This report was prepared by the Institute of Marine Research, Norway, for the Norwegian Environment Agency, as part of Norway´s contribution to OSPAR. The report summarizes the latest knowledge on species and habitats associated with seamounts in the OSPAR Regions I, IV and V. Knowledge was sought from published literature, reports and online marine data archives.
The global bathymetry model of Harris et al. (2014) predicts that 161 seamounts occurr within the OSPAR maritime area. Not all of these have been charted or studied, and the literature and bathymetry database review in this report resulted in a much shorter list of 100 seamounts or seamount-like features within the deep seas of OSPAR, i.e. regions I, IV and V. Published literature from the OSPAR area documented that there is knowledge of planktonic organisms for 11 seamounts, information on benthic species for 24 seamounts, and of fish from 16 seamounts. The best described component is the benthos with a total of 49 peer-reviewed papers.
The global knowledge of seamounts (and a few studies from the NE Atlantic) suggests that seamounts are inhabited by species from the regional species pool within the biogeographical zones they occur. At individual seamounts the structure of species assemblages and production patterns are variable over time and modified by factors such as the local and regional hydrography and circulation which are sometimes modified by the seamounts themselves. Other significant factors causing variability are the varying depths of slopes and summits in relation to the depth of the euphotic zone, the depth of summits relative to mesopelagic scattering layers, and presumably the distance from continents, islands and wider areas such banks and ridges. Furthermore, seamounts represent isolated shallows in the deep-sea and may have several important local and regional functions. However, within the OSPAR maritime area, few studies have produced more than descriptive data, hence a major shortage is the lack of quantitative information on species occurrences as well as studies measuring processes and documenting functions. The roles of seamounts at regional scales, e.g. as stepping stones for species across wider ocean areas, have only been incompletely studied.
Seamounts apparently constitute patches of suitable habitats for aggregating commercially valuable fish species that are relatively easy to locate and target, and if not properly controlled, to overexploit. This is known from the OSPAR region V, e.g. from historical depletion of orange roughy west of the British Isles and sharp declines of alfonsino aggregations on seamounts north of the Azores. Also, seamounts are features likely to have Vulnerable Marine Ecosystems (VMEs) (sensu FAO, 2009), primarily in the form of structure-forming coral and sponge aggregations. These require special protective action such as called for by the UN General Assembly and OSPAR, and accordingly several nations, the EU and the Northeast Atlantic Fisheries Commission (NEAFC) implemented measures to prevent further signficant adverse impacts from bottom fishing. Studies at many seamounts in OSPAR have shown that many summits have rich occurrence of VME indicator taxa and probably VMEs. Significant adverse impacts of past bottom fishing have been well documented in some slope and shelf habitats, but studies on seamounts have been scattered, and as yet there is not enough information to assess the overall status of VMEs on seamounts in Region I, IV and V.
1 - Introduction
Seamounts are mountains in the sea that can rise thousands of meters from their base at the surrounding seafloor. Most seamounts are old volcanoes generated near the mid-ocean spreading ridges, in areas with upwelling mantle plumes, or in subduction zones (i.e. arc trench systems) (Staudigel and Clague 2010). Seamounts are most commonly defined as isolated geological features of conical form, that rise at least 1000 m from the surrounding sea-floor without protruding the surface of the sea (Menard 1964, International Hydrographic Organization 2008). Seamounts can be higher than 1000 m, often up to 3000-4 00 m when rising from abyssal depths.
Seamounts impact oceanic circulation at local and sometimes regional scales and generate enhanced hydrodynamic activity. The distinct physical flow-features generated by the seamounts can have a large impact on the biological life around the seamount. The doming of density layers above the seamounts, sometimes resulting in so-called Taylor cones, can bring nutrient rich deep-water into the euphotic zone enhancing photosynthetic productivity at shallow summits and slopes. Increased vertical mixing due to amplified tidal motion, acceleration of oceanic flow around the seamount, and creation of internal waves can enhance the availability of seston and particulate organic matter and thereby nourish communities of sessile suspension feeders in the deeper water layers (White et al. 2007). Seamounts are therefore often characterized by high biomass aggregations of corals, sponges and crinoids (McClain et al. 2010, Etnoyer 2010, Schlacher et al. 2014). These assemblages create a habitat for other invertebrates, micro-organisms and fish and are generally regarded as hot spots for deep-sea biodiversity. If the summits reach into the epipelagic or mesopelagic zones, vertically migrating surface-feeding zooplankton and micronekton impinging on the summits at daytime create favorable feeding conditions for many suspension feeders as well as aggregating benthic and benthopelagic fish species.
Seamounts are distributed in all the world´s oceans. Global estimates of seamount numbers range between 10 000 and 30 000 (Wessel 2001, Yesson et al. 2011, Harris et al. 2014) depending on what data types have been used for the assessment (global bathymetry vs. satellite altimetry) and the mathematical model used to distinguish the seamounts from other elevations on the sea floor (geometry, overall height, height to width ratio etc.). The Pacific Ocean has most seamounts, by far. For example, the modelling effort of Harris et al. (2014) indicate the presence of roughly 1700 seamounts in the Atlantic, compared with 6900 in the Pacific.
2 - OSPAR maritime area
2.1 - Geography, oceanography & biology
In 1992 the OSPAR Convention for the protection of the environment of the Northeast Atlantic was adopted, and since then the OSPAR Commission has been mandated to promote pertinent agreements and actions on behalf of the fifteen governments (Belgium, Denmark, Finland, France, Germany, Iceland, Ireland, Luxembourg, the Netherlands, Norway, Portugal, Spain, Sweden, Switzerland and United Kingdom) and the European Union. The OSPAR maritime area comprises approximately 13 500 000 km2 of the Atlantic Ocean and adjacent marginal seas and coastal zones with the boundaries defined by the OSPAR convention text (https://www.ospar.org/site/assets/files/1290/ospar_convention_e_updated_text_in_2007_no_revs.pdf). The maritime area is split into 5 regions (Figure 1). Below a brief characterization is provided of the geography, oceanography, biology and human activities in Regions I- Arctic Waters, IV- Bay of Biscay and Iberian Coast, and V- Wider Atlantic. These are the three OSPAR regions that have seamounts.
Region I, Arctic Waters, is the most northerly of the OSPAR regions. It covers roughly 5 530 000 km2 and constitutes about 40% of the OSPARs maritime area. The region is characterized by very large seasonal differences in light and temperature and by its northern areas being completely ice covered. The inflow of relatively saline Atlantic Water and, to a lesser extent, the Norwegian Coastal Current bring warm surface water northwards through the eastern parts of the Norwegian Sea into the Barents Sea and along the west coast of Svalbard. Sinking of cold saline water occurs at high latitudes and in the Greenland Sea. Along the Greenland east coast cold and dense water flows southwards at depth, eventually overflowing to the North Atlantic proper across the Scotland to Greenland ridges and contributing to global deepwater formation. Below 600-1000 m, the deeper basins of the Norwegian and Greenland Seas have Norwegian Sea Deep-water with permanent sub-zero temperature (-0.9oC), and low salinity. In eastern areas with Atlantic inflow at the surface, there is a permanent strong vertical stratification and arctic conditions in the deeper parts. Prominent geomorphological features of the region are the wide continental shelves and slopes, the deep basins with abyssal depths, and the major extension of the mid-ocean ridge system from Iceland into the Arctic Ocean. The biological communities are rich and diverse in the epipelagic zone and shelf waters. The Barents Sea, as the major marginal sea of this region, supports the most productive fisheries in the North Atlantic. In addition to fishing, petroleum and gas exploration and production are significant activities in the Norwegian and Barents Sea. Arctic tourism and marine transport are growing industries in the region.
Region IV, Bay of Biscay and Iberian Coast, constitutes a rather small region, and comprises roughly 540 000 km2 of ocean area along the French, Spanish and Portuguese continental shelves, slope and partly abyssal plain. The circulation along the northern Iberian Peninsula and the Bay of Biscay is mainly driven by winds and shows a high seasonality, with interchanging upwelling and downwelling periods. Mean circulation is week compared to the rest of the North Atlantic basin. During summer northerly trade-winds generate southerly flowing surface water along the Iberian Peninsula and up-welling in the top 200-300 m. From 400 to 1300 m the Mediterranean out flow water, moving toward west, dominates. Deeper than this the North Atlantic Deep Water is found which is characterized by very slow flows. The region is characterized by a highly diversified bottom topography and supports complex benthic ecosystems and a large number of fish species. The region supports a rich fishing industry in addition to maritime transport and tourism.
Region V, Wider Atlantic,comprises nearly 50% of the OSPAR maritime area. The area is 6 350 000 km2 encompassing the deep waters of the southern parts of the NE Atlantic. Most of the region is deep ocean extending across the abyssal plain and the mid-Atlantic ridge. The habitats span from the highly homogenous abyssal plains to the geomorpologically more complex mid-Atlantic ridge with its slopes, central valley and multiple fracture zones, as well as the continental rise, slope and bank areas of the European continent. Seamounts in the region are associated with abyssal, ridge and continental slope areas. A major oceanographic feature of Region V is the northeastward flowing East Atlantic Drift originating off of North America as an extension of the Gulf Stream. A branch of this major current flows northwards in to the Nordic Seas. Circulation and watermasses are well described for Region V, including some important frontal features that are significant for the regional biogeography. A major oceanic one is the Sub-polar Front dividing the area into a northern area dominated by cool-temperate waters and southern area dominated by warm-temperate waters. The region supports both inshore and offshore fisheries mostly associated with islands, seamounts and continental banks, and has also other significant maritime industries such as tourism and transport.
Figure 1. Map showing the extent of the OSPAR maritime area that comprises a total of 5 regions (Region I, II, III, IV and V), of which Regions I, IV and V have significant deep-sea areas. The orange and blue patches represent the Atlantic and Arctic deep-sea biogeographic zones after Dinter (2001), and blue-grey dots and small patches are seamounts, as predicted by Harris et al. (2014).
2.2 - Biogeographic provinces
The OSPAR biogeographic classification (Dinter, 2001) divides the OSPAR maritime area into zones supporting characteristic fauna and reflecting depth, geomorphological and oceanographic conditions (Figures 1 and 4, Table 1). Briefly, the classification first divides the area into the benthal and neritopelagical areas less than 1000 m deep, and the deeper areas that are more than 1000 m deep. These two regions roughly represent the continental shelves and upper slopes, and the deep-sea, respectively. Each of these two are further split into an Arctic and an Atlantic subregion.
Within the continental shelf and slope region, 6 provinces are recognised in the Arctic subregion and 11 provinces in the Atlantic subregion. Within the deep-sea there is merely a split into an Arctic and an Atlantic sub-region which encompasses only the North Atlantic province in the Atlantic sub-region. The Dinter (2001) classification may not fully capture the full range of biogeographic features of the deep-sea. This is significant when making judgements on the biogeographical affinity of faunas associated with seamounts that are oceanic and mostly deep features. Few biogeographic accounts focused specifically on the biogeography of seamounts in the North Atlantic, but an exception is the collection of papers in Mironov et al. (2006).
Table 1. Biogeographic provinces of the OSPAR maritime area (Dinter 2001).
3 - Definition and classification of seamounts
In the literature a range of definitions and classifications of seamounts have been presented, each satisfying different scientific disciplines. Staudigel and Clague (2010) presented a broad definition of seamounts as isolated geological features on the sea floor and classified these into six evolutionary stages that are structurally very distinct based on age and size. In their effort to model the global distribution of seamounts Yesson et al. (2011) used the criteria of seamounts being isolated geological features and made the distinction between seamounts and knolls in that isolated topographic features being 1000 m or higher from the surrounding seafloor are seamounts and features between 100 and 1000 m are knolls. Morato et al. (2013) in their later effort to model NE Atlantic seamounts used the same height distinction but defined them small and large seamounts. They argued that there is no biological reason to support the traditional size limit of 1000 m for a seamount (Pitcher et al. 2007, Wessel 2007) and small topographic features may be functionally equally important in the deep-sea as larger ones (Koslow et al. 2001). For this report the definition of seamounts will be that of IHO (2008) where seamounts are defined as a distinct, generally equidimensional, elevation greater than 1000 m above the surrounding relief as measured from the deepest isobath that surrounds the feature and distinguished from other characteristic elevations of the seafloor, such as knolls, guyots, mounds and banks based on height, shape, origin, location, and isolation (see Table 2). This definition is still the most widely used definition of seamounts.
Table 2. Classification of elevated features of the seafloor from the digital gazetteer of names and geographic position of generic features of the seafloor, www.gebco.com.
In this report, seamount will thus be used in its geological sense and not as ecological unit. We use the work of Harris et al. (2014) to present modelled, but hithero unchartered seamounts. The modelled seamounts (Harris et al. 2014) are taken from a new digital, global seafloor geomorphic features map (GSFM) that has been created using a combination of manual and ArcGIS methods based on the analysis and interpretation of a modified version of the SRTM30_PLUS global bathymetry grid. In that account seamounts are defined as isolated features with conical form (length:width ratio <2) extending at least 1000 m from the surrounding seafloor. The modelled seamounts are further classified based on their physical characteristics i.e. 1) basal area, 2) the height of the seamount, 3) peak depth, 4) proximity, i.e. the distance to the nearest seamount or the shelf break based on geodesic distance and 5) percent escarpment, i.e. the proportion of each seamount feature with slope greater than 5 degrees, yielding a total of 11 different seamount morphotypes (Macmillan-Lawlerand & Harris 2016). These are further divided into five broad groups including: 1) Very large and tall seamounts with low escarpment - morphotype 6, 2) Large and tall seamounts with shallow peak - morphotype 9 and 10, 3) Intermediate seamounts - morphotype 3, 5 and 11, 4) Small seamounts with deep peaks - morphotype 1, 2 and 3, and 5) Small and short seamounts with very deep peaks - morphotype 7 and 8.
4 - Distributions of seamounts in the OSPAR regions
Exploration of a range of sources led us to consider three different (but complementary) classes of seamounts reflecting the types, quantity and quality of data available: charted seamounts, studied seamounts, and modelled seamounts. Below, the occurrence of seamount within each OSPAR region will be presented for all three classes individually, after which a comparison of the three will be made to gain a more comprehensive knowledge on the occurrence and distribution of seamounts within OSPAR maritime area.
4.1 - Charted seamounts
Information on charted seamounts was collected from the digital gazetteer of names and geographic position of generic features of the seafloor (IHO_IOC GEBCO Gazetteer of Undersea Feature Names) managed by the sub-committee Undersea Feature Names (SCUFN) of GEBCO (General Bathymetric Chart of the Oceans) (www.gebco.net). Scrutinization of this database identified 71 seamount features within the OSPAR Region I, IV and V (see Figure 2 and associated table). In total, 22 seamounts are registered in Region I, with the majority being associated with the mid-Atlantic ridge systems Knipovich, Molloy and Gakkel. Three seamounts are registered associated with ridges and valleys of the continental slope in Region IV. 46 seamounts are registered in Region V. These are mainly associated with the mid-Atlantic ridge and occur in very high numbers around the Azores archipelago.
4.2 - Studied seamounts
Information on studied seamounts was harvested from the database of literature prepared for this report (see methods sections for the different taxonomic groups in each chapter). Information on planktonic organisms was found for 11 seamount or seamount-like features. Of these, one lies in Region I, four lie in Region IV, and six in Region V. Information on the benthic community composition was found for 24 seamounts, i.e. three from Region I, two from Region IV and 19 from Region V. Fish communities have been studied on 16 seamounts or seamount-like features. Of these, one lies in Region I, two in Region IV and thirteen in Region V (Table 4).
Overall, the benthic fauna seemed to be the most well studied component of the seamount biota with a total of 49 references in our literature database. Roughly half as many references, i.e. 26, were found for studies of fish. Investigations to study plankton communities were more infrequent. The data above are indicative of the emphasis in past studies, but we recognise that our data base could lack relevant historical records. The extensive historical record of biogeographers pioneer studies in the 20th and 19th century, e.g. as illustrated by Mironov et al. (2006), has not re-explored here.
Table 3. Seamounts and seamount-like features of the OSPAR Region I, IV and V that have been subjected to biological investigations yielding information of plankton communities, benthic fauna, fish or marine mammals. Note that this list is conservative in the sense that shallow features that would not satisfy the stringent definition of seamounts are included, e.g. banks such as Hatton, Faroe and Galicia. *For marine mammals, presence is geographic only, i.e. irrespective of whether species observed are capable of diving to depths that suggest direct interaction with the seamount ecosystem (see specific section on marine mammal diving depths below).
4.3 - Modelled seamounts
The distribution of modelled seamounts (Figures 1 & 2) was extracted from the work of Harris et al. (2014) who used the SRTM30 PLUS global bathymetry grid (Becker et al. 2009) supplemented with the EMODnet (2013) data to create geomorphic feature maps of the world´s oceans. The work identified a total of 161 seamounts within the OSPAR maritime area. Within Region I (Arctic Waters) 16 seamounts covering a total area of 6 253 km2 were identified. Three of the seamounts lie within the North Atlantic deep-sea biogeographical province while the remaining 13 seamounts lie in the Arctic deep-sea subregion. Of the 16 seamounts situated in Region I, 61% belong to the group of small seamounts with deep peaks, while 39% belong to the group of intermediate seamounts.
Within Region IV, Bay of Biscay and Iberian Coast, the modelling effort of Harris et al. (2014) identified two seamounts covering a total area of 745 km2 located within the same biogeographical province (i.e. the North Atlantic Deep-sea province). Both seamounts belong to the group of small and short seamounts with very deep peaks.
The bulk of OSPAR seamounts occur in Region V ‘Wider Atlantic’ and 143 were detected by Harris et al. (2014). This is 91% of all the seamounts of the OSPAR maritime area. Within this region, seamounts cover an area of 89 820 km2. This is 1.4% of the total area of the region. All seamounts occur within the North Atlantic Deep-sea province. Of the 143 seamounts in Region V 53% belong to the group of small seamounts with deep peaks, 35% belong the the intermediate seamounts, 10% belong to the group of small and short seamounts with very deep peaks and only 2% belong to the large seamounts with shallow peaks. Most of the seamounts in Region V are associated with the mid-Atlantic Ridge and the Azores archipelago, but there are others that are off-ridge and more isolated features, e.g. the Milne complex, and Altair and Antialtair. Others occur very close to islands or the continental slopes, e.g. the seamounts west of Scotland.
A minority of the modelled seamounts (i.e. 27%) lie within the Exclusive Economic Zones (EEZs) of the OSPAR member counties. Five seamounts are found within Norwegian EEZ, four in the Danish EEZ, three in the Spanish EEZ and 48 in the Portuguese EEZ. However, a majority of the seamounts (i.e. 66%) lie on the extended continental shelf (ECS) claim areas of OSPAR member counties (Figure 3), with e.g. 51 seamounts within the Portuguese ECS, 7 in the Icelandic ECS, and 5 in the Norwegian ECS.
4.4 - Chartered, studied and modelled seamounts
Our literature search (studied seamounts) and the GEBCO database (charted seamounts) identified 100 unique seamount like features within OSPAR regions I, IV and V for which there is at least some geophysical or biological data (Figure 2). Information of the benthic communities is available for 49% of these seamounts. However, 29% of the seamounts for which there is information on the composition of the benthic communities do not have an official name accepted by GEBCO Gazetteer of Undersea Feature Names.
Furthermore, 24% of the seamounts that are referred to as seamounts in the literature are in the Gazetteer of Undersea Feature Names referred to as banks, 4% are referred to as knolls, and 2% are referred to as hills. Region I have 21 of these 100 seamount-like features, while region IV have 6. The bulk of the charted and studied seamounts are thus found in Region V (Wider Atlantic), i.e. 73 features.
A comparison of the seamounts predicted by Harris et al. (2014) and seamounts listed in the IHO IOC GEBCO Gazetteer of Undersea Feature Names demonstrates a big discrepancy in the polar areas of Region I, i.e. north of 78oN (Figure 2). The model of Harris et al. (2014) does not register seamounts in the Arctic region while the Gazetteer has 15 seamounts including Eistla, Atla, Gjalp, Koldewey, Danilcuck, Korotaev, Johannsen, Cagni, Robert Perry, Aref´yev, Bukmeyer, Agafonov, Zheglov, Afanasenkov, Pyle and Vladimirov seamounts. There is a large cluster of modelled seamounts on the Ægir ridge in the Norwegian Sea, as well as a smaller cluster of seamounts off of the SE coast of Greenland that are not chartered nor been subjected to targeted studies.
The IHO IOC GEBCO Gazetteer of Undersea Feature Names lists 3 seamounts in Region IV. Among these is the Vigo Seamount which is not among the mounds predicted by Harris et al. (2014). In addition, our search of published literature identified 3 features that were cited to be seamounts but not detected in the modelling effort of Harris et al. (2014) nor categorized as seamounts in the IHO IOC GEBCO Gazetteer. These are Porto Hill, Le Danois Bank and Gascone Knoll.
The overlay of modelled and studied/charted seamounts (Figure 2) revealed that only a small proportion of the modelled seamounts in the OSPAR maritime areas have been the subject of scientific investigations of any kind, either biological, geographical or oceanographic. This may partly be explained by a poor precision in the geographical positioning of the seamounts which may have yielded deviating recording from one study to the next. Many seamounts were first detected using single beam echo-sounders prior to the introduction of today’s precise GPS capability on research vessels. Since then multi-beam sounders have been introduced and the precision of the geographical positioning is greatly enhanced. Furthermore, many seamount have not been charted by ships but are only mapped from remote sensing data (sea surface altimetry). There is a degree of uncertainty associated with satellite altimetry data as well as with the necessary spatial interpolation included in the models used to delineate and distinguish seamounts. This adds uncertainty to the positioning, shapes and summit sizes of different features (Wessel et al. 2010). An important explanation for the relative sparsity of seamount data from targeted research studies is largely due to the high costs involved in running cruises to their often remote locations.
Figure 2. Locations of seamounts in OSPAR Region I (Arctic Waters), IV (Bay of Biscay and Iberian Coast) and V (Wider Atlantic), modelled by Harris et al. (2014) (red shapes) and occurring in our list of charted and/or studied seamounts (circle and a number). Orange and blue patches represent the Atlantic and Arctic deep-sea biogeographic zones after Dinter (2001). For the names of the seamounts see the text table 4 on the next page.
Figure 3. Occurrence of modelled seamounts (from Harris et al. 2014) in the exclusive economic zones and extended continental shelf claim areas (ECS) of OSPAR member counties. EEZ and ECS boundaries are the outer limit lines and points displayed on the UNEP Shelf Programmes data inventory map and were downloaded from http://www.continentalshelf.org/ecs-shapefiles2.aspx?src=http://tuvalu.grida.no/cgi-bin/ecs.pl?subm=prt_44_2009;&type=line
Figure 4. Locations of seamounts (red shape) in OSPAR Regions I (Arctic Waters), IV (Bay of Biscay and Iberian Coast) and V (Wider Atlantic), predicted by Harris et al. (2014) and the biogeographic regions of the area (after Dinter 2001).
5 - The diversity of planktonic organisms near seamounts
In this chapter published literature on planktonic organisms from seamounts in the OSPAR Region I, IV and V has been reviewed to identify planktonic species observed at and near seamounts. Most studies have reported occurrence, densities or distributions of phytoplankton and zooplankton taxa, but also other types of information such as biomass, production and chlorophyll a concentration has been recorded. We have emphasized reviewing relevant literature and studies with dedicated plankton sampling but note that the taxa lists are not necessarily exhaustive.
5.1 - Data collection
The literature search was mainly based on standardized searches in the scientific database Web of Science. As a first step to gather relevant information about planktonic organisms, all seamounts listed in the table accompanying Figure 2 were examined using the search terms: 1) the name of the seamount, plankton, and seamount, 2) The name of the seamount, and plankton, 3) the name of the seamount, and seamount. Secondly, seamounts were googled to search for web pages and papers that could provide relevant data and additional references. Thirdly, reference lists have been examined to reveal additional relevant papers. Peer-reviewed articles have been emphasized, but to some extent ‘gray literature’ and web pages have also been examined.
5.2 - Species
The general conclusion is that very few dedicated planton studies have been made at OSPAR seamounts, hence little information is available. Exceptions are Condor the Terra Seamount, Joao de Castro Bank, Sedlo Seamount, and Le Danois Bank, where some dedicated plankton sampling has been carried out. However, in different seamounts the sampling has often focused on different components of the planktonic community. For example, decapods (macro-zooplankton) were studied in detail at Le Danois Bank, the phytoplankton community was best studied at Condor the Terra seamount and at Gorringe Bank (with its two summits Gettysburg and Ormonde), whereas a detailed taxa list of fish larvae is available from Joao de Castro Bank. Therefore, taxa lists will not necessarily be comparable between seamounts and assessments of differences in diversity between seamounts becomes impossible.
The most well-studied seamounts regarding planktonic communities are described below. In Table 5 information from all named seamounts in OSPAR region I, IV, and V is summarized, and whenever taxa lists were too extensive to include here, those have been listed in separate appendix tables.
Condor the Terra Seamount (Region V) located very close to the islands Fiala and Pico in the Azores, is one of the best studied seamounts regarding planktonic organisms. Zooplankton composition and taxonomical diversity were described based on several surveys. In total, 19 phyla were recorded (Carmo et al. 2013, Colaço at al. 2013), and the Condor hosts at least 147 taxa and developmental stages (Appendix Table 1). Crustacea were the most abundant (73%), mainly represented by Copepoda (61%). Other important taxa were Urochordata (17%), Protozoa (5%) and Mollusca (4%). Copepoda, especially Calanoida, was the most diverse taxon. Plankton sampling at Condor has also been carried out in relation to food web structures (Colaço at al. 2013), with recording and analysis of 7 species/taxa: various scyphozoans, ctenophores, copepods, euphausids, decapod larvae, chaetognaths and pyrosomes. The phytoplankton community was described based on a number of surveys, and the seamount hosts at least 106 taxa (Appendix Table 2), observed by Santos et al. (2013). In addition, chlorophyll a (presumed to reflect phytoplankton biomass) has been measured.
At Joao de Castro Bank (Region V) the zooplankton, including fish larvae, was studied (Cardigos et al. 2005; Sobrinho-Gonçalves and Cardigos, 2006). The invertebrate zooplankton was dominated by copepods (61 %), followed by siphonophores (21.6 %), salps (7.4 %), chaetognaths (4.0 %), decapods (3.7 %), and minor components of mollusca, euphausids, amphipods and polychaetes (all < 0.6 %). Concerning fish larvae, 35 taxa belonging to seven families were present at Joao de Castro (Appendix Table 3). The community was largely dominated (89% of the samples) by Myctophidae, mainly Ceratoscopelus maderensis, and Gonostomatidae. Mesopelagic and bathypelagic taxa dominated completely, while neritic taxa were rare (< 0.1 %).
From the Sedlo Seamount (Region V) information about zooplankton taxa, biomass in different size classes, metabolism, stable isotopes and fatty acids analysis, and chlorophyll a concentration is available (Hirsch et al. 2009a, Hirsch et al. 2009b, Kiriakoulakis et al. 2009, Martin and Christiansen 2009). The zooplankton community consisted of Copepoda (6 taxa), Ostracoda (1 taxon), Euphausiacea (1 taxon), Pteropoda (1 taxon) and Chaetognatha (3 taxa) (Hirsch et al. 2009a).
A cluster of seamounts in the southern/central Norwegian Sea (Region I) were predicted by Harris et al. (2014). No literature was found for these seamounts as they have not been studied previously. However, transects sampled by Continuous Plankton Recorder on merchant ships are operated from Norway to Iceland and provide some data (Reid et al. 2003). Information about the planktonic community in surface water exactly above the group of seamounts were extracted and are presented here. The data comprise primarily mesozooplankton records as macroplankton are not sampled well due to a small sampling volume. The data originate from from 2008 to 2016 and the sampling depth is about 7 m below the surface. In a limited geographical area (-4.1° to -0.7° W, 67.7° to 68.4° N), corresponding to the seamount positions, 12 taxa were observed (Appendix Table 5). The most common taxon was Copepoda, and 6 copepod species were found, in addition to unidentified copepod nauplii. The other taxa recorded were Appendicularia, Chaetognatha, Euphausiacea and Hyperiidea (Amphipoda). In an extended geographical area (-4° to 0° W, 67° to 69° N) around the seamount locations, a total of 25 taxa were registered. In total 12 copepod species were observed, in addition to unidentified copepod egg and nauplii. In addition, Foraminifera, Appendicularia, Thecosomata, Radiolaria, echinoderm larvae, Euphausiacea, Chaetognatha, bivalve larvae, and Hyperiidea occurred. Of these, echinoderm and bivalve larvae may be most related to the seamounts, as they are pelagic stages of benthic organisms.
At Le Danois Bank (region IV) information of macro-zooplankton are available, from studies focusing on decapod crustaceans (Cartes et al. 2007). A total of 56 species was identified (Appendix table 4): Decapoda (47 species), Euphausiacea (4 species), Mysidacea (5 species). The most dominant species was Pagurus alatus, Polycheles typhlops, Parapagurus pilosimanus, Bathynectes maravigna, Anapagurus laevis, Pagurus excavatus, Pontophilus norvegicus, P. spinosus, Sergia robusta, Munida tenuimana, and Geryon trispinosus.
Table 5. Seamounts (and banks) for which plankton information exists and what plankton taxa have been registered.
The mid-Atlantic Ridge(MAR) has been rather extensive studied for assessments of primary production, phytoplankton, zooplankton and micronekton from the full depth range (e.g. Gaard et al. 2008; Stemmann et al. 2008; Hosia et al. 2008, 2017; Youngbluth et al. 2008; Pierrot-Bultz 2008; Letessier et al. 2011). These studies did not focus on specific seamounts and will hence not be reported on in any detail here, but these ridge studies as well as others with focus on oceanic deep-water habitats provide information that may be relevant for future seamount studies.
5.3 - Conclusions
Very few seamounts in the OSPAR Regions I, IV and V have been subjected to extensive and systematic studies on plankton diversity. Information on plankton communities was only found from 11 out of 101 seamounts in the region. The different studies have focused on different taxonomic groups, from mesoplankton and phytoplankton, to macroplankton and fish larvae. In addition, there has been a large variation between studies in sampling methods and taxonomic precision. Due to the lack of comparative studies it is not possible to assess whether seamounts are areas of increased plankton diversity. On the contrary, lower pelagic diversity was found at the Le Danois Bank compared to surroundings, due to the absence of meso-bathypelagic species over the summit (Cartes et al. 2007), and reduced zooplankton biomass was observed at Sedlo seamount and Joao de Castro bank.
The zooplankton species found at individual seamounts are widely distributed taxa, and the species composition at a specific site mainly reflects the zooplankton fauna associated with the relevant biogeographic region in which the seamount is located, as well as the water masses surrounding the seamount. Although there is limited data on plankton communities at or near individual seamounts, extensive information on the diversity and distributions of zooplankton in the OSPAR region exists. This includes full depth data generated at oceanic features such as the mid-Atlantic Ridge and epipelagic data over wide areas of the North Atlantic from the Continuous Plankton Recorder (CPR) Survey (https://www.cprsurvey.org/).
6 - Benthic invertebrates and biotopes
In this chapter published literature on benthic invertebrates from seamounts in the OSPAR Regions I, IV and V has been reviewed. Aims have been to consider to what extent the avialable information suggest that seamounts in these regions function as biodiversity hotspots and as areas particularly suitable for settlement and growth of taxa that may need particular protection from anthropogenic disturbance, such as cold-water corals and sponges.
6.1 - Data collection
To capture as many relevant citations as possible our search included a range of scientific databases, internet engines and reference lists of published papers. As a first step ASFA and Web of Science were scrutinized using the search terms: 1) seamount*, and coral*, and north east Atlantic */NE Atlantic* orth Atlantic, not Mediterranean*, 2) seamount* and porifera*/sponge* and North East Atlantic */NE Atlantic* orth Atlantic, not Mediterranean*, 3) seamount* and VME* and North East Atlantic */NE Atlantic* orth Atlantic, not Mediterranean*, 4) seamount* and stylasterid* and North East Atlantic */NE Atlantic* orth Atlantic, not Mediterranean* and 5) seamount* and benthos*/macro fauna*/fauna* and North East Atlantic */NE Atlantic* orth Atlantic, not Mediterranean*. This search resulted in 155 and 113 citations from Web of Science and ASFA, respectively, published in the years 1945 to 2018 for Web of Science and 1971 to 2018 for ASFA. Thereafter, the internet engines Google and Google scholar, as well as Research Gate, were searched for web pages and papers that could provide relevant data and additional references. The potential relevance of all citations was evaluated and irrelevant citations were excluded (e.g. studies of cold-water ‘coral mounds’ as they represent a different type of feature, studies outside the OSPAR Regions I, IV and V and papers not containing the species information required for the review). Seamounts Online was scrutinized yielding occurrence data on 331 species entries from 12 seamounts from the OSPAR regions. References for these studies were acquired. The remaining list of papers was compared to the latest review of OSPAR seamounts, Kvile (2011) and Morato et al. (2013), for a final check for possibly lacking data. After this step our list included 49 papers for the inclusion in the review with information on benthic species. These originated from 29 unique seamounts or seamount-like features of the OSPAR maritime area (listed in Table 2 Appendix 1).
6.2 - Species
Among benthic invertebrates, the most well studied phylum was the Cnidaria, encompassing e.g. the Alcyonaceans (gorgonians and soft corals), Actiniaria, Cerantharia, Pennatulacea, Antipatharia, Scleractinia (Caryophilliidae) and Stylasterida corals. Overall, the occurrence of coral taxa had been documented from 25 seamounts within the OSPAR maritime area (Table 6). The phylum Porifera was also fairly well studied. Documentation of species/taxa occurrence (mainly in the groups Haxactinellidae and Demospongia) was found from 16 seamounts. The majority of the coral and sponge studies has had a focus on large and conspicuous species. These were studied either by underwater video surveys, ROV sampling or by identifying by-catch from the long-line fishery.
The seamounts and banks from around the Azores Islands, i.e. Gigante, Cavala, Ferradura, Acor, Princesa Alice, Condor de Terra, Voador, Mar de Prata and Formigas, are particularly well studied for coral species (Braga-Henriques et al. 2013). Here, the coral fauna is dominated by Alcyonaceans (57%), Scleractinians (24%), Antipatharians (10%) and Stylasterids (9%). Overall, 164 coral species have been described from these seamounts and the Azores island slopes. Surveys for the sampling of Porifera fauna have been carried out on the Schultz Massif Seamount in Region I (Torkildsen 2013, Roberts et al. 2018), Rosemary Bank Seamount in Region V (McIntyre et al. 2016) and Ormonde seamount in Region IV (Xavier & Van Soest 2007, Cristobo et al. 2015, Ramos et al. 2015). The seamounts and banks west of the British Isles, i.e. Anton Dohrn Seamount, Herbrides Terrace Seamount, George Bligh Bank and Rosemary Bank Seamount have been subjected to a number of surveys to assess the occurrence of epibenthic biotope forming species (Howell et al. 2010, Narayanaswamy et al. 2013, Henry et al. 2014, Henry & Roberts 2014, Davies et al. 2015). These studies have yielded detailed information on the occurrence of Cnidaria, Porifera and Retaria taxa and the biotopes they construct.
One of the most well studied seamounts is the Ormonde Seamount (Region IV) with a total of 5 publications describing the benthos and with registrations of species occurrences from 7 different phyla (Table 4). A total of 59 mega benthic taxa were described from Ramos et al. (2016) where Cnidaria (19 taxa), Porifera (10 taxa) and Echinodermata (9 taxa) were dominant benthic phyla. Ormonde seamount hosts at least 12 coral species/taxa from the groups Anthipatharia, Gorgonia, Stylasterida and Pennatulacea and 8 sponge species from Demospongia (e.g. Cladorhizidae) and Hexactinellidae. Furthermore, the gastropod fauna has been described (Ávila et al. 2003).
Table 6. Summarization of the most commonly studied phyla on the seamounts of the OSPAR regions.
Condor the Terra Seamount (Region V) is also well studied with 6 publications describing benthic species and communities (Braga-Henriques et al. 2013, Colaco et al. 2013, Zeppelli et al. 2013, in addition to Tempera et al. 2011, Braga-Henriques et al. 2011 and Pereira et al. 2011) from 8 different phyla. Dominant benthic phyla at this seamount were Cnidaria (27 taxa), Porifera (27 taxa) and Echinodermata (11 taxa) (Pereira et al. 2011). The porifera fauna is rather poorly described with regards to the taxonomy but much work has been put into taxonomical descriptions of corals. Within the Cnidaria most species occur within the group Alcyonacea (11 taxa) and with Actiniaria, Cerantharia, Pennatulacea, Antipatharia being represented with 1 taxon each. Scleractinia were represented by 2 taxa and Stylasterida with 3 species (Braga-Henriques et al. 2011). Biotopes of particular interest are the Viminella flagellum and Dentomuricea sp. coral gardens in coexistence with the hydrozoan cf. Polyplumaria flabellata and the Pheronema sponge aggregations. Pheronema sponges and Dentomiricea corals colonize both hard and soft substrates while Viminella flagellum and cf. Polyplumaria flabellata hydrozoan were most common on hard substrate (Tempera et al. 2011).
Anton Dohrn Seamount is one of the most well studied seamounts in Region V. The megabenthos of this seamount seems to be dominated by specimens from phyla Cnidaria, Porifera, Echinodermata and Retaria. Among the corals the groups Antipatharia and Scleractinians dominate with 5 genera/taxa each, i.e. Antipathes sp., Leiopathes sp., Sticopathes sp., Paranthipathes sp. Bathypathes sp. for the Antipatharians and Lophelia pertusa, Madrepora oculata, Solenosmilia fragilis, Carophyllidae and Flabellum sp. for the scleractinians. Furthermore Actinaria, Gorgonia, Alcyonacea, Pennatulacea and Bamboo corals occur on the seamount. The Xenophyophore Syringammina fragilissima occurred in characteristic xenophyophore aggregations and the glass sponge Aphrocallistes sp. was a dominant component of sponge aggregations. Overall, 30 morphospecies were described by Howell et al. (2010) while Davies et al. (2015) described 13 biotopes from this seamount.
The different seamount studies reviewed had different aims, from documenting the occurrence of all species within one specific genera of Mollusca at one seamount (Ávila et al. 2003), to describing the large dominating habitat forming epifauna at another seamount (Davies et al. 2015). The tools used to sample fauna also differs greatly, as well as the precision of the taxonomical work. Therefore, the absence of a specific taxonomic group from a seamount does not necessarily mean that the taxa is absent from that seamount. It is equally or even more likely that the absence is due to undersampling and that there has not been targeted effort to sample that specific group of organisms. Performing analyses for distinguishing the suitability of different types of seamount for different benthic communities or taxa is therefore not possible.
6.3 - Biotopes
From the Anton Dohrn Seamount Davies et al. (2015) described 13 different biotopes which included several different types of coral gardens, coral reefs, sponge aggregations or xenophyophore communities. These were found mainly in connection with steep escarpments and small topographic elevations within the seamount. Also, the Rosemary Bank is known to host highly diverse sponge aggregations. At 1200 to 1440 m depth McIntyre et al. (2016) identified both boreal ostur sponge grounds dominated by demosponges from the Geodia genus as well as hexactinellid sponge grounds dominated by Pheronema sponges. Ramos et al. (2016) documented single species and multi species coral gardens with Viminella flagellum on hard bottom, as well as coral gardens composed of Paramuricea clavata and Callogorgia verticillata. Deep-sea sponge aggregations on the Ormonde seamounts occur deeper than 200 m and are composed of Pachastrellidae, Geodiidea, Axinellidae and Polymastidae sponges. In the Hebrides Terrace Solenosmilia coral reefs and xenophyophore aggregations have been described (Cross et al. 2014).
These studies confirm the suitability of seamounts for colonization by filter and suspension feeding coral and sponges. Overall, 10 of the 29 seamounts for which benthic species have been studied are confirmed to host coral gardens and/or sponge aggregations. Moreover, the occurrence of coral taxa has been described from another 16 seamounts within the OSPAR regions with sponges also occurring on 7 of these. Scleractinian, Alcyonacean, Gorgonian, Antipatharian and Stylasterid corals together with some types of sponges are all listed as groups that often contain sensitive and potentially vulnerable species that often contribute to forming VMEs sensu FAO (2009), and these taxa feature amongst the VME indicators recognized by the International Council for the Exploration of the Sea (ICES) to be recorded in the ‘ICES VME database’ (http://www.ices.dk/marine-data/data-portals/Pages/vulnerable-marine-ecosystems.aspx). Our data collation shows that all studied seamounts in the OSPAR regions contains VME indicator species and are rightly regarded as VME elements (FAO 2009). It´s probable that future quantitative studies of more seamounts would substantially increase the number of OSPAR seamounts documented to harbour VMEs. This would generate important input data to facilitate management advice.
Table 7. This table shows where quantitative surveys have been carried out that enable the characterization of biotopes. This table shows where VME indicating biotopes have been described.
It should be noted that the biotopes referred to above are not uniquely associated with seamounts. There is a range of other geomorphological features, such as knolls, banks, hills, ridges, canyons that provide similar environmental conditions and that harbor a similar set of biotopes including coral gardens, coral reefs and sponge aggregations (Table 6 & 7, and other references).
6.4 - Diversity
Seamounts are generally regarded as hotspots for deep-sea biodiversity. Seamounts are often characterized by very rich communities of filter and suspension feeding corals and sponges (see e.g. Etnoyer 2010, McIntyre et al. 2016), biotopes that are well known to host increased numbers of benthic species (see e.g. Klitgaard 1995, Freiwald et al. 2012). Coral gardens, coral reefs and sponge aggregations are patchily distributed in the deep-sea, primarily due to the need of hard substrate for settlement. However, the bed-rock of the seamounts offer suitable substrate for settlement for many species of coral and sponge larvae. Furthermore, seamounts offer a range of other bottom substrates suitable for colonization of different sets of fauna such as patches of sand and mud, and coral framework that contribute to the overall high species richness of the seamount. A peak in the richness in benthic invertebrates on a seamount off the British Isles (at 1300-1400 m and 1500-1600 m) has been linked to the interface between warm and cold waters, which could harbor both cold water and temperate water adapted fauna, i.e. cool Arctic and warm Lusitanian biogeographic provinces (Henry et al. 2010). It is likely that similar oceanographic features could be of importance in regulating small scale fauna occurrences on seamounts. Beyond the diversity of substrate and water mass properties, seamounts possess a range of other properties that could elevate species abundance and biomass. Accelerated bottom currents around the topographic elevation increases flow and hence food availability. Due to upwelling and enhanced mixing surface productivity and sedimentation rates of organic particles can be elevated increasing the amount of food supplied to the area even further. Jointly, these factors presumably contribute to the potential of seamounts to support high diversity and high abundance of benthic communities. However, such properties may not be unique to seamounts. Recent studies have demonstrated that biodiversity and abundance is equally high in other topographically complex elevated features in the deep-sea, such as banks, hills, knolls and canyons (Howell et al. 2010 and references in that paper). Biodiversity in these types of ecosystems can be (but is not always) elevated compared to the abyssal plains, e.g. Durden et al. (2015) found three times higher benthic fauna biomass on abyssal hills compared to abyssal plains. The large ecosystem studies of the mid-Atlantic ridge undertaken by the ECOMAR and MAR-ECO projects found that the MAR provides an extensive relatively shallow oceanic habitat and distribution area for species well known from continental margin settings. However, there was little support for regarding MAR as having an added effect on benthic biodiversity or biomass beyond what would be expected given the opportunity it offers in terms of area available for colonization and production (Priede et al. 2013).
6.5 - Conclusions
Due to the lack of quantitative studies it is difficult to assess whether seamounts are areas of increased benthic biodiversity or not in the OSPAR Maritime Area. Only one study, i.e. Howell et all. (2010), has systematically studied this and demonstrated that biodiversity and abundance on seamounts off the British Isles was similar to that on banks and hills, not higher. Studies from seamounts in the OSPAR Regions I, IV and V, however, confirm that seamounts are highly suitable for settlement and growth of sessile filter-feeding fauna such as cold-water corals and sponges. On 10 of the 29 seamounts for which benthic species had been studied coral reefs, coral gardens or sponge aggregations were found. Corals have been recorded on another 16 seamounts but the quantitative data needed for defining such habitats is lacking. It is likely that future quantitative studies of seamounts would substantially increase the number of OSPAR seamounts documented to harbour VMEs.
7 - Fish communities
In this chapter published literature from seamounts in the OSPAR Region I, IV and V has been explored to describe the diversity of fish on seamounts and the significance of seamounts as fishing areas.
7.1 - Data collection
The literature search for information on fish assemblages at seamounts in the OSPAR region included scientific databases, internet search engines and reference lists of published papers. Web of Science, Sciencedirect and Google Scholar were used with the following search terms:
Seamount*, and fish*, and northeast Atlantic */NE Atlantic* orth Atlantic*
The name of individual seamounts and fish*
Thereafter, the internet engine Google was used to search for web pages and papers that could provide relevant data and additional references. The database Seamounts Online yielded occurrence data on 58 species entries from 5 seamounts from the OSPAR regions. References for these studies were acquired. The remaining list of papers was compared to the latest review of OSPAR seamounts, Kvile (2011) and Morato et al. (2013), for a final check for possibly lacking data. After excluding papers that were not relevant for this report (studies outside the OSPAR Regions I, IV and V and papers not containing the species information required for the review), we were left with 26 papers containing information on fish species from 15 seamounts or seamount-like features of the OSPAR maritime area (listed in Table 4).
7.2 - Species
Region V was, by far, the best studied region with species data from 13 different seamounts and seamount-like features; compared to only one seamount-like feature from Region I and three seamounts and seamount-like features from Region IV. Comparing abundances between seamounts was not possible due to lack of abundance data from several of the seamounts. The species list compiled is therefore a presence-absence list (Appendix 3). The degree to which the different seamounts were studied also varied greatly. Condor de Terra and Galicia were of the most studied seamounts and/or seamount like features and also contained the highest number of species records. The low number of species on some of the seamounts may be a result of the seamounts being studied less exhaustively.
7.2.1 - Region I
The only study included from Region I is from the Faroe Bank which is located very close the border between Region I and V, hence the fish fauna is unlikely to be typical for Arctic seamounts. The fish species list from Faroe Bank consists of 59 species representing 13 orders, 30 families and 52 genera (Magnussen, 2002). The most abundant species is Atlantic cod (Gadus Morhua, Gadiformes), Haddock (Melanogrammus aeglefinus, Gadiformes), Saithe (Pollachius virens, Gadiformes) and Grey gurnard (Eutrigla gurnardus, Scorpaeniformes), which made up more than 60% of total abundance and biomass. The best represented order is the Gadiformes with 17 species from 5 families. The family Gadidae alone had 8 species from 8 genera. The fish assemblage on the Faroe Bank is mainly boreal, with some occurrence of both Arctic and Mediterranean species, presumably reflecting the openness and position of the bank at the border between provinces. The fish fauna on Faroe Bank is similar to that of neighboring areas. All but one species found on the Faroe bank is also found in the northern North Sea, and 23 of 66 fish species on the Rockall Trough and 17 of 34 species on the slope of the eastern Norwegian Sea were also found on Faroe Bank. Only a small number of the species are utilized commercially at this traditional and productive fishing ground (Magnussen 2002).
7.2.2 - Region IV
The fish species list from Le Danois Bank consist of 74 species from17 orders, 43 families and 66 genera (Serrano et al. 2005). The most abundant species, by biomass, were Blackmouth catshark (Galeus melastomus, Carcharhiniformes) and Rabittfish (Chimaera monstrosa, Chimaeriformes). The more abundant species by numbers was Bluntnose smooth-head (Xenodermichthys copei, Osmeriformes). The more prominent order was Gadiformes with 18 species from 7 families. Macrouridae alone was represented with 7 species from 7 genera. A study comparing Le Danois Bank samples with samples from the same depth on the adjacent continental shelf, found biomass estimates that were three times larger and species richness that was twice as high (Serrano et al. 2005). Nevertheless, Le Danois Bank samples were not found to be significantly more diverse than continental shelf samples.
Gorringe ridge - Gettysberg and Ormond seamounts are the best studied seamounts in region IV. The fish species list consists of 44 species representing 11 orders, 27 families and 35 genera (Abecasis et al. 2009, Gonçalves et al. 2004, Maul 1976, OCEANA, 2005). The most common species were all from the order Perciformes: Canary damsel (Abudefduf luridus), Mediterranean rainbow wrasse (Coris julis) and Ornate wrasse (Thalassoma pavo). The best represented order was Perciformes with 25 species/taxa from 12 families. Best represented family were Labridae with 6 species from 4 genera. Abecasis et al. (2009) found that 56% of the species found at Gorringe ridge were common on both summits.
7.2.3 - Region V
The fish species from Anton Dohrn seamount consist of 27 species from 10 orders, 18 families and 25 genera (Neat et al. 2008, WWF 2001). The most abundant species is the black scabbardfish (Aphanopus carbo, Perciformes), rabbitfish (Chimarea monstrosa, Chimaeriformes), common Atlantic grenadier (Nezumia equalis, Gadiformes) and North Atlantic codling (Lepidion eques, Gadiformes). These four species made up 77%, by abundance. Best represented order was Gadiformes with 9 species from 4 families. Best represented families were Macrouridae and Moridae, both with 3 species from 3 genera.
At Rosemary’s bank the fish species list consists of 25 species, diversified in 9 orders, 17 families and 25 genera (Howell et al. 2007, Neat et al. 2008). Best represented order was Gadiformes with 9 species from 3 families. Most abundant species were blue whiting (Micromesistius poutassou, Gadiformes), Baird’s slickhead (Alepocephalus bairdii, Osmeriformes), slender codling (Halargyreus johnsonii, Gadiformes) and roundnose grenadier (Coryphaenoides rupestris, Gadiformes) which made up 53% of numerical abundance. Also of note were the relatively high numbers of deep-water redfish. The preliminary analyses from surveys which have collected information from the seamounts suggest that they are indeed dynamic ecosystems that may well differ from the shelf slope (Neat et al. 2008).
The fish species list from Condor de Terra seamount in the Azores consists of 135 species from 21 orders, 62 families and 105 genera (Colaço et al. 2013, Giacomello & Menezes 2011). Species richness was high but the demersal catches were highly dominated by Blackspot seabream (Pagellus bogaraveo, Perciformes), Blackbelly rosefish (Helicolenus dactylopterus, Scorpaeniformes) and Common mora (Mora moro, Gadiformes) which made up 65% (numbers) and 53% (biomass) of the catch (Menezes et al. 2011). The most abundant mesopelagic species/taxa were Cyclothone (Stomiiformes), Dofleini’s lanternfish (Lobianchia dofleini, Myctophiformes), white-spotted lanternfish (Diaphus rafinesquei, Myctophiformes), pygmy lanternfish (Lampanyctus pussillus, Myctophiformes) and scaly dragonfish (Stomias boa ferox, Stomiiformes) (Porteiro et al. 2011). Best represented order was Perciformes with 37 species from 17 families, while best represented familiy were Myctophidae with 15 species from 13 genera. The species number, catches per unit of effort, and zonation with depth found at the Condor de Terra seamount were in general agreement with that observed for the demersal fish community of the Azores (Menezes & Giacomello 2013).
At Galicia Bank 139 species representing 23 orders, 63 families and 111 genera occurred. The most abundant species were Mediterranean slimehead (Hoplostethus mediterraneus, Beryciformes) and Mediterranean codling (Lepidion lepidion, Gadiformes) (Bañon et al. 2016). Best represented order was Gadiformes with 24 species from 7 families. Best represented families were Macrouridae (12 species), followed by Moridae, Stomiidae and Sternoptychidae (7 species each). Of the 139 fish species reported in Bañon et al. (2016), over 70% have been reported in the continental shelf and slope of Galician waters. The lack of observation of the remaining species is likely due to a less intensive sampling on the deep-water areas of the Galician coast, compared to Galicia bank. Many of these species have been reported in other areas of the North-eastern Atlantic, indicating the lack of endemic species on the bank.
The fish species list from Formigas Dollabarat consists of 41 species from 7 orders, 18 families and 37 genera (Afonso et al. 2018). Most common species, by occurrence in trawl catches, were all from the order Perciformes: ornate wrasse (Thalassoma pavo), parrotfish (Sparisoma cretense), Mediterranean rainbow wrasse (Coris julis), Canary damsel (Abudefduf luridus) and Azores chromis (Chromis limbata). Best represented order were Perciformes with 27 species from 10 families. Best represented families were Carangidae and Labridae, both with 6 species from 5 genera.
The lowest number of species was found at George Bligh bank. The species list consists of 13 species from 8 orders, 10 families and 10 genera (Narayanaswamy et al. 2013). Most common order was Gadiformes with 5 species from 3 families.
Hatton bank is one of the best studied seamount-like features in Region V, but different from many of the others because it is very extensive and rather to be regarded as a continental shelf and slope habitat. The fish species list consists of 29 species, diversified in 11 orders, 19 families and 23 genera (Howell et al. 2007, Narayanaswamy et al. 2013, OASIS 2003). Most abundant species were roundnose grenadier (Coryphaenoides rupestris, Gadiformes) and rabbitfish (Chimaera monstrosa, Chimaeriformes) (Howell et al. 2007). The most common order was Gadiformes with 10 species/taxa from 4 families.
Extensive studies of pelagic and demersal fish were conducted on the mid-Atlantic Ridge, including on e.g. the Faraday seamount with a species list consisting of 133 species and is diversified in 19 orders, 47 families and 105 genera (Bergstad et al. 2008; OASIS 2003; Sutton et al. 2008). The most dominant species were glacier lanternfish (Benthosema glaciale, Myctophiformes) which made up 51% and 28% of the catch, in abundance and biomass respectively. Best represented order were Stomiiformes with 25 species/taxa from 5 families. Best represented family were Myctophidae (from Myctophiformes) with 20 species from 15 genera. The mid-Atlantic Ridge (MAR) has numerous seamounts and seamount-like features and the Faraday may not be representative for more than a subset of these. For the pertinent section of the MAR, Bergstad et al. (2008) and Sutton et al. (2008) offer comprehensive accounts of demersal and pelagic fishes, respectively. A full check-list of fishes recorded on the mid-Atlantic Ridge between the Azores and Iceland was recently published (Porteiro et al. 2017), including extensive literature references to older studies in this area.
Hebrides Terrace seamount is one of the seamounts with the lowest number of species; only 21 species, diversified in 8 orders, 10 families and 11 genera (Milligan et al. 2016). Most abundant species were North Atlantic codling (Lepidion eques, Gadiformes), False boarfish (Neocyttus helgae, Zeiformes) and roundnose grenadier (Coryphaenoides rupestris, Gadiformes) which together made up nearly 60% of observed abundance. Best represented order were Gadiformes with 8 species from 3 families. Best represented family was Macrouridae with 4 species from 3 genera. Milligan et al. (2016) found significant variation in community composition between transects on Hebrides Terrace and reefs at Rockall Bank but were open to the possibility that these differences could be a result of stochastic variation caused by the low densities of deep-sea fish.
The fish species list from Sedlo consists of 80 species from 20 orders, 43 families and 65 genera (Menezes et al. 2009, Menezes et al. 2012). According to Menezes et al. (2009), the most abundant species were splendid alfonsino (Beryx splendens, Beryciformes), black cardinal fish (Epigonus telescopus, Perciformes) and common mora (Mora moro, Gadiformes) which made up 79% and 58% of the catch, by abundance and biomass, respectively. Best represented order was Gadiformes with 16 species from 4 families. Best represented family was Macrouridae with 9 species 7 genera.
The fish species list from Josephine bank consist of29 species from 11 orders, 21 families and 28 genera (SeamountsOnline; Froese & Sampang 2004b). Most common order was Perciformes with 10 species/taxa from 8 families.
The fish species list from Joao de Castro bank consists of 33 species from 8 orders, 22 families and 29 genera (Cardigos et al. 2005, Santos et al. 2010). The most common order was Perciformes with 21 species/taxa from 12 families. Most common families were Labridae and Carangidae with 4 species from 3 and 4 genera, respectively.
7.3 - Diversity
Our review of fish assemblage studies on seamounts and seamount-like features in the OSPAR Regions I, IV and V resulted in a list of 456 species, representing 29 orders and 132 families. This is ulikely to be exhaustive, and it must be noted that there is a high diversity of studies with different methodologies. Also, the list includes both demersal and pelagic fishes, but to a varying degree between studies and sites. We have not compared this number to overall species lists for the relevant OSPAR Regions, but the rather high number of species recorded shows that seamounts are habitats for a high proportion of the regional species pool.
The most common (by number of observations) and diverse orders were Perciformes and Gadiformes. Perciformes was represented by 102 taxa from 27 families and made up 22% of all presence/absence registrations over all seamounts combined. Gadiformes was represented by 52 taxa from 7 families and made up 17% of the observations. The majority of the dominant species from the different seamounts belonged to one of these two orders. Other important orders were Stomiiformes, Squaliformes, Osmeriformes and Myctophiformes (in order of importance).
The species list represents only 1.3% of the fish species on Earth, but 25% of all families and 45% of all orders. This means that the seamount fishes in the OSPAR region represent a genetic diversity higher than what the number of species alone would suggest. This result is consistent with similar studies done for over 60 seamounts worldwide (Froese & Sampang 2004). On the seamounts studied in this report, the number of species and number of genera is nearly the same, meaning that the majority of species are the only representative of their genus.
Seamounts are regarded as hotspots of pelagic biodiversity. Morato et al. (2010) observed higher species richness in association with seamounts in the central and western Pacific Ocean than in coastal or oceanic areas. Seamounts were found to have higher species diversity within 30–40 km of the summit. The information about biodiversity at seamounts in the OSPAR regions compared to adjacent areas is sparse and does not allow similar comparisons and analyses. The limited amount of information found indicates both a higher diversity at the seamount (i.e. Le Danois Bank) and the lack of such a difference (i.e. Condor de Terra), but further analyses would be needed to resolve these issues.
7.4 - Fisheries
Le Danois bank: There is no constant fishery on Le Danois bank. Only a few vessels work sporadically using gillnets and targeting monkfish (Lophius spp.), or long lines targeting Beryx spp., forkbeard (Phycis blennoides) and red sea-bream (Pagellus bogaraveo) (Serrano et al. 2005).
Anton Dohrn and Rosemarys bank: Fisheries on the Anton Dorhn are targeting blue ling (Molva dypterygia) while deep-water redfish (Sebastes) and blue ling are targeted at Rosemary’s bank. Landings of all the major commercial species in this region have declined in the last decade. No formal assessments are made, but CPUE data suggest declining abundance in many species (Neat et al. 2008).
Condor de Terra seamount: The demersal fisheries on Condor de Terra Seamount was closed for the first time in June 2010 after a pronounced decline in abundance of targeted species and has remained closed since then. This area is one of the most accessible seamounts for the scientific community and is an internationally recognized study area of reference (Menezes et al. 2011). Previous fisheries were multi-specific longline and handline fisheries. The main targets were blackspot seabream (Pagellus bogaraveo) and Atlantic wreckfish (Polyprion americanus).
Galicia bank: Fishing activity on the bank has progressively decreased. The low fishing pressure and absence of bottom trawling have led to well-preserved deep-sea biotopes of conservation importance such as coral communities. Nowadays, only 3 vessels are sporadically moving to the bank, targeting Anglerfish (Lophius piscatorius) with gillnets (Bañon et al. 2016).
Gorringe ridge: There is no official information about the type and intensity of fishing, but it is general knowledge that the Gorringe Bank is exploited by several deep-water commercial fleets, including long-liners targeting scabbardfish (Lepidopus caudatus) and other species (Gonçalves et al. 2004).
Hatton bank: The upper slopes of this extensive bank area have been important for mixed-species fisheries by an international trawler and longliner fleet, fishing both inside and outside EEZs. Trawlers have mainly targeted roundnose grenadier (Coryphaenoides rupestris), and deep-sea bottom longline fisheries targeted Greenland halibut and gadoids, with a significant bycatch of deepwater sharks (Bensch et al. 2009, ICES WGDEEP). The fisheries are regulated by catch quotas and spatial measures by the North East Atlantic Fisheries Commission (NEAFC) and the EU, and a ban on targeting of elasmobranchs are in force. In order to protect vulnerable marine ecosystems the North East Atlantic Fisheries Commission (NEAFC) and the European Union (EU) have closed parts of the Hatton Bank to bottom fishing (EC 2009, NEAFC 2010), and restricted fishing to certain subareas.
Dom João de Castro bank: Dom Joao de Castro is an important fishing ground both for demersal fish, such as the black-spot seabream (Pagellus bogaraveo) and the blue-mouth (Helicolenus dactylopterus), and tuna (Santos et al. 2010).
On the mid-Atlantic Ridge rather extensive fisheries were conducted in the past, but these have declined to low levels (see https://www.neafc.org/international/22299) and are now managed by a set of restrictive spatial measures and total allowable catch limits maintained by NEAFC, the EU and states. There is a small fishery for orange roughy being conducted on Faraday Seamount by the Faroe Islands, and some fishing for roundnose grenadier on the Reykjanes Ridge, but past fisheries for e.g. alfonsino on seamounts in the high seas near the Azores ceased several decades ago.
7.5 - Conclusions
Seamounts are habitats for fish from the regional pool of fishes, hence there are large differences in the fish assemblage identity amongst seamounts within the OSPAR Maritime Area. Species compositions and vertical distribution patterns seem to correspond with that of adjacent island and continental slopes. On a higher taxonomic level species diversity is high. The species list presented in this report represents only 1.3% of all fish species on Earth, but 25% of all families and 45% of all orders. Open ocean seamounts constitute small and presumably somewhat isolated oceanic living-spaces for species that occur on continental slopes and island slopes. This widens the ranges of some species and assemblages, but too few studies of connectivity have been carried out to draw general concludions on present connectivity and significance of seamounts as ‘stepping stones’.
Typical features of seamount summits, especially in Region V, are shoals of e.g. alfonsino, orange roughy and grenadiers, as well as mesopelagic scattering layers of a mixture of invertebrates and fish that impinge on the summits and slopes during daytime. The significance of seamounts in OSPAR for wide-ranging tunas and pelagic sharks is unclear. Some of the oceanic seamounts with aggregating benthopelagic species of commercial value (e.g. alfonsino, grenadiers, orange roughy, redfish, Greenland halibut) attracted extensive trawl fisheries in the past, but few of these fisheries persisted beyond an exploratory phase. Exceptions are some of the fisheries on the slopes of banks near continental Europe still being conducted but now under stricter management. More artisanal and locally very important fisheries are conducted on many seamounts relatively close to islands and continents. With few excpections, such as the Faroe Bank, seamounts are insignificant as fishing areas in Region I, but remain somewhat more important in Regions V and IV, at least if major banks near continents are considered.
8 - Marine mammals
In this chapter published literature and online research databases have been searched for documentation to explore to what extent marine mammals are associated with seamounts or seamount-like features in the OSPAR maritime area.
8.1 - Species occurrence
While substantial efforts have been made to relate marine mammal occurrence to steep topographies, the literature addressing specific association with seamounts is scarce. Morato et al. (2008) undertook a comprehensive study of the abundance of key marine organisms, including some marine mammal species, on seamounts within the Azores EEZ, relative to the abundance in deeper adjacent waters. The data were obtained during regular fisheries surveys, and in terms of marine mammals, they were limited to the most common visitors: common dolphins (Delphinus delphis), spotted dolphins (Stenella frontalis), bottlenose dolphins (Tursiops truncatus) and sperm whales (Physeter macrocephalus). Results indicated that only common dolphins showed some degree of association with seamounts (Morato et al. 2008). However, most dolphins are not generally considered deep divers and would only benefit from shallow seamount summits, although a purported maximum dive depth of 700 meters have been reported for striped dolphins (Stenella coeruleoalba, Archer 2002, Ringelstein et al. 2006). Of the four species included in the study by Morato et al. (2008), only sperm whales are known as a deep diver, regularly diving to depths of >700 meters and often exceeding 1000 meters (e.g. Watkins et al. 2002, Watwood et al. 2006). This species thus has the capability of reaching and potentially exploiting ecosystems in direct association with many of the deeper seamounts identified in this region.
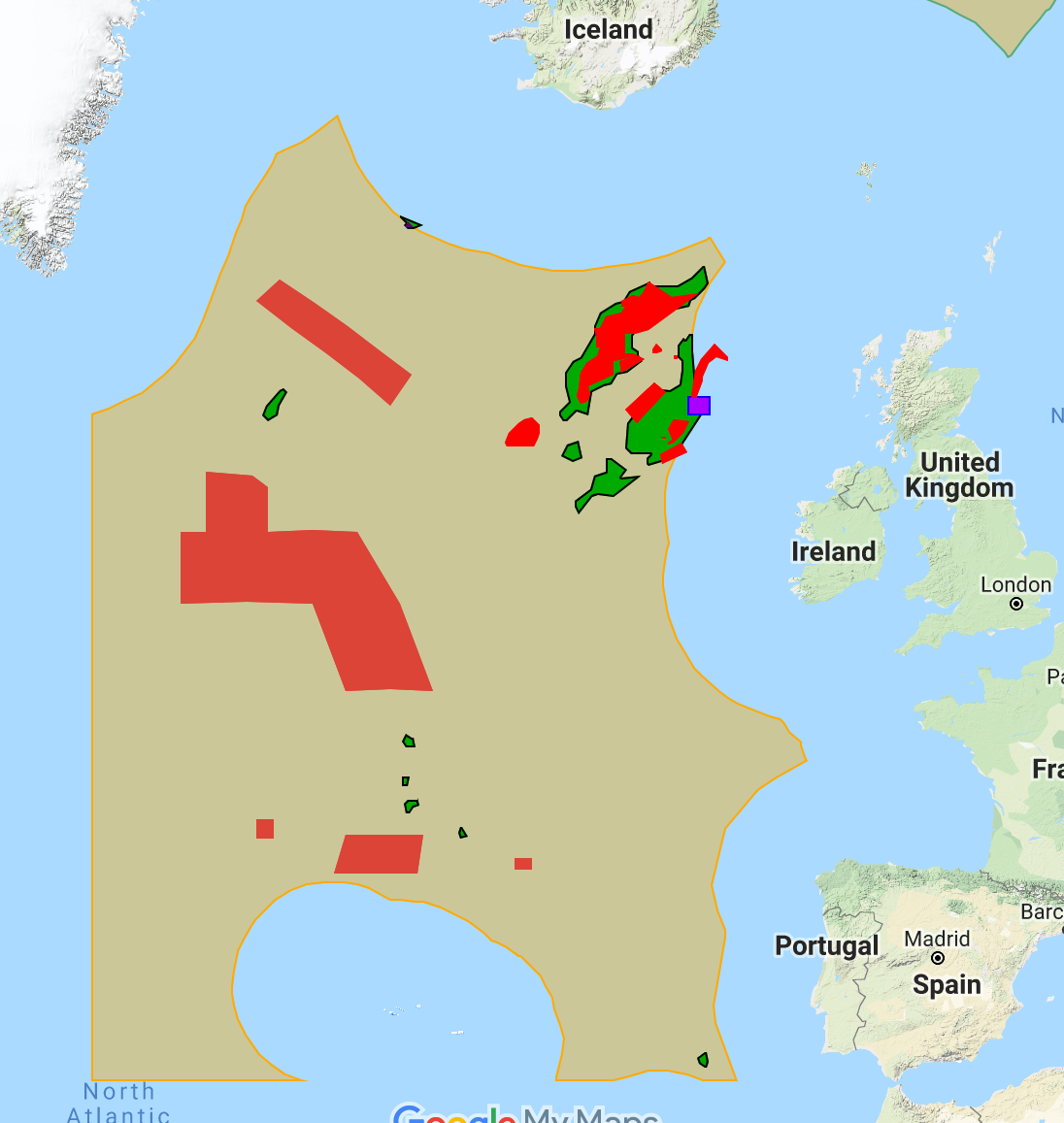
Since Morato et al. (2008) published their findings, the online repository OBIS (Ocean Biogeographic Information System, http://www.iobis.org/) has grown significantly, and now constitutes an invaluable resource from which to extract geographic data for a range of marine organisms. We extracted all observational data available in OBIS for marine mammals within the OSPAR region, constituting 124,167 unique observations of whales and 5,803 for seals. These represent a combination of direct human observation (e.g. visual surveys from ships or airplanes) or machine observation (e.g. via satellite telemetry from animal-borne transmitters and dataloggers). The spatial distribution of these observations across the OSPAR region is shown in Figure 5, which suggests a strong bias in observations towards coastal areas, or areas surrounding oceanic archipelagos such as the Azores.
Figure 5. Modelled seamounts within the OSPAR region (yellow), and all marine mammal sightings available for this region through the Ocean Biogeographic Information System (OBIS). Whale observations are represented by red dots, while seal sightings are indicated by green diamonds.
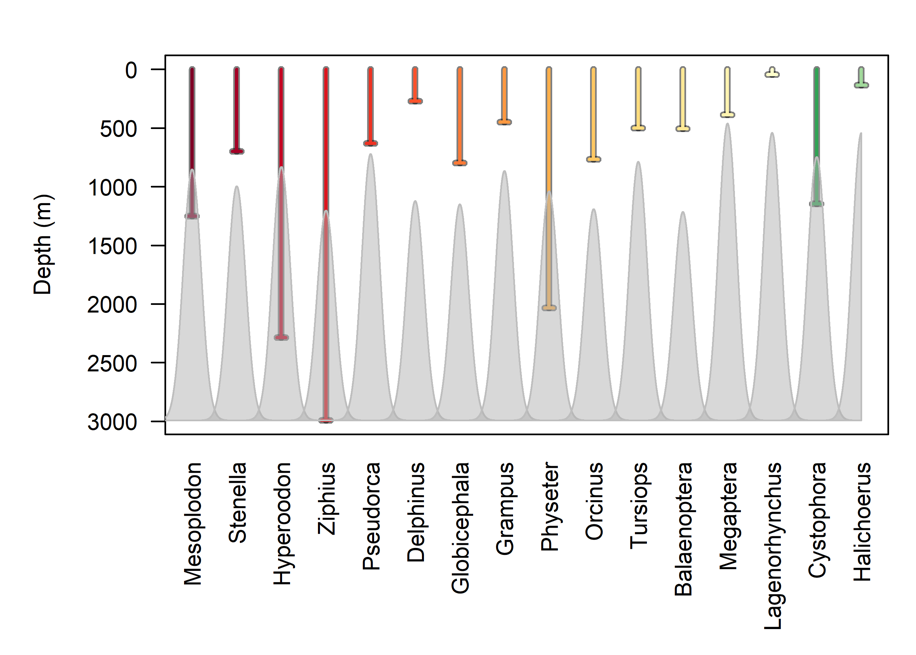
Even when marine mammals are present in areas geographically associated with seamounts, this does not necessarily mean that they associate with, or are dependent on, the seamount associated ecosystems. Since marine mammals are air-breathing divers, they must return regularly to the surface to breathe. This limits their ability to exploit resources at greater depths, and their ability to come into direct contact with seamount-associated ecosystems depends greatly on their diving capability. Diving capabilities varies greatly among and within marine mammal taxa. While most delphinids are not known for deep-diving abilities, striped dolphins Stenella coeruleoalba are known to feed partially on deep-dwelling prey, and based on stomach contents they are believed to be able to dive to at least 700 m (Ringelstein et al. 2006). False killer whales (Pseudorca crassidens) have also been shown to be capable of deep dives to ~600m (Minamikawa, Watanabe, and Iwasaki 2013). While killer whales (Orcinus orca) are not generally thought of as deep divers, they have been shown to be capable of very deep dives, down to more than 700 m (Reisinger et al. 2015). Similarly, while bottlenose dolphins (Tursiops truncatus) generally dive to relatively shallow depths, they have been shown to be capable of performing dives to depths of at least 450 m (Klatsky, Wells, and Sweeney 2007). Among other odontocetes (toothed whales), long-finned pilot whales (Globicephala melas) are capable of long and deep dives, regularly to below 600 m with maximum depths in excess of 800 m (Heide-Jørgensen et al. 2002, Aoki et al. 2017). Sperm whales (Physeter macrocephalus) are commonly referred to as the true deep-diving whales and are indeed capable of dives to at least 1185 m (Watkins et al. 1993). However, recent studies have revealed that the true champions among diving whales belong to the family Ziphiidae (beaked whales and bottlenose whales). The deepest known dive of any marine mammal, 2 992 meters, was reported for a Cuvier’s beaked whale (Ziphius cavirostris). While this study was conducted of the Southern California coast, this species occurs in all the world’s oceans, including the Northeast Atlantic. Similarly, northern bottlenose whales (Hyperoodon ampullatus) occur throughout the northern hemisphere, and have been reported to dive to depths of at least 1 453 meters (Hooker and Baird 1999), and possibly well over 2 000 meters (Rune R. Hansen pers. comm). For Mesoplodon, Stenella, Hyperoodon and Ziphius 12-6% of all observations of the species occur within 10 nautical miles of a seamount (Figure 6).
Figure 6. Proportion of all marine mammal observations included in the OBIS database that occur within 10 nm of a seamount, summarized per genus.
Several of these deep-diving species are known to occur within the OSPAR region and are therefore more than likely to interact with seamounts within this region. Figure 7 shows the diving capability of marine mammal species groups known to visit seamounts in the OSPAR region, and the average depth of the seamounts which they are known to visit.
Figure 7. Maximum observed diving capabilities of marine mammals known to occur at seamounts in the OSPAR region, relative to the average peak depth of visited seamounts.
8.2 - Conclusions
Available marine mammal data revealed that several groups of deep diving whales and seals (i.e. Mesoplodon, Hyperoodon, Ziphius, Physeter and Cystophora) are commonly sighted at seamounts and either only the summit or the full seamount are within their diving ranges. There is potential for deep divers such as the beaked and bottlenose whales (Hyperoodon and Ziphius) and sperm whales (Physeter) to use the summit and the slopes of the relatively deep seamounts as foraging grounds. Most sightings of sea mammals are, however, from coastal areas of the European mainland and the large islands and island groups of the NE Atlantic (i.e. Svalbard, Azores and Island) and the relative significance of seamounts and other features to marine mammals in the OSPAR Regions I, IV and V is still unclear.
9 - Ecosystem structure & function
9.1 - Enhanced productivity and biomass
A number of distinct physical flow-features are generally associated with seamounts, i.e. doming of density layers, increased vertical mixing, flow acceleration and internal waves. These can contribute to bringing nutrient-rich deep-water into the euphotic zone resulting in enhanced photosynthetic production. This may in turn increase the flow of seston and particulate organic matter to deep water layers (White et al. 2007). Hence, compared with surrounding oceanic waters, seamounts potentially sustain elevated populations of both zooplankton and benthic invertebrates, as well as pelagic and demersal fish (McClain et al. 2010, Etnoyer 2010, Schlacher et al. 2014 and many others). However, there is a large variation between seamounts and conditions associated with them, hence to what extent these general features and characteristics apply to seamounts within the OSPAR maritime area, is not known.
We explored the literature collated for this report to consider four common claims/hypotheses:
Seamounts are characterized by distinct physical flow-features such as doming of density layers, increased vertical mixing, flow acceleration and internal waves
Phyto- and zooplankton production is increased on summits and adjacent to seamounts
Biomass, abundance or cover of sessile benthic fauna is enhanced on and around seamounts
Biomass of pelagic and demersal fish is enhanced around seamounts
There are only a few seamounts studied in sufficient detail to carry out meaningful explorations of these hypotheses. We found comprehensive documentation of oceanography, plankton, benthos and fish from two seamounts (Sedlo and Condor de Terra) and two banks (Le Danois and Don Joao de Castro) within the OSPAR maritime area. Results from those studies are summarized below (Table 8).
Table 8. Summary of available information on physical flow features, primary and secondary productivity and biomass of phytoplankton, zooplankton, benthos and fish from two seamounts and two banks in the OSPAR maritime area.
Dom Joao de Castro bank has a shallow summit with hydrothermal activity in the euphotic zone. Reduced zooplankton biomass over the seamount (“a biomass hole”) has been detected but with an increased abundance of Ceratoscopelus maderensis fish larvae (Sobrinho-Goncalves & Cardigos 2006). There is not quantitative information on neither benthos or fish that would allow comparisons of secondary production with adjacent areas of similar topography. Cold-water coral and sponges have been described from the area, however, quantifications allowing the classification of biotopes are lacking.
Sedlo seamount was studied in detail in the OASIS project, with several cruises dedicated to the studies of oceanography, plankton, benthos and fish during the year 2002-2005. The studies confirmed the existence of an anti-cyclonic flow above the seamount likely due to the generation of a Taylor cone retaining particles as well as larvae around the seamount (Mohn et al. 2009). The presence of high-quality suspended particulate organic matter present around the seamount (Kiriakoulakis et al. 2009) could provide an important food source to the biological communities of seamount. Benthic surveys indicate the seamount is inhabited by a highly diverse community of sessile megabenthos, of mainly Hexacorallia and sponges (Santos et al. 2010), however, no quantitative estimations allowing comparisons with adjacent areas of similar topography have been published neither for benthos nor fish.
Condor de Terra seamount was studied in detail from 2008 to 2011 within the CONDOR project (CONDOR - Observatory for long-term study and monitoring of Azorean seamount ecosystems), collecting multiannual and seasonal samples on oceanography, plankton, benthos as well as fish. Condor de Terra is by far the most well studied seamount in the OSPAR maritime area, and it is easily accessible close to islands in the Azores. The oceanographic conditions are characterized by enhanced mixing, upwelling–downwelling processes and closed circulation structures over the seamount making is distinct from the surrounding ocean (Bashmachnikov et al. 2011). However, over the three years examined the study found no effect of the seamount on the abundance of phytoplankton or chlorophyll a (Santos et al. 2013) or the abundance or diversity of zooplankton (Carmo et al. 2013). Neither were abundances or biomasses of pelagic and demersal fish enhanced around the seamount. Benthic biotopes of particular interest are the Viminella flagellum and Dentomuricea sp. coral gardens in coexistence with the hydrozoan Polyplumaria flabellata and the Pheronema sponge aggregations (Tempera et al. 2011). No quantitative descriptions of the benthos were found, neither any comparisons of cover or biomass of large sessile epifauna between the slopes of the seamount and slopes of nearby islands, hence hypothesis regarding enhances biomass of epibenthos on seamounts was not possible.
Le Danois bank was studied in detail in 2003 and 2004 within the ECOMARG project, collecting samples on bathymetry, oceanography, plankton, benthos as well as fish. This bank is one of the most well studied seamounts like features in the OSPAR maritime area. Circulation patterns have been studied and identified the formation of a weak Taylor column over the bank (González-Pola et al. 2012). Lower pelagic diversity over the seamount has been detected compared to the surrounding ocean and well as a lower abundance of mesopelagic decapods, mysids, and fishes while an elevated abundance of euphausiids (Cartes et al. 2007b). Diversity was similar on the seamount as compared to the continental shelf. Catches of demersal fish as well as sponges and species from Echinodermata were substantially higher on the bank compared to adjacent shelf areas (Serrano et al. 2005a).
Overall, this exersice indicates that, despite rather extensive research efforts to collect data on oceanography and to measure rates of biological processes during at least the last two decades, very little quantitative information exsists from seamount or seamount-like features in the OSPAR martime area. Some studies have demonstrated that seamounts are characterized by distinct physical flow-features such as formation of Taylor column, closed circulation cells and often experience enhanced vertical mixing. However, this rarely leads to enhanced phytoplankton or zooplankton productivity. When measured, zooplankton abundance is similar or lower compared to adjacent areas. Where processes in the water column have been measured there seems to be a lack of concurrent quantitative surveys of both benthos and fish. Assessing the link between pelagic and benthic productivity is therefore not possible.
9.2 - Endemicity & connectivity
The early literature frequently referred to seamounts as isolated biological “islands” in the ocean serving as areas of speciation and having high rates of endemism (presence of species unique to a defined geographic location) (Hubbs 1959, Stocks & Hart 2007). A different view has been that seamounts function as stepping stones for species dispersal, in particular for species with limited larval life spans. Using seamounts as stepping stones might facilitate widening of distributions across ocean basins of e.g. the NE Atlantic. This literature review found little evidence to support the proposal of higher levels of endemism on seamounts, and rather contrasting results for different taxa. Without a broad sampling of adjacent and regionally distributed seamounts it is not possible to conclude that new species found and described at a particular seamount are endemic to that seamount or not. A few examples below illustrate the problem.
Comparisons of seamounts and seamount-like features in the North Atlantic indicate that endemism amongst coral taxa at seamounts is low, or non-existent (Howell et al. 2010, Thoma et al. 2009). Indeed, seamounts are normally inhabited by species/taxa with of a wide geographical distribution. The extensive study of corals around the Azores Islands and seamounts of Braga-Henriques et al. (2013) documented the occurrence of 164 coral species and with 23 species being restricted to the Azores region. However, none of the species were restricted to seamounts. Studies of other animal groups confirm that levels of endemism in the Azores region compared to other areas of the Atlantic can be high. Ávila et al. (2012) documented that 45% of the Rissoidae species (Gastropoda, Mollusca) in the Azores are endemic to that region while in e.g. Greenland and Scandinavia the ratio of endemism is much lower (6-3%).
Many studies of seamounts have found high proportions of fish species thought to be endemic. On the Great Meteor seamount in the North Atlantic (but not in the OSPAR maritime area), 9% of the fishes found were endemic (Fock et al. 2002), and on Hawaiian seamounts the rate was 5% for fishes (Stocks 2002). However, some scientists are questioning the “island hypothesis” by pointing out that many seamounts may have apparently high rates of endemism because the full range of the species is not known. In a recent review by Kvile et al. (2014) a total of 155 actual values of endemism were compiled for 107 different seamounts. Among these values, 112 opposed the idea of seamounts as centers of endemism while 32 supported the idea. The literature search for data on fish assemblages on seamounts in the OSPAR region yielded no information on endemic species. On the contrary, several of the papers stated that the fish assemblages were comparable to adjacent areas (Magnussen 2002, Menezes et al. 2011, Bañon et al. 2016).
Overall, there seems to be more support for the idea that seamounts act as stepping stones for species dispersal, rather than isolated biological “islands” serving as areas of increased speciation, in particular for fish. E.g. the species compositon of fish communities on the summits and upper slopes between the seamounts around the Azores islands and the Horseshoe seamounts is highly similar. Indeed, seamounts are often inhabited by species/taxa with of a wide geographical distribution. The large ecosystem assessments of the mid Atlantic ridge undertaken by the ECOMAR and MARECO projects did not detect elevated endemism at the mid Atlantic ridge, instead species represented there were species also known from continental margins of the northern Atlantic (Bergstad et al. 2008, Priede et al. 2013). For Rissoidae snails (Mollusca, Gastropoda) there is evidence of species dispersal between the seamounts Gorringe, Josephine, Ampère and Seine, however, these seamounts do not act as a stepping stones between Portugal mainland and the Madeira archipelago. Instead larvae dispersal seems to occur between the Portugal mainland and the Madeira archipelago (Avila et al. 2012). In order to model how seamounts could facilitate species dispersal for different sets of species between different sets of seamounts detailed knowledge on oceanographic flow, reproductive cycles, larval distributions and longevity is required. Genetic samples can potentially help verify results from the larval dispersal models.
9.3 - Vulnerable Marine Ecosystems
9.3.1 - Defining vulnerable marine ecosystems
Vulnerable Marine Ecosystems (VMEs), as used here, consists of groups of species, communities and habitats that may be particularly vulnerable to impacts from bottom fisheries. The VME concept stems from a series of United Nations General Assembly (UNGA) resolutions regarding sustainable fisheries requesting states and regional fisheries management organizations/arrangements to take immediate actions to protect VMEs from destructive fishing practices in areas beyond national jurisdiction (UNGA 2004, 2006 and 2009). The definition of VMEs was widely discussed during the development of actions, including in several expert workhops convened by the FAO in 2006-2008. In a Technical Consultation in 2008, requested by the FAO Committee on Fisheries (COFI), the International Guidelines for the Management of Deep-sea Fisheries in the High Seas (FAO 2009) were adopted. These guidelines are instruments of reference to help States and RFMOs/As in formulating and implementing measures for the management of deep-sea fisheries in the high seas and they recommend specific conservation and management measures for the protection of VMEs. The focus is on the high seas, but it is customary that States are attentive to actions taken in the high seas and endeavour to introduce similar actions within their respective jurisdictions.
In the FAO guidelines efforts are made to define and exemplify VMEs, and briefly, recommends that VMEs are classified as vulnerable based on the following characteristics: 1) uniqueness or rarity, 2) functional significance, 3) fragility, 4) species life history traits making recovery difficult, and 5) structural complexity (FAO 2009, Article 42). In the Annex to the guidelines (FAO 2009), scleractinian, alcyonacean, gorgonian, antipatharian and stylasterid corals are listed as groups that often contain sensitive and potentially vulnerable species that often contribute to forming VMEs. The list also contains some types of sponge dominated communities, chemosynthetic seep and vent communities with high level of endemism and communities composed of dense aggregations of large xenophyophores, hydroids and bryozoans. Furthermore, the guidelines list a range of specific topographical, hydro-physical and geological features that potentially support VME taxa. These include; 1) summits and flanks of seamounts, guyots, knolls, hills and banks, 2) submerged edges and slopes, 3) canyons and trenches, 4) hydrothermal vents, and 5) cold seeps.
The guidelines clarify that risks of adverse impacts are determined by vulnerability, the probability of a threat occurring, and the mitigation means applied to the threat. This means that not all VMEs necessarily need the same level of protection. Accordingly, in order to further assist management, the guidelines encourage evaluation of risk of adverse impacts and defines ‘significant adverse impacts (SAI)’ (in Para. 17-20). SAIs are those that compromise ecosystem integrity (structure or function) in a manner that: 1) impairs the ability of the affected populations to replace themselves; 2) degrades the long-term productivity of habitats; and 3) causes, on more than a temporary basis, significant loss of species richness, habitat or community types. Impact should be evaluated individually, in combination and cumulatively. The guidelines furthermore list a series of criteria to be considered when making SAI evaluations.
9.3.2 - Vulnerable species and habitats associated with seamounts
A range of vulnerable benthic invertebrates and biotopes are commonly associated with seamounts, including coral gardens, coral reefs, sponge aggregations and xenophyophore aggregations (Table 7). On the Anton Dohrn seamount alone Davies et al. (2015) mapped 13 different biotopes of which 10 could be considered vulnerable marine ecosystems sensu FAO (2009), i.e. different types of coral gardens, coral reefs, sponge aggregations or xenophyophore communities. All the coral taxa listed in the annex of the FAO Guidelines (as groups that often contain sensitive and potentially vulnerable species that often contribute to forming VMEs) have been registered on numerous OSPAR seamounts (Appendix 2). Sponge dominated communities (of both Hexactinellidae and Demospongia) are commonly encountered on seamounts (McIntyre et al. 2014, Ramos et al. 2016, Roberts et al. 2019) as are xenophyophores (Cross et al. 2014). At least one seamount like feature in the OSPAR area, i.e. Dom Joao de Castro bank, has a well described area of hydrothermal venting (Colaco et al. 2006).
The management actions called for by UNGA in relation to VMEs, and the guidance offered in FAO (2009), concern specifically the need to avoid significant adverse impacts of fishing on VMEs where they are known or likely to occur, and this is one of several aims of sustainable fisheries management. Another key aim is to ensure lasting fishing opportunities, and the UNGA resolutions also concern this aspect. It is fully recognized that the harvested target resources (and by-catches) may also be ‘vulnerable’ but facilitating fisheries and conserving resources into the future require a range of additional analyses and actions than those included in the FAO Guidelines on Deep-Sea Fisheries in the High Seas (FAO 2009).
Several seamounts have in the past been subjected to a boom and bust fisheries for aggregating fish (see chapter on Seamount fishing and management). The reason for the ‘boom and bust’ characteristic of seamount trawling fisheries lies in the life history traits of fish species associated with seamounts. Many seamount fishes have significantly longer lifespans, higher age at maturation, slower growth, and lower natural mortality compared to non-seamount fishes (Morato et al. 2004), rendering them far more vulnerable to exploitation (Jennings et al. 1998, Musick 1999, Denney et al. 2002) than other species. In addition, species that display aggregation behavior such as shoaling and schooling may have higher vulnerability because of increased catchability (Pitcher and Parrish 1993). Simulations by Morato et al. (2004) indicated that exploitation rates higher than 5% would not be sustainable for many seamount species.
Data on fisheries and the species targeted on seamounts and in other deepwater areas are compiled by national laboratories and used by ICES (International Council for the Exploration of the Sea) in the annual assessments forming the basis of advice to national and intergovernmental fisheries management and e.g. OSPAR. The vulnerability of chondricthyans and some teleost fishes is well known and recognized by ICES and management bodies. Management advice reflects this recognition but is rarely very precise due to data deficiency causing assessments to be inadequate for many species. In this situation management advice is mostly based on precautionary approaches.
Of the 456 fish species on our list, 18 (4%) are listed as threatened by the IUCN. Another 10 species are listed as “Near-threatened”. Of the 18 threatened species, 12 belong to the class Chondrichthyes. The proportion of threatened species ranged from 0 to 10%. These listings serve mainly to raise awareness, whereas the ICES advice provides concrete evaluations of states and variability and proposed management action.
9.3.3 - Spawning, nursery & feeding grounds for fish
Seamounts in the OSPAR maritime area presumably function as nurseries, feeding and spawning areas for a range of fish species, but there are rather few studies focusing on these roles, hence the significance of seamounts for the completion of life-cycles is not fully understood. To some species, that are known to aggregate at seamounts as shoals or schools, the association is rather obvious, and it must be assumed that seamounts are highly important, e.g. alfonsinos (Hareide 1995, Hareide & Garnes 2001). The same may be the case for orange roughy, although that species also occurs in other continental slope areas. To some other species common at seamounts, e.g. roundnose grenadier (Macrouridae), the significance of seamounts is less clear because major aggregations also occur along continental slopes, on mid-ocean ridges, and even in deep fjords and shelf troughs. A recent paper suggests that the Hebrides Terrace and potentially also other seamounts in the area serve as nursery grounds for deep-water skates (Henry et al. 2016). The observation of spawning blackmouth catshark Galeus melastomus in the cold-water coral reef Mingulay furthermore indicate that seamount harboring Lophelia pertusa cold-water coral reefs could act as nursery ground for elasmobranchs (Henry et al. 2013).
9.3.4 - Seamount fishing and management
In the Northeast Atlantic, states and intergovernmental fisheries management bodies have for a long time been concerned about the sustainability of deep-sea fisheries, including those conducted on seamounts and oceanic banks and ridges. Severe declines in some fisheries in the 1990s (e.g. redfishes, orange roughy, Greenland halibut and blue ling) following an overly optimistic and almost unregulated expansion in international deep-sea fisheries in the 1980s and earlier (e.g. see multiple papers in Hopper 1995) gradually facilitated greater focus on the need for adequate targeted management measures based on relevant scientific advice. In the mid-1990s the ICES established an expert group dealing with deep-sea fish and invertebrate stocks. Despite chronic shortage of data, ICES soon issued advice pointing to the vulnerability of the target resources due to their life-history characteristics, their tendency to aggregate, and their likely slow recovery rates following depletion (e.g. Gordon 2005, Large and Bergstad 2005). This was also recognized worldwide more or less at the same time (e.g. NAFO 2001, FAO 2005, 2008) and sparked a lot of research on various aspects of deep-sea fish biology and fisheries, including in the NE Atlantic where the information available to science and advisory processes gradually increased (e.g. Bergstad 2013). The worldwide research effort and literature on the vulnerability of resources on seamounts, ridges and slopes, is very extensive (e.g. Clark et al. 2007 and recently e.g. Rogers 2018, Victorero et al. 2018). As noted ICES provides regular advice on deep-sea species and fisheries in the NE Atlantic (http://ices.dk/community/advisory-process/Pages/Latest-Advice.aspx), and as time-series of data have grown and the biological understanding has increased, the advice statements have become more robust and relevant. For many typical seamount species that were once considered major promising targets, the advice is now much more precautionary, with either recommending very low catches compatible with low production rates, or advice to ban fisheries or targeting of certain species/stocks/life stages (e.g. spawning fish and juveniles), and even to minimize by-catches in fisheries targeting other species. For orange roughy a zero catch has been recommended for many years. For alfonsinos the recommended total allowable catch has been reduced to a level essentially only compatible with a limited fishery, mostly in the Azores where the species is fished in handline and longline fisheries. Trawl fisheries in international waters for that species have ceased.
10 - Threats
The concept of ‘Vulnerable Marine Ecosystems’ as referred to above originated in the context of fisheries, i.e. in the UNGA resolutions on sustainable fisheries, and reflects the specific concern that fisheries may adversely impact other ecosystem components. The term ‘vulnerable’ refers to sensitivity to a particular activity, i.e. fishing, and not all other disturbances that may occur due to human activity in the sea. This is often not fully appreciated. Species, habitats and seamounts ecosystems are, however, affected by a number of stressors in addition to fishing, e.g. ocean warming, ocean acidification, pollution, as well as emerging industries such as mineral extraction, and possibly oil and gas production. The impacts of any stressor causing reduced fitness in seamount biota must, however, be evaluated and viewed in combination with other stressors acting on the seamount ecosystems. Where in-situ information on the health condition of seamount ecosystems are lacking, laboratory studies (exposing species that are commonly associated with seamounts to anthropogenic stressors) can be used to harvest important information on the probable impact of e.g. changing climate and anthropogenic industrial activity on seamount ecosystem health condition.
10.1 - Fishing
The significance of fishing as a threat has been described above and will not be repeated extensively here. Since the 1960s onwards, many seamounts worldwide have been subject to bottom trawling, often targeting aggregating long-lived fish species with comparatively low fecundity and slow growth, which has resulted in a boom and bust fishery with very high catches at first which then quickly reduce to almost zero (Victorero et al. 2018). Such fisheries often had two major consequences; 1) the depletion of target and non-target fish species and 2) the destruction of structurally complex and fragile benthic habitats such as coral reef, coral gardens and sponge aggregations. The relatively recent management efforts to reduce the risk of such adverse impacts of fisheries seamount biota within the OSPAR maritime area are described in a subsequent chapter. Compared with the expansive and exploratory period in the 1960s to 1990s, the attitude in the fishing industry and society at large has changed considerably, and undesirable practices have largely been abandoned. Deep-sea fisheries, including most on seamounts, declined to low levels, for multiple reasons. In some cases the reason was a decline in abundance of target resource, in other cases deep-sea fisheries proved unprofitable or ceased for other socioeconomic and political reasons. A primary example is the pronounced and abrupt decline in the major operations conducted by the USSR and eastern European fleets prior to the political changes in the late 1980s. A similar decline happened much later for western European fleets that largely pulled out of the deep-sea fisheries after 2000 when the EU and states such as Norway, Iceland and the Faroes introduced new management measures. The threat to seamount biota from fishing is thus likely to be much lower at present than previously when fisheries were more profitable and under weaker management.
10.2 - Ocean acidification
As CO2 levels in the oceans are rising, pH and carbonate ion (CO23-) availability are gradually reducing. Carbonate chemistry is changing in the entire global ocean, but with polar areas being affected at a higher pace than other areas. This due to to the fact that CO2 is more easily absorbed into cold water and that polar waters are experiencing an increasing fresh water input due to melting ice, which reduces the buffering capacity of the ocean. Within OSPAR region I (i.e. the Norwegian & Icelandic Seas) changes are already now occurring at measurable rates and seamounts within this area are therefore likely among the OSPAR seamount that will be first hit by ocean acidification. Rapid expansion of areas undersaturated with respect to calcium carbonate have been detected (Olafsson et al. 2009, AMAP 2013, Qi et al. 2017) and pH levels in the Norwegian Sea are decreasing at one of the highest rates globally (Chierici et al. 2017). The aragonite saturation horizon in the Iceland Sea is shoaling with 4 m per year (Olafsson et al. 2009). Although not assessed in the field these changes will likely cause severe impacts to the fitness as well as the distribution of fauna building skeletal structures from aragonite and spognes that are simple organisms with very limited capacity for acid base regulation (de Bruin 2017). Interestingly, modelling efforts have shown that under future projected ocean pH and aragonite saturation seamount summits and upper slopes may provide a refugia for e.g. cold-water scleractinian corals (Tittensor et al. 2010). While the seamount summits are likely to be impacted by changes in carbon chemistry, they are less affected, and they consistently provide a more suitable habitat than the surrounding seafloor, mainly because they lie in shallower waters with a higher aragonite saturation state. The potential of seamounts to work as a refuge for ocean acidification is dependent upon the ability of larvae and adults of mobile species to disperse vertically. Carefully monitoring of health condition of these northern seamounts are desirable.
10.3 - Ocean warming
CO2 emissions are causing both atmosphere and oceans to warm. Heat content of the North Atlantic surface layers has increased significantly in the last 60 years (Lee et al. 2011). However, during the last two decades water at 300 to 1500 m has also been warming (Chen & Tung 2014, Somavilla et al. 2016) potentially also exposing seamount species and ecosystems to waters of warmer temperatures. Ocean warming is expected to have a profound impact on all marine ecosystems as increased temperature can affect the fitness of specimens, competitive interactions between species and eventually the geographical distributions of species (see e.g. Frainer et al. 2017, Pessarrodona et al. 2018). However, seamounts generally have extensive depth profiles which could provide refugia from higher temperatures. Indeed, paleontological records demonstrate that in past history coral growth on seamounts has varied to a great extent with periods of gapid growth ingerchanged with period of growth hiatuses. Rapid coral growth on SW Rockall Bank and in Porcupine Seabight are clearly related to overall climatic warm phases (Frank et al. 2009). Long-term monitoring of health condition of speciments as well as the vertical species distrubitons of the OSPAR seamounts in light of climate change are desirable.
10.4 - Mineral extraction
Seamounts offer a significant source of minerals in the deep-sea. Cobalt-rich ferromanganese crusts are commonly associated with exposed hard rock on seamounts (Hein et al. 2010). Furthermore, polymetallic sulfide deposits resulting from hydrothermal venting can be found along the Atlantic and Arctic Mid Ocean Ridge as well as the Azores archipelago and seamounts (Boschen et al. 2013, Pedersen et al. 2010). To date there is no commercial extraction of minerals in the deep-sea. However, the interest for the activity is growing. Deep-sea mining, if commenced, is likely to cause severe local impact to deep-sea habitats. The loss of hard substrate may cause substantial shifts in benthic community composition (Gollner et al. 2017) and life-history characteristics of fauna associated with seamounts (e.g. slow growth and infrequent recruitment) imply that recovery of damaged habitats may take very long (Schlacher et al. 2014). Suspended particulate waste generated during excavation can be transported to more distant locations and affect marine life both when suspended and when settling to the sea floor. It´s well known that sessile filter feeders such as sponges and corals possess mechanisms to deal with temporarily increased suspended particle loads and sedimentation rates, e.g. reduced or arrested pumping and mucus production, but that these can be costly to apply and result in reduced energy stores and growth (Larsson and Purser 2011, Larsson et al. 2013, Tjensvoll et al. 2013, Kutti et al. 2015). Furthermore, deep-sea corals are highly sensitive to heavy metals that can be elevated in the sediment plumes generated during excavation (Martins et al. 2018).
10.5 - Litter
Litter is emerging as an issue in the deep-sea as well as in coastal areas. Woodall et al. (2015) analysed video data from a range of seamounts in the Atlantic and Indian Ocean and found, to some surprise, litter on all surveyed seamounts. The litter was composed mainly of plastic, metal, fishing gear and glass and was patchily distributed, with a mean of 4.5 items of litter per hectar in Atlantic seamounts. Most litter is generally found on the summits of the seamounts with an average of 14 items of litter per hectar on the Condor seamount (Pham et al. 2013) and 0.04 items per hectar on the Gorringe bank (Vieira et al. 2015). While plastics is the most commonly found type of litter in the NE Atlantic in general, litter on seamounts is prevailingly of lost fishing gear (lines and weights) (Pham et al. 2013, 2014, Vieira et al. 2015).
10.6 - Cumulative impact
Cumulative impacts to seamount biota are predicted to occur mainly as a result of climate change related pressures (e.g. ocean acidification and warming) in combination with industrial activities such as fishing and mineral and energy excavation/production. Predicting the cumulative impacts from human activities in the deep-sea is difficult as there is limited knowledge on the effect of the different stressors in isolation and only very few studies have assessed how two, let alone three, stressors interact. It is generally assumed that cumulative impacts are additive (Agbayani et al. 2015), however, this is not always the case. For the sponge Geodia atlantica Scanes et al. (2018) found that elevated suspended sediments caused cellular stress and reduced respiration rates. However, a combined treatment of increased suspended sediments and warming did not affect the levels of cellular stress and increased respiration. For the cold-water coral Primnoa resedaeformis Scanes et al. (2018) found that suspended sediments reduced O:N ratios, while warming increased respiration, nitrogen excretion, and cellular stress which resulted in lower O:N ratios. Suspended sediment and warming can act alone or interact to cause significant negative impact to a range of deep-sea species, however, responses are likely to be species-specific.
11 - Knowledge gaps
In the relatively recent review of seamounts in the OSPAR maritime area (Morato et al. 2013) in-situ information on either geology, biology or bathymetry from scientific cruises was found from 37 seamounts or seamount like features. We collated information on biology only and found data from 28 seamounts or seamount like features. 33 papers documenting the biological life around seamounts within the OSPAR maritime area had been published between 2013 and 2019, indicating a slow but continuous increase in knowledge. However, the overlay of modelled and studied seamounts revealed that in total only 17% of the modelled seamounts in the OSPAR maritime areas have been the subject to biological investigations of any kind illustrating the overall poor documentation of OSPAR seamounts. For the seamounts that have been subjected to scientific investigations the majority are single discipline studies looking into one or few components of the ecosystem. Total biodiversity is unknown for most seamounts and density estimates are rare. There is a major shortage of information on processes and functions and the roles of seamounts at regional scales, e.g. as stepping stones for species across wider ocean areas. Such knowledge is critically needed to inform conservation and management strategies.
Within the OSPAR maritime area 100 chartered and 161 modelled seamounts are present, indicating that only 62% of the potential seamounts in the area have been documented by ship-based surveys. Furthermore, for many there is a discrepancie between described postion of chartered and modelled seamounts. Accurate information on location of seamounts is chritically needed for spatial planning and management. Improved data collection on topography from ship based multi-beam surveys is therefore needed.
Species, biotopes and seamounts ecosystems can be subjected to anthropogenic disturbance caused by fishing and pollution, emerging industries such as mineral extraction, as well as climate change. Appart from a few studies documenting the occurrence of litter on seamounts and fishing disturbance there is little in-situ knowledge of health status of seamount ecosystems and no long-term monitoring of seamount ecosystems. Establishing long-term monitoring in a few selected seamounts would allow the evaluation of changes in the health condition and inform mamangments accordingly.
12 - Management and conservation of seamounts
The mandate to manage human activities in seamount habitats in the relevant subarea of the Atlantic Ocean is divided amongst 1) national governments managing activity their respective areas under national jurisdiction, and 2) intergovernmental organizations (IGOs) managing activity beyond national jurisdiction (ABNJ). Several national governments have submitted extended continental shelf claims to the UN, and these may affect the management of seamount features associated with the seabed even if the features are currently under the mandate of an IGO. Human activities that are or may become particularly relevant for seamounts in the OSPAR maritime area include fishing, seabed mineral extraction and bioprospecting. The latter two are still in an exploratory and prospecting phase. In the following a brief account is given for fishing and mineral extraction.
12.1 - Living marine resources
Fishing on seamounts has a long history and has attracted increasing management attention during the last two decades. National authorities or the EU regulate fishing activity on seamounts inside the Exclusive Economic Zones (EEZs). This is facilitated by a range of measures including technical regulations, total allowable catches (quotas), and spatial management in the form of temporal and permanent fishing closures. The EU also sets catch limits for union vessels fishing outside Union waters. However, beyond the EEZs, the intergovernmental regional fisheries management organizations (RFMOs) have the mandate to regulate fisheries, and they implement legally binding measures applying to all contracting parties as well as third parties. Most pertinent to the issues of habitats and biota are the RFMOs dealing with bottom fishing, i.e. fishing for benthic and benthopelagic resources with gear that is likely to contact the seafloor during the normal course of fishing operations.
In the Northeast Atlantic, the most relevant RFMO is the Northeast Atlantic Fisheries Commission (NEAFC). NEAFC has been attentive to international calls (e.g. reflected in UN General Assembly resolutions) for enhanced protection of benthos and fisheries resources often associated with seamounts. Since 2004 the Commission, with the support of scientific advice from the International Council for the Exploration of the Sea (ICES), progressed towards the present set of regulations protecting Vulnerable Marine Ecosystems (VMEs) (sensu FAO 2009) as well as deep-sea fisheries resources. The updated list of regulations are available on https://www.neafc.org/managing_fisheries/measures/current, and the more pertinent for VME-protection is the Rec. 19:2014, further illustrated on the site https://www.neafc.org/managing_fisheries/vmec. Included in Rec. 19:2014 are measures restricting bottom fishing to designated limited subareas named ‘existing fishing areas’ that are open to fishing under certain conditions set by other ‘resource-specific measures’ (incl. those pertinent to all deep-sea species, and specific measures for blue ling, macrourids, orange roughy, sharks, rays, chimaeras, a.o., and also a general ‘gillnet ban’ for deep-sea fisheries). Other subareas are only open to pre-evaluated exploratory fishing accepted by the Commission. The third category are subareas fully closed to bottom fishing. These multiple fishing closures, introduced in both previously fished and unfished areas, were designated to protect known VMEs or representative geomorphological features likely to have VMEs, e.g. seamounts and mid-ocean ridge sections. On Figure 8 the present set of ‘existing fishing areas’ and fishing closures are shown as green and red areas, respectively. The fishing closures include a number of seamounts such as Altair, Antialtair, numerous seamounts on the mid-Atlantic Ridge, and the Edora Bank to the southwest of the Rockall-Hatton banks where large bank and slope areas have been closed. The Rec. 19:2014 also applies to the parts of the NEAFC ‘regulatory area’ in the Norwegian Sea, the Barents Sea and the Arctic Ocean but there are no seamount fisheries in those areas.
It should be noted that one of the ‘existing fishing areas’ maintained by NEAFC encompasses the Josephine Seamount in the very southeastern corner of the ‘Regulatory Area’. Despite advice from ICES suggesting that the area is likely to have VMEs, the commission did not yet include it amongst the fishing closures listed in Rec. 19:2014.
ICES is requested to provide advice on the need for further action should new science reveal a need for amendments, and the VME-closures were recently reviewed by ICES and found appropriate. The Rec 19:2014 includes a review clause, i.e. a date by which the VME-closures shall be evaluated. The rule is, however, that closures should be maintained unless there is evidence suggesting they are redundant.
Bottom fishing in deeper parts of the NEAFC ‘Regulatory area’ has declined to very low levels (see https://www.neafc.org/international/22299 ). Furthermore, most remaining fisheries are conducted with midwater gears that at most touch the bottom accidentally. This is also the case for the few and small fisheries that are currently conducted on seamounts.
Figure 8. The NEAFC ‘Regulatory area’, i.e. the area beyond EEZs in the North Atlantic with subareas designated as ‘existing fishing areas’ (green), and as fishing closures (red) (NEAFC Recommendation 19:2014). The yellow area is only open to exploratory fishing if accepted by the commission, but no such exploration was ever conducted and the subarea has proven de facto closed. The remainder of the NEAFC ‘Regulatory Area’ in the Norwegian Sea, Barents Sea and the Arctic Ocean are also included in Rec. 19:2014 but are not as relevant for this report because no seamount fisheries are conducted in those areas. The purple box is the ‘haddock box’ where bottom fishing is restricted to protect Rockall haddock.
As mentioned above, bottom fishing within EEZs are regulated by either the Common Fisheries Policy (for EU members) or national regulations, and some of the national regulations mimic the NEAFC measures. These are relevant for many seamounts in the OSPAR Maritime Area, but further details on individual actions taken by states have not been included here. In 2016, the European Parliament agreed to implement new fisheries legislations for Union vessels which among other provisions included a ban on bottom trawling in waters deeper than 800 m, with the objective to provide protection to vulnerable marine species and habitats.
Pelagic fisheries are major operations in the OSPAR maritime area, i.e. targeting species such as herring, mackerel, blue whiting and the redfishes. These fisheries are regulated by national and NEAFC measures. Seamounts do not appear to be particularly significant for the target species nor the fisheries for them.
Inasmuch as seamounts are significant for high-seas fisheries for highly migratory species such as tuna and tuna-like species, including pelagic sharks, these fisheries are regulated by the International Commission for the Conservation of Atlantic Tunas (ICCAT, https://www.iccat.int/en/# ). Similarly, marine mammal management is the mandate of the International Whaling Commission (IWC). The North Atlantic Marine Mammal Commission (NAMMCO) advises the nations regulating whaling operations being conducted in the area relevant for this report.
12.2 - Mineral extraction activity
Mining activity on seamounts in the OSPAR Maritime Area is a potential rather than a realized activity. In areas beyond national jurisdiction, the International Seabed Authority (ISA) has the mandate to regulate exploration and, should it emerge, exploitation. Otherwise, national legislation will apply. The issues of regional activity planning, regional Strategic Environmental Plans, and Environmental Plans for specific projects are being discussed nationally and in the ISA, and this activity is also significant for future activity on seamounts. ISA has granted exploratory licenses for an area on the mid-Atlantic Ridge south of the Azores, but not for any seamounts in the OSPAR Maritime Area.
12.3 - OSPAR Marine protected areas
At the Ministerial Meeting in Sintra in 1998, OSPAR Ministers agreed to promote the establishment of a network of marine protected areas. Following a period of preparatory work, the 2003 OSPAR Ministerial Meeting in Bremen adopted Recommendation 2003/3 on a network of marine protected areas with the purpose of establishing an ecologically coherent network of well-managed MPAs in the North-East Atlantic. Since then, many MPAs have been established within EEZs and some outside, and a subset of these MPAs encompass seamounts.
OSPAR raises awareness on activities that may adversely affect seamount habitats or biota. This is one of the primary functions of the MPAs that have been established. Management objectives are being developed and efforts are made to facilitate implementation of effective measures. However, OSPAR does not have the mandate to regulate activities such as fishing or mining, and in order to facilitate the implementation of binding measures OSPAR must work with states and international organizations that have the relevant mandates. An example of such processes is the near parallel introduction of NEAFC VME closures and OSPAR high-seas MPAs in the mid-Atlantic. The NEAFC closures (and other spatial measures) predated the MPAs, but subsequently there has been a lot of interactions between the organizations regarding new measures. Two seamount features selected as MPAs are not within NEAFC VME closures, e.g. the Milne and Josephine Seamounts. This probably reflects that NEAFC has given first priority to areas with active fishing such as the Rockall-Hatton, less to seamounts where only pre-evaluated fishing is allowed and where past activity has more or less ceased.
NEAFC and OSPAR has signed a MoU that facilitates discussions and awareness. OSPAR and NEAFC furthermore jointly promotes collaborative links with other IGOs with mandates to regulate shipping (IMO) and mining (ISA) in the Northeast Atlantic and invites other organizations to join the ‘Collective Arrangement’.
Many seamounts inside EEZs have been established as national MPAs and feature in the OSPAR MPA repository (http://mpa.ospar.org/home_ospar ). The detailed national management actions introduced for these areas have not been recorded here. Given that activity levels are demonstrably higher inside than outside EEZs, measures applying to seamounts under national jurisdiction are highly relevant and important. An example of MPAs established within EEZs is the Formigas Dollabarat in the Azores with many commercial species, such as blackspot Seabream (Pagellus bogaraveo), conger (Conger conger) and forkbeard (Phycis phycis). This was declared as an MPA in 2006, prohibiting all commercial and recreational fishing. The declaration as an MPA has, however, had a marginal effect. Overall, a lack of response or even a reduction in abundance and fish size was the most common trend, with few cases of positive responses (Afonso et al. 2018). Landings of major commercial species on the seamounts Anton Dohrn and Rosemarys declared by the UK as MPAs has declined and although no formal assessments are made, CPUE data from surveys suggest declining abundance in many species (Neat et al. 2008). On Condor de Terra, again in the Azores, all demersal fisheries have been closed since 2010 after a pronounced decline in targeted species, such as blackspot seabream (Pagellus bogaraveo) and Atlantic wreckfish (Polyprion americanus). This area is known as the most accessible seamounts for the scientific community and is an internationally recognized study area of reference (Menezes et al. 2011) offering an opportunity for studying effects of no-take areas as a means of regulating fisheries and rebuilding/maintaining resources.
13 - Significance of seamounts for threatened species
Nine species of fish that weere recorded on the OSPAR seamounts (Appendix 3) are on the OSPAR list of threatened and/or declining species. These are Thunnus thynnus (bluefin tuna) found on the Condor Seamount, Squalus acanthias (spurdog) caught on the Faroe Bank, Rostroraja alba (white skate) caught on the Josephine Bank, Raja clavata (Thornback skate) caught on Condor Seamount, Faroe Bank, Josephine Bank and Le Danois Bank, Hoplostethus atlanticus (orange roughy) caught on Le Danois Bank, Gorringe ridge, Faraday Seamount and Sedlo Seamount, Gadus morhua (cod) caught on the Faroe Bank, Dipturus batis (common skate) caught on the Condor Seamount, Gorringe ridge and Le Danois Bank, Centrophorus granulosus (Gulper shark) caught on the Sedlo Seamount and Gorringe ridge and Centroscymnus coelolepis (Portuguese dogfish) that has been caught on the Hatton Bank, Condor Seamount, Sedlo Seamount, Gorringe ridge, George Bligh Bank and Le Danios Bank.
Furthermore, 18 of the species that we registered on the OSPAR seamounts are listed as threatened by the IUCN: Sphyrna zygaena (Smooth hammerhead) on Joao de Castro, Bodianus scrofa (Barred hogfish) on Formigas Dollabarat and Dom Joao de Castro, Thunnus obesus (Bigeye tuna) on Condor de Terra and Dom Joao de Castro, Thunnus thynnus (Blue-fin tunny) on Condor de Terra, Mycteroperca fusca (Island grouper) on Formigas Dollabarat, Carcharhinus longimanus (Oceanic whitetip shark) on Condor de Terra, Galeorhinus galeus (Tope shark) on Faroe Bank and Condor de Terra, Isurus oxyrinchus (Shortfin mako) on Condor de Terra and Galicia, Mobula birostris (Giant manta) on Formigas Dollabarat and Gorringe bank, Mobula tarapacana (Chilean devil ray) on Formigas Dollabarat, Dipturus batis (Blue skate) on Condor de Terra, Galicia Bank and Le Danois bank, Leucoraja circularis (Sandy ray) on Faroe Bank and Le Danois bank, Raja maderensis (Madeiran ray) on Gorringe bank, Rostroraja alba (Bottlenosed skate) on Josephine seamount, Centrophorus squamosus (Leafscale gulper shark) on Condor de Terra, Sedlo and Galicia bank, Dalatias licha (Kitefin shark) on Condor de Terra, Sedlo, Galicia bank and Le Danois Bank, Centroscymnus owstonii (Roughskin dogfish) on Condor de Terra and Sedlo and Squalus acanthias (Piked dogfish) on Faroe bank. Cartilaginous fish are overrepresented on the list of both OSPAR and IUCN (6 out of 9 and 12 out of 18, respectively). Cartilaginous fish are particularly vulnerable to overexploitation due to their slow growth and low reproduction rates. In the North Atlantic region cartilaginous fish are both directly and indirectly taken by numerous commercial fisheries and incidental capture is one of the main sources of mortality for these species (Bonanomi et al. 2017). Many of the mentioned species are affected by fishing and subjected to management by an international or national fisheries authority. In the North East Atlantic, thornback ray (Raja clavata), spurdog (Squalus acanthias), leafscale gulper shark (Centrophorus squamosus) and Portuguese dogfish (Centroscymnus coelolepis) are known to constitute a significant bycatch component of direct fisheries (Bonanomi et al. 2017).
Four habitats associated with OSPAR seamounts, i.e., hydrothermal vents, Lophelia pertusa reefs, deep-sea sponge aggregations and coral gardens, are on the OSPAR list of threatened and/or declining habitats. Hydrothermal venting is known to occur on the Dom Joao de Castro Bank and Gigante seamount. Coral gardens have been documented on the Acor Bank, Anton Dohrn Seamount, Condor de Terra Seamount, Galicia Bank, Josephine Bank and the Ormonde Seamount. Lophelia pertusa coral reefs have been documented from Anton Dohrn Seamount, Galicia Bank and George Bligh Bank and deep-sea sponge aggregations have been documented from the Anton Dohrn Seamount, Condor de Terra Seamount, Galicia Bank, Josephine Bank, Ormonde Seamount, Rosemary Seamount and Schultz massif. In addition, sea-pens (Pennatulacea) are found on Anton Dohrn Seamount, Condor de Terra Seamount, George Bligh Bank and Ormonde seamount. Whether they form sea-pen and burrowing megafauna communities is unknown since density estimations and extent of distribution from these areas are not known. Furthermore, xenophyophore assemblages are documented from the Anton Dohrn Seamount, Condor de Terra Seamounts, Hebrides Terrace and Sedlo Seamount and communities of stylasterids can be found on the Acor Bank, Condor de Terra Seamount, Josephine Bank and the Ormonde Seamount. Although not on the OSPAR list of threatened and declining habitats, both taxa are considered highly sensitive to physical disturbance and occur on the list of groups of species that often contribute to forming vulnerable marine ecosystems (FAO 2009).
The OSPAR list of threatened and/or declining mollusc species are mainly shallow water molluscs, hence only seamounts with shallow peaks are likely to harbor species from the list. Only 2% of the OSPAR seamounts modelled by Harris et al. (2014) belong to this group. Ormonde peak on the Gorringe ridge, Princess Alice Bank, Dom Joao de Castro Bank and Formigas Bank in the Azores archipelago are examples of seamounts with shallow peaks. Mollusc fauna has been studied explicitly on Dom Joao de Castro Bank and Formigas Bank yielding documentation on the occurrence of Patella aspera on both (Avila & Azevedo 1997, Avila et al. 2004). None of the Molluscs on the OSPAR list of threatened and/or declining species and habitats, i.e. Patella aspera, Ostera edulis, Nucella lapillus and Arctica islandica, have been documented on the shallow Ormonde peak (Ramos et al. 2015 and the oceana project: https://eu.oceana.org/sites/default/files/reports/seamounts_gorringe_bank_eng.pdf). One crustacean species, i.e. Megabalanus azoricus, an abundant barnacle found mainly on Azores islands (but also Madeira and the Canary Islands) were it is harvested for human consumption, is also listed as threatened and declining. The list of threatened and declining species does not contain any cnidaria or porifera species (i.e. coral and sponges). Due to the limited knowledge on taxonomy, occurrence and distribution of all but the very common deep-sea coral and sponges it is virtually impossible to evaluate their status. The most prominent large, common and structure forming deep-sea corals and sponges are represented on the list of threatened and declining species and habitats though the habitats they construct, i.e. coral gardens, coral reefs and deep-sea sponge aggregations. Exceptions to this are Solenosmilia variabilis and Madrepora oculata reefs that are not included as a coral reef habitat.
14 - Conclusions
This review demonstrated that despite a rather extensive research effort the last two decades (33 biological papers from OSPAR seamounts published between 2013 and 2019) there is still a lack of quantitative information on plankton, benthos, fish and mammals even from the most well studied seamounts. Comparisons between seamount ecosystems and adjacent areas are almost completely lacking and overall, most seamounts in the OSPAR maritime area have not been subjected to any scientific investigations at all.
Based on existing published studies we conclude that species composition of seamounts, when it comes to plankton, benthos, fish and mammals, reflects that of the regional species pool. Open ocean seamounts constitute small and presumably isolated living-spaces for species that occur on the slopes of the continental and islands. This widens the ranges of some species, but too few studies of connectivity have been carried out to draw any conclusions regarding the significance of seamounts as ‘stepping stones’. Species richness may be elevated compared to the surrounding sea-bed primarily due to a large habitat heterogeneity but is similar to other geomorphologically heterogenous features such as e.g. ridges and canyons. The distinct physical flow-features generally associated with seamounts does not seem to result in a locally elevated zooplankton abundance. However, many seamount are indeed characterized by large communities of sessile filter and suspension feeding organisms, such as corals and sponges. Typical features of seamount summits, especially in Region V, are shoals of benthopelagic fish such as alfonsino, orange roughy and grenadiers, as well as mesopelagic scattering layers of a mixture of invertebrates and fish that impinge on the summits and slopes during daytime. Several groups of deep diving whales and seals (i.e. Mesoplodon, Hyperoodon, Ziphius, Physeter and Cystophora) are commonly sighted at seamounts, however, most sightings of mammals are from coastal areas of the European mainland and the large islands and island groups of the NE Atlantic hence it is unlikely that seamount ecosystems are highly significant habitats for these species.
Seamount fisheries have, in the past, represented a serious threat to several species of commercially targeted fish, by-catch species and vulnerable benthic habitats but fishing is now largely strictly managed and has become a much smaller threat to seamount ecosystems as compared to 10 to 20 years ago. A major concern is, however, the slow recovery of fish stocks in previously exploited seamounts now protected from fishing. A range of threatened species (such as orange roughy, several species of elasmobranchs and corals) utilize seamount as a habitat and are likely to benefit from the protection of seamounts from adverse impacts of anthropogenic activity. Long-term monitoring or at least repeat studies are, however, needed to evaluate recovery, that may take decades or longer.
15 - Reference lists
15.1 - References used in the plankton chapter:
Arístegui J., Mendonça A., Vilas JC., Espino M., Polo I., Montero MF., Martins A. 2009. Plankton metabolic balance at two North Atlantic seamounts. Deep-Sea Research II, 56: 2646-2655
Pierrot-Bults AC (2008). A short note on the biogeographic patterns of the Chaetognatha fauna in the North Atlantic. Deep Sea Research II, 55:137-141.
Cardigos F., Colaço A., Danda P.R., et al. 2005. Shallow water hydrothermal vent field fluids and communities of the D. Joao de Castro Seamount (Azores). Chemical Geology, 224: 153-168
Carmo V., Santos M., Menezes G.M., Loureiro C., Lambardi P., Martins A. 2013. Space and time variability of zooplankton communities at Condor seamount, Azores (NE Atlantic). Deep-Sea Research II, 98: 63-74
Cartes JE., Serrano A., Velasco F., Parra S., Sánchez F. 2007. Community structure and dynamics of deep-water decapod assemblages from Le Danois Bank: Influence of environmental variables and food availability. Progress in Oceanography, 75: 797-816
Colaço A., Giacomello E., Porteiro F., Menezes G.M. 2013. Trophodynamic studies on the Condor seamount (Azores, Portugal, North Atlantic). Deep-Sea Research II, 98: 178-189
Gaard E, Gislason A, Falkenhaug T, Søiland H, Musaeva E, Vereshchaka A, Vinogradov G (2008) Horizontal and vertical copepod distribution and abundance on the Mid-Atlantic Ridge in June 2004. Deep Sea Research II, 55:59-71.
González-Pola C, del Río G.D, Ruiz-Villarreal M, Sánche R.Fz, Mohn C. (2012). Circulation patterns at Le Danois Bank, an elongated shelf-adjacent seamount in the Bay of Biscay, Deep Sea Research Part I: Oceanographic Research Papers, 60: 7-21.
Heger A., Ieno, E.N., King NJ., Morris KJ., Bagley PM., Priede IG. 2008. Deep-sea pelagic bioluminescence over the Mid-Atlantic Ridge. Deep-Sea Research, 55: 126-136.
Hesthagen, I. H. 1970a. On the near-bottom plankton and benthic invertebrate fauna of the Josephine Seamount and the Great Meteor Seamount. Meteor Forschungsergebnisse D8: 61-70, 1970
Hesthagen, Ivar H. 1970b. The near-bottom plankton and benthic invertebrate fauna of Josephine and Great Meteor Seamounts (Table 2). PANGAEA, https://doi.org/10.1594/PANGAEA.611095
Hirch S., Kiriakoulakis K., Koppelmann R., Peters J., Christiansen B. 2009a. Food sources and diets of zooplankton at two seamounts in the NE Atlantic: Are there seamount effects? In: Hirch S., Trophic interactions at seamounts, PhD Dissertation, University Hamburg.
Hirch S., Martin B., Christiansen B. 2009b. Zooplankton metabolism and carbon demand at two seamounts in the NE Atlantic. Deep-Sea Research II, 56: 2656-2670
Hosia A, Stemmann L, Youngbluth M (2008) Distribution of net-collected planktonic cnidarians along the northern Mid-Atlantic Ridge and their associations with the main water masses. Deep Sea Research II, 55:106-118
Hosia A, Falkenhaug T, Baxter EJ, Pagès F. (2017) Abundance, distribution and diversity of gelatinous predators along the northern Mid-Atlantic Ridge: A comparison of different sampling methodologies. PLoS ONE 12(11): e0187491.
Kiriakoulakis K., Vilas JC., Blackbird SJ, Arístegui J., Wolff GA. 2009. Seamounts and organic matter – Is there an effect? The case of Sedlo and Seine seamounts, Part 2. Composition of suspended particulate organic matter. Deep-Sea Research II, 56: 2631-2645
Letessier TB, Falkenhaug T, Debes H, Bergstad, OA, Brierley A (2011). Abundance patterns and species assemblages of euphausiids associated with the Mid-Atlantic Ridge, North Atlantic. Journal of Plankton Research 33:1510-1525.
Martin B., Christiansen B. 2009. Distribution of zooplankton biomass at three seamounts in the NE Atlantic. Deep-Sea Research II, 56: 2671-2682
Olivieira AP., Coutinho TP., Cabeçadas G., et al. 2016. Primary production enhancement in a shallow seamount (Gorringe – northseat Atlantic). Journal of Marine systems, 164:13-29
Reid PC., Colebrook JM., Matthews JBL., Aiken J. 2003. The Continuous Plankton Recorder: concepts and history, from plankton indicator to undulating recorders. Progress in Oceanography, 58: 117-173.
Santos M., Moita M.T., Bashmachnikov I., Menezes G.M., Carmo V., Loureiro CM., Mendonça A., Silva AF., Martins A. 2013. Phytoplankton variability and Oceanographic conditions at Condor seamount, Azores (NE Atlantic). Deep-Sea Research II, 98: 52-62.
Sobrinho-Gonçalves L. and Cardigos F. 2006. Fish larvae around a seamount with shallow hydrothermal vents from the Azores. Thalassas-An International Journal of Marine Science, 22: 19-28
Tokarev, Yu. N., Bityukov, E. P., Williams, R. 2003. Influence of seamounts at the Atlantic Ocean on modification of the bioluminescence and plankton characteristics. Marine Ecological Journal 2:46-58. http://repository.marine-research.org/handle/299011/697
Stemmann, L, A. Hosia, M.J. Youngbluth, H. Søiland, M. Picheral and G. Gorsky 2008. Vertical distribution (0–1000 m) of macrozooplankton, estimated using the Underwater Video Profiler, in different hydrographic regimes along the northern portion of the Mid-Atlantic Ridge. Deep Sea Research II, 55: 94-105
Weigmann, R. 1974a. Untersuchungen zum Vorkommen der Euphausiaceen (Crustacea) im Bereich der Großen Meteorbank. Meteor Forschungsergebnisse, Deutsche Forschungsgemeinschaft, Reihe D Biologie, Gebrüder Bornträger, Berlin, Stuttgart, D17, 17-32
Weigmann, R. 1974b. Relative abundance of euphausiids (Crustacea) in water samples of the Josephine Seamount, Atlantic Ocean (Table 8). PANGAEA, https://doi.org/10.1594/PANGAEA.611406
Youngbluth, M., T. Sørnes, A. Hosia and L. Stemmann 2008. Vertical distribution and relative abundance of gelatinous zooplankton, in situ observations near the Mid-Atlantic Ridge. Deep Sea Research II, 55: 119-125.
15.2 - References used in the benthos chapter:
Becker JJ, Sandwell DT, Smith WHF, Braud J, Binder B, Depner J, Fabre D, Factor J, Ingalls S, Kim SH, Ladner R, Marks K, Nelson S, Pharaoh A, Trimmer R, Von Rosenberg J, Wallace G, Weatherall P (2009) Global Bathymetry and Elevation Data at 30 Arc Seconds Resolution: SRTM30_PLUS, Marine Geodesy, 32:355-371, DOI: 10.1080/01490410903297766
Durden JM, Bett BJ, Jones DOB, Huvenne VAI, Ruhl HA (2015) Abyssal hills – hidden source in increased habitat heterogeneity, benthic megafaunal biomass and diversity in the deep-sea. Progress in Oceanography 137: 209-218.
EMODNet, 2013. European Marine Observation and Data Network, Digital Terrain Model data derived from the EMODnet Hydrography portal. http://www.emodnet- hydrography.eu.
FAO. 2009. International Guidelines for the Management of Deep-sea Fisheries in the High Seas / Directives internationales sur la gestion de la pêche profonde en haute mer / Directrices Internacionales para la Ordenación de las Pesquerías de Aguas Profundas en Alta Mar. Rome/Roma. 73p. www.fao.org/fishery/topic/166308/en.
FAO. 2016. Vulnerable Marine Ecosystems: Processes and Practices in the High Seas, by Anthony Thompson, Jessica Sanders, Merete Tandstad, Fabio Carocci and Jessica Fuller, eds. FAO Fisheries and Aquaculture Technical Paper No. 595. Rome, Italy. www.fao.org/3/a-i5952e.pdf.
Freiwald A, Beuck L, Wisshak M (2012) Korallenriffe im kalten Wasser des Nordatlantiks - Entstehung, Artenvielfalt und Gefährdung. In: Beck E (ed) Die Vielfalt des Lebens. Wiley-VCH, Weinheim, pp 89-96.
Henry LA, Moreno-Nava J, Hennige SJ, Wicks, LC, Vad J, Roberts JM (2013) Cold-water coral reef habitats benefit recreationally valuable sharks. Biological Conservation. 161:67-70.
Henry LA, Stehmann MFW, De Clippele L, Findlay HS, Golding N, Roberts MJ (2016). Seamount egg-laying grounds of the deep-water skate Bathyraja richardsoni. Journal of Fish Biology 89: 1473-1481.
Klitgaard AB (1995) The fauna associated with outer shelf and upper slope sponges (Porifera, Demospongiae) at the Fareo Islands, northeastern Atlantic. Sarsia 80:1-22.
Magnussen E (2002) Demersal fish assemblages of Faroe Bank: Species composition, distribution, biomass spectrum and diversity. Marine Ecoloy Progress Series 238:211-225.
Priede IG, Bergstad OA, Miller PI, Vecchione M, Gebruk A, et al. (2013). Does Presence of a mid-ocean ridge enhance biomass and diversity) PLoS ONE 8(5): e61550, doi:10.1371/journal.pone.0061550
Roberts JM, Henry LA, Long D, Hartley J (2008) Cold-water coral reef frameworks, megafaunal communities and evidence for coral carbonate mounds on the Hatton Bank, north east Atlantic. Facies 54:297-316.
Scanes E, Kutti T, Fang JKH, Johnston EL, Ross PM, Bannister R (2018) Mine waste and acute warming induce energetic stress in the deep-sea sponge Geodia atlantica and coral Primnoa resedeaformis; results from a mesocosm study. Frontiers in Marine Science.
Thoma JN, Pante E, Brugler MR, France SC (2009). Deep-sea octocorals and antipatharians show no evidence of seamount-scale endemism in the NW Atlantic. Marine Ecology Progress Series, 397: 25-35.
Tittensor DP, Baco AR, Hall-Spencer JM, Orr JC, Rogers AD (2010) Seamounts as refugia from ocean acidification for cold
15.3 - References used in the fish chapter:
Abecasis, D., Cardigos, F., Almada, F., & Gonçalves, J. M. S. (2009). New records on the ichthyofauna of the Gorringe seamount (Northeastern Atlantic). Marine Biology Research, 5(6), 605-611. doi:10.1080/17451000902729696
Afonso, P., Schmiing, M., Fontes, J., Tempera, F., Morato, T., & S. Santos, R. (2018). Effects of marine protected areas on coastal fishes across the Azores archipelago, mid-North Atlantic. Journal of Sea Research, 138, 34-47. doi:https://doi.org/10.1016/j.seares.2018.04.003
Bañon, R., Arronte, J. C., Rodriguez-Cabello, C., Piñeiro, C.-G., Punzon, A., & Serrano, A. (2016). Commented checklist of marine fishes from the Galicia Bank seamount (NW Spain). Zootaxa, 4067(3), 293-333.
Bensch, A., Gianni, M., Gréboval, D., Sanders, J. S., & Hjort, A. (2009). Worldwide review of bottom fisheries in the high seas. FAO Fisheries and Aquaculture Technical Paper 522, 145p.
Bergstad, O. A., Menezes, G., & Høines, Å. S. (2008). Demersal fish on a mid-ocean ridge: Distribution patterns and structuring factors. Deep Sea Research Part II: Topical Studies in Oceanography, 55(1), 185-202. doi:https://doi.org/10.1016/j.dsr2.2007.09.005
Cardigos, F., Colaço, A., Dando, P. R., Ávila, S. P., Sarradin, P. M., Tempera, F., . . . Serrão Santos, R. (2005). Shallow water hydrothermal vent field fluids and communities of the D. João de Castro Seamount (Azores). Chemical Geology, 224(1), 153-168. doi:https://doi.org/10.1016/j.chemgeo.2005.07.019
Colaço, A., Giacomello, E., Porteiro, F., & Menezes, G. M. (2013). Trophodynamic studies on the Condor seamount (Azores, Portugal, North Atlantic). Deep Sea Research Part II: Topical Studies in Oceanography, 98, 178-189. doi:https://doi.org/10.1016/j.dsr2.2013.01.010
Council regulation (EU) 2018/2025 of 17 December 2018 fixing for 2019 and 2020 the fishing opportunities for Union fishing vessels for certain deep-sea fish stocks.
Council regulation (EU) 2016/2285 of 12 December 2016 fixing for 2017 and 2018 the fishing opportunities for Union fishing vessels for certain deep-sea fish stocks and amending Council Regulation (EU) 2016/72.
Froese, R., & Sampang, A. (2004). Taxonomy and biology of seamount fishes. In T. Morato & D. Pauly (Eds.), Seamounts: Biodiversity and Fisheries. Fisheries Research Services Internal Report No 02/08. Pp 25-32.
Froese, R., & Sampang, A. (2004b). Appendix2: Preliminary list of Seamount Fishes with Localities. In T. Morato & D. Pauly (Eds.), Seamounts: Biodiversity and Fisheries. Fisheries Research Services Internal Report No 02/08.
Menezes, G. M & Giacomello, E. 2013. Spatial and temporal variability of demersal fishes at Condor seamount (Northeast Atlantic). Deep-Sea Research II 98 (2013):101–113.
Giacomello, E; Menezes, G.M. and Bergstad, O.A. (eds.) (2013). An integrated approach for studying seamounts: CONDOR Observatory. Deep-Sea Research II 98: 1-217.
Gonçalves, J., Bispo, J., & Augusto Silva, J. (2004). Underwater survey of ichthyofauna of Eastern Atlantic Seamounts: Gettysburg and Ormond (Gorringe Bank) (Vol. 51).
Howell, K. L., Davies, J. S., Hughes, D. J., & Narayanaswamy, B. E. (2007). Strategic Environmental Assessment /Special Area for Conservation: Photographic Analysis Report. Department of Trade and Industry, Strategic Environmental Assessment Report, UK. 163p.
Magnussen, E. (2002). Demersal fish assemblages of Faroe Bank: species composition, distribution, biomass spectrum and diversity. Marine Ecology and Progress Series, 238, 211-225.
Maul, G. E. (1976). The Fishes Taken in Bottom Trawls by R.V. Meteor During the 1967 Seamounts Cruises in the Northeast Atlantic. Meteor Forschungsergebnisse (Reihe D 22), 1-69.
Menezes GM and Giacomello E. 2013. Spatial and temporal variability of demersal fishes at Condor seamount (Northeast Atlantic). Deep-Sea Research II 98: 101-113.
Menezes, G., Giacomello, E., Catarino, D., & Rosa, A. (2011). Demersal fish assemblages of the Condor seamount. In E. Giacomello & G. Menezes (Eds.), CONDOR Observatory for long-term study and monitoring of Azorean seamount ecosystems (Vol. Final Project Report. Arquivos do DOP, Série Estudos 1/2012. , pp. 129-147).
Menezes, G. M., Rosa, A., Melo, O., & Pinho, M. R. (2009). Demersal fish assemblages off the Seine and Sedlo seamounts (northeast Atlantic). Deep Sea Research Part II: Topical Studies in Oceanography, 56(25), 2683-2704. doi:https://doi.org/10.1016/j.dsr2.2008.12.028
Menezes, G. M., Rosa, A., Melo, O., & Porteiro, F. (2012). Annotated list of demersal fishes occurring at Sedlo Seamount, Azores north-east central Atlantic Ocean. Journal of Fish Biology, 81(3), 1003-1018. doi:doi:10.1111/j.1095-8649.2012.03355.x
Milligan, R. J., Spence, G., Roberts, J. M., & Bailey, D. M. (2016). Fish communities associated with cold-water corals vary with depth and substratum type. Deep Sea Research Part I: Oceanographic Research Papers, 114, 43-54. doi:https://doi.org/10.1016/j.dsr.2016.04.011
Morato, T., Hoyle, S. D., Allain, V., & Nicol, S. J. (2010). Seamounts are hotspots of pelagic biodiversity in the open ocean. Proceedings of the National Academy of Sciences, 107(21), 9707-9711. doi:10.1073/pnas.0910290107
Narayanaswamy, B. E., Hughes, D. J., Howell, K. L., Davies, J., & Jacobs, C. (2013). First observations of megafaunal communities inhabiting George Bligh Bank, Northeast Atlantic. Deep Sea Research Part II: Topical Studies in Oceanography, 92, 79-86. doi:https://doi.org/10.1016/j.dsr2.2013.03.004
NEAFC 2014: Recommendation on the protection of vulnerable marine ecosystems in the NEAFC Regulatory Area. North-east Atlantic Fisheries Commision (NEAFC). Available at: https://www.neafc.org/managing_fisheries/measures/current (accessed 08.02.2019)
Neat, F., Burns, F., & Drewery, J. (2008). The deepwater ecosystem of the continental shelf slope and seamounts of the Rockall Through: A report on the ecology and biodiversity based on FRS scientific surveys.
OASIS. (2003). Seamounts of the North-East Atlantic. Retrieved from Hamburg and WWF Germany, Frankfurt am Main, November 2003: 38p.
OCEANA. (2005). The seamounts of Gorringe Bank. Available at https://eu.oceana.org/en/publications/reports/seamounts-gorringe-bank-1 (accessed December 2018).
Porteiro, F. M., Giacomello, E., Carmo, V., Graça, G., Cascão, I., Morais, A., . . . Menezes, G. (2011). Mesopelagic fishes around the Condor seamount. In E. Giacomello & G. Menezes (Eds.), CONDOR Observatory for long-term study and monitoring of Azorean seamount ecosystems (pp. 81-91): Arquivos do DOP, Série Estudos 1/2012.
Santos, R., Tempera, F., Colaco, A., Cardigos, F., & Morato, T. (2010). Mountains in the Sea: Dom João de Castro Seamount, Azores (Vol. 23). Oceanography 23 (1): 200-201.
Serrano, A., E Sánchez, J., Cartes, J., Sorbe, S., Parra, I., Frutos, I., . . . Preciado, I. (2005). ECOMARG Project: A multidisciplinary study of Le Danois Bank (Cantabrian Sea, N Spain). ICES CM 2005/P:11.
Sutton, T. T., Porteiro, F. M., Heino, M., Byrkjedal, I., Langhelle, G., Anderson, C. I. H., . . . Bergstad, O. A. (2008). Vertical structure, biomass and topographic association of deep-pelagic fishes in relation to a mid-ocean ridge system. Deep Sea Research Part II: Topical Studies in Oceanography, 55(1), 161-184. doi:https://doi.org/10.1016/j.dsr2.2007.09.013
WWF, 2001. Implementation of the EU Habitats Directive Offshore: Natura 2000 Sites for Reefs and Submerged Sandbanks: Vol. IV The Reefs Inventory.
15.4 - References used in the mammals chapter:
Aoki, Kagari, Katsufumi Sato, Saana Isojunno, Tomoko Narazaki, and Patrick J. O. Miller. 2017. “High diving metabolic rate indicated by high-speed transit to depth in negatively buoyant long-finned pilot whales.” The Journal of Experimental Biology 220 (20): 3802–11. doi:10.1242/jeb.158287.
Archer, F.I. 2002. “Striped dolphin.” In Encyclopedia of Marine Mammals, edited by W.F. Perrin, 1201–3. New York: Academic Press.
Heide-Jørgensen, M.P., D. Bloch, E. Stefansson, L. H. Ofstad, and R. Dietz. 2002. “Diving behaviour of long-finned pilot whales Globicephala melas around the Faroe Islands Diving behaviour of long-finned pilot whales Globicephala melas around the Faroe Islands.” Wildl. Biol. 8 (4): 307–13.
Hooker, Sascha K, and Robin W Baird. 1999. “Deep-diving behaviour of the northern bottlenose whale, Hyperoodon ampullatus (Cetacea: Ziphiidae).” Proceedings of the Royal Society B-Biological Sciences 1 (January): 671–76.
Klatsky, L. J., R. S. Wells, and J. C. Sweeney. 2007. “OFFSHORE BOTTLENOSE DOLPHINS ( TURSIOPS TRUNCATUS ): MOVEMENT AND DIVE BEHAVIOR NEAR THE BERMUDA PEDESTAL.” J. Mammal. 88 (1): 59–66.
Minamikawa, S., H. Watanabe, and T. Iwasaki. 2013. “Diving behavior of a false killer whale , Pseudorca crassidens , in the Kuroshio – Oyashio transition region and the Kuroshio front region of the western North Pacific.” Marine Mammal Science 29 (1): 177–85. doi:10.1111/j.1748-7692.2011.00532.x.
Morato, Telmo, Divya Alice Varkey, Carla Damaso, Miguel Machete, Marco Santos, Rui Prieto, Ricardo S. Santos, and Tony J. Pitcher. 2008. “Evidence of a seamount effect on aggregating visitors.” Marine Ecology Progress Series 357 (Fonteneau 1991): 23–32. doi:10.3354/meps07269.
Reisinger, Ryan R, Mark Keith, Russel D Andrews, and P J N De Bruyn. 2015. “Journal of Experimental Marine Biology and Ecology Movement and diving of killer whales ( Orcinus orca ) at a Southern Ocean archipelago.” Journal of Experimental Marine Biology and Ecology 473. Elsevier B.V.: 90–102. doi:10.1016/j.jembe.2015.08.008.
Ringelstein, Julien, Claire Pusineri, Sami Hassani O, Laureline Meynier P, and Vincent Ridoux P. 2006. “Food and feeding ecology of the striped dolphin , Stenella coeruleoalba , in the oceanic waters of the north-east Atlantic.” J. Mar. Biol. Ass. UK 86: 909–18.
Watkins, W.A., M.A. Daher, K. M. Fristrup, T. J. Howald, and G. Notarbartolo di Sciara. 1993. “SPERM WHALES TAGGED WITH TRANSPONDERS AND TRACKED UNDERWATER BY SONAR.” Marine Mammal Science 9 (1): 55–67.
Watkins, William A., Mary Ann Daher, Nancy A. Dimarzio, Amy Samuels, Douglas Wartzok, Kurt M. Fristrup, Paul W. Howey, and Romaine R. Maiefski. 2002. “Sperm whale dives tracked by radio tag telemetry.” Marine Mammal Science 18 (1): 55–68. doi:10.1111/j.1748-7692.2002.tb01018.x.
Watwood, Stephanie L., Patrick J O Miller, Mark Johnson, Peter T. Madsen, and Peter L. Tyack. 2006. “Deep-diving foraging behaviour of sperm whales (Physeter macrocephalus).” Journal of Animal Ecology 75 (3): 814–25. doi:10.1111/j.1365-2656.2006.01101.x.
15.5 - References used in the other chapters:
Afonso, P., et al. (2018). Effects of marine protected areas on coastal fishes across the Azores archipelago, mid-North Atlantic. Journal of Sea Research 138: 34-47.
Avila SP, Azevedo JMN (1997) Shallow-water molluscs from the Rormigas Islets, Azoresm collected during the Santa Maria e Formigas 1990 scientific expedition, Azoreana 8: 323-330.
Bañon, R., et al. (2016). Commented checklist of marine fishes from the Galicia Bank seamount (NW Spain). Zootaxa 4067(3): 293-333.
Bergstad, O.A. 2013. North Atlantic demersal deep-water fish distribution and biology: present knowledge and challenges for the future. Review paper. Journal of Fish Biology 83: 1489-1507. doi:10.1111/jfb.12208.
Boschen RE, Rowden AA, Clark MR, Gardner JPA (2013) Mining of the deep-sea seafloor massive sulfides: A review of the deposits, their benthic communities, impacts from mining, regulatory frameworks and management strategies. Ocean & Coastal Managements 84: 54-67.
Clark, M. (2001). Are deepwater fisheries sustainable? — the example of orange roughy (Hoplostethus atlanticus) in New Zealand. Fisheries Research 51: 123-135.
Clark, M. R., Vinnichenko, V. I., Gordon, J. D. M., Beck-Bulat, G. Z., Kukharev, N. N. & Kakora, A. F. (2007). Large-scale distant-water trawl fisheries on seamounts. In: Seamounts: Ecology, Fisheries, and Conservation (Pitcher, T. J., Morato, T., Hart, P. J. B., Clark, M. R., Haggan, N. & Santos, R. S., eds), pp. 361–399. Oxford: Blackwell Publishing.
de Bruin T (2017). Sub-leathal stress responses of a deep-water boreal sponge to future climate change scenarios. Master Thesis. University of Amsterdam Nov 2017. 29 pp.
FAO (2005). Deep Sea 2003: Conference on the Governance and Management of Deep-sea Fisheries. FAO Fisheries Proceedings, 3/1 and 3/2. 718p + 487p.
FAO (2008). Worldwide review of bottom fisheries in the high seas. FAO Fisherie and Aquaculture Technical Paper, 522: 145pp.Fock, H., et al. (2002). Biodiversity and species-environment relationships of the demersal fish assemblage at the Great Meteor Seamount (subtropical NE Atlantic), sampled by different trawls. Marine Biology Research 141: 185-199.
Frank N, Ricard E, Lutringer-Paquet A, van der Land C, Colin C, Blamart D, Foubert A, Van Rooij D, Henriet J-P, de Haas H, van Weering T (2009) The Holocene occurrence of cold water corals in the NE Atlantic: Implications for coral carbonate mound evolution. Marine Geology 266: 129-142.
Gordon, J.D.M. 2005. Environmental and biological aspects of deepwater demersal fishes. P. 70-88 in Shotton, R. (Ed.) Deep Sea 2003: Conference on the Governance and Management of Deep-sea Fisheries Part 1. Conference reports. Queenstown, New Zealand, 1-5 december 2003. FAO Fisheries Proceedings. No. 3/1. Rome, FAO. 2005. 718p.
Grüss, A., et al. (2014). Conservation and fisheries effects of spawning aggregation marine protected areas: What we know, where we should go, and what we need to get there. ICES Journal of Marine Science 71(7): 1515-1534.
Hareide, N.-R. (1995). Comparisons between longlining and trawling for deep-water species – selectivity, quality and catchability. P. 227-234 in: Hopper, A. G. (ed.) 1995. Deep-Water Fisheries of the North Atlantic Oceanic Slope. NATO ASI Series, Series E: Applied sciences, vol 296. London: Kluwer Academic Publishers, 420p.
Hareide, N.-R. & Garnes, G. (2001). The distribution and catch rates of deep water fish along the Mid-Atlantic Ridge from 43 to 61N. Fisheries Research 51, 297–310.
Hein JR, Conrad TA, Staudigel A (2010) Seamount mineral deposits: A source of rare metals for high-technology industries. Oceanography 23:184-189.
Hopper, A. G. (ed.) 1995. Deep-Water Fisheries of the North Atlantic Oceanic Slope. NATO ASI Series, Series E: Applied sciences, vol 296. London: Kluwer Academic Publishers, 420p.
Hubbs, C. L. (1959). Initial discoveries of fish faunas on seamounts and offshore banks in the eastern Pacific. Pacific Science 13: 311-316.
Kvile, K. Ø., et al. (2014). A global assessment of seamount ecosystems knowledge using anecosystem evaluation framework. Biological Conservation 173: 108-120.
Large, P.A. and Bergstad, O.A. 2005. Deepwater fish resources in the northeast Atlantic: fisheries, state of knowledge on biology and ecology, and recent developments in stock assessment and management. P. 149-161 in Shotton, R. (Ed.) Deep Sea 2003: Conference on the Governance and Management of Deep-sea Fisheries Part 1. Conference reports. Queenstown, New Zealand, 1-5 december 2003. FAO Fisheries Proceedings. No. 3/1. Rome, FAO. 2005. 718p.
Magnussen, E. (2002). Demersal fish assemblages of Faroe Bank: species composition, distribution, biomass spectrum and diversity. Marine Ecology and Progress Series 238: 211-225.
Martins I, Godinho A, Goulart J, Carreiro-Silva M (2018) Assessment of Cu sub-lethal toxicity (LC50) in the cold-water gorgonian Dentomuricea meteor under a deep-sea mining activity scenario. Environmental Pollution 240: 903-907.
Menezes, G., et al. (2011). Demersal fish assemblages of the Condor seamount. CONDOR Observatory for long-term study and monitoring of Azorean seamount ecosystems. E. Giacomello and G. Menezes. Final Project Report. Arquivos do DOP, Série Estudos 1/2012. : 129-147.
Mironov, A.N., Gebruk, A.V. and Southward, A.J. (Eds). Biogeography of the North Atlantic seamounts. Moscow: KMK Scientific Press Ltd. 2006. 196 p.
NAFO (2003). Symposium on Deep-sea fisheries, 12-14 September 2001 (eds. J.A.Mooore and J.D.M. Gordon). Journal of Northwest Atlantic Fisheries Science, 31: 1-466.
Neat, F., et al. (2008). The deepwater ecosystem of the continental shelf slope and seamounts of the Rockall Through: A report on the ecology and biodiversity based on FRS scientific surveys.
OASIS (2003). Seamounts of the North-East Atlantic. Hamburg and WWF Germany, Frankfurt am Main, November 2003: 38.
Pham CK, Gomes-Pereira JN, Isidro EJ, Santos RS, Morato T (2013) Abundance of litter on Condor seamount (Azores, Portugal, Northeast Atlantic). Deep Sea Research Part II: Topical Studies in Oceanography 98:204-208.
Pham CK, Ramirez-Llodra E, Alt CHS, Amaro T, Bergmann M et al. (2014) Marine litter distribution and density in European seas, from the shelves to deep basins. PLoS One 9 (4)
Porteiro F, Sutton T, Byrkjedal, I, Orlov A, Heino M, Menezes G and OA Bergstad. 2017. Fishes of the Northern Mid-Atlantic Ridge collected during the MAR-ECO cruise in June-July 2004. An annotated checklist. Arquipelago, Suppl. 10, 1-125.
Rogers, A. D. (2018). The Biology of Seamounts: 25 years on. Advances in Marine Biology 79: 137-224.
Serrano, A., E Sánchez, J., Cartes, J., Sorbe, S., Parra, I., Frutos, I., . . . Preciado, I. (2005). ECOMARG Project: A multidisciplinary study of Le Danois Bank (Cantabrian Sea, N Spain).
Stocks, K. (2002). Using SeamountsOnline, a biogeographic information system for seamounts, to examine patterns in seamount endemism. Proceedings of the 2002 International Council for the Exploration of the Seas Annual Science Conference. Copenhagen, Denmark.
16 - Appendix 1 - Plankton registrations
Table 1
Carmo et al. 2013. Zooplankton at Condor seamount.
Taxonomic groups
Species
PROTOZOA
Foraminifera
Foraminifera (unidentified)
Radiolaria
Acantharia
Dyctiocoryne spp.
Phaeodaria
Polycystina
Radiolaria (unidentified)
Radiolaria colony
Spongaster spp.
Spongodiscidae
Ciliata
Ciliata colony
Tintinnidea
Tintinnidea
ANIMALIA
Invertebrate
Egg (unidentified)
Trocophora larva
Cnidaria
Cnidaria (unidentified)
Hydrozoa
Hydromedusae
Planula larva
Siphonophora
Scyphozoa
Ephyra
Scyphozoa (unidentified)
Ctenophora
Ctenophora (unidentified)
Platyhelminthes
Muller's larva
Nemertea
Nemertea (unidentified)
Bryozoa
Cyphonaute larva
Mollusca
Heteropoda
Atlanta oxygyrus
Atlanta spp.
Pterotrachea spp.
Pteropoda
Cavolinia spp.
Creseis spp.
Diacria spp.
Limacina spp.
Cymbulioidea
Desmopterus spp.
Peraclis spp.
Embrionic shell
Gastropoda
Echinospira larva
Gastropoda Veligera larva
Bivalvia
Bivalvia Veligera larva
Cephalopoda
Teuthida
Polychaeta
Nectochaeta larva
Polychaeta sedentaria larva
Polychaeta larva
Tomopteridae
Vanadis spp.
Crustacea
Crustacea nauplius (unidentified)
Egg w/ nauplius (unidentified)
Cladocera
Cladocera (unidentified)
Evadne spp.
Evadne spp. (partnogenetic ♀)
Evadne spp. (♀ w/ egg)
Pseudevadne spp.
Pseudevadne spp. (partnogenetic ♀)
Pseudevadne spp. (♀ w/ egg)
Ostracoda
Conchoecilla spp.
Myodocopa
Ostracoda (unidentified)
Copepoda
- Calanoida
Acartia spp.
Aetideidae
Aetideopsis spp.
Aetideus spp.
Calanoida (unidentified, incl. copepodites)
Calanus spp. group (1)
Calocalanus spp.
Candacia spp.
Centropages abdominalis
Centropages spp.
Centropages typicus
Centropages violaceus
Eucalanidae
Eucalanus (monachus)?
Euchaeta spp.
Euchirella spp.
Gaetanus spp.
Heterorhabdidae
Lucicutia spp.
Metrididae
Pleuromamma spp.
Pontellidae
Pontellina spp.
Pseudocalanus acuspes
Rhincalanus spp.
Rhincalanus spp. Nauplius
Scaphocalanus spp.
Scollecithrix bradyi
Spinocalanus spp.
Temora spp.
Undeuchaeta spp.
- Poecilostomatoida
Copilia spp.
Corycaeidae
Oncaeidae
Poecilostomatoida (unidentified)
Sapphirinidae
- Cyclopoida
Cyclopoida (unidentified)
Oithona spp.
- Harpacticoida
Clytemenestra spp.
Distioculus spp.
Euterpina spp.
Harpacticoida (unidentified)
Harpacticoida nauplius
Microsetella spp.
Cirripedia
Cirripedia nauplius (unidentified)
Cypris larva
Sacculina spp.
Isopoda
Isopoda
Amphipoda
Gammaridea
Hyperidea
Vibilia spp.
Mysidacea
Mysidacea
Euphausiacea
Calyptopis larva
Euphausia spp.
Euphausiacea Juvenile (unidentified)
Furcilia larva
Metanauplius
Nematobrachion flexipes
Stylocheiron spp. furcilia larva
Stylocheiron spp. Juvenile
Thysanoessa spp. Juvenile
Decapoda
Crab zoea larva
Decapoda Juvenile (unidentified)
Decapoda larva (unidentified)
Luciferidae
Megalopa larva
Metazoea larva
Mysis larva
Penaeidae
Protozoea larva
Scylarus arctus phyllosoma larva
Sergestes spp. larva
Zoea larva
Echinodermata
Echinodermata larva (unidentified)
Asteroidea
Brachiolaria larva
Echinoidea
Echinopluteus larva
Holothuroidea
Auricularia larva
Doliolaria larva
Ophiuroidea
Ophiopluteus larva
Chaetognatha
Chaetognatha (unidentified)
Phoronida
Actinotrocha larva
Hemichordata
Enteropneusta tornaria larva
Urochordata
Appendicularia
Appendicularia
Doliolida
Doliolida (unidentified)
Doliolida larva
Doliolida old nurse
Salpida
Salpida
Cephalochordata
Amphioxus
Vertebrata
Teleostei
Fish egg
Fish larva
Myctophidae
Table 2
Santos et al. 2013. Phytoplankton at Condor seamount.
Hesthagen, Ivar H (1970): The near-bottom plankton and benthic invertebrate fauna of Josephine and Great Meteor Seamounts (Table 2). PANGAEA, https://doi.org/10.1594/PANGAEA.611095
Weigmann, Renate (1974): Relative abundance of euphausiids (Crustacea)
in water samples of the Josephine Seamount, Atlantic Ocean (Table 8).
PANGAEA, https://doi.org/10.1594/PANGAEA.611406,
Main taxonomic groups
Relative abundance of euphausiids
Porifera
Euphausia brevis
Hydrozoa
Meganyctiphanes norvegica
Hydrozoa
Stylocheiron suhmii
Scyphozoa
Euphausia hemigibba
Scyphozoa
Thysanopoda subaequalis
Anthozoa, larvae
Nematoscelis megalops
Nematoda
Stylocheiron longicorne
Echiurida
Stylocheiron carinatum
Sipunculida
Polychaeta
Ostracoda
Copepoda
Cirripedia
Cirripedia
Mysidacea
Amphipoda
Amphipoda
Euphausiacea
Decapoda
Decapoda
Decapoda, larvae
Cumacea
Pycnogonida
Solenogastres
Gastropoda
Cephalopoda
Bivalvia
Chaetognatha
Bryozoa
Crinoidea
Asteroidea
Ophiuroidea
Echinoidea
Appendicularia
Seriocarpa rhizoides
Ascidiacea
Thaliacea
Pisces
17 - Appendix 2 - Benthos tables
References for papers on the infauna composition of known seamounts, banks and knolls
PORIFERA: Colaco A, Raghukumar C, Mohandrass C, Cardigos F, Santos RS (2006) Effect of shallow-water venting in Azores on a few marine biota. Cah. Biol. Mar. 47:359-364
Cristobo J, Rios P, Pomponisa SA, Xavier J (2015) A new carnivorous sponge, Chondrocladia robertballardi sp. nov. (Porifera: Cladorhizidae) from two north-east Atlantic seamounts. Journal of the Marine Biological Association of the United Kingdom. 95:1345-1352.
Davies JS, Stewart HA, Narayanaswamy BE, Jacobs C, Spicer J, Golding N, et al. (2015) Benthic Assemblages of the Anton Dohrn Seamount (NE Atlantic): Defining Deep-Sea Biotopes to Support Habitat Mapping and Management Efforts with a Focus on Vulnerable Marine Ecosystems. PLoS ONE 10(5): e0124815. doi:10.1371/journal.pone.0124815
Henry LA. et al. Environmental variability and biodiversity of megabenthos on the Hebrides Terrace Seamount (Northeast Atlantic). Sci. Rep. 4, 5589; DOI:10.1038/srep05589 (2014).
McIntyre FD, Drewery J, Eerkes‐Medrano D, Neat FC (2016) Distribution and diversity of deep‐sea sponge grounds on the Rosemary Bank Seamount, NE Atlantic. Mar Biol 163:143 DOI 10.1007/s00227-016-2913-z
Narayanaswamy BE, Hughes DJ, Howell KJ, Davies J, Jacobs C (2013) First observations of megafaunal communities inhabiting George Bligh Bank, Northeast Atlantic Deep-Sea Research II 92: 79-86
Ramos M, Bertocci I, Tempera F, Calado G, Albuquerque M, & Duarte P (2015). Patterns in megabenthic assemblages on a seamount summit (Ormonde Peak, Gorringe Bank, Northeast Atlantic) Marine Ecology 37:1057-1072
Serrano A, González-Irusta JM, Punzón A, García-Alegre A, Lourido A, Ríos P, Blanco M, Gómez-Ballesteros M, Druet M, Cristobo J, Cartes JE (2017) Deep-sea benthic habitats modeling and mapping in a NE Atlantic seamount (Galicia Bank). Deep-Sea Research Part I 126: 115-127.
van Haren H, Hanz U, de Stigter H, Mienis F, Duineveld G (2017) Internal wave turbulence at a biologically rich Mid-Atlantic seamount. PLoS ONE 12(12): e0189720.
Roberts EM, Mienis F, Rapp HT, Hanz U, Meyer HK, Davies AJ (2019) Oceanographic setting and short-timescale environmental variability at an Arctic seamount sponge ground, Deep-Sea Research Part I, https://doi.org/10.1016/j.dsr.2018.06.007
Santos RS, Tempera F, Menezes G, Porteiro F, Morato T (2010) Mountains in the sea: Sedlo Seamount, Azores. Oceanography 23:202-203.
Henry LA, Roberts M (2014). Applying the OSPAR habitat definition of deep-sea sponge aggregations to verify suspected records of the habitat in UK waters. JNCC Report 508.
Xavier JR, Van Soest RWM (2007) Demosponge fauna of Ormonde and Gettysburg Seamounts (Gorringe Bank, Northeast Atlantic): diversity and zoogeographical affinities. Journal of the Marine Biological Association of the United Kingdom 87:1643-1653.
Torkildsen, M. M. (2013). Diversity of hexactinellid sponges (Porifera, Hexactinellida) on an Arctic seamount, the Schultz Massif. M.Sc. Thesis, University of Bergen.
Plotkin, A., Gerasimova, E., & Rapp, H. T. (2017). Polymastiidae (Porifera: Demospongiae) of the Nordic and Siberian Seas. Journal of the Marine Biological Association of the United Kingdom.
Hestetun, J. T., Tompkins-Macdonald, G., & Rapp, H. T. (2017). A review of carnivorous sponges (Porifera: Cladorhizidae) from the Boreal North Atlantic and Arctic. Zoological Journal of the Linnean Society, 181(1), 1–69. https://doi.org/10.1093/zoolinnean/zlw022
Cárdenas, P., Rapp, H. T., Klitgaard, A. B., Best, M., Thollesson, M., & Tendal, O. S. (2013). Taxonomy, biogeography and DNA barcodes of Geodia species (Porifera, Demospongiae, Tetractinellida) in the Atlantic boreo-arctic region. Zoological Journal of the Linnean Society, 169(2), 251–311. https://doi.org/10.1111/zoj.12056
Henrich R, Freiwald A, Betzler C, Bader B, Schäfer P, Samtleben C, Brachert TC, Wehrmann A, Zankl H, Kühlmann DHH (1995). Controls on modern carbonate sedimentation on warm-temperate to arctic coasts, shelves and seamounts in the Northern Hemisphere: Implications for fossil counterparts. Facies 32:71-108.
Cherkis, N.Z., Steinmetz, S., Schreiber, R. et al. (1994) Vesteris Seamount: An enigma in the Greenland BasinMar Geophys Res16: 287. https://doi.org/10.1007/BF01224746
Colaco et al. 2013
RETARIA (foraminifera): Davies et al. 2015, Henry et al. 2014, Narayanaswamy et al. 2013
CNIDARIA: Duineveld GCA, Lavaleye MSS, Berghuis EM (2004) Particle flux and food supply to a seamount cold- water coral community (Galicia Bank, NW Spain). Marine Ecology Progress Series 277:13-23.
Frank N (2008) Short Cruise Report M151 Atlantic Thermocline Ocean and Ecosystems Dynamic during Natural Climate Change. Ponta Delgada (Portugal) - Funchal (Portugal) October 6 - 31, 2018. RV Meteor.
Braga-Henriques A, Porteiro FM, Ribeiro PA, de Matos V, Sampaio I, Ocana O, Santos RS (2013) Diversity, distribution and spatial structure of the cold-water coral fauna of the Azores (NE Atlantic). Biogeosciences 10:4009-4036, doi:10.5194/bg-10-4009-2013
Howell KL, Mowles SL, Foggo A (2010) Mounting evidence: near-slope seamounts are faunally indistinct from an adjacent bank. Marine Ecology 31:52-62.
Howell KL, Bullimore RD, Foster NL (2014) Quality assurance in the identification of deep-sea taxa from video and image analysis: response to Henry and Roberts. ICES Journal of Marine Science 71:899-906.
Cross T, Howell KL, Hughes E, Seeley, July R (2014) Analysis of seabed imagery from the Hebrides Terrace Seamount (2013). JNCC Report No. 510.
Moura CJ (2015) The hydrozoan fauna (Cnidaria: Hydrozoa) from the peaks of the Ormonde and Gettysburg seamounts (Gorringe Bank, NE Atlantic). Zootaxa 3972:148-180. doi: 10.11646/zootaxa.3972.2.2.
Magnussen E (2002) Demersal fish assemblages of Faroe Bank: Species composition, distribution, biomass spectrum and diversity. Marine Ecoloy Progress Series 238:211-225.
Roberts JM, Henry LA, Long D, Hartley J (2008) Cold-water coral reef frameworks, megafaunal communities and evidence for coral carbonate mounds on the Hatton Bank, north east Atlantic. Facies 54:297-316.
Colaco et al. 2013, Davies et al. 2015, Henry et al. 2014, Narayanaswamy et al. 2013, Ramos et al. 2015, Serrano et al. 2017, van Haren et al. 2017, Santos et al. 2010, Henry & Roberts 2014, ICES 2015, Molodtsova et al. 2008
BRYOZOA: Berning B, Harmelin JG, Bader B (2017) New Cheilostomata (Bryozoa) from NE Atlantic seamounts, islands, and the continental slope: evidence for deep-sea endemism. European Journal of Taxonomy 347:1-51. https://doi.org/10.5852/ejt.2017.347
Souto J, Berning B, Ostrovsk AN (2016) Systematics and diversity of deep-water Cheilostomata (Bryozoa) from Galicia Bank (NE Atlantic). Zootaxa 4067:401-459.
Colaco et al. 2013
MOLLUSCA:Oliviero M, Gofas S (2006) Coralliophiline diversity at mid Atlantic seamounts, Neogastropoda, Muricidae, Coralliophilinae. Bulletin of Marine Science 69: 205-230.
Avila SP, Malaquias MAE (2003). Biogeographical relationships of the Molluscan fauna of the Ormonde seamount (Gorringe Bank, North-East Atlantic Ocean). Journal of Molluscan Studies 69:145-150.
Avila SP, Cardigos SPF, Santos RS (2004) D. João de Castro Bank, a shallow water hydrothermal-vent in the Azores: checklist of the marine molluscs. Life and Marine Sciences 21A:75-80.
Colaco et al. 2013, Ramos et al. 2015, Serrano et al. 2017, Zepilli et al. 2017
ANNELIDA: Surugiu V, Dauvin JC, Gillet P, Ruellet T (2008) Can seamounts provide a good habitat for polychaete annelids? Example of the northeastern Atlantic seamounts. Deep-Sea Research I 55:1515-1531.
Colaco et al. 2013, Davies et al. 2015, Narayanaswamy et al. 2013, Ramos et al. 2015, Zepilli et al. 2017
ARTHROPODA: Cartes JE, Serrano A, Velasco F, Parra S, Sánchez F (2007) Community structure and dynamics of deep-water decapod assemblages from Le Danois Bank (Cantabrian Sea, NE Atlantic): Influence of environmental variables and food availability Progress in Oceanography 75: 797-816.
Colaco et al. 2013, Davies et al. 2015, Narayanaswamy et al. 2013, Ramos et al. 2015, Serrano et al. 2017, Zepilli et al. 2017
ECHINODERMATA: Colaco et al. 2013, Davies et al. 2015, Henry et al. 2014, Narayanaswamy et al. 2013, Ramos et al. 2015, Serrano et al. 2017
ASCIDIACEA: Narayanaswamy et al. 2013, Ramos et al. 2015
BRACHIOPODA: Narayanaswamy et al. 2013
ECHIURA: Narayanaswamy et al. 2013
NEMATODA:Zeppilli D, Bongiorni L, Cattaneo A, Danovaro R, Santos RS (2013). Meiofauna assemblages of the Condor Seamount (North-East Atlantic Ocean) and adjacent deep-sea sediments . Deep-Sea Research II 98:87-100.
Benthic taxa registered at seamounts and seamount like features in the OSPAR maritime area.
Presence/absence data of fish species at different seamounts in the OSPAR area: Hatton Bank (HB), Formigas Dollabarat (FD), Faroe Bank (FB), Condor de Terra (CT), Sedlo (S), Anton Dohrn (AD), Rosemary's bank (RB), Gorringe bank (GB), Faraday (F), Galicia bank (GB), Josephine (J), Hebrides Terrasse (HT), Georges Bligh bank (GBB), Le Danois bank (LDB), Joao De Castro (JDC). Act=Actinopterygii, cho=Chondrichthyes.
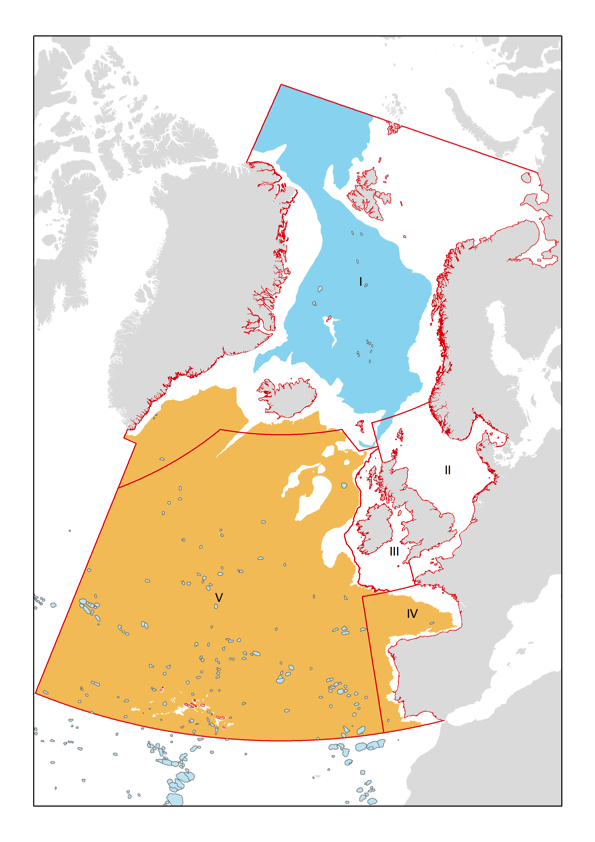
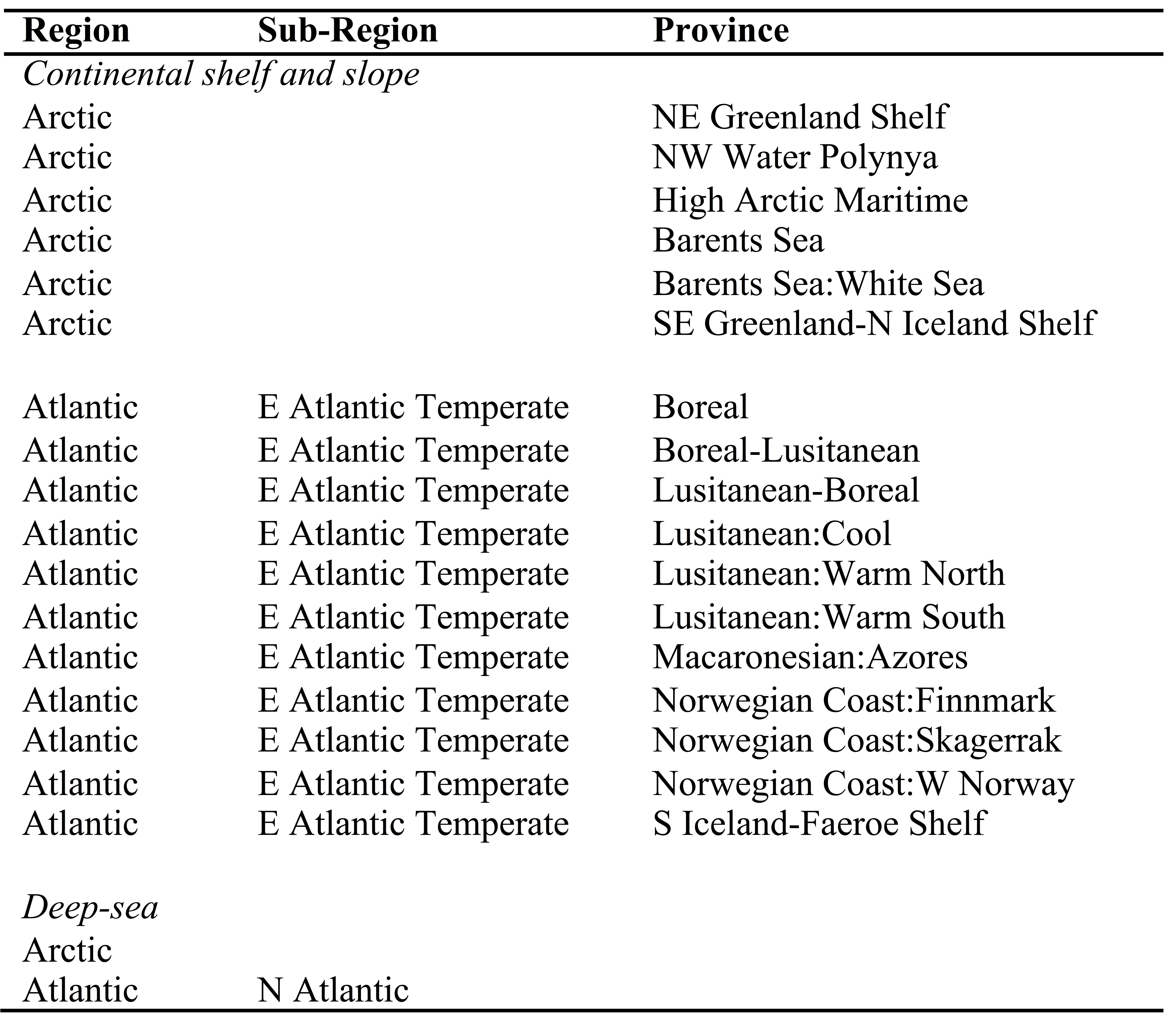


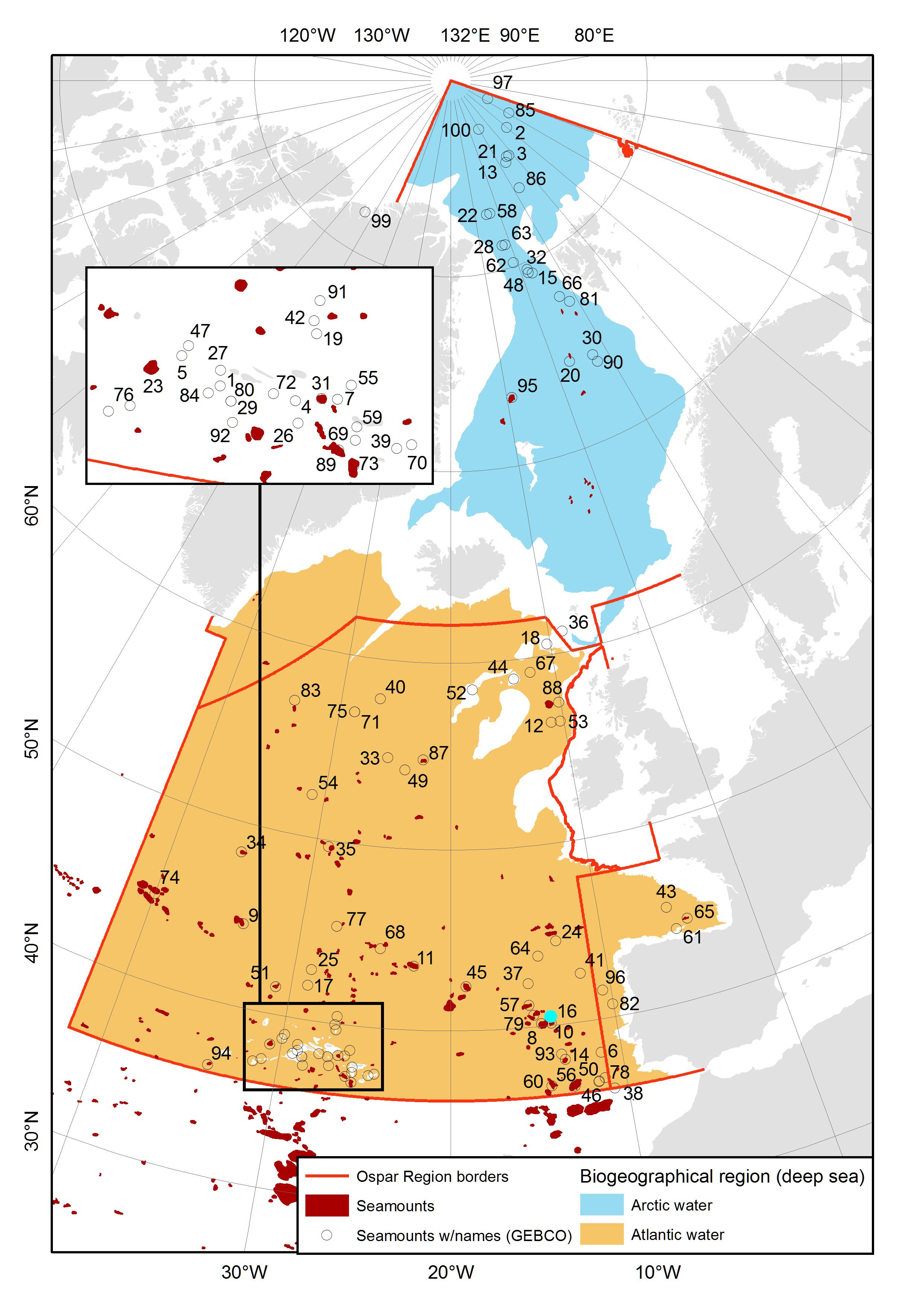

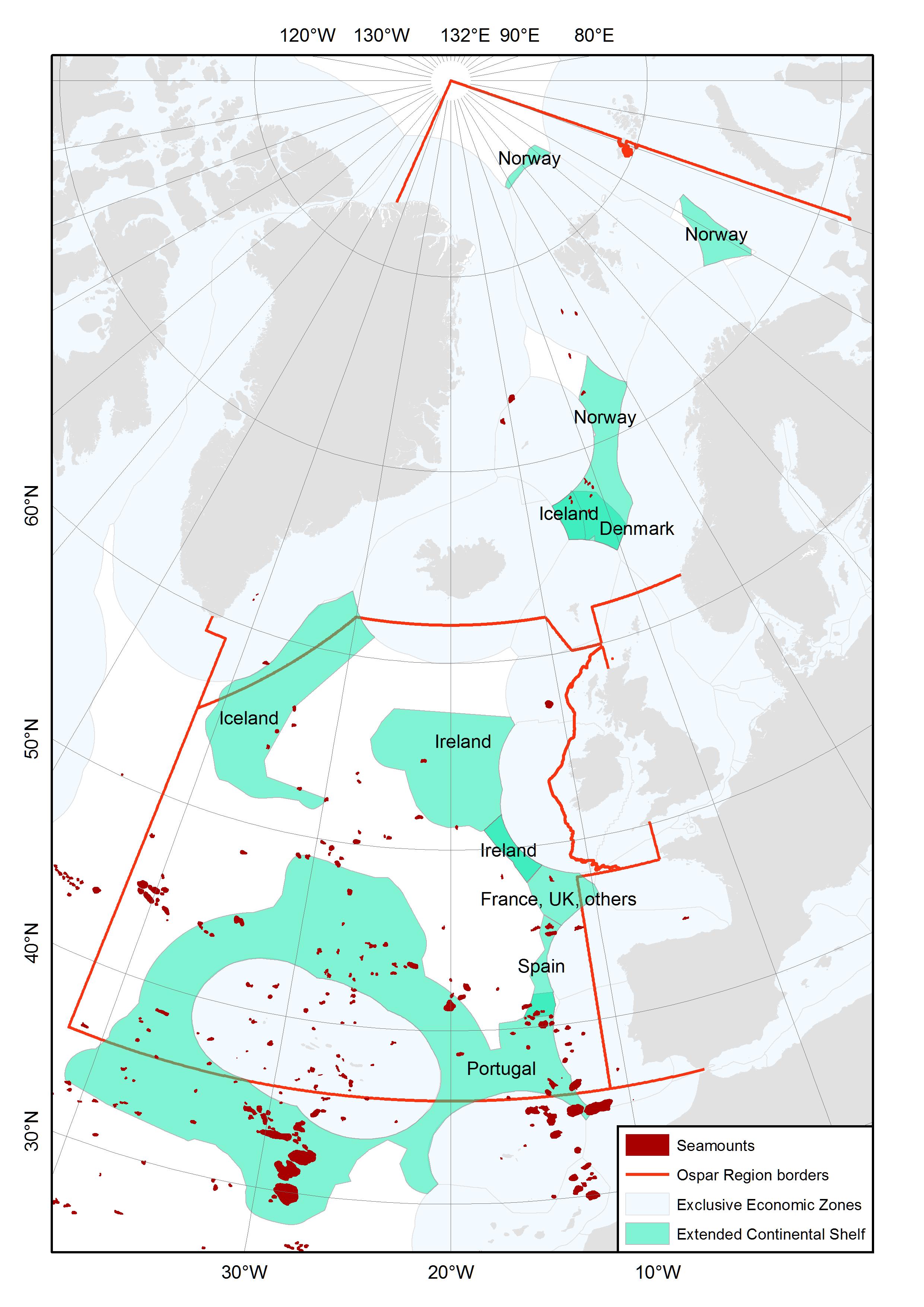
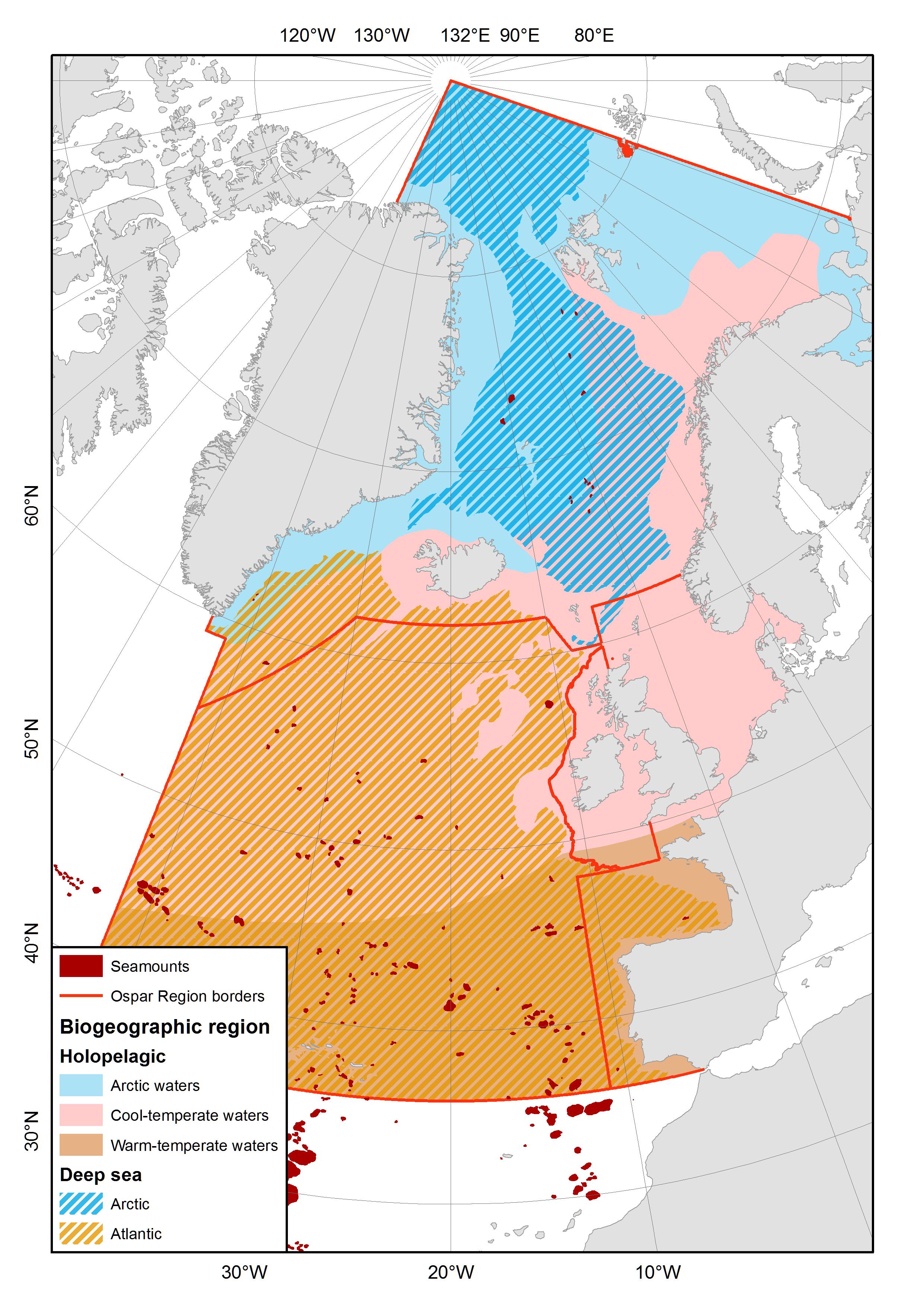

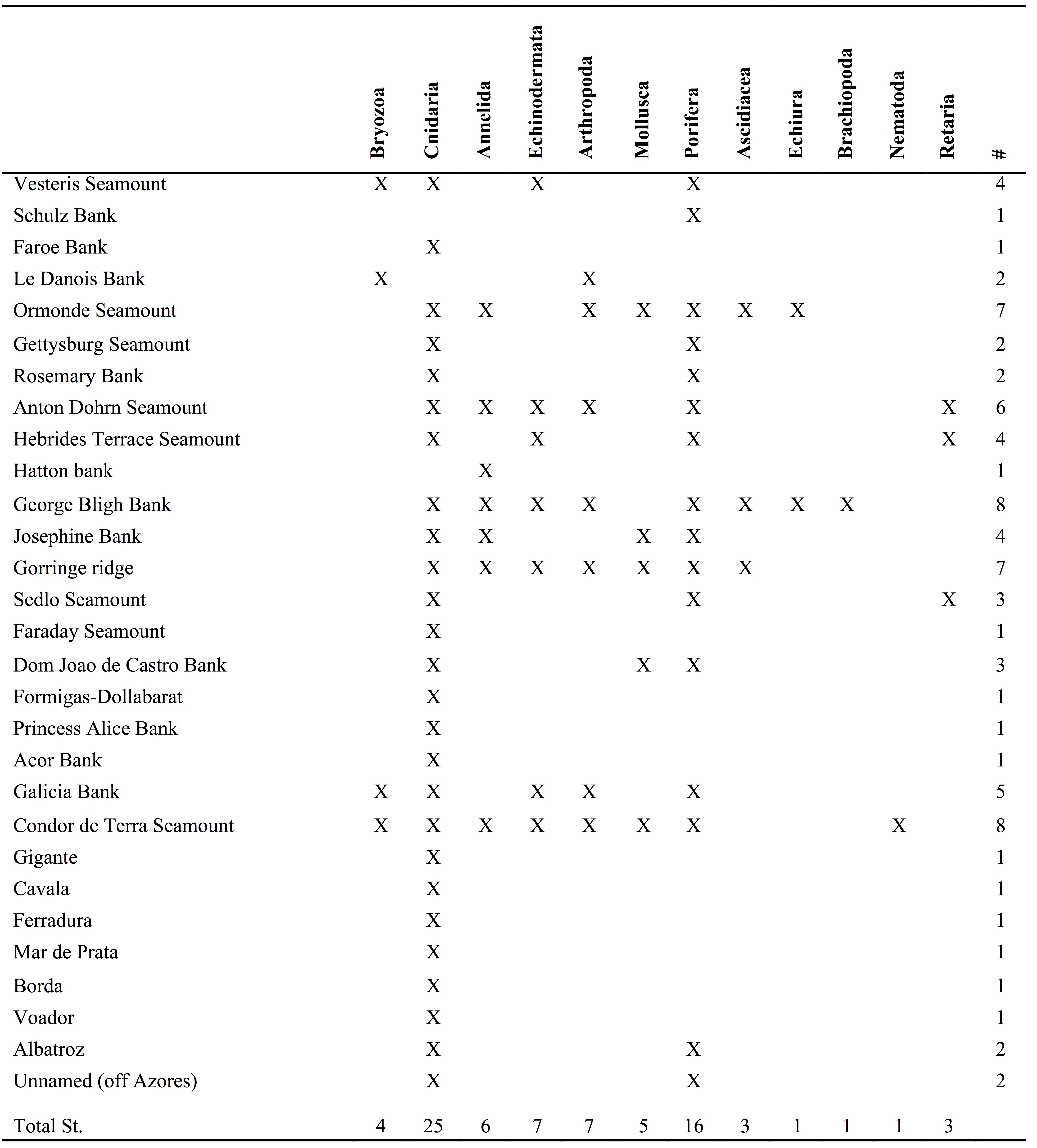
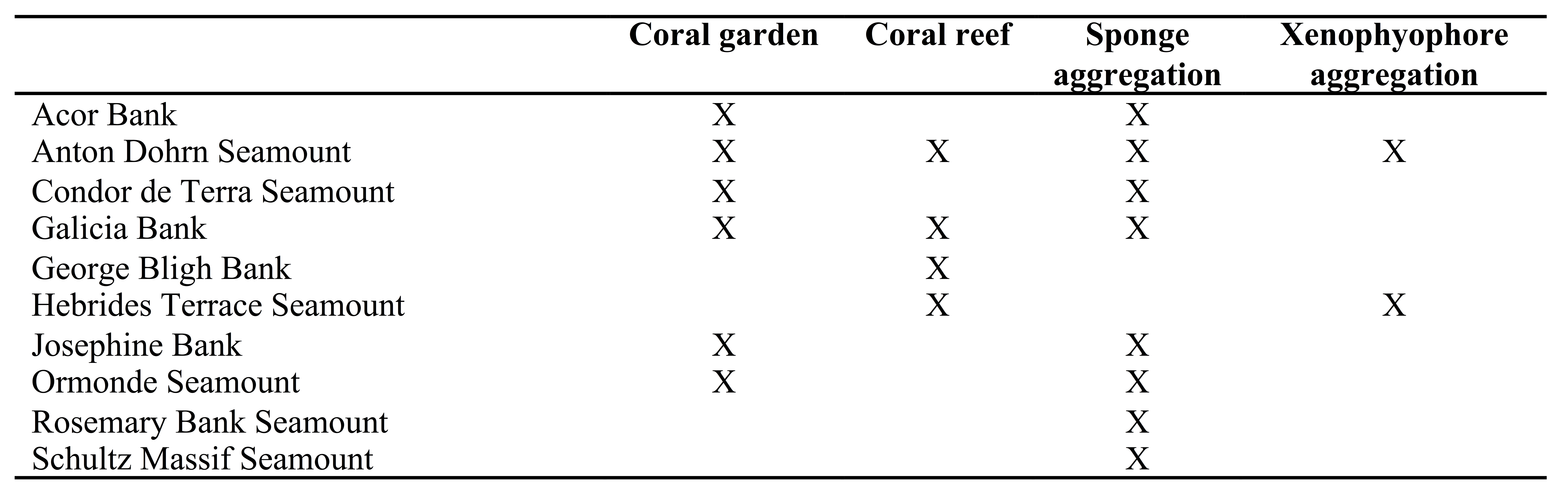
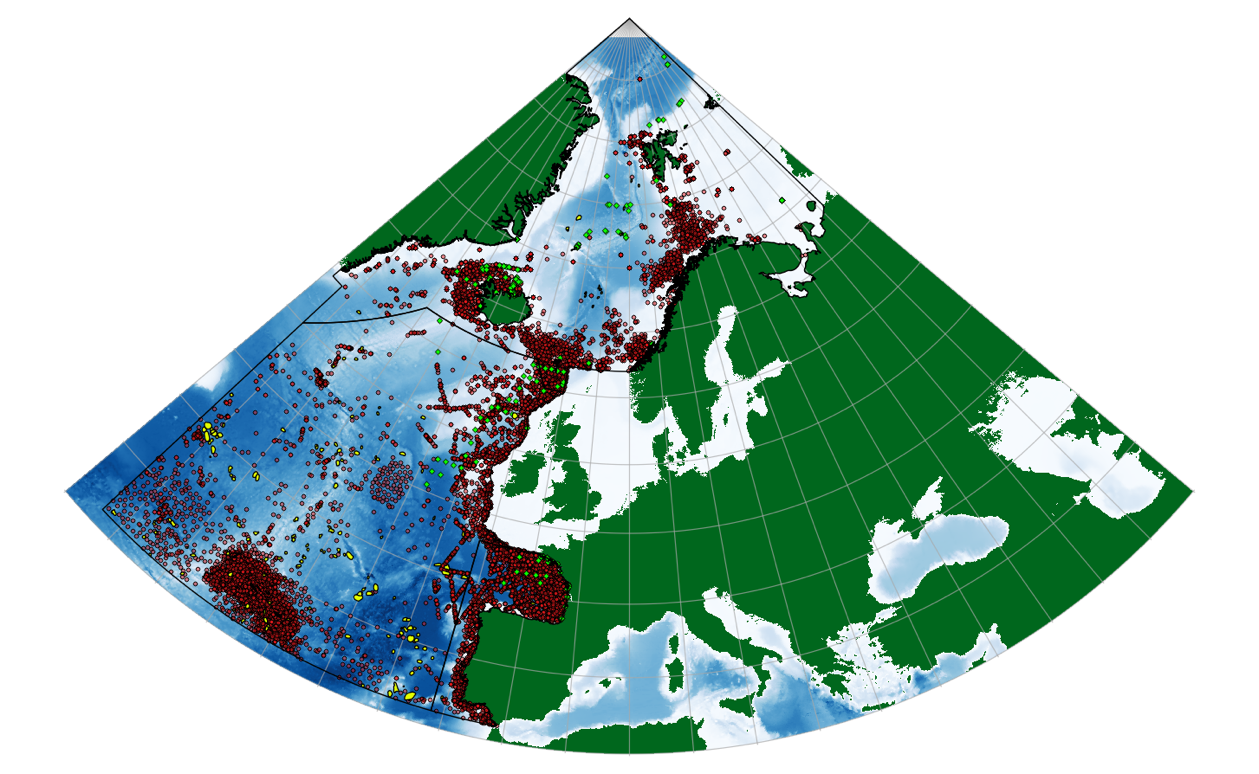
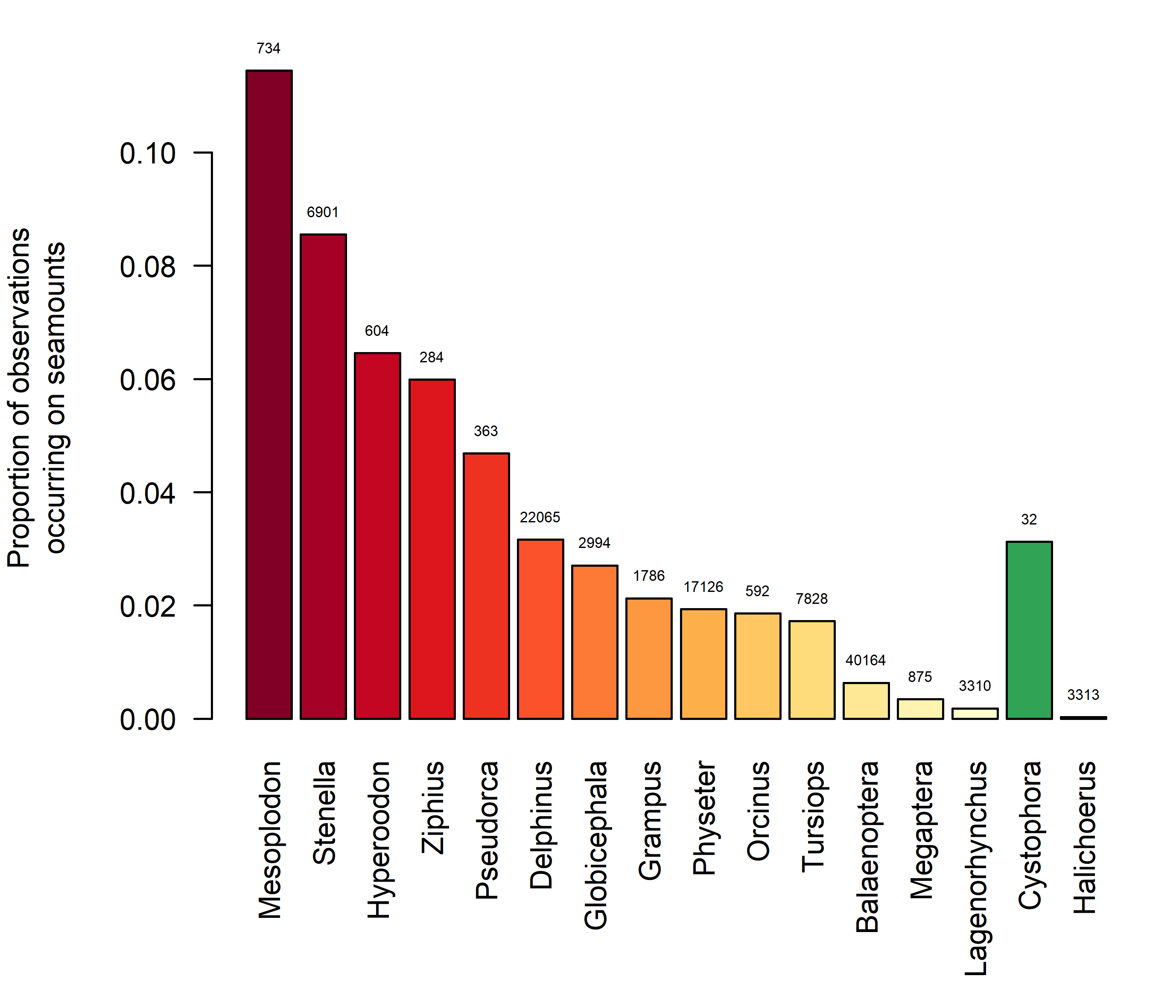 Figure 6. Proportion of all marine mammal observations included in the OBIS database that occur within 10 nm of a seamount, summarized per genus.
Figure 6. Proportion of all marine mammal observations included in the OBIS database that occur within 10 nm of a seamount, summarized per genus.