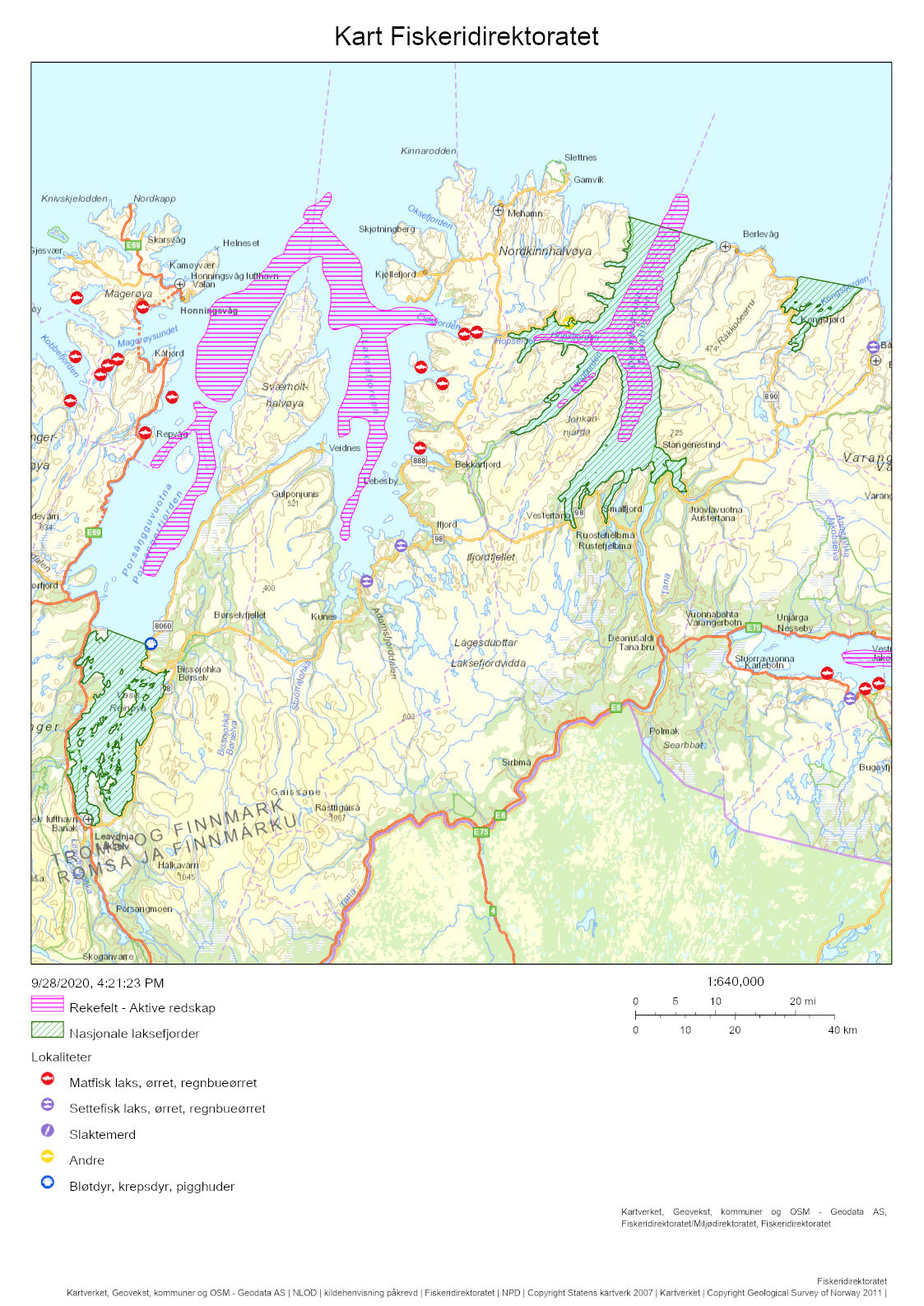
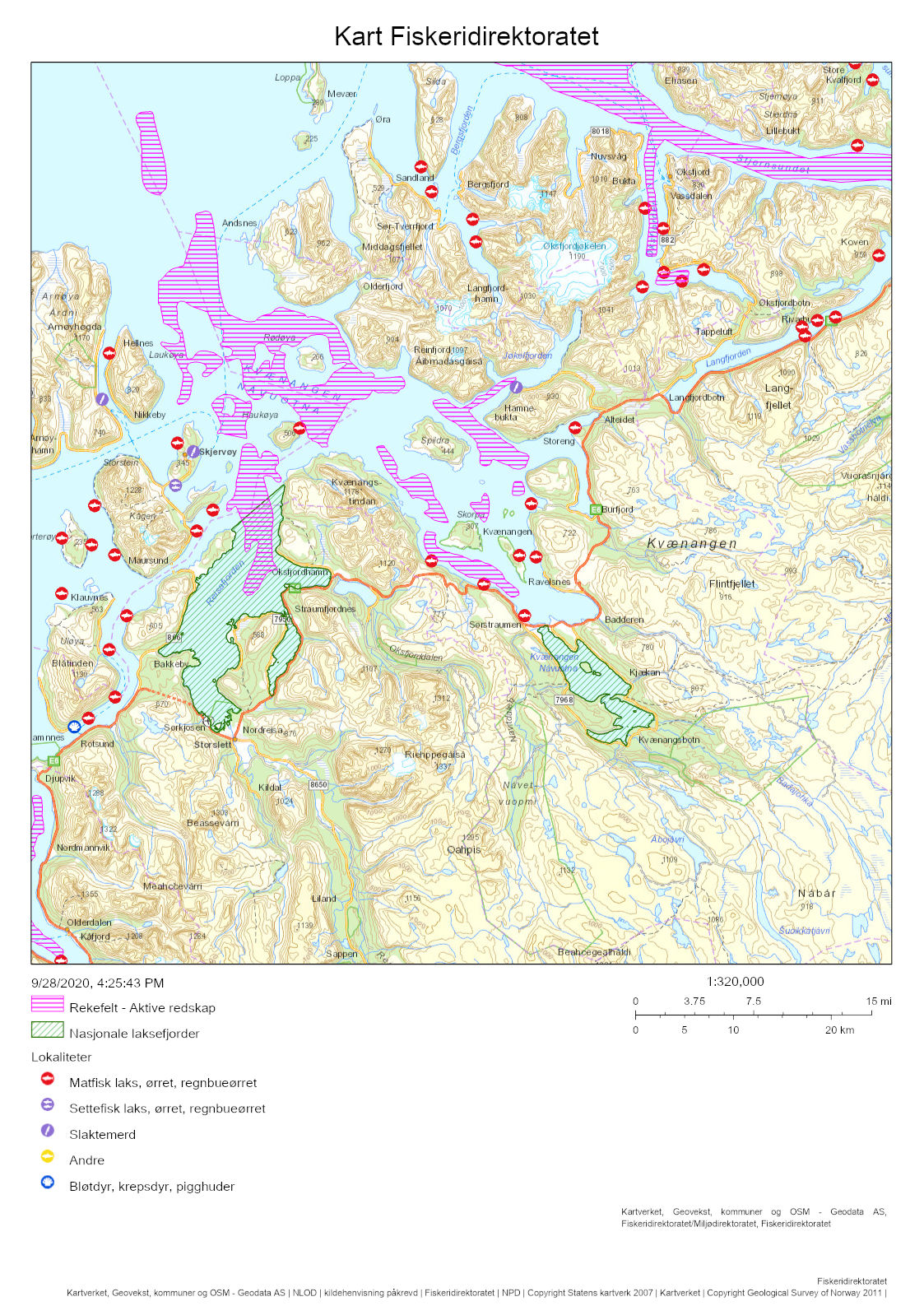
After having been closed for shrimp trawling since the early 1970s, a reopening of the Porsanger and Tana Fjords is currently considered. The Institute of Marine Research (IMR) was tasked by the Norwegian Directorate of Fisheries to assess the ecosystem in the two untrawled fjords. Kvænangen was selected as a reference fjord with an active shrimp trawl fishery. Mapping cruises were carried out in 2018-2019, three with shrimp trawls, and two with shrimp pots. A beam trawl survey was carried out in the Porsanger Fjord in May 2019, as a continuation of a time series started in 2007. The inner part of the Porsanger Fjord southeast of Reinøya is presented separately due to the special environmental conditions in these basins, with very low bottom temperature and very high density of northern shrimp. A total of 158 different species and species groups were found on the cruises carried out with shrimp trawls in 2018 and 2019. The highest species diversity was found in Kvænangen, while in the inner Porsanger Fjord the highest number of unique species was registered. Genetic studies show that shrimp in the Barents Sea and along the Norwegian coast from Trøndelag to Troms (including Kvænangen) constitute two different biological populations, while shrimp in the fjords in Finnmark are a genetic mixture of the coastal and Barents Sea populations. However, our results indicate that the shrimp stocks in Kvænangen, the outer Porsanger Fjord and the Tana Fjord are largely comparable. Shrimp density is at about the same level, with the highest average value in Kvænangen and the lowest in the outer Porsanger Fjord, and the shrimp show faster growth and larger sizes in Kvænangen compared to the two unfished Finnmark fjords. In contrast, the shrimp in the inner Porsanger Fjord stand out clearly with the highest density and a deviating population structure. Cod is a key species in all three fjords. Based on the otoliths, a large proportion of the aged cod (2 years and older) was identified as coastal cod, and only a minority as migrating Northeast Arctic cod (“skrei”). Of all species, haddock was registered most often and with the highest density. A large part of the registered haddock was fry and juvenile fish, and the results suggest that haddock uses the fjords mainly as a nursery area. Herring does the same, and juvenile herring are present all year round in all the fjords. The cruise data indicate that there is little saithe at the bottom of the three fjords, most likely because saithe is more pelagic. Capelin was most abundant in the Porsanger Fjord, while golden redfish, also juveniles, was present in Kvænangen and the Tana Fjord in particular. Stomach contents show that shrimp have very different main predators in the three fjords. In Kvænangen and the outer Porsanger Fjord, shrimp are important for both small and large cod, redfish and small haddock. In the Tana Fjord, large cod is the most important predator, while shrimp in the inner Porsanger Fjord are mainly preyed upon by sculpins and small cod. Vulnerable biotopes typically have large and fragile species of sponges, corals and sea feathers, which are easily destroyed by bottom trawls. The beam trawl surveys in the Porsanger Fjord registered the largest quantities of sponges along the western side of the fjord. Inside the sill north of Lille-Tamsøya, cauliflower corals are found in relatively small quantities in areas deeper than 170 m (current trawl limit). The basins along the east coast are therefore not particularly vulnerable to bottom trawling, while the deep areas in the northwestern part of the fjord have a lot of sponges and can be vulnerable to bottom trawling. Previously conducted video surveys registered corals on the eastern side of the outer Porsanger Fjord outside the sill at Lille-Tamsøya, which shows that there are vulnerable biotopes in the outermost Porsanger Fjord. There are no corresponding surveys from the Tana Fjord. It is crucial that vulnerable areas are mapped before opening for any bottom trawling, as vulnerable bottom fauna is protected and should not be exposed to bottom trawling. For the last three years, there has been a commercial pot fishing for shrimp in the inner Porsanger Fjord. The purpose of the pot surveys carried out as part of IMR’s assessment was to investigate shrimp distribution on non-trawlable bottom, as well as compare catch rates between pots and trawls on trawlable bottom. Shrimp catches were registered on only 46% of 181 pot stations, and catch rates were higher on trawlable bottom. Catch rates in the inner Porsanger Fjord were significantly higher than in the other areas, and the results indicate that pot fishing for shrimp with the current technology and methods will give a low yield outside the special conditions in the inner Porsanger Fjord. IMR recommends that only one of the two closed fjords should be re-opened for shrimp trawling to maintain a protected area that can act as future reference for trawling effects on fjord ecosystems. Precautionary catch options are presented. Additionally, a cautious opening of a possible shrimp trawl fishery is advised to allow for bottom mapping and for building up time series on shrimp. If the Porsanger Fjord is opened, this should only apply to the area outside the sill ("Ryggen") north of Lille-Tamsøya as Porsanger municipality, as part of its plan for rebuilding demersal fish stocks to historic levels, wants to close the inner and eastern part for commercial fishing. A future shrimp trawl fishery should be spatially regulated such that conflicts with the ongoing pot fishing for king crab are minimized, especially in the Porsanger Fjord that has produced the largest landings of king crab over the past years. Temporal closures of areas are recommended if by-catch of commercially important fish are above precautionary limits. The results from the survey show that a mandatory sorting grid is very effective in reducing by-catches, but that fry and juveniles below 10-15 cm remain vulnerable. This may be a particular issue for cod, haddock and redfish when locally occurring in high densities, but bycatches of capelin and juvenile herring should also be monitored.
Kartlegging av fjordøkosystemene i Tana- og Porsangerfjorden
— Råd og kunnskapsbidrag fra Havforskningsinstituttet i forbindelse med vurdering av en eventuell åpning av direktefiske etter reker med bunntrål i Tana- og Porsangerfjorden
Rapportserie:
Rapport fra havforskningen 2020-39
ISSN: 1893-4536
Publisert: 27.10.2020
Prosjektnr: 15619-03
Oppdragsgiver(e): Fiskeridirektoratet
Referanse: 16/7477
Forskningsgruppe(r):
Bentisk økologi
,
Økosystemprosesser
Tema:
Reke – Barentshavet,
Reke – kyst og fjord
Program:
Kystøkosystemer
Godkjent av:
Forskningsdirektør(er):
Geir Huse
Programleder(e):
Jan Atle Knutsen
English summary
Sammendrag
Porsanger- og Tanafjorden har siden tidlig på 1970-tallet vært stengt for reketråling, men vurderes nå åpnet igjen. I den anledning ba Fiskeridirektoratet Havforskningsinstituttet utarbeide en plan for kartlegging av økosystemet i de to utrålte fjordene. Kvænangen ble valgt ut som en referansefjord med et aktivt reketrålfiske. Kartleggingstokt ble gjennomført i 2018-2019, tre med reketrål, og to med reketeiner. Bomtrålundersøkelser ble gjennomført i Porsangerfjorden i mai 2019, som en fortsettelse av en tidsserie fra 2007. Den indre delen av Porsangerfjorden innenfor og øst for Reinøya har spesielle miljøforhold med svært lav bunntemperatur og svært stor tetthet av dypvannsreke. Resultater for dette området presenteres derfor separat. Totalt 158 forskjellige arter og artsgrupper ble funnet på tråltoktene gjennomført høsten 2018, samt våren og høsten 2019. Den høyeste artsdiversiteten ble funnet i Kvænangen, mens indre Porsangerfjorden har flest unike arter. Torsk og hyse ble oftest registrert. Dypvannsreke ligger på en tredjeplass. Rekebestandene i Kvænangen, ytre Porsangerfjorden og Tanafjorden er i stor grad sammenlignbare, mens rekene i indre Porsangerfjorden skiller seg tydelig ut med den høyeste tettheten og en avvikende bestandsstruktur. I de tre andre fjordområdene ligger reketettheten på samme nivå, med den høyeste gjennomsnittsverdien i Kvænangen og den laveste i ytre Porsangerfjorden. Rekene vokser hurtigere og blir større i Kvænangen sammenlignet med de to ufiskede finnmarksfjordene. Genetiske studier viser at reker i Barentshavet og fra kyststrekningen Trøndelag til Troms (inkludert Kvænangen) utgjør to forskjellige biologiske populasjoner, mens fjordene i Finnmark inneholder en blanding av reker fra kysten og fra Barentshavet. Torsk er en nøkkelart i alle tre fjordene. Basert på otolittene ble en stor andel av alderslest torsk (2 år og eldre) identifisert som kysttorsk og kun et mindretall som skrei. Hyse var den arten som ble registrert oftest og med høyest tetthet. En stor del av hysen som ble registrert, var yngel og juvenil fisk, noe som antyder at hyse bruker fjordene hovedsakelig som oppvekstområde. Det samme gjør silda, og ungsild er til stede hele året i alle fjordene. Sei står mer pelagisk, og toktdataene indikerer at det finnes lite sei på bunnen i de tre fjordene. Lodde er det mest av i Porsangerfjorden, mens vanlig uer var særlig til stede i Kvænangen og Tanafjorden, også ungfisk. Analyser av mageinnhold viser at reker har veldig forskjellige hovedpredatorer i de tre fjordene. I Kvænangen og ytre Porsangerfjorden spises reker av både små og stor torsk, uer og liten hyse. I Tanafjorden er stor torsk den viktigste predatoren, mens reker i indre Porsangerfjorden hovedsakelig spises av ulker og små torsk. Selv om de nye toktdataene fra de tre fjordene tillater en karakterisering av fiskesamfunn og dietter, trenger økosystemmodeller (som Ecopath) mer presise data om bunndyr, sjøpattedyr, fugler, pelagisk fisk, plankton og primærprodusenter for å gi en pålitelig beskrivelse av disse fjordøkosystemene. Sårbare biotoper har typisk store og skjøre arter av svamper, koraller og sjøfjær som blir lett ødelagt av bunntrål. Bomtrålundersøkelsene i Porsangerfjorden viser at de største mengdene med svamp ble registrert på vestkysten. Innenfor terskelen ved Lille-Tamsøya finnes blomkålkoraller i relativt små mengder på bunn dypere enn 170 m (nåværende trålegrense). Bassengene langs østkysten er derfor relativt lite sårbare overfor bunntråling, mens de dype områdene nordvest i fjorden har mye svamp og derfor kan være sårbare overfor bunntråling. Det har tidligere blitt registrert koraller i Porsangerfjorden utenfor terskelen ved Lille-Tamsøya, og høy biodiversitet av bl.a. svamp nord for Lille-Tamsøya, som viser at det finnes sårbare biotoper i denne ytterste delen av Porsangerfjorden. Det finnes ikke tilsvarende undersøkelser fra Tanafjorden. Ingen sårbare områder bør åpnes for bunntråling. De tre siste årene har det foregått et kommersielt teinefiske etter reker i indre Porsangerfjorden. Formålet med de gjennomførte teineundersøkelsene var å undersøke mulige rekeforekomster på ikke-trålbar bunn og sammenligne fangstrater mellom teiner og trål. På 181 teinestasjoner var det reker på kun 46 %. Fangstratene i indre Porsangerfjorden var betydelig høyere enn i de andre områdene, og resultatene indikerer at teinefiske etter reker vil gi et lavt utbytte utenfor de spesielle forholdene her. Havforskningsinstituttet anbefaler at bare én av de to stengte fjordene gjenåpnes for reketrålfiske. Forsiktige fangstnivåer er presentert. Blir Porsangerfjorden åpnet, bør dette bare gjelde området utenfor terskelen ved Lille-Tamsøya da Porsanger kommune, som ønsker å gjenoppbygge bunnfiskbestandene til historiske nivåer, ønsker å stenge den innerste og østlige delen for kommersielt fiske. Et fremtidig reketrålfiske kan komme i konflikt med det pågående teinefisket etter kongekrabbe, spesielt i Porsangerfjorden som har de største landingene de siste par årene. Havforskningsinstituttet anbefaler en forsiktig åpning av et eventuelt reketrålfiske, som vil gi tid til bunnkartlegging og oppbygging av tidsserier på reke. Resultatene fra kartleggingen viser at en relativ liten andel av kommersielt viktige fiskebestander som hyse og torsk er sårbar for reketråling. Det er hovedsakelig torske-, hyse- og ueryngel (< 10-15 cm) som kan bli påvirket, men et rekefiske må også overvåkes med hensyn til bifangster av lodde og ungsild.
1 - Bakgrunn
Porsanger- og Tanafjorden har siden tidlig på 1970-tallet vært stengt for reketråling. Årsaken til at fjordene i sin tid ble stengt, var bl.a. stor innblanding av fisk og fiskeyngel i rekefangstene og konflikter mellom redskapsgrupper. Sorteringsristen kom inn i kystrekefisket fra 1990, og dermed ble bifangstproblematikken redusert vesentlig. I tillegg stenger Fiskeridirektoratets sjøtjeneste rekefelt dersom det blir for mye yngel av fisk og/eller reker i reketrålfangstene. Disse tiltakene bidrar til at deler av det grunnlaget som fjordene opprinnelig ble stengt på, er falt bort. Fiskeridirektoratet vurderer derfor muligheten for igjen å kunne høste av rekeressursene i disse to fjordene ved bruk av bunntrål.
En arbeidsgruppe nedsatt av Fiskeridirektøren har gjennomgått forvaltningen av kystrekefisket nord for 62° N. Arbeidsgruppen vurderte også åpning av Porsanger- og Tanafjorden for direktefiske etter reker med trål. I den anledning så direktoratet en mulighet for å innhente ny kunnskap. I brev av 18.09.2017 fra Fiskeridirektoratet til Havforskningsinstituttet ba direktoratet Havforskningsinstituttet komme med tilbakemelding om hvorvidt instituttet ønsket å være med i dette arbeidet, og evt. hva vi kunne bidra med. Dette resulterte i en prosjektbeskrivelse til direktoratet i oktober 2017 (oppdatert i mai 2018) med en plan for kartlegging av fjordøkosystemene i de to stengte fjordene, samt i en referansefjord hvor det hele tiden har pågått et aktivt reketrålfiske (Kvænangen). Fire dedikerte tokt ble gjennomført i 2018-2019 for å hente inn data. Disse dataene har blitt sammenstilt med data fra Havforskningsinstituttet sitt årlige kystressurstokt, som fra og med 2017 har økt antallet bunntrålstasjoner i Finnmark og Troms sammenlignet med tidligere år. Innsamlede data har videre blitt sammenstilt med allerede eksisterende data fra bunndyrsovervåking i Porsangerfjorden og data fra instituttets årlige kongekrabbetokt i Finnmarksfjordene. En foreløpig rapport ble sendt til Fiskeridirektoratet i slutten av oktober 2019. Denne versjonen er den endelige rapporten fra Havforskningsinstituttet sitt arbeid med kartlegging av fjordøkosystemene og fiskeriressursene i Tanafjorden, Porsangerfjorden og Kvænangen.
Det finnes få eller ingen uutnyttede rekebestander av kommersiell størrelse i Nord-Atlanteren (et mulig unntak er Øst-Grønland). Bortsett fra felt som er for små eller med for dårlig bunn til å tråles, og felt som er stengt eller forlatt pga. lav reketetthet, så utnyttes sannsynligvis alle kjente, trålbare rekefelt. I mangel på referanseområder upåvirket av tråling, har vi derfor lite kunnskap om hvordan et kommersielt fiskeri påvirker bestandsstrukturen og tettheten til rekebestander. Rekefiskere hevder bl.a. at rekefelt må tråles kontinuerlig («pløyes») for å gi gode fangster. Vi kjenner ikke til noen vitenskapelige studier spesifikt på reker som viser dette. En forklaring kan være at tråling roter opp bunnen og slik frigjør næringsstoffer fra sedimentet (Dounas mfl., 2005, 2007). De ufiskede rekebestandene i Tana- og Porsangerfjorden er derfor unike.
Kongekrabben har etablert seg i Finnmarksfjordene, og forvaltes i Porsanger- og Tanafjorden med mål om maksimalt bærekraftig langtidsutbytte. I 2018 ble det rapportert fanget 921 tonn i Porsangerfjorden og 160 tonn i Tanafjorden. Pga. mulige konflikter mellom teinefisket etter kongekrabbe og et fremtidig trålfiske etter dypvannsreke har vi tatt med et kapittel om kongekrabben i denne rapporten.
Havforskningsinstituttet har gjennomført overvåking av bunndyrsamfunnene i Porsangerfjorden med ujevne mellomrom (avhengig av prosjektfinansiering) for å studere effekten av den introduserte kongekrabben. På slutten av 2000-tallet ble studier på effekten av kongekrabbens beiting på bunnsamfunn gjennomført som del av EPIGRAPH-prosjektet (Pedersen mfl. 2018). De to utrålte fjordene presenterte en gylden mulighet for å studere effekten av fravær av bunntråling på bunndyrsamfunn (selv om ulik tilstedeværelse og forvaltning av kongekrabbe øst og vest for 26 ˚Ø muligens kunne gjøre en slik sammenligning vanskelig). Det viste seg imidlertid at den kommersielle reketrålen som ble benyttet under kartleggingstoktet, fanget bunndyr svært dårlig. Datatilfanget på bunndyr ble derfor mye mindre enn vi opprinnelig hadde sett for oss. Våre bunndyrresultater er derfor hovedsakelig basert på den tidligere overvåkingen i Porsangerfjorden som ble supplert med nye data fra et av toktene i 2019, samt resultater fra kystressurstoktet og noen videotransekt fra Mareano.
I tillegg til det pågående teinefisket etter kongekrabbe i Finnmarksfjordene (Sundet mfl. 2019), fiskes det nå også etter reker med teiner. Parallelt med tråltoktene høsten 2018 og våren 2019 gjennomførte Havforskningsinstituttet teineundersøkelser i de tre fjordene. Basert på tokt- og fiskeridata kommer instituttet med råd i denne rapporten vedrørende den videre utviklingen av teinefisket.
I prosjektet har vi inkludert deler av en tidligere prosjektskisse «Blandingsfiskeri som reguleringsmodell» (Anon. 2012). Vi har relatert dette til fiskeri i Porsanger- og Tanafjorden med fokus på rekefiske. I en totalvurdering må man også ta i betraktning at fjordtorskbestandene er på et historisk lavt nivå og vurdere bestandenes behov for mat når de skal bygges opp igjen. Vi har derfor inkludert en beskrivelse av prosjektet «Porsangerfjorden tilbake til livet» som ble initiert av Porsanger kommune i 2001. Hensikten med dette prosjektet er å få kunnskap om årsaker til den vedvarende kraftige reduksjonen av fiskebestandene i fjorden, samt å finne fram til tiltak som kan bringe bestandene tilbake til historiske nivåer. Våre råd for Porsangerfjorden har tatt hensyn til føringene som dette prosjektet gir.
Formålet med kartleggingen har vært å:
- Sammenligne de ufiskede rekebestandene i Porsanger- og Tanafjorden med rekebestanden i Kvænangen, hvor det hele tiden har foregått et kommersielt rekefiske, med hensyn på tetthet, utbredelse, størrelses- og stadiefordeling, og genetisk bestandsstruktur.
- Registrere artsrikdom og funksjonell biodiversitet, og eventuelle sårbare marine økosystemer i Tana- og Porsangerfjorden (overfor bunntråling), og sammenligne med Kvænangen.
- Studere blandingsfiske som reguleringsmodell for økt verdiskaping.
- Etablere et faglig grunnlag som kan bidra til å sikre en god rådgivning for et maksimalt bærekraftig langtidsutbytte (MSY) for marine ressurser i fjordene, som også tar hensyn til resten av økosystemet.
2 - Forsøksområde
2.1 - Porsangerfjorden
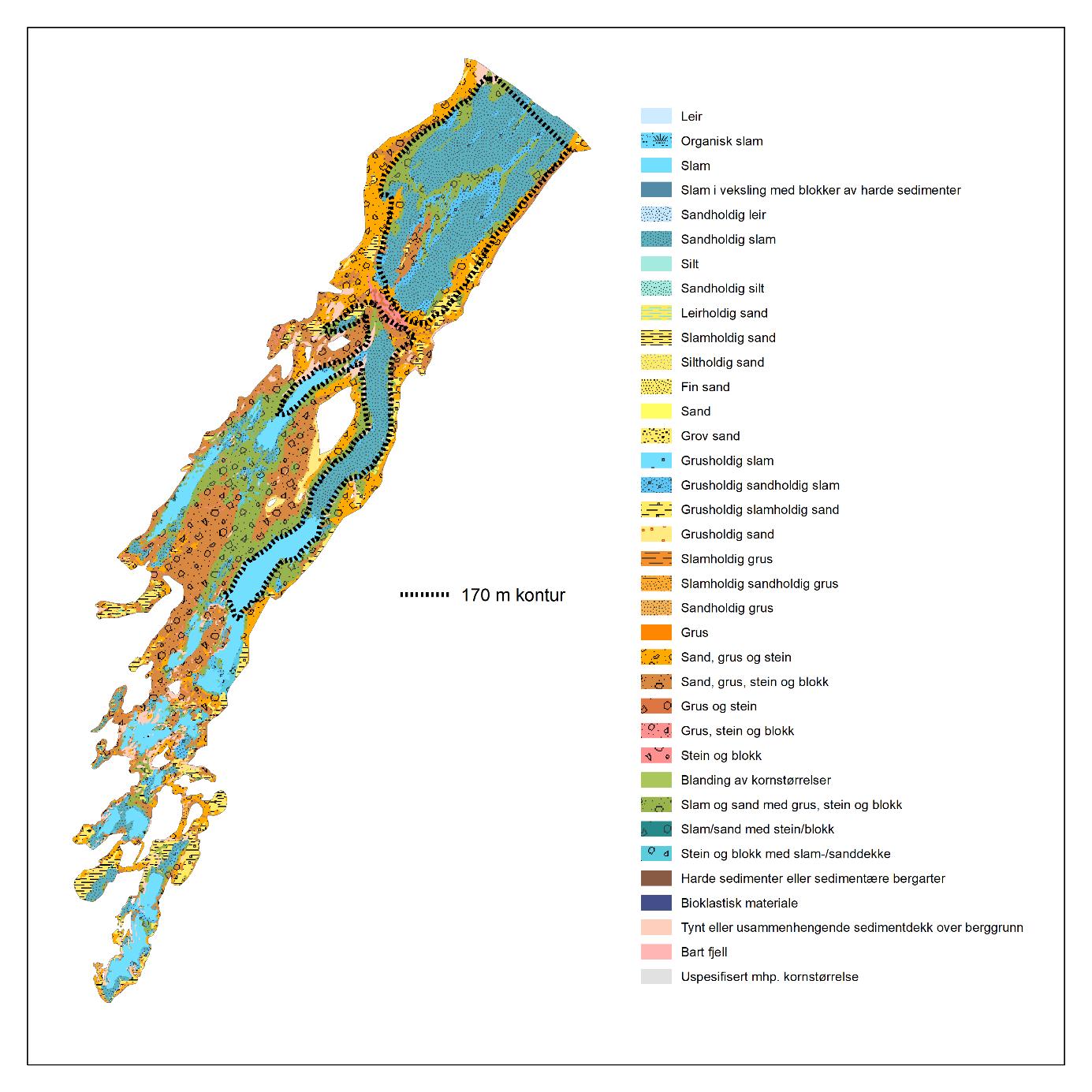
Porsangerfjorden er en av de største fjordene i Nord-Norge med en lengde på 120 km (Figur 2.1). Vestsiden av fjorden er grunn, østsiden har dypere bassenger, mens den indre delen innenfor Reinøya preges av en kompleks bunntopografi. I den østlige indre delen, innenfor et gruntområde, ligger de to bassengene Østerbotn og Rotenbottenjarga med dyp ned til 110 m, mens den vestre indre delen har en mer variert batymetri med dyp ned til 90 m. Den midterste delen av fjorden er adskilt fra den ytre delen av en 100 m dyp terskel («Ryggen») rett nord for Lille-Tamsøya, mens den ytre delen er åpen ut mot Barentshavet. Her finnes den dypeste delen av fjorden, med dyp ned til 300 m. Den innerste delen av fjorden innenfor Reinøya er etablert som nasjonal laksefjord. En detaljert kartlegging av bunnforhold i hele Porsangerfjorden har blitt gjennomført av Norges geologiske undersøkelse (NGU) (geo.ngu.no/kart/marin/MARINEKART.html?kart=4&latlon=74.55,29.6&zoom=5#) (Figur 2.2). De kommersielle rekefeltene i fjorden finnes i de dypere delene øst i den midterste delen og i den ytre delen. Det var et aktivt rekefiske i Porsangerfjorden fra 1931 (Hjort og Ruud 1938) frem til stengingen av fjorden tidlig på 1970-tallet. Det finnes to oppdrettsanlegg for laks og ørret i fjorden.
Selv om vi her beskriver tre deler av Porsangerfjorden, indre, midterste og ytre, vil vi i denne rapporten som regel bruke betegnelsen «ytre Porsangerfjorden» om området utenfor Reinøya. Pga. de spesielle miljøforholdene i området innenfor Reinøya (avsnitt 4.2), presenteres resultater fra Porsangerfjorden separat for den indre og ytre delen (Figur 3.2). I de tilfellene der vi med «ytre Porsangerfjorden» mener området utenfor terskelen «Ryggen», spesifiseres dette.
2.2 - Tanafjorden
Tanafjorden er mindre enn Porsangerfjorden og ligger ca. 80 km øst for denne (Figur 2.1). Tanafjorden er ca. 64 km lang. Den ytre delen har et stort dypområde som strekker seg inn i Barentshavet, med dyp ned til 250 m. Den indre delen forgreiner seg i tre fjordarmer med en mer kompleks bunntopografi. Fjorden har ingen terskler. Hele fjorden er en nasjonal laksefjord og har derfor ingen lakseoppdrettsanlegg. Rekefeltene dekker det meste av den ytre delen og strekker seg innover i de tre fjordarmene. Fjorden hadde et aktivt reketrålfiske fra 1931 (Hjort og Ruud 1938) frem til stengingen av fjorden tidlig på 1970-tallet.
2.3 - Kvænangen
En referansefjord måtte oppfylle to kriterier: 1) den måtte ha et aktivt reketrålfiske og 2) den måtte ha noenlunde like klima- og miljøforhold som Tana- og Porsangerfjorden. Laksefjorden, Varangerfjorden og Kvænangen ble vurdert som aktuelle kandidater. Laksefjorden ville ha vært ideell der den ligger mellom de to ufiskede fjordene, men rekefisket i denne fjorden er ofte stengt pga. for høy yngelinnblanding. Varangerfjorden er mer en havbukt enn en fjord, og mye tokttid ville ha gått med til steaming frem og tilbake til dette området. Valget falt derfor på Kvænangen, som er en av Nord-Norges viktigste rekefjorder. Kvænangen ligger 165 km sørvest for Porsangerfjorden og er ca. 60 km lang (Figur 2.3). Fjorden har ingen ytre terskel. Den ytre delen av fjorden forgreiner seg i to fjordarmer, hvorav den nordlige er Kvænangen, mens den sørlige er Reisafjorden. Reisafjorden ble inkludert i referansefjorden «Kvænangen» for å øke det totale arealet. De to fjordene samlet vil heretter bli omtalt som kun «Kvænangen». Den ytre delen av fjorden har en renne med dyp ned til 400 m. Innover i de to fjordarmene finnes flere dype bassenger. Fjordbunnen har ellers en kompleks bunntopografi. Den innerste delen av Kvænangen innenfor Sørstraumen, en av Norges sterkeste tidevannsstrømmer, er en nasjonal laksefjord. Hele Reisafjorden er også en nasjonal laksefjord. Rekefeltene i fjorden finnes i alle de dype bassengene. Kvænangen og Reisafjorden har hatt et aktivt rekefiske siden 1931 (Hjort og Ruud 1938), med årlige landinger på rundt 180 tonn de siste 10 år (2009-2019). Det finnes tolv lakseoppdrettsanlegg i sjø i de to fjordene.
3 - Tokt
3.1 - Kartleggingstokt med reketrål og -teiner i 2018 og 2019
3.1.1 - Stratasystem
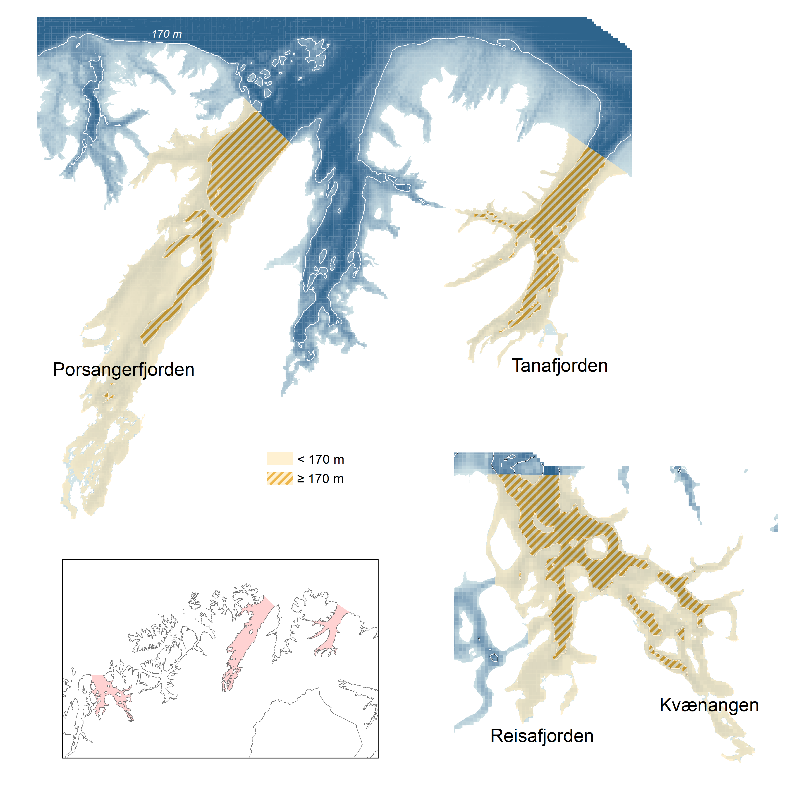
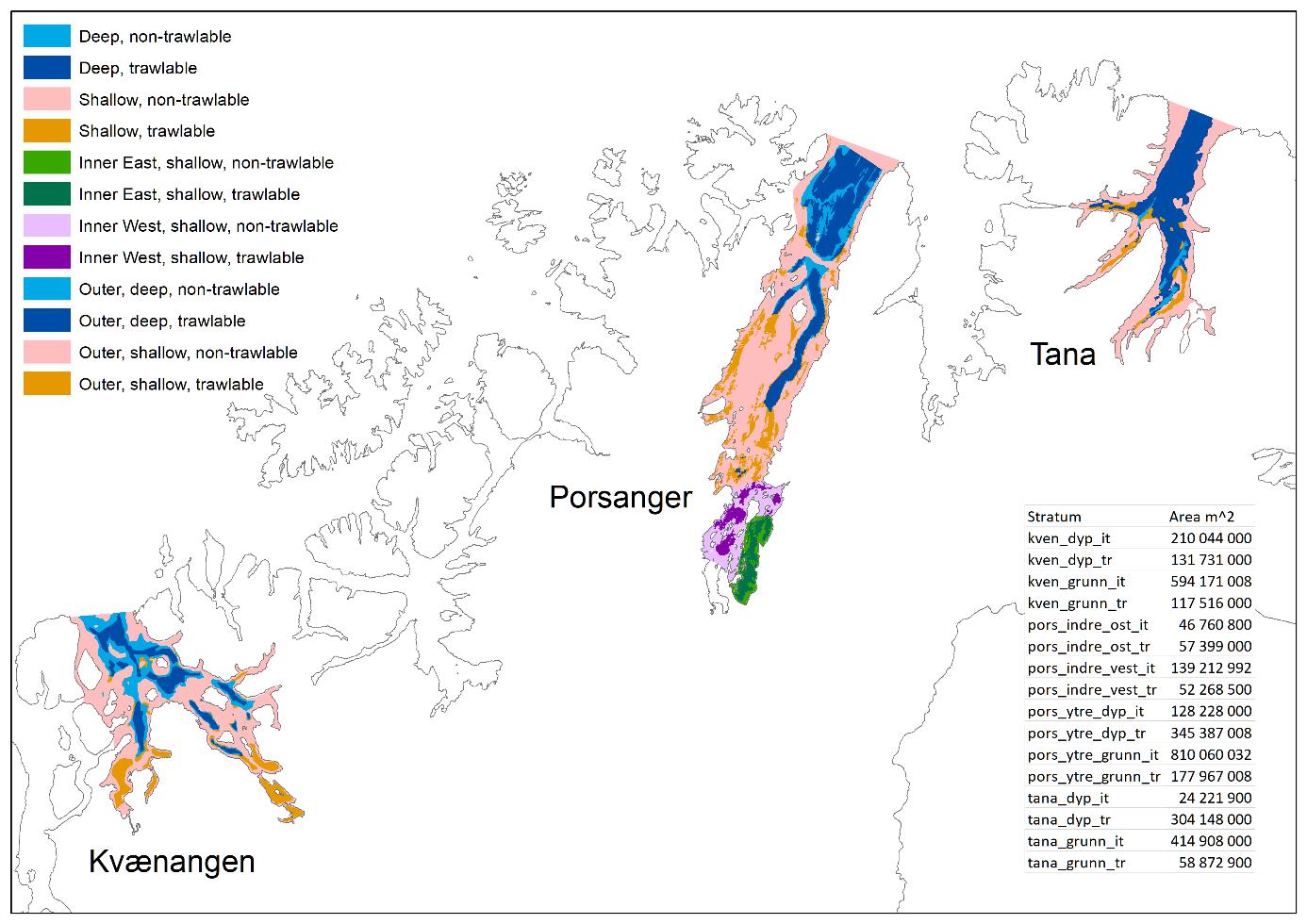
Et stratasystem for de tre fjordene ble utarbeidet basert på følgende kriterier: 1) områder vi vil ha ut resultater/rapporter for (f.eks. enkeltfjorder), 2) økosystemforhold (f.eks. det indre området av Porsangerfjorden (se under)), 3) områder grunnere og dypere enn 170 m (nåværende trålegrense) og 4) trålbare og ikke-trålbare områder (for å kunne trekke ut tilfeldige trålstasjoner).
Inndeling av Porsangerfjorden i trålbare og ikke-trålbare områder ble basert på Norges geologiske undersøkelse (NGU) sin kartlegging av bunnforhold (Figur 2.2), der slam og sand ble vurdert som trålbare sedimenter, mens alt annet (berggrunn, grus og stein) ble vurdert som ikke-trålbart. Områder som ikke ble dekket av kartleggingen (hovedsakelig svært grunne områder), ble antatt å være ikke-trålbare. Tilsvarende kart finnes ikke for Kvænangen og Tanafjorden, så her brukte vi kartlagte rekefelt sammen med skyggerelieffkart fra Fiskeridirektoratets karttjeneste til å identifisere trålbare områder. Både trålbare og ikke-trålbare områder ble delt i områder dypere og grunnere enn 170 m (Figur 3.1). Dette resulterte i fire forskjellige strata i Tanafjorden og Kvænangen (Figur 3.2). I Porsangerfjorden ble den indre delen delt i en vestlig og en østlig del da tettheten av dypvannsreke varierer enormt fra den grunne, vestlige delen hvor det nesten ikke finnes reke, til den østlige, dypere delen med en eksepsjonell høy reketetthet (se under). Den vestlige og østlige indre delen ble igjen delt i hhv. trålbare og ikke-trålbare områder, slik at Porsangerfjorden totalt ble delt inn i åtte strata.
Ved beregning av rekebiomasse ble gjennomsnittlig reketetthet ganget opp med areal av trålbar bunn, da vi regner med at det aller meste av dypvannsreken står på bløtbunn. For bunnfisk, derimot, ganget vi gjenommsnittlig tetthet med arealet av hele fjordbunnen da vi regner med at de forskjellige bunnfiskartene lever i mange forskjellige typer habitat (i større og mindre grad). Rekebiomassen kan derfor være underestimert, mens biomassen av de forskjellige bunnfiskartene sannsynligvis er overestimert.
3.1.2 - Gjennomføring av tråltokt med «Katla»
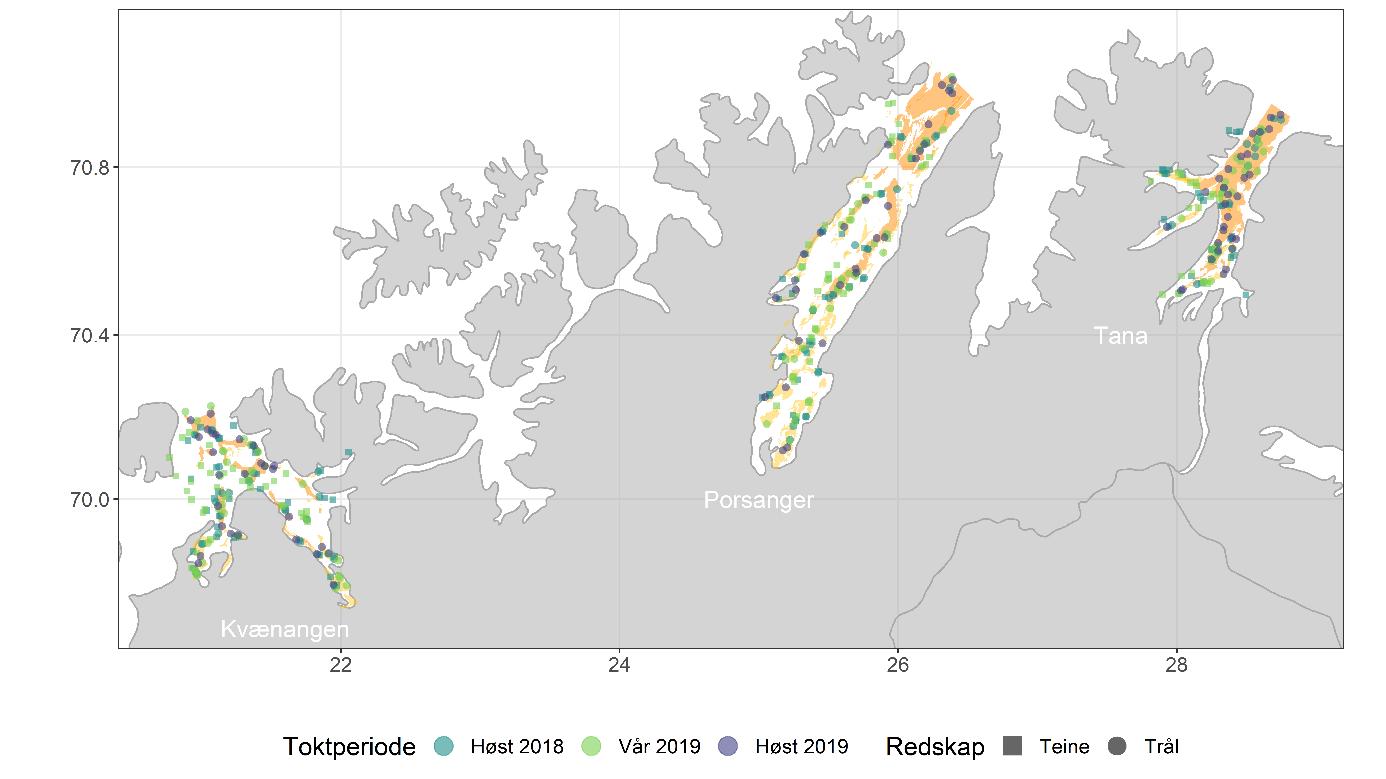
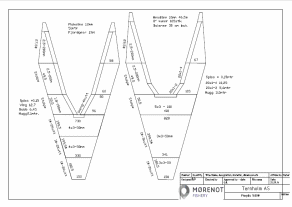
Det ble gjennomført tre tråltokt: høsten 2018, våren 2019 og høsten 2019 (Figur 3.3, Tabell 3.1). Det ble satt av sju dager til tråling i hver fjord på hvert tokt. Reketråleren «Katla» (LK7560, 14,95 m) ble leid inn til å gjennomføre tråltoktene. Trålen som ble brukt, var en reketrål 1600 ma uten fiskerist, med 15 mm masker i fiskeposen og 35 m sveiper (Figur 3.4). Bunngiret var en bobbinslenke med rullende elementer med diameter på 15 cm. Sveipebredden ble satt til 20 og 25 m for hhv. reke og bunnfisk (lengde-uavhengig sveipebredde for fisk).
Trålstasjonene ble trukket ut tilfeldig, der antallet stasjoner per stratum var proporsjonalt med arealet av stratumet (Figur 3.2). Ute på sjøen ble bunnforholdene for hvert trålhal inspisert før tråling. Om bunnforholdene var dårlige, flyttet vi trålhalet opptil 1 km vekk fra den opprinnelige posisjonen. Alternativt kunne stasjonen forkastes og erstattes av en stasjon på en liste med tilfeldig uttrukne ekstrastasjoner. Tauetid var 15 minutter og tauehastighet var 1,5-1,7 knop. Følgende ble registrert på hver trålstasjon:
- Totalvekt og antall av all fisk, alle rekearter og alle bunndyr i trålen.
- Individprøve (lengde, kjønn, modning, vekt) av inntil 20 stk. torsk, hyse, sei, vanlig ulke, brosme, gråsteinbit, flekksteinbit, vanlig uer, kveite og rødspette.
- Otolitt- og mageprøver ble tatt av inntil 10 individer av artene torsk, hyse, sei og vanlig uer.
- Lengdemåling og stadiebestemmelse (kjønn og modningsstadium av hunnreker) av en tilfeldig rekeprøve på opptil 300 reker.
- På de to tråltoktene i 2019 ble bunntemperatur målt vha. en temperaturlogger festet på headlinen.
| Porsangerfjorden | Tanafjorden | Kvænangen | |||||
| Toktnr. | Dato | Stasjoner | Dato | Stasjoner | Dato | Stasjoner | Totalt |
| 2018006 | 01.-07.10. | 25 | 26.10.-01.11. | 19 | 17.-23.10. | 24 | 68 |
| 2019824 | 04.-10.04. | 28 | 27.03.-02.04. | 22 | 18.-24.03. | 32 | 82 |
| 2019825 | 09.-15.10. | 28 | 17.-23.10. | 29 | 29.09.- 06.10. | 27 | 84 |
3.1.3 - Gjennomføring av teinetokt med «Henriette» og «Rind»
Det ble gjennomført to teinetokt, høsten 2018 og våren 2019, parallelt med tråltoktene med «Katla» (Figur 3.3, Tabell 3.2). Sjarken «Henriette» (LK4631, 10,31 m) ble leid inn til å gjennomføre første del av toktet høsten 2018 (Porsangerfjorden) og hele toktet våren 2019. Fiskeridirektoratets sjøtjeneste stilte fartøyet «Rind» (LEXO, 21,67 m) til disposisjon for gjennomføring av andre og tredje del av teinetoktet høsten 2018 (Kvænangen og Tanafjorden). Målet med toktene var å 1) sammenligne fangstrate mellom teiner og reketrål, og 2) undersøke utbredelsen av dypvannsreke på ikke-trålbar bunn.
Teinene som ble brukt, var reketeiner (70x50x75 cm med plastkalver) designet til bruk i den innerste delen av Porsangerfjorden (Figur 3.5). De var dekket med netting (maskevidde 10 mm) og veide 3,5 kg. Blytau ble montert i bunn for å gjøre dem tyngre (4,8 kg). Inngangsåpningen hadde en diameter på 12,5 cm, og laksefôr ble brukt som agn. Etter erfaringer i løpet av toktet i Porsangerfjorden høsten 2018 ble det montert 4 kg ekstra blytau i bunnen for å gjøre teinene enda tyngre og mer stabile. Teinene var satt sammen i lenker à 8 stykker. En temperaturlogger ble festet til hver lenke. Den opprinnelige planen var å la teinene stå ute ett døgn; i praksis varierte ståtiden mellom ett og to døgn. Data ble registrert per teine:
- Totalt antall dypvannsreker. Vi fikk ikke registrert totalvekt av rekene da vekten ikke fungerte i sjøgang.
- Lengdemåling (CL) av opptil 20 reker.
- All bifangst (art/artsgruppe, antall).
Den opprinnelige planen var at det skulle settes teiner i alle trålposisjoner samt på tilfeldig utvalgte posisjoner på ikke-trålbar bunn, til sammen 8 lenker (stasjoner) per dag. I løpet av toktet høsten 2018 ble dette redusert til 6 lenker per dag pga. tidsbegrensning, og stasjoner på ikke-trålbar bunn ble nedprioritert. Vi mistenkte raskt at teinene var for lette til å bruke på strømsterke områder, og teinestasjonene her ble derfor kuttet. Tidsbegrensning, strøm og til dels mye vind og bølger gjorde altså at vi ikke klarte å gjennomføre kartleggingen med teiner i alle de på forhånd uttrukne posisjonene.
| Porsangerfjorden | Tanafjorden | Kvænangen | |||||
| Toktnr. | Dato | Stasjoner | Dato | Stasjoner | Dato | Stasjoner | Totalt |
| 2018006 | 30.09.-06.10. | 29 | 24.-30.10. | 21 | 16.-23.10. | 37 | 87 |
| 2019824 | 02.-10.04. | 42 | 27.03.-02.04. | 21 | 17.-24.03. | 31 | 94 |

3.2 - Fjord i Nord (FiN) tokt med R/V Kristine Bonnevie i Porsangerfjorden, mai 2019
3.2.1 - Gjennomføring
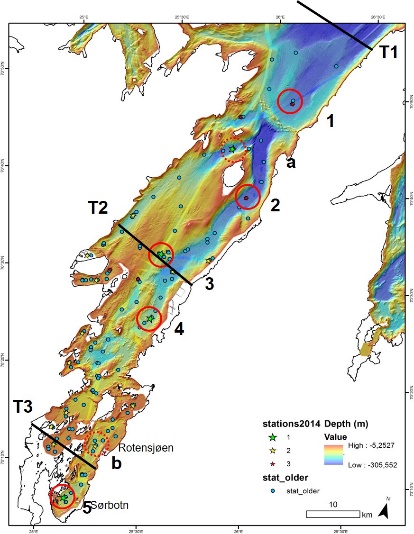
Et Fjord i Nord (FiN) tokt med R/V Kristine Bonnevie ble gjennomført i Porsangerfjorden 16.-21. mai 2019. Målet med toktet var å kartlegge økosystemet og studere forandringer i faunasammensetningen i den indre, midtre og ytre delen av fjorden før en eventuell åpning for reketrålfiske.
Totalt 18 stasjoner ble dekket. På noen av stasjonene ble flere redskap benyttet. Tre transekter i den ytre, midtre og innerste delen av fjorden (T1, T2, T3) ble valgt ut (Figur 3.6). På disse tre transektene ble det samlet inn data på vannmiljø (CTD med vannhentere) og zooplankton (WP2). Det ble tatt fem stasjoner med boxcorer (infauna og sediment) og sju trålhal med hhv. bomtrål og Campelen-trål (bunnfisk og bunndyr). Posisjonene til stasjonene samsvarte med posisjoner fra tidligere bomtrålundersøkelser i fjorden, samt faste bunntrålstasjoner på kystressurstoktet. Pelagisk tråling ble gjennomført med Harstad-trål under transittetapper (tre hal).
3.3 - Havforskningsinstituttets årlige kystressurstokt
3.3.1 - Gjennomføring
Kystressurstoktet gjennomføres årlig i oktober og november. I 2017 ble antallet bunntrålstasjoner økt for å få en bedre dekning av rekefeltene i fjordene i Troms og Finnmark. Kystressurstoktet bruker en Campelen-trål med maskevidde i fiskeposen på 21 mm. Standard tauetid er 30 minutter og tauehastighet er 3 knop. Følgende registreres på alle bunntrålhal på kystressurstoktet:
- Totalvekt og antall av all fisk og alle rekearter i trålen.
- Lengdemåling og aldersbestemmelse av et representativt utvalg av fisk.
- Lengdemåling og stadiebestemmelse (kjønn og modningsstadium av hunnreker) av en tilfeldig rekeprøve på ca. 300 reker.
4 - Fysisk oseanografi
4.1 - Strøm
Kartene i dette kapittelet med havstrømmer og saltholdighet langs bunnen og i overflaten viser kvartalsvise gjennomsnitt fra 2019 og er basert på resultater fra en hydrodynamisk modell som bruker ROMS med en horisontal oppløsning på 160 m x 160 m. Disse fjordmodellene dekker Kvænangen/Porsangerfjorden og Tanafjorden / det østlige Finnmark i separate modellrutenett. Kystmodellen som gir input langs de åpne grensene, NorKyst800, er forklart i Asplin mfl. (2020). NorKyst800 og fjordmodellene (160 m x 160 m) utgjør Havforskningsinstituttet sine viktigste havmodeller for beskrivelse av det fysiske miljøet i kystsonen.
Bunnstrømmen i Kvænangen og Porsangerfjorden var ikke så sterk i 2019, men det var mye strøm i den ytre delen av Tanafjorden, særlig inn fjorden langs vestkysten (Figurer 4.1, 4.2, 4.3). Strøm i overflaten varierte mellom årstidene og mellom fjordene i 2019. I Kvænangen var det til dels sterk strøm i overflaten ut fjorden langs nordsiden i 2. og 3. kvartal. I Porsangerfjorden var det også en tilsvarende overflatestrøm ut av fjorden i 2. og 3. kvartal. Tanafjorden skilte seg ut blant de tre fjordene ved at det i hele 2019 var en sterk overflatestrøm ut langs fjordens østside grunnet avrenningen fra Tanaelven.
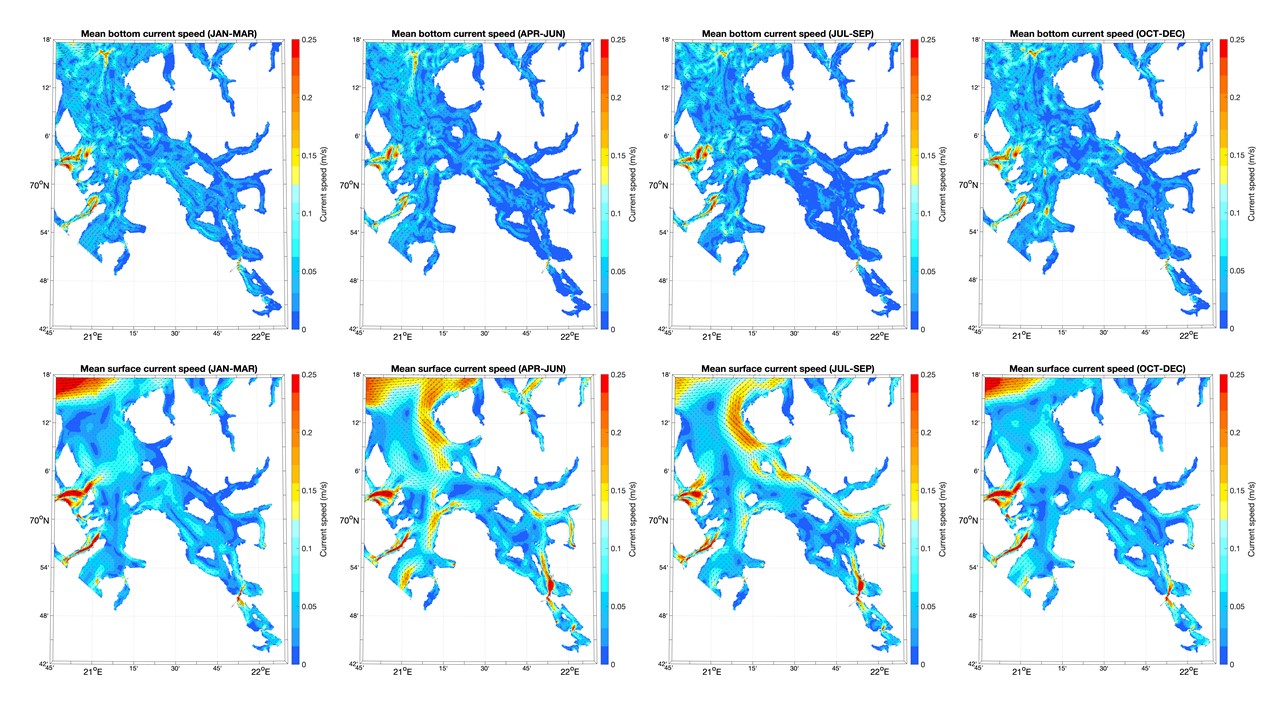
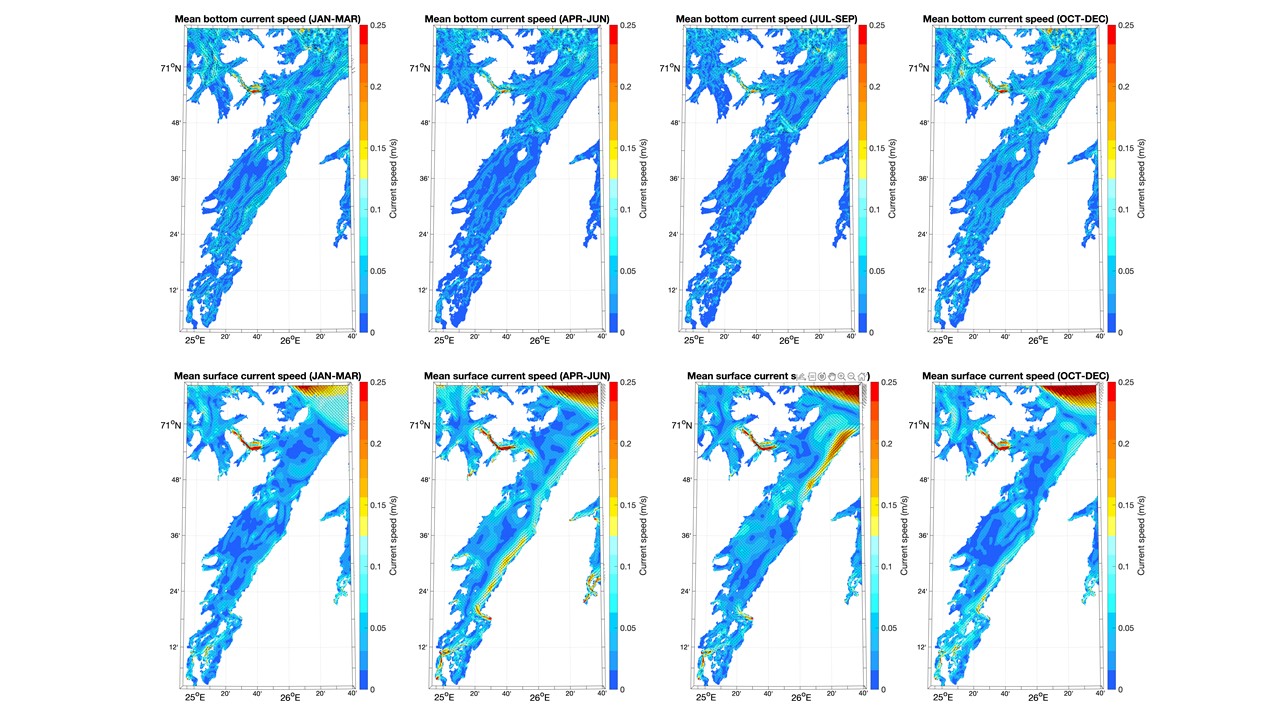
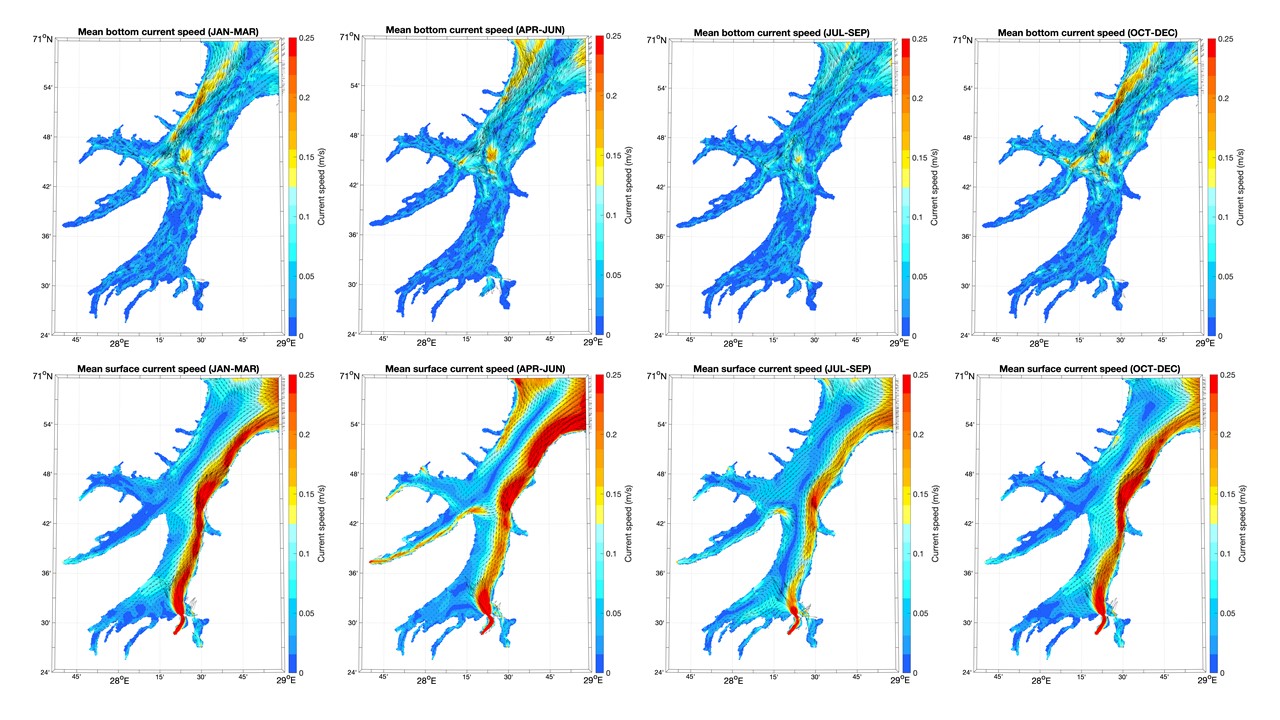
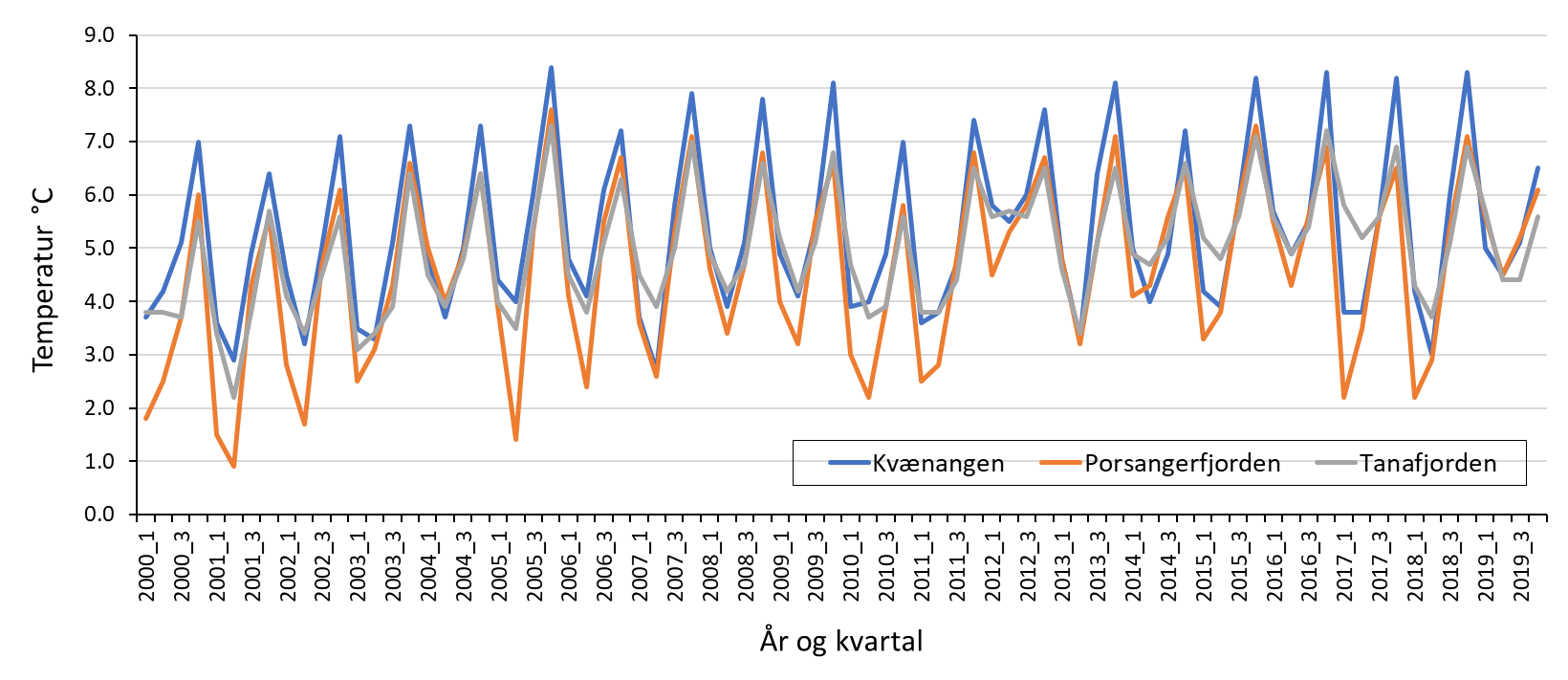
4.2 - Temperatur, saltholdighet og oksygeninnhold i vannet
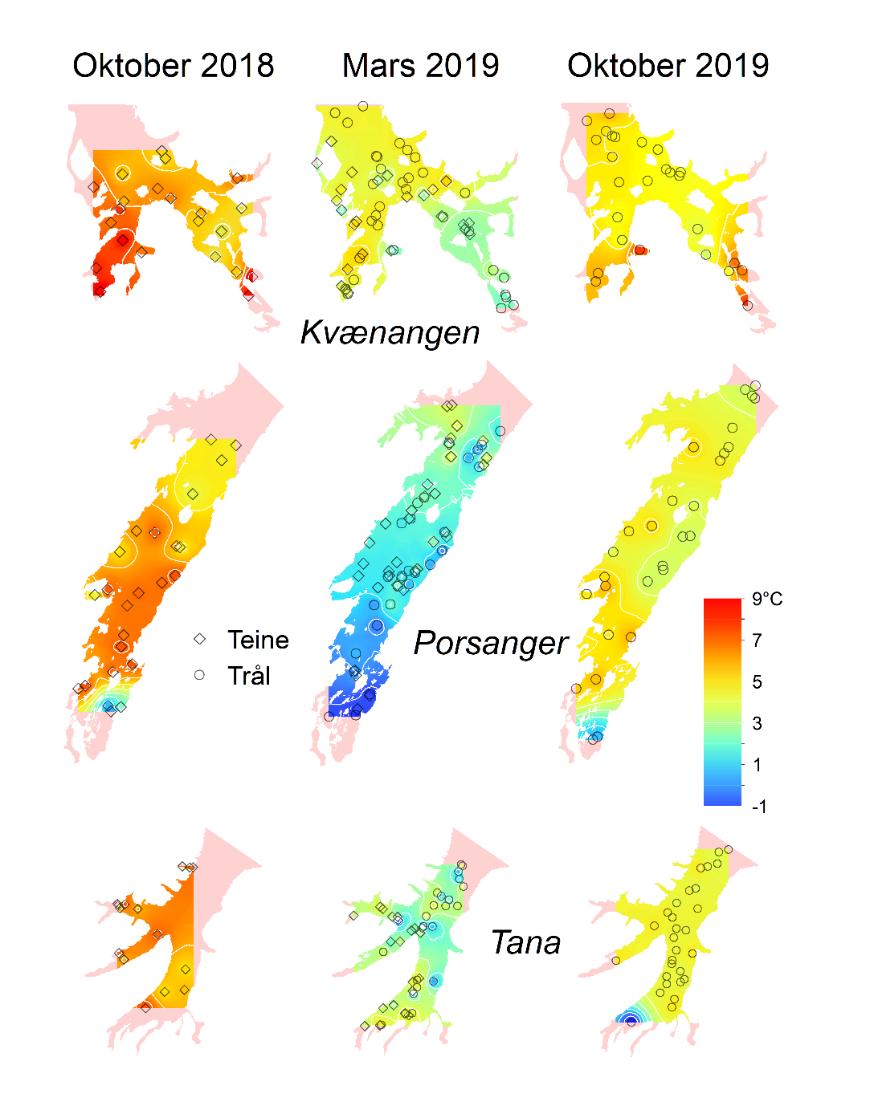
Temperaturmålingene fra oktober 2018, mars/april 2019 og oktober 2019 viste at bunntemperaturen varierte mye mellom og innad i fjordene, og mellom årstider og år (Figur 4.4). Bunntemperaturen var høyest i oktober, opp i 9 °C enkelte steder, og lavest i mars. I disse to årene var Porsangerfjorden den kaldeste av de tre fjordene. De laveste temperaturene ble målt i de indre østlige bassengene i Porsangerfjorden, hvor det var 0 til -1 °C hele året. Også innerst i Tanafjorden ble det i oktober 2019 målt temperaturer ned i -1 °C. Modellerte kvartalsvise bunntemperaturer fra de tre kartleggingsfjordene tilbake til 2000 viser noe av det samme som målingene i 2018 og 2019, nemlig at bunnvannet er kaldest i 1. og 2. kvartal (Figur 4.5). Denne tidsserien viser også det samme som Figur 4.4 indikerer, nemlig at av de tre fjordene er Porsangerfjorden den kaldeste og Kvænangen den varmeste fjorden.
Fra Porsangerfjorden finnes det oseanografiske data fra FiN-toktet i mai 2019 og fra tokt med Håkon Mosby sommeren 2015 (Stramska mfl. 2018). Målinger i hele vannsøylen langs snitt både på tvers (Figur V1.1 i Vedlegg 1) og langs (Figur V1.2) av Porsangerfjorden (upubliserte data) viser at i august 2015 lå bunntemperaturen i hele den midtre og ytre delen av fjorden på 5-6 °C. I mai 2019 ble det målt temperaturer ned i -1 °C i de innerste bassengene, mens bunntemperaturen i resten av fjorden lå på 3-5 °C (Figur V1.3). De forskjellige målingene indikerer at bunntemperaturen i Porsangerfjorden er lavest i 1. kvartal og høyest i 4. kvartal, slik tidsserien fra 2000 til 2019 også viser (Figur 4.5). Saltholdigheten i Porsangerfjorden i august 2015 og mai 2019 var høyest på bunn i de dypeste områdene i munningen av fjorden, lavere innover i fjorden og lavest i overflaten (Figurer V1.2, V1.4). Dette henger sammen med at relativt salt og varmt atlanterhavsvann (dypvann) og kystvann (overflaten) (34-34.5‰) kommer inn Porsangerfjorden i nordvest og møter ferskvannspåvirket overflatevann. Det oppstår en klassisk fjordgradient i saltholdighet der verdiene øker utover i fjorden i overflaten samt at noe av det lavsaltholdige vannet blandes ned inne i fjorden. Dette medfører også at saltholdigheten i dypvannet inne i fjorden er lavere enn på samme dyp lenger ute.
Oksygennivået ved bunn i august 2015 var høyere i den midterste delen av fjorden og minket utover i fjorden (Figur V1.2). Målinger fra mai 2019 viste noe av det samme, overflatevannet samt bunnvannet i de innerste østlige bassengene hadde det høyeste oksygeninnholdet, mens bunnvannet i fjordmunningen hadde det laveste innholdet av oksygen (Figur V1.5).
Vi har ingen målinger av saltholdighet eller oksygeninnhold fra Kvænangen eller Tanafjorden. Modellerte kvartalsvise nivåer fra 2019 viser at saltholdigheten ved bunn varierte gjennom året, spesielt i Kvænangen, med de høyeste verdiene i 2. og 3. kvartal (Figur V1.6). I de grunnere områdene av Kvænangen lå saltholdigheten i bunnvannet i januar-mars på ca. 33,7 ‰, mens bunnvannet i hele Tanafjorden holdt mer enn 34 ‰ gjennom hele året, med unntak av området ved munningen av Tanaelven.
5 - Bestander
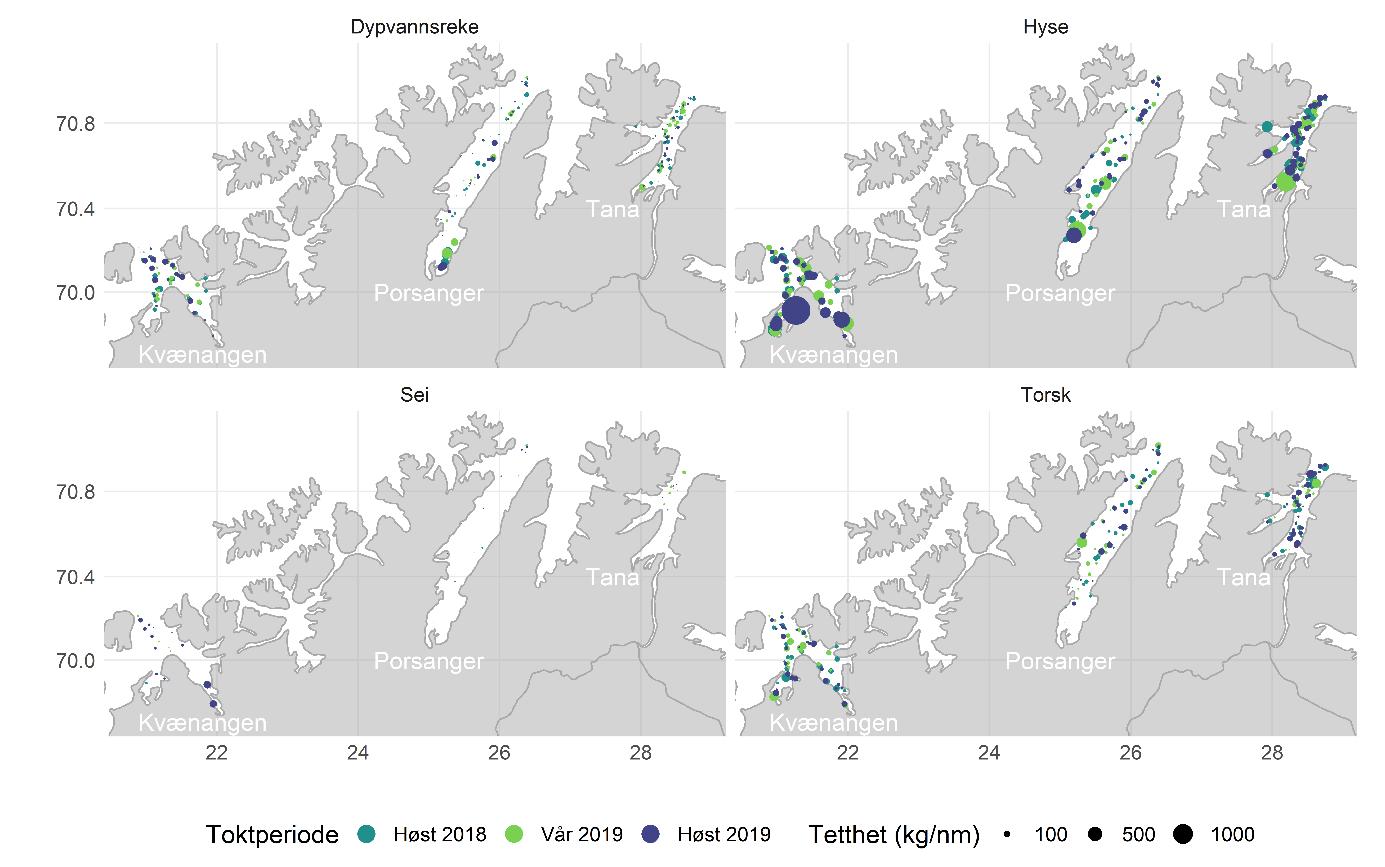
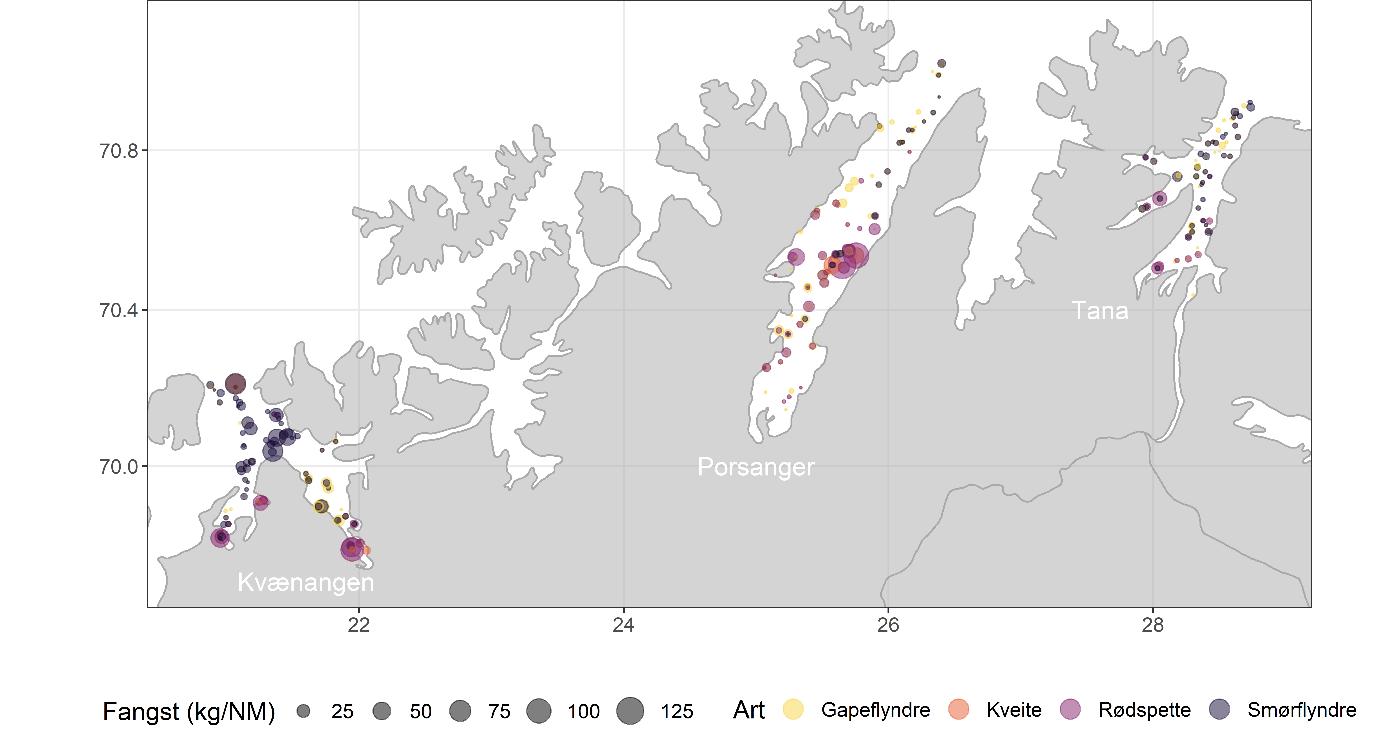
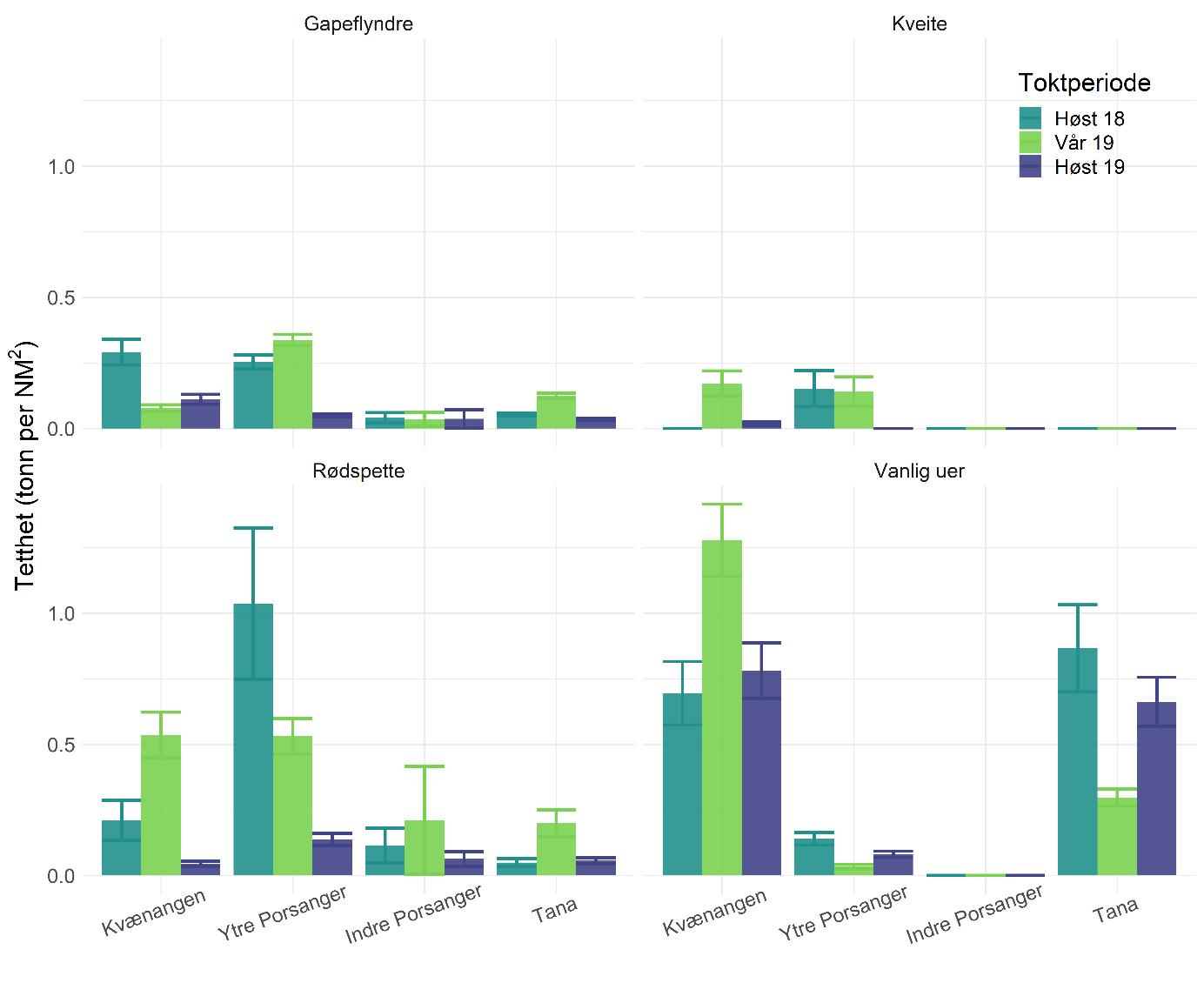
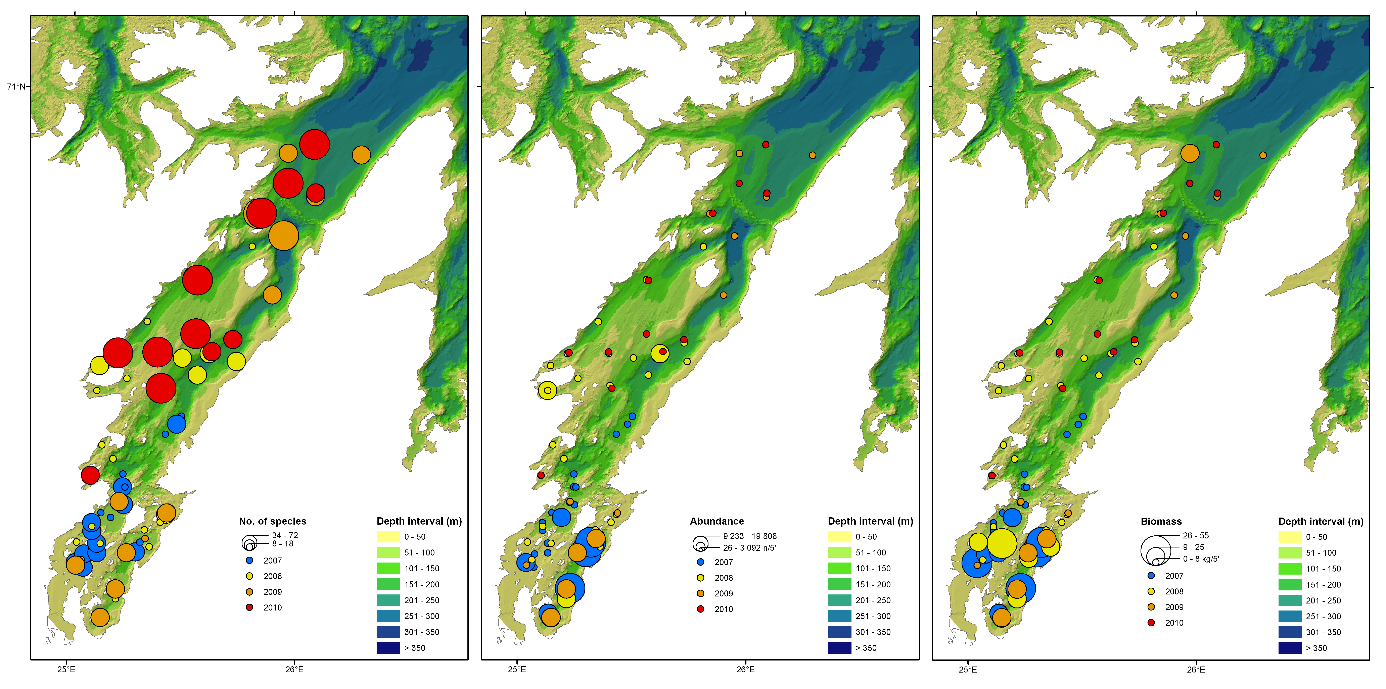
Total 158 forskjellige arter og artsgrupper ble funnet i løpet av de tre tråltoktene med «Katla» (Tabell 5.1). Den høyeste artsdiversiteten ble funnet i Kvænangen, mens den indre delen av Porsangerfjorden har flest unike arter. Artene som ble oftest registrert, er hyse og torsk som begge i gjennomsnitt var til stede på 94 % av alle stasjoner. Dypvannsreke ligger på en tredjeplass med 85 %, og deretter følger brunalger, gapeflyndre og sild. Dette gjenspeiles på oversiktskartet over registrerte fangster (Figur 5.1) som viser at dypvannsreke, hyse og torsk forekom i nesten alle områder av de tre fjordene i betydelige mengder. Spesielt hyse forekom i veldig høye tettheter i alle fjordene og toktperiodene (Figur 5.2). Dette blir tydelig når man sammenligner forekomstene av disse tre artene med andre arter som f.eks. sei. Sei er også en relevant kommersiell art i undersøkelsesområdet, men ble registrert på relativt få stasjoner og hovedsakelig i små mengder.
Estimert tetthet av hyse i alle fjordene er betydelig høyere enn tettheten av alle andre arter (Figur 5.2). Dypvannsreke og torsk ligger omtrent på samme nivå med noe regionale forskjeller, mens sei finnes hovedsakelig i Kvænangen og der kun med høyere forekomster høsten 2019. Et unntak fra dette mønsteret er indre Porsangerfjorden. Tetthet av dypvannsreke i den østlige, indre delen av Porsangerfjorden er mye høyere enn i alle andre undersøkte områder, mens omtrent ingen reker ble funnet i den vestlige, indre delen. Fordelingen er motsatt for hyse og torsk, som kun ble registrert i den vestlige delen.
Detaljerte resultater for dypvannsreke, hyse, sei, torsk og andre bunnfisk og bunndyr blir presentert i de følgende avsnittene.
| Toktperiode | Fjord | Antall arter |
| Høst 2018 | Kvænangen | 63 |
| Tanafjorden | 46 | |
| Ytre Porsangerfjorden | 65 | |
| Indre Porsangerfjorden | 47 | |
| Vår 2019 | Kvænangen | 66 |
| Tanafjorden | 45 | |
| Ytre Porsangerfjorden | 39 | |
| Indre Porsangerfjorden | 26 | |
| Høst 2019 | Kvænangen | 55 |
| Tana | 38 | |
| Ytre Porsangerfjorden | 47 | |
| Indre Porsangerfjorden | 20 | |
| Totalt | Alle | 158 |
5.1 - Dypvannsreke
5.1.1 - Beskrivelse av arten
Dypvannsreke (Pandalus borealis) er utbredt fra tempererte til arktiske strøk (Shumway mfl. 1985, Bergstrøm 2000). I Nordøst-Atlanteren finnes den fra Skagerak og Nordsjøen i sør til nord for Svalbard. De store kommersielle bestandene i norske farvann finnes i Skagerak, Norskerennen og i Barentshavet. Langs hele norskekysten lever reken på en mosaikk av større og mindre bløtbunnsområder (Zimmermann mfl. 2019). De store tetthetene finner man på bløtbunn, men dypvannsreken kan også trekke inn på mer steinete bunn (Shumway mfl. 1985). I Nordøst-Atlanteren lever reken i vann med temperaturer fra 7-9 °C i sør til under null innerst i Porsangerfjorden. Dypvannsreken skifter kjønn fra hann til hunn etter at den har reprodusert som hann i 1-2 sesonger. Etter paring om høsten bærer hunnene eggene mellom svømmeføttene inntil de klekker neste vår. En hunn kan reprodusere flere ganger. I nordlige, kalde områder reproduserer en del av hunnrekene bare annethvert år. De pelagiske larvene driver med havstrømmene i opptil tre måneder (avhengig av temperaturen) før de bunnslår. Forskjellige aldersgrupper (størrelser) av reker finnes gjerne i forskjellige områder og på forskjellige dyp. Juvenile reker står ofte grunnere enn eldre reker (Shumway mfl. 1985). Mange steder er det observert at hunner trekker inn på grunnere vann før eggklekkingen om våren. Da reken må skifte skall for å vokse, mangler den harde strukturer som alder kan leses ut ifra. Klare modaltopper i lengdefrekvensfordelinger for de 2-4 første aldersgruppene gjør imidlertid aldersbestemmelse mulig.
5.1.2 - Rekebestandene i Kvænangen, Porsanger- og Tanafjorden
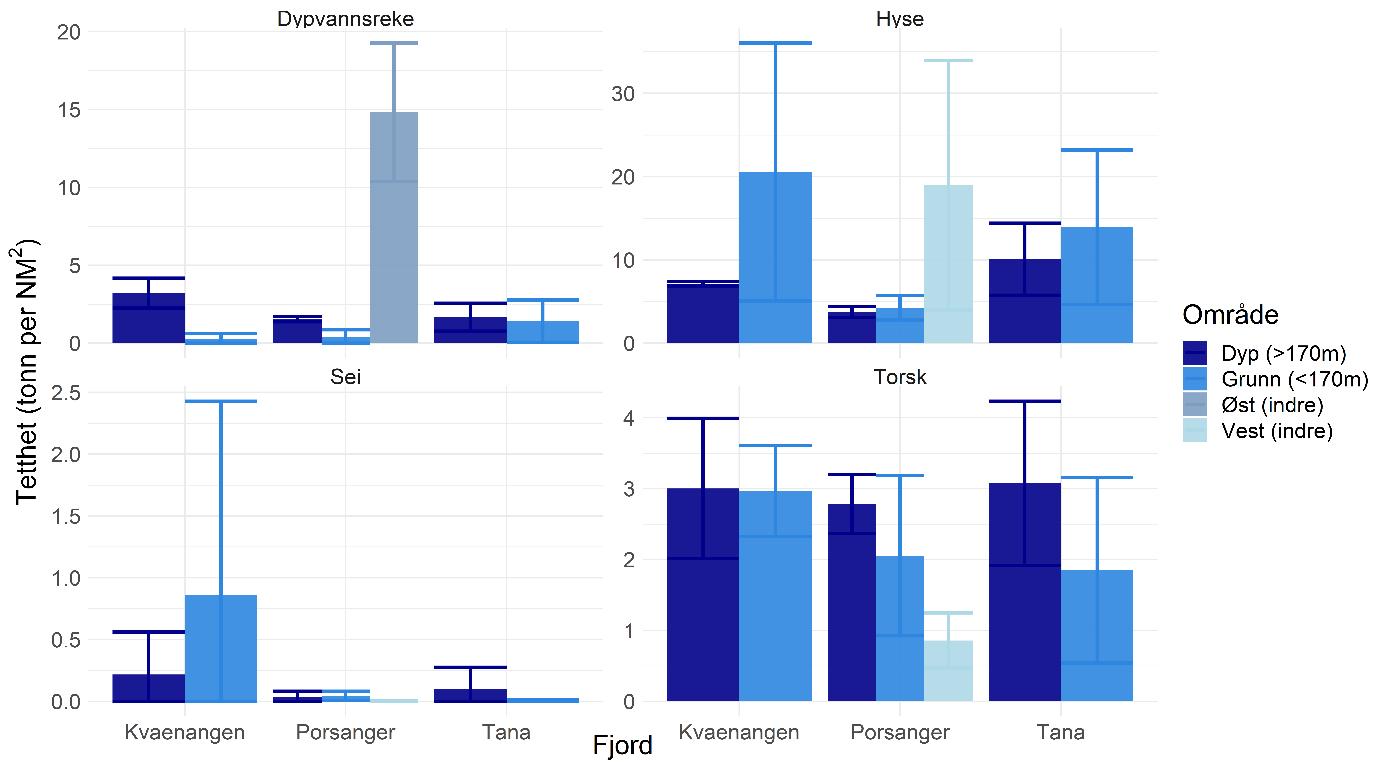
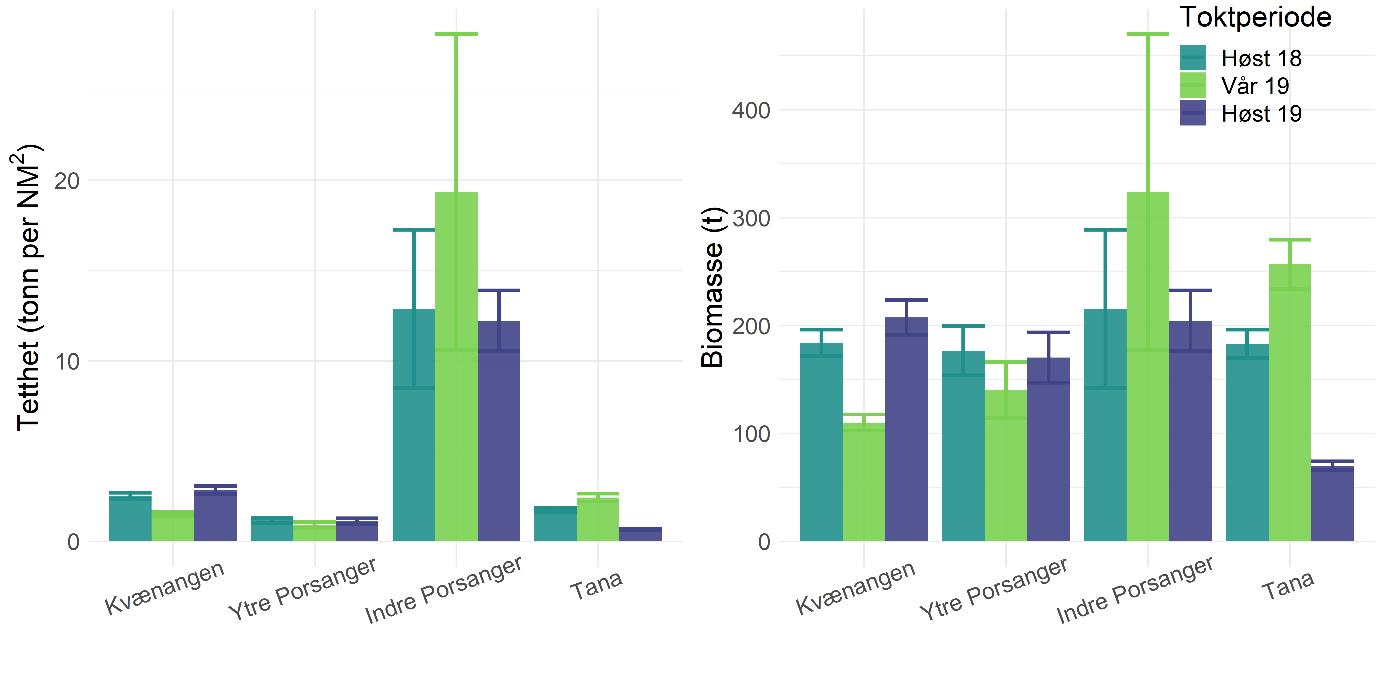
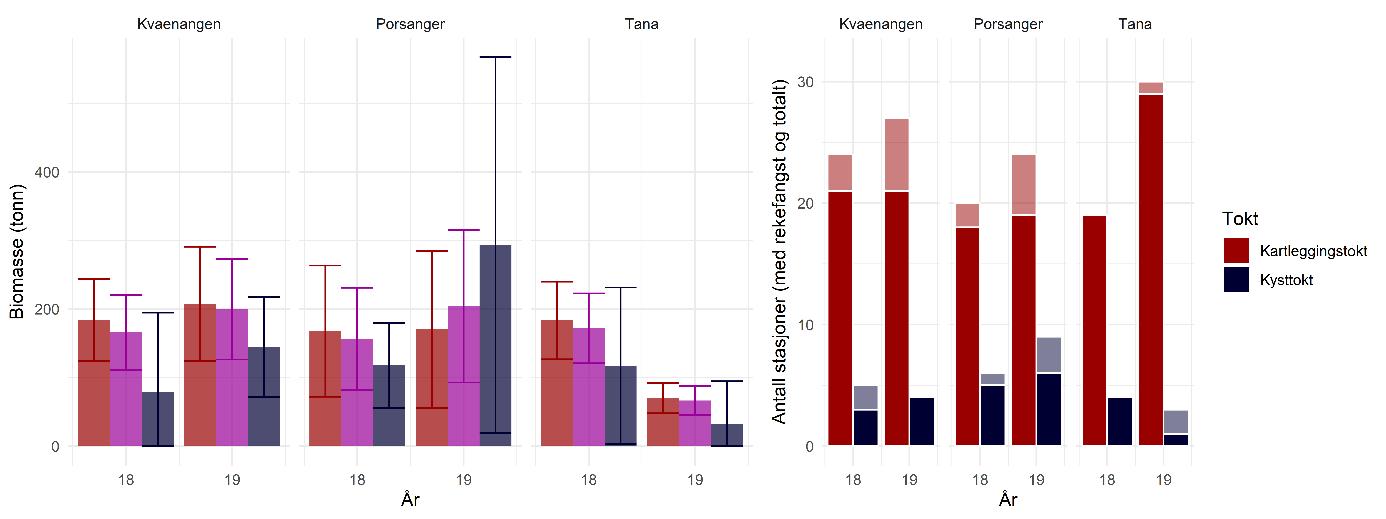
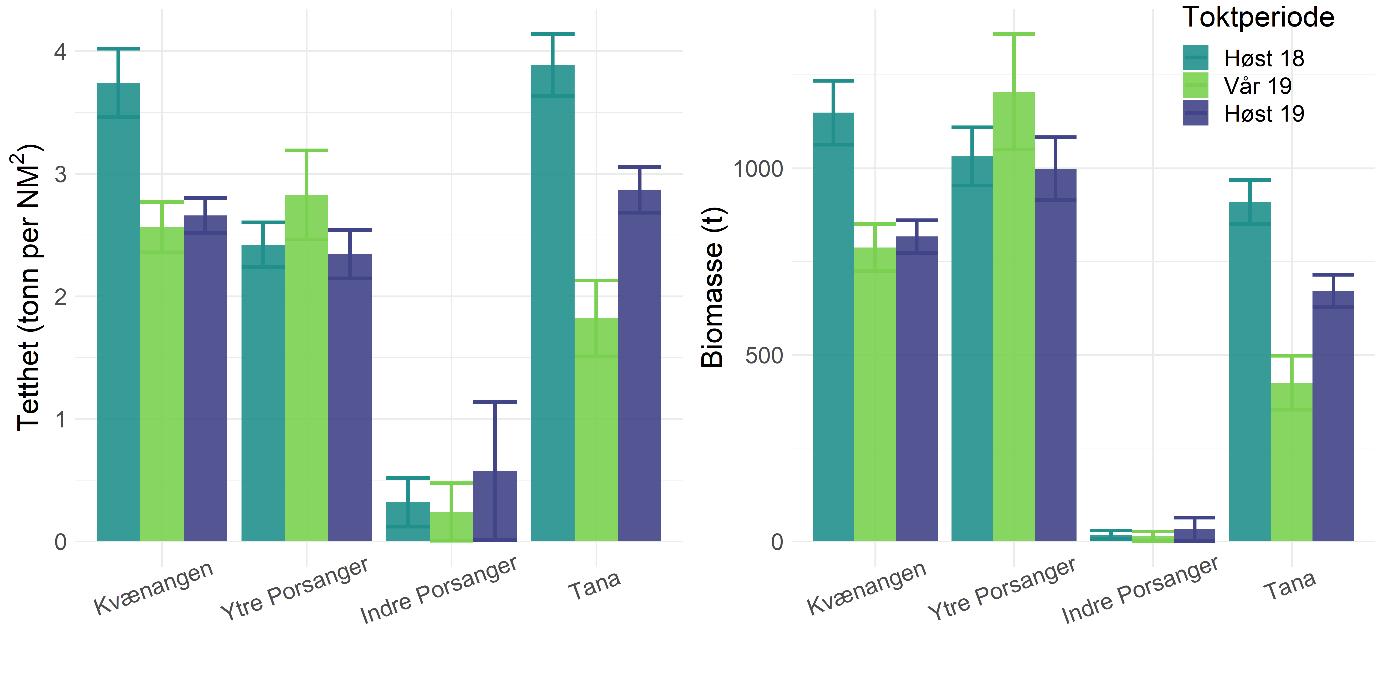
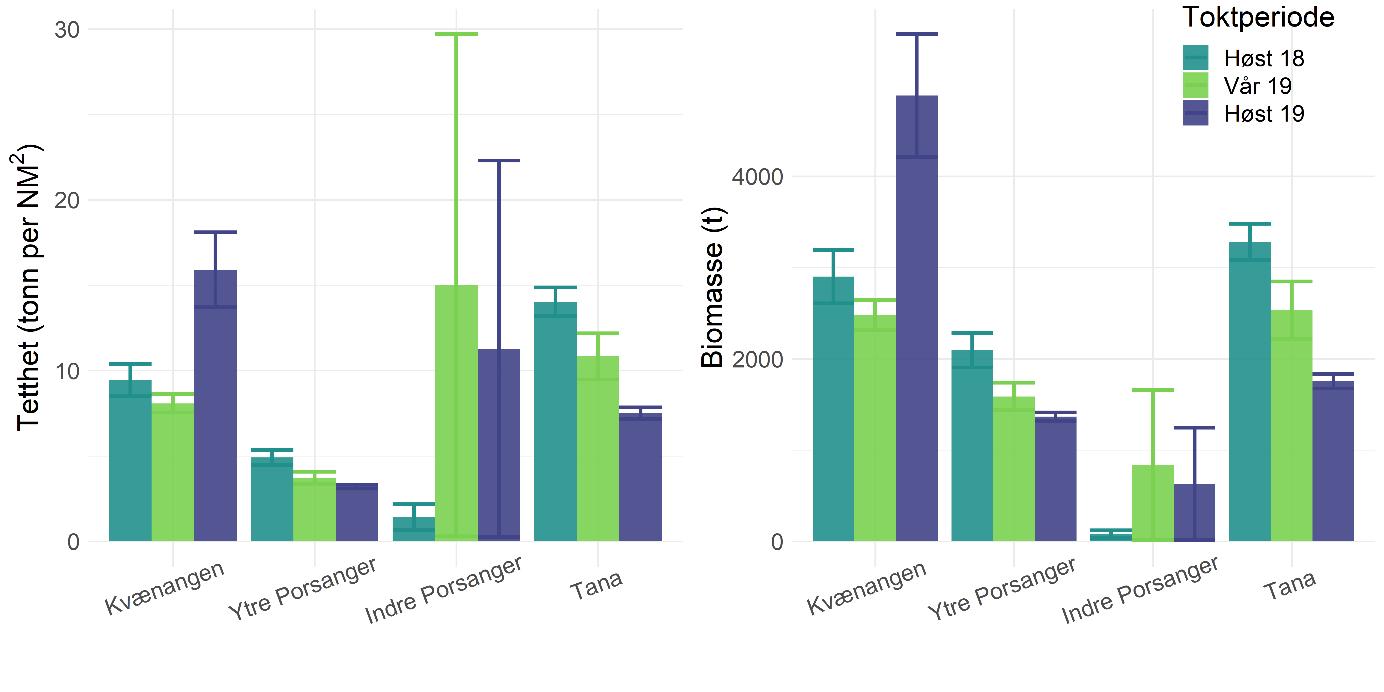
Rekebestandene i Kvænangen, ytre Porsangerfjorden og Tanafjorden er i stor grad sammenlignbare, mens rekene i indre Porsangerfjorden skiller seg tydelig ut. Her registrerte vi både den høyeste tettheten (Figur 5.3) og en avvikende bestandsstruktur (avsnitt 5.1.5) gjennom alle de tre toktperiodene. Total biomasse er et produkt av tetthet og arealstørrelse, og da det finnes reker kun i et lite område i de to østlige bassengene i den indre delen av Porsangerfjorden, resulterer det i en biomasse i dette området som er på samme nivå som i de andre fjordområdene. Det begrensede arealet betyr i tillegg få trålstasjoner (ca. to per toktperiode) siden antall stasjoner per stratum var proporsjonalt med areal av stratumet, og som en følge av dette har vi den største usikkerheten rundt estimatene fra indre Porsangerfjorden. I de tre andre fjordområdene er den estimerte reketettheten på samme nivå, med den høyeste gjennomsnittsverdien i Kvænangen og den laveste i ytre Porsangerfjorden. Biomassen i de ulike fjordene er estimert som tetthet ganger areal, noe som resulterer i omtrent de samme rekebiomassene i Kvænangen, ytre Porsangerfjorden og Tanafjorden.
Habitatstørrelse for reker baserer seg på estimert areal av trålbar bløtbunn (Figur 3.2), dvs. at det antas at reker finnes kun på bløtbunn, selv om de kan forekomme i lavere tetthet også på andre bunntyper. Det betyr at den totale rekebiomassen sannsynligvis er et underestimat av den faktiske biomassen. I tillegg foreligger det en mye mer nøyaktig bunnkartlegging av Porsangerfjorden enn av Tanafjorden og Kvænangen, noe som kan gjøre det problematisk å sammenligne biomasser direkte mellom fjordene. Tall på totalbiomasser må derfor tolkes med varsomhet, og det er viktig å huske på de underliggende begrensninger. Generelt er tetthet en bedre indikator på bestandsforekomst fordi biomassen da oppgis i forhold til et standard areal. Det er mye bedre å sammenligne tetthet mellom forskjellige områder enn biomasse.
Tetthet og biomasse endrer seg over tid i alle fjordområder, men uten en unik trend (Figur 5.3). Det finnes sesongsvingninger i alle områder, spesielt tydelig og signifikant i Kvænangen og Tanafjorden, men svingningene går i motsatt retning: Mens biomassen er høyere om våren enn om høsten i Tanafjorden og indre Porsangerfjorden, er det motsatt i Kvænangen og ytre Porsangerfjorden. I tillegg finnes det en tydelig nedgang i biomassen i Tanafjorden fra høsten 2018 til høsten 2019.
Tettheten i Kvænangen er lik eller høyere enn i de to andre fjordene, og det gjelder også individstørrelse, men generelt finnes det ikke noen stor forskjell mellom den fiskete rekebestanden i Kvænangen og de ufiskete bestandene i Porsanger- og Tanafjorden. Det er kontraintuitivt ikke kun fordi vi sammenligner en fisket med en ufisket bestand, men også fordi undersøkelser av genetisk bestandsstruktur (avsnitt 5.1.4) og livshistorie (avsnitt 5.1.5, 5.16) viser at det dreier seg om forskjellige populasjoner med potensielt store forskjeller i dynamikk. Det er uklart hvordan dette kan forklares. Den veldig høye tettheten i indre Porsangerfjorden er sannsynligvis knyttet til de spesielle miljø- og økosystemforhold der, og ikke minst det at det nesten ikke finnes rekepredatorer der. Tetthet av stor fisk, som torsk, er imidlertid veldig likt på tvers av de andre fjordområdene, og de lavere tetthetene i Porsanger- og Tanafjorden kan derfor ikke forklares ved et høyere predasjonstrykk der. Det betyr at det må være andre faktorer enn predasjon som påvirker rekene i disse tre fjordene, mest sannsynlig en kombinasjon av tetthetseffekter og forskjell i økosystemproduktivitet og bunnsamfunn, potensielt knyttet til forskjeller i økosystemsammensetning (f.eks. forekomst av kongekrabbe).
5.1.3 - Tidsserier
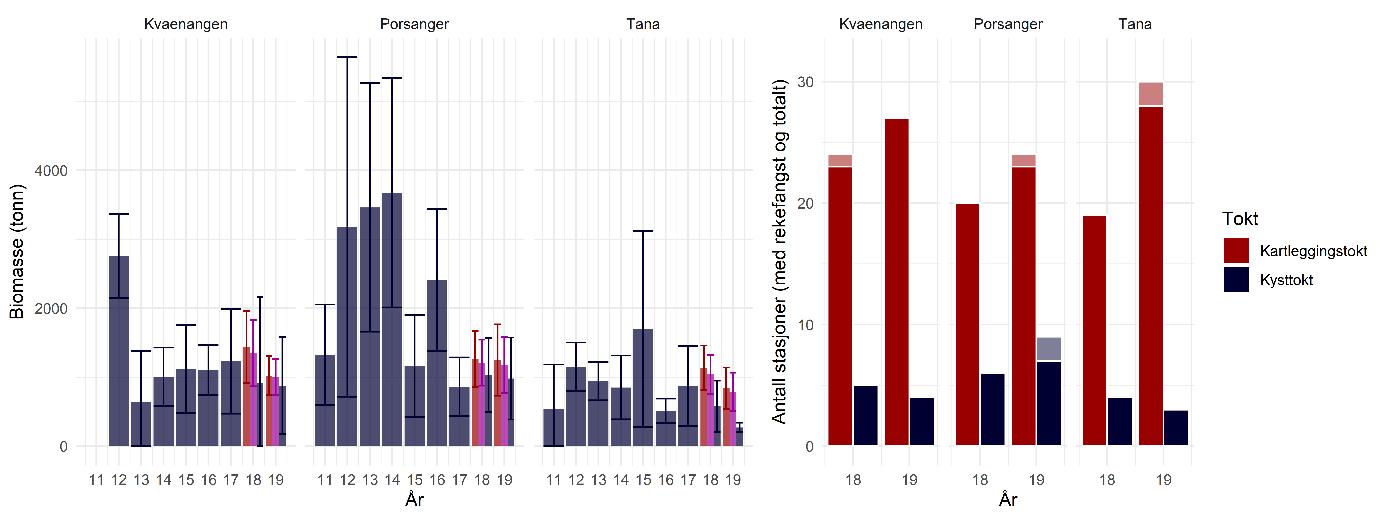
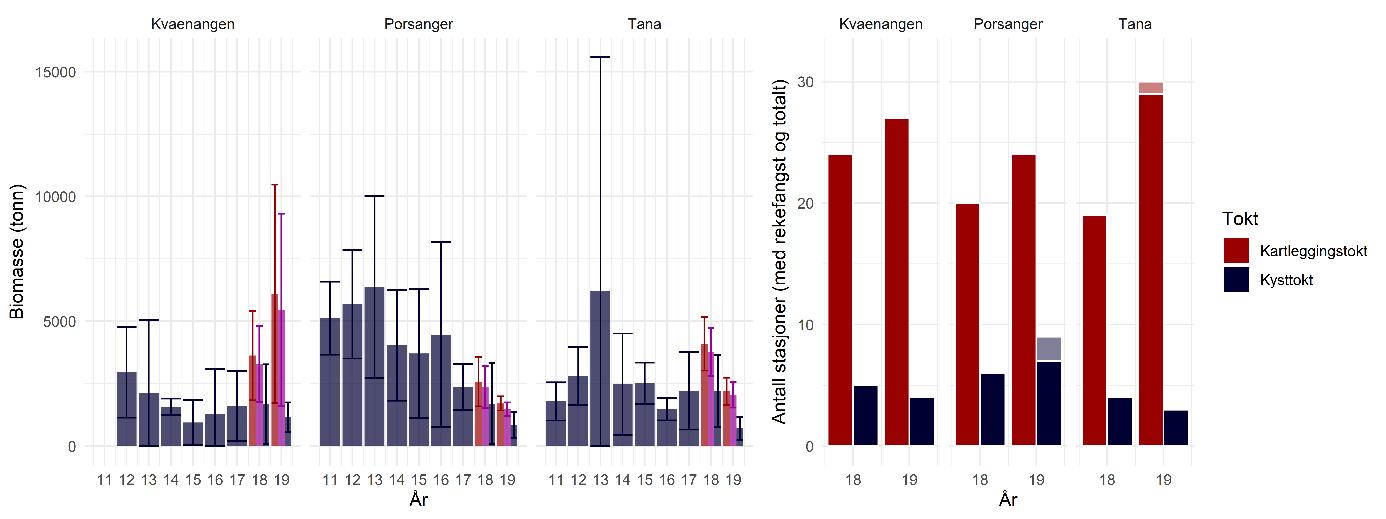
Biomasseestimater av dypvannsreke basert på kartleggingstokt- og kysttoktdata er i stor grad sammenlignbare og viser ingen statistisk signifikant forskjell (Figur 5.4). Det årlige kystressurstoktet dekker de kartlagte fjordene hver høst, men med færre stasjoner per fjord. Resultatene fra kartleggingstoktet er derfor sannsynligvis mer nøyaktige pga. det høye antallet stasjoner og prøver, og fordi dette toktet var målrettet mot dypvannsreke.
Før 2017 var dypvannsreke ikke en prioritet art i kystressurstoktet og datagrunnlaget er derfor ikke bra nok til å lage en troverdig bestandsindeks lenger tilbake i tid. Selv om antall stasjoner i fjordene har blitt økt fra og med 2017 for å forbedre dekningen av rekefelt, er det fortsatt et lavt antall per år og fjord sammenlignet med kartleggingstoktet (Figur 5.4, høyre side). I tillegg ble forskjellig metodikk brukt på de to toktene: på kartleggingstoktet ble det benyttet en modifisert reketrål mens kysttoktet bruker en Campelen-trål og høyere trålfart. Uansett samsvarer resultatene for høsten 2018 og høsten 2019 da toktene overlappet i tid, noe som indikerer at kystressurstoktet sannsynligvis gir et representativt bilde av bestandstilstanden til reke. Men med et lavt antall stasjoner vektes hver enkelt prøve mer, noe som betyr mer variasjon og usikkerhet. For at kystressurstoktet skal kunne brukes til å bygge opp en tidsserie for overvåking av bestandsutviklingen i fremtiden, anbefales det derfor en økning av prøvetakingsinnsatsen målrettet mot dypvannsreke.
5.1.4 - Genetisk bestandsstruktur
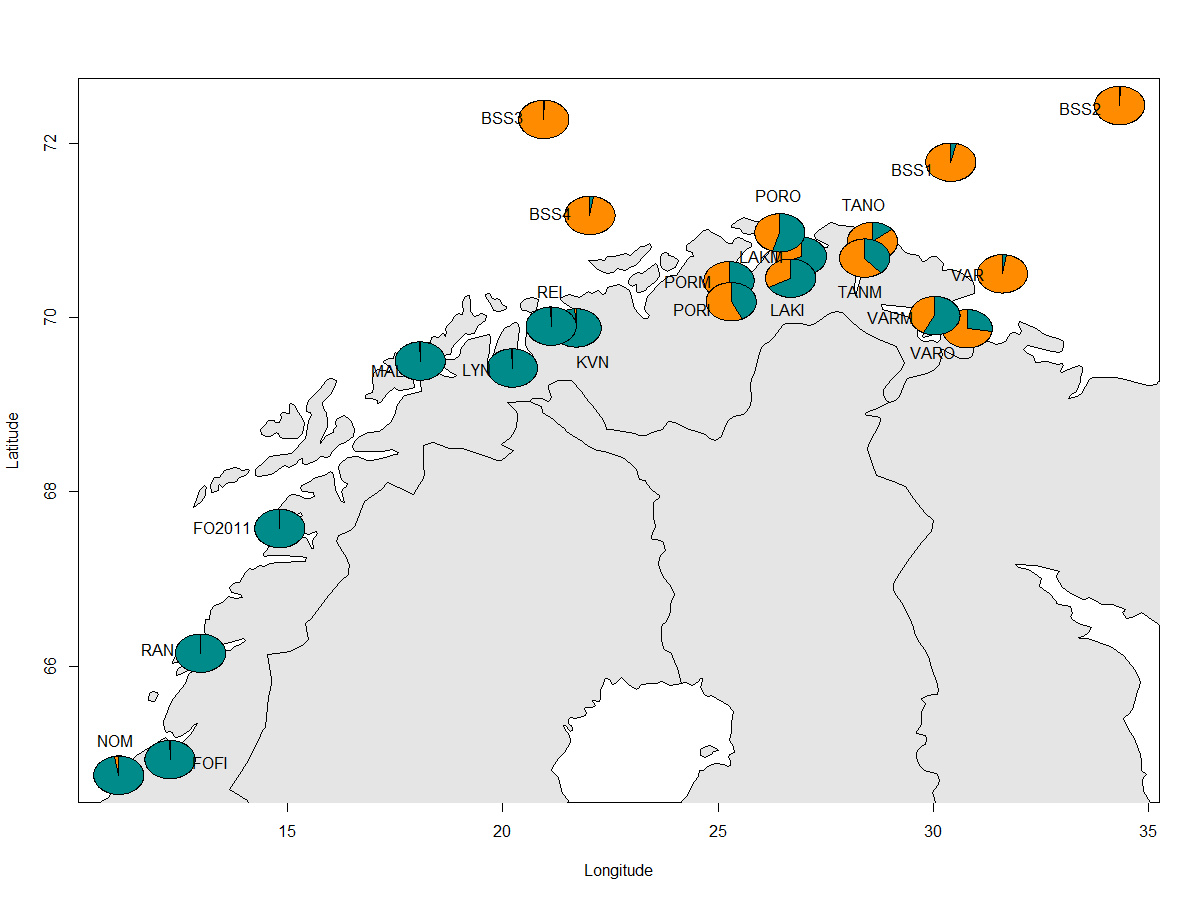
Den genetiske bestandsstrukturen til dypvannsreke langs norskekysten (Trøndelag til Varanger) ble kartlagt ved hjelp av 10 mikrosatellitter (Hansen 2020). Rekeprøver ble samlet inn fra forskjellige tokt og ved hjelp av rekefiskere i perioden 2010-2018. Prøver av kystreker ble analysert sammen med rekeprøver fra det sørlige Barentshavet for å sette resultatene inn i en større sammenheng (Figur 5.5). Det ble funnet signifikante forskjeller mellom reker i Barentshavet og reker fra kyststrekningen Trøndelag til Troms (Tabell 5.2), og de respektive områdene viste hver sin homogene trend ved å fordele seg i hver sin gruppe. Fjordene i Finnmark viste en mer blandet trend ved å inneholde reker både fra kysten (Trøndelag-Troms) og fra Barentshavet. Den klareste grensen ble funnet mellom reker i Barentshavet og på kyststrekningen Trøndelag-Troms, men også reker på indre lokaliteter i Øst-Finnmark (Varangerfjorden og Tanafjorden) i tillegg til i fjordene vest for Tanafjorden viste signifikante genetiske forskjeller fra rekene i Barentshavet. Rekeprøven tatt rett på yttersiden av Vardø viste lik genetisk struktur som prøver tatt i Barentshavet, noe som indikerer at grensen mellom kysten og Barentshavet ligger nært kysten i dette området. Forskjeller i genetisk struktur mellom fjordene ble funnet mellom reker i Kvænangen og i Porsangerfjorden, noe som tyder på at grensen mellom de rene kystrekene og de blandete Finnmarksrekene befinner seg et sted mellom disse lokalitetene. Alle fjordene sør for Porsangerfjorden var signifikant forskjellig fra fjordene i Finnmark (Tabell 5.2), med unntak av indre lokaliteter i Varangerfjorden og Laksefjorden. I Varangerfjorden ble det funnet signifikante forskjeller mellom den indre og den ytre lokaliteten (Tabell 5.2), med en større andel kystreker i indre deler av fjorden (Figur 5.5). Porsangerfjorden viste ingen signifikante genetiske forskjeller mellom ytre og indre deler av fjorden, til tross for den grunne terskelen mellom de indre, østlige bassengene og ytre deler av fjorden som indikerer en isolert bestand, og til tross for forskjellige miljøforhold med svært lave bunntemperaturer om vinteren i den indre delen av fjorden (Myksvoll mfl. 2012).
Fjorder i Norge er generelt karakterisert av høye og bratte fjell, dype fjordbasseng og terskler (Myksvoll mfl. 2013). Til forskjell fra dette er fjordene i Finnmark generelt vide, og vannmassene i fjordene har ofte god utveksling med de utenforliggende vannmassene (Wassmann mfl. 1996). Den mer åpne naturen til Finnmarksfjordene kan være med å forklare den genetiske strukturen vi observerer, og da særlig forskjellen mellom kyststrekningen Trøndelag-Troms og Finnmark. Havstrømmer er viktige for drift og fordeling av reker i larvefasen (Pedersen mfl. 2003). Samtidig har temperatur vist seg å ha en større effekt på den genetiske populasjonsstrukturen enn larvedrift har (Jorde mfl. 2015). Over et større område i Nord-Atlanteren korrelerte temperatur positivt med populasjonsstrukturen som ble funnet.
Tidligere studier har vist at reker fra Trøndelag (prøven NOM) hører til den samme genetiske populasjonen som reker fra Nordsjøen og Skagerrak (Jorde mfl. 2015, Knutsen mfl. 2015). Den genetiske bestandsstrukturen av dypvannsreke langs norskekysten sammenfaller altså ikke med de nåværende forvaltningsenhetene (reker hhv. nord og sør for 62 °N).
| BSS1 | BSS2 | BSS3 | BSS4 | VARO | VARM | VAR | TANO | TANM | LAKM | LAKI | PORO | PORM | PORI | KVN | REI | LYN | MAL | FO2011 | RAN | FOFI | NOM | |
| BSS1 | 0,179 | 0,064 | 0,072 | 0,121 | <0,001 | 0,086 | 0,161 | 0,015 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | |
| BSS2 | 0,001 | 0,705 | 0,17 | 0,179 | <0,001 | 0,161 | 0,406 | 0,003 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | |
| BSS3 | 0,003 | -0 | 0,304 | 0,025 | <0,001 | 0,12 | 0,17 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | |
| BSS4 | 0,003 | 0,002 | 0,001 | 0,135 | <0,001 | 0,304 | 0,453 | 0,01 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | |
| VARO | 0,002 | 0,001 | 0,004 | 0,003 | 0,024 | 0,357 | 0,517 | 0,775 | <0,001 | 0,008 | 0,03 | 0,111 | 0,137 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | |
| VARM | 0,011 | 0,01 | 0,015 | 0,01 | 0,003 | <0,001 | 0,006 | 0,4 | 0,658 | 0,754 | 0,866 | 0,651 | 0,714 | 0,018 | 0,097 | 0,007 | 0,021 | 0,034 | 0,105 | 0,008 | 0,004 | |
| VAR | 0,002 | 0,001 | 0,002 | 0,001 | 0,001 | 0,007 | 0,567 | 0,027 | <0,001 | <0,001 | <0,001 | 0,001 | 0,002 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | |
| TANO | 0,002 | 0,001 | 0,002 | 0 | 0 | 0,004 | 0 | 0,237 | <0,001 | <0,001 | 0,004 | 0,001 | 0,011 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | |
| TANM | 0,003 | 0,005 | 0,009 | 0,005 | -0 | 0 | 0,003 | 0,001 | 0,007 | 0,176 | 0,434 | 0,348 | 0,796 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | |
| LAKM | 0,013 | 0,017 | 0,02 | 0,016 | 0,007 | 0 | 0,011 | 0,007 | 0,004 | 0,315 | 0,199 | 0,01 | 0,046 | 0,043 | 0,021 | 0,008 | 0,006 | 0,017 | 0,027 | 0,023 | 0,018 | |
| LAKI | 0,01 | 0,012 | 0,015 | 0,011 | 0,004 | -0 | 0,007 | 0,007 | 0,001 | 0,001 | 0,52 | 0,472 | 0,686 | 0,002 | 0,124 | 0,003 | 0,007 | 0,004 | 0,051 | 0,065 | 0,208 | |
| PORO | 0,008 | 0,01 | 0,015 | 0,01 | 0,003 | -0 | 0,008 | 0,004 | 0 | 0,001 | 0 | 0,318 | 0,481 | 0,003 | 0,011 | <0,001 | <0,001 | <0,001 | 0,013 | 0,002 | 0,05 | |
| PORM | 0,009 | 0,008 | 0,013 | 0,009 | 0,002 | 0 | 0,005 | 0,006 | 0,001 | 0,004 | 0 | 0,001 | 0,341 | <0,001 | 0,002 | 0,001 | <0,001 | <0,001 | 0,002 | <0,001 | <0,001 | |
| PORI | 0,006 | 0,006 | 0,011 | 0,009 | 0,001 | -0 | 0,004 | 0,003 | -0 | 0,002 | -0 | 0 | 0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | 0,002 | |
| KVN | 0,024 | 0,029 | 0,035 | 0,028 | 0,015 | 0,003 | 0,022 | 0,017 | 0,009 | 0,003 | 0,005 | 0,004 | 0,007 | 0,007 | 0,652 | 0,81 | 0,642 | 0,843 | 0,359 | 0,756 | 0,171 | |
| REI | 0,022 | 0,025 | 0,029 | 0,022 | 0,013 | 0,002 | 0,018 | 0,015 | 0,007 | 0,003 | 0,002 | 0,003 | 0,005 | 0,005 | 0 | 0,523 | 0,622 | 0,566 | 0,243 | 0,768 | 0,317 | |
| LYN | 0,027 | 0,03 | 0,036 | 0,028 | 0,016 | 0,004 | 0,023 | 0,019 | 0,01 | 0,004 | 0,004 | 0,006 | 0,006 | 0,008 | -0 | 0 | 0,349 | 0,4 | 0,774 | 0,555 | 0,065 | |
| MAL | 0,024 | 0,028 | 0,035 | 0,027 | 0,015 | 0,003 | 0,022 | 0,017 | 0,01 | 0,004 | 0,004 | 0,005 | 0,007 | 0,006 | 0 | 0 | 0,001 | 0,52 | 0,063 | 0,52 | 0,191 | |
| FO2011 | 0,026 | 0,03 | 0,038 | 0,03 | 0,016 | 0,003 | 0,023 | 0,021 | 0,011 | 0,004 | 0,004 | 0,006 | 0,008 | 0,007 | -0 | 0 | 0,001 | 0 | 0,097 | 0,358 | 0,017 | |
| RAN | 0,021 | 0,024 | 0,03 | 0,022 | 0,012 | 0,002 | 0,019 | 0,015 | 0,006 | 0,003 | 0,002 | 0,003 | 0,005 | 0,006 | 0,001 | 0,001 | -0 | 0,002 | 0,002 | 0,297 | 0,454 | |
| FOFI | 0,025 | 0,03 | 0,034 | 0,028 | 0,016 | 0,004 | 0,023 | 0,019 | 0,01 | 0,003 | 0,002 | 0,005 | 0,006 | 0,007 | -0 | -0 | 0 | 0 | 0,001 | 0,001 | 0,157 | |
| NOM | 0,016 | 0,022 | 0,027 | 0,023 | 0,01 | 0,004 | 0,016 | 0,014 | 0,006 | 0,004 | 0,001 | 0,003 | 0,006 | 0,004 | 0,002 | 0,001 | 0,002 | 0,001 | 0,004 | 0 | 0,002 |
5.1.5 - Bestandsstruktur
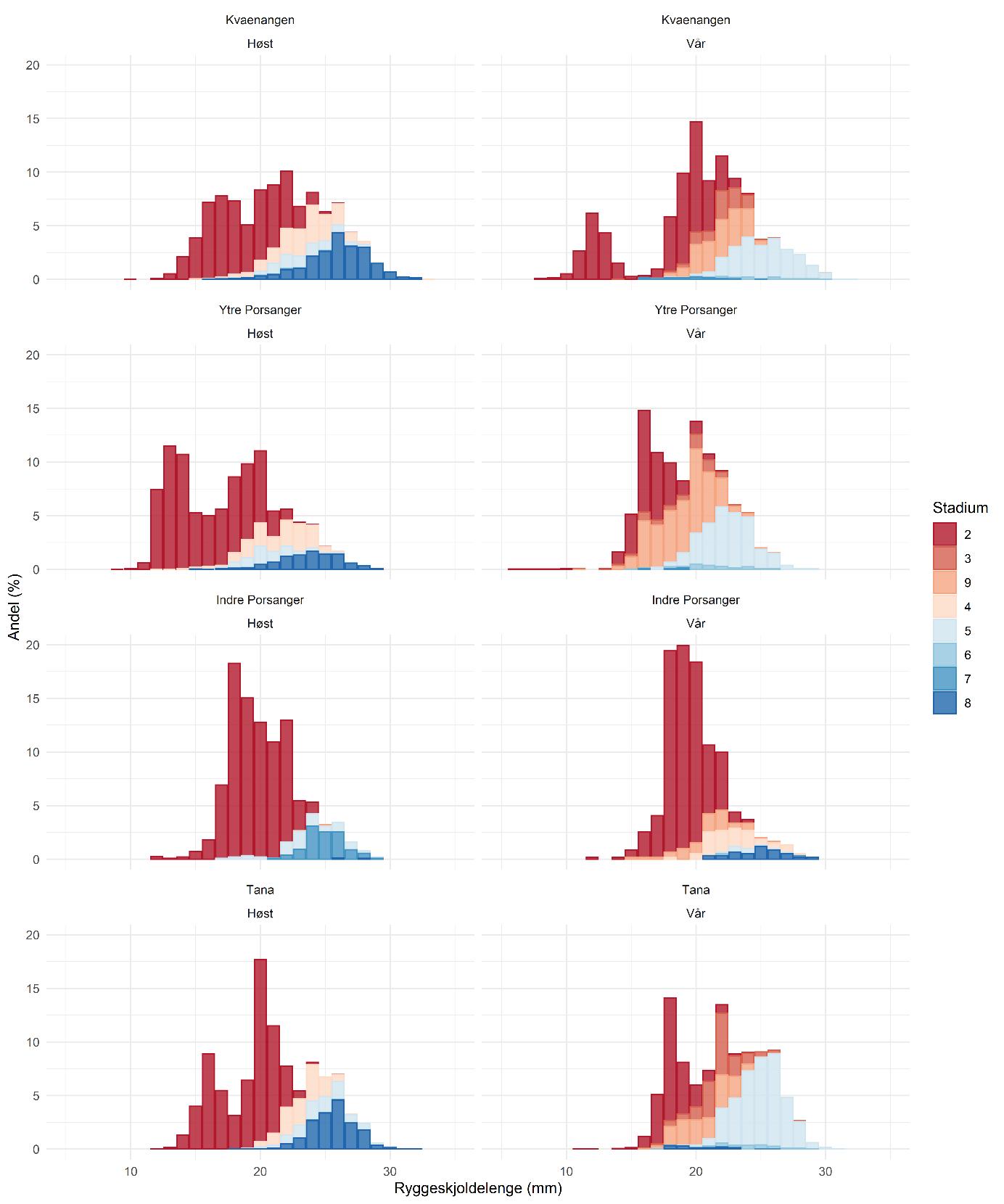
Lengdefrekvensfordelingene fra de tre fjordene hadde to til tre tydelige modaltopper (Figur 5.6) som viser aldersklasser. I mange rekepopulasjoner opptrer juvenile reker sammen med eldre reker fra de er rundt 1 år gamle (Rasmussen 1953, Shumway mfl. 1985, Nilssen og Hopkins 1991). Vi antar derfor at den første modaltoppen i lengdefrekvensfordelingene består av 1-åringer. Pluss-gruppen består av de eldste årsklassene som ikke kan skilles fra hverandre pga. overlappende lengde. Som allerede nevnt, ble de største rekene funnet i Kvænangen.
Det følgende gjelder for rekebestandene i Kvænangen, Tanafjorden og ytre Porsangerfjorden. Rekene i den indre delen av Porsangerfjorden omtales for seg til slutt.
Hannrekene begynner å skifte kjønn til hunn når de er 2 år gamle (Figur 5.6), men en del skifter ikke kjønn før de er 3 år gamle. Kjønnsskiftet foregår om våren. De fleste hunnrekene har hoderogn i oktober (stadier 4 og 8), mens en mindre andel allerede har fått utrogn (stadium 5). Paringen foregår altså i oktober/november. I mars har de aller fleste hunnrekene fremdeles utrogn, slik at larvene sannsynligvis klekker i april/mai.
Mens en tydelig modaltopp av 1-åringer sees i lengdefordelingen fra mars 2019 i Kvænangen, fikk vi omtrent ikke 1-åringer i trålen i mars i hverken Tana- eller Porsangerfjorden. De små rekene kan ha vært på ikke-trålbare områder. Det kan også være at 1-åringene faktisk kun finnes i et veldig lavt antall i disse to fjordene, men at de vandrer inn fra andre områder i løpet av sommeren. Om høsten er 1-åringene tilstede i alle de tre fjordene. I rekebestanden i Skagerrak og Norskerennen ser vi noe tilsvarende (Søvik og Thangstad 2020). I områdene vest av Lindesnes mangler 1-åringene i januar/februar, mens de alltid finnes inne i Skagerak. Da 2-åringene alltid er tilstede ute i Norskerennen migrerer de sannsynligvis dit ut fra Skagerrak.
I indre Porsangerfjorden kan vi gå ut ifra at veksten er svært langsom pga. den lave temperaturen, vi ser heller ingen klare modaltopper i lengdefrekvensfordelingene. Populasjonen er totalt dominert av hanner. I mars 2019 fant vi nesten ingen hunner med utrogn, men en god del hunner med hoderogn. Kanskje betyr det at de parer seg om våren eller sommeren? Om høsten var det en del hunner med utrogn. De aller fleste hunnene om høsten var imidlertid i stadium 7, dvs. de hadde reprodusert tidligere, men ikke utviklet hoderogn den høsten. Det kan tyde på at hunnene bare reproduserer annethvert år, eller kanskje hvert tredje år? Produktiviteten og veksten er altså lav, men tettheten er svært høy. Dette må skyldes fraværet av predatorer og fiske i de indre østlige bassengene. Biomassen må ha akkumulert over mange år.
5.1.6 - Vekst og dødelighet
Vekst og naturlig dødelighet ble undersøkt for dypvannsreke i Tanafjorden, Porsangerfjorden og Kvænangen (Båtevik 2020). Resultatene fra de nordnorske fjordene ble sammenlignet med tilsvarende resultater fra den (delvis) utrålte Gullmarsfjorden på Sveriges vestkyst. Gitt det positive forholdet mellom temperatur og vekst (Shumway mfl. 1985), ble det forventet en høyere vekst i den sørlige populasjonen i Gullmarsfjorden sammenlignet med fjordpopulasjonene i Finnmark (Båtevik 2020). Da rask vekst innebærer et kortere livsløp og derfor en høyere årlig dødelighet (Shumway mfl. 1985, Nilssen og Hopkins 1991), ble det også forventet en høyere naturlig dødelighet i Gullmarsfjorden. Gullmarsfjorden ble stengt for reketråling i 1990. I 1996-1997 undersøkte man effekten av tråling på bunnhabitater. Tre områder ble trålt månedlig for å simulere kommersiell bunntråling, mens tre utrålte områder utgjorde et kontrollområde. Fjorden var derfor ikke fullstendig utrålt under datainnsamlingen.
Total dødelighet er summen av fiskedødelighet og naturlig dødelighet (Z = F + M). Naturlig dødelighet er viktig i bestandsvurderinger da denne parameteren bestemmer produktiviteten til den modellerte bestanden (Clark 1999, Aanes mfl. 2007, Williams 2011) og dermed det bærekraftige høstingsnivået (ICES 2016). Naturlig dødelighet er imidlertid vanskelig å estimere (Vetter 1988). En ufisket bestand utgjør en sjelden mulighet til å estimere M direkte. De tre stengte fjordene Porsangerfjorden, Tanafjorden og Gullmarsfjorden gjorde det mulig å estimere M direkte for populasjoner på forskjellige breddegrader i hele utbredelsesområdet til dypvannsreke i Nordøst-Atlanteren. Men som nevnt over, ble Gullmarsfjorden vurdert til å ikke være helt ufisket. De indre bassengene i Porsangerfjorden med bunntemperaturer ned mot 0 °C ble vurdert til å kunne representere de aller nordligste leveområdene til reke. Imidlertid viste det seg at dataene for denne lokale populasjonen var av for dårlig kvalitet til å kunne estimere vekst og dødelighet; det ble tatt forholdsvis få trålhal og den lave temperaturen fører til langsom vekst med overlappende lengdeintervaller for de forskjellige aldersgruppene. Resultatene for Porsangerfjorden i dette avsnittet gjelder derfor bare for reke i den midtre og ytre delen av fjorden.
Klare modaltopper for de 2-4 første aldersgruppene i lengdefrekvensfordelinger gjør at alder kan bestemmes. Vekst og dødelighet ble estimert vha. electronic length frequency analysis (ELEFAN) (Pauly og Sparre 1991) og linearized length converted catch curve analysis (Sparre og Venema 1998). Vekst ble antatt å følge von Bertalanffys vekstfunksjon.
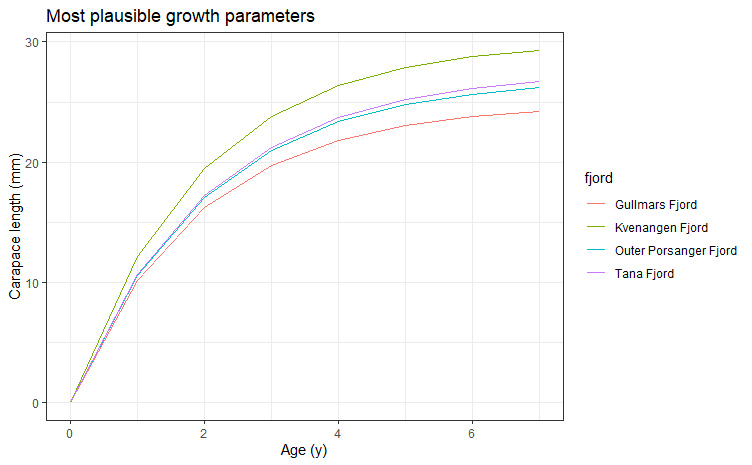
Vekstparametrene for hver fjord (Tabell 5.3) ble valgt ut fra en rekke modellkjøringer, basert på den høyeste Rn-verdien (goodness of fit) (Pauly og David 1981) samt en vurdering av hvor godt von Bertalanffy vekstkurvene fulgte modaltoppene i lengdefrekvensfordelingene (Båtevik 2020). Vekstkurvene viste at det var 6-7 aldersgrupper (inkludert 0-gruppen) i fjordene. Sammenligning av ϕ-verdiene (vekstindeks) (ϕ = ln(K) + 2ln (L∞)) (Tabell 5.3) og de fjordspesifikke vekstkurvene (Figur 5.7) viste at veksten var høyest i Kvænangen, etterfulgt av Tana- og Porsangerfjorden, og lavest i Gullmarsfjorden, altså motsatt av hva som var forventet. Årlig gjennomsnittlig bunntemperatur (± standardavvik) i Gullmarsfjorden i 1998 (6,9 ± 0,8 °C) var høyere enn årlige gjennomsnittlige bunntemperaturer i de nordnorske fjordene i 2018-2019 (Båtevik 2020), men temperatur er ikke nødvendigvis den viktigste driveren bak variasjoner i livshistorie hos reke (Nilssen og Hopkins 1991, Bergström 1992). Et lavt oksygennivå i bunnvannet i Gullmarsfjorden kan ha ført til den lave veksten. Dette har blitt observert for flere arter av fisk og krepsdyr (Green 2009, Seidman og Lawrence 2009, Adelman og Smith 2011). Bunnvannet i denne terskelfjorden ble ikke fornyet hverken våren 1995 eller våren 1997 (Hansson mfl. 1997) noe som førte til et oksygennivå ned i 1-2 ml/l (Båtevik 2020). I de åpne fjordene i Troms og Finnmark er det sannsynligvis en jevnlig fornying av bunnvannet (kapittel 4). På FiN-tokt i Porsangerfjorden i august 2015 ble det målt et oksygeninnhold mellom 5 og 5,5 ml/l i bunnvannet (Figurer V1.1, V1.2). Vekst i Kvænangen var mye høyere enn i de ufiskede finnmarksfjordene. Fiskede bestander kan ha høyere produksjon, og dermed vekst, fordi den lavere bestandstettheten kan føre til mindre konkurranse for mat (Nilssen og Hopkins 1991). Men tettheten av reke var høyere i Kvænangen enn i de andre fjordene (med unntak av indre Porsangerfjorden) (Figur 5.3), som også er motsatt av hva som var forventet. Dette stemmer med det enkelte rekefiskere hevder, nemlig at rekefelt må “tråles” for å gi gode fangstrater. En mulig forklaring kan være at tråling roter opp bunnen og slik frigjør næringsstoffer fra sedimentet (Dounas mfl. 2005, 2007).
| Fjord | Lꝏ (mm) | K (y-1) | t_anchor | Rn | Φ |
| Gullmarsfjorden | 24,8 (24,50, 26,17) | 0,53 (0,32, 0,55) | 0,09 (0,06, 0,43) | 0,57 | 2,51 (2,35, 2,52) |
| Porsangerfjorden | 27,0 (26,99, 27,01) | 0,50 (0,51, 0,50) | 0,38 (0,37, 0,41) | 0,78 | 2,56 (2,56, 2,57) |
| Tanafjorden | 27,5 (27,15, 28,51) | 0,50 (0,47, 0,53) | 0,13 (0,08, 0,20) | 0,93 | 2,58 (2,57, 2,60) |
| Kvænangen | 30,1 (29,96, 30,28) | 0,53 (0,50, 0,54) | 0,23 (0,14, 0,26) | 0,89 | 2,66 (2,65, 2,69) |
Alle fjordpopulasjonene viste en minkende dødelighet med økende alder. Dødelighet ble derfor estimert for både yngre (Zy) og eldre reker (Zo). Årlig total dødelighet for yngre reker (aldersgrupper 2-5 år avhengig av bestand) varierte mellom 0,83 og 1,05, mens total dødelighet for eldre reker (aldersgrupper 3-7 år avhengig av bestand) varierte mellom 0,62 og 1,01 (Tabell 5.4). Både Zy og Zo var høyest i Kvænangen (Tabell 5.4). Det hefter stor usikkerhet ved aldersestimering av de eldste rekene da forholdet mellom lengde og alder blir mer usikkert. En stor kroppsstørrelse kan skyldes både høy alder og rask vekst (Sparre og Venema 1998). Det er derfor bedre å sammenligne alder for de yngre rekene.
Vi har ikke prøvd å tallfeste fiskedødeligheten i Kvænangen eller Gullmarsfjorden pga. stor usikkerhet i biomasseestimatene og også i offisielle landinger fra Kvænangen. Vi kan likevel konkludere med at M i Kvænangen må være lavere enn i Tana- og Porsangerfjorden, og M i Gullmarsfjorden er i hvert fall ikke høyere enn i de ufiskede finnmarksfjordene. At M ikke er betydelig høyere i Gullmarsfjorden sammenlignet med de nordlige fjordene er motsatt av hva som var forventet. Lav dødelighet i Gullmarsfjorden er imidlertid ikke uventet gitt den lave veksten (Nilssen og Hopkins 1991). I tillegg til temperatur og vekst påvirker predasjonstrykket naturlig dødelighet (Simpfendorfer, Bonfil og Latour 2005), men vi vet ikke om predasjonstrykket i Gullmarsfjorden var mye lavere i 1997 sammenlignet med dagens predasjonstrykk i de nordnorske fjordene.
Total dødelighet i de tre nordnorske fjordene ligger på noenlunde samme nivå. I de ufiskede fjordene skyldes all dødelighet naturlige årsaker, mens i den fiskede Kvænangen skyldes mye av dødeligheten fisket.
| Fjord | ZY | ZO | MY | MO |
| Gullmarsfjorden | 0,92 ± 0,03 | 0,70 ± 0,01 | ||
| Porsangerfjorden | 0,83 ± 0,05 | 0,81 ± 0,04 | 0,83 ± 0,05 | 0,81 ± 0,04 |
| Tanafjorden | 0,94 ± 0,05 | 0,62 ± 0,03 | 0,94 ± 0,05 | 0,62 ± 0,03 |
| Kvænangen | 1,05 ± 0,03 | 1,01 ± 0,01 |
5.2 - Torsk
Torsk er en nøkkelart i alle tre fjordene. Det finnes to torskebestander i dette området: lokale fjordpopulasjoner av kysttorsk og Barentshavtorsk eller skrei, som hovedsakelig forekommer i Barentshavet, men som også delvis vandrer inn i disse fjordområdene, spesielt i sammenheng med gyte- og næringsvandring om vinteren og våren. Torsk, og spesielt skrei, er veldig viktig for lokale kystfiskere (avsnitt 6.3) og også for dypvannsreke, da torsk er en hovedpredator på reke. Det finnes derfor en direkte økologisk sammenheng mellom torske- og rekebestander, og fiske på en av de to artene kan indirekte påvirke den andre.
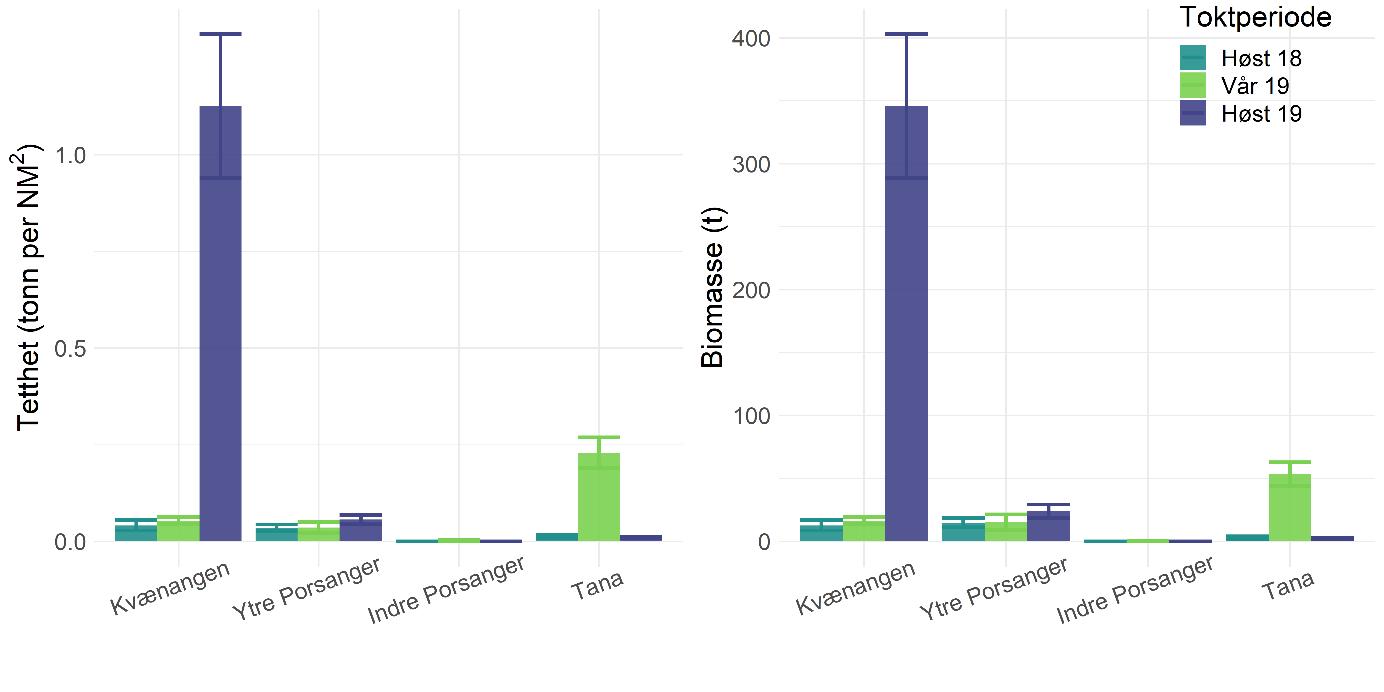
Tetthet og biomasse av torsk var stort sett konsistent på tvers av fjord, dyp og toktperiode (Figur 5.8). Unntaket er indre Porsangerfjorden hvor torskeforekomstene var mye lavere og begrenset til den nordvestlige delen, mens ingen torsk ble funnet i de østlige, kalde bassengene. Vi fant ikke betydelige sesongforskjeller, med unntak av i Tanafjorden hvor tettheten våren 2019 var signifikant lavere enn tettheten høsten 2018 og 2019. Endringer gjennom toktperiodene og år viser ikke en klar trend.
Tetthet og biomasse estimert fra data fra kystressurstoktet samsvarer i stor grad med estimatene fra kartleggingstoktet (Figur 5.9). Et signifikant avvik finnes kun for Tanafjorden i 2019. Kysttoktdata fra tidligere år indikerer generelt ikke store endringer. Selv om det kan se ut som om det er en økning i biomassen i 2012-2014 i Porsangerfjorden, er usikkerheten i kysttoktdataene veldig stor, hovedsakelig pga. det lave antallet stasjoner per fjord. I tillegg ble arealet av hele fjorden brukt til å beregne biomassen, og det resulterer sannsynligvis i en overestimering av totalbiomasse. Biomassene som vises i Figurene 5.8 og 5.9 bør derfor tolkes først og fremst som en indeks som viser endringer over tid i et område, og ikke som en nøyaktig representasjon av den faktiske biomassen.
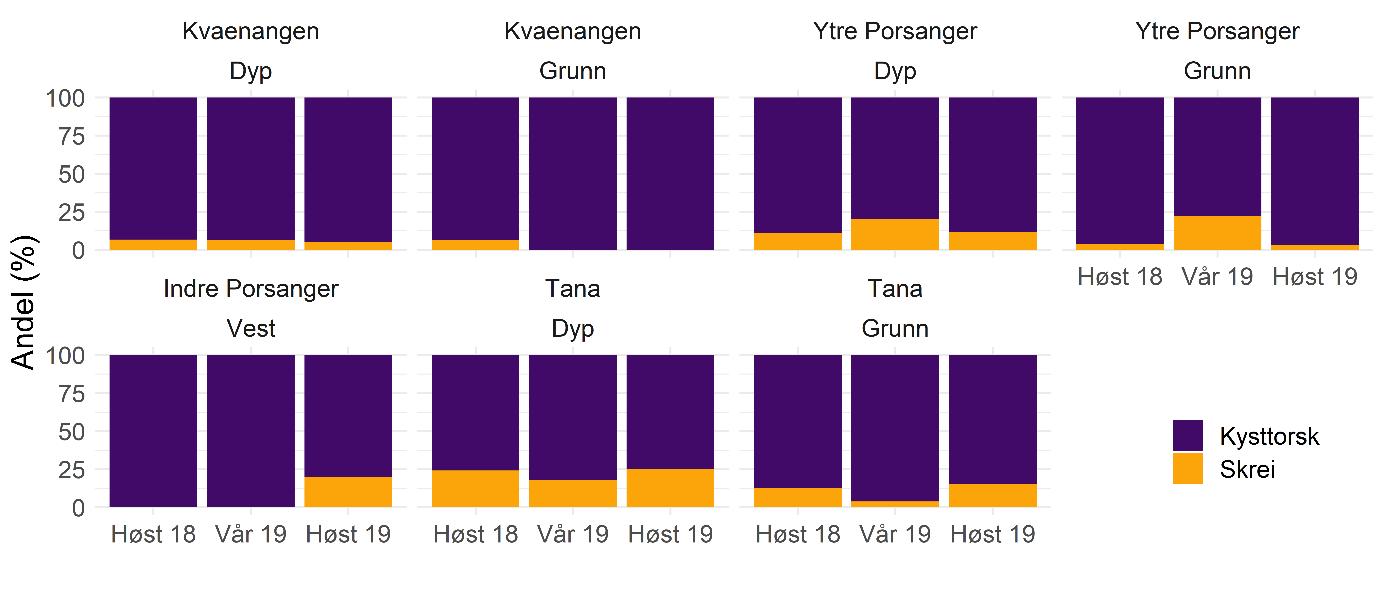
Basert på otolittene ble en stor andel av alderslest torsk (2 år og eldre) fra kartleggingstoktet identifisert som kysttorsk og kun et mindretall som skrei (Figur 5.10). Andelen av skrei var høyest i Tanafjorden (21 % i gjennomsnittet) mens den var lavest i Kvænangen og indre Porsangerfjorden (hhv. 5 % og 6 %). Kun i ytre Porsangerfjorden finnes det en tydelig økning om våren, da andelen gikk opp til rundt 21 % sammenlignet med ca. 9 % om høsten. I tillegg til otolitter ble også genetikkprøver (av stor torsk) tatt, noe som vil gi et mer nøyaktig bilde av bestandsstrukturen til voksen fisk. Genetikkanalysene skal ferdigstilles i løpet av 2021.
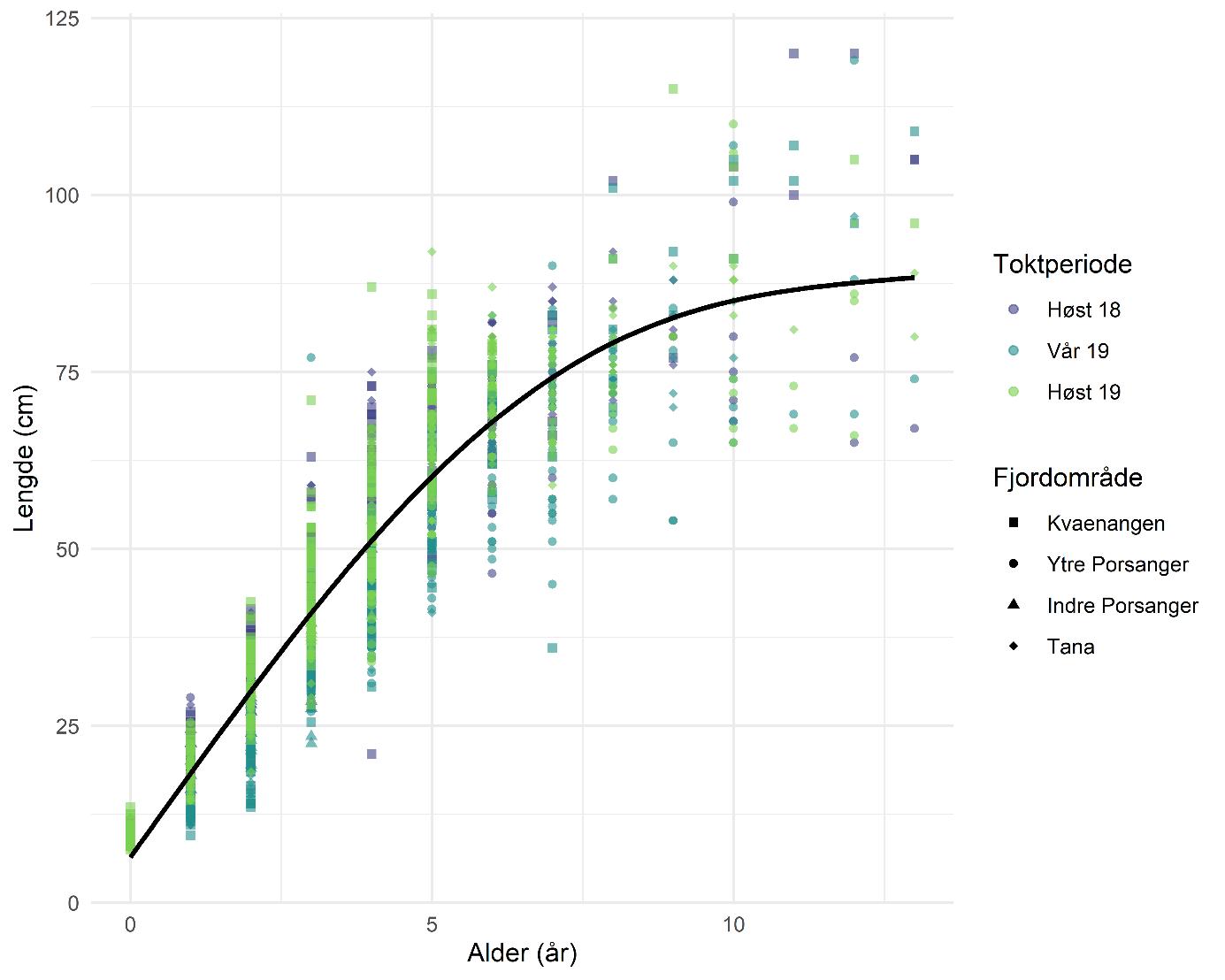
Torskens alders- og størrelsessammensetning var stor sett lik i alle toktperiodene og fjordene (Figur 5.11). Det finnes noen variasjoner, men uten en klar eller signifikant trend. Det tydeligste unntaket er indre Porsangerfjorden hvor ingen torsk over 5 år ble registrert. Det kan skyldes den generelt lave torskeforekomsten og det lave antallet trålhal i indre Porsangerfjorden.
5.3 - Hyse
Sammen med torsk er hyse en sentral art i Barentshavet og i kystområdene i Finnmark og Troms. I motsetning til torsk blir hyse langs kysten og i Barentshavet forvaltet som én bestand, nordøstarktisk hyse. Flere av hysens gyteområder ligger i kystområdene i Nord-Norge. Hyse har en livssyklus og økologisk betydning som er sammenlignbar med torsk, og hyse spiser også reker, selv om reker har en litt mindre betydning i hysens diett enn i torskens diett.
Hyse var den arten som ble registrert oftest, på tvers av alle stasjoner og toktperioder under kartleggingstoktet, og arten hadde i gjennomsnitt også den høyeste tettheten (Figur 5.12). Tettheten av hyse var i de fleste fjordområdene og toktperiodene høyere enn 5 tonn per nautisk kvadratmil. Tettheten var veldig konsistent på tvers av trålstasjonene innenfor hver fjord og toktperiode og har derfor lav usikkerhet, unntatt i indre Porsangerfjorden hvor hyse forekommer hovedsakelig i det nordvestlige området. Sammen med et lavt antall stasjoner i indre Porsangerfjorden resulterte det i stor variasjon. Høy tetthet skalerer opp til høye biomasseestimater, som viser at det finnes en betydelig hysebestand i disse fjordene. I tillegg ser vi også signifikant variasjon over tid, med en klar økning i Kvænangen mens tetthet og biomasse minker i ytre Porsanger- og Tanafjorden. Vi fant ikke noe negativt forhold mellom forekomst av hyse og dypvannsreke, og det finnes heller ikke mer hyse i områder hvor dypvannsreke ikke blir fisket.
En mulig forklaring på endringene over tid er endringer i romlig fordeling av bestanden. En stor del av hysen som ble registrert under toktene, var yngel og juvenil fisk, og det antyder at hyse bruker fjordene hovedsakelig som oppvekstområde og vandrer ut til Barentshavet når de blir eldre. Observerte tettheter er også knyttet til trender i hele bestanden, som viser en nedgang i gytebestanden over de siste årene mens årsklassen i 2016 var veldig sterk, noe som kan ha bidratt til den høye tettheten av juvenil hyse i fjordene. I tillegg forklarer det hvorfor vi registrerte en høyere tetthet i områder grunnere enn 170 m, da ung hyse hovedsakelig oppholder seg i grunne områder.
Estimater fra kartleggingstoktet samsvarer generelt bra med estimater fra kystressurstoktet (Figur 5.13) som viser trender over en lengre tidsperiode. Men det finnes også forskjeller, spesielt høsten 2019 fant vi en signifikant lavere biomasse basert på kysttoktdata. Det antyder at kysttoktet ikke klarer å fange opp alle svingninger i regional tetthet pga. et lavere antall stasjoner, men likevel klarer å gi en god nok indikasjon av endringer over tid.
5.4 - Sei
All sei som finnes langs norskekysten nord for 62 °N og i Barentshavet, blir forvaltet som én bestand. Seibestanden har omtrent samme størrelse og betydning som nordøstarktisk hyse. Men i motsetning til hyse og torsk er sei ikke hovedsakelig en bunnfisk, men er delvis pelagisk. Derfor finner man ofte konsentrasjoner av sei høyere opp i vannsøylen og delvis med stor tetthet, spesielt i strømrike områder og andre områder med gode beitemuligheter. Dette betyr at sei har mindre interaksjoner med dypvannsreke enn torsk og hyse, og at det lokalt kan finnes sei i veldig store tettheter mens tettheten over større områder kan være relativt lav. I tillegg blir ikke sei fanget i bunntrål hvis de ikke står på bunnen. Det betyr at bunntrålingen under kartleggingstoktet sannsynligvis ga et ufullstendig bilde av seibestanden.
Estimater fra kartleggingstoktet bekrefter seiens pelagiske natur, med stort sett veldig lave tettheter og biomasser på tvers av de tre fjordene og toktperiodene (Figur 5.14). Data indikerer at det finnes lite sei på bunnen i disse fjordene, og at sei er nesten fraværende fra noen områder, spesielt indre Porsangerfjorden. Men betydelige tettheter ble registrert på flere stasjoner innerst i Kvænangen høsten 2019, og det viser at det kan finnes lokalt betydelige mengder. Vi antar at en god del sei ikke ble observert under kartleggingstoktet fordi de ikke stod på bunnen. Kystressurstoktet overvåker derfor sei med akustisk mengdemåling. Disse dataene viser at det finnes mer sei i studieområdet enn det ble registrert under kartleggingstoktet, men dataene viser også at seiforekomstene generelt ikke er veldig store i kystområdet (Staby mfl. 2018).
5.5 - Andre bunnfiskarter
I tillegg til de store bestandene i torskefamilien, spesielt torsk og hyse, finnes det en blanding av mange forskjellige bunnfiskarter i Kvænangen, Porsanger- og Tanafjorden, f.eks. uer, flatfisk som gapeflyndre, kveite, rødspette og smørflyndre, brosme, breiflabb og ulker, spesielt i indre Porsangerfjorden. På kartleggingstoktet hadde vi også sporadiske registeringer av skater som kloskate og hvitskate, lange, og ringbuk- og langebarnarter.
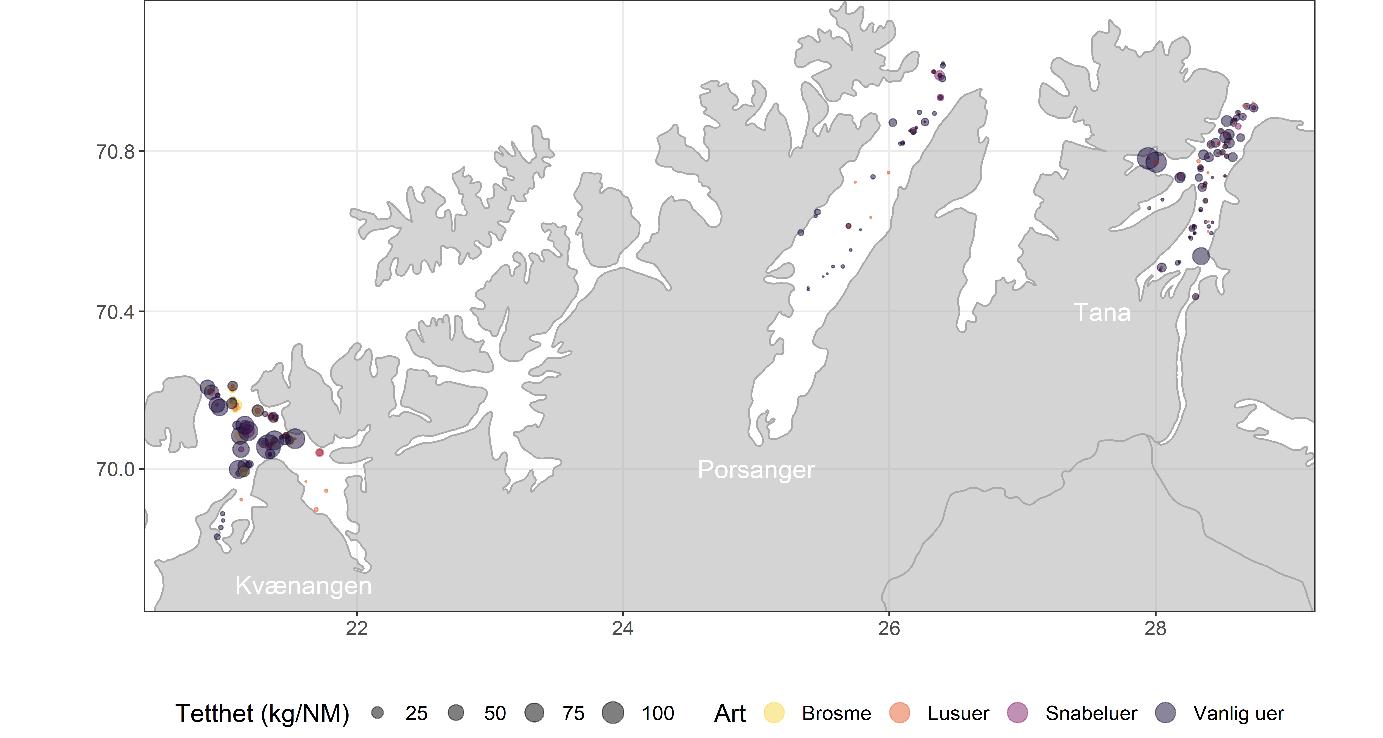
Data fra kartleggingstoktet viser at bunnfisk ble hovedsakelig funnet i de ytre delene av fjordene (Figur 5.15), spesielt i Kvænangen og Tanafjorden, mens flatfisk var mer jevnt fordelt innenfor og på tvers av fjordene (Figur 5.16). Vanlig uer dominerte i Kvænangen og Tanafjorden, mens rødspette hadde den høyeste tettheten i Porsangerfjorden (Figur 5.17). Fangsten av uer bestod hovedsakelig av vanlig uer, men det ble også registrert lusuer og snabeluer. Uerfangsten bestod i stor grad av yngel og juvenil fisk. Det indikerer at deler av de undersøkte fjordene kan være oppvekstområde for vanlig uer. Generelt viste alle disse bunnfiskartene relativt stor variasjon over tid og område, noe som kan skyldes ofte lave prøvemengder som påvirker resultatene sterkt. Dette gjelder spesielt kveite fordi kveite kun forekom som noen få, men store individ i våre fangster. Tilfeldige enkeltfangster påvirker da estimatene i stor grad.
Feltundersøkelser og laboratoriestudier i Porsangerfjorden har vist at ulker er den viktigste predatoren på nylig bunnslått yngel, og at kombinasjonen av ulke som angriper nedenfra og ungtorsk som angriper ovenfra, er en spesielt dødelig kombinasjon for yngelen. Kartleggingstoktene fanget bare ulker i indre Porsangerfjorden, men at ikke vanlig ulke ble fanget andre steder skyldes nok at det ikke ble fisket i de grunne habitatene til denne arten. I de innerste, kalde bassengene i Porsangerfjorden var det mye av den arktiske arten glattulke, og her ble det også registrert tiskjegg, krokulke og ringbuk.
5.6 - Sild
Vintersildfisket etter gytemoden sild som hadde overvintret i Norskehavet, kollapset fra fangstnivåer på opp mot 1 million tonn på 1950-tallet til 15 000 tonn i 1969 (Christensen 2014). En del av flåten gikk etter kollapsen inn i de nordnorske fjordene og fortsatte fisket, også på småsild, til også dette ble stoppet etter et par år.
Sild har alltid vært en viktig art i alle de tre fjordøkosystemene, både som byttedyr og som predator. Strand mfl. (2020) beskriver dette godt. Den lokale sildestammen med innslag av mye stor sild forsvant fra Porsangerfjorden på begynnelsen av 1990-tallet. Det er likevel mye sild i fjorden i dag, men det er trolig ungsild av Norsk Vårgytende Sild (NVG) siden den vandrer ut av fjordsystemene som 1-2 åringer. Det vil derfor være et mål å få tilbake den lokalt store silda i Porsangerfjorden, og trolig er dette også situasjonen for de to andre fjordene.
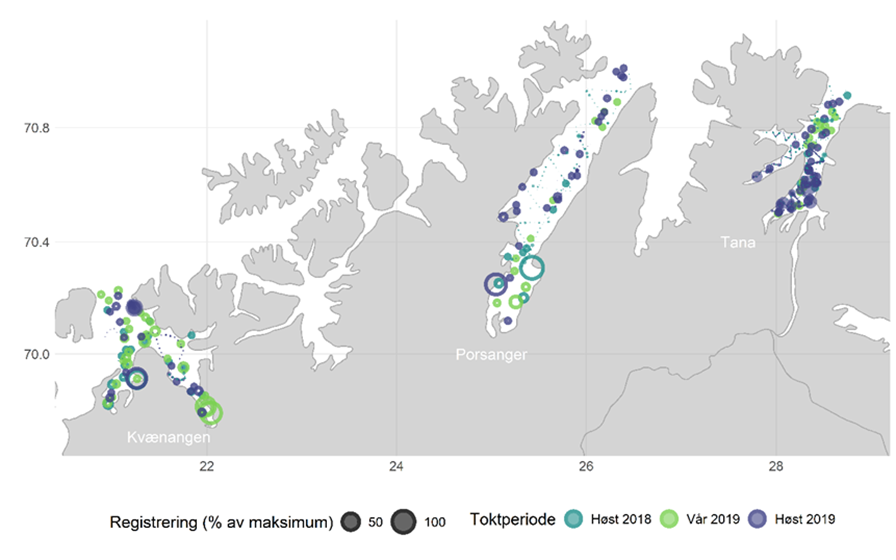
Figur 5.18 viser bunntrålfangster og akustisk mengdemåling i 2018-2019. Begge metodene har sine begrensninger når det gjelder representative målinger av småsild, men gir kombinert kanskje et mer eller mindre representativt bilde. Sildetettheten er stort sett høyest i de innerste delene av fjordene og nær kysten, i grunne områder.
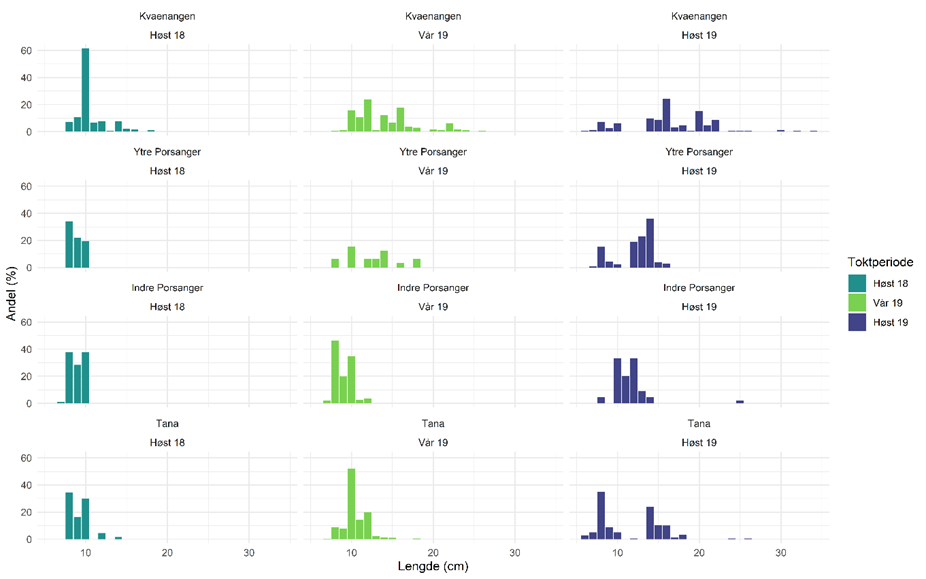
Figur 5.19 viser lengdefordelinger av sild i trålfangstene på kartleggingstoktet i de tre fjordene høsten 2018, våren 2019 og høsten 2019. De siste årene har det i Kvænangen vandret inn store mengder NVG-sild om vinteren før gyting. Dette har samlet både hval og fiskere, og resultert i et svært godt fiskeri. Denne silda hadde vandret ut igjen før kartleggingstoktet i mars/april 2019. Lengdefordelingene viser at fjordene ellers i året er dominert av 1-3 årsklasser og hvordan årsklassene varierer mellom fjordene/områdene. Lengdefordelingene viser også vekstforskjell mellom sild i ytre og indre Porsangerfjorden.
5.7 - Bunndyr
Bunndyr inkluderer et betydelig antall arter. Noen av disse er sårbare overfor bunntråling, spesielt koraller, svamper og andre arter som strekker seg opp fra havbunnen og som kan ødelegges ved fysisk kontakt. Norge har internasjonale forpliktelser til å ta vare på sårbare habitater og økosystem. En kartlegging av bunndyr på trålbare områder er derfor viktig når man vurderer en gjenåpning av bunntråling i ufiskete områder.
Den opprinnelige planen var å kartlegge utbredelsen av forskjellige typer bunndyr som del av kartleggingstoktet. Imidlertid viste det seg at reketrålen på «Katla» fanget bunndyr svært dårlig. Vi har derfor basert oss på bomtråldata fra FiN-toktet i mai 2019 samt tidligere innsamlet bomtråldata. Vi har også brukt data fra Campelen-trålen på kystressurstoktet høsten 2019 og data fra videotransekter fra et MAREANO tokt i 2011. Vi vurderte opprinnelig også filming av bunn med go-pro kamera fastmontert til trål. Dette ble prøvd ut under FiN-toktet, men pga. svært dårlig sikt på bunn, ga det ingen brukbare resultater. For Porsangerfjorden finnes det data fra NGU fra tidligere kartlegging av bunnsediment, som viser sammensetningen av sedimenttype (Figur 2.2).
Bunndyrs- og sedimentdata mangler for Tanafjorden. Fra Kvænangen finnes det registreringer fra Campelen-trålen fra kystressurstoktet høsten 2019. Det følgende kapittelet gir en oversikt over den kunnskapen vi har om bunndyr i Porsangerfjorden og Kvænangen, basert på de nevnte datakildene.
5.7.1 - Bomtrålundersøkelser av bunndyrssamfunn - fordeling i tid og rom
Totalt 65 stasjoner med en liten bomtrål (2 m) har blitt tatt i Porsangerfjorden i perioden 2007-2019 (Tabell 5.5). 2019-dataene er fra FiN toktet i mai 2019. Innsamling med bomtrål ble gjennomført i 2008 også, men materialet ble bare bestemt til dyregrupper (ingen artsidentifisering). Data fra 2008 presenteres derfor kun i noen oversiktskart og er ikke inkludert i de artsbaserte analysene. Bomtrålen er en liten bunntrål med meier og en duk av kjetting på undersiden som hvirvler opp bunndyr, og fanger disse inni et nett med maskevidde på 4 mm.
| 2007 | 2009 | 2010 | 2011 | 2014 | 2016 | 2019 |
| 9 | 6 | 4 | 9 | 22 | 6 | 6 |
Fordelingen av bunndyr over tidsperioden 2007-2010 viser at artsrikdommen er høyest i de midtre og ytre delene av Porsangerfjorden (Figur 5.20). Antall individer og biomasse er høyest i den indre, kalde delen av fjorden, med noe økt biomasse i de ytre delene. De dominerende dyregruppene er pigghuder (ECD i Figur 5.21) i de indre delene av fjorden, krepsdyr (ART i Figur 5.21) i de sentrale delene, og svamper (POR) og nesledyr (CNI) i de vestlige og ytre delene av fjorden.
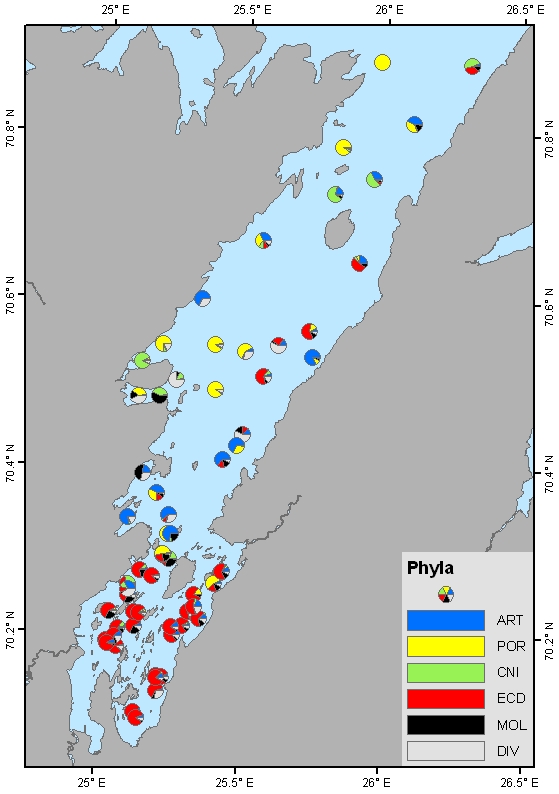
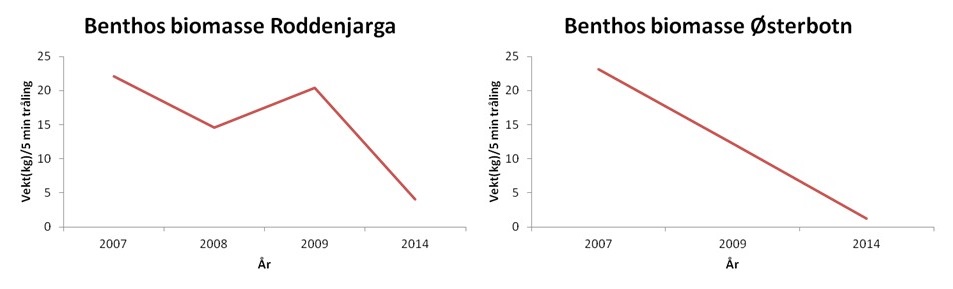
Fra 2007 til 2014 kan det se ut som om biomassen i de indre bassengene i Porsangerfjorden har minket (Figur 5.22). Det er usikkert hva grunnen til dette er. Økt temperatur, beiting fra nye arter (kongekrabbe), eller endring i toktet (fangstmåte) (bytte av forskningsskip fra R/V Johan Ruud til R/V Johan Hjort) kan være mulige årsaker.
En samfunnsanalyse ble gjennomført for å beskrive bunndyrsvariasjonen fra område til område i Porsangerfjorden og for å identifisere samfunn sårbare for bunntråling (Vedlegg 3). Det ble valgt en oppløsning på ti samfunn og ut av disse ble åtte identifisert som sårbare overfor bunntråling.
5.7.2 - Sårbarhet overfor bunntråling – bomtråldata fra Porsangerfjorden
Det finnes flere metoder for å gradere sårbarhet av bunndyr overfor bunntråling (se f.eks. FAO, OSPAR, ICES arbeidsgrupper). I dette studiet tas det utgangspunkt i taxa som finnes i Porsangerfjorden og som kan stå mer enn 10 cm oppreist fra havbunnen og derfor kan bli ødelagt av bunntråling (Tabell 5.6). Sårbare taxa forekommer på de fleste bomtrålstasjonene i Porsangerfjorden (Figur 5.23) og med fra én og opptil flere forskjellige taxa på samme lokalitet.
| Dyregruppe | Taxa |
| Porifera | Svamp (Antho dichotoma, Axiniella sp., Crania sp., Geodia sp., Halichondria sp., Myxilla sp., Phakellia sp., Pseudosuberites sp., Suberites sp.) |
| Cnidaria | Hydroider (Lafoea fruticosa) |
| Cnidaria | Blomkålkoraller (Nephtheidae) |
| Cnidaria | Sjøanemoner (Tealia sp.) |
| Echinodermata | Medusahode (Gorgonacephalus sp.) |
| Echinodermata | Sjøliljer (Heliometra sp.) |
| Bryozoa | Bryozoa (mosedyr) (Reteporella sp., Securiflustra securifrons) |
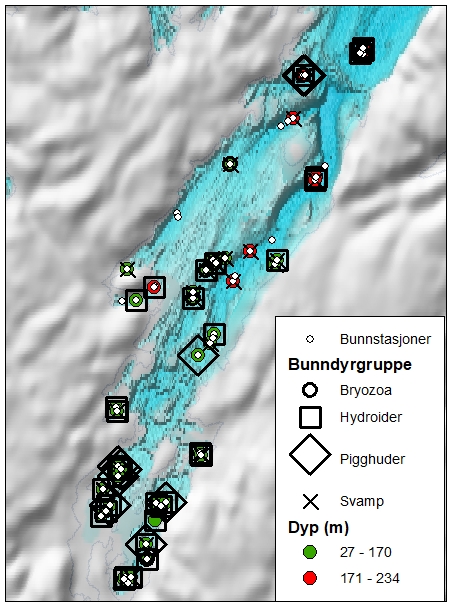
Sårbare biotoper har typisk store og skjøre arter av svamper, koraller og sjøfjær. Disse blir lett ødelagt av fiskeredskap med bunnkontakt, spesielt bunntrål. Oppreiste svamp og koraller brukes som sårbare arter av FAO og OSPAR. I Porsangerfjorden ble svamp registrert på alle de nordlige stasjonene og på alle stasjoner dypere enn 170 m (bortsett fra ytre Olderfjorden på vestkysten) (Figur 5.24). De største mengdene ble registrert på vestkysten. Blomkålkoraller ble funnet mest i sørlige deler av Porsangerfjorden og kun i relativt små mengder på stasjoner dypere enn 170 m (Figur 5.24). Kartleggingen som har pågått opp til 2019, tilsier at bassengene dypere enn 170 m langs østkysten av Porsangerfjorden er relativt lite sårbare overfor bunntråling, mens de dype områdene nordvest i fjorden har mye svamp og derfor kan være sårbare overfor bunntråling.
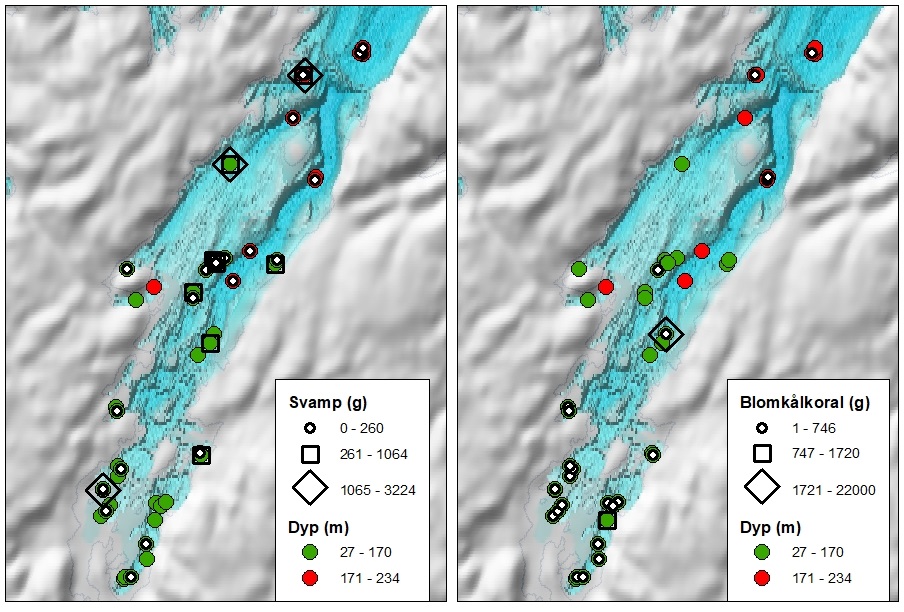
5.7.3 - Bunndyrsdata fra Mareano video-transekter nord i Porsangerfjorden
På tokt 2011105 i Barentshavet i 2011 kjørte Mareano fire videostasjoner med campod i ytre Porsangerfjorden (utenfor terskelen «Ryggen») i forbindelse med værligge i fjorden (Figur 5.25, Tabell V4.1, V4.2 i Vedlegg 4). To stasjoner var kystnært inn mot fjellsiden på østsiden av fjorden (732 og 733), og to andre var lenger inn i fjorden rett nord for Lille-Tamsøya (734 og 735). Stasjon 734 var en svært lang videolinje over og på begge sidene av terskelen nord for Lille-Tamsøya. Alle stasjonene hadde et gjennomsnittsdyp dypere enn trålegrensen på 170 m (Tabell V4.1).
Stasjonene ble ikke etter-opparbeidet slik Mareano vanligvis gjør for å sikre god kvalitet da stasjonene ikke var en del av Mareano-oppdraget. Observasjonene er derfor kun real-time-logget ombord i henhold til Mareanos feltrutiner. Mareano-prosjektet kan ikke bekrefte at registreringene er fullstendige. Dataene angir tilstedeværelse av taxa av epibentisk megafauna.
De fire videostasjonene ligger innenfor tre hovedtyper av sediment: 1) grove partikler (sand, grus), 2) grus, småstein og steinblokker og 3) sand, grus og småstein.
På begge stasjonene i nordøst (732 og 733) ble det registret risengrynskorall (Primnoa resedaeformis), sjøtre (Paragorgia arborea) og blomkålkorall (Drifa glomerata), samt den sårbare biotopen «svampspikelbunn» bestående av Geodia svamper (Figur 5.26). På transektene nord for Lille-Tamsøya ble det registrert flere individer enn på transektene i nordøst, og mange forskjellige svamper.
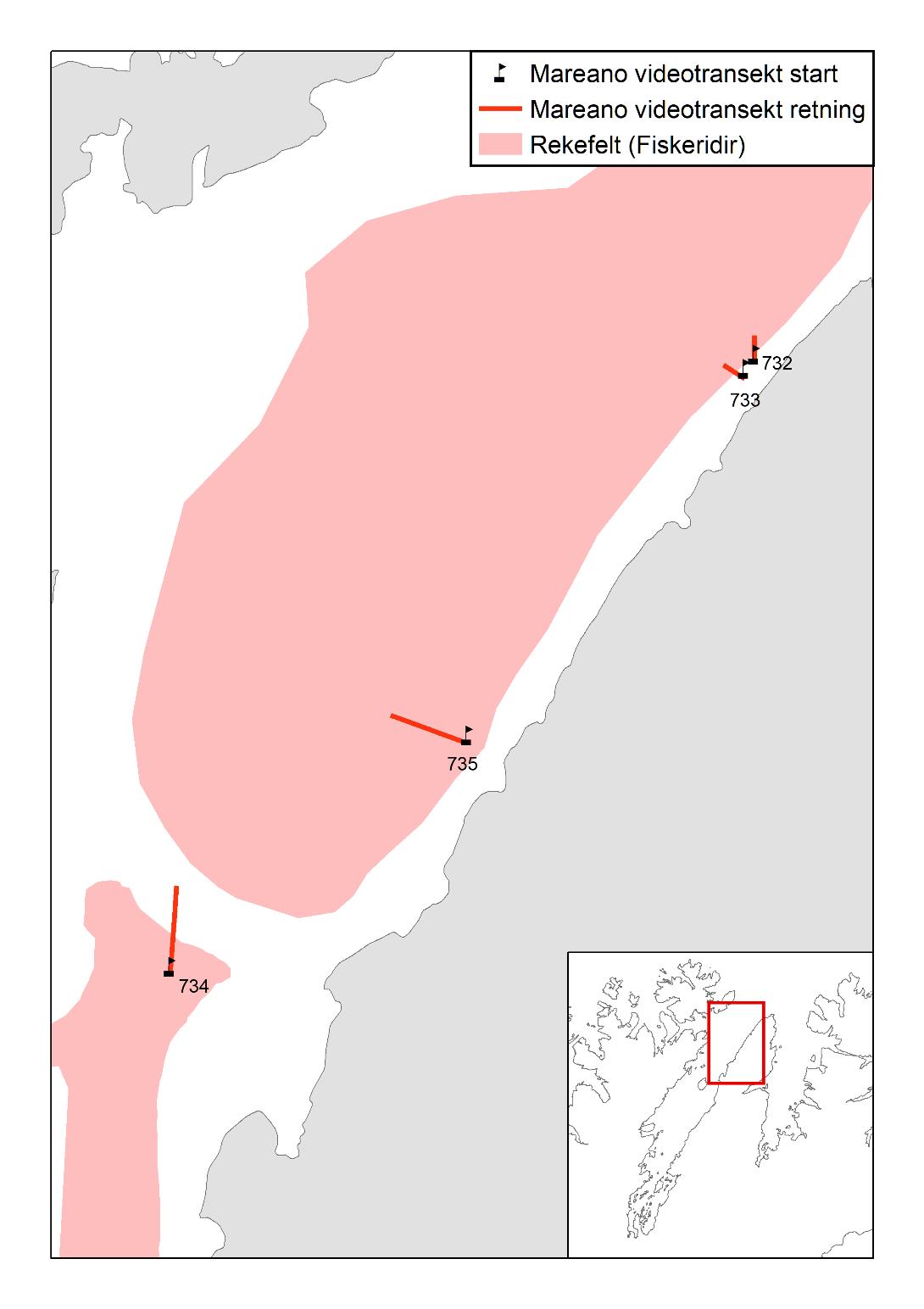
MAREANO registrerte koraller i ytre Porsangerfjorden (utenfor terskelen «Ryggen») langs østkysten, og høy biodiversitet og mange individregisteringer av bl.a. svamp nord for Lille-Tamsøya. Disse funnene indikerer at havbunnen i ytre Porsangerfjorden kan være sårbar for bunntråling, og en detaljert bunnkartlegging anbefales her.

5.7.4 - Bunndyrsdata fra kystøkotoktet i Porsangerfjorden og Kvænangen høsten 2019
På Havforskningsinstituttets kystressurstokt høsten 2019 ble det registrert bunndyr fra trålstasjoner i Porsangerfjorden og Kvænangen, men ikke fra Tanafjorden. Toktet (toktnummer 2019629) ble gjennomført med R/V Kristine Bonnevie 2.-24. oktober (Tabell V5.1 i Vedlegg 5). Arter og taxonomiske grupper av bunndyr ble registrert med antall og vekt i kg per trålstasjon (serienummer) (Figurer V5.1, V5.2, Tabeller V5.2, V5.3).
Det ble registrert høyere biomasse av bunndyr på trålstasjonene i Kvænangen sammenlignet med Porsangerfjorden, men det ble registrert flest sårbare arter på trålstasjonene i Porsangerfjorden (Figur 5.27, Tabeller V5.2, V5.3).
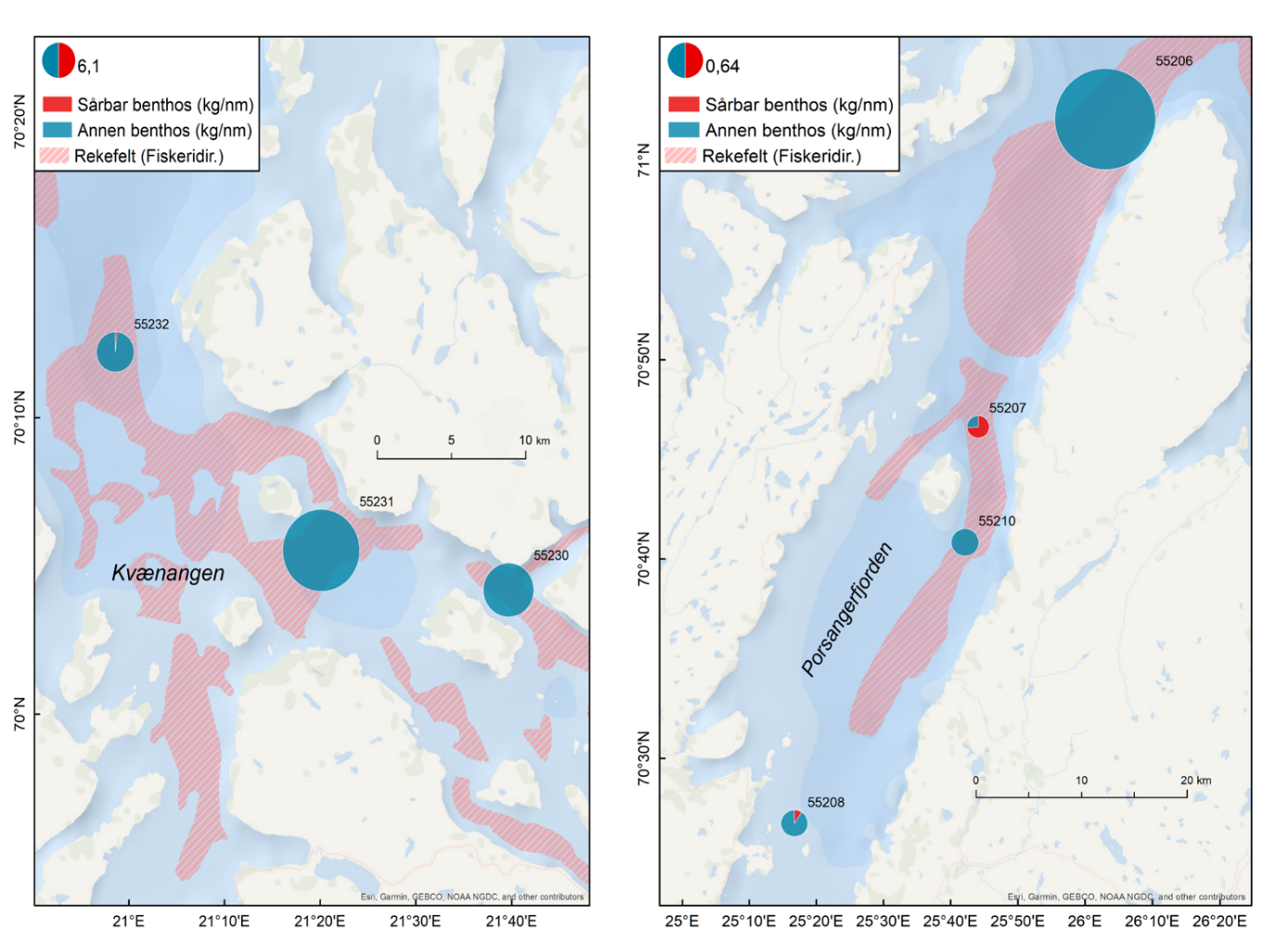
5.8 - Kongekrabbe
5.8.1 - Biologi, bestand og reguleringer
Kongekrabben (Paralithdodes camtschaticus, kamtsjatkakrabbe) er en kaldtvannsart som har sitt naturlige utbredelsesområde i det nordlige Stillehavet, men ble satt ut i Murmanskfjorden på Kolahalvøya på 1960-tallet av russiske forskere for å bygge opp en ny næring i området. I 1977 dukket de første individene opp i norsk farvann. Utsettingen av arten ble starten på et lønnsomt fiskeri i både russisk og norsk sone, og i dag er teinefiske etter kongekrabbe en viktig næring i Øst-Finnmark. Etter hummer er kongekrabben den best betalte arten i norsk fiskeri.
Kongekrabben kan oppnå en anselig størrelse. I dens opprinnelige leveområder har man funnet individer på opptil 10 kg. I norske farvann blir den sjelden tyngre enn 8 kg, og med en ryggskjoldlengde på opptil 23 cm. Kongekrabben er en kaldtvannsart og foretrekker temperaturer mellom 2 og 7 ºC. Arten finnes fra et par meters dyp og ned til 500 m, avhengig av alder, kjønn og årstid. De unge krabbene trives best på grov grus eller steinete underlag, mens de voksne individene foretrekker sand- og mudderbunn.
Om våren samler de voksne krabbene seg på grunt vann for å gyte og pare seg. Etter 2-4 måneder vandrer de gradvis mot dypere vannmasser, trolig for å finne mat. Krabbene overvintrer på dypt vann (300-500 m) før de igjen trekker mot grunnere områder på senvinteren og tidlig om våren.
Fra 1994 og frem til 2007 ble krabben i Barentshavet forvaltet som én bestand. Fra og med 2008 har Norge og Russland forvaltet krabben som to separate bestander i de respektive havområdene. Kongekrabben er på samme tid både en uønsket art og en verdifull fiskeressurs. Utbredelsesområdet til kongekrabben i norsk farvann er derfor delt i to, et kvoteregulert område øst for Nordkapp og et område med fritt fiske vest for Nordkapp. Innenfor det kvoteregulerte området ønsker forvaltningen et økonomisk bærekraftig fiske, noe som forutsetter en nokså stor bestand. Samtidig er det ønskelig å begrense spredningen ut av dette området, og fisketrykket settes derfor ganske høyt.
Havforskningsinstituttet overvåker bestanden av kongekrabbe (Figur 5.28). For å bestemme bestandsstørrelsen i det kvoteregulerte området gjennomføres et tokt med trål og teiner i fjordene i Finnmark, og et tokt med teiner i åpne havområder. For å overvåke spredningen vestover, gjennomføres det et teinetokt i fjordene vest for Nordkapp.
Både Porsangerfjorden og Tanafjorden ligger innenfor det kvoteregulerte området for kongekrabbe, og det er derfor interessant å evaluere mulige bestandseffekter av en eventuell åpning av disse fjordene for reketråling.
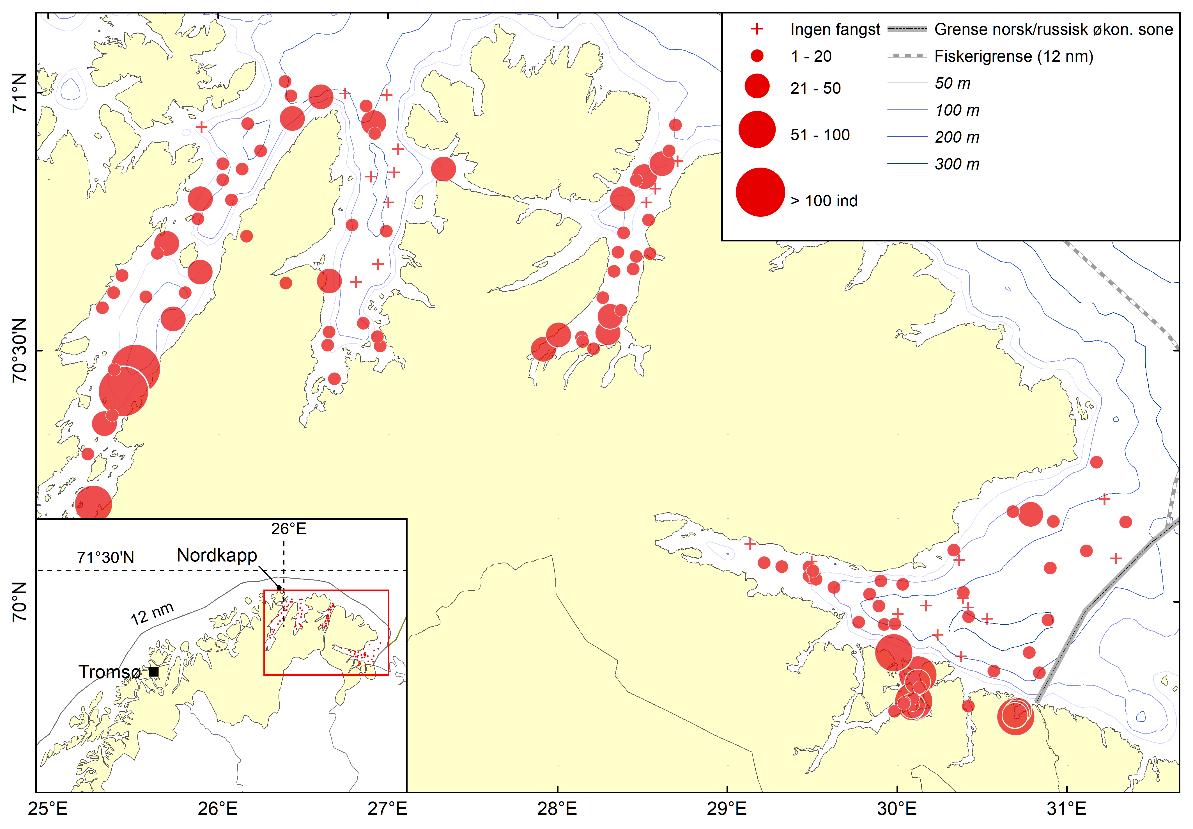
5.8.2 - Kongekrabbe som bifangst i rekefiske
Kartleggingstoktet med reketrål uten fiskerist i Porsanger- og Tanafjorden i 2018 og 2019 viste generelt veldig lite fangst av krabber sammenlignet med kongekrabbetoktet og kystressurstoktet, hvor det brukes hhv. Agassiz- og Camepelentrål (Figur 5.29). Dette forholdet gjelder også aktuelt reketrålingsdyp, dvs. områder dypere enn 170 m, hvor det med reketrålen på «Katla» bare ble fanget hhv. 0,2 og 0,3 krabber per trålte time i de to fjordene. Snittvekten på de fangede krabbene var ca. 1 kg.
Ved en eventuell gjenåpning av fjordene for reketråling vil det benyttes fiskerist, slik at kongekrabber i praksis ikke vil bli fanget i dette fiskeriet. Men selv om krabber i liten grad går inn i selve trålen, vil det være risiko for at trålen kjører over dem. For å få et estimat på antall krabber som muligens kan bli overkjørt per tråltime, ble det i Porsangerfjorden høsten 2018 gjennomført seks trekk med en ekstra trålpose under hovedtrålen. Den mulige overkjøringsraten viste seg å være mye høyere enn fangstraten, med et snitt på 17 krabber overkjørt per trålte time, og 18 krabber om man utelukkende legger til grunn de to stasjonene dypere enn 170 m.
En ytterligere indikasjon på overkjøringsraten får man ved å se på fangstratene til forskningsskipet R/V Johan Hjort, som foretok seks tråltrekk i Porsangerfjorden i samme periode som forsøkene med ekstra sekk ble gjennomført med «Katla» (oktober 2018). Det ble i disse seks trekkene fanget 514 krabber med snittvekt på 1 kg. Om vi sammenlikner antall krabber fanget i ekstra-sekken til «Katla» med antall krabber i trålen til R/V Johan Hjort, per trålte nautiske mil, er tallene henholdsvis 10 og 58. Forskjellene i fangstrate mellom de ulike tråltypene holder seg om man utvider sammenlikningsgrunnlaget.
Dette indikerer at ekstra-sekken på «Katla» ga et realistisk, men kanskje underestimert anslag av overkjøringsraten, og at denne er i størrelsesorden 50-100 ganger høyere enn fangstraten til krabber som ender i trålposen, når det ikke benyttes skillerist.
Vi har imidlertid ikke data på om krabber blir skadet eller drept når en reketrål dras over dem. Det viktigste trålfisket etter reker innenfor forvaltningsområdet for kongekrabber foregår i Varangerfjorden. Om vi sammenlikner tettheten av kongekrabber dypere enn 170 m i de fire fjordene Porsanger-, Lakse-, Tana- og Varangerfjorden finner vi den laveste tettheten i sistnevnte (Figur 5.30).
Oppsummert kan vi med stor sikkerhet slå fast at fiske med reketrål i de aktuelle fjordene vil føre til overkjøring av mange ganger flere krabber, enn antallet som havner i trålen, selv uten at det benyttes skillerist. Vi har imidlertid ikke grunnlag for å konkludere hvorvidt overkjøring vil skade eller drepe krabbene. Lavere fangster på dyp større enn 170 m i Varangerfjorden kan f.eks. skyldes at krabbeinvasjonen kom østfra og derfor har (over)belastet matressursene lengst i dette området, bifangst i annet fiskeri enn rekefisket, eller andre forhold vi ikke har kunnskap om. Men i og med at kongekrabbefisket er svært lukrativt, kan mulige nye faktorer som kan påvirke lønnsomheten i Porsanger- og Tanafjorden være verdt å se nærmere på.
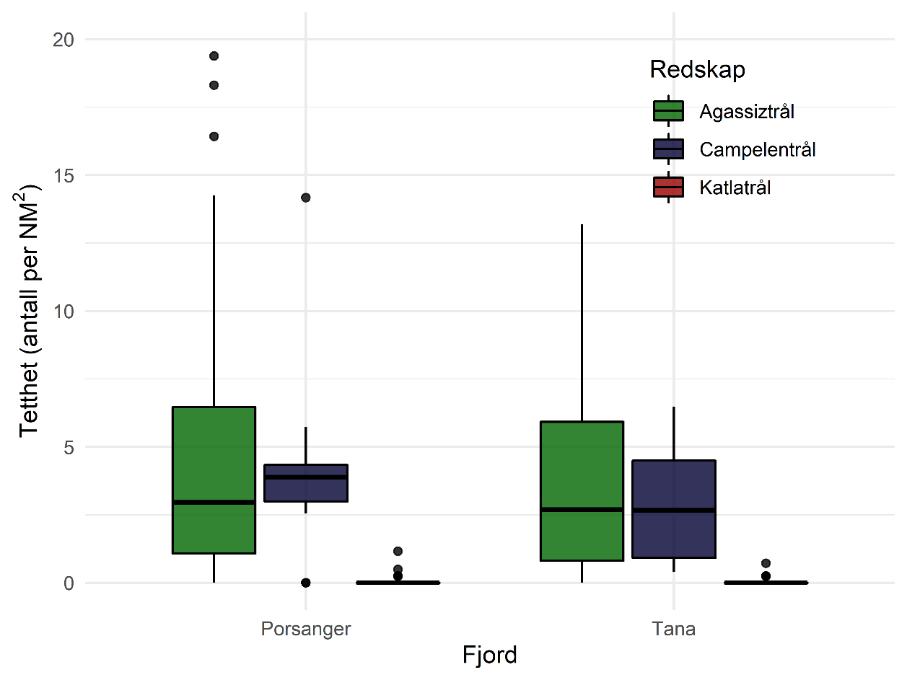
5.9 - Lodde – mulig lokal bestand i Porsangerfjorden
Lodde er torskens foretrukne byttedyr (ICES 2018) fordi den er energirik og lett å fange. Mye tyder på at Porsangerfjorden har sin egen bestand, som tidligere var viktig som mat for torsken og for de stedene fiskeriet etter torsk foregikk.
Lokale fiskere har identifisert områdene utenfor Sandvik og Prestøya som gyteområder for lodde (røde sirkler, Figur 5.31). Om våren kunne hunder vasse ut på fjære sjø rundt Trollholmene, og komme på land med levende lodde. Loddas gytefelter er plassert lenger ut enn torskens (blå sirkler), og de nyklekte larvene vil derfor sannsynligvis ikke fanges i strømvirvler på samme måte, men heller transporteres innover mot de kalde, indre overvintringsdelene av fjorden. Noen uker etter gytelodda går sannsynligvis unglodda ut og møter våren lenger ute i fjorden. Fisker Hedly Josefsen anslår området nord til Brenna/Kjæs på østsiden som aktuell nordlig utbredelse for lodde, fordi de fisket mye torsk med lodde i magen fra dette området og innover om våren og tidlig på sommeren. Denne observasjonen stemmer godt overens med de tetteste registreringene av de sannsynligvis lokale loddeforekomstene i fjorden (Figur 5.32). Høsten 2019 ble det filmet betydelige mengder loddeyngel i de indre, østlige bassengene i fjorden, noe som ytterligere underbygger fiskernes og fjordbrukernes oppfatning av en stedegen loddebestand.
Lodda er som sagt torskens foretrukne byttedyr, fordi den er energirik og lett å fange. Det er i utgangspunktet en livshistoriestrategi som borger for selvutslettelse. Men lodda øker sannsynligvis sjansen for å overleve fordi den er tilpasset et liv i kaldt vann. I de innerste delene av Porsangerfjorden, og nord i Barentshavet, er det områder med temperatur rundt 0 °C året rundt. Om torsken er sulten nok følger den etter, men fordøyelsen og omsetningen av byttet til vekst og nye gyteprodukter går tregt, og den vil lete etter alternative byttedyr. Analyser av øresteinene til torsk fanget i de kalde, indre bassengene av Porsangerfjorden, viste at de vokste seinere enn torsk fanget i de varmere delene lenger ut (Larsen 2010). Fiskernes og fjordbrukernes observasjoner tyder på at storseien, som i tidligere tider gikk inn i fjorden om somrene, i stor grad unngikk de indre, østlige, kalde og lodderike bassengene (Figur 5.31). Det kalde vannet gir sannsynligvis lodda en slags pause.
Undersøkelsene fra Epigraph-perioden (2008–2011) viser bare beskjedne loddeforekomster der i dag, men til gjengjeld finner man en betydelig rekebestand (Pedersen mfl. 2018). Våre kartleggingsdata fra 2018/2019 bekrefter dette bildet (Figur 5.33). Figur 5.34 viser tettheter og biomasse av lodde.
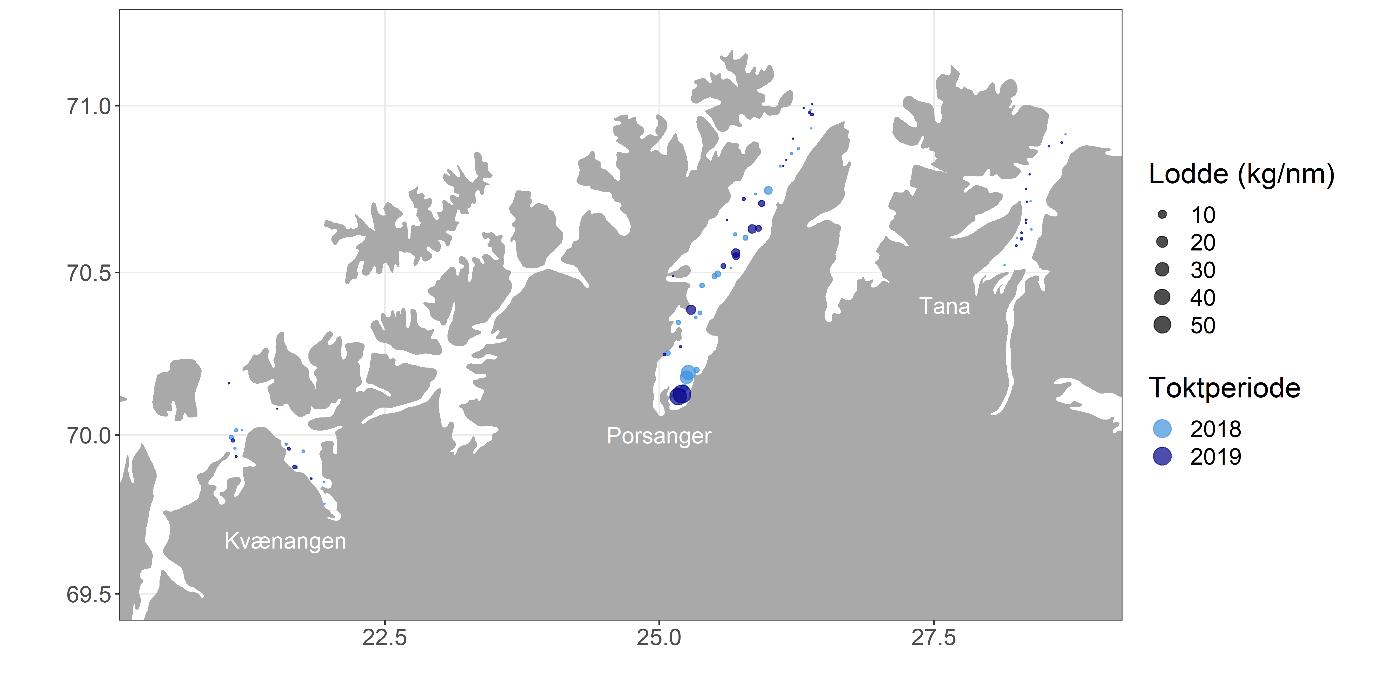
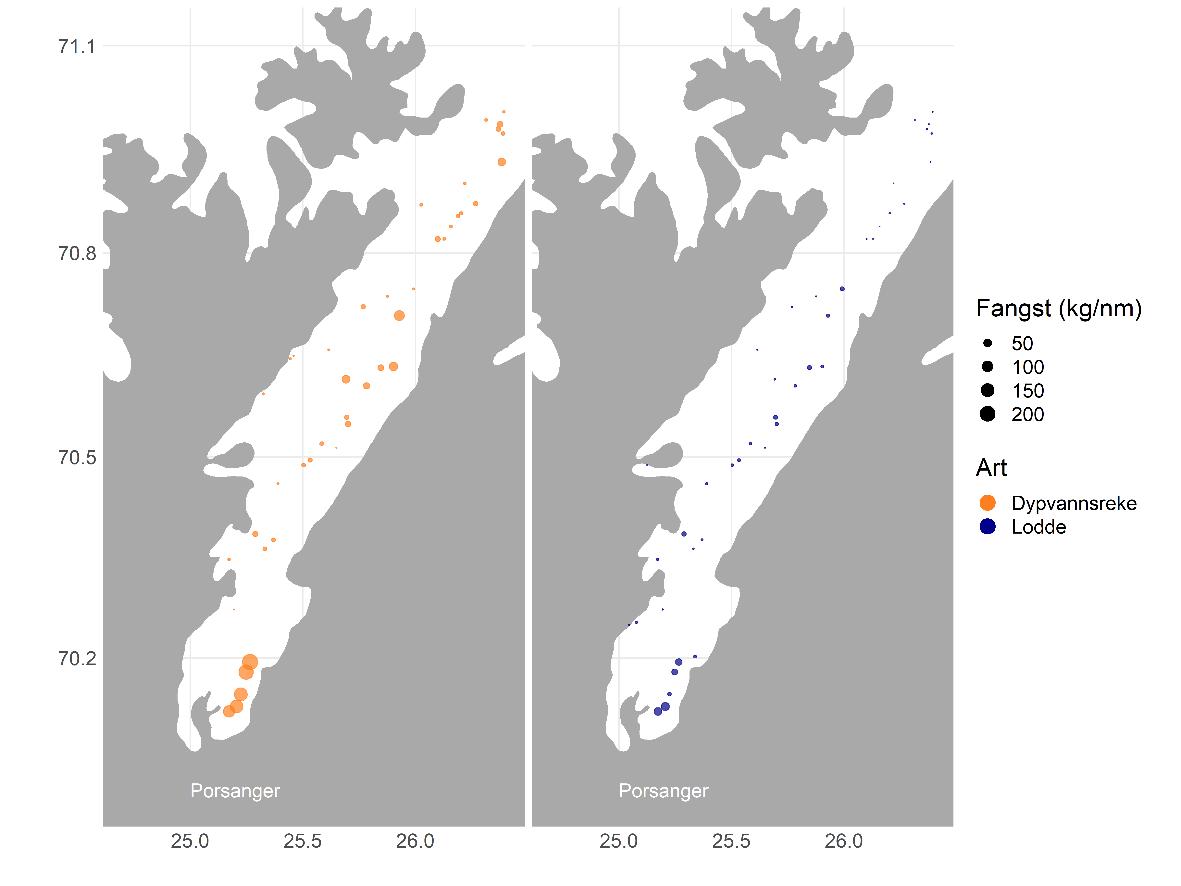
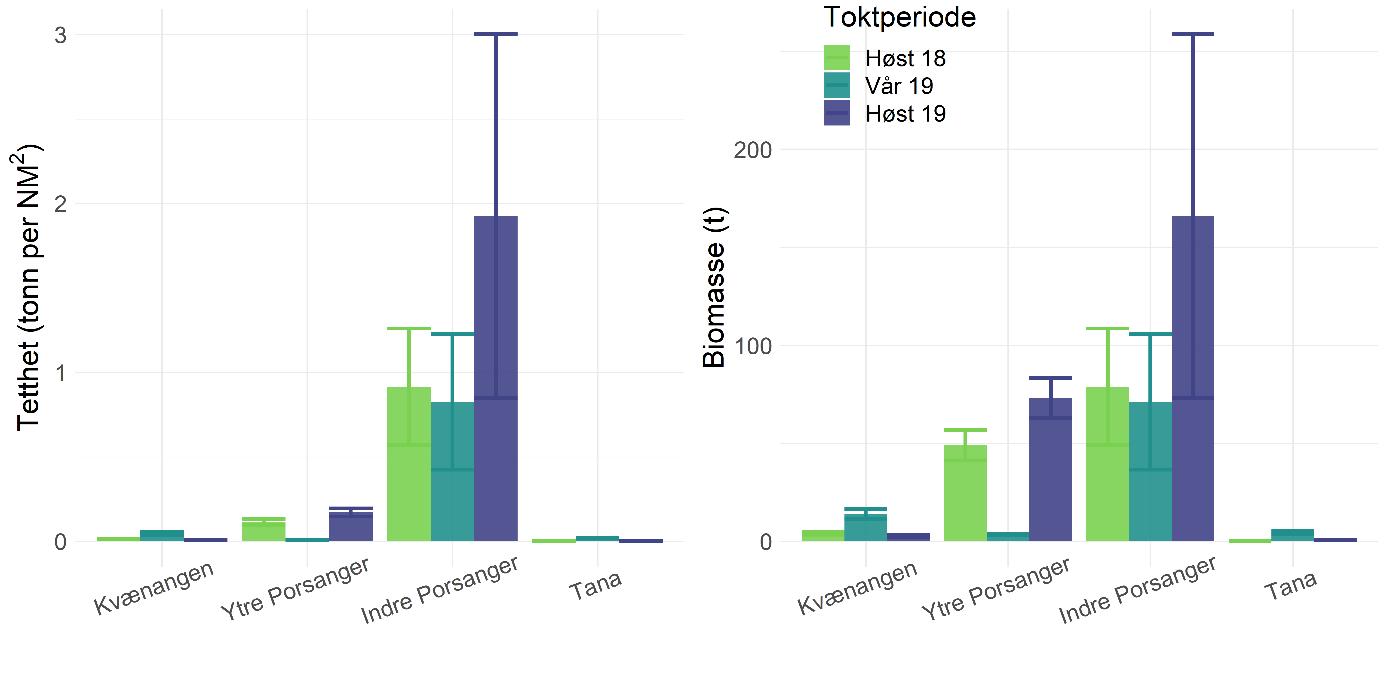
6 - Fiskerier
Data fra sluttseddelregisteret viser at landinger i alle de tre fjordene er dominert av torsk (Figur 6.1) og deretter følger sei og hyse. Alle andre arter bidrar relativt lite til totallandingene, selv om de er delvis regionalt viktige. Fiskerier som bør nevnes, er rekefisket i Kvænangen med en betydelig innsats hvert år (avsnitt 6.1.1), og som var grunnen til at Kvænangen ble valgt ut som referansefjord i denne undersøkelsen. I tillegg har et teinefiskeri på reke etablert seg i indre Porsangerfjorden over de siste årene (avsnitt 6.1.2), mens rekebestanden i Tanafjorden er helt ufisket. Regionalt relevant er også fisket etter kongekrabbe, spesielt i Porsangerfjorden som er et veldig viktig fangstområde for denne arten (avsnitt 6.2), og kveite. Andre bunnfisk som blir landet, er hovedsakelig brosme og uer, og i mindre grad en blanding av blålange, rødspette, skater og steinbit (avsnitt 6.3). Det foregår generelt veldig lite pelagisk fiske i disse fjordene, men de siste årene har NVG-sild flyttet overvintringsområdet sitt delvis inn i Kvænangen og Reisafjorden og det har resultert i betydelige sildelandinger derfra (avsnitt 6.4).
6.1 - Dypvannsreke
6.1.1 - Trålfiske i Kvænangen
6.1.2 - Teinefiske i Porsangerfjorden
De tre siste årene har det foregått et kommersielt teinefiske etter reker i Porsangerfjorden. Landingene presentert i denne rapporten er registrert på fangstfelt 80324 hos Råfisklaget, som inkluderer Vedbotn, Smørfjorden, Porsangen og Kåfjord. Kåfjord peker ut mot Magerøysundet, mens Vedbotn og Smørfjorden er sidefjorder i Porsangerfjorden. I praksis foregår fisket hovedsakelig i de indre, kalde bassengene i Porsangerfjorden, på høyde med og innenfor Reinøya, der det sjelden er dypere enn omtrent 100 meter. Teinefiske etter reker er et typisk vinterfiske, med hovedsesong fra oktober til årsskiftet (Figur 6.3).
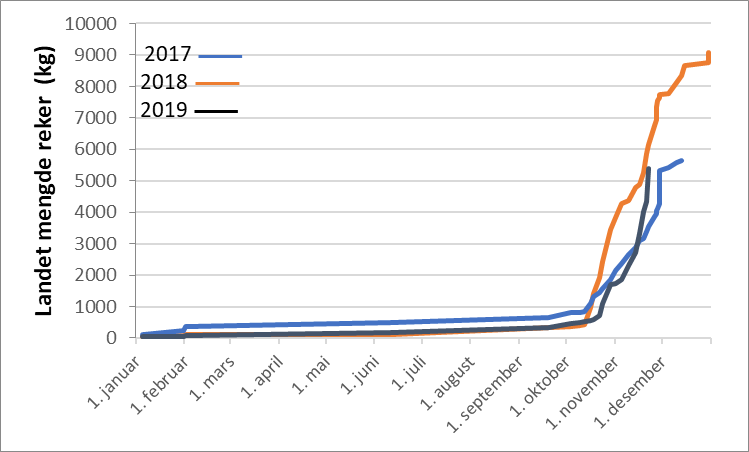
Ser man på fangstvolum, peker 2018 seg ut som det store teineåret i Porsangerfjorden, men året står ikke like sterkt med tanke på verdien av fiskeriet. I 2017 ble det landet i overkant av 5000 kg reker til en verdi av 450 000 kroner, noe som ga fiskerne en kilopris på rundt 80 kr. Året etter økte totalfangsten til litt over 9000 kg, men verdien av fangsten sank til under 400 000 kroner, og en kilopris på bare rundt 40 kroner. En mulig årsak til prisnedgangen fra 2017 til 2018 er bruk av teiner med finere maskevidde og dermed større innslag av dårligere betalte småreker. Med tall på andel småreker vil man kunne si noe sikrere om prisen for Porsangerreker er påfallende lav, men slike tall har vi ikke i dag. I 2019 var totalfangsten nede på nivået fra 2017 igjen, men kiloprisen var fortsatt bare halvparten av det som ble oppnådd det første fangståret.
Mens det ble fisket med inntil flere hundre teiner per båt i 2017 og 2018, innførte Fiskeridirektoratet en begrensning i innsats til 100 teiner per båt gjeldende fra 2019. Om man tar som forutsetning at hver landing representerer ett sjøvær, påvirket ikke begrensningen i antall teiner fangsteffektiviteten dramatisk (Figur 6.4). Mens hver båt i gjennomsnitt fisket 270 kg reker per sjøvær i 2018, ble dette tallet redusert til 260 kg reker i 2019. Antall båter som deltok i teinefisket etter reker i Porsangerfjorden varierte fra to båter i 2017, til syv båter i 2018, før antallet falt til tre båter i 2019.
Oppsummert ser det ut som om interessen for å delta i rekefisket med teiner har dabbet av, men at fangsten per sjøvær de to siste årene har holdt seg relativt stabil.
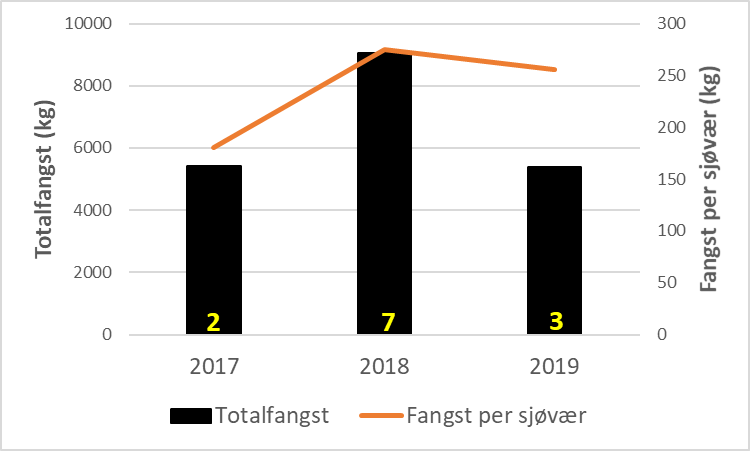
6.1.3 - Bifangst av fisk i reketrål
Tidligere var bifangst et stort problem i rekefisket pga. finmasket trålnett som fisket en stor mengde fisk og bunndyr. Reguleringstiltak, spesielt innføring av en obligatorisk fiskerist, men også midlertidig stenging av rekefelt med høye bifangstrater, har redusert problemet betydelig de siste 30 årene. Reketråling kan likevel påvirke fiskebestander negativt fordi fiskeyngel kan bli fanget til tross for skilleristen. Det er hovedsakelig bunnfisk under 12 cm som er sårbar, fordi de har samme størrelse som reker og derfor går gjennom fiskeristen og inn i trålposen. Slik bifangst avtar med størrelsen på fiskeyngelen opp til ca. 18 cm hvor all fiskeyngel sorteres ut og overlever.
Lengdebaserte bestandsestimater i Porsanger- og Tanafjorden bekrefter at en relativ liten andel av kommersielt viktige fiskebestander som hyse og torsk er sårbar for reketråling (Figur 6.5). Det er hovedsakelig torske- og ueryngel under 10 cm som kan bli påvirket, fordi kartleggingstoktet viser betydelige forekomster av disse i Porsanger- og Tanafjorden. Ved en størrelse under 10 cm går nesten alle fisk gjennom fiskeristen og blir derfor fisket av trålen. Toktdata viser at det er et mindre problem for hyse fordi andelen hyse med sårbar lengde var lavere på trålbar bunn (dyp ≥ 170 m).
Estimerte gjennomsnittlige bifangstrater i en reketrål tilsvarende den som ble brukt på «Katla» men med skillerist, ligger for hyse og torsk stort sett under 0,5 kg per nautisk mil og enda lavere for uer (Figur 6.6). Det er omtrent 1 % eller mindre av fangstratene for de to artene som ble registrert uten skillerist, og betyr at risten er veldig effektiv når det gjelder å redusere uønsket bifangst av fisk. Men det finnes også betydelig variasjon på tvers av fjordområder, toktperioder og trålstasjoner. Det gjelder også mht. forholdet mellom estimert yngelbifangst og rekefangsten (Tabell 6.1). Prosentandelen av hver av de tre artene ligger stort sett under 2 % av rekefangsten, men det finnes flere høyere andeler, spesielt i Tanafjorden hvor bifangstandelen av hyseyngel i gjennomsnitt var 11,5 %. Det viser at veldig høye tettheter av hyse i Tanafjorden, spesielt høsten 2018, resulterte i betydelig høyere bifangstrater, noe som understreker at yngelbifangsten kan være betydelig ved høye tettheter av fiskeyngel i et område. I tillegg kan bifangst av fiskeyngel ha negative effekter selv om fangstraten i absolutt vekt er lav. Det er spesielt relevant mht. uer fordi den er en langtlevende art som er veldig avhengig av enkelte store årsklasser, og hvis bifangstdødelighet reduserer rekrutteringen, kan det skade bestanden på lang sikt.
Resultatene bekrefter at skilleristen generelt klarer å holde fiskebifangsten på et lavt nivå, men at eventuelle store tettheter av fiskeyngel på rekefelt likevel kan resultere i betydelige bifangstmengder. Det understrekes at Fiskeridirektoratets praksis med å midlertidig stenge rekefelt med høye bifangstrater av fiskeyngel er et viktig verktøy for å holde fiskebifangsten på et lavt nivå.
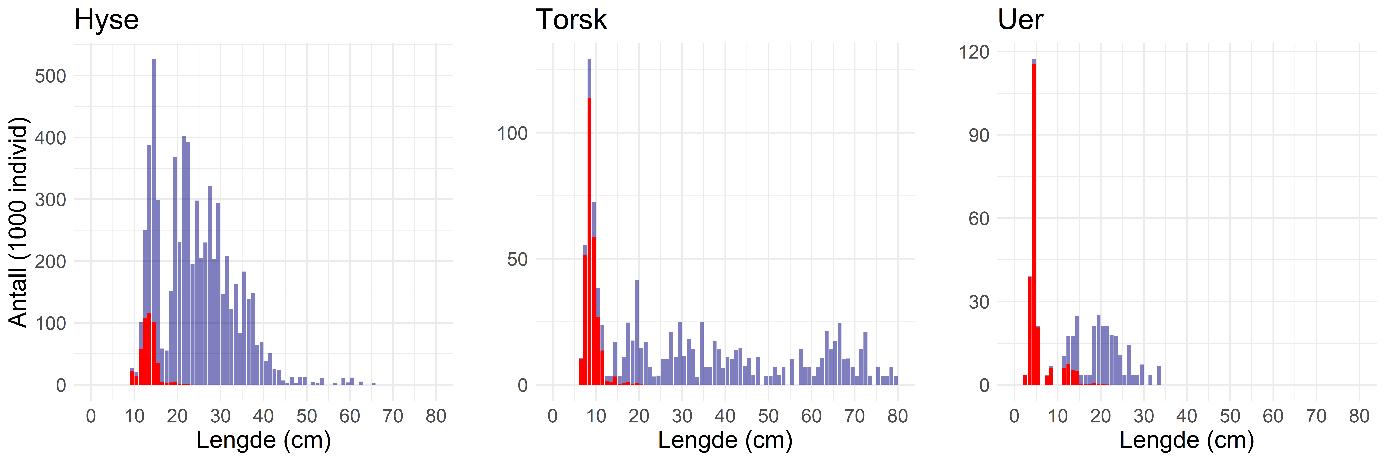
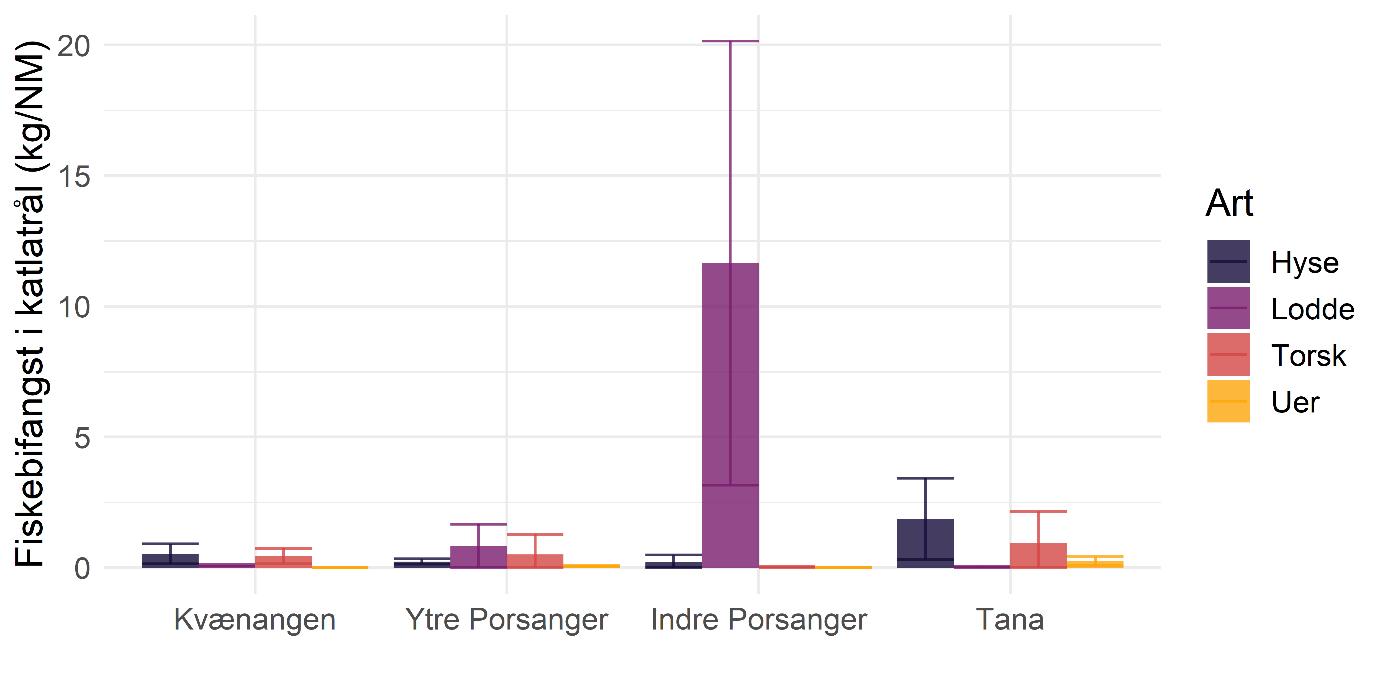
| Art | Fjordområde | Bifangst (kg/NM) | Rekefangst (kg/NM) | Bifangstrate (vekt %) | Bifangstrate (fisk per kg reke) |
| Hyse | Kvænangen | 0,53 | 24,24 | 2,17 | 0,8 |
| Tanafjorden | 1,86 | 16,13 | 11,55 | 5,41 | |
| Ytre Porsangerfjorden | 0,23 | 11,41 | 2,06 | 0,81 | |
| Indre Porsangerfjorden | 0,21 | 88,52 | 0,24 | 0,05 | |
| Lodde | Kvænangen | 8,30 | 0,09 | 0,38 | 0,34 |
| Tanafjorden | 2,42 | 0,03 | 0,16 | 0,15 | |
| Ytre Porsangerfjorden | 74,53 | 0,82 | 7,18 | 6,53 | |
| Indre Porsangerfjorden | 11,66 | 88,52 | 13,17 | 11,97 | |
| Torsk | Kvænangen | 0,44 | 24,24 | 1,83 | 1,7 |
| Tanafjorden | 0,95 | 16,13 | 5,87 | 6,98 | |
| Ytre Porsangerfjorden | 0,5 | 11,41 | 4,42 | 7,36 | |
| Indre Porsangerfjorden | 0,02 | 88,52 | 0,02 | 0,03 | |
| Uer | Kvænangen | 0,01 | 24,24 | 0,02 | 0,01 |
| Tanafjorden | 0,26 | 16,13 | 1,59 | 1,06 | |
| Ytre Porsangerfjorden | 0,06 | 11,41 | 0,54 | 0,4 | |
| Indre Porsangerfjorden | 0 | 0 | 0 |
6.2 - Kongekrabbe
Det kommersielle fisket etter kongekrabbe startet i 2002. I oppstarten ble det kun fisket etter hannkrabber. I 2008 ble det åpnet for fiske etter hunnkrabber. Antall deltagende fartøyer i det kvoteregulerte fisket ble mer enn fordoblet fra 2007 til 2019 og utgjør nå nærmere 650 fartøyer. Disse fartøyene skal dele en kvote som i samme periode har variert mellom 1000 og 2600 tonn. Myndighetene har varslet at adgangen til å delta i krabbefisket skal utvides til også å gjelde fiskere bosatt i Vest-Finnmark, slik at krabberessursen framover vil gi et bidrag til inntjeningen for enda flere båter.
Varangerfjorden har inntil nylig vært det viktigste fangstfeltet. De siste par årene har imidlertid de største landingene kommet fra Porsangerfjorden, og disse økte til nesten 900 tonn i 2018, før de falt til rundt 600 tonn i 2019 (Figur 6.7). Landinger fra Tanafjorden er betydelig lavere, mellom 100 og 155 tonn over de siste årene. Fiskedødeligheten i kongekrabbefisket økte utover 2000-tallet og særlig etter 2008 har den vært høy. Fiskerne får høyest pris for de største krabbene, noe som sannsynligvis bidrar til at det er de største hannkrabbene som hele tiden blir tatt først. Dette kan være årsaken til at gjennomsnittsstørrelsen av hannkrabber over minstemålet er blitt betydelig lavere etter år 2000. En stadig større andel av krabbene blir omsatt levende. Dette gir gode priser til fiskerne, og verdien av dette fiskeriet har gjort det svært viktig både for enkelte lokalsamfunn og for hele Øst-Finnmark.
Kongekrabben i Porsangerfjorden hadde tidligere en tydelig årstidsvandring mellom grunne og dype områder. I desember kom gjerne store hanner helt grunt, og fiskeriene fulgte etter nesten inn i fjæresteinene. Fra mars og utover var det gyting og skallskifte hos hunner og ikke-kjønnsmodne hanner i de grunnere delene av fjorden. Store, kjønnsmodne hanner, som skal pare seg, skifter ikke skall før parringen er over. Det betød at fra desember og utover på nyåret var det mye krabber som stod grunt, og fra mars/april og utover var det mange krabber som i tillegg akkurat hadde skiftet skall og derfor hadde dårlig kjøttfylde. Fra september og utover trakk gjerne krabbene dypt igjen, ned på det som ble beskrevet som «leira». Kvaliteten på krabbene var på dette tidspunktet god, og fiskeriene konsentrerte seg ofte i de dypeste områdene av fjorden.
I dag synes vandringsdynamikken mellom dype og grunne områder å være mindre regelmessig. Det kan skyldes at det generelt er mindre mat tilgjengelig for krabbene, som bunndyrundersøkelsene i denne rapporten kan tyde på (Figur 5.22). De store hannkrabbene skifter gjerne ikke skall hvert år. Hyppigheten av skallskifte vil sannsynligvis i stor grad påvirkes av fødetilgangen, og det vil igjen kanskje påvirke vandringen. Om høsten vil i tillegg kanskje de dype områdene friste mindre for enkelte deler av populasjonen, enn grunnere områder lenger inne i fjorden, som ble senere invadert og derfor sannsynligvis inneholder mer mat. Med mindre mat tilgjengelig vil sannsynligvis flere hannkrabber bruke lenger tid mellom skallskiftene, og dermed vil de vandre mer. I tillegg kan man paradoksalt nok oppleve å få krabber av god kvalitet på alle tider på året. For å oppsummere, så har de største krabbene tidligere stått dypt fra september og fram mot jul, mens bildet er mer sammensatt i dag.
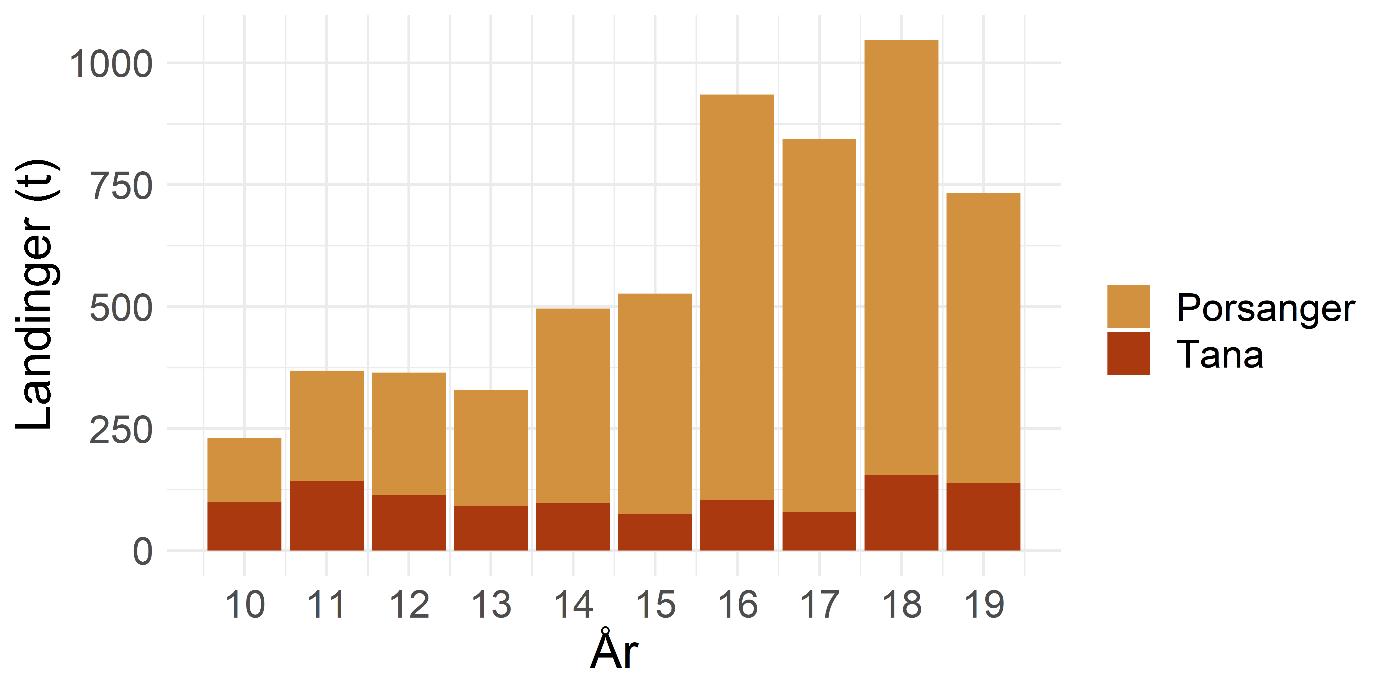
6.3 - Bunnfisk
Bunnfisk utgjør den største andelen av landinger på tvers av de tre fjordene, og for alle år mellom 2010 og 2019, gjennomsnittlig fra 76 % (Kvænangen) opp til 93 % (Tanafjorden). Torsk er den viktigste av alle bunnfiskarter (Figur 6.8) og bidrar i gjennomsnitt med mer enn halvparten av alle bunnfisklandinger, opptil 80 % i noen år i Tanafjorden, og med mer enn 4000 tonn per år i Porsangerfjorden over de siste tre år. Sei og hyse er de to andre artene med betydelige landinger som ligger omtrent på det samme nivået i Porsangerfjorden (gjennomsnittlig 15-16 % av alle landinger) og Tanafjorden (12-13 %), mens det finnes mye mer sei enn hyse i Kvænangen (hhv. 16 % og 5 %). Det har vært en ganske stor variasjon i landingene gjennom de siste 10 årene, spesielt for sei som f.eks. bidro med mer enn 50 % av landingene i 2013 sammenlignet med faktisk 0 % de siste årene. Landinger av alle andre bunnfisk er marginale og hovedsakelig et resultat av bifangst i torskefisket. Mest relevant i denne sammenhengen er brosme, mens alle andre arter forekommer i veldig små kvantum. Unntaket er kveite som ikke blir tatt i store mengder, men som likevel er en kommersielt viktig art, spesielt i Porsangerfjorden.
Torskefisket viser store sesongmessige svingninger, mens landinger av andre bunnfisk er mye mer jevnt fordelt gjennom året (Figur 6.9). Dette er knyttet til gytevandring av skrei, som resulterer i en stor økning av fangster om vinteren og våren, spesielt i mars og april. Torskelandingene er høyest i Porsangerfjorden, og der blir mesteparten av torsken fanget på nordvestsiden av ytre delen (fra Smørfjorden og utover) og i Magerøysundet. Magerøysundet er ikke en del av Porsangerfjorden, slik vi har definert dette fjordområdet (Figur 3.2), men er inkludert i den samme statistiske lokasjon. (Statistisk lokasjon Porsangerfjorden (03-24) omfatter hele Porsangerfjorden samt Magerøysundet.) Det er derfor ikke mulig å skille landinger i Porsangerfjorden fra landinger i Magerøysundet i de offisielle landingsdataene. Men landingsdata per mottaksstasjon viser at nesten all torsk har blitt levert til mottaksstasjonen i Nordvågen ved Honningsvåg, og det kan antas at disse kommer hovedsakelig fra Magerøysundet.
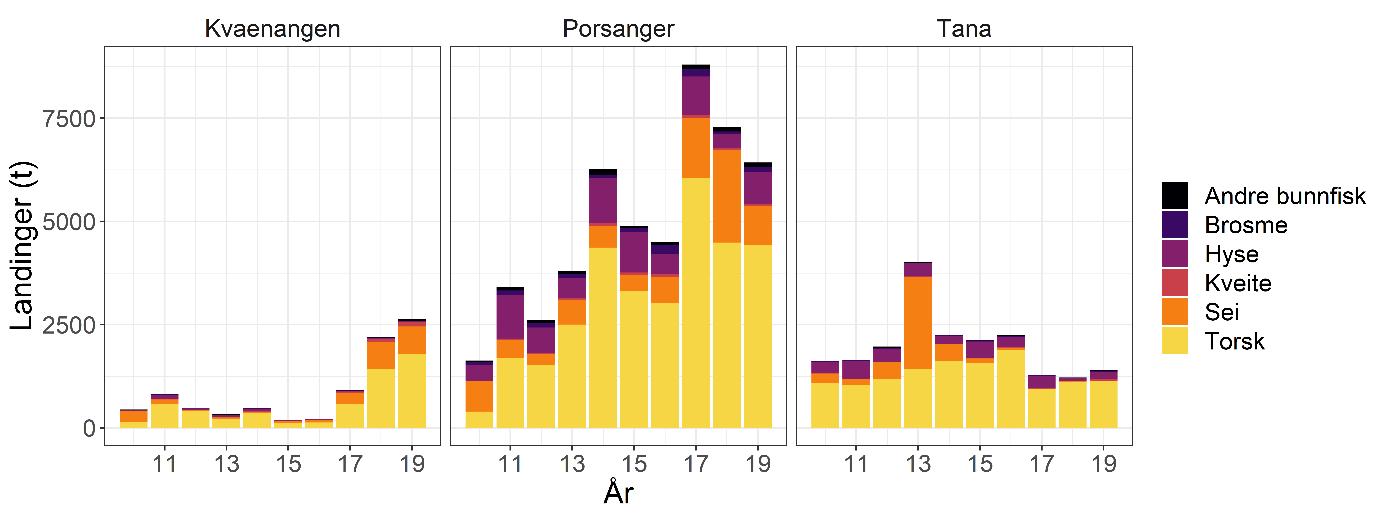
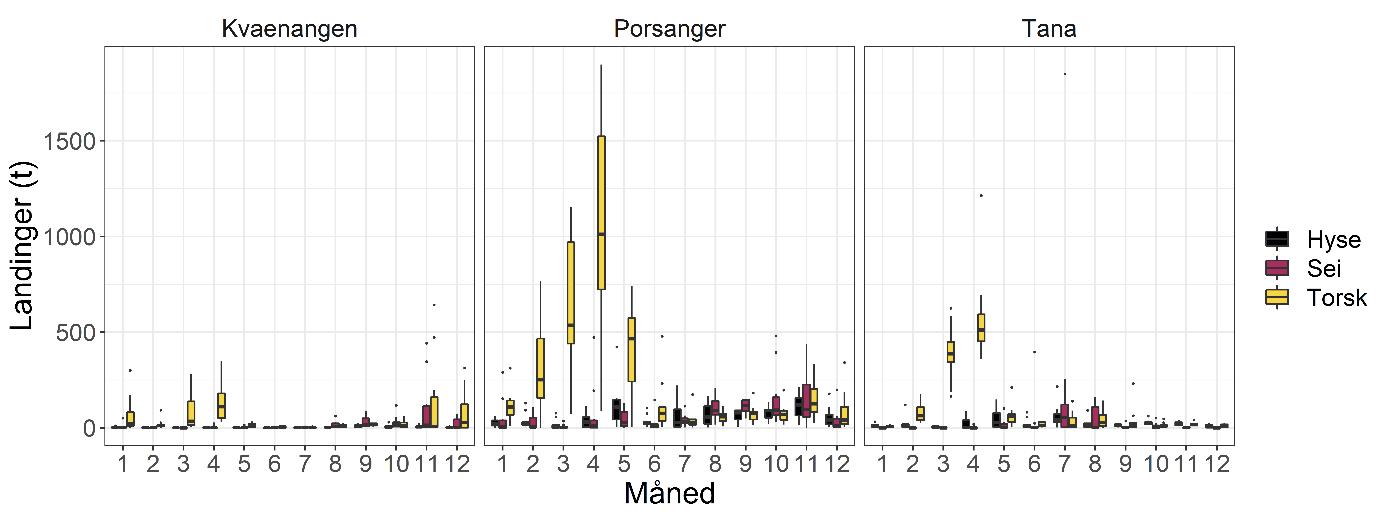
6.4 - Pelagisk fisk
Pelagisk fiske er stor sett helt ubetydelig i de tre fjordene. Unntaket er fisket etter NVG-sild som har delvis overvintret i Kvænangen de siste tre årene. Landingene har da økt enormt, opp til 107 100 tonn i 2019. Endringer i overvintringsdynamikken til NVG-sild fører til forandringer også i kystfisket på bestanden og resulterer i en midlertidig sterk vekst i landinger rundt overvintringsområdet. Men dette er en eksepsjonell fase og derfor ikke representativt for et vanlig fiske i en fjord over et lengre tidsrom. Bortsett fra NVG-sild finnes det veldig lite landinger av pelagisk fisk i de tre fjordene. Den eneste arten med landinger av betydning de siste årene er rognkjeks med rundt 176 tonn i Kvænangen og 103 tonn i Porsangerfjorden i 2019. Det ble også landet 133 tonn og 170 tonn av lodde i hhv. Kvænangen og Porsangerfjorden i 2001, men deretter har fisket etter lodde forsvunnet fra fjordene.
7 - Resultater fra teineundersøkelser
Teineundersøkelsene ble kjørt parallelt med kartleggingstoktet med trål høsten 2018 og våren 2019. Hovedformålet med disse undersøkelsene var å undersøke mulige rekeforekomster på ikke-trålbar bunn utenfor de kommersielle rekefeltene og sammenligne fangster der med teinefangst på trålbar bløtbunn. På bløtbunn ble teinestasjonene plassert i de samme posisjonene som trålstasjonene for å kunne gjøre en direkte sammenligning av fangstrater mellom teiner og trål. Totalt ble 181 teinestasjoner tatt, 87 høsten 2018 og 94 våren 2019, men kun på 46 % av teinestasjonene ble det registrert reker (Figur 7.1), noe som resulterte i et stort antall nullfangster.
Fangstraten i teinene var generelt lav og lå i gjennomsnitt på under 10 reker per teinedøgn på tvers av hele toktet, unntatt i indre Porsangerfjorden (Figur 7.2). Fangstratene i de to østlige bassengene i indre Porsangerfjorden var med gjennomsnittlig 70 reker per teinedøgn, og et maksimum på 361 reker per teinedøgn, betydelig høyere enn i alle de andre områdene. Fangstratene på trålbar bunn var gjennomført høyere sammenlignet med ikke-trålbar bunn. Resultatene viser også en nedgang i fangstrate fra høst til vår, men da de to toktperiodene ikke dekket de samme stedene er det ikke mulig å konkludere om forskjellen er pga. årstiden eller prøvetakingen (avsnitt 7.5). Statistisk analyse viser at fjordområde, toktperiode og bunndyp best forklarer variasjonen i dataene, og av disse er fjordområde den klart viktigste faktoren. Sammenhengen mellom fangst og bunndyp er positiv og signifikant, men svak og påvirket av problemer med å bruke teiner på stasjoner dypere enn 200 m.
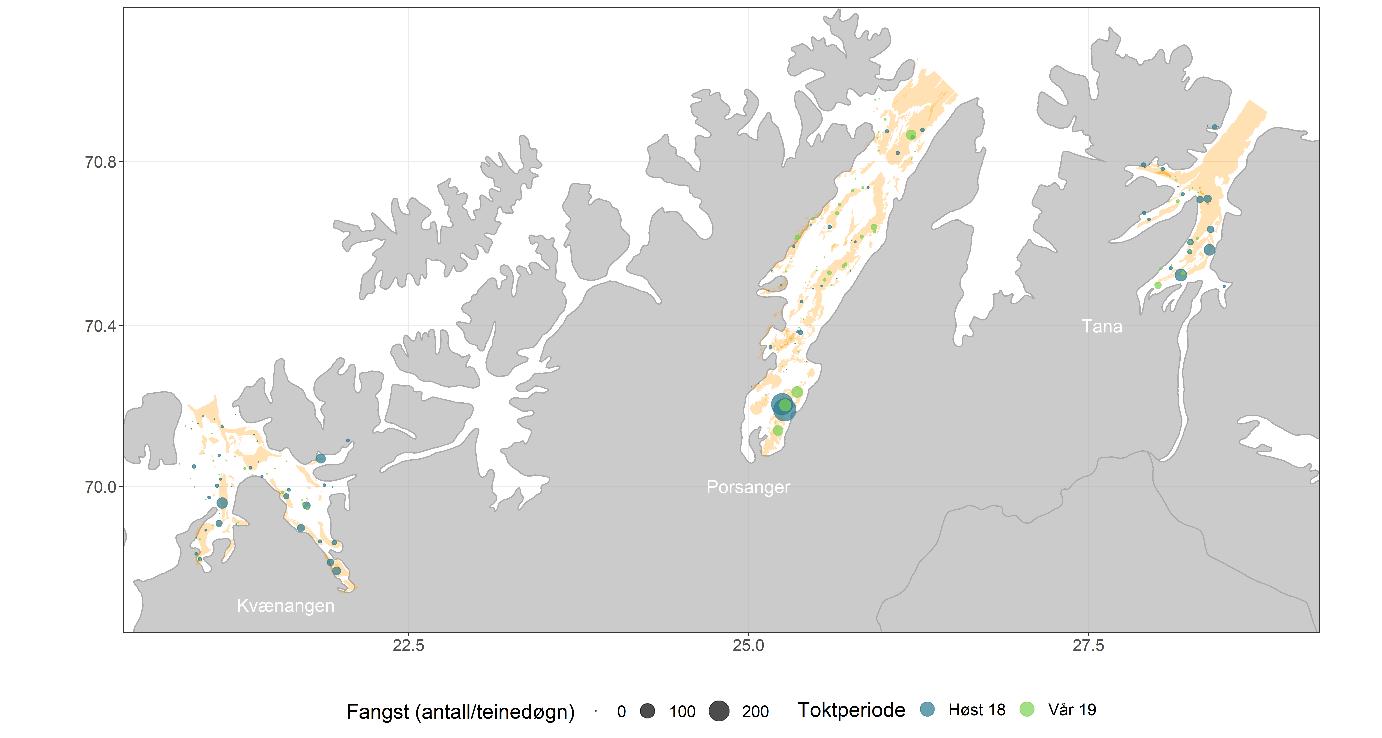
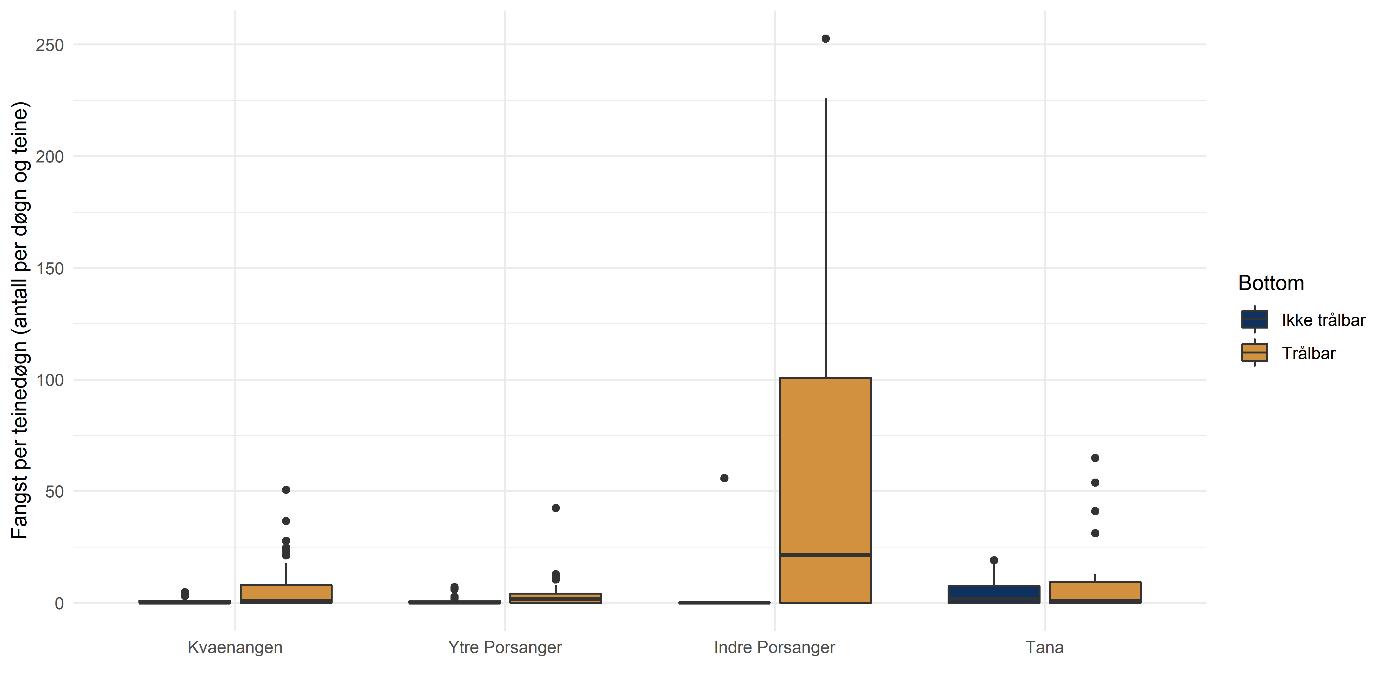
7.1 - Nye potensielle områder for et teinefiske etter reke
Teineundersøkelsen viser mye lavere fangstrater i alle undersøkte områder sammenlignet med indre Porsangerfjorden. Dette indikerer at teinefiske etter reker vil gi et lavt utbytte utenfor de spesielle forholdene i indre Porsangerfjorden. Men enkelte betydelige fangster, spesielt i Tanafjorden, viser at flere undersøkelser er nødvendig. Det ser ut til å være en trend at fangstratene er høyere i indre, beskyttede fjordområder, dvs. i områder som er velegnet for teinefiske. Denne trenden kan delvis forklares med problemer under teineundersøkelsene på dype og utsatte områder hvor teinene sannsynligvis ble negativt påvirket av strøm (veltet eller dradd avgårde). Dagens erfaring med teinefiske etter reker er fortsatt begrenset, og forbedringer i design og bruk av teiner kan derfor øke fangstpotensialet. Mer forskning er nødvendig for å vurdere om andre områder enn indre Porsangerfjorden har potensiale for et teinefiske etter dypvannsreke.
7.2 - Fangstrate
En god del av teinestasjonene ble lokalisert på samme sted eller i umiddelbar nærhet av trålstasjonene. Det gjorde det mulig å direkte sammenligne fangstratene på reke i hhv. trål og teiner, og derfor vurdere om teinefangster samsvarer med rekefangster i trålen, som er antatt å gi et representativt bilde av reketetthet. Dette er et relevant spørsmål siden teiner kan benyttes på ikke-trålbare områder som er utilgjengelige for tradisjonelle fiskerifaglige bestandsvurderingsmetoder.
Toktdata viser at det finnes en signifikant positiv korrelasjon mellom teine- og trålfangster på samme sted (Figur 7.3). Men resultatene antyder også at sammenhengen ikke er veldig nøyaktig pga. stor variasjon og fordi resultatet hovedsakelig er bestemt av noen veldig høye fangster i indre Porsangerfjorden, mens korrelasjonen i alle andre områder ikke er veldig tydelig. En grunn er en stor andel av nullfangster på mange teinestasjoner. Konklusjonen er derfor at teinefangster kan gi en pekepinn på rekeforekomst og -tetthet i et område, men at resultatene er beheftet med stor variasjon og usikkerhet og derfor må tolkes forsiktig.
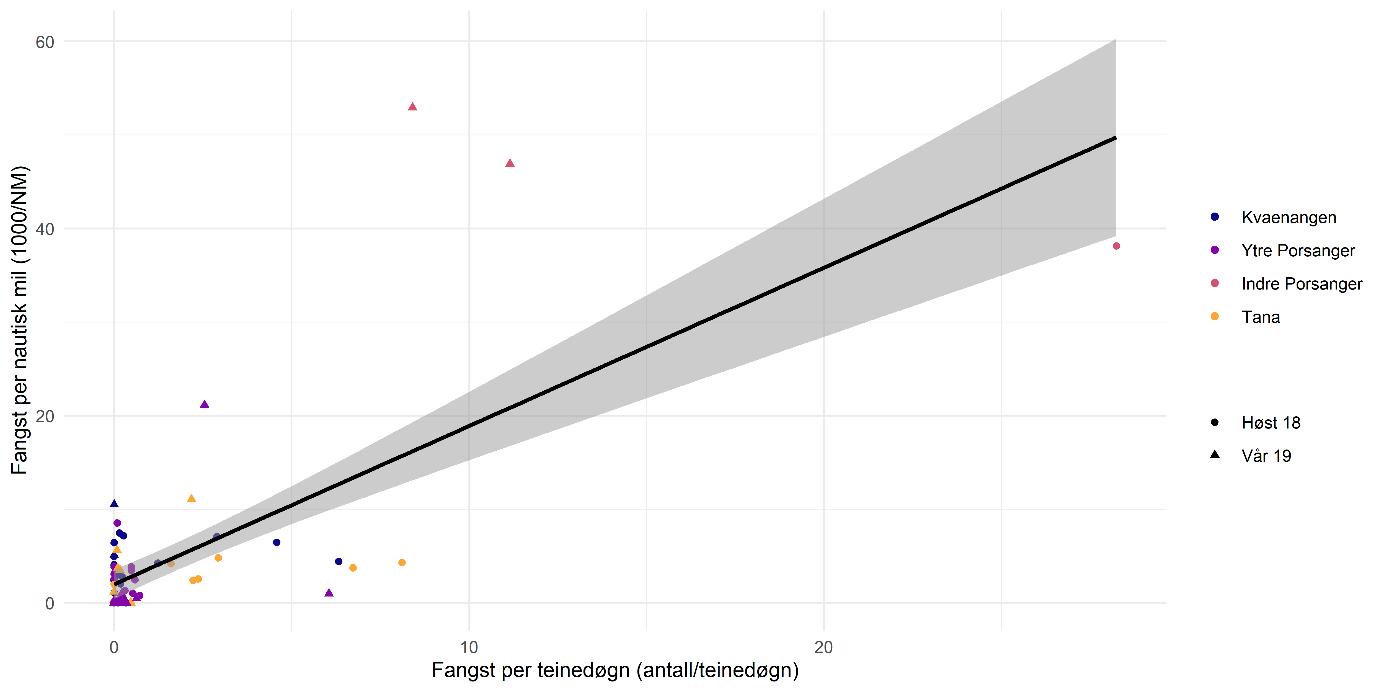
7.3 - Innsatsbegrensning – indre Porsangerfjorden
Kystnært teinefiske etter reke i kommersiell skala er et relativt nytt fenomen i norsk sammenheng. Det betyr at teiner, agn og fiskemetoder er under utvikling, og fisket må forventes å kunne gi økte fangster framover, og kanskje også å kunne utvides til områder som i dag ikke er kommersielt interessante pga. for lave fangster. F.eks. annonserer Frøystad med et nyutviklet agn, som ifølge produsenten gir økte fangster i forhold til tradisjonelle agntyper som pellets og sild. Teinefiskere i Finnmark eksperimenterer også med å heve teinene opp fra bunnen for å øke fangstraten. Det må forventes at næringen innoverer ytterligere før metoder og fangstrater stabiliseres noenlunde gitt et kjent bestandsgrunnlag.
Foreløpig finner det bare sted et beskjedent uttak av reker gjennom teinefisket i Porsangerfjorden. Ettersom dette er en ny næring, er det også usikkert om det vil etableres en verdikjede som er økonomisk bærekraftig på sikt. Etter vår vurdering er det bestandsgrunnlag for å øke uttaket, og vi foreslår også at maksimalt antall teiner som kan benyttes per båt, økes fra dagens 100 til 150 teiner, og videre at man følger med på utviklingen i bestanden og fangstene framover. Se ellers kapittel 10 for råd fra Havforskningsinstituttet om teinefisket etter reker.
7.4 - Bifangst av andre arter i reketeiner
Teineundersøkelsene viser at teinetypen og agnet som ble brukt, fanger reker veldig selektivt (Figur 7.4). Mer enn 95 % av de registrerte individene ble identifisert som dypvannsreke eller reker i familien Pandalidae i begge toktperiodene og alle fjordområdene. I de fleste tilfellene bestod 99-100 % av fangsten av pandalide reker inkl. dypvannsreke. Det var veldig lite bifangst, og bifangsten var hovedsakelig en blanding av enkelte individer av bunndyr, bl.a. kongekrabber (totalt 4 stk.), småkrabber (pyntekrabbe og eremittkreps), kongsnegl, trollhummer og slangestjerne. Bifangsten inkluderte veldig lite fisk, hovedsakelig ulker og kun fire torskeyngel og én brosme. Konklusjonen er altså at denne typen reketeiner er veldig rekespesifikke og med lite bifangst. Unntaket er andre pandalide reker, f.eks. blomsterreker, med en sammenlignbar fangbarhet. En detaljert tabell med alle registrerte arter fra reketeinene finnes i Vedlegg 2 (Tabell V2.3).
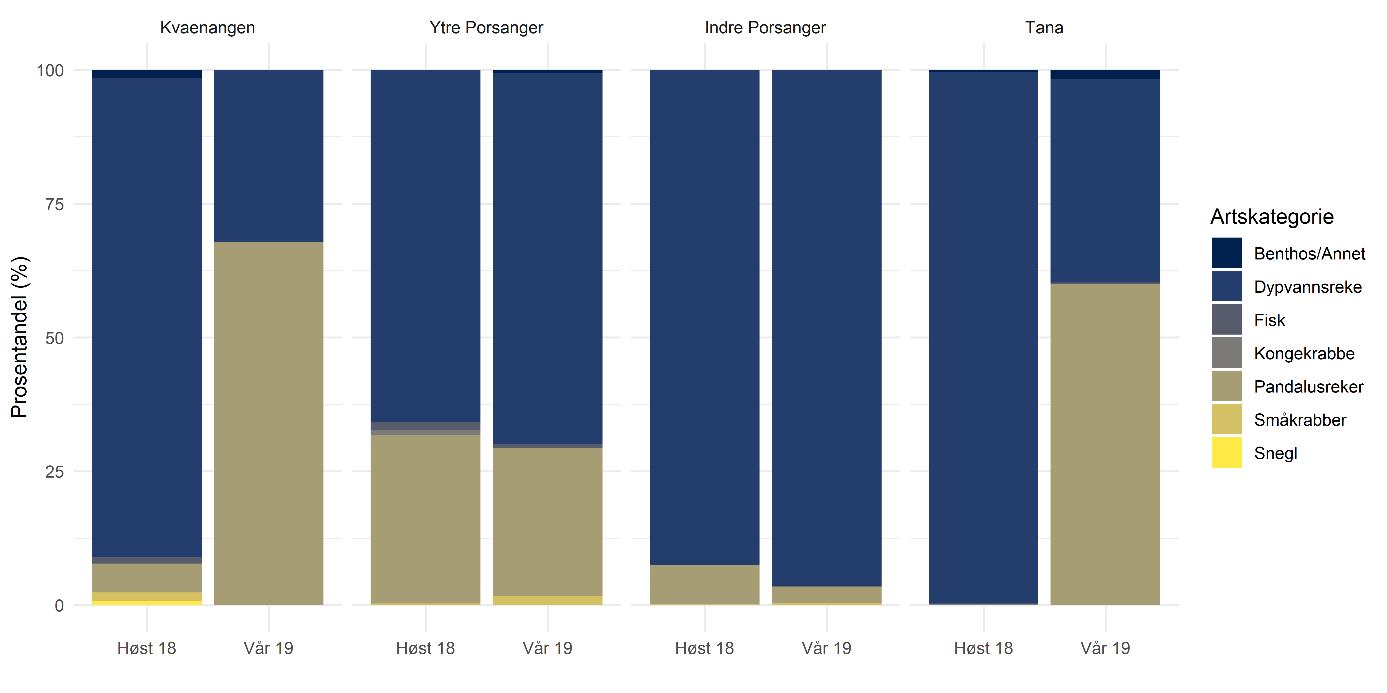
7.5 - Datakvalitet
Teineundersøkelsene ble kjørt parallelt med kartleggingstoktet høsten 2018 og våren 2019, med fokus på å sammenligne fangstrater av reke mellom trål og teiner. Måten dataene ble samlet inn på, gjør det ikke mulig å utforske andre spørsmål systematisk, f.eks. hvordan ståtid av teiner påvirker fangstraten. De samme stedene ble ikke dekket på de to teinetoktene. Dessuten viste det seg at på det første toktet i Porsangerfjorden var teinene for lette til å kunne settes i dype eller strømutsatte områder, noe som påvirket datakvaliteten negativt høsten 2018. Sammenlagt fører alle disse faktorene til at datasettet er relativt lite, ubalansert og med manglende kontrast, og at det derfor ikke kan brukes for å skille mellom forskjellige effekter på fangstrate som f.eks. område, årstid og bunndyp.
I tillegg er teinefiske på reke fortsatt i utvikling, og det er uklart hvordan forskjellige parametre som plassering over bunn eller agn påvirker fangstratene. Det anbefales derfor videre undersøkelser for å analysere relevante spørsmål rundt teinefiske på reke på en mer systematisk måte.
8 - Porsangerfjorden tilbake til livet (Porsangerfjorden 2.0)
Porsanger kommune tok initiativ til prosjektet «Porsangerfjorden tilbake til livet» i 2001. Hensikten var å få kunnskap om årsaker til den vedvarende kraftige reduksjonen av fiskebestandene i fjorden, samt å finne fram til tiltak som kunne bringe bestandene tilbake til historiske nivåer.
I begynnelsen ble oppmerksomheten i stor grad viet den mulige sammenhengen mellom kråkebollenes nedbeiting av tareskogen langs kysten av Nord-Norge, og dårlig overlevelse av 0-gruppe sei og torsk i fjordene. Det var den gang en ny problemstilling. Fiskeri- og kystminister Helga Pedersen uttalte følgende om initiativet: Foruten den betydning Porsangerfjordprosjektet har i seg selv, ser jeg at prosjektet kan ha stor overføringsverdi for andre kommuner og regioner som står ovenfor tilsvarende utfordringer.
Forskningsprosjektet Epigraph (2008-2011) ble igangsatt som en oppfølging til Porsanger-initiativet, og konkluderte bl.a. med at torskebestanden i fjorden var redusert til omtrent en femtedel av nivået på 1980-tallet. Dette anslaget er i god overensstemmelse med en nylig utgitt ekspertrapport, der det anslås at gytebestanden av kysttorsk nord for 62 °N i dag er omtrent en tredjedel av nivået på begynnelsen av 1990-tallet (Aglen mfl. 2020).
Porsanger kommune er fortsatt prosjekteier og pådriver for forvaltningsinitiativet som har en målsetting om å bygge opp igjen fjordbestandene, og få fisk inn i tidligere fiskerike områder som har vært forlatt i tiår. Det faglige grunnlaget er siden oppstarten utvidet med fjordspesifikke rapporter som Porsangerfjorden 2.0 – en mulighetsstudie (Strand 2019), samt felt- og laboratoriestudier utført ved Havforskningsinstituttets stasjon i Finnmark (Pedersen mfl. 2020, Strand mfl. 2020). I samarbeid med fiskere er det også laget en rapport, som knytter publisert teori enda nærmere opp mot faktiske hendelser i fjorden enn det som er tilfelle i Porsangerfjord 2.0-rapporten. (Rapporten er foreløpig ikke publisert, men kan fås tilsendt ved henvendelse til hansks@hi.no.) Kommunens prosjekt har tatt opp i seg navnet etter mulighetsstudien, og kalles i dag Porsangerfjorden 2.0, eller forkortet P20. Et enstemmig kommunestyre har bedt departement og direktorat om å utsette avgjørelsen om en eventuell åpning av Porsangerfjorden for reketråling, til rammen for P20-prosjektet er fastlagt.
For å nå målet om mer torsk og flere større individer er fjorden foreslått delt inn i tre soner, der det kommersielle fisket foregår som før i én av dem (sone 3) og forbys i de to andre (sone 1 og 2) (Figur 8.1). Sonen der det kommersielle fisket foreslås å foregå som før (sone 3) inkluderer området der mesteparten av det kommersielle fisket foregår også i dag, slik at ulempen for fiskerne skal bli så liten som mulig.
Dagens kommersielle fiske foregår i tilknytning til gytefjorden Smørfjorden i perioden februar til og med april, og må derfor antas å i hovedsak beskatte fjordtorsk. I den ene av de to sonene som foreslås vernet mot kommersielt fiske, tar P20-prosjektet mål av seg til å stimulere egg- og yngelproduksjonen gjennom å etablere en gytemerd med oppfôret lokal torsk, samt en tarepark for nylig bunnslått yngel.
Tareparken vil bestå av 10-15 m høye kunstig anlagte taretrær (Figur 8.2), av en type som i dag er under utvikling ved Havforskningsinstituttets stasjon i Holmfjord. Studier i fjorden har vist at bunnlevende ulke er den viktigste predatoren på nylig bunnslått yngel, og at kombinasjonen av ulke som angriper nedenfra og ungtorsk som angriper ovenfra er en spesielt dødelig kombinasjon for yngelen. Ved å løfte store deler av produksjonen utenfor ulkas rekkevidde har vi tro på at yngeloverlevelsen vil øke.
Etter at torsken er ferdiggytt i den vestlige gytefjorden (sone 3), vandrer deler av bestanden over til østsiden (sone 2), hvor de bl.a. beiter på lokale lodde- og rekeforekomster. I P20-prosjektet foreslås denne østlige delen av fjorden som det andre området vernet mot kommersielt fiske, slik at sannsynligheten for flere store flergangsgytere i bestanden øker.

9 - Økosystemvurderinger
9.1 - Magedata
Under de tre tråltoktene med «Katla» (høst 2018, samt vår og høst 2019) ble mageprøver av et representativt utvalg av bunnfisk samlet inn for å kartlegge hvem som spiser hvem i økosystemet i de tre fjordene. Instruksen var å samle inn mager fra inntil 10 fisk per stasjon av hver av følgende arter (samt andre tallrike arter): torsk, hyse, sei, vanlig ulke, brosme, gråsteinbit, flekksteinbit, vanlig uer, kveite og rødspette. Materiale ble samlet inn for artene torsk, hyse og vanlig uer, samt noen rødspette, og et fåtall hvitting, sei, brosme og vanlig ulke. Alt materiale vil bli opparbeidet og publisert, men i denne rapporten fokuseres det på resultatene fra innsamlingen høsten 2018, med vekt på dietten til stor (≥ 35 cm) og liten (< 35 cm) torsk og hyse samt vanlig uer. Det er også disse diettdataene som har blitt brukt i Ecopath-modelleringen (se under). Detaljerte tabeller finnes i Vedlegg 6, og de viktigste byttedyrgruppene vises grafisk nedenfor (Figurer 9.1, 9.2, 9.3).
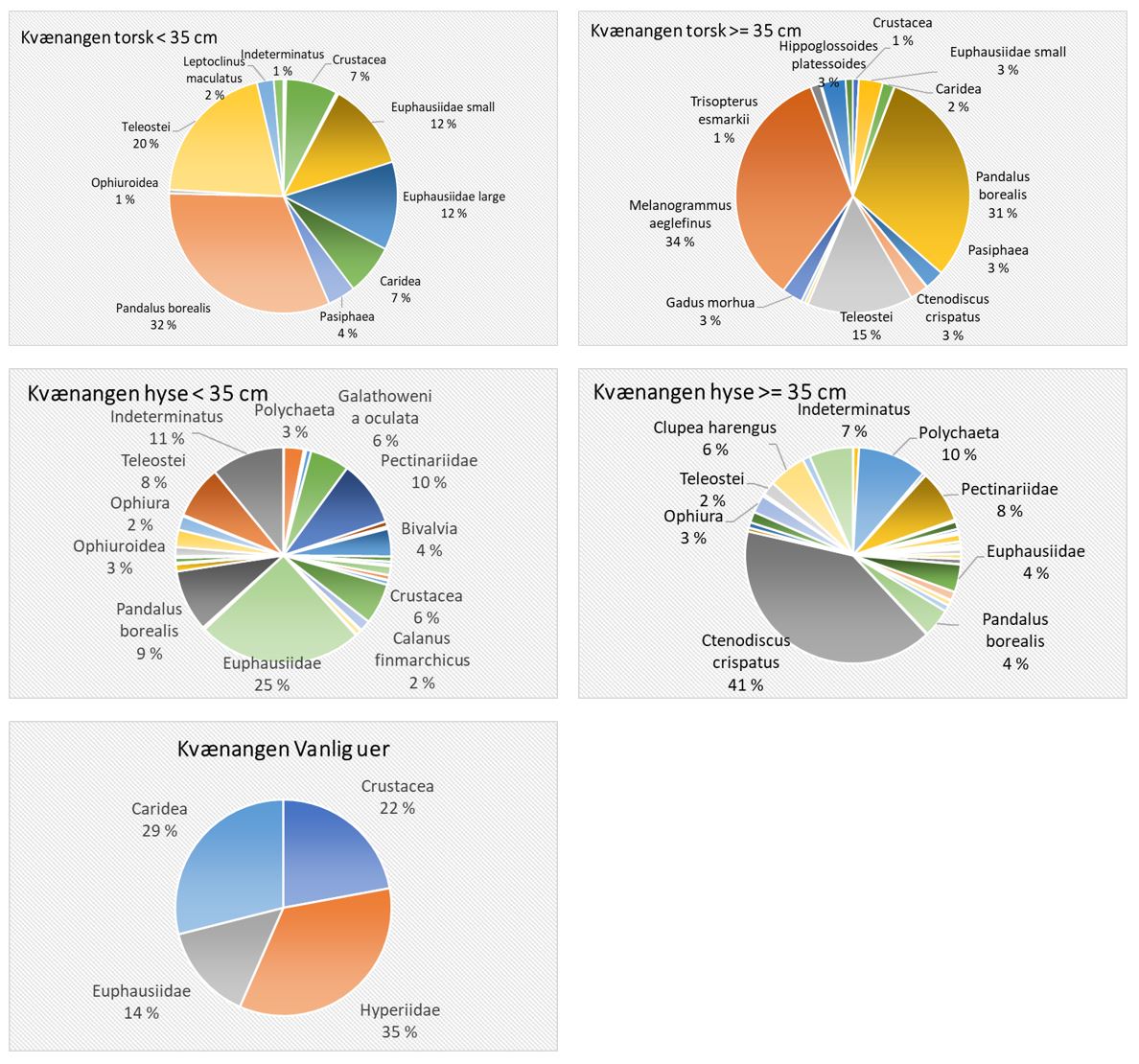
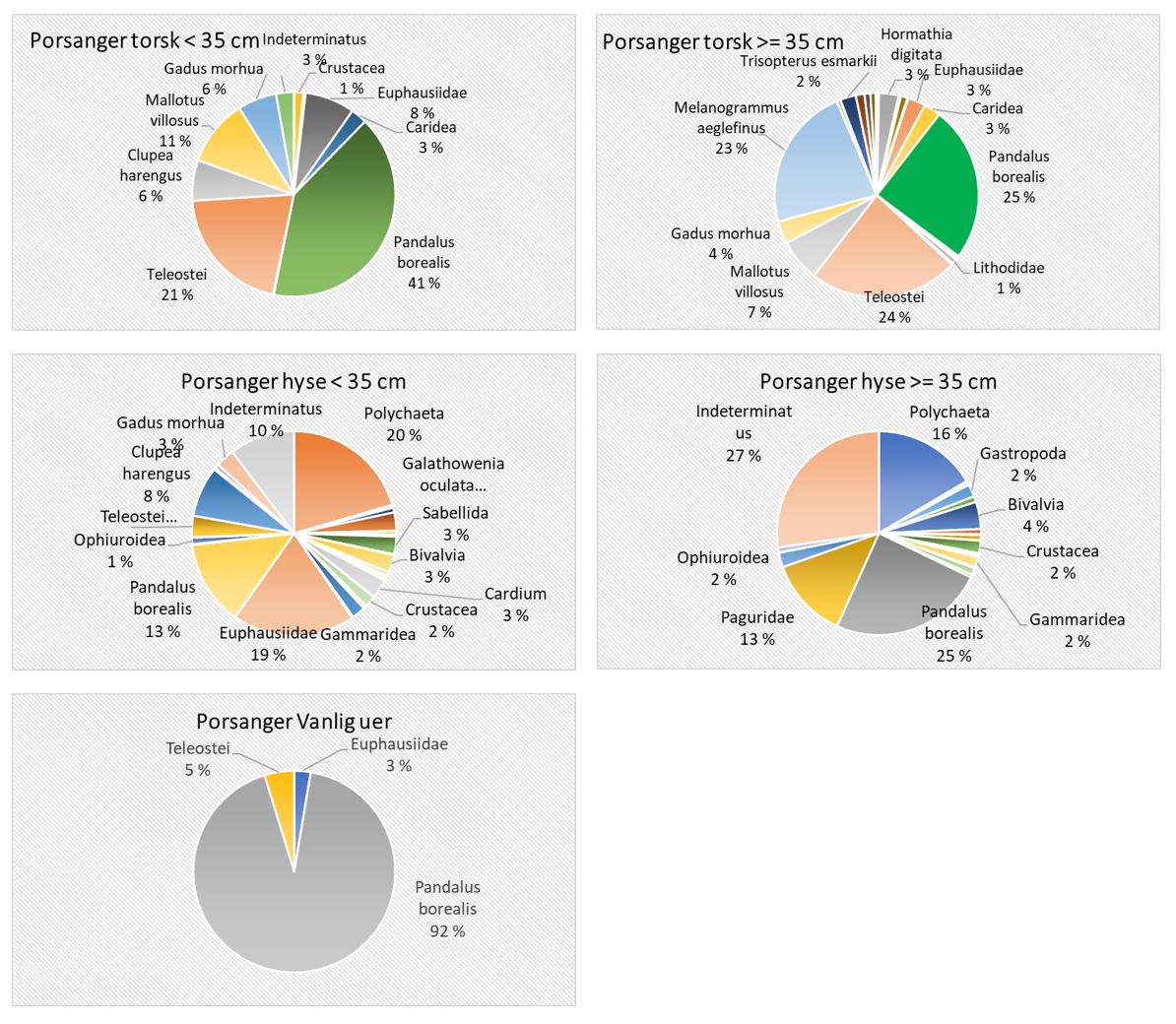
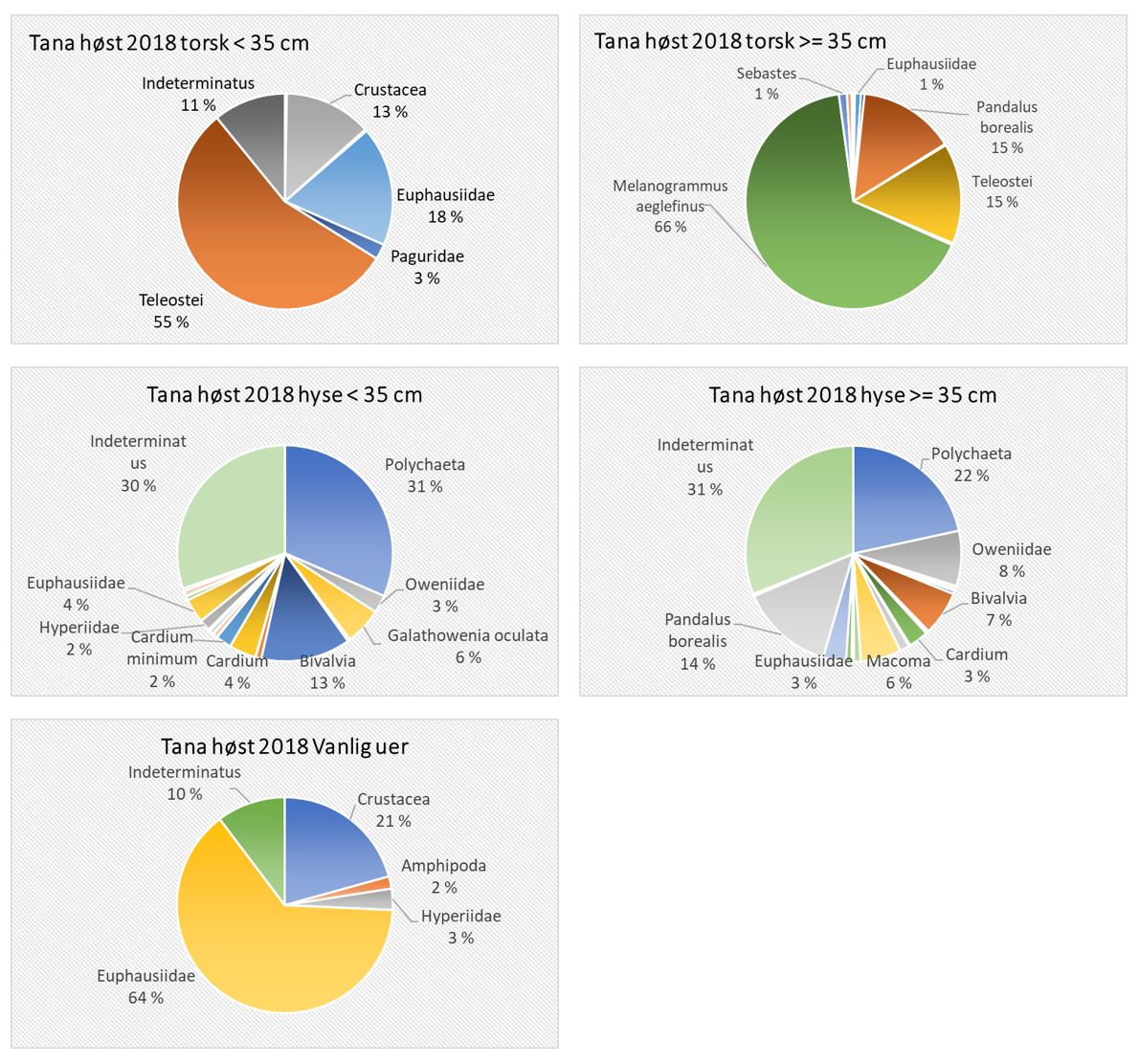
Materialet innsamlet våren 2019 er også opparbeidet, og nedenfor vises torskedietten i Tanafjorden våren 2019 som et eksempel (Figur 9.4). For den mindre torsken består dietten av mindre fisk og flere dyregrupper om våren enn om høsten, bl.a. mer reker om våren, og for den større torsken betyr reker mer om våren enn om høsten, og hyse om høsten er byttet ut med lodde om våren som det viktigste fiskebyttedyret. En endelig økosystemmodell må ta hensyn til slike sesongvariasjoner i dietten.

9.2 - Food web / Ecopath
9.2.1 - Ecopath og Ecosim og Porsangerfjorden
Ecopath, utviklet av Jeff Polovina (Polovina 1984) er sannsynligvis det mest populære numeriske modelleringsverktøyet som brukes i dag for å representere flukser av masse eller energi i marine matnett (Colléter mfl. 2013). Ecopath er basert på to ligninger som styrer hovedprosessene i modellen. Den første ligningen styrer produksjonen:
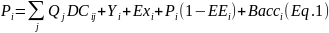
Produksjon måles i gram karbon per m² og år. For en art/bestand i, deler produksjonen P seg i 1) det som spises av predatoren j, 2) fangstene Y i fiskeriene, 3) eksporten ut fra systemet Ex, 4) annen dødelighete P(1-EE) hvor EE, kalt «ecotrophic efficiency», er den delen av produksjonen som går til predasjon, fangster eller eksport, og 5) biomasseakkumulering Bacc som representerer en årlig vekst eller reduksjon i arten/bestanden. Den andre ligningen er forbruk og sier at forbruket C er lik produksjonen P pluss den ikke-assimilerte og respirerte massen: C = P + Resp + ikke-assimilert masse (Eq.2, Figur 9.5).
En modell av Porsangerfjorden ble nylig utviklet av Pedersen mfl. (2018). Den består av fem modeller som representerer fem delområder: tre deler i den ytre fjorden (utenfor Reinøya) og to i den indre fjorden (en østlig og en vestlig del). Denne modellen var sammensatt av 63 grupper som inkluderte sjøpattedyr (kobbe, hval, oter), fugler, pelagisk fisk, bunnfisk, forskjellige bunndyr og primærprodusenter som planteplankton og makroalger (Figur 9.6). Målet med arbeidet var å utforske påvirkningen fra kongekrabbe (Paralithodes camtschaticus) på matnettet i Porsangerfjorden.
I 2019 justerte en masterstudent, Birte Schuppe, modellen til Pedersen mfl. for å studere effekten av en potensiell åpning av et reketrålfiske på biomassen av reker, torsk, hyse og sei i økosystemet i Porsangerfjorden (Schuppe 2019). Schuppe brukte bare tre delområder: et dypt stratum > 170 m, et grunt stratum som strakte seg fra fjordmunningen og halvveis inn i fjorden, og én indre del. Hennes tre modeller ble beregnet basert på modellen til Pedersen mfl. og biomassedata innsamlet høsten 2018 og våren 2019 under kartleggingstoktet med «Katla».
Arbeidet vårt er en videreføring og utvidelse av arbeidet til Pedersen mfl. (2018) og Schuppe (2019) til å også omfatte Tanafjorden og Kvænangen. Målet er å sammenligne økosystemene i de tre fjordene og studere økosystemeffekter av en potensiell åpning av et reketrålfiske. Vår modell inneholder åtte delmodeller: et dypt og et grunt stratum per fjord, samt et østlig og et vestlig stratum i indre Porsangerfjorden (Figur 3.2). Sammenlignet med den opprinnelige modellen fra 2018, fjernet vi aldersklasser for kongekrabbe, og slo sammen Ctenodiscus crispatus (muddersjøstjerne) med den andre detritivore pigghudgruppen. Videre slo vi sammen biomassen av glassreker med Pandalidae reker fordi glassrekene på enkelte trålstasjoner i Kvænangen hadde en like stor biomasse som dypvannsreker.
9.2.2 - Oppbygging av modellene
Data som går inn i modellene
For å løse Ecopath-ligning (Eq. 1) trenger vi for hver art/artsgruppe data på produksjon, forbruk, diett, fangster, eksport, «ecotrophic efficiency» og akkumulering av biomasse. Vanligvis anses biomasseakkumulering som null (dvs. at arten/bestanden er stabil over tid). I vårt tilfelle ignorerer vi også eksporten, som gir oss fem parametere som vi trenger data for.
Produksjon og forbruk er vanskelige variabler å måle, men forholdet, P/B (tilsvarer total dødelighet) og Q/B (summen av forbruket), kan estimeres. Vi brukte Pedersen mfl. (2018) sine verdier for P/B og Q/B, som kom fra eksisterende litteratur om Porsangerfjorden, lignende fjorder, eller fra Norskehavs- og/eller Barentshavsregionene. For dypvannsreker brukte vi dødelighet estimert av Båtevik (2020) for de tre fjordene.
Diettdata har vi tatt fra Pedersen mfl. (2018), med unntak av dietten for stor og liten torsk, hyse og annen bunnfisk (hovedsakelig uer) hvor vi brukte magedata fra kartleggingstoktet høsten 2018.
Ecopath kan bruke parametre for en art til å estimere en ukjent parameter. Ideelt sett lar vi modellen fikse EE («ecotrophic efficiency») ved å gi den biomassedata. For å lage en riktig representasjon av økosystemet, ble bare høstbiomasser brukt i denne omgang. De samsvarte med ferdig opparbeidet magedata for høsten 2018 (diettdata for våren og høsten 2019 vil bli lagt inn senere). Sjøpattedyrobservasjoner ble beskrevet i Tanafjorden av Kjell T. Nilsen (pers. komm.) og brukt til å beregne biomasse (gC/m²) av havert og steinkobbe i den fjorden. For de to andre fjordene og for sjøfugl ble biomassedata hentet fra Pedersen mfl. (2018). Biomassedata var tilgjengelig fra fjordkartleggingen høsten 2018 for de fleste fiskearter i modellen. For bunndyr brukte vi data fra fangster i Campelen-trålen på kystressurstoktet i Kvænangen høsten 2019. Ettersom de fleste bunndyrarter ikke blir fanget ordentlig av en Campelen-trål, valgte vi å beholde Pedersen mfl. sin biomasse for bunndyr for Porsangerfjorden. Tanafjorden fiskes på samme måte som Porsangerfjorden, derfor antok vi at bunndyr ble funnet i lignende tettheter her som i Porsangerfjorden. Ingen kvantitativ informasjon var tilgjengelig for primærprodusenter eller pelagisk fisk i Tanafjorden og Kvænangen, så vi beholdt Pedersen mfl. sine verdier for Porsangerfjorden for disse gruppene.
Balansering av modellene
Når parametrene er bestemt for hver art, verifiserer Ecopath-rutinen om modellen er balansert, dvs. om alle artsbiomassene er høye nok til å støtte predasjon og fiskeridødelighet (med en EE > 1). Dette trinnet er veldig viktig og interessant da det avslører uoverensstemmelser mellom de forskjellige datasettene (vanligvis biomasse vs. dietter). I dette arbeidet stoler vi på biomassestimatene og har derfor prøvd å endre dem så lite som mulig. Hvis biomassedata fra 2018-kartleggingstoktet ikke tillot en EE < 1, prøvde vi data fra den forrige Porsanger-modellen (Schuppe (2019) eller Pedersen mfl. (2018)). Hvis dette alternativet heller ikke balanserte modellen, kunne vi enten 1) fikse EE (0,9 for de fleste av artene, 0,5 for primærprodusenter som tang som for det meste ender i detritus, og 0 for ufiskede topppredatorer) ved å la modellen estimere biomassen, eller 2) endre dietten til predatorene. Når det gjelder bunndyr, pelagisk fisk og primærprodusenter, brukte vi stort sett det første alternativet pga. manglende kvalitetssikrede data for biomassen deres. For bunnfisk og reker modifiserte vi heller diettene (basert på mageinnhold), da de bare viser et øyeblikksbilde av en predators diett og kan påvirkes av fangstforholdene (f.eks. beiting inne i trålen). Samlet sett var megabentiske artsgrupper, små gadoider (spesielt sei) og bunnfisk ofte ubalanserte, og fremhevet uoverensstemmelsen mellom den estimerte biomassen av disse gruppene, predatorbiomassen, og den estimerte betydningen av disse byttedyrene i dietten til predatorene. Videre studier bør fokusere på å forbedre mengdeestimatene av disse gruppene.
Vi ønsker å understreke at ingen usikkerhetsvurdering ble gjort innen rapporten ble skrevet. Fremtidig utvikling av modellene vil inkludere dette.
9.2.3 - Resultater og diskusjon
Lignende biomassetettheter (gC/m²) ble funnet i de tre fjordene, og også i grunne og dype områder, til tross for forskjellige tilførsels- og modellestimerte biomasser. På den annen side er netto primærproduksjon større i grunne områder i Kvænangen sammenlignet med de to andre fjordene (Figur 9.7). Store biomasser av toppredatorer, torsk, hyse og særlig sei (Figur 9.8), førte til høy dødelighet av sild i Kvænangen. For å balansere dødeligheten med produksjonen, lot vi modellen estimere hele næringskjeden fra planteplankton til sildebiomassen, som nesten doblet dem, derav den høyere primærproduksjonen. Fremtidig prøvetaking i Kvænangen bør undersøke nøyere den pelagiske delen og primærproduksjonen for å bekrefte dette.
Som nevnt i Pedersen mfl. (2018), er den østre, indre delen av Porsangerfjorden forskjellig fra resten av denne fjorden og også fra de andre fjordene ved sine arktiske egenskaper: lav bunntemperatur, isdekke om vinteren, høyt oksygeninnhold og lavere saltholdighet (Vedlegg 1). De to indre, østlige bassengene inneholder et økosystem med store tettheter av små bløtdyr, planteetende kråkeboller, store muslinger, sjøstjerner og andre konsumenter som sild og ulker. De fleste av disse biomassene ble estimert av modellen, og bekreftes av bomtrålundersøkelsene i området (Figur 9.8). Som observert i fangstene fra kartleggingstoktet, inneholder dette området også store tettheter av dypvannsreker. I modellene har den ytre, dype delen av Porsangerfjorden enda høyere tettheter av reker, men dette skyldes en oppskalering av biomassen med en faktor på 10 og en reduksjon av rovdyrdietten under balanseringsprosessen.
Det er vanskelig å peke på ytterligere forskjeller mellom de resterende lagene i de tre fjordene. Tanafjorden ser ut til å inneholde store mengder av crangonide reker, men de ble estimert av modellen og kan skyldes den høye biomassen av stor torsk og hyse som ble fanget der (selv om de ikke er veldig forskjellige fra fangstene i Kvænangen) og den store andelen av crangonide reker i dietten til hyse (15 % av mageinnholdet).
Sammen med biomassedata for predatorene, viser analyser av mageinnhold at reker har veldig forskjellige hovedpredatorer i de forskjellige områdene. I Kvænangen og ytre Porsangerfjorden spises reker av både små og stor torsk, uer og liten hyse. I Tanafjorden er stor torsk den viktigste predatoren, mens reker i indre Porsangerfjorden hovedsakelig beites på av ulker og små torsk (Figur 9.9). Tre hovedpredasjonssystemer trer altså frem: Kvænangen og ytre Porsangerfjorden, indre Porsangerfjorden, og Tanafjorden.
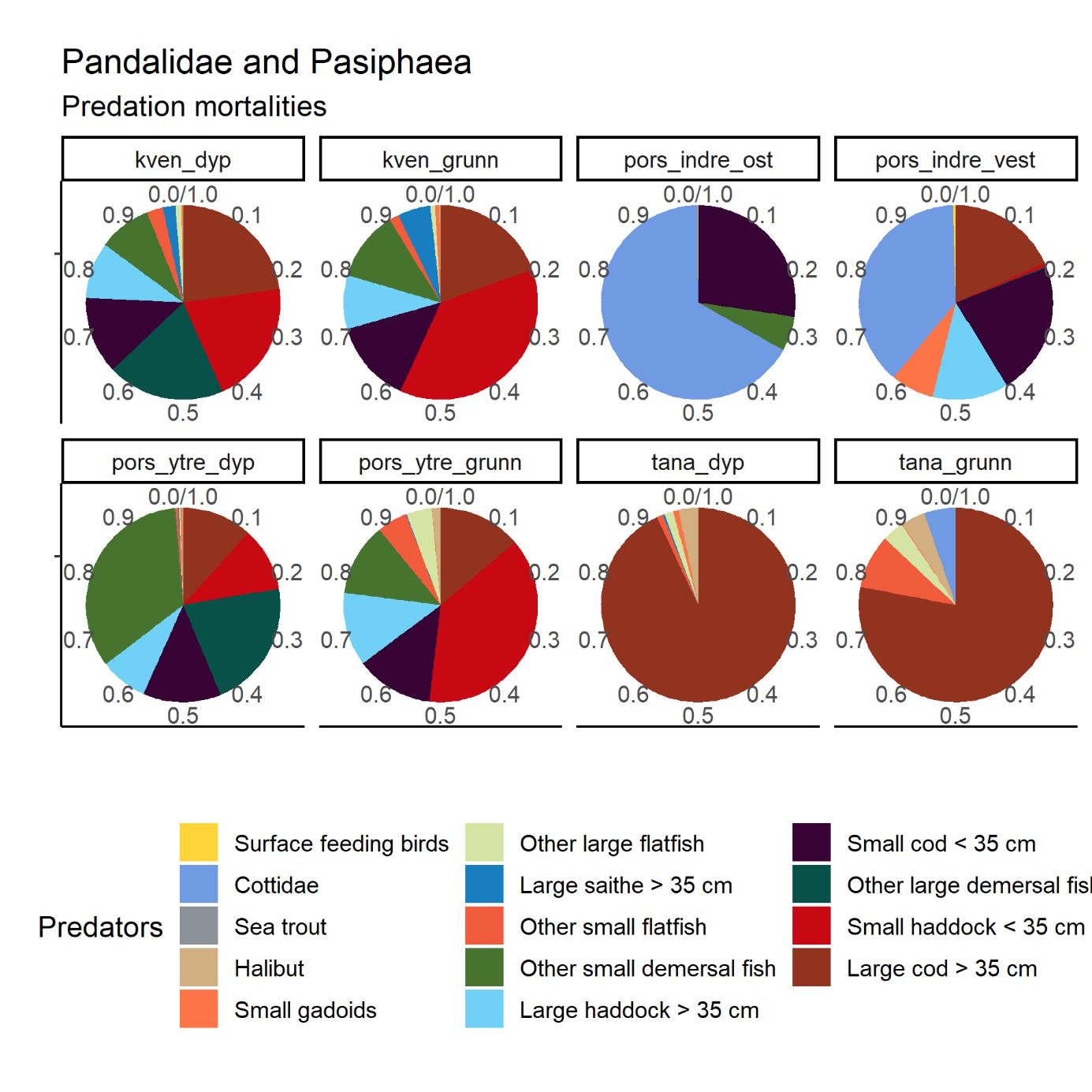
Det var ikke mulig å simulere påvirkningen av et rekefiske for denne rapporten. Schuppe (2019) utviklet modellen for den ytre dype delen av Porsangerfjorden til en Ecosim prediksjonsmodell. Hun brukte en basis fiskeinnsats på reker tilsvarende den vesle beskatningen som tråltoktene med «Katla» påførte rekeressursen, dvs. F ~ 0,0002 (total dødelighet Z ~ 1,2). Fiskeinnsatsen ble så økt for å utforske 1) maksimal fiskedødelighet hvor det ikke observeres noen endringer i nivået på rekebiomassen, og 2) økning av fiskeinnsatsen til F ~ 1. For en fiskeinnsats opp til 1000 ganger høyere enn basis (F ~ 0,2, Z ~ 1,4) ble det ikke observert noen endringer i rekebiomassen. Men med en fiskeinnsats 5500 ganger høyere enn basis (F ~ 1,0, Z ~ 2,0) kollapset rekebiomassen til 0 i løpet av tolv år.
Selv om de nye dataene fra kartleggingstoktet gjør det mulig å karakterisere fiskesamfunn og dietter i de tre fjordene, trenger Ecopath-modeller mer presise data på bunndyr, sjøpattedyr, fugler, pelagisk fisk, plankton og primærprodusenter for å gi en pålitelig beskrivelse av disse økosystemene.
10 - Konklusjoner og råd på rekebeskatning i Porsanger- og Tanafjorden
Havforskningsinstituttets råd i forbindelse med en eventuell åpning av Tana- og Porsangerfjorden for reketrålfiske (bunntråling) presenteres i dette kapitlet.
Vitenskapelige problemstillinger og verneverdier
Et utrålt område er unikt og har høy verdi, både fra et verne- og fra et vitenskapelig perspektiv. Klimaframskrivinger peker på økt viktighet av nordlige kyst- og havområder – for nærings- og gytevandringer, gyte- og oppvekstområder for kommersielt viktige fiskebestander. Intakt økosystemintegritet og minimal negativ påvirkning på rekruttering i tilstøtende kystområder vil være gunstig i et scenario i tråd med framskrivingene.
Sett fra et vitenskapelig perspektiv vil utrålte/bunntrålfrie områder være svært verdifulle. Det gjør at vi kan sammenligne trålte og utrålte områder og dermed øke vår forståelse av hvordan bunntråling påvirker fiskesamfunn og økosystem.
- Dersom fjordene vurderes åpnet for bunntråling anbefaler Havforskningsinstituttet at man kun åpner Porsangerfjorden utenfor terskelen («Ryggen» litt nord for Lille-Tamsøya), og at Tanafjorden forblir stengt for bunntråling.
Porsangerfjorden tilbake til livet (P2.0)
P2.0-prosjektet, som eies av Porsanger kommune, har som mål å bygge opp igjen bunnfiskbestandene i Porsangerfjorden. Hvorvidt det er mulig å kombinere bunntråling og gjenoppbygging av bestander av langlivete bunnfiskarter – i samme økosystem – er usikkert. Derimot vet vi fra Øresund at et område med langvarig bunntrålforbud var i stand til å bevare bred alders- og størrelsessammensetning hos torsk, i samme tidsrom som torskebestander kollapset i tilgrensende områder (Lindegren mfl. 2010, Sundelöf mfl. 2013). P2.0 ønsker at den indre delen (innenfor Reinøya) og den østlige delen av midtre Porsangerfjorden forblir stengt. De trålbare delene av midtre Porsangerfjorden ligger i den østlige delen.
- Havforskningsinstituttet tilrår at Porsangerfjorden innenfor terskelen («Ryggen») forblir stengt for bunntråling.
Sårbare habitater og økosystem (koraller og svamp)
Norge har internasjonale forpliktelser til å ta vare på sårbare habitater og økosystem (som koraller og svamp), f.eks. gjennom Konvensjonen for biologisk mangfold. Vi har ganske god kunnskap om utbredelsen av disse samfunnene i Porsangerfjorden etter mange år med bomtrålundersøkelser, samt data fra Havforskningsinstituttets kysttokt og noen Mareano-transekter. I Tanafjorden derimot, har det ikke vært gjennomført hverken bomtrålundersøkelser eller bunnkartlegging. I den ytre delen av Porsangerfjorden utenfor terskelen («Ryggen») er det påvist koraller langs de østlige skråningene, samt bunnsamfunn med høy biodiversitet og biomasse rett nord for Lille-Tamsøya, men størrelsen av de lokale sårbare habitatene er ikke kjent.
- Det er Havforskningsinstituttets klare råd at områder med sårbare arter og bunnsamfunn ikke bør åpnes for tråling.
- Havforskningsinstituttet anbefaler at det i forkant av en eventuell åpning for reketråling i et område gjennomføres en grundig habitat- og bunnkartlegging av både bunndyrssamfunn og sedimenter.
- Havforskningsinstituttet understreker at dette er spesielt viktig dersom det bestemmes at Tanafjorden skal åpnes. Om ytre Porsangerfjorden åpnes, må forekomster av sårbare habitater registreres også her.
Manglende tidsserier for reke
Vi har per i dag ikke dokumentert kunnskap som viser hvordan en ufisket rekebestand reagerer på fiske i finnmarksfjordene. Vi kjenner ennå ikke rekrutteringen til disse rekebestandene, dvs. frekvensen av gode årsklasser, og det er også viktig at reke som mat for bunnfisk sikres. Havforskningsinstituttet anbefaler derfor et svært forsiktig rekeuttak ved oppstart av et eventuelt reketrålfiske. Tabell 10.1 gir en oversikt over mulige fangster for et trålfiske basert på estimerte biomasser og forskjellige F-verdier (fiskedødelighet), presentert for Tanafjorden og ytre Porsangerfjorden separat.
- Havforskningsinstituttets råd er at dersom det åpnes for reketrålfiske, utformes dette som et prøvefiske med en totalkvote der aktørene plikter å rapportere posisjon, fangst, innsats og redskapstype fra alle trålhal, og at de også kan pålegges å foreta hal i på forhånd utplukkede områder.
Kongekrabbefiske
Porsangerfjorden er p.t. det viktigste området for kongekrabbefisket i Finnmark. Vi vet at det tas lite krabbe i en kommersiell reketrål. Vi regner med at trålen vil treffe mange krabber, men vi vet ikke om eller hvor skadelige slike sammenstøt er.
- Havforskningsinstituttet tilrår at man i forbindelse med eventuell åpning av et reketrålfiske iverksetter undersøkelser for å kvantifisere eventuelle skader på kongekrabbe.
- Havforskningsinstituttet anbefaler at det settes av områder til hhv. trål og teiner for slik å unngå redskapskonflikter.
Økosystemeffekter og bifangstbestemmelser
Et rekefiske med trål må ikke ødelegge oppbyggingen av lokale bestander som vanlig uer og kysttorsk, eller den stedegne loddebestanden i Porsangerfjorden. Havforskningsinstituttet anbefaler derfor at bifangstbestemmelsene bør innskjerpes. Resultatene fra kartleggingstoktene har vist at de fleste større torsk i trålen var kysttorsk (ikke skrei), men vi har ikke resultater for torskeyngel. Tidligere ble det fisket stor hyse innerst i Porsangerfjorden, men den har nå forsvunnet. Dette taler for at også hyse trenger et vern.
- Dersom det åpnes for reketrålfiske tilrår Havforskningsinstituttet at følgende bifangstbestemmelser innføres i området som åpnes (antall yngel/individer per 10 kg reke i fangsten):
Torsk: 3 yngel
Vanlig uer: 3 yngel
Hyse: 8 yngel
Lodde: 50 individer
- Havforskningsinstituttet anbefaler at det i et tidlig stadium av et evt. gjenåpnet fiskeri blir tatt DNA-prøver av torskeyngelen (0+), og at grensen kan økes til 8 yngel per 10 kg trålte reker, om det viser seg at det er snakk om skrei.
Anbefalinger om høsting av reke med teiner
Rekepopulasjonen i de indre østlige bassengene av Porsangerfjorden er unik i norsk sammenheng. På grunn av fravær av fiske i fire tiår og få predatorer er den høye biomassen sannsynligvis akkumulert over mange år. Det kalde bunnvannet og en langsom vekst og reproduksjon tilsier en svært lav produksjon, og taler for at ved en eventuell desimering vil det ta svært lang tid for bestanden å bygge seg opp igjen. Pga. lite kunnskap om denne bestanden og en fortsatt utvikling av teinefisket er det vanskelig å komme med eksakte råd om uttak. Imidlertid tilsier den lange oppbyggingstiden for rekebestanden i disse fjordbassengene at det bør velges et forsiktig uttak av reker.
- Havforskningsinstituttet anbefaler at det settes et tak på de årlige fangstene fra indre Porsangerfjorden på 20 tonn, og et tak på 5 tonn i et eventuelt tilsvarende teinefiske i Tanafjorden.
- Havforskningsinstituttet anbefaler en grense på 150 teiner per båt for både Porsanger- og Tanafjorden.
- Havforskningsinstituttet anbefaler at teinefisket og bestandsutviklingen overvåkes gjennom et prøvefiske med krav om registrering av fangster, innsats og posisjon.
- Havforskningsinstituttet anbefaler at maskevidden i teinene økes og at det innføres påbud om fluktåpning med råtnetråd.
| Ytre Porsangerfjorden | Tanafjorden | |||||||||
| Gjennomsnitt H2018, V2019, H2019 | Variasjons-bredde maks-min | Gjennomsnitt H2018, V2019, H2019 | Variasjons-bredde maks-min | |||||||
| Biomasse (tonn) | N*w= | 162 | 182 | 143 | 170 | 264 | 76 | |||
| Beskatningsgrad | F= | 0,3 | 0,3 | 0,3 | 0,3 | 0,3 | 0,3 | |||
| Naturlig dødelighet | M= | 0,8 | 0,8 | 0,8 | 0,8 | 0,8 | 0,8 | |||
| Total dødelighet | Z= | 1,1 | 1,1 | 1,1 | 1,1 | 1,1 | 1,1 | |||
| Fangst (tonn) | C= | 29 | 33 | 26 | 31 | 48 | 14 | |||
| Konstant Z=1,1 | F= | 0,1 | 0,1 | 0,1 | 0,1 | 0,1 | 0,1 | |||
| M= | 1 | 1 | 1 | 1 | 1 | 1 | ||||
| C= | 10 | 11 | 9 | 10 | 16 | 5 | ||||
| F= | 0,2 | 0,2 | 0,2 | 0,2 | 0,2 | 0,2 | ||||
| M= | 0,9 | 0,9 | 0,9 | 0,9 | 0,9 | 0,9 | ||||
| C= | 20 | 22 | 17 | 21 | 32 | 9 | ||||
| F= | 0,3 | 0,3 | 0,3 | 0,3 | 0,3 | 0,3 | ||||
| M= | 0,8 | 0,8 | 0,8 | 0,8 | 0,8 | 0,8 | ||||
| C= | 29 | 33 | 26 | 31 | 48 | 14 | ||||
| F= | 0,4 | 0,4 | 0,4 | 0,4 | 0,4 | 0,4 | ||||
| M= | 0,7 | 0,7 | 0,7 | 0,7 | 0,7 | 0,7 | ||||
| C= | 39 | 44 | 35 | 41 | 64 | 18 | ||||
| F= | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 | ||||
| M= | 0,6 | 0,6 | 0,6 | 0,6 | 0,6 | 0,6 | ||||
| C= | 49 | 55 | 43 | 52 | 80 | 23 | ||||
| F= | 0,6 | 0,6 | 0,6 | 0,6 | 0,6 | 0,6 | ||||
| M= | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 | ||||
| C= | 59 | 66 | 52 | 62 | 96 | 28 | ||||
Bakgrunnsinformasjon til Tabell 10.1 (Havforskningsinstituttets kvoteråd for beskatning av rekeressursen i Porsanger- og Tanafjorden):
- I samsvar med Porsanger kommune sitt forvaltningsinitiativ P2.0 som har en målsetting om å bygge opp igjen fjordbestandene av bunnfisk, så legges det til grunn at et evt. reketrålfiske i Porsangerfjorden avgrenses til området (≥ 170 m) utenfor terskelen nord for Lille-Tamsøya, og våre beskatningsalternativer for Porsangerfjorden er derfor bare gitt for dette området (i tillegg kommer fiske med reketeiner i den indre delen av fjorden). Vi har lagt til grunn hele den estimerte rekebiomasse i midtre og ytre Porsangerfjorden (dvs. utenfor Reinøya) da vi kan anta at rekebestanden henger sammen over terskelen (og dybdegrensen). For Tanafjorden har vi lagt til grunn rekebiomassen i hele fjorden.
- Biomasse av reke (gjennomsnitt, maksimum og minimum) er beregnet over alle tre toktperioder av kartleggingstoktet (høst 2018, vår 2019 og høst 2019) for trålbar bunn dypere enn 170 m. Biomasseestimatene fra kartleggingstoktet er sannsynligvis underestimater fordi 1) areal som reketetthet ganges opp med, bare inkluderer bunnarealet som vi ut ifra bunntopografi og bunnsedimentkart har definert som trålbar bunn, 2) fangstevnen til trålen og 3) det faktum at det p.t. fiskes 150-200 tonn reker i Kvænangen fra en estimert toktbiomasse på ca. 200 tonn.
- Basert på estimert naturlig dødelighet i Porsanger- og Tanafjorden og foreløpige økosystembetraktninger ved hjelp av Ecopath/Ecosim modellering antar vi at en bærekraftig total dødelighet Z (naturlig dødelighet (M) pluss fiskedødelighet (F)) ligger på omtrent Z = 1,1. Naturlig dødelighet vil trolig bli redusert når vi starter et fiske (og dermed kunne man ha fisket mer innenfor samme totale dødelighet), men i oppstarten vil vi legge mest vekt på opsjonen med naturlig dødelighet lik den vi har estimert uten fiske (M=0,8). Data fra Kvænangen antyder at naturlig dødelighet reduseres proporsjonalt med økende fiskedødelighet til en viss grense. Vi har her benyttet denne fremgangsmåten for å beregne mulige rekefangster, men det er veldig usikkert hvor mye og hvor raskt naturlig dødelighet kan endre seg.
11 - Takk
Tusen takk til skipperne Dag Olav Mollan og Espen Ottem, og mannskap på «Katla» og «Henriette» for den store innsatsen de alle la ned både før, under og etter toktene i Troms og Finnmark. Tusen takk til Fiskeridirektoratets sjøtjeneste ved Rolf Harald Jensen og Tor Johansen, for at de vederlagsfritt stilte «Rind» til disposisjon for kartleggingstoktet høsten 2018. Mannskapet gjorde en fantastisk jobb med teineregistreringene, og toktpersonell fra Havforskningsinstituttet fikk bo om bord. Jo Inge Hesjevik leide velvillig ut reketeiner til toktene. Takk til Sigrid Elvenes på Norges geologiske undersøkelse (NGU) for hjelp med kartløsningen deres. Torstein Pedersen ved Norges arktiske universitet (UiT) har kommet med mange gode råd i forbindelse med Ecopath-modelleringen. Edvin Fuglebakk hjalp oss med å plukke ut tilfeldige trål- og teinestasjoner i forkant av toktgjennomføringen. Takk til Knut Yngve Børsheim for at vi fikk bruke upubliserte oseanografiske data fra Porsangerfjorden. Ida Vee og Ragni Olssøn opparbeidet data på bunndyr fra kystressurstoktet i 2019, og Gjertrud Jensen og Genoveva Gonzalez-Mirelis hjalp oss med Mareano-dataene. Bjarte Bogstad hjalp oss med sammenstillingen av magedataene fra tråltoktene. Hege Øverbø Hansen, Siri Aaserud Olsen, Tone Vollen, Marita Larsen, Birte Schuppe, Jostein Saltskår, Felicia Keulder-Stenevik, Josefina Johansson, Anne Kari Sveistrup, Reidar Johannesen og Sindre Nygård Larsen deltok på ett eller flere av toktene og gjorde sitt til at gjennomføringen gikk knirkefritt. Tone Vollen og Maria Jenssen hjalp til med logistikken rundt toktene i Tromsø. Forskningsgruppene Dyphavsarter og bruskfisk og Fiskeridynamikk lånte velvillig ut lengdemålingsbrett og vekter til toktene. Maria Jenssen, Marita Larsen og Hanna Ellerine Helle Danielsen har hjulpet med formatteringen av rapporten. Fiskeridirektoratet og Fjord i Nord (FiN) finansierte deler av toktgjennomføringen, og NFR prosjektnr. 299554 CoastRisk har finansiert deler av prosjektet.
12 - Referanser
Adelman I.R., Smith L.L. 2011. Effect of Oxygen on Growth and Food Conversion Efficiency of Northern Pike, The Progressive Fish-Culturist. Taylor & Francis Group, 32(2): 93–96. doi: 10.1577/1548-8640(1970)32[93:EOOOGA]2.0.CO;2.
Anon. 2012. Blandingsfiske som reguleringsmodell – skisse til forprosjekt. Havforskningsinstituttet. 10 pp.
Aglen A., Nedreaas K., Knutsen J.A., Huse G. 2020. Kysttorsk nord for 62-grader nord — Vurdering av status og forslag til forvaltningstiltak og ny gjenoppbyggingsplan. Fisken og havet, https://www.hi.no/hi/nettrapporter/fisken-og-havet-2020-2.
Asplin L., Albretsen J., Johnsen I.A., Sandvik A.D. 2020. The hydrodynamic foundation for salmon lice dispersion modeling along the Norwegian coast. Ocean Dynamics. doi: 10.1007/s10236-020-01378-0 (https://link.springer.com/article/10.1007/s10236-020-01378-0)
Bergström B. 1992. Growth, growth modelling and age determination of Pandalus borealis. Marine ecology progress series, 83:167–183.
Bergström B.I. 2000. The Biology of Pandalus. Advances in Marine Biology, 38: 55¬–244.
Båtevik T. 2020. Estimating natural mortality rates for northern shrimp (Pandalus borealis) from a wide range of its latitudinal distribution in the Northeast Atlantic. Masteroppgave. Universitetet i Bergen. Det matematisk-naturvitenskapelige fakultet. 79 pp.
Christensen P. 2014. Norges fiskeri- og kysthistorie : B. 4 : Havet, fisken og oljen : 1970-2014. Bergen: Fagbokforlaget.
Clark, W.G. 1999. Effects of an erroneous natural mortality rate on a simple age-structured stock assessment. Canadian Journal of Fisheries and Aquatic Sciences. National Research Council of Canada, 56(10): 1721–1731. doi: 10.1139/f99-085.
Colléter M., Valls A., Guitton J., Lyne M., Sánchez F.A., Christensen V., Gascuel D., Pauly D. 2013. EcoBase/: A repository solution to gather and communicate information from EwE models. University of British Columbia. 60 pp. https://hal-agrocampus-ouest.archives-ouvertes.fr/hal-01103442
Dounas C.G., Davies I.M., Hayes P.J., Arvanitidis C.D., Koulouri P.T. 2005. The effect of different types of otter trawl ground rope on benthic nutrient releases and sediment biogeochemistry. American Fisheries Society, 41: 539–544. Available at: https://www.researchgate.net/publication/216187171 (Accessed: 12 June 2020).
Dounas C., Davies I., Triantafyllou G., Koulouri P., Petihakis G., Arvanitidis C., Sourlatzis G., Eleftheriou, A. 2007. Large-scale impacts of bottom trawling on shelf primary productivity, Continental Shelf Research. Pergamon, 27(17), pp. 2198–2210. doi: 10.1016/j.csr.2007.05.006.
Green J. 2009. Growth, size and reproduction in daphnia (Crustacea: Cladocera), Proceedings of the Zoological Society of London, 126(2), pp. 173–204. doi: 10.1111/j.1096-3642.1956.tb00432.x.
Hansen A. 2020. Genetic population structure of the northern shrimp (Pandalus borealis) along the Norwegian coast. Masteroppgave. Norges arktiske universitet. Fakultet for biovitenskap, fiskeri og økonomi. 60 pp.
Hansson M., Valentinsson D., Ulmestrand M., Lindahl A. 1997. Räktrålningens effekter i Gullmarsfjorden. Lysekil, Sweden: Fiskeriverket, Havsfiskelaboratoriet, Institute of Marine Research.
Hjort J., Ruud J. 1938. Rekefisket som naturhistorie og samfundssak. Report on Norwegian Fishery and Marine Investigations. Bergen, Norway: Directorate of Fisheries, 4(4): 1–158.
ICES 2018. Report of the Arctic Fisheries Working Group (AFWG), 18–24 April 2018, Ispra, Italy. ICES CM 2018/ACOM, 06, 857 pp.
Jorde P.E., Søvik G., Westgaard J.I., Albretsen J., André C., Hvingel C., Johansen T. 2015. Genetically distinct populations of northern shrimp, Pandalus borealis, in the North Atlantic: adaption to different temperatures as an isolation factor. Molecular Ecology, 24: 1742-1757.
Knutsen H., Jorde P.E., Gonzalez E.B., Eigaard O.R., Pereyra R.T., Sannæs H., Dahl M., André C., Søvik G. 2015. Does population genetic structure support present management regulations of the northern shrimp (Pandalus borealis) in Skagerrak and the North Sea? ICES Journal of Marine Science 72(3): 863-871.
Larsen T.H. 2010. Population dynamics of cod, Gadus morhua, in Porsangerfjord, Northern Norway-Analysis of back-calculated length at age from otoliths in 1992 and 2009. Universitetet i Tromsø.
Lindegren, M., Diekmann, R., Möllmann, C. 2010. Regime shifts, resilience and recovery of a cod stock. Marine Ecology Progress Series, 402: 239-253. doi: 10.3354/meps08454
Myksvoll M.S., Sandvik A.D., Skarðhamar J., Sundby S. 2012. Importance of high resolution wind forcing on eddy activity and particle dispersion in a Norwegian fjord. Estuarine, Coastal and Shelf Science, 113: 293-304.
Myksvoll M.S., Sandvik A.D., Asplin L., Sundby S. 2013. Effects of river regulations on fjord dynamics and retention of coastal cod eggs. ICES Journal of Marine Science, 71/4): 943-956.
Nilssen E.M., Hopkins C. 1991. Population parameters and life histories of the deep-water prawn Pandalus borealis from different regions. International Council for the Exploration of the Sea. Tromsø, Norway, pp. 1–27.
Pauly D., David N. 1981. ELEFAN I, a BASIC program for the objective extraction datal of growth parameters from length-frequency data. Berichte der Deutschen Wissenschaftlichen Kommission für Meeresforschung, 28(32): 205–211.
Pauly D., Sparre P. 1991. A note on the development of a new software package, the FAO-ICLARM Stock Assessment Tools (FiSAT). Fishbyte, Newsletter of the Network of Tropical Fisheries Scientists, 9(1): 47–49.
Pedersen O.P., Aschan M., Rasmussen T., Tande K.S., Slagstad, D. 2003. Larval dispersal and mother populations of Pandalus borealis investigated by a Lagrangian particle-tracking model. Fisheries Research, 65: 173–190.
Pedersen T, Fuhrmann M.M., Lindstrøm U., Nilssen E.M., Ivarjord T., Ramasco V., Jørgensen L.L., Sundet J.H., Sivertsen K., Källgren E. 2018. Effects of the invasive red king crab on food web structure and ecosystem properties in an Atlantic fjord. Marine Ecology Progress Series, 596: 13–31.
Pedersen T., Strand H.K., Christie H., Moy F. 2020. Predation mortality from ambush and cruising predators on newly-settled 0-group gadoids. Journal of Experimental Marine Biology and Ecology, 529: 151396.
Polovina J.J. 1984. Model of a coral reef ecosystem. Coral Reefs, 3(1): 1–11. https://doi.org/10.1007/BF00306135
Pritchard J.K., Stephens M., Donelly P. 2000. Inference of population structure using multilocus genotype data. Genetics, 155(2): 945–959.
Rasmussen B. 1953. On the Geographical Variation in Growth and Sexual Development of the Deep Sea Prawn (Pandalus borealis Kr.). Reports on Norwegian Fishery and Marine Investigations. Directorate of Fisheries, 5(3): 1–160.
Schuppe, B 2019. How does shrimp fishery affect the ecosystem in a previous closed fjord for bottom trawling? Master thesis at the Department of Geography of RWTH Aachen University, Germany, 21 November 2019. 141 pp.
Seidman E.R., Lawrence A.L. 2009. Growth, feed, digestibility, and proximate body composition of juveline Penaeus vannamei and Penaeus monodon grown at a different dissolved oxygen level. Journal of the World Mariculture Society. John Wiley & Sons, Ltd, 16(1–4): 333–346. doi: 10.1111/j.1749-7345.1985.tb00214.x.
Shumway S.E., Perkins H.C., Schick D.F., Stickney A.P. 1985. Synopsis of Biological Data on the Pink Shrimp, Pandalus borealis Krøyer, 1838. 144 pp.
Simpfendorfer C.A., Bonfil R., Latour R.J. 2005. Mortality estimation. Management techniques for elasmobranch fisheries. Rome: Food and Agriculture organization of the united nations, pp. 127–142.
Sparre P., Venema S.C. 1998. Introduction to tropical fish stock assessment. Part 1. Manual. FAO Fisheries Technical Paper, 306(1), rev 2. 407 pp.
Staby A., Mehl S., Berg E., Aglen A., Korsbrekke K. 2018. Akustisk mengdemåling av sei og kysttorsk Finnmark-Møre høsten 2018. Toktrapport, Havforskningsinstituttet. Toktrapport/Havforskningsinstituttet/ISSN 15036294/Nr. 6–2020.10.06
Stramska M., Børsheim K.Y., Jankowski A., Søiland H., Cieszyńska A. 2018. Observations of coastal ocean currents in the Barents Sea (Porsangerfjord) during the summers of 2014 and 2015. Estuarine, Coastal and Shelf Science 211: 6-22.
Strand H.K. 2019. Porsangerfjorden 2.0 — En mulighetsstudie. Rapport fra Havforskningen, 2019-7. ISSN:1893-4536. 44 pp.
Strand H.K., Pedersen T., Christie H., Moy F. 2020. Synergism between cruising cod and ambush sculpin predators on 0-group gadoids is modified by daylight cycle and presence of aggressive wolffish. Journal of Experimental Marine Biology and Ecology 526:151356.
Strand H.K., Daleng, J., Eliassen, T., Follesø, T., Gjøsæter, H., Hansen, H.O., Johansen, H., Josefsen, H., Larsen, D., Martinsen, T., Nedreaas, K., Pedersen, T., Skern-Mauritzen, M., Strand, M. 2020. Fiskerier i Porsangerfjorden - Et møte mellom fjordbrukere og forskere. Havforskningsinstituttets rapportserie. In prep .
Sundelöf, A., Wennhage, H., Svedäng, H. 2013. A red herring from the Öresund (ICES40G2): the apparent recovery of the Large Fish Indicator (LFI) in the North Sea hides a non-trawled area. ICES Journal of Marine Science, 70: 1081–1084.
Sundet J.H., Hvingel C., Hjelset A.M. 2019. Kongekrabbe i norsk sone. Bestandstaksering og rådgiving 2019. Kongekrabberådgivingen for 2019, Havforskningsinstituttet. https://www.hi.no/resources/Bestandsvurderinger-av-kongekrabbe-for-2020-2.pdf.
Søvik G., Thangstad T. 2020. Results of the Norwegian Bottom Trawl Survey for Northern Shrimp (Pandalus borealis) in Skagerrak and the Norwegian Deep (ICES Divisions 3.a and 4.a east) in 2020. NAFO SCR Doc, 20(001), 31 pp.
Vetter, E.F. 1988 Estimation of natural mortality in fish stocks: a review, in Collected reprints. California: National marine fisheries service, Southwest fisheries science centre, pp. 25–40. doi: 10.1016/S0140-6736(00)85147-8.
Wassmann P., Svendsen H., Keck A., Reigstad M. 1996. Selected aspects of the physical oceanography and particle fluxes in fjords of northern Norway. Journal of Marine Systems, 8: 53–71.
Zimmermann F., Søvik G., Thangstad T.H. 2019. Kunnskapsstatus rekefelt langs norskekysten. Bestilling fra Fiskeridirektoratet. Rapport fra Havforskningen nr. 2019-15. ISSN:1893-4536. 13 pp.
13 - Vedlegg
13.1 - Vedlegg 1. Oseanografi
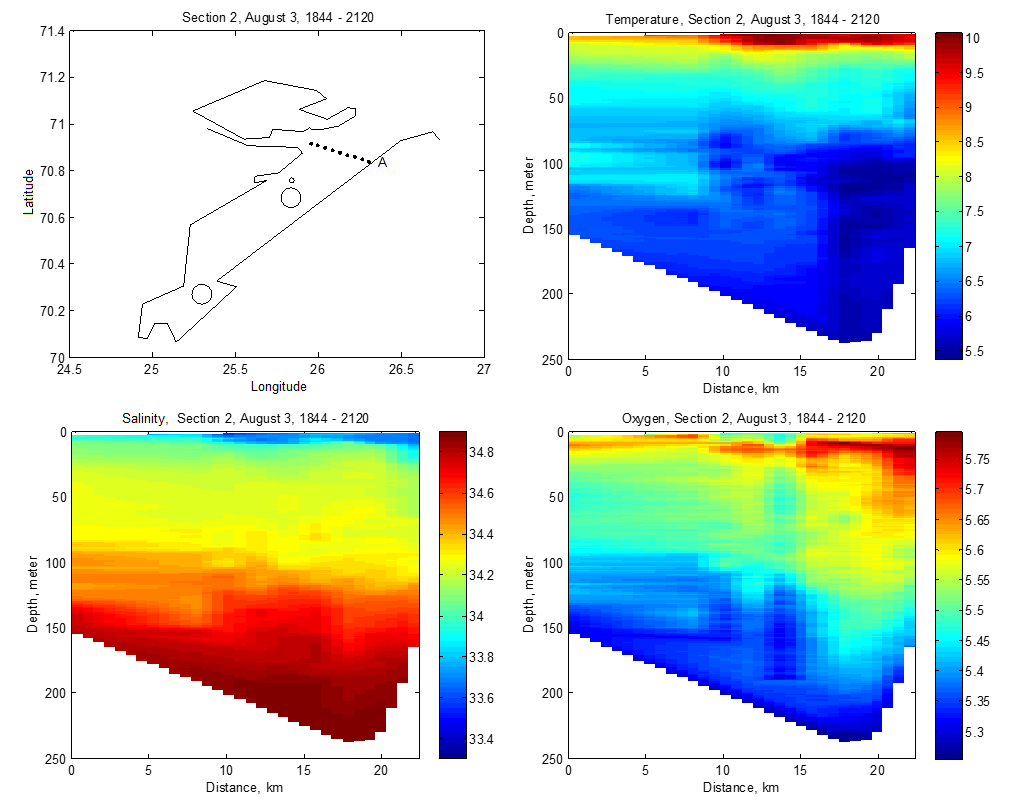
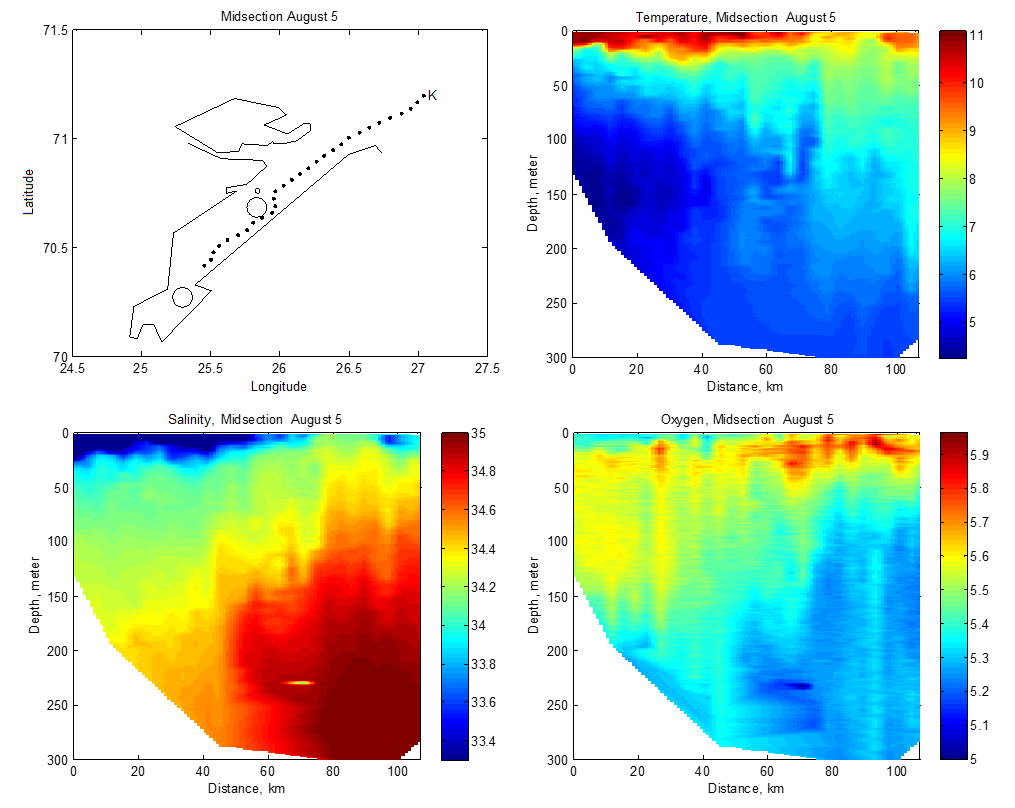
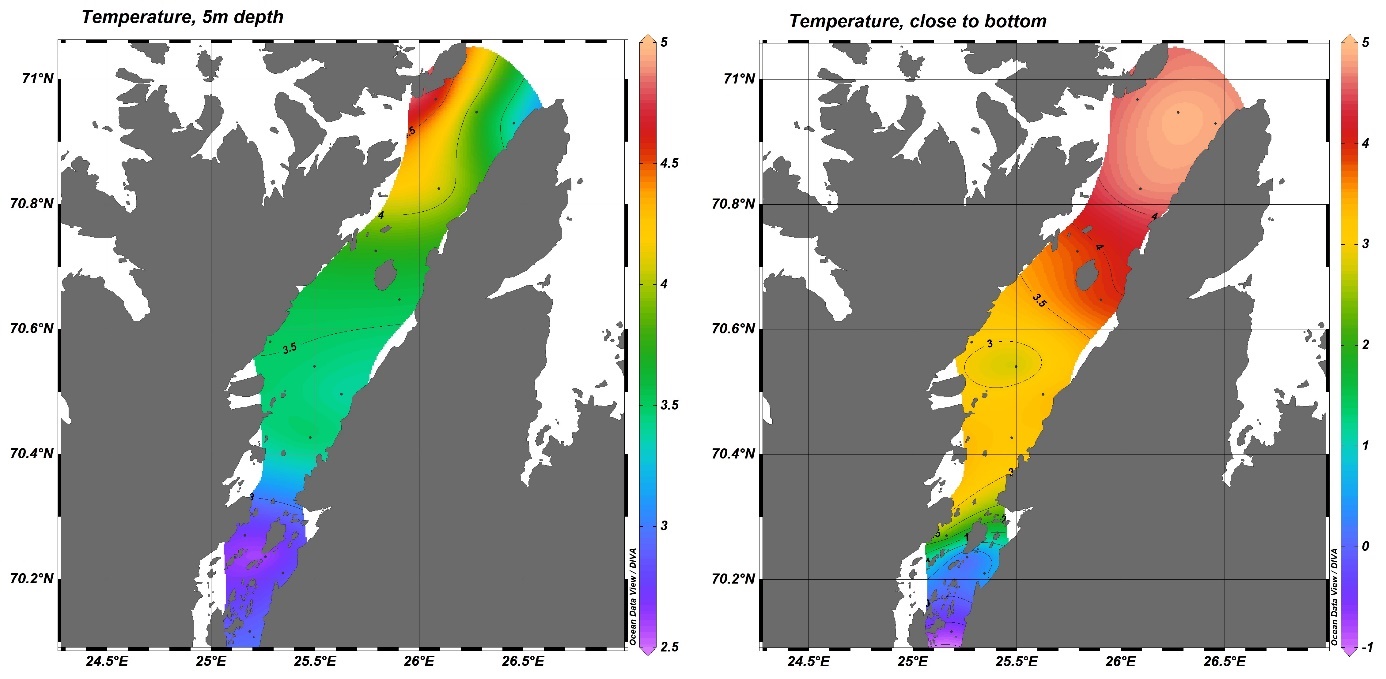
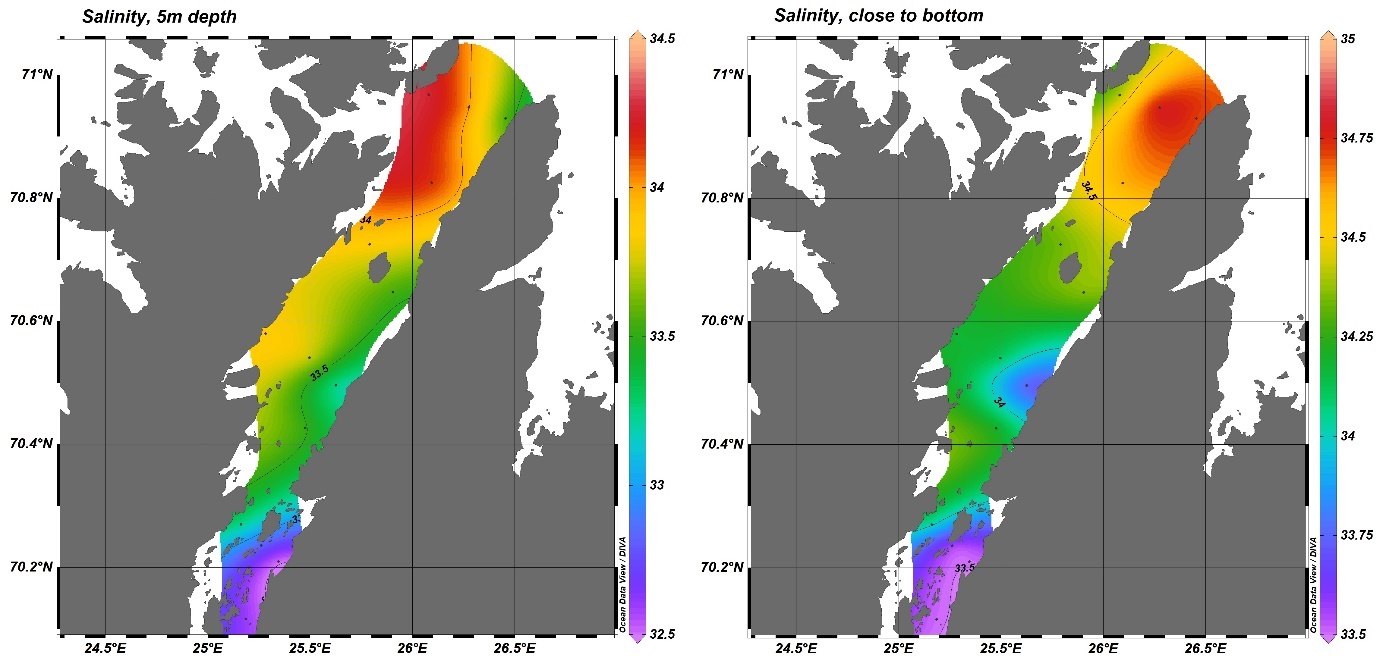
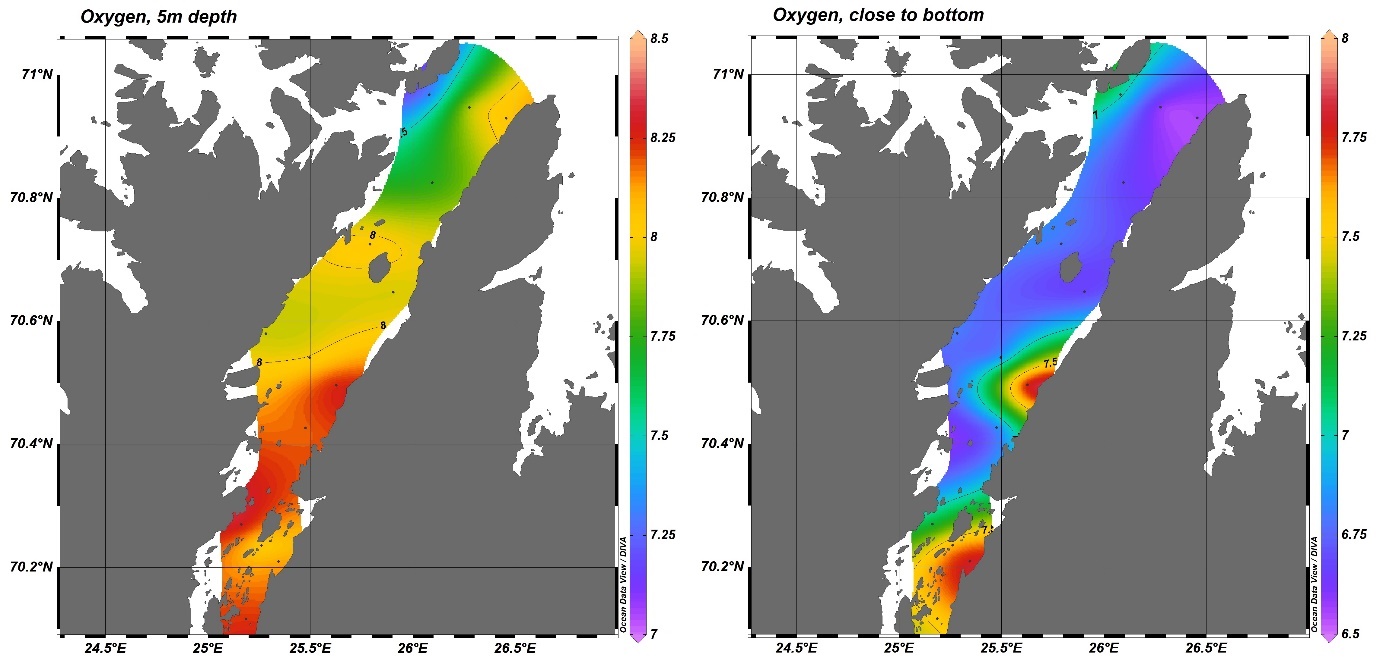
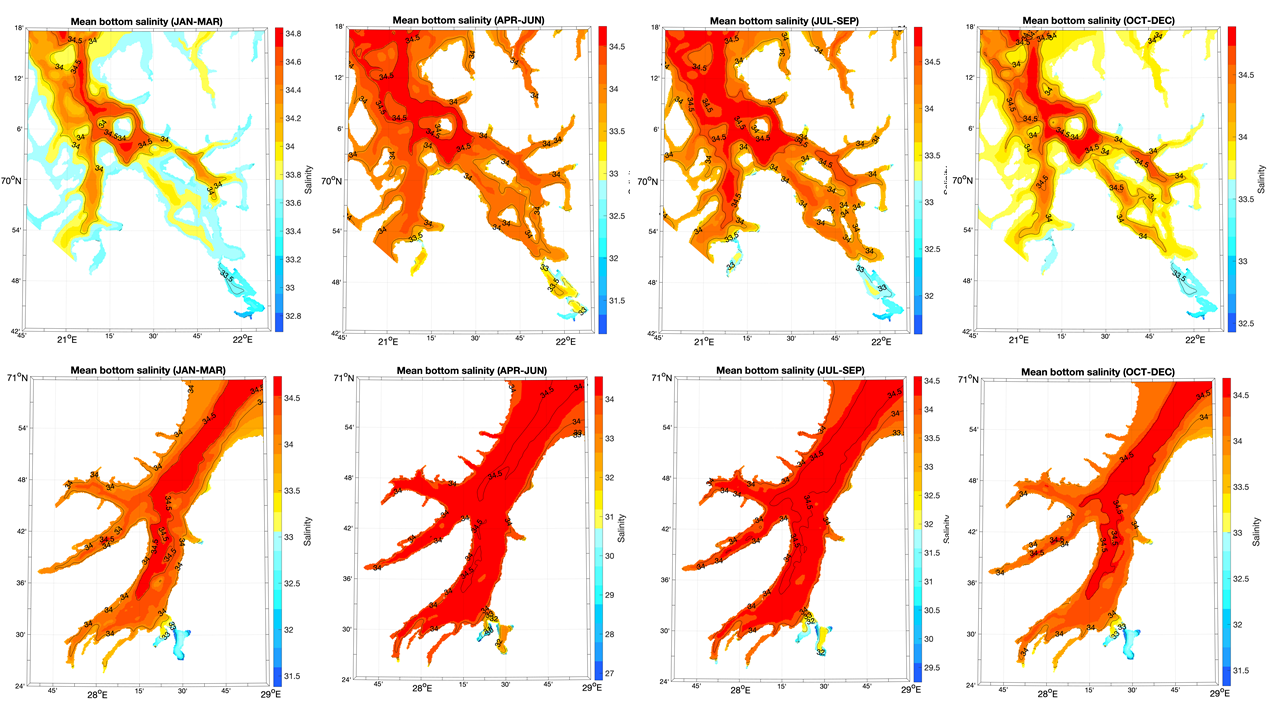
13.2 - Vedlegg 2. Artsregisteringer fra kartleggingstokt 2018 og 2019
| Art (eller artsgruppe) | Kvænangen | Ytre Porsangerfjorden | Indre Porsangerfjorden | Tanafjorden | ||||||||
| H 18 | V 19 | H 19 | H 18 | V 19 | H 19 | H 18 | V 19 | H 19 | H 18 | V 19 | H 19 | |
| Aega sp. | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| armføttinger | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 |
| A. propinqvus | 0 | 31 | 26 | 0 | 4 | 4 | 0 | 0 | 0 | 0 | 5 | 0 |
| åttearmete blekkspruter | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| blå brennmanet | 12 | 0 | 19 | 0 | 0 | 8 | 0 | 0 | 25 | 5 | 0 | 0 |
| blæretang | 0 | 0 | 0 | 63 | 0 | 0 | 67 | 0 | 0 | 47 | 0 | 0 |
| blåskjell | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 11 | 0 | 0 |
| blomkålkorall | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| blomkålkoraller | 0 | 0 | 0 | 11 | 4 | 0 | 83 | 100 | 0 | 0 | 0 | 0 |
| blomsterreke | 0 | 0 | 4 | 21 | 0 | 4 | 17 | 25 | 0 | 0 | 5 | 7 |
| børsteormer | 0 | 22 | 4 | 5 | 0 | 21 | 17 | 0 | 0 | 0 | 0 | 0 |
| breiflabb | 0 | 3 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 9 | 0 |
| brennmanet | 83 | 0 | 74 | 58 | 0 | 42 | 33 | 0 | 0 | 47 | 0 | 27 |
| brosme | 12 | 12 | 26 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| brunalger | 0 | 75 | 67 | 63 | 88 | 92 | 100 | 100 | 100 | 84 | 86 | 83 |
| brunpølse | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 0 |
| Crangon affinis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 0 |
| Crangonidae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 0 |
| Ctenodiscus sp. | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 0 |
| D. bonnieri | 0 | 34 | 26 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 |
| Drøbakskråkebolle | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 |
| dypvannsreke | 88 | 66 | 78 | 95 | 79 | 79 | 50 | 75 | 75 | 100 | 100 | 97 |
| eremittkreps | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| fingertare | 0 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| fireflekket var | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| firetrådet tangbrosme | 8 | 3 | 0 | 11 | 4 | 0 | 0 | 0 | 0 | 0 | 9 | 0 |
| fremmedlegeme | 0 | 22 | 15 | 0 | 12 | 17 | 0 | 25 | 0 | 0 | 41 | 10 |
| Fucus sp. | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| gapeflyndre | 67 | 62 | 44 | 95 | 96 | 75 | 100 | 50 | 25 | 79 | 100 | 50 |
| Gersemia sp. | 0 | 0 | 0 | 0 | 0 | 0 | 67 | 0 | 0 | 0 | 0 | 0 |
| glassmanet | 21 | 0 | 0 | 68 | 0 | 0 | 50 | 0 | 0 | 5 | 0 | 0 |
| glassmaneter | 67 | 3 | 33 | 0 | 4 | 88 | 0 | 0 | 50 | 0 | 0 | 7 |
| glassreke | 29 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| glassreker | 29 | 59 | 52 | 11 | 4 | 21 | 0 | 0 | 0 | 16 | 23 | 10 |
| glassvar | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 |
| glattulke | 0 | 0 | 0 | 0 | 0 | 0 | 83 | 25 | 50 | 0 | 0 | 0 |
| Gonatus sp. | 0 | 0 | 0 | 16 | 0 | 0 | 0 | 0 | 0 | 11 | 0 | 0 |
| Gonatus fabricii | 0 | 0 | 0 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| G. arcticus | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 |
| grisetang | 0 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Harmothoe sp. | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 |
| havålebrosme | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| havmus | 8 | 22 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| havsil | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hormathia sp. | 0 | 0 | 0 | 5 | 0 | 0 | 33 | 0 | 0 | 0 | 0 | 0 |
| hvitskate | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| hvitting | 58 | 72 | 78 | 53 | 54 | 96 | 33 | 25 | 50 | 32 | 64 | 77 |
| hydroider og småmaneter | 0 | 6 | 33 | 0 | 8 | 62 | 0 | 0 | 50 | 0 | 0 | 53 |
| hyse | 100 | 84 | 100 | 100 | 96 | 100 | 33 | 25 | 50 | 100 | 100 | 97 |
| isopoder | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 |
| kameleonslangestjerne | 0 | 0 | 0 | 11 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 |
| kloskate | 8 | 6 | 11 | 11 | 8 | 4 | 17 | 0 | 0 | 11 | 5 | 7 |
| knurr | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 9 | 0 |
| kolmule | 12 | 9 | 22 | 0 | 0 | 8 | 0 | 0 | 0 | 0 | 5 | 7 |
| kongekrabbe | 0 | 0 | 0 | 21 | 4 | 17 | 50 | 100 | 0 | 5 | 0 | 7 |
| koralldyr | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| kråkeboller | 0 | 9 | 7 | 0 | 4 | 0 | 0 | 25 | 0 | 0 | 0 | 0 |
| krill | 42 | 41 | 56 | 53 | 25 | 50 | 33 | 0 | 50 | 0 | 68 | 70 |
| krokulke | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 25 | 0 | 0 | 0 | 0 |
| kveite | 0 | 12 | 4 | 5 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| kystringbuk | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 |
| laksesild | 4 | 50 | 15 | 11 | 83 | 25 | 0 | 0 | 0 | 26 | 91 | 73 |
| lange | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| langfinnet snottfisk | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| langhalet langebarn | 0 | 3 | 0 | 11 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 |
| Liparis sp. | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 |
| lodde | 42 | 41 | 26 | 84 | 62 | 71 | 83 | 75 | 100 | 37 | 68 | 33 |
| lomre | 4 | 3 | 4 | 21 | 4 | 0 | 0 | 0 | 0 | 5 | 14 | 3 |
| lusuer | 54 | 12 | 41 | 26 | 12 | 8 | 0 | 0 | 0 | 74 | 41 | 67 |
| lysing | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 |
| lysprikkfisker | 0 | 53 | 52 | 0 | 0 | 46 | 0 | 0 | 0 | 84 | 59 | 0 |
| maneter | 0 | 3 | 15 | 16 | 0 | 0 | 50 | 0 | 0 | 68 | 0 | 0 |
| medusahoder | 0 | 0 | 11 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 |
| Meganyctiphanes sp. | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| mudderbunnsjørose | 0 | 9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| mudderreker | 0 | 12 | 22 | 5 | 0 | 8 | 0 | 0 | 0 | 0 | 0 | 7 |
| muddersjøstjerne | 0 | 19 | 11 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 |
| nesledyr | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 |
| nordlig knurrulke | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 |
| nordlig lysprikkfisk | 50 | 0 | 0 | 32 | 29 | 0 | 0 | 0 | 0 | 0 | 0 | 73 |
| norsk storkrill | 0 | 0 | 0 | 16 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| øyepål | 54 | 41 | 44 | 42 | 71 | 46 | 0 | 0 | 0 | 84 | 73 | 70 |
| Pandalidae | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| panserreker | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 0 |
| Pentamera calcigera | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 |
| Periphylla periphylla | 33 | 31 | 41 | 5 | 12 | 12 | 0 | 0 | 0 | 26 | 41 | 20 |
| Phaeocystaceae | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 |
| Ptycogena sp. | 0 | 0 | 0 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ptycogena sp. | 0 | 0 | 0 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| pukkelringbuk | 0 | 0 | 0 | 5 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 |
| pyntekrabber | 0 | 3 | 4 | 0 | 12 | 17 | 17 | 25 | 50 | 5 | 9 | 0 |
| ribbemaneter | 0 | 0 | 0 | 16 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 |
| rødalger | 0 | 19 | 4 | 5 | 0 | 4 | 17 | 0 | 0 | 0 | 23 | 0 |
| rødpølse | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| rødspette | 8 | 28 | 11 | 47 | 62 | 29 | 67 | 25 | 75 | 21 | 23 | 13 |
| rognkjeks | 4 | 9 | 4 | 0 | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 |
| rur | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 |
| sagtang | 0 | 0 | 0 | 16 | 0 | 0 | 100 | 0 | 0 | 26 | 0 | 0 |
| sandflyndre | 0 | 6 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sandpyntekrabbe | 0 | 0 | 0 | 5 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 |
| sei | 17 | 16 | 44 | 42 | 4 | 21 | 0 | 25 | 0 | 21 | 32 | 10 |
| sekkedyr | 0 | 0 | 0 | 0 | 0 | 8 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sertulariidae | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sildG03 | 79 | 91 | 59 | 26 | 29 | 88 | 50 | 100 | 75 | 21 | 82 | 83 |
| sjøanemoner | 0 | 0 | 11 | 5 | 0 | 4 | 0 | 0 | 0 | 0 | 5 | 0 |
| sjøgress | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 |
| sjøkjeks | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sjønellik | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 |
| sjøpølser | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 3 |
| sjøroser | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sjøstjerner | 17 | 25 | 33 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sjøtre | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sjustripeskjell | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| skjellbrosme | 8 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 0 |
| skrubbe | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| slangestjerner | 0 | 3 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 |
| slimål | 0 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| småflekket rødhai | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 |
| smørflyndre | 88 | 78 | 70 | 32 | 42 | 17 | 0 | 0 | 0 | 47 | 73 | 43 |
| snabeluer | 0 | 28 | 0 | 26 | 0 | 8 | 0 | 0 | 0 | 5 | 23 | 0 |
| snegler | 0 | 6 | 0 | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 0 |
| sølvtangbrosme | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sølvtorsk | 58 | 56 | 59 | 32 | 58 | 29 | 0 | 0 | 0 | 68 | 82 | 50 |
| Spirontocaris sp. | 0 | 3 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| stormaneter | 0 | 0 | 0 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| strømsild | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Strongylocentrotus sp. | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sukkertare | 0 | 0 | 0 | 53 | 0 | 0 | 100 | 0 | 0 | 5 | 0 | 0 |
| svamper | 0 | 3 | 4 | 11 | 0 | 8 | 50 | 0 | 0 | 5 | 14 | 3 |
| sypike | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 |
| tang | 83 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| tangsprell | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 0 |
| tare | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| T. inermis | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| tiarmete blekkspruter | 0 | 3 | 0 | 0 | 0 | 12 | 0 | 0 | 0 | 0 | 0 | 7 |
| tiskjegg | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 25 | 0 | 0 | 0 | 0 |
| torsk | 100 | 81 | 100 | 100 | 92 | 96 | 100 | 50 | 25 | 100 | 100 | 93 |
| tverrhalet langebarn | 0 | 0 | 4 | 5 | 12 | 17 | 33 | 25 | 25 | 0 | 23 | 0 |
| uerslekten | 0 | 0 | 0 | 0 | 0 | 38 | 0 | 0 | 0 | 0 | 0 | 50 |
| urskjell | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| vanlig ålebrosme | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 |
| vanlig uer | 46 | 47 | 52 | 68 | 29 | 42 | 0 | 0 | 0 | 100 | 82 | 73 |
| vanlig ulke | 0 | 0 | 0 | 0 | 0 | 0 | 33 | 25 | 50 | 0 | 0 | 0 |
| vassild | 21 | 28 | 19 | 26 | 29 | 38 | 0 | 0 | 0 | 53 | 64 | 73 |
| Art (eller artsgruppe) | Kvænangen | Ytre Porsangerfjorden | Indre Porsangerfjorden | Tanafjorden | ||||||||
| H 18 | V 19 | H 19 | H 18 | V 19 | H 19 | H 18 | V 19 | H 19 | H 18 | V 19 | H 19 | |
| Aega sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| armføttinger | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A, propinqvus | 0 | 0,1 | 0,1 | 0 | 0,3 | 0 | 0 | 0 | 0 | 0 | 0,1 | 0 |
| åttearmete blekkspruter | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| blå brennmanet | 0,9 | 0 | 1,1 | 0 | 0 | 0 | 0 | 0 | 0,1 | 0,3 | 0 | 0 |
| blæretang | 0 | 0 | 0 | 0,4 | 0 | 0 | 1,1 | 0 | 0 | 0,1 | 0 | 0 |
| blåskjell | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| blomkålkorall | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| blomkålkoraller | 0 | 0 | 0 | 0 | 0,1 | 0 | 0,1 | 1 | 0 | 0 | 0 | 0 |
| blomsterreke | 0 | 0 | 0 | 0,2 | 0 | 0 | 0,4 | 0 | 0 | 0 | 0 | 0 |
| børsteormer | 0 | 0 | 0,1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| breiflabb | 0 | 7,4 | 0 | 0 | 6,7 | 0 | 0 | 0 | 0 | 0 | 8,5 | 0 |
| brennmanet | 25,7 | 0 | 7 | 9,7 | 0 | 14,1 | 18,2 | 0 | 0 | 11,5 | 0 | 2,2 |
| brosme | 14,3 | 8,7 | 12,6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| brunalger | 0 | 0,5 | 1,1 | 0,4 | 4,1 | 3,5 | 6,8 | 94,3 | 8,7 | 1,5 | 0,5 | 0,7 |
| brunpølse | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1,9 | 0 | 0 | 0 | 0 |
| Crangon affinis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Crangonidae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ctenodiscus sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| D. bonnieri | 0 | 0,2 | 0,3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Drøbakskråkebolle | 0 | 0 | 0 | 0 | 0 | 0 | 0,1 | 0 | 0 | 0 | 0 | 0 |
| dypvannsreke | 31,2 | 24,9 | 39,7 | 13,2 | 12,5 | 15,2 | 185,6 | 139,5 | 88,1 | 18,7 | 26,2 | 7,4 |
| eremittkreps | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| fingertare | 0 | 0,1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| fireflekket var | 0,5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| firetrådet tangbrosme | 0,4 | 0,1 | 0 | 0,1 | 0,4 | 0 | 0 | 0 | 0 | 0 | 0,2 | 0 |
| fremmedlegeme | 0 | 0,3 | 0,1 | 0 | 0,1 | 0,2 | 0 | 0 | 0 | 0 | 0 | 0,1 |
| Fucus sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| gapeflyndre | 5,9 | 1,7 | 3,4 | 4,2 | 4,7 | 0,9 | 2,8 | 0,9 | 2 | 0,9 | 1,7 | 0,9 |
| Gersemia sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| glassmanet | 0,5 | 0 | 0 | 1,2 | 0 | 0 | 0,6 | 0 | 0 | 0,3 | 0 | 0 |
| glassmaneter | 51,9 | 0,1 | 0,7 | 0 | 0,1 | 1,6 | 0 | 0 | 1,7 | 0 | 0 | 0,2 |
| glassreke | 9,5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| glassreker | 8,7 | 27,6 | 4,8 | 0 | 0,3 | 0,3 | 0 | 0 | 0 | 1,3 | 0,8 | 1,7 |
| glassvar | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1,1 |
| glattulke | 0 | 0 | 0 | 0 | 0 | 0 | 0,6 | 3,5 | 0,7 | 0 | 0 | 0 |
| Gonatus sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Gonatus fabricii | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| G. arcticus | 0 | 0 | 0 | 0 | 0 | 0 | 11,3 | 0 | 0 | 0 | 0 | 0 |
| grisetang | 0 | 0,3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Harmothoe sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| havålebrosme | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| havmus | 1,3 | 4,5 | 9,9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| havsil | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hormathia sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0,1 | 0 | 0 | 0 | 0 | 0 |
| hvitskate | 0 | 20,4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| hvitting | 14,4 | 12,6 | 16,4 | 0,8 | 1,9 | 1,8 | 23,3 | 0,2 | 4,8 | 0,5 | 5,8 | 1,5 |
| hydroider og småmaneter | 0 | 21,7 | 126,3 | 0 | 0 | 35,8 | 0 | 0 | 122,3 | 0 | 0 | 12,2 |
| hyse | 127,5 | 129,3 | 214,6 | 66,7 | 52,5 | 43,3 | 58,8 | 809,9 | 304,1 | 189,3 | 146,1 | 104,8 |
| isopoder | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| kameleonslangestjerne | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| kloskate | 2,2 | 3,3 | 3,8 | 2,3 | 2,4 | 3,2 | 1,9 | 0 | 0 | 0,6 | 3,2 | 1 |
| knurr | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,7 | 0 |
| kolmule | 5,2 | 2,8 | 1,8 | 0 | 0 | 0,9 | 0 | 0 | 0 | 0 | 0,1 | 0,4 |
| kongekrabbe | 0 | 0 | 0 | 16,3 | 1,8 | 3 | 15,1 | 3,2 | 0 | 3,1 | 0 | 3,1 |
| koralldyr | 0 | 8,3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| kråkeboller | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| krill | 8,1 | 1,7 | 4,9 | 0,5 | 2,9 | 0,4 | 0,7 | 0 | 0,3 | 0 | 2,8 | 1,9 |
| krokulke | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| kveite | 0 | 18,6 | 6,7 | 38,9 | 45,4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| kystringbuk | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| laksesild | 0 | 0 | 0 | 0 | 0,7 | 0 | 0 | 0 | 0 | 0 | 0,4 | 0 |
| lange | 0 | 8,8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| langfinnet snottfisk | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| langhalet langebarn | 0 | 0 | 0 | 0,1 | 0 | 0 | 0,1 | 0 | 0 | 0 | 0 | 0 |
| Liparis sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| lodde | 0,4 | 1,5 | 0,5 | 1,8 | 0,2 | 3,3 | 14,8 | 14,9 | 26 | 0 | 0,4 | 0,1 |
| lomre | 2,8 | 0,6 | 3,6 | 0,9 | 1,4 | 0 | 0 | 0 | 0 | 0,7 | 1,2 | 1,6 |
| lusuer | 0,9 | 3,6 | 3,3 | 1,3 | 0,2 | 0,8 | 0 | 0 | 0 | 0,4 | 0,4 | 0,4 |
| lysing | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,6 | 0 |
| lysprikkfisker | 0 | 0,2 | 0,5 | 0 | 0 | 0,2 | 0 | 0 | 0 | 0,1 | 0,1 | 0 |
| maneter | 0 | 0 | 11 | 13,8 | 0 | 0 | 144,1 | 0 | 0 | 273,7 | 0 | 0 |
| medusahoder | 0 | 0 | 1,1 | 0 | 0 | 2,3 | 0 | 0 | 0 | 0 | 0 | 0 |
| Meganyctiphanes sp. | 0,4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| mudderbunnsjørose | 0 | 0,6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| mudderreker | 0 | 0,1 | 0,1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| muddersjøstjerne | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| nesledyr | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| nordlig knurrulke | 0 | 0 | 0 | 0 | 0 | 0 | 0,1 | 0 | 0 | 0 | 0 | 0 |
| nordlig lysprikkfisk | 0,8 | 0 | 0 | 0 | 0,9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| norsk storkrill | 0 | 0 | 0 | 1,7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| øyepål | 0,7 | 1,8 | 6,3 | 4,1 | 1,4 | 2,8 | 0 | 0 | 0 | 13,9 | 1,1 | 1,6 |
| Pandalidae | 13,3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| panserreker | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,1 | 0 | 0 | 0 | 0 |
| Pentamera calcigera | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Periphylla periphylla | 10,5 | 7,7 | 9,8 | 1,9 | 4 | 5,8 | 0 | 0 | 0 | 2,1 | 3,1 | 2,9 |
| Phaeocystaceae | 0 | 0 | 0 | 0 | 0 | 0 | 0,4 | 0 | 0 | 0 | 0 | 0 |
| Ptycogena sp. | 0 | 0 | 0 | 0,3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ptycogena sp. | 0 | 0 | 0 | 0,1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| pukkelringbuk | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| pyntekrabber | 0 | 0,1 | 0,3 | 0 | 0 | 0 | 0,3 | 1,3 | 0 | 0 | 0 | 0 |
| ribbemaneter | 0 | 0 | 0 | 2155,6 | 0 | 0 | 16,3 | 0 | 0 | 0 | 0 | 0 |
| rødalger | 0 | 0,1 | 0,1 | 0,2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| rødpølse | 0 | 0 | 0,7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| rødspette | 33,9 | 25,7 | 5,4 | 30,2 | 0 | 6,4 | 3,9 | 11,4 | 1,1 | 3,2 | 11,8 | 5,7 |
| rognkjeks | 13,7 | 7,4 | 4 | 0 | 0 | 0 | 0 | 0 | 2,5 | 0 | 0 | 0 |
| rur | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sagtang | 0 | 0 | 0 | 0,3 | 0 | 0 | 2,1 | 0 | 0 | 0 | 0 | 0 |
| sandflyndre | 0 | 7 | 0 | 0,9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sandpyntekrabbe | 0 | 0 | 0 | 0 | 0 | 0 | 0,2 | 0 | 0 | 0 | 0 | 0 |
| sei | 3,3 | 4,6 | 34,2 | 1,1 | 11,6 | 3,6 | 0 | 0,1 | 0 | 0,8 | 9,7 | 1,4 |
| sekkedyr | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sertulariidae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sildG03 | 11,6 | 23,8 | 16,1 | 0,1 | 0,1 | 0,7 | 122,7 | 16 | 91,9 | 0,2 | 0,7 | 0,5 |
| sjøanemoner | 0 | 0 | 0,5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,4 | 0 |
| sjøgress | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sjøkjeks | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sjønellik | 0 | 0 | 0 | 0 | 0 | 0 | 3,2 | 0 | 0 | 0 | 0 | 0 |
| sjøpølser | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,1 |
| sjøroser | 0,1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sjøstjerner | 0 | 0,2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sjøtre | 0 | 0 | 142,9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sjustripeskjell | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| skjellbrosme | 4 | 5,4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| skrubbe | 0 | 1,4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| slangestjerner | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| slimål | 0 | 0,1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| småflekket rødhai | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1,8 | 0 |
| smørflyndre | 5,6 | 14,8 | 4,1 | 1,6 | 3,5 | 2,1 | 0 | 0 | 0 | 4,3 | 3,2 | 2,8 |
| snabeluer | 0 | 0 | 0 | 4,6 | 0 | 0,3 | 0 | 0 | 0 | 2,6 | 1,1 | 0 |
| snegler | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sølvtangbrosme | 0 | 0,4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sølvtorsk | 0,9 | 5,1 | 1,3 | 0,9 | 0,7 | 0,1 | 0 | 0 | 0 | 0,6 | 8,2 | 1,1 |
| Spirontocaris sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| stormaneter | 0 | 0 | 0 | 4488,7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| strømsild | 0,4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Strongylocentrotus sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| sukkertare | 0 | 0 | 0 | 4,7 | 0 | 0 | 1,5 | 0 | 0 | 0 | 0 | 0 |
| svamper | 0 | 0 | 0,3 | 2 | 0 | 0,1 | 0,6 | 0 | 0 | 0,5 | 2,6 | 0 |
| sypike | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,4 | 0 |
| tang | 1,3 | 0 | 0 | 0,2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| tangsprell | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,1 | 0 | 0 | 0 | 0 |
| tare | 3,4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Thysanoessa inermis | 0 | 0 | 0 | 0,3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| tiarmete blekkspruter | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| tiskjegg | 0 | 0 | 0 | 0 | 0 | 0 | 0,1 | 0 | 0 | 0 | 0 | 0 |
| torsk | 0 | 42,6 | 35,9 | 44,4 | 41,6 | 33 | 4,7 | 6,5 | 31 | 52,5 | 24,5 | 41,5 |
| tverrhalet langebarn | 0 | 0 | 0 | 0 | 0 | 0 | 0,1 | 0 | 0 | 0 | 0 | 0 |
| uerslekten | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,1 |
| urskjell | 0,1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| vanlig ålebrosme | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| vanlig uer | 20,5 | 36,8 | 20,3 | 2,8 | 1,6 | 2,6 | 0 | 0 | 0 | 11,7 | 4,9 | 12,2 |
| vanlig ulke | 0 | 0 | 0 | 0 | 0 | 0 | 0,2 | 1,4 | 1,3 | 0 | 0 | 0 |
| vassild | 4,9 | 3,7 | 23,3 | 0,7 | 0,2 | 0,6 | 0 | 0 | 0 | 0,5 | 0,3 | 0,3 |
| Kvænangen | Ytre Porsangerfj. | Indre Porsangerfj. | Tanafjorden | |||||
| Art | Stasjoner | Antall | Stasjoner | Antall | Stasjoner | Antall | Stasjoner | Antall |
| blomsterreke | 4 | 73 | 18 | 94 | 4 | 35 | 12 | 417 |
| dypvannsreke | 28 | 1536 | 28 | 654 | 5 | 3359 | 21 | 2457 |
| Hormathia sp. | 1 | 3 | 0 | 0 | 1 | 1 | 0 | 0 |
| Pagurus pubescens | 0 | 0 | 0 | 0 | 1 | 2 | 0 | 0 |
| pyntekrabber | 1 | 2 | 4 | 4 | 2 | 5 | 0 | 0 |
| reker | 6 | 29 | 26 | 183 | 3 | 190 | 3 | 7 |
| sandpyntekrabbe | 0 | 0 | 0 | 0 | 1 | 3 | 0 | 0 |
| vanlig ulke | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 |
| Atlantopandalus propinqvus | 4 | 9 | 0 | 0 | 0 | 0 | 3 | 8 |
| Colus sp. | 2 | 6 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ctenodiscus sp. | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Epizoanthus sp. | 3 | 6 | 0 | 0 | 0 | 0 | 0 | 0 |
| eremittkreps | 6 | 25 | 3 | 8 | 0 | 0 | 1 | 1 |
| havedderkopper | 2 | 3 | 0 | 0 | 0 | 0 | 0 | 0 |
| høyrygget skolest | 1 | 2 | 0 | 0 | 0 | 0 | 0 | 0 |
| kongsnegl | 3 | 7 | 0 | 0 | 0 | 0 | 0 | 0 |
| muslinger | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| sjøanemoner | 2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 |
| sjøstjerner | 2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 |
| sjustripeskjell | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| slangestjerner | 2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 |
| slimål | 1 | 18 | 0 | 0 | 0 | 0 | 0 | 0 |
| Spirontocaris sp. | 2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 |
| torsk | 2 | 4 | 2 | 3 | 0 | 0 | 0 | 0 |
| trollhummer | 1 | 1 | 3 | 4 | 0 | 0 | 6 | 13 |
| brosme | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| kongekrabbe | 0 | 0 | 2 | 3 | 0 | 0 | 1 | 1 |
| krokulke | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| panserreker | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| uerslekten | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 3 |
| ukjent | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 9 |
| annen fisk | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 |
| langfinnet snottfisk | 0 | 0 | 2 | 2 | 0 | 0 | 0 | 0 |
| ringbukfamilien | 0 | 0 | 2 | 4 | 0 | 0 | 0 | 0 |
13.3 - Vedlegg 3. Samfunnsanalyse av data fra bomtrålundersøkelser.
Da to forskjellige forskningsskip og flere typer vinsjer er blitt brukt i tidsperioden 2007-2019 er det ikke mulig å sammenligne bomtrålhalene kvantitativt. Biomassen av alle artene ble derfor omgjort til andel av totalbiomasse per trålhal. Disse tallene er brukt i analysene her.
For å identifisere faunistiske sammenliknbare stasjoner ble en multivariat «samfunnsanalyse» valgt (clusteranalyse med group-average og Bray-Curtis). Data fra alle år ble slått sammen (Figur V3.1). Totalt 214 evertebrater (bestemt til art eller slekt) som lever på bunn (bunndyr), ble inkludert i analysen etter standardisering av artsnavn mellom år og områder. Det ble valgt en oppløsning på 10 samfunn (cutting level ~40 %) til å beskrive bunndyrsvariasjonen fra område til område i Porsangerfjorden.
Stasjoner dypere enn 170 m ble kun tatt fra midtre Porsangerfjorden og utover mot åpent hav. Representative bilder av bunndyr er vist i Figurene V3.2, V3.3, V3.4.
Bunndyrssamfunn med risiko for bunntråling dypere enn 170 m
Samfunn 2 ble tatt i flere dype bassenger (99-234 m, gjennomsnittlig 140 m) fra den grunnere, indre delen til den dype, ytre delen av Porsangerfjorden utenfor terskelen («Ryggen») (Figurer V3.5, V3.6, Tabell V3.1). Den kommersielle dypvannsreken (Pandalus borealis) ble registret i alle trålhal med 2-67 % av totalbiomassen. Alle trålhalene tatt i de to indre østlige bassengene (Østerbotn og Rotenbottenjarga) var karakterisert ved forekomsten av panserreke, som ikke forekom i den ytre og midterste delen av fjorden. Derimot fantes skjellet Arctinula greenlandica i bassengene i den ytre og midterste delen, men ikke i de indre bassengene. Mens faunaen i trålhalene tatt i bassengene i indre Porsangerfjorden var ganske lik i årene 2007, 2009 og 2014, utviklet den seg til å bli mer lik bunndyrssamfunnene i bassengene i den midtre og ytre delen av Porsangerfjorden fra 2014 til 2019.
Samfunn 5 var, som Samfunn 2, utbredt i bassengene i hele fjorden (90-209 m), men også langs vestkysten (47-70 m) i den indre og midterste delen av fjorden. Alle, bortsett fra ett trålhal hadde opp til 99 % av biomassen bestående av kongekrabbe (Paralithodes camtschatica). I de indre delene av fjorden besto 44 % av biomassen i trålhalene av medusahodet Gorgonacephalus sp. og opptil 20 % av blomkålkorall (Nephtheidae). Trålhalene fra indre Porsangerfjorden liknet også på halene fra vestsiden av fjorden ved at de inneholdt sjøanemonen Hormathia sp. og sporadiske fangster av svamp. Trålhalene tatt hovedsakelig på østsiden av midtre Porsangerfjorden var karakterisert av dypvannsreke. Her ble det ikke tatt medusahode eller blomkålkoraller, og svamp ble sjelden tatt.
Samfunn 33 (218 og 233 m dyp) henholdsvis øst og vest for Tamsøya var dominert av Pandalidae reker (evt. dypvannsreken Pandalus borealis).
Samfunn 7 var utbredt på både grunt (48-71 m) og dypt (104-220 m) vann, hovedsakelig i midtre Porsangerfjorden. Ofte ble mange typer svamp (opptil 76 % av fangstbiomassen) og sjøanemonen Hormathia sp. (opp til 58 % av fangstbiomassen) fangstet med trålen. Under halvparten av fangstene hadde dypvannsreker, og mengdene var forholdsvis små.
Samfunn 43 var lokalisert i Smørfjorden på 57 m dyp, og i den ytre delen av Olderfjorden på 221 m dyp på vestsiden av midtre Porsangerfjorden. Disse to lokalitetene var like pga. forekomst av polychaetene Pectinaria sp. (opptil 90 % av fangstbiomassen) og Lumbrineris sp. (opptil 4 % av fangstbiomassen). Kun den dype stasjonen i Olderfjorden hadde 12 % biomasse bestående av Pandalidae reker (Pandalus montagui).
Samfunn 48 lokalisert på vestsiden (159 m) og østsiden (74 og 251 m) av midtre Porsangerfjorden var totalt dominert av polychaeter (flere arter) og med opptil 4 % dypvannsrekebiomasse i fangstene.
Samfunn 61 ble registrert på én stasjon nordvest for Tamsøya på 218 m dyp. Denne stasjonen var dominert av sjøanemonen Metridium senile (82 % av biomassen). Dypvannsreke utgjorde mindre enn 2 % av biomassen.
Samfunn 62 besto av bare én stasjon som lå nord for Tamsøya på 221 m dyp. Denne var dominert av sneglen Bulbus smithi (80 % av biomassen) og av tanglopper innen slekten Gammarus sp. (10 %).
Bunndyrssamfunn uten risiko for bunntråling
Samfunn 1 ble registret kystnært i indre deler av Porsangerfjorden, på grunt vann mellom 27 og 44 m dyp (gjennomsnittlig 36 m dyp). Her dominerte kråkeboller (Strongylocentrotus sp.), haneskjell (Chlamys islandica), og panserreken (Sclerocrangon sp.).
Samfunn 32 (én stasjon på 155 m dyp) var dominert av en blanding av blomkålkorallen Nephtheidae (90 % av fangstbiomassen) og Pandalidae reker (opptil 2 % av biomassen).



| Porsangersamfunn | Arter funnet i alle trålhal | Arter funnet i mange trålhal, hvor biomassen dominerte totalbiomassen |
| Samfunn 1 | Strongylocentrotus sp. | Buccinidae indet |
| 6 stasjoner | Chlamys islandica | Ophiura sarsi |
| 68 arter | Sclerocrangon sp. | Pagurus pubescens |
| 36 m | Pandalus montagui | Nephtys sp. |
| Indre Porsangerfj. | ||
| Samfunn 2 | Pandalus borealis | Ctenodiscus crispatus |
| 14 stasjoner | Ciliatocardium ciliatum | |
| 82 arter | Bathyarcha glacialis | |
| 140 m | Ophiura sarsi | |
| Vidt utbredt | Sabinea septemcarinata | |
| Samfunn 5 | Paralithodes camtschatica (0-99 %) | |
| 14 stasjoner | Gorgonacephalus sp. (0-44 %) | |
| 98 arter | Pandalus borealis | |
| 123 m | Ciliatocardium ciliatum | |
| Indre og midtre Porsangerfj. | Nephtheidae | |
| Samfunn 7 | Porifera | |
| 18 stasjoner | Hormathia digitata | |
| 168 arter | Pagurus pubescens | |
| 117 m | ||
| I hovedsak midtre Porsangerfj. | ||
| Samfunn 32 | Nephthyidae (mangebørsteorm) (97 %) | |
| 1 stasjon | ||
| 26 arter | ||
| 155 m | ||
| Midtre Porsangerfj. | ||
| Samfunn 33 | Pandalidae (80%) | |
| 2 stasjoner | ||
| 28 arter | ||
| 155 m | ||
| Ytre Porsangerfj. | ||
| Samfunn 43 | Pectinaria sp. (opptil 90 %) | Yoldiella sp. (opptil 30 %) |
| 2 stasjoner | Lumbrineris sp. | |
| 20 arter | Nephtys sp. | |
| 139 m | ||
| Vest i midtre Porsangerfj. | ||
| Samfunn 48 | Maldane sarsi | Sabellidae sp. |
| 3 stasjoner | Ampharetidae | |
| 40 arter | Nephtys sp. | |
| 162 m | Myriochele oculata | |
| Midtre Porsangerfj. | ||
| Samfunn 61 | Metridium senile (82 %) | |
| 1 stasjon | Ampharetidae | |
| 31 arter | ||
| 219 m | ||
| Ytre Porsangerfj. | ||
| Samfunn 62 | Bulbus smithi (80 %) | |
| 1 stasjon | Gammarus sp. (10 %) | |
| 24 arter | ||
| 221 m | ||
| Ytre Porsangerfj. |
13.4 - Vedlegg 4. Registrering av bunnfauna fra Mareano videotransekter i 2011
| Startposisjon | Hele transektet | ||||
| Stasjon | Breddegrad | Lengdegrad | Gjennomsnittsdyp | Dyp_maks | Dyp_min |
| 732 | 70 56.21 | 26 28.68 | 213 | 238 | 128 |
| 733 | 70 55.78 | 26 28.68 | 207 | 247 | 12 |
| 735 | 70 50.12 | 26 10.86 | 197 | 310 | 69 |
| 734 | 70 45.95 | 26 00.06 | 189 | 232 | 81 |
| 732 | 733 | 734 | 735 | |
| Total antall artsobservasjoner | 40 | 36 | 42 | 40 |
| Total antall individobservasjoner | 92 | 67 | 121 | 166 |
| SVAMP | ||||
| Porifera | 9 | 6 | 20 | 15 |
| Axinella / Phakellia sp. | 4 | 2 | 10 | 12 |
| Geodia sp. | 3 | 2 | 3 | |
| Craniella / Mycale sp. | 5 | 5 | ||
| Flabellum like | 1 | |||
| Mycale lingua | 1 | 3 | 10 | |
| Tethia Craniella axinella | 7 | 10 | ||
| Totalt: | 16 | 12 | 48 | 52 |
| Koraller | ||||
| Primnoa resedaeformis | 3 | 1 | ||
| Drifa glomerata | 2 | 3 | 2 | |
| Drifa / Phakellia sp. | 1 | |||
| Paragorgia sp. | 1 | |||
| Paragorgia arborea | 1 | 1 | ||
| Totalt: | 6 | 6 | 3 |
| 732 | 733 | 734 | 735 | |
| Andre dyregrupper | ||||
| Bryozoa | 3 | 2 | ||
| Cerianthidae | 3 | 5 | 2 | |
| Actiniaria | 4 | 6 | 10 | 16 |
| Amphipoda | 1 | |||
| Antho dichotoma | 2 | 2 | ||
| Aplysilla sulfurea | 1 | 1 | 4 | |
| Ascidiacea colonial | 1 | |||
| Astarte sp. | 1 | |||
| Asteroidea | 1 | 1 | 4 | 3 |
| Aunice like | 1 | |||
| Brachiopoda | 2 | 1 | 1 | |
| Brosme brosme | 2 | |||
| Ceramaster granularis | 1 | 2 | 7 | 13 |
| Clupea harengus | 1 | |||
| Craniella hornera | 1 | |||
| Ceramaster sp. | 1 | |||
| død lodde | 4 | 3 | 9 | |
| død fisk | 4 | 1 | 2 | |
| Echinoidea | 1 | |||
| Echinus sp. | 3 | 1 | 1 | |
| feather? | 1 | |||
| Foraminifera agglutinated | 1 | 2 | ||
| torskefisk | 1 | |||
| Gadus morhua | 1 | 4 | 1 | |
| Gastropoda | 1 | |||
| håkjerring | 1 | |||
| Henricia sp. | 5 | 4 | 10 | 24 |
| Heteranomia sp. | 1 | |||
| Hippasteria phrygiana | 1 | 4 | 2 | |
| Hippolitidae | 1 | |||
| Holothuriidae | 1 | |||
| Hormathiidae | 3 | |||
| Horneria | 1 | 6 | ||
| Hyas sp. | 2 | |||
| Hydrozoa | 2 | |||
| Isops cf. | 1 | |||
| Latrunculia sp. | 1 | |||
| Melanogrammus aeglefinus | 2 | 1 | 4 | |
| Modiollula | 1 | |||
| Munida sp. | 1 | 1 | ||
| Natantia | 2 | |||
| Neptunea sp. | 1 | |||
| Bolocera | 1 | |||
| Ophiuorida | 3 | 1 | 3 | 4 |
| Pagurus sp. | 6 | 2 | 3 | |
| Pandalidae | 6 | 5 | 3 | |
| Pandalus borealis | 4 | |||
| Paralithodes | 1 | |||
| Parastichopus tremulus | 2 | 3 | 5 | |
| Pectinidae | 1 | |||
| Pelocinae | 1 | |||
| Pleuronectiformes | 2 | 1 | 1 | |
| Polymastidea | 1 | |||
| Pollachius virens | 1 | |||
| Polychaeta / Sabellida | 1 | 1 | ||
| Polymastia sp. | 4 | 1 | 7 | 6 |
| Porania cf. | 2 | |||
| Porania pulvillus | 2 | |||
| Psolus sp. | 3 | |||
| Pterasteridae | 1 | |||
| Pycnogonidae | 1 | |||
| Reteporella beaniana | 1 | 1 | ||
| rundskate | 1 | |||
| Sabellidae | 1 | 1 | ||
| Scatte | 1 | |||
| Sebastes sp. | 5 | 4 | 2 | 1 |
| Serpulidae | 1 | 1 | ||
| Serpulidae / Astarte | 1 | |||
| Stichastella cf. | 1 | |||
| Stylaster sp. | 1 | |||
| Teleostei | 1 | 2 | 3 | 1 |
| Trisopterus esmarkii | 1 | |||
| Tunicata colonial | 2 | |||
| Zooide | 1 |
13.5 - Vedlegg 5. Registreringer av bunnfauna fra kystressurstoktet høsten 2019
| Startposisjon | |||||||
| Fjord | Serienr. | Dato | Breddegrad | Lengdegrad | Dyp_maks | Dyp_min | Distanse |
| Porsangerfj. | 55206 | 05.10.2019 | 70 58.06 | 026 25.19 | 284 | 278 | 1,05 |
| 55207 | 05.10.2019 | 70 43.85 | 025 57.07 | 294 | 280 | 1,69 | |
| 55208 | 06.10.2019 | 70 25.61 | 025 18.80 | 140 | 136 | 0,88 | |
| 55210 | 06.10.2019 | 70 38.19 | 025 51.88 | 238 | 230 | 1,61 | |
| Kvænangen | 55230 | 13.10.2019 | 70 02.43 | 021 43.10 | 348 | 298 | 1,08 |
| 55231 | 13.10.2019 | 70 04.49 | 021 23.77 | 346 | 335 | 1,58 | |
| 55232 | 13.10.2019 | 70 11.94 | 021 04.00 | 393 | 372 | 1,58 | |
| Serienr. | Fylum | Klasse | Art /dyregruppe | Antall | Vekt (kg) |
| 55207 | Annelida | Polychaeta | Polychaeta | 3 | 0,001 |
| 55206 | Arthropoda | Malacostraca | Pontophilus norvegicus | 55 | 0,1 |
| 55206 | Arthropoda | Malacostraca | Atlantopandalus propinqvus | 11 | 0,01 |
| 55206 | Arthropoda | Malacostraca | Pandalus borealis | ||
| 55207 | Arthropoda | Malacostraca | Pontophilus norvegicus | 41 | 0,09 |
| 55207 | Arthropoda | Malacostraca | Pandalus borealis | ||
| 55208 | Arthropoda | Malacostraca | Sabinea septemcarinata | 206 | 0,17 |
| 55208 | Arthropoda | Malacostraca | Atlantopandalus propinqvus | 34 | 0,03 |
| 55208 | Arthropoda | Malacostraca | Sabinea sarsi | 17 | 0,03 |
| 55208 | Arthropoda | Malacostraca | Pandalus borealis | ||
| 55210 | Arthropoda | Malacostraca | Sabinea septemcarinata | 193 | 0,19 |
| 55210 | Arthropoda | Malacostraca | Pandalus borealis | ||
| 55210 | Arthropoda | Malacostraca | Pontophilus norvegicus | 193 | |
| 55208 | Cnidaria | Anthozoa | Hormathia digitata nodosa | 1 | 0,02 |
| 55208 | Cnidaria | Anthozoa | Hormathia digitata | 3 | 0,03 |
| 55207 | Echinodermata | Asteroidea | Ctenodiscus crispatus | 3 | 0,004 |
| 55206 | Echinodermata | Holothuroidea | Stichopus tremulus | 39 | 5,17 |
| 55208 | Echinodermata | Ophiuroidea | Ophiura sarsii | 1 | 0,002 |
| 55206 | Mollusca | Bivalvia | Pseudamussium peslutrae | 11 | 0,06 |
| 55207 | Mollusca | Bivalvia | Pseudamussium peslutrae | 2 | 0,008 |
| 55206 | Mollusca | Cephalopoda | Bathypolypus sp. | 1 | 0,004 |
| 55207 | Mollusca | Cephalopoda | Bathypolypus bairdii | 1 | 0,008 |
| 55208 | Mollusca | Cephalopoda | Bathypolypus sp. | 1 | 0,004 |
| 55207 | Porifera | Demospongiae | Phakellia ventilabrum* | 0,18 | |
| 55207 | Porifera | Demospongiae | Axinella rugosa* | 0,03 | |
| 55207 | Porifera | Demospongiae | Artemisina foliata* | 0,02 | |
| 55207 | Porifera | Porifera* | 0,08 | ||
| 55208 | Porifera | Demospongiae | Phakellia sp.* | 1 | 0,009 |
| 55208 | Porifera | Porifera* | 0,02 |
| Serienr. | Fylum | Klasse | Art /dyregruppe | Antall | Vekt (kg) |
| 55231 | Annelida | Polychaeta | Laetmonice filicornis | 7 | 0,58 |
| 55231 | Annelida | Polychaeta | Maldanidae | 16 | 0,02 |
| 55230 | Arthropoda | Malacostraca | Pandalus borealis | ||
| 55231 | Arthropoda | Malacostraca | Pasiphaea spp. | 165,65 | |
| 55231 | Arthropoda | Malacostraca | Pandalus borealis | ||
| 25532 | Arthropoda | Malacostraca | Lithodes maja | 2 | 0,81 |
| 25532 | Arthropoda | Malacostraca | Pagurus sp. | 8 | 0,05 |
| 25532 | Arthropoda | Malacostraca | Pandalus borealis | ||
| 25532 | Arthropoda | Malacostraca | Pasiphaea spp. | 92,70 | |
| 25532 | Arthropoda | Malacostraca | Pontophilus norvegicus | 0,02 | |
| 25532 | Arthropoda | Malacostraca | Atlantopandalus propinqvus | 0,52 | |
| 25532 | Arthropoda | Malacostraca | Spirontocaris sp. | 0,01 | |
| 25532 | Arthropoda | Malacostraca | Dichelopandalus bonnieri | 4,26 | |
| 55230 | Cnidaria | Anthozoa | Liponema multicorne | 1 | 0,24 |
| 55230 | Cnidaria | Anthozoa | Bolocera tuediae | 1 | 0,14 |
| 55230 | Cnidaria | Anthozoa | Actinostola callosa | 1 | 0,57 |
| 55231 | Cnidaria | Anthozoa | Hormathia digitata | 8 | 0,21 |
| 25532 | Cnidaria | Anthozoa | Hormathia d. parasitica | 8 | 0,008 |
| 25532 | Cnidaria | Anthozoa | Actinostola callosa | 39 | 0,06 |
| 25532 | Cnidaria | Anthozoa | Bolocera tuediae | 32 | 0,06 |
| 25532 | Cnidaria | Anthozoa | Caryophyllia smithii* | 0,14 | |
| 55230 | Echinodermata | Asteroidea | Ctenodiscus crispatus | 900 | 12,68 |
| 55231 | Echinodermata | Asteroidea | Ctenodiscus crispatus | 14777 | 45,92 |
| 25532 | Echinodermata | Asteroidea | Ctenodiscus crispatus | 1199 | 4,70 |
| 55230 | Echinodermata | Asteroidea | Hippasteria phrygiana | 1 | 0,04 |
| 25532 | Echinodermata | Echinoidea | Echinus esculentus | 8 | 0,03 |
| 55230 | Echinodermata | Echinoidea | Brisaster fragilis | 15 | 1,05 |
| 25532 | Echinodermata | Echinoidea | Brisaster fragilis | 8 | 0,05 |
| 55231 | Echinodermata | Ophiuroidea | Ophiura spp. | 64 | 0,11 |
| 25532 | Echinodermata | Ophiuroidea | Ophiura sp. | 24 | 0,03 |
| 25532 | Echinodermata | Ophiuroidea | Ophiurooidea | 24 | 0,008 |
| 55230 | Mollusca | Bivalvia | Astarte sp. | 4 | 0,008 |
| 55231 | Mollusca | Bivalvia | Astarte crenata | 32 | 0,16 |
| 55230 | Mollusca | Bivalvia | Pseudamussium peslutrae | 56 | 0,27 |
| 55231 | Mollusca | Bivalvia | Pseudamussium peslutrae | 352 | 2,63 |
| 25532 | Mollusca | Bivalvia | Pseudamussium peslutrae | 229 | 1,42 |
| 55230 | Mollusca | Gastropoda | Scaphander punctostriatus | 4 | 0,004 |
| 55231 | Mollusca | Gastropoda | Buccinum finnmarkianum | 16 | 0,28 |
| 55231 | Mollusca | Gastropoda | Berringius turtoni | 16 | 0,28 |
| 55231 | Mollusca | Gastropoda | Neptunea despecta | 3 | 0,38 |
13.6 - Vedlegg 6. Byttedyrsammensetning i magene til torsk, hyse og vanlig uer
| Kvænangen høsten 2018 | |||
| torsk < 35 cm | Vekt % | Ecopath-gruppe | Endelig fordeling i Ecopath (Vekt %) |
| Rhodophyceae | 0,02 | Kelp | 0,02 |
| Polychaeta | 0,12 | Detrivorous polychaetes | 0,12 |
| 0,12 | Predatory polychaetes | 0,12 | |
| Eunicidae | 0,08 | Detrivorous polychaetes | 0,08 |
| 0,08 | predatory polychaetes | 0,08 | |
| Crustacea | 7,27 | Small benthic crustaceans | 7,27 |
| Isopoda | 0,02 | Small benthic crustaceans | 0,02 |
| Gammaridea | 0,18 | Small benthic crustaceans | 0,18 |
| Hyperiidae | 0,15 | Small benthic crustaceans | 0,15 |
| Euphausiidae small | 12,22 | Small krill | 12,56 |
| Euphausiidae large | 12,22 | Large krill | 12,56 |
| Caridea | 7,05 | Pandalid and pasiphae | 7,05 |
| Pasiphaea | 3,94 | Pandalid and pasiphae | 3,94 |
| Pandalus borealis | 31,95 | Pandalid and pasiphae | 32,63 |
| Ophiuroidea | 0,52 | Other predatory benthic invertebrates | 0,52 |
| Teleostei | 20,33 | Other small demersal fish | 20,33 |
| Leptoclinus maculatus | 2,40 | Other small demersal fish | 2,40 |
| Indeterminatus | 1,36 | ||
| 100,00 | 100,00 | ||
| Kvænangen høsten 2018 | |||
| torsk ≥ 35 cm | Vekt % | Ecopath-gruppe | Endelig fordeling i Ecopath (Vekt %) |
| Phaeophyceae | 0,01 | Kelp | 0,04 |
| Virgulariidae | 0,01 | Large epibenthic suspension feeders | 0,04 |
| Polychaeta | 0,22 | Detrivorous polychaetes | 0,25 |
| predatory polychaetes | 0,03 | ||
| Maldanidae | 0,00 | Detrivorous polychaetes | 0,03 |
| Mytilidae | 0,01 | Large bivalves | 0,04 |
| Astarte | 0,12 | Large bivalves | 0,15 |
| Scaphopoda | 0,00 | Small molluscs | 0,03 |
| Crustacea | 0,82 | Small benthic crustaceans | 0,85 |
| Gammaridea | 0,00 | Small benthic crustaceans | 0,03 |
| Hyperiidae | 0,01 | Small benthic crustaceans | 0,04 |
| Euphausiidae small | 3,29 | Small krill | 1,68 |
| Euphausiidae large | Large krill | 1,68 | |
| Caridea | 1,61 | Crangonid shrimps | 1,64 |
| Spirontocaris sp. | 0,01 | Crangonid shrimps | 0,04 |
| Spirontocaris liljeborgii | 0,05 | Crangonid shrimps | 0,08 |
| Pontophilus norvegicus | 0,02 | Crangonid shrimps | 0,05 |
| Pandalus borealis | 30,46 | Pandalid and pasiphae | 30,49 |
| Pasiphaea | 2,67 | Pandalid and pasiphae | 2,70 |
| Munida sp. | 0,05 | Other large crustaceans | 0,08 |
| Munida sarsi | 0,03 | Other large crustaceans | 0,06 |
| Ctenodiscus crispatus | 2,51 | Detrivorous echinoderms | 2,54 |
| Teleostei | 14,55 | ||
| Clupea harengus | 0,46 | Herring | 2,31 |
| Mallotus villosus | 0,49 | Small pelagic | 2,34 |
| Maurolicus muelleri | 0,04 | Small pelagic | 1,89 |
| Gadus morhua | 2,83 | Small cod | 4,68 |
| Melanogrammus aeglefinus | 33,92 | Small haddock | 35,77 |
| Trisopterus esmarkii | 1,31 | Small gadoids | 3,16 |
| Leptoclinus maculatus | 0,18 | Other small demersal fish | 2,03 |
| Hippoglossoides platessoides | 3,35 | Other small flatfish | 5,20 |
| Indeterminatus | 1,00 | ||
| 100,03 | 100,00 | ||
| Porsangerfjorden høsten 2018 | |||
| torsk < 35 cm | Vekt % | Ecopath-gruppe | Endelig fordeling i Ecopath (Vekt %) |
| Polychaeta | 0,09 | Predatory polychaetes | 0,05 |
| Detrivorous polychaetes | 0,05 | ||
| Yoldiella sp. | 0,01 | Large bivalves | 0,01 |
| Crustacea | 1,42 | Small benthic crustaceans | 1,42 |
| Amphipoda | 0,06 | Small benthic crustaceans | 0,06 |
| Gammaridea | 0,21 | Small benthic crustaceans | 0,21 |
| Acanthonotozomatidae | 0,07 | Small benthic crustaceans | 0,07 |
| Hyperiidae | 0,01 | Small benthic crustaceans | 0,01 |
| Euphausiidae | 7,91 | Large krill | 3,96 |
| Small krill | 3,96 | ||
| Caridea | 2,58 | Pandalus and pasiphae | 2,58 |
| Pandalus borealis | 40,83 | Pandalus and pasiphae | 41,54 |
| Munida sarsi | 0,10 | Other large crustaceans | 0,10 |
| Teleostei | 20,74 | ||
| Clupea harengus | 6,43 | Herring | 14,05 |
| Mallotus villosus | 10,55 | Small pelagic | 18,17 |
| Gadus morhua | 6,19 | Cod | 13,81 |
| Indeterminatus | 2,82 | ||
| 100,02 | 100,02 |
| Porsangerfjorden høsten 2018 | |||
| torsk ≥ 35 cm | Vekt % | Ecopath-gruppe | Endelig fordeling i Ecopath (Vekt %) |
| Hydrozoa | 0,01 | Large epibenthic suspension feeders | 0,01 |
| Actinaria | 0,35 | Large epibenthic suspension feeders | 0,35 |
| Hormathia digitata | 3,10 | Large epibenthic suspension feeders | 3,10 |
| Polychaeta | 0,23 | Predatory polychaetes | 0,12 |
| Detrivorous polychaetes | 0,12 | ||
| Pectinariidae | 0,01 | Detrivorous polychaetes | 0,01 |
| Astarte sp. | 0,08 | Large bivalves | 0,08 |
| Cardium sp. | 0,03 | Large bivalves | 0,03 |
| Cephalopoda | 0,01 | Other large predatory benthic invertebrates | 0,01 |
| Crustacea | 1,07 | Small benthic crustaceans | 1,07 |
| Cumacea | 0,01 | Small benthic crustaceans | 0,01 |
| Lysianassidae | 0,02 | Small benthic crustaceans | 0,02 |
| Hyperiidae | 0,11 | Small benthic crustaceans | 0,11 |
| Euphausiidae | 2,79 | Large krill | 1,40 |
| Small krill | 1,40 | ||
| Caridea | 2,68 | Pandalus and pasiphae | 2,68 |
| Pandalus borealis | 24,72 | Pandalus and pasiphae | 24,72 |
| Sabinea sp. | 0,31 | Crangonid shrimps | 0,31 |
| Sabinea septemcarinata | 0,04 | Crangonid shrimps | 0,04 |
| Paralithodes camtschatica | 0,31 | King carb | 0,31 |
| Lithodidae | 0,74 | Other large crustaceans | 0,74 |
| Munida sp. | 0,10 | Other large crustaceans | 0,10 |
| Munida sarsi | 0,11 | Other large crustaceans | 0,11 |
| Brachyura | 0,04 | Other large crustaceans | 0,04 |
| Ophiuroidea | 0,00 | Other pred, benthic invertebrates | 0,16 |
| Teleostei | 23,67 | ||
| Mallotus villosus | 6,68 | Small pelagic fish | 9,64 |
| Gadus morhua | 3,61 | Small cod | 6,57 |
| Melanogrammus aeglefinus | 22,76 | Small haddock | 25,72 |
| Gadidae | 0,62 | Small gadoids | 3,58 |
| Trisopterus esmarkii | 2,48 | Small gadoids | 5,44 |
| Lycodes vahlii | 1,38 | Small demersal fish | 4,34 |
| Cottidae | 0,93 | Cottidae | 3,89 |
| Hippoglossoides platessoides | 0,84 | Other small flatfish | 3,80 |
| Indeterminatus | 0,16 | ||
| 100,00 | 100,00 |
| Tanafjorden høsten 2018 | |||
| Torsk < 35 cm | Vekt % | Ecopath-gruppe | Endelig fordeling i Ecopath (Vekt %) |
| Polychaeta | 0,28 | Predatory polychaetes | 1,50 |
| Detrivorous polychaetes | 1,50 | ||
| Crustacea | 13,09 | Small benthic crustaceans | 14,45 |
| Hyperiidae | 0,29 | Small benthic crustaceans | 1,65 |
| Euphausiidae | 17,90 | Large krill | 10,31 |
| Small krill | 10,31 | ||
| Paguridae | 2,31 | Other large crustaceans | 3,67 |
| Teleostei | 55,25 | Small demersal fish | 56,61 |
| Indeterminatus | 10,88 | ||
| 100,00 | 100,00 | ||
| Tanafjorden høsten 2018 | |||
| torsk ≥ 35 cm | Vekt % | Ecopath-gruppe | Endelig fordeling i Ecopath (Vekt %) |
| Bivalvia | 0,02 | Large bivalves | 0,04 |
| Crustacea | 0,09 | Small benthic crustaceans | 0,11 |
| Ostracoda | 0,00 | Small benthic crustaceans | 0,02 |
| Hyperiidae | 0,11 | Small benthic crustaceans | 0,13 |
| Euphausiidae | 0,94 | Large krill | 0,49 |
| Small krill | 0,49 | ||
| Caridea | 0,51 | Pandalus and pasiphae | 0,53 |
| Pandalus borealis | 14,46 | Pandalus and pasiphae | 14,48 |
| Brachyura | 0,20 | Other large crustaceans | 0,30 |
| Teleostei | 15,03 | ||
| Gadus morhua | 0,29 | Small cod | 4,05 |
| Melanogrammus aeglefinus | 66,19 | Small haddock | 69,95 |
| Sebastes | 1,21 | Small demersal fish | 4,97 |
| Hippoglossoides platessoides | 0,69 | Small flatfish | 4,45 |
| Indeterminatus | 0,26 | ||
| 100,00 | 100,00 |
| Kvænangen høsten 2018 | |||
| Hyse < 35 cm | Vekt % | Ecopath-gruppe | Endelig fordeling i Ecopath (Vekt %) |
| Actinaria | 0,09 | Large epibenthic suspension feeders | 0,34 |
| Polychaeta | 2,97 | Predatory polychaetes | 1,73 |
| Detrivorous polychaetes | 1,73 | ||
| Nothria sp. | 0,35 | Detrivorous polychaetes | 0,60 |
| Maldanidae | 0,76 | Detrivorous polychaetes | 1,01 |
| Galathowenia oculata | 5,88 | Detrivorous polychaetes | 6,13 |
| Pectinariidae | 9,85 | Detrivorous polychaetes | 10,10 |
| Terebellidae | 0,78 | Detrivorous polychaetes | 1,03 |
| Gastropoda | 0,30 | Predatory gastropoda | 0,55 |
| Scaphander sp. | 0,05 | Predatory gastropoda | 0,30 |
| Bivalvia | 4,14 | Large bivalves | 4,39 |
| Yoldiella sp. | 0,77 | Large bivalves | 1,02 |
| Pectinidae | 0,46 | Large bivalves | 0,71 |
| Chlamys sp. | 0,08 | Large bivalves | 0,33 |
| Chlamys septemradiata | 0,02 | Large bivalves | 0,27 |
| Delectopecten vitreus | 0,09 | Large bivalves | 0,34 |
| Astarte sp. | 0,03 | Large bivalves | 0,28 |
| Cardium sp. | 1,35 | Large bivalves | 1,60 |
| Cuspidaria sp. | 0,01 | Large bivalves | 0,26 |
| Arctinula greenlandica | 0,71 | Small molluscs | 0,96 |
| Bathyarca sp. | 0,06 | Small molluscs | 0,31 |
| Dacrydium vitreum | 0,02 | Small molluscs | 0,27 |
| Scaphopoda | 0,64 | Small molluscs | 0,89 |
| Crustacea | 6,10 | Small benthic crustaceans | 6,35 |
| Calanus finmarchicus | 1,65 | Small benthic crustaceans | 1,90 |
| Cumacea | 0,11 | Small benthic crustaceans | 0,36 |
| Amphipoda | 0,02 | Small benthic crustaceans | 0,27 |
| Gammaridea | 0,83 | Small benthic crustaceans | 1,08 |
| Hyperiidae | 0,15 | Small benthic crustaceans | 0,40 |
| Euphausiidae | 24,76 | Small krill | 12,63 |
| Large krill | 12,63 | ||
| Spirontocaris liljeborgii | 0,35 | Crangonid shrimps | 0,60 |
| Pandalus borealis | 9,24 | Pandalus and pasiphae | 9,49 |
| Paguridae | 1,06 | Other large crustaceans | 1,31 |
| Munida sp. | 0,23 | Other large crustaceans | 0,48 |
| Brachyura | 0,72 | Other large crustaceans | 0,97 |
| Hyas sp. | 0,25 | Other large crustaceans | 0,50 |
| Asteroidea | 0,00 | Predatory asteroids | 0,25 |
| Ctenodiscus crispatus | 1,36 | Detrivorous echinoderms | 1,61 |
| Ophiuroidea | 2,60 | Other predatory benthic invertebrates | 2,85 |
| Ophiura sp. | 2,09 | Other predatory benthic invertebrates | 2,34 |
| Echinozoa | 0,19 | Detrivorous echinoderms | 0,44 |
| Holothuroidea | 0,00 | Detrivorous echinoderms | 0,25 |
| Teleostei | 7,94 | Small pelagic fish | 8,19 |
| Indeterminatus | 10,95 | ||
| 100,01 | 100,01 |
| Kvænangen høsten 2018 | |||
| Hyser ≥ 35 cm | Vekt % | Ecopath-gruppe | Endelig fordeling i Ecopath (Vekt %) |
| Porifera | 0,02 | Large epibenthic suspension feeders | 0,15 |
| Hydrozoa | 0,03 | Large epibenthic suspension feeders | 0,16 |
| Actinaria | 0,01 | Large epibenthic suspension feeders | 0,14 |
| Hormathia digitata | 0,84 | Large epibenthic suspension feeders | 0,97 |
| Polychaeta | 10,37 | Predatory polychaetes | 5,31 |
| Detrivorous polychaetes | 5,31 | ||
| Maldanidae | 0,41 | Detrivorous polychaetes | 0,54 |
| Oweniidae | 0,05 | Detrivorous polychaetes | 0,18 |
| Galathowenia oculata | 0,02 | Detrivorous polychaetes | 0,15 |
| Pectinariidae | 7,89 | Detrivorous polychaetes | 8,02 |
| Terebellidae | 0,31 | Detrivorous polychaetes | 0,44 |
| Sabellida | 1,12 | Large epibenthic suspension feeders | 1,25 |
| Gastropoda | 0,45 | Predatory gastropoda | 0,58 |
| Scaphander sp. | 0,24 | Predatory gastropoda | 0,37 |
| Clione limacina | 0,14 | Other benthic invertebrates | 0,27 |
| Bivalvia | 1,06 | Large bivalves | 1,19 |
| Yoldiella sp. | 0,05 | Large bivalves | 0,18 |
| Pectinidae | 0,43 | Large bivalves | 0,56 |
| Chlamys sp. | 0,02 | Large bivalves | 0,15 |
| Chlamys septemradiata | 0,30 | Large bivalves | 0,43 |
| Astarte sp. | 0,08 | Large bivalves | 0,21 |
| Cardium sp. | 0,13 | Large bivalves | 0,26 |
| Bathyarca sp. | 0,10 | Small molluscs | 0,23 |
| Musculus sp. | 0,00 | Small molluscs | 0,13 |
| Nucula sp. | 0,06 | Small molluscs | 0,19 |
| Hiatella sp. | 0,00 | Small molluscs | 0,13 |
| Scaphopoda | 0,71 | Small molluscs | 0,84 |
| Crustacea | 0,67 | Small benthic crustaceans | 0,80 |
| Ostracoda | 0,00 | Small benthic crustaceans | 0,13 |
| Cumacea | 0,01 | Small benthic crustaceans | 0,14 |
| Isopoda | 0,00 | Small benthic crustaceans | 0,13 |
| Amphipoda | 0,02 | Small benthic crustaceans | 0,15 |
| Gammaridea | 0,78 | Small benthic crustaceans | 0,91 |
| Ampelisca sp. | 0,01 | Small benthic crustaceans | 0,14 |
| Hyperiidae | 0,03 | Small benthic crustaceans | 0,16 |
| Euphausiidae | 4,15 | Small krill | 2,20 |
| Large krill | 2,20 | ||
| Caridea | 1,31 | Crangonid shrimps | 1,44 |
| Spirontocaris sp. | 0,10 | Crangonid shrimps | 0,23 |
| Pasiphaea sp. | 0,79 | Pandalus and pasiphae | 0,92 |
| Pasiphaea sp. | 0,95 | Pandalus and pasiphae | 1,08 |
| Pandalus borealis | 4,27 | Pandalus and pasiphae | 4,40 |
| Munida sp. | 0,06 | Other large crustaceans | 0,19 |
| Branchiopoda | 0,07 | Other large crustaceans | 0,20 |
| Ctenodiscus crispatus | 40,53 | Detrivorous echinoderms | 40,66 |
| Echinozoa | 0,54 | Detrivorous echinoderms | 0,67 |
| Holothuroidea | 0,82 | Detrivorous echinoderms | 0,95 |
| Ophiuroidea | 1,62 | Other predatory benthic invertebrates | 1,75 |
| Ophiura sp. | 2,68 | Other predatory benthic invertebrates | 2,81 |
| Ophiura sarsi | 0,37 | Other predatory benthic invertebrates | 0,50 |
| Teleostei | 2,04 | ||
| Clupea harengus | 5,63 | Herring | 6,78 |
| Benthosema glaciale | 1,13 | Small pelagic fish | 2,28 |
| Indeterminatus | 6,58 | ||
| 100,00 | 100,00 |
| Porsangerfjorden høsten 2018 | |||
| Hyser < 35 cm | Vekt % | Ecopath-gruppe | Endelig fordeling i Ecopath (Vekt %) |
| Actinaria | 0,01 | Large epibenthic suspension feeders | 0,01 |
| Polychaeta | 20,48 | Predatory polychaetes | 12,82 |
| Detrivorous polychaetes | 12,82 | ||
| Onuphidae | 0,41 | Detrivorous polychaetes | 0,41 |
| Nothria sp. | 0,03 | Detrivorous polychaetes | 0,03 |
| Spiochaetopterus typicus | 0,01 | Detrivorous polychaetes | 0,01 |
| Oweniidae | 0,76 | Detrivorous polychaetes | 0,76 |
| Galathowenia oculata | 2,87 | Detrivorous polychaetes | 2,87 |
| Pectinariidae | 0,1 | Detrivorous polychaetes | 0,1 |
| Terebellidae | 0,5 | Detrivorous polychaetes | 0,5 |
| Sabellidae | 0,33 | Detrivorous polychaetes | 0,33 |
| Sabellida | 2,79 | Detrivorous polychaetes | 2,79 |
| Gastropoda | 0,03 | Predatory gastropoda | 0,03 |
| Scaphander sp. | 0,08 | Predatory gastropoda | 0,08 |
| Clione limacina | 0,01 | Other benthic invertebrates | 0,01 |
| Bivalvia | 2,82 | Large bivalves | 2,82 |
| Nucula tenuis | 0,02 | Small molluscs | 0,02 |
| Yoldiella sp. | 0,53 | Large bivalves | 0,53 |
| Bathyarca sp. | 0,12 | Small molluscs | 0,12 |
| Mytilidae | 0,01 | Large bivalves | 0,01 |
| Mytilus edulis | 0,04 | Large bivalves | 0,04 |
| Dacrydium vitreum | 0,44 | Small molluscs | 0,44 |
| Pectinidae | 0,02 | Large bivalves | 0,02 |
| Chlamys septemradiata | 0,03 | Large bivalves | 0,03 |
| Arctinula greenlandica | 0,15 | Small molluscs | 0,15 |
| Astarte sp. | 0,04 | Large bivalves | 0,04 |
| Cardium sp. | 3,05 | Large bivalves | 3,05 |
| Cuspidaria sp. | 0,02 | Large bivalves | 0,02 |
| Scaphopoda | 0,03 | Large bivalves | 0,03 |
| Crustacea | 1,81 | Small benthic crustaceans | 1,81 |
| Cumacea | 0,07 | Small benthic crustaceans | 0,07 |
| Apseudes sp. | 0,03 | Small benthic crustaceans | 0,03 |
| Isopoda | 0,02 | Small benthic crustaceans | 0,02 |
| Amphipoda | 0,42 | Small benthic crustaceans | 0,42 |
| Gammaridea | 2,13 | Small benthic crustaceans | 2,13 |
| Lysianassidae | 0,03 | Small benthic crustaceans | 0,03 |
| Hyperiidae | 0,18 | Small benthic crustaceans | 0,18 |
| Euphausiidae | 19,34 | Small krill | 9,67 |
| Large krill | 9,67 | ||
| Pandalus borealis | 13,47 | Pandalus and pasiphae | 16,05 |
| Munida sp. | 0,01 | Other large crustaceans | 0,01 |
| Branchiopoda | 0,10 | Other large crustaceans | 0,10 |
| Ophiuroidea | 1,01 | Other predatory benthic invertebrates | 1,01 |
| Ophiura sarsi | 0,01 | Other predatory benthic invertebrates | 0,01 |
| Holothuroidea | 0,07 | Detrivorous echinoderms | 0,07 |
| Teleostei | 3,35 | ||
| Clupea harengus | 7,99 | Herring | 11,41 |
| Mallotus villosus | 0,27 | Small pelagic | 1,11 |
| Maurolicus muelleri | 0,68 | Small pelagic | 1,52 |
| Gadus morhua | 2,96 | Small cod | 3,80 |
| Indeterminatus | 10,32 | ||
| 100,00 | 100,00 |
| Hyser ≥ 35 cm | Vekt % | Ecopath-gruppe | Endelig fordeling i Ecopath (Vekt %) |
| Polychaeta | 16,48 | Predatory polychaetes | 9,33 |
| Detrivorous polychaetes | 9,33 | ||
| Galathowenia oculata | 0,24 | Detrivorous polychaetes | 1,33 |
| Pectinariidae | 0,40 | Detrivorous polychaetes | 1,49 |
| Gastropoda | 2,00 | Predatory gastropoda | 3,09 |
| Clione limacina | 0,92 | Other benthic invertebrates | 2,01 |
| Bivalvia | 4,31 | Large bivalves | 5,40 |
| Yoldiella sp. | 0,84 | Large bivalves | 1,93 |
| Dacrydium vitreum | 0,11 | Small molluscs | 1,20 |
| Cardium sp. | 0,87 | Large bivalves | 1,96 |
| Cuspidaria sp. | 0,02 | Large bivalves | 1,11 |
| Crustacea | 1,98 | Small benthic crustaceans | 3,07 |
| Cumacea | 0,15 | Small benthic crustaceans | 1,24 |
| Isopoda | 0,13 | Small benthic crustaceans | 1,22 |
| Amphipoda | 0,36 | Small benthic crustaceans | 1,45 |
| Gammaridea | 1,59 | Small benthic crustaceans | 2,68 |
| Hyperiidae | 0,18 | Small benthic crustaceans | 1,27 |
| Euphausiidae | 1,10 | Large krill | 1,64 |
| Small krill | 1,64 | ||
| Meganyctiphanes norvegica | 0,45 | Large krill | 1,54 |
| Pandalus borealis | 24,62 | Pandalus and pasiphae | 25,71 |
| Paguridae | 12,86 | Other large crustaceans | 13,95 |
| Ophiuroidea | 2,31 | Other predatory benthic invertebrates | 3,40 |
| Holothuroidea | 0,01 | Other predatory benthic invertebrates | 1,10 |
| Teleostei | 0,87 | Small demersal fish | 1,96 |
| Indeterminatus | 27,20 | ||
| 100,00 | 100,00 |
| Tanafjorden høsten 2018 | |||
| Hyser < 35 cm | Vekt % | Ecopath-gruppe | Endelig fordeling i Ecopath (Vekt %) |
| Annelida | 0,32 | Other benthic invertebrates | 1,40 |
| Polychaeta | 31,38 | Predatory polychaetes | 16,77 |
| Detrivorous polychaetes | 16,77 | ||
| Oweniidae | 2,67 | Detrivorous polychaetes | 3,75 |
| Galathowenia oculata | 5,55 | Detrivorous polychaetes | 6,63 |
| Gastropoda | 0,00 | Predatory gastropoda | 1,08 |
| Scaphander sp. | 0,38 | Predatory gastropoda | 1,46 |
| Bivalvia | 13,39 | Large bivalves | 14,47 |
| Yoldiella sp. | 0,85 | Large bivalves | 1,75 |
| Astarte sp. | 0,04 | Large bivalves | 1,93 |
| Cardium sp. | 4,05 | Large bivalves | 1,11 |
| Cardium minimum | 2,35 | Large bivalves | 1,12 |
| Cuspidaria sp. | 0,02 | Large bivalves | 5,13 |
| Dacrydium vitreum | 0,03 | Small molluscs | 3,43 |
| Nucula tenuis | 0,67 | Small molluscs | 1,10 |
| Scaphopoda | 0,08 | Small molluscs | 1,16 |
| Crustacea | 0,01 | Small benthic crustaceans | 1,09 |
| Calanoida | 0,59 | Small benthic crustaceans | 1,67 |
| Cumacea | 0,01 | Small benthic crustaceans | 1,09 |
| Amphipoda | 0,07 | Small benthic crustaceans | 1,15 |
| Gammaridea | 0,20 | Small benthic crustaceans | 1,28 |
| Hyperiidae | 1,68 | Small benthic crustaceans | 2,76 |
| Euphausiidae | 3,63 | Large krill | 2,89 |
| Small krill | 2,89 | ||
| Crangonidae | 0,55 | Crangonid shrimps | 1,63 |
| Ophiuroidea | 0,10 | Other large crustaceans | 1,18 |
| Holothuroidea | 0,78 | Detrivorous echinoderms | 1,86 |
| Teleostei | 0,40 | Small demersal fish | 1,48 |
| Indeterminatus | 30,20 | ||
| 100,00 | 100,00 |
| Tanafjorden høsten 2018 | |||
| Hyser ≥ 35 cm | Vekt % | Ecopath-gruppe | Endelig fordeling i Ecopath (Vekt %) |
| Polychaeta | 21,60 | Predatory polychaetes | 11,87 |
| Detrivorous polychaetes | 11,87 | ||
| Oweniidae | 8,26 | Detrivorous polychaetes | 9,33 |
| Galathowenia oculata | 0,39 | Detrivorous polychaetes | 1,46 |
| Pectinariidae | 0,38 | Detrivorous polychaetes | 1,45 |
| Ditrupa sp. | 0,02 | Predatory gastropoda | 1,09 |
| Scaphander sp. | 0,46 | Predatory gastropoda | 1,53 |
| Bivalvia | 6,64 | Large bivalves | 7,71 |
| Yoldiella sp. | 0,22 | Large bivalves | 1,29 |
| Pectinidae | 0,05 | Large bivalves | 1,12 |
| Astarte sp. | 0,19 | Large bivalves | 1,26 |
| Cardium sp. | 2,96 | Large bivalves | 4,03 |
| Cardium minimum | 0,20 | Large bivalves | 1,27 |
| Dacrydium vitreum | 0,01 | Small molluscs | 1,08 |
| Nucula tenuis | 1,46 | Small molluscs | 2,53 |
| Macoma sp. | 6,05 | Small molluscs | 7,12 |
| Scaphopoda | 0,01 | Small molluscs | 1,08 |
| Crustacea | 1,06 | Small benthic crustaceans | 2,13 |
| Ostracoda | 0,00 | Small benthic crustaceans | 1,07 |
| Calanoida | 0,06 | Small benthic crustaceans | 1,13 |
| Cumacea | 0,00 | Small benthic crustaceans | 1,07 |
| Amphipoda | 0,00 | Small benthic crustaceans | 1,07 |
| Gammaridea | 0,26 | Small benthic crustaceans | 1,33 |
| Hyperiidae | 0,83 | Small benthic crustaceans | 1,90 |
| Euphausiidae | 3,42 | Large krill | 2,78 |
| Small krill | 2,78 | ||
| Pandalus borealis | 14,04 | Crangonid shrimps | 15,11 |
| Ophiuroidea | 0,08 | Other large crustaceans | 1,15 |
| Holothuroidea | 0,23 | Detrivorous echinoderms | 1,30 |
| Indeterminatus | 31,12 | ||
| 100,00 | 100,00 |
| Kvænangen høsten 2018 | |||
| Vanlig uer | Vekt % | Ecopath-gruppe | Endelig fordeling i Ecopath (Vekt %) |
| Crustacea | 22,10 | Small benthic crustaceans | 22,10 |
| Hyperiidae | 34,50 | Small benthic crustaceans | 34,50 |
| Euphausiidae | 14,46 | Large krill | 7,23 |
| Small krill | 7,23 | ||
| Caridea | 28,94 | Pandalus and pasiphae | 28,94 |
| 100,00 | 100,00 | ||
| Porsangerfjorden høsten 2018 | |||
| Vanlig uer | Vekt % | Ecopath-gruppe | Endelig fordeling i Ecopath (Vekt %) |
| Euphausiidae | 2,62 | Small krill | 1,31 |
| Large krill | 1,31 | ||
| Pandalus borealis | 92,62 | Pandalus and pasiphae | 92,62 |
| Teleostei | 4,77 | Small demersal fish | 4,77 |
| 100,01 | 100,01 | ||
| Tanafjorden høsten 2018 | |||
| Vanlig uer | Vekt % | Ecopath-gruppe | Endelig fordeling i Ecopath (Vekt %) |
| Crustacea | 20,71 | Small benthic crustaceans | 22,77 |
| Amphipoda | 1,83 | Small benthic crustaceans | 3,89 |
| Hyperiidae | 3,18 | Small benthic crustaceans | 5,24 |
| Euphausiidae | 63,98 | Large krill | 34,05 |
| Small krill | 34,05 | ||
| Indeterminatus | 10,30 | ||
| 100,00 | 100,00 | ||