).
Nordøstarktisk hyse (hyse) er en torskefisk som finnes langs hele kysten nord for Stad og inne i Barentshavet, inkludert vestsiden av Svalbard. En ny genetisk studie tyder på at hyse er utbredt fra Lofoten og nordover, og at hyse lenger sør er lokale bestander mer i slekt med nordsjøhyse (Berg mfl. 2020). Hovedgytefeltene til hyse er i motsetning til torsken ute langs kontinentalskråningen (de foreslåtte SVO-ene Eggakanten sør og Eggakanten nord), men den gyter også i foreslått SVO Kystsonen Lofoten (F6 i figur 8.8.1) (Bergstad mfl. 1987, Solemdal mfl. 1997, Jakobsen and Ozhigin, 2011, Castano-Primo mfl. 2014). Gytingen foregår fra mars til juli, med hovedtyngde i slutten av april.
Overvintringsområdene for norsk vårgytende sild (sild) er svært dynamisk og fra midten av 1990-tallet til midt på 2000 tallet overvintret mye av silda innerst i Vestfjordområdet. Etter dette forsvant den fra Vestfjorden om vinteren og begynte å overvintre ute i Norskehavet (Huse mfl. 2010). Huse mfl. viste at silda endrer overvintringsområde når det kommer sterke årsklasser inn i den voksne bestanden. Dette fenomenet kalles numerisk dominans og skyldes at den nye sterke årsklassen «styrer» hvor resten av silda svømmer siden de dominerer i bestanden.
Hovedgyteområder for sild er på kystbankene (Dragesund mfl. 1980 , Sætre mfl. 2002, Sundby mfl. 2013) som er dekket i andre SVO-er (foreslåtte SVO-er Kystsonen Norskehavet sør og nord), men også i foreslått SVO Kystsonen Lofoten er det store gytefelt vest for Lofoten og Vesterålen. Selv om noe sildegyting også er observert i Vestfjorden, er det ikke ansett som så sentralt som kystbankene (F8, figur 8.8.1). Sild gyter på bunn og er avhengig blant annet av riktig bunnsubstrat (Runnstrøm, S. 1941). Sildelarvene vil etter klekking i perioden mars–juli drive nordover langs norskekysten (Sætre mfl. 2002). Sildeeggene og larvene er viktig næringskilde for fisk og andre organismer som beiter på eggene i perioden etter gyting. En gytebestand på 10 millioner tonn legger ca. 2 millioner tonn gyteprodukter hvert år (Toresen 1991 , Høines og Bergstad 1999 , Varpe mfl. 2005) . Dette er en stor matkilde for dyr langs kysten for rovfisk som torsk, sei og annen bunnfisk, i tillegg til hval om våren og sommeren.
Vi har tre arter av uer i Norge, vanlig uer (Sebastes norwegicus), snabeluer (Sebastes mentella) og lusuer (Sebastes viviparus). Vanlig uer er vurdert som sterkt truet (EN) og er på et historisk lavmål og regnes for å ha redusert rekrutteringskapasitet på grunn av sterkt redusert gytebestand. Nord for 62°N er det forbudt å fiske uer, med unntak av juksafiske fra båt under 15 m lengde i perioden 1. juni–31. august. En del av ynglingen av vanlig uer foregår vest og nord for Lofoten (Husa mfl. 2020). Uer parrer seg om høsten og føder levende yngel i mars–juni. Yngelen transporteres i hovedsak i de øverste 50 m av sjøen, men kan strekke seg ned mot 100 m dybde i transport fra gytefelt til Barentshavet om våren. Denne yngeltransporten knytter sammen kystnære SVO-er oppover hele kysten og inn mot Barentshavet. Vanlig uer kan bli én meter lang, opptil 15 kg og kan bli opptil 60 år gammel, og har derfor lav restitueringsevne (Husa mfl. 2020). Arten lever på 100–500 meters dyp på kontinentalsokkelen, langs kysten og noen steder inne i fjordene.
Øyepål (Trisopterus esmarkii) er en ganske liten fisk i torskefamilien med maksimal individvekt på 0,1 kg og med en levetid som sjelden går over 3 år. Øyepål opptrer i store stimer, som regel over mudderbunn. Den spiser hovedsakelig planktoniske krepsdyr, som krill og raudåte (Sundby mfl. 2013) . Øyepål utgjør igjen en del av næringsgrunnlaget for andre større fisk som torsk, hvitting og sei, men også sjøpattedyr. Siden 1995 har fiskedødeligheten vært på et lavere nivå enn tidligere. Utbredelsen strekker seg også nordover Norskekysten og inn i det sørlige Barentshavet. Hovedgytefeltene er spredt ut i den nordlige delen av Nordsjøen. De nordlige gytefeltene som har relevans for egg-, larve- og yngeldrift gjennom Lofoten–Vesterålen omfatter kysten av Møre, Haltenbanken, Vestfjorden, rundt Røst og utover langs vestligste delen av kontinentalsokkelen utenfor Vesterålen (Sundby mfl. 2013) .
Isgalt (Macrourus berglax) gyter hovedsakelig på Røstbanken og Trænabanken (Savvatimsky 1985) mens det er noe gyteaktivitet nordover i det foreslåtte SVO Tromsøflaket (Savvatimsky 1985). Gytingen foregår fra midt-vinter til tidlig vår med høydepunkt i desember-januar (Eliassen 1983, Savvatimsky 1985).
Brugde (Cetorhinus maximus) samler seg i kystområder av kontinentalsokkel dominert av overgangsvann mellom lagdelte og blandede vannmasser. Årlige observasjoner av brugde er godt dokumentert (Sims 2008). Lofotenområdet er identifisert som et viktig beiteområde for brugde (observasjonsdata fra https://havfunn.dugnadforhavet.no og Junge mfl. in prep). Brugde spiser dyreplankton, med k alanoide k opepod er som den viktigste byttedyrgruppen (se kapittel ovenfor om plankton). Brugde har vist seg å respondere på dyreplanktongradienter, og viser at de er selektive filter spisere som v elger de rikeste og mest lønnsomme planktonkonsentrasjonene (Sims og Quayle 1998).
Bunnsamfunn
M areano har kartlagt benthos med video og innsamlingsredskaper i dette området. Observasjoner fra video er sammen med resultater fra den oseanografiske modellen Norkyst 800, sedimentkart og topografi brukt til å modellere utbredelsen av sårbare habitater. Antallet observerte taxa av bunndyr gir en god indikasjon på biomangfoldet på havbunnen. Området har rike benthosforekomster som inkluderer et av verdens største kaldtvannskorallrevkomplekser (Røstrevet og Hola). Til sammen er det registrert mer enn 400 individuelle korallrev. Området omfatter også Andfjorden som har forekomster av bambuskorallskog (Isidella lofotensis), sjøfjærbunn og det store Steinaværrevet. Utenfor Andøya, ligger Bleiksdjupet, en av Europas største undervannsraviner.
Syv ulike sårbare naturtyper karakterisert av fastsittende megafauna er blitt modellert basert på Mareanos observasjoner. Modellen over sannsynlig utbredelse av dyphavssjøfjær av arten Umbellula encrinus dekker det største arealet. Disse forekommer i relativt lav tetthet og på dyp mellom 1000 og 2000 m. Svampskog (svampsamfunn på hardbunn) og bløtbunns-svampsamfunn (også kalt Svampspikelbunn) har også en vid modellert utbredelse. Bløtbunnssvampene forekommer som et bånd langs den vestlige og sørlige del av Røsttunga, mens svampskogen forekommer på grunnere områder med steinbunn fra Røst til Langøya. Hardbunnskorallskog er modellert med et mer begrenset areal som et parallelt bånd med Røstrevet, noe dypere enn korallrevene der. Områdene med modellert utbredelse av sjøfjær indikerer at disse forekommer hyppigst i renner som krysser sokkelen (Kvalnesdjupet, Malangsdjupet og Fugløydjupet).
I den omfattende kartleggingen av alle virvelløse dyr langs hele Norge, er det et særlig høyt mangfold mellom Vestfjorden og Troms, på grunn av overlappende temporære og arktiske arter i dette overgangsområdet mellom to klimasoner (Brattegard og Holte 1997, Brattegard 2011).
Dypvannsreke finnes på en mosaikk av små og store rekefelt langs norskekysten. Tidligere fantes det er aktivt reketrålfiske i Vesterålen (data fra Fiskeridirektoratet). Se også omtalen av dypvannsreke under foreslått SVO Tromsøflaket (Kap. 8.7.5).
Sjøpattedyr
Steinkobben (Phoca vitulina), vår mest tallrike kystsel, samles på land i forbindelse med forplantning og hårfelling. De legger seg helst opp på litt beskyttede lokaliteter i skjærgården (skjær og sandbanker som tørrlegges ved fjære sjø) (Bjørge og Øien 1999). I yngle- og hårfellingstiden fra midten av juni til begynnelsen av september er steinkobbene svært stedbundne og knyttet til stedet der de ble født. Fra andre del av september til tidlig juni, er det næringssøk som har høyeste prioritet. I denne perioden vil de oppsøke steder med god tilgang på mat, ofte mange titalls kilometer fra stedet der de ble født. Områder rundt Øksnes og Andøya er viktige habitater for steinkobbe og bestanden teller i underkant av 1850 dyr (Nilssen og Bjørge 2019).
Spekkhogger (Orcinus orca) er sosiale dyr som opptrer i familiegrupper av varierende størrelse, vanligvis om lag 5–15 individer. Den er en marin topp-predator, og beiter på et bredt register av byttedyr som omfatter andre hvalarter, sel, mange fiskearter, blekksprut og sjøfugl (Leonard og Øien 2020). I norske farvann er det sannsynligvis makrell og sild som dominerer matseddelen (Leonard og Øien 2020). Det er særlig om vinteren de kan utgjøre store ansamlinger i forbindelse med overvintringen av sild (Similä mfl. 1995). Etter at norsk vårgytende sild hadde bygget seg opp igjen etter kollapsen, fulgte den i flere tiår et mønster der den overvintret innerst i Vestfjorden (Ofotfjorden og Tysfjorden) sammen med anslagsvis 500 spekkhoggere som beitet på den. For ca. 10 år siden endret silda overvintringslokalitet til kystområdene og til dels langt inn i fjordsystemene fra Andøya og nordover. Området var da et viktig beiteområde for spekkhoggerne om vinteren når store flokker av spekkhoggere følger silda på dens vandringer til overvintringsområdet og gytefelt. For tida vil dette i særlig grad gjelde foreslått SVO Tromsøflaket, og historisk for de foreslåtte SVO-ene Kystsonen Lofoten og Kystsonen Norskehavet sør. Utenom overvintringsperioden, har spekkhoggerne i kystnære farvann sannsynligvis en mer variert diett.
Sjøfugl
Dette området er overvintringsområde for flere sjøfuglarter, særlig kystbundne arter som for eksempel ærfugl (Somateria mollissima), storskarv (Phalacrocorax carbo) og toppskarv (Phalacrocorax aristotelis) (Systad mfl. 2019). Området er et av de viktigste områdene i Norge for hekkende sjøfugl, med de store fuglefjellene Røst, Værøy, Fuglenyken og Bleiksøy, som alle er hekkekolonier for lunde (Fratercula arctica) (Fauchald mfl. 2015). Røst har vært den desidert største lundekolonien i Norge fram til årtusenskiftet, men antallet har falt kraftig (Anker-Nilssen mfl. 2015, Anker-Nilssen mfl. 2018). De kolonihekkende sjøfuglene er avhengige av at fiskelarver driver forbi koloniene til rett tid (Sandvik mfl. 2016), noe som ikke har slått til over lenger tid på Røst.
Også andre arter som lomvi (Uria aalge) og alke (Alca torda) finnes i flere av disse koloniene. Lofoten og Vesterålen er i tillegg et svært viktig overvintringsområde for bl.a. gulnebblom (Gavia adamsii) (nær truet-NT, Henriksen og Hilmo 2015), teist (Cepphus grylle) og praktærfugl (Somateria spectabilis). En stor andel av verdensbestanden av gulnebblom bruker området under trekk og overvintring (Birdlife International 2021). I økende grad brukes området som rasteplass under vårtrekket for arktiske gjess (Tombre mfl. 2019, Oudman mfl. 2020).
Data samlet inn bl.a. gjennom Seapop viser at beiteområdene for sjøfugl gjerne strekker seg ut til 100 km fra koloniene (Systad mfl. 2018).
Figur 8.8.3 Kart viser fordeling av hekkende sjøfugl (i farger) i foreslått SVO Kystsonen Lofoten og nærliggende områder. A) Hekkende sjøfugl med pelagiske beiteområder (skravert felt). B), C) og D) Hekkende sjøfugl med kystnære beiteområder. Beiteområdene til disse artene er mer kystnære enn utstrekningen for området (Systad mfl. 2018). De anbefalte grensene for SVO Kystsonen Lofoten og inntilliggende og overlappende foreslåtte SVO-er er vist med røde streker.
Det har vært omfattende hekkesvikt samt langvarig bestandsnedgang hos flere sjøfuglarter i området, særlig pelagisk tilknyttede arter som lomvi, lunde og krykkje (Rissa tridactyla). Krykkje på Anda i Vesterålen har vært et unntak. Fuglene er avhengig av at fiskeyngel driver forbi til rett tidspunkt og med rett størrelse. Dersom de pelagiske bestandene gyter lenger nord eller tidligere i sesongen, kan det være at yngelen først når riktig størrelse lenger nord og øst enn i Lofoten–Vesterålen.
Lomvi hekker ennå på Fuglenykene, Anda og Bleik i Vesterålen. De relativt store lundekoloniene på Fuglenykene og Bleik var kraftig redusert ved siste telling i 2015. Lomvi viser klare tegn til framgang på andre øyer i Røstarkipelaget, der den hekker i skjul, slik at den unngår predasjon og forstyrrelser fra havørn (Haliaeetus albicilla). Selv om bestanden er liten, forklarer dette at det i siste tiårsperioden (2007–2017) likevel har vært en årlig bestandsøkning på 10 prosent på nøkkellokaliteten Røst (som omfatter flere øyer) (Anker-Nilssen mfl. 2018).
Torskeegg fra Moskenesstraumen. Foto: M.E. Johannesen, HI
Koraller og svamp i Andfjorden. Foto: Mareano / HI
8.9 - Forslag til nytt SVO Det sentrale Barentshavet (BH7)
8.9.1 - Sammendrag
Områdebeskrivelse og strukturer
Forslag til nytt SVO Det sentrale Barentshavet (BH7) er karakterisert av den sørlige delen av den grunne (< 200 m dyp) Sentralbanken, deler av den litt dypere Thor Iversen-banken (< 250 m dyp) sør for dette og det dypere bassenget (> 300 m dyp) som ligger sørøst av disse mot Novaja Semlja.
Atlanterhavsstrømmen som kommer inn fra sørvest deler seg i sørvestlig del av området, og to strømgrener går østover sør av Sentralbanken. I tillegg strømmer det inn kaldere arktiske vannmasser, og om vinteren is, til nordlig del av området. Styrken og omfanget av tilstrømning av kaldere vann og is varierer med vindforholdene.
Miljøverdier
Området innehar en stor andel sjøfugl, da sjøfugl fra hekkekolonier rundt hele Barentshavet (både de arktiske øyene og fastlandet) samles her. Området er særlig viktig etter hekkesesongen for sjøfugl (lomvi og polarlomvi) som svømmer inn i området for å beite, mens de myter og ikke kan fly, og området er også viktig for overvintring for disse artene. I tillegg dekker dette deler av høst- og overvintringsområdene til lunde og polarmåke, samt krykkjebestandene på vårparten. De nevnte artene bruker større deler av Barentshavet, men i dette området samler flere av bestandene seg store deler av året. En rekke sjøfuglarter som lomvi, polarlomvi, krykkje, lunde, og havhest er rødlistet.
Om våren og sommeren fører havstrømmene dyreplankton, egg, yngel og fiskelarver inn i området. Derfor er dette et beiteområde for voksen fisk, sjøfugl og til dels sjøpattedyr. Torske- og hyseyngel bunnslår seg om vinteren, mens yngel av andre arter holder seg i øverste vannmasser. Det er store forekomster av spesielle og sjeldne Haploops-samfunn på Thor Iversen-banken.
Samlet sett er dette et unikt og viktig område, som et særlig attraktivt svømmebeiteområde for sårbare og rødlistede arter i den atlantiske delen av Barentshavet, med stor betydning for biologisk produktivitet og mangfold.
Bunnsamfunn nær Svalbard. Foto: Mareano, HI
Figur 8.9.1. Kart over forslag til nytt SVO Det sentrale Barentshavet (BH7). De foreslåtte grensene for SVO-et er markert med tykk rød strek, mens grensene for det overlappende foreslåtte SVO Iskantsonen (BH2) og andre nærliggende foreslåtte SVO-er er markert med tynn rød strek. Grensene for forvaltningsplanområdet er vist med brun linje. Kartet viser utbredelsen av viktige miljøverdier som er av betydning for området. Pilene illustrerer svømmetrekkruter for forskjellige bestander av alkefugl inn i området etter hekkesesongen.
Beitende alkefugl på sjøen. Foto: H. Strøm, NP
8.9.2 - Vurdering i forhold til EBSA-kriteriene – Forslag til nytt SVO Det sentrale Barentshavet (BH7)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til nytt
SVO Det sentrale Barentshavet (BH7)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet /sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Dyreplankton Fisk
Sjøfugl
Bunnsamfunn
Området har en rik forekomst av raudåte som er mat for fiskelarver (Ellertsen mfl. 1989, Hernandez 2019, Zhou mfl. 2009).
Det er et særlig sentralt område for bunnslåing av fiskeyngel (sild, torsk, hyse og uer) (Dragesund mfl. 2008, Vikebø mfl. 2005, Skagseth mfl. 2015, Jakobsen og Ozhigin 2011).
Det er store forekomster av spesielle og sjeldne Haploops-samfunn på Thor Iversen-banken (Lien mfl. 2018).
Området innehar en stor andel sjøfugl nasjonalt og internasjonalt, da mytende sjøfugl fra hekkekolonier rundt hele Barentshavet samles her på svømmebeite om høsten (Systad mfl. 2019).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Fisk Bunnsamfunn Sjøfugl
Området er en viktig passasje for fiskeegg og larver (sild, torsk, hyse og uer) (Dragesund mfl. 2008, Vikebø mfl. 2005, Skagseth mfl. 2015, Jakobsen og Ozhigin 2011). Området er et svært viktig yngelområde for lodde, sild, torsk, hyse gjennom sommeren (Bergstad mfl. 1987, Hylen mfl. 2008, Sundby og Nakken 2008, Jakobsen og Ozhigin 2011, Castano-Primo mfl. 2014, Eriksen mfl. 2017). Torsk og hyseyngel bunnslår seg om vinteren, mens yngel av andre arter holder seg i øverste vannmasser (Eriksen mfl. 2011).
Det er store forekomster av Haploops-samfunn på Thor Iversen-banken (Lien mfl. 2018).
Loggerdata viser at området er viktig gjennom store deler av året for sjøfugl (SEATRACK.no).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Bunnsamfunn Sjøfugl
Utbredelsen av Haploops-samfunnet i Barentshavet for øvrig er lite kjent, og OSPAR utreder hvorvidt Haploops-habitatet skal vurderes som truet, ettersom det foregår stor fiskeriaktivitet med bunntrål i området (Buhl-Mortensen og Buhl-Mortensen 2018).
Området er viktig for en rekke pelagisk beitende sjøfuglarter som rødlistet lomvi, polarlomvi, krykkje, lunde og havhest i høst- og vintersesongen (Systad mfl. 2019).
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Sjøfugl
Bunnsamfunn
De store forekomster av Haploops-samfunn på Thor Iversen-banken lever i bunnkonstruksjoner som er svært sårbare for bunnforstyrrelser og samtidig veldig avhengig av rett bunntype og strømmer (Lien mfl. 2018).
De aktuelle sjøfuglartene i området er alle rødlistet og sårbare for menneskelig påvirkning. Området er svært viktig for de samme artene i høst- og vintersesongen (Systad mfl. 2019).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Plante-plankton
Dyreplankton Fisk
Når det gjelder planteplankton foreligger noe data, men de bør reanalyseres.
Høy produksjon av dyreplankton i dette området er viktig for planktonspisende fisk i området (Dalpadado og Mowbray 2010, Orlova mfl. 2010, Prokopchuk 2019, ICES 2019). Høy predasjon fører til lave biomasser i disse bankområdene.
Området har betydning som egg, larve- og yngelområde for fiskeressursene i Norskehavet og Barentshavet (Jakobsen og Ozhigin 2011, Sundby mfl. 2013, Eriksen mfl. 2017). Torsk og hyseyngel bunnslår seg om vinteren, mens yngel av andre arter (lodde, sild osv.) holder seg i øverste vannmasser, hvor de er tilgjengelige for større fisk, sjøpattedyr og sjøfugl (Boitsov mfl. 2013, Systad mfl. 2019).
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Planteplankton
Sjøfugl Bunnsamfunn Pattedyr
Fisk
Artsrikdommen for fisk i det sentrale Barentshavet er større enn i områdene rundt (Johannesen mfl.2021).
Området har et stort mangfold av sjøfugl (Systad mfl. 2019). Det er dessuten beiteområde for kvitnos, vågehval og tidvis nise (Leonard og Øien 2020). Det biologiske mangfoldet antas være knyttet til planteplankton, men dette er ikke dokumentert.
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
8.9.3 - Lokalisering
Forslag til SVO Det sentrale Barentshavet er et nytt SVO som er avgrenset på grunnlag av betydningen området har for en rekke pelagisk beitende sjøfuglarter i høst- og vintersesongen.
Beiteområdet er lokalisert i den sentrale delen av Barentshavet, mellom ca. 71° og 75°N og mellom ca. 28° og 44°Ø, men selve SVO-et er foreslått avgrenset mot øst ved grensen til Russland ved ca. 37°Ø. Området omfatter den sørlige delen av Sentralbanken, deler av den litt dypere Thor Iversen-banken sør for dette og det dypere bassenget som ligger sørøst for disse mot Novaja Semlja (figur 8.9.1).
Det er ingen verneområder i området.
8.9.4 - Introduksjon til området
Det sentrale Barentshavet er karakterisert av den sørlige delen av den grunne (< 200 m dyp) Sentralbanken, deler av den litt dypere Thor Iversen-banken sør for dette og det dypere bassenget (> 300 m dyp) som ligger sørøst av disse mot Novaja Semlja.
Atlanterhavsstrømmen som kommer inn fra sørvest er en viktig komponent i havsirkulasjonen (Loeng, 1991). Denne strømmen deler seg i sørvestlig del av området (figur 8.9.2). To strømgrener går østover sør av Sentralbanken (Loeng 1991), og begge grenene bringer om våren og sommeren med seg dyreplankton og fiskelarver inn til området (Eriksen mfl. 2019). Nordlig del av området kan være dekket av is om vinteren, men dette var mer vanlig før når Barentshavet var kaldere (Barton mfl. 2018, Årthun mfl. 2012) .
Figur 8.9.2. Gjennomsnittlig overflatestrøm (piler) og bunndyp (farger) i forslag til nytt SVO Det sentrale Barentshavet og omkringliggende områder.
8.9.5 - Beskrivelse av miljøverdiene
Plankton
Store deler av det foreslåtte SVO Det sentrale Barentshavet er dominert av vannmasser av atlantisk opprinnelse (Ozhigin mfl. 2011). Dyreplanktonet er i stor grad knyttet til de dominerende atlantiske vannmassene i området. Gjennomsnittsbiomassen om høsten i perioden 1989–2017 på Thor Iversen-banken, som dekker mesteparten av det foreslåtte SVO Det sentrale Barentshavet, var 6,0 g tørrvekt per m2 (Dalpadado mfl. 2020). Undersøkelser som er foretatt på et snitt (Vardø nord) gjennom området fra mars til juli viser små endringer i biomasse (5,0 g tørrvekt per m2) over tid og den atlantiske raudåten (Calanusfinmarchicus) utgjorde opptil 90 prosent av dyreplanktonbiomassen, fulgt av Metridia longa (Arashkevich mfl. 2002). Denne studien viser at små kopepoder, Oithona, Microcalanus og Pseudocalanus-arter, dominerte i antall. Vertikalfordelingen endret seg med sesongen, i mars var mesteparten av dyreplanktonet lokalisert dypere enn 100 m i hele området, i mai i de øvre 100 m, og i juli hadde hovedmengden migrert mot dypet i den sørlige del av området (Arashkevich mfl. 2002). De lave biomassene vi generelt ser i bankområdene i forhold til øvrige deler av Barentshavet kan skyldes predasjon fra lodde, sild, og 0-gruppe fisk, som det finnes store konsentrasjoner av i dette området (Dalpadado og Mowbray 2013, Orlova mfl. 2010, Prokopchuk 2019, ICES 2019).
Fisk
Lodde (Mallotus villosus) lever hele sitt korte liv i Barentshavet (Gjøsæter 1998, Orlova mfl. 2005, 2011). Den modnende delen av bestanden beiter også i området. Denne bestanden som består av 3–5 år gammel fisk vandrer mot kysten (inn i foreslått SVO Kystsonen Finnmark) for å gyte, som oftest i begynnelsen av mars (Gjøsæter 1998, Gjøsæter mfl. 2009). Det foreslåtte SVOet er også et svært viktig område for transport av larver og yngel av lodde, torsk (Gadus morhua), sild (Clupea harengus), hyse (Melanogrammus aeglefinus) og delvis sei (Pollachius virens), fra gytefelt til Barentshavet om våren (Olsen mfl. 2010, Eriksen mfl. 2017). I hovedsak foregår driften i de øverste 50 m av sjøen, men kan strekke seg ned mot 100 m. Det er den beitende fisken som er grunnlaget for de beitende sjøfuglene på ettersommer og høst.
Det er i det foreslåtte SVO Det sentrale Barentshavet diversiteten av fiskearter er høyest (Johannesen mfl. 2021).
Bunnsamfunn
Sør for Sentralbanken, på den atlantiske siden av polarfronten er det gjennom Mareano-programmet nylig innsamlet store forekomster av Haploops-samfunn på Thor Iversen-banken (Lien mfl. 2018). De vanligste artene er H. tubicola, men også H. similis er registrert, og innsamlingene viser en tetthet på 100–300 individer per m2. Utbredelsen av dette samfunnet i Barentshavet er lite kjent, og det er for tiden under utredning i OSPAR hvorvidt dette habitatet skal vurderes som truet, ettersom det foregår stor fiskeriaktivitet med bunntrål i området hvor Haploops-samfunnene er registrert (Buhl-Mortensen og Buhl-Mortensen 2018).
Dypvannsreke (Pandalus borealis) er utbredt i sentrale deler av Barentshavet, og denne rekebestanden er den største og økonomisk viktigste i norske farvann (Garcia 2007, NAFO, ICES 2020 a). Reken i Barentshavet og ved Svalbard utgjør en egen genetisk bestand, helt ulik rekene langs norskekysten (Jorde mfl. 2015, Hansen 2020). Larvedriftsmodellering viser at hunnreker i hele Barentshavet bidrar til rekrutteringen (Pedersen mfl. 2003). Bestanden som overvåkes årlig, er i god forfatning og har siden 1990-tallet fluktuert uten trend (NAFO, ICES 2020 a). ICES gir årlige kvoteråd, men det settes ingen kvoter. Dypvannsreke er en viktig art i økosystemet og er viktig som mat for bunnfisk, særlig torsk (Berenboim mfl. 2000, Holt mfl. 2019). Utbredelsen av bestanden har de siste årene flyttet seg mot nordøst, og det har blitt mindre reke på de tradisjonelle fiskefeltene i vest. Landingene har økt de siste tre årene, både fordi Russland igjen har begynt å fiske reker og fordi tredjeland lander mer enn tidligere (NAFO, ICES 2020 a). Tredjeland har bare mulighet til å fiske reker i Svalbardsonen og i Smutthullet. Se også omtalen av dypvannsreke under foreslått SVO Havområdene rundt Svalbard (Kap. 8.3.5).
Sjøpattedyr
Det er ingen spesifikke sjøpattedyr som er knyttet til det sentrale Barentshavet. Området tjener likevel som et generelt beiteområde først og fremst for kvitnos (Lagenorhynchus albirostris), men også for vågehval (Balaenoptera acutorostrata) og knølhval (Megaptera novaeangliae) (Leonard og Øien 2020). Nise (Phocoena phocoena) som holder til langs kysten av Kola og Nord-Norge kan også ha beitevandringer ut til Sentralbanken.
Sjøfugl
Området er endepunkt for flere av bestandene under svømmetrekket for lomvi (Uria aalge) og polarlomvi (Uria lomvia) fra de arktiske øyene og fra fastlandet (bl.a. Erikstad mfl. 2018), basert på data fra SEATRACK (http://seatrack.seapop.no/). Lomvi som hekker i kolonier sørover langs norskekysten drar til dette området på høsten. Området er også viktig for overvintring for disse artene. I tillegg dekker dette deler av høst- og overvintringsområdene til lunde (Fratercula arctica) og polarmåke (Larus hyperboreus), samt krykkjebestandene (Rissa tridactyla) på vårparten (Fauchald mfl. in prep).
De nevnte artene bruker større deler av Barentshavet, men i dette området samler flere av bestandene seg store deler av året. Det er særlig viktig som endepunkt for svømmetrekket etter hekkesesongen for lomvi og polarlomvi, og som overvintringsområde for flere arter.
9 - Resultater for Norskehavet
9.1 - Oversikt over særlig verdifulle områder i Norskehavet
Det er beskrevet åtte særlig verdifulle områder, NH1–NH8, i Norskehavet (figur 9.1, tabell 9.1). Lokalisering, beskrivelser og miljøverdivurderingene for hvert av disse er beskrevet i kapitlene 9.3.–9.10.
Det er gjennom vurderingene foretatt nye avgrensninger av eksisterende SVO-er, bl.a. gjennom utvidelser av kandidatområder, og noen forskyvninger mellom eksisterende områder. I tillegg er to nye områder, NH4 og NH8, identifisert. Avgrensningene omtales nærmere under hvert område.
Figur 9.1. Forslag til særlig verdifulle og sårbare områder (SVO) i Norskehavet. To eksisterende SVO-er er uendret og ligger nord og vest i Norskehavet (NH1-Havis Framstredet og NH2-Vesterisen). To forslag til nye SVO-er ligger i det sentrale Norskehavet (NH4-Midtatlantisk rygg, NH8-Dyphavsområdene i Norskehavet), ett foreslått endret SVO ligger rundt Jan Mayen (NH3-Jan Mayen), ett foreslått endret SVO ligger langs kontinentalskråningen (NH5-Eggakanten nord) og to foreslått endrete SVO-er ligger langs kysten av Midt-Norge (NH6-Kystsonen Norskehavet nord, NH7-Kystsonen Norskehavet sør). Grensene for forvaltningsplanområdet er vist med brune linjer.
Forslag SVO-kode Miljøverdi-rapport 2021
SVO-kode Meld. St. 2019-2020
SVO-navn Meld. St. 2019-2020
Forslag SVO-navn Miljøverdirapport 2021
Forslag til endringer
NH1
NH11
Havis Framstredet
Havis Framstredet
Uendret
NH2
NH12
Vesterisen
Vesterisen
Uendret
NH3
NH8
Jan Mayen
Jan Mayen
Utvidet med Kandidatområdet
NH4
NH6
Den arktiske front
Midtatlantisk rygg
Nytt område. Overlapper delvis med SVO Den arktiske front som ikke lenger anses som et SVO
NH5
NH7
Eggakanten, sør
Eggakanten, sør
Starter i nord ved grensen til forvaltningsplan-området for Barentshavet, ellers uendret
NH6
NH1
Iverryggen
Kystsonen Norskehavet nord
Inkludert i Kystsonen Norskehavet nord
NH6
NH2
Haltenbanken
Kystsonen Norskehavet nord
Inkludert i Kystsonen Norskehavet nord
NH6
NH3
Sklinnabanken
Kystsonen Norskehavet nord
Inkludert i Kystsonen Norskehavet nord
NH6
NH5
Froan med Sularevet
Kystsonen Norskehavet nord
Inkludert i Kystsonen Norskehavet nord
NH6/NH7
NH9
Kystsonen Norskehavet
Kystsonene Norskehavet sør/ nord
Inkludert i Kystsonen Norskehavet sør/nord
NH6
NH10
Remman
Kystsonen Norskehavet nord
Inkludert i Kystsonen Norskehavet nord
NH7
NH4
Mørebankene
Kystsonen Norskehavet sør
Inkludert i Kystsonen Norskehavet sør. Utvidet nordover slik at området ligger inntil Kystsonen Norskehavet nord
NH8
Dyphavsområdene i Norskehavet
Nytt område
NH6
Den arktiske front
Utelatt, opprinnelig begrunnelse med forhøyet produksjon er ikke tilstrekkelig dokumentert
Tabell 9.1 Samlet oversikt over koder for foreslåtte SVO-er i foreliggende rapport, sammenlignet med koder og navn på eksisterende SVO-er i Norskehavet (fra Meld. St. 20 (2019-2020)), navneforslag for de foreslåtte SVO-ene og kort beskrivelse av forslag til endringer i grensesetting.
9.2 - Generell beskrivelse av økosystem Norskehavet
Utstrekning
Norskehavet ligger i det nordøstlige Atlanterhavet. Mot sør grenser det til den grunnere Nordsjøen langs 62˚N breddegraden mellom Norge og Færøyene, og mot nordøst grenser det til det grunnere Barentshavet (figur 9.2). Norskehavet dekker mer enn 1,1 millioner km 2 , og består av to dype bassenger, Norskehavsbassenget og Lofotenbassenget. Disse er mellom 3000 og 4000 m dype, og er adskilt av Vøringplatået (mellom 1000 og 3000 m dypt). Mot vest danner bassenghellingen overgangen til det grunnere Islandshavet i sør og Grønlandshavet i nord. Det er hovedsakelig tre vannmasser i Norskehavet: atlanterhavsvann, arktisk vann og kystvann. Hvert eneste sekund strømmer om lag 8 millioner tonn varmt og salt vann inn i Norskehavet fra det nordlige Atlanterhavet.
Figur 9.2. Kart over Norskehavet med bunntopografi og viktigste havstrømmer (atlanterhavsvann - rødt, arktisk vann - blått, kystvann - grønt).
Havstrømmer og vannmasser
Lengst vest i De nordiske hav strømmer det kaldt og ferskere vann sørover med Øst-Grønlandsstrømmen. Denne strømmen avgir arktisk vann til sidegrener inn mot Grønlandshavet og Islandshavet, og dette fører til at det sørlige Norskehavet har relativt kaldt og ferskt vann i vest. Der atlantiske og arktiske vannmasser møtes, dannes det en front, den arktiske fronten, hvor det er store temperaturendringer på tvers av fronten. Variasjoner i egenskapen og mengden av atlanterhavsvann og arktisk vann som strømmer inn i Norskehavet har stor betydning for klimaet i Norskehavet. Innstrømmende atlanterhavsvann bringer med seg varme og store mengder næringssalter som sammen med næringssaltene som strømmer ut av Polhavet og vertikalblandingen i havområdet om vinteren, utgjør grunnlaget for den biologiske produksjonen i området. Langs norskekysten, fra overflaten og ned til 50–100 m dyp, strømmer Den norske kyststrømmen nordover (figur 9.2). Den har sin opprinnelse hovedsakelig fra Østersjøen og ferskvannsavrenning langs norskekysten, og dette gjør at kystvannet har relativ lav saltholdighet. Kystvannet har også mindre næringssalter sammenlignet med atlanterhavsvannet. Havstrømmene og klimaet i Norskehavet er i stor grad påvirket av storskala atmosfæreforhold. I det sørlige Norskehavet er for eksempel vestlig utbredelse av atlanterhavsvann (dvs. beliggenheten av den arktiske fronten) knyttet til styrken av sørvestlige vinder (Blindheim mfl. 2000). År med mye sørvestlige vinder gir mindre vestlig utbredelse av atlanterhavsvann og mer arktisk vann.
Kort om økosystem og -ressurser
Omrøring av vannmasser med ulik temperatur og saltholdighet på grunn av vind og vinterblanding, har stor betydning for utbredelsen av plankton og fisk i området. Dyreplankton som raudåte og krill, finnes i store mengder og er viktig mat for fiskebestandene vi driver fiske på; norsk vårgytende sild (sild) (Clupea harengus), kolmule (Micromesistius poutassou) og makrell (Scomber scombrus). Etter gyting vandrer makrellen til Norskehavet for å beite og blir der hele sommeren før den trekker seg tilbake. Makrellen har i flere av de siste årene hatt hovedområder under den aktive beiteperioden om sommeren mer sentralt og lenger nord i Norskehavet enn tidligere perioder, inkludert havområdene sør og sørøst av Jan Mayen og sørvest av Svalbard. Kolmulen gyter i stor grad vest av Storbritannia og beiter i Norskehavet. Silda beiter i Norskehavet om sommeren, men gyter langs norskekysten. S ildeyngel vokser for det meste opp i Barentshavet og 3 – 5 årige sild kommer tilbake til den store norskehavsbestanden. Sjøfuglene som hekker ved dette havområdet, er avhengig av produksjonen av dyreplankton, fiskelarver og yngel. Hval og sel beiter også på den rike plankton- og fiskeproduksjonen. Endringene i fordeling og mengde av dyreplankton vil kunne påvirke bl.a. beitevandringen til pelagiske fisk og generell mattilgang til høyere trofiske nivåer.
Plankton i forvaltningsplanområdet
Oppblanding av næringssalter fra dypvannet gir grunnlag for økt biomasse av planteplankton. Også frontsystemer vil kunne gi grunnlag for økt biomasse og produksjon, da «sammenstøt» av ulike vannmasser kan resultere i innblanding av næringssalter til overflaten. I Norskehavet er dette særlig aktuelt over den midtatlantiske ryggen og over sokkelen innenfor Eggakanten. Der fronter og topografiske forhold sammenfaller vil man kunne få særlig stor produksjon og biomasse av planteplankton. I slike områder vil også planteplanktonproduksjon være mer stabilt høy, sammenlignet med åpne havområder. Inne ved kysten vil man også ha tilførsel av næringssalter fra land, sammen med ferskvann, som resulterer i tidlig og stor planteplanktonproduksjon.
Planteplanktonproduksjon lokalt og adveksjon (forflytning) fra omkringliggende områder vil være en viktig miljøverdi i alle hav- og kystområder som basis i marine næringsnett. Som primærprodusent vil de i stor grad påvirkes av fysiske forhold (lagdeling, temperatur, lysforhold), kjemiske forhold (næringssalter) og biologiske interaksjoner (Skjoldal og Rey 1989). For sistnevnte vil blant annet beitetrykket fra dyreplankton være en viktig faktor som påvirker planteplanktonet negativt ved at de blir spist, men det vil også kunne være positivt for enkelte arter gjennom regenerering av næringssalter. Planteplankton som element, har en økosystemfunksjon som basis for marine næringskjeder. Planteplankton er til stede i alle våre havområder og vil gjennomgå markante endringer gjennom året både når det gjelder biomasse og sammensetning og vil også vise stor variasjon mellom årene (Dale mfl. 1999). Planteplanktonartene vil være til stede over store områder (større enn et SVO), har høy vekstrate i perioder og vil til enhver tid utnytte de fysiske og kjemiske forholdene. Arter som er knyttet til is er unike i forhold til åpent isfritt hav. På grunn av elementet «Planteplankton» sin store variasjon i tid og rom og deres fritt flytende pelagiske levesett, er det vanskelig å benytte i en utredning omkring miljøverdier inne i enkeltområder.
Dyreplankton fungerer som et ledd mellom planteplankton på den ene siden og fisk, hval og andre organismer høyere opp i næringskjeden på den andre (Melle mfl. 2004). Herbivort dyreplankton, som bl.a. inkluderer Calanus-artene og en rekke andre arter av hoppekreps, spiser planteplankton og overfører biomasse dannet ved fotosyntese til dyreplanktonbiomasse. Det finnes også en rekke karnivore dyreplanktonorganismer som lever av annet dyreplankton, eksempler er amfipoder, pilorm og maneter. Krill regnes i hovedsak som omnivore, dvs. den spiser både plante- og dyreplankton. Dyreplanktonet i Norskehavet inngår i et komplekst nettverk av næringskjeder, der for eksempel karnivort dyreplankton fungerer som næring for høyere trofiske nivåer, men samtidig konkurrerer med høyere trofiske nivåer om å beite på herbivort dyreplankton (Skjoldal 2004).
Raudåte (Calanus finmarchicus), som utgjør hovedmengden av dyreplanktonbiomassen i Norskehavet, finnes i atlantiske- og atlantisk/arktiske vannmasser, men har de dype havbassengene i sentrale Norskehavet som sine kjerneområder (Sundby 2000, Broms mfl. 2009, Melle mfl. 2014). Raudåte fra de dypekjerneområdene transporteres med strømmene til omkringliggende hav- og sokkelområder, inkludert Barentshavet, Nordsjøen og Norskekysten. Den årlige produksjonen av raudåte starter tidligst i kystvannet i sørøstlige Norskehavet, og senere i atlantisk og arktisk vann mot nordvest (Broms mfl. 2007, Bagøien mfl. 2012). I de arktiske vannmassene vest i Norskehavet er det store mengder C. hyperboreus, som har Grønlandshavet som sitt kjerneområde (Hirche 1991, 1997). Krill er en annen viktig gruppe, og arten Meganyctiphanes norvegica (storkrill) har utbredt fordeling, men med høyest antall i atlanterhavsvann i østlige Norskehavet og kontinentalskråningen. Thysanoessa inermis (småkrill) er hovedsakelig en sokkelart og finnes over den norske kontinentalsokkelen og i kaldere subarktisk vann i vestlige områder ved Jan Mayen. T. longicaudata (småkrill) er vanlig over det meste av Norskehavet, den finnes særlig i atlantisk og subarktisk vann i sør og vest. Amfipoden Themisto abyssorum finnes i atlantiske og subarktiske vannmasser, mens den større T. libellula er en typisk arktisk art som finnes i vestlige Norskehavet. Andre viktige dyreplanktongrupper er pilorm, maneter, ribbemaneter, snegler, pelagiske reker, vannlopper og muslingkreps (for detaljert beskrivelse, se Melle mfl. 2004).
Biomassen av dyreplankton i Norskehavet om våren har generelt vært redusert fra tidlig på 2000-tallet, og var på det laveste nivået i 2010 før den har stabilisert seg, med en mulig økning i løpet av de siste ti årene (ICES 2020 a, 2021). Reduksjonen har vært størst i de vestlige arktiske delene av Norskehavet, og storskalafordelingen av biomasse har endret seg fra å være høyest i vest, til nå å være jevnere fordelt mellom vannmassene (ICES 2020 a, 2021). Mengden raudåte og C. hyperboreus har gått ned i sørlige deler av Norskehavet (Dupont mfl. 2017, Kristiansen mfl. 2019), det er ikke gjort tilsvarende undersøkelser i nordlige områder. Nedgangen i dyreplankton sammenfaller med høyere varmeinnhold i Norskehavet og redusert tilstrømning av arktisk vann i det sørvestlige Norskehavet (Skagseth og Mork 2012, Kristiansen mfl. 2019, ICES 2020 a, 2021). Store bestander av planktonspisende fisk kan også være en årsak til nedgangen i dyreplanktonbiomassen (Huse mfl. 2012). I Norskehavet forekommer tempererte arter foreløpig i lavt antall og vil i liten grad påvirke økosystemet (Miljostatus.no).
9.2.1 - Pågående og framtidige endringer i Norskehavet
Generell informasjon om klimaendringer, med konsekvenser for temagruppene, samt forsuring er gitt i kap. 5.1. I Norskehavet ventes det høyere overflatetemperatur i dyphavsområdene som følge av klimaendringer fra økt drivhuseffekt, men naturlige variasjoner vil i stor grad dominere over klimasignalet de neste 10–20 årene (Hanssen-Bauer mfl. 2015). Global oppvarming vil kunne føre til at Atlanterhavsstrømmen langs eggakanten svekkes, men observasjoner viser så langt ikke noen tegn på en reduksjon av strømmen (Hanssen-Bauer mfl. 2015, Oziel mfl. 2020, Tsubouchi mfl. 2021). På grunn av Norskehavets store dyp og tilførsel av vann både fra sørvest og nord, er det ventet at temperaturendringer vil skje saktere i de store og dype bassengene enn i sokkelhavene Nordsjøen og Barentshavet.
Over de siste tiårene har det vært en rask nedgang i havis i Framstredet. Dette er en av flere viktige indikatorer på klimaendringer og global oppvarming (Carmack mfl. 2015). Isdekket reduseres både i utbredelse og tykkelse, mengden flerårsis minker raskt og smeltesesongen utvides (Spreen mfl. 2011, Carmack mfl. 2015, Onarheim mfl. 2018, Spreen mfl. 2020). Isdekket blir også mer mobilt (Rampal mfl. 2009, Kwok mfl. 2013), og det vil trolig fortsette å være stor utstrømming av havis i Framstredet i de kommende år. Det er svært sannsynlig at den nedadgående trenden for havis vil fortsette, gitt utviklingen i utslipp av CO2, og at dette vil fortsette å endre klimaet i Arktis (IPCC 2019) . Klimaindusert endring i sesongmessig utbredelse av havis og lagdeling i havet endrer vilkårene for en rekke arktiske og boreale arter, inkludert marine pattedyr, og har stor påvirkning på økosystemene.
Isbiota
Se kapittel 9.3.6 under foreslått SVO Havis Framstredet.
Plankton
Dyreplanktonbiomassen i Norskehavet har virket stabil de siste ti årene (ICES 2019, 2021), og det vil derfor trolig ikke være vesentlige endringer i biomasse eller produksjon i nær fremtid. Det har tidligere vært en reduksjon i både biomassen generelt i Norskehavet og mengden Calanus finmarchicus i sørlige deler av Norskehavet, der sistnevnte er basert på studier fra Svinøysnittet og områder nord for Færøyene (Dupont mfl. 2017, Kristiansen mfl. 2019), men fra ca. 2010 virker det derimot å ha vært en økning igjen i biomassen, og mengden C. finmarchicus er antatt å øke noe i nær fremtid (Kjesbu mfl. innsendt). I sørlige Norskehavet lever Calanus helgolandicus og C. finmarchicus i utkanten av sitt utbredelsesområde, og økt havtemperatur vil kunne gi redusert mengde C. finmarchicus og økt mengde C. helgolandicus, tilsvarende situasjonen i Nordsjøen (Beaugrand mfl. 2002). Dette vil kunne påvirke produksjonen og sesongmønsteret. I foreslått SVO Kystsonen Norskehavet sør er det registrert lav forekomst av varmekjære arter (Miljostatus.no), og en kan anta det samme for foreslått SVO Kystsonen Norskehavet nord. Dersom havtemperaturen fortsetter å stige som den har gjort de siste ti-årene, vil mengden og antall arter av varmekjært dyreplankton øke.
Fisk
Mens gytefelter vil forandre seg lite, kan vanntemperatur, transport og matforsyning bli påvirket av klimaendringer, enten av større eller hyppigere uoverensstemmelse av vårblomstring og gytetid eller av en endret atlanterhavsstrøm og derfor raskere eller tregere transport langs kysten og eggakanten (Solemdal mfl. 1997, Loeng 2006, Hylen mfl. 2008, Dragesund mfl. 2008, Sundby and Nakken, 2008, Sundby mfl. 2013). Om ikke dyreplankton av rett kvalitet (rett livsstadie av tilstrekkelig næringsrik art) er til stede, vil larvene ha lav overlevelse (Cushing 1990).
Mesopelagisk fauna og bunnsamfunn
Klimaendringer vil særlig påvirke bunnlevende organismer i dype områder i relativ nær fremtid direkte gjennom temperaturøkninger, hypoksi og havforsuring. Dette er beskrevet i kapittel 5.
Sjøpattedyr
Mindre is og dårligere iskvalitet kan føre til videre nedgang i bestandene av grønlandssel (Pagophilus groenlandicus) og klappmyss (Cystophora cristata) (Haug og Biuw 2019).
Narhval (Monodon monoceros) foretrekker kalde arktiske vannmasser (0,6–1,5 °C; Heide-Jørgensen mfl. 2020) og studier av effekter av tidligere tiders klimaendringer på fylogeografi (sammenheng mellom genetisk fordeling og artsutvikling formet gjennom lang tids geografiske endringer) og demografi, tyder på at denne arten responderte kraftig på temperaturendringer og endringer i isutbredelse og vil antakelig gjøre det igjen som følge av dagens endringer i disse parameterne (Louis mfl. 2020). Andre utfordringer for narhvalene som følge av redusert isdekke er økt predasjon fra spekkhoggere, noe som allerede er dokumentert i Arktisk Canada (Kovacs mfl. 2011, Breed mfl. 2017, Lefort mfl. 2020), og forstyrrelser fra økt skipstrafikk (Reeves mfl. 2014).
De pågående økningene i havtemperaturen samtidig med reduksjon i isutbredelsen kan også bli kritiske for grønlandshvalene (Balaena mysticetus) både på grunn av tap av habitat og også termisk stress, i tillegg til en økning i predasjon fra spekkhoggere (Orcinus orca), noe som allerede er et alvorlig problem for grønlandshvalbestanden i Okhotskhavet (Kovacs mfl. 2011, Shpak og Paramonov 2018).
Sjøfugl
Sjøfugl er sårbare for endret næringstilgang. Om dyreplankton, fiskelarver og -yngel ikke opptrer i rett tid for sjøfuglenes hekkesesong, vil det ha negativ effekt og lav ungeoverlevelse. Framtidsutsiktene for sjøfugl er derfor tett knyttet til dyreplankton og fiskens valg av gyteplass- og -tid.
9.2.2 - Løpende kartleggings- og overvåkingsaktivitet
Norskehavet
Forslag SVO
Plankton
Fisk
Bunnsamfunn
Mesopelagisk fauna
Sjøfugl
Sjøpattedyr
Frekvens
Nasjonale program
Mareano-programmet (Mareano.no)
NH3-NH8
X
Årlig, rullerende
SEAPOP-programmet (Seapop.no)
NH1, NH3 NH5-NH7
X
Årlige
SEATRACK-programmet (seatrack.seapop.no)
NH1, NH3 NH5-NH7
X
Årlige
Havforskningsinstituttet
Overvåkingstokt
Økosystemtoktet Norskehavet
NH2-NH8
X
X
X
Årlig
Dyp-pelagisk tokt i Norskehavet
NH8
X
Hvert 3. år
Kysttokt
NH6-NH7
X
X
X
Årlig
Gytetokt, NVG-sild
NH6-NH7
X
Årlig
Sildelarvetokt langs kysten
NH6-NH7
X
Årlig
Eggtokt, kyst
NH6-NH7
X
Årlig
Int. gytetokt kolmule (IBWSS)
NH4, NH8
X
Årlig
Makrellmerketokt på gytefelt
NH5-NH6
X
Årlig
Egga sør
NH5
X
Hvert 2. år
Biologisk prøvetaking av grønlandssel
NH1-NH2
X
Hvert 5. år
Bestandsestimering av grønlandssel og klappmyss
NH1-NH2
X
Hvert 5. år
Økologisk overvåking av grønlandssel-bestanden
NH1-NH2
X
Hvert 3. år
Tareundersøkelser
NH6-NH7
X
Årlig, rullerende
Oseanografiske snitt
Svinøy, nordvest
NH4, NH7-NH8
X
5x årlig
Norsk Polarinstitutt
Oseanografisk, inkl. havisovervåking
NH1
Kontinuerlig
Fram Strait Arctic Outflow Observatory
NH1
Kontinuerlig
Tabell 9.2. Tabellen viser et utvalg av løpende kartleggings- og overvåkingsprogram, tokt og oseanografiske snitt, der foreslåtte SVO-er i Norskehavet helt eller delvis inngår i kartleggingen/overvåkingen. Foreslåtte SVO-er og miljøverdier som kartlegges/overvåkes er ført opp i tabellen.
9.3 - SVO Havis Framstredet (NH1)
9.3.1 - Sammendrag
Områdebeskrivelse og strukturer
Det eksisterende SVO Havis Framstredet (NH1) ligger i den østlige delen av Framstredet, nærmere Svalbard. Miljøforholdene i Framstredet er dominert av Øst-Grønlandstrømmen med utstrømmende ferske og kalde vannmasser fra Polhavet på vestsiden og nordgående innstrømning av varmt og salt atlanterhavsvann på østsiden. Havis strømmer også ut med Øst-Grønlandstrømmen. Det er også noe resirkulering av atlanterhavsvann i Framstredet.
Havisen i SVO Havis Framstredet har ulik opprinnelse og er derfor veldig variabel. Isen i østlige deler ligner i Barentshavet, mens i nord og vest er isen eldre og tykkere.
Miljøverdier
Primærproduksjonen i isen starter tidligere enn i vannmassene, noe som bidrar til å forlenge den produktive sesongen i området. Planteplanktonproduksjon og biomasse vil i perioder være høye sammenlignet med nærliggende åpne havområder. Stor variasjon i istyper medfører også mange ulike typer av issamfunn. Atlantisk dyreplankton som blir transportert med atlantiske vannmasser fra Norskehavet til Arktis via Framstredet, kan være viktig for mesopelagiske planktonspisende predatorer i det arktiske bassenget. Iskantsonen i Framstredet er viktig for den truede ismåken før hekkesesongen starter og om høsten. Både grønlandshval (kritisk truet–CR) og narhval (sterkt truet–EN) finnes i drivisen i Framstredet året rundt. Spitsbergenbestanden av grønlandshval har nordvestre del av Framstredet som et forplantingsområde. Flere unike populasjoner av marine pattedyr er avhengig av havis i Framstredet. Utryddelse av disse artene vil representere tap av viktig biologisk mangfold. Isen og området i iskantsonen er i veldig liten grad direkte påvirket av menneskelig aktivitet.
Grønlandshval. Foto: N. Cobbing, NP
Figur 9.3.1. Kart over SVO Havis Framstredet (NH1). Grensene for SVO-et er vist med tykk rød strek, mens grenser for overlappende og nærliggende foreslåtte SVO-er er vist med tynnere rød eller oransje strek. Grensene for forvaltningsplanområdet er vist med brun linje. Kartet viser utbredelsen av utvalgte miljøverdier som er av betydning for SVO-et. Kun grønlandshval er markert på kartet, men området har også stor verdi for flere andre sjøpattedyr.
9.3.2 - Vurdering i forhold til EBSA-kriteriene - SVO Havis Framstredet (NH1)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i SVO Havis Framstredet (NH1)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet/sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Sjøfugl
Isbiota Sjøpattedyr
Issamfunn har en egenverdi i seg selv både mht samfunnstyper (Syvertsen 1991, Bluhm mfl. 2017a) og artssammensetning, samt betydning for produksjon som kanaliseres videre i næringsnettet (Søreide mfl. 2010, 2013, Leu mfl. 2011, Kohlbach mfl. 2016, 2017, Wang mfl. 2015). At havisen forekommer over dyphav og har eldre is enn størstedelen av SVO Iskantsonen (von Quillfeldt mfl. 2018) fører dessuten til helt særegne samfunn og forekomst av flere endemiske isavhengige arter, f.eks. arter av nematoder og amfipoder (Horner mfl. 1992, Werner og Gradinger 2002, Arndt og Swadling 2006, Tschesunov 2006, Poulin mfl. 2011, Bluhm mfl. 2017b, Hop mfl. 2020).
Drivis i Framstredet er eneste kjente forplantningsområde for Spitsbergenbestanden av grønlandshval og er viktig habitat for narhval gjennom hele året (Stafford mfl. 2018, Ahonen mfl. 2019).
Dette er et svært viktig område for ismåke fra store deler av Arktis (Gilg mfl. 2010).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Isbiota Sjøfugl Sjøpattedyr
Større andel av flerårsis sammenlignet med Barentshavet gjør dette området særdeles viktig for arter med hele livssyklusen i eller på undersiden isen (f.eks. hjuldyr, amfipoder, kopepoder m.m.) (Arndt og Swadling 2006, Bluhm mfl. 2017b, Hop mfl. 2020).
Drivis i Framstredet er eneste kjente forplantningsområde for Spitsbergenbestanden av grønlandshval (Stafford mfl. 2018).
Dette er et svært viktig område for ismåke fra store deler av arktis (Gilg mfl. 2010).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Isbiota Sjøfugl Sjøpattedyr
Isbiota har havis som habitat. Både type og mengde havis er gjenstand for raske endringer som følge av klimaendringer (Hansen mfl. 2013, Hansen mfl. 2014, Renner mfl. 2014, Webster mfl. 2014, Krumpen mfl. 2016, von Quillfeldt mfl. 2018) og påvirker dermed forekomst og overlevelse av arter, samt biologiske prosesser i isen (Fernández-Méndez mfl. 2015, Bluhm mfl. 2017b, Bluhm mfl. 2018, Kauko mfl. 2018, von Quillfeldt mfl. 2018, Hop mfl. 2020).
Drivis i Framstredet er eneste kjente forplantningsområde for Spitsbergenbestanden av grønlandshval og et viktig habitat for narhval gjennom hele året (Stafford mfl. 2018, Ahonen mfl. 2019). Begge disse artene er rødlistet.
Området er viktig for flere sjøfugl og særlig for ismåke (sårbar–VU på rødlista for Svalbard).
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Sjøfugl
Isbiota Sjøpattedyr
Isbiota, særlig arter i flerårsis vil være utsatt ved klimaendringer. Iskonsentrasjonen i SVO Havis Framstredet har minket med om lag -10 % per tiår i september og -8 % per tiår i mars i perioden 1979–2016 (Onarheim mfl. 2018 – deres figur 1a og 1b). Tidspunkt for issmelting har blitt forskjøvet med 0 til 5 eller 10 dager tidligere per tiår, avhengig av lokasjon i området (Stroeve og Notz 2018 – deres figur 3). Tilsvarende har tidspunkt for isdannelse blitt forskjøvet med 5 til 20 dager senere per tiår. Redusert isutbredelse, sen isdannelse/tidlig smelting gir kortere produktiv sesong for isbiota (Wassmann og Reigstad 2011, Hop mfl. 2013, Barber mfl. 2015), men tynnere is kan gi bedre lysforhold noe som vil kunne øke produksjonen (Leu mfl. 2015). Mengde isbiota som transporteres via den transpolare driften gjennom Framstredet påvirkes også (Gradinger mfl. 2009, Soltwedel mfl. 2016, Krumpen mfl. 2019). Iseksporten gjennom Framstredet har økt (Smedsrud mfl. 2017) noe som kan påvirke oppholdstid for isamfipoder i Framstredet (Arndt og Pavlova 2005) samt eksport av ismeiofauna inn i Grønlandshavet (Gradinger mfl. 1999), men det kan være store variasjoner i iseksport mellom år og gjennom året (Ricker mfl. 2018).
Klimaendringer påvirker havis i stor grad i Framstredet og dermed reduseres kvaliteten på habitatet til arktisk endemiske marine pattedyr direkte og indirekte via potensialet for industriell aktivitet i området (Laidre mfl. 2015, Reeves mfl. 2014).
Området er svært viktig for ismåke, særlig vår og høst (Gilg mfl. 2010).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Planteplankton
Isbiota
Primærproduksjonen i isen starter tidligere enn i vannmassene noe som bidrar til å forlenge den produktive sesongen i et område (Hegseth 1998, Leu mfl. 2011, 2015).
Planteplanktonproduksjon og biomasse vil i perioder være høye sammenlignet med nærliggende åpne havområder (Nöthing mfl. 2020).
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Isbiota Sjøpattedyr
Stor variasjon i istyper (Krumpen mfl. 2016, 2019) fører til mange ulike typer av issamfunn i Framstredet (von Quillfeldt mfl. 2018). Isbiota utgjør en rekke ulike samfunnstyper og består av et høyt antall arter fra en rekke ulike artsgrupper (Syvertsen 1991, Horner mfl. 1992, Gradinger mfl. 2010, Poulin mfl. 2011, Bluhm mfl. 2011, Bluhm mfl. 2017a, b, Hop mfl. 2020).
Flere unike populasjoner av marine pattedyr er avhengig av havis i Framstredet (Laidre mfl. 2015). Utryddelse av disse artene vil representere tap av viktig biologisk mangfold.
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
Sjøfugl
Isbiota Sjøpattedyr
Med unntak av de pågående klimaendringer, så er isbiota lite påvirket av menneskelig aktivitet (Eamer mfl. 2013).
Isdekkede områder i Arktis er ganske uberørte marine pattedyrhabitater - essensielle for arktiske endemiske marine pattedyr og sjøfugl sterkt knyttet til iskantsonen, som ismåke (Gilg mfl. 2010).
9.3.3 - Lokalisering
SVO Havis Framstredet er tidligere definert i Meld. St. 20 (2019-2020). Området omfatter havisen i nordlig del av Norskehavet, og er en fortsettelse vestover av SVO Iskantsonen i Barentshavet figur 9.3.1). Havisen i dette SVO-et er dessuten en del av et større område med havis i Framstredet som strekker seg helt til Grønland og som også SVO Vesterisen er en del av.
Avgrensningen av havisen i Framstredet, som er et særlig verdifullt og sårbart område, er i forvaltningsplanen for Norskehavet satt der det forekommer havis 15 prosent av dagene i april (15 prosent isfrekvens), basert på satellittobservasjon av isutbredelse fra 30-årsperioden 1988–2017. Denne grensen kan oppdateres med nye isdata ved senere oppdateringer av forvaltningsplanene (Meld. St. 20 (2019-2020)). Fagekspertgruppen foreslår ingen endring i denne definisjonen for avgrensning for SVO Havis Framstredet.
SVO Havis Framstredet overlapper med Molloydypet, en del av foreslått nytt SVO Midtatlantisk rygg, og ligger tett opp mot grensen til de foreslått endrete SVO-ene Iskantsonen og Havområdene rundt Svalbard (figur 9.3.1).
9.3.4 - Introduksjon til området
Den vestlige delen av Framstredet er dominert av havis som blir eksportert sørover fra Polhavet, med Øst-Grønlandsstrømmen (Kwok 2004, Spreen mfl. 2020). Denne isen er ofte en blanding av is fra sibirske havområder (Kara- og Laptevhavet og Øst-Sibir), den transpolare driften, Beaufort-virvelen i Polhavet og regionalt dannet havis (Hansen mfl. 2013, 2014, 2015, Zamani mfl. 2019). Derfor består isen av istyper med ulik alder fra ung is til gammel flerårsis. Flerårsis har lavere saltholdighet enn førsteårsis (Notz 2009). Særlig eldre istyper kan ha skrugarder av betydelig tykkelse. Isen i iskantsonen kan være påvirket av smelteprosesser, som når den møter det varme nordgående atlanterhavsvannet. Siden isdynamikken i Framstredet er stor, kan isen være utsatt for smelting i relativt kort tid (få dager til få uker) før den driver ut av området, sammenlignet med Barentshavet. Isen i Framstredets iskantsone er i gjennomsnitt tykkere enn i Barentshavet (Vinje mfl. 1998, Kwok 2009). Havisen i nordlig del av Norskehavet dekker langt dypere områder enn i Barentshavet. Innblandet i havisen er isfjell fra kalvende breer på Grønland.
Generelt viser isen i Framstredet større snøtykkelse enn isen i Barentshavet. Det kan være flere faktorer som bidrar til dette, både regionale forskjeller i nedbør, og fordeling av istyper med mer flerårsis i Framstredet som kan samle mer snø om høsten enn årsis som dannes først i løpet av vinter og vår (Kwok og Cunningham 2008, Forsström mfl. 2017, Webster mfl. 2019, Merkouriadi mfl. 2020).
I Grønlandshavet og vestlige del av Framstredet er det atlantiske vannmasselaget betydelig kaldere enn lenger øst, og tetthetsforskjellen mellom det arktiske og atlantiske laget er mindre markant enn i Barentshavet. Men også i Framstredet kan det dannes et ferskt, stabilt overflatelag i smeltesesongen, med varierende tykkelse og grad av isolasjon fra de dypereliggende vannmassene.
Utbredelsen av havis i Framstredet har de fleste år et maksimum i april og minimum i september. Hovedutviklingen er at isutbredelsen har en klar negativ trend gjennom perioden med satellittovervåkning (1979–2020), og minker med −6,0 prosent og −10,0 prosent per dekade for henholdsvis april og september (data fra www.mosj.no). Isutbredelsen er også karakterisert med stor variasjon mellom år. Det året isutbredelsen var størst, april 1986, var to år etter en markert nedgang i april 1984. Siden den gang har mønsteret generelt sett vært at år med stor isutbredelse (større enn middelet) avløses av år med isutbredelse mindre enn middelet, samtidig som år-til-år-variasjonen minker. Den minste isutbredelsen for april i SVO Havis Framstredet var i 2004. Det er også stor år-til-år variasjon for september måned, i hele perioden med satellittovervåkning. Den minste isutbredelsen for SVO Havis Framstredet var i september 2004 og 2017, med nesten like verdier. For mer informasjon se www.mosj.no/no/klima/hav/havisutbredelse.
Figur 9.3.2. Gjennomsnittlig overflatestrøm (piler) og bunndyp (fargekodet) i SVO Havis Framstredet og omkringliggende områder.
pH i området varierer fra sør til nord, fra rundt 8,1 til 8,15, i overflatevannet. Metningsgraden for aragonitt og kalsitt ligger på hhv. 1,8 og 2,8 i de sørlige delene, og avtar nordover og vestover, til 1,5 og 2,5. Måleserier fra Grønlandshavet viser en signifikant trend i pH på -2,2 milli-pH per år siden 1990-tallet, mens i Framstredet er trenden litt sterkere, -2,5 milli-pH per år (Fransner mfl.mfl. 2020). Aragonitt- og kalsittmetning avtar også. Totalt har havforsuring redusert pH i overflatevannet med 0,12 enheter siden den industrielle revolusjon, og metningsgraden for aragonitt og kalsitt med 0,5 og 0,9 (Lauvset mfl.mfl. 2016, Fransner mfl.mfl. 2020). Tilgjengelige data (Olsen mfl.mfl. 2020) viser at metningshorisonten til aragonitt ligger på rundt 2000 m, mens vannsøylen er overmettet med kalsitt helt til bunns.
9.3.5 - Beskrivelse av miljøverdiene
Isbiota
SVO Havis Framstredet (og SVO Vesterisen) er en liten del av et større område med havis i Framstredet, og isbiotaen i den delen som utgjør SVO-et skiller seg ikke særlig fra nærliggende områder. Se foreslått endret SVO Iskantsonen i Barentshavet for mer omtale av isbiota (Kap. 8.4.5).
Dannelsen av et overflatelag er med på å fremme våroppblomstringen i iskantsonen (Cherkasheva mfl. 2014). Denne er dominert av kiselalger, særlig på sokkelen (Krisch mfl. 2020, Hop mfl. 2006), antageligvis som en konsekvens av adveksjon av silikatrikt vann fra Polhavet. Om sommeren er primærproduksjonen i området begrenset av tilgang på næringssalter, særlig nitrat, fordi vannet som strømmer ut av Polhavet har underskudd på dette i forhold til andre næringssalter (Krisch mfl. 2020).
Mer enn 2000 arter (både encellete og flercellete) fra en rekke artsgrupper (f.eks. virus, encellede bakterier, archaea, mikroalger, sopp, ciliater, krageflagellater, amøber, foraminiferer, flercellete hjuldyr, nematoder, hoppekreps, amfipoder m.m.) har tilknytning til havis i hele eller deler av sin livssyklus (Bluhm mfl. 2017 a,b, Hassett mfl. 2019, Ehrlich mfl. 2020, Gradinger 2020), men det er lite informasjon om biologien i akkurat den delen som utgjør SVO Havis Framstredet. Isforhold er bestemmende for forekomst av issamfunn (Horner mfl. 1992, Gradinger mfl. 1999, Gradinger mfl. 2010, Bluhm mfl. 2017a og b, Hop mfl. 2020). Stor variasjon i istyper fører derfor til at man kan forvente mange ulike typer av issamfunn i Framstredet. Ulike isalgearter dominerer flerårsis sammenlignet med årsis, og det er ofte andre arter knyttet til skrugarder enn på undersiden av isen (Hop mfl. 2020). Totalt 16 arter av amfipoder, hoppekreps og flerbørstemark er rapportert fra is og nærliggende vannmasser i Framstredet og i Barentshavet (Arndt mfl. 2009). I tillegg forekommer flere arter av amfipoder med tilknytning til havis gjennom hele livssyklusen. Senere forskning viser imidlertid at noen av disse artene også kan overleve i pelagiske vannmasser i større grad enn tidligere antatt (Berge mfl. 2012, Kunisch mfl. 2020). Det er også variasjon i artssammensetning og produksjon gjennom året og mellom år (Bluhm mfl. 2017 b), noe som for eksempel har vært dokumentert i Framstredet for hjuldyr (Gradinger mfl. 1991, Friedrich og De Smet 2000). Skrugarder har helt særegne issamfunn (Fernandez-Mendez mfl. 2018) og har stor betydning for gjennomføring av livssyklus for enkelte artsgrupper når undersiden av isen smelter om sommeren (Gradinger mfl. 2010).
Mindre sediment i isen som kommer via den transpolare driften gjennom Framstredet og samtidig tynnere is, bidrar til bedre lysforhold og dermed også mulighet for økt primærproduksjon i isen (Krumpen mfl. 2019). Økt issmelting fører også til redusert transport av isfauna fra de grunne og kystnære sibirske sokkelområdene til Framstredet, sammenlignet med tidligere (Gradinger mfl. 2009, Soltwedel mfl. 2016, Krumpen mfl. 2019, Ehrlich mfl. 2020) og påvirker også hvor mye som frigjøres fra isen til vannsøyle/havbunn og eventuelle nye issamfunn (Olsen mfl. 2017). Adveksjon av «varmt» atlanterhavsvann fremmer smelting av havis og påvirker også sammensetningen av isalger og forklarer hvorfor Melosira arctica, en typisk art i eldre og mer komplekse isalgesamfunn, kan være så godt som fraværende nord for Framstredet (til forskjell fra tidligere). Generelt vil mer årsis og ung is og mindre flerårsis resultere i mindre komplekse samfunn, noe som har vært observert i flere arktiske områder (Melnikov mfl. 2002, Krumpen mfl. 2019, Hop mfl. 2020).
Isalger utgjør en konsentrert og næringsrik matkilde for både dyr som lever i isen og pelagiske beitere (Søreide mfl. 2010, Leu mfl. 2011, Wang mfl. 2015, Kohlbach mfl. 2016). Til forskjell fra store deler av foreslått endret SVO Iskantsonen i Barentshavet, ligger havisen i SVO Havis i Framstredet over store havdyp, noe som normalt vil redusere mengden av isbiota som eksporteres til havbunnen, men også mulighet for rekruttering av arter fra havbunnen til isen (Lund-Hansen mfl. 2020). I enkelte deler av Arktis er det imidlertid observert at Melosira arctica kan eksporteres til flere tusen meters dyp og være en viktig matkilde for bunnsamfunn (Boetius mfl. 2013).
Plankton
Studier av planteplanktonbiomasse viser at det kan være stor variasjon i løpet av året, med maksimal biomasse om våren og sommeren (Nöthig mfl. 2015, Vernet mfl. 2019). Produksjonen vil i perioder være høy, der issituasjonen (Wassmann mfl. 2010) og vannmassetransport vil ha en betydning for den totale produksjon og biomassen (Vernet mfl. 2019). Det foreligger flere studier med estimert primærproduksjon for dette området (Hop mfl. 2006, Reigstad mfl. 2011) som kan fungere som referanser til framtidig utvikling. Nöthig mfl. (2020) fant at det var noe høyere biomasse av planteplankton assosiert med Vestspitsbergenstrømmen, sammenlignet med Øst-Grønlandstrømmen. I de eksisterende tidsseriene er det registrert en mindre økning i klorofyll a i deler av området (Nöthing mfl. 2019, Reigstad mfl. 2011), som er foreslått knyttet til temperatur og stratifisering samt isdekke. Høyere temperatur fører ikke nødvendigvis til økt biomasse (målt som klorofyll), men stratifisering er viktig.
Atlantisk dyreplankton blir transportert med atlantiske vannmasser fra Norskehavet til Arktis via Framstredet. Enkeltstudier har vist at mengden som blir transportert gjennom Framstredet varierer gjennom året, og kan være høyest om vinteren (Basedow mfl. 2018). Dyreplankton advektert via Framstredet til det arktiske bassenget kan være viktig for mesopelagiske planktonspisende predatorer i det arktiske bassenget (Basedow mfl. 2018).
SVO Havis Framstredet er ikke inkludert i Havforskningsinstituttets standard overvåkingsprogram, og det finnes begrenset informasjon angående dyreplankton, men det finnes noe informasjon fra enkeltstudier. Dominerende kopepoder i området er Oithona similis, Calanus finmarchicus, C. glacialis, C.hyperboreus, Microcalanus arte. og Pseudocalanus arter, og sammensetningen av disse varierer med sesongen (Basedow mfl. 2018, Svensen mfl. 2019). Små kopepoder og protozooplankton gir et betydelig bidrag til dyreplankton-antall og biomasse om høsten, når de større kopepodene har vandret ned fra de øvre 200 meterne av vannsøylen (Svensen mfl. 2011). Av karnivore dyreplankton er chaetognatha (pilorm, hovedsakelig Eukrohnia hamata) observert å dominere i antall og biomasse (Blachowiak-Samolyk mfl. 2007). Forekomst av plante- eller dyreplankton er ikke avgjørende for definisjonen av dette SVO-et.
Cliona limacina. Foto: C.Brohms, HI
Fisk
Selv om det er polartorsk (Boreogadus saida) og andre arktiske arter som har beiteutbredelse i dette området (Joiris mfl. 2014), er det i seg selv ikke regnet som tilstrekkelig relevant til at fisk kan brukes som begrunnelse for dette SVO-et.
Bunnsamfunn
Havbunnen i dette SVO-et er for det meste bløt arktisk dyphavsleire, og de bunnlevende dyrene er kjent fra kun sporadiske studier, ettersom det er svært vanskelig å ta prøver av bunnen fra en båt som ligger i is. Nedfall av død isbiota er ofte en stor andel av fødetilgangen til de bunnsamfunnene som lever under helt og delvis isdekke. Studier fra området (Hoffmann mfl. 2018) viser at det er høyere tetthet av bunndyr i områder med høyere isdekke, men at også havdyp spiller inn. Slangestjerner, flerbørstemark, hyperbentiske krepsdyr og snegler er blant de mest typiske artsgruppene på høyarktiske bløte havbunner. Slike arter er ofte begrenset av vanntemperatur i tillegg til mattilgang. Flere av disse dyrene er føde for høyere trofiske nivå, og de frigjør slik den energien som har falt ned til bunnen tilbake i vannsøylen. Ingen av disse gruppene kan bevege seg langt i løpet av en generasjon. Bunndyrsamfunnene er ikke vesentlige i forhold til definisjonen av dette SVO-et.
Sjøpattedyr
Spitsbergenbestanden av grønlandshval (Balaena mysticetus) (M1, figur 9.3.1) ble nesten utryddet som følge av hard beskatning under den første kommersielle hvalfangsten som startet i begynnelsen av 1600-tallet på Svalbard. Den lille gjenværende bestanden kombinert med disse hvalenes lange levealder (over 200 år) og langsomme livshistorie, gjør at bestanden er svært truet i dag og klassifiseres som «Kritisk truet–CR» både på den norske rødlisten og av IUCN (Vacquié-Garcia mfl. 2017b, Cooke og Reeves 2018). Akustisk overvåkning i drivisen i Framstredet mellom Svalbard og Grønland har vist at arten er til stede her året rundt med avansert sangaktivitet døgnet rundt i vintermånedene, noe som tyder på at dette området er et forplantningsområde for denne bestanden (Moore mfl. 2012, Stafford mfl. 2012, 2018, Ahonen mfl. 2017). Opportunistiske observasjoner bekrefter konsentrasjoner av beitende grønlandshval om våren/tidlig sommer langs iskanten i Framstredet rundt 76–77˚N (Joiris mfl. 2014, de Boer mfl. 2019) og nye sporingsstudier viser at dette området er viktig furasjeringshabitat sommer og høst. Disse hvalene overvintrer i den nordlige delen av sitt utbredelsesområde dypt inne i isen nord for iskantsonen (Lydersen mfl. 2012, Kovacs mfl. 2020).
Narhvalen (Monodon monoceros) i Barentsregionen har fått svært lite oppmerksomhet. De få studiene som har vært gjort, viser at deres foretrukne habitat er inne i drivisen i Framstredet og den nordlige Barentsregionen. Andre nar (M1, figur 9.3.1) ble nesten utryddet som følge av hard beskatning under den første kommersielle hvalfangsten som startet i begynnelsen av 1600-tallet på Svalbard. Den lille gjenværende bestanden kombinert med disse hvalenes lange levealder (over 200 år) og langsomme livshistorie, gjør at bestanden er svært truet i dag og klassifiseres som «Kritisk truet–CR» både på den norske rødlisten og av IUCN (Vacquié-Garcia mfl. 2017b, Cooke og Reeves 2018). Akustisk overvåkning i drivisen i Framstredet mellom Svalbard og Grønland har vist at arten er til stede her året rundt med avansert sangaktivitet døgnet rundt i vintermånedene, noe som tyder på at dette området er et forplantningsområde for denne bestanden (Moore mfl. 2012, Stafford mfl. 2012, 2018, Ahonen mfl. 2017). Opportunistiske observasjoner bekrefter konsentrasjoner av beitende grønlandshval om våren/tidlig sommer langs iskanten i Framstredet rundt 76–77˚N (Joiris mfl. 2014, de Boer mfl. 2019) og nye sporingsstudier viser at dette området er viktig furasjeringshabitat sommer og høst. Disse hvalene overvintrer i den nordlige delen av sitt utbredelsesområde dypt inne i isen nord for iskantsonen (Lydersen mfl. 2012, Kovacs mfl. 2020).
Narhvalen i Barentsregionen har fått svært lite oppmerksomhet. De få studiene som har vært gjort viser at deres foretrukne habitat er inne i drivisen i Framstredet og den nordlige Barentsregionen. Andre narhvalbestander tilbringer sommeren i ofte isfrie fjorder og bukter med brefronter og overvintrer i offshore områder med tett isdekke (f.eks. Heide-Jørgensen mfl. 2015). Men både en flytelling (Vacquié-Garcia mfl. 2017) og passiv akustisk overvåkning (Ahonen mfl. 2019) bekrefter at narhvalene i Barentsregionen oppholder seg i drivisen hele året og foretrekker områder med opptil 90 prosent isdekke. Genetiske studier viser liten genetisk diversitet mellom narhval fra ulike områder, men narhval fra Svalbard og Øst-Grønland skiller seg fra andre bestander (Louis mfl. 2020). De har en ekstremt dyp dykkeadferd (Heide-Jørgensen mfl. 2015). Narhval er vurdert til å være en av de arktiske marine pattedyrartene som er mest utsatt for effekter av pågående klimaendringer (Laidre mfl. 2008, Reeves mfl. 2014). Tallrikheten av finnhval basert på tellinger i perioden 2014–2018 er beregnet til 11 387 (CV=0,17, 95 % konfidensintervall: 8 072-16 063) og den viktigste utbredelsen i norske farvann er fra Finnmark til nordvestspissen av Svalbard, men signifikant økende tetthet mot iskanten i Framstredet.
Isbjørn (Ursus marinus) forekommer spredt i området, men er ikke avgjørende for definisjon av området som et SVO.
Sjøfugl
Iskantsonen er generelt svært viktig for ismåke (Pagophila eburnea), særlig vår og høst (se Gilg mfl. 2010). Store deler av den russiske, hele Svalbard- og store deler av den grønlandske bestanden trekker gjennom dette området. I tillegg er området (dvs. iskantsonen/isdekte områder) viktig for den grønlandske hekkebestanden. Også polarlomvi (Uria lomvia), alkekonge (Alle alle) og teist (Cepphus grylle) går inn i isen når de beiter i hekkesesongen, men trekker i stor grad ut av dette området vinterstid. Sjøfuglforekomstene er svært relevant i forhold til definisjonen av dette SVO-et, særlig for ismåke (Joiris mfl. 2014, Gilg mfl. 2010).
9.3.6 - Pågående og framtidige endringer
Barentshavregionen er det området i Arktis hvor isreduksjonen har vært mest dramatisk de senere årene, men samme trend observeres også i Framstredet (von Quillfeldt mfl. 2018). Trenden viser at isen trekker seg nordover over tid, både for sommer- og vintersesongen, men det er store mellomårlige variasjoner. Også istykkelsen i Framstredet har blitt mindre siden overvåkingen begynte tidlig på 1990-tallet (Hansen mfl. 2013, Hansen mfl. 2014, Renner mfl. 2014, Krumpen mfl. 2016), og det er antydning til at snødybden har blitt mindre (for eksempel Webster mfl. 2014). Disse trendene ventes å fortsette, og vil også få innvirkning på isbiota, både artssammensetning, biomasse og produksjon (Fernández-Méndez mfl. 2015, Bluhm mfl. 2017b, Bluhm mfl. 2018, Kauko mfl. 2018, von Quillfeldt mfl. 2018, Hop mfl. 2020), og det er sannsynlig at det vil bli flere smeltedammer og mer isbiota i infiltrasjonslaget (overgangssonen mellom is og snø) (Fernández-Méndez mfl. 2018). Imidlertid mangler det lange tidsserier for isbiota, særlig i iskantsonen, og det er derfor ikke mulig å forutsi omfanget av disse endringene på isbiota.
Thenisto compressa Foto: C.Brohms, HI
9.4 - SVO Vesterisen (NH2)
9.4.1 - Sammendrag
Områdebeskrivelse og strukturer
Det eksisterende SVO Vesterisen (NH2) er et drivisområde som er påvirket av den kalde sørgående Øst-Grønlandstrømmen som frakter is og kaldt vann fra Polhavet. Vesterisen endres mye i utstrekning fra år til år og kan ligge meget langt øst om våren. På 1960- og 1970-tallet kunne den strekke seg til og øst for Jan Mayen, men de siste ti-årene har iskanten derimot vært mye lenger vest, og det har vært en nedadgående trend i utbredelse av sjøis de siste tiårene.
Miljøverdier
Området er et kjerneområde for yngling for klappmyss som er sterkt truet (EN). Også grønlandssel har dette som viktig beite- og yngleområde. G rønlandssel og klappmyss er begge endemiske i Nord-Atlanteren. Problemene til klappmyssen kan ha sammenheng med reduksjonen i isdekket. Mindre drivis kombinert med færre og tynnere drivisflak forringer kvaliteten på yngleområdene. Flere arter (f.eks. ismåke og polarlomvi) trekker gjennom Vesterisen på vei fra Svalbard til sine overvintringsområder. Ismåke og flere av sjøfuglene på Jan Mayen er rødlistet. Iskantsonen er lite påvirket av menneskelig aktivitet til tross for et mindre unntak av fangstvirksomhet.
Grønlandssel med unge. Foto: Kjell Arne Fagerheim, HI
Figur 9.4.1. Kart over SVO Vesterisen (NH2). Grensene for SVO-et er vist med tykk rød strek, mens grenser for overlappende og nærliggende foreslåtte SVO-er er vist med tynnere rød strek. Grensene for forvaltningsplanområdet er markert med brun linje. Kartet viser utbredelsen av utvalgte miljøverdier av betydning for SVO-et, dvs. kasteområde for klappmyss (M1-2) og kaste- og hårfellingsområde for grønlandssel (M-2). Hårfellingsområde for klappmyss (M-1-1) litt lenger nord er også vist.
9.4.2 - Vurdering i forhold til EBSA-kriteriene – SVO Vesterisen (NH2)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i SVO Vesterisen (NH2)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet/sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Sjøpattedyr
Eneste ungeproduksjonsområde for grønlandssel og klappmyss i sentrale Nord-Atlanteren (Haug og Biuw 2019).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Sjøfugl
Sjøpattedyr
Klappmyss og grønlandssel føder ungene på isen som dannes her i mars–april (Haug og Biuw 2019).
Området er antatt å ha betydning som beiteområde for sjøfugl som hekker på Jan Mayen, og en del arter trekker gjennom Vesterisen (Gilg mfl. 2010). Hvordan hekkefuglene på Jan Mayen bruker området er ukjent.
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Sjøpattedyr
Området er viktig for klappmyssbestanden og grønlandssel som yngler her. I rødlista for 2010 endret klappmyss status til sterkt truet (EN) fordi bestanden antas å ha blitt redusert med mer enn 50 prosent på mindre enn tre generasjoner (Haug og Biuw 2019).
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Sjøpattedyr
Sjøpattedyrhabitatet trues av klimaendringer. Reproduksjonen er nedadgående (Haug og Biuw 2019).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Sjøpattedyr
Grønlandssel og klappmyss er avhengige av isdannelsen her for den årlige ungeproduksjonen (Haug og Biuw 2019). Tap av havis i området vil derfor føre til tap av viktig biologisk mangfold i området.
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
Sjøpattedyr
Iskantsonen er lite påvirket av menneskelig aktivitet, til tross for et mindre unntak av fangstvirksomhet (Haug og Biuw 2019).
9.4.3 - Lokalisering
SVO Vesterisen er tidligere definert i Meld. St. 20 (2019-2020). Området omfatter havisen nord- og vestover fra Jan Mayen og er en del av et større område med havis i Framstredet som strekker seg helt til Grønland og som også SVO Havis Framstredet er en del av (figur 9.4.1).
Vesterisen som særlig verdifullt og sårbart område er avgrenset i vest av grensen for forvaltningsplanen for Norskehavet, og overlapper så vidt med foreslått SVO Jan Mayen. Området overlapper også så vidt med en liten del av foreslått nytt SVO Midtatlantisk rygg.
Det er ingen verneområder i området.
9.4.4 - Introduksjon til området
Drivisbeltet som dannes utenfor Grønlands østkyst blir kalt «Vesterisen». Området er hovedsakelig påvirket av den kalde sørgående Øst-Grønlandsstrømmen som transporterer is og kalde vannmasser sørover fra Polhavet langs med Grønlands østkyst (figur 9.4.2, Helland-Hansen og Nansen 1909). Selv om noe av sjøisen i Grønlandshavet blir dannet lokalt, kommer mesteparten av sjøisen fra Polhavet med Øst-Grønlandsstrømmen. Vesterisen endres mye i utstrekning fra år til år og kan ligge meget langt øst om våren (Gabrielsen mfl. 1997). De mellomårlige variasjoner i isutbredelse påvirkes i stor grad av endringer i det atmosfæriske trykkfeltet (Deser mfl. 2000). Det var særlig stor utbredelse av Vesterisen i siste halvdel av 1800-tallet og det er blitt rapportert at isen til tider har ligget midtveis mellom Jan Mayen og Norge, noe som var tilfellet i 1858 og 1881 (Iversen 1936). Sjøisen som ble ført med Jan Mayen-strømmen på 1960- og 1970-tallet kunne strekke seg østover fra Øst-Grønlandskysten til nord for Jan Mayen og ble kalt ”Isodden”, en tunge med sjøis (Loeng og Vinje 1979, Vinje 2001). De siste ti-årene har iskanten derimot vært mye lengre vest og det vært en nedadgående trend i utbredelse av sjøis de siste tiårene (Onarheim mfl. 2018). Sammenlignet med årene 1870–1920 har iskanten i 1989–2002 flyttet seg 250–375 km vestover (Divine og Dick 2006). I gjennomsnitt er det maksimum isdekke i april og minimum isdekke i september (Kvingedal 2005). Området er viktig for både eksisterende og foreslått endret SVO Jan Mayen ved at Jan Mayen-strømmen transporter drivis og arktiske vannmasser til området.
I SVO Vesterisen ligger dagens pH-verdier i overkant av 8,15 i overflatevannet. Metningsgraden av aragonitt er rundt 1,7 (Fransner mfl. 2020, Lauvset mfl. 2016). Dette er lavere enn i SVO-ene langs Norskekysten, og dette skyldes først og fremst temperatur: kalkmineraler blir mer løselig i kaldere vann. Metningshorisonten ligger rundt 2000 m. Havforsuringen har redusert pH-en i overflaten med 0,09–0,1 enheter. Dette er noe mindre enn langs Norskekysten som har varmere vann og dermed tar opp mer menneskeskapt CO2 (Maier-Reimer og Hasselmann 1987).
Figur 9.4.2. Kart over gjennomsnittlig overflatestrøm (piler) og bunndyp (fargekodet) i SVO-ene Vesterisen og Jan Mayen og omkringliggende områder.
9.4.5 - Beskrivelse av miljøverdiene
Isbiota
SVO Vesterisen er en svært liten del av et større område med havis i Framstredet, og isbiotaen i den delen som utgjør SVO-et, skiller seg ikke særlig fra tilsvarende områder i Framstredet (Se SVO Havis Framstredet, Kap 9.3.5). Tatt områdets størrelse i betraktning og det at området ofte kan ha lite eller ingen is – og dersom det forekommer is, er det ofte færre og tynnere drivisflak enn tidligere, er imidlertid ikke isbiota avgjørende for definisjon av området som SVO.
Plankton
Det finnes lite informasjon når det gjelder produksjon og biologisk mangfold av plankton i SVO Vesterisen. Den årlige primærproduksjonen i dette området (fra 72°N) samt for områder lenger nordover, er estimert basert på satellittdata (Børsheim 2017). Produksjonen i Øst-Grønlandsstrømmen ble funnet å være lav og økte østover inn i Grønlandshavet. Primærproduksjonen har økt siden 2003, med lavest økning i Øst-Grønlandsstrømmen og høyere i Grønlandshavet (Børsheim 2017). Studie basert på håvtrekk under isen i/nær SVO-et og andre deler av Øst-Grønlandsstrømmen har observert at kopepoder generelt og særlig Calanus spp. utgjorde hoveddelen av biomassen i Øst-Grønlandsstrømmen. I tillegg utgjorde Hymenodora (pelagisk reke), Parathemisto (amfipode), Euchaeta (kopepod) og Eukrohnia (pilorm) mye av biomassen (Hopkins 1969). Samme undersøkelse viste også at hoveddelen av dyreplanktonet var konsentrert i de øvre 500 meterne, også om vinteren. Det er også rapportert at Oithona similis, Calanus spp., Metridia longa, samt nauplii av kopepoder dominerte dyreplanktonet, som ellers bestod av chaetognatha, amphipoda, appendikularia, euphausiids og cnidaria (Alcaraz mfl. 2010). SVO-et ligger utenfor de områdene hvor det regelmessig tas prøver ifm. Havforskningsinstituttets plante- og dyreplanktonovervåkning i Norskehavet. Plankton er ikke vesentlig for definisjonen av dette SVO-et.
Fisk
Selv om det er polartorsk (Boreogadus saida) og andre arktiske arter som har beiteutbredelse i dette området (Joiris mfl. 2014), er det i seg selv ikke regnet som tilstrekkelig relevant til at fisk kan brukes som begrunnelse for dette SVO-et.
Bunnsamfunn
Bunndyrene i dette området som er assosiert med Jan Mayen bruddsone er diskutert spesifikt under forslag til SVO NH4 Midtatlantisk rygg under.
Det øvrige området er dype, bløte kaldtvannssletter. Utenom spredte kartleggingslokaliteter er det ingen spesifikke studier av disse dyphavsslettenes bunndyrsfauna, men slike bunnforhold har ofte bunndyrsamfunn som omfatter slangestjerner, flerbørstemark, snegler og hyperbentiske krepsdyr (Mareano.no). I et område med tidvis isdekke vil mye av føden bestå av nedfalls-isbiota, men i hvilken grad avhenger av dyp. Flere av disse dyrene er føde for høyere trofiske nivåer, og de frigjør slik den energien som har falt ned til bunnen tilbake i vannsøylen. Flere av artsgruppene er representert med arter som har smale temperaturnisjer, og som er knyttet til kalde vannmasser. Bunndyrsamfunnene er ikke vesentlige i forhold til definisjonen av dette SVO-et.
Sjøpattedyr
Vesterisen er et kjerneområde for kasting for klappmyss (Cystophora cristata) (sterkt truet-EN, 2010) og grønlandssel (se figur 9.4.1). Grov ‘gammel-is’ og årsis var et viktig yngleområde for grønlandssel og klappmyss som begge er endemisk i Nord-Atlanteren. Den grove ‘gammel-isen’ var særlig viktig for klappmyss. På grunn av klimaendringer dannes ikke Vesteris-odden seg på samme måte som tidligere og mengden grov ‘gammel-is’ har blitt mindre. Ynglingen for begge artene foregår nå lenger vest og i drivisstrømmen sørover langs Øst-Grønland. Etter en lang periode med bestandsøkning hos grønlandssel, fra 1870-tallet til tidlig på 2000-tallet , kan dette ha snudd da et svært lavt estimat av ungeproduksjonen basert på tellinger i 2018 gir en estimert totalbestand i 2019 på 426 808 dyr (95 % konfidensintervall: 313 004–540 613), noe som er betydelig lavere enn tidligere estimater på 2000-tallet (Haug og Biuw 2019). For klappmyss er bestandsanslaget på 76 623 dyr (95 % konfidensintervall: 58 299–94 947) i 2019. Alt tyder på at klappmyssbestanden i Vesterisen har avtatt betydelig i størrelse i perioden fra slutten av 1940-tallet og fram til rundt 1980. Muligens har bestanden fortsatt å avta noe også etter 1980, og dagens nivå er antakelig ikke mer enn knapt 7 prosent av nivået for rundt 70 år siden (Haug og Biuw 2019). De siste årenes nedgang er trolig ikke fangstrelatert. Den har trolig større kopling til mindre is og is av dårligere kvalitet.
Sjøfugl
Vesterisen er ikke kartlagt med hensyn på sjøfugl. Sporing av ismåke (Pagophila eburnea) ved bruk av satellittsendere viser at fugler fra Svalbard (og Grønland og Canada) trekker gjennom dette området på vei til overvintringsområdet mellom Grønland og Canada (Labradorhavet og Davis Strait; Gilg mfl. 2010). Ismåke beiter på polartorsk, isamfipoder, åtsler og rester etter sel tatt av isbjørn eller mennesker, og har sterk tilknytning til isen. Polarlomvi (Uria lomvia) fra Svalbard trekker også gjennom dette området. Hekkende sjøfugl (blant annet lomvi (Uria aalge), polarlomvi, alkekonge (Alle alle), teist (Cepphus grylle), havhest (Fulmarus glacialis) og krykkje (Rissa tridactyla)) fra Jan Mayen beiter i SVO Vesterisen.
9.5 - Forslag til endret SVO Jan Mayen (NH3)
9.5.1 - Sammendrag
Områdebeskrivelse og strukturer
Forslag til endret SVO Jan Mayen (NH3) har spesielle topografiske forhold, fordi den ligger oppe på den midtatlantiske ryggen og som en isolert øy i et stort hav. To vannmasser, den kalde fra Øst-Grønlandsstrømmen og den varme fra atlanterhavsstrømmen møtes ved Jan Mayen. Dette SVO-et er foreslått utvidet med Kandidatområdet.
Miljøverdier
Det er særlig sjøfugl som gjør dette området verdifullt. Jan Mayen er i norsk sammenheng et enestående viktig hekkeområde for sjøfugl, med 15 arter som hekker i 22 sjøfuglkolonier med mer enn 300 000 hekkende par sjøfugl. De mest tallrike artene er havhest, alkekonge og polarlomvi, men også alke, teist, lomvi, lunde, polarmåke og krykkje hekker her, i tillegg til mer sørlige arter som sildemåke og gråmåke. Også tyvjo og storjo er relativt tallrike. De pelagiske artene som dominerer på øya, beiter grovt sett ut til 100 km fra koloniene. Flere av sjøfuglartene på Jan Mayen er rødlistet, men koloniene på Jan Mayen virker relativt robuste.
Næringsrike vannmasser stimulerer høy og stabil produksjon i området. Området inneholder dyreplankton knyttet til ulike habitater og vil dermed kunne ha stor artsdiversitet. Høy biomasse av dyreplankton er blitt påvist for området. Krysningen mellom ulike havstrømmer på den midtatlantiske rygg legger til rette for en rekke fiskearter. Det er registrert 71 fiskearter i området hvorav ålebrosmearten Lycenchelys platyrhina ikke er registrert i andre norske havområder. De grunne kystnære bunnsamfunnene nær Jan Mayen er i et 4 km bredt belte preget av vulkanutbruddet i 1970, der submarine områder ned til 30 m ble dekket av lava.
Den avsidesliggende posisjonen gjør at foreslått endret SVO Jan Mayen i svært liten grad er påvirket av mennesker og derfor anses som et område med høy grad av naturlighet.
Jan Mayen. Foto: Harald Næss, HI
Figur 9.5.1. Kart over forslag til endret SVO Jan Mayen (NH3). Den foreslåtte grensen for SVO-et er vist med tykk rød strek, mens grenser for overlappende og nærliggende eksisterende og foreslåtte SVO-er er vist med tynn rød strek. Grensene for forvaltningsplanområdet er markert med brun linje. Kartet viser utbredelsen av utvalgte miljøverdier av betydning for området, dvs. kasteområde for klappmyss (M1-2), kaste- og hårfellingsområde for grønlandssel (M-2) og beiteområde for pelagisk beitende sjøfugl (SF1). Hårfellingsområde for klappmyss (M-1-1) litt lenger nord er også vist.
9.5.2 - Vurdering i forhold til EBSA-kriteriene – Forslag til endret SVO Jan Mayen (NH3)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til endret SVO Jan Mayen (NH3)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet/sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Planteplankton
Dyreplankton Sjøfugl Fisk Bunnsamfunn
Området ser ut til å ha høyere primærproduksjon enn omkringliggende havområder (Pálsson mfl. 2012). Topografiske forhold, samt frontsystemer, muliggjør tilgang på næringsrikt vann som stimulerer høy og mer stabil produksjon i området.
Høy biomasse av dyreplankton er påvist for områder ved Jan Mayen, og krill og amfipoder er funnet i betydelige mengder (Dalpadado mfl. 1998, Melle mfl. 2004, ICES 2020, 2021). Dyreplanktonarter som lever i ulike habitater er vanlige i området (Broms mfl. 2009, Fossum mfl. 2012).
Ålebrosmearten, Lycenchelys platyrhina, er registrert som endemisk for området (Fossum mfl. 2012, Wienerroither mfl. 2011). Eneste sted i norske havområder hvor det er mulig å følge retablering av grunne kystnære bunnsamfunn etter et vulkanutbrudd (Gulliksen mfl. 2003).
De grunne kystnære bunnsamfunnene nær Jan Mayen er i et 4 km bredt belte preget av vulkanutbruddet i 1970, der submarine områder ned til 30 m ble dekket av lava. Suksesjonsstudier av dette området (Gulliksen mfl. 2004) viser at det tar lengre tid enn i sammenlignbare, men mer sørlige områder, å tilbake-etablere faunaen fra de uberørte områdene.
Isolert-liggende Jan Mayen er et viktig hekkeområde for sjøfugl (seapop.no).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Sjøfugl
Beiteområder 100 km ut fra Jan Mayen er viktig for sjøfugl som hekker på øya (www.seapop.no). Dette gjelder særlig polarlomvi, lomvi, alkekonge, havhest og krykkje, der særlig de to siste kan bruke enda større områder ut fra kolonien (f.eks. Christensen-Dalsgaard mfl. 2018).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Sjøfugl
Flere av sjøfuglartene på Jan Mayen er i nedgang totalt sett eller rødlistet, men koloniene på Jan Mayen virker relativt robuste (seapop.no). Kolonien kan således virke som et refugium for arter som ellers går tilbake. Rødlistede arter på Jan Mayen omfatter alke (sterkt truet-EN), lomvi (sårbar–VU), polarlomvi (nær truet–NT), sabinemåke og ismåke (VU), krykkje (NT), havhest (NT) og praktærfugl (NT). Av de pelagiske artene er det kun alkekonge som ikke er på rødlisten (Henriksen og Hilmo 2015).
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Sjøfugl
Flere av sjøfuglartene på Jan Mayen er i nedgang totalt sett eller rødlistet, men koloniene på Jan Mayen virker relativt robuste (seapop.no). Betydningen av dette området er viktig med flere livskraftige bestander av pelagiske sjøfugl som ellers går tilbake, selv om både lomvi og polarlomvi har hatt en negativ utvikling siste 10 år på øya (Anker-Nilssen mfl. 2020).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Planteplankton
Dyreplankton
Det foreligger ingen systematisk informasjon angående planteplankton, men ut fra strøm og topografi er det indikasjoner på at området etter isløsning har høyere primærproduksjon enn omkringliggende havområder (Fossum mfl. 2012, Pálsson mfl. 2012).
Området har høy biomasse av dyreplankton (ICES 2020, 2021) og det er funnet høye konsentrasjoner av krill og amfipoder (Fossum mfl. 2012).
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Planteplankton
Fisk
Dyreplankton Sjøfugl Bunnsamfunn
Det foreligger ingen systematisk informasjon angående planteplankton, men tidsavgrensede prosjekter tyder på at området har høyere primærproduksjon enn omkringliggende havområder (Pálsson mfl. 2012).
Flere ulike vannmasser og habitater som inneholder ulike dyreplanktonarter møtes i området (Fossum mfl. 2012).
71 fiskearter er registrert i området hvorav én (Lycenchelys platyrhina) ikke er registrert i andre norske havområder (Wienerroither mfl. 2011).
En studie av mange havbørstemark (Bakken mfl. 2010) mer enn doblet antallet kjente havbørstemark-arter fra Jan Mayen-området (nå 121 navngitte arter registrert innenfor studieområdet). Eneste sted i norske havområder hvor det er mulig å følge retablering av grunne kystnære bunnsamfunn etter et vulkanutbrudd (Gulliksen mfl. 2003).
Det hekker relativt mange sjøfuglarter på Jan Mayen ift. de andre arktiske øyene, med både arktiske og mer sørlige arter (Systad mfl. 2019).
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
Sjøfugl
Bunnsamfunn
De grunne kystnære bunnsamfunnene nær Jan Mayen er i et 4 km bredt belte preget av vulkanutbruddet i 1970, der submarine områder ned til 30 m ble dekket av lava. Suksesjonsstudier av dette området (Gulliksen mfl. 2003) viser at det tar lengre tid enn i sammenlignbare, men mer sørlige områder, å tilbake-etablere faunaen fra de uberørte områdene.
Jan Mayen er et viktig hekkeområde for sjøfugl, med 15 arter som hekker i 22 sjøfuglkolonier med totalt 300 000 hekkende par sjøfugl. Øya er uendret som viktig hekkeområde for sjøfugl (seapop.no).
9.5.3 - Lokalisering
Forslag til endret SVO Jan Mayen, dekker det eksisterende SVO Jan Mayen, sist oppdatert i Meld. St. 20 (2019-2020), men grensene for SVO-et er utvidet for å inkludere kandidatområdene hvor beitearealene for pelagisk beitende sjøfugl strekker seg 100 km ut fra de viktigste hekkekoloniene på Jan Mayen (figur 9.5.1).
Forslag til endret SVO Jan Mayen ligger i skjæringspunktet mellom Norskehavet, Grønlandshavet og Islandshavet. Området overlapper så vidt med SVO Vesterisen og med deler av det foreslått nye SVO Midtatlantisk rygg. Området overlapper også delvis med eksisterende SVO Den arktiske front som fagekspertgruppen foreslår blir fjernet.
Forslag til endret SVO Jan Mayen dekker også den marine delen av Jan Mayen naturreservat.
9.5.4 - Introduksjon til området
Jan Mayen ligger i et område hvor nordgående varmt atlanterhavsvann møter kalde arktiske vannmasser fra en sidegren av den sørgående Øst-Grønlandsstrømmen. Frontsystemet som dannes gir grunnlag for store forekomster av dyreplankton, fisk, sjøfugler og sjøpattedyr. Det er tidligere skrevet en rapport om miljø og levende ressurser ved Jan Mayen for Olje- og energidepartementet (Fossum mfl. 2012).
Jan Mayen danner et felles grensepunkt mellom Norskehavet, Grønlandshavet og Islandshavet, og sirkulasjonen og fordelingen av vannmassene er i stor grad bestemt av bunntopografien (Helland-Hansen og Nansen 1909, Voet mfl. 2010). Om det er de kalde arktiske eller varmere atlantiske vannmassene som dominerer ved Jan Mayen, vil variere både i tid og rom (se figur 9.4.2. i Kap. 9.4. SVO Vesterisen). Denne dynamikken gjenspeiles også til en viss grad i det biologiske mangfoldet der ulike arktiske og nordlige arter befinner seg i området avhengig av hvilke vannmasser som dominerer (Fossum mfl. 2012). Siden den arktiske fronten som regel ligger litt øst for Jan Mayen, er området rundt Jan Mayen dominert av arktiske vannmasser (Gabrielsen mfl. 1907). Om vinteren kan temperaturene i luft og sjø bli så lave at hele området fryser. Selv om noe av sjøisen blir dannet lokalt, kommer mesteparten av sjøisen fra Polhavet med Øst-Grønlandsstrømmen. I gjennomsnitt er det maksimum isdekke i april og minimum isdekke i september (Kvingedal 2005). I tidligere år, som på 1960- og 1970-tallet, kunne sjøis som ble ført med Jan Mayen-strømmen strekke seg østover fra Øst-Grønlandskysten til nord og øst for Jan Mayen (Vinje 2001). Denne tungen med sjøis nord for Jan Mayen ble kalt ”Isodden”, en tunge med sjøis (Nansen 1906). De siste tiårene har det derimot vært en nedadgående trend i utbredelse av sjøis i Grønlandshavet og Isodden har ikke dannet seg på mange år (Onarheim mfl. 2018).
I de arktiske vannmassene er pH-en litt høyere enn i de atlantiske, i overkant av 8,15 vs i overkant av 8,1. Dette er i stor grad drevet av temperatur, siden lavere temperaturer gir høyere pH (Fransner mfl. 2020). Havforsuringen har redusert pH-verdiene i området med 0,1 enhet, og denne nedgangen er like sterk i både arktisk- og atlanterhavsvann (Fransner mfl. 2020, Lauvset mfl. 2016). Vannet er mettet med kalsitt over hele vannsøylen, mens for aragonitt ligger metningshorisonten ved rundt 2000 m. Pga. havforsuringen er dette et par hundre meter grunnere enn i førindustriell tid (Fransner mfl. 2020).
9.5.5 - Beskrivelse av miljøverdiene
Plankton
Det foreligger ingen systematisk informasjon angående planteplankton, men tidsavgrensede prosjekter tyder på at området har høyere primærproduksjon enn omkringliggende havområder (Pálsson mfl. 2012) .
Høy biomasse av dyreplankton er blitt påvist for områder ved Jan Mayen, og overvåkningstidsserien i mai viser verdier opp til 20 g per m 2 (Melle mfl. 2004, ICES 2020 a, 2021). Særlig i perioden 1995 – 2015 ble det registrert høye verdier. En enkeltstudie understøttet overvåkingsserien og viste at planktonbiomassen varierte mellom 12 – 22 g (tørrvekt per m 2 ; Fossum mfl. 2012).
I områdene omkring Jan Mayen forekommer tre Calanus-arter (C. finmarchicus , C. hyperboreus og C. glacialis) der C. finmarchicus dominerer. I tillegg finnes en rekke andre k opepodearter og slekter, der mengden vil variere sterkt gjennom sesongen (Broms mfl. 2009 , Fossum mfl. 2012). Krill har blitt registrert i betydelige mengder omkring Jan Mayen, og artene Meganyctiphanes norvegica , Thysanoessa inermis og T. longicaudata finnes i området (Dalpadado mfl. 1998). Biomassen av amfipoder vurdert i Norskehavet, Grønlandshavet og Islandshavet, har blitt funnet å være høyest i området vest og nord for Jan Mayen (Melle mfl. 2004). Amfipoden Themisto libellula, den største arten i Themisto - slekten og en typisk arktisk art, har blitt funnet i høye konsentrasjoner ved Jan Mayen (Dalpadado mfl. 1998 , Olsen mfl. 2006), mens T. abyssorum også har blitt funnet å dominere i området (Fossum mfl. 2012). Vingesneglen Limacina helicina og pilorm Eukrohnia spp . har blitt registrert å være svær t vanlige i området (Fossum mfl. 2012). Ingen av de oven n evnte artene er særegne for Jan Mayen-området, og finnes utstrakt i arktiske og atlantiske vannmasser, men siden disse to vannmassene møtes ved Jan Mayen, vil området inneholde dyreplankton knyttet til ulike habitater og dermed kunne ha stor artsdiversitet. Videre er det indikasjoner på at mengden dyreplankton i området om høsten er sammenlignbare med mengdene ved Norskekysten og østlige Norskehavet. Imidlertid blir ikke dyreplanktonet ved Jan Mayen overvåket like systematisk som andre områder i Norskehavet, noe som begrenser muligheten til å vurdere mengder og produksjon, samt sesongutvikling og mellomårlige variasjoner (Fossum mfl. 2012).
Fisk
Posisjonen av foreslått endret SVO Jan Mayen i krysningen mellom ulike havstrømmer på den midtatlantiske rygg legger til rette for et mangfold av fiskearter. Wienerroither mfl. (2011) har publisert en mangfoldig artsliste med alle fiskearter som er registrert rundt Jan Mayen. Dataene kom fra vitenskapelige samlinger, litteratur og Havforskningsinstituttets databaser. Mellom 1877 og 2009 ble 71 fiskearter registrert i området, hvorav én (Lycenchelys platyrhina) ikke var registrert i andre norske havområder.
De første opplysningene om torsk (Gadus morhua) ved Jan Mayen stammer fra 1929 og kommer fra ansatte i Statens havnevesen som skulle undersøke muligheten for å bygge havn på Jan Mayen. Tidlig på 1930-tallet ble det gjennomført fiskeriundersøkelser for å finne ut hvor mye torsk det var ved Jan Mayen. Fangstene varierte sterkt, og det ble aldri satt i gang noe omfattende fiske i dette området. Seinere har det også dukket opp torsk i forbindelse med rekeundersøkelser ved Jan Mayen, og det har kommet rapporter om innblanding av torsk under rekefiske på 1980-tallet, samt under linefiske i 2018 og 2019 (https://www.hi.no/hi/nyheter/2019/august/forskar-vidare-pa-jan-mayen-torsken). På ”grunt” vann nær selve Jan Mayen er det en egen torskebestand (Fossum mfl. 2011). Genetiske undersøkelser viser at denne torskebestanden ikke er i slekt med barentshavstorsken, men er mest sannsynlig rekruttert fra Island.
Det er flere loddebestander på den nordlige halvkulen (Rose, 2005). Loddebestanden ved Island – Øst-Grønland og Jan Mayen er en av disse, og denne har beiteområdene sine mellom Nord-Island, Grønland og Jan Mayen. Om sommeren har lodda (Mallotus villosus) tradisjonelt trukket nordover mot Jan Mayen for å beite i de planktonrike områdene langs polarfronten, men det skjedde en drastisk endring i vandringsmønster ved starten av 2000-tallet, trolig i sammenheng med temperaturøkning (Carscadden mfl. 2013, Jansen mfl. 2021). Beitevandringen i senere år har i større grad gått til områder lenger vest, tettere inn mot østkysten av Grønland.
Makrell (Scomber scombrus) står utbredt i sentrale og nordlige deler av Norskehavet inkludert sør–sørøst av Jan Mayen på ettersommeren. Makrellen som mellom 2012 og 2019 ble fanget ved Jan Mayen, er stor og feit, med mager fulle av større amfipoder. Makrellen er litt varmekjær og foretrekker temperaturer over 6 °C, men kan også vandre inn i kalde vannmasser (Nøttestad mfl. 2020).
Bunnsamfunn
Mye av bunnsamfunnene i dette området er beskrevet i foreslått nytt SVO Midtatlantisk rygg, der området Jan Mayen Bruddsone er diskutert (Kap. 9.6.5). Ut fra dagens kunnskap om bunnsamfunn på grunnere vann ved Jan Mayen, er det ikke funnet at disse har særlig miljøverdi (Mareano.no), men områdene er ikke særlig grundig undersøkt.
De grunne kystnære bunnsamfunnene nær Jan Mayen er i et 4 km bredt belte preget av vulkanutbruddet i 1970, der submarine områder ned til 30 m ble dekket av lava. Suksesjonsstudier av dette området (Gulliksen mfl. 2004) viser at det tar lengre tid enn i sammenlignbare, men mer sørlige områder, å tilbake-etablere faunaen fra de uberørte områdene. Generelt er den sublittorale (ned til 30 m) faunaen en “fattigere” utgave av sublittorale soner i Norge og på Svalbard, med lite fastsittende fauna ned til 15 m, noe som er vanlig i områder med isskuring av strender.
Dypere bløtbunnsområder har hatt lite fokuserte faunistiske studier, med unntak av en studie av mangebørstemark (Bakken mfl. 2010) som mer enn doblet antallet kjente polychaet-arter fra Jan Mayen-området (nå 121 navngitte arter registrert innenfor studieområdet).
På ”grunt” vann nær selve Jan Mayen er det en egen rekebestand (Pandalus borealis) (Fossum mfl. 2012) som utgjør en egen genetisk populasjon (Jorde mfl. 2015). Populasjonen karakteriseres av langsom vekst, kjønnsmodning og kjønnsskifte ved høy alder og lang levetid (Nilssen og Aschan 2009). Rekene er store sammenlignet med for eksempel rekene i Barentshavet.
Sjøpattedyr
Området rundt Jan Mayen ser ikke ut til å være av spesiell betydning for hvalartene. Et mulig unntak er nebbhval (Hyperoodon ampullatus) som ser ut til å være knyttet til den midtatlantiske ryggen fra Jan Mayen til Spitsbergen, men dette blir omtalt under foreslått nytt SVO Midtatlantisk rygg.
For 20–30 år siden var den omtalte “Isodden” nordøstover fra Jan Mayen kjerneområdet for ungefødsler for grønlandssel og klappmyss. De siste tiårene har det imidlertid vært en nedadgående trend i utbredelse av sjøis i Grønlandshavet og Isodden har ikke dannet seg på mange år (Onarheim mfl. 2018).
Sjøfugl
Jan Mayen er et viktig hekkeområde for sjøfugl, med 15 arter som hekker i 22 sjøfuglkolonier med totalt 300 000 hekkende par sjøfugl. Øya er uendret som viktig hekkeområde for sjøfugl. Området ble kartlagt med tanke på overvåking i regi av Seapop-programmet (https://seapop.no/) i 2010 og 2011. Overvåking startet for fullt i 2012 og ble noe utvidet i 2013. Estimert antall hekkende par i 2013 av havhest (Fulmarus glacialis) var 180 000, av alkekonge (Alle alle) 90 000 og av polarlomvi (Uria lomvia) 110 000 (Fauchald mfl. 2015). Undersøkelsene i 2010 viste ellers at alke (Alca torda), teist (Cepphus grylle), lomvi (Uria aalge) og lunde (Fratercula arctica) hekker på Jan Mayen, men er relativt fåtallige sammenlignet med alkekonge, polarlomvi og havhest. Lomvi og polarlomvi har gått tilbake i hele undersøkelsesperioden, mens arter som havhest og polarmåke (Larus hyperboreus) har vært stabile og storjo (Stercorarius skua) øker (Anker-Nilssen mfl. 2020). I tillegg hekker følgende arter på Jan Mayen: polarmåke, svartbak (Larus marinus), sildemåke (Larus fuscus), gråmåke (Larus argentatus), krykkje (Rissa tridactyla) og sabinemåke (Xema sabini), samt tyvjo (Stercocarius parasiticus) som sammen med storjo er relativt tallrike. De pelagiske artene som dominerer på øya beiter grovt sett ut til 100 km fra koloniene, noe som er grunnlaget for utvidelsen av dette området i forhold til opprinnelig SVO (Systad mfl. 2019).
Klappmyss. Foto: Michael Poltermann, HI
9.6 - Forslag til nytt SVO Midtatlantisk rygg (NH4)
9.6.1 - Sammendrag
Områdebeskrivelse og strukturer
Forslag til nytt SVO Midtatlantisk rygg (NH4) består av flere ulike rygger og bruddsoner som strekker seg over et langt område. De store havstrømmene styres av ryggen, og den arktiske fronten har en relativt fast lokalisering over ryggen. Den komplekse topografien skaper økte vertikale strømninger langs ryggen, som også består av spredte passive og aktive varme kilder. Dette er en ultra-saktespredende spredningsrygg, og derfor blant de eldste kjente varme havkildebunnøkosystemene i verden med ekstremt lav restitueringsevne. Helt i nord munner området ut i Molloydypet, det dypeste bassenget i havområdet.
Miljøverdier
Området har mye endemisk bunnfauna som er unik og spesialisert til å tåle høye temperaturer. Mange av artene er kjemosyntetiske eller lever i symbiose med kjemosyntetiske mikroorganismer, som betyr at de ikke er avhengige av en næringskjede som starter med sollys og primærproduksjon. Området er særlig produktivt med bunnsamfunn bygget av svamp og korall og undervannsfjell som kan fungere som gyte- og oppvekstområder for populasjoner av langsomtvoksende fisk, og som står på OSPARs liste over truede og minkende habitat. I tillegg har området hardbunnskorallskog som er nær truet (NT) ifølge den norske rødlisten for naturtyper. Dette er typisk saktevoksende, skjøre habitater, med en helt unik artssammensetning. Filtrerende arter, særlig svamp, kan være sensitive for forhøyede mengder suspenderte partikler i vannet. Flere arter er avhengig av spesielle bunnsubstrat, og restitusjonsevne av et ødelagt habitat er meget lav. Dette forslaget til nytt SVO har også flere andre sårbare naturtyper.
Enkeltstudier har ikke vist høyere produksjon av planteplankton i det eksisterende SVO Den arktiske front, men høye konsentrasjoner av plante- og dyreplankton i nærheten av fronten er observert og forklart med mesoskala fysiske prosesser. Det er gjort for få studier av dette SVO-et og selve fronten til å si noe sikkert angående produktiviteten av plankton, men det er så langt ikke dokumentert generell forhøyet produksjon i frontområdet sammenlignet med områdene rundt. Høy produktivitet var et viktig argument for SVO Den arktiske front, men siden dette viser seg å ikke være godt nok undersøkt, blir SVO Den arktiske front ikke lenger anbefalt som et selvstendig SVO. Fronten kan nok likevel ha en betydning som habitatsgrense for ulike arter, samt fungere som et aggregeringsområde på ulike trofiske nivåer grunnet horisontalt strømmønster og det faktum at det er en artsdiversitet av blant annet byttedyr på tvers av fronten. Det er indikasjoner på at den midtatlantiske ryggen kan være et viktig sommerbeiteområde for nebbhval.
Figur 9.6.1. Kart over forslag til nytt SVO Midtatlantisk rygg (NH4). De foreslåtte grensene for SVO-et er vist med tykk rød strek, mens grenser for overlappende og nærliggende eksisterende og foreslåtte SVO-er er vist med tynn rød eller oransje strek. Grensene for forvaltningsplanområdet er markert med brun linje. De ulike delene av det nye forslaget til SVO er vist med ulike fargefelt.
9.6.2 - Vurdering i forhold til EBSA-kriteriene – Forslag til nytt SVO Midtatlantisk rygg (NH4)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til nytt SVO Midtatlantisk rygg (NH4)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet/sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Bunnsamfunn
Spredningsryggen og Jan Mayen-bruddsonen omfatter flere aktive varme kilder som Lokeslottet, Ægirs Kilde og Fåvne, samt utdødde kilder som Copper Hill og Mohns Treasure. Både de aktive og de utdødde varme kildene har mye endemisk fauna og mikroorganismer (Kongsrud og Rapp 2012, Kongsrud mfl. 2017, Ramirez-Llodra mfl. 2020, Steen mfl. 2016, Tandberg mfl. 2012, 2013 og 2018), både artene og sammensetningen av de høyere taksa er unik sammenlignet med andre varme kilder. Flere av artene er vurdert som kvalifisert til å bli rødlistet (Rødlista 2021, under forberedelse).
Den ultrasakte spredningen av Knipovichryggen gjør at vi har i verdenssammenheng svært gamle varme kilder (Lokeslottet er beregnet til ca. 10 000 år). Økosystemene her er derfor mye eldre enn varme kilde-systemer i resten av verden.
Gasshydratsystemet på Vestnesaryggen har en unik kjemosyntetisk avhengig makrofauna og bakteriell fauna, som også viser sammenhenger til områder i direkte nærhet til de varme kildene langs Mohnsryggen (Siboglinidae-samfunn) (Olssen mfl. 2016, Åström mfl. 2017).
De geologiske strukturene påvirker oseanografien - og lokalt vil det oppstå “seamounts fenomen” som retensjon og lokal oppstrømning (McClain mfl. 2010, Etnoyer 2010, Schlacher mfl. 2014, White mfl. 2007). Tette samfunn av filtrerende dyr, deriblant svampeskoger og korallhager (Burgos mfl. 2020, Meyer mfl. 2019, Roberts mfl. 2018) vil være knyttet til disse områdene.
Rett ved Molloyryggen er Molloydypet, det dypeste punktet i Nord-Atlanteren (5600 muh).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Fisk
Bunnsamfunn
Den kjemosyntetiske faunaen og de virvelløse dyrene som lever i symbiose med kjemosyntetiske bakterier ved de varme havbunnskildene er fullstendig avhengige av de varme havbunnskildene og det spesielle kjemiske og fysiske miljøet rundt kildene (f.eks. Kongsrud og Rapp 2012, Tandberg mfl. 2012, Steen mfl. 2016). De unike økosystemene vi finner ved de aktive kildene vil kun finnes så lenge kildene er aktive, og vil erstattes med andre, unike, økosystem når kilden er utdødd (sulfitthaug-system). Nye kilder som oppstår vil relativt raskt koloniseres hvis det finnes nærliggende aktive kilder, slik at gravide hunndyr eller larver kan spre seg dit (Eilertsen mfl. 2018).
De kalde gasshydrat-områdene (Vestnesaryggen) har også en rik endemisk fauna som er avhengig av hydratsystemet. Det vurderes om kalde gassoppkommer kan være i bruk som “stepping stones” for varm-kilde-endemisk fauna. I så fall er dette området ikke bare viktig for seg selv, men også for Mohnsryggen og ryggene videre sørover (Åström mfl. 2017).
Det er kjent fra andre spredningsrygger og undersjøiske fjell at de kan fungere som viktige gyte- og oppvekstområder for noen fiskearter (Hareide 1995, Hareide og Garnes 2001, Henry mfl. 2013, 2016). Dette er ikke blitt studert ved den arktiske delen av den midtatlantiske ryggen eller undersjøiske fjell.
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Sjøpattedyr
Bunnsamfunn
Svampeskoger og korallhager er funnet på flere lokaliteter og modellert over store deler av den arktiske del av den midtatlantiske ryggen (Roberts mfl. 2018, Meyer mfl. 2019, Burgos mfl. 2020). De er vurdert som truet av OSPAR-kommisjonen (OSPAR 2008) og står på den norske rødlisten for naturtyper (Artsdatabanken 2018).
Faunaen ved de aktive og passive kildene er ofte delvis endemisk. Habitatene er på norsk rødliste (varm havkildebunn M12, LC) (Artsdatabanken 2018), og artene er vurdert som kvalifisert til å bli rødlistet (Rødlista 2021, under forberedelse).
Kalde gassoppkommer står som livskraftig (LC) i norsk rødliste for naturtyper (M11), men det foreligger veldig begrenset informasjon om naturtypen slik av dette kan forandres raskt hvis ny informasjon tilkommer (Artsdatabanken, 2018). Kilde-endemisk fauna har et svært begrenset leveområde og må slik anses som truet så lenge habitatet (kildene) er truet.
Tilgjengelige observasjonsdata antyder at utbredelsen av nebbhval i Nord-Atlanteren er knyttet til den midtatlantiske rygg, men det er behov for økt kunnskap.
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Bunnsamfunn
Varm havkildebunn (M12) står som intakt (LC) på den norske rødlisten for naturtyper. Svampeskoger og korallområder har flere NIN-kategorier som alle strekker seg fra NT (nær truet) til EN (sterkt truet) (Artsdatabanken 2018).
Både svamper og koraller er saktevoksende arter som kan bli svært gamle, men har dårlig evne til å restituere seg (Doughty mfl. 2014, Lacharité og Metaxas 2013, Mortensen og Buhl-Mortensen 2005, Rogers 2004, Roberts mfl. 2009). Filtrerende arter, særlig svamp, kan være meget sensitive for forhøyede mengder suspenderte partikler i vannet (Fang mfl. 2018, Kutti mfl. 2015, Scanes mfl. 2018). Mange arter ved varme og kalde kilder samt i sulfitthaugene ved utdødde kilder er totalt avhengige av det spesielle bunnsubstratet, og restitusjonsevnen av et ødelagt habitat er meget lav (Ramirez-Llodra mfl. 2020).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Plante-plankton Dyreplankton
Bunnsamfunn
Enkelte områder i de øvre vannlag har vist høyere biomasse av dyreplankton (ICES 2020, 2021). Enkeltstudier har ikke vist høyere produksjon av planteplankton i selve fronten (Erga mfl. 2014), men høye konsentrasjoner av plante- og dyreplankton i nærheten av fronten er observert og forklart med mesoskala fysiske prosesser (C. Broms pers. medd.). Det kan imidlertid være mellomårlige variasjoner, og det er gjort for få studier av SVO-et og selve fronten til å si noe sikkert ang. produktiviteten av plankton.
Svampskoger og korallskoger er høyproduktive områder som spiller en svært viktig rolle for karbonsyklusen på bunn (Cathalot mfl. 2015, Kutti mfl. 2013). Det er for eksempel beregnet at korallrevene på norsk kontinentalsokkel er ansvarlig for 30 prosent av all omsetning av organisk materiale ved bunnen til tross for at de dekker kun 1 prosent av arealet på den norske sokkelen.
Kombinasjonen av at de store havstrømmene styres av ryggene samt at særlig de aktive varme kildene beriker vannet i sin umiddelbare nærhet med oppløste mineraler som er egne og uavhengige næringskilder for hele økosystem. Det er endemisk mikrobielle samfunn i aktive hydrotermale områder (Olsen mfl. 2016, Orcutt mfl. 2020), og tette samfunn av filtrerende dyr som bygger svampskoger og korallhager (Fossum mfl. 2012, Burgos mfl. 2020, Meyer mfl. 2019, Roberts mfl. 2018). Slike endemiske samfunn viser en meget høy lokal produktivitet langs de forskjellige delene av den midtatlantiske ryggen stedvis er særdeles høy (van der Meeren mfl. 2021).
Knipovichryggen strekker seg så langt nord at den i deler av året overlapper med iskantsonen, som er produktiv pelagisk (Hegseth 1998, Leu mfl. 2011, 2015).
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Dyreplankton
Bunnsamfunn
Da ulike vannmasser og habitater som inneholder forskjellige dyreplanktonarter og utviklingsstadier møtes ved fronten, kan det ikke utelukkes at diversiteten er høyere i fronten enn andre nærliggende områder (Melle mfl. 2004).
Svampskoger og korallskoger er levested for en rekke andre arter, fra mikroorganismer til fisk, og er ofte assosiert med en kraftig forhøyet biodiversitet (Klitgaard 1994, Freiwald mfl. 2012). Både arktiske dype hardbunnslokaliteter og ekstremt dype bløtbunnslokaliteter vil mest sannsynlig ha unik fauna. Dette er vist på ryggene både tilgrensende nord og tilgrensende sør for Molloyryggen (Jakobsen 2020, Mareano.no).
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
Bunnsamfunn
Bunnområdene er relativt urørt i de dypere delene av området (Mareano.no).
9.6.3 - Lokalisering
Forslag til SVO Midtatlantisk rygg er et nytt SVO som er basert på ny kunnskap og løpende kartlegging.
Det nye forslaget SVO Midtatlantisk rygg omfatter Jan Mayen-ryggen, Jan Mayen-bruddsonen, Mohnsryggen, Knipovichryggen og Molloyryggen/Molloydypet og skaper skjæringspunktene mellom Grønlandshavet i nord, Islandshavet i sørvest samt Norskehavet i sørøst (figur 9.6.1). Området begrenses ved hjelp av geomorfologisk tolking av havbunnsdata og kart over undersjøiske rygger, fjell, spredningsrygger og riftdaler fra Harris mfl. (2014).
Forslag til nytt SVO Midtatlantisk rygg overlapper delvis med eksisterende forslag til utvidetendret SVO Jan Mayen som fagekspertgruppen foreslår at utvides noe og med eksisterende SVO Den arktiske front som fagekspertgruppen foreslår blir fjernet. Området overlapper også så vidt med det eksisterende SVO Vesterisen.
Det er ikke opprettet vernesoner i dette området.
9.6.4 - Introduksjon til området
I riften mellom den nordamerikanske og den eurasiske kontinentalplaten nord fra Island mot Polhavet strekker den midtatlantiske rygg seg ut i en del som ofte blir kalt AMOR (Arctic Mid Ocean Ridge). AMOR består av flere forskjellige rygger og bruddsoner, med de sentrale delene i norske havområder. Lengst i sør strekker Kolbeinsryggen seg fra Island og opp til Jan Mayen-bruddsonen. Jan Mayen-bruddsonen strekker seg så østover frem til, og forbi øya Jan Mayen til den går over i Mohnsryggen. Tretti km vest for Bjørnøya gjør AMOR en 80 graders sving og går over i Knipovichryggen. Denne ryggen strekker seg 550 km i nordlig retning og ender i Molloydypet, med det største dyp både for norske havområder og i hele Arktis, rett vest for de nordlige delene av Spitsbergen. Den korte Molloyryggen strekker seg videre nordover mot Svalbardbruddsonen og Lenadypet og går over i Gakkelryggen som strekker seg tvers over Polhavet (figur 9.6.2).
Figur 9.6.2. Figur 1 fra Pedersen og Bjerkgård 2016. Oversikt over aktive (røde) og passive (oransje) varme kilder, sulfid-hauger (grønne) og varme utstrømninger (gule) langs den midtatlantiske ryggen nord for Island.
Bunnstrømmene i området er til en viss grad bestemt ut fra topografien, der ryggen får stor dominans (Voet mfl. 2010). I grove trekk kan vi si at øst for den midtatlantiske ryggen i Norskehavet er vannet dominert av varme atlantiske vannmasser, mens på vestsiden av ryggen i Islandshavet og Grønlandshavet er det dominert av kaldere arktiske vannmasser. Dette skaper en skarp front i det øvre laget, den arktiske fronten, som har en relativ fast lokalisering over ryggen. Det er store temperatur- og saltholdighetsendringer på tvers av fronten, og nord for Jan Mayen gir dette relative sterke strømmer langs fronten, og i kjernen av fronten er strømhastighetene rundt 50 cm per sek (Bosse og Fer 2019). Sør for Jan Mayen vil derimot temperatur- og saltholdighetsendringene kompensere hverandre med hensyn til tetthetsendringene på tvers av frontene, noe som medfører at strømmene langs fronten her er svakere og gir mindre vertikale strømninger (Fossum mfl. 2012). I dypet er strømmen svakere, men nord for Jan Mayen er det relativt høye hastigheter, 10–20 cm per sek, langs ryggen (Bosse og Fer 2019). I kombinasjon med den røffe bunntopografien bidrar dette til økte vertikale strømmer og blanding i dypet langs Mohns- og Knipovichryggen (Bosse og Fer 2019, Nikurashin og Ferrari 2011). I Molloydypet har fraksjoner av dypvann fra Grønlandshavet, Norskehavet og Polhavet variert over tid, men i de senere årene har Molloydypet vært dominert av dypvann fra Polhavet (Langehaug og Falck 2012).
Figur 9.6.3. Kart over gjennomsnittlig overflatestrøm (piler) og bunndyp (fargekoder) i SVO Midtatlantisk rygg og omkringliggende områder.
Den midtatlantiske rygg er i liten grad kartlagt (figur 9.6.4). Beskrivelsen er derfor basert på all tilgjengelig kunnskap, men det er store ubeskrevne arealer som gjenstår å kartlegge.
Fig 9.6.4. Kart over Mareanos kartlegging inkludert Norskehavsundersøkelsene 2019 (gule kvadrater), med undersøkte lokaliteter fra Den midtatlantiske rygg og ellers i Norskehavet og Barentshavet (Kart fra Mareano.no). Her vises ikke kartlag for miljøverdier da kartleggingen fremdeles ikke har nok detaljer om utbredelse av de fleste miljøverdiene som omtales.
Jan Mayen-ryggen (BT1, figur 9.6.1) strekker seg rett sørover fra øya Jan Mayen i Norskehavet. Området preges av varmt vann på østsiden (fra den norske atlanterhavsstrømmen) og kaldt arktisk vann på vestsiden (fra sidegrener til den sørgående strøm på vestsiden (Øst-Grønlandsstrømmen). Bunntopografien styrer sirkulasjonen av vannmassene, og den arktiske front følger her Jan Mayen-ryggen. Seks undervannsfjell er blitt identifisert på Jan Mayen-ryggen ved å studere batymeteriske data (Gaina mfl. 2017).
Vest og øst for den aktive vulkanen Jan Mayen finner man en aktiv forkastningssone på omtrent 530 km kalt Jan Mayen-bruddsonen (BT2, figur 9.6.1). I denne sonen er den midtatlantiske ryggen forskjøvet. Den vestre delen av bruddsonen er omtrent 130 km lang og er assosiert med mange undervannsfjell, varme kilder og områder med hydrotermal aktivitet. Seks undervannsfjell er blitt identifisert. En banke assosiert med bruddsonen (Marø Bank: 09°24,79’W, 71°07,96’N, 170 m) har navn og er blitt kartlagt ved hydrografiske undersøkelser (IHO_IOC GEBCO Gazetteer of Undersea Feature Names). Seks undervannsfjell er blitt identifisert på Jan Mayen-ryggen ved å studere batymeteriske data (Gaina mfl. 2017), men de er ikke blitt undersøkt verken hydrografisk eller biologisk.
Mohnsryggen (BT3, figur 9.6.1) strekker seg 550 km østover i omtrent 60 graders retning fra Jan Mayen og frem til “hjørnet” Mohns-Knipovich-svingen, 300 km vest av Bjørnøya. Området preges av Jan Mayen-strømmen som fører kaldt arktisk vann østover på nordsiden av Mohnsryggen og varmt atlanterhavsvann som strømmer østover på sørsiden av Mohnsryggen og skaper en skarp arktisk front. Ryggen er generelt dårlig studert, men er assosiert med mange undervannsfjell, varme kilder og områder med hydrotermal aktivitet. Langs Mohnsryggen er det flere varme kilder og områder med hydrotermal aktivitet, og med fortsatt fokusert toktaktivitet oppdages det stadig nye områder med geotermisk påvirkning. Førtitre undersjøiske fjell er her blitt identifisert ved å studere batymetriske data (Gaina mfl. 2017). Det største kjente undervannsfjellet er Trollveggen–Soria Moria-formasjonen (Gaina mfl. 2017) som også er kjent for sine forekomster av hydrotermale kilder (Pedersen mfl. 2010). To banker assosiert med Mohnsryggen, Myrsethbanken (02°52,27’W, 71°26,14’N, 2000 m) og Louise A. Boyd-banken (02°50,02’E, 72°39,86’N, 1424 m), har navn og er blitt kartlagt ved hydrografiske undersøkelser (IHO_IOC GEBCO Gazetteer of Undersea Feature Names). I overgangssonen mellom Mohns- og Knipovichryggen finner vi Schulzbanken (07°29,70’E, 73°51,99’N).
Knipovichryggen (BT4, figur 9.6.1) er et omtrent 550 km langt segment av den arktiske midtatlantiske ryggen som strekker seg i rett nordlig retning fra Mohns-Knipovich-svingen i sør til Molloydypet i nord. Ryggen er assosiert med en mengde undersjøiske fjell, der åtte er blitt navngitt og kartlagt ved hydrografiske undersøkelser (IHO_IOC GEBCO Gazetteer of Undersea Feature Names), men det finnes ikke publiserte data av biologisk karakter fra noen av disse undersjøiske fjellene. I sørlige deler av Knipovichryggen finner vi Boytsov Seamount (minimumsdyp på 965 m), Dibner Seamount (minimumsdyp på 990 m) og Chichagov Seamount (minimumsdyp på 990 m). Her finner vi også Gnom Knoll med et minimumsdyp på 2721 m. I de nordlige delene av Knipovichryggen finner vi Pogrebitsky seamount (minimumsdyp på 990 m), Litvin Seamount (minimumsdyp på 840 m) og Gorynych Hills en kjede av 3 topper med det minimumsdyp på 1754 m. Aller lengst nord karakteriseres Knipovichryggen av Svyatogo Rise (med minimumsdyp på 1498 m) i syd og Molloybruddsonen i nord.
Molloyryggen er en kort rygg på omtrent 40 km som strekker seg mellom Molloydypet (BT5, figur 9.6.1) i sør til Svalbardbruddsonen i nord, omtrent 130 km vest om det nordligste punktet på Spitsbergen. Det inntilliggende Molloydypet er på 5569 m og er det dypeste punkt i norske havområder. Molloyryggen er assosiert med flere undersjøiske fjell. På Molloyryggen har tre undersjøiske fjelltopper blitt navngitt og kartlagt ved hydrografiske undersøkelser (IHO_IOC GEBCO Gazetteer of Undersea Feature Names), dvs. Atla Seamount, Eistla seamount og Gjalp seamount. Molloyryggen strekker seg opp fra ca. 2500 m fra omkringliggende havbunn til 1330 m på sitt høyeste punkt. Det er blitt observert en plume (vannstrøm) fra en varm kilde, men selve kilden er enda ikke funnet og derfor ikke kartlagt.
pH-verdiene i overflatevannet er på rundt 8,1 til 8,15 langs hele ryggen, mens det er store gradienter i metningsgrad for kalkmineraler på tvers; metningsgraden er mindre på vestsiden, i det arktiske vannet, og høyere på østsiden (Fransner mfl., 2020, Lauvset mfl. 2016). Dette skyldes at det arktiske vannet er kaldere, har høyere konsentrasjoner av løst uorganisk karbon, samt lavere alkalinitet enn det atlantiske vannet i øst. For aragonitt er metningsgraden rett over to på østsiden, og rett under to på vestsiden. For kalsitt er metningsgraden høyere, i overkant av tre på østsiden og i underkant av tre på vestsiden. Årsaken til at vi ikke ser igjen disse gradientene i pH, er at pH er sterkt påvirket av temperatur, den øker med lavere temperaturer, som i det arktiske vannet.
Tilgjengelige måledata (Olsen mfl. 2020) viser at hele vannsøylen over den midtatlantiske ryggen er overmettet med kalkmineralet kalsitt, og verdiene avtar fra overflatevannet til rett i overkant av 1 på rundt 3000 meters dyp. På 1000 m er metningsgraden rundt 2. Det finnes ikke data fra Molloydypet. Kalkmineralet aragonitt er mer løselig enn kalsitt, og metningsgraden er derfor lavere. Metningshorisonten ligger i dag på 2000 meter, men vil bli grunnere etter hvert som havet tar opp mer CO2.
9.6.5 - Beskrivelse av miljøverdiene
Plankton
Det er ingen generell økning i dyreplanktonbiomassen i pelagialen og øvre vannlag over den midtatlantiske ryggen på våren (mai) eller sensommeren (juli/august), som er tidspunktene for Havforskningsinstituttets storskala overvåkningstidsserier, sammenliknet med omkringliggende områder. Enkeltområder kan derimot ha høyere biomasse en gjennomsnittet i forslaget til nytt SVO- (ICES 2020 a, 2021), bl.a. ved Jan Mayen (se forslag til endret SVO Jan Mayen). Det foreslåtte SVO-et dekker et utstrakt område i nord-sør-retning, og det vil derfor være tidsforskjeller i produksjonen (Paasche 1960 , Rey 2004 , Broms og Melle 2007). Selv om det i Norskehavet er en generell forsinkelse i planktonproduksjon fra norskekysten mot åpent hav og a tlantisk e og a rktiske vannmasser (dvs. en forsinkelse fra øst mot vest; Rey 2004 , Broms og Melle 2007 , Bagøien mfl. 2012), kan SVO-et ha en tidligere produksjonsstart sammenliknet med sentrale deler av Norskehavet (Rey 2004).
Den arktiske fronten har en relativt fast lokalisering over den midtatlantiske ryggen, og generelt vil frontsystemer kunne være mer produktive enn omgivende vannmasser dersom vertikal blanding av vannmassene bringer næringsrikt vann mot overflaten. Det er generelt lite informasjon angående planktonproduksjon langs fronten over det foreslåtte SVO-et . Økt produksjon ble imidlertid ikke observert i et undersøkt område av fronten sør for Jan Mayen, hvor temperatur- og saltholdighetsegenskapene til vannmassene ved fronten førte til tetthetskompensering og dermed en svak horisontal tetthetsgradient. Dette førte igjen til en svak vertikal blanding av vannmassene og fronten gav ingen stimulerende effekt på produksjonen av planteplankton (Erga mfl. 2014). Undersøkelser i Norskehavet sommeren (juni) 1952, 1953 og 1954, indikerer ikke høyere planteplanktonbiomasse eller -produksjon i pelagialen over det foreslått nye SVO-et nordøst for Jan Mayen (Paasche 1960 , Ramsfjell 1960). Estimat av primærproduksjon basert på satellittinformasjon for perioden 1998-2018 indikerer også at den arktiske fronten ikke har høyere produksjon enn omkringliggende områder (Henson mfl. 2009 , Mayot mfl. 2020). Rey mfl. (2000) fant stor grad av variasjon i perioden 1993 – 1995 i området, men høyere produksjon i Grønlandshavet enn i selve fronten. På lik linje med planteplankton, ble det heller ikke observert noen økning i biomassen av dyreplankton i undersøkelser av den arktiske fronten lokalisert over ryggen sør for Jan Mayen. Høye konsentrasjoner av planteplankton og meso-dyreplankton ble derimot observert nær fronten og knyttet til virvelaktivitet (Broms mfl. under utarbeidelse). Vi kan derfor ikke se bort fra at fronten kan bidra til økt produksjon av plankton f.eks. ved mesoskala fysiske prosesser som virvelaktivitet i dette nye forslaget til SVO. Frontområdene kan også være mer stabile og forutsigbare når det gjelder planktonets produksjonssesong, og det kan være horisontal adveksjon av plankton inn mot frontene som resulterer i høyere biomasse. Frontområdene kan derfor være viktige beiteområder og føre til høyere produksjon oppover i næringskjeden, selv om det ikke er vist høyere planktonproduktivitet i området. De fysiske strukturene i fronten over ryggen kan variere mellom år, og ulike deler av fronten kan opptre forskjellig og dermed gi variasjon i produktiviteten til plankton. Det er gjort for få studier av selve fronten til å si noe sikkert angående planktonets produktivitet i området, men fordi det så langt ikke er dokumentert generell forhøyet produksjon i frontområdet sammenlignet med områdene rundt (høy produktivitet var et viktig argument for det eksisterende SVO Den arktiske front), blir SVO Den arktiske front ikke lenger anbefalt som et selvstendig SVO.
Da atlantiske og arktiske vannmasser dominerer hhv. øst og vest for ryggen, blir de pelagiske vannmassene over ryggen et møtested for ulike habitater som inneholder ulike dyreplanktonsamfunn. Det vil derfor kunne være høyere diversitet i området. Calanus finmarchicus dominerer dyreplanktonbiomassen i atlantiske vannmasser, og finnes også i store mengder i a tlantisk og a rktisk blandingsvann, mens C. hyperboreus dominerer i a rktiske vannmasser (Hirche 1991 , Melle mfl. 2004 , Broms mfl. 2009). Flere av krillartene har en utbredt fordeling og finnes i vannmassene på begge sider av ryggen og fronten. Biomassen av amfipoder er høyest på den arktiske siden av fronten pga. høye konsentrasjoner av den store arktiske arten Themisto libellula (Dalpadado mfl. 1998). Sesongutviklingen av C. finmarchicus er forsinket på den arktiske siden av fronten sammenliknet med den atlantiske siden, noe som fører til ulik sammensetning av stadier på tvers av fronten (Broms mfl. under utarbeidelse). Et område på tvers av ryggen vil dermed kunne inneholde større diversitet av utviklingsstadier sammenliknet med andre områder. En rekke arter av ulike dyreplanktongrupper er observert, men det er ikke utført diversitetsundersøkelser som kan si om området har høyere diversitet enn områdene rundt. I tilkn ytning til Jan Mayen er det derimot observert høy primærproduksjon og høy biomasse av ulike dyreplanktonarter, s e forslått endret SVO Jan Mayen (Kap. 9.5.5) .
Fisk
Sild (Clupea harengus) beiter i dette området gjennom sommeren, men hovedbeite foregår i de t sørvestlige og sentrale Norskehavet. Sild kan nyttiggjøre seg av store mengder C. hyperboreus i frontområdet mellom Norskehavet og Grønlandshavet nord for Jan Mayen (Broms 2007), til tross for at den synes å unngå arktiske vannmasser pga. for lave temperaturer tidlig i beitesesongen (Misund mfl. 1997 , 1998). Observasjoner fra dette området viser individer med fulle mager, og det er antatt at beiteintensiteten er begrenset av magevolum og tømmingsrate (Holst mfl. 2004). I disse sentrale områdene kan variasjoner i posisjonen av den arktiske fronten påvirke utbredelsen av norsk vårgytende sild og makrell i beiteperioden om sommeren/høsten. Områder vest av fronten er karakterisert av arktiske vannmasser som disse pelagiske fiskeartene i stor grad unngår.
Fiskesamfunnet i abiotisk sone er rikt, og en ålebrosmeart (Lycenchelys platyrhina) som er endemisk for Jan Mayen-ryggen er blitt funnet (Fossum mfl. 2012), også nevnt under SVO Dypområdene i Norskehavet (Kap. 10.5.5).
Mesopelagisk fauna
Ut fra dagens kunnskap er det ikke funnet at mesopelagisk fauna utgjør en særlig miljøverdi i dette området.
Bunnsamfunn
Bunnsamfunn grunnere enn 200 m (“Grunt vann”) opptrer nær eksisterende/forslag til endret SVO Jan Mayen og er beskrevet der (Kap. 9.5.5). Videre beskrivelse av bunnsamfunn er fra den abiotiske sonen fra 200 m og ned til mer enn 5000 m dyp.
Områder med hardt substrat på Den midtatlantiske rygg ser ut å være kolonisert av tette samfunn av filtrerende dyr (Burgos mfl. 2020). Filtrerende svamp og korall bygger tredimensjonal struktur på bunnen. Dette fungerer som habitat for mange andre arter, og det har en viktig funksjon i kretsløpet av organisk karbon og næringssalter på bunn. Samfunn bygget av svamp og korall (“coral gardens” og “sponge grounds”) står på OSPARs liste over truede og minkende habitat. Undervannsfjell (såkalte “seamounts”) står også på den listen fordi de ofte er kolonisert av samfunn av korall og svamp og fordi de kan fungere som gyteområder for populasjoner av langsomtvoksende fisk. Hardbunnskorallskog står også som nær truet (NT) på den norske rødlisten for naturtyper.
Den midtatlantiske ryggen har både aktive og utdødde (passive) varme havbunnskilder, og det er også flere frittliggende sulfitthauger som gjenstår etter tidligere varme kilder. Disse har mye endemisk fauna som er både spesialisert i forhold til å tåle høye temperaturer og ikke minst ved at mange av artene er kjemosyntetiske eller lever i symbiose med kjemosyntetiske mikroorganismer, noe som betyr at de ikke er avhengige av en næringskjede som starter med sollys og primærproduksjon. Både artene og sammensetningen av de høyere taksa er unik sammenlignet med andre varme kilder i verden. Det er forventet at nærmere undersøkelser vil vise flere varme kilder. Varm havkildebunn står som nær truet (NT) på den norske rødlisten for naturtyper.
Jan-Mayen ryggen (BT1, figur 9.6.1) er topografisk sett relativ lite kompleks og bunnen består i større deler av leireblandet mudder med spredte forekomster av enslige steiner som er blitt ført til området av istransport (såkalte drop stones). Disse steinene og annet hardt substrat er ofte kolonisert av fastsittende fauna som svamp, mosdyr, hydroider, anemoner, bløtkoraller, fjærstjerner og sjøfjær i høye tettheter. På områder grunnere enn 800 m er det blitt observert samfunn med tette forekomster av fjærstjerner, bløtkorallen Gersemia rubiformis, svampen Stylocordyla borealis og slangestjerner. Også dyphavssjøfjærene Umbellula encrinus og Virgularia arter forekom på noen stasjoner med veldig høye tettheter. Totalt er det blitt registrert 120 forskjellige taksa (Fossum mfl. 2012), og området er å betrakte som relativt artsrikt. Prediktiv habitatmodellering har vist at tette samfunn av svampene Geodia parva, G. hentscheli, blomkålkoraller og sjøfjær med høy sannsynlighet forekommer på deler av Jan Mayen-ryggen (Burgos mfl. 2020).
I bruddsonen vest for øya Jan Mayen (BT2, figur 9.6.1) er det funnet flere aktive varme kilder. Den svært grunne Seven Sisters Vent Field (Marques mfl. 2020) på 140 m dyp med en ut-vanns temperatur på ~200 °C (Olsen mfl. 2016) rett vest av øya Jan Mayen, samt Jan Mayen Vent Field (JMVF) like nord for øya Jan Mayen. JMVF består av de separate kildene Perle og Bruse (Stensland mfl. 2019), Trollveggen (Schander mfl. 2010) og Soria Moria (Schander mfl. 2010), alle på dyp mellom 500 og 750 m. Kildene på JMVF har en ut-vanns temperatur på 260–270 °C (Pedersen mfl. 2005). Det er lite endemisk eller spesifikt kjemosyntetisk fauna på de “grunne” (grunnere enn 1000 m) varme kildene som ligger i Jan Mayen-bruddsonen, men området er betegnet av en svært rik og divers fauna (Schander mfl. 2010). Nyere undersøkelser på tilsvarende dype varme kilder sør for Island viser at mer spesifikke undersøkelser av overgangssonene mellom bakgrunnsfaunistiske områder og kilde-påvirkete områder avslører mulig endemisk fauna (Taylor mfl. in press). Prediktiv habitatmodellering indikerer en rik fauna med tette samfunn av filtrerende svamper, som Geodia parva og G. hentscheli, blomkålkorall og sjøfjær assosiert med banker og undervannsfjell i området. Jan Mayen-bruddsonen har spredte forekomster av sjeldne, sårbare og livshistorisk viktige økosystem (f.eks. varm havkildebunn) med endemiske arter samt spredte forekomster av undervannsfjell (seamounts) med sårbare økosystem bygget av svamp og korall (Burgos mfl. 2020).
Langs Mohnsryggen (BT3, figur 9.6.1) er det flere varme kilder og områder med hydrotermal aktivitet, og med fortsatt fokusert toktaktivitet oppdages det stadig nye områder med geotermisk påvirkning. De mange aktive kildene langs Mohnsryggen kommer i tillegg til inaktive (“døde”) kilder, der den gjenstående kildestrukturen (mound) ofte er habitat for en helt egen fauna, noe vi har for eksempel på Mohns Treasure (Paulsen 2017, Ramirez-Llodra mfl. 2020). Av de varme kildene langs Mohnsryggen er Lokeslottet den best studerte, med flere beskrevne endemiske makrofaunaarter (e.g. Kongsrud mfl. 2017, Kongsrud og Rapp 2012, Tandberg mfl. 2012, 2018) og særegne mikrobielle systemer (se Steen mfl. 2016). I motsetning til faunaen ved varme kilder beskrevet i andre deler av verden, er de dype varme kildene langs AMOR dominert av amfipoder (i motsetning til overveiende dekapoder ved andre varme kilder) og rissoide snegler, med en polychaetfauna som er dominert av taksa av mye mindre størrelse enn de kjente “gigant-polychaetene” ved sørligere kilder. I senere tid er også de varme kildene Ægirs Kilde (Pedersen og Bjerkgård 2016) og Fåvne (Sivertsen 2020) oppdaget og begynt kartlagt, begge med mye, sannsynlig endemisk og kilde-spesialisert makrofauna. De to sulfid-avsetningene Copper Hill og Mohns Treasure er også rikt bebodd av makrofauna (Ramirez-Llodra mfl. 2020). Det diskuteres om kolonalisering av de i geologisk sammenheng temporære varme kildene kan skje gradvis etter “stepping-stone”-prinsippet, med makrobiologisk nedfall (“fall biota”) og kalde gassoppkommer som mellomsteg (se f.eks. Smith 1989, Tandberg 2013). Dette er særlig viktig for taksa som ikke har planktoniske stadier som kan hjelpe spredning til andre plasser (f.eks. Amfipoda som har direkte utvikling). Schulzbanken nær Knipovichryggen er karakterisert av høye tettheter og diversitet av filtrerende svamper (Roberts mfl. 2018). På toppen av banken dominerer glassvamper fra slektene Schaudinnia og Asconema (med tettheter på opptil 200 individ per bilde) mens den dypeste delen av banken (fra 1000 til 1400 m dyp) er dominert av massive svamper som Geodia hentscheli og Stelletta rhaphidiophora. Schultzbanken blir regelmessig oversvømt av varmt og oksygenrikt vann fra overflaten og er karakterisert av strømninger som fører til retensjon av partikler rundt banken slik at tilgangen på mat til bunndyr blir forhøyet (Roberts mfl. 2018, Meyer mfl. 2019). Slike hydrodynamiske faktorer ser ut til å være et generelt trekk ved undervannsfjell (White mfl. 2007) og en hovedfaktor til de tette samfunnene av filtrerende svamp og korall som vanligvis blir funnet på undervannsfjell (McClain mfl. 2010, Etnoyer 2010, Schlacher mfl. 2014). Mohnsryggen er relativt lite studert, og Schultzbanken er den eneste banke eller undervannsfjell det finnes publisert biologisk og oseanografisk informasjon om. Sårbare samfunn av svampene Geodia parva og G. hentscheli er blitt funnet på Schultzbanken i området Mohns-Knipovichsvingen og modellert langs hele Knipovichryggen.
E-sygynea Loki. Foto: KG Jebsen, UiB
Knipovichryggen (BT4, figur 9.6.1) er relativt lite studert sammenlignet med de andre delene av den arktiske del av den midtatlantiske ryggen. Connelly mfl. (2007) detekterte signaler i vannsøylen som tyder på hydrotermal aktivitet på havbunn sør på Knipovichryggen, men den eksakte lokaliteten til kilden er enda ikke kjent. Knipovichryggen har spredte forekomster av sjeldne, sårbare og livshistorisk viktige økosystem (f.eks. kalde gassoppkommer og varm havkildebunn) med endemiske arter. Prediktiv habitatmodellering har vist at tette samfunn av svampene Geodia parva og G. hentscheli med høy sannsynlighet forekommer over store deler av Knipovichryggen (Burgos mfl. 2020).
Gasshydratsystemet på Vestnesaryggen ligger kun kilometer fra den midtatlantiske spredningsryggen, øst for Molloydypet og nord for Molloybruddsonen, og er en av de best studerte kalde gassoppkommene i norske farvann. Der er det blitt dokumentert et egenartet bunndyrsamfunn, karakterisert av bakteriematter og børstemark (fra familien Siboglinoidea) med endemiske kjemosyntetiske symbiotiske bakterier, som tydelig skiller seg tydelig fra omkringliggende områder gjennom å være mer artsrik og ha større tetthet og biomasse av små bunndyr (Åström mfl. 2017). De ligner med det på de bløte bunner ved barittfeltene i direkte nærhet til varme kilder langs Mohnsryggen (Olsen mfl. 2016). På harde gasshydratskorper i oppkommet er det blitt identifisert fastsittende bryozoa, hydroida og porifera (Olsen mfl. 2016). Langsom utlekking av lave mengder metan fra reservoarer av gasshydrater har også blitt detektert lenger sør på Knipovichryggen (Johnson mfl. 2015), men biologiske samfunn rundt de kildene er ikke blitt studert.
Svært lite er kjent om de spesifikke bunndyrsamfunnene langs Molloyryggen og Molloydypet (BT5, figur 9.6.1). Det finnes ikke publiserte data av biologisk karakter fra dette området, men sårbare samfunn av svampene Geodia parva og G. hentscheli er modellert langs hele Molloyryggen og er blitt funnet på undersjøiske fjell både sør og nord for Molloyryggen. Det har nylig (sommeren 2020) vært utført tokt som har samlet prøver i både Molloydypet og langs Molloyryggen (Mareano.no), men ingen data er opparbeidet til en publiserbar standard enda. Kvalitative beskrivelser nevner store mengder av karnivore svamp fra slekten Asbestopluma dyphavssjøpølser fra slekten Elpidia, samt store mengder av “små krepsdyr” (som vanligvis vil være føde for Asbestopluma) på de dypeste områdene (Jakobsen 2020). Pedersen og Bjerkgård (2016) nevner at det er observert en plume (vannstrøm) fra en varm kilde på Molloyryggen, men selve kilden er ikke funnet og derfor ikke kartlagt. Basert på det vi kjenner av varme kilder lengre sør langs AMOR, samt det vi vet om observasjoner på varme kilder lenger nord på den tilstøtende Gakkelryggen (Boetius mfl. 2016, Hacon tokt 2019 (ingen publisert toktrapport til nå)) vil varme kilder langs AMOR ha tilhørende spesialisert fauna.
Sjøpattedyr
Geografisk fordeling av observasjoner under hvaltelletoktene i perioden 2014 – 2018 rapportert av Leonard og Øien (2020) indikerer at den midtatlantiske ryggen kan være viktig sommerbeiteområde, særlig for nordlig nebb hval (Hyperoodon ampullatus) , mens spermhval (Physeter macrocephalus) , spekkhogger (Orcinus orca), knølhval (Megaptera novaeangliae) og blåhval (Balaenoptera musculus) u tnytter store deler av Norskehavet i tillegg. Dette foreslåtte nye SVO-et anses vanligvis for å være et transittområde for vandrende bardehval mellom sørlige områder og de nordlige beiteområdene. Utbredelsen og tallrikheten av nebbhval er dårlig kjent, men den informasjonen som foreligger, antyder at nebbhvalen har en tilknytning til den midtatlantiske ryggen fra Jan Mayen til Spitsbergen (Nils Øien pers. medd.). Fra satellittmerkinger (Folkow med flere 1996, 2010) vet vi at klappmyss beiter i kantene til dyphavsområdene i Norskehavet, ikke minst langs den dem midtatlantiske ryggen (se figur 9.10.3).
Sjøfugl
Deler av dette området tilknyttet den arktiske fronten, er overvintringsområde for flere sjøfuglarter, særlig pelagiske arter som havhest (Fulmarus glacialis), polarlomvi (Uria lomvia) og alkekonge (Alle alle), i overensstemmelse med data fra SEATRACK (http://seatrack.seapop.no/). Avgrenset område dekker kun en mindre andel av det totale leveområdet for sjøfuglene, og tetthetene er lave. Områdene ved Jan Mayen er viktige for sjøfugl, og dekkes av den foreslåtte utvidelsen av SVO Jan Mayen. I denne sammenhengen er beiteområdet en mindre del av SVO Midtatlantisk rygg, og miljøverdien Sjøfugl er derfor beskrevet i foreslått endret SVO Jan Mayen (Kap. 9.5.5).
9.6.6 - Pågående og framtidige endringer
Havforsuring
Som følge av økt innhold av CO2 i atmosfæren, vil havforsuringen kunne endre løseligheten av metaller i havet. Med redusert pH minker hydroksid- og karbonationer, og disse endringene vil påvirke fordeling, frigjøring og tilgjengelighet av metaller som danner sterke kompleks med disse ionene (Millero mfl. 2009). Marine organismer trenger spormetaller for å vokse, for eksempel jern, mangan og sink. Havforsuring kan også føre til endret frigjøring og toksisitet av ulike tungmetaller slik som kvikksølv, bly, kadmium, nikkel og kobolt (Stockdale mfl. 2016). Metylkvikksølv er den organiske formen av kvikksølv som er giftig for levende organismer. Organisk bundet kobber og kadmium vil minke med 10 prosent ved økt havforsuring, mens organisk bundet kobolt og nikkel vil øke. Det er sannsynlig at klimaendringer og påfølgende havforsuring vil endre metallsyklusene i havet (Hoffmann mfl. 2012), noe som kan gi en betydelig effekt på marine mikroorganismer (Stockdale mfl. 2016).
Åpning av utredninger for mineralutvinning
Flere av de kartlagte og potensielle varme kildene og gassoppkommene har mineraloppsamlinger (massive sulfidforekomster rundt de varme kildene), og det er et økende press for å utvinne mineralene herfra. Det er ventet store økologiske effekter av mineralutvinning både fordi man ved utvinning vil ta bort all naturlig havbunn i utvinningsområdene, og vil suspendere store mengder uorganiske partikler som kan påvirke både de fastsittende og filtrerende dyrenes mulighet til å effektivt ta opp føde, og primærproduksjonen lokalt. I høringssvar på forslaget til konsekvensutredningsprogram for mineralutvinning i området har Havforskningsinstituttet pekt på stor mangel på kunnskap, både med hensyn til biomangfold og økologiske forhold, men også når det gjelder teknologien som vil anvendes for utvinningen (van der Meeren mfl. 2021).
Barittfelt og børstemark, Lokeslottet. Foto: KG. Jebsen, Stiftelse, UiB
9.7 - Forslag til endret SVO Eggakanten sør (NH5)
9.7.1 - Sammendrag
Områdebeskrivelse og strukturer
Forslag til endret SVO Eggakanten sør (NH5) har topografi som påvirker strømmønstre og vanntransport. Sokkelskråningen fører til sterk, ensrettet og smal atlanterhavsstrøm langs Eggakanten. Temperaturen i vannet avtar nordover som følge av varmetap til atmosfæren og blanding med tilstøtende vannmasser. Bunnen kommer opp som en bratt vegg med raviner og kløfter.
Eggakanten strekker seg fra Stad til nordvestspissen av Svalbard. Her beskrives det foreslått endrete SVO Eggakanten sør, fra Stad til grensen mot forvaltningsplanområdet for Barentshavet. Den delen av Eggakanten som ligger i forvaltningsplanområdet for Barentshavet, bekrives som det foreslått endrete SVO Eggakanten nord, beskrives under Barentshavet.
Miljøverdier
I likhet med foreslåtte SVO Eggakanten nord har dette området flere sårbare naturtyper: dyphavsjøfjær, Lophelia-rev, hardbunnskorallskog, og kaldtvanns-svampsamfunn. Antallet kjente korallrev er høyere i foreslått SVO Eggakanten sør enn i foreslått SVO Eggakanten nord. Storneset har eneste sikre observasjon av Madrepora-rev i Norge. Eggakanten og områdene omkring har større mengder av mesopelagiske fisk enn mer vestlige deler av Norskehavet, inkludert forslåtte nye SVOene Midtatlantisk rygg og Dyphavsområdene i Norskehavet. Området inneholder viktige gyteområder for flere fiskearter som f.eks. hyse, og flere dypvannsarter, blant annet vanlig uer som er sterkt truet (EN) samt mesopelagisk fisk.
Tilgangen av plankton og fisk i ulike livsstadier og størrelser gjør dette til et viktig beiteområde. God tilgang på dyreplankton legger til rette for overlevelse av en rekke fiskearters tidligste livsstadier, bl.a. norsk vårgytende sild og torsk som driver nordover langs eggakanten. Eggakanten er imidlertid også viktig beiteområde for en rekke andre fiskearter. Dette er også et svært viktig beiteområde, særlig i hekketida, for pelagisk beitende sjøfugl, deriblant flere rødlistede arter, inkludert alkefugler som lomvi og lunde, som beiter på fiskelarver som driver med strømmen, og for sjøpattedyr, som klappmyss og spermhval.
Svampskog ved Storegga. Foto: Mareano/HI
Figur 9.7.1. Kart over foreslått endret SVO Eggakanten sør (NH5). Grensene for det anbefalte SVO-et er vist med tykk rød strek, mens grenser for overlappende og nærliggende foreslåtte SVO-er som foreligger i dette arbeidet er vist med tynnere rød eller oransje strek. Grensene for forvaltningsplanområdet er markert med brun linje. Kartet viser utbredelsen av utvalgte miljøverdier som er av betydning for utstrekning av foreslått SVO. Miljøverdier knyttet til fisk (ynglefelt), sjøfugl (beiteområde) og bunnsamfunn er vist.
9.7.2 - Vurdering i forhold til EBSA-kriteriene - Forslag til endret SVO Eggakanten sør (NH5)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til endret SVO Eggakanten sør (NH5)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet/sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Bunnsamfunn
Korallverneområdet Storneset inneholder den eneste sikre observasjonen av korallrev dominert av Madrepora oculata (Sundahl mfl. 2020).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Bunnsamfunn
Fisk Sjøpattedyr Sjøfugl
Viktig gyteområde for hyse (Sundby mfl. 2013). Dypvannsarter som vanlig uer (sterkt truet-EN), snabeluer og vassild har viktige gyteområder i området (Cadrin mfl. 2010). Tidlige livsstadier av norsk vårgytende sild, torsk, vanlig uer og snabeluer driver nordover langs eggakanten (Sætre mfl. 2002, Sundby mfl. 2013). Eggakanten og områdene omkring har større mengde av de mesopelagiske fiskene nordlig lysprikkfisk, laksesild, og liten laksetobis, og dette er trolig viktigere gyteområde, sammenliknet med mer vestlige deler av Norskehavet (Dalpadado mfl. 1998, Salvanes 2004).
Eggakanten og øvre sokkelskråningen er viktig beiteområde for spermhval og klappmyss (Folkow og Blix 1999).
Eggakanten er et svært viktig beiteområde for pelagisk beitende sjøfugl, særlig i hekketida (se f.eks. Christensen-Dalsgaard mfl. 2018).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Bunnsamfunn
Fisk Sjøfugl
Vanlig uer (sterkt truet-EN) har viktige gyteområder her (Cadrin mfl. 2010).
Ueren har dessuten preferanse for de til dels unike korallrevene, som er saktevoksende og har sen regenerering (Sundahl mfl. 2020).
Eggakanten er et svært viktig beiteområde for flere rødlistede pelagisk beitende sjøfugl, særlig i hekketida (se f.eks. Christensen-Dalsgaard mfl. 2018).
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Fisk
Bunnsamfunn
Saktevoksende og sent kjønnsmodne vanlig uer (sterkt truet-EN) har viktige yngleområder her (Cadrin mfl. 2010).
I likhet med foreslått SVO Eggakanten nord har dette området flere sårbare naturtyper: Dyphavsjøfjær (Umbellula encrinus), Lophelia-rev, hardbunnskorallskog, og kaldtvanns-svampsamfunn (dominert av glassvamp) (Mareano.no). De til dels unike korallrevene er saktevoksende og har sen regenerering (Sundahl mfl. 2020).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Plante-plankton
Dyreplankton
Fisk
Eggakanten er et stort område som mest sannsynlig vil ha stor variasjon i produksjon av planteplankton. Selve de topografiske forholdene legger til rette for oppstrømning som vil kunne stimulere planteplanktonproduksjon inne på sokkelen (Hanssen-Bauer mfl. 2015).
Dyreplanktonet i området anses som viktig da eggakanten er et viktig gyte- og larvedriftsområde (Sundby mfl. 2003).
Viktig beiteområde for en rekke fiskebestander, inkludert mesopelagiske arter som laksesild og perlemorsfisk (Bergstad mfl. 1999, Bjelland mfl. 2000, Sundby mfl. 2013, Dalpadado mfl. 1998, Salvanes 2004).
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Dyreplankton
Fisk Bunnsamfunn Sjøfugl
En rekke arter og grupper av plankton har Eggakanten inkludert i sine utbredelsesområder (Torgersen mfl. 1997, Dalpadado mfl. 1998), og arter som har sokkelen med kystvann eller oseaniske områder med atlantisk vann som sine habitater, vil kunne «møtes» i området ved Eggakanten. Det foreligger derimot for lite informasjon til å konkludere angående mangfold av dyreplankton.
Tilgangen av plankton og fisk i ulike livsstadier og størrelser gjør området viktig for et større mangfold, der sjøfugl og sjøpattedyr finner mat. Relativt viktig område for fiskelarver og yngel (Sætre mfl. 2002, Sundby mfl. 2013).
Antallet kjente korallrev er høyere i foreslått SVO Eggakanten sør enn i foreslått SVO Eggakanten nord. Særlig er konsentrasjonen av korallrev stor langs Storegga (Mareano.no). Korallverneområdet Storneset inneholder den eneste sikre observasjonen av korallrev dominert av Madrepora oculata (Sundahl mfl. 2020).
Eggakanten er et svært viktig beiteområde for pelagisk beitende sjøfugl, særlig i hekketida (se f.eks. Christensen-Dalsgaard mfl. 2018).
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
Bunnsamfunn
Eggakanten sør har i de dypeste delene tilsynelatende helt urørt bunn (Mareano.no).
9.7.3 - Lokalisering
Avgrensing av SVO Eggakanten ble gjort i Meld. St. 20 (2019-2020) og er ikke endret i forhold til dette. Den foreslåtte endringen er imidlertid å dele inn i SVO-er som blir liggende i hvert sitt forvaltningsområde, forslag til SVO Eggakanten sør som beskrives her, og foreslått SVO Eggakanten nord som beskrives under Barentshavet. Den foreslåtte SVO Eggakanten strekker seg fra Stad til nord av Lofoten, og stopper sør for Lofoten ved grensen for forvaltningsplanområdet for Norskehavet. Området omfatter hele sokkelskråningen og et stykke inn på sokkelen. Det er ulik bredde på området, avhengig av hvor bratt skråningen er (figur 9.7.1).
Området overlapper delvis med eksisterende SVO Mørebankene, som fagekspertgruppen foreslår utvides med tilstøtende eksisterende SVO-er og Kandidatområder og skifter navn til SVO Kystsonen Norskehavet sør.
9.7.4 - Introduksjon til området
Foreslått SVO Eggakanten sør har en sokkelskråning med 200-1000 meter dyp med skråninger videre nedover til 3000 m dyp. Denne skråningen er assosiert med en sterk, ensrettet og smal atlanterhavsstrøm som går langs skråningen (Orvik og Niiler 2002) med strømhastigheter 30-100 cm per sek (Anderson mfl. 2011) og relativt høye sjøtemperaturer, 8 °C (Mork og Skagseth 2010). Det er ulik bredde på området, avhengig av hvor bratt skråningen er. Atlanterhavsstrømmen bringer med seg næringssalter, plankton og fiskeyngel, slik at forrslått SVO Eggakanten sør, i likhet med foreslått SVO Eggakanten nord, er et viktig transport- og gjennomstrømningsområde for marine organismer på vei til Barentshavet og havområdene vest og nord av Svalbard (Rey 2012, Dalpadado mfl. 2012, Sundby mfl. 2013). Sjøtemperaturene i foreslått SVO Eggakanten sør varierer i stor grad med temperaturene i atlanterhavsstrømmen. Ved Stad er temperaturen i kjernen av det atlantiske vannet, ca. 100-200 m dyp, 8 °C. Temperaturen avtar nordover som følge av varmetap til atmosfæren og blanding med tilstøtende vannmasser, og vest av Lofoten har den avtatt til ca. 7 °C (Mork og Blindheim 2003). Sesongvariasjonen i 100-200 m dyp er ca. 1 °C, men i overflaten varierer temperaturen derimot med flere grader på grunn av soloppvarming i et 50 m tynt overflatelag om sommeren (Mork og Skagseth 2010). Under det atlantiske laget, i det intermediære laget og dyphavet, er derimot temperaturen mye lavere, og på 600 m dyp og dypere er temperaturen under 0 °C (Mork og Skagseth 2010). Temperaturen i det atlantiske vannet har hatt en hovedsakelig positiv trend gjennom de siste 40 årene (Skagseth og Mork 2012, González- Pola mfl. 2019).
Figur 9.7.2. Kart over gjennomsnittlig overflatestrøm (piler) og bunndyp (fargekoder) i foreslått SVO Eggakanten sør og omkringliggende områder.
pH i overflatevannet er i underkant av 8,1 over hele området (Fransner mfl. 2020, Lauvset mfl. 2016, Jones mfl. 2020). I 2019 ble verdier mellom 8,04 og 8,02 målt på stasjoner over eggakanten på Svinøysnittet. I den sørligste delen av foreslått SVO Eggakanten har det blitt gjort pH målinger siden midten av 1990-tallet. Disse viser signifikante trender på rundt -2,7 milli-pH per år i overflaten (Fransner mfl. 2020). Trenden avtar nedover i vannsøylen, til rundt 0,8 milli-pH mellom 1000 og 2000 m. Under dette er ikke trenden i pH signifikant (Fransner mfl. 2020). Metningsgraden av aragonitt og kalsitt minker også, rundt -0,012 og -0,02 per år i overflatevannet. Dagens verdier ligger rundt 2,3 til 2,1 for aragonitt og 3,5 til 3,3 for kalsitt, og er avtagende nordover. Vannet under 2000 m er undermettet med hensyn på aragonitt.
9.7.5 - Beskrivelse av miljøverdiene
Plankton
Eggakanten har topografiske forhold som legger til rette for oppstrømning som vil kunne stimulere planteplanktonproduksjon inne på sokkelen (Hanssen-Bauer mfl. 2015) .
Storskala overvåkningsdata viser at det er relativ lav dyreplanktonbiomasse både om våren (mai) og sensommeren (juli/august) langs Eggakanten (ICES 2019, 2021). Sammenliknet med hele Norskehavet og tilgrensende havområder, er det registrert lave verdier av både krill og amfipoder langs Eggakanten (Dalpadado mfl. 1998), men på sokkelen generelt er det stedvis høye konsentrasjoner av krill (Melle mfl. 2004). Meganyctiphanes norvegica (storkrill) finnes det mest av i varmt atlantisk- og kystvann. Det er lite Thysanoessa longicaudata (småkrill) i nordlige deler av Eggakanten, men høyere konsentrasjoner i sørlige deler av området. Undersøkelser tyder ikke på at Eggakanten har høyere konsentrasjon av Calanus spp. sammenliknet med andre områder (Melle mfl. 2004, Broms mfl. 2009, Strand mfl. 2020). Langs Svinøytransektet, som krysser Eggakanten ved Mørebankene, er det registrert 50-80 prosent nedgang i C. finmarchicus bestanden fra 1995-2011. Det er registrert lav forekomst av varmekjære arter langs kysten (Miljostatus.no).
Fisk
Tidlige livsstadier av norsk vårgytende sild (Clupea harengus) og torsk (Gadus morhua) driver nordover langs Eggakanten (Sætre mfl. 2002, Sundby mfl. 2013) . Området er også et viktig gyteområde for hyse (Melanogrammus aeglefinus) i mars til juni (Sundby mfl. 2013). Blåkveite (Reinhardtius hippoglossoides) gyter i løpet av høst og vinter (Sundby mfl. 2013). Vassild har også viktige gyteområder her (Cadrin mfl. 2010). Andre dypvannsarter som vanlig uer (Sebastes norvegicus; sterkt truet-EN) og snabeluer (Sebastes mentella) føder levende yngel i april-mai. Snabelueren er utbredt langs kontinentalskråningen mot Norskehavet fra 400 til 600 meters dyp fra Shetland og nordover til Nord-Spitsbergen (Cadrin mfl. 2010). Det har vært en gradvis økning i den fiskbare delen av blåkveitebestanden fram til 2014, etter det har bestanden stabilisert seg (ICES 2020 a, 2021). I tillegg er eggakanten viktig beiteområde for en rekke fiskearter, både de ovennevnte og andre som kolmule, sei, hyse mfl. (Sundby mfl. 2013) .
Mesopelagisk fauna
Eggakanten og områdene omkring har større mengde av de mesopelagiske fiskene nordlig lysprikkfisk (Benthosema glaciale), laksesild (Maurolicus muelleri), og liten laksetobis (Arctozenus risso) sammenliknet med mer vestlige deler av Norskehavet (Dalpadado mfl. 1998, Salvanes 2004). Dette er trolig et viktigere gyteområde enn ute i dyphavet (Wienerroither, pers medd.).
Virvelløse dyr, som semipelagiske krepsdyr (krill, hoppekreps, og amfipoder) samt blekksprut og geleplankton (maneter), utgjør den største mesopelagiske biomassen, med 70 prosent, der fisken kun utgjør 10 prosent (Skjoldal mfl. 2004). Dette er viktig næringstilgang for dypdykkende sjøpattedyr, og kanskje særlig spermhval.
Bunnsamfunn
I likhet med SVO Eggakanten nord har dette området flere sårbare naturtyper: Dyphavsjøfjær (Umbellula encrinus), Lophelia-rev, hardbunnskorallskog, og kaldtvanns-svampsamfunn (dominert av glassvamp). Utbredelsen av disse er ikke modellert for SVO Eggakanten sør, men observasjoner gjort i felt er presentert i Mareano sin karttjeneste (Mareano.no). Antallet kjente korallrev er høyere i foreslått SVO Eggakanten sør enn i foreslått SVO Eggakanten nord. Særlig er konsentrasjonen av korallrev stor langs Storegga. Revene i dette området har vært eksponert for en høy fiskeri-intensitet, og det er her observert områder hvor skadene påført fra bunntråling er totale (Fosså mfl. 2002). To områder er vernet mot fiske med aktive bunnredskaper: Storneset og Aktivneset. Særlig for Storneset er forekomst av korallrev dominert av steinkorallen Madrepora oculata. Dette er den eneste sikre observasjonen av Madrepora-rev i Norge (Sundahl mfl. 2020).
Ut fra innsamlede data og modellert resultat, er det dokumentert 832 korallrev, og modellert i overkant av 200 000 korallrev i kartutsnittet (Mareano.no) (figur 9.7.3).
Figur 9.7.3. Kartet viser dokumenterte og modellerte korallrev for de foreslåtte SVO-ene, Eggakanten sør og Kystsonen Norskehavet nord den nordlige delen av foreslått SVO Kystsonen Norskehavet sør (Mareano.no) .
Sjøpattedyr
Eggakanten og den øvre sokkelskråningen er viktig beiteområde for klappmyss (Cystophora cristata) (Folkow og Blix 1999) og spermhval (Physeter macrocephalus) . Blåkveite og andre meso/batypelagiske arter er trolig viktige byttedyr for klappmyss langs E ggakanten og sokkelskråningen (Folkow og Blix 1999). Bestanden av klappmyss i Nordøst-Atlanteren er beregnet til 76 623 dyr (95 % konfidensintervall: 58 299-94 947) i 2019 (Haug og Biuw 2019).
Sjøfugl
Eggakanten er beiteområde for sjøfugl langs hele kysten. Dette er behandlet under de respektivt foreslåtte SVO-ene Kystsonen Norskehavet nord og Kystsonen Norskehavet sør. Krykkjer (Rissa tridactyla) som hekker på Helgelandskysten, er påvist å beite aktivt på Eggakanten i hekketiden (Christensen-Dalsgaard mfl. 2018). Sjøfuglene beiter på fiskelarver som driver med kyststrømmen (f.eks. Sandvik mfl. 2016).
Hornkoraller. Foto: Mareano / HI
9.8 - Forslag til endret SVO Kystsonen Norskehavet nord (NH6)
9.8.1 - Sammendrag
Områdebeskrivelse og strukturer
Det foreslått endrete SVO Kystsonen Norskehavet nord (NH6) har utpregete retensjonsområder med relativt lang residenstid over forholdsvis store grunne områder, dannet på grunn av mange sund mellom øyer og skjær, der noen er langt utenfor fastlandet og ut mot Eggakanten.
Miljøverdier
Samlet sett har området et høyt mangfold av habitater og arter, ikke minst i tareskogene. Området dekker særlig viktige gytefelt for en rekke fiskearter, ikke minst i mars–april for kommersielt store og viktige arter som nordøstarktisk torsk og norsk vårgytende sild, men også øyepål og vanlig uer (sterkt truet-EN). Gytingen pågår senvinter og vår, med påfølgende larvedrift nordover utover forsommeren. Retensjonsområder har opphopning av plankton og dermed høyere biomasse som beites av fiskelarver og -yngel. Det er også et viktig beiteområde for brugde (sterkt truet-EN) og håbrann (sårbar-VU).
Området er viktig hekke- og beiteområde for sjøfugl, særlig for kystbundne arter som ærfugl, toppskarv, storskarv, teist, svartbak og sildemåke, men også noen pelagiske arter, for eksempel krykkje og tyvjo. Flere er rødlistet. Gruntvannsområdene er et viktig myteområde for ærfugl og er i tillegg viktige som overvintringsområder for blant annet ærfugl, teist, skarver, stormåker, lommer og dykkere. I dette SVO-et finnes majoriteten av den norske bestanden av nordlig sildemåke som har vært i tilbakegang en lengre periode. Sklinna huser også en av verdens største kolonier av toppskarv.
Det foreslåtte SVO-et har et stort antall korallrev og sårbare naturtyper, hvorav de aller fleste er intakte uten tegn til påvirkning av bunnfiske. Korallrevene forekommer både kystnært og ute på kontinentalsokkelen, men er særlig konsentrert til Sularevet og Iverryggen.
Vanlig uer i korallskog. Foto: Mareano / HI
Figur 9.8.1. Kart over foreslått endret SVO Kystsonen Norskehavet nord (NH6). Grensene for det foreslåtte SVO-et er vist med tykk rød strek, mens grenser for overlappende og inntilliggende forslag til SVO-er er vist med tynn rød strek. Grensene for forvaltningsplanområdet er markert med brun linje. Kartet viser utbredelsen av utvalgte miljøverdier av betydning for det foreslåtte SVO-et. Miljøverdier knyttet til gytefelt for fisk/ynglefelt for vanlig uer, samt beiteområder for hekkende, pelagisk beitende sjøfugl er markert i venstre kart og miljøverdier knyttet til bunnsamfunn og sjøpattedyr er markert i høyre kart.
Lomvi. Foto: Mari Skuffedal Myksvoll, HI
9.8.2 - Vurdering i forhold til EBSA-kriteriene – Forslag til endret SVO Kystsonen Norskehavet nord (NH6)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til endret SVO Kystsonen Norskehavet nord (NH6)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet/sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Bunnsamfunn
Sjøfugl
Innen det foreslåtte SVO-et er det registrert et stort antall korallrev og sårbare naturtyper. Indikatorarter for svampskog (hovedsakelig Axinellide svamper) er hyppig i dette området, og hardbunnskorallskog (Paragorgia, Primnoa og Paramuricea) er ikke uvanlig. Korallrevene forekommer både kystnært og ute på kontinentalsokkelen, men er særlig konsentrert til to områder på sokkelen: Sularevet og Iverryggen (Mortensen mfl. 2001, Freiwald 2002, Fosså mfl. 2015).
I dette området finnes majoriteten av den norske bestanden av nordlig sildemåke Larus f. fuscus (seapop.no).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Fisk
Sjøpattedyr Sjøfugl
Tidlige livsstadier av norsk vårgytende sild, torsk, vanlig uer og snabeluer og andre arter driver nordover langs norskekysten (Sætre mfl. 2002, Sundby mfl. 2013). Viktig beiteområde for brugde (sterkt truet-EN) og håbrann (sårbar-VU) (Sims 2008)
Området har vært landets viktigste ynglehabitat for havert (Sørlie mfl. 2019, Nilssen mfl. 2019) og kasteplasser for steinkobber (Arneberg og Jelmert 2017). (Arneberg og Jelmert 2017).
I dette området finnes majoriteten av den norske bestanden av nordlig sildemåke Larus f. fuscus (seapop.no), som har vært i tilbakegang en lengre periode (Fauchald mfl. 2015). Lomvibestanden (sårbar-VU) på Sklinna er en av få kolonier som øker i Norge (seapop.no), og området er et av det viktigste for ærfugl i Norge gjennom hele året, både som hekke-, myte og overvintringsområde. Storskarv, toppskarv og teist har også viktige leveområder her både i hekkesesongen og gjennom vinteren (seapop.no).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Fisk Bunnsamfunn
Sjøpattedyr
Sjøfugl
Viktig beiteområde for brugde (sterkt truet-EN) og håbrann (sårbar-VU) (Sims 2008).
Innen det foreslåtte SVO-et er det registrert et stort antall korallrev og sårbare naturtyper (Mortensen mfl. 2001, Freiwald 2002, Fosså mfl. 2015).
Området har vært viktige kasteplasser for steinkobber, og selv om totalbestanden av steinkobbe i Norge har vært økende de siste årene, har bestanden gått noe tilbake i Trøndelag og Froan (Arneberg og Jelmert 2017).
Lomvibestanden (sårbar-VU) på Sklinna er en av få kolonier som øker i Norge (seapop.no), området er et av det viktigste for ærfugl (nær truet-NT) i Norge og hovedutbredelse for nordlig sildemåke (Larus f. fuscus) i Norge (Fauchald mfl. 2015).
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Fisk
Bunnsamfunn Sjøfugl
Brugde og håbrann er særlig sårbare arter pga sen modning, få avkom og langsom vekst; fare for bifangst (Sims 2008).
Innen SVO-et er det registrert et stort antall korallrev og sårbare naturtyper (Mortensen mfl. 2001, Freiwald 2002, Fosså mfl. 2015).
For sjøfugl er sokkelområdet og Eggakanten svært viktige beiteområder gjennom hele året (f.eks. Christensen-Dalsgaard mfl. 2018), og mange av bestandene er i tilbakegang (Fauchald mfl. 2015).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Dyreplankton
Fisk Bunnsamfunn
Det er ikke registrert høyere dyreplanktonproduksjon i området basert på Havforskningsinstituttets storskala overvåkningsdata i perioden mai og juli/august (ICES 2020 a og b, 2021), men det vil kunne være store lokale forskjeller der retensjonsområder har opphopning av dyreplankton og dermed høyere biomasse. Dyreplanktonet i området er viktig for fiskelarver og -yngel.
Området er viktig gyteområde for norsk vårgytende sild i februar og mars (Dragesund mfl. 1980). Her finnes det også gytefelt for torsk, vanlig uer og øyepål (Husa mfl. 2020). Det foreslåtte SVO-et er også et viktig fiskeegg/larver/yngel område (sild, torsk, vanlig uer og øyepål) (Sætre mfl. 2002, Sundby mfl. 2013).
Stortareskogen (Laminaria hyperborea) bidrar til høy produksjon av både biomasse (Abdullah mfl. 2004), tryggere oppvekst av tidlige livsstadier av fisk og en rekke andre arter alger og virvelløse dyr (Marstein 1997, Andersen 2007, Norderhaug 2012, Christensen-Dalsgaard mfl. 2020).
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Fisk
Bunnsamfunn
Området inneholder store mengder av egg-, larve- og yngel av mange fiskearter, som er viktig mat for mange sjøfugl -, fiske- og sjøpattedyrarter og bidrar til biologisk biomangfold i området (Sætre mfl. 2002, Sundby mfl. 2013, Systad mfl. 2019).
Kaldtvannskorallrev, svampsamfunn (Svampspikelbunn og svamphage, samt høyt biomangfold observert med video danner komplekse leveområder for fisk og bunndyr (Mortensen mfl. 2001, Freiwald 2002, Fosså mfl. 2015, Mareano.no), Stortareskogen (Laminaria hyperborea) bidrar til høy produksjon av både biomasse (Abdullah mfl. 2004), tryggere oppvekst av tidlige livsstadier av fisk og en rekke andre arter alger og virvelløse dyr (Marstein 1997, Andersen 2007, Norderhaug 2012, Christensen-Dalsgaard mfl. 2020, Norderhaug mfl. 2020).
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
Bunnsamfunn
Innen det foreslåtte SVO-et er det registrert et stort antall korallrev og sårbare naturtyper (Mortensen mfl. 2001, Freiwald 2002, Fosså mfl. 2015). De aller fleste er intakte uten tegn til påvirkning av bunnfiske.
9.8.3 - Lokalisering
Forslag til endret SVO Kystsonen Norskehavet nord er et større SVO som dekker flere eksisterende, mindre SVO-er sist oppdatert i Meld. St. 20 (2019-2020), dvs. SVO Iverryggen, SVO Haltenbanken, SVO Sklinnabanken, SVO Froan med Sularevet, SVO Remman samt den midtre delen av SVO Kystsonen Norskehavet (tabell 9.1). Det er ikke naturlige skiller til stede som i stor nok grad forsvarer å opprettholde disse områdene som separate SVO-er. Forslag til endret SVO Kystsonen Norskehavet nord inkluderer også kandidatområdene for sjøfugl i områdene fra Vikna til og med Vega samt ut til og med Sklinnabanken, og området er i tillegg utvidet inn mot kysten for å inkludere beiteområder for sjøfugl og kystnære gyteområder for fisk.
Forslag til endret SVO Kystsonen Norskehavet nord ligger utenfor kysten av Møre og Romsdal, Trøndelag og Nordland. Området strekker seg fra renna mellom Grip og Smøla i sør og grenser opp mot Vestfjorden i nord (Fig. 9.8.1). Området grenser til det foreslått endrete SVO Kystsonen Norskehavet sør.
Forslag til endret SVO Kystsonen Norskehavet nord dekker dermed marine verneområdet Remman, samt flere områder som er foreslått som marine verneområder, Iverryggen, Froan-Sularevet og Borgan-Frelsøy. Verdensarvområdet Vega ligger også i dette området.
9.8.4 - Introduksjon til området
Foreslått endret SVO Kystsonen Norskehavet nord er sammensatt av mange ulike økosystem: små, lukkede poller og fjorder med grunne terskler inne på sokkelen utenfor Trøndelag og Helgeland, nærhet til store fjorder, som Trondheimsfjorden og Ransfjorden og åpen, eksponert kyst ut mot Eggakanten. En variert topografi gir rom for mange ulike naturtyper og leveområder, fra grunne til dype områder og fra meget beskyttede til sterkt eksponerte områder. Området innehar også flere banker som Haltenbanken og Sklinnabanken der topografiske strømvirvler genereres slik at vannmassene oppholder seg der over lengre tid (Sætre mfl. 2002) (Fig. 9.8.2). Slike retensjonsområder med relativt lang residenstid fungerer som midlertidige samlingsplasser for drivende egg, larver og yngel. Det fysiske og kjemiske miljøet i SVO Kystsonen Norskehavet nord er hovedsakelig påvirket av kyststrømmen og vannmasser i skjærgård og fjorder, bestemt av topografiske forhold som terskler og bassengdyp. Kjennetegn på kyststrømmen er først og fremst relativ lav saltholdighet, ~34 g per kg. Typiske hastigheter i kyststrømmen er 20-50 cm per sek, men den kan bli over 100 cm per sek og vil variere med endringer i tetthet og vind (Sætre 2007, Skagseth mfl. 2011). Kyststrømmen er viktig for transport nordover av fiskeegg og larver (Vikebø mfl. 2005, Skagseth mfl. 2015). Det er observert økt temperatur i kystsonen (~0,8°C) over de siste ~40 år, blant annet som følge av innblanding av varmere atlanterhavsvann (Albretsen mfl. 2012, González- Pola mfl. 2019). Under kystvannet er området mer påvirket av det saltere atlanterhavsvannet som blir advektert inn på sokkelen.
De ytre delene av området har pH-verdier rundt 8,05-8,1 (Lauvset mfl. 2016, Fransner mfl. 2020, Jones mfl. 2020). Det er lite data fra kystnære strøk, men som ellers, forventer vi lavere verdier mot kysten. Metningsgraden for aragonitt (ΩAr) er i underkant av to, mens den er litt over tre for kalsitt (ΩCa) (Lauvset mfl. 2016, Fransner mfl. 2020, Jones mfl. 2020), basert på sommerdata. Hele vannsøylen er overmettet med hensyn til både kalsitt og aragonitt, siden området er ganske grunt.
Figur 9.8.2. Kart over gjennomsnittlig overflatestrøm (piler) og bunndyp (fargekode) i foreslått SVO Kystsonen Norskehavet nord og omkringliggende områder.
9.8.5 - Beskrivelse av miljøverdiene
Plankton
Generelt vil det være høyere produksjon av planteplankton langs kysten pga økt tilførsel av næringssalter. Det finnes lite informasjon omkring planteplanktonproduksjon og biologisk mangfold i foreslått endret SVO Kystsonen Norskehavet nord. Topografiske forhold sikrer oppstrømning av næringssalter som stimulerer stabil og høy planteplanktonproduksjon gjennom sesongen. Det er dessuten en rekke ikheter med forslag til endrete SVO-er Kystsonen Lofoten er det mange likheter med forhold omtalt i Tromsøflaket og SVO Eggakanten nord. Ved Skrova pågår det fast overvåkning av blant annet planteplankton, som viser den generelle sesongmessige variasjonen i området, som dekker både den sørlige delen av foreslått SVO-er Området rundt Lofoten ogkystsonen Norskehavet nord (Naustvoll mfl. 2020).
Forslag til endret SVO Kystsonen Norskehavet nord er sammensatt av mange ulike økosystem, og dyreplanktonproduksjonen og artssammensetningen vil dermed variere i ulike deler av området. Generelt sett er det ikke høyere biomasse i dette området sammenliknet med andre områder i Norskehavet, basert på storskala overvåkningstidsserier fra mai og juli/august (ICES 2019, 2020 a, 2021). Det kan likevel være lokale forskjeller der retensjonsområder har opphopning av plankton og dermed høyere biomasse. Dyreplanktonbiomassen ved og oppstrøms for de grunne bankene Sklinnabanken (6 -16 g tørrvekt per m2) og i området ved Iverryggen (ca 8-12 g tørrvekt per m2) i mai er noe høyere sammenlignet med omkringliggende områder. Høyere verdier har ikke blitt observert i juli/august. Eksempelvis har områdene Remman, Froan og Haltenbanken noe lavere biomasse sammenliknet med ovennevnte områder, og forekomstene er på mellom 2 og 10 g tørrvekt per m2 i disse tidsperiodene.
Generelt langs kysten, inkludert det foreslåtte SVO Kystsonen Norskehavet nord, utgjør Calanus finmarchicus (raudåte) hovedmengden av biomassen. Denne arten, samt andre kopepodearter, er særdeles viktige som mat for larver og yngel til en rekke fiskearter som gyter i området eller har avkom som driver gjennom området (Sundby mfl. 2013). Produksjonen av C. finmarchicus i kystområdet starter i mars, men antallet holder seg lavt frem til april/mai da det er en stor økning av yngre stadier (Broms og Melle 2007, Bagøien mfl. 2012). Mai er også tidspunktet hvor konsentrasjonen av voksne individer er størst. I juni finnes et stort antall individer av både yngre og eldre stadier av den nye generasjonen. Konsentrasjonen avtar utover i juli, men i august fører produksjonen av enda ny(e) generasjon(er) til en ny økning (Broms og Melle 2007, Bagøien mfl. 2012, Strand mfl. 2020). C. finmarchicus har generelt én generasjon i året, men kan ha to eller flere generasjoner i kystvannet, som er relativt varmt, og produksjonssesongen blir dermed forlenget (Broms og Melle 2007, Bagøien mfl. 2012). Planktonets sesongutvikling er tidligere i kystvannet (2-6 uker) sammenliknet med atlantiske og arktiske vannmasser i Norskehavet (Broms og Melle 2007, Bagøien mfl. 2012).
Da forslag til endret SVO Kystsonen Norskehavet nord ligger nedstrøms for forslag til endret SVO Kystsonen Norskehavet sør, kan en anta at området har hatt tilsvarende endringer i raudåtebestanden, som vil tilsi en nedgang på mellom 50 prosent (i atlanterhavsvann vest for området) og 80 prosent (i kystvann) i perioden 1995-2011 (Dupont mfl. 2017, se forslag til endret SVO Kystsonen Norskehavet sør, Kap. 9.9). Det er ikke gjort analyser for perioden etter 2011, men generelt i Norskehavet har den totale mengden dyreplankton vært på et stabilt og antakelig noe økende nivå etter 2011. Det foreligger også opparbeidede prøver fra Svinøysnittet sør for foreslått endret SVO Kystsonen Norskehavet nord som inkluderer andre dyreplanktonarter, men disse dataene er ikke analysert for langtidstrender. Det foreligger lite informasjon om artsdiversitet, men enkeltundersøkelser fra Froan har registrert høyere antall kopepoder på Mausundbanken i Froan sammenliknet med områder utenfor banken (Fragoso mfl. 2019). En rekke arter av amfipoder, isopoder og tifotkreps som lever på tareskogen er registrert i Froan (Skadeheim og Rinde 1995), men undersøkelsen inkluderte ikke frittsvømmende plankton.
Fisk
Sild (Clupea harengus) gyter på bunn og er avhengig blant annet av riktig bunnsubstrat for vellykket gyting og vil således gyte langs store deler av norskekysten (Runnstrøm 1941). Gyteområdene for norsk vårgytende sild varierer over tid, men etter Mørebankene finner vi i dette SVO-forslaget noen av de viktigste gyteområdene for norsk vårgytende sild (Dragesund mfl. 1980, Sætre mfl. 2002, Sundby mfl. 2013). Sild vandrer inn fra overvintringsområder i havet og i norske fjorder, og gyter på Haltenbanken og Sklinnabanken i perioden februar/mars. Etter gyting vandrer silda vestover på beitevandring og vil ikke i særlig grad finnes i området utenom gyteperioden (Holst mfl. 2004). En gytebestand på 10 millioner tonn legger ca. 2 millioner tonn gyteprodukter hvert år, men mengden kan svinge kraftig. Beregnet gytebestand i 2020 var 3,5 millioner tonn (ICES 2020 b, 2021). Denne produksjonen av gyteprodukter som silda bidrar med, er en viktig næringskilde for fisk og andre organismer som beiter på eggene og larvene vår og sommer (Toresen 1991, Høines og Bergstad 1999, Varpe mfl. 2005). Sildelarvene vil etter klekking drive nordover med kyststrømmen langs kysten, og videre inn i oppvekstområdene i Barentshavet tidlig på sommeren. Larvene vil ofte ha en viss oppholdstid på gytebankene på grunn av strømforhold (Sætre mfl. 2002). Forslag til SVO Kystsonen Norskehavet nord er derfor også et viktig beiteområde for sildelarvene den første tiden etter klekking. Sild har stor betydning for økosystemene langs kysten opp til Barentshavet. Den beiter på raudåte og er selv en viktig matressurs for sjøfugl, rovfisk som torsk, sei og annen bunnfisk i tillegg til hval. Store flokker av spekkhoggere følger silda på dens vandringer.
Vi har tre arter av uer i Norge, vanlig uer (Sebastes norvegicus), snabeluer (Sebastes mentella) og lusuer (Sebastes viviparus). Vanlig uer er vurdert som sterkt truet (EN) på grunn av en sterk nedgang i bestanden langs norskekysten (Husa mfl. 2020). Bestanden er på et historisk lavmål og regnes for å ha redusert rekrutteringskapasitet på grunn av sterkt redusert gytebestand. Nord for 62°N er det forbudt å fiske uer, med unntak av juksafiske fra båt under 15 m lengde i perioden 1. juni-31. august.
Vanlig uer er langtlevende fisk og kan bli opptil 60 år gammel. Arten lever på 100-500 meters dyp på kontinentalsokkelen, langs kysten og noen steder inne i fjordene i dette SVO-et. Uer parrer seg om høsten og føder levende yngel i mars-juni. Den vokser langsomt og blir sent kjønnsmoden (11-15 år). SVO Kystsonen Norskehavet nord er et viktig yngleområde som strekker seg langs Eggakanten og kontinentalsokkelen fra Shetland og nordover til Andøya, med Storegga, Haltenbanken og Vesterålen som de viktigste områdene (Husa mfl. 2020).
Isgalt (Macrourusberglax) gyter hovedsakelig på Røstbanken og Trænabanken innenfor forslaget til SVO Kystsonen Norskehavet nord (Savvatimsky 1985), mens det er noe gyteaktivitet nordover i forslag til endret SVO Tromsøflaket (Savvatimsky 1985). Gytingen foregår fra midt-vinter til tidlig vår med høydepunkt i desember-januar (Eliassen 1983, Savvatimsky 1985).
Øyepål (Trisopterus esmarkii) er en ganske liten fisk i torskefamilien med en levetid som sjelden går over tre år. Øyepål opptrer i store stimer, som regel over mudderbunn. Den spiser hovedsakelig planktoniske krepsdyr, som krill og raudåte (Sundby mfl. 2013). Øyepål utgjør igjen en del av næringsgrunnlaget for andre større fisk som torsk, hvitting og sei, men også sjøpattedyr. Siden 1995 har fiskedødeligheten vært på et lavere nivå enn tidligere. Utbredelsen strekker seg også nordover norskekysten og inn i det sørlige Barentshavet. Hovedgytefeltene er spredt ut i den nordlige delen av Nordsjøen. De nordlige gytefeltene som har relevans for egg-, larve- og yngeldrift i forslag til endrete SVOene Kystsonen Norskehavet nord og Kystsonen Norskehavet sør omfatter kysten av Møre, Haltenbanken, Vestfjorden, rundt Røst og utover langs vestligste delen av kontinentalsokkelen utenfor Vesterålen (Sundby mfl. 2013). Bestandsstørrelsen varierer mye fra år til år på grunn av variabel rekruttering og et kort livsløp. I 2018 er gytebestanden beregnet til å være over anbefalt grense for biomasse (Bpa).
Brugde (Cetorhinus maximus) og håbrann (Lamna nasus) er særlig sårbare arter pga sen modning, få avkom og langsom vekst, og er listet som henholdsvis truet og sårbar på norsk rødliste. Områder langs kysten i overgangen mellom Møre og Romsdal og Trøndelag er identifisert som viktige områder for begge arter, særlig om sommeren for brugde (observasjonsdata fra https://dugnadforhavet.no og Junge mfl. in prep) og litt senere for håbrann (omtrent sep/okt). Brugde forekommer langs habitater på kontinentalsokkelen der fronter ofte er til stede for å oppkonsentrere plankton (f.eks. Sims 2008). Brugde spiser dyreplankton, med kalanoide kopepoder som den viktigste byttedyrgruppen (som Calanus arter). For håbrann er dette området identifisert som beiteområde pga sild (González Triginer 2020, masteroppgave).
Bunnsamfunn
Innen det foreslåtte SVO-et er det registrert et stort antall korallrev (Fig. 9.8.3) og andre sårbare naturtyper som svampskog (Fig. 9.8.4). Det er dokumentert 832 korallrev, og modellert i overkant av 200 000 korallrev i kartutsnittet som er vist i Fig. 9.7.3 (Mareano.no). Indikatorarter for svampskog (hovedsakelig Axinellide svamper) er hyppig i dette området, og hardbunnskorallskog (Paragorgia, Primnord og Paramuricea) er ikke uvanlig. Korallrevene forekommer både kystnært og ute på kontinentalsokkelen, men er særlig konsentrert til to områder på sokkelen: Sularevet og Iverryggen (Fig. 9.8.1). Sularevet er et revområde med rundt 500 individuelle korallrev som forekommer langs en rundt 20 km lang rygg. Disse revene er store (opp til ca 30 m i høyde, 500 m i lengde), gamle (opp mot 9500 år), og i stor grad intakte (bortsett fra på Iverryggen) (Mortensen mfl. 2001, Freiwald 2002, Fosså mfl. 2015). Både Sularevet og Iverryggen er vernet mot fiske med aktive bunnredskaper.
Figur 9.8.3. Kartet viser dokumenterte korallrev i foreslått endret SVO Kystsonen Norskehavet nord, sentrale deler av SVO Eggakanten sør og nordlig del av SVO Kystsonen Norskehavet sør (Mareano.no)
Figur 9.8.4 Mareano-observasjoner (fra video) av svampskog (svampsamfunn på hardbunn) i tillegg til hardbunnskorallskog i de foreslåtte SVOene Kystsonene Norskehavet sør og nord, samt foreslått endret SVO Eggakanten sør (Kart fra Mareano.no).
Sjøkrepsbestandene nordover langs norskekysten er lite kjent. Sjøkreps (Nephrops norvegicus) finnes ofte på samme type bunn som dypvannsreke, slik at Fiskeridirektoratets kartlagte rekefelt (Fiskeri (fiskeridir.no)) gir en pekepinn på utbredelsen av sjøkreps. Norskekysten utgjør den nordligste delen av artens utbredelsesområde (Sjøkreps | Havforskningsinstituttet (hi.no)). Sammenlignet med Nordsjøen (ICES 2020 c, 2021) skiller norskekysten seg ut med særdeles store individer. Et kystnært teinefiske etter sjøkreps, både kommersielt og fritidsfiske, har vokst frem de siste årene (Søvik mfl. 2017). Det er per dags dato vanskelig å verdisette de enkelte sjøkrepsfeltene langs norskekysten, og vi kjenner heller ikke bestandsstatus til denne arten langs kysten.
Heller ikke bestandsstatus til dypvannsreken (Pandalus borealis) langs store deler av norskekysten fra Vestland til Vestfjorden, inkludert SVO Kystsonen Norskehavet nord (Trøndelag/Helgelandskysten) er kjent. Fiskeridirektoratets kartlagte rekefelt viser at rekene finnes på en mosaikk av store og små rekefelt langs hele kysten. Rekene finnes til dels også utenfor disse kommersielle feltene. Temperatur, substrat, saltholdighet og dyp bidrar til å bestemme utbredelse og tetthet (Shumway mfl. 1985, Zimmermann mfl. 2019). Dypvannsreken fra Skagerrak og nord til Kvænangen utgjør én genetisk bestand (Jorde mfl. 2015, Hansen 2020), men reker i indre deler av fjordene kan muligens utgjøre egne genetiske populasjoner (Knutsen mfl. 2015).
Stortareskogen (Lamiaria hyperborea) som opptrer i Norskehavets SVO kystsoner er viktige biotoper for hele økosystem (Marstein 1997, Christie mfl. 2009, Norderhaug mfl. 2020), for produktiviteten i økosystemet, som tryggere leveområde for tidlige livsstadier (Christensen-Dalsgaard mfl. 2020), helårsleveområder for en rekke virvelløse dyr og ved å tilføre området næringssalter når gamle algedeler brytes ned (Andresen 2007). Jo mer eksponert tareskogen er mot havet, jo høyere artsmangfold er det som påvekst og assositerte arter (Norderhaug mfl. 2012). Samlet bidrar tareskogen med økt produktivitet i SVO-et (Abdullah og Fredriksen 2004).
Sjøpattedyr
Havert (Halichoerus grypus) gjennomfører hele sin livssyklus ved kysten og lever spredt i SVO-et. Haverten danner kolonier, særlig i forbindelse med ungekasting (fødsel) og parring (september–desember) og hårfelling (februar–april), men lever spredt resten av året (Bjørge og Øien 1999).
Steinkobbene (Phoca vitulina) samles på land spredt gjennom hele det foreslåtte SVO-et i forbindelse med forplantning (juni-juli) og hårfelling (august), og de legger seg helst opp på litt beskyttede lokaliteter i skjærgården (skjær og sandbanker som tørrlegges ved fjære sjø) (Sørlie mfl. 2020, Nilssen mfl. 2019). Yngle- og hårfellingstid er fra juni til begynnelsen av september.
Spekkhoggere (Orcinus orca) trekker inn i området når det er sildegyting her, for å beite på silda (Leonard og Øien 2020, Similä mfl. 1995).
Sjøfugl
Helgelandskysten og området ned til Smøla er et stort og fuglerikt område, tidligere beskrevet under flere SVO-er (Remman, Froan, Haltenbanken, Sklinnabanken) (Systad mfl. 2019). Området er svært heterogent, med store arealer med tareskog og store gruntvannsområder som gir grunnlag for god næringstilgang for mange ulike arter av sjøfugl.
Strekningen Vikna-Vega inkluderer blant annet Sklinna, Horta og Horsvær, samt viktige kystnære marine beiteområder for sjøfugl (Systad mfl. 2019, Christensen-Dahlsgaard 2017). Utløpet av Trondheimsfjorden er viktig som overvintringsområde for havdykkender, og Froan, Frøya, Hitra og Smøla huser store bestander av hekkende sjøfugl, primært kystnære arter (bl.a. Systad mfl. 2019).
Sklinna huser en av verdens største kolonier av toppskarv (Phalacrocorax aristotelis), som i gode år teller over 3000 hekkende par. I tillegg hekker også lomvi (Uria aalge), teist (Cepphus grylle), lunde (Fratercula arctica), ærfugl (Somateria mollissima), gråmåke (Larus argentatus) og svartbak (Larus marinus). Sklinna er dermed den eneste sjøfuglkolonien med et komplett utvalg av de vanlige fuglefjellsartene mellom Runde og Røst, og er således Midt-Norges viktigste sjøfuglkoloni. Siden 2010 har hekkende toppskarv på Sklinna blitt instrumentert med GPS- og dykkeloggere for å registrere hvor de drar for å finne mat (Christensen-Dalsgaard mfl. 2018, 2020). Resultatene viser at toppskarven bruker områder med rike tareforekomster opp til 46 km fra kolonien, slik at Horta og Vikna er viktige funksjonsområder for hekkende toppskarv fra Sklinna. Undersøkelser gjort vha. GPS-instrumentering av teist på Sklinna i 2018, viste at det er tilsvarende habitater som også er viktige for denne arten (SEAPOP, upubliserte resultater).
Undersøkelser gjort på funksjonsområdene til krykkje (Rissa tridactyla) som hekker på Sør-Gjæslingan, sørvest for Vikna, har vist at også området ved Vikna blir brukt til å lete etter mat (Christensen-Dalsgaard mfl. 2018). I tillegg ble det vist at hekkende krykkjer fra Sør-Gjæslingan bruker Eggakanten som beiteområde. Krykkjen flyr altså til områder opptil 300 km fra kolonien for å finne mat, hvilket underbygger tidligere vurderinger av Eggakanten som et svært viktig område for pelagiske sjøfugler.
Den største kjente kolonien av nordlig sildemåke (Larus fuscus) i Norge ligger på Horsvær (Bustnes 2010), og i tillegg hekker denne arten i flere kolonier i området mellom Vikna og Vega. Det finnes også flere hekkelokaliteter for teist (Cepphus grylle), storskarv (Phalacrocorax carbo), rødnebbterne (Sterna paradisaea) og den i Norge sterkt truede (EN) makrellterne (Sterna hirunda) i området (Lorentsen mfl. 2012).
Figur 9.8.6 Kart viser fordeling av pelagiske sjøfuglarter (A: havsule, krykkje, lomvi, lunde, alke), kystnære fiskespisende arter (B: toppskarv, storskarv, teist), kystnære bentisk beitende arter (C: ærfugl) og kystnære overflatebeitende arter (D: gråmåke, sildemåke, tyvjo og E: svartbak) i foreslått endret SVO Kystsonen Norskehavet nord og nærliggende områder. Skravert felt i A) viser beiteområder for pelagisk beitende sjøfugl. Grensene for det foreslåtte SVO-et er vist med tykk rød strek, mens grenser for inntilliggende foreslått endrete SVO-er er vist med tynn rød strek.
Gruntvannsområdene er også identifisert som et viktig myteområde for ærfugl (Christensen-Dalsgaard mfl. 2010) og er i tillegg viktige som overvintringsområder for blant annet ærfugl (Somateria mollissima), teist, skarver, stormåker, lommer og dykkere.
På Sklinna har hekkebestanden av toppskarv (Phalacrocorax aristotelis) vært i sterk vekst siden overvåkingen startet i 1984. Årsaken er delvis at et nytt hekkeområde har blitt tilgjengelig etter at en ny steinmolo ble bygget i 1990. De siste 10 årene ser bestanden på Sklinna ut til å ha stabilisert seg. Etter at krykkje forsvant fra Sklinna begynte SEAPOP å overvåke toppskarv på Sør-Gjæslingan i Vikna i stedet (SEAPOP.no). Også her er arten i sterk tilbakegang. For øvrig har både alke og lomvi en positiv bestandsvekst på Sklinna med hhv sju og seks prosent årlig de siste ti årene. Lunde derimot har hatt en årlig nedgang på 11 prosent i samme periode. Teist har hatt en årlig vekst på fem prosent, sildemåke tre prosent i året og svartbak to prosent i året. Gråmåke har hatt en betydelig nedgang på 15 prosent i året. Også storskarv, toppskarv og ærfugl har hatt en nedgang på hhv 10 prosent, fem prosent og 16 prosent i året i perioden 2007-2017.
9.9 - Forslag til endret SVO Kystsonen Norskehavet sør (NH7)
9.9.1 - Sammendrag
Områdebeskrivelse og strukturer
I forslag til endret SVO Kystsonen Norskehavet sør (NH7) møtes atlanterhavsvann og kyststrømmen og gir særlig næringsrike forhold. Topografien er helt spesiell, med kort distanse fra Eggakanten til fastland i den sørligste delen, og med grunne flater, skjær og øyer i den nordlige delen som gir retensjonsområder for vannet med relativt lang residenstid. Storvokste tareskoger bidrar til strømmønsteret.
Miljøverdier
Mørebankene, som vil vøre inkludert i forslaget til i SVO K ystsonen Norskehavet sør er et kjerneområde for gyting og tidlig oppvekst hos norsk vårgytende sild og sei, men er også viktig for torsk, både kysttorsk og tidligere også skrei, hyse, øyepål og vanlig uer (sterkt truet-NE). Gytingen av kysttorsk i SVO-et foregår både ute på kysten og inne i beskyttede fjordarmer, i relativ nærhet til oppvekstområdene for yngelen. Mørebanken er viktig særlig om sommeren, for brugde (sterkt truet-NE). Det er ikke registrert høyere dyreplanktonproduksjon generelt i området, men det vil kunne være store lokale forskjeller der retensjonsområder har opphopning av plankton og dermed høyere biomasse. Dyreplanktonet i området er viktig for fiskelarver og -yngel.
Foreslått endret SVO Kystsonen Norskehavet sør er et viktig hekke-, fjærfellings-, trekk- og overvintringsområde for sjøfugl. Flere lokaliteter innenfor området har nasjonal verdi som hekkelokalitet og/eller myteområde. Området er et viktig beiteområde for bl.a. havsule, lomvi, lunde og alke og også flere kystnære arter beiter i de grunne områdene av det foreslåtte SVO-et, hvorav ærfugl, toppskarv, gråmåke og sildemåke har gode forekomster. Runde har fuglefjell med det høyeste artsmangfoldet av sjøfugl i Norge i hekketiden, og huser en rekke arter som er sårbare og truede. Runde har også det største lundefjellet sør for Røst og havsulekolonien her utgjør mer enn 50 prosent av den totale norske bestanden. Det foreslåtte SVO-et er også et viktig overvintringsområde for alkekonge og gulnebblom. Langs hele kysten i det foreslåtte SVO-et har steinkobbe kolonier, de største ved Sandøy og Haram i Møre og Romsdal.
Forslag til det endrete SVO-et har flere sårbare naturtyper, blant annet korallrev, hardbunnskorallskoger, sjøfjærsamfunn og svampskog. Svampskog er relativt vanlig i den mest kystnære delen av dette forslaget til SVO. Hardbunnskorallskog er vanligere i fjordområder og nær sokkelkanten (der området overlapper med foreslått endret SVO Eggakanten sør). Kombinasjonen av strømsystem, tilgang på dyreplankton og svampskog som sammen med korallrev i seg selv er artsrike og tilbyr leveområder for mange frittsvømmende arter, gir samlet et stort artsmangfold.
Figur 9.9.1. Kart over forslag til endret SVO Kystsonen Norskehavet sør (NH7). Grensene for det foreslåtte SVO-etSVO-et er vist med tykk rød strek, mens grenser for overlappende og inntilliggende foreslått endrete SVOSVO-er er vist med tynn rød strek. Grensene for forvaltningsplanområdet er markert med brun linje. Kartet viser utbredelsen av utvalgte miljøverdier av betydning for det foreslåtte SVOSVO-et. Miljøverdier knyttet til gytefelt for fisk/ ynglefelt for vanlig uer, samt beiteområder for pelagisk beitende sjøfugl er markert i venstre kart og miljøverdier knyttet til gytefelt for NVG-sild, bunnsamfunn og sjøpattedyr er markert i høyre kart.
Sildelaver fra sildelarvetokt. Foto: Karsten Hansen, HI
9.9.2 - Vurdering i forhold til EBSA-kriteriene - Forslag til endret SVO Kystsonen Norskehavet sør (NH7)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til endret SVO Kystsonen Norskehavet sør (NH7)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet/sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Sjøfugl Bunnsamfunn
Fisk
På Mørebankene er det miljøforhold som legger til rette for det viktigste gyteområdet for norsk vårgytende sild (Dragesund mfl. 1980).
Det er dokumentert og analysert en stor tetthet av korallrev i området (Mareano.no).
Runde er den største sjøfuglkolonien i Sør-Norge med lunde som dominerende art. Fuglefjellet har et høyt artsmangfold av sjøfugl i hekketiden, deriblant lunde, lomvi, alke, toppskarv og storjo (seapop.no). Krykkje og havhest var tidligere vanlig, men er nå ekstinkt i kolonien. Det finnes imidlertid mindre krykkjekolonier ennå i mindre kolonier på øyer og i tettsteder i området (seapop.no).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Bunnsamfunn
Fisk Sjøfugl
Mørebankene er det viktigste gyteområdet for norsk vårgytende sild. Silda gyter på bunn og gytingen foregår i februar-mars (Dragesund mfl. 1980, Sætre mfl. 2002, Sundby mfl. 2013). Første fødeopptak for sildelarver skjer i området (Sætre mfl. 2002). Viktig beiteområde for brugde (sterkt truet-EN) (f.eks. Sims 2008).
Stortareskog fungerer som skjul og beiteplass for tidlige livsstadier av bunndyr, og bidrar til strøm og bølgedemping som bidrar til retensjon av vann og beitetiden av fiskelarver (Andersen 2007, Christie mfl. 2009, Norderhaug mfl. 2112)
Mørebanken er det viktigste området for havsule i Norge, der kolonien på Runde er den desidert største i Norge (Fauchald mfl. 2015). Lundekolonien på Runde er regionalt viktig. De grunnere og mer kystnære områdene er viktige for kystnære arter som ærfugl, teist og toppskarv, både i hekketiden og som overvintringsområde (seapop.no).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Fisk
Bunnsamfunn
Sjøfugl
Viktig beiteområde for brugde (sterkt truet-EN) (f.eks. Sims 2008).
Det er dokumentert og analysert en stor tetthet av korallrev i området (Mareano.no).
Runde er den største sjøfuglkolonien i Sør-Norge, der en rekke arter, som lunde (sårbar-VU), lomvi (kritisk truet-CR) og ærfugl (nær truet-NT) har hatt sterk nedgang siden 70-tallet (seapop.no). Krykkje (sterkt truet-EN) er forsvunnet fra kolonien på 2000-tallet, det samme gjelder havhest (sterkt truet-EN). Krykkjene hekker ennå i kolonier i Nordfjord og flere steder i tettstedene ved kysten. Lunde er forsvunnet fra kolonier helt i sør (Utvær ved Sognefjorden), men finnes ennå i koloniene Veststeinen og Einevarden i Nordfjord (seapop.no). Makrellterne (sterkt truet-EN) har et av de viktigste hekkeområdene i Norge her (Systad mfl. 2007, seapop.no).
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Fisk
Sjøfugl
Området er et viktig fiskeegg/larver/yngel område (sild, torsk, vanlig uer og øyepål). Brugde er en særlig sårbar art pga sen modning, få avkom og langsom vekst; fare for bifangst (f.eks. Sims 2008).
Tareskogområdene i de kystnære områdene er viktig for en rekke kystnære sjøfuglarter i tilbakegang, med lav rekrutteringsrate, som alkefugler som legger ett eller få egg og er svært avhengig av tilstrekkelig næringstilgang for å fø opp kyllingene (seapop.no). Beiteområdene på sokkelen og ved eggakanten er viktige for de pelagiske artene som hekker i regionen, men også for overvintrende bestander, og de grunnere områdene inne ved kysten er viktige for kystnære arter som ærfugl og måker (Seapop.no).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Dyreplankton
Bunnsamfunn Sjøfugl
Fisk
Det er ikke registrert høyere dyreplanktonproduksjon generelt i området (ICES 2020, 2021), men det vil kunne være store lokale forskjeller der retensjonsområder har opphopning av dyreplankton og dermed høyere biomasse. Dyreplanktonet i området er viktig for fiskelarver og -yngel.
Mørebankene er det viktigste gyteområdet for norsk vårgytende sild (Dragesund mfl. 1980). Sildeegg er viktig næringskilde for andre organismer (Toresen 1990, Høines og Bergstad 1999, Varpe mfl. 2015). Området er også et viktig gytefelt for torsk (skrei) og hyse, samt særlig viktig både som gyte- og tidlig oppvekstområde for sei (Sundby mfl. 2013).
Det er i forhold til Nordsjøen, særlig storvokst sjøkreps i området (Søvik mfl. 2016). Stortareskogen (Laminaria hyperborea) bidrar til høy produksjon av både biomasse (Abdullah mfl. 2004), tryggere oppvekst av tidlige livsstadier av fisk og en rekke andre arter alger og virvelløse dyr (Andersen 2007, Norderhaug 2012, Christensen-Dalsgaard mfl. 2020). Tareskoger bidrar med økt oppløst organisk materie, som næring til primær og sekundærproduskjon (Abdullah og Fredriksen 2004)
Når silda gyter på Mørebankene, er dette området viktig beiteområde for sjøfugl (Seapop.no).
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Fisk
Sjøfugl Bunnsamfunn
Kombinasjonen av strømmer, opp over Eggakanten og kyststrømmen sørfra gir et mangfold av dyreliv på sokkelflatens grunne områder. En rekke arter og grupper av plankton fra Eggakanten (Torgersen mfl. 1997, Dalpadado mfl. 1998), og arter som har sokkelen med kystvann eller oseaniske områder med atlantisk vann som sine habitater, vil kunne «møtes» ytterst i området, nær Eggakanten. Tareskog på hardbunn og åpen sandbunn er leveområder for en rekke bunndyr, fra koraller og svamp, krepsdyr og andre virvelløse arter, i tillegg til mange frittsvømmende fiskearter (Brattegard og Holte 1997, Mareano.no).
Stortareskogen (Laminaria hyperborea) bidrar til høy produksjon av både biomasse (Abdullah mfl. 2004), tryggere oppvekst av tidlige livsstadier av fisk og en rekke andre arter alger og virvelløse dyr (Andersen 2007, Norderhaug 2012, Christensen-Dalsgaard mfl. 2020, Norderhaug mfl. 2020).
Sjøfuglsamfunnet på Runde har det høyeste artsmangfoldet i Norge i hekketiden (seapop.no). Mørebanken som overvintringsområde er viktig for mange kystnære og pelagiske sjøfuglarter (seapop.no).
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
9.9.3 - Lokalisering
Forslag til SVO Kystsonen Norskehavet sør er et endret SVO som dekker flere eksisterende SVO-er, sist oppdatert i i Meld. St. 20 (2019-2020), dvs. SVO Mørebankene, SVO Bremanger og Ytre Sula (i forvaltningsplanområdet for Nordsjøen), samt sørlig del av SVO Kystsonen Norskehavet (Tabell 9.1 og 10.1). Sammenslåingen av disse SVOene er begrunnet med at alle har de samme miljøverdier knyttet til gytefelt og beiteområder for sjøfugl og sjøpattedyr. Kandidatområdene for sjøfugl ved det eksisterende SVO Mørebankene der beitearealene for pelagisk beitende sjøfugl strekker seg sørover til nordre del av Sogn og Fjordane er inkludert i dette forslaget til endret SVO, og området er i tillegg utvidet inn mot kysten for å inkludere de helt kystnære gytefeltene og sjøfuglenes beiteområder. Det er en naturlig sammenheng mellom de eksisterende SVOene som nå er foreslått samlet i SVO Kystsonen Norskehavet sør.
Det foreslåtte SVO-et Kystsonen Norskehavet sør ligger utenfor kysten av Vestland og Møre og Romsdal og strekker seg fra Bremanger i sør til renna mellom Grip og Smøla i nord (Fig. 9.9.1). Området grenser til foreslått endret SVO Kystsonen Norskehavet nord, og overlapper delvis med en sørlig del av det eksisterende SVO Eggakanten som fagekspertgruppen foreslår endret, så Eggakanten i Norskehavetat skifter navn til SVO Eggakanten sør.
Foreslått endret SVO Kystsonen Norskehavet sør dekker en rekke eksponerte verneområder i kommunene Bremanger, Flora, Solund, Askvoll og Gulen, og flere områder som er foreslått som marine verneområder, Griphølen, Giske.
9.9.4 - Introduksjon til området
Foreslått endret SVO Kystsonen Norskehavet sør har variert topografi med flere banker som påvirker strømforholdene i området og som gir rom for mange ulike naturtyper og leveområder. Kyststrømmen som passerer det foreslåtte SVO-et Kystsonen Norskehavet sør, genererer topografiske strømvirvler over bankene slik at vannmassene oppholder seg der over lengre tid (Sætre mfl. 2002) (Fig. 9.9.2). Det fysiske og kjemiske miljøet er hovedsakelig påvirket av kyststrømmen som i de øvre 50-100 m dyp følger kysten nordover. Den har sin opprinnelse hovedsakelig fra Østersjøen og ferskvannsavrenning langs norskekysten. Kyststrømmen påvirker, og påvirkes av vannmasser i skjærgård og fjorder, bestemt av topografiske forhold som terskler og bassengdyp. Kjennetegn på kyststrømmen er først og fremst relativ lav saltholdighet, ~34 g/kg. Typiske hastigheter i kyststrømmen er 20-50 cm per sek men den kan bli over 100 cm per sek og vil variere med endringer i tetthet og vind (Sætre 2007, Skagseth mfl. 2011). Det er observert økt temperatur i kystsonen (~0,8°C) over de siste ~40 år, blant annet som følge av varmere atlanterhavsvann (Albretsen mfl. 2012, González-Pola mfl. 2019). Under kystvannet og lengre vest i området, dvs. nærmere eggakanten, er området mer påvirket av det saltere atlanterhavsvannet som blir ført (advektert) inn på sokkelen.
Dagens pH ligger rundt 8,05-8,1 (Lauvset mfl. 2016, Fransner mfl. 2020 Jones mfl. 2020), med laveste verdier nærmest kysten. Metningsgraden for aragonitt (ΩAr) er i underkant av to, mens den er litt over tre for kalsitt (ΩCa) (Lauvset mfl. 2016, Fransner mfl. 2020, Jones mfl. 2020), basert på sommerdata. Siden starten av den industrielle revolusjon har havforsuringen redusert pH med ca 0,11 enheter, og ΩAr og ΩCa er redusert med hhv. 0,6 og 0,9 (Lauvset mfl. 2016, Fransner mfl. 2020). Inne på sokkelen er hele vannsøylen overmettet med hensyn til både aragonitt og kalsitt.
Figur 9.9.2. Kart over gjennomsnittlig overflatestrøm (piler) og bunndyp (fargekoder) i forslag til endret SVO Kystsonen Norskehavet sør og omkringliggende områder.
9.9.5 - Beskrivelse av miljøverdiene
Plankton
Det foreligger lite informasjon om planteplankton fra området. Området har en relativt lav biomasse av dyreplankton i mai og juli/august (ca 4-8 g tørrvekt per m 2), som er tidspunktene for storskala overvåkningstidsseriene i Norskehavet innsamlet årlig av Havforskningsinstituttet (ICES 2020 a, 2021).
Generelt langs kysten, inkludert forslag til endret SVO Kystsonen Norskehavet sør, utgjør Calanus finmarchicus (raudåte) hovedmengden av biomassen. Denne arten, samt andre kopepodearter, er særdeles viktige som mat for larver og yngel av en rekke fiskearter som gyter i området eller har avkom som driver gjennom området (Sundby mfl. 2013). Det grunne sokkelområdet har lave tettheter av C. finmarchicus etter vinteren, antakelig pga. høyere predasjonsdødelighet (Bagøien mfl. 2001) og at individer blir skylt bort med vannstrømmene om vinteren (Slagstad og Tande 1996). Tilførsel av C. finmarchicus fra havområdene utenfor er dermed viktig for å forsyne kystområdet. Gjennom sesongen er det en egenproduksjon av C. finmarchicus i kystområdet, men det er ikke kjent hvor stor denne er i forhold til tilførselen. Produksjonen av C. finmarchicus i kystområdet starter i mars, men antallet holder seg lavt frem til april/mai da det er en stor økning av yngre stadier (Broms og Melle 2007, Bagøien mfl. 2012). Mai er også tidspunktet hvor konsentrasjonen av voksne individer er størst. I juni finnes et stort antall individer av både yngre og eldre stadier av den nye generasjonen. Konsentrasjonen avtar utover i juli, men i august fører produksjonen av enda ny(e) generasjon(er) til en ny økning (Broms og Melle 2007, Bagøien mfl. 2012, Strand mfl. 2020). C. finmarchicus har generelt én generasjon i året, men kan ha to eller flere generasjoner i kystvannet, som er relativt varmt, og produksjonssesongen blir dermed forlenget (Broms og Melle 2007, Bagøien mfl. 2012). Planktonets sesongutvikling er tidligere i kystvannet (2-6 uker) sammenliknet med atlantiske og arktiske vannmasser i Norskehavet (Broms og Melle 2007, Bagøien mfl. 2012).
Det har vært en reduksjon i mengden raudåte (C. finmarchicus) fra 1995 til 2011, basert på prøvetaking langs Svinøysnittet, et overvåkningstransekt som går fra Herøy i Møre og Romsdal og 195 nautiske mil mot nordvest. Nedgangen har vært størst i kystvannet innerst mot kysten (ca. 81 prosent) og lavere i ytre deler av Mørebankene (ca 60 prosent). I atlanterhavsvann vest for bankene (dvs. lengre fra kysten) har nedgangen vært på ca 50 prosent (Dupont mfl. 2017). Det er ikke gjort analyser av raudåte for perioden etter 2011, men generelt i Norskehavet har den totale mengden dyreplankton vært på et stabilt og antakelig noe økende nivå etter 2011.
Det foreligger også opparbeidede prøver fra Svinøysnittet som inkluderer andre dyreplanktonarter i området, men disse dataene er ikke analysert for langtidstrender. Det er ikke gjort analyser av den totale artssammensetningen i området, men basert på de opparbeidede prøvene er det ikke rapportert om unike arter. Kyststrømmen med vannmasser fra områder lengre sør, strømmer over Mørebankene og videre nordover langs kysten, og sørlige arter av dyreplankton som er vanlige i Nordsjøen eller lengre sør blir transportert inn i området. Området ligger også i utbredelsesgrensen til tempererte arter som f.eks. C. helgolandicus (Beaugrand mfl. 2002). Eventuelle endringer i mengden av sørlige arter som tidligere ikke har hatt sitt tradisjonelle leveområde langs norskekysten eller i Norskehavet, vil dermed tidlig gjøre seg gjeldende i dette foreslått endrete SVO-et. Fra 2006 ble det observert en økning i sørlige arter i området, men etter 2011 synes det å ha vært en nedgang. De sørlige artene forekommer i lavt antall og vil i liten grad påvirke økosystemet i området (Miljostatus.no).
Fordi Svinøysnittet blir innsamlet flere ganger i året, og det foreligger historiske data tilbake til 1990-tallet, er området ved dette snittet et viktig overvåkningsområde mht. endringer i planktonets artssammensetning og mengde. Da dette er et område i Norskehavet der effekter av klimaendringer tidlig kan observeres, bl.a. i form av endringer i mengden av tempererte arter (f.eks. C. helgolandicus) versus sub-arktiske arter (f.eks. C. finmarchicus), er det viktig å opprettholde overvåkningen.
Fisk
Norsk vårgytende sild (Sild, Clupea harengus) hører til den atlantoskandiske sildestammen sammen med to andre bestander, islandsk sommergytende og islandsk vårgytende sild. Sild gyter på bunn og er avhengig blant annet av riktig bunnsubstrat for vellykket gyting og vil således gyte langs store deler av norskekysten (Runnstrøm 1941). Gyteområdene for norsk vårgytende sild varierer over tid, men Mørebankene, som anbefales inkludert i forslag til endret SVO K ystsonen Norskehavet sør, er et særlig viktig gyteområde der den største andelen av silden gyter (Dragesund mfl. 1980 , Sætre mfl. 2002 , Sundby mfl. 2013). Sild a vandrer inn fra overvintringsområder i havet og i norske fjorder, og gyter på Mørefeltene i perioden februar/mars. Etter gyting vandrer silda vestover på beitevandring og vil ikke i særlig grad finnes på Mørebankene utenom gyteperioden (Holst mfl. 2004). Gyteproduktene er en stor matkilde for dyr langs kysten om våren og sommeren, og er beregnet i 2020 til å utgjøre over et halvt tonn (ICES 2020b, 2021). Sildeeggene ligger på bunn i ca. tre uker før klekking og vil i den perioden være viktig næringskilde for fisk og andre organismer som beiter på eggene (Toresen 1991 , Høines og Bergstad 1999 , Varpe mfl. 2005). Sildelarvene vil etter klekking drive nordover med kyststrømmen langs kysten, og driver inn i oppvekstområdene i Barentshavet tidlig på sommeren. Larvene vil ofte ha en viss oppholdstid på gytebankene, både på selve Mørebanken, men også oppover langs kysten nord for det foreslåtte SVO-et (Sætre mfl. 2002). Mørebankene er derfor et viktig beiteområde for sildelarvene den første tiden etter klekking. Sild har stor betydning for økosystemene langs kysten og i Barentshavet. Denne larve- og yngeltransporten langs store deler av norskekysten er et viktig moment i begrunnlsen for å anbefale at alle de foreslåtte kystnære SVO-er grenser til hverandre, fra Bremanger til og med Finnmarkskysten.. Silda beiter på raudåte og er selv en viktig matressurs for sjøfugl, rovfisk som torsk, sei og annen bunnfisk i tillegg til hval. Store flokker av spekkhoggere følger silda på dens vandringer. Gytebestanden av norsk vårgytende sild nådde en foreløpig topp i 2008. Etter 2009 har gytebestanden minket og er i 2020 estimert til å være 3,5 millioner tonn. Rekruttering fra unge årsklasser til gytebestanden har vært dårlig i mange år, men 2016-årsklassen er foreløpig estimert til å være over middels og ventes å bidra til å bremse nedgangen i gytebestanden dersom forvaltningsplanen blir fulgt (ICES 2020b, 2021).
Øyepål (Trisopterus esmarkii) er en ganske liten fisk i torskefamilien med maksimal individvekt på 0,1 kg og med en levetid som sjelden går over tre år. Øyepål opptrer i store stimer, som regel over mudderbunn. Den spiser hovedsakelig planktoniske krepsdyr, som krill og raudåte (Sundby mfl. 2013) . Øyepål utgjør igjen en del av næringsgrunnlaget for andre større fisk som torsk, hvitting og sei, men også sjøpattedyr. Siden 1995 har fiskedødeligheten vært på et lavere nivå enn tidligere. Utbredelsen strekker seg også nordover n orskekysten og inn i det sørlige Barentshavet. Hovedgytefeltene er spredt ut i den nordlige delen av Nordsjøen. De nordlige gytefeltene som har relevans for egg-, larve- og yngeldrift i de foreslåtte SVO-ene Kystsonen Norskehavet sør og Kystsonen Norskehavet nord omfatter kysten av Møre, Haltenbanken, Vestfjorden, rundt Røst og utover langs vestligste delen av kontinentalsokkelen utenfor Vesterålen (Sundby mfl. 2013) . Bestandsstørrelsen varierer mye fra år til år på grunn av variabel rekruttering og et kort livsløp.
Mørebankene er også et viktig gytefelt for hyse (Melanogrammus aeaglefinus) fra mars til juni og for sei (Pollachius virens) om vinteren med topp i februar. Sei har dette som et av sine kjerneområder for gyting og tidlig oppvekst (Sundby mfl. 2013).
Kysttorsken (Gadus morhua) gyter her i mars-april, og også nordøstarktisk torsk (skrei) har tidligere hatt gytefelt her. Gytingen av kysttorsk i området foregår både ute på kysten og inne i beskyttede fjordarmer, i relativ nærhet til oppvekstområdene for yngelen. Merkeforsøk har vist at kysttorsk gjerne oppholder seg innenfor rekkevidden av en enkelt fjord (Jakobsen 1987, Karlsson og Mork 2003) og gytingen skjer på samme sted fra år til år.
Brugde (Cetorhinus maximus) er en særlig sårbar art pga sen modning, få avkom og langsom vekst, og er listet som truet på norsk rødliste. Mørebanken er identifisert som viktig område særlig om sommeren (observasjonsdata fra https://dugnadforhavet.no, pers medd. Runde Miljøstasjon, Junge mfl. in prep). Brugde forekommer langs habitater på kontinentalsokkelen der fronter ofte er til stede for å oppkonsentrere plankton (f.eks. Sims 2008). Brugde spiser dyreplankton, med kalanoide kopepoder (Calanus arter) som den viktigste byttedyrgruppen.
Bunnsamfunn
Mareano har kartlagt bunnsamfunn og har både dokumentert og modellert antall koralllrev i og ved overlappsonen for de foreslått endrete SVOene Kystsonene Norskehavet sør og nord samt i Eggakanten sør (se Fig. 9.7.3). Det er dokumentert 832 korallrev, og modellert i overkant av 200 000 korallrev i kartutsnittet som er vist i Fig. 9.7.3 (Mareano.no).
Det er lite kunnskap om bestanden av dypvannsreke (Pandalus borealis) nordover langs norskekysten sammenlignet med bestandene i Nordsjøen/Skagerrak og i Barentshavet. Dette gjelder særlig for rekene langs kysten i Vestland, Møre og Romsdal og Trøndelag fylker. Bestandsstatus til dypvannsreken ved Mørebankene er derfor ikke kjent.
Sammenlignet med Nordsjøen (ICES 2020c, 2021) skiller de foreslåtte kystnære SVOene ved norskekysten seg ut med særdeles store individer av sjøkreps. Et kystnært teinefiske etter sjøkreps (Nephrops norvegicus), både kommersielt og fritidsfiske, har vokst frem de siste årene (Søvik mfl. 2017). Landingene fra det kommersielle teinefisket på Møre (statistisk område 07) har økt de siste fem årene, med de hittil høyeste landingene i 2020 (data fra Fiskeridirektoratet). Det er per dags dato vanskelig å verdisette de enkelte sjøkrepsfeltene langs norskekysten, og vi kjenner heller ikke bestandsstatus til denne arten i dette foreslåtte SVO-et.
Det foreslåtte SVO-etSVO-et overlapper med forslag til endret SVO Eggakanten sør. Korallrev og andre sårbare naturtyper langs kontinentalsokkelen og øvre del av skråningen er omtalt under beskrivelsen av det foreslåtte SVO-etSVO-et Eggakanten sør (Kap. 9.7). Utenom revene langs Storegga (Eggakanten sør) forekommer de fleste korallrevene innen SVO Kystsonen Norskehavet sør kystnært og i Breisunddjupet (korallverneområde), nordvest av Ålesund. Mindre skader påført av bunnfiske-redskaper er observert i flere tilfeller (Fosså mfl. 2015). Mareano har modellert sannsynlig utbredelse av generelle biotoper (identifisert fra analyse av sammensetning av megafauna observert fra videoopptak) (Fig. 9.8.6, Kap. 9.8). Tre av disse generelle biotopene tyder på at sårbare og verdifulle naturtyper er til stede. Både video-observasjoner og kart over de modellerte generelle biotopene indikerer at svampskog er relativt vanlig i den mest kystnære delen av SVO Kystsonen Norskehavet sør.
Stortareskogen (Laminaria hyperborea) som opptrer i det foreslåtte SVO-et, er viktige biotoper for hele økosystem (Christie mfl. 2009, Norderhaug mfl. 2020), for produktiviteten i økosystemet, som tryggere leveområde for tidlige livsstadier (Christensen-Dalsgaard mfl. 2020), helårsleveområder for en rekke virvelløse dyr og ved å tilføre området næringssalter når gamle algedeler brytes ned (Andresen 20007, Jo mer eksponert tareskogen er mot havet, jo høyere artsmangfold er det som påvekst og assositerte arter (Norderhaug mfl. 2012). Samlet bidrar tareskogen med økt produktivitet i SVO-et (Abdullah og Fredriksen 2004).
Sjøpattedyr
Spekkhogger (Orcinus orca) spiser sannsynligvis mest makrell og sild (Nøttestad mfl. 2014). Langs norskekysten opptrer spekkhogger gjennom hele året (Christensen 1982, Øien 1988), men det er særlig om vinteren de kan utgjøre store ansamlinger i forbindelse med overvintringen av sild (Similä mfl. 1995). For tida vil dette i særlig grad gjelde det foreslått endrete SVO Tromsøflaket, og historisk sett for de foreslått endrete SVO-ene Kystsonen Norskehavet sør og Kystsonen Lofoten. Utenom overvintringsperioden, har spekkhoggerne i kystnære farvann sannsynligvis en mer variert diett.
Haverten (Cystophora cristata) er flokkdyr som danner kolonier, særlig i forbindelse med ungekasting (fødsel) og parring (september–desember) og hårfelling (februar–april). I resten av året er de spredt langs store deler av kysten. I forvaltningsplanområdet for Norskehavet har ungeproduksjonen på strekningen Lofoten – Stad gått ned fra 943 unger før 2008 til 398 unger etter 2014 (Nilssen mfl. 2020).
Steinkobben (Phoca vitulina), vår mest tallrike kystsel, er en liten selart. Steinkobbene samles på land i forbindelse med forplantning (juni-juli) og hårfelling (august). Ungene blir født med korthåret voksenpels og går i vannet etter noen timer. I yngle- og hårfellingstiden fra midten av juni til begynnelsen av september er steinkobbene svært stedbundne og knyttet til stedet der de ble født. Fra andre del av september til tidlig juni, er det næringssøk som har høyeste prioritet. I denne perioden vil de oppsøke steder med god tilgang på mat, ofte mange titalls kilometer fra stedet der de ble født. Bestanden har gått noe tilbake i Trøndelag og sørlige deler av Nordland. De største koloniene i SVO-et er på Sandøy og Haram i Ålesund kommune, Møre og Romsdal (Bjørge og Nilssen 2019).
Klappmyss (Cystophora cristata) kan beite ute i dyphavet og har et hot-spot ved Shetland, men det er ikke kjent at de trekker helt inn i dette foreslåtte SVO-et, i hvert fall ikke i nyere tid (Folkow mfl. 1996, Folkow mfl. 2010)
Sjøfugl
Forslaget til endret SVO Kystsonen Norskehavet sør er også begrunnet i at det er viktig beiteområde for mange sjøfuglarter, herunder havsule (Morus bassanus), lomvi (Uria aalge), lunde (Fratercula arctica) og alke (Alca torda). I dette området hekker disse artene primært på Runde, men det finnes også kolonier på Veststeinen der lunde er den dominerende arten og på Einevarden der krykkje (Rissa tridactyla) er den dominerende arten), begge koloniene i Sogn og Fjordane. De to siste koloniene hører til i området fra Bremanger til Ytre Sula, men de beiter også på Mørebanken (Systad mfl. 2019).
Runde er en nøkkellokalitet i Seapop. Fuglefjellet har det høyeste artsmangfoldet for sjøfugl i Norge, og huser en rekke arter som er sårbare og truede. Runde er det største lundefjellet sør for Røst (>50 000 par), havsulekolonien her er på mer enn 3500 par, mer enn 50 prosent av den totale norske bestanden (Seapop.no). Storjo (Stercorarius skua) er tett koblet mot havsulene og parasitterer denne arten for mat. Storjokolonien er den største på fastlandet i Norge, og sammenlignbar med kolonien på Bjørnøya, med ca. 100 hekkende par.
Figur 9.9.3. Kart viser fordeling av pelagiske sjøfuglarter (Havsule, krykkje, lomvi, lunde, alke), kystnære fiskespisende arter (Toppskarv, storskarv, teist), kystnære bentisk beitende arter (Ærfugl) og kystnære overflatebeitende arter (Gråmåke, sildemåke, tyvjo og svartbak) i forslag til endret SVO Kystsonen Norskehavet sør og nærliggende områder. Skravert felt i A) viser beiteområder for pelagisk beitende sjøfugl. Grensene for forslag til endret SVO-et er vist med tykk rød strek, mens grenser for inntilliggende og overlappende forslag til endrete SVO-er er markert med tynn rød strek.
De siste ti årene har havsule og storjo her hatt en årlig vekst på hhv. åtte prosent og fem prosent. Bestanden av lomvi har hatt en nedgang på 14 prosent i den siste tiårsperioden. Lunde har hatt en årlig nedgang på seks prosent og nedgangen i toppskarvbestanden (Phalacrocorax aristotelis) som var på over 5 000 par, har vært særlig stor etter 2012, med fullstendig hekkesvikt fra dette året og framover. Det virker imidlertid som at hekkingen har vært vellykket på andre deler av øya enn i overvåkningsfeltene, slik at situasjonen sannsynligvis er litt bedre enn det overvåkingsresultatene kan tyde på (www.miljostatus.no). Toppskarv finnes for øvrig i en rekke kolonier langs kysten i dette foreslåtte SVO-et.
Foreslått endret SVO Kystsonen Norskehavet sør er også et viktig overvintringsområde for alkekonge (Alle alle) og gulnebblom (Gavia adamsii). Også flere kystnære arter beiter i de grunne områdene av SVO Kystsonen Norskehavet sør, og det er gode forekomster av ærfugl (Somateria mollissima), toppskarv (Phalacrocorax aristotelis), gråmåke (Larus argentatus) og sildemåke (Larus fuscus).
Fisk over tareskog. Foto: Frithjof Moy, HI
9.10 - Forslag til nytt SVO Dyphavsområdene i Norskehavet (NH8)
9.10.1 - Sammendrag
Områdebeskrivelse og strukturer
Forslag til nytt SVO Dyphavsområdene i Norskehavet (NH8) gjelder to adskilte bassenger, Norskehavsbassenget i sør (3600 m dypt) og Lofotenbassenget (3200 m dypt) i nord, på hver side av Jan Mayen bruddsonen (se SVO Midtatlantisk rygg). De dekker de to dypeste bassengene sentralt i Norskehavet og har en tydelig syklonisk sirkulasjon (retning mot klokken) i dypet med svake strømmer i de flate og indre områdene av bassengene sammenlignet med ytterkantene av bassengene. Det er derfor en stor grad av resirkulering og lang oppholdstid av vann inne over bassengflatene. Det fysiske og kjemiske miljø i foreslått nytt SVO Dyphavsområdene i Norskehavet er bestemt av fordelingen mellom atlantisk og arktisk vann i det øvre laget, fra overflaten ned til ca. 200-300 m dyp i Norskehavsbassenget, og av atlanterhavsvann ned til ca. 600-800 m dyp i Lofotenbassenget. Topografien i de dype bassengene i Norskehavet viser mange ekstremt bratte undervannsfjell som stikker høyt opp fra dyphavsslettene og som innrammer dyphavsbassengene. Ægirryggen strekker seg tvers over den sørlige del av området.
Miljøverdier
Dyphavsområdene i Norskehavet er særdeles viktige som overvintringsområder og reservoar for Calanus-artene, og helt sentrale for å opprettholde de store populasjonene av Calanus spp. Slike dyp er helårs leveområde for mesopelagiske arter, som laksesild, liten laksetobis og nordlig lysprikkfisk, i tillegg til krill, amfipoder, hoppekreps og blekksprut, som igjen er viktig føde for dypdykkende hval, som spermhval, finnhval og nebbhval. Dessuten er det rødlistede, skjøre og sent restituerende bunnsamfunn som korallskoger og svampeskoger i området. På de dype mudderslettene er antagelig særlig svampeområdene oppvekstområde for mange andre arter. Områder med bløtbunn innimellom de harde sidene gjør at sidene av Ægir-ryggen blir særlig artsrike. I øvre sone er dyreplankton, særlig Calanus-arter, sentrale i dietten til pelagisk fisk som sild, kolmule og makrell som igjen inngår i dietten til vågehval. Disse planktonreservoarene har også betydning for produktivitet og reproduksjon av fisk og sjøfugl i eggakantens og kystsonens SVO-er. På så store dyp er menneskelig påvirkning svært liten, så for fisk og dyreplankton er området ansett som naturlig.
Raudåte, Calanus finnmarchicus . Foto: T. van der Meeren, HI
Gonatus-art. Foto: CT. Broms, HI
Figur 9.10.1. Kart over foreslått nytt SVO Dyphavsområdene i Norskehavet (NH8). Grensene for området er vist med tykk rød strek, mens grenser for nærliggende, foreslått endrete SVO-er er vist med tynn rød eller oransje strek . Grensene for forvaltningsplanområdene er markert med brun linje. Kartet viser utbredelsen av utvalgte miljøverdier (forhøyet produksjon av plankton) av betydning for det foreslåtte SVO-et.
9.10.2 - Vurdering i forhold til EBSA-kriteriene – Forslag til nytt SVO Dyphavsområdene i Norskehavet (NH8)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til nytt SVO Dyphavsområdene i Norskehavet (NH8)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet/sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Fisk
Mesopelagisk fauna Bunnsamfunn
Dyreplankton
De to dype havområdene er særdeles viktige som bestandsreservoar, som opprettholder de store populasjonene av Calanus spp., som overvintrer i dypet (Torgersen og Huse 2005). Syklonisk sirkulasjon fører til stor grad av retensjon av Calanusfinmarchicus i områdene (Torgersen og Huse 2005, Melle mfl. 2014), som brer seg inn over sokkelen langs Eggakanten fra Stad til Svalbard.
Store bestander av mesopelagiske arter og andre dypvannsfisk lever her i dypet (Salvesen 2004). SVO Dyphavsområdene i Norskehavet er dypt nok til at det er helårs leveområde for mesopelagiske arter, som laksesild (Maurolicusmuelleri), liten laksetobis (Arctozenusrisso) og nordlig lysprikkfisk (Benthosema glaciale) (Dalpadado mfl. 1998; Salvanes 2004).
Hardbunnsområdene langs kanten av Ægirryggen innehar tette områder med bløtkoraller og svamper. Dette er økosystem som regnes som unike og som truet av OSPAR-kommisjonen (OSPAR 2008).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Fisk Bunnsamfunn Sjøpattedyr
Dyreplankton
Dyphavsområdene er overvintringshabitat for Calanus spp. og særdeles viktige for å opprettholde populasjonene (Melle mfl. 2014).
Derfor er de også viktige beiteområder for de tre pelagiske fiskeartene norsk vårgytende sild, makrell og øyepål, gjennom sommeren (Sætre mfl. 2002, Holst mfl. 2004). Den nordlige del av området er et viktig beiteområde for snabeluer gjennom sommeren (ICES 2019, 2021). Det er helårs leveområde for mesopelagiske fiskearter, som laksesild, liten laksetobis og nordlig lysprikkfisk (Dalpadado mfl. 1998, Salvanes 2004).
På de dype mudderslettene er antagelig særlig svampeområdene oppvekstområde for mange andre arter (Brix mfl. innsendt).
Sommerbeite for vågehval (pelagisk fisk) og spermhval (krepsdyr, blekksprut, mesopelagisk fisk) (Skjoldal mfl. 2004, Nøttestad mfl. 2015).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Bunnsamfunn
Svampskoger og korallrevene er vurdert som truet av OSPAR-kommisjonen (OSPAR 2008) og står på den norske rødlisten over naturtyper (Artsdatabanken 2018).
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Bunnsamfunn
Korall- og svampeskogene i skråningene langs Ægirryggen er begge økosystem som regnes som skjøre. De er vurdert som truet av OSPAR-kommisjonen (OSPAR 2008) og står på den norske rødlisten over naturtyper (Artsdatabanken 2018).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Bunnsamfunn
Dyreplankton
Calanus spp. overvintrer i dype havområder og områdene er dermed avgjørende for å opprettholde høy Calanus-produksjon i Norskehavet. Populasjonen fra Norskehavsbassengene forsyner omkringliggende hav- og sokkelområder med Calanus, og er dermed viktig for sekundærproduksjonen i bl.a. Barentshavet, Nordsjøen og Norskekysten (Melle mfl. 2014, Skjoldal 2004).
Svampeskoger og korallhager er begge økosystem som skaper leveområder for høye tettheter av assosiert fauna (Brix mfl. innsendt).
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Fisk Bunnsamfunn
Det er en rekke arter fisk, mange sjeldent sett, i dypet under lyssonen, inkludert laksesild og lysprikkfisk (Dalpadado mfl. 1998, Salvanes 2004), i tillegg til blekksprut, isopoder, krill og Calanus, som igjen er viktig føde for fisk og sjøpattedyr (Skjoldal mfl. 2004).
Ægirryggen, og særlig de bratte sidene der har store områder med tette svampe- og bløtkorallområder. Disse har en rikt assosiert fauna (Brix mfl. under vurdering 2021). Områder med bløtbunn innimellom de harde sidene gjør at sidene av ryggen blir særlig artsrike.
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
Dyreplankton Fisk Mesopelagisk fauna Bunnsamfunn
Dette gjelder særlig under lyssonen, fra den mesopelagiske sonen og ned til bunnen der menneskelige aktiviteter antas å ha lite påvirkning på økosystemet. Klimaendringer og forsuring antas å skje saktere på større dyp.
9.10.3 - Lokalisering
Foreslått SVO Dyphavsområdene i Norskehavet er et nytt SVO, basert på ny informasjon som ikke var til stede da de nå eksisterende SVOene ble utredet.
Dyphavsområdene omfatter to dype havområder i Norskehavet, det sørlige Norskehavsbassenget og det nordlige Lofotenbassenget med dybder ned til henholdsvis 3600 m og 3200 m dyp (Fig. 9.10.1). Områdene er avgrenset av ekstremt bratte og høye undervannsfjellsider som innrammer dyphavsbassengene.
Det er ikke etablerte verneområder i dette forslått nye SVO-et.
9.10.4 - Introduksjon til området
Begge dyphavsområdene har en tydelig syklonisk sirkulasjon (retning mot klokken) med svake strømmer i de flate og indre områdene av bassengene sammenlignet med ytterkantene av bassengene. Strømhastighetene i dypet er henholdsvis <1 cm per sek s det indre og opp til 5 cm per sek i ytterkantene (Voet mfl. 2010). Dette medfører at vannmassene i Dyphavsområdene i Norskehavet har stor grad av resirkulering og lengre oppholdstid sammenlignet med andre områder med sterkere strømmer. Det fysiske og kjemiske miljø i Dyphavsområdene er bestemt av fordelingen mellom atlantisk og arktisk vann i det øvre laget, fra overflaten ned til ca. 200-300 m dyp i Norskehavsbassenget og av atlanterhavsvann ned til ca. 600-800 m dyp i Lofotenbassenget (Blindheim og Østerhus 2005). Arktisk vann er mest fremtredende i de vestlige områdene av Norskehavsbassenget. Under det øvre laget er det et kaldere arktisk intermediært lag med temperaturer rundt nullpunktet, og under det igjen ligger dypvannet med temperaturer under 0°C. Om vinteren blir det øvre laget utsatt for sterk avkjøling og kraftige vinder som medfører at vinterblandingen rekker ned til 300 m og 600 m dyp for henholdsvis Norskehavs- og Lofotenbassenget (Nilsen og Falck 2006). Nye undersøkelser har vist at topografien i de dype bassengene i Norskehavet er mye mer kompleks enn man tidligere trodd, med mange undervannsfjell som stikker høyt opp fra dyphavsslettene og med ekstremt bratte og høye fjellsider som innrammer dyphavsbassengene. Den gamle, inaktive Ægirryggen strekker seg tvers over den sørlige del av området.
pH i overflatevannet ligger rundt 8,1, og er litt høyere i Lofotenbassenget enn i Norskehavsbassenget, denne forskjellen er hovedsakelig temperaturdrevet (Fransner mfl. 2020, Lauvset mfl. 2016). Overflatevannet er overmettet for både kalsitt og aragonitt, i Norskehavsbassenget ligger metningsgraden for kalsitt i overkant av tre mens for aragonitt er den i overkant av to. I Lofotenbassenget er metningsgraden litt høyere for begge kalkmineralene, mye fordi det er større innslag av arktiske vannmasser i sør. Metningsgraden avtar nedover i dypet, og vi finner metningshorisonten for aragonitt på rundt 2000 m, mens vannet er overmettet med kalsitt helt til bunns. Måleserier både fra forskningstokt og tidsserien på Stasjon M (2°Ø, 66°N) viser signifikante trender i havforsuringsparametre ned til 1000 m i begge basseng (Fransner mfl. 2020, Jones mfl. 2020, Skjelvan mfl. 2008).
Figur 9.10.2. Venstre kart) Simulert overflatestrøm i Norskehavet for 19. mars 2021 kl. 13:00. (Datakilde: OSCAR/Earth & Space Research (https://www.esr.org/research/oscar/ og https://earth.nullschool.net). Høyre kart) Gjennomsnittlig overflatstrøm (piler) og bunndyp (fargekoder) i foreslått nytt SVO Dyphavsområdene i Norskehavet og omkringliggende områder.
9.10.5 - Beskrivelse av miljøverdiene
Plankton
Den sterke vinterblandingen medfører at næringsstoffer fra dypet blir transportert til overflaten slik at tilgangen på næringsstoffer ikke er en begrensende faktor for våroppblomstringen (Blindhiem og Østerhus 2005).
Dyphavsområdene i Norskehavet er særdeles viktige for å opprettholde de store populasjonene av Calanus-artene, da artene overvintrer i dypet og høye tettheter synes å være knyttet til steder med dype overvintringsområder. Syklonisk sirkulasjon i Norskehavsbassenget og Lofotenbassenget fører til stor grad av retensjon av plankton i disse områdene sammenliknet med områder med sterkere strøm, som blant annet langs Norskekysten (Torgersen og Huse 2005). Calanus-artene (C. finmarchicus, C. hyperboreus, C. glacialis, C. helgolandicus) utgjør hovedmengden av dyreplanktonet i de norske havområdene Norskehavet, Nordsjøen og Barentshavet, og i de tilgrensende havområdene Islandshavet og Grønlandshavet, samt tilhørende sokkelområder. I Norskehavet er det C. finmarchicus (raudåte), som er mest utbredt av disse artene (Broms mfl. 2009). Biomassen av raudåte 1. juli i Norskehavet, som er modellert til å være tidspunktet for produksjonstoppen, er estimert til 33 millioner tonn (for et område på 1,2 millioner km2) og den årlige produksjonen i samme område er estimert til 190-290 millioner tonn (Skjoldal mfl. 2004, Hjøllo 2012).
De dype havbassengene i Norskehavet og den subpolare gyren i Labrador-/Irmingerhavet er antatt å være raudåtas hovedsenter (Melle mfl. 2014). Av bassengene i Norskehavet er det særlig Norskehavsbassenget som anses som hovedovervintringsområde (Sundby 2000, Broms mfl. 2009).
Raudåta er funnet å overvintre mellom 600 og 1000 m dyp i sørøstlige Norskehavet, hovedsakelig rundt 800 m ved Stasjon M (2°Ø, 66°N), hovedsakelig mellom 700 og 800 m i Lofotenbassenget, mellom 800 og 1200 m vest for Tromsøflaket, og i Færøy–Shetland-kanalen er overvintringen hovedsakelig funnet å være i de øvre lagene av Norskehavsdypvannet mellom 600 og 1200 m (Dale mfl. 1999, Heath mfl. 2000, Halvorsen mfl. 2003, Melle mfl. 2004). Overvintringsdypet varierer geografisk i Norskehavet, og er generelt dypere i nord og øst enn i sør og vest siden den overvintrer under det atlantiske overflatevannet og står i arktisk-intermediære vannmasser. Det vil derfor være vannmassene mellom ca 400 og 1500 meters dyp på senhøsten og vinteren som er særdeles viktige som SVO for dyreplankton.. Raudåta vandrer ned til dype vannmasser på sensommeren og høsten, og etter overvintringen starter oppvandringen på senvinteren. Raudåte kommer til overflaten igjen utover våren (Bagøien mfl. 2012, Melle mfl. 2014).
De dype havbassengene fungerer som bestandsreservoarer og fordelingssentre for Calanus spp, og høye konsentrasjoner av raudåte i områder som ligger kystnært Norskehavet er sannsynligvis avhengige av tilførsel fra Norskehavsbassenget. Dette gjelder kontinentalsokkelen der grunnere overvintringsdyp og sannsynligvis høyere dødelighet pga predasjon (Bagøien mfl. 2001), samt mulighet for at individer blir skylt bort med vannstrømmene om vinteren (Slagstad og Tande 1996), fører til lave tettheter etter vinteren. Tilførsel av raudåte fra havområdene utenfor er dermed viktig for å forsyne larver og yngel av en rekke fiskearter som gyter langs Norskekysten. Selv om overvintring av Calanus arter. i dype fjorder er viktig for lokale fiskebestander, har fjordene tilførsel av atlanterhavsvann fra Norskehavet. Dyreplankton, inkludert Calanus-arter, blir transportert nordover og østover fra Norskehavet inn i Barentshavet, og en stor del av raudåta som finnes i Barentshavet er transportert inn fra Norskehavet (Skaret mfl. 2014).
Raudåta er en viktig næringskilde for mange arter. Dyrespisende dyreplankton (karnivore dyreplankton) som for eksempel amfipoder (Themisto spp.) og krill, beiter på raudåta. Raudåta utgjør en stor del av dietten til de pelagiske fiskeartene sild, makrell og ung kolmule. Yngre stadier av raudåte er viktig mat for larver og yngel av en rekke fiskearter, blant andre torsk og sild. Samtidig som amfipoder og krill er konkurrenter til sild, makrell og kolmule, fordi de alle beiter på raudåta, så er amfipoder og krill også selv mat til de pelagiske fiskeartene. Slik inngår raudåta som en viktig del av et komplekst næringsnett (se Skjoldal mfl. 2004).
Fisk
På grunn av planktonressursen fra dypet er foreslått nytt SVO Dyphavsområdene i Norskehavet også viktig beiteområde for de tre pelagiske fiskeartene (sild, makrell og kolmule) gjennom sommeren. Etter gyting drar den norske vårgytende silda (Clupea harengus) ut i Norskehavet på en lang vandring for å finne mat. Den beiter på raudåte hele sommeren over store deler av havet, men særlig i sentrale og vestlige deler (Sætre mfl. 2002). Den er selv en viktig matressurs for hval i åpent hav i Norskehavet, men beiter primært i de øverste 30 meterne av vannsøylen gjennom døgnet og jaktes derfor særlig av vågehval og spekkhogger.
Makrellen (Scomber scombrus) står utbredt i sentrale og nordlige deler av Norskehavet inkludert sør–sørøst av Jan Mayen på sildetoktene. Den blir selv spist av stor fisk (makrellstørje (Thunnus thynnus) og hval (spekkhoggere, finhval og knølvhal) (Nøttestad mfl. 2014, Nøttestad mfl. 2020, Løviknes mfl. 2021).
Kolmule (Micromesistius poutassou) er en liten torskefisk som hovedsakelig holder til i Nordøst-Atlanteren og i Middelhavet. Om sommeren befinner kolmula seg betydelig dypere enn makrellen og silda ute i Norskehavet. Større voksen kolmule finnes vestover i de åpne havområdene over svært store områder av Norskehavet. De kan stå mellom 80 og 400 m dyp, avhengig av område, vannmasser og tid på døgnet.
Den nordlige del av foreslått nytt SVO Dyphavsområdene i Norskehavet er et viktig beiteområde for snabeluer gjennom sommeren (ICES 2019, 2021).
Mesopelagisk fauna
Foreslått nytt SVO Dyphavsområdene i Norskehavet er dypt nok til at det er helårs leveområde for mesopelagiske arter, som laksesild (Maurolicus muelleri), liten laksetobis (Arctozenus risso) og nordlig lysprikkfisk (Benthosema glaciale) (Dalpadado mfl. 1998, Salvanes 2004). De antas å være blant de mest tallrike fiskeartene i Nordatlanteren og er viktig bytte for pelagiske fiskebestander. De finnes også lokalt i dype fjorder og langs Eggakanten siden de er tett knyttet til dypet og bare trekker opp om natten (Dalpadado mfl. 1998, Salvanes 2004). Det finnes også betydelige mengder mesopelagiske krepsdyr og blekksprut, som akkar (Thodarodes sagittatus) og Gonatus-arter ute i havdypet i Norskehavet, med økende biomasse i den østre delen av SVO-et (Skjoldal mfl. 2004).
Bunnsamfunn
Den sørøstligste delen av dette området ble undersøkt bentisk av IceAGE3-toktet sommeren 2020 (Brix mfl. sendt inn 01-2021). De dype mudderslettene er visuelt dominert av sjøpølser (Elpidia arter), sjøliljer (Bathycrinus arter), svamp (dominert av Caulophacus cf. arcticus) og sjøstjerner (Hymenaster pellucinus). Særlig svampeområdene virker å være oppvekstområde for mange andre arter, og mye assosiert krepsdyrfauna ble observert på nesten hver svamp. Fysiske prøver med halte (hyperbentisk slede) og gravende (grabb/kjerne) prøvetakere fra mudderslettene viser et rikt samfunn av små (> 10mm) evertebrater dominert av mange børstemark (56 prosent) og hyperbentiske krepsdyr, blant annet Tanaidacea, Isopoda, Amphipoda og Harpacticoida (bunnlevende og helårs-stedbundne kopepoder). Åtte sledeprøver har resultert i litt over 10 000 individer. Ægir-ryggen som strekker seg S/SØ gjennom området er en ekstinkt spredningsrygg (33 mill år gammel, og har derfor et tynt sedimentlag over hard skorpe). Den har en flat, sedimentdekket topp, med relativt bratte skråninger og mange lokale raviner, og her finnes mye hardbunnsfauna med et rikt assosiert dyreliv, mye tilsvarende det som finnes på sjøfjell. Områder med bløtbunn innimellom de harde sidene gjør at sidene av ryggen blir særlig artsrike. Tette områder med bløtkoraller og svamper ble observert på de få stasjonene som ble utført, og anemoner, snegler, slangestjerner, sjøstjerner og krepsdyr var organismegrupper som dominerte i habitatene som bløtkoraller og svamper lager (Brix mfl. sendt inn 04-2021).
Sjøpattedyr
Disse områdene har en viss betydning som beiteområder for vågehval (Balaenoptera acutorostrata) som beiter på de pelagiske fiskebestandene. Vågehval er tallrik i Nord-Atlanteren, og har en allmenn utbredelse om sommeren i Nordøst-Atlanteren. Dessuten er spermhval knyttet til dyphavsområdene i Norskehavet der de beiter på blekksprut og mesopelagisk fisk. Fra satelittmerking er det vist at klappmyss (Cystophora cristata) beiter i kantene til dyphavsområdene i Norskehavet (Folkow mfl. 1996, 2010).
Figur 9.10.3 Beitevandring for satelittmerket klappmyss i Norskehavet (Fra Folkow mfl. 2010)
Sjøfugl
Dyphavsområdene i Norskehavet er ikke typisk sjøfuglhabitat, men pelagisk overflatebeitende sjøfugl som havhest (Fulmarus glacialis) og andre stormfugler bruker også disse områdene (SEATRACK.seapop.no). Polarlomvi fra blant annet kolonien på Bjørnøya overvintrer i området avgrenset sør for Jan Mayen (SEATRACK.seapop.no), men forekomstene av sjøfugl er langt fra avgjørende for definisjonen av dette forslaget til nytt SVO-et.
10 - Resultater for Nordsjøen og Skagerrak
10.1 - Oversikt over særlig verdifulle områder i Nordsjøen/Skagerrak
Det er beskrevet fire særlig verdifulle områder, NS1-NS4, i Nordsjøen og Skagerrak (Fig. 10.1). Lokalisering, beskrivelser og miljøverdivurderingene for hvert av disse områdene er beskrevet i kapitlene 10.3 - 10.6.
Det er gjennom vurderingene foretatt nye avgrensninger av eksisterende SVO-er, bl.a. gjennom utvidelser av Kandidatområder og noen forskyvninger og sammenslåinger av eksisterende SVO-er. I tillegg er et nytt område (NS3) identifisert. Avgrensningene omtales nærmere under hvert område-kapittel.
Figur 10.1. Forslag til særlig verdifulle og sårbare områder (SVO) i Nordsjøen og Skagerrak. To områder ligger langs kysten av Sør-Norge (NS1-Boknafjorden og Jærstrendene, NS4-Ytre Oslofjord), ett ligger i deler av Norskerenna (NS3- Norskerenna), og ett består av flere mindre områder spredt over sentrale deler av Nordsjøen (NS2-Tobisfelt). I tillegg er foreslått SVO Kystsonen Norskehavet sør (NH7) vist i kartet siden deler av dette foreslåtte SVO-et ligger i Nordsjøen. Grensene for forvaltningsplanområdet er vist med brune linjer.
Forslag SVO-kode Miljøverdi-rapport 2021
SVO-kode Meld. St. 2019-2020
SVO-navn Meld. St. 2019-2020
Forslag SVO-navn Miljøverdirapport 2021
Foreslåtte endringer
NH7
NS7
Bremanger til Ytre Sula
Kystsonen Norskehavet sør
Inkludert i Kystsonen Norskehavet sør
NS1
NS4
Karmøyfeltet
Boknafjorden og Jærstrendene
Inkludert i Boknafjorden og Jærstrendene
NS1
NS8
Boknafjorden og Jærstrendene
Boknafjorden og Jærstrendene
Utvidet med Karmøyfeltet, og litt inn i fjorden.
NS2
NS11
Vikingbanken
Tobisfelt
Inkludert i Tobisfelt
NS2
NS5
Tobisfelt sør
Tobisfelt
Inkludert i Tobisfelt
NS3
NS2
Transekt Skagerrak
Norskerenna
Delvis inkludert i Norskerenna
NS3
NS3
Listastrendene og Siragrunnen
Norskerenna
Siragrunnen inkludert i Norskerenna
NS3
NS10
Skagerrak
Norskerenna
Inkludert i Norskerenna
NS4
NS6
Ytre Oslofjord
Ytre Oslofjord
Utvidet med Kandidat-området for bedre å dekke vinterbeite for sjøfugl
NS9
Korsfjorden
Utelatt, fyller kriteriene, men kun for kystmiljøverdier
NS1
Gytefelt makrell
Utelatt, fyller ikke kriteriene
Tabell 10.1 Samlet oversikt over foreslåtte SVO-koder i foreliggende rapport, sammenlignet med koder og navn på eksisterende SVO-er i Nordsjøen og Skagerrak (fra Meld. St. 20 (2019-2020)), navneforslag på de foreliggende SVOene og begrunnelse for forslag til endring i navn og grensesetting.
Akustikk, havsil som stiger opp fra bunnen. Foto: E. Johnsen, HI
10.2 - Generell beskrivelse av økosystem Nordsjøen og Skagerrak
Utstrekning
Nordsjøen, særlig den sørlige delen, er grunt, to tredjedeler grunnere enn 100 m. Et unntak er Norskerenna, som har dybder på over 700 m. Dybdeforholdene er viktige for sirkulasjonen, fordi topografien i stor grad bestemmer hvordan vannmassene beveger seg. Sokkelområdet er dekket av et flere kilometer tykt sediment lag avleiret fra de omkringliggende landområdene. Bunnen består ellers hovedsakelig av sand, skjellsand og grus på grunt vann, og mudder i de dypere områdene. Nordsjøen strekker seg fra nordøst av Storbritannia til den engelske kanal, den tyske og danske vestkysten, mens Skagerrak strekker seg mellom Norge og Danmarks vestkyst, fra Nordsjøen i vest til Kattegat og norskekysten lengst øst og nord.
Havstrømmer og vannmasser
Den nordlige delen av Nordsjøen er kjennetegnet av innstrømning av relativt salt atlanterhavsvann fra flere retninger (Fig. 10.2). Nord for Shetland dreier en del av Den norske atlanterhavsstrøm (ekstra tykk, rød pil) sørover, langs skråningen på vestre side (tykk, rød pil). Denne innstrømningen er relativt stabil og følger topografien langs hele Norskerenna. Hoveddelen av innstrømningen finner man på 100-200 m dyp. I områdene rett øst for Shetland og mellom Orknøyene og Shetland finner man også en relativt stabil innstrømning av atlanterhavsvann (tynne, røde piler). Her er dybdeforholdene grunnere slik at innstrømningen foregår nærmere overflaten. Dette medfører også at innstrømningen er avhengig av de dominerende vindforholdene og varierer mer i styrke og retning enn dypvannsinnstrømningen langs skråningen til Norskerenna. Hele vannsøylen i det nordvestlige Nordsjøen vest for Norskerenna er stort sett dominert av det salte atlanterhavsvannet.
Figur 10.2. Overflatestrømmer i Nordsjøen.
Det sentrale og sørlige Nordsjøen er relativt grunt med dyp hovedsakelig mindre enn 50 m. Man finner stort sett Nordsjøvannmasser med noe lavere saltholdighet enn atlanterhavsvannet. I vinterhalvåret vil vannmassene stort sett være gjennomblandet, mens det om sommeren dannes et varmere overflatelag på 10-20 m. Vinden i det sentrale Nordsjøen har mest innflytelse på strømretningen, og man har en østgående drift i dette området inn mot Jylland og Skagerrak. Sirkulasjon i området er ellers kjennetegnet ved en sirkulasjon langs kysten som stort sett går mot klokken (Fig. 10.2, tynne, grønne piler). Det er en liten kilde av atlanterhavsvann gjennom Den engelske kanal i sør, men dette utgjør kun noen få prosent av all tilførsel av atlanterhavsvann i Nordsjøen, og dette vannet blandes raskt inn i Nordsjøvannet. De største tilførselskildene av ferskvann kommer fra de store kontinentale elvene, som f.eks. Rhinen og Elben. Denne utstrømningen av ferskvann opprettholder en nordgående kyststrøm langs Jyllands vestkyst som stort sett dreier inn østover i Skagerrak sammen med driften fra den sentrale Nordsjøen.
Vannmassene i Skagerrak har sin opprinnelse i innstrømningen av atlanterhavsvann fra nordvestlige Nordsjøen, Nordsjøvann fra sentrale Nordsjøen, mindre salt kystvann fra sørlige Nordsjøen, brakkvann fra Østersjøen gjennom Kattegat og ferskvannstilførsel lokalt fra land. Sirkulasjonen går for det meste mo t klokken på grunn av denen østgående innstrømning av ulike vannmasser sør for Norskerenna. Overflatevannet blandes med Østersjøvann nord for Kattegat og dreier nordover langs Sveriges vestkyst og vestover langs sørlandskysten. Dette er starten på Den norske kyststrøm (tykk, grønn pil). Kyststrømmen er drevet av tetthetsforskjellene mellom det ferskere vannet nærmest kysten og det saltere vannet utenfor, men den er også i stor grad også vindstyrt. Atlanterhavsvannet som følger skråningen til Norskerenna på dansk side, fortsetter norsk side under kyststrømmen, til tross for innblanding av mindre salte vannmasser. Skagerrakbassenget har dyp over 700 m og har en slags terskel vest for Utsira som er rundt 270 m dyp. Bunnvannet i Skagerrak består stort sett av atlanterhavsvann, men det kan gå flere år mellom hver utskiftning. Enten kommer det inn tungt, saltere atlanterhavsvann langs sørvestlige skråning av Norskerenna som skyver vekk det eksisterende bunnvannet, eller så op pstår det episoder hvor vinteravkjølingen av overflatevann i sentrale Nordsjøen er så kraftig at det har sunket ned i og fylt opp Skagerrakbassenget. Sistnevnte utskiftning med avkjølt Nordsjøvann skjedde sist vinteren 2010, mens siste utskiftning med atlanterhavsvann skjedde våren 2018 (https://www.hi.no/hi/temasider/hav-og-kyst/klimaet-i-havet/klimastatus/nordsjoen-og-skagerrak).
Det kjemiske miljøet med næringssalttilførsel, oksygen, næringssaltkonsentrasjoner og havforsuringsparametere (pH, pCO2, alkalinitet, DIC og kalkmetning) er hovedsakelig drevet av fysiske forhold som variasjoner i kystrømmen, atlanterhavsvann og ferskvann, og biologisk produksjon (Omar mfl., 2019, Jones mfl. 2020). Forholdet mellom driverne vil variere fra år til år, som et resultat av vær og vind, og kan dermed tilsløre eventuelle endringer i næringssaltstilførsel som er viktig for biologisk produksjonen og øvrig kjemisk miljø som havforsuring som stammer fra økt CO2-opptak fra atmosfæren. Nordsjøen har relativt høyt CO2-opptak og spiller en rolle med å transportere CO2 fra luft til mellomdype lag i Nord-atlanteren, og dette er delvis styrt av bunntopografien som påvirker biologisk produksjon og respirasjon (Thomas mfl. 2004). I perioden 2011 til 2019 har pH og kalkmetning i dypvannet i Skagerrak minket med henholdsvis 0,013 og 0,019 per år, hvilket er mange ganger høyere enn en global middel-pH minking på 0,002 per år. Dette skyldes først og fremst innstrømming av atlanterhavsvann (Jones mfl. 2020).
Kort om økosystem og -ressurser
De dominerende pelagiske fiskene i Nordsjøen er gjennom hele året nordsjøsild og brisling. Silda gyter sør i Nordsjøen etter sommeren og beiter i norsk sektor. Det er også historiske gytefelt for norsk vårgytende sild i den nordlige delen av Nordsjøen, langs Rogalandskysten, som Boknafjorden og Karmøyfeltet. Makrell og hestmakrell er i hovedsak til stede om sommeren når de entrer Nordsjøen fra sør og nordvest. Disse bestandene har gytefelt fra Atlanterhavet vest av Storbritannia og inn i Norskehavet og Nordsjøen. De dominerende torskefiskene er nordsjøtorsk, nordsjøhyse, hvitting og nordsjøsei. Flyndrefiskene rødspette, gapeflyndre, sandflyndre, tunge og lomre er viktige bunnfisk. Tobis, øyepål og brisling er nøkkelarter i Nordsjøen både som fiskeressurs, men også som byttedyr for en rekke større fiskearter, pattedyr og fugl.
Bunnfaunaen varierer geografisk, og har sammenheng med vannmassepåvirkning og sedimentenes sammensetning. Også dyp, temperaturvariasjon og strømforhold virker inn på artssammensetningen, bl.a. fordi de fleste bunnlevende artene har larver som transporteres med vannmassene (Heip og Craeymeersch 1995). Bunnfaunaen er viktig som føde for fisk som torsk, hyse og flyndre, i tillegg til å ha betydning i omsetningen av sedimentert organisk materiale (se Ramirez-Llodra mfl. 2016). Sammensetningen av evertebrater som lever på bunnen i Nordsjøen viser et skille mellom en sørlig artssammensetning dominert av frittlevende organismer, mens den nordlige komponenten er mer dominert av fastsittende bunnorganismer. Tallet på arter er høyere i nord enn i sør og biomassen er større nær kysten enn lengre ute. Artsmangfold og -tetthet er dessuten koblet til bunntype og ser ut til å påvirkes av endringer i havtemperaturen. Forekomst av habitatdannende arter i afotisk sone er lite kjent til tross for at Nordsjøen ansees å være et av de best studerte havområder i verden. Forekomstene av korallrev i Østfold (Hvaler/Tisler) er relativt godt kjent (Järnegren og Kutti 2014), men for områdene mellom Rogaland og Østfold er det ingen sikre registreringer. Kunnskap om utbredelse av andre koraller (hornkoraller), sjøfjær og svamp er også lite kjent. Imidlertid har Havforskningsinstituttet de senere år startet registrering av sjøfjær og hornkoraller i bifangst under det årlige reketråltoktet.
Fordelingen av sjøfugl varierer geografisk, der flere ulike bestander oppholder seg i Nordsjøen gjennom året. Skagerrak og Kattegat er dessuten kjente overvintringsområder for alkefugl fra Storbritannia (Systad mfl. 2019). Om vinteren og i hekketiden oppholder de fleste sjøfugler seg på kysten eller i relativt kystnære farvann. Flere pelagisk beitende arter - som havsule, havhest, krykkje og alkefugler - har beiter i åpent hav gjennom det meste av vinterhalvåret (Systad mfl. 2019, Fauchald mfl. 2006). Hekkefuglene samler seg ved hekkekolonien på ettervinteren så tidlig som februar for de fleste fuglefjellsartene (Systad mfl. 2019). Under svømmetrekket den første måneden etter hekkingen kan det være et betydelig antall flygeudyktige alkefugler i åpent hav i Nordsjøen og på bankene. Også kystnære arter som ærfugl oppholder seg lengst mulig ut fra kysten under fjærskiftet på høsten, da de er flygeudyktige og særlig sårbare av den grunn. Nordsjøen er ellers et kjent overvintringsområde for store måker fra hele Nord-Atlanteren (Fauchald mfl. 2006).
Vågehval, nise og kvitnos opptrer tallrikt i Nordsjøen. Disse finnes over store deler av havområdet og beiter på fisk som tobis, sild og makrell, men også på dyreplankton. Nise og kvitnos må regnes som relativt stedegne arter innen Nordsjøområdet, mens vågehvalen foretar næringsvandringer om sommeren til nordområdene fra vinteroppholdssteder på sørligere breddegrader. Nordsjøen er det viktigste habitatet for denne arten i østlige Atlanterhavet (Hammond mfl. 2017). Variasjon i fordeling av hval knyttes til endringer i byttedyrsituasjonen. Tumlere og en rekke andre hvalarter er også til stede i Nordsjøen (Weir 2001). To arter sel, havert og steinkobbe, har fast tilhold året gjennom i Nordsjøen. Streifdyr av arktiske seler, særlig klappmyss, forekommer.
Plankton i forvaltningsområdet
Planteplankton som element har en økosystemfunksjon, som basis for marine næringskjeder. Planteplankton er til stede i alle våre havområder og vil gjennomgå markante endringer gjennom året både når det gjelder biomasse og sammensetning. Planteplankton vil også vise stor variasjon mellom årene (Skogen og Moll 2000). Planteplanktonartene vil være til stede over store områder (større enn de eksisterende og foreslåtte SVO-ene), har høy vekstrate i perioder og vil til enhver tid utnytte de fysiske og kjemiske forholdene. På grunn av elementet «Planteplankton» sin store variasjon i tid og rom og deres fritt flytende pelagiske levesett, er det vanskelig å benytte i en utredning omkring miljøverdier inne i enkeltområder.
Nordsjøen har generelt høy primærproduksjon, produksjonen av planteplankton. Dette er i stor grad på grunn av små dybder og den effektive omrøringen av vannmasser som bringer næringsrikt bunnvann opp i lyset. Eutrofiering (overgjødsling) har medført problematisk høye algekonsentrasjoner i enkelte områder. Dette har minket de senere årene, sannsynligvis pga. lavere tilførsel til hele Nordsjøen av næringssalter fra de store europeiske elvene og andre landbaserte kilder. Primærproduksjonen foregår i de øvre vannlagene, ned til 20-30 m i atlanterhavsvann. Konsentrasjonen av planteplankton er størst under våroppblomstringen, med topp i mars-april ved kysten og april-mai i åpne havområder. Høstoppblomstringen har en topp i september - oktober, men er av mindre omfang. Den dominerende algegruppen i Nordsjøen, som i alle andre nordiske havområder, er kiselalgene (diatomeene), som er særlig viktige under våroppblomstringen. Eventuelle høstoppblomstringer vil kunne domineres av fureflagellater og/eller kiselalger avhengig av de fysiske og kjemiske forholdene. Små flagellater er til stede mer eller mindre hele året, men er mest tallrike i sommerperioden. Artssammensetningen og tettheten om våren er antatt å ha en stor betydning for hvor mye energi som bindes og som blir tilgjengelige for planteetere og predatorer resten av året. Våroppblomstringen vil også bidra med betydelig karbontransport til dypvannet gjennom sedimentasjon. Endringer i havklima vil kunne få betydning for algeutbredelsen ved at mer varmekjære arter etablerer seg i norske farvann. Generelt vil det være høyere produksjon og større biomasse av planteplankton i de kystnære områdene, områder som på grunn av topografiske forhold gir oppstrømning av dypvann eller fronter mellom ulike vannstrømmer/vannmasser. Planteplanktonproduksjon vil finne sted innen hele havområdet og transporteres med havstrømmene fra et område inn i et annet. Mindre områder (inkludert eksisterende og foreslåtte SVO-er) vil påvirkes av lokal produksjon og produksjon i utenforliggende områder.
Dyreplankton er viktig næringsgrunnlag for flere kommersielle fiskearter, sjøpattedyr og sjøfugl. Viktige arter er raudåte (Calanus finmarchicus), krill og amfipoder. Den vanligste hoppekrepsen nord i Nordsjøen er raudåte som er svært viktig for dyreplanktonspisende fisk. Sesongmessig produksjonssyklus og produktivitet av C. finmarchicus varierer mellom år, og mye tyder på at bestanden i Nordsjøen og Skagerrak avhenger av tilførsel fra Norskehavet (Reid mfl. 2003, Gao mfl. 2021). Sør i Nordsjøen er det små altetende arter som dominerer, med kort livssyklus og flere generasjoner hvert år. Dyreplanktonet ved samtlige SVO-er i Nordsjøen/Skagerrak avspeiler den generelle utviklingen i Nordsjøen som hovedsakelig er drevet av klimatiske variasjoner. De siste 50 årene har man observert en rekke endringer i både mengde og artssammensetning av dyreplankton i Nordsjøen og Skagerrak som følge av stigende havtemperaturer (Beaugrand mfl. 2002, O’Brien 2013, Edwards mfl. 2016). Det har skjedd en gradvis økning i forekomst og utbredelse av sørlige, varmekjære arter, mens kaldtvannsarter (boreale arter) har avtatt. Mengden av raudåte (C. finmarchicus) er redusert med 70 prosent siden 1960-tallet. Samtidig har den varmekjære arten Calanus helgolandicus, utvidet sin nordlige grense. Raudåte gyter tidlig på våren, slik at maksimumstettheten av raudåte sammenfaller med tidspunktet for maksimumstetthet av pelagiske fiskelarver, som beiter på disse. En økning i dyreplanktonarter med senere gytetidspunkt (for eksempel C. helgolandicus) kan gi et misforhold mellom fiskelarver og deres byttedyr (Arneberg mfl. 2018). Skiftet i retning av mer sørlige arter bidrar også til en generelt lavere produksjon av dyreplankton (Edwards mfl. 2016). I tillegg til temperatur har innstrømming av atlanterhavsvann tidlig på året stor betydning for å opprettholde bestanden av raudåte i nordlige Nordsjøen og Skagerrak. Pseudocalanus regnes for å være den viktigste hoppekrepsen i næringskjeden i Skagerrak/Nordsjøen, etter raudåte. Denne har vist en nedgang med 80 prosent fra 2003 til 2009 og har vært under gjennomsnittet siden (Bakketeig mfl. 2016, Arneberg mfl. 2018, O’Brien 2013) Nedgangen er særlig fremtredende på sensommeren, slik at den vanlige «sekundære» oppblomstringen av hoppekreps i august–september er redusert de siste årene.
Økte havtemperaturer øker også overlevelsesevnen til fremmede arter som ikke hører naturlig hjemme i systemet, for eksempel Amerikansk ribbemanet Mnemiopsis leidyi som nå er vanlig langs hele kysten av Skagerrak og Nordsjøen (Hosia og Falkenhaug 2015, Bakketeig mfl. 2017, Artsdatabanken 2018).
10.3.1 - Pågående og framtidige endringer i Nordsjøen og Skagerrak
Generell informasjon om klimaendringer, med konsekvenser for miljøverdigruppene, samt forsuring er gitt i kap. 5.1. Trendene relatert til temperaturøkning i Nordsjøen og Skagerrak kan sees i sammenheng med det som registreres både langs norskekysten og i Norskehavet/Atlanterhavet. Nordsjøen og Skagerrak har hatt en markert temperaturstigning, som er forventet å fortsette (Arneberg mfl. 2018, Engelhard mfl. 2014).
Plankton
Dyreplanktonet i området avspeiler den generelle utviklingen i Nordsjøen som hovedsakelig er drevet av klimatiske variasjoner. Dersom havklima fortsetter å utvikle seg mot et varmere hav, kan man forvente en videre økning av varmekjære arter med lavere produksjon og endret sesongmønster (Arneberg mfl. 2018, Edwards mfl. 2016). I tillegg kan man forvente økt forekomst av fremmede arter (Artsdatabanken 2018).
Raudåte (Calanus finmarchicus) overvintrer på store dyp og høyere temperaturer vil føre til økt metabolisme, og forkorte overvintringsperioden. Dette gjør populasjonen i dette området ekstra sårbar for endringer i miljøforhold. Stigende temperaturer samt lave oksygennivåer i Norskerenna som følge av klimaendringer (miljostatus.no) vil redusere mulighetene for overvintring av raudåte i dette området.
Med stigende havtemperatur som følge av klimaendringer er det forventet en fortsatt økning i varmekjære arter, og en nedgang i forekomst av Calanus finmarchicus. Dette vil føre til redusert sekundærproduksjon, lavere total zooplanktonbiomasse samt et endret sesongmønster. Reduksjon i zooplanktonbiomasse vil særlig være merkbar i vårperioden (april-mai) på grunn av nedgangen i raudåte.
Stigende havtemperaturer øker også sannsynligheten for at flere fremmede arter etablerer seg i havområdet. Økt havtemperatur forventes å øke reproduksjon hos den fremmede arten Mnemiopsis leidyi (Amerikansk lobemanet) som er utbredt i kyststrømmen mellom Oslofjord og Trøndelag.
Klimaendringer forventes å øke ferskvannsavrenning fra land, som kan gi en formørkning av kystvannet. Reduserte lysforhold vil påvirke hele det planktoniske økosystemet, og kan gi en økning i ikke-visuelle predatorer som f.eks. maneter (Aksnes mfl. 2009).
Fisk
Studier peker på at fiskesamfunnet i Skagerrak endres i perioder med lavere eller høyere temperaturer (Barceló mfl. 2016). Noen fiskearter vil kunne antas å påvirkes negativt av klimaendringene/ havoppvarmingen. Høy overflatetemperatur i sommermånedene gjør at torsk (Gadus morhua) oppholder seg i dypere vannmasser (Freitas mfl. 2016). Perioder med mindre tilgjengelig dyreplankton i form av kaldtvannsarten raudåte er korrelert med svakere rekruttering av torsk i kystnære områder (Johannessen mfl. 2012).
Havsil vil bli påvirket grunnet to forhold. For det første vil varmere vann påvirke dyreplanktonsammensetningen og mengden egnet dyreplankton for havsil (Van Deurs mfl. 2009). Dette vil kunne påvirke både larveoverlevelse, veksthastighet og gytebiomasse av havsil. For det andre vil nedgravd havsil få en høyere metabolisme i varmere vann som reduserer sannsynlighet for vinteroverlevelse og også medfører lavere mengde energireserve til gyting (Van Deurs mfl. 2009, Eliasen 2013). I tillegg viser Johnsen mfl. (2017) at større tettheter med havsil bidrar til stimformasjonene og predasjonsunnvikelse. Totalt sett, grunnet den viktige rollen havsil har i Nordsjøen, betyr dette at tobisfeltene vil få en enda større verdi for å kunne sikre et bærekraftig økosystem.
På grunn av god tilførsel av atlantisk dypvann representerer områdene i Skagerrak, som foreslått SVO Ytre Oslofjord, kystnære «miljørefugier» (Keppel mfl. 2012) som vil kunne tilby gode miljøforhold for fiskebestander og biologisk mangfold inn i en tid med større variabilitet i kystvannet (se Frigstad mfl. 2020).
Dyreplankton, mest hoppekreps, i tobisfelt. Foto: HI
Bunnfauna
Bunnfaunaen varierer geografisk, og har sammenheng med vannmassepåvirkning og sedimentenes sammensetning. Også dyp, temperaturvariasjon og strømforhold virker inn på artssammensetningen, bl.a. fordi de fleste bunnlevende artene har larver som transporteres med vannmassene (Heip og Craeymeersch 1995). Artsmangfold og -tetthet ser ut til å påvirkes av endringer i havtemperaturen.
Sjøpattedyr
Sjøpattedyrene i Nordsjøen og Skagerrak er sel og hval med en bred temperaturtoleranse, men de vil bli negativt påvirket om beitegrunnlaget svekkes (Nilssen og Bjørge 2019).
Sjøfugl
Sjøfuglene i Nordsjøen og Skagerrak er, i likhet med fisk, avhengig av tilstrekkelig tilgang til næring. Om plankton og fiskebestandene endres og svekkes, vil dette ha en negativ påvirkning på hekkesuksess og vinteroverlevelse (Henriksen og Hilmo 2015).
10.3.2 - Løpende kartleggings- og overvåkingsaktivitet
Nordsjøen
Foreslått SVO
Plankton
Fisk
Bunn-samfunn
Meso-pelagisk fauna
Sjø-fugl
Sjø-pattedyr
Frekvens
Nasjonale program
Mareano-programmet (Mareano.no)
NS1-NS4
X
Årlig, rullerende
SEAPOP-programmet (Seapop.no)
NS1-NS4
X
Årlig
SEATRACK-programmet (seatrack.seapop.no)
NS1-NS4
X
Årlig
Havforskningsinstituttet
Overvåkingstokt
Overvåking bunnfisk og pelagisk fisk (IBTS-tokt)
NS2
X
Årlig
Overvåking sild og brisling i Nordsjøen
NS3-NS4
X
Årlig
Akustisk gyting sei
NS2
X
Årlig
Akustisk tobistokt Nordsjøen
NS2
X
Årlig
Økosystemtoktet Nordsjøen/Skagerrak
NS2-NS3
X
X
X
X
X
Årlig
Kysttorsk sør, vinter
NS1, NS4
X
Årlig
Overvåking reke sør, Nordsjøen-Skagerrak
NS3-NS4
X
X
Årlig
Eggtokt, kyst
NS1, NS4
X
Årlig
Kystsel
NS1, NS4
X
Hvert 5. år
Høstundersøkelser Skagerrak
NS4
X
Årlig
Strandnot, feltarbeid
NS4
X
X
Årlig
Oseanografiske snitt
Utsira V
NS1-NS2
X
4x årlig
Torungen-Hirtshals
NS3
X
9-12x årlig
Jomfruland-Skagen
NS4-NS4
X
12-22x årlig
Torbjørnsskjær-Jomfruland
NS4-NS4
X
12-22x årlig
Tabell 10.2. Tabellen viser et utvalg av løpende kartleggings- og overvåkingsprogram, tokt og oseanografiske snitt, der foreslåtte SVO-er i Nordsjøen helt eller delvis inngår i kartleggingen/overvåkingen. Foreslåtte SVO-er og miljøverdier som kartlegges/overvåkes er oppført i tabellen.
GO Dannvig klar til Nordsjøtokt. Foto: HI
10.4 - Forslag til endret SVO Boknafjorden og Jærstrendene (NS1)
10.4.1 - Sammendrag
Områdebeskrivelse og strukturer
Foreslått endret SVO Boknafjorden og Jærstrendene (NS1) har stor variasjon i geologi og økologi – fra åpne sjøarealer mot Nordsjøen og Norskerenna i vest, og til grunne tareskogsområder. Kyststrømmen fra Skagerrak har en betydelig påvirkning samtidig som det i en stor grad er åpent eksponert mot nordlige Nordsjøen. Topografi, fjorder, øyer og nærheten til Norskerenna, gir opphav til lokale strømmønstre og retensjonsområder.
Miljøverdier
Utveksling av vann fra kyststrømmen og atlanterhavsvann fra de dypere lagene av Norskerenna bidrar til høyt mangfold av dyreplankton. Den nære tilknytningen til Norskerenna fører også til relativt høy biomasse av dyreplankton sammenlignet med sentrale Nordsjøen. Området har høyere forekomst av raudåte sammenlignet med områdene lenger vest.
Kun gytefelt for kystnær torsk, og norsk vårgytende sild er kartfestet i det foreslåtte SVO-et, men det er funnet nygytte egg fra torskefisk som torsk, hyse, sei og hvitting, samt flyndrefisker. Sanddynene langs Jærstrendene er særlig gode gytefelt for flatfisk. Historisk var dette viktige gytefelt for sild, men selv om de ikke er like viktige i dag, kan de igjen bli viktige.
Området er et viktig sjøfuglområde med relativt store hekkebestander av en underart av storskarv, toppskarv, ærfugl og store bestander av sildemåke og gråmåke. Det er også viktig hekkeområde for den i Norge sterk truede (EN) makrellterna (Sterna hirunda) i juli. Det foreslåtte SVO-et har sørligste hekkeplass for krykkje, lomvi, lunde i Norge. Jærkysten er i tillegg viktig som overvintringsområde for lommer, dykkere og havdykkender fra store deler av Arktis og Fennoskandia. Dette er også eneste kjente faste yngleområde for havert sør for Stadt og yngle- og hårfellingsområde for steinkobbe.
Karmøyfeltet er også viktig i økosystemsammenheng på grunn av store forekomster av reke – der reke er en nøkkelart i økosystemet. Svamper er funnet i større ansamlinger i Karmøyfeltet og det er forekomster av sårbare arter som den endemiske bambuskorallen i norske farvann.
Figur 10.3.1. Kart over foreslått endret SVO Boknafjorden og Jærstrendene (NS1). Grensene for foreslått SVO-etdet foreslåtte SVO-et er vist med tykk rød strek, mens grensen for det nærliggende forslaget til nytt SVO Norskerenna (NS3) er vist med tynn rød strek. Grensen for forvaltningsplanområdet inn mot kysten er vist med brun linje. Kartet viser utbredelsen av utvalgte miljøverdier med betydning for området. Miljøverdier knyttet til fisk (gytefelt), sjøpattedyr og sjøfugl er vist.
Makrellterne. Foto: Jan de Lange, HI
10.4.2 - Vurdering i forhold til EBSA-kriteriene – Forslag til endret SVO Boknafjorden og Jærstrendene (NS1)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til endret SVO Boknafjorden og Jærstrendene (NS1)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet/sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Sjøpattedyr
Sjøfugl
Den sørligste kjente ynglekoloni av havert er ved Kjørholmane naturreservat i Sola kommune (Nilssen og Bjørge 2019).
Sørligste hekkeplass for lomvi og lunde i Norge med nokså små bestander (Gasbjerg mfl. 2011).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Fisk
Sjøfugl
Sjøpattedyr
Kun gytefelt for kysttorsk i indre deler av Boknafjorden og fjordene i Ryfylke (Aglen mfl. 2016) og norsk vårgytende sild vest og sør for Karmøy (Aglen mfl. 2012) er kartlagt, men nygytte egg fra torskefisk som torsk, hyse, sei og hvitting, samt flyndrefisker (Aglen mfl. 2016) er funnet i området. Forekomst av reker, en nøkkelart i økosystemet, har lenge vært kjent som en viktig art, særlig på Karmøyfeltet (Hjort og Ruud 1938).
Eneste kjente faste yngleområde for havert sør om Stad. Yngle- og hårfellingsområde for steinkobbe (Nilssen og Bjørge 2019).
Området er viktig vinterbeiteområde for kystnære sjøfugl som ærfugl, storskarv, toppskarv, gråmåke og sildemåke (seapop.no, Gasbjerg mfl. 2011). Det er relativt store hekkebestander av gråmåke og sildemåke i området, og det har vært et av de viktigere hekkeområdene for makrellterne i Norge (Systad mfl. 2007). Toppskarv og storskarv av den kontinentale underarten er nokså tallrike i dette området (Lorentsen mfl. akseptert).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Sjøfugl
Området er et av de viktigere hekkeområdene for makrellterne i Norge (Systad mfl. 2007), men bestanden er nesten utradert på grunn av hekkesvikt de siste tiårene (Systad pers medd.). Ellers er det flere rødlistede sjøfugl som overvintrer i området, f.eks. svartand (nær truet-NT), bergand, sjøorre og horndykker (sårbar-VU) (Systad mfl. 2007).
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Bunnsamfunn
Sjøfugl
Svamper er funnet i større ansamlinger i Karmøyfeltet og det er forekomster av sårbare arter som den endemiske bambuskorallen i norske farvann (Mareano.no).
For sjøfugl er området sårbart ut fra artsrikheten, flere rødlistede arter i et eksponert område med mye menneskelig aktivitet (Gasbjerg mfl. 2011, Systad mfl. 2007). Dette gjelder i stor grad kystnære arter.
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Plante-plankton
Dyreplankton
Fisk Sjøpattedyr
Området vil i perioder ha oppstrømning av dypvann på innsiden av kyststrømmen som gir grunnlag for høyere primærproduksjon (Aure og Stigebrandt 1989), men det foreligger lite data som kan benyttes til å vurdere hvorvidt området er mer produktivt enn omkringliggende havområder.
Området ligger i nær tilknytning til den dype Norskerenna, noe som fører til relativt høy biomasse av dyreplankton sammenlignet med sentrale Nordsjøen (Bakketeig 2017). Området har høyere forekomst av raudåte sammenlignet med områdene lenger vest (Strand mfl. 2020).
Nygytte egg fra torskefisk som torsk, hyse, sei og hvitting, samt flyndrefisker (Aglen mfl. 2016) og gytefelt for norsk vårgytende sild vest og sør for Karmøy (Aglen mfl. 2012), viser at det er høy produktivitet i området.
Tilgang til ungekastingsplasser og fisk fra fjord, kyst og hav bidrar til både havert og steinkobbekolonier (Nilssen og Bjørge 2019).
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Dyreplankton Bunnsamfunn Sjøpattedyr
Sjøfugl
Utveksling av vann fra kyststrømmen og atlanterhavsvann fra de dypere lagene av Norskerenna bidrar med høyt mangfold av dyreplankton som hoppekreps, krill og pelagiske reker (Bakketeig mfl. 2017).
Svamper er funnet i større ansamlinger i Karmøyfeltet og det er forekomster av sårbare arter som den endemiske bambuskorallen i norske farvann (Mareano.no)
Norges sørligste yngleplass for havert bidrar til at det kan antas at det generelle biologiske mangfoldet er høyere i dette området enn i omkringliggende områder, uten at det er dokumentert.
Viktig sjøfuglområde med relativt store hekkebestander og overvintringsområde for en rekke sjøfugl, der nordlige og sørlige arter møtes. Området har en høy artsrikhet gjennom hele året (seapop.no).
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
10.4.3 - Lokalisering
SVO Boknafjorden og Jærstrendene er tidligere definert i Meld. St. 20 (2019-2020), men det foreslått endrete SVO-et er utvidet i nord for å dekke deler av det eksisterende SVO Karmøyfeltet som fagekspertgruppen nå foreslår å fjerne som eget SVO. Området er i tillegg utvidet noe innover i fjorden for å dekke viktige, godt kartlagte områder for sjøfugl og sel.
Boknafjorden er en fjord i Rogaland som trenger seg inn mot nordøst mellom Jæren og Karmøy, og Jærstrendene er et langstrakt strandområde langs kysten av Jæren i Rogaland. Boknafjorden og Jærstrendene som særlig verdifullt område strekker seg fra Jærstrendene i sør til og med Utsira og Karmøy i nord, og inkluderer områder utenfor kysten og et stykke innover i Boknafjorden (Fig. 10.3.1).
Foreslått endret SVO Boknafjorden og Jærstrendene dekker det marine verneområdet Jærkysten og flere mindre naturvernområder.
10.4.4 - Introduksjon til området
Foreslått endret SVO Boknafjorden og Jærstrendene er et særegent område med store grunne partier med sand- og steinbunn. Området er preget av lavsaltholdig kystvann med kyststrømmen som medfører relativt sterk, persistent strøm sørfra, innslag av saltere atlanterhavsvann lenger til havs samt noe påvirkning av fjordvann fra utvekslingen med de innerste delene av Boknafjorden (Sætre 2007, Fig. 10.3.2). I vestlandsfjorder gir primærproduksjon på våren høy pH og aragonittmetning (indikator for kalsifisering) og lavt uorganisk karbon (DIC). Med økt dyp øker CO2 og gir lav pH og metningsgrad grunnet respirasjon av organisk materiale (Findlay mfl. 2008, 2013, 2014, Kitidis mfl. 2012, Jones mfl. 2018). Det nedbrytbare materialet kommer opp til overflaten igjen ved vertikal omblanding i form av næringssalter til neste års produksjon og ved at CO2 transporteres fra dypet til overflaten.
Figur 10.3.2. Gjennomsnittlig overflatestrøm (piler) og bunndyp (farger) i foreslått SVO Boknafjorden og Jærstrendene og omkringliggende områder.
Vindforholdene gjennom året har stor innvirkning på hvordan dynamikken i overflaten endrer seg samt hvilke vannmasser som gjør seg mest gjeldende (Sætre 2007). Det fysiske miljøet under kyststrømmen på rundt 50 m og nedover er sterkt preget av atlanterhavsvann og vil derfor være utsatt for mer langperiodiske variasjoner i saltholdighet og temperatur. I tillegg til stor variasjon i strøm er dette området også utsatt for en del vind og derfor høy bølgeaktivitet. Utveksling av vann fra kyststrømmen bidrar med næringssalter og oksygen til fjordene og indikerer høy primærproduksjon (Aure og Stigebrandt 1989). CO2-opptak fra atmosfæren og havforsuringstilstanden er sterkt knyttet til vannutveksling og ferskvannstilførsel. Økt ferskvann vil gi økt havforsuring med lav aragonittmetning (Chierici og Fransson 2009). Store deler av området er svært eksponert og er utsatt for betydelig vertikal omrøring av vannmassene. I tillegg er det en vesentlig adveksjon av vannmasser både langs kysten som en del av kyststrømmen og på tvers som en del av vannutvekslingen mellom kysten og fjordene. Atlanterhavsvannet vil også gjøre seg gjeldende inn mot kysten og under overflatelaget.
10.4.5 - Beskrivelse av miljøverdiene
Plankton
Området vil i perioder ha oppstrømning av dypvann på innsiden av kyststrømmen som gir grunnlag for estimert høyere produksjon (Aure og Stigebrandt 1989). I den midtre delen av det foreslåtte SVO-et har det pågått overvåkning av planteplanktonbiomasse og artssammensetning gjennom programmet ØKOKYST (eks. Kaurin mfl. 2020) og tidligere overvåkingsprogram (Norderhaug mfl. 2012). For Boknafjord-systemet er det vist at tilførselen av ferskvann er viktig for planteplanktonproduksjon, gjennom økt stabilitet og næringssaltdynamikk (Erga 1989). Det foreligger lite data som kan benyttes til å vurdere hvorvidt SVO-et er mer produktivt enn omkringliggende havområder.
Det foreligger lite informasjon om dyreplankton fra Boknafjorden. Advektive prosesser fører til stor variabilitet i dyreplanktonforekomster, og det er registrert relativt høye konsentrasjoner av zooplankton i de indre delene av fjorden sammenlignet med ytre områder (Johannessen mfl. 1995). De ytre områdene ved Karmøy har en relativt høy biomasse av dyreplankton sammenlignet med sentrale Nordsjøen (6-13 g tørrvekt per m2) i april/mai (Bakketeig mfl. 2017 og Falkenhaug upubl. data). Dette er basert på overvåkningsdata fra Havforskningsinstituttets faste snitt «Utsira-StartPoint». Området har også høyere forekomst av raudåte (Calanus finmarchicus) sammenlignet med områdene lenger vest (Strand mfl. 2020). Dette har sammenheng med nærheten til det dype området i Norskerenna (250 m), der raudåte og makroplankton (krill, pelagiske reker mm) forekommer i de dypere lagene (Falkenhaug upubl. data). Man antar Norskerenna er en viktig transportvei for raudåte som advekteres inn i Nordsjøen med innstrømmende atlanterhavsvann (Madden mfl. 1999).
Boknafjorden har god vannkontakt med utenforliggende havområder, og det er grunn til å anta at dyreplanktonet i dette området har gjennomgått de samme storskala endringene som i øvrige Nordsjøen. med en reduksjon i mengden raudåte (C. finmarchicus) og en økning i varmekjære arter som C. helgolandicus. Dette har ført til lavere sekundærproduksjon og endret sesongmønster (Edwards mfl. 2016).
Den fremmede arten Mnemiopsis leidyi (Amerikansk lobemanet) har vært tallrik i kyststrømmen fra og med år 2005 (Hosia og Falkenhaug 2015). Arten kan ha stor innvirkning på planktonsamfunnet gjennom høy predasjon på zooplankton som medfører økning i mengden av planteplankton (Granhag mfl. 2011).
Fisk
Sild (Clupea harengus) gyter eggene sine på bunnen, og er derfor avhengig av et spesielt bunnsubstrat. Norsk vårgytende sild har gytefelt vest og sør for Karmøy, og i perioden 1930-1959 var disse viktige gytefelt i mars og april (Aglen mfl. 2012). I perioden 1960-1988 ble det ikke observert gyting her, men i 1989-2002 og 2009-2010 ble det igjen observert noe gytende sild her. Disse gytefeltene kan igjen bli viktige for norsk vårgytende silda
Gytefelt for kystnær torsk (Gadus morhua) er kartlagt i indre deler av Boknafjorden og fjordene i Ryfylke gjennom nasjonalt program for kartlegging av marine naturtyper (Aglen mfl. 2016). Det er funnet nygytte egg fra torskefisk som torsk, hyse, sei og hvitting, samt flyndrefisker (Aglen mfl. 2016). Disse gytefeltene er ikke kartfestet ut over gytefelt for kysttorsk. Enkelte gytefelter i midtre deler av fjordene vil kunne avgi egg og larver til ytre områder, men disse vil i hovedsak drive med kyststrømmen nordover forbi Karmøy. Fisk som bunnslår og vokser opp i kystsonen av Jærstrendene vil ha sin opprinnelse i gytefelt lengre øst (Huserbråten mfl. 2018).
Bunnsamfunn
De dypere og sentrale deler av Boknafjorden er hovedsakelig bløtbunn. Sidene av fjorden og lokale terskelområder har større innslag av hardbunn. Geografisk spesifikk informasjon om bunnfauna eksisterer først og fremst i form av en rekke rapporter fra miljøundersøkelser i forbindelse med industritiltak (resipientundersøkelser, akvakultur-utbygging etc). Disse undersøkelsene er for det meste utført med bunngrabb og beskriver infauna (organismer som lever nede i sedimentene). Så langt vi kjenner til eksisterer det ingen oversikt og sammenstilling av slike undersøkelser fra området. I 2020 startet prosjektet «Marine grunnkart i kystsonen», et samarbeid mellom Kartverket, Norges geologiske undersøkelse og Havforskningsinstituttet. Som del av dette prosjektet er det gjennomført flere tokt med visuell kartlegging. Resultater fra disse undersøkelsene vil bli publisert i rapportform og gjennom karttjeneste i løpet av 2021. I Midtsund, innenfor Sandsfjorden, nordvest for SVO Boknafjorden og Jærstrendene ligger det eneste kjente korallrevet i Rogaland. Dette området er etablert som verneområde for korallrev (Mareano.no).
Karmøyfeltet er også viktig i økosystemsammenheng på grunn av store forekomster av reke (Pandalus borealis) – der reke er en nøkkelart i økosystemet. Svamper er funnet i større ansamlinger i Karmøyfeltet.
Dypvannsreker er nærmere omtalt under SVO Norskerenna, som fungerer som et reservoar for bestanden (Kap. 10.5.5). Hjort og Ruud (1938) beskriver Høgsfjorden (en av de sørligste fjordene i Boknafjordområdet) som en av de fineste rekefjorder i landet.
Sjøpattedyr
Haverten (Halichoerus grypus)er flokkdyr som danner kolonier, særlig i forbindelse med ungekasting (fødsel) og parring (september–desember) og hårfelling (februar–april). I resten av året er de spredt langs store deler av kysten. I SVO Boknafjorden og Jærstrendene er det en ynglekoloni av havert ved Kjørholmane naturreservat i Sola kommune (Nilssen og Bjørge 2019).
Steinkobben (Phoca vitulina), vår mest tallrike kystsel, er en liten selart. Steinkobbene samles på land i forbindelse med forplantning (juni-juli) og hårfelling (august), og de legger seg helst opp på litt beskyttede lokaliteter i skjærgården (skjær og sandbanker som tørrlegges ved fjære sjø) (Bjørge og Øien 1999). Noen av de største koloniene er ved Kvitsøy og Randaberg kommuner i SVO Boknafjorden og Jærstrendene. Fra andre del av september til tidlig juni, er det næringssøk som har høyeste prioritet. I denne perioden vil de oppsøke steder med god tilgang på mat, ofte mange titalls kilometer fra stedet der de ble født.
Sjøfugl
Området er et viktig sjøfuglområde med relativt store hekkebestander av underarten Phalacrocorax carbo sinensis av storskarv, toppskarv (Phalacrocorax aristotelis), ærfugl (Somateria mollissima) og store bestander av sildemåke (Larus fuscus) og gråmåke (Larus argentatus) (Fig. 10.3.3). Jærkysten er i tillegg viktig som overvintringsområde for lommer, dykkere og havdykkender fra store deler av Arktis og Fennoskandia (Systad mfl. 2007). Kjørholmane er Norges sørligste hekkeplass for lomvi (Uria aalge) og lunde (Fratercula arctica) (Gasbjerg mfl. 2011), men kolonien er kun på noen titalls par. Også krykkje (Rissa tridactyla) har sin sørligste hekkeplass i Norge her, i Nevlunghavn på Karmøy (Seapop.no). Forekomster av havhest (Fulmarus glacialis) i området er forsvunnet på 2000-tallet (Seapop.no).
Figur 10.3.3. Kart viser fordeling av pelagiske sjøfuglarter (A: lomvi, lunde, krykkje, alke), kystnære fiskespisende og bentisk beitende arter (C: toppskarv, storskarv, ærfugl) og kystnære overflatebeitende arter (B: sildemåke, rødnebbterne, og D: tyvio og gråmåke) i foreslått endret SVO Boknafjorden og Jærstrendene og nærliggende områder. Skravert felt i C) viser beiteområder for kystnære fiskespisende og bentisk beitende arter. Grensene for det foreslåtte SVO-et er markert med rød strek.
10.5 - Forslag til endret SVO Tobisfelt (NS2)
10.5.1 - Sammendrag
Områdebeskrivelse og strukturer
Forslag til endret SVO Tobisfelt (NS2) har et særegent bunnhabitat bestående av grov sand og fin grus med gode oksygenforhold på ikke alt for store dyp. Feltene er spredt utover Nordsjøen, men det største arealet samlet sett ligger i norsk økonomisk sone. Det spesielle habitatet dannes i områder med særlig sedimentkvalitet på bunnen, som gjør det mulig for tobis (havsil) å grave seg ned.
Miljøverdier
Tobisfeltene er gyte- og leveområde for tobis (dekker flere arter sil, hvorav havsil er den vanligste i våre farvann). Havsil er ved siden av å være kommersielt viktig, en nøkkelart i økosystemet i Nordsjøen. Havsilen er svært stedbunden fordi den har strenge krav til sjøbunnen. Individer eldre enn et halvt år oppholder seg nedgravd i sanden store deler av tiden og ved gyting klistres eggene til sand og grus. Det er bare mellom larveklekking i februar/mars og bunnslåing som yngel i mai/juni at havsil ikke er helt avhengig av passende bunnforhold. Den sterke tilknytningen til egnet bunnhabitat medfører at havsilen er svært flekkvis fordelt i Nordsjøen og dette foreslåtte SVO-et dekker alle tobisområder i norsk økonomisk sone.
Området er svært viktig for sjøfugl siden havsil er et av de viktigste byttedyrene for alkefugler og måker. De sentrale delene av Nordsjøen er et viktig overvintringsområde for havhest og til dels krykkje. Lomvi og alke bruker også området, og alkekonge og lunde er innom i mindre grad. Dette er også viktige beiteområder for havert og vågehval. Bestandsnedgang for tobis vil føre til sviktende næringstilgang til sjøfugl, fisk og sjøpattedyr i Nordsjøen og langs kysten av Norge og Storbritannia.
Enkelte av tobisområdene ligger innenfor områder hvor det har vært registrert høyere planteplanktonbiomasse. Områdene er karakterisert av høyere konsentrasjoner av dyreplankton, sammenlignet med omkringliggende områder, særlig i vår- og sommerperioden.
Havsil. Foto: E. Svendsen
Figur 10.4.1. Kart over foreslått endret SVO Tobisfelt (NS2). Grensene for det foreslåtte SVO-et er vist med tykk rød strek, mens grenser for de øvrige SVO-ene i Nordsjøen er vist med tynn rød strek. Grensene for forvaltningsplanområdet er markert med brun linje. Kartet viser utbredelsen av tobisfeltene som er grunnlaget for avgrensingen av det foreslåtte SVO-et. Tobisfelt utenfor det foreslåtte SVOet og norsk økonomisk sone er markert i kartet og viser at de feltene som ligger innenfor norsk sone utgjør hovedarealet.
Havhest. Foto: Erik Olsen, HI
10.5.2 - Vurdering i forhold til EBSA-kriteriene – Forslag til endret SVO Tobisfelt (NS2)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til endret SVO Tobisfelt (NS2)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet/sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Dyreplankton
Fisk
Bunntopografien kan bidra til spesielle hydrografiske forhold, med påfølgende generelt høyere primær- og sekundærproduksjon på grunn av oppstrømming og frontsystemer (Kiørboe mfl. 1988, Jonasdottir og Koski 2011, Richardson mfl. 2000). Områdene er karakterisert av høyere konsentrasjoner av dyreplankton, sammenlignet med omkringliggende områder, særlig i vår og sommerperioden (Krause 1995).
Områdene er helt sentrale for bestandsutviklingen av havsil i norsk sektor av Nordsjøen (Sundby mfl. 2017, ICES 2017). Havsilen som forflytter seg lite, lever her i beite-, overvintrings-, og gyteperioden der eggene legges på bunnen her. Bunnhabitatet består i hovedsak av grus og grov sand som er meget viktig for havsil (Wright mfl. 2000, Holland mfl. 2005, Jensen mfl. 2011, ICES 2017).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Sjøpattedyr
Sjøfugl
Fisk
Områdene er helt sentrale for bestandsutviklingen av havsil i norsk sektor av Nordsjøen (Sundby mfl. 2017, ICES 2017). Havsilen som forflytter seg lite, lever her i beite-, overvintrings-, og gyteperioden der eggene legges på bunnen her. Bunnhabitatet består i hovedsak av grus og grov sand som er meget viktig for havsil (Wright mfl. 2000, Holland mfl. 2005, ICES 2017, Jensen mfl. 2011).
Viktige beiteområder for havert (Hammond og Wilson 2016) og vågehval (Olsen og Holst 2001).
Området er svært viktig for sjøfugl siden havsil er et av de viktigste byttedyrene for alkefugler og måker. De sentrale delene av Nordsjøen er et viktig overvintringsområde for havhest og til dels krykkje. Lomvi og alke bruker også området, og alkekonge og lunde er innom i mindre grad (Fauchald mfl. 2007, Systad mfl. 2019).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Sjøfugl
Fisk
Havsilbestanden i sentrale og sørlige deler av norsk sektor har vært flere år på svært lavt nivå og var før 2013 listet på rødlista som sårbar (VU). Den har hatt et oppsving de siste årene da man har klart å bygge opp gytebestanden og hatt en høyere rekruttering enn på begynnelsen av 2000-tallet (Johnsen 2020). Havsilbestanden på Vikingbanken har ikke hatt tilsvarende oppsving, og bestanden der er fremdeles kritisk lav (ICES 2020, Johnsen 2020).
Området har stor betydning for en rekke rødlistede sjøfuglarter oppover Norskekysten, men også som beiteområde for bestander av rødlistede sjøfuglarter i området, f.eks. havhest (sterkt truet-EN) fra Vestlandet gjennom hele året (seapop.no).
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Fisk
Med unntak av at området er et viktig leveområde for havsil og viktig for dens predatorer er ikke habitatet særlig følsomt (Furness 1999, Furness 2002).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Plante-plankton
Dyreplankton
Fisk
De grunne bankområdene i Nordsjøen har generelt en høyere primær- og sekundærproduksjon på grunn av oppstrømming og frontsystemer (Kiørboe mfl. 1988, Jonasdottir og Koski 2011, Richardson mfl. 2000), i enkelte av områdene er det registrert høyere planteplanktonbiomasse (Edwards 2001). Området har høye konsentrasjoner av dyreplankton i vår- og sommerperioden (Krause 1995). Dyreplankton er viktig byttedyr for fisk i området (van Deurs 2009).
Havsil beiter på plankton (Macer 1966, Van Deurs mfl. 2014) og er en nøkkelart når det gjelder å overføre energi til høyere trofisk nivå i og utenfor SVO-grensene (fisk, sjøfugl og sjøpattedyr) (Furness 1990, Furness 2002).
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Fisk Sjøfugl
Havsil er et viktig byttedyr for sjøfugl, fisk og hval, også utenfor det foreslåtte SVO-et (Furness 1999, Furness 2002). Mattilgangen gjør området til viktige beiteområder for havert (Hammond og Wilson 2016) og vågehval (Olsen og Holst 2001).
Området er svært viktig for sjøfugl siden havsil er et av de viktigste byttedyrene for alkefugler og måker. De sentrale delene av Nordsjøen er et viktig overvintringsområde for havhest og til dels krykkje. Lomvi og alke bruker også området, og alkekonge og lunde er innom i mindre grad (Systad mfl. 2019, Fauchald mfl. 2007).
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
10.5.3 - Lokalisering
Foreslått endret SVO Tobisfelt dekker de eksisterende SVO-ene Tobisfelt sør og Tobisfelt nord (Vikingbanken), sist oppdatert i Meld. St. 20 (2019-2020). Begge disse SVO-ene omfatter viktige områder for tobis, og er her anbefalt slått sammen til ett SVO Tobisfelt.
Foreslått endret SVO Tobisfelt omfatter de viktigste tobisområdene i norsk sektor av Nordsjøen. De er fordelt mellom Vikingbanken i nord, sentrale områder rundt Ling, Midtbanken, Austbanken og Engelsk Klondyke, og de mer sørlige områdene ved Inner Shoal, Vestbanken og Ytterbanken (Fig. 10.4.1). Området overlapper ikke med andre eksisterende eller foreslåtte SVO-er.
Det er ingen marine verneområder som også dekker tobisfeltene.
10.5.4 - Introduksjon til området
Det fysiske miljøet i foreslått endret SVO Tobisfelt er preget av innstrømmende atlanterhavsvann. Vannmassene på sokkelen i nordlige deler av Nordsjøen kommer enten nordfra rundt Shetland eller vestfra mellom Shetland og Orknøyene og er derfor sterkt preget av egenskapene til atlanterhavsvannet (Fig. 10.4.2). Områdene er svært eksponerte og utsatt for betydelig vertikal omrøring, og derfor er vannmassene i disse sokkelområdene relativt homogene uten klare vertikale gradienter. Unntaket er at overflatevannet kan oppnå noe høyere temperatur gjennom sommeren. Strømforholdene er sterkt avhengig av vindforholdene, men generelt er det en drift sørover ved de nordlige tobisfeltene og en mer østlig drift ved de sørligste tobisfeltene.
Figur 10.4.2. Kart over gjennomsnittlig overflatestrøm (piler) og bunndyp (fargekoder) i foreslått endret SVO Tobisfelt og omkringliggende områder.
10.5.5 - Beskrivelse av miljøverdiene
Plankton
Det foreligger ingen systematisk datainnsamling av planteplankton i det aktuelle området. Reid mfl. (1990) viser til høyere produksjon i de kystnære områdene i sentrale, sørlige og østlige deler av Nordsjøen, med noe lavere produksjon i nordlige deler. En studie av McQuatters-Gollop mfl. (2007) indikerte høyere produksjon i de kystnære delene av Nordsjøen enn i de åpne havområdene, med en økende trend i biomasse (klorofyll) etter 80-tallet. Edwards mfl. (2001) fant at det var forskjeller i de ulike regionene av Nordsjøen der de sentrale og sørlige delene har høy produksjon, og der områdene i de nordlige delene har lavere produksjon. Enkelte av de indikerte tobisområdene vil ligge innenfor de områder der man har registrert høyere planteplanktonbiomasse. Topografiske forhold i de grunne delene av Nordsjøen kan medføre oppstrømning og høyere produksjon lokalt.
Foreslått SVO Tobisfelt er preget av stor utskifting av vannmasser med atlantisk opprinnelse. Dyreplanktonet i dette området av Nordsjøen domineres av raudåte (Calanus finmarchicus) i vårperioden, og et større innslag av C. helgolandicus og Pseudocalanus spp senere på året. Det er registrert forhøyede konsentrasjoner av dyreplankton i området for foreslått SVO Tobisfelt, på sokkelområdet sørvest av norskerenna i vår- og sommerperioden (Krause 1995). Det er ikke kjent om dette er et resultat av økt lokal produksjon eller om det skyldes tilførselsprosesser. De grunne bankområdene i Nordsjøen regnes for å ha generell høy primær- og sekundærproduksjon. Dette skyldes stor vertikal omrøring, som fører til økt oppstrømning av næringssalter og derved økt primær- og sekundærproduksjon. I overgangen mellom de vertikalt homogene grunne områdene, og de omkringliggende stratifiserte vannmassene kan det oppstå et frontsystem med økt konsentrasjon av dyreplankton. Dette er dokumentert ved andre områder i Nordsjøen med lignende bunntopografi (Kiørboe mfl. 1988, Jonasdottir og Koski 2011, Richardson mfl. 2000).
Det er grunn til å anta at dyreplanktonet i dette området har gjennomgått de samme storskala endringene som i det øvrige Nordsjøen, med en reduksjon i mengden raudåte (C. finmarchicus) og en økning i varmekjære arter som C. helgolandicus (Beaugrand mfl. 2002). Dette har ført til lavere sekundærproduksjon og endret sesongmønster over tid (Edwards 2016).
Fisk
Foreslått endret SVO Tobisfelt er gyte-og leveområder for tobis som er en samlebetegnelse for flere arter sil, hvorav havsil er det vanligste i norske farvann. Havsil er, ved siden av å være kommersielt viktig, en nøkkelart i økosystemet i Nordsjøen (Furness 2002). Havsil beiter på plankton (Macer 1966, Van Deurs mfl. 2014) og er en nøkkelart i forhold til å overføre energi til høyere trofisk nivå i og utenfor SVOgrensene (fisk, sjøfugl og sjøpattedyr) (Furness 1990, Furness 2002). En rekke torskefiskarter, makrell, sild og flyndrer beiter på de forskjellige sil-artene (Greenstreet mfl. 1998, Høines og Bergstad 1999, Adlerstein og Welleman 2000, Furness 2002, Fredriksen mfl. 2006). Havsil finnes over større områder, men de definerte områdene i foreslått SVO Tobisfelt er de områdene som historisk har hatt de største biomassene (ICES 2016). Etter en langsom oppbygging av gytebiomassen har de midtre og sørlige områdene fått en jevnlig rekruttering de siste årene (ICES 2020 b) noe som er viktig for å forsyne store områder med larver, og bidra til store mengder mat til hval, fisk og sjøfugl i økosystemet. I det nordlige området er situasjonen annerledes, og på tross av over to tiår med fiskestopp på havsil så er fremdeles mengden havsil på Vikingbanken kritisk lav (ICES 2020 b). Til tross for denne forskjellen, er det ikke naturlige forskjeller mellom tobisfeltene i sentrale Nordsjøen og det nordlige på Vikingbanken. Derfor er det er naturlig å se alle tobisfeltene som et felles SVO.
Områdene har sedimentforhold som er helt sentrale for bestandsutviklingen av havsil i norsk sektor av Nordsjøen (Sundby mfl. 2017). Havsilen er svært stedbunden siden den har strenge krav til sjøbunnen, og lever her i beite-, overvintrings, og gyteperioden der eggene legges på bunnen, klistret til sand og grus. Bunnhabitatet består i hovedsak av grus og grov sand som er meget viktig for havsil når den graver seg ned i skjul om dagen. Individer eldre enn et halvt år oppholder seg nedgravd i sanden store deler av tiden, og det er bare mellom larveklekking i februar/mars og bunnslåing som yngel i mai/juni at havsil ikke er helt avhengig av passende bunnforhold (Wright mfl. 2000, Holland mfl. 2005, Jensen mfl. 2011).
Bunnsamfunn
Bunnsamfunnet i havsilhabitat karakteriseres av mobile arter og meiofauna (Kröncke og Bergfeld 2003). Fastsittende fauna er mindre vanlig. Betydningen av havsil for bunnsamfunnet er lite kjent, men man må anta at eggene utgjør en sesongmessig fødekilde.
Sjøpattedyr
Foreslått endret SVO Tobisfelt er et viktig beiteområde for sjøpattedyr, særlig havert (Halichoerus grypus) og vågehval (Balaenoptera acutorostrata). I Nordsjøen er havsil dominerende i dietten til havert (Hammond og Wilson 2016) og vågehval (Olsen og Holst 2001).
Sjøfugl
Området er svært viktig for sjøfugl siden et av de viktigste byttedyrene til alkefuglene, fiskender og måker er havsil. Betydningen av denne ressursen strekker seg langt utover grensene for området av den grunn. De sentrale delene av Nordsjøen er et viktig overvintringsområde for havhest (Fulmarus glacialis) og til dels krykkje (Rissa tridactyla). Lomvi (Uria aalge) og alke bruker også området, og alkekonge (Alle alle) og lunde (Fratercula arctica) er innom i mindre grad (Systad mfl. 2019, Fauchald mfl. 2007).
10.6 - Forslag til nytt SVO Norskerenna (NS3)
10.6.1 - Sammendrag
Områdebeskrivelse og strukturer
Foreslått nytt SVO Norskerenna (NS3) er en viktig dyprenne som går gjennom Skagerrak parallelt med kysten. Med sitt betydelige dyp i et ellers grunt sokkelhav, har Norskerenna lys-, temperatur-, strøm- og fysiske miljøforhold som er helt unike i havområdet. Området er mørkere, kaldere og saltere enn de grunnere områdene ellers i Nordsjøen.
Miljøverdier
Norskerenna skiller seg fra resten av Nordsjøen/Skagerrak, med stor andel av mesopelagiske dyreplanktonarter som ellers ikke er vanlige i de grunne områdene av Nordsjøen, som for eksempel pelagiske reker, geleplankton, krill og store hoppekreps. Norskerenna har overvintrende populasjoner av raudåte som er en nøkkelart i den pelagiske næringskjeden. Fordi Norskerenna er det eneste området i Nordsjøen/Skagerrak hvor det forekommer overvintrende raudåte kan dette ha betydning for forekomst av raudåte langs kysten av Nordsjøen/Skagerrak, særlig i vårperioden da raudåta vandrer opp til overflaten for å gyte.
Norskerenna er eneste sted i Nordsjøen der dypvannsfisk har naturlig biotop, og laksesild er dominerende art. Skagerrak er viktig som oppvekstområde for dypvannsreke i sør og området kan tenkes å komme til å utgjøre et klimarefugium for dypvannsreke i dette området når havet varmes opp ytterligere. Dyreplankton, reker og flere fiskearter er viktig føde for arter med tilhold i Norskerenna, men også fisk, sjøfugl og sjøpattedyr i grunnere områder beiter på arter som opprinnelig kommer fra Norskerenna.
Vestlig del av de dype områdene i Skagerrak/Norskerenna har høy tetthet av bambuskorall og sjøfjær. Forekomstene i Norskerenna er de rikeste forekomstene av bambuskorall i Norge utenom forekomster i fjorder. Det spesielle dyresamfunnet i Norskerenna er i seg selv et fungerende økosystem med et unikt biologisk mangfold, som også i stor grad er beskyttet mot menneskelig aktivitet.
Detalj av bambuskorall. Foto: EPIGRAPH/HI
Figur 10.5.1. Kart over foreslått nytt SVO Norskerenna (NS3). Grensene for foreslått nytt SVO-et er markert med tykk rød strek, grenser for de nærliggende foreslåtte endrete SVO-ene (Boknafjorden og Jærstrendene (NS1) og Ytre Oslofjord (NS4)) er markert med tynnere røde streker og grensene for forvaltningsplanområdet er vist med brun linje. Kartet viser utbredelsen av utvalgte miljøverdier med betydning for området. Utbredelse av overvintringsområdet for hoppekreps i Norskerenna og de tilstøtende kjerneområdene for bambuskorall (Isidella lofotensis) og sjøfjær (Funiculina, Kophobelemnon, Halipteris) er inkludert iforeslått nytt SVO-et. Kartet er basert på relative mengder i trålbifangst på reketokt i perioden 2017-2020 (Buhl-Mortensen og Thangstad, in prep). Kjerneområder for sjøfjær sammenfaller med områder med store fangster av dypvannsreke (Søvik mfl. 2019).
Meganyctiphanes norvegica. Foto: Cecilie Brohms, HI
10.6.2 - Vurdering i forhold til EBSA-kriteriene – Forslag til nytt SVO Norskerenna (NS3)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til nytt SVO Norskerenna (NS3)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet/sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Bunnsamfunn
Dyreplankton Fisk
Norskerenna er et unikt dypområde i et ellers grunt havområde (Nordsjøen/Skagerrak) der innstrømning av atlanterhavsvann i de dypere vannlag fører med seg planktonarter fra utenforliggende havområder (Bakketeig 2017, Bergstad 1991). Norskerenna er habitat for overvintrende raudåte (Heath mfl. 2004, Melle mfl. 2014, Falkenhaug pers obs) og for pelagiske reker og krill (Meganyctiphanes norvegica).
Det er eneste sted i Nordsjøen der dypvannsfisk har naturlig biotop (Bergstad 1991).
Vestlig del av de dype områdene i Skagerrak/Norskerenna har høy tetthet av bambuskorall og sjøfjær (Artsdatabanken 2018, Buhl-Mortensen og Thangstad, under forberedelse).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Dyreplankton
Bunnsamfunn Fisk
Norskerenna er det eneste området i Nordsjøen/Skagerrak der det forekommer overvintrende raudåte (Calanus finmarchicus) (Heath mfl. 2004, Melle mfl. 2014 Falkenhaug pers obs). Dette kan ha lokal betydning for forekomst av raudåte langs kysten av Nordsjøen/Skagerrak, særlig i vårperioden da raudåta vandrer opp til overflaten for å gyte.
Mesopelagiske fiskearter er avhengig av dypet (Bergstad 1991).
Skagerrak er viktig som oppvekstområde for dypvannsreke i sør (Hjort og Ruud 1938, Søvik og Thangstad 2021). Det er i SVO-et særlig gode forhold for bambuskorall (Buhl-Mortensen og Thangstad, under forberedelse).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Dyreplankton
Raudåte (C. finmarchicus) har vist en nedgang i Nordsjøen/Skagerrak de siste 50 år (Edwards 2016). Norskerenna er habitat for overvintrende populasjoner av raudåte, og det eneste området som er tilstrekkelig dypt for overvintring av raudåte (Heath mfl. 2004, Melle mfl. 2014, Falkenhaug pers obs)
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Dyreplankton
Fisk Bunnsamfunn
Mangler kunnskap om overvintrende populasjoner av raudåte, men det er grunn til å tro at stigende temperaturer samt lave oksygennivåer i Norskerenna som følge av klimaendringer vil redusere mulighetene for overvintring av raudåte i dette området (Heath mfl. 2004, Melle mfl. 2014).
Norskerenna har Nordsjøens eneste forekomster av mesopelagisk fauna, som pelagiske krepsdyr, blekksprut, geleplankton og fisk, der laksesild (Maurolicus muelleri) er dominerende art (Bergstad 1991).
De dypere områdene av Skagerrak kan tenkes å komme til å utgjøre et klimarefugium for dypvannsreke i sør når havet varmes opp ytterligere. Dypvannsreke har sin sørligste utbredelsesgrense i Nordøst-Atlanteren nettopp i Nordsjøen og Skagerrak (NAFO, ICES 2019), og i den vestligste delen er det forekomster av bambuskorall (Buhl-Mortensen og Thangstad, under forberedelse).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Dyreplankton Mesopelagisk fana
Fisk Bunnsamfunn
Norskerenna har overvintrende populasjoner av raudåte som er en nøkkelart i den pelagiske næringskjeden (Heath mfl. 2004, Melle mfl. 2014, Falkenhaug pers obs).
Norskerenna inneholder store forekomster av krill som er svært viktig for sei og kolmule. Skolest er trolig avhengig av overvintringen til Calanus og dypvannsreke, mens vassild synes helst å utnytte rike forekomster av geléplankton i dypet (Bergstad 1991). Norskerenna har Nordsjøens eneste forekomster av mesopelagisk fauna, inkl. fisk, og laksesild (Maurolicus muelleri) er dominerende art (Bergstad 1991).
Bestanden av dypvannsreke i Skagerrak/Norskerenna er mye større enn de små geografisk adskilte rekefeltene langs norskekysten (Hjort og Ruud 1938). Slik sett utgjør rekebestanden i Norskerenna/Skagerrak et viktig reservoar for rekene nordover langs kysten.
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Fisk
Dyreplankton Bunnsamfunn Mesopelagisk fauna
Norskerenna er det eneste området i Nordsjøen/Skagerrak der det forekommer overvintrende raudåte (C.finmarchicus, Heath mfl. 2004, Melle mfl. 2014, Falkenhaug pers obs). Denne overvintringspopulasjonen kan ha betydning for forekomst av raudåte langs kysten av Nordsjøen/Skagerrak, særlig i vårperioden da raudåta vandrer opp til overflaten for å gyte.
Norskerenna har Nordsjøens eneste forekomster av mesopelagisk fisk, og laksesild (Maurolicus muelleri) er dominerende art (Bergstad 1991).
Norskerenna skiller seg fra resten av Nordsjøen/Skagerrak, med stor andel av mesopelagiske dyreplanktonarter som ellers ikke forekommer vanlig i de grunne områdene av Nordsjøen, som for eksempel pelagiske reker, geleplankton, krill, store hoppekreps (Calanus hyperboreus, Paraeuchaeta norvegica) (Bergstad 1991, Bakketeig mfl. 2017).
I den vestligste delen særlige korallforekomster (Bergstad 1991, Bakketeig mfl. 2017, Buhl-Mortensen og Thangstad, under forberedelse).
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
10.6.3 - Lokalisering
Foreslått SVO Norskerenna er et nytt område basert på ny kunnskap om områdets betydning for plankton og bunnsamfunn. I tillegg til arealer som ikke tidligere har vært definert som SVO, dekker området hele eller deler av tre eksisterende SVO-er som fagekspertgruppen foreslår å fjerne som egne SVO-er. Dette gjelder SVO Listastrendene og Siragrunnen, SVO Transekt Skagerrak og SVO Skagerrak, sist oppdatert i Meld. St. 20 (2019-2020).
Norskerenna strekker seg langs hele sørvest-Norge og inn mot Oslofjorden, er over 700 m dyp utenfor Arendal og har en terskel på ca. 270 m utenfor Stavanger. Norskerenna som særlig verdifullt område inkluderer hele Norskerenna innenfor norsk sektor, samt viktige områder for bambuskorall og sjøfjær utenfor kysten av sørvest-Norge (Fig. 10.5.1). Området ligger i nordøst tett opp mot SVO Ytre Oslofjord som fagekspertgruppen foreslår blir utvidet.
Foreslått nytt SVO Norskerenna overlapper med det marine verneområdet Transekt Skagerrak, og ligger tett opp til Listastrendene landskapsvernområde.
10.6.4 - Introduksjon til området
De kystnære delene av Norskerenna er nær overflaten preget av lavsaltholdig kystvann samt relativt sterke strømmer. Vest-Jyllandstrømmen kommer fra sørlige Nordsjøen og dreier østover inn i Skagerrak. Denne møter utstrømningen fra Østersjøen og Kattegat og definerer oppstarten på Den norske kyststrømmen som går mot klokka langs Skagerrak-kysten og videre nordover (Sætre, 2007). Noe saltere vannmasser strømmer også i overflaten inn fra vest og blander seg med kystvannet. I tillegg er det en vesentlig resirkulering av kystvann fra kyststrømmen som øker oppholdstiden til organismer i Skagerrak i hele vannsøylen. Under kyststrømmen finner man noe saltere vannmasser som er blandet fra flere kilder, samt at man har mer stagnerende bunnvann under 3-400 m dyp.
Figur 10.5.2. Kart over gjennomsnittlig overflatestrøm (piler) og bunndyp (fargekoder) i foreslått nytt SVO Norskerenna og omkringliggende områder.
Det fysiske miljøet i Norskerenna er hovedsakelig dominert av kyststrømmen på norsk side med lavsaltholdig kystvann og relativt sterke sørvestgående strømmer (Sætre 2007). Områdene sentralt i Skagerrak er mer preget av både saltere atlanterhavsvann, nordsjøvann og østersjøvann. Vindforholdene har stor innvirkning på hvordan dynamikken i overflaten endrer seg samt hvilke vannmasser som gjør seg mest gjeldende. Det fysiske miljøet under kyststrømmen på rundt 50 m og nedover til 2-300 m dyp er sterkt preget av atlanterhavsvann og vil derfor være utsatt for mer langperiodiske variasjoner i saltholdighet og temperatur. Under dette intermediære vannlaget finner man mer stagnerende vannmasser ettersom utskiftningsperioden i bunnvannet i Norskerenna er på mellom 1 og 5 år.
Norskerenna strekker seg langs hele Sørvest-Norge og inn mot Oslofjorden, er over 700 m dyp utenfor Arendal og har en terskel på ca. 270 m utenfor Stavanger. Av den grunn kan bunnvannet i Skagerrak betraktes på samme måte som i en fjord der man har stagnerende vannmasser under terskelnivå og med mer eller mindre jevnlig utskiftning av tyngre vann. Utskiftningen i Skagerrak skjer med ett eller flere (opptil 4-5) års mellomrom og normalt i perioden mars-april. Etter 1990 er det registrert 15 utskiftninger, der den siste skjedde våren 2018 (Klimastatus.hi)
10.6.5 - Beskrivelse av miljøverdiene
Plankton
I områdene ved kysten og i kyststrømmen er det høyere produksjon og biomasse av planteplankton enn i de utenforliggende områdene. Publikasjoner omtalt under foreslått SVO Tobisfelt (Kap 10.4) vil i stor grad være gjeldende også for dette forslaget til SVO-et for storskala dynamikk. Data fra transektet mellom Norge og Danmark (Torungen-Hirtshals) indikerer avtakende produksjon fra de kystnære områdene til de dypereliggende delene (Naustvoll, upubl). Data fra Økokyst, og tidligere program, beskriver forholdene i de kystnære områdene (f.eks. Frigstad mfl. 2018, Naustvoll mfl. 2020).
Fordeling av vannmasser og forflytnings- (advektive) prosesser har stor betydning for dyreplanktonsamfunnet i dette området. I de øvre vannlag er dyreplanktonet dominert av store herbivore hoppekreps som raudåte (Calanus finmarchicus) i vårperioden april-mai, mens C. helgolandicus og små hoppekreps dominerer senere på sesongen. Mengden av raudåte i området er sannsynligvis avhengig av en årlig tilførsel med havstrømmer fra nordlige Nordsjøen og er således følsom for klimaendringer. I de øvre vannlag av Skagerrak har man observert en økning i mengden C. helgolandicus de siste 20 år mens raudåte har store mellomårsvariasjoner uten en klar langtidstrend (Falkenhaug mfl. 2014). Dette kan ha sammenheng med nærheten til lokale overvintringspopulasjoner i Norskerenna samt variasjoner i advektiv tilførsel. Den mikrobielle næringskjeden, med beiting fra protozooplankton (flagellater og ciliater) kan ha stor betydning, særlig i de sentrale delene av Skagerrak, samt i høstperioden (Maar mfl. 2004).
Innstrømming av atlanterhavsvann i de dypere vannlag av Norskerenna fører med seg planktonarter fra utenforliggende havområder. Artssammensetningen av dyreplankton i de dypere deler av Norskerenna skiller seg derfor fra resten av Nordsjøen/Skagerrak, med stor andel av arter som ellers ikke forekommer vanlig i de grunne områdene av Nordsjøen, som for eksempel pelagiske reker, geleplankton, krill og store hoppekreps (Calanus hyperboreus, Paraeuchaeta norvegica) (Bergstad 1991, Bakketeig mfl. 2017). I Norskerenna er det registrert store mengder av krill-arten Meganyctiphanes norvegica som er svært viktig føde for flere fiskearter i området (Bergstad 1991).
Norskerenna er det eneste området i Nordsjøen/Skagerrak der det forekommer overvintrende raudåte (C. finmarchicus, Heath mfl. 2004, Melle mfl. 2014, Falkenhaug pers obs). Denne overvintringspopulasjonen kan ha betydning for forekomst av raudåte langs kysten av Nordsjøen/Skagerrak, særlig i vårperioden da raudåta vandrer opp til overflaten for å gyte. Man antar at mengden av raudåte i Nordsjøen/Skagerrak er avhengig av en årlig tilførsel av raudåte fra havområdene utenfor Nordsjøen. Denne innstrømningen av raudåte foregår i Norskerenna, der atlanterhavsvann strømmer inn fra nord og følger vestskråningen av Norskerenna inn i Skagerrak.
Raudåte (C. finmarchicus) overvintrer på store dyp, med redusert metabolisme (diapause) for å spare på energireserver. I Norskerenna overvintrer raudåte ved temperaturer på 7-8ºC (Heath mfl. 2004), som er adskillig høyere enn foretrukket temperatur for overvintring (3–6°C). Dette gjør populasjonen i dette området ekstra sårbar for endringer i miljøforhold (kap. 10.2.1). Stigende temperaturer samt lave oksygennivåer i Norskerenna som følge av klimaendringer (miljostatus.no) vil redusere mulighetene for overvintring av raudåte i dette området.
Fisk
Foreslått nytt SVO Norskerenna har spesielle miljøforhold som er styrt av den kontinuerlige koblingen til Norskehavet og Atlanterhavet ved innstrømningen av atlanterhavsvann. Innstrømningen og dybdeforholdene legger til rette for advektiv produksjon i tillegg til den lokale produksjonen. Produksjonen av kopepoder langs vestkanten av Norskerenna og innerst i Skagerrak er viktig beiteområder for de epipelagiske bestandene (sild, (Clupea harengus) og makrell (Scomber scombrus)), men også for bentopelagiske arter som øyepål (Trisopterus esmarkii), kolmule (Micromesistius poutassou) og skolest (Coryphaenoides rupestris). I tillegg er krill viktig, og ingen andre steder i Nordsjøen er det like store mengder Meganyctiphanes norvegica som er svært viktig for sei (Pollachius virens) og kolmule.
Mesopelagisk fauna
Foreslått nytt SVO Norskerenna er også dypt nok til å ha forekomster av vassild (Argentina silus) og skolest (Coryphaenoides rupestris) som er utelukket fra resten av nordsjøbassenget, men forekommer i dype fjorder og langs skråningene i Atlanterhavet. Skolestbestanden er isolert, mens vassildforekomstene er kontinuerlige fra indre Skagerrak til Nord-Norge. Begge disse artene er langtlevende og aggregerende kommersielle målarter, og de er derfor sårbare. ICES har i flere år anbefalt 0-fangst for skolest og direkte fiske har vært stoppet siden 2006. Begge arter gyter i Skagerrak. Skolest er trolig avhengig av overvintringen til Calanus og dypvannsreke (Pandalus borealis), mens vassild synes helst å utnytte rike forekomster av geléplankton i dypet (Bergstad 1991). I tillegg har Norskerenna også Nordsjøens eneste forekomster av laksesild (Maurolicus muelleri) (Bergstad 1991).
Bunnsamfunn
Dypere enn 200 m er det hovedsakelig mudderbunn og mudderbunnsbunndyr, forskjellig fra sand-, grus- og hardbunnsområdene på Nordsjøplatået (Bergstad 1991).
Informasjon om utbredelse av koraller (hornkoraller og korallrev), sjøfjær og svamp er sparsom fra området. Imidlertid har Havforskningsinstituttet de senere år startet registrering av sjøfjær og hornkoraller i bifangst under det årlige reketoktet i Norskerenna/Skagerrak. Her presenteres en oversikt over slike funn fra perioden 2017-2019. Disse funnene gir grunnlag for å avgrense deler av Skagerrak som verdifulle områder spesifikt med tanke på bunnfauna. Bambuskorall (Isidella lofotensis) er en hornkorall som med få og usikre unntak kun er registrert i norske hav- og kystområder. Forekomstene i Norskerenna er de rikeste forekomstene i Norge utenom forekomster i fjorder (spesielt Hardangerfjorden, Trondheimsfjorden og Andfjorden) (Buhl-Mortensen og Thangstad, under forberedelse) (Fig. 10.5.1).
Dypvannsreke utgjør én genetisk bestand fra Skagerrak og nordover til Kvænangen (Jorde mfl. 2015, Knutsen mfl. 2015). Rekene i fjordene i Skagerrak er delvis genetisk forskjellige fra rekene ute i Skagerrak (Knutsen mfl. 2015). Rekebestanden i Skagerrak og Norskerenna er økonomisk viktig, og bestandsstatus bestemmes årlig gjennom et eget reketokt og bestandsvurdering i regi av ICES (ICES 2021). Rekrutteringen har vært lav siden 2006-2007, og bestanden har de siste årene ligget under føre-var nivået (ICES 2021).
Dypvannsreke har sin sørligste utbredelsesgrense i Nordøst-Atlanteren nettopp i Nordsjøen og Skagerrak. Skagerrak er viktig som oppvekstområde for dypvannsreke i sør. Mesteparten av de ett-årige rekene finnes i dette området (Søvik og Thangstad 2021). Ettåringene driver/migrerer etter hvert vestover og bidrar slik til rekruttering til den voksne rekebestanden i Norskerenna vest av Lindesnes.
Norskerenna vest av Rogaland utgjør sannsynligvis et viktig område for produksjon av rekelarver som har betydning for rekefeltene videre nordover langs kysten, da preliminære driftsmodelleringsstudier viser at larver som klekker her, driver nordover med den norske kyststrømmen (upubliserte resultater). Et viktig moment er at bestanden i Skagerrak/Norskerenna er så mye større enn de små geografisk adskilte rekefeltene langs norskekysten (Hjort og Ruud 1938). Slik sett utgjør rekebestanden i Norskerenna/Skagerrak en viktig kilde for produksjon og spredning av rekeyngel nordover langs kysten.
Sjøkreps (Nephrops norvegicus) er avhengig av bløtbunn for å grave sine huler i bunnsedimentet (Johnson mfl. 2013). Store bløtbunnsområder sør og øst i Skagerrak gjør at det finnes en stor og produktiv sjøkrepsbestand i dette området, men hovedsakelig utenfor det foreslåtte SVO-et og hovedsakelig i svensk og dansk farvann (ICES 2020 a). I norsk farvann i Skagerrak finnes det større områder med sjøkrepsbunn langs territorialgrensen til Danmark samt i Ytre Oslofjord og ved Hvaler. Videosledeundersøkelser tyder på at bestanden er i god forfatning (ICES 2020 b).
Store deler av norsk sone i Nordsjøen (Norskerenna) består også av bløtbunn og dermed ideelt sjøkrepshabitat. Den biologiske produksjonen i sjøkrepsbestanden i norsk sone i Nordsjøen (Norskerenna) synes likevel å ha avtatt de senere årene (ICES 2020 a). Sjøkrepslandingene herfra har minket fra mer enn 1000 tonn til rundt 200 tonn i 2016-2019 (ICES 2020 a), men dette skyldes også forandringer i forvaltning og reguleringer (Søvik mfl. 2017). Denne bestanden har vi lite kunnskap om. Det tidligere trålbaserte fisket er nå delvis erstattet av et kystnært teinefiske (Søvik mfl. 2017).
Sjøpattedyr
Området omfatter beiteområder for mindre kolonier av steinkobbe (Phoca vitulina), men det oppfyller ikke kriteriene for å begrunne SVO-statusen basert på sjøpattedyr (Nilssen og Bjørge 2019).
Sjøfugl
Området er del av et større område som er viktig for flere sjøfuglarter, men isolert sett har ikke dette området stor betydning. Det foreslåtte SVO-et berører områder viktig for overvintrende alkefugl og fiskemåke (Larus canus), samt kystområder viktig for mytende ærfugl (Somateria mollissima) og hekkende storskarv (Phalacrocorax carbo) (Systad mfl. 2007). For sjøfugl er likevel de overflatebeitende bestandene av sildemåke (Larus fuscus), gråmåke (Larus argentatus) og de i Norge sterk truede (EN) makrellternene (Sterna hirunda), de viktigste for dette foreslåtte SVO-et (Systad mfl. 2007, Gasbjerg mfl. 2011). Disse artene er beregnet å bruke områdene ut til mellom 60-100 km fra koloniene i hekketiden (Systad mfl. 2018).
10.7 - Forslag til endret SVO Ytre Oslofjord (NS4)
10.7.1 - Sammendrag
Områdebeskrivelse og strukturer
Forslag til endret SVO Ytre Oslofjord (NS4) er i høy grad påvirket av kyststrømmen, som her tar opp ferskvann fra store elver, vann som kommer ut fra Kattegat og Østersjøen og Nordsjøvann som kommer opp langs vestkysten av Danmark. Det er derfor helt spesielle fysisk/kjemiske og klimatiske forhold. Dette er også grunnlaget for de marine naturparkene Tjøme og Hvaler, som i stor grad preges av de samme vannkvalitetene. Størrelsen på SVO-et er valgt for å dekke områder som i større grad enn nasjonalparkene, omfatter miljøverdier for økosystemet i Skagerrak.
Miljøverdier
Området er et viktig hekke-, trekk- og overvintringsområde for en rekke sjøfugler, deriblant flere sårbare og truede arter. Kystnære arter som ærfugl og siland beiter her gjennom hele året, og forekomstene av ærfugl er nasjonalt viktige. Gråmåke, svartbak og fiskemåke er viktige arter utenom hekkesesongen. Området er også viktig for alkekonge om høsten. Området dekker deler av overvintringsområdet til britiske lomvi, men sannsynligvis også for de sørlige forekomstene av lomvi i Norge. Også betydelige bestander av fiskemåke overvintrer her. Deler av området er dessuten viktig for steinkobbe hele året (beite- og kasteområde).
Området har korallrev som er unike for Skagerrak/Nordsjøen og som representerer et av de største kystnære korallrevområdene i verden. Det finnes også store sammenhengende bløtbunnsområder (Svennerbanken) der reke trives. De store bløtbunnsområdene øst i Skagerrak (Ytre Oslofjord og Hvaler i norsk farvann) gjør at det finnes en stor og produktiv sjøkrepsbestand her
Området har høy biomasse av planteplankton og høyere primærproduksjon sammenlignet med Indre Skagerrak (hav) og andre fjordsystemer.
Planteplankton. Foto: planktonlaboratoriet, HI
Figur 10.6.1. Kart over foreslått utvidet SVO Ytre Oslofjord (NS4). Grensene for det foreslått utvidete SVO-et er vist med tykk rød strek, grenser for det nærliggende forslag til nytt SVO Norskerenna (NS3) er vist med tynn rød strek, og grensene for forvaltningsplanområdet er vist med brun linje. Kartet viser utbredelsen av utvalgte miljøverdier med betydning for området. Miljøverdier knyttet til fisk (gytefelt), bunnsamfunn, sjøpattedyr og sjøfugl er vist.
Sjøkreps . Foto: EPIGRAPH/HI
10.7.2 - Vurdering i forhold til EBSA-kriteriene – Forslag til endret SVO Ytre Oslofjord (NS4)
CBD EBSA kriterier
Beskrivelse
Rangering av kriteriets relevans i forslag til endret SVO Ytre Oslofjord (NS4)
Ikke tilstrekkelig informasjon
Lav
Middels
Høy
Unikhet/sjeldenhet
Området inneholder enten (i) unike (“den eneste av sitt slag"), sjeldne (opptrer kun i få lokaliteter) eller endemiske arter, popula-sjoner eller samfunn, og/eller (ii) unike, sjeldne eller distinkte habitater eller økosystem, og/eller (iii) unike eller uvanlige geomorfologiske eller oseanografiske egenskaper.
Sjøfugl
Bunnsamfunn
Koster/Hvalerområdet har korallrev som er unike for Skagerrak/Nordsjøen og som representerer et av de største kystnære korallrevområdene i verden (Fosså mfl. 2015).
Dette er et helårlig viktig sjøfuglområde, som hekkeområde for ærfugl, makrellterne samt flere kystnære arter, og ikke minst som vinterbeite for ærfugl og lomvi (Systad mfl. 2007, Ottersen mfl. 2010).
Livshistorisk viktige områder
Områder som kreves for at en populasjon skal overleve eller trives.
Fisk Bunnsamfunn Sjøpattedyr
Sjøfugl
Det er to genetisk ulike torskestammer, “fjordtorsk” og “nordsjøtorsk”, der nordsjøtorsk antas ha gytefelt lenger ute i skjærgården enn fjordtorsk (Andre mfl. 2016, Knutsen mfl. 2018).
Sjøkreps er avhengig av bløtbunn for å grave sine huler i bunnsedimentet (Johnson mfl. 2013) og det er en stor bestand i bløtbunnsområdene øst i Skagerrak (Ytre Oslofjord og Hvaler i norsk farvann).
Deler av området er dessuten viktig for steinkobbe hele året (beite- og kasteområde) (Nilssen og Bjørge 2019).
Hele området i Ytre Oslofjord nedover kysten til Risør er et viktig hekke-, trekk-, og overvintringsområde for en rekke sjøfuglarter (Systad mfl. 2007, Ottersen mfl. 2010). Området er for eksempel viktig for ærfuglbestanden gjennom hele året. (Systad mfl. 2019). Makrellterne har et av de viktigste hekkeområdene i Norge her (Gasbjerg mfl. 2011).
Viktighet for truede eller nedadgående arter og/eller habitater
Område som inneholder habitat for overlevelse og restitusjon av truede eller nedadgående arter eller områder med betydelig ansamling av slike arter.
Sjøfugl Bunnsamfunn
Atlanterhavspåvirkningen er stor og stabil i Koster/Hvalerområdet og gir også miljøforhold for korallrev (Desmophylllum pertusum, tidligere kalt Lophelia pertusa) (Fosså mfl. 2015). Sjøkreps er avhengig av bløtbunn for å grave sine huler i bunnsedimentet (Johnson mfl. 2013). Dessuten er det vist i langtidsovervåking en reduksjon i antall arter og individer siden 1950 (Oug mfl. 2015, Walday mfl. 2019).
Området er viktig for ærfuglbestanden (nær truet-NT) gjennom hele året (Systad mfl. 2019). Makrellterne (sterkt truet-EN) er også en viktig art i Nordsjøen og Skagerrak, særlig i utløpet av Oslofjorden (Systad mfl. 2007, Gasbjerg mfl. 2011, Henriksen og Hilmo 2015).
Sårbarhet, skjørhet, følsomhet eller lav restitusjons-evne
Områder som inneholder en relativt høy andel av følsomme habitater, biotoper eller arter som er funksjonelt skjøre (høy sjanse for forringelse eller utryddelse ved menneskelig aktivitet eller ved naturlige hendelser) eller med sen restitusjon.
Bunnsamfunn
Sjøfugl
Foreslått endret SVO Ytre Oslofjord er et område hvor flere fjordsystemer og vannmasser møtes. Atlanterhavspåvirkningen er stor og stabil i Koster/Hvalerområdet og gir miljøforhold for korallrev (Desmophylllum pertusum, tidligere kalt Lophelia pertusa) (Fosså mfl. 2015).
Området er viktig for ærfuglbestanden (nær truet-NT) gjennom hele året, og dekker deler av overvintringsområdet til britiske lomvi og alke. Makrellterne (sterkt truet-EN) som beiter både pelagisk og kystnært har et av de viktigste hekkeområdene i Norge her (Systad mfl. 2019). Andre arter som primært har tilknytning til kysten (og ferskvannsområdene innenfor) er hettemåke (sårbar-VU) og fiskemåke (nær truet-NT) (Henriksen og Hilmo 2015).
Viktighet for biologisk produktivitet
Området inneholder arter, populasjoner eller samfunn med relativt høyere naturlig biologisk produktivitet
Dyreplankton
Plante-plankton Bunnsamfunn
Området har høy biomasse av planteplankton og høyere primærproduksjon sammenlignet med indre Skagerrak (hav) og andre fjordsystemer (Kristiansen 1997, Skjoldal mfl. 1997).
Strømmønster og forekomster av krill og raudåte tyder på at også dyreplankton møter dette kriteriet, men det er ikke dokumentert.
Ytre Oslofjord finnes det store sammenhengende bløtbunnsområder der dyphavsreken trives (Svennerbanken) (Hjort og Ruud 1938). De store bløtbunnsområdene øst i Skagerrak (Ytre Oslofjord og Hvaler i norsk farvann) gjør at det finnes en stor og produktiv sjøkrepsbestand her. Tilførsel av tarerester utnyttes av reker og andre krepsdyr på dypt vann (Ramirez-Llodra mfl. 2016).
Viktighet for biologisk mangfold
Området inneholder relativt høyere mangfold av økosystemer, habitater, samfunn eller arter, eller har et høyere genetisk mangfold.
Sjøfugl
Bunnsamfunn
Korallrev og tareskog bidrar som habitatbyggere til høyt biologisk mangfold (Fosså mfl. 2015, Ramirez-Llodra mfl. 2016, Prøis 2018, Reitan 2020).
Området har stort mangfold i sjøfuglbestanden, de fleste med kysttilknytning, men også pelagisk beitende arter (Systad mfl. 2019, Gasbjerg mfl. 2011). Kystnære arter dominerer mht. biologisk mangfold (Reitan 2020).
Naturlighet
Område med en relativt høyere grad av naturlighet som følge av mangel på eller lavt nivå av menneskeskapte forstyrrelser eller forringelse.
10.7.3 - Lokalisering
SVO Ytre Oslofjord er tidligere definert i Meld. St. 20 (2019-2020), men området er foreslått utvidet sørover for å inkludere kandidatområdet for sjøfugl i forlengelsen av SVO-et. Sjøfugl fra de britiske øyer og Norskekysten overvintrer i dette området, bl.a. britiske lomvi. De beiter pelagisk, og området er utvidet for å dekke (norske deler av) det viktige området som strekker seg nedover i Kattegat. Ved denne utvidelsen inkluderes også flere viktige kolonier for steinkobbe.
Foreslått endret SVO Ytre Oslofjord omfatter ytre del av Oslofjorden og den norske delen av havområdet sør for dette mellom svenskegrensen i sørøst og Risør i sørvest (Fig. 10.6.1). Området ligger tett opp mot foreslått nytt SVO Norskerenna.
Foreslått endret SVO Ytre Oslofjord dekker flere marine verneområder, Ytre Hvaler nasjonalpark, Færder nasjonalpark og Jomfruland nasjonalpark.
10.7.4 - Introduksjon til området
Foreslått endret SVO Ytre Oslofjord er nær overflaten preget av lavsaltholdig kystvann med flere brakk- og ferskvannskilder som møtes. Området ligger nær starten av kyststrømmen slik at de ytterste områdene er dominert av strømmer som medfører relativt sterk, vedvarende strøm fra sør som dreier mot vest. Under kyststrømmen finner man noe saltere vannmasser som også er blandet fra flere kilder, samt at man har mer stagnerende bunnvann under 3-400 m dyp.
Figur 10.6.2. Kart over gjennomsnittlig overflatestrøm (piler) og bunndyp (fargekoder) i foreslått endret SVO Ytre Oslofjord og omkringliggende områder.
Det fysiske miljøet i Ytre Oslofjord er i stor grad påvirket av prosesser i havområdene utenfor fjorden, og særlig kan områdene Nordsjøen og Skagerrak i enkelte år eller perioder av året ha stor betydning for forholdene. Ytre Oslofjord er et åpent fjordsystem som inkluderer åpne kystområder, fjorder og et stort estuaria på østsiden ved Hvaler. De topografiske forholdene i fjorden gjør at området er oppdelt i flere større og mindre bassenger og fjordområder. Overflatevannet i Ytre Oslofjord (øvre 30 m) er en blanding av tilførsler fra sørlige og sentrale deler av Nordsjøen og Kattegat. Avhengig av vindforhold vil også varierende mengder vann fra Tyskebukta iblandes. Flere store vassdrag drenerer ut i Ytre Oslofjord og påvirker de øvre 0-5 m i særlig grad.
Mens store deler av Ytre Oslofjord er eksponert og dermed utsatt for betydelig vertikal omrøring av vannmassene, så er områdene nært land mindre påvirket av vind og bølger. Kyststrømmen vil hovedsakelig hente inn (advektere) vannmasser fra sør og mot vest, samt at der vil være god vannkontakt i hele området. Vassdragene vil ha størst innvirkning nær elvemunningene, men særlig Glomma influerer hele Hvalerområdet og områdene vest- og nordover. Atlanterhavsvannet vil helst gjøre seg gjeldende under overflatelaget.
Kjemisk miljø som havforsuring er for en stor del drevet av de fysiske forhold og advekterte vannmasser og ferskvann. I den indre delen av Oslofjorden vises hver vinter en svært lav aragonittmetning som er knyttet til ferskvannstilførsel. Til tider vises det undermetning av aragonitt, hvilket betyr begrensninger i kalkskalldannelse. Det betyr at indre Oslofjord er mer følsom for havforsuring enn den mer marine Ytre Oslofjord (Jones mfl. 2020).
10.7.5 - Beskrivelse av miljøverdiene
Plankton
Ytre del av Oslofjorden har høy planteplanktonproduksjon sammenlignet med indre Skagerrak (hav). Årsaken til økt produksjon henger sammen med tilførsel av næringssalter fra vassdrag i regionen, da særlig knyttet til flomsituasjoner (Kristiansen 1997), og transport fra utenforliggende områder. Området overvåkes i regi av Havforskningsinstituttet og innen Miljødirektoratets Økokyst program (f.eks. Fagerli mfl. 2020). Det er tidligere foretatt estimater av primærproduksjon i ulike fjordsystemer fra Oslofjorden til Vestlandet, der den ytre delen av Oslofjorden kom ut med betydelig høyere produksjon enn andre fjordsystemer (Skjolddal mfl. 1997).
Området er påvirket både av kyststrømmen i de øvre vannlag, samt innstrømmende atlanterhavsvann fra den nordlige Nordsjøen i de dypere vannlag. Dette setter sitt preg på dyreplanktonsamfunnet med stor vertikal variasjon i artssammensetning. Området har god vannkontakt med utenforliggende havområder, og dyreplanktonsamfunnet skiller seg i liten grad fra omkringliggende områder (Salt 2019). Ved fast overvåkingsstasjon i Ytre Oslofjord (OF2) domineres dyreplanktonet av hoppekreps som Calanus finmarchicus, C. helgolandicus og Pseudocalanus. I de dypere lagene forekommer krill, pelagiske reker samt C. hyperboreus, knyttet til innstrømming av atlanterhavsvann. Den fremmede arten Mnemiopsis leidyi ble første gang observert i Oslofjorden i 2005. Maneten observeres hvert år i Oslofjorden i perioden juli-oktober, og har en stor evne til å beite ned dyreplanktonet (Hosia og Falkenhaug 2015).
Fisk
I foreslått endreet SVO Ytre Oslofjord finner vi to genetisk ulike ”økotyper” av torsk (Gadus morhua) som vi refererer til som "fjord-" og "Nordsjø-" torsk (Andre mfl. 2016, Knutsen mfl. 2018). Andelen av de to komponentene varierer med sted og år, men generelt er det høyere andel av fjordtypen inne i fjordene og av nordsjøtypen ute i skjærgården. Det må bemerkes at nordsjøtypen kan være fisk som er gytt og lever hele livet på norskekysten og ikke nødvendigvis er fra Nordsjøen. Det er ingen klar romlig segregering av typene og generelt opptrer de sammen, både som juveniler og som eldre. Det antas at de vil segregere til ulike gyteplasser under gyting, siden de opprettholder genetiske forskjeller og i naturen ikke ser ut til å krysse seg med hverandre i noen større grad. En hypotese er at fjordtypen primært vil gyte inne i fjordene mens nordsjøtypen vil gyte ute i skjærgården eller trekke lenger ut i Nordsjøen. Havforskningsinstituttet har gjennomført kartlegging av gytefelter for kysttorsk, men denne kartleggingen har i hovedsak fokusert på indre områder og fjordene. De kartlagte gytefeltene tilknyttet ytre Oslofjord vil derfor være funnet inne i fjorder, men det utelukker ikke at det kan være viktige gytefelt i ytre områder for mer mobile populasjoner av torsk. Hovedgytingen er i februar–mars, og eggene klekkes etter to til tre uker. Den første vinteren når yngelen en lengde på 13–27 cm.
Sei (Pollachius virens) i Nordsjøen gyter fra januar til mars på 150 og 300 meter dyp på eggakanten fra vest av Shetland, Tampen og til Vikingbanken. Størstedelen av yngelen er på kysten av Vestlandet, men sterke årsklasser blir også observert langs Skagerrakkysten. Seibestanden i Nordsjøen er fortsatt over de kritiske nivåene fastsatt for bærekraftig forvaltning, men også her er rådet at kvoten går ned med 25 prosent i 2021 sammenlignet med kvoter for 2020. Selv om foreslått endret SVO Ytre Oslofjord har gode gytefelt for torskefisk, vil de ikke falle inn under EBSA-kriteriene for livshistorisk viktige områder og produktivitet. Det foreslåtte SVO-et som ligger i overgangen mellom fjorder, Kattegat og Skagerrak har bredt artsmangfold hos fisk (Kraufvelin mfl. 2017, Skjæveland 2020). Havforskningsinstituttets strandnotserie har stasjoner innaskjærs i området. Analyser av datasettet viser variasjoner i artssammensetningen av fiskesamfunnet på grunt vann i perioder preget av varmere vann (Barceló mfl. 2016). Fiskesamfunnet på grunt vann i det foreslåtte SVO-et Ytre Oslofjord er preget av fravær av større individer av fiskespisende topp-predatorer, og domineres av meso-predatorer av mindre størrelse (Synnes 2020).
Bunnsamfunn
Foreslått endret SVO Ytre Oslofjord er et område hvor flere fjordsystemer og vannmasser møtes. Grunne eksponerte hardbunnsområder har forekomster av tareskog med høy artsrikhet av sublitorale makroalger (Reitan 2020). Atlanterhavspåvirkningen er stor og stabil i Koster/Hvalerområdet og gir også miljøforhold for korallrev korallrev (Desmophylllum pertusum, tidligere kalt Lophelia pertusa) (Fosså mfl. 2015). Disse revene er unike for Skagerrak/Nordsjøen, og området representerer et av de største kystnære kaldtvannskorallrevområder i verden (Fig. 10.6.1). I 2019 startet prosjektet “Frisk Oslofjord”, et samarbeid mellom blant andre Kartverket, Norges geologiske undersøkelse og Havforskningsinstituttet. Som del av dette prosjektet er det gjennomført flere tokt med visuell kartlegging. I Hvalerrenna ble det observert hardbunnskorallskog og svampsamfunn. Resultater fra disse undersøkelsene vil bli publisert i rapportform og gjennom karttjeneste i løpet av 2021.
Ellers er det lange tidsserier med registrering av biologisk mangfold i bunnsamfunn i det foreslått endrete SVO-et som viser at det er en nedadgående trend i både artsmangfold og individtetthet (Oug ml. 2015, Walday mfl. 2019). Beskrivelse av det nåværende biologiske mangfoldet i tareskogen er presentert i to nylige mastergrader (Prøis 2018, Reitan 2020). Tareskog viste seg dessuten å bli utnyttet av krepsdyr på dypere vann (under 300 m), særlig av dypvannsreker (Pandalus borealis), men også amfipoder når taren var mer nedbrutt (Ramirez-LLodra mfl. 2016).
Det norske reketrålfisket startet i Oslofjorden helt på slutten av 1800-tallet (Hjort og Ruud 1938). Bunntrålfisket etter reker drives i dag på dyp større enn 60 m fra Jærens Rev til svenskegrensen. I Ytre Oslofjord finnes det store sammenhengende bløtbunnsområder der reken (Pandalus borealis) trives (Svennerbanken) (Hjort og Ruud 1938). Nylig gjennomførte sammenligninger med historiske data på bunnsamfunnet i Skagerrak og Kattegat viser endringer til et mer homogent bunnsamfunn med tap av sjeldne arter (Obst mfl. 2018). Se ellers nærmere omtale av dypvannsreken under foreslått nytt SVO Norskerenna (Kap. 10.5.5).
Sjøkreps (Nephrops norvegicus) er avhengig av bløtbunn for å grave sine huler i bunnsedimentet (Johnson mfl. 2013). De store bløtbunnsområdene øst i Skagerrak (Ytre Oslofjord og Hvaler i norsk farvann) gjør at det finnes en stor og produktiv sjøkrepsbestand her. Se ellers nærmere omtale av sjøkrepsen og sjøkrepshabitat i Skagerrak under foreslått nytt SVO Norskerenna (Kap. 10.5.5).
Sjøpattedyr
Steinkobben (Phoca vitulina), den mest tallrike kystselen, har yngle- og hårfellingstid fra midten av juni til begynnelsen av september (Nilssen og Bjørge 2019) . Da er de svært stedbundne og knyttet til stedet der de ble født. Fra andre del av september til tidlig juni, er det næringssøk som har høyeste prioritet. I denne perioden vil de oppsøke steder med god tilgang på mat. Bestanden har økt de siste årene på grunn av en gjenvekst i Ytre Oslofjord og langs Skagerrakkysten der bestandene nærmest ble utradert etter to store virusepidemier i 1988 og 2002. De største koloniene er blant annet ved Kragerø, Færder, Grenland og Hvaler. Deler av området er viktig for steinkobbe hele året (beite- og kasteområde).
Sjøfugl
Hele området i foreslått endret SVO Ytre Oslofjord nedover kysten til Risør er et viktig hekke-, trekk-, og overvintringsområde for sjøfugl. Forekomstene av ærfugl (Somateria mollissima) i ytre Oslofjord er nasjonalt viktige (Systad mfl. 2007, Ottersen mfl. 2010). Arten er utsatt for næringssvikt, forstyrrelser og jakt i dette området (Hanssen mfl. 2020). Makrellterne (Sterna hirundo), som i Norge er rødlistet som sterkt truet (EN), er også en viktig art i Nordsjøen og Skagerrak, særlig i utløpet av Oslofjorden (Systad mfl. 2007, Gasbjerg mfl. 2011). Gråmåke (Larus argentatus), svartbak (Larus marinus) og fiskemåke (Larus canus) er viktige arter utenom hekkesesongen. Fiskemåke er rødlistet som nær truet (NT) i Norge, og betydelige mengder av den norske bestanden overvintrer i dette havområdet. Vinterstid er området viktig for bl.a. britiske lomvi (Uria aalge), hvor arten ikke er rødlistet, men sannsynligvis også for de sørlige forekomstene av lomvi i Norge, der arten har vært i en langvarig tilbakegang og rødlistet som kritisk truet (CR) (Henriksen og Hilmo 2015). Området er også viktig for alkekonge (Alle alle) om høsten.
Ærfugl, sildemåke og storskarv (Phalacrocorax carbo) er definert som indikatorer i forvaltningsplan-sammenheng. Hekkebestandene av ærfugl og sildemåke går tilbake i dette området. Ærfugl har hatt en nedgang på to prosent per år i tiårsperioden 2007-2017 (Anker-Nilssen mfl. 2018). Storskarvbestanden som hekker i dette området, er av underarten Phalacrocorax carbo sinensis (mellomskarv). Denne hekkebestanden var stabil i tiårsperioden 2007-2017 (Anker-Nilssen mfl. 2018).
Det er påvist hendelser med stor dødelighet på våren for ærfugl i området, grunnene er ikke avklart, men inkluderer sult/matmangel. Det ble også funnet blyhagl i en stor andel av fuglene, noe som tyder på utstrakt skadeskyting under den lokale jakten (Hanssen mfl. 2020).
Figur 10.6.3. Kart viser fordeling av hekkende sjøfugl (i farger) med kystnære beiteområder i foreslått endret SVO Ytre Oslofjord og nærliggende områder. Grensene for det foreslåtte SVO-et er markert med rød strek. Skravert felt i nederste venstre figur viser beiteområder for pelagisk beitende sjøfugl.
Steinkobbe. Foto: Carl Freitas Brandt, HI
11 - Referanser generell innledning
Aarflot, JM., Skjoldal, HR., Dalpadado, P., Skern-Mauritzen, M. 2018. Contribution of Calanus species to the mesozooplankton biomass in the Barents Sea. ICES J. Mar. Sci. 75, 2342–2354. https://doi.org/10.1093/icesjms/fsx221
Barrett, R., Lorentsen, S-H., Anker-Nilssen, T. 2006. The status of breeding seabirds in mainland Norway. Atlantic Seabirds 8(3), 97-126.
Beaugrand G., Reid PC., Ibañez F., Lindley JA., Edwards M. 2002. Reorganization of North Atlantic Marine Copepod Biodiversity and Climate. Science, 296, 1692-1694.
Berge J., Varpe Ø., Moline MA., Wold A., Renaud PE., Daase M., Falk-Petersen S. 2012. Retention of ice-associated amphipods: possible consequences for an ice-free Arctic Ocean. Biology Letters 8, 1012-1015. doi:10.1098/rsbl.2012.0517.
Brattegard, T. 2011. Endringer i norsk marin bunnfauna 1997-2010. DN-Utredning 8-2011. 112 s.
Chierici, MA. Fransson, A. 2018. Arctic chemical oceanography at the edge: focus on carbonate chemistry (chapter 13), In: At the edge, Ed. P. Wassmann.ISBN:978-82-8104-299-5, ss.343.
Deschutter, Y., De Schamphelaere, KD., Everaert, G., Mensens, C., De Troch, M. 2019. Seasonal and spatial fatty acid profiling of the calanoid copepods Temora longicornis and Acartia Clausi linked to the environmental stressors in the North Sea. Marine Environmental Research, 144, 92-101. https://doi.org/10.1016/j.marenvres.2018.12.008
Edwards, M., Helaouet, P., Alhaija, RA., Batten, S., Beaugrand, G., Chiba, S., Horaeb, RR., Hosie, G., Mcquatters-Gollop, A., Ostle, C., Richardson, AJ., Rochester, W., Skinner, J., Stern, R., Takahashi, K., Taylor, C., Verheye, HM., Wootton, M. 2016. Global Marine Ecological Status Report: results from the global CPR Survey 2014/2015. SAHFOS Technical Report, 11: 1-32. Plymouth, U.K. ISSN 1744-0750.
Engelhard, HGH., Righton, D., Pinnegar, J. 2013. Climate change and fishing: a century of shifting distribution in North Sea cod. Global Change Biology (2014) 20, 2473-2483. doi: 10.1111/gcb.12513
Eriksen, E., Skjoldal, HR., Gjøsæter, H., Primicerio, R. 2017. Spatial and temporal changes in the Barents Sea pelagic compartment during the recent warming. Progress in Oceanography 151, 206-226. https://doi.org/10.1016/j.pocean.2016.12.009
Faglig forum for norske havområder 2019. Særlig verdifulle og sårbare områder - Faggrunnlag for revisjon og oppdatering av forvaltningsplanene for norske havområder. M-1303/2019
Faglig forum for norske havområder 2019. Om kandidatområder for særlig verdifulle og sårbare områder. M-1471/2019
Fauchald, P., Anker-Nilssen, T., Barrett, RT., Bustnes, JO., Bårdsen, B-J., Christensen-Dalsgaard, S., Descamps, S., Engen, S., Erikstad, KE., Hanssen, SA. mfl. 2015. The status and trends of seabirds breeding in Norway and Svalbard – NINA Report 1151. 84 s.
Fossheim, M, Primicerio, R., Johannesen, E., Ingvaldsen, RB. Aschan, MM. Dolgov. AV. 2015. Recent warming leads to a rapid borealization of fish communities in the Arctic . Nat. Clim. Change 5, 673-677.
Fransner, F., Fröb, F., Tjiputra, T., Chierici, M., Fransson, A., Jeansson, E., Johannessen, T., Jones, E., Lauvset, SK., Ólafsdottír, S. mfl. 2020. Nordic Seas Acidification. Biogeosciences Discussions. Doi: 10.5194/bg-2020-339.
González-Pola, C., Larsen, KMH., Fratantoni, P. og Beszczynska-Möller, A. (Red.). 2020. ICES Report on Ocean Climate 2019. ICES Cooperative Research Reports No. 350. 136 s. https://doi.org/10.17895/ices.pub.7537
Hamilton, CD., Kovacs, KM., Ims, RA., Aars, J., Lydersen, C. 2017. An Arctic predator-prey system in flux: climate change impacts on coastal space use by polar bears and ringed seals. J Anim Ecol 86: 1054-64.
Hamilton, CD., Kovacs, KM., Ims, RA., Lydersen, C. 2018. Haul-out behaviour of Arctic ringed seals: Inter-annual patterns and impacts of current environmental change. Polar Biol. 41: 1063-1082.
Hamilton, CD., Vacquié-Garcia, J., Kovacs, KM., Ims, RA., Lydersen, C. 2019a. Contrasting changes in space use induced by climate change in two Arctic marine mammal species. Biol Lett 15, 20180834. doi:10.1098/rsbl.2018.0834.
Hamilton, CD., Kovacs, KM., Lydersen, C. 2019b. Sympatric seals use different habitats in an Arctic glacial fjord. Mar Ecol Prog Ser 615, 205-220.
Hanssen-Bauer, I., Førland, EJ., Hisdal, H., Mayer, S., Sandø, AB., Sorteberg, A. 2018. Climate in Svalbard 2100– a knowledge base for climate adaptation. M-1242 | 2018. NCCS report no. 1/2019.
ICES 2020. Working Group on the Integrated Assessments of the Norwegian Sea (WGINOR; outputs from 2019 meeting). 52s. ICES Scientific Reports. 2:29. 46 s. http://doi.org/10.17895/ices.pub.5996
IPCC 2019: IPCC Special Report on the Ocean and Cryosphere in a Changing Climate [H-O. Pörtner, DC.
Jiang, L-Q., Carter, BR., Feely, RA., Lauvset, SK., Olsen, A. 2019. Surface ocean pH and buffer capacity: past, present and future. Scientific Reports 9, 18624 (2019). https://doi.org/10.1038/s41598-019-55039-4
Jørgensen, LL., Ljubin, P., Skjoldal, HR., Ingvaldsen, RB., Anisimova, N., Manushin, I. 2015 Distribution of benthic megafauna in the Barents Sea: baseline for an ecosystem approach to management. ICES Journal of Marine Science 72 (2), 595-613.
Kunisch EH., Bluhm BA., Daase M., Gradinger R., Hop H., Melnikov IA., Varpe Ø., Berge J. 2020. Pelagic occurrences of the ice amphipod Apherusa glacialis throughout the Arctic. Journal of plankton research, 42(1), 73-86.
Laidre, KL., Stirling, I., Lowry. L., Wiig, Ø., Heide-Jørgensen, MP., Ferguson, S. 2008. Quantifying the sensitivity of arctic marine mammals to climate-induced habitat change. Ecol Appl 18(2), S97-S125.
Lauvset, SK., Gruber, N., Landschützer, P., Olsen, A., Tjiputra, J. 2015. Trends and drivers in global surface ocean pH over the past 3 decades. Biogeiscience, 12, 1285-1298. doi:10.5194/bg-12-1285-2015
Lauvset, SK., Key, RM., Olsen, A., van Heuven, S., Velo, A., Lin, X., Schirnick, C., Kozyr, A., Tanhua, T., Hoppema, M., Jutterström, S., Steinfeldt, R., Jeansson, E., Ishii, M., Perez, FF., Suzuki, T. og Watelet, S. 2016.: A new global interior ocean mapped climatology: the 1° × 1° GLODAP version 2, Earth Syst. Sci. Data, 8, 325–340, https://doi.org/10.5194/essd-8-325-2016 .
Manno, C., Bednaršek, N., Tarling, GA., Peck, VL., Comeau, S., Adhikari, D., Bakker, DCE., Bauerfeind, E., Bergan, AJ., Berning, MI. mfl. 2017. Shelled pteropods in peril: Assessing vulnerability in a high CO2 ocean. Earth-Science Reviews, 169, 132-145. https://doi.org/10.1016/j.earscirev.2017.04.005
Manushin, IE., Strelkova, NA., Luybin, PA., Zhuravleva, NE., Zakharov, DV., Vyaznikova, VS. 2021. Long-Term Dynamics of the Macrozoobenthos Biomass in the Eastern Barents Sea, 1924-2014. Biology Bulletin (2020) 47, 1088-1098. https://doi.org/10.1134/S1062359020090083
Meld. St. 10 (2010-2011). Oppdatering av forvaltningsplanen for det marine miljø I Barentshavet og havområdene utenfor Lofoten. Tilråding fra Miljøverndepartementet av 11. mars 2011, godkjent i statsråd samme dag. (Regjeringen Stoltenberg II).
Meld. St. 20 (2014–2015) Oppdatering av forvaltningsplanen for Barentshavet og havområdene utenfor Lofoten med oppdatert beregning av iskanten. Tilråding fra Klima- og miljødepartementet 24. april 2015, godkjent i statsråd samme dag. (Regjeringen Solberg). https://www.regjeringen.no/no/dokumenter/meld.-st.-20-2014-2015/id2408321/
Meld. St. 20 (2019–2020) Helhetlige forvaltningsplaner for de norske havområdene — Barentshavet og havområdene utenfor Lofoten, Norskehavet, og Nordsjøen og Skagerrak. Tilråding fra Klima- og miljødepartementet 24. april 2015, godkjent i statsråd samme dag. (Regjeringen Solberg). https://www.regjeringen.no/no/dokumenter/meld.-st.-20-20192020/id2699370/
Meld. St. 29 (2020-2021). Heilskapleg nasjonal plan for bevaring av viktige områder i marin natur. Tilråding fra Klima- og miljødepartementet, 9. april 2021, godkjend i statsråd same dag. (Regjeringen Solberg). https://www.regjeringen.no/no/dokumenter/meld.-st.-29-20192020/id2715224/
Niemi, A., Bednaršek, N., Michel, C., Feely, RA., Williams, W., Azetsu-Scott, K., Walkusz, W., Reist JD. 2021. Biological Impact of Ocean Acidification in the Canadian Arctic: Widespread Severe Pteropod Shell Dissolution in Amundsen Gulf. Front. Mar. Sci. 8:600184. doi: 10.3389/fmars.2021.600184
Nilssen, KT., Bjørge, A. 2019. Status for kystsel – Anbefaling av jaktkvoter 2020. S. 56-69 in A. Bjørge (ed). Forskerutvalg om sjøpattedyr 2019 – Anbefalinger om forskning og forvaltning. Rapport fra Havforskningen Nr. 2020-19.
NP 2019. Avgrensning av SVO Havområdene rundt Svalbard, inkludert Bjørnøya - Notat til Faglig Forum for norske havområder. Norsk Polarinstitutt 2019. 20 s.
O ’Brien, TD., Wiebe, PH., Falkenhaug, T. (Eds). 2013. ICES Zooplankton Status Report 2010/2011. ICES Cooperative Research Report No. 318. 208 pp.
Olsen, E., von Quillfeldt, CH. (red.) 2003. Identifisering av særlig verdifulle områder i Barentshavet og Lofoten. Havforskningsinstituttet/Norsk Polarinstitutt.
Ottersen, G. Auran, JA. (red.) 2007. Arealrapport med miljø og ressursbeskrivelse. Fisken og havet 6/2007, Havforskningsinstituttet.
Ottersen, G., Postmyr, E., Irgens, M. (red.) 2010. Faglig grunnlag for en forvaltningsplan for Nordsjøen og Skagerrak: Arealrapport. Fisken og Havet nr. 6-2010/TA-nummer 2681/2010, Havforskningsinstituttet, Bergen. 196 s.
Rogne, S., Johnsen, E. 2019. LoVe: våre øyne og ører under vann. Hentet fra https://www . hi.no/hi/nyheter/2019/januar/love-vare-oyne-og-orer-under-vann
Shackell, NL., Bundy, A., Nye, JA., Link, JS. 2012. Common large-scale responses to climate and fishing across Northwest Atlantic ecosystems. ICES J Mar Sci 69, 151–162.
Stige LC., Eriksen E., Dalpadado P., Kotaro O. 2019. Direct and indirect effects of sea ice cover on major zooplankton groups and planktivorous fishes in the Barents Sea. ICES Journal of Marine Science, in press. https://doi.org/10.1093/icesjms/fsz063
St. John, MA., Borja, A., Chust, G., Heath, M., Grigorov, I., Mariani, P., Martin, AP., Santos, RS. 2016. A Dark Hole in Our Understanding of Marine Ecosystems and Their Services: Perspectives from the Mesopelagic Community. Front. Mar. Sci. 17. https://doi.org/10.3389/fmars.2016.00031
St.meld. nr. 8 (2005-2006) Helhetlig forvaltning av det marine miljø i Barentshavet og havområdene utenfor Lofoten (forvaltningsplan). Tilråding fra Miljøverndepartementet av 31. mars 2006, godkjent i statsråd samme dag. (Regjeringen Stoltenberg II). https://www.regjeringen.no/no/dokumenter/stmeld-nr-8-2005-2006-/id199809/
Sutton, TT., Clark, MR., Dunn, DC., Halpin, PN., Rogers, AD., Guinotte, J., Bograd, SJ., Angel, JAA., Wishner, K., Haedrich, R.L. mfl. 2017. A gobal biogeographic classification of the mesopelagic zone. Deep Sea Research Part 1: Oceanographic Research Papers 126, 85-102. https://doi.org/10.1016/j.dsr.2017.05.006
Systad, GHR., Fauchald, P., Descamps, S., Christensen-Dalsgaard, S., Strøm, H., Tarroux, A. 2019. Identifisering av viktige områder for sjøfugl i norske havområder – innspill til forvaltningsplanarbeidet 2018. NINA Rapport 1627. Norsk institutt for naturforskning.
van Leeuwe, M A., Tedesco, L., Arrigo, K.R., Assmy, P., Campbell, K., Meiners, K.M., Rintala, J-M., Selz, V., Thomas, D.N., Stefels, J. 2018. Microalgal community structure and primary production in Arctic and Antarctic sea ice: A synthesis. Elem Sci Anth, 6: 4. DOI: https://doi.org/10.1525/elementa.267
von Quillfeldt, CH. (red.) 2018. Miljøverdier og sårbarhet i iskantsonen. Norwegian Polar Institute, Brief report no. 047, 262 s.
Wenneck, TdL. 2005. Mengdeindekser for 19 “ikke-kommersielle” arter i Barentshavet basert på bunnfisketokt - “Vintertoktet” - i perioden 1981-2003. Fisken og Havet nr. 2 2005. Havforskningsinstituttet, Bergen. 70 s.
12 - Referanser Barentshavet og områder utenfor Lofoten
Al-Habahbeh, AK., Kortsch, S., Bluhm, BA., Beuchel, F., Gulliksen, B., Ballantine, C., Cristini, D. and Primicerio, R., 2020. Arctic coastal benthos long-term responses to perturbations under climate warming. Philosophical Transactions of the Royal Society A, 378(2181). s. .20190355.
Aarflot, J.M., Aksnes, D., Opdal, A.F., Skjoldal, H.R., Fiksen, Ø. 2019. Caught in broad daylight. Topographic constrains of zooplankton depth distributions. Limnol. Oceanogr. 64, 849–859. https://doi.org/10.1002/lno.11079 .
Aarflot, J.M., Skjoldal, H.R., Dalpadado, P., Skern-Mauritzen, M. 2018. Contribution of Calanus species to the mesozooplankton biomass in the Barents Sea. ICES J. Mar. Sci. 75, 2342–2354. https://doi.org/10.1093/icesjms/fsx221
Aars J. 2013. Variation in detection probability of polar bear maternity dens. Polar Biol 36: 1089-96.
Aars J, Marques TA, Buckland ST, Andersen M, Belikov S, Boltunov A, Wiig O. 2009. Estimating the Barents Sea polar bear subpopulation size. Mar Mamm Sci 25: 35-52.
Aars J, Marques TA, Lone K, Andersen M, Wiig Ø, Bardalen Fløystad IM, Hagen SB, Buckland ST. 2017. Polar bear population structure and trend in the western Barents Sea. Polar Res 36: 1374125.
Ahonen H, Stafford KM, de Steur L, Lydersen C, Wiig Ø, Kovacs KM. 2017. The underwater soundscape in western Fram Strait: breeding ground of Spitsbergen’s endangered bowhead whales. Mar Poll Bull 123: 97-112.
Ahonen H, Stafford KM, Lydersen C, de Steur L, Kovacs KM. 2019. A multi-year study of narwhal occurrence in the western Fram Strait—detected via passive acoustic monitoring. Polar Res. 38: 3468.
Albert O.T. 2003. Migration from nursery to spawning area in relation to growth and maturation of Greenland Halibut (Reinhardtius hippoglossoides) in the Northeast Arctic. Journal of Northwest Atlantic Fisheries Science. 3 1: 113- 125.
Albert, OT., Nilssen, EM., Stene, A., Gundersen, A. C., Nedreaas, K. H. 2001. Maturity classes and spawning behaviour of Greenland halibut (Reinhardtius hippoglossoides). Fisheries Research, 51(2-3), 217-228. doi:10.1016/s0165-7836(01)00247-8.
Albert, OT., Vollen, T. 2015. A major nursery area around the Svalbard archipelago provides recruits for the stocks in both Greenland halibut management areas in the Northeast Atlantic. ICES Journal of Marine Science: Journal du Conseil, 72(3), 872-879. doi:10.1093/icesjms/fsu191
Albretsen J, Aure J, Sætre R. Danielssen DS, 2012. Climatic variability in the Skagerrak and coastal waters of Norway. ICES J. Mar. Sci., 69, 758-763. doi: 10.1093/icesjms/fsr187
Allen, JT, Brown L, Sanders R, Moore CM, Mustard A, Fielding S, Lucas M, Rixen M, Savidge G, Henson S, Mayor D, 2005. Diatom carbon esport enhanced by silicate upwelling in the northeast Atlantic. Nature. 437, 728-732.
Ambrose WG., von Quillfeldt CH., Clough LM., Tilney PVR., Tucker T. 2005. The sub-ice algal community in the Chukchi sea: large- and small-scale patterns of abundance based on images from a remotely operated vehicle. Polar Biology 28, 784-795, doi:10.1007/s00300-005-0002-8.
Amélineau, F., Grémillet, D., Harding, AMA., Walkusz, W., Choquet, R., Fort, J. 2019. Arctic climate change and pollution impact little auk foraging and fitness across a decade. Scientific Reports 9(1), DOI: 10.1038/s41598-018-38042-z
Andersen, LW., Born, EW., Gjertz, I., Wiig, Ø., Holm, L-E., Bendixsen, C. 1998. Population structure and gene flow of the Atlantic walrus (Odobenus rosmarus rosmarus) in the eastern Atlantic Arctic based on mitochondrial DNA and microsatellite variation. Molecular Ecol 7: 1323–1336.
Andersen, LW., Jacobsen, MW., Lydersen, C., Semenova, V., Boltunov, A., Born, EW., Wiig, Ø., Kovacs, KM. 2017. Walruses (Odobenus rosmarus rosmarus) in the Pechora Sea in the context of contemporary population structure of Northeast Atlantic walruses. Biol J Linn Soc 122: 897–915.
Andersen, LW., Lydersen, C., Frie, AK., Rosing-Asvid, A., Hauksson E., Kovacs, KM. 2011. A population on the edge: genetic diversity and population structure of the world’s northernmost harbour seals (Phoca vitulina). Biol J Linn Soc 102: 420-439.
Andersen, M., Derocher, AE., Wiig, O., Aars, J. 2012. Polar bear (Ursus maritimus) maternity den distribution in Svalbard, Norway. Polar Biol 35: 499-508.
Anderson LG., Falck. E., Jones, EP., Jutterström, S., Swift, JH. 2004. Enhanced uptake of atmospheric CO 2 during freezing of seawater: A field study in Storfjorden, Svalbard. J Geophys Res, Oceans Vol 109, C6, https://doi.org/10.1029/2003JC002120
Anderson, M., Orvik, KA., LaCasce, JH., Koszalka, I., Mauritsen, C. 2011. Variability of the Norwegian Atlantic Current and associated eddy field from surface drifters. J Geophys Res, Oceans Vol 116, C08032, doi: 10.1029/2011JC007078.
Anker-Nilssen, T., Barrett, RT., Lorentsen, S-H., Strøm, H., Bustnes, JO., Christensen-Dalsgaard, S., Descamps, S., Erikstad, KE., Fauchald, P., Hanssen, SA., Lorentzen, E., Moe, B., Reiertsen, TK., Systad, GH. 2015. SEAPOP. De ti første årene. Nøkkeldokument 2005-2014. SEAPOP. Norsk institutt for naturforskning, Norsk Polarinstitutt & Tromsø Museum – Universitetsmuseet. Trondheim, Tromsø. 58 s.
Anker-Nilssen, T., Bustnes,JO.; Benjaminsen, S., Christensen-Dalsgaard, S. Dehnhard, N., Descamps, S., Erikstad, KE., Fauchald, P., Hanssen, SA., Langset, M., Lorentsen, SH., Lorentzen, E., Moe, B., Reiertsen, TK., Strøm, H., Systad, GH., Bruset, B., Sivertsen, K. Sjøfugl i Norge 2019. Trondheim: SEAPOP 2020.
Archer, SD., Verity, PG., Stefels, J. 2000. Impact of microzooplankton on the progression and fate of the spring bloom in fjords of northern Norway. Aquat Microb Ecol. 22, 27–41.
Ardyna, M., Mundy, C.J., Mayot, N., Matthes, LC., Oziel, L., Horvat, C., Leu, E., Assmy, P., Hill, V., Matrai, PA., Gale, M., Melnikov, IA. Arrigo, KR. 2020. Under-Ice Phytoplankton Blooms: Shedding Light on the “Invisible” Part of Arctic Primary Production. Front. Mar. Sci. 7:608032. doi: 10.3389/fmars.2020.608032
Arndt CE., Swadling KM. 2006. Crustacea in Arctic and Antarctic sea ice: distribution, diet and life history strategies. Adv. Mar. Biol. 51, 197-315.
Arneberg, P., Jelmert, A. (Red) 2017. Status for miljøet i Barentshavet og ytre påvirkning– rapport fra Overvåkingsgruppen 2017. Fisken og Havet, særnummer 1b-2017, Havforskningsinstituttet.
Assmy P., Ehn J.K., Fernández-Méndez M., Hop H., Katlein C., Sundfjord A., Bluhm K., Daase M., Engel A., Fransson A., Granskog M.A., Hudson S.R., Kristiansen S., Nicolaus M., Peeken I., Renner A.H.H., Spreen G., Tatarek A., Wiktor J. 2013. Floating Ice-Algal Aggregates below Melting Arctic Sea Ice. PLoS ONE 8, e76599.
Assmy P., Fernández-Méndez M., Duarte P., Meyer A., Randelhoff A., Mundy CJ., Olsen LM., Kauko HM., Bailey A., Chierici M., Cohen L., Doulgeris AP., Ehn JK., Fransson A., Gerland S., Hop H., Hudson SR., Hughes N., Itkin P., Johnsen G., King JA., Koch BP., Koenig Z., Kwasniewski S., Laney SR., Nicolaus M., Pavlov AK., Polashenski CM., Provost C., Rösel A., Sandbu M., Spreen G., Smedsrud LH., Sundfjord A., Taskjelle T., Tatarek A., Wiktor J., Wagner PM., Wold A., Steen H., Granskog MA. 2017. Leads in Arctic pack ice enable early phytoplankton blooms below snow-covered sea ice. Scientific Reports 7, 40850.
Barber DG., Hop H., Mundy CJ., Else B., Dmitrenko IA., Tremblay J-E., Ehn JK., Assmy P., Daase M., Candlish LM., Rysgaard S. 2015. Selected physical, biological and biogeochemical implications of a rapidly changing Arctic Marginal Sea Ice Zone. Progress in Oceanography 139, 122-150.
Barton, BI., Lenn, YD., Lique, C. 2018. Observed Atlantification of the Barents Sea Causes the Polar Front to Limit the Expansion of Winter Sea Ice. J. Phys. Oceanogr. 48, 8, 1849–1866.
Basedow SL., McKee D., Lefering I., Gislason A., Daase M., Trudnowska E., Egeland ES., Choquet M., Falk-Petersen S. 2019. Remote sensing of zooplankton swarms. Scientific reports, DOI:10.1038/s41598-018-37129-x.
Basedow, S.L., Sundfjord, A., von Appen, W-J., Halvorsen, E., Kwasniewski, S., Reigstad, M. 2018. Seasonal variation in transport of zooplankton into the Arctic Basin through the Atlantic gateway, Fram Strait. Front. Mar. Sci. 5:194. doi: 10.3389/fmars.2018.00194
Becker M., A. Olsen, P. Landschützer, A. Omar, G. Rehder, C. Rödenbeck og I. Skjelvan. 2020 The northern European shelf as increasing net sink for CO2. Biogeosciences Discuss. https://doi.org/10.5194/bg-2019-480 , in review 2020.
Belkin, IM., Cornillon, PC., Sherman, K, 2009. Fronts in large marine ecosystems. Prog Oceanogr. 81, 223–236.
Beuchel, F., Gulliksen, B. and Carroll, M.L., 2006. Long-term patterns of rocky bottom macrobenthic community structure in an Arctic fjord (Kongsfjorden, Svalbard) in relation to climate variability (1980–2003). Journal of Marine Systems, 63(1-2). .35-48.
Berenboim, BI., Dolgov, AV., Korzhev, VA., Yaragina, NA. 2000. The impact of cod on the dynamics of Barents Sea shrimp (Pandalus borealis) as determined by multispecies models. Journal of Northwest Atlantic Fishery Science 27, 69-75.
Berg, PR, Jorde PE, Glover KA,Dahle G, Taggart,JB Korsbrekke K, Dingsør GE, Skjæraasen JE, WrightPJ, Cadrin S, Knutsen H, Westgaard JI, Genetic structuring in Atlantic haddock contrasts with current management regimes, ICES Journal of Marine Science, fsaa204, https://doi.org/10.1093/icesjms/fsaa204
Berge J., Varpe Ø., Moline M.A., Wold A., Renaud P.E., Daase M., Falk-Petersen S. 2012. Retention of ice-associated amphipods: possible consequences for an ice-free Arctic Ocean. Biology Letters 8, 1012-1015. doi:10.1098/rsbl.2012.0517.
Bergstad OA, Johannesen E, Høines Å, Ellingsen KE, Lien VS, Byrkjedal I, Yoccoz NG, Tveraa T, Wienerrother R, Langhelle G, Wenneck TdL, 2017. Dermersal fish assemblages in the boreo-Arctic shelf waters around Svalbard during the warm period 2007-2014. Polar Biol, 41, 125-142. https://doi.org/10.1007/s00300-017-2176-2
Bergstad, OA., Jørgensen, T., Dragesund, 0. 1987. Life history and ecology of the gadoid resources of the Barents Sea. Fisheries Research. 5(2-3): 119-161.
BirdLife International 2021. Important Bird Areas factsheet: Lofoten. Downloaded from http://www.birdlife.org on 21/04/2021.
Bjørge, A., Øien, N. 1999. Statusrapport for Havforskningsinstituttets overvåkning av kystsel. Havforskningsinstituttet, Rapport SPS-9904. 35 s.
Bjørke, H., Sundby, S. 1987. Distribution and abundance indices of postlarval and 0-group cod. I: Loen, H. (red.) The effect of oceanographic conditions on distribution and population dynamics of commercial fish stocks in the Barents Sea. Proceedings of the third Soviet-Norwegian Symposioum, Murmansk, 26-28 May 1986. Havforskningsinstituttet, Bergen 1987.
Blachowiak-Samolyk K, Søreide JE, Kwasniewski S, Sundfjord A, Hop H, Falk-Petersen S, Hegseth EN. 2008. Hydrodynamic control of mesozooplankton abundance and biomass in northern Svalbard waters (79–81°N). Deep Sea Research Part II: Topical Studies in Oceanography 55(20-21): 2210-2224. doi.org/10.1016/j.dsr2.2008.05.018.
Blanchet MA, Lydersen C, Ims RA, Kovacs KM. 2015. Seasonal, oceanographic and atmospheric drivers of diving behaviour in a temperate seal species living in the High Arctic. PLoS ONE 10: e0132686. doi: 10.1371/journal.pone.0132686.
Blanchet MA, Lydersen C, Ims RA, Lowther AD, Kovacs KM. 2014. Harbour seal Phoca vitulina movement patterns in the high-Arctic archipelago of Svalbard, Norway. Aquat Biol 21: 167-181.
Bluhm, B.A., Gebruk, AV., Gradinger, R., Hopcroft, RR., Huettmann, F., Kosobokova, KN., Sirenko, BI., Weslawski,, JM. 2011. Arctic marine biodiversity: an update of species richness and examples of biodiversity change. Oceanography, 24(3), s. 232-248.
Bluhm, BA., Gradinger, R. 2008. Regional variability in food availability for Arctic marine mammals. Ecological Applications, 18(sp2), s. S77-S96.
Bluhm, BA., Hop, H., Melnikov, IA., Poulin, M., Vihtakari, M., Collins, E. Gradinger, R., Juul-Pedersen, T., von Quillfeldt, C. 2017b. Sea-ice biota. S. 33-62 in CAFF State of the Arctic Marine Biodiversity Report. Conservation of Arctic Flora and Fauna International Secretariat, Akureyri, Iceland. 978-9935-431-63-9.
Bluhm, BA., Hop, H., Vihtakari, M., Gradinger, R., Iken, K., Melnikov, IA., Søreide, JE. 2018. Sea ice meiofauna distribution on local to pan‐Arctic scales. Ecology and evolution 8(4), s.2350-2364.
Bluhm, B.A., Janout, M.A., Danielson, S.L., Ellingsen, I., Gavrilo, M., Grebmeier, J.M., Hopcroft, R.R., Iken, K., Ingvaldsen, R.B. Jørgensen, L.L., Kosobokova, K.N., Kwok, R., Polyakov, I.V., Renaud, P.E., Carmack, E.C., 2020. The pan-Arctic continental slope: sharp gradients of physical processes affect pelagic and benthic ecosystems. Front. Mar. Sci., 20 Nov 2020 | https://doi.org/10.3389/fmars.2020.544386
Bluhm, BA., Swadling, K., Gradinger, R. 2017a. Chapter 16: Sea ice as habitat for macrograzers. S. 394-414 in Brown, T. A. mfl. 2017. "Transfer of ice algae carbon to ice-associated amphipods in the high-Arctic pack ice environment." Journal of Plankton Research 39 (4): 664-674. Thomas D.N. (ed.). Sea ice 3rd edition, Wiley Blackwell.
Boetius, A., Albrecht, S., Bakker, K., Bienhold, C., Felden, J., Fernandez-Mendez, M., Hendricks, S., Katlein, C., Lalande, C., Krumpen, T., Nicolaus, M., Peeken, I., Rabe, B., Rogacheva, A., Rybakova, E., Somavilla, R., Wenzhofer F. 2013. Export of algal biomass from the melting Arctic sea ice. Science, 339(6126), 1430-1432.
Boetius, A., Anesio, AM., Deming, JW., Mikucki, JA., Rapp, J.Z. 2015. Microbial ecology of the cryosphere: sea ice and glacial habitats. Nat Rev Micro 13:677-690.
Boitsov, VD., Dolgov, AV., Krysov, AI., Seliverstova, EI., Shevelev, MS. (Red) 2013. Polar cod of the Barents Sea. PINRO Press, Murmansk (in Russian)
Brattegard, T. 2011. Endringer i norsk marin bunnfauna 1997-2010. DN-Utredning 8-2011. 112 s.
Broms, C. 2015. Dyreplankton i Norskehavet. S. 100-101 i Bakketeig, I.E., Gjøsæter, H., Hauge, M., Sunnset, B.H., Toft, K.Ø. (red.). Havforskningsrapporten. Ressurser, miljø og akvakultur på kysten og i havet. Fisken og Havet, særnummer 1-2015.
Broms, C., 2016. Dyreplankton i Norskehavet. S. 98-99 i Bakketeig, I.E., Hauge, M., Kvamme, C., Sunnset, B.H., Toft, K.Ø. (red.) Havforskningsrapporten. Ressurser, miljø og akvakultur på kysten og i havet. Fisken og Havet, særnummer 1 -2016
Broms, C., 2017. Dyreplankton i Norskehavet. S. 98-99 i Bakketeig, I.E., Hauge, M., Kvamme, C (red.) Havforskningsrapporten. Ressurser, miljø og akvakultur på kysten og i havet. Fisken og Havet, særnummer 1 –2017.
Brown TA., Assmy P., Hop H., Wold A., Belt ST. 2017. Transfer of ice algae carbon to ice-associated amphipods in the high-Arctic pack ice environment. Journal of Plankton Research 39(4): 664-674. https://doi.org/10.1093/plankt/fbx030..
Båtevik, T. 2020. Estimating natural mortality rates for northern shrimp (Pandalus borealis) from a wide range of its latitudinal distribution in the Northeast Atlantic. Masteroppgave. Universitetet i Bergen. Det matematisk-naturvitenskapelige fakultet. 79 s.
Cadrin, S. X., Bernreuther, M., Daníelsdóttir, A. K., Hjörleifsson, E., Johansen, T., Kerr, L., Kristinsson, K., Mariani, S., Nedreaas, K., Pampoulie, C., Planque, B., Reinert, J., Saborido-Rey, F., Sigurðsson, T., Stransky, C. 2010. Population structure of beaked redfish, Sebastes mentella: evidence of divergence associated with different habitats. – ICES Journal of Marine Science, 67: 1617–1630.
CAFF. 2017. State of the Arctic Marine Biodiversity Report. Conservation of Arctic Flora and Fauna International Secretariat, Akureyri, Iceland. 978-9935-431-63-9.
Carmack, E., Barber, D., Christensen, J., Macdonald, R., Rudels, B., Sakshaug, E., 2006. Climate variability and physical forcing of the food webs and the carbon budget on panarctic shelves. Progress in Oceanography, 71 (2006), 145-181.
Carscadden, J.E., Gjøsæter, H., Vilhjálmsson, H., 2013a. Recruitment in the Barents Sea, Icelandic, and Eastern Newfoundland/Labrador Capelin (Mallotus villosus) Stocks. Progress in Oceanography , 114, 84-96
Carscadden, J.E., Gjøsæter, H., Vilhjálmsson, H., 2013b. A comparison of recent changes in distribution of capelin (Mallotus villosus) in the Barents Sea, around Iceland and in the Northwest Atlantic. Progress in Oceanography , 114, 64-83.
Carstensen, J., Olszewska, A., Kwasniewski, S. 2019. Summer mesozooplankton biomass distribution in the West Spitsbergen Current 2001-2014. Front. Mar. Sci. 6:202. doi: 10.3389/fmars.2019.00202
Carroll ML., Carroll J. 2003. The Arctic Seas. S. 127-156 in Black K., Shimmield G. (Red.) Biogeochemistry of Marine Systems, Blackwell Publ Ltd, Oxford.
Carroll ML., Denisenko SG., Renaud PE., Ambrose WG. 2008. Benthic infauna of the seasonally ice-covered western Barents Sea: Patterns and relationships to environmental forcing. Deep-Sea Res. II 55:2340-2351.
Castano-Primo, R., Vikebø, F. B., Sundby, S. 2014. A model approach to identify the spawning grounds and describing the early life history of Northeast Arctic haddock (Melanogrammus aeglefinus). ICES Journal of Marine Science, 71: 2505-2514.
Cheng B, Zhang Z, Vihma T, Johansson M, Bian L, Li Z, Wu H, 2008. Model experiments on snow and ice thermodynamics in the Arctic Ocean with CHINARE 2003 data. J Geophys Res 113, C09020. doi:10.1029/2007JC004654
Cheng B, Vihma T, Rontu L, Kontu A, Kheyrollah Pour H, Duguay C, Pulliainen J, 2014. Evolution of snow and ice temperature, thickness and energy balance in Lake Orajärvi, northern
Cherkasheva, A., Bracher, A., Melsheimer, C., Köbler, EM., Nöthing, EM., Bauerfield, E., Boetius, A. 2014. Influence of the physical enviroment on polar phytoplankton blooms: a case study in the Fram Strait. J.Mar,Sys, 132:196-207. doi.org/10.1016/j.jmarsys.2013.11.008
Chierici, M., Fransson, A. 2009. Calcium carbonate saturation in the surface water of the Arctic Ocean: undersaturation in freshwater influenced shelves. Biogeoscience, 6, 2421-2432.
Chierici, M., Fransson, A. 2018. Arctic chemical oceanography at the edge: focus on carbonate chemistry (chapter 13), I: At the edge, Ed. P. Wassmann. ISBN:978-82-8104-299-5, ss.343.
Chierici, M., Skjelvan, I., Norli, M., Jones, E., Børsheim, K., Lauvset, S., Lødemel, H., Sørensen, K., King, A. og Johannessen, T. 2017. Overvåking av havforsuring i norske farvann i 2016, Rapport, Miljødirektoratet, M-776|2017, Tech. Rep.
Chierici, M., Vernet, M., Fransson, A., Børsheim, Y. 2019. Net community production and carbon exchange from winter to summer in the Atlantic Water inflow to the Arctic Ocean. Frontiers in Marine Science, doi: 10.3389/fmars.2019.00528.
C hristensen-Dalsgaard, S., Anker-Nilssen, T., Dehnhard, N., Strøm, H. (redaktører), Bustnes, J.O., Benjaminsen, S., Descamps, S., Erikstad, K.E., Fauchald, P., Hanssen, S.A., Langset, M., Lorentsen, S.-H., Lorentzen, E., Moe, B., Reiertsen, T.K., Systad, G.H.R. 2020. Sjøfugl i Norge 2019. Resultater fra SEAPOP-programmet. SEAPOP: 28 s.
Christensen-Dalsgaard, S., May, R.., Lorentsen, S-H. 2018. Taking a trip to the shelf: Behavioural decisions are mediated by the proximity to foraging habitats in the black-legged kittiwake. Ecology and Evolution 8: 866-878. https://doi.org/10.1002/ece3.3700 .
Christiansen JS. 2012. The TUNU programme: Euro-Arctic marine fishes – diversity and adaptation. S. 35-50 i Verde C og di Prisco G (eds.) Adaptation and evolution in marine environments: from pole to pole. Springer-Verlag, Berlin Heidelberg.
Cochrane, S., Pearson, T., Greenacre, M., Costelloe, J., Ellingsen, I., Dahle, S., Gulliksen, B. (2012). Benthic fauna and functional trails along a Polar Front transect in the Barents Sea - Advancing tool for ecosystem-scale assessments. Journal of Marine Systems 204-217.
Cooke J., Reeves R. 2018. Balaena mysticetus (East Greenland-Svalbard-Barents Sea subpopulation). The IUCN Red List o f Threatened Species 2018: e.T2472A50348144. doi.org/10.2305/IUCN.UK.2018-1.RLTS.T2472A50348144.en. downloaded on 15 January 2020.
Daase M., Eiane K. 2007. Mesozooplankton distribution in northern Svalbard waters in relation to hydrography. Polar Biology, 30, 969– 981.
Daase, M., Eiane, K., Aksnes, DL. Vogedes, D. 2008. Vertical distribution of Calanus spp. And Metridia longa at four Arctic locations. Mar. Biol. Res. 4:193-207
Daase, M., Falk-Petersen, S., Varpe, O., Darnis, G., Soreide, JE., Wold, A., Leu, E., Berge, Philippe, B., Fortier, L. 2013. Timing of reproductive events in the marine copepod Calanus glacialis: a pan-Arctic perspective Canadian Journal of Fisheries and Aquatic Sciences, 70, s. 871-884.
Dahlke, S., Hughes, NE., Wagner, PM., Garland, S., Wawrzyniak, T., Ivanov, B., Maturilli, M. 2020. The observed recent surface air temperature development across Svalbard and concurring footprints on local sea ice cover. Int. J. Climatol. 2020: 1-20.
Dalpadado, P., Arrigo, KR., Hjøllo, SS., Rey, F., Ingvaldsen, RB., Sperfeld, E., van Dijken, G., Stige, LC., Olsen, A., Ottersen, G., 2014. Productivity in the Barents Sea - Response to Recent Climate Variability. PLoS ONE 9(5): e95273. doi:10.1371/journal.pone.0095273.
Dalpadado, P. Arrigo, KR., van Dijken, G.L., Skjoldal, HR., Bagøien, E., Dolgov, AV., Prokopchuk, IP., Sperfeld, E, 2020. Climate effects on temporal and spatial dynamics of phytoplankton and zooplankton in the Barents Sea. Prog. Oceanogr. 185:1-20.
Dalpadado, P., Ellertsen, B., Melle, W., Skjoldal, H.R. 1998. Summer distribution patterns and biomass estimates of microzooplankton and micronekton in the Nordic Seas. Sarsia, 83:2, 103-116.
Dalpadado, P., Ingvaldsen, RB., Stige, LC., Bogstad, B., Knutsen, T., Ottersen, G., Ellertsen, B., 2012. Climate effects on Barents Sea ecosystem dy namics. ICES J. Mar. Sci. 69, 1303–1316. https://doi.org/10.1093/icesjms/fss063
Dalpadado, P., Mowbray, F. 2013. Comparative analysis of feeding ecology of capelin from two shelf ecosystems, off Newfoundland and in the Barents Sea. Prog. Oceanogr. 114, 97–105. https://doi.org/10.1016/j.pocean.2013.05.007.
Dalsgaard, J., St. John, M., Kattner, G,, Müller-Navarra, D, Hagen, W.2003. Fatty acid trophic markers in the pelagic marine environment. Advances in Marine Biology 46.. 227318.
Degen R., Jørgensen L., Ljubin P., Ellingsen IH., Pehlke H., Brey T. 2016. Patterns and drivers of megabenthic secondary production on the Barents Sea shelf. Marine Ecology Progress Series, 546, s. 1-16. doi: 10.3354/meps11662
Degerlund, M., Eilertsen, HC. 2010. Main Species Characteristics of Phytoplankton Spring Blooms in NE Atlantic and Arctic Waters (68–80° N). Estuaries and Coasts 33,242–269. DOI 10.1007/s12237-009-9167-7
de Moura Neves, B., Edinger, E., Hayes, VW., Devine, B., Wheeland, L. Layne, G. 2018. Size metrics, longevity, and growth rates in Umbellula encrinus (Cnidaria: Pennatulacea) from the eastern Canadian Arctic. Arctic Science, 4(4), s.722-749.
Derocher, A., Andersen, M., Wiig, Ø., Aars, J., Hansen, E., Biuw, M. 2011. Sea ice and polar bear den ecology at Hopen Island, Svalbard. Mar Ecol Prog Ser 441: 273-79.
Derocher AE, Wiig Ø, Andersen M. 2002. Diet composition of polar bears in Svalbard and the western Barents Sea. Polar Biol 25: 448-52.
Docquier D., Fuentes-Franco R., Koenigk T., Fichefet T. 2020. Sea Ice—Ocean Interactions in the Barents Sea Modeled at Different Resolutions. Frontiers in Earth Science 8, 172.
Dolgov, AV., Orlova, E., Johannesen, E., Bogstad, B., Rudneva, G., Dalpadado, P., Mukhina, N., 2011. Planktivorous fishes, in: Jakobsen, T., Ozhigin, VK. (Red.), The Barents Sea. Ecosystem, Resources, Management. Half a Century of Russian–Norwegian Cooperation. Tapir Press, Trondheim, Norway, s. 438–454.
Dolgov, AV., Smirnov, OV. 2001. The role of Greenland halibut, Reinhardtius hippoglossoides (Walbaum), in the ecosystem of the Barents Sea. Deep-sea Fisheries: Symp./NAFO/ICES/CSIRO (12–14 September 2001): Abstracts. Paper 1.10.
Dragesund, O, Hylen, A, Olsen, S, Nakken, O. 2008. The Barents Sea 0-group surveys; a new concept of pre-recruitment studies. I: Nakken, O. (Red.), Norwegian spring-spawning herring and Northeast Arctic cod – 100 years of research and management, Tapir Academic Press, Trondheim, s 119-136.
Drevetnyak, K., Nedreaas, KH. 2009. Historical movement pattern of juvenile beaked redfish (Sebastes mentella Travin) in the Barents Sea as inferred from long-term research survey series. Marine Biology Research, 5(1), 86-100. doi:10.1080/17451000802534865
Durner, GM., Douglas, DC., Nielson, RM., Amstrup, SC., McDonald, TL., Stirling, I., Mauritzen, M., Born, EW., Wiig, Ø., DeWeaver, E., mfl. 2009. Predicting 21st-century polar bear habitat distribution from global climate models. Ecol Monographs 79: 25-58.
Eamer, J., Donaldson, GM., Gaston, AJ., Kosobokova, KN., Larusson, KF., Melnikov, IA., Reist, JD., Richardson, E., Staples, L., von Quillfeldt, C.H. 2013. Life Linked to Ice: A guide to sea-ice associated biodiversity in this time of rapid change. CAFF Assessment Series No. 10. Conservation of Arctic Flora and Fauna, Iceland.
Edvardsen, A., Pedersen, JM., Slagstad, D., Semenova, T., Timonin, A. 2006. Distribution of overwintering Calanus in the North Norwegian Sea. Ocean Sci. 2:87–96.
Ehrlich J., Schaafsma F.L., Bluhm B.A., Peeken I., Castellani G., Brandt A., Flores H. 2020. Sympagic Fauna in and Under Arctic Pack Ice in the Annual Sea-Ice System of the New Arctic. Frontiers in Marine Science, 7.
Eliassen, J.E., 1983. Occurrence, gonad development and age distribution of the roughhead grenadier (Macrocurusberglax Lacepde) (Gadiformes , Macrouridae) along the Continental slope of northern Norway. ICES CM 1983/G:41, 19 s.
Ellertsen, B., Fossum, P., Solemdal, P., Sundby, S. 1989. Relation between temperature and survival of eggs and first feeding larvae of northeast Arctic cod (Gadus morhua L.) Rapp.P.-v. Reun. Cons. int. Explor. Mer, 191: 209-219.
Ericson Y., Falck, E., Chierici, M., Fransson, A., Kristiansen, S. 2019. Marine CO 2 system variability in a high arctic tidewater-glacier fjord system, Tempelfjorden, Svalbard. Continental Shelf Research, 181, 1-13, https://doi.org/10.1016/j.csr.2019.04.013 .
Eriksen, E., Benzik, AN., Dolgov, AV., Skjoldal, HR., Vihtakari, M., Johannesen, E., Prokhorova, TS., Keulder-Stenevik, F., Prokopchuk, I., Strand, E. 2020. Diet and trophic structure of fishes in the Barents Sea: the Norwegian-Russian program “Year of stomachs” 2015 - establishing a baseline. Progress in Oceanography. https://doi.org/10.1016/j.pocean.2019.102262
Eriksen, E, Bogstad, B, Nakken, O. 2011. Ecological significance of 0-group fish in the Barents Sea ecosystem. Polar Biology 34:647–657, doi:10.1007/s00300-010-0920-y.
Eriksen, E., Huserbråten, M., Gjøsæter, H., Vikebø, F., Albretsen, J. 2019. Polar cod egg and larval drift patterns in the Svalbard archipelago. Polar Biology https://doi.org/10.1007/s00300-019-02549-6
Eriksen, E., Ingvaldsen, RB., Nedreaas, K., Prozorkevich, D. 2015. The effect of recent warming on polar cod and beaked redfish juveniles in the Barents Sea. Regional Studies in Marine Science 2: 105–112
Eriksen, E, Ingvaldsen, R, Stiansen, JE, Johansen, OG. 2012. Thermal habitat for 0-group fishes in the Barents Sea; how climate variability impacts their density, length and geographical distribution. ICES J. Mar. Sci. 69(5): 870–879, doi:10.1093/icesjms/fsr210.
Eriksen, E., Skjoldal, HR., Dolgov, AV., Dalpadado, P., Orlova, EL., Prozorkevich, DV. 2016. The Barents Sea euphausiids: methodological aspects of monitoring and estimation of abundance and biomass. ICES Journal of Marine Science, 73:1533–1544.
Eriksen, E., Skjoldal, HR., Gjøsæter, H., Primicerio, R. 2017. Spatial and temporal changes in the Barents Sea pelagic compartment during the recent warming. Progress in Oceanography 151, 206-226. https://doi.org/10.1016/j.pocean.2016.12.009
Erikstad, KE., Benjaminsen, S., Reiertsen, TK, Ballesteros, M., Strøm, H. 2018. Modellering av svømmetrekket til lomvi (Uria aalge) fra Bjørnøya til norskekysten. Utvikling av ny metodikk for bruk av lysloggere (GLS), dykkeloggere (TDR) og sjøtemperaturer (SST) til kartlegging av svømmetrekket hos alkefugl. NINA rapport 1546, Norsk institutt for naturforskning.
Espinasse B., Basedow SL., Tverberg V., Hattermann T., Eiane K. 2016. A major Calanus finmarchicus overwintering population inside a deep fjord in northern Norway: implications for cod larvae recruitment success. Journal of Plankton Resource, 38:604-609.
Faglig forum for norske havområder. 2019. Særlig verdifulle og sårbare områder - Faggrunnlag for revisjon og oppdatering av forvaltningsplanene for norske havområder M-1303 2019, 304 s.
Falk-Petersen, S., Pavlov, V., Timofeev, S., Sargent, JR. 2007. Climate variability and possible effects on arctic food chains: The role of Calanus . I: Ørbæk J.B., Kallenborn R., Tombre I,., Hegseth, EN., Falk-Petersen, S., Hoel, AH. (Red) Arctic Alpine Ecosystems and People in a Changing Environment. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-540-48514-8_9
Falk-Pedersen, J., Renaud, P., Anisimova, N. 2011. Establishment and ecosystem effects of the alien invasive red kung crab (Paralithoodes camtschaticus) in the Barents Sea - a review. ICES Journal of Marine Science, 68 (2011), 479-488. https://doi.org/10.1093/icesjms/fsq192
Fall, J., Johannesen, E., Englund, G., Johansen, GO., Fiksen, Ø. 2021. Predator-prey overlap in three dimensions: cod benefit from capelin coming near the seafloor Ecography in press. Finland. Tellus A 66, 21564. http://dx.doi.org/10.3402/tellusa.v66.21564
Fall, J., Skern-Mauritzen, M. 2014. White-beaked dolphin distribution and association with prey in the Barents Sea, Marine Biology Research, 10:10, 957-971, DOI: 10.1080/17451000.2013.872796
Fauchald, P., Anker-Nilssen, T., Barrett, RT., Bustnes, JO., Bårdsen, B-J., Christensen-Dalsgaard, S., Descamps, S., Engen, S. Erikstad, KE., Hanssen, SA., Lorentsen, S-H., Moe, B., Reiertsen, TK., Strøm, H., Systad GH., 2015. The status and trends of seabirds breeding in Norway and Svalbard – NINA Report 1151. 84 s.
Fer, I, Drinkwater, KF, 2014. Mixing in the Barents Sea Polar Front near Hopen in spring. J Mar Sys. 130, 206–218.
Fernández-Méndez, M., Katlein, C., Rabe, B., Nicolaus, M., Peeken, I., Bakker, K., Flores, H., Boetius, A. 2015. Photosynthetic production in the central Arctic Ocean during the record sea-ice minimum in 2012. Biogeosciences 12, 3525-3549.
Fernández-Méndez, M., Olsen, LM., Kauko, HM., Meyer, A., Rösel, A., Merkouriadi, I., mfl. 2018. Algal hot spots in a changing Arctic Ocean: Sea-ice ridges and the snow-ice interface. Front. Mar. Sci. 5, 75. doi: 10.3389/fmars.2018.00075.
Flores, H., David, C., Ehrlich, J., Hardge, K., Kohlbach, D., Lange, BA. mfl. 2019. Sea-ice properties and nutrient concentration as drivers of the taxonomic and trophic structure of high-Arctic protist and metazoan communities. Polar Biol. 42, 1377-1395.
Folkow, LP., Blix, AS. 1999. Diving behaviour of hooded seals (Cystophora cristata) in the Greenland and Norwegian Seas. Polar Biology 22: 61-74.
Forsberg, H. 2018. Morphological description of egg capsules of skates (Rajidae and Arhynchobatidae) and potential nursery grounds in the Barents Sea. Bacheloroppgave UiT
Fort, J., Steen, H., Strom, H., Tremblay, Y., Grønningsæter, E. mfl. 2013. Energetic consequences of contrasting winter migratory strategies in a sympatric Arctic seabird duet. Journal of Avian Biology, Wiley, 44, pp. 255-262. doi:10.1111/j.1600-048X.2012.00128.x
Fossheim, M., Primicerio, R., Johannesen, E., Ingvaldsen, RB. Aschan, MM. Dolgov. AV. 2015. Recent warming leads to a rapid borealization of fish communities in the Arctic . Nat. Clim. Change, 5 (2015), s. 673-677.
Frainer, A., Primicerio, R., Kortsch, S., Aune, M., Dolgov, AV., Fossheim, M., Aschan, MM. 2017. Climate-driven changes in functional biogeography of Arctic marine fish communities. Proc. Natl. Acad. Sci. 114, 12202-12207.
Franks, PJS., 1992. Sink or swim – Accumulation of biomass at fronts. Mar Ecol Prog Ser. 82, 1-12.
Fransner, F., Fröb, F., Tjiputra, J., Chierici, M., Fransson, A., Jeansson, E., Johannessen, T., Jones, E., Lauvset, SK., Olafsdottir,KS., Omar, A., Skjelvan I., Olsen. A. 2020. Nordic Seas Acidification, sendt til Biogeosciences, 2020.
Fransson, A, Chierici, M, Anderson, LG, Bussman, I, Jones, EP, Swift, JH. 2001. The importance of shelf processes for the modification of chemical constituents in the waters of the eastern Arctic Ocean: implication for carbon fluxes. Cont Shelf Res, 21, 225-242.
Fransson, A., Chierici, M., Hop, H., Findlay, H., Kristiansen, S., Wold, W. 2016. Late winter- to-summer change in ocean acidification state in Kongsfjorden with implications for calcifying organisms. Polar Biology. 39: 1841-1857.
Fransson, A., Chierici, M., Nojiri, Y. 2009. New insights into the spatial variability of the surface water CO 2 in varying sea ice conditions in the Arctic Ocean. Cont. Shelf Res., 29, 1317-1328, doi:10.1016/j.csr.2009.03.008.
Fransson, A., Chierici, M., Nomura, D., Granskog, MA., Kristiansen, S., Martma, T., Nehrke, G. 2015. Effect of glacial drainage water on the CO2 system and ocean acidification state in an Arctic tidewater-glacier fjord during two contrasting years. Journal of Geophysical Research-Oceans, 120, doi:10.1002/2014JC010320.
Fransson A., Chierici, M., Nomura, D., Granskog, MA., Kristiansen, S., Martma, T., Nehrke, G. 2020. Influence of glacial water and carbonate minerals on wintertime sea-ice biogeochemistry and the CO 2 system in an Arctic fjord in Svalbard. Annals of Glaciology, https://doi.org/10.1017/aog.2020.52 .
Fransson A., Chierici, M., Skjelvan, I., Olsen, A., Assmy, P., Peterson, A., Spreen, G., Ward, B. 2017. Effect of sea-ice and biogeochemical processes and storms on under-ice water fCO 2 during the winter-spring transition in the high Arctic Ocean: implications for sea-air CO 2 fluxes. JGR- Oceans, N-ICE special issue, doi: 10.1002/2016JC012478.
Freitas, C., Kovacs, KM., Andersen, M., Aars, J., Sandven, S., Skern-Mauritzen, M., Pavlova, O., Lydersen, C. 2012. Importance of fast ice and glacier fronts for female polar bears and their cubs during spring in Svalbard, Norway. Mar Ecol Prog Ser 447: 289-304.
Freitas, C., Kovacs, KM., Ims, RA., Fedak, MA., Lydersen, C. 2008a. Ringed seal post-moulting movement tactics and habitat selection. Oecologia 155: 193-204.
Freitas, C., Kovacs, KM., Ims, RA., Fedak., MA., Lydersen, C. 2009. Deep into the ice: over-wintering and habitat selection in Atlantic walruses. Mar Ecol Prog Ser 375: 247-261.
Førland, EJ., Benestad, R., Hanssen-Bauer, I., Haugen, JE. og Skaugen, TE. 2011. Temperature and Precipitation Development at Svalbard 1900–2100, Advances in Meteorology, 2011, 893790, doi:10.1155/2011/893790, 2011.
Gammelsrød T, Leikvin O, Lien V, Budgell P, Loeng H, Maslowski W, 2009. Mass and heat transport in the NE Barents Sea: Observations and models. J. Mar. Syst. 75, 56-69.
Garcia, EG. 2007. The northern shrimp (Pandalus borealis) offshore fishery in the Northeast Atlantic. Advances in Marine Biology 52, 147-266.
Geoffrey, M., Daae, M., Cusa, M., Darnis, G., Graeve, M., Hernández, NS., Berge, J., Renaud, PE., Cottier, F., Falk-Petersen, S. 2019. Frontiers in Marine Science 2019, https://doi.org/10.3389/fmars.2019.00364
Gerland, S., Hall, R. 2006. Variability of fast-ice thickness in Spitsbergen fjords. Annals of Glaciology 44, 231-239. doi:10.3189/172756406781811367.
Gerland, S., Gascard, JC., Ivanov, B., Nielsen, CP., Nilsen, F., Pavlova, O., Leu, E., Tverberg, V., Barrault, S, 2008. Fast ice evolution in Kongsfjorden compared with other Svalbard fjords. Extended abstract. Proceedings of the NySMAC Seminar: Ny-Ålesund and IPY, Cambridge, U.K., 16-17 October 2007. Polarnet, NRC, Italy, pp 44-46
Gilg, O., Strøm, H., Aebischer, A., Gavrilo, MV., Volkov, AE., Miljeteig, C., Sabard, B. 2010. Post-breeding movements of northeast Atlantic ivory gull Pagophila eburnea populations. J. Avian Biol. doi: 10.1111/j.1600-048X.2010.05125.x
Gilg, O., Kovacs, KM., Aars, J., Fort, J., Gauthier, G., Gramillet, D., Ims, RA., Meltofte, H., Moreau, J., Post, E., Schmidt, NM., Yannic, G., Bollache, L. 2012. Climate change and the ecology and evolution of Arctic vertebrates. Ann NY Acad Sci 1249: 166-190.
Gjertz, I. 1991. The narwhal, Monodon monoceros , in the Norwegian High Arctic. Mar Mamm Sci 7: 402-408.
Gjertz, I., Kovacs, KM., Lydersen, C., Wiig, Ø. 2000. Movements and diving of bearded seal (Erignathusbarbatus) mother and pups during lactation and post-weaning. Polar Biol 23: 559-566.
Gjevik B, Nost E, Straume T, 1994. Model simulations of the tides in the Barents Sea. J Geophys Res, 99 (C2): 3337-3350.
Gjøsæter, H. 1998. The population biology and exploitation of capelin (Mallotus villosus) in the Barents Sea. Sarsia 83: 453-496.
Gjøsæter, H., Bogstad, B., Tjelmeland, S. 2009. Ecosystem effects of the three capelin stock collapses in the Barents Sea. Marine Biology Research 5: 40 - 53.
Gjøsæter, H., Hallfredsson, EH., Mikkelsen, N., Bogstad, B., Pedersen, T., 2016. Predation on early life stages is decisive for year-class strength in the Barents Sea capelin (Mallotus villosus) stock. ICES J. Mar. Sci. 73, 182-195.
Gjøsæter, H., Huserbråten, M., Vikebø, F., Eriksen, E. 2020. Key processes regulating the early life history of Barents Sea polar cod. Polar Biol 43, 1015–1027 (2020). https://doi.org/10.1007/s00300-020-02656-9
Gjøsæter, H., Ushakov, NG., Prozorkevich, DV. 2011. Capelin. I: Jakobsen, T., Ozhigin, V. (Red), The Barents Sea ecosystem: Russian-Norwegian cooperation in science and management. Tapir Academic Press, Trondheim, Norway, s. 201-214.
Gluchowska, M., Dalpadado, P., Beszczynska-Möller, A., Olszewska, A., Ingvaldsen, R. B., Kwasniewski, S. 2017. Interannual zooplankton variability in the main pathways of the Atlantic water flow into the Arctic Ocean (Fram Strait and Barents Sea branches). ICES J. Mar. Sci. 74, 1921–1936. doi: 10.1093/icesjms/fsx033
González-Pola, C., Larsen, KMH., Fratantoni, P., Beszczynska-Möller, A. (Red.). 2019. ICES Report on Ocean Climate 2018. ICES Cooperative Research Report No. 349. 122 s.
Gosselin, M., Levasseur, M., Wheeler, P.A., Horner, R.A., Booth, B.C., 1997. New measurements of phytoplankton and ice algal production in the Arctic Ocean. Deep-Sea Research II 44, 1623–1644.
Gradinger R. 2020. Life in Arctic Sea Ice. In: Goldstein M.I., DellaSala, D.A. (Red.), Encyclopedia of the World's Biomes, vol. 2. Elsevier, s. 507–514.
Gradinger, RR., Bluhm, BA. 2004. In situ observations on the distribution and behavior of amphipods and Arctic cod (Boreogadus saida) under the sea ice of the high Arctic Canadian Basin. Polar Biology 27, 595-603.
Gradinger, R., Bluhm, BA. 2020. First Arctic sea ice meiofauna food web analysis based on abundance, biomass and stable isotope ratios of sea ice metazoan fauna from near-shore Arctic fast ice. Marine Ecology Progress Series 634:29-43 https://doi.org/10.3354/meps13170
Gradinger, R., Bluhm, B., Iken, K. 2010. Arctic sea-ice ridges-Safe havens for sea-ice fauna during periods of extreme ice melt? Deep Sea Research Part II: Topical Studies in Oceanography, 57 (1-2), 86-95.
Gradinger, RR., Kaufman, MM., Bluhm, BA. 2009. Pivotal role of sea ice sediments in the seasonal development of near-shore Arctic fast ice biota. Marine Ecology 394, 49–63.
Grebmeier, JM., Overland, JE., Moore, SE., Farley, EV., Carmack, EC., Cooper, LW., Frey, KE., Helle, JH., McLaughlin, FA., McNutt, L. 2006. A major ecosystem shift in the northern Bering Sea. Science 311: 1461-1464.
Grémillet, D., Fort, J., Amélineau, F., Zakharova, E., le Bot, T., Sala, T., Gavrilo, M. (2015). Arctic warming: nonlinear impacts of sea-ice and glacier melt on seabird foraging. Global Change Biology 21, 1116-1123. https://doi.org/10.1111/gcb.12811
Gulliksen, B. 1984. Under-ice fauna from Svalbard waters. Sarsia 69(1), 17-23.
Halbach, L., Vihtakari, M., Duarte, P., Everett, A., Granskog, M., Hop, H., Kauko, HM., Kristiansen, S., Myhre, P. I., Pavlov, A. K., Pramanik, A., Tatarek, A., Torsvik, T., Wiktor, JM., Wold, A., Wulff, A., Steen, H., Assmy, P. 2019. Tidewater Glaciers and Bedrock Characteristics Control the Phytoplankton Growth Environment in a Fjord in the Arctic, Front. Mar. Sci., 6, 254, https://doi.org/10.3389/fmars.2019.00254.
Hallfredsson, EH., Pedersen, T. 2009. Effects of predation from juvenile herring (Clupea harengus) on mortality rates of capelin (Mallotus villosus) larvae. Canadian Journal of Fisheries and Aquatic Sciences 66, 1693-1706.
Halvorsen, E., Tande, KS., Edvardsen, A., Slagstad, D., Pedersen, OP. 2003. Habitat selection of overwintering Calanus finmarchicus in the NE Norwegian Sea and shelf waters off Northern Norway in 2000–02. Fisheries Oceanography 12:4/5, 339-35.
Hamilton, CD., Kovacs, KM., Ims, RA., Aars, J., Lydersen, C. 2017. An Arctic predator-prey system in flux: climate change impacts on coastal space use by polar bears and ringed seals. J Anim Ecol 86: 1054-64.
Hamilton, CD., Kovacs, KM., Ims, RA., Lydersen, C. 2018. Haul-out behaviour of Arctic ringed seals: Inter-annual patterns and impacts of current environmental change. Polar Biol. 41: 1063-1082.
Hamilton CD., Kovacs, KM., Lydersen, C. 2015a. Year-round haul-out behaviour of walruses (Odobenus rosmarus) in the Northern Barents Sea. Mar Ecol Prog Ser 519: 251-263.
Hamilton, CD., Lydersen, C., Ims, RA, Kovacs, KM. 2015b. Predictions replaced by facts in a changing Arctic: a keystone species’ responses to declining sea ice. Biol Lett 11: 20150803. http://dx.doi.org/10.1098/rsbl.2015.0803
Hamilton, CD., Vacquié-Garcia, J., Kovacs, KM., Ims, RA., Lydersen, C. 2019a. Contrasting changes in space use induced by climate change in two Arctic marine mammal species. Biol Lett 15: 20180834. doi:10.1098/rsbl.2018.0834.
Hamilton, CD., Kovacs, KM., Lydersen, C. 2019b. Sympatric seals use different habitats in an Arctic glacial fjord. Mar Ecol Prog Ser 615: 205-220.
Hamilton CD., Lydersen C., Aars J., Biuw M. mfl. 2021. Marine mammal hotspots in the Greenland and Barents Seas. Mar Ecol Prog Ser 659:3-28. https://doi.org/10.3354/meps13584
Hamilton, CD., Lydersen, C., Fedak, MA., Freitas, C., Hindell, MA., Kovacs, KM. 2019c. Behavioural ontogeny of bearded seals Erignathus barbatus through the first year of life. Mar Ecol Prog Ser 627: 179-194.
Hamilton, CD., Lydersen, C., Ims, RA., Kovacs, KM. 2014. Haul-out behaviour of the world's northernmost population of harbour seals (Phoca vitulina) throughout the year. PLoS One 9: e86055.
Hammill, MO., Kovacs, KM., Lydersen, C. 1994. Local movements by nursing bearded seal (Erignathus barbatus) pups. Polar Biol 14: 569-570.
Hansen, A. 2020. Genetic population structure of the northern shrimp (Pandalus borealis) along the Norwegian coast. Masteroppgave. Norges arktiske universitet. Fakultet for biovitenskap, fiskeri og økonomi. 60 s.
Hansen, AS., Nielsen, TG., Levinsen, H., Madsen, S.D., Thingstad, TF. Hansen, BW. 2003. Impact of changing ice cover on pelagic productivity and food web structure in Disko Bay, West Greenland: A dynamic model approach. Deep Sea Research I 50: 171-87.
Hansen, E., Ekeberg, O-C., Gerland, S., Pavlova, O., Spreen, G. mfl. 2014. Variability in categories of Arctic sea ice in Fram Strait. Journal of Geophysical Research: Oceans, 119, 7175–7189. https://doi.org/10.1002/2014JC010048 .
Hansen, E., Gerland, S., Granskog, MA., Pavlova, O., Renner, AHH., Haapala, J. mfl. 2013. Thinning of Arctic sea ice observed in Fram Strait: 1990-2011. Journal of Geophysical Research: Oceans, 118, 5202–5221. https://doi.org/10.1002/jgrc.20393 .
Hansen, E., Gerland, S., Høyland, KV., Pavlova, O. mfl. 2015. Time variability in the annual cycle of sea ice thickness in the transpolar, drift. Journal of Geophysical Research: Oceans, 120, 8135–8150. https://doi.org/10.1002/2015JC011102 .
Hansen, HØ., Aschan, M. 2000. Growth, size- and age-at-maturity of shrimp, Pandalus borealis , at Svalbard related to environmental parameters. J ournal of Northwest Atlantic Fishery Science 27, 83-91.
Hanssen-Bauer, I., Førland, EJ., Hisdal, H., Mayer, S., Sandø, AB., Sorteberg A. 2018. Climate in Svalbard 2100– a knowledge base for climate adaptation. M-1242 | 2018. NCCS report no. 1/2019.
Hasle, GR., Heimdal, BR. 1998. The net phytoplankton in Kongsfjorden, Svalbard, July 1988, with general remarks on species composition of Arctic phytoplankton. Polar Res. 17: 31–52.
Hasle, GR., Syvertsen, EE., von Quillfeldt, CH. 1996. Fossula arctica gen. nov., spec. nov., a marine Arctic araphid diatom. Diatom Research 11(2): 261-272.Hassel, A., Skjoldal, HR., Gjøsæter, H., Loeng, H., Omli, L. 1991. Impact of grazing from capelin (Mallotus villosus) on zooplankton: A case study in the northern Barents Sea in august 1985. Polar Res . 10: 371– 388. doi: 10.1111/j.1751-8369.1991.tb00660.x
Hassett, BT., Gradinger R. 2016. Chytrids dominate arctic marine fungal communities. Environmental microbiology, 18(6), s. 2001-2009.
Hatam, I., Lange, B., Beckers, J., Haas, C, Lanoil, B. 2016. Bacterial communities from Arctic seasonal sea ice are more compositionally variable than those from multi-year sea ice. The ISME Journal 10:2543-2552.
Haug, T., Bogstad. B., Chierici, M., Gjøsæter, H., Hallfredsson EH., Høines ÅS., Hoel, AH., Ingvaldsen, RB., Jørgensen LL., Knutsen, T., Loeng, H., Naustvoll, L-J., Røttingen, I., Sunnanå, K. 2017. Future harvest of living resources in the Arctic Ocean north of the Nordic and Barents Seas: A review of possibilities and constraints. Fisheries Research 188 (2017) 38–57.
Hegseth, EN. 1998: Primary production in the northern Barents Sea. Polar Research 17(2), 113-123.
Hegseth, EN., Assmy, P., Wiktor, JM., Wiktor, JJ., Kristiansen, S., Leu, E. mfl. 2019. Phytoplankton Seasonal Dynamics in Kongsfjorden, Svalbard and the Adjacent Shelf. I: Hop, H., Wiencke, C. (Red) The Ecosystem of Kongsfjorden, Svalbard. Advances in Polar Ecology, 2. Springer, Cham. https://doi.org/10.1007/978-3-319-46425-1_6
Hegseth E., Svendsen H., Quillfeldt C. 1995. Phytoplankton in fjords and coastal waters of northern Norway: environmental conditions and dynamics of the spring bloom. I: Skjoldal, HR., Hopkins, C., Erikstad, KE., Leinaas, HP. (Red.) Ecology of Fjords and Coastal Waters.
Heimdal, BR. 1989. Arctic Ocean Phytoplankton. I Herman Y. (Red.), The Arctic Seas. Climatology, 860 Oceanography, Geology, and Biology, Van Nostrand Reinhold Company, New-York, 193-222.
Helland-Hansen, B, Nansen, F, 1909. The Norwegian Sea: Its Physical Oceanography Based Upon the Norwegian Researches 1900-1904.
Heide-Jørgensen, MP., Blackwell, SB., Williams, TM., Sinding, MHS., Skovrind, M., Tervo, OM., Garde, E., Hansen, RG., Nielsen, NHL., Ngô, MN., Ditlevsen, S. 2020. Some like it cold: Temperature-dependent habitat selection by narwhals. Ecol Evol 00:1–18. doi: 10.1002/ece3.6464.
Hernández, NS. 2019. A Patch of Calanus finmarchicus in the Lofoten-Vesterålen Region. Characteristics and determining factors. BIO-3950 Master thesis in Biology. Faculty of Biosciences, Fisheries and Economics, University of Tromsø.
Hjelset, AK. (red) 2014. Report from the workshop: Workshop on king- and snow crabs in the Barents Sea. Rapport fra Havforskningen nr. 18-2014. 83 s.
Hobbs, RC., Reeves, RR., Prewitt, JS., Desportes, G., Breton-Honeyman, K., Christensen, T., Citta, JJ., Ferguson, SH., Frost, KJ., Garde, E. mfl. 2020. Global review of the conservation status of monodontid stocks. Mar Fish Rev 81 (3-4): 1-53.
Hodal, H., Falk-Petersen, S., Hop, H. mfl. 2012. Spring bloom dynamics in Kongsfjorden, Svalbard: nutrients, phytoplankton, protozoans and primary production. Polar Biol 35: 191–203. https://doi.org/10.1007/s00300-011-1053-7
Hollowed, AB., Barange, M., Beamish, RJ., Brander, K., Cochrane, K., Drinkwater, K., Foreman, MGG., Hare, JA., Holt, J., Ito, S-I., Kim, S., King, JR., Loeng, H. MacKenzie, BR., Mueter, FJ., Okey, TA., Peck, MA., Radchenko, VI., Rice, JC. Schirripa, MJ., Yatsu, A., Yamanaka, Y. 2013a. Projected impacts of climate change on marine fish and fisheries. ICES Journal of Marine Science; doi:10.1093/icesjms/fst081
Hollowed, AB., Planque, B., Loeng, H. 2013b. Potential movement of fish and shellfish stocks from the sub-Arctic to the Arctic Ocean. Fish. Oceanogr. 22:5, 355-370.
Hop, H., Wold, A., Vihtakari, M., Daase, M., Kwasniewski, S., Gluchowska, M. mfl. 2019a. “Zooplankton in Kongsfjorden (1996-2016) in relation to climate change,” in The Ecosystem of Kongsfjorden, Svalbard, eds H. Hop and C. Wiencke (Cambridge: Springer), 229–302. doi: 10.1007/978-3-319-46425-1_7
Hop, H., Assmy, P., Wold, A., Sundfjord, A., Daase, M., Duarte, P., Kwasniewski, S., Gluchowska, M., Wiktor, J. M., Tatarek, A., Wiktor, J., Kristiansen, S., Fransson, A., Chierici, M., Vihtakari, M. 2019b. Pelagic Ecosystem Characteristics Across the Atlantic Water Boundary Current From Rijpfjorden, Svalbard, to the Arctic Ocean During Summer (2010–2014). Front. Mar. Sci., 6, 181, https://doi.org/10.3389/fmars.2019.00181 .
Hop, H., Bluhm, BA., Daase, M., Gradinger, R., Poulin, M. 2013. Arctic sea ice biota. Arctic Report Card: Update for 2013
Hop, H., Gjosaeter, H. 2013. Polar cod (Boreogadus saida) and capelin (Mallotus villosus) as key species in marine food webs of the Arctic and the Barents Sea. Marine Biology Research 9 (9): 878-894.
Hop, H., Pearson, T., Hegseth, EN., Kovacs, KM., Wiencke, C., Kwasniewski, S., Eiane, K., Mehlum, F., Gulliksen, B., Wlodarska-Kowalczuk, M., Lydersen, C., Weslawski, JM., Cochrane, S., Gabrielsen, GW., Leakey, RJG., Lønne, OJ., Zajaczkowski , M ., Falk-Petersen, S., Kendall, M., Wängberg, S-Å., Bischof, K., Voronkov, AY., Kovaltchouk, NA., Wiktor, JM., Poltermann, M., di Prosco, G., Papucci, C., Gerland, S. 2002. The marine ecosystem of Kongsfjorden, Svalbard. Polar Res. 21(1):167-208.
Hop, H., Poltermann, M., Lønne OJ., Falk-Petersen, S., Korsnes, R., Budgell, WP. 2000. Ice amphipod distribution relative to ice density and under-ice topography in the northern Barents Sea. Polar Biology, 23(5), s. 357-367.
Hop, H., Vihtakari, M., Bluhm, B.A., Assmy, P., Poulin, M., Gradinger, R., Peeken, I., von Quillfeldt, CH., Olsen, LM., Zhitina, L., Melnikov, IA. 2020. Changes in sea-ice protist diversity with declining sea ice in the Arctic Ocean from the 1980s to 2010s. Frontiers Marine Science, doi: 10.3389/mars2020.00243.
Hopkins, CCE., Nilssen, EM. 1990. Population biology of the deep-water prawn (Pandalus borealis) in Balsfjord, northern Norway: I. Abundance, mortality, and growth, 1979-1983. Journal du Conseil International pour l’Exploration de la Mer 47, 148-166.
Hopwood, M. J., Carroll, D., Dunse, T., Hodson, A., Holding, J. M., Iriarte, J. L., Ribeiro, S., Achterberg, E. P., Cantoni, C., Carlson, D. F., Chierici, M., Clarke, J. S., Cozzi, S., Fransson, A., Juul-Pedersen, T., Winding, M. S., Meire, L. 2020. Review Article: How does glacier discharge affect marine biogeochemistry and primary production in the Arctic? The Cryosphere, 14, 1347–1383, https://doi.org/10.5194/tc-14-1347-2020 .
Horner, R., Ackley, SF., Dieckmann, GS., Gulliksen, B., Hoshiai, T., Melnikov, I.A., Reeburgh, W.S., Spindler, M., Sullivan C.W. 1992. Ecology of Sea Ice Biota. 1. Habitat and terminology. Polar Biology 12, 417–427.
Hovde, S.C., Albert, OT. og Nilssen, EM. 2002. Spatial, seasonal and ontogenetic variation in diet of Northeast Arctic Greenland halibut (Reinhardtius hippoglossoides). ICES Journal of Marine Science, 59: 421–437.
Hunt Jr., GL., Bakken, V., Mehlum, F., 1996. Marine birds in the marginal ice zone of theBarents Sea in late winter and spring. Arctic 49. 53–61
Hunt, GL., Blanchard, AL., Boveng, P., Dalpadado, P., Drinkwater, KF., Eisner, L., Hopcroft, RR., Kovacs, KM., Norcross, BL., Renaud, P. mfl. 2013. The Barents and Chukchi Seas: Comparison of two Arctic shelf ecosystems. J. Mar. Syst. 109–110, 43–68. https://doi.org/10.1016/j.jmarsys.2012.08.003
Husa, V., Gonzalez-Mirelis, G., Kutti, T., Mortensen, P., Søvik G., Jensen, G., Bakkeplass K., Skjoldal H.R. 2020. Marinbiologisk mangfold i Andfjorden marine verneområde. Rapport fra Havforskningen 2020-30 ISSN:1893-4536, Publisert: 28.09.2020.
Huse, G., Fernö, A., Holst, JC. 2010. Establishment of new wintering areas in herring co-occurs with peaks in the ‘first time/repeat spawner’ ratio. Marine Ecology Progress Series 409, 189-198.
Huse, G., Toresen, R. 2000. Juvenile herring prey on Barents Sea capelin larvae. Sarsia 85, 385-391.
Huseby S. 2002. The phytoplankton spring bloom dynamics in the Lofoten area, Northern Norway. Degree of Cand. Scient. Thesis. UiT.
Hylen, A, Nakken, O, Nedreaas, K. 2008. Northeast Arctic cod: fisheries, life history, fluctuations and management. I: Norwegian Spring-Spawning Herring and Northeast Arctic Cod-100 Years, s. 41–82. O. Nakken (Red). Tapir Academic Press, Trondheim, Norway.
ICES 2005. Spawning and life history information for North Atlantic cod stocks. Prepared by the ICES/GLOBEC Working Group on Cod and Climate Change. Editor Keith Brander. ICES Cooperative Research Report No. 274, June 2005. 152s.
ICES 2017. Report of the Working Group on the Integrated Assessments of the Barents Sea (WGIBAR). ICES CM 2017/SSGIEA:04, Murmansk, Russia.
ICES 2018. Interim Report of the Working Group on the Integrated Assessments of the Barents Sea (WGIBAR). WGIBAR 2018 REPORT 9-12 March 2018. Tromsø, Norway. ICES CM 2018/IEASG:04. 210 s.
ICES 2019. The Working Group on the Integrated Assessments of the Barents Sea (WGIBAR). ICES Scientific Reports. 1:42. 157 s. http://doi.org/10.17895/ices.pub.5536
ICES 2020 a. Report of the Working Group on Integrated Assessments of the Barents Sea (WGIBAR).
Ingvaldsen, RB., Asplin, L., and Loeng, H. 2004. Velocity field of the western entrance to the Barents Sea. Journal of Geophysical Research, 109, C03021 , doi:10.1029/2003JC001811.
Ingvaldsen, RB., Gjøsæter, H., 2013. Impact of marine climate variability on the spatial distribution of Barents Sea capelin. Marine Biology Research, 9(9):867-877.
Iversen, M., Aars, J., Haug, T., Alsos, IG., Lydersen, C., Bachmann, L., Kovacs, KM. 2013. The diet of polar bears (Ursus maritimus) from Svalbard, Norway, inferred from scat analysis. Polar Biol 36: 561-71.
Järnegren, J., Kutti, T. 2014. Lophelia pertusa in Norwegian waters. What have we learned since 2008? - NINA Report 1028. 40 pp. ISBN: 978-82-426-2640-0.
Jakobsen, T., Ozhigin, V (Red) 2011. The Barents Sea – ecosystem, resources, management. Half a century of Russian-Norwegian cooperation. Tapir Academic Press, Trondheim, Norway, 825 s.
Johannesen, E., Lindstrøm, U., Michalsen, K., Skern - Mauritzen, M., Fauchald, P., Bogstad, B., Dolgov, AV.. 2012. Feeding in a heterogeneous environment: spatial dynamics in summer foraging Barents Sea cod. Marine Ecology Progress Series 458:181-197.
Johannesen, E., Mørk, HL., Korsbrekke, K., Wienerroither, R., Eriksen, E., Fossheim, M., d. L. Wenneck, T., Dolgov, A., Prokhorova T., Prozorkevich, D. 2017. Arctic fishes in the Barents Sea 2004-2015: Changes in abundance and distribution.
Johannesen, E., Wienerroither, R., Mørk, HL., Husson, B., Holmin, AJ., Johnsen, E., Dolgov, A., Prokhorova, T. 2021. Fish diversity data from the Barents Sea Ecosystem Survey 2004-2019. Rapport fra Havforskningen 2021-15. 45 s. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-en-2021-15
Johnsen, G., Norli M., Moline M., Robbins I, von Quillfeldt C., Sørensen K., Cottier F. Berge J. 2018. The advective origin of an under-ice spring bloom in the Arctic Ocean using multiple observational platforms. Polar Biology. https://doi.org/10.1007/s00300-018-2278-5
Johnsen, G., Volent, Z., Sakshaug, E., Sigernes, F., Pettersson, LH. 2009. Remote sensing in the Barents Sea. In “Ecosystem Barents Sea” Sakshaug, E., Johnsen, G., Kovacs. KM. (Red.). Tapir Academic Press.
Jones, E., Chierici, M., Skjelvan, I., Norli, M., Børsheim, KY., Lødemel, HH., Kutti, T., Sørensen, K., King, AL., Lauvset, S., Jackson, K., de Lange, T., Johannessen, T., Mourgues, C. 2019. Monitoring ocean acidification in Norwegian seas in 2018, Rapport, Miljødirektoratet, M-1417|2019.
Jones, E., Chierici, M., Skjelvan, I., Norli, M., Frigstad, H., Børsheim, K., Lødemel, H., Kutti, T., King, A., Sørensen, K., Lauvset, S., Jackson-Misje, K., Apelthun, L., de Lange, T., Johannsessen, T., Mourgues, C., Bellerby, R.2020. Monitoring ocean acidification in Norwegian seas in 2019, Rapport, Miljødirektoratet, M-1735, Tech. rep., 2020.
Jorde, PE., Søvik, G., Westgaard, J-I., Albretsen, J., André, C., Hvingel, C., Johansen, T., Sandvik, AD., Kingsley, M., Jørstad, KE. 2015. Genetically distinct populations of northern shrimp, Pandalus borealis , in the North Atlantic: adaptation to different temperatures as an isolation factor. Molecular Ecology 24, 1742-1757.
Jourdain, E., Vongraven, D. 2017. Humpback whale (Megaptera novaeangliae) and killer whale (Orcinus orca) feeding aggregations for foraging on herring (Clupea harengus) in Northern Norway. Mammalian Biology 86: 27-32.
Jørgensen, LL., Bakke, G., Hoel, AH. 2020. Responding to global warming: New fisheries management measures in the Arctic. Progress in Oceanography, 188, p.102423.
Jørgensen, LL, Ljubin, P, Skjoldal, HR, Ingvaldsen, RB, Anisimova, N, Manushin, I. 2015a. Distribution of benthic megafauna in the Barents Sea: baseline for an ecosystem approach to management. ICES Journal of Marine Science; 72 (2): 595-613
Jørgensen, LL., Pecuchet, L, Ingvaldsen, R, Primicerio, R (under forberedelse). Benthos in the Atlantic gateway to a changing Arctic Ocean. (to be submitted to Progress in Oceanography)
Jørgensen, LL, Planque, B, Thangstad TH, Certain G 2015b. Vulnerability of megabenthic species to trawling in the Barents Sea. ICES Journal of Marine Science. DOI: 10.1093/icesjms/fsv107.
Jørgensen, LL., Primicerio, R., Ingvaldsen, RB., Fossheim, M., Strelkova, N., Thangstad, TH., mfl. 2019. Impact of multiple stressors on sea bed fauna in a warming Arctic. Marine Ecology Progress Series, 608, 1-12.
Karnovsky, NJ., Kwazniewski, S., Weslawski, JM.., Walkusz, W, Beszczynska-Möller, A. 2003. Foraging behavior of little auks in a heterogeneous environment, Mar. Ecol. Prog. Ser., 253, 289– 303.
Kędra, M., Renaud, PE., Andrade, H., Goszczko, I. and Ambrose, WG. 2013. Benthic community structure, diversity, and productivity in the shallow Barents Sea bank (Svalbard Bank). Marine Biology, 160(4), 805-819.
King, J., Skourup, H., Hvidegaard, SM., Rösel, A., Gerland, S., Spreen, G., mfl. 2018. Comparison of freeboard retrieval and ice thickness calculation from ALS, ASIRAS, and CryoSat‐2 in the Norwegian Arctic to field measurements made during the N‐ICE2015 expedition. Journal of Geophysical Research: Oceans, 123(2), 1123-1141.
King J, Spreen G, Gerland S, Haas C, Hendricks S, Kaleschke L, og Wang C, 2017. Sea‐ice thickness from field measurements in the northwestern Barents Sea. Journal of Geophysical Research: Oceans, 122(2), 1497-1512.
Kjesbu, OS., Bogstad, B., Devine, JA., Gjøsæter, H., Ingvaldsen, R., Nash, RDM. Stiansen, JE. 2014. Synergies between climate and management for Atlantic cod fisheries at high latitudes. Proc Natl Acad Sci U.S.A. 111, 3478–3483. doi: 10.1073/pnas.1316342111
Klepikofsky, NN., Øien, N. 2017. Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea and adjacent waters, August-October 2016. IMR/PINRO Joint Report Series, 2-2017.
Knutsen, T., Wiebe, P.H., Ingvaldsen, R.B. Lien G. 2017. High latitude epipelagic and mesopelagic scattering layers- A reference for future Arctic Ecosystem change. Front. Mar: Sci. 4:1-21
Kohlbach, D.., Ferguson, SH.., Brown, TA., Michel, C. 2019. Landfast sea ice-benthic coupling during spring and potential impacts of system changes on food web dynamics in Eclipse Sound, Canadian Arctic. Marine Ecology Progress Series 627, 33-48.
Kohlbach, D., Graeve, M., Lange, BA., David, C., Peeken, I., Flores, H. 2016. The importance of ice algae-produced carbon in the central Arctic Ocean ecosystem: food web relationships revealed by lipid and stable isotope analyses. Limiology and Oceanography 61, 2027-2044.
Kohlbach, D., Hop, H., Wold, A., Schmidt, K., Smik, L., Belt, ST., Keck Al-Habahbeh, A., Woll, M., Graeve, M., Dabrowska, AM., Tatarek, A, Atkinson, A., Assmy, P. 2021. Multiple Trophic Markers Trace Dietary Carbon Sources in Barents Sea Zooplankton During Late Summer. Front. Mar. Sci. 7:610248. doi: 10.3389/fmars.2020.610248
Kohlbach, D., Schaafsma, F.L., Graeve, M., Lebreton, B., Lange, B.A., David, C., Vortkamp, M., Flores, H. 2017. Strong linkage of polar cod (Boreogadus saida) to sea ice algae-produced carbon: evidence from stomach content, fatty acid and stable isotope analyses. Progress in Oceanography, 152, s. 62-74.
Kortsch, S., Primicerio, R., Beuchel, F., Renaud, P.E., Rodrigues, J., Lønne, O.J. and Gulliksen, B., 2012. Climate-driven regime shifts in Arctic marine
Kortsch, S., Primicerio, R., Fossheim, M., Dolgov, A., Aschan, M. 2015. Climate change alters the structure of arctic marine food webs due to poleward shifts of boreal generalists. Proceedings of the Royal Society of London B: Biological Sciences 282(1814): 2015.1546. Ecography 41: 1–14, 2018 doi: 10.1111/ecog.03443
Kortsch, S., Primicerio, R., Aschan, M., Lind, S., Dolgov, A.V. and Planque, B., 2019. Food‐web structure varies along environmental gradients in a high‐latitude marine ecosystem. Ecography , 42 (2), pp.295-308.
Kosobokova, K., Hopcroft, RR., Hirche, H-J. 2011. Patterns of zooplankton diversity through the depths of the Arctic’s central basins. Marine Biodiversity 41(1):29-50
Kovacs, KM. 2018. Bearded seal (Erignathus barbatus). S. 83-86. I: Bernd Würsig, J.G.M. Thewissen og Kit M. Kovacs (Red). Encyclopedia of Marine Mammals, 3rd ed. Academic Press/Elsevier, San Diego, CA, USA.
Kovacs, KM., Aars, J., Lydersen, C. 2014. Walruses recovering after 60+ years of protection at Svalbard, Norway. Polar Res 33: 26034. doi.org/10.3402/polar.v.33.26034.
Kovacs, KM., Aguilar, A., Aurioles, D., Burkanov, V., Campagna, C., Gales, N., Gelatt, T., Goldsworthy, S., Goodman, SJ., Hofmeyr, GJG., Härkönen, T., Lowry, L., Lydersen, C., Schipper, J., Sipilä, T., Southwell, C., Stuart, S., Thompson, D., Trillmich, F. 2012. Global threats to pinnipeds. Mare Mamm Sci 28: 414-436.
Kovacs, KM., Krafft, B., Lydersen, C. 2020b. Bearded seal (Erignathus barbatus) pup growth - body size, behavioral plasticity and survival in a changing climate. Mar Mamm Sci 36: 276-284. doi.10.1111/mms.12647.
Kovacs, KM., Lydersen, C., Overland, JE., Moore, SE. 2011 Impacts of changing sea-ice conditions on Arctic marine mammals. Mar Biodiv 41: 181–194.
Kovacs. KM., Lydersen, C., Vacquiè-Garcia, J., Shpak, O., Glazov, D., Heide-Jørgensen, MP. 2020a. The endangered Spitsbergen bowhead whales’ secrets revealed after hundreds of years in hiding. Biol Lett 20200148. http://dx.doi.org/10.1098/rsbl.2020.0148 .
Krafft, BA., Kovacs, KM., Andersen, M., Aars, J., Lydersen, C., Ergon, T., Haug, T. 2006. Abundance of ringed seals (Pusa hispida) in the fjords of Spitsbergen, Svalbard, during the peak molting period. Mar Mamm Sci 22: 394-412.
Krysov, AI., Røttingen, I. 2011. Herring. s. 215-224. I: Jakobsen, T., Ozhigin, V. (Red) The Barents Sea – ecosystem, resources, management. Half a century of Russian-Norwegian cooperation. Tapir Academic Press, Trondheim, Norway, 825 s.
Kunisch, EH., Bluhm, BA., Daase, M., Gradinger, R., Hop, H., Melnikov IA., Varpe, Ø., Berge, J. 2020. Pelagic occurrences of the ice amphipod Apherusa glacialis throughout the Arctic. Journal of plankton research, 42(1), s. 73-86.
Kvingedal, B. 2005. Sea-ice extent and variability in the Nordic Seas, 1967—2002. GMS , 158 , 39-49.
Kwasniewski, S., M. Głuchowska, W. Walkusz, NJ. Karnovsky, D. Jakubas, K. Wojczulanis-Jakubas, AMA. Harding, I. Goszczko, M. Cisek, A. Beszczyńska-Moeller, W. Walczowski, JM. Weslawski, W., Stempniewicz, L. 2012. Inter-annual changes in zooplankton on the West Spitsbergen Shelf in relation to hydrography, and their consequences for the diet of planktivorous seabirds. ICES J. Mar. Sci., 69, s. 890-901
Kwok, R., 2008. Outflow of Arctic Ocean sea ice into the Greenland and Barents Seas: 1979 2007. J Clim 22:2438-2457
Kwok, R. 2018. Arctic sea ice thickness, volume, and multiyear ice coverage: losses and coupled variability (1958–2018). Environmental Research Letters, 13(10), 105005.
Kwok, R., Maslowski, W., Laxon, SW., 2005. On large outflows of Arctic sea ice into the Barents Sea. Geophys Res Lett 32, L22503. http://dx.doi.org/10.1029/2005GL024485
Laidre, KL., Stern, H., Kovacs, KM., Lowry, L., mfl. 2015. Arctic marine mammal population status, sea ice habitat loss, and conservation recommendations for the 21 st century. Conserv Biol 29: 724-737 https://doi.org/10.1111/cobi.12474
Laidre, KL., Stirling, I., Lowry. L., Wiig, Ø., Heide-Jørgensen, MP., Ferguson, S. 2008. Quantifying the sensitivity of arctic marine mammals to climate-induced habitat change. Ecol Appl 18(2): S97-S125.
Lauvset, SK., Chierici, M., Counillon, F., Omar, A., Nondal, G., Johannessen, T., Olsen, A. 2013 Annual and seasonal fCO 2 and air–sea CO 2 fluxes in the Barents Sea. Journal of Marine Systems, 113–114, Pages 62-74, https://doi.org/10.1016/j.jmarsys.2012.12.011
Lauvset, SK., Key, RM., Olsen, A., van Heuven, S., Velo, A., Lin, X., Schirnick, C., Kozyr, A., Tanhua, T., Hoppema, M., Jutterström, S., Steinfeldt, R., Jeansson, E., Ishii, M., Perez, F. F., Suzuki, T. og Watelet, S. 2016.: A new global interior ocean mapped climatology: the 1° × 1° GLODAP version 2, Earth Syst. Sci. Data, 8, 325–340, https://doi.org/10.5194/essd-8-325-2016 .
Lefort, KJ., Garroway, CJ., Ferguson, SH. 2020. Killer whale abundance and predicted narwhal consumption in the Canadian Arctic. Global Change Biol 26: 4276-4283. doDoi.10.1111/gcb.15152.
Le Fèvre J, 1986. Aspects of the biology of frontal systems. Adv Mar Biol. 23, 163–299.
Leonard, DM., Øien, NI. 2020. Estimated Abundances of Cetacean Species in the Northeast Atlantic from Norwegian Shipboard Surveys Conducted in 2014 – 2018. NAMMCO Scientific Publications 11 https://doi.org/ 10.7557/3.4694
Leu, E., Brown, TA., Graeve, M., Wiktor, J., Hoppe, CJM., Chierici, M., Fransson, A., Verbiest, S., Kvernvik, AC., Grennacre, J. 2020. Spatial and temporal variability of ice algal trophic markers – with recommendations about their application. Journal of Marine Science and Engineering 8, 676, doi:10.3390/jmse8090676.
Leu, E., Mundy, CJ., Assmy, P., Campbell, K., Gabrielsen, TM., Gosselin, M., Juul-Pedersen, T., Gradinger, R. 2015. Arctic spring awakening - Steering principles behind the phenology of vernal ice algal blooms. Progress in Oceanography 139, 151-170.
Leu, E., Søreide, JE., Hessen, DO., Falk-Petersen, S., Berge, J. 2011. Consequences of changing sea-ice cover for primary and secondary producers in the European Arctic shelf seas: Timing, quantity, and quality. Progress in Oceanography 90(1-4), 18-32.
Leu, E., Wiktor, J., Søreide, JE., Berge, J., Falk-Petersen, S. 2010. Increased irradiance reduces food quality of sea ice algae. Mar. Ecol. Prog. Ser. 4011, 49-60.
Lewis, KM., van Dijken, GL., Arrigo, KR., 2020. Changes in phytoplankton concentration now drive increased Arctic Ocean primary production. Science 369, 198–202.
Li, WKW., McLaughlin, FA., Lovejoy, C., Carmack, EC. 2009. Smallest algae thrive as the Arctic Ocean freshens. Science 326, 539-539.
Lien VS (red.), 2018. Polarfrontens fysiske beskaffenhet og biologiske implikasjoner – en verdi- og sårbarhetsvurdering av polarfronten i Barentshavet. Fisken og Havet, nr. 8-2018. Havforskningsinstituttet. ISSN 0071-5638.
Lind, S., Ingvaldsen, R.B., 2012. Variability and impacts of Atlantic Water entering the Barents Sea from the north. Deep-Sea Research , I 62 (2012), 70-88. doi:10.1016/j.dsr.2011.12.007.
Lind, S., Ingvaldsen, RB., Furevik, T. 2018. Arctic warming hotspot in the northern Barents Sea linked to declining sea-ice import. Nature climate change , 8 (7), 634-639.
Loeng, H., 1991, Features of the physical oceanographic conditions of the Barents Sea. Polar Res. 10 (1), 5–18.
Loeng, H., Drinkwater, K. 2007 An overview of the ecosystems of the Barents and Norwegian Seas and their response to climate variability. Deep-Sea Research II 54: 2478–2500.
Loeng, H., Ottersen, G. 2011. Klimaendringer og effekter på økosystemet i Barentshavet. Ottar 2011 (4) s. 29-33.
Lone, K., Aars, J., Ims, RA. 2013. Site fidelity of Svalbard polar bears revealed by mark-recapture positions. Polar Biol 36: 27-39.
Lone, K., Hamilton, CD., Aars, J., Lydersen, C., Kovacs, KM. 2019. Summer habitat selection by ringed seals (Pusa hispida) in the drifting sea ice of the northern Barents Sea. Polar Res 38: 3483, http://dx.doi.org/10.33265/polar.v38.3483 .
Lone, K., Merkel, B., Lydersen, C., Kovacs, KM., Aars, J. 2018a. Sea ice resource selection models for polar bears of the Barents Sea subpopulation. Ecography 41: 567–78
Louis, M., Skovrind, M., Samaniego Castruita, JA., Garilao, C., Kaschner, K., Gopalakrishnan, S., Haile, JS., Lydersen, C., Kovacs, KM., Garde, E., Heide-Jørgensen, MP., Postma, L., Ferguson, SH., Willerslev, E., Lorenzen, ED. 2020. Influence of past climatic change on phylogeography and demographic history of narwhals, Monodon monoceros Proc Roy Soc B: Biol Sci 287:1925. doi:10.1098/rspb.2019.2964.
Lovejoy, C., von Quillfeldt, CH., Hopcroft, R., Poulin, M., Thaler, M., Arendt, K., Debes, H., Gíslason, Á, Kosobokova, K. 2017. Plankton Ss. 63-84 i CAFF. 2017. State of the Arctic Marine Biodiversity Report. Conservation of Arctic Flora and Fauna International Secretariat, Akureyri, Iceland. 978-9935-431-63-9.
Lowther, AD., Kovacs, KM., Griffiths, D., Lydersen, C. 2015. Identification of motivational state in a free-ranging marine mammal inferred from changes in behaviour. Mar Mamm Sci 31(4): 1291-1313.
Lydersen, C. 1998. Status and biology of ringed seals (Phoca hispida) in Svalbard. NAMMCO Sci. Publ 1: 46-62.
Lydersen, C., Aars, J., Kovacs, KM. 2008. Estimating the number of walruses in Svalbard from aerial surveys and behavioural data from satellite telemetry. Arctic 61: 119-128.
Lydersen, C., Assmy, P., Falk-Petersen, S., Kohler, J., Kovacs, KM., Reigsta,d M., Steen, H., Strøm, H., Sundfjord, A., Varpe, Ø., Walczowski, W., Weslawski, JM., Zajaczkowski, M. 2014. The importance of tidewater glaciers for marine mammals and seabirds in Svalbard, Norway. J Mar Syst 129: 452-471.
Lydersen, C., Freitas, C., Wiig, Ø., Bachmann, L., Heide-Jørgensen, MP., Swift, R., Kovacs, KM. 2012. Lost highway not forgotten: Satellite tracking of a bowhead whale (Balaena mysticetus) from the critically endangered Spitsbergen Stock. Arctic 65: 76-86.
Lydersen, C., Martin, T., Gjertz, I., Kovacs, KM. 2007. Satellite tracking and diving behaviour of sub-adult narwhals (Monodon monoceros) in Svalbard, Norway. Polar Biol 30: 437-442.
Lydersen, C., Martin, AR., Kovacs, KM., Gjertz, I. 2001. Summer and autumn movements of white whales (Delphinapterus leucas) in Svalbard, Norway. Mar Ecol Progr Ser 219: 265-274.
Lydersen, C., Nøst, OA., Lovell, P., McConnell, BJ., Gammelsrød, T., Hunter, C., Fedak, MA., Kovacs, KM. 2002. Salinity and temperature structure of a freezing Arctic fjord - monitored by white whales (Delphinapterus leucas). Geophys Res Lett 29: 2119. doi: 10.1029/2002GL015462.
Lydersen, C., Vacquié-Garcia, J., Lydersen, E., Christensen, GN., Kovacs, KM. 2017. Novel terrestrial haul-out behaviour by ringed seals (Pusa hispida) in Svalbard, in association with harbour seals (Phoca vitulina). Polar Res 36: 1374124, doi:10.1080/17518369.2017.1374124. 6 s.
Lynghammar, A. 2014. Chondrichthyan fishes in the Arctic Ocean and adjacent seas – do we know our species? PhD thesis. UiT The Arctic University of Norway, Tromsø.
Lønne, O., Gabrielsen, G. 1992. Summer diet of seabirds feeding in sea-ice-covered waters near Svalbard. Polar Biology 12. 685-692.
Lønne, OJ., Gulliksen, B. 1991. Sympagic macro-fauna from multiyear sea-ice near Svalbard. Polar Biology 11(7), 471-477.
Marquardt, M., Majaneva, S., Pitusi, V., Søreide, JE. 2018. Pan-Arctic distribution of the hydrozoan Sympagohydra tuuli ? First record in sea ice from Svalbard (European Arctic). Polar Biology 41(3), pp. 583-588.
Martínez Llobet, S., Ahonen, H., Lydersen, C., Berge, J., Ims, R., Kovacs, KM. Innsendt manus. Bearded seal vocalisations across seasons and habitat types in Svalbard, Norway.
McGovern, M., Pavlov, AK., Deininger, A., Granskog, MA., Leu, E., Søreide, JE., Poste, A. 2020. Terrestrial Inputs Drive Seasonality in Organic Matter and Nutrient Biogeochemistry in a High Arctic Fjord System (Isfjorden, Svalbard). Frontiers in Marine Science, 7, 747.
McMahon, KW., Ambrose Jr, WG., Johnson, BJ., Sun, MY., Lopez, GR., Clough, LM., Carroll, ML. 2006. Benthic community response to ice algae and phytoplankton in Ny Ålesund, Svalbard. Marine Ecology Progress Series, 310, s. 1-14.
McMinn, A., Hegseth EN. 2004. Quantum yield and photosynthetic parameters of marine microalgae from the southern Arctic Ocean, Svalbard. J. Mar. Biol. Assoc. UK 84, 865-871.
Mecklenburg, C., Lynghammar, A., Johannesen, E., Byrkjedal, I., Christiansen, J., Dolgov, A., Karamushko, O., Mecklenburg, T., Møller, P., Steinke, D. 2018. Marine Fishes of the Arctic Region. Conservation of Arctic Flora and Fauna. Akureyri, Iceland.
Meld. St. 20 (2019–2020) Helhetlige forvaltningsplaner for de norske havområdene — Barentshavet og havområdene utenfor Lofoten, Norskehavet, og Nordsjøen og Skagerrak. Tilråding fra Klima- og miljødepartementet 24. april 2015, godkjent i statsråd samme dag. (Regjeringen Solberg). https://www.regjeringen.no/no/dokumenter/meld.-st.-20-20192020/id2699370/
Meld. St. 29 (2020-2021). Heilskapleg nasjonal plan for bevaring av viktige områder I marin natur. Tilråding fra Klima- og miljødepartementet, 9. april 2021, godkjend i statsråd same dag. (Regjeringen Solberg). https://www.regjeringen.no/no/dokumenter/meld.-st.-29-20192020/id2715224/
Melle W., Ellertsen B., Skjoldal HR. 2004. In: The Norwegian Sea Ecosystem, H. R. Skjoldal (ed.). Tapir academic press. Trondheim.
Melnikov IA., Kolosova EG., Welch HE., Zhitina LS. 2002. Sea ice biological communities and nutrient dynamics in the Canada Basin of the Arctic Ocean. Deep-Sea Res. 49, 1623–1649.
Menze, S., Ingvaldsen, R.B., Haugan, P., Fer, I., Sundfjord, A., Beszczynska-Möller, A, Falk-Petersen, S., 2019. Atlantic Water Pathways Along the North-Western Svalbard Shelf Mapped Using Vessel-Mounted Current Profilers. Journal of Geophysical Research: Oceans, DOI: 10.1029/2018JC014299.
Menze, S., Ingvaldsen, RB., Nikolopoulos, A., Hattermann, T., Albertsen, JA., Gjøsæter, H. 2020. Productive detours- Atlantic water inflow and acoustic backscatter in the major troughs along the Svalbard shelf. Prog. Ocean. 188:1-14
Meredith, MM., Sommerkorn, S., Cassotta, C., Derksen, A., Ekaykin, A., Hollowed, G., Kofinas, A., Mackintosh, J., Melbourne-Thomas, M., Muelbert, MC., Ottersen, G., Pritchard, H., Schuur, EAG. 2019: Polar Regions. In: IPCC Special Report on the Ocean and Cryosphere in a Changing Climate [H.-O. Pörtner, D.C. Roberts, V. Masson-Delmotte, P. Zhai, M. Tignor, E. Poloczanska, K. Mintenbeck, A. Alegría, M. Nicolai, A. Okem, J. Petzold, B. Rama, N.M. Weyer (Red.)]. IPCC, Gland Switzerland.
Merkel, B., Lydersen, C., Yoccoz, NG., Kovacs, KM. 2013. The world's northernmost harbour seal population - how many are there? PLoS ONE 8: e67576.
Michalsen, K., Hvingel, C., Orlova, EL., Bogstad, B., Dolgov, AV., Høines, Å., McBride, MM. 2007. Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea August-September 2007. Vol. 2.
Moore, SE., Stafford, KM., Melling, H., Berchok, C., Wiig, Ø., Kovacs, KM., Lydersen, C., Richter-Menge, J. 2012. Comparing marine mammal acoustic habitats in Atlantic and Pacific sectors of the High Arctic: year-long records from Fram Strait and the Chukchi Plateau. Polar Biol 35: 475-480.
Moran, SB., Lomas, MW., Kelly, RP., Gradinger, R., Iken, K., Mathis, JT. 2012. Seasonal succession of net primary productivity, particulate organic carbon export, and autotrophic community composition in the eastern Bering Sea. Deep Sea Research Part II: Topical Studies in Oceanography, 65–70, 84–97.
Mork, KA., Skagseth, Ø., 2010. A quantitative description of the Norwegian Atlantic Current by combining altimetry and hydrography, Ocean Sci., 6, 901-911, doi:10.5194/os-6-901-2010, 2010.
Muilwijk, M., Smedsrud, LH., Ilicak, M., Drange, H. 2018. Atlantic Water heat transport variability in the 20th century Arctic Ocean from a global ocean model and observations. Journal of Geophysical Research: Oceans, 123, 8159–8179. https://doi.org/10.1029/2018JC014327
Mukhina, N.V., Drevemyak KV. Dvinina, EA. 1992. Redfish spawning grounds in the Barents Sea and adjacent waters. ICES Document CM 2003/G:54. 21 s.
Munk P, Fox CJ, Bolle LJ, van Damme CJG, Fossum P, Kraus G, 2009. Spawning of North Seafishes linked to hydrographic features. Fish. Oceanogr. 18, 458–468.
NAFO, ICES. 2020. NAFO/ICES Pandalus Assessment Group Meeting, 26 to 30 October 2020. By WebEx. NAFO SCS Doc. 20/21. 89 pp.
NAMMCO & IMR 2019. Report of Joint IMR/NAMMCO International Workshop on the Status of Harbour Porpoise in the North Atlantic. Tromsø, Norway. 235 s.
Naustvoll, LJ., Thormar, J., Lundsør, E., Kroglund, T., Norderhaug, KM., Moy, F. 2020. ØKOKYST-Delprogram Klima, Årsrapport 2019. Miljødirektoratet M-1612/2020.
Nilsen, F., Skogseth, R., Vaardal-Lunde, J., Inall, M., 2016. A simple shelf circulation model: intrusion of Atlantic Water on the West Spitsbergen Shelf. Journal of Physical Oceanography, 46,4.
Nilssen, KT., Bjørge, A . 2019. Status for kystsel – Anbefaling av jaktkvoter 2020. S 56-69 in A. Bjørge (ed). Forskerutvalg om sjøpattedyr 2019 – Anbefalinger om forskning og forvaltning. Rapport fra Havforskningen Nr. 22020-19.
Nilssen, KT., Lindstrøm, U., Westgaard, JI., Lindblom, L., Blencke, T-R., Haug, T. 2019. Diet and prey consumption of grey seals (Halichoerus grypus) in Norway. Marine Biology Research 15(2). https://doi.org/10.1080/17451000.2019.1605182
Nizovtsev G.P. 1989. Recommendations on Rational Exploitation of Greenland Halibut Stocks in the Barents and Norwegian Seas. PINRO Press, Murmansk. 93 s. (På Russisk).
Nordeide, JT., Båmstedt, U. 1998. Coastal cod and north-east Arctic cod - do they mingle at the spawning grounds in Lofoten?, Sarsia, 83:5, 373-379, DOI: 10.1080/00364827.1998.10413696
Nöthig, E.-M., Bracher, A., Engel, A, Metfies, K. Niehoff, B., Peeken, I., Bauerfeind, E., Cherkasheva, A., Gäbler-Schwarz S., Hardge, K., Kilias, E., Kraft A., Mebrahtom Kidane, Y., Lalande, C., Piontek J., Thomisch, K., Wurst, M. 2015. Summertime plankton in Fram Strait – a compilation of long- and short-term observations. Polar Research 34: 23349, doi:10.3402.
Nöthig, E.-M., Ramondenc, S., Haas, A., Hehemann, L., Walter, A., Bracher, A., mfl. 2020. Summertime in situ chlorophyll a and particulate organic carbon standing stocks in surface waters of the Fram Strait and the Arctic Ocean (1991–2015). Front. Mar. Sci . 7:350. doi: 10.3389/fmars.2020.00350
O'Corry-Crowe GM, Lydersen C, Heide-Jørgensen MP, Hansen L, Mukhametov LM, Dove O, Kovacs KM. 2010. Population genetic structure and evolutionary history of North Atlantic beluga whales (Delphinapterus leucas) from West Greenland, Svalbard and the White Sea . Polar Biol . 33 , 1179–1194. (10.1007/s00300-010-0807-y) Oljedirektoratet. 2020. Faktakart, https://factmaps.npd.no/factmaps/3_0/
Olli, K., Riser, CW., Wassmann, P., Ratkova, T., Arashkevich, E., Pasternak, A., 2002. Seasonal variation in vertical flux of biogenic matter in the marginal ice zone and the central Barents Sea. Journal of Marine Systems, 38(1-2), s. 189-204.
Olsen, E., Aanes, S., Mehl, S., Holst, J.C., Aglen, A., Gjøsæter, H., 2010. Cod, haddock, saithe, herring, and capelin in the Barents Sea and adjacent waters: a review of the biological value of the area. ICES J. Mar. Sci., 67, 87-101.
Olsen, LM., Laney, SR., Duarte, P., Kauko, HM., Fernández-Méndez, M., Mundy, CJ., Rösel, A., Meyer, A., Itkin, P., Cohen, L., Peeken, I., Tatarek, A., Wiktor, J., Taskjelle, T., Pavlov, AK., Hudson, SR., Granskog, MA., Hop, H., Assmy, P. 2017. The role of multiyear ice in seeding ice algae blooms in Arctic pack ice. J. Geophy. Res, doi:10.1002/2016JG003626.
Onarheim, IH., Eldevik, T., Smedsrud, LH., Stroeve, JC. 2018. Seasonal and Regional Manifestation of Arctic Sea Ice Loss. Journal of Climate 31, 4917-4932. https://doi.org/10.1175/JCLI-D-17-0427.1
Onarheim, IH., Årthun M. 2017. Toward an ice-free Barents Sea, Geophys. Res. Lett., 44, 8387–8395, doi:10.1002/ 2017GL074304.
Orlova, EL., Boitsov, VD., Dolgov, AV., Rudneva, GB., Nesterova, VN. 2005. The relationship between plankton, capelin, and cod under different temperature conditions. ICES Journal of Marine Science, 62: 1281-1292.
Orlova, EL. Dalpadado, P.., Knutsen, t., V. N. Nesterova, VN., Prokopchuk. 2011. Zooplankton. Pages 90–119inT.Jakobsen and V. K. Ozhigin, editors. The Barents Sea.Ecosystem, resourc
Orlova, EL., Boitsov, VD., Nesterova, VN. 2010b. The impact of hydrographic conditions on the structure and functioning of the trophic complex of plankton – pelagic fishes - cod. Pinro press, Murmansk, 190 sider. (På russisk).
Orlova, EL., Dolgov, AV., Renaud, PE., Boitsov, VD., Prokopchuk, IP., Zashihina, MV. 2013. Structure of the macroplankton–pelagic fish–cod trophic complex in a warmer Barents Sea. Marine Biology Research 9, 851-866.
Orlova, EL., Dolgov, AV., Renaud, PE., Greenacre, M., Halsband, C., Ivshin, VA. 2015. Climatic and ecological drivers of euphasiid community structure vary spatially in the Barents Sea: relationships from a long time series (1952-2009). Frontiers of Marine Science, https://doi.org/10.3389/fmars.2014.00074
Orlova, EL., Rudneva, G.B., Renaud, P.E., Eiane, K., Savinov, V., Yurko, A.S. 2010a. Climate impacts on feeding and condition of capelin Mallotus villosus in the Barents Sea: evidence and mechanisms from a 30 year data set. Aquat. Biol. 10:105-118.
Orvik KA og Niiler PP, 2002. Major pathways of the Atlantic water in the northern North Atlantic and Nordic Seas toward Arctic, Geophys. Res. Lett. , 29 (19), 1896, doi: 10.1029/2002GL015002 .
Oudman, T., Laland, K., Ruxton, G., Tombre, I., Shimmings, P., Proup, J. 2020. Young Birds Switch but Old Birds Lead: How Barnacle Geese Adjust Migratory Habits to Environmental Change. Frontiers in Ecology and Evolution 7, 1-15.
Owrid, G., Socal, G., Civitarese, G., Luchetta, A., Wiktor, J., Nothig, EM., Andreassen, I., Schauer, U., Strass, S. 2000. Spatial variability of phytoplankton, nutrients and new production estimates in the waters around Svalbard. Polar Research 19(2). 155-171.
Ozhigin, VK., Ingvaldsen, RB., Loeng, H., Boitzov, VD., Karsakov, AL. 2011. Kap.2. The Barents Sea. Side 39-76 I Jakobsen T. Ozhigin, VK. (red.) The Barents Sea. Ecosystem, resources, management. Half a century of Russian-Norwegian cooperation. Tapir Academic Press, Trondheim.
Oziel, L., Neukermans, G., Ardyna, M., Lancelot, C., Tison, J-L., Wassmann, P., Sirven, J., Ruiz-Pino, D., Garscard, J-C. 2017 Role of Atlantic inflows and sea ice loss on shifting phytoplankton blooms in the Barents Sea. Journal og Geophysical Research: Oceans. doi/full/10.1002/2016JC012582
Oziel, L., Baudena, A., Ardyna, M., Massicotte, P., Randelhoff, A., Sallee, J.-B. mfl. 2020. Faster Atlantic currents drive poleward expansion of temperate phytoplankton in the Arctic Ocean. Nat. Commun. 11:1705. doi: 10.1038/s41467-020-15485-5
Pavlova, O., Gerland, S. Hop, H. 2019. Changes in sea-ice extent and thickness in Kongsfjorden, Svalbard (2003-2016). In: Hop H, Wiencke C (Red) The ecosystem of Kongsfjorden, Svalbard. Advances in Polar Ecology 2, Springer Verlag
Pavlova, O., Pavlov, V., Gerland, S. 2014 The impact of winds and sea surface temperatures on the Barents Sea ice extent, a statistical approach, J. Mar. Syst. 130: 248–255, doi:10.1016/j.jmarsys.2013.02.011.
Pedersen, C-E., Falk, K. 2001 Chick diet of dovekies Alle alle in Northwest Greenland. Polar Biol. 24(1), 53-58.
Pedersen, OP., Aschan, M., Rasmussen, T., Tande, KS., Slagstad, D. 2003. Larval dispersal and mother populations of Pandalus borealis investigated by a Lagrangian particle-tracking model. Fisheries Research 65, 173-190.
Pitusi V. 2019. Seasonal abundance and activity of sympagic meiofauna in van Mijenfjorden, Svalbard. Master thesis, UiT The University of Norway.
Pizzolato, L., Howell, SEL., Dawson, J., Laliberte, F., Copland, L. 2016. The influence of declining sea ice on shipping activity in the Canadian Arctic. Geophysical Research Letters, 43 (23), 12,146– 12,154, doi:10.1002/2016gl071489.
Poltermann M., 1998. Abundance, biomass and small-scale distribution of cryopelagic amphipods in the Franz Josef Land area (Arctic). Polar Biology 20 (2), 134-138.
Polyakov, IV., Pnyushkov, AV., Alkire, MB., Ashik, IM., Baumann, TM., Carmack, EM., fl.2017. Greater role for Atlantic inflows on sea-ice loss in the Eurasian Basin of the Arctic Ocean. Science 356, 285–291. oi: 10.1126/science.aai8204
Popova, E., Yool, A., Aksenov, Y., Coward, A. 2013. Role of advection in Arctic Ocean lower trophic dynamics: a modeling perspective. J. Geophys. Res. Oceans 118, 1571–1586. doi: 10.1002/jgrc.20126
Poulin, M., Daugbjerg, N., Gradinger, R., Ilyash, L., Ratkova, T., von Quillfeldt, CH. 2011. The pan-Arctic biodiversity of marine pelagic and sea-ice unicellular eukaryotes: A first-attempt assessment Marine Biodiversity. Marine Biodiversity DOI 10.1007/s12526-010-0058-8./41 (1), 13-28.
Prokopchuk, I., 2019. Feeding ecology of immature herring Clupea harengus in the Barents Sea. In: Shamray, E., Huse, G., Trofimov, A., Sundby, S., Dolgov, A., Skjoldal, H. R., Sokolov, K., Jorgensen, L. L., Filin, A., Haug, T., Zabavnikov, V. (Red.), Influence of Ecosystem Changes on Harvestable Resources at High Latitudes. The Proceedings of the 18th Russian-Norwegian Symposium, Murmansk, Russia, 5–7 June 2018, IMR/PINRO Joint Report Series, No. 1-2019, s. 151–160 (217 s).
Rasmussen, B. 1953. On the geographical variation in growth and sexual development of the deep sea prawn (Pandalus borealis Kr.). Fiskeridirektoratets skrifter, Serie Havundersøkelser Vol. X. No 3. 160 s.
Reder, S., Lydersen, C., Arnold, W., Kovacs, KM. 2003. Haulout behaviour of High Arctic harbour seals (Phoca vitulina vitulina) in Svalbard, Norway. Polar Biology 27(1), 6-16. DOI: 10.1007/s00300-003-0557-1
Reeves, RR., Ewins, PJ., Agbayani, S., Heide-Jørgensen, MP., Kovacs, KM., Lydersen, C., Suydam, R., Elliot, W., Polet, G., Dijk, Y., Blijleven, R. 2014. Distribution of endemic cetaceans in relation to hydrocarbon development and commercial shipping in a warming Arctic. Mar.Policy 44: 375-389.
Reiertsen, TK., Erikstad, KE., Anker-Nilssen, T., Barrett, RT., Boulinier, T., Frederiksen, M., González-Solís, J., Gremillet, D., Johns, D., Moe, B., mfl. 2014. Prey density in non-breeding areas affects adult survival of black-legged kittiwakes Rissa tridactyla . Marine Ecology ProgressSeries, 509: 289-302. doi: 10.3354/meps10825.
Reigstad M., Carroll J., Slagstad, D., Ellingsen, I., Wassmann, P. 2011. Intra-regional comparison of productivity, carbon flux and ecosystem composition within the northern Barents Sea. Progress in Oceanography 90(1-4), 33-46.
Renaut, S., Devred, E., Babin M. 2018. Northward expansion and intensification of phytoplankton growth during the early ice-free season in Arctic. Geophys. Res. Lett. 45, 10590-10598.
Ressler, PH., Dalpadado, P., Macaulay, GJ., Handegard, N., Skern-Mauritzen, M. 2015. Acoustic surveys of euphausiids and models of baleen whale distribution in the Barents Sea. Mar. Ecol. Prog. Ser. 527:13-29
Rey, F, 2012. Declining silicate concentrations in the Norwegian and Barents Seas. ICES J Mar Sci, 69: 1-5.
Rodrigues, J., 2008. The rapid decline of the sea ice in the Russian Arctic. Cold Reg. Sci. and Tech. 54, 124–142.
Rodrigues, J., 2009. The increase in the length of the ice-free season in the Arctic. Cold Reg. Sci. and Tech. 59, 78–101.
Runnstrøm, S. 1941. Quantitative investigations on herring spawning and its yearly fluctuations at the west coast of Norway. – Fiskeridirektoratets Skrifter Serie Havundersøkelser 6(8), 5-71.
Rusten, M. (red) 2014. Development of methodology for calculations of environmental risk for the marginal ice zone – a joint project between Akvaplan-niva and DNV GL. Report No.: 2014-0545, Rev00.
Rybakova, E., Kremenetskaia, A., Vedenin, A., Boetius, A., Gebruk A. 2019. Deep-sea megabenthos communities of the Eurasian Central Arctic are influenced by ice-cover and sea-ice algal falls. bioRxiv:515015.
Sakshaug E. 2004. Primary and secondary production in the Arctic Sea, in Stein, R., Macdonald, R.W. (eds): The organic carbon cycle in the Arctic Ocean. Springer, Berlin, pp. 57-81.
Sakshaug, E., Johnsen, G., Kristiansen, S., von Quillfeldt, CH., Rey, F., Slagstad, D., Thingstad, F. 2009. Phytoplankton and primary production. S. 167 – 208 in Sakshaug E., Johnsen G., Kovacs K. (Red.) Ecosystem Barents Sea. Tapir Academic Press, Trondheim, Norway.
Savvatimsky, PI. 1985. Features of spawning concentrations of roughhead grenadier Macrocurusberglax L. at the Norwegian coast. ICES CM 1985/G:6, 34 s.
Scott, CL., Falk-Petersen, S., Gulliksen, B., Lønne OJ., Sargent, JR. 2001. Lipid indicators of the diet of the sympagic amphipod Gammarus wilkitzkii in the Marginal Ice Zone and in open waters of Svalbard (Arctic). Polar Biology, 24(8), s. 572-576.
Shpak, OV., Paramonov, AY. 2018 The bowhead whale, Balaena mysticetus Linnaeus, 1758, in the western Sea of Okhotsk 2009–2916. distribution pattern, behaviour and threats. Russian J Mar Biol 44: 210–218.
Shumway, SE., Perkins, HC., Schick, DF., Stickney, AP. 1985. Synopsis of Biological Data on the Pink Shrimp, Pandalus borealis Krøyer, 1838, NOAA Technical Report NMFS 30, FAO Fisheries Synopsis No. 144. 65 pp.
Similä, T., Holst, JC., Christensen, I. 1996. Occurrence and diet of killer whales in northern Norway: seasonal patterns relative to the distribution and abundance of Norwegian spring-spawning herring. Can. J. Fish. Aquat. Sci. 53 (4):769-779.
Similä, T., Ugarte, F. 1996. Surface and underwater observations of cooperatively feeding killer whales in northern Norway. Canadian Journal of Zoology: 71(8): 1494-1499. https://doi.org/10.1139/z93-210
Sims, DW. 2008. Sieving a living: a review of the biology, ecology and conservation status of the plankton‐feeding basking shark Cetorhinus maximus . Advances in marine biology, 54, s.171-220.
Sims, DW., Quayle, VA., 1998. Selective foraging behaviour of basking sharks on zooplankton in a small-scale front. Nature 393, 460–464.
Skagseth Ø., Drinkwater, KF., Terrile, E. 2011. Wind- and buoyancy-induced transport of the Norwegian Coastal Current in the Barents Sea. Journal of Geophysical Research, 116, C08007, doi:10.1029/2011JC006996.
Skagseth, Ø., Eldevik, T., Årthun, M., Asbjørnsen, H., Lien, VS., Smedsrud, LH. 2020. Reduced efficiency of the Barents Sea cooling machine. Nat. Clim. Change 10, 661–666 .
Skagseth, Ø. Mork, KA. 2012. Heat content in the Norwegian Sea, 1995-2010. ICES J. Mar. Sci., 69, 826-832, doi: 10.1093/icesjms/fsss026Skjoldal H.R. (Ed.) 2004. The Norwegian Sea Ecosystem, Tapir Academic Press, Trondheim, Norway, 559 s.
Skardhamar, J., Skagseth, Ø., Albretsen, J., 2015. Diurnal tides on the Barents Sea continental slope. Deep-Sea Research I 97, 40-51.
Skardhamar, J., Svendsen, H., 2005. Circulation and shelf-ocean interaction off North Norway. Continental Shelf Research, 25, 1541-1560.
Skern‐Mauritzen, M., Johannesen, E., Bjørge, A., Øien, N. 2011. Baleen whale distributions and prey associations in the Barents Sea. Mar. Ecol. Prog. Ser . 426: 289– 301. doi: 10.3354/meps09027
Skern-Mauritzen, M., Skaug, H., Øien, N. 2009. Line transects, environmental data and GIS: Cetacean distribution, habitat and prey selection along the Barents Sea shelf edge. In: Øien N, Pike D, editors. North Atlantic Sighting Surveys. Counting the Whales in the North Atlantic, 1987–2001. Tromsø: NAMMCO Scientific Publications 7: 179–200.
Skogen, MD., Olsen, A., Børsheim, KY., Sandø, AB., Skjelvan, I., 2014 Modelling ocean acidification in the Nordic and Barents Seas in present and future climate. J Mar Sys, 131:10-20.
Skogseth, R., Olivier, LL, Nilsen, F., Falck, E., Fraser, N., Tverberg, V., mfl. 2020. Variability and decadal trends in the Isfjorden (Svalbard) ocean climate and circulation an indicator for climate change in the European Arctic. Progress in Oceanography, 102394.
Slagstad, D., Tande, K., Wassmann, P. (1999). Modelled carbon fluxes as validated by field data on the north Norwegian shelf during the productive period in 1994. Sarsia 84, 303–317. doi: 10.1080/00364827.1999.10420434
Smedsrud, LH, Esau, I, Ingvaldsen, RB, Eldevik, T, Haugan, PM. mfl. 2013 The role of the Barents Sea in the Arctic climate system. Reviews of Geophysics, 51 : 415–449.
Solemdal, P., Knutsen, T., Bjørke, H. 1989. Spawning areas and spawning period of the Northeast Arctic haddock (Melanogrammus aeglefinus L.). Havforskningsinstituttets egg- og larveprogram (HELP) No. 25/1989. 40 s. http://hdl.handle.net/11250/115257
Solemdal, P., Mukhina, N. V., Knutsen, T., Bjørke, H., Fossum, P. 1997. Maturation, spawning and egg drift of Arcto-Norwegian haddock (Melanogrammus aeglefinus). Poster presented at the Fisheries Society of the British Isles Annual Symposium, 8–11 July 1997. University College, Galway, Ireland. http://hdl.handle.net/11250/195225
Sonne, C., Letcher, RJ., Jenssen, BM, Desforges, JP., Eulaers, I., Andersen-Ranberg, E., Gustavson, K., Styrishave, B, Dietz, R. 2017. A veterinary perspective on One Health in the Arctic. Acta Veterinaria Scandinavica 59(1): 1-11.
Spreen, G., de Steur, L., Divine, D., Gerland, S., Hansen, E., Kwok, R. 2020. Arctic sea ice volume export through Fram Strait from 1992 to 2014. Journal of Geophysical Research: Oceans , 125 (6), e2019JC016039.
Stafford, KM., Moore, SE., Berchok, CL., Wiig, Ø., Lydersen, C., Hansen, E., Kalmbach, D., Kovacs, KM. 2012. Spitsbergen’s endangered bowhead whales sing through the polar night. Endang Spec Res 18: 95-103. doi: 10.3354/esr00444
Stern H, Laidre KL. 2016. Sea-Ice indicators of Polar Bear Habitat. The Cryosphere 10: 1-15.
Stige, LC., Eriksen, E., Dalpadado P., Kotaro O. 2019. Direct and indirect effects of sea ice cover on major zooplankton groups and planktivorous fishes in the Barents Sea. ICES Journal of Marine Science, in press. https://doi.org/10.1093/icesjms/fsz063
Storrie, L., Lydersen, C., Andersen, M., Wynn, RB., Kovacs, KM. 2018. Determining the species assemblage and habitat use of cetaceans in the Svalbard Archipelago, based on observations from 2002-2014. Polar Res 37: 1463065. doi.org/10.1080/17518369.2018.1463065.
Strand, E., Bagøien, E., Edwards, M., Broms, C., Klevjer, T. 2020. Spatial distribution and seasonality of four Calanus species in the Northeast Atlantic. Progress in Oceanography, 185: 102344.
Strøm, H., Bakken, V., Skoglund, A., Descamps, S., Fjeldheim, V. and Steen, H. 2020 . Population status and trend of the threatened ivory gull Pagophila eburnea in Svalbard. Endangered Species Research 43:435-445. DOI: 10.3354/esr01081
Sundby, S. 2000. Recruitment of Atlantic cod stocks in relation to temperature and advection of copepod populations. Sarsia, 85:277-298.
Sundby, S. (red.) 2013. Kunnskapsinnhenting Barentshavet–Lofoten–Vesterålen (KILO). Fisken og Havet 3-2013. Havforskningsinstituttet.
Sundby, S., Nakken, O., 2008. Spatial shifts in spawning habitats of Arcto-Norwegian cod related to multidecadal climate oscillations and climate change. ICES J Mar Sci 65: 953–962
Svendsen, H., Beszczynska-Møller, A., Hagen, JO., Lefauconnier, B., Tverberg, V., Gerland. S., Ørbæk, JB., Bischof, K., Papucci, C., Zajaczkowski, M., Azzolini, R., Bruland, O., Wiencke, C., Winther. J-G., Dallmann, W., 2002. The physical environment of Kongsfjorden-Krossfjorden, an Arctic fjord system in Svalbard. Polar Res 21:133–166.
Systad, GHR., Bjørgesæter, A., Brude, OW., Skeie, GM. 2018. Standardisering og tilrettelegging av sjøfugldata til bruk i konsekvens- og miljørisikoberegninger. NINA Rapport 1509. Norsk institutt for naturforskning.
Systad, GHR., Fauchald, P., Descamps, S., Christensen-Dalsgaard, S., Strøm, H., Tarroux, A. 2019. Identifisering av viktige områder for sjøfugl i norske havområder – innspill til forvaltningsplanarbeidet 2018. NINA Rapport 1627.
Syvertsen, EE. 1991. Ice algae in the Barents Sea: types of assemblages, origin, fate and role in the ice-edge phytoplankton bloom. Polar Res. 10(1), 277-288.
Szymanski, A., Gradinger, R. 2016. The diversity, abundance and fate of ice algae and phytoplankton in the Bering Sea. Polar Biology 39, 309-325.
Sømme JD. 1934. Animal Plankton of the Norwegian Coast Waters and the open Sea. Fiskeridirektoratets Skrifter, Serie Havundersøkelser, IV, NO.9.
Søreide, JE., Carroll, ML., Hop, H., Ambrose Jr, WG., Hegseth, EN., Falk-Petersen, S. 2013. Sympagic-pelagic-benthic coupling in Arctic and Atlantic waters around Svalbard revealed by stable isotopic and fatty acid tracers. Marine Biology Research 9(9), 831-850.
Søreide, JE., Falk-Petersen, S., Hegseth, EN., Hop, H., Carroll, ML., Hobson, KA., Blachowiak-Samolyk, K. 2008. Seasonal feeding strategies of Calanus in the high-Arctic Svalbard region. Deep-Sea Research Part II Topical Studies in Oceanography, 55: 2225–2244.
Søreide, JE., Leu, E., Berge, J., Graeve, M., Falk-Petersen, S. 2010. Timing in blooms, algal food quality and Calanus glacialis reproduction and growth in a changing Arctic. Global Change Biology 16, 3154–3163.
Søvik, G., Nedreaas, K., Zimmermann, F., Husson, B., Strand, HK., Jørgensen, LL., Strand, M., Thangstad, TH., Hansen, A., Båtevik, T., Albretsen, J., Staby, A. 2020. Kartlegging av fjordøkosystemene i Tana- og Porsangerfjorden. Råd og kunnskapsbidrag fra Havforskningsinstituttet i forbindelse med vurdering av en eventuell åpning av direktefiske etter reker med bunntrål i Tana- og Porsangerfjorden. Rapport fra Havforskningen 2020-39, ISSN:1893-4536. 140 s. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-2020-39
Tande, KS. 1989. Calanus in North Norwegian fjords and in the Barents Sea. S. 38Y-407 I: Sakshaug. E., Hopkins, CCE., Øritsland. NA. (Red.): Proceedings of the Pro Mare Symposium on Polar Marine Ecology. Trondheim. 12-16 May 1990. Polar Research.
Tedesco, L., Vichi, M., Scoccimarro, E. 2019. Sea-ice algal phenology in a warmer Arctic. Science Advances 5, eaav4830.
Tombre, IM., Oudman, T., Shimmings, P., Griffin, L., Prop, J. 2019. Northward range expansion in spring-staging barnacle geese is a response to climate change and population growth, mediated by individual experience. Glob. Chang. Biol. 23, 3680–3693. doi: 10.1111/gcb.14793
Townhill, BL, Holt, RE, Bogstad, B., Durant, JM., Pinnegar, JK., Dolov, AV., Yaragina, NA., Johannesen, E., Ottersen, G. 2020. Diets of the Barents Sea cod from the 1930s to the present day. Earth System Science Data. https://doi.org/10.5194/essd-2020-96.pdf
Trudnowska, E., Gluchowska, M., Beszczynska-Möller, A., Blachowiak-Samolyk K., Kwasniewski, S. 2016. Plankton patchiness in the Polar Front region of the West Spitsbergen Shelf. Mar Ecol Prog Ser 560.1-18. https://doi.org/10.3354/meps11925
Tverberg, V., Skogseth, R., Cottier, F., Sundfjord, A., Walczowski, W., Inall, M., Falck, E., Pavlova, O., Nilsen, F. 2019. The Kongsfjorden Transect: seasonal and inter-annual variability in hydrography. I: H. Hop, C. Wiencke (Red.): The ecosystem Kongsfjorden, Svalbard. Berlin: Springer.
Vacquie-Garcia, J., Lydersen, C., Biuw, M., Haug, T., Fedak, MA., Kovacs, K.M. 2017a. Hooded seal Cystophora cristata foraging areas in the Northeast Atlantic Ocean - investigated using three complementary methods. PLoS ONE 12, e0187889, 23 pp
Vacquie-Garcia, J., Lydersen, C., Ims, RA., Kovacs, KM. 2018. Habitats and movement patterns of white whales Delphinapterus leucas in Svalbard, Norway in a changing climate. Movement Ecol 621: 1-12.
Vacqui é -Garcia, J., Lydersen, C., Kovacs, KM. 2019. Diving behavior of adult male white whales, Delphinapterus leucas , in Svalbard, Norway. Polar Res 41: 253-263.
Vacquié-Garcia, J., Lydersen, C., Marques, TA., Aars, J., Ahonen, H., Skern-Mauritsen, M., Øien, N., Kovacs, KM. 2017b. Late summer distribution and abundance of ice-associated whales in the Norwegian High Arctic. Endang Spec Res 32: 59-70. doi: 10.3354/esr00791.
Vacquié-Garcia, J., Lydersen, C., Marques, TA., Andersen, M., Kovacs, KM. 2020. First abundance estimate for white whales (Delphinapterus leucas) in Svalbard, Norway. Endang Spec Res 41: 253-263.
VanWormer, E., Mazet, JAK., Hall, A., Gill, VA., Boveng, PL., London, JM., Gelatt, T., Fadely, BS., Lander, ME., Sterling, J., Burkanov, VN., Ream, RR., Brock, PM., Rea, LD., Smith, BR., Jeffers, A., Henstock, M., Rehberg, M.J., Burek-Huntington, K.A., Cosby. S,L,, Hammond, J.A. and Goldstein, T. 2019. Viral emergence in marine mammals in the North Pacific may be linked to Arctic sea ice reduction. Sci Rep 9: 1-11.
Velvin, R., Christensen, G., Mannvik, H-P., Eikrem, W., Hermansen, S., Aasen, AT., Dahl-Hansen, G., Ivarjord, T., Tobiesen, A., Bryntesen, T., Larsen, G., Kistenich, S. 2020. ØKOKYST Delprogram Norskehavet Nord (I), Årsrapport 2019. Miljødirektoratet M-1608
Vernet, M., Ellingsen, IH., Seuthe, L., Slagstad, D., Cape, MR. Matrai, PA. 2019. Influence of Phytoplankton Advection on the Productivity Along the Atlantic Water Inflow to the Arctic Ocean. Front. Mar. Sci. 6:583. doi: 10.3389/fmars.2019.00583
Vikebø, F., Sundby, S., Ådlandsvik, B., Fiksen, Ø. 2005. The combined effect of transport and temperature on distribution and growth of larvae and pelagic juveniles of Arcto-Norwegian cod. ICES J. Mar. Sci., 62: 1375e1386.
Vikebø, FB., Strand, KO., Sundby, S. 2019. Wind Intensity Is Key to Phytoplankton Spring Bloom Under Climate Change. Front. Mar. Sci. 6:518. doi: 10.3389/fmars.2019.00518
Vinje, T., 1998. On the variation during the past 400 years of the Barents Sea ice edge position and Northern Hemisphere temperatures. In: Proceedings of the WCRP Symposium on Polar Processes and Global Climate, Rosario, Orcas Island, WA, USA, s. 271–273.
Vinje, T., 2001. Anomalies and trends of sea-ice extent and atmospheric circulation in the Nordic Seas during the period 1864–1998. J. Clim. 14, 255–267.
Vinje, T., Kvambekk, AS., 1991. Barents Sea drift ice characteristics. Pol. Res. 10(1), 59-68. von Quillfeldt C.H. (red.) 2018. Miljøverdier og sårbarhet i iskantsonen. Norwegian Polar Institute, Brief report no. 047, 262.
Vollen, T., Albert, OT., Nilssen, EM. 2004. Diet composition and feeding behaviour of juvenile Greenland halibut (Reinhardtius hippoglossoides) in the Svalbard area. Journal of Sea Research, 51 (3–4): 251–259.
Vonnahme, T., Leroy, M. Thoms, S., van Oevelen, D., Harvey, R., Kristiansen, S., Gradinger, R., Dietrich, U.. Völker, C., 2021. Modeling silicate–nitrate–ammonium co-limitation of algal growth and the importance of bacterial remineralization based on an experimental Arctic coastal spring bloom culture study. Biogeosciences ISSN 1726-4170.s doi: 10.5194/bg-18-1719-2021.
von Quillfeldt CH. (red.) 2018. Miljøverdier og sårbarhet i iskantsonen. Norwegian Polar Institute, Brief report no. 047, 262 s.
Wadhams, P. 1986. The seasonal ice zone, in The Geophysics of Sea Ice, edited by N. Untersteiner, s. 825–991, Plenum, N.Y.
Walczowski, W., Beszczynska-Möller, A., Wieczorek, P., Merchel, M., Grynczel, A., 2017. Oceanographic observations in the Nordic Sea and Fram Strait in 2016 under the IOPAN long term monitoring program AREX. Oceanologia 59:187–194
Walczowski, W., Piechura, J., Goszczko, I., Wieczorek, P., 2012. Changes in Atlantic water properties: an important factor in the European Arctic marine climate. ICES J Mar Syst 69:864 869.
Walday, M. Christensen, G., Eikrem, W., Fagerli, CW., Frigstad, H., Kaurin, MR, S, A., Trannum, H. 2020. Evaluering av ØKOKYST - stasjonsnett og klassegrenser. Miljødirektoratet rapport M-1784|2020. 74 s.
Walkusz, W., Storemark, K., Skau, T., Gannefors Krafft, C., Lundberg, M. 2003. Zooplankton community structure; a comparison of fjords, open water and ice stations in the Svalbard area. Polish Polar Research. 24.
Wang C, Cheng B, Wang K, Gerland S, Pavlova O, 2015. Modelling snow ice and superimposed ice on landfast sea ice in Kongsfjorden, Svalbard. Polar Res 34, 20828. http://dx.doi.org/10.3402/polar.v34.20828
Wang, SW., Budge, SM., Iken, K., Gradinger, RR., Springer, AM., Wooller, MJ. 2015. Importance of sympagic production to Bering Sea zooplankton as revealed from fatty acid-carbon stable isotope analyses. Marine Ecology Progress Series, 518, s. 31-50.
Wassmann, P., Kosobokova, KN., Slagstad, D., Drinkwater, KF., Hopcroft, RR., Moore, SE. mfl. 2015. The contiguous domains of Arctic Ocean advection: trails of life and death. Progr. Oceanogr. 139, 42–65. doi: 10.1016/j.pocean.2015.06.011
Wassmann, P., Reigstad, M. 2011. Future Arctic Ocean seasonal ice zones and implications for pelagic-benthic coupling. Oceanogr. 24, 220-231.
Wassmann, P., Reigstad, M., Haug, T., Rudels, B., Carroll, ML., Hop, H., Gabrielsen, GW., Falk‐Petersen, S., Denisenko, SG., Arashkevich, E., Slagstad, D., Pavlova, O. 2006. Food webs and carbon flux in the Barents Sea. Progress in Oceanography 71, 232‐287.
Wassmann, P., Slagstad, D., Ellingsen, I., Ross, RM. 2019. Advection of Mesozooplankton Into the Northern Svalbard Shelf Region. Front. Mar. Sci. 6, 1–10.
Wathne, JA., Haug, T., Lydersen, C. 2000. Prey preferences and niche overlap of ringed seals Phoca hispida and harp seals P. groenlandica in the Barents Sea. Mar Ecol Prog Ser 194: 233-239.
Wenneck, TdL. 2005. Mengdeindekser for 19 «ikke-kommersielle» arter i Barentshavet basert på bunnfisktokt «Vintertoktet» -i perioden 1981-2003. FISKEN OG HAVET nr 2-2005.
Werner, I., Auel, H., 2005. Seasonal variability in abundance, respiration and lipid composition of Arctic under-ice amphipods. Marine Ecology Progress Series, 292, s. 251-262.
Węsławski, JM., Adamski, P. 1987. Cold and warm years on South-Spitsbergen coastal marine ecosystems. Polar. Res. 8, 95-106.
Weslawski, JM., Dragańska‐Deja , K., Legeżyńska , J., Walczowski, W. 2018. Range extension of a boreal amphipod Gammarus oceanicus in the warming Arctic. Ecology and Evolution. DOI: 10.1002/ece3.4281
Weslawski, JM., Hacquebord, L, Stempniewicz, L, Malinga. M. 2000. Greenland whales and walruses in the Svalbard food web before and after exploitation. Oceanologia, 42: 37-56.
Węsławski, JM., Kwaśniewski, S., Wiktor, J., Zajączkowski, M. 1993. Observations on the fast ice biota in the fjords of Spitsbergen. Polish Polar Research, pp. 331-343.
Węsławski, JM., Stempniewicz, L., Mehlum, F., Kwaśniewski, S. 1999. Summer feeding strategy of the little auk (Alle alle) from Bjørnøya, Barents Sea. Polar Biology 21 (3), 129-134.
Weslawski, JM., Wiktor, J., Kotwicki, L. 2010. Increase in biodiversity in the arctic rocky littoral, Sorkappland, Svalbard, after 20 years of climate warming. Marine Biodiversity, 40(2), s. 123-130. DOI 10.1007/s12526-010-0038-z
Wiborg KF. 1954. Investigations on zooplankton in Coastal and Offshore waters of Western and Northwestern Norway. Fiskeridirektoratets skrifter, serie Havundersøkelser, XI, no 1.
Wiedmann, I., Ershova, E., Bluhm, BA., Nöthig E-M., Gradinger, RR., Kosobokova, K., Boetius, A. 2020. What Feeds the Benthos in the Arctic Basins? Assembling a Carbon Budget for the Deep Arctic Ocean. Frontiers in Marine Science 7, 224.
Wienerroither, R., Johannesen, E., Dolgov, AV., Byrkjedal, I., Bjelland, O., Drevetnyak, K., Eriksen, KB., Høines, Å., Langhelle, G., Langøy, H., Prokhorova, T., Prozorkevich, D., Wenneck, T. 2011. Atlas of the Barents Sea Fishes. IMR/PINRO Joint Report Series 1-2011. 272 s.
Wiig Ø, Derocher, AE., Belikov, SE. 1999. Ringed seal (Phoca hispida) breeding in the drifting pack ice of the Barents Sea. Mar Mamm Sc. 15: 595-598.
Zakharov, DV., Jørgensen, LL. 2017. New Species of the Gastropods in the Barents Sea and Adjacent Waters. Rusian Journal of Biological INvasions, 8(3) s. 226-231. ISSN 2075-1117.
Zakharov, DV., Jørgensen, LL., Manushin, IE., Strelkova, NA. 2020. Barents Sea megabenthos: Spatial and temporal distribution and production. Marine Biological Journal, 5(2), s.19-37. https://mbj.marine-research.org ; doi: 10.21072/mbj.2020.05.2.03
Zeyl, E., Aars, J., Ehrich, D., Wiig, Ø. 2009. Families in space: relatedness in the Barents Sea population of polar bears (Ursus maritimus). Mol Ecol 18: 735-49.
Zhang, J., Schweige,r A., Steele, M., Stern, H. 2015. Sea ice floe size distribution in the marginal ice zone: Theory and numerical experiments, J. Geophys. Res. Oceans, 120, doi:10.1002/2015JC010770.
Zhou, M., Tande, KS., Zhu, Y., Basedow, S., 2009. Productivity, trophic levels and size spectra of zooplankton in northern Norwegain shelf regions. Deep Sea Res. II: Topical studies in Oceanography, 56:1934-1944.
Zimmermann, F., Søvik, G., Thangstad, TH. 2019. Kunnskapsstatus rekefelt langs norskekysten. Bestilling fra Fiskeridirektoratet. Rapport fra Havforskningen nr. 2019-15. ISSN:1893-4536. 13 s.
Øien, N. (2009). Distribution and abundance of large whales in Norwegian and adjacent waters based on ship surveys 1995-2001. NAMMCO Scientific Publications , 7 , 31-47. https://doi.org/10.7557/3.2704
Øren, K., Kovacs, KM., Yoccoz, NG., Lydersen, C. 2018. Assessing site-use and sources of disturbance at walrus haul-outs using monitoring cameras. Polar Biol 41: 1737-1750.
Ådlandsvik, B, Loeng, H, 1991. A study of the climatic system in the Barents Sea. Pol. Res. 10, 45–49.
Årthun, M., Eldevik, T., Smedsrud, L.H. 2019. The role of Atlantic heat transport in future Arctic winter sea ice loss. Journal of Climate, 32 (11), 3327-3341.
Årthun, M., Eldevik T., Smedsrud LH., Skagseth Ø., Ingvaldsen RB. 2012. Quantifying the influence of Atlantic heat on Barents Sea ice variability and retreat. Journal of Climate, 25(13), s. 4736-4743.
Årthun, M., Ingvaldsen, RB., Smedsrud, LH., Schrum, C. 2011. Dense water formation and circulation in the Barents Sea. Deep-Sea Res I 58: 801-817.
13 - Referanser Norskehavet
Abdullah, MI., Fredriksen, S. 2004. Production, respiration and exudation of dissolved organic matter by the kelp Laminaria hyperborea along the west coast of Norway. Journal of the Marine Biological Association of the United Kingdom 84, 887–894.
Albretsen, J., Aure, J., Sætre, R., Danielssen, DS. 2012. Climatic variability in the Skagerrak and coastal waters of Norway. ICES J. Mar. Sci. 69, 758-763. doi: 10.1093/icesjms/fsr187
Alcaraz, M., Almeda, R., Calbet, A., Saiz, E., Duarte, CM., Lasternas, S., Agustí, S., Santiago, R., Movilla, JI., Alonso, A. 2010. The role of arctic zooplankton in biogeochemical cycles: respiration and excretion of ammonia and phosphate during summer. Polar Biology 33, 1719-173.
Anderson, M., Orvik, KA., LaCasce, JH., Koszalka, I., Mauritsen C. 2011. Variability of the Norwegian Atlantic Current and associated eddy field from surface drifters. J Geophys Res, Vol 16, C08032, doi: 10.1029/2011JC007078.
Anker-Nilssen, T., Bustnes, JO., Benjaminsen, S., Christensen-Dalsgaard, S., Dehnhard, N., Descamps, S., Erikstad, KE., Fauchald, P., Hanssen, SA., Langset, M., Lorentsen, SH., Lorentzen, E., Moe, B., Reiertsen, TK., Strøm, H., Systad, GH., Bruset, B., Sivertsen, K. 2020. Sjøfugl i Norge 2019. Trondheim: SEAPOP 2020.
Arndt, CE., Gulliksen, B., Lønne, OJ. Berge, J. 2009. Ice fauna. S. 303-322. I: Sakshaug, E., Johnsen, G., Kovacs, K. (Red.) Ecosystem Barents Sea. Tapir Academic Press, Trondheim, Norway.
Arndt, CE., Pavlova, O. 2005. Origin and fate of ice fauna in the Fram Strait and Svalbard area. Mar. Ecol. Prog. Ser. 301, 55-66.
Arndt, CE., Swadling, KM. 2006. Crustacea in Arctic and Antarctic sea ice: distribution, diet and life history strategies. Adv. Mar. Biol. 51, 197-315.
Bagøien, E., Kaartvedt, S., Aksnes, DL., Eiane, K. 2001. Vertical distribution and mortality of overwintering Calanus. Limnology and Oceanography 46, 1494-1510.
Bagøien, E., Melle, W., Kaartvedt, S. 2012. Seasonal development of mixed layer depths, nutrients, chlorophyll and Calanus finmarchicus in the Norwegian Sea – A basin-scale habitat comparison. Progress in Oceanography 103: 58-79.
Bakken, T., Kongsrud, JA., Oug, E., Cochrane, SKJ., Moen, TL., Solbakken, BEB. 2010. Polychaetes from Jan Mayen (Annelida, Polychaeta). Polar Research 29, 1-21. https://doi.org/10.1111/j.1751-8369.2009.00132.x
Barber, DG., Hop, H., Mundy, CJ., Else, B., Dmitrenko, IA., Tremblay, J-E., Ehn, JK., Assmy, P., Daase, M., LM., Rysgaard, S. 2015. Selected physical, biological and biogeochemical implications of a rapidly changing Arctic Marginal Sea Ice Zone. Progress in Oceanography 139, 122-150.
Basedow, SL., Sundfjord, A., von Appen, W-J., Halvorsen, E., Kwasniewski, S., Reigstad, M. 2018. Seasonal variation in transport of zooplankton into the Arctic Basin through the Atlantic gateway, Fram Strait. Frontiers in Marine science 5:194. doi: 10.3389/fmars.2018.00194
Beaugrand, G., Reid, PC., Ibañez, F., Lindley, JA., Edwards, M. 2002. Reorganization of North Atlantic Marine Copepod Biodiversity and Climate. Science 296: 1692-1694
Berge, J., Varpe, Ø., Moline, MA., Wold, A., Renaud, PE., Daase, M., Falk-Petersen, S. 2012. Retention of ice-associated amphipods: possible consequences for an ice-free Arctic Ocean. Biology Letters 8, 1012-1015. doi:10.1098/rsbl.2012.0517.
Bergstad, OA., Bjelland, O., Gordon, JDM. 1999. Fish communities on the slope of the eastern. Norwegian Sea. Sarsia 84:67-78.
Bjelland, O., Bergstad, OA., Skjæraasen, JE., Meland, K. 2000. Trophic ecology of deep-water fishes associated with the continental slope of the eastern Norwegian Sea. Sarsia 85:101-117.
Blachowiak-Samolyk, K., Kwasniewski, S., Dmoch, K., Hop, H., Falk-Petersen, S. 2007. Trophic structure of zooplankton in the Fram Strait in spring and autumn 2003. Deep Sea Research 54: 2716-2728.
Blindheim, J., Østerhus, S. 2005. The Nordic seas, main oceanographic features. I: Drange, H., Dokken, T., Furevik, T., Gerdes, R., Berger, W. (Red.) The Nordic Seas: An Integrated Perspective, Geophys. Monogr. Ser., vol. 158, s. 11–37, American Geophysical Union, Washington, D. C.
Bluhm, BA., Gebruk, AV., Gradinger, R., Hopcroft, RR., Huettmann, F., Kosobokova, KN., Sirenko, BI., Weslawski, JM. 2011. Arctic marine biodiversity: an update of species richness and examples of biodiversity change. Oceanography 24(3), s. 232-248.
Bluhm, BA., Hop, H., Melnikov IA., Poulin, M., Vihtakari, M., Collins, E. Gradinger, R., Juul-Pedersen, T., von Quillfeldt, C. 2017b. Sea-ice biota. S. 33-62 in CAFF. 2017b. State of the Arctic Marine Biodiversity Report. Conservation of Arctic Flora and Fauna International Secretariat, Akureyri, Iceland. 978-9935-431-63-9.
Bluhm, BA., Hop, H., Vihtakari, M., Gradinger, R., Iken, K., Melnikov, IA., Søreide, J.E. 2018. Sea ice meiofauna distribution on local to pan‐Arctic scales. Ecology and evolution 8(4), s.2350-2364.
Bluhm, BA., Swadling, K., Gradinger, R. 2017a. Chapter 16: Sea ice as habitat for macrograzers. S. 394-414 in Thomas D.N. (Red.). Sea ice 3rd edition, Wiley Blackwell.
Boetius, A. 2016. Hot and Cold at the Gakkel Ridge of the Central Arctic Basin. PS101 Weekly report no 3. 26 September-2 October 2016. Alfred-Wegener-Institut.
Boetius, A., Albrect, S., Bakker, K., Bienhold, C., Felden, J., Fernandez-Mendez, M., Hendricks, S., Katlein, C., Lalande, C., Krumpen, T. mfl. 2013. Export of algal biomass from the melting Arctic sea ice. Science 126: 1430-1432.
Bosse, A., Fer, I., 2019. Mean structure and seasonality of the Norwegian Atlantic Front Current along the Mohn Ridge from repeated glider transects. Geophysical Research Letters 46(22), 13170-13179.
Brix, S., Kaiser, S., Lörz, AN., LeSaout, M., Schumacher, M., Bonk, F., Egilsdottír, H., Ólafsdóttir, S., Tandberg, AHS., Taylor, J. mfl. Sendt inn 04-2021. Habitat variability and faunal zonation at the Ægir Ridge, a canyon-like structure in the deep Norwegian Sea. PeerJ special Issue WCMB2020 .
Brix, S., Taylor, J., LeSaout, M., Mercado-Salas, NF., Kaiser, S., Lörz, A-N. mfl. Sendt inn. 01-2021. Depth transect and connectivity along gradients in the North Atlantic and Nordic Seas in the frame of the IceAGE project (Icelandic marine Animals: Genetics and Ecology). Cruise No. SO276 (MerMet17-06). 22.06.2020-26.07.2020, Emden (Germany) - Emden (Germany). IceAGE3. Senckenberg am Meer Cruise Report 2020.
Broms, 2007. Distribution and life cycle of Calanus species in relation to the seasonal phytoplankton development and predator-prey interactions with Norwegian spring spawning herring in the physical setting of the Norwegian Sea ecosystem. Dissertation for the degree philosophiae doctor (Ph.D.). University of Oslo.
Broms, C., Melle, W. 2007. Seasonal development of Calanus finmarchicus in relation to phytoplankton bloom dynamics in the Norwegian Sea. Deep-Sea Research II 54:2760-2775.
Broms, C., Melle, W., Kaartvedt, S. 2009. Oceanic distribution and life cycle of Calanus species in the Norwegian Sea and adjacent waters. Deep Sea Research Part II: Topical Studies in Oceanography 56, 1910-1921.
Burgos, J., Buhl-Mortensen, L., Buhl-Mortensen, P., Ólafsdóttir, SH., Steingrund, P., Ragnarsson, SÁ., Skagseth, Ø. 2020. Predicting the distribution of Indicator Taxa of Vulnerable Marine Ecosystems in the Arctic and Sub-arctic Waters of the Nordic Seas. Frontiers in Marine Science 7:131 https://doi.org/10.3389/fmars.2020.00131
Bustnes, JO. 2010. Studies of lesser black-backed gulls in northern Norway in 2009. SEAPOP Short Report 10-2010.
Børsheim, KY. 2017. Bacterial and primary production in the Greenland Sea. Journal of Marine Systems 176, 54-63.
Cadrin, SX., Bernreuther, M., Daníelsdóttir, AK., Hjörleifsson, E., Johansen, T., Kerr, L., Kristinsson, K., Mariani, S., Nedreaas, K., Pampoulie, C. mfl. 2010. Population structure of beaked redfish, Sebastes mentella : evidence of divergence associated with different habitats. – ICES Journal of Marine Science 67, 1617–1630.
Carmack, E., Polyakov, I., Padman, L., Fer, I., Hunke, E., Hutchings, J., Melling, H. 2015. Toward quantifying the increasing role of oceanic heat in sea ice loss in the new Arctic. Bulletin of the American Meteorological Society , 96 (12), 2079-2105.
Cathalot, C, Van Oevelen, D., Cox, TJS., Kutti, T., Lavaleye, M., Duineveld, G. and Meysman, FJR. 2015. Cold-water coral reefs and adjacent sponge grounds: hotspots of benthic respiration and organic carbon cycling in the deep sea. Front. Mar. Sci. 2:37. https://doi.org/10.3389/fmars.2015.00037
Cherkasheva, A., Bracher, A., Melsheimer, C., Koberle, C., Gerdes, R., Nothig, E.M., Bauerfeind, E., Boetius, A. 2014. Influence of the physical environment on polar phytoplankton blooms: A case study in the Fram Strait. Journal of Marine Systems 132, 196-207.
Christensen-Dalsgaard, S., Lorentsen, S.-H., Dahl, E.L., Follestad, A., Hanssen, F., Systad, G.H. 2010. Offshore vindenergianlegg – sjøfugl, havørn, hubro og vadere. NINA Rapport 557. Norsk institutt for naturforskning.
Christensen-Dalsgaard, S., Mattisson, J., Norderhaug, KM., Lorentsen, S-H. 2020. Sharing the neighbourhood: assessing the impact of kelp harvest on foraging behaviour of the European shag. Mar Biol 167, 136. https://doi.org/10.1007/s00227-020-03739-1
Christensen-Dalsgaard, S., May, RF., Lorentsen, S-H. 2018. Taking a trip to the shelf: Behavioural decisions are mediated by the proximity to foraging habitats in the black-legged kittiwake. Ecology and Evolution 8: 866-878. https://doi.org/10.1002/ece3.3700 .
Christie, H., Norderhaug KM., Fredriksen, S. 2009. Macrophytes as habitat for fauna. Marine Ecology Progress Series 396, 221–233.
Connelly, DP., German, CR., Asada, M., Okino, K., Egorov, A., Naganuma, T., Pimenov, M., Cherkasev, G., Tamaki, K. 2007. Hydrothermal activity on the ultra-slow spreading southern Knipovich Ridge. Geochem. Geophys. Geosyst. G3, 8, Q08013.
Cushing, DH. 1990. Plankton production and year-class strength in fish populations: an update of the match/mismatch hypothesis. Advances in marine biology 26, 249-293.
Dale, T., Bagøien, E., Melle, W., Kaartvedt, S. 1999. Can predator avoidance explain varying overwintering depth of Calanus in different oceanic water masses? Mar Ecol Prog Ser 179,113- 121.
Dale, T., Rey, F., Heimdal, BR. 1999. Seasonal development of phytoplankton at a high-latitude oceanic site. Sarsia 84. 419-435.
Dalpadado, P., Ellertsen, B., Melle, W., Skjoldal, HR. 1998. Summer distribution patterns and biomass estimates of microzooplankton and micronekton in the Nordic Seas. Sarsia 83:2, 103-116.
de Boer, MN., Janinhoff, N., Nijs, G., Verdaat, H. 2019. Encouraging encounters: unusual aggregations of bowhead whales Balaena mysticetus in the western Fram Strait. Endang Species Res 39,51-62. https://doi.org/10.3354/esr00948
Dragesund, O., Hamre, J., Ulltang, Ø. 1980. Biology and population dynamics of the Norwegian spring-spawning herring. Rapp P-v Réun Cons int Explor Mer 177, 43-71.
Dupont, N., Bagøien, E., Melle, W. 2017. Interannual variability in spring abundance of adult Calanus finmarchicus from the overwintering population in the southeastern Norwegian Sea. Progress in Oceanography 152, 75-85.
Eamer, J., Donaldson, GM., Gaston, AJ., Kosobokova, KN., Larusson, KF., Melnikov, IA., Reist, JD., Richardson, E., Staples, L., von Quillfeldt, CH. 2013. Life Linked to Ice: A guide to sea-ice associated biodiversity in this time of rapid change. CAFF Assessment Series No. 10. Conservation of Arctic Flora and Fauna, Iceland.
Ehrlich, J., Schaafsma, FL., Bluhm, BA., Peeken, I., Castellani, G., Brandt, A., Flores, H. 2020. Sympagic Fauna in and Under Arctic Pack Ice in the Annual Sea-Ice System of the New Arctic. Frontiers in Marine Science, 7:452. https://doi.org/10.3389/fmars.2020.00452
Eilertsen, MH., Georgieva, MN., Kongsrud, JA., Wiklund, H., Glover, AG., Rapp HT. 2018. Genetic connectivity from the Arctic to the Antarctic: Sclerolinum contortum and Nicomache lokii (Annelida) are both widespread in reducing environments. Sci. Rep. 8:4810. https://doi.org/10.1038/s41598-018-23076-0
Eliassen, JE. 1983. Occurrence, gonad development and age distribution of the roughhead grenadier (Macrocurus berglax Lacepde) (Gadiformes, Macrouridae) along the Continental slope of northern Norway. ICES CM 1983/G:41, 19 s.
Erga, SR., Ssebiyonga, N., Hamre, B., Frette, Ø., Hovland, E., Hancke, K., Drinkwater, K., Rey, F. 2014. Environmental control of phytoplankton distribution and photosynthetic performance at the Jan Mayen Front in the Norwegian Sea. Journal of Marine systems 130, 193-205.
Etnoyer, P. 2010. Deep-sea coral on seamounts. Oceanography 23, 128-129.
Fang, JKH., Rooks, CA., Krogness, CM., Kutti, T., Hoffmann, F., Bannister, RJ. 2018. Impact of particulate sediment, bentonite and barite (oil-drilling waste) on net fluxes of oxygen and nitrogen in Arctic-boreal sponges. Environ. Pollut. 238, 948-958. https://doi.org/10.1016/j.envpol.2017.11.092
Fernández-Méndez, M., Katlein, C., Rabe, B., Nicolaus, M., Peeken, I., Bakker, K., Flores, H., Boetius, A. 2015. Photosynthetic production in the central Arctic Ocean during the record sea-ice minimum in 2012. Biogeosciences 12(11), 3525-3549.
Fernández-Méndez, M., Olsen, LM., Kauko, HM., Meyer, A., Rösel, A., Merkouriadi, I., Mundy, CJ., Ehn, JK., Johansson, AM., Wagner, PM., Ervik Å. 2018. Algal hot spots in a changing Arctic Ocean: Sea-ice ridges and the snow-ice interface. Frontiers in Marine Science 5:75.
Folkow, LP., Blix, A.S. 1999. Diving behaviour of hooded seals (Cystophora cristata) in the Greenland and Norwegian Seas. Polar Biology 22, 61-74.
Folkow, LP., Mårtensson, P-E., Blix, AS. 1996. Annual distribution of hooded seals (Cystophora cristat a) in the Greenland and Norwegian Seas. Polar Biol. 16, 179-189.
Folkow, LP., Nordøy, ES., Blix, AS. 2010. Remarkable development of diving performance and migrations of hooded seals (Cystophora cristata) during their first year of life. Polar Biol. 33, 433-441.
Fossum, P., Mork, KA., Tverberg, V., Boitsov S., Heldal, HE., Bagøien, E., Rønning, J., Slotte, A., Vollen, T., Wienerroither, R. mfl. 2012. Beskrivelse av miljø og levende marine ressurser i havområdene ved Jan Mayen. Rapport fra Havforskningen nr. 16-2012, 119 s.
Fosså, JH., Kutti, T., Buhl-Mortensen, P., Skjoldal, HR. 2015. Vurdering av norske korallrev. Rapport fra Havforskningen nr. 8–2015, 64 s.
Fosså, JH., Mortensen, PB., Furevik, DM. 2002. The deep-water coral Lophelia pertusa in Norwegian waters: Distribution and fishery impacts. Hydrobiologia 471, 1-12.
Fragoso, GM., Davies, EJ., Ellingsen, I., Chauton, MS., Fossum, T., mfl. 2019. Physical controls on phytoplankton size structure, photophysiology and suspended particles in a Norwegian biological hotspot. Progress in Oceanography, 175: 284-299
Fransner, F., Fröb, F., Tjiputra, T., Chierici, M., Fransson, A., Jeansson, E., Johannessen, T., Jones, E., Lauvset, SK., Ólafsdottír, S., Omar, A., Skjelvan, I., Olsen., A. 2020. Nordic Seas Acidification. Biogeosciences Discussions. Doi: 10.5194/bg-2020-339
Freiwald, A., Beuck L., Wisshak, M. 2012. Korallenriffe im kalten Wasser des Nordatlantiks – Entstehung, Artenvielfalt und Gefährdung. I: Beck, E. (red.) Die Vielfalt des Lebens. Wiley-VCH, Weinheim, pp 89-96.
Freiwald, A., Hühnerbach, V., Lindberg, B., Wilson, JB., Campbell, J. 2002. The Sula reef complex, Norwegian Shelf. Facies 47, 179-200.
Friedrich, C., De Smet, WH. 2000. The rotifer fauna of arctic sea ice from the Barents Sea, Laptev Sea and Greenland Sea. Hydrobiologia 432, 73-89.
Gabrielsen, GW., Brekke, B., Alsos, IG., Hansen, JR. 1997. Natur- og kulturmiljøet på Jan Mayen. Meddelser Nr. 144, Norsk Polarinstitutt.
Gaina, C., Blischke, A., Geissler, WH., Kimbell, GS., Erlendsson, Ö. 2017 Seamounts and oceanic igneous features in the NE Atlantic: a link between plate motions and mantle dynamics. Geological Society London Special Publications. 447, 419-442.
Gilg, O., Strøm, H., Aebischer, A., Gavrilo, MV., Volkov, AE., Miljeteig, C. and Sabard, B. 2010. Post-breeding movements of northeast Atlantic ivory gull Pagophila eburnea populations. J. Avian Biol. doi: 10.1111/j.1600-048X.2010.05125.x
Gradinger, R. 2020. Life in Arctic Sea Ice. I: Goldstein MI., DellaSala DA. (Red.) Encyclopedia of the World's Biomes, vol. 2. Elsevier, s. 507–514.
Gradinger, R., Bluhm, B., Iken, K. 2010. Arctic sea-ice ridges-Safe havens for sea-ice fauna during periods of extreme ice melt? Deep Sea Research Part II: Topical Studies in Oceanography 57 (1-2), 86-95.
Gradinger, R., Friedrich, C., Spindler, M. 1999. Abundance, biomass and composition of the sea-ice biota of the Greenland Sea pack ice. Deep Sea Research Part II: Topical Studies in Oceanography 46 (6-7), 1457-1472.
Gradinger, RR., Kaufman, MM., Bluhm, BA. 2009. Pivotal role of sea ice sediments in the seasonal development of near-shore Arctic fast ice biota. Marine Ecology 394, 49–63.
Gradinger, RR., Spindler, M., Henschel, D. 1991. Development of Arctic sea-ice organisms under graded snow cover. I: Sakshaug, E., Hopkins, CCC, Øritsland, NA. (Red.) Proceedings of the Pro Mare Symposium on Polar Marine Ecology, Trondheim, 12-16 May 1990. Polar Res. 10(1), 295-307.
Gulliksen, B., Beuchel, F., Brattegard, T., Palerud, R. 2004. The Marine Sublittoral Fauna of Jan Mayen Island. Zoogeography and Succession on “New” Lava Grounds. Kap 15 (p 159-171) I: Skreslet, S. (Red.) Jan Mayen Island in Scientific Reports. Kluwer Academic Publishers, The Netherlands. https://doi.org/10.1007/978-1-4020-2957-8_15
Halvorsen, E., Tande, KS., Edvardsen, A., Slagstad, D., Pedersen, OP. 2003. Habitat selection of overwintering Calanus finmarchicus in the NE Norwegian Sea and shelf waters off Northern Norway in 2000–02. Fisheries Oceanography 12(4-5), 339-351.
Hansen, A. 2020. Genetic population structure of the northern shrimp (Pandalus borealis) along the Norwegian coast. Masteroppgave. Norges arktiske universitet. Fakultet for biovitenskap, fiskeri og økonomi. 60 s.
Hansen, E., Ekeberg, OC., Gerland, S., Pavlova, O., Spreen, G., Tschudi, M. 2014. Variability in categories of Arctic sea ice in Fram Strait. Journal of Geophysical Research: Oceans 119, 7175-7189.
Hansen, E., Gerland, S., Granskog, MA., Pavlova, O., Renner, AHH., Haapala, J., Løyning, TB., Tschudi, M. 2013. Thinning of Arctic sea ice observed in Fram Strait: 1990-2011. Journal of Geophysical Research: Oceans 118, 5202–5221. ht tps://doi.org/10.1002/jgrc.20393
Hansen, E., Gerland, S., Høyland, KV., Pavlova, O., Spreen, G. 2015. Time variability in the annual cycle of sea ice thickness in the Transpolar Drift. Journal of Geophysical Research: Oceans 120, 8135–8150. https://doi.org/10.1002/2015JC011102
Hanssen-Bauer, I., Førland, EJ., Haddeland, I., Hisdal, H., Mayer, S., Nesje, A., Nilsen, JEØ., Sandven, S., Sandø, AB., Sorteberg. A., Ådlandsvik, B. 2015. Klima i Norge 2100 Kunnskapsgrunnlag for klimatilpasning oppdatert i 2015. NCCS report, NCCS, Oslo, Norway, 203 s.
Hareide, N-R. 1995. Comparisons between longlining and trawling for deep-water species – selectivity, quality and catchability – A review. p. 227-234 I: Hopper, AG. (Red.) Deep-Water Fisheries of the North Atlantic Oceanic Slope. NATO ASI Series (Series E: Applied Sciences), vol 296. Springer, Dordrecht.
Hareide, N-R., Garnes, G. 2001. The distribution and catch rates of deep water fish along the Mid-Atlantic Ridge from 43 to 61°N. Fisheries Research 51, 297–310.
Haug, T., Biuw, M. 2019. Ishavssel: fangst, bestandssituasjon og forskning. Notat til Forskerutvalg om sjøpattedyr, Tromsø, 24.-25. oktober 2019.
Heath, MR., Fraser, JG., Gislason, A., Hay, SJ., Jonasdottir, SH., Richardson, K. 2000. Winter distribution of Calanus finmarchicus in the Northeast Atlantic, ICES Journal of Marine Science 57, 1628- mic Publishers, 420p.
Hegseth E.N. 1998. Primary production in the northern Barents Sea. Polar Research 17, 113-123.
Heide-Jørgensen, MP., Hjort Nielsen, N., Hansen, RG., Schmidt, HC., Blackwell, SB. Jørgensen, OA. 2015. The predictable narwhal: Satellite tracking shows behavioural similarities between isolated subpopulations. Journal of Zoology 297 (1). DOI: 10.1111/jzo.12257
Helland-Hansen, B., Nansen, F. 1909. The Norwegian Sea - its physical oceanography based upon the Norwegian researches 1900-1904. Det Mallingske Bogtrykkeri.
Henry, LA., Stehmann, MFW., De Clippele, L., Findlay, HS., Golding, N., Roberts, MJ. 2016. Seamount egg-laying grounds of the deep-water skate Bathyraja richardsoni. Journal of Fish Biology 89, 1473-1481.
Henson, SA., Dunne, PD, Sarmiento, JL. 2009. Decadal variability in North Atlantic phytoplankton blooms. Journal of Geophysical Research: Oceans Vol 114. https://doi.org/10.1029/2008JC005139
Hirche, HJ. 1991. Distribution of dominant calanoid copepod species in the Greenland Sea during late fall. Polar Biology 11, 351-362.
Hjøllo, SS., Huse, G., Skogen, MD., Melle, W. 2012. Modeling secondary production in the Norwegian Sea with a fully coupled physical/primary production/individual-based Calanus finmarchicus model system. Marine Biology Research 8, 508-526.
Hoffmann, R., Braeckman, U., Hasemann, C., Wenzhöfer, F. 2018. Deep-sea benthic communities and oxygen fluxes in the Arctic Fram Strait controlled by sea-ice cover and water depth, Biogeosciences 15, 4849–4869. https://doi.org/10.5194/bg-15-4849-2018
Holst, JC., Røttingen, I., Melle, W. 2004. 7. The Herring. I: Skjoldal, HR. (Red.) The Norwegian Sea Ecosystem, Tapir Academic Press, Trondheim, s. 203-226.
Hop, H., Bluhm, BA., Daase, M., Gradinger, R., Poulin, M. 2013. Arctic sea ice biota [in Arctic Report Card 2013], https://arctic.noaa.gov/Report-Card
Hop, H., Falk-Petersen, S., Svendsen, H., Kwasniewski, S., Pavlov, V., Pavlova, O., Soreide, JE. 2006. Physical and biological characteristics of the pelagic system across Fram Strait to Kongsfjorden. Progress in Oceanography 71(2-4), 182-231.
Hop, H., Vihtakari, M., Bluhm, BA., Assmy, P., Poulin, M., Gradinger, R., Peeken, I., von Quillfeldt, CH., Olsen, LM., Zhitina, L., Melnikov, IA. 2020. Changes in sea-ice protist diversity with declining sea ice in the Arctic Ocean from the 1980s to 2010s. Frontiers Marine Science, doi: 10.3389/mars2020.00243.
Hopkins, TL. 1969. Zooplankton Biomass Related to Hydrography along the Drift Track of Arlis II in the Arctic Basin and the East Greenland Basin. Journal of the Fisheries Research Board of Canada 26 (2), 305-310.
Horner, R., Ackley, SF., Dieckmann, GS., Gulliksen, B., Hoshiai, T., Melnikov, IA., Reeburgh, WS., Spindler, M., Sullivan, CW. 1992. Ecology of Sea Ice Biota. 1. Habitat and terminology. Polar Biology 12, 417–427.
Høines, ÅS., Bergstad, OA. 1999. Resource sharing among cod, haddock, saithe and pollack on a herring spawning ground. Journal of Fish Biology, 55, 1233–1257.
ICES 2019 a. Report of the Working Group on Integrated Ecosystem Assessments for the Norwegian Sea (WGINOR). ICES WGINOR REPORT 2018 26-30 November 2018. Reykjavik, Iceland. ICES CM 2018/IEASG:10. 123 pp
ICES 2020 a. Working Group on the Integrated Assessments of the Norwegian Sea (WGINOR; outputs from 2019 meeting). ICES Scientific Reports. 2:29. 46 s. http://doi.org/10.17895/ices.pub . 5996
ICES 2020 b. Herring (Clupea harengus) in subareas 1, 2, and 5, and in divisions 4.a and 14.a, Norwegian spring-spawning herring (the Northeast Atlantic and the Arctic Ocean). I: Report of the ICES Advisory Committee, 2020. ICES Advice 2020. https://doi.org/10.17895/ices.advice.5876
ICES 2020 c. ICES Working Group on the Assessments of Demersal Stocks in the North Sea and Skagerrak (WGNSSK). ICES Scientific Reports. 2:61. 1249 pp. http://doi.org/10.17895/ices.pub.6092
IPCC, 2019: IPCC Special Report on the Ocean and Cryosphere in a Changing Climate [H.-O. Pörtner, D.C. Kvingedal, B. 2005. Sea-Ice Extent and Variability in the Nordic Seas, 1967-2002. I: Drange, H., Dokken, T., Furevik, T., Gerdes, R., Berger, W. (Red.) b. 158 The Nordic Seas: An Integrated Perspective, Oceanography, Climatology, Biogeochemistry, and Modeling, s. 39-50, American Geophysical Union, Washington DC, USA.
Johnson, JE., Mienert, J., Plaza-Faverola, A., Vadakkepuliyambatta, S., Knies, J., Bünz, S., Andreassen, K., Ferré, B. 2015. Abiotic methane from ultraslow-spreading ridges can charge Arctic gas hydrates. Geology 43 (5), 371–374. https://doi.org/10.1130/G36440.1
Joiris, CR., Falck, E., D'Hert, D., Jungblut, S., Boos, K. 2014. An important late summer aggregation of fin whales Balaenoptera physalus , little auks Alle alle and Brünnich’s guillemots Uria lomvia in the eastern Greenland Sea and Fram Strait: influence of hydrographic structures. Polar Biology 37, 1645-1657. https://doi.org/10.1007/s00300-014-1551-5
Jones, E., Chierici, M., Skjelvan, I., Norli, M., Frigstad, H., Børsheim, KY., Lødemel, HH., Kutti, T., King, AL., Sørensen, K., Lauvset SK., Jackson-Misje, K., Apelthun, LB., de Lange, T., Johannessen, T., Mourgues, C., Bellerby, R. 2020. Monitoring ocean acidification in Norwegian Seas 2019. Rapport, Miljødirektoratet, M-1735.
Jorde, PE., Søvik, G., Westgaard, J-I., Albretsen, J., André, C., Hvingel, C., Johansen, T., Sandvik, AD., Kingsley, M., Jørstad, KE. 2015. Genetically distinct populations of northern shrimp, Pandalus borealis , in the North Atlantic: adaptation to different temperatures as an isolation factor. Molecular Ecology 24, 1742-1757.
Kauko, HM., Olsen, LM., Duarte, P., Peeken, I., Granskog, MA., Johnsen, G., Fernández-Méndez, M., Pavlov, AK., Mundy, CJ., Assmy, P. 2018. Algal colonization of young Arctic Sea ice in spring. Frontiers in Marine Science 5, 199.
Knutsen, H., Jorde, PE., Gonzalez, EB., Eigaard, OR., Pereyra, RT., Sannæs, H., Dahl, M., André, C., Søvik, G. 2015. Does population genetic structure support present management regulations of the northern shrimp (Pandalus borealis) in Skagerrak and the North Sea? ICES Journal of Marine Science 72(3), 863-871.
Kohlbach, D., Graeve. M., Lange, BA., David, C., Peeken, I., Flores, H. 2016. The importance of ice algae‐produced carbon in the central Arctic Ocean ecosystem: Food web relationships revealed by lipid and stable isotope analyses. Limnology and Oceanography 61(6), 2027-2044.
Kohlbach, D., Schaafsma, FL., Graeve, M., Lebreton B., Lange BA., David C., Vortkamp M., Flores H., 2017. Strong linkage of polar cod (Boreogadus saida) to sea ice algae-produced carbon: evidence from stomach content, fatty acid and stable isotope analyses. Progress in Oceanography 152, 62-74.
Kongsrud, JA., Eilertsen, MH., Alvestad, T., Kongshavn, K., Rapp, HT. 2017. New species of Ampharetidae (Annelida: Polychaeta) from the Arctic Loki Castle vent field. Deep-Sea Research II 137, 232-245. https://doi.org/10.1016/j.dsr2.2016.08.015
Kongsrud, JA., Rapp, HT. 2012. Nicomache (Loxochona) lokii arter nov. (Annelida: Polychaeta: Maldanidae) from the Loki’s Castle vent field: an important structure builder in an Arctic vent system. Polar Biology 35, 161-170. https://doi.org/10.1007/s00300-011-1048-4
Krisch, S., Browning, TJ., Graeve, M., Ludwichowski, KU., Lodeiro, P., Hopwood, MJ., Roig, S., Young, J-C., Kanzow, T, Achterberg, EP. 2020. The influence of Arctic Fe and Atlantic fixed N on summertime primary production in Fram Strait, North Greenland Sea. Scientific Reports 10, 15230.
Kristiansen, I., Hátún, H., Petursdottir, H., Gislason, A., Broms, C., Melle, W., Jacobsen, JA., Eliasen, SK., Gaard E. 2019. Decreasing influx of Calanus spp. Into the south-western Norwegian Sea since 2003. Deep Sea Research 149, 103048.
Krumpen, T., Belter, HJ., Boetius, A., Haas, C., Hendricks, S., Nicolaus, M., Nöthig, E-M., Paul, S., Peeken, I., Ricker, R., Stein, R. 2019. Arctic warming interrupts the Transpolar Drift and affects long-range transport of sea ice and ice-rafted matter. Scientific Reports 9, 5459. doi: 10.1038/s41598-019-41456-y.
Krumpen, T., Gerdes, R., Haas, C., Hendricks, S., Herber, A., Selyuzhenok, L., Smedsrud, L., Spreen, G. 2016. Recent summer sea ice thickness surveys in Fram Strait and associated ice volume fluxes. The Cryosphere, 10, 523-534.
Kunisch, EH., Bluhm, BA., Daase, M., Gradinger, R., Hop, H., Melnikov, IA., Varpe, Ø., Berge, J. 2020. Pelagic occurrences of the ice amphipod Apherusa glacialis throughout the Arctic. Journal of plankton research, 42(1), 73-86.
Kutti, T., Bannister, RJ., Fossa, JH. 2013. Community structure and ecological function of deep-water sponge grounds in the Traenadypet MPA-Northern Norwegian continental shelf. Cont. Shelf Res. 69, 21–30. https://doi.org/10.1016/j.csr.2013.09.011
Kutti, T., Bannister, RJ., Fosså, JH., Krogness, CM., Tjensvoll, I., Søvik, G., 2015. Metabolic responses of the deep-water sponge Geodia barretti to suspended tailings, drill cuttings and bottom sediment. Journal of Experimental Marine Biology and Ecology 473, 64-72.
Kvingedal, B. 2005. Sea-Ice Extent and Variability in the Nordic Seas, 1967-2002. I: Drange, H., Dokken, T., Furevik, T., Gerdes, R., Berger, W. (Red.) b. 158 The Nordic Seas: An Integrated Perspective, Oceanography, Climatology, Biogeochemistry, and Modeling, s. 39-50, American Geophysical Union, Washington DC, USA.
Kwok, R. 2009. Outflow of Arctic Ocean Sea ice into the Greenland and Barents Seas: 1979–2007. Journal of Climate 22, 2438–2457. https://doi.org/10.1175/2008JCLI2819.1
Kwok, R., Spreen, G, Pang, S. 2013. Arctic sea ice circulation and drift speed: Decadal trends and ocean currents. Journal of Geophysical Research: Oceans, 118, 2408–2425. https://doi.org/10.1002/jgrc.20191
Lacharité, M., Metaxas, A. 2013. Early Life History of Deep-Water Gorgonian Corals May Limit Their Abundance. PLoS ONE 8(6): e65394.
Langehaug, H., Falck, E. 2012. Changes in the properties and distribution of the intermediate and deep waters in the Fram Strait. Progress in Oceanography 96: 56-76. https://doi.org/10.1016/j.pocean.2011.10.002
Lauvset, SK., Key, RM., Olsen, A., van Heuven, S., Velo, A., Lin, X., Schirnick, C., Kozyr, A., Tanhua, T., Hoppema, M. mfl. 2016. A new global interior ocean mapped climatology: the 1 o x 1 o GLODAP version 2. Earth System Science Data 8, 325-340.
Leonard, DM., Øien, NI. 2020. Estimated abundance of cetacean species in the Northeast Atlantic from Norwegian shipboard surveys conducted in 2014-2018. NAMMCO Scientific Publications 11. https://doi.org/10.7557/3.4694 .
Leu, E., Mundy, CJ., Assmy, P., Campbell, K., Gabrielsen, TM., Gosselin, M., Juul-Pedersen, T., Gradinger, R. 2015. Arctic spring awakening - Steering principles behind the phenology of vernal ice algal blooms. Progress in Oceanography 139, 151-170.
Leu, E., Søreide, JE., Hessen, DO., Falk-Petersen, S., Berge, J. 2011. Consequences of changing sea-ice cover for primary and secondary producers in the European Arctic shelf seas: Timing, quantity, and quality. Progress in Oceanography 90(1-4), 18-32.
Loeng, H., Vinje, T. 1979. On the sea ice conditions in the Greenland and Barents Seas. p.163-176, I: (ANON, Red.) POAC 79 . Proceedings of the fifth international conference on Port and Ocean Engineering under Arctic Conditions. The Norwegian Institute of Technology, Trondheim 1979.
Lorentsen, S-H. (Red.), Christensen-Dalsgaard, S., Follestad, A., Langset, M., May, R., Dahl, EL., Hamre, Ø. 2012. Fagrapport til strategisk konsekvensutredning av fornybar energiproduksjon til havs - sjøfugl. NINA Rapport 825. Norsk institutt for naturforskning.
Louis, M., Skovrind M., Samaniego Castruita, JA., Garilao, C., Kaschner, K., Gopalakrishnan, S., Haile, JS., Lydersen, Ch., Kovacs, KM., Garde, E. mfl. 2020. Influence of past climatic change on phylogeography and demographic history of narwhals, Monodon monoceros . Proceedings of the Royal Society B. 287, 20192964. https://doi.org/10.1098/rspb.2019.2964
Lund-Hansen, LC., Haubjerg Søgaard, D., Sorrell, B., Gradinger, R., Meiners, KM. 2020. Arctic sea ice ecology - Seasonal Dynamics in Algal and Bacterial Productivity. Springer. Doi: https://doi.org/10.1007/978-3-030-37472-3
Løviknes, S., Jensen, KH., Krafft, BA., Anthonypillai, V., Nøttestad, L. 2021. Feeding hotspots and distribution of Fin and Humpback whales in the Norwegian Sea From 2013 to 2018. Frontiers in Marine Science, 22 February 2021. https://doi.org/10.3389/fmars.2021.632720
Maier-Reimer, E., Hasselmann, K. 1987. Transport and storage of CO2 in the ocean – an inorganic ocean circulation carbon cycle model. Climate Dynamics 2, 63-90.
Marques, APA., Roerdink, DL., Baumberger, T., de Ronde, CEJ., Ditchburn, RG., Denny, A., Thorseth, IH., Okland, I., Lilley, MD., Whotehouse, MJ., Pedersen, RB. 2020. The Seven Sisters Hydrothermal System: First Record of Shallow Hybrid Mineralization Hosted in Mafic Volcaniclasts on the Arctic Mid-Ocean Ridge. Minerals 10, 439. https://doi.org/10.3390/min10050439
Marstein, AC. 1997. Epiphytic algae on kelp stipes from Vega – an area with varying densities of sea urchins. Blyttia 3, 123-129.
Mayot, N., Matrai, PA., Arjona, A., Belanger, S., Marchese, C., Jaegler, T., Ardyna, M., Steele, M. 2020. Springtime export of Artic Sea ice influences phytoplankton production in the Greenland Sea. JGR Oceans, https://doi.org/10.1029/2019JC015799
McClain, CR., Lundsten, L., Barry, J., DeVogelaere, A. 2010. Assemblage structure, but not diversity or density, change with depth on a northeast Paci fi c seamount. Marine Ecology 31, 14-25.
Meld. St. 29 (2020-2021). Heilskapleg nasjonal plan for bevaring av viktige områder i marin natur. Tilråding fra Klima- og miljødepartementet, 9. april 2021, godkjend i statsråd same dag. (Regjeringen Solberg). https://www.regjeringen.no/no/dokumenter/meld.-st.-29-20192020/id2715224/
Melle, W., Ellertsen, B., Skjoldal, HR. 2004. Zooplankton: The link to higher trophic levels. I: Skjoldal HR (Red.), The Norwegian Sea Ecosystem. Tapir Academic Press, Trondheim. 137-202.
Melle, W., Runge, J., Head, E., Plourde, S., Castellani, C., Licandro, P., Pierson, J., Jonasdottir, S. Johnson, C., Broms, C. mfl. 2014. The North Atlantic Ocean as habitat for Calanus finmarchicus : Environmental factors and life history traits. Progress in Oceanography 129, 244-284. https://doi.org/10.1016/j.pocean.2014.04.026
Melnikov, IA., Kolosova, EG., Welch, HE., Zhitina, LS. 2002. Sea ice biological communities and nutrient dynamics in the Canada Basin of the Arctic Ocean. Deep-Sea Res. 49, 1623–1649.
Merkouriadi, I., Liston, GE., Graham, RM., Granskog, MA. 2020. Quantifying the potential for snow-ice formation in the Arctic Ocean. Geophysical Research Letters 47, e2019GL085020. https://doi.org/10.1029/2019GL085020
Meyer, HK., Roberts, EM., Rapp, HT., Davies, AJ. 2019. Spatial patterns of arctic sponge ground fauna and demersal fish are detectable in autonomous undervater (AUV) imagery. Deep Sea Research I 153, 103137 https://doi.org/10.1016/j.dsr.2019.103137
Misund, OA., Melle, W., Fernö, A. 1997. Migration behavior of Norwegian spring spawning herring when entering the cold front in the Norwegian Sea. Sarsia 82, 107-112.
Misund, OA., Vilhjálmsson, H., Jákupsstovu, SH., Røttingen, I., Belikov, S., Asthorsson, O., Blindheim, J., Jónsson, J., Krysov, A., Malmberg, SA., Sveinbjørnsson, S. 1998. Distribution, migration and abundance of Norwegian spring spawning herring in relation to the temperature and zooplankton biomass in the Norwegian Sea as recorded by coordinated surveys in Springand Summer 1996. Sarsia 83, 117-127.
Mork, KA., Skagseth, Ø. 2010. A quantitative description of the Norwegian Atlantic Current by combining altimetry and hydrography. Ocean Science 6, 901-911. doi:10.5194/os-6-901-2010.
Mortensen, PB., Buhl-Mortensen, L. 2005. Morphology and growth of the deep-water gorgonians Primnoa resedaeformis and Paragorgia arborea . Marine Biology 147, 775-788.
Mortensen, PB., Hovland, MT., Fosså, JH., Furevik, DM. 2001. Distribution, abundance and size of Lophelia pertusa coral reefs in mid-Norway in relation to seabed characteristics. Journal of the Marine Biological Association of the UK 81, 581-597.
Nansen, F. 1906. Northern Waters: Captain Roald Amundsen’s oceanographic observations in the Arctic Seas in 1901. Videnskabs-Selskapets Skrifter. I. Mathematisk-Naturvitenskabelig Klasse, No 3. Christiania, Dybwad.
Nikurashin, M., Ferrari, R. 2011. Global energy conversion rate from geostrophic flows into inernal lee waves in the deep ocean. Geophysical Research Letters 38, L08610. https://doi.org/10.1029/2011GL046576
Nilsen, JEO., Falck, E. 2006. Variations of mixed layer properties in the Norwegian Sea for the period 1948–1999. Prog. Oceanogr. 70, 58–90. https://doi.org/10.1016/j.pocean.2006.03.014
Nilssen, EM., Aschan, MM. 2009. Catch, survey and life-history data for shrimp (Pandalus borealis) off Jan Mayen. Deep-Sea Research II 56, 2023-2036.
Nilssen, KT., Bjørge, A. 2019. Status for kystsel – Anbefaling av jaktkvoter 2020. Pp 56-69 i A. Bjørge (Red.). Forskerutvalg om sjøpattedyr 2019 – Anbefalinger om forskning og forvaltning. Rapport fra Havforskningen Nr. 2020-19.
Nilssen, KT., Bjørge, A. Biuw, M. 2020. Status for kystsel. Anbefaling av jaktkvoter 2021. Rapport til Forskerutvalg om sjøpattedyr, Tromsø, 22.-23. oktober 2020.
Nilssen, KT., Lindstrøm, U., Westgaard, JI., Lindblom, L., Blencke, T-R. Haug, T. 2019. Diet and prey consumption of grey seals (Halichoerus grypus) in Norway. Marine Biology Research 15(2), 137-149. https://doi.org/10.1080/17451000.2019.1605182
Norderhaug, KM., Christie, H., Andersen, GS., Bekkby, T. 2012. Does the diversity of kelp forest macrofauna increase with wave exposure? Journal of Sea Research 69, 36–42.
Norderhaug, KM., Filbee-Dexter, K., Freitas, C., Birkely, SR., Christensen, L., Mellerud, I., Thormar, J., van Son, T., Moy, F., Vázquez Alonso, M., Steen, H. 2020. Ecosystem-level effects of large-scale disturbance in kelp forests. Marine Ecology Progress Series 656, 163-180. https://doi.org/10.3354/meps13426
Nöthig, E-M., Bracher, A., Engel, A, Metfies, K., Niehoff, B., Peeken, I., Bauerfeind, E., Cherkasheva, A., Gäbler-Schwarz, S., Hardge, K. mfl. 2015. Summertime plankton in Fram Strait – a compilation of long- and short-term observations. Polar Research 34, 23349. https://doi.org/10.3402/polar.v34.23349
Nöthig, EM, Ramondenc, S, Haas, A, Hehemann, L, Walter, A, Bracher, A, Lalande, C, Metfies, K, Peeken, I, Bauerfeind, E and Boetius, A 2020. Summertime Chlorophyll a and Particulate Organic Carbon Standing Stocks in Surface Waters of the Fram Strait and the Arctic Ocean (1991–2015). Frontiers in Marine Sciences 7, 350. doi: 10.3389/fmars.2020.00350
Nøttestad, L. Boge, E., Ferter, K. 2020. Comeback at Atlantic bluefin tuna (Thunnus thynnus) to Norwegian waters. Fisheries Research 231, 105689. https://doi.org/10.1016/j.fishres.2020.105689
Nøttestad, L. Krafft, B., Anthonypillai, V., Bemasconi, M., Langård, L. Mørk, H. L., Fernö, A. 2015. Recent changes in distribution and relative abundance of cetaceans in the Norwegian Sea and their relationship with potential prey. Frontiers in Ecology and Evolution 2, 83. doi: 10.3389/fevo.2014.00083.
Nø ttestad, L., Ólafsdóttir, AH., Anthonypillai, V. Homrum, E., Jansen, T., Wieland K. mfl. 2020. Cruise report from the International Ecosystem Summer Survey in the Nordic Seas (IESSNS) 1st July – 4th August 2020. Working Group Document to ICES Working group on Widely Distributed Stocks (WGWIDE, No. 3), ICES HQ, Copenhagen, Denmark, (digital meeting) 26. august – 1. september 2020. 55 s.
Nøttestad, L., Sivle, LD., Krafft, BA., Langård, L., Anthonypillai, V., Bernasconi, M., Langøy, H., Fernø, A. 2014. Prey selection of offshore killer whales Orcinus orca in the Northeast Atlantic in late summer: spatial associations with mackerel. Marine Ecology Progress Series 499, 275-283. DOI:10.3354/meps10638
Olsen, RE., Knutsen, T., Melle, W. 2006. Nye marine fórressurser (NYMAR). Internrapport, Havforskningsinstituttet, 74 s.
Olsen, LM., Laney, SR., Duarte, P., Kauko, HM., Fernández-Méndez, M., Mundy CJ., Rösel, A., Meyer, A., Itkin, P., Cohen, L. mfl. 2017. The seeding of ice algal blooms in Arctic pack ice: The multiyear ice seed repository hypothesis. J. Geophys. Res. Biogeosci. 122, 1529-1548. doi:10.1002/2016JG003668.
Olsen, A., Lange, N., Key, RM., Tanhua, T., Bittig, HC., Kozyr. A., Álvarez, M., Azetsu-Scott, K., Becker, S., Brown, PJ. mfl. 2020. An updated version of the global interior biogeochemical data product, GLODAPv2.2020. Earth System Science Data 12, 3653-3678. DOI: 10.5194/essd-12-3653-2020
Olsen, BR., Økland, IE., Thorseth, IH., Pedersen, RB., Rapp, HT. 2016. Environmental challenges related to offshore mining and gas hydrate extraction. Miljødirektoratets rapportserie M-532, 28 s.
Onarheim, IH., Eldevik T., Smedsrud LH., Stroeve JC. 2018. Seasonal and Regional Manifestation of Arctic sea ice loss. Journal of Climate, 31(12), 4917-4932.
Orvik, KA., Niiler, PP. 2002. Major pathways of the Atlantic water in the northern North Atlantic and Nordic Seas toward Arctic. Geophys. Res. Lett. 29(19), 1896. doi:10.1029/2002GL015002.
Paasche, E. 1960. Phytoplankton investigations in the Norwegian Sea in June, 1954, related to hydrography and compared with primary production data. Fiskeridirektoratets Skrifter, Serie Havundersøkelser 2, 77 s.
Paulsen, E. 2017. Community structure and biodiversity of the benthic megafauna at the inactive hydrothermal site Mohn's Treasure, on the Mohn's Ridge, Arctic Mid-Ocean Ridge (AMOR). M.Sc.-thesis, NTNU, Trondheim.
Pedersen. RB., Bjerkgård, T. 2016. Sea-floor massive sulphides in Arctic Waters. Chapter 5 in Mineral Resources in the Arctic. s. 210-215.
Pedersen, RB., Rapp, HT., Thorseth, IH., Lilley, MD., Barriga, FJAS., Baumberger, T., Flesland, K., Fonseca, R., Früh-Green, GL., Jørgensen, SL. 2010. Discovery of a black smoker vent field and vent fauna at the Arctic Mid-Ocean Ridge. Nature Communications 126, 1124. https://doi.org/10.1038/ncomms1124
Pedersen, RB., Thorseth, IH., Hellevang, B, Schultz, A., Taylor, P., Knudsen. HP., Steinsbu, BO. 2005. Two vent fields discovered at the ultraslow spreading Arctic ridge system. Eos Trans. American Geophysical Union 86(52), Fall Meet. Suppl. (2005).
Poulin, M., Daugbjerg, N., Gradinger, R., Ilyash, L., Ratkova, T., von Quillfeldt, CH. 2011. The pan-Arctic biodiversity of marine pelagic and sea-ice unicellular eukaryotes: A first-attempt assessment. Marine Biodiversity 41 (1), 13-28. https://doi.org/10.1007/s12526-010-0058-8
Ramirez-Llodra, E., Hilario, A., Paulsen, E., Costa, CV., Bakken, T., Johnsen, G., Rapp, HT. 2020. Benthic Communities on the Mohn’s Treasure Mound: Implications for Management of Seabed Mining on the Arctic Mid-Ocean Ridge. Frontiers in Marine Science 7, 490. https://doi,org/10.3389/fmars.2020.00490
Rampal, P., Weiss, J., Marsan, D. 2009. Positive trend in the mean speed and deformation rate of Arctic sea ice, 1979–2007. Journal of Geophysical Research: Oceans, 114, C05013. https://doi.org/10.1029/2008JC005066
Ramsfjell, E. 1960. Phytoplankton distribution in the Norwegian Sea in June, 1952 and 1953. Fiskeridirektoratets Skrifter, Serie Havundersøkelser 10, 39 s.
Reigstad, M., Carroll, J., Slagstad, D., Ellingsen, I., Wassmann, P. 2011. Intra-regional comparison of productivity, carbon flux and ecosystem composition within the northern Barents Sea. Progress in Oceanography 90(1-4), 33-46.
Renner, AH., Gerland, S., Haas, C., Spreen, G., Beckers, JF., Hansen E., Nicolaus, M., Goodwin, H. 2014. Evidence of Arctic sea ice thinning from direct observations. Geophysical Research Letters, 41(14), 5029-5036.
Rey, F. 2012. Declining silicate concentrations in the Norwegian and Barents Seas. ICES J. Mar. Sci. 69, 1-5.
Rey, F., Noji, TT., Miller, A. 2000. Seasonal phytoplankton development and new production in the central Greenland Sea. Sarsia 85, 329-344.
Rey, F. 2004. Phytoplankton: the grass of the sea. I: The Norwegian Sea Ecosystem, HR. Skjoldal (Red.). Tapir academic press. Trondheim.
Ricker, R., Girard-Ardhuin, F., Krumpen, T., Lique, CJTC. 2018. Satellite-derived sea ice export and its impact on Arctic ice mass balance. The Cryosphere 12(9), 3017-3032.
Roberts, EM., Mienis, F., Rapp, HT., Hanz, U., Meyer, HK., Davies, AJ. 2018. Oceanographic setting and short-timescale environmental variability at an Arctic seamount sponge ground. Deep-Sea Research Part I 138, 98-113.
Spreen, G., de Steur, L., Divine, D., Gerland, S., Hansen, E., Kwok, R. 2020. Arctic sea ice volume export through Fram Strait from 1992 to 2014. Journal of Geophysical Research: Oceans, 125, e2019JC016039. https://doi.org/10.1029/2019JC016039
Spreen, G., Kwok, R., Menemenlis, D. 2011. Trends in Arctic sea ice drift and role of wind forcing: 1992–2009. Geophysical Research Letters 38, L19501. https://doi.org/10.1029/2011GL048970
Sætre, R. (Red.) 2007. The Norwegian Coastal Current—Oceanography and Climate. Tapir Academic Press, Trondheim, 159 s.
Sætre, R., Toresen, R., Søiland, H., Fossum, P. 2002. The Norwegian spring-spawning herring-spawning, larval drift and larval retention. Sarsia 87(2), 167-178.
Salvanes, AG. 2004. Mesopelagic Fish. I: HR. Skjoldal (Red). The Norwegian Sea Ecosystem. Tapir Academic Press 2004, Trondheim, s. 301-314.
Sandvik, H., Barrett, R., Erikstad, KE., Myksvoll, MS., Vikebø, F., Yoccoz, NG., Anker-Nilssen, T., Lorentsen, SH., Reiertsen, TK., Skarðhamar, J., Skern-Mauritzen, M., Systad, GH.: «Modelled drift patterns of fish larvae link coastal morphology to seabird colony distribution». Nature Communications 2016. doi: 10.1038/ncomms11599
Savvatimsky, PI., 1985. Features of spawning concentrations of roughhead grenadier (Macrocurus berglax L. at the Norwegian coast. ICES CM 1985/G:6, 34 s.
Scanes, E., Kutti, T., Fang, JKH., Johnston, EL, Ross, PM. Bannister, RJ. 2018. Mine Waste and Acute Warming Induce Energetic Stress in the Deep-Sea Sponge Geodia atlantica and Coral Primnoa resedeaformis ; Results From a Mesocosm Study. Front. Mar. Sci. 5:129. https://doi.org/10.3389/fmars.2018.00129
Schander, C., Rapp, HT., Kongsrud, JA., Bakken, T. mfl. 2010. The fauna of hydrothermal vents on the Mohn Ridge (North Atlantic). Marine Biodiversity Research 6(2): 155-171. https://doi.org/10.1080/17451000903147450
Schlacher, T., Baco, AR., Rowden, AA., O'Hara, T., Clark, MR., Kelley, C., mfl. 2014. Seamount benthos in a cobalt-rich crust region of the central Pacific: conservation challenges for future seabed mining. Diversity and Distributions 20: 491-502.
Shumway, SE., Perkins, HC., Schick, DF., Stickney, AP. 1985. Synopsis of Biological Data on the Pink Shrimp, Pandalus borealis Krøyer, 1838, NOAA Technical Report NMFS 30, FAO Fisheries Synopsis No. 144. 65 pp.
Sims, DW. 2008. Sieving a living: a review of the biology, ecology and conservation status of the plankton‐feeding basking shark Cetorhinus maximus . Advances in marine biology, 54, s.171-220.
Sivertsen, A. 2020. Success in seeking seabed assets. Norwegian Continental Shelf 1: 18-20.
Skagseth, Ø., Slotte, . Stenevik, EK., Nash, R. 2015. Characteristics of the Norwegian Coastal Current during years with high recruitment of Norwegian Spring Spawning Herring (Cluepa harengus L). PLoS ONE 10 (12). e0144117 DOI: 10.1371/journal.pone.0144117
Skagseth, Ø., Mork, KA. 2012. Heat content in the Norwegian Sea, 1995-2010. ICES J. Mar. Sci., 69, 826-832, doi: 10.1093/icesjms/fsss026.
Skaret, G., Dalpadado, P., Hjøllo, SS., Skogen, MD., Strand, E. 2014. Calanus finmarchicus abundance, production and population dynamics in the Barents Sea in a future climate. Progress in Oceanography, 125: 26-39.
Skjelvan, I., E. Jeansson, M. Chierici, A. Omar, A. Olsen, S. Lauvset, T. Johannessen. 2014. Ocean acidification and uptake of anthropogenic carbon in the Nordic Seas, 1981-2013, Report (på norsk), Norwegian Environment Agency, M244-2014.
Skjelvan, I., Falck, E., Rey, F., Kringstad, S. B. (2008). Inorganic carbon time series at Ocean Weather Station M in the Norwegian Sea. Biogeosciences, 5 (2), 549-560.
Skjoldal, HR.., Dalpadado, P., Dommasnes, A. 2004. 17. Food webs and trophic interactions. I: Skjoldal HR (Red) The Norwegian Sea ecosystem. Tapir Academic Press, Trondheim 2004, s 447-506.
Slagstad, D., Tande, K. 1996. The importance of seasonal vertical migration in across shelf transport of Calanus finmarchicus . Ophelia 44: 189-205.
Smedsrud L.H., Halvorsen M.H., Stroeve J.C., Zhang R., Kloster K.J.T.C. 2017. Fram Strait sea ice export variability and September Arctic sea ice extent over the last 80 years. 11(1), 65-79.
Soltwedel T., Bauerfeind, E., Bergamnn, M., Bracher., A., mfl. 2016. Natural variability or anthropogenically-induced variation? Insights from 15 years of multidisciplinary observations at the arctic marine LTER site HAUSGARTEN. Ecological Indicators 65, 89–102.
Søreide, JE., Carroll, ML., Hop, H., Ambrose Jr, WG., Hegseth, EN., Falk-Petersen, S. 2013. Sympagic-pelagic-benthic coupling in Arctic and Atlantic waters around Svalbard revealed by stable isotopic and fatty acid tracers. Marine Biology Research 9(9), 831-850.
Søreide, JE., Leu, E., Berge, J., Graeve, M., Falk-Petersen, S. 2010. Timing in blooms, algal food quality and Calanus glacialis reproduction and growth in a changing Arctic. Global Change Biology 16, 3154–3163.
Stafford KM, Moore SE, Berchok CL, Wiig Ø, Lydersen C, Hansen E, Kalmbach D, Kovacs KM. 2012. Spitsbergen’s endangered bowhead whales sing through the polar night. Endang Spec Res 18: 95-103. doi: 10.3354/esr00444
Stafford, KM, Lysersen, C., Wiig, Ø., Kovacs, K. 2018. Extreme diversity in the songs of Spitsberen’s bowhead whales. Biology Letters 14 (4). https://doi.org/10.1098/rsbl.2018.0056
Steen, IH., Dahle, H., Stokke, R., Roalkvam, I., Daae, L., Rapp, HT., Pedersen, RB., Thorseth, IH. 2016. Novel Barite Chimneys at the Loki’s Castle Vent Field Shed Light on Key Factors Shaping Microbial Communities and Functions in Hydrothermal Systems. Frontiers in Microbiology 6: 1510 https://doi.org/10.3389/micb.2015.01510
Stensland, A., Baumberger, T., Lilley, MD. mfl. 2019. Transport of carbon dioxice and heavy metals from hydrothermal vents to shallow water by hydrate-coated gas bubbles. Chemical Geology 513: 120-132. https://doi.org/10.1016/j.chemgeo.2018.12.021
Strand, E., Bagøien, E., Edwards, M., Broms, C., Klevjer, T. 2020. Spatial distribution and seasonality of four Calanus species in the Northeast Atlantic. Progress in Oceanography, 185: 102344.
Stroeve, J., Notz, D. 2018. Changing state of Arctic sea ice across all seasons. Environ. Res. Lett. 13 103001.
Svensen, C., Halvorsen, E., Vernet, M., mfl., 2019. Zooplankton communities associated with new and regenerated primary production in the Atlantic inflow north of Svalbard. Frontiers in Marine Science. https://doi.org/10.3389/fmars.2019.00293
Svensen, C., Seuthe, L., Vasilyeva, Y., Pasternak, A., Hansen, E. 2011. Zooplankton distribution across Fram Strait in autumn: Are small copepods and protozooplankton important? Progress in Oceanography, 91:534-544.
Systad, GHR., Fauchald, P., Descamps, S., Christensen-Dalsgaard, S., Strøm, H., Tarroux, A. 2019. Identifisering av viktige områder for sjøfugl i norske havområder - innspill til forvaltningsplan-arbeidet 2018. NINA Rapport 1627.
Syvertsen, EE. 1991. Ice algae in the Barents Sea: types of assemblages, origin, fate and role in the ice-edge phytoplankton bloom. Polar Res. 10(1), 277-288.
Søvik, G., Furevik, D., Jørgensen, T., Bakke, S, Larssen, WE., Thangstad, TH., Woll, AK. 2017. The Norwegian Nephrops fishery – history, exploitation and management. In: Thu, BJ., Gundersen, AC. (eds.). Sustainable bio-resources. Management, product development and raw material quality. Orkana Akademisk, Stamsund, pp: 95-118. ISBN 978-82-8104-290-2.
Tandberg, AHS., Rapp, H.T, Schander, C., Vader, W., Sweetman, AK., Berge, J. 2012. Exitomelita sigynae gen. et sp. nov.: a new amphipod from the Arctic Loki Castle vent field with potential gill ectosymbionts. Polar Biology 35: 705-716. https://doi.org/10.1007/s00300-011-1115-x
Tandberg, AHS., Rapp, HT., Schander, C., Vader, W. 2013. A new species of Exitomelita (Amphipoda: Melitidae) from a deep-water wood fall in the northern Norwegian Sea. Journal of Natural History 47: 25-28. https://doi.org/10-1080/00222933.2012.725778
Tandberg, AHS., Vader, W., Olsen, BR., Rapp, HT. 2018. Monoculodes bousfieldi sp.n. from the Arctic hydrothermal vent Loki’s Castle. Marine Biodiversity 48: 927-937. https://doi.org/10.1007/s12526-018-0869-6
Taylor, J., Devey, C., Le Saout, M. mfl. (i trykken). The geological and faunal composition of Steinahóll Vent Sites, Reykjanes Ridge, Iceland. Frontiers in Marine Biology.
Toresen, R. 1991. Predation on the eggs of Norwegian spring-spawning herring (Clupea harengus L.) on a spawning ground on the west coast of Norway. ICES J. Mar. Sci. 48(1): 15–21.
Torgersen, T., Huse, G. 2005. Variability in retention of Calanus finmarchicus in the Nordic Seas. ICES Journal of Marine Science, 62, 1301-1309.
Torgersen, T., Kaartvedt, S., Melle, W., Knutsen, T. 1997. Large scale distribution of acoustical scattering layers at the Norwegian continental shelf and the eastern Norwegian Sea. Sarsia, 82:87-9
Triginer, VG. 2020. Distribution range, occurrence and bycatch risk of a large pelagic shark, the porbeagle (Lamna nasus), in Norwegian waters. MSc thesis. University of Oslo. http://urn.nb.no/URN:NBN:no-82861
Tschesunov, AV. 2006. Biology of marine nematodes. Moscow, KMK Scientific Partnership Publishers, 367 p. (På russisk).
Vacquié-Garcia, J., Lydersen, C., Marques, TA., Aars, A., Ahonen, H., Skern-Mauritzen, M., Øien, N., Kovacs, KM. 2017. Late summer distribution and abundance of ice-associated whales in the Norwegian High Arctic. Endang Spec Res 32: 59-70. doi: 10.3354/esr00791.
Varpe, Ø., Fiksen, Ø., Slotte, A. 2005. Meta-ecosystems and biological energy transport from ocean to coast: the ecological importance of herring migration. Oecologia, 146(3):443–451.
Vinje, T., Nordlund, N., Kvambekk, Å. (1998). Monitoring ice thickness in Fram Strait. Journal of Geophysical Research: Oceans, 103(C5),
Voet, G., Quadfasel, D., Mork, KA., Soiland, H. 2010. The mid-depth circulation of the Nordic seas derived from profiling float observations. Tellus, 62(4), 516–529.
von Quillfeldt, CH. (red.) 2018. Miljøverdier og sårbarhet i iskantsonen. Norwegian Polar Institute, Brief report no. 047, 262 s.
Wang, S.W., Budge SM., Iken, K., Gradinger RR., Springer AM., Wooller MJ., 2015. Importance of sympagic production to Bering Sea zooplankton as revealed from fatty acid-carbon stable isotope analyses. Marine Ecology Progress Series, 518, s.31-50.
Wassmann, P., Reigstad, M. 2011. Future Arctic Ocean seasonal ice zones and implications for pelagic-benthic coupling. Oceanogr. 24, 220-231.
Wassmann, P., Reigstad, M., Haug, T., Rudels, B., Carroll, ML., Hop, H., Gabrielsen, GW., Falk‐Petersen, S., Denisenko, SG., Arashkevich, E., Slagstad, D., Pavlova, O. 2006. Food webs and carbon flux in the Barents Sea. Progress in Oceanography 71, 232‐287.
Webster, MA., Parker, C., Boisvert, L. Kwok, R. 2019. The role of cyclone activity in snow accumulation on Arctic sea ice. Nature Communications, 10,5285. https://doi.org/10.1038/s41467-019-13299-8
Werner, I., Gradinger, R. 2002. Under-ice amphipods in the Greenland Sea and Fram Strait (Arctic): environmental controls and seasonal patterns below the pack ice. Marine Biology 140(2), 317-326.
White, M., Bashmachnikov, I., Arístegui, J., Martins, AR. 2007. Physical processes and seamount productivity, I: Pitcher, T.J. mfl. (Red.) Seamounts: ecology, fisheries & conservation. Fish and Aquatic Resources Series, 12: s. 65-84
Wienerroither, RM., Nedreaas, KH., Uiblein, F., Christiansen, JS., Byrkjedal, I. Karamushko, O. 2011. The marine fishes of Jan Mayen Island, NE Atlantic – past and present. Marine Biodiversity 41:395–411
Zamani, B., Krumpen, T., Smedsrud, LH., Gerdes, R. 2019. Fram Strait sea ice export affected by thinning comparing high-resolution simulations and observations. Climate Dynamics , 53 (5-6), 3257-3270. https://link.springer.com/article/10.1007/s00382-019-04699-z
Zimmermann, F., Søvik, G., Thangstad, TH. 2019. Kunnskapsstatus rekefelt langs norskekysten. Bestilling fra Fiskeridirektoratet. Rapport fra Havforskningen nr. 2019-15. ISSN:1893-4536. 13 s.
Øien, N. 1988. The distribution of killer whales (Orcinus orca) in the North Atlantic based on Norwegian catches, 1938-1981, and incidental sightings, 1967-1987. Rit. Fiskideildar 11: 65-78.
Åström, EKL., Carroll, M.L, Ambrose, WG jr., Sen, A., Silyakova, A. Carroll, JL. 2017. Methane cold seeps as biological oases of the high-Arctic deep sea. Limnology and Oceanography 63: S209-S231. https://doi.org/10.1002/lno.10732
14 - Referanser Nordsjøen og Skagerrak
Adlerstein, S.A. og Welleman, H.C. (2000) Diel variation ofstomach contents of North Sea cod (Gadus morhua) during a 24-h fishing survey: an analysis using generalized additivemodels. Canadian Journal of Fishery and Aquatics Sciences,57, 2363–2367.
Aglen, A., Nedreaas, K., Moland, E., Knutsen, H., Kleiven, A.R., Johannessen, T., Wehde, H., Jørgensen, T., Espeland, S.H., Olsen, E.M., Knutsen, J. A.Kunnskapsstatus kysttorsk i sør (Svenskegrensa - Stadt) 2016. Fisken og Havet 4/2016.
Aksnes, DL., Dupont, N., Staby, A., Fiksen, O., Kaartvedt, S Aure, J. 2009.Coastal water darkening and implications for mesopelagic regime shifts in Norwegian fjords. Marine Ecology-Progress Series, 387, 39–49.
André, C., Svedäng, H., Knutsen, H., Dahle, G., Jonsson, P., Ring, A-K., Sköld, M., Jorde, PE. 2016. Population structure in Atlantic cod in the eastern North Sea‑Skagerrak‑Kattegat: early life stage dispersal and adult migration. BMC Research Notes 9, 63. https://www.researchgate.net/publication/292986183
Aure, J., Stigebrandt, A. 1989. On the influence of topographic factors upon the oxygen consumption rate in sill basins of fjords. Estuarine, Coastal and Shelf Science, 28, 1, 59-69.
Bakketeig IE., Hauge M. Kvamme C (red) 2017. Havforskningsrapporten 2018. Fisken og Havet, særnr 1-2017
Barceló, C., Ciannelli, L., Olsen, E. M., Johannessen, T., Knutsen, H. 2016. Eight decades of sampling reveal a contemporary novel fish assemblage in coastal nursery habitats. Global Change Biology. 22(3), 1155 - 1167. https:/doi.org/10.1111/gcb.13047.
Beaugrand, G., Reid, P., Ibañez, F., Lindley, J., Edwards, M. 2002. Reorganization of North Atlantic Marine Copepod Biodiversity and Climate. Science (New York, N.Y.). 296. 1692-4. 10.1126/science.1071329.
Bergstad, OA. 1991. Distribution and trophic ecology of some gadoid fish of the Norwegian deep, Sarsia, 75:4, 315-325, DOI: 10.1080/00364827.1991.10413456
Chierici, M. Fransson, A. 2009. CaCO3 saturation in the surface water of the Arctic Ocean: undersaturation in freshwater influenced shelves. Biogeosciences, 6, 2421-2432. www.biogeosciences.net/6/2421/2009
Edwards, M., Helaouet, P., Alhaija, R.A., Batten, S., Beaugrand, G., Chiba, S., Horaeb, R.R., Hosie, G., Mcquatters-Gollop, A., Ostle, C. mfl. 2016. Global Marine Ecological Status Report: results from the global CPR Survey 2014/2015. SAHFOS Technical Report, 11: 1-32. Plymouth, U.K. ISSN 1744-0750.
Edwards, M., Reid, P., Planque, B. 2001. Long-term and regional variability of phytoplankton biomass in the Northeast Atlantic (1960–1995), ICES Journal of Marine Science 58 (1), Pages 39–49, https://doi.org/10.1006/jmsc.2000.0987 .
Eliasen, K. 2013. Sandeel, Ammodytes spp., as a Link between Climate and Higher Trophic Levels on the Faroes Shelf (Doctoral dissertation, PhD Thesis, Faroe Marine Research Institute).
Erga, SR. 1989. Ecological studies on the phytoplankton of Boknafjorden, western Norway. 1. The effect of water exchange processes and environmental factors on temporal and vertical variability of biomass, Sarsia, 74:3, 161-176, DOI: 10.1080/00364827.1989.10413425
Fagerli, CW., Trannum, HC., Staalstrøm, A., Eikrem, W., Sørensen, K., Marty, S., Frigstad, H., Gitmark, J. 2020. Økokyst Delprogram Skagerrak. Miljødirektoratet M-1603. 2020.
Falkenhaug, T., Bagøien, E., Broms, C., Albretsen, J. 2014. Dynamics of co-occurring Calanus finmarchicu s and C. helgolandicus in Skagerrak. Poster at ICES/PISCES 6 th Zooplankton Symposium, Bergen 2014.
Fauchald, P., Lorentsen, S-H., Systad, G.H. Tveraa, T. 2006. Utbredelsen av sjøfugl i Skagerrak, Kattegat og Nordsjøen . - NINA Rapport 171.
Findlay, HS., Artioli, Y., Moreno Nava, J., Hennige, SJ., Wicks, LC., Huvenne, EMS. Woodward, EMS., Roberts, JM. 2013. Tidal downwelling and implications for the carbon biogeochemistry of cold-water corals in relation to future ocean acidification and warming. Glob. Change Biol. 19, 2708-2719, doi: 10.1111/gcb.12256
Findlay, HS., Hennige, SJ., Wicks, LC., Navas, JM., Woodward, EMS., Roberts, JM. 2014. Fine-scale nutrient and carbonate system dynamics around cold-water coral reefs in the northeast Atlantic. Sci Rep 4, 3671. https://doi.org/10.1038/srep03671 .
Findlay, HS, Tyrrell, T, Bellerby, RGJ, Merico, A, Skjelvan, I, 2008. Carbon and nutrient dynamics of the Norwegian Sea. Biogeosci., 5, 1395-1410.
Fosså, JH., Kutti, T., Buhl-Mortensen, P., Skjoldal, HR. 2015. Vurdering av norske korallrev. Rapport fra Havforskningen Nr. 8–2015, 64 s.
Fredriksen, M., Edwards, M., Richardson, AJ., Hallliday, NC., Wanless, S. 2006. From plankton to top predators: bottom-up control of a marine food web across four trophic levels. Journal of Animal Ecology 75. S. 1259-1268.
Freitas, C., Olsen, E. M., Knutsen, H., Albretsen, J., Moland, E. 2016. Temperature‐associated habitat selection in a cold‐water marine fish. Journal of Animal Ecology 85(3), 628-637.
Frigstad, H., Andersen, GS., Trannum, HC., Naustvoll, LJ., Kaste, Ø., Hjermenn, DØ. 2018. Synthesis of climate relevant result from selected monitoring programs in the coastal zone. Part 2: Quantitative analyses. NIVA rapport 7311-2018.
Frigstad, H., Kaste, Ø., Deininger, A., Kvalsund, K., Christensen, G., Bellerby, RGJ., Sørensen, K., Norli, M., King, A. 2020. Influence of riverine input on Norwegian coastal systems. Frontiers in Marine Science 7, 332. doi:10.3389/fmars.2020.00332
Furness, RW. 1990. A preliminary assessment of the quantities of Shetland sandeels taken by seabirds, seals, predatory fish and the industrial fishery in 1981–83. Ibis, 132: 205–217.
Furness, RW. 2002. Management implications of interactions between fisheries and sandeel-dependent seabirds and seals in the North Sea. ICES Journal of Marine Science, 59: 261–269.
Gasbjerg, G., Christensen-Dalsgaard, S., Lorentsen, S-H., Systad, GH. Anker-Nilssen, T. 2011. Tverrsektoriell vurdering av konsekvenser for sjøfugl. Grunnlagsrapport til en helhetlig forvaltningsplan for Nordsjøen og Skagerrak. - NINA Rapport 733. 139 s.
Gao, S., Hjøllo, SS., Falkenhaug, T., Strand, E., Edwards, M., Skogen, MD. 2021. Overwintering distribution, inflow patterns and sustainability of Calanus finmarchicus in the North Sea. Progress in Oceanography 194 (2021)
Granhag L., Møller LF Hansson LJ. 2011. Size-specific clearance rates of the ctenophore Mnemiopsis leidyi based on in situ gut content analyses. Journal of Plankton Research 33: 1043–1052.
Greenstreet, SPR., McMillan, JA., Armstrong, A. 1998. Seasonal variation in the importance of pelagic fish in the diet of piscivorous fish in the Moray Firth, NE Scotland: a response to variation in prey abundance. ICES Journal of Marine Science, 55, 121–133.
Hammond, PS. Wilson, L.J. 2016. Grey seal diet composition and prey consumption. Scottish Marine and Freshwater Science Report7 (20): 1-52.
Hammond. PS., Lacey, C., Gilles, A., Viquerat, S., Börjesson, P., Herr, H., Macleod, K., Ridoux, V., Santos, MB., Scheidat, M., Teilmann, J., Vingada, J.2017. Estimates of cetacean abundance in European Atlantic waters in summer 2016 from the SCANS-III aerial and shipboard surveys. Environmental Science. 40 s.
Hanssen, SA., Christensen-Dalsgaard, S., Moe, B., Langset, M. Anker-Nilssen, T. 2020. Økt vinterdødelighet hos ærfugl i ytre Oslofjord og Agder. Statusrapport høsten 2020. NINA Rapport 1862. Norsk institutt for naturforskning.
Heath, MR., Boyle, PR., Gislason, A., Gurney, WCS., Hay, SJ., Head, EJH., Holmes, S., Ingvarsdóttir, A., Jónasdóttir, SH., Lindeque, P., Pollard, RT., Rasmussen, J., Richards, K., Richardson, K., Smerdon, G., Speirs, D. 2004. Comparative ecology of over-wintering Calanus finmarchicu s in the northern North Atlantic, and implications for life-cycle patterns, ICES Journal of Marine Science, Volume 61, Issue 4, 2004, Pages 698–708, https://doi.org/10.1016/j.icesjms.2004.03.013
Heip, C., Craeymeersch, JA. 1995. Benthic community structures in the North Sea. Helgolander Meeresunters 49, 313–328. https://doi.org/10.1007/BF02368359
Hjort, J., Ruud, JT. 1938. Rekefisket som naturhistorie og samfundssak. Fiskeridirektoratets skrifter, Serie Havundersøkelser Vol. V, No. 4. 158 s.
Holland, G. J., Greenstreet, SP., Gibb, IM., Fraser, HM., Robertson, MR. 2005. Identifying sandeel Ammodytes marinus sediment habitat preferences in the marine environment. Marine Ecology Progress Series, 303, 269-282.
Huserbråten, MBO., Moland, E., Albertsen, J. 2018. Cod at drift in the North Sea. Progress in oceanography 167, 116-124. https://doi.org/10.1016/j.pocean.2018.07.005
Høines, ÅS., Bergstad, OA. 1999. Resource sharing among cod, haddock, saithe and pollack on a herring spawning ground. Journal of Fish Biology, 55, 1233–1257.
ICES. 2017. Report of the Benchmark on Sandeel (WKSand 2016), 31 October - 4 November 2016, Bergen, Norway. ICES CM 2016/ACOM:33. 319 s.
ICES 2020 a. Working Group on the Assessments of Demersal Stocks in the North Sea and Skagerrak (WGNSSK). ICES Scientific Reports. 2:61. 1249 pp. http://doi.org/10.17895/ices.pub.6092
ICES 2020 b. Herring Assessment Working Group for the Area South of 62°N (HAWG). ICES Scientific Reports. 2:60. 1054 s. http://doi.org/10.17895/ices.pub.6105
ICES 2020 c. International ecosystem survey in Nordic Sea (IESNS) in May-June 2020. 16-18 June 2020, on Teams.
Järnegren, J., Kutti, T. 2014. Lophelia pertusa in Norwegian waters. What have we learned since 2008? - NINA Report 1028. 40 pp. ISBN: 978-82-426-2640-0.
Jensen, H., Rindorf, A., Wright, PJ., Mosegaard, H. 2011. Inferring the Location and Scale of Mixing between Habitat Areas of Lesser Sandeel through Information from the Fishery. Ices Journal of Marine Science 68: 43–51. https://doi.org/10.1093/icesjms/fsq154 .
Johannessen, A., Blom, G., Folkvord, A Svendsen, H. 1995. The effect of local wind on the distribution of early Norwegian spring spawning herring (Clupea harengus L.) larvae. In: Ecology of Fjords and Coastal Waters. H. Skjoldal, C. Hopkins, K. Erikstad. H. Leinaas (Red) Amsterdam: Elsevier Science Ltd., s. 365–384.
Johannessen, T., Dahl, E., Falkenhaug, T., Naustvoll, LJ. 2012. Concurrent recruitment failure in gadoids and changes in the plankton community along the Norwegian Skagerrak coast after 2002. ICES Journal of Marine Science 69,795-801. http://doi.org/10.1093/icesjms/fsr194
Johnsen, E. 2020. Råd for tobisfiskeriet i norsk sone for 2020 og rapport for tobistokt i Nordsjøen 23. april – 13. mai. Toktrapport/Havforskningsinstituttet/ISSN 1503 6294/Nr.9–2020
Johnsen, E., Rieucau, G., Ona, E., Skaret, G. 2017. Collective Structures Anchor Massive Schools of Lesser Sandeel to the Seabed, Increasing Vulnerability to Fishery. Marine Ecology Progress Series 573, 229–36. https://doi.org/10.3354/meps12156.
Johnson, MP., Lordan C., Power, AM. 2013. Habitat and ecology of Nephrops norvegicus . Advances in Marine Biology 64, 27-63.
Jónasdóttir, S., Koski, M. 2011 Biological processes in the North Sea: comparison of Calanus helgolandicus and Calanus finmarchicus vertical distribution and production, Journal of Plankton Research, Volume 33, Pages 85–103, https://doi.org/10.1093/plankt/fbq085
Jones, E., Chierici, M., Skjelvan, I., Norli, M., Frigstad, H., Børsheim, KY., Lødemel, HH., Kutti, T., King, AL., Sørensen, K., Lauvset SK., Jackson-Misje, K., Apelthun, LB., de Lange, T., Johannessen, T., Mourgues, C., Bellerby, R. 2020. Monitoring ocean acidification in Norwegian Seas 2019. Rapport, Miljødirektoratet, M-1735.
Jorde, PE., Søvik, G., Westgaard, J-I., Albretsen, J., André, C., Hvingel, C., Johansen, T., Sandvik, AD., Kingsley, M., Jørstad, KE. 2015. Genetically distinct populations of northern shrimp, Pandalus borealis , in the North Atlantic: adaptation to different temperatures as an isolation factor. Molecular Ecology 24, 1742-1757.
Keppel, G., Van Niel, KP., Wardell‐Johnson, GW., Yates, CJ., Byrne, M., Mucina, L., Schut, AGT., Hopper, SD., Franklin, SE. 2012. Refugia: identifying and understanding safe havens for biodiversity under climate change. Global Ecology and Biogeography 21, 393-404. https://doi.org/10.1111/j.1466-8238.2011.00686.x
Kitidis, Hardman-Mountford, VN., Litt, E., Brown, I. mfl. 2012: Seasonal dynamics of the carbonate system in the Western English Channel. 2012. Continental Shelf Research, 42, 30-40, http://dx.doi.org/10.1016/j.csr.2012.04.012
Kiørboe, T., Munk, P., Richardson, K., Christensen, V., Paulsen, H. 1988. Plankton dynamics and herring larval growth, drift and survival in a frontal area. Mar. Ecol. Prog Ser. 44: 205-219.
Knutsen, H., Jorde, PE., Gonzalez, EB., Eigaard, OR., Pereyra, RT., Sannæs, H., Dahl, M., André, C., Søvik, G. 2015. Does population genetic structure support present management regulations of the northern shrimp (Pandalus borealis) in Skagerrak and the North Sea? ICES Journal of Marine Science 72(3), 863-871.
Knutsen, H., Jorde, P. E., Hutchings, J. A., Hemmer‐Hansen, J., Grønkjær, P., Jørgensen, K. E. M., André, A., Sodeland, M., Albretsen, J., Olsen, E. M. 2018. Stable coexistence of genetically divergent Atlantic cod ecotypes at multiple spatial scales. Evolutionary Applications 11, 1527-1539.
Kraufvelin, P., Svensson, F., Fredriksson, R., Bergström, L., Karlsson, M., Wennhage, H., Wikström, A., Bergström, U. 2017. Inventering och modellering av fisk-och kräftdjurssamhällen i Kosterhavets nationalpark. Länsstyrelsen Västra Götaland, Göteborg. Pp: 29-41; 65-75.
Kröncke, I., Bergfeld, C. 2003. North sea benthos: a review. Senckenbergiana maritima 33. 205–268. https://doi.org/10.1007/BF03043049
Macer, CT. 1966. Sandeels (Ammodytidae) in the south-western North Sea: their biology and Fishery. MAFF Fishery Invest. London Ser. II 24(6):1-55.
Madden, H., Beare, D., Heath, MR., Fraser, JG., Gallego, A. 1999. The spring/early summer distribution of Calanus arter in the northern North Sea and adjacent areas. Fish. Oceanogr. 8 (Suppl. 1): 138 – 152.
Meld. St. 29 (2020-2021). Heilskapleg nasjonal plan for bevaring av viktige områder I marin natur. Tilråding fra Klima- og miljødepartementet, 9. april 2021, godkjend i statsråd same dag. (Regjeringen Solberg). https://www.regjeringen.no/no/dokumenter/meld.-st.-29-20192020/id2715224/
Melle, W., Runge, J., Head, E., Plourde, S., Castellani, C., Licandro, P., Pierson, J., Jonasdottir, S., Johnson, C., Broms, C. mfl. 2014. The North Atlantic Ocean as habitat for Calanus finmarchicu s: Environmental factors and life history traits. Progress in Oceanography, 129: 244-284. https://doi.org/10.1016/j.pocean.2014.04.026
Maar, M., Nielsen, TG., Gooding, S., Tonnesson, K. mfl. 2004. Trophodynamic function of copepods, appendicularians and protozooplankton in the late summer zooplankton community in the Skagerrak. Mar Biol 144: 917–933.
Naustvoll, LJ., Thormar, J., Lundsør, E., Kroglund, T., Norderhaug, KM., Moy, F. 2020. ØKOKYST-Delprogram Klima, Årsrapport 2019. Miljødirektoratet M-1612/2020.
Nilssen, KT. Bjørge, A. 2019. Status for kystsel – Anbefaling av jaktkvoter 2020. Pp 56-69 i A. Bjørge (Red). Forskerutvalg om sjøpattedyr 2019 – Anbefalinger om forskning og forvaltning. Rapport fra Havforskningen Nr. 2020-19.
Norderhaug, KM., Christie, H., Andersen, GS. og Bekkby, T. 2012. Does the diversity of kelp forest macrofauna increase with wave exposure? Journal of Sea Research 69: 36-42.
O’Brien, TD., Wiebe, PH. og Falkenhaug, T. (Red). 2013. ICES Zooplankton Status Report 2010/2011. ICES Cooperative Research Report No. 318. 208 s.
Obst, M., Vicario, S., Lundin, K., Berggren, M., Karlsson, A., Haines, R., Williams, A., Goble, C., Mathew, C., Güntsch, A. 2018. Marine long-term biodiversity assessment suggests loss of rare species in the Skagerrak and Kattegat region. Marine Biodiversity 48:2165-2176. doi:10.1007/s12526-017-0749-5
Olsen, E., Holst, JC. 2001. A note on common minke whale (Balaenoptera acutorostrata) diets in the Norwegian Sea and the North Sea. Journal of Cetacean Research and Management 3(2). S. 179-183.
Omar, AM., Thoas, H., Olsen, A., Becker, M., Skjelvan, I., Reverdin, G. 2019. Trends in ocean acidification and p CO2 in the Northern North Sea, 2003-2015. Journal of Geophysical Research: Biogeoscience 124, https://doi.org/10.1029/2018JGO04992
Oug, E., Christiansen, M.E., Dobbe, K., Rønning, A-H., Bakken, T., Kongsrud, J.A. 2015. Mapping of marine benthic invertebrates in the Oslofjord and the Skagerrak: sampling data of museum collections from 1950-1955 and from recent investigations. Fauna Norvegica, 35, 35-45. https://doi.org/10.5324/fn.v35i0.1944
Prøis, A. 2018. Ytre Hvaler nasjonalpark: Makroalger og assosiert fauna i fjæresonen, Mastergrad Universitetet i Oslo. https://www.duo.uio.no/bitstream/handle/10852/64228/Alis-Pr-is---Masteroppgave.pdf?sequence=1&isAllowed=y
Ramirez-Llodra, E., Rinde, E., Gundersen, H. mfl. 2016. A snap shot of the short-term response of crustaceans to macrophyte detritus in the deep Oslofjord. Sci Rep 6, 23800. https://doi.org/10.1038/srep23800
Reid, PC., Lancelot, C., Gieskes, WWC., Hagmeier, E., Weichart, G. 1990. Phytoplankton of the North Sea and its dynamics: A review. Netherlands Journal of Sea Research 26(2-4), 295-331.
Reitan, SJ. 2020. Sublitorale makroalger i Ytre Hvaler Nasjonalpark. Masteroppgave, Institutt for biovitenskap, UiO. sunnivareitanmaster.pdf (uio.no)
Richardson, K., Visser, A Pedersen, F. 2000. Subsurface phytoplankton blooms fuel pelagic production in the North Sea. J. Plankton Res., 22, 1663–1671.
Salt 2019. Kunnskapsstatus Oslofjorden. Salt Rapport nr 1036
Skjæveland, SB. 2020. Kvalitativ og kvantitativ inventering av fiskesamfunnet i Ytre Oslofjord – en beskrivelse av diversitet og generelle trekk ved fiskesamfunnet i Færder- og Ytre Hvaler nasjonalparker for perioden 2017-2019. MSc - Masteroppgave ved Universitet i Agder.
Skogen, M, Moll, A. 2000. Interannual variability of the North Sea primary production: comparison from two model studies. Continental Shelf Research 20. 129-151.
Strand, E., Bagøien, E., Edwards, M., Broms, C., Klevjer, T. 2020. Spatial distributions and seasonality of four Calanus species in the Northeast Atlantic. Progress in Oceanography 185: (2020) 102344, ISSN 0079-6611, https://doi.org/10.1016/j.pocean.2020.102344 .
Sundby, S., Kristiansen, T., Nash, R., Johannessen, T. 2017. Dynamic Mapping of North Sea Spawning–Report of the KINO Project.
Synnes, A.E.W. 2020. Seascape ecology of Atlantic cod (Gadus morhua) in coastal Skagerrak: population structure, connectivity, and role in fish assemblage. PhD - Doktorgradavhandling, Universitetet i Agder.
Systad, GHR., Bjørgesæter, A., Brude, OW., Skeie, GM. 2018. Standardisering og tilrettelegging av sjøfugldata til bruk i konsekvens- og miljørisikoberegninger. NINA Rapport 1509. Norsk institutt for naturforskning.
Systad, GHR., Fauchald, P., Descamps, S., Christensen-Dalsgaard, S., Strøm, H. Tarroux, A. 2019. Identifisering av viktige områder for sjøfugl i norske havområder – innspill til forvaltningsplanarbeidet 2018. NINA Rapport 1627. Norsk institutt for naturforskning.
Systad, G., Hanssen, SA., Anker-Nilssen, T. og Lorentsen, S-H. 2007. Særlig Verdifulle Områder (SVO) for sjøfugl i Nordsjøen og Norskehavet - NINA Rapport 230. 54 s. Norwegian Institute for Nature Research (NINA), Trondheim.
Sætre, R. (Red.) 2007. The Norwegian Coastal Current—Oceanography and Climate. Tapir Academic Press, Trondheim. 159 s.
Sørlie, M., Nilssen, KT., Bjørge, A. Freitas, C. 2020. Diet composition and biomass consumption of harbour seals in Telemark and Aust-Agder, Norwegian Skagerrak. Marine Biology Research 16(2). https://doi.org/10.1080/17451000.2020.1751205
Søvik, G., Furevik, D., Jørgensen, T., Bakke, S, Larssen, WE., Thangstad, TH., Woll, AK. 2017. The Norwegian Nephrops fishery – history, exploitation and management. In: Thu, BJ., Gundersen, AC. (eds.). Sustainable bio-resources. Management, product development and raw material quality. Orkana Akademisk, Stamsund, pp: 95-118. ISBN 978-82-8104-290-2.
Søvik, G., Thangstad, TH. 2021. Results of the Norwegian Bottom Trawl Survey for Northern Shrimp (Pandalus borealis) in Skagerrak and the Norwegian Deep (ICES Divisions 3.a and 4.a east) in 2021. NAFO SCR Doc. 21/001, Serial No. N7157. 38 pp. https://www.nafo.int/Portals/0/PDFs/sc/2021/scr21-001.pdf
Thomas, H., Bozec, YB., Elkaya, K., de Baar, JW. 2004. Enhanced Open Storage of CO 2 from Shelf Sea Pumping. Science 304 (5673), s. 1005-1008. DOI: 10.1126/science.1095491
Walday, M., Borgersen, G., Beylich, B., Eikrem, W., Gitmark, J., Naustvoll, LJ., Selvik, JR. 2019. Overvåking av Ytre Oslofjord i 2014-2018. NIVA rapport l. nr. 7423-2019. 106 s.
van Deurs, M., van Hal, R., Tomczak, MT., Jonasdottir, SH., Dolmer, P. 2009. Recruitment of Lesser Sandeel Ammodytes Marinus in Relation to Density Dependence and Zooplankton Composition. Marine Ecology-Progress Series 381: 249–58. https://doi.org/10.3354/meps07960.
van Deurs, M., Koski, M., Rindorf., A.2014. Does Copepod Size Determine Food Consumption of Particulate Feeding Fish? ICES Journal of Marine Science 71, 1: 35–43. https://doi.org/10.1093/icesjms/fst090 .
Weir, CR. 2001. Sightings of marine mammals and other animals recorded from offshore installations in the North Sea. Pp 93-103 in A.W. Thorpe (Red) North Sea Bird Club 21st Anniversary report.
Wright, PJ., Jensen, H., Tuck, I. 2000. The Influence of Sediment Type on the Distribution of the Lesser Sandeel, Ammodytes marinus . Journal of Sea Research 44: 243–56.
15 - Fagekspertgrupper
Forkortninger
Prosjektleder – PL; Institusjonskoordinator – IK; Fagkoordinatorer – FK; Referansegruppe – RG; Havforskningsinstituttet - HI; Norsk Polarinstitutt - NP; Universitetet i Bergen - UiB; Universitetet i Oslo - UiO; Universitetet i Tromsø - UiT; Norsk institutt for naturforskning - NINA; Akvaplan-niva - A-N; Norges geologiske undersøkelse - NGU; Miljødirektoratet - Mdir.
Fokusere på kriteriesettet og da spesielt langlevede arter
Margaret Dolan
NGU
Den Midtatlantiske ryggen
Børge Holte
HI
Kystsonen, Barentshavet
Tina Kutti
HI
Den Midtatlantiske ryggen
Lis Lindal Jørgensen
HI
Barentshavet-økotokt
Henning Steen
HI
Tarsekog, Norskehavet og Nordsjøen
Guldborg Søvik
HI
Reke og sjøkreps
Torsten Hugo Struck
UiO
Biodiversitet, Nordsjøen
Anne Helene Solberg Tandberg
UiB
Den Midtatlantiske ryggen
Gro I. van der Meeren
HI
Krepsdyr og grurnnere vann
Andrey Voronkov
HI
RG Barentshavet - kyst, Mareano
Sjøfugl
Geir Systad
NINA
FK
Signe Christensen-Dalsgaard
NINA
Sebastian Descamps
NP
Nordlig del av hhv. Barentshavet og Norskehavet
Per Fauchald
NINA
Hallvard Strøm
NP
Nordlig del av hhv. Barentshavet og Norskehavet
Sjøpattedyr
Arne Bjørge
HI
FK
Kit Kovacs
NP
Nordlig del av hhv. Barentshavet og Norskehavet
Martin Biuw
HI
Christian Lydersen
NP
Nordlig del av hhv. Barentshavet og Norskehavet
Kjell T. Nilssen
HI
Nils Øien
HI
Øystein Wiig
UiO
RG
Jon Aars
NP
Isbjørn
Kart
Elena Eriksen
HI
SVO oversiktskart, temakart
Jon Albretsen
HI
Strømkart
Gjermund Hartviksen
Kystverket
RG Kart
Gerhard Heggebø
Kartverket
RG Kart
Per Arne Horneland
HI
Ragnvald Larsen
Mdir
RG Kart
Are Olsen
UiB
Forsuringskart
Redaksjon
Elena Eriksen
HI
FK Kunnskasgrunnlag, arbeidsmøter
Gro I. van der Meeren
HI
FK Rapportredaksjon
Hanne Johnsen
NP
Redaksjon
Bente M. Nilsen
HI
Redaksjon
Cecilie H. von Quillfeldt
NP
Redaksjon
Redaksjonsstøtte
Mimi Stokkeland
UiB
Referanselisterydding, annen informasjon
Susanne Zazzera
UiB
Referanselisterydding, annen informasjon
Redaksjon, rapportvurdering
Harald Loeng
HI
Fagfellevurdering
Vedlegg 1 Marine verneområder og andre effektive arealbaserte bevaringstiltak
Nylig kom nasjonal plan for bevaring av viktige områder for marin natur (Meld. St. 29 (2020-2021)). Meldingen omtaler kunnskap, virkemidler og status for arbeidet med marine verneområder og andre effektive arealbaserte bevaringstiltak, både nasjonalt og internasjonalt. Meldingen gir en oversikt over områder med ulike bevaringstiltak i Norge. Mange av disse ligger helt eller delvis i et eksisterende SVO. I stedet for å omtale dette under hvert SVO-forslag er det her gjort en kortfattet oppsummering med utgangspunkt i meldingen. Meldingen understreker at bevaring av viktige områder for marin natur er en integrert del av en bærekraftig havøkonomi. I den sammenheng pekes det også på den pågående SVO-prosessen som et viktig bidrag til en mer systematisk tilnærming for å vurdere hvordan ulike tiltak kan bidra til en effektiv bevaring av viktige områder i norske havområder.
Marine verneområder omfatter både opprettelse av marine verneområder, som er en egen vernekategori etter naturmangfoldloven, og for eksempel nasjonalparker og naturreservat med marint areal. Formålet er langsiktig bevaring mot påvirkning på tvers av sektorer.
Andre effektive arealbaserte bevaringstiltak er sektortiltak som gir positiv og langvarig bevaringseffekt for biodiversitet og økosystemtjenester i et område. Dette kan for eksempel være samfunn med sjeldne eller truete arter, representative naturlige økosystemer, arter med begrenset utbredelse, områder med kritiske økosystemfunksjoner og -tjenester eller områder med betydning for økologiske sammenhenger.
Et rådgivende utvalg nedsatt av daværende Miljøverndepartementet i samarbeid med Fiskeridepartementet kom i 2004 med en tilrådning om 36 kandidatområder til en marin verneplan som samlet representerte et utvalg av undersjøisk natur fra kysten og skjærgården (Figur 1). 15 av disse er senere opprettet som marine verneområder etter naturmangfoldloven. Tre av områdene som ligger helt eller delvis utenfor territorialgrensen er opprettet som marine beskyttede områder etter havressursloven. Det er meldt oppstart av verneprosess etter naturmangfoldloven for flere områder. Samtidig som at arbeidet med den marine verneplanen fortsetter, forberedes det en fase 2 som inkluderer områder lenger ut fra kysten. Marint areal inngår også i flere nasjonalparker, som ytre Hvaler nasjonalpark, Færder nasjonalpark og Raet nasjonalpark (hvor også et av verneområdene er lokalisert). Samlet sett utgjør vern etter naturmangfoldloven om lag 3,6 prosent av territorialfarvannet ved fastlands-Norge.
Territorialfarvannet rundt Jan Mayen er en del av Jan Mayen naturreservat. Om lag 87 prosent av territorialfarvannet rundt Svalbard er en del av verneområder etter svalbardmiljøloven (Figur 2). Norge har også meldt disse områdene inn som «Marine Protected Areas» (MPA) til OSPAR-konvensjonen om bevaring av det marine miljøet i Nordøst-Atlanteren. Denne innmeldingen har ikke konsekvenser for rettslig status for verneområdet nasjonalt. OSPAR har et mål om et felles nettverk av marine verneområder, som også skal være økologisk sammenhengende og representativt.
Selv om meldingen peker på behov for en mer systematisk tilnærming til hvordan fiskeritiltak og tiltak i andre sektorer, som petroleumsvirksomheten og nye næringer, kan regnes som andre effektive arealbaserte bevaringstiltak, finnes det allerede flere eksempler på andre effektive arealbaserte bevaringstiltak. Blant annet er atten kjente korallrevområder langs kysten opprettet som marine beskyttete områder etter havressursloven og er dermed beskyttet mot skade fra fiskeriaktivitet (Figur 3). Et annet eksempel er vedtaket om ikke å iverksette petroleumsvirksomhet, inkludert leting og seismikkskyting på Froan-Sularevet og Iverryggen som er en områdespesifikk ramme for petroleumsvirksomhet i Meld. St. 20 (2019-2020). Det er også opprettet hummerfredningsområder på Skagerrakkysten hvor høsting ikke er tillatt, noe som har vist seg å bidra til styrking av hummerbestanden lokalt. Videreutvikling av reglene i forskrift om regulering av fiske med bunnredskap i Norges økonomiske sone, fiskerisonen rundt Jan Mayen og i fiskevernsonen ved Svalbard har ført til forbud mot fiske med bunnredskap i ti områder ved Svalbard (Figur 4). Bakgrunnen for stengingen er at områdene ut fra tilgjengelig kunnskap inneholder forekomster av sårbare arter og habitat, og at det ikke, eller bare i liten grad, har vært fisket der tidligere. Det er også forbudt med vanlig fiske dypere enn 1000 meter sør for Bjørnøya og dypere enn 800 meter nord for Bjørnøya, med unntak av bl.a. Yermakplatået nord for Spitsbergen hvor grensen er 300 meter (Nye fiskeområder). Også grunnere områder rundt Nordaustlandet, kystarealet øst for Svalbard og området rundt Kong Karls Land inngår i nye fiskeområder. I de nye fiskeområdene er ikke fiske tillatt uten at det er gitt spesiell tillatelse.
Figur 1. Eksisterende og planlagte marine verne områder. Kilde: Miljødirektoratet, Kartverket og Fiskeridirektoratet (Fra: Meld. St. 29 (2020-2021)).
Figur 2. Verneområder ved Svalbard og Jan Mayen. Kilde: Miljødirektoratet og Fiskeridirektoratet (Fra: Meld. St. 29 (2020-2021)).
Figur 3. Beskyttelse av korallrev mot ødeleggelser fra fiskeriaktivitet. Kilde: Miljødirektoratet, Kartverket, Fiskeridirektoratet (Fra: Meld. St. 29 (2020-2021)).
Figur 4. Kategorier av områder ved Svalbard som ikke er åpne for vanlig fiske. Kilde: Fiskeridirektoratet (Fra: Meld. St. 29 (2020-2021)).