[Text]
Negative effekter på kysttorskbestander som følge av torskeoppdrett i åpne merder i sjø
— Risikovurdering og kunnskapsstatus 2025
Rapportserie:
Rapport fra havforskningen 2025-78
ISSN: 1893-4536
Risikorapport norsk fiskeoppdrett
Publisert: 09.12.2025
Prosjektnr: 16203
Oppdragsgiver(e): Fiskeridirektoratet
Referanse: 25/5093
Forskningsgruppe(r):
Smittespredning og sykdom
,
Populasjonsgenetikk
,
Reproduksjon og utviklingsbiologi
,
Dyrevelferd
,
Bunnfisk
,
Bunnsamfunn
,
Økosystemprosesser
,
Dyphavsfisk, hai og skalldyr
,
Kystoseanografi
,
Bentisk økologi
Tema:
Torskeoppdrett
Program:
Miljøeffekter av akvakultur
Godkjent av:
Forskningsdirektør(er):
Geir Lasse Taranger
Programleder(e):
Mari Skuggedal Myksvoll
English summary
Sammendrag
I denne risikovurderingen vurderer vi risiko for «Negative effekter på kysttorskbestander som følge av torskeoppdrett i åpne merder i sjø». Dette er en oppdatering og videreutvikling av risikovurderingen som Havforskningsinstituttet publiserte i 2022. Vurderingen er gjort for de to nordligste forvaltningsområdene for kysttorsk, mellom 62°N og 67°N og nord for 67°N. Området sør for 62°N inkluderes ikke i årets vurdering da aktiviteten i området er svært lav og det foreligger få søknader om økt kapasitet.
Kysttorskbestandene sør for 67°N vurderes fortsatt å være i dårlig tilstand, særlig sør for 62°N, og det har vært knyttet bekymring til økt produksjon av oppdrettstorsk, blant annet fra fiskerinæringen. Nord for 67°N har gytebiomassen økt kraftig og kysttorskbestanden anses nå for å være i god tilstand. Siden 2022 har produksjonen av oppdrettstorsk økt jevnt fra 4976 til 14 683 tonn i 2024 og ved utgangen av september 2025 er det slaktet 13 765 tonn. Per oktober 2025 utgjør godkjente lokaliteter og søknader til sammen en kapasitet på 165 000 tonn. I denne risikovurderingen er det lagt til grunn en kapasitetsutnyttelse på 40 % av den maksimale kapasiteten. Tidsperspektivet er satt til perioden 2026-2030. Om situasjonen eller premissene skulle endre seg i vesentlig grad innen 2030, vil det være behov for å gjøre en ny vurdering.
Vi konkluderer med at risikoen for «Negative effekter på kysttorskbestander som følge av torskeoppdrett i åpne merder i sjø» vurderes å være lav nord for 67°N og moderat i området mellom 62°N og 67°N, med en estimert produksjon på henholdsvis 13 000 og 49 000 tonn. Selv om produksjonen av oppdrettstorsk i området mellom 62°N og 67°N ikke vurderes som høy, er tilstanden til kysttorskbestanden i området svak og dermed antatt å være lite motstandsdyktig mot eventuell genetisk innkrysning fra oppdrettstorsk. Det er derfor en bekymring for at innkrysning med oppdrettstorsk kan medføre en generell reduksjon i genetisk variasjon som kan gi negative konsekvenser for bestandenes evne til å tilpasse seg endrede miljøbetingelser.
Som i vurderingen fra 2022, anser vi fortsatt rømming og gyting i merd som de største utfordringene i torskeoppdrett. Antallet oppdrettstorsk som har rømt de siste årene har vært dominert av fåtallige men store hendelser. Basert på erfaringer med rømmingstall fra lakseoppdrett, er det i tillegg grunn til å tro at antall rømte torsk er noe høyere enn det som er innrapportert. Status på kjønnsmodning viser at det er en betydelig andel som når stadiet for gyting i merd. Overlevelse fra egg gytt i merd til kjønnsmoden torsk er påvist, men kunnskapen om dette er svak og fragmentert. Hvis slik overlevelse er av en viss størrelsesorden kan gyting i merd være betydelig mer alvorlig enn rømming i et regionalt perspektiv.
Mengden oppdrettstorsk som får gyte i merd og antall fisk som rømmer, vurdert i forhold til tilstanden på kysttorskbestanden, vil være avgjørende for hvor stor den genetiske påvirkningen på kysttorskbestandene kan bli. Det mangler dokumentert kunnskap rundt hvilke endringer i egenskaper hos oppdrettstorsken som vil overføres til villtorsk ved en innkrysning. Det er derfor stor usikkerhet knyttet til hvilke konsekvenser dette eventuelt kan få for kysttorskbestandene på lengre sikt. Forhindring av rømming og bedre kontroll på gyting i merd er problemstillinger det nå arbeides med både fra forskningen og næringen sin side, gjennom optimalisering av lysstyring, forbedring av oppdrettsnøter og oppdrettsteknologi, inkludert nedsenkbar merdteknologi og lukkede anlegg.
Av økologiske interaksjoner er det negative endringer i adferd og fysiologi hos kysttorsk, samt økt sykdomsforekomst som følge av smitte fra oppdrettstorsk til villtorsk, det er satt søkelys på. Med dagens oppdrettsvolum vurderes det å være lav sannsynlighet for at disse hendelsene vil bidra til risiko for negative effekter på kysttorskbestandene. Det er likevel verdt å merke seg at med en økning i produksjon tilsier all erfaring at sykdomssituasjonen vil endre seg. Man må derfor anta at også oppdrettstorsken, i takt med at produksjonen øker, vil møte på utfordringer med sykdom. Det er ikke gitt at det vil være de samme kjente agensene og sykdommene som vil gjøre seg gjeldende i framtida, man må også være forberedt på at ukjente og/eller nye varianter av kjente agens vil skape utfordringer. Denne type overraskelser med mulig katastrofale konsekvenser og høy usikkerhet i form av manglende kunnskap, har vi valgt å markere med sorte svaner i risikovurderingen.
Til tross for intensivert forskningsaktivitet de siste årene, er det fortsatt mangel på kunnskap for en rekke av risikokildene og hendelsene som er identifisert i denne risikovurderingen. Mangel på kunnskap skaper usikkerhet og resultater som bygger på svak kunnskap, kan skjule deler av risikobildet gjennom eksempelvis feilaktige antakelser. Vi har fått mer kunnskap om modning og gyting i merd gjennom målrettet forskning, men fortsatt er kunnskapen svak i forhold til hvorvidt egg fra gyting i merd vil ende opp i gytende individer, som igjen vil kunne krysse seg inn i de lokale kysttorskbestandene. «Bruk av kunstig belysning for å utsette kjønnsmodning» er en ny risikokilde vi har valgt å ta med i årets vurdering. Vi antar at kunstig belysning kan bidra til at kysttorsk tiltrekkes oppdrettsanlegg og endrer adferd, men det er foreløpig lite kunnskap om dette. Å bygge opp denne type kunnskap tar tid og det vil være nødvendig å identifisere og prioritere de viktigste kunnskapshullene i den videre utvikling av næringen.
Det vil forekomme tekstlikhet og gjenbruk av test fra tidligere utgaver av Havforskningsinstituttet sin risikovurdering av torskeoppdrett, inkludert tilhørende kunnskapstatus.
1 - Risikovurdering
1.1 - Innledning
1.1.1 - Problemstilling
Både politikere og myndigheter har ytret ønske om å diversifisere havbruksnæringen som en del av en påtenkt fremtidig produksjonsøkning, og torsk antas å være en av artene med størst potensial. I 2022 kom Havforskningsinstituttet med den første risikovurderingen av effekter av torskeoppdrett på kysttorsk, med tilhørende rapport for kunnskapsstatus. Siden 2022 har produksjonen av oppdrettstorsk økt jevnt, og i 2024 ble det slaktet 14 683 tonn rund vekt oppdrettstorsk. Ved utgangen av september 2025 er det slaktet 13 765 tonn oppdrettstorsk (Fiskeridirektoratet, 20.10.2025).
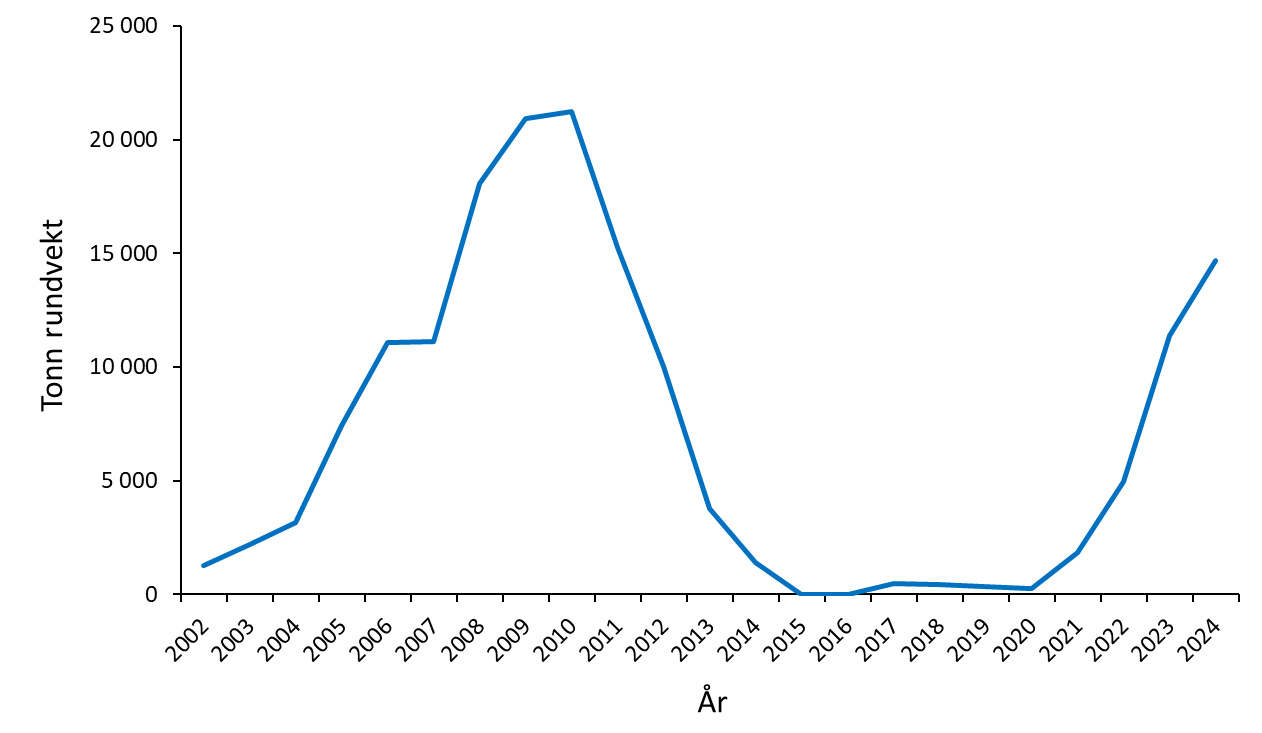
I 2024 og 2025 har det vært 11-12 aktive sjølokaliteter hvor det produseres oppdrettstorsk. Disse har en samlet total kapasitet på 41 335 tonn. Ved utgangen av august 2025 var det gitt tillatelse til en økning i produksjon med drøyt 12 000 tonn fordelt på fire lokaliteter, hvorav én er en utvidelse av allerede eksisterende lokalitet. Per 9. oktober 2025, foreligger det søknader om 18 nye lokaliteter med åpne merder i sjø til behandling. Disse har en samlet omsøkt biomasseøkning på 56 915 tonn. Sammen med nåværende godkjente kapasitet på 108 291 tonn, vil dette utgjøre en total kapasitet på 165 206 tonn, hvis alle søknadene godkjennes (tabell 1.1).
| Forvaltnings-område: | Kapasitet samlet oversikt for matfisk (tonn): | |||
| Godkjent kapasitet pr. 09.10.2025 ** | I drift pr. 09.10.2025 | Søknader* | Samlet: | |
| Sør for 62°N | 7 340 | 5 780 | 4 379 | 11 719 |
| 62-67°N | 84 033 | 30 955 | 37 879 | 121 912 |
| Nord for 67°N | 16 918 | 4 600 | 14 657 | 31 575 |
| SUM: | 108 291 | 41 335 | 56 915 | 165 206 |
I utgangspunktet vil miljøeffektene fra torskeoppdrett være tilsvarende som for lakseoppdrett, men siden villtorsken lever hele sitt liv i det marine miljø, med større fysisk nærhet til oppdrettsanleggene enn villaksen, vil noen av risikokildene, hendelsene og konsekvensene skille seg fra de vi har identifisert for laks.
Det er bestandene av kysttorsk, ikke nordsjøtorsk og den nordøstarktiske torsken (heretter skrei), som vurderes som mest utsatt for negativ påvirkning fra torskeoppdrett.
Frem til 2021 ble kysttorsk forvaltet i to geografiske områder; nord og sør for 62°N. Etter en metoderevisjon i 2021 ble det nordligste området delt i to ved 67°N og vi har nå tre forvaltningsområder; sør for 62°N (Skagerrak og Vestlandet), 62°N – 67°N (fra Sildegapet like sør for Stadlandet til Fugløyfjorden sørvest for Bodø) og nord for 67°N (nordlige del av Nordland inkludert Lofoten og Vesterålen samt Troms og Finnmark) (figur 1.1).
I forbindelse med innføring av trafikklyssystemet som verktøy for regulering av vekst i oppdrettsnæringen, ble norskekysten delt inn i 13 ulike produksjonsområder (PO). Produksjon av torsk eller søknader om torskeoppdrett er ikke jevnt fordelt mellom disse. Ved utgangen av august 2025 foreligger det ingen godkjente torskelokaliteter eller søknader om torskeoppdrett i PO1, 2, 11 og 13. I PO3, 7, 9, 11 er det per i dag ingen aktive lokaliteter med fisk i sjø (produksjon av torsk), men det foreligger godkjente lokaliteter i tillegg til at det er nye søknader til behandling (tabell 6.1 kunnskapsstatus). Forvaltningsområdet sør for 62°N, mellom 62°N til 67°N, og nord for 67°N overlapper delvis med ulike produksjonsområder (PO); hhv PO 1-4, PO 5-8, og PO 9-13.
Det har vært knyttet bekymring til økt produksjon av oppdrettstorsk, blant annet fra fiskerinæringen. Selv om det har vært gjennomført ny forskning siden forrige risikovurdering i 2022, er det fortsatt mangel på kunnskap som gjør vurderingen av påvirkninger fra dagens og morgendagens torskeoppdrett utfordrende. Dette gjelder både for denne risikovurderingen og for forvaltningen som skal ta beslutningene om det er forsvarlig å øke produksjonen av torsk i oppdrett fra dagens nivå. Kunnskapshull og manglende innsikt skaper usikkerhet og bidrar dermed til økt risiko, noe vi utdyper for hver av de aktuelle risikokildene.
1.1.2 - Mål og omfang
Målsetningen med denne risikovurderingen er å vurdere risiko for «Negative effekter på kysttorskbestander som følge av torskeoppdrett i åpne merder i sjø» med hovedvekt på endringer i adferd, fysiologi, økt sykdomsforekomst og genetisk påvirkning.
Utslipp av næringssalter, partikulært organisk materiale, miljøgifter eller legemidler er ikke inkludert i denne vurderingen, men vil i all hovedsak ha samme miljøeffekter som beskrevet for lakseoppdrett. Velferd hos oppdrettstorsk vurderes ikke, men vi henviser til vurderingen som ble gjort i 2022, Risikovurdering av velferd til oppdrettstorsk. En ny risikovurdering av fangstbasert akvakultur (FBA) og mellomlagring av torsk inngår ikke i denne rapporten. Dette er vurdert som en stabil næring og mengden torsk i denne typen anlegg har gått ned siden forrige risikovurdering. Inntil premissene for vurderingen som ble gjort i 2022 endrer seg, henviser vi til «Risikovurdering av effekter på ville kysttorskebestander ved mellomlagring og fangstbasert akvakultur (FBA)».
I denne risikovurderingen har vi valgt å sette søkelys på kysttorsk og ikke inkludere skrei av to hovedgrunner. Den ene er de svake bestandsnivåene for kysttorsk sør for 67°N, den andre er at kysttorsken er mer stasjonær enn skreien, noe som gjør at sårbarheten for lokal påvirkning øker. I tillegg er det lettere å vurdere effekter fra lokale/regionale oppdrettsanlegg på bestander som gyter og oppholder seg i området det meste eller hele året.
Risikovurderingen omfatter oppdrett av torsk i åpne merder i sjø. Med åpne merder i sjø menes anlegg hvor det benyttes nøter for hold av fisken og det er fri gjennomstrømming av vannmassene. Risikoen for negative effekter av oppdrettsaktiviteten avhenger av produksjonsintensiteten (tildelt kapasitet av torsk i oppdrett innenfor en aktuell region). I risikovurderingen av torskeoppdrett fra 2022 tok vi utgangspunkt i samlet tillatelseskapasitet i eksisterende og omsøkte anlegg, det vil si den maksimale mulige produksjonen. I årets risikovurdering tar vi utgangspunkt i nåværende produksjon, utviklingen som har vært siden forrige risikovurdering og med et tidsperspektiv på fem år frem i tid (2026-2030).
Det er et uttalt mål fra torskenæringen (Fagseminar i regi av CodCluster 12.05.25) at det produseres 75 000 tonn torsk innen 2030, men næringen er også klare på at denne målsetningen per nå ikke ligger an til å nås.
Mesteparten av tillatt produsert biomasse per i dag er uutnyttet og ikke i drift. Disse tillatelsene, slik vi ser det, kan i prinsippet tas i bruk når som helst. Selv om det er stor usikkerhet knyttet til om og når disse vil tas i bruk, må de likevel hensyntas i en risikovurdering siden produksjonsvolum er en viktig driver i risikobildet. Det vil aldri stå maksimal biomasse i sjøen til enhver tid. For å sikre en mest mulig jevn utnyttelse av maksimalt tillatt biomasse (MTB) og jevnlig utslakting, settes fisk ut på ulike lokaliteter til ulike tider. Det også krav om en brakkleggingsperiode på minimum to måneder mellom utsett. På bakgrunn av dette antar vi en kapasitetsutnyttelse på 40 %, som er lagt til grunn for estimering av produksjon i de respektive forvaltningsområdene. Basert på tallene i tabell 1.1. for samlet kapasitet, utgjør dette henholdsvis 4 688 tonn sør for 62°N, 48 765 tonn mellom 62°N og 67°N og 12 630 tonn nord for 67°N, rundet opp til nærmeste hele 1000 tonn i risikokartene. Hovedtyngden av oppdrettsaktiviteten foregår i forvaltningsområdet mellom 62°N og 67°N (tabell 1.2, figur 1.1).
Det foreligger to søknader om lukkede anlegg i sjø. Nærmere beskrivelse av hvilken merdteknologi som er planlagt for disse er ikke tilgjengelig, men vi velger likevel å ikke inkludere kapasiteten i disse søknadene i risikovurderingen.
| Forvaltnings-område: | Antall lokaliteter matfisk | |||
| Totalt antall lokalitets-tillatelser 09.10.2025 | I drift pr. 09.10.2025 | Søknader om nye lokaliteter * | Samlet: | |
| Sør for 62°N | 2 | 1 | 2 | 4 |
| 62-67°N | 29 | 9 | 12 | 41 |
| Nord for 67°N | 8 | 2 | 4 | 12 |
| SUM: | 39 | 12 | 18 | 57 |
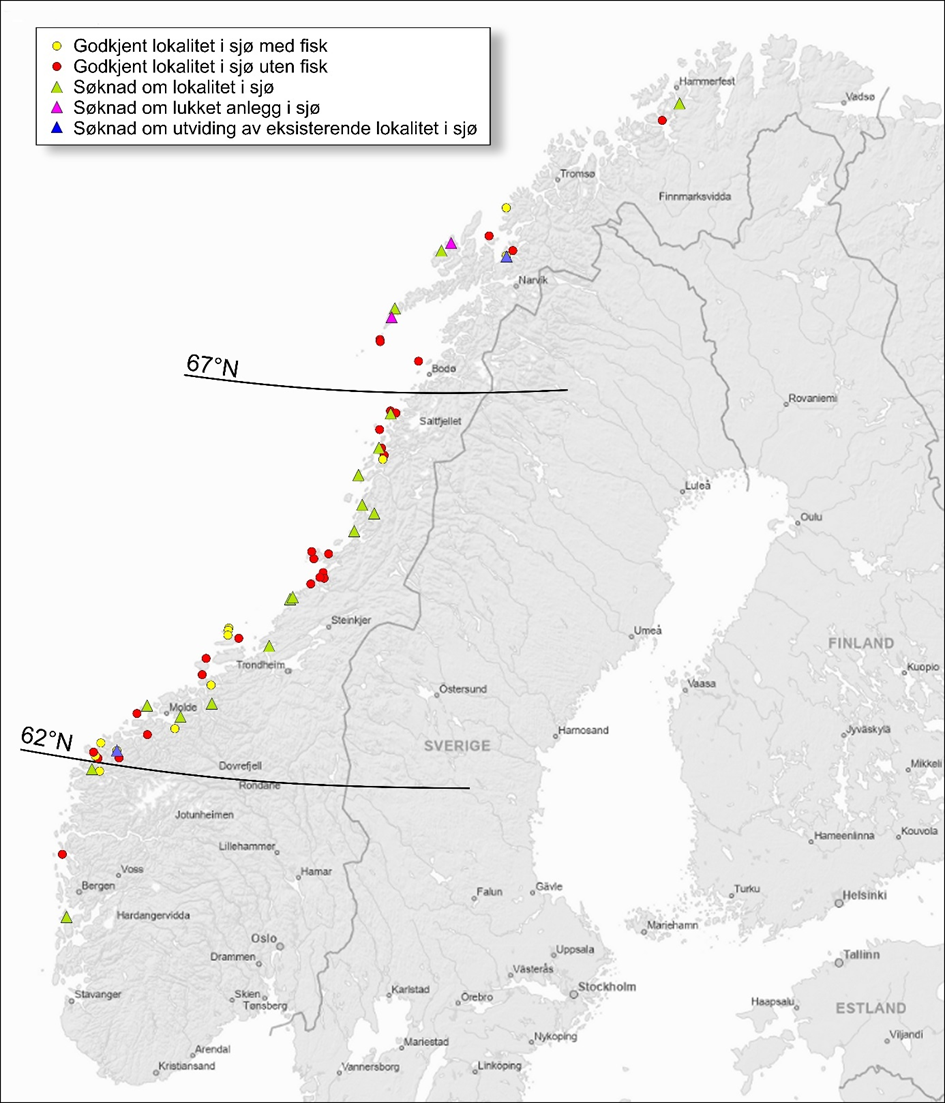
I 2025 er det kun ett torskeoppdrettsanlegg i drift i forvaltningsområdet sør for 62°N og det foreligger kun to søknader om torskeoppdrett i dette området. Det vurderes derfor som lite trolig at det blir noen vesentlig økning i produksjon i dette forvaltningsområdet de neste årene. Det er derfor ikke gjort en risikovurdering for forvaltningsområdet sør for 62°N.
Om situasjonen og premissene for årets risikovurdering skulle endre seg i vesentlig grad innen 2030, vil det være behov for å gjøre en ny vurdering.
1.2 - Metodikk for risikovurdering
I denne risikovurderingen kartlegges og analyseres risiko for «Negative effekter på kysttorskbestander som følge av torskeoppdrett i åpne merder i sjø».
Metodikken som ligger til grunn for risikoresultatene presentert i denne rapporten er utviklet av Havforskningsinstituttet og publisert i Andersen mfl. (2022). I dette kapittelet oppsummeres hovedpunktene i metodikken med fokus på å forstå resultatene av risikovurderingen, hvordan de er fremkommet og hva de bygger på. For detaljer rundt grunnleggende elementer i metodikken og hvordan risikovurderingen gjennomføres henviser vi til artikkelen.
I takt med forskningsfronten innen risikofaget finnes ingen objektive og korrekte risikotall som utgjør en form for fasit, det finnes kun usikkerhet knyttet til hva som ligger foran oss. Og det er nettopp denne usikkerheten fagekspertene sier noe om i denne rapporten. Målet med risikovurderingen er å skape risikoforståelse og risikoerkjennelse hos forvaltningen og andre interessenter som utgangspunkt for prioriteringer og beslutninger om veivalg og tiltak. Det er samtidig avgjørende at beslutningstakere og andre som skal benytte resultatene fra risikovurderingen, forstår at ufullstendig informasjon, utilstrekkelig kunnskap, hypoteser og antakelser er en del av, og i stor grad karakteriserer en slik analyse. Vi har valgt å visualisere resultatet av risikovurderingene i form av grafiske hierarkiske strukturer (risikokart) som er ment å gi en hurtig og intuitiv forståelse for risikokilder, hendelser og mulige konsekvenser med tilhørende usikkerhet knyttet til næringens aktivitet. Det er imidlertid argumentasjonen som ligger til grunn for risikokartene som skal skape tillit og eventuelt overbevise leseren om at risikovurderingen gir mening.
Fundamentale forhold knyttet til risikoanalysefaget som terminologi, definisjoner og metodisk tilnærming er forankret i Society for Risk Analysis, glossary (SRA, 2018); «Risk, Surprises and Black Swans – Fundamental Ideas and Concepts in Risk Assessment and Risk Management» (Aven, 2014) og NS 5814:2021 + AC:2023.
Risiko defineres i tråd med (SRA, 2018) som følger:
Med risiko menes konsekvenser (C) av aktiviteten med tilhørende usikkerhet (U)
Risikobegrepet introduserer dermed «konsekvenser» og «usikkerhet» - (C,U) som to sentrale aspekter som henger sammen. Med «usikkerhet» menes her en kunnskapsbasert usikkerhet (epistemisk) som knyttes til det å ikke vite fremtidige konsekvenser. Risikovurderingene i denne rapporten måler og beskriver usikkerhet ved hjelp av subjektive sannsynligheter og kunnskapsstyrke knyttet til negative effekter på kysttorskbestander som følge av påvirkning fra torskeoppdrett. Vi måler og beskriver risiko ved (C’,Q,K) der C’ er spesifikke konsekvenser av aktiviteten, Q er et verktøy for å måle usikkerhet, og K er bakgrunnskunnskapen som vurderinger av C’ og Q bygger på. Vi velger å splitte opp de spesifikke konsekvensene C’, i risikokilder RS’, og hendelser A’. Risiko beskrives da ved de fem variablene (RS’,A’,C’,Q,K). Vi måler usikkerheten Q ved hjelp av subjektive sannsynligheter P og vurderinger av kunnskapsstyrke SoK (Strength of Knowledge). Betraktninger om P og SoK hviler på bakgrunnskunnskapen K, og vi skriver Q (P,SoK,K).
For denne aktuelle problemstillingen relateres usikkerheten eksempelvis til:
-
Genetiske endringer i kysttorskbestandene
-
Status på kysttorskbestandene
-
Negative endringer i fysiologi og adferd hos kysttorsk
-
Sykdomsutbrudd i oppdrett og smittespredning til villfisk
Usikkerheten knyttes til hva som kan skje, hvor ofte, samt til omfang og alvorlighetsgrad av konsekvensene. Vi kan måle denne usikkerheten ved hjelp av sannsynlighetsbetraktninger og forventningsverdier forankret i historiske data. Imidlertid vil en tilnærming der vi utelukkende bruker sannsynligheter som mål på usikkerhet, kunne føre til uheldige forenklinger og tap av viktige nyanser. I denne risikovurderingen søker forskerne å legge all tilgjengelig kunnskap til grunn, samt karakterisere denne, for å øke innsikt og forståelse. Søkelys på usikkerhet, herunder manglende kunnskap knyttet til mulige kausale sammenhenger og konsekvenser fremover i tid, bidrar til effektiv utnyttelse av all tilgjengelig innsikt og stimulerer til videre utvikling av kunnskapsgrunnlaget.
Styrken til bakgrunnskunnskapen (i forhold til RS’, A’, og C’) vurderes eksplisitt ved SoK, det vil si en vurdering av hvorvidt kunnskapen som danner grunnlaget for en bestemt konklusjon, er sterk eller svak. Resultater som bygger på svak kunnskap, kan skjule deler av risikobildet gjennom eksempelvis feilaktige antakelser. Ved å inkludere en redegjørelse for kunnskapsstyrken i analysen, gis beslutningstakere mulighet til å ta stilling til hvorvidt kunnskapen på et område er tilstrekkelig som fundament for strategiske og operasjonelle veivalg. I motsatt fall vil det være naturlig å vurdere en føre-var-strategi og diskutere tiltak som bidrar til å styrke kunnskapsgrunnlaget.
I arbeidet med denne rapporten har vi systematisert og strukturert risikokildene (RS’,A’,C’) knyttet til negative effekter på kysttorskbestander som følge av påvirkning fra torskeoppdrett i åpne merder. Det er lagt vekt på å inkludere all tilgjengelig kunnskap som utgangspunkt for å uttrykke og formidle risiko. Kunnskapsgrunnlaget varierer mellom de ulike risikokildene, avhengig av om det er gjennomført mye vitenskapelige undersøkelser og om det eksisterer gode rapporteringssystemer og/eller overvåkningsprogrammer i tillegg til tidligere erfaringer i lignende situasjoner.
Bayesianske nettverk er valgt som et kvalitativt verktøy for å visualisere risiko, herunder; kausale sammenhenger mellom RS’, A’ og C’; sannsynligheter P knyttet til hvorvidt disse inntreffer, og styrken på kunnskapen SoK som disse vurderingene bygger på (figur 1.2.1). Hensikten er at de grafiske strukturene (kalt risikokart) og tilhørende argumentasjon skal bidra til hurtig og intuitiv risikoforståelse.

I risikokartene benyttes fargekodene grønn, gul og rød farge på en node for å illustrere, lav, moderat og høy sannsynlighet. Fargen på sirkelen rundt noden illustrerer hvorvidt denne vurderingen bygger på sterk (grønn), moderat (oransje) eller svak (rød) kunnskap (SoK) (figur 1.2).
Forskerne benytter all tilgjengelig kunnskap, samt refleksjoner rundt kunnskap vi eventuelt mangler i arbeidet med å identifisere RS’, A’ og C’ og fastsette sannsynligheter for at de inntreffer. Eksempelvis er sannsynligheten for at «Høyt antall oppdrettstorsk rømmer» i figur 1.2 vurdert som moderat og fargekode gul benyttes på noden. Det er kunnskapsbasert usikkerhet knyttet til omfanget av feil og mangler i rapporteringen og forskerne argumenterer for at sannsynlighetsvurderingen av risikokilden bygger på moderat kunnskap. Fargekoden oransje benyttes på sirkelen rundt noden.
Et annet eksempel som vist i figur 1.2. er risikokilden «Høyt antall sykdomsutbrudd og smittespredning i merd» der sannsynlighetsvurderingen baseres på kunnskap om i hvilken grad det er grunn til å tro at det vil forekomme et høyt antall sykdomsutbrudd og spredning av smitte fra oppdrettstorsk til kysttorsken. I eksempelet vurderes sannsynligheten som lav for omfattende smitte fra oppdrettstorsk til kysttorsk og fargekode grønn benyttes. Det er ikke full oversikt over sykdomsutbrudd i torskeoppdrett og kunnskapsstyrken vurderes som moderat. Fra tid til annen forekommer det utbrudd av «nye», hittil ukjente sykdommer, eller sykdommer som er kjente, men nye for en gitt art eller i et område. Konsekvensene vil derfor variere, men i enkelte tilfeller kan de bli svært alvorlige for både norsk oppdretts- og villfisk. Denne typen hendelser kan gi opphav til en potensiell overraskelse med alvorlige konsekvenser og er visualisert med svarte svaner i risikokartet.
Hendelsen «Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettstorsk» påvirkes i all hovedsak av tre underliggende risikokilder der sannsynligheten for at disse skal inntreffe varierer fra moderat til lav. For å forstå hvordan hendelsen «Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettstorsk» kan vurderes som lite sannsynlig (har lav sannsynlighet) må man lese forskernes argumentasjon. I dette tilfellet er forklaringen at produksjonen vil være fordelt på relativt få antall lokaliteter og anlegg og er spredt over et stort geografisk område. Det totale smittepresset innenfor forvaltningsområdet vurderes derfor til, generelt sett, å være lavt. Dette veier da tyngre enn de to andre risikokildene. På samme vis må leseren studere forskernes argumentasjon for å forstå hvordan en samlet vurdering av kunnskapsstyrken knyttet til underliggende risikokilder summeres opp til kunnskapsstyrken for en overliggende hendelse.
Vurderingen av konsekvensen, sammen med sannsynlighet for uønsket hendelse og kunnskapsstyrke, ligger til grunn for konklusjoner om risiko. Manometeret i figuren uttrykker i dette tilfellet moderat risiko for «Negative effekter på kysttorskbestander som følge av torskeoppdrett i åpne merder i sjø». Risikokartene er et nyttig redskap for å visualisere risikokilder og kausale sammenhenger, mens karakterisering og argumentasjon i teksten gir grunnlaget for vurdering og måling av konsekvenser, sannsynligheter, kunnskapsstyrke og risiko. Det er denne teksten som skal skape tillit og eventuelt overbevise leseren om at resultatet av risikovurderingen gir mening.
1.3 - Faktorer som medfører «Negative effekter på kysttorskbestander som følge av torskeoppdrett i åpne merder i sjø»
Faktorer som påvirker omfanget av «Negative effekter på kysttorskbestander som følge av påvirkning fra torskeoppdrett i åpne merder i sjø» er i hovedsak «Negative endringer i adferd hos kysttorsk», «Negativ endring i fysiologi hos kysttorsk», «Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettstorsk», og «Genetisk påvirkning hos kysttorskbestander» (figur 1.4).

«Negativ endring i adferd hos kysttorsk» omfatter endring i kysttorskens beite, gyte- eller vandringsadferd og påvirkes i hovedsak av de to risikokildene «Bruk av kunstig belysning for å utsette kjønnsmodning» og «Tiltrekning for beiting på spillfôr, bunndyrsamfunn og andre organismer».
“Bruk av kunstig belysning for å utsette kjønnsmodning». Lysstyring benyttes for å utsette kjønnsmodning hos oppdrettstorsk i merd. Lys er kjent for å ha en sterk tiltrekningskraft på ulike organismer i havet og benyttes blant annet i forbindelse med fiskeriaktivitet. Hvordan bruken av denne typen lys vil påvirke fisk og andre organismer i nærheten av oppdrettsanlegg er ukjent, men det antas at bruk av lys vil tiltrekke seg dyreliv av ulike arter som plankton og fisk. Denne tiltrekningen kan være både direkte (tiltrekning som følge av selve lyset) og indirekte (tiltrekning av predatorer som følge av ansamling av byttedyr).
Påvirkningen av denne typen lys vil avhenge av hvor langt ut i vannmassene rundt et oppdrettsanlegg lyset når. Dette vil variere med årstid og sikten (turbiditeten) i vannmassene. Påvirkningen vil være relativt lokal rundt det enkelte oppdrettsanlegg, men mange anlegg innenfor et mindre geografisk område vil kunne påvirke et større område som helhet, f.eks. et fjordsystem.
Det er ikke gjort vitenskapelige studier på hvordan bruk av kunstig belysning for å utsette kjønnsmodning av oppdrettstorsk vil påvirke adferden til kysttorsken. Vi antar likevel, basert på kunnskap fra andre studier på bruk av lys, at dette kan være en kilde til risiko for kysttorsken.
«Tiltrekning for beiting på spillfôr, bunndyrsamfunn og andre organismer». Det er kjent at mange dyr, også fisk og kysttorsk, tiltrekkes av fysiske installasjoner, inkludert kunstige rev. Et torskeoppdrettsanlegg kan fungere som et kunstig rev og tilby både beskyttelse og mat for vill torsk. Tettheten av fisk som samler seg rundt et oppdrettsanlegg kan henge sammen med fôrintensitet og fôringstider, og torsk kan spise betydelige mengder spillfôr. Feltstudier viser at ca. 20 % av torsken fanget i områder med stor tetthet av lakseoppdrettsanlegg hadde spesialisert seg på å spise spillfôr i form av pellets. Torsk kan også beite på dyr som samler seg rundt anlegget og oppholde seg der over lengre perioder, vandre mellom anlegg, regelmessig komme innom anlegg og også veksle mellom opphold i naturlige habitat og nært torskeoppdrettsanlegg.
Denne endringen i adferd kan føre til at kysttorsken eksempelvis velger å oppholde seg i nærheten av oppdrettsanlegg for å beite, heller enn å vandre til lokale gytefelt for å gyte. Det kan også være at kysttorsk ikke vil oppholde seg i et godt oppvekstområde som følge av oppdrettsaktivitet.
Ett studie har vist at villtorsk på gyteplassene har spist oppdrettsfôr, noe som dokumenterer at en andel av villtorsken som har beitet rundt oppdrettsanlegg, likevel vandrer til gyteplassene for å gyte. Det er behov for mer kunnskap om hvordan «Tiltrekning for beiting på spillfôr, bunndyrsamfunn og andre organismer» endrer kysttorskens adferd og hva konsekvensene av denne endringen er.
«Negative endringer i fysiologi hos kysttorsk» kan oppstå hvis kysttorsk beiter på spillfôr, bunndyr og andre organismer med endret fettsyresammensetning. Selv om det antas at individer av vill torsk ikke tar direkte skade av å spise plantebasert fôr fra oppdrettsanlegg på kort sikt, så er det sannsynlig at fôrkilder av ikke-marint opphav kan ha negative konsekvenser for villfiskens helse og reproduksjon på lengre sikt. Spesielt de essensielle fettsyrene ARA (arakidonsyre, 20:4n-6), EPA (eikosapentaensyre, 20:5n-3) og DHA (dokosaheksaensyre, 22:6n-3) er viktige for eggutvikling, befruktning, larveoverlevelse og vekst. Det er likevel stor usikkerhet knyttet til om kysttorskens beiting rundt oppdrettsanlegg har negative effekter på kysttorskbestander, eller om det kan føre til en positiv effekt på lokale torskebestander, fordi økt mattilgang og påvist økt kondisjon hos fisken kan gi økt reproduksjon.
«Negative endringer i fysiologi hos kysttorsk» påvirkes i hovedsak av «Tiltrekning for beiting på spillfôr, bunndyrsamfunn og andre organismer». Hovedattraksjonen for aggregering av villfisk rundt oppdrettsanlegg er trolig det ekstra næringstilskuddet fra spillfôr, noe som vil gi et annet næringsinnhold sammenlignet med naturlig føde. Dette kan være positivt for individuell vekst hos fisken, men kan også fungere som en økologisk felle. Økologiske feller defineres som et kunstig habitat i det naturlige miljøet som tiltrekker seg fisk, og siden fører til redusert overlevelse eller reproduksjon hos fisken.
Marine pellets til torskeoppdrett produseres for å gi maksimal vekst med minimale negative effekter på helsen til fisken. Fôrpellets som brukes til torskeoppdrett inneholder et høyere nivå av marine ingredienser og er mindre fettrike enn fôrpellets som brukes i lakseoppdrett (ca. 17% vs. 37%). Men selv om fôrprodusentene fremholder at torsken tåler høye nivå av vegetabilske ingredienser og sammensetningen av oppdrettsfôr for torsk er tilpasset en marin fisk, er det likevel vesentlig forskjellig fra torskens naturlige diett, både i fettmengde og sammensetning. Mengden av terrestriske fettsyrer som oljesyre (18:1n-9), linolsyre (18:2n-6) og α-linolensyre (18:3n-3) er høyere sammenlignet med hva som vanligvis finnes i marine dyr.
Ved Havforskningsinstituttet har det vært gjennomført eksperimentelle studier som skal kartlegge hvordan forhøyede mengder terrestriske fettsyrer fra kommersielt marint torskefôr påvirker gonademodning, fekunditet og larveutvikling hos torsk, og resultatene fra denne studien vil være klar i løpet av 2026.
Mengde pellets som utfôres ved de ulike lokalitetene er direkte avhengig av antall og størrelsen på fisken (biomasse) som står i anlegget på et gitt tidspunkt. I oppdrettsnæringen fôres fisken etter appetitt og det benyttes kameraovervåkning for å ha best mulig kontroll på utfôringen og for å unngå overfôring og spillfôr Det antas at prosentvis mengde spillfôr fra et oppdrettsanlegg er relativt likt uavhengig av art (torsk, laks) og geografisk område. Forbruk av fôr i torskeoppdrett er ikke oppgitt i Fiskeridirektoratets biomassestatistikk for matproduksjon av torsk.
Feltstudier viser at ca. 20 % av torsken fanget i områder med stor tetthet av lakseoppdrettsanlegg hadde spesialisert seg på å spise spillfôr i form av pellets. Selv om det ikke finnes dokumentasjon, antas det at det samme vil gjelde for torskeoppdrettsanlegg.
«Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettstorsk» vil kunne forekomme dersom sykdomsutbrudd i åpne merder fører til forhøyet smittepress på mottakelig villfisk enten gjennom horisontal eller vertikal smitte. Om mange nok individer (del av gytebestanden, eller yngelen) blir kraftig svekket eller dør av sykdom som følge av smitte fra oppdrett, kan det i ytterste konsekvens føre til økt dødelighet ut over den naturlige dødeligheten i populasjonen og dermed negative effekter på kysttorskbestandene. Dette gjelder særlig for smittespredning av nye og endrede agens. Fra tid til annen forekommer det utbrudd av «nye», hittil ukjente sykdommer, eller sykdommer som er kjente, men nye for en gitt art eller i et område. Konsekvensene vil derfor variere, men i enkelte tilfeller kan de bli svært alvorlige for både oppdretts- og villfisk. Denne typen hendelser gir opphav til en potensiell overraskelse med alvorlige konsekvenser og er visualisert med svarte svaner i risikokartet (figur 1.3). Økt sykdomsforekomst avhenger av om det er et «Høyt antall sykdomsutbrudd og smittespredning i merd», om det forekommer «Tiltrekning for beiting på spillfôr, bunndyrsamfunn og andre organismer» og hvorvidt et «Høyt antall oppdrettstorsk rømmer».
«Høyt antall sykdomsutbrudd og smittespredning i merd». Som hos et hvert annet husdyr, vil det også i oppdrett av fisk forekomme sykdom og smittespredning. Under et sykdomsutbrudd i merd vil syk fisk skille ut smittestoff og det oppstår et økt smittepress i merden, anlegget og i omgivelsene. Smitte kan spres med vannmassene til mottakelig villfisk i nærhetene av oppdrettsmerdene (horisontal smitte).
Lukkede anlegg i sjø vurderes som en løsning for mange av miljøutfordringene innen oppdrett, inkludert smittespredning. Slike anlegg vil kreve god vanngjennomstrømning og vil ikke nødvendigvis være lukket i den forstand at de vil hindre vannbåren smitte å slippes ut til omgivelsene. Dette vil være avhengig av i hvilken grad avløpsvannet fra et lukket merdanlegg filtreres og desinfiseres. I den nye ordningen for lukkede anlegg som nylig er lagt frem av regjeringen, er det ikke krav om rensing/desinfisering av avløpsvann.
Sannsynligheten for at det vil komme et sykdomsutbrudd med tilhørende smittespredning i en oppdrettsmerd med torsk vurderes å være uavhengig av geografisk område. Antallet sykdomsutbrudd og dermed også smittespredning som kan forventes innenfor et gitt geografisk område vil avhenge av antallet oppdrettsanlegg og mengde fisk i sjø.
«Tiltrekning for beiting på spillfôr, bunndyrsamfunn og andre organismer». Som ved all horisontal smittespredning forutsetter dette at det er overlapp i tid og rom mellom en mottakelig vert og høy nok smittedose. Kysttorsk som tiltrekkes og beiter ved torskeoppdrettsanlegg kan dermed være utsatt for smitte ved sykdomsutbrudd og bidra til å spre smitte til kysttorskbestandene i området. Et eksempel kan være smitte til torskeyngel som er stasjonære i de lokale oppvekstområdene over tid. Dette kan føre til økt dødelighet ut over den naturlige dødeligheten i populasjonen. Økt smittepress til miljøet vil kunne bidra til oppbygging av reservoar. De mulige negative effektene som smitte fra torskeoppdrett kan ha i omgivelsene, vil avhenge av typen smitte (agens), forekomst av mottakelige arter i området og ulike miljøfaktorer (f.eks. temperatur, saltholdighet og strøm). Overlevelse av et agens i miljøet er en av nøkkelfaktorene for spredningspotensialet, og dette varierer mellom ulike agens.
«Høyt antall oppdrettstorsk rømmer». Antallet oppdrettstorsk som har rømt de siste årene har vært dominert av fåtallige men store hendelser. I 2022 ble det meldt inn ca. 87 000 rømt torsk til Fiskeridirektoratet, i 2023 var det 900, og i 2024 ble det meldt inn 30 000 rømt torsk. For 2025 er det per medio oktober rapportert inn 6 rømmingshendelser med torsk, men antall rømte fisk er ikke slått fast enda. Basert på erfaringer med rømmingstall fra lakseoppdrett, er det grunn til å tro at antall rømte torsk er noe høyere enn det som er innrapportert.
Det er bred enighet om at laksenøter ikke egner seg til produksjon av torsk. For å minske risiko for rømming har det vært mye fokus på å optimalisere typen not til bruk i oppdrett av torsk. Dette gjelder spesielt kvaliteten på selve notlinen, måten noten er sydd sammen på (forsterkede sømmer og sikringer) og at man unngår løse tråder som torsken har vist seg å kunne nappe i.
Annen merdteknologi, som bruk av lukkede anlegg i sjø vil sannsynligvis hindre drypprømming av torsk ved at den ikke kan beite seg vei ut gjennom notveggen, men det vil ikke hindre rømming om det skulle forekomme et havari av merden.
Dersom rømt oppdrettstorsk er smittebærer eller syk, vil den kunne spre smittestoff direkte ut i vannmassene eller overføre smittestoff om den kommer i nærkontakt med eller blir spist av kysttorsk. Det samme gjelder om syk oppdrettstorsk dør etter rømming. Det er også mulig med vertikal overføring av enkelte agens, som nodavirus, dersom smittet oppdrettstorsk, som er bærer av viruset, gyter med frisk kysttorsk.
«Genetisk påvirkning på kysttorskbestandene». Vill kysttorsk har en komplisert bestandsstruktur med lokale bestander som i større eller mindre grad skiller seg genetisk fra hverandre. Genetisk påvirkning på ville bestander skjer når oppdrettstorsk gyter sammen med villtorsk og får levedyktig avkom som blir en del av den ville bestanden. Det er påvist at befruktede egg fra oppdrettstorsk vil kunne unnslippe merdene og overleve til kjønnsmoden torsk. Det er også påvist at denne torsken gyter med hverandre, men det er ikke sikkert påvist at torsk med opphav fra gyting i merd faktisk gyter med villtorsk. Data fra en undersøkelse indikerer at krysninger mellom vill- og oppdrettstorsk kan ha funnet sted, men dette mangler det solid dokumentasjon på. Adferdsstudier i tankforsøk og merkeforsøk på gytefelt med oppdrettstorsk og villtorsk har imidlertid vist at det er sannsynlig at gyting kan skje mellom vill- og oppdrettstorsk. Basert på erfaring fra andre arter kan avkom av avlet fisk forventes å ha redusert overlevelse i naturen, men dette er foreløpig ikke dokumentert for oppdrettstorsk. Sannsynligheten for «Genetisk påvirkning på kysttorskbestandene» avhenger av hvorvidt et «Høyt antall oppdrettstorsk rømmer», om det forekommer «Betydelig gyting i merd», om det er «Betydelig overlevelse av avkom gytt i merd til gytende individer» og hvorvidt det er en «Svak tilstand hos kysttorskbestandene».
«Høyt antall oppdrettstorsk rømmer». Rømt oppdrettstorsk kan spre seg raskt utover relativt store områder rundt anlegget og kan oppsøke lokale gyte- og oppvekstområder, spesielt i løpet av gyteperioden for lokale torskebestander. Torsk som rømmer som småfisk er i større grad utsatt for predasjon fra vill torsk, sei og andre arter rundt anleggene og kan ha høy dødelighet. Oppdrettsanleggene for torsk ligger i umiddelbar nærhet til naturlige oppvekst/leveområder, og relativt nært gytefelt for ville artsfrender. Dette er i kontrast til laks som har vesentlige hindre å passere (må vandre til og opp elver, endre miljø fra salt til ferskvann) for å kunne gyte. Derfor er det for torsk et større potensiale enn hos laks for genetisk påvirkning på ville bestander ved en rømmingshendelse.
«Betydelig gyting i merd». Mengden egg gytt i merd vil være avhengig av hvor mange oppdrettstorsk som kjønnsmodner i et oppdrettsanlegg. Med utgangspunkt i data fra Fiskeridirektoratets overvåkning og forsøk ved Havforskningsinstituttet har man begynt å få oversikt over andelen oppdrettstorsk som modner og gyter i merd. Det er vist at andelen gytende og utgytte torsk i merd og på slakteri, med tilfeldig utvelgelse, var henholdsvis 16 og 30 % i gjennomsnitt i perioden desember 2024 til juli 2025. Innenfor disse gruppene var det en overvekt av hanner, men likevel en andel på 21 % hunner (i merdene) og 34 % hunner (på slakteri).
Bruk av nedsenkbar merdteknologi for produksjon av torsk kan gi en mer jevn og stabil produksjonstemperatur og man vil unngå høye temperaturtopper i sommerhalvåret. Dette kan gjøre at temperaturen gjennom produksjonssyklusen blir mer optimal i forhold til gonadeutvikling (økt sannsynlighet for uønsket vellykket gyting i merd). Samtidig vil det være en fordel med nedsenktbare merder i forhold til lysstyring, siden fisken i mindre grad vil eksponeres for naturlig lys.
Lukkede anlegg i sjø vil kreve god vanngjennomstrømning og vil ikke nødvendigvis være lukket i den forstand at de vil hindre egg og larver fra gyting i merd i å slippe ut til omgivelsene. Dette vil være avhengig av i hvilken grad avløpsvannet fra et lukket merdanlegg filtreres og desinfiseres. Hvis vannet inne i merdene har høyt nok saltinnhold kan egg, på grunn av oppdrift, til en viss grad fanges inne i slike merder. Men dette vil ikke være tilfelle for larvene etter at eggene har klekket. Bruk av lystett duk og tak på slike merder kan potensielt gi bedre mulighet for kontroll med kjønnsmodning ved bruk av lysstyring, men det er så langt ingen erfaringer med slike anlegg med hensyn til gyting i merd.
«Betydelig overlevelse av avkom gytt i merd til gytende individer». Egg gytt fra villfisk har stor dødelighet, og normalt er det kun brøkdelen av en promille som overlever til voksen fisk. Mengden av avkom fra oppdrettstorsk som overlever til gytende individer avhenger i hovedsak av tre faktorer: (i) at egg gytt i merd har tilstrekkelig eggkvalitet slik at de overlever til yngelstadiet, (ii) at gyteproduktene spres til oppvekstområder for villtorsk og (iii) at avkommene overlever til kjønnsmodning.
Fôret som oppdrettstorsken spiser i vekstfasen, består i høy grad av plantebaserte ingredienser, og dette er forskjellig fra stamfiskfôr som er optimalisert med hensyn til god embryo- og larveutvikling. Det er vist at egg gytt i merd av en domestisert torsk er av dårligere kvalitet og har en lavere klekkeprosent enn egg fra villtorsk. Overlevelsen fra egg til plommesekklarve er vist å være vesentlig lavere for avlet oppdrettstorsk (29 %) enn for vill torsk (76 %). Ubalanse i makro- og mikronæringsstoffer kan bidra til redusert levedyktighet av egg som gytes i merd, men det mangler dokumentasjon på dette.
Ved gyting i merd vil havstrømmene kunne føre egg og larver bort fra merden. Mengden av egg og larver fra oppdrettstorsk som spres til gunstige oppvekstområder for kysttorsk avhenger av mengden gyteprodukter og strømforholdene. Strømforhold og hydrografi vil være forskjellig på ulike lokaliteter, og sannsynligheten for spredning til gunstige oppvekstområder vil derfor variere.
Vi vet lite om hva som skjer med torsken fra yngelstadiet til voksne individer, men fysiske miljøforhold, predasjon og tilgang på næring vil være regulerende faktorer for hvor mye torsk som overlever og vokser opp. Videre overlevelse til kjønnsmodne individer forventes også å være lavere for oppdrettstorsk enn for villtorsk, men dette er ikke undersøkt.
«Svak tilstand hos kysttorskbestandene». Sør for Stadt (62°N) har fiskepresset over lang tid vært høyere enn bestandene har tålt, og det har vært nødvendig med sterke fiskerestriksjoner. Til tross for dette er det fortsatt en negativ trend for torskebestandene sør for 62°N.
Nord for Stadt har størrelsen på kysttorskbestandene variert mye fra år til år uten en klar trend. En gjenoppbyggingsplan ble foreslått i 2011 men har så langt ikke vært vellykket. Derfor har en ny vurdering av status for kysttorsk nord for Stadt blitt utført i regi av det internasjonale havforskningsrådet (ICES). Her ble kysttorsk nord for 62°N, som før var en forvaltningsenhet, delt i to ulike enheter med separat rådgivning: fra 62°N til 67°N (PO 5-8), og nord for 67°N (PO 9-13). For kysttorskbestanden mellom 62°N til 67°N anses bestanden å være svak, mens bestanden nord for 67°N anses å være i god forfatning.
Størrelse (biomasse) og tilstand (gytebestandsmål) kan brukes til å si noe om robustheten av bestandene i de ulike forvaltningsenhetene, men for kysttorsk i de to sørligste sonene er det knyttet stor usikkerhet rundt tilstanden. En liten bestand med få individer og lav robusthet kan være vanskelig å dokumentere. Kartlegging av gytefelt for kysttorsk indikerer at det kan finnes mange små lokale gytefelt for små lokale bestander langs hele kysten.
1.4 - Risikovurdering «Negative effekter på kysttorskbestander som følge av påvirkning fra torskeoppdrett i åpne merder i sjø»
1.4.1 - Negative effekter på kysttorskbestander mellom 62°N og 67°N som følge av torskeoppdrett i åpne merder i sjø ved en produksjon på 49 000 tonn

«Bruk av kunstig belysning for å utsette kjønnsmodning». Lysstyring brukes i torskeoppdrett for å forsinke og redusere graden av kjønnsmodning. Det finnes for lite kunnskap til at vi i denne analysen gjør en vurdering av denne risikokilden. Vi velger likevel å ha med noden da vi antar at den kan ha en påvirkning på kysttorskens adferd. Manglende datagrunnlag og dokumentasjon visualiseres med en hvit node med en rød ring.
«Tiltrekning for beiting på spillfôr, bunndyrsamfunn og andre organismer». I områder med mange oppdrettsanlegg er det observert at rundt 20 % av kysttorsken spesialiserer seg på beiting under anleggene. Det vurderes derfor at det er høy sannsynlighet for at kysttorsk vil tiltrekkes til oppdrettsanlegg og beite på spillfôr og byttedyr. Det er lite eller ingen dokumentasjon på tiltrekning av kysttorsk til torskeoppdrett, men basert på en rekke studier på tiltrekning til lakseoppdrettsanlegg som viser dette, vurderes kunnskapsstyrken som sterk.
«Negative endringer i fysiologi hos kysttorsk». Selv om det kan påvises endringer i fettsyresammensetningen vektlegges det at torskefôr er utviklet for å tilfredsstille ernæringsbehovet til torsk. I tillegg må det tas i betraktning at med en estimert produksjon av torsk på 49 000 tonn, vil den totale mengden spillfôr være lav. Det vurderes derfor at det er lav sannsynlighet for at villfisk som beiter på spillfôr og andre organismer rundt torskeoppdrettsanlegg vil føre til en «Negativ endringer i fysiologi hos kysttorsk».
Foreløpige resultater viser at oppdrettstorsk som spiser marine pellets er i stand til å gyte levedyktige egg, selv om befruktningsratene og utviklingssuksessen til embryoene var lavere enn hos referansegruppen av villtorsk. Der er derimot lite kunnskap om konsekvensene av beiting på torskefôr og om dette kan skade reproduksjonssuksessen hos disse fiskene. Kunnskapsstyrken vurderes derfor totalt sett som moderat.
«Negativ endring i adferd hos kysttorsk». Selv om det antas at sannsynligheten for tiltrekning av kysttorsk til torskeoppdrettsanlegg i forbindelse med beiteaktivitet er høy, vektlegges det at antallet og tettheten av torskeoppdrettsanlegg mellom 62°N og 67°N (som omfatter rundt 1/3 av Norskekysten) er lav, med ni anlegg per 2025 og en antatt økning til 21 anlegg i løpet av de neste fem årene. Hvordan bruk av kunstig belysning påvirker adferden til kysttorsk kan ikke vurderes ut fra nåværende kunnskap. Basert på at det er få torskeanlegg, vurderes det å være lav sannsynlighet for «Negativ endring i adferd hos kysttorsk».
Det er god kunnskap om at kysttorsk tiltrekkes oppdrettsanlegg, selv om det er lite kunnskap om tiltrekning spesifikt til torskeanlegg. Det er svak kunnskap om bruk av kunstig belysning og hvordan dette påvirker villtorskens adferd. Det mangler også studier på hvorvidt kysttorsk som beiter ved torskeoppdrettsanlegg velger bort viktige habitater knyttet til gyte- eller oppvekstområder. Kunnskapsstyrken vurderes derfor totalt sett som moderat.
«Høyt antall sykdomsutbrudd og smittespredning i merd». Antallet sykdomsutbrudd og mengden smittespredning fra oppdrettsfisk vil avhenge av antallet oppdrettsanlegg og mengden fisk i sjø. Med en estimert produksjon på 49 000 tonn oppdrettstorsk, vil antallet utbrudd være fordelt på relativt få anlegg og lokaliteter (21). Det vurderes derfor at det vil være lav sannsynlighet for et «Høyt antall sykdomsutbrudd og smittespredning i merd» i området mellom 62°N og 67°N.
Selv om det ikke er fullstendig oversikt over annet enn de meldepliktige sykdommene (sykdommer på liste F), gir fiskehelserapporten en god oversikt over sykdomssituasjonen i norsk fiskeoppdrett. Siden vi ikke kan si med sikkerhet hvilke(n) sykdom(mer) som vil gjøre seg gjeldene, hvor utbruddene vil forekomme eller omfanget av disse, vurderes kunnskapsstyrken som moderat.
«Høyt antall oppdrettstorsk rømmer». Datagrunnlaget for å vurdere sannsynligheten for store rømminger av oppdrettstorsk i et gitt geografisk område er begrenset. Dersom vi betrakter rapporterte rømminger over de siste 5 årene fra hele landet ser vi at størrelsen på rømminger varierer voldsomt, med flere små hendelser og tre store rømminger. Totalt antall rømte oppdrettstorsk de siste årene er dermed dominert av disse få store rømmingene. Siden det per nå er få (12) produksjonsanlegg i drift, har andelen store rømminger per anlegg derfor vært ganske høy. Det vurderes derfor at det er moderat sannsynlighet at det høyt antall oppdrettstorsk rømmer mellom 62°N og 67°N.
Store rømningshendelser skjer sporadisk og er ikke jevnt fordelt i tid og rom. De forventede rømningstallene for fremtiden anses som usikre. Basert på erfaringer med rømmingstall fra lakseoppdrett, er det også grunn til å tro at antall rømte torsk er noe høyere enn det som er innrapportert. I tillegg er det usikkerhet rundt hvordan rømningstiltakene næringen jobber med vil resultere i mindre rømming. Kunnskapsstyrken vurderes derfor totalt sett som moderat.
«Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettstorsk». Basert på en antagelse om at tiltrekning av villfisk til oppdrettsanleggene er høy, og at antallet rømte oppdrettstorsk vil være moderat, vektlegges det at det er få lokaliteter spredt over et stort geografisk område noe som gjør at det totale smittepresset innenfor forvaltningsområdet vil være lavt. Sannsynligheten vurderes derfor som lav for «Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettstorsk».
Siden det er usikkerhet knyttet til antallet sykdomsutbrudd og hvilke sykdommer som vil gjøre seg gjeldene, samt hvor mye oppdrettstorsk som vil rømmer vurderes kunnskapsstyrken som moderat.
«Betydelig gyting i merd». Det er vist store variasjoner i andelen oppdrettstorsk som blir kjønnsmodne og gyter i oppdrettsmerdene (målt til 1-35 % i merd, og 2-40 % på slakteri). Av disse er andelen mellom hanner og hunner ikke jevnt fordelt. Hunner modner seinere enn hannfisk og utgjorde ca. 21 % (i merd) 34% (på slakteri). Siden det er hunner som produserer egg er det antall gytende og eventuelt utgytte hunner som er relevant for denne vurderingen. Vi vurderer derfor sannsynligheten for betydelig gyting i merd som moderat.
Det er stor variasjon i modningsgrad gjennom året og mellom fiskegrupper. Det er også usikkert om hvordan utslakting av fisk som er i modning vil påvirke andelen som faktisk gyter i merd. Kunnskapsstyrken vurderes som moderat.
«Betydelig overlevelse av avkom gytt i merd til gytende individer». Selv om det finnes noe kunnskap om befruktningsgrad og overlevelse i larvefasen fra labforsøk, finnes det ingen undersøkelser om hvorvidt disse larvene fra oppdrettstorsk overlever i naturen til gytende individer. Det vurderes derfor at det ikke finnes nok kunnskap til å gjøre en sannsynlighetsvurdering av overlevelsen av avkom gytt i merd til gytende individer. Vi velger likevel å ha med denne risikokilden da den teoretisk sett kan føre til genetisk påvirkning. Manglende datagrunnlag og dokumentasjon visualiseres med en hvit node med en rød ring.
«Svak tilstand hos kysttorskbestandene». Vurdering av status for kysttorskbestanden mellom 62°N og 67°N gjort av ICES, konkluderer med lav biomasse og redusert gytebestandsmål for bestanden og det vurderes følgelig at det er høy sannsynlighet for at kysttorskbestanden er svak. Vi vet at det er statistisk usikkerhet i estimatene, men selv med denne usikkerheten vurderes det å være høy sannsynlighet for at bestanden er i dårlig forfatning. Vi vurderer derfor kunnskapsstyrken som moderat.
«Genetisk påvirkning på kysttorskbestander». Med antatt moderat sannsynlighet for et høyt antall rømt oppdrettstorsk, moderat sannsynlighet for betydelig gyting i merd med usikker overlevelse og svak tilstand på kysttorskbestandene i området, vurderes det totalt sett å være moderat sannsynlighet for «Genetisk påvirkning på kysttorskbestandene».
Det er manglende kunnskap rundt antallet oppdrettsfisk som rømmer og mengden egg som tilføres miljøet fra oppdrettstorsk gjennom gyting i merd. Det finnes ingen kunnskap om hvor stor andel av disse individene som vil vokse opp og bli gytende individer. Det er vist at oppdrettstorsk kan gyte med vill torsk, men vi har lite kunnskap om i hvilken grad gener med bakgrunn fra oppdrett vil etableres i kysttorskbestandene. Kunnskapsstyrken vurderes samlet sett som moderat.
«Negative effekter på kysttorskbestander mellom 62°N og 67°N som følge av torskeoppdrett i åpne merder i sjø ved en produksjonsintensitet på 49 000 tonn». Det vurderes å være lav sannsynlighet for negative endringer i fysiologi og adferd, samt økt sykdomsforekomst. Det vurderes derimot at det er moderat sannsynlighet for genetisk påvirkning på kysttorskbestander som følge av torskeoppdrett nord for 62°N og sør for 67°N. Kysttorskbestanden vurderes å være svak og dermed lite robust til å motstå en eventuell genetisk innkrysning fra oppdrettstorsk. På lengre sikt er det derfor en bekymring for at innkrysning med oppdrettstorsk medfører en generell reduksjon i genetisk variasjon og genetisk ensretting mellom kysttorskbestandene som kan gi negative konsekvenser for bestandenes evne til å tilpasse seg endrede miljøbetingelser. Vurderingen av «Genetisk påvirkning på kysttorskbestandene» vektlegges mer enn de tre andre hendelsene og sannsynligheten for negative effekter på kysttorskbestandene mellom 62°N og til 67°N vurderes totalt sett som moderat.
Det mangler kunnskap om flere av de underliggende risikokildene og hendelsene og kunnskapsstyrken vurderes derfor totalt sett som moderat.
Moderat sannsynlighet kombinert med moderat kunnskapsstyrke gir noe usikkerhet grunnet manglende kunnskap. Forventet økning av torskeproduksjonen de neste fem årene vurderes som moderat og risikoen for ««Negative effekter på kysttorskbestander mellom 62°N og 67°N som følge av torskeoppdrett i åpne merder i sjø ved en produksjonsintensitet på 49 000 tonn» vurderes derfor som moderat.
1.4.2 - Negative effekter på kysttorskbestander nord for 67°N som følge av torskeoppdrett i åpne merder i sjø ved en produksjon på 13 000 tonn

«Bruk av kunstig belysning for å utsette kjønnsmodning». Lysstyring brukes i torskeoppdrett for å forsinke og redusere graden av kjønnsmodning. Det finnes for lite kunnskap til at vi i denne analysen gjør en vurdering av denne risikokilden. Vi velger likevel å ha med noden da vi antar at den kan ha en påvirkning på kysttorskens adferd. Manglende datagrunnlag og dokumentasjon visualiseres med en hvit node med en rød ring.
«Tiltrekning for beiting på spillfôr, bunndyrsamfunn og andre organismer». I områder med mange oppdrettsanlegg er det observert at rundt 20 % av kysttorsken spesialiserer seg på beiting under anleggene. Det vurderes derfor at det er høy sannsynlighet for at kysttorsk vil tiltrekkes til oppdrettsanlegg og beite på spillfôr og byttedyr. Det er lite eller ingen dokumentasjon på tiltrekning av kysttorsk til torskeoppdrett, men basert på en rekke studier på tiltrekning til lakseoppdrettsanlegg som viser dette, vurderes kunnskapsstyrken som sterk.
«Negative endringer i fysiologi hos kysttorsk». Selv om det kan påvises endringer i fettsyresammensetningen vektlegges det at torskefôr er utviklet for å tilfredsstille ernæringsbehovet til torsk. I tillegg må det tas i betraktning at produksjonen av torsk er lav og at dermed også den totale mengden spillfôr vil være lav. Det vurderes derfor at det er lav sannsynlighet for at villfisk som beiter på spillfôr og andre organismer rundt torskeoppdrettsanlegg vil føre til en «Negativ endringer i fysiologi hos kysttorsk».
Foreløpige resultater viser at oppdrettstorsk som spiser marine pellets er i stand til å gyte levedyktige egg, selv om befruktningsratene og utviklingssuksessen til embryoene var lavere enn hos referansegruppen av villtorsk. Der er derimot lite kunnskap om konsekvensene av beiting på torskefôr og om dette kan skade reproduksjonssuksessen hos disse fiskene. Kunnskapsstyrken vurderes derfor totalt sett som moderat.
«Negativ endring i adferd hos kysttorsk». Selv om det antas at sannsynligheten for tiltrekning av kysttorsk til torskeoppdrettsanlegg i forbindelse med beiteaktivitet er høy, er tetthet av torskeoppdrettsanlegg nord for 67° N lav, med to anlegg per 2025 og en antatt økning til seks anlegg i løpet av de neste fem årene. Hvordan bruk av kunstig belysning påvirker adferden til kysttorsk kan ikke vurderes ut fra nåværende kunnskap. Basert på at det er få torskeanlegg, vurderes det å være lav sannsynlighet for «Negativ endring i adferd hos kysttorsk».
Det er god kunnskap om at kysttorsk tiltrekkes oppdrettsanlegg, selv om det er lite kunnskap om tiltrekning spesifikt til torskeanlegg. Det er svak kunnskap om bruk av kunstig belysning og hvordan dette påvirker villtorskens adferd. Det mangler også studier på hvorvidt kysttorsk som beiter ved torskeoppdrettsanlegg velger bort viktige habitater knyttet til gyte- eller oppvekstområder. Kunnskapsstyrken vurderes derfor totalt sett som moderat.
«Høyt antall sykdomsutbrudd og smittespredning i merd». Antallet sykdomsutbrudd og mengden smittespredning fra oppdrettsfisk vil avhenge av antallet oppdrettsanlegg og mengden fisk i sjø. Med en estimert produksjon på 13 000 tonn oppdrettstorsk, vil antallet utbrudd være fordelt på relativt få anlegg og lokaliteter (6). Det vurderes derfor at det vil være lav sannsynlighet for et «Høyt antall sykdomsutbrudd og smittespredning i merd» i området nord for 67°N.
Selv om det ikke er fullstendig oversikt over annet enn de meldepliktige sykdommene (sykdommer på liste F), gir fiskehelserapporten en god oversikt over sykdomssituasjonen i norsk fiskeoppdrett. Siden vi ikke kan si med sikkerhet hvilke(n) sykdom(mer) som vil gjøre seg gjeldene, hvor utbruddene vil forekomme eller omfanget av disse, vurderes kunnskapsstyrken som moderat.
«Høyt antall oppdrettstorsk rømmer». Datagrunnlaget for å vurdere sannsynligheten for store rømminger av oppdrettstorsk i et gitt geografisk område er begrenset. Dersom vi betrakter rapporterte rømminger over de siste 5 årene fra hele landet ser vi at størrelsen på rømminger varierer voldsomt, med flere små hendelser og tre store rømminger. Totalt antall rømte oppdrettstorsk de siste årene er dermed dominert av disse få store rømmingene. Det er i 2025 kun to anlegg i drift og fire søknader til behandling i dette forvaltningsområdet. Det vurderes derfor at det er lav sannsynlighet for at et høyt antall oppdrettstorsk rømmer nord for 67°N.
Store rømningshendelser skjer sporadisk og er ikke jevnt fordelt i tid og rom. De forventede rømningstallene for fremtiden anses som usikre. Basert på erfaringer med rømmingstall fra lakseoppdrett, er det også grunn til å tro at antall rømte torsk er noe høyere enn det som er innrapportert. I tillegg er det usikkerhet rundt hvordan rømningstiltakene næringen jobber med vil resultere i mindre rømming. Kunnskapsstyrken vurderes derfor totalt sett som moderat.
«Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettstorsk». Basert på en antagelse om at tiltrekning av villfisk til oppdrettsanleggene er høy, og at antallet rømte oppdrettstorsk vil være moderat, vektlegges det at det er svært få lokaliteter spredt over et stort geografisk område noe som gjør at det totale smittepresset innenfor forvaltningsområdet vil være lavt. Sannsynligheten vurderes derfor som lav for «Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettstorsk».
Siden det er usikkerhet knyttet til antallet sykdomsutbrudd og hvilke sykdommer som vil gjøre seg gjeldene, samt hvor mye oppdrettstorsk som vil rømmer vurderes kunnskapsstyrken som moderat.
«Betydelig gyting i merd». Det er vist store variasjoner i andelen oppdrettstorsk som blir kjønnsmodne og gyter i oppdrettsmerdene (målt til 1-35 % i merd, 2-40 % på slakteri). Av disse er andelen mellom hanner og hunner ikke jevnt fordelt. Hunner modner seinere en hannfisk og utgjorde ca. 21 % (merd) 34% (slakteri). Siden det er hunner som produserer egg er det antall gytende og eventuelt utgytte hunner som er relevant for denne vurderingen. Det vektlegges at forvaltningsområdet for 67°N har få anlegg og lav torskeproduksjon. Vi vurderer derfor sannsynligheten for betydelig gyting i merd som lav. Det er stor variasjon i modningsgrad gjennom året og mellom fiskegrupper. Det er også usikkert om hvordan utslakting av fisk som er i modning vil påvirke andelen som faktisk gyter i merd. Kunnskapsstyrken vurderes som moderat.
«Betydelig overlevelse av avkom gytt i merd til gytende individer». Selv om det finnes noe kunnskap om befruktningsgrad og overlevelse i larvefasen fra labforsøk, finnes det ingen undersøkelser om hvorvidt disse larvene fra oppdrettstorsk overlever i naturen fram til de selv blir til gytende individer. Det vurderes derfor at det ikke finnes nok kunnskap til å gjøre en sannsynlighetsvurdering av overlevelsen av avkom gytt i merd til gytende individer. Vi velger likevel å ha med denne risikokilden da den teoretisk sett kan føre til genetisk påvirkning. Manglende datagrunnlag og dokumentasjon visualiseres med en hvit node med en rød ring.
«Svak tilstand hos kysttorskbestandene». Tatt i betraktning at fisketrykket er på vei ned og at kysttorskbestanden er vurdert som bra nord for 67°N, vurderes det at det er lav sannsynlighet for at kysttorskbestanden er svak i dette forvaltningsområdet. Kunnskapsstyrken vurderes som god.
«Genetisk påvirkning på kysttorskbestandene». Med antatt lav sannsynlighet for et høyt antall rømt oppdrettstorsk, moderat sannsynlighet for betydelig gyting i merd og god tilstand på kysttorskbestandene i området, vurderes det å være lav sannsynlighet for «Genetisk påvirkning på kysttorskbestandene».
Det er manglende kunnskap rundt antallet oppdrettsfisk som rømmer og mengden egg som tilføres miljøet fra oppdrettstorsk gjennom gyting i merd. Det finnes ingen kunnskap om hvor stor andel av disse individene som vil vokse opp og bli gytende individer. Det er vist at oppdrettstorsk kan gyte med vill torsk, men vi har lite kunnskap om i hvilken grad gener med bakgrunn fra oppdrett etableres i kysttorskbestandene. Kunnskapsstyrken vurderes samlet sett som moderat.
«Negative effekter på kysttorskbestander nord for 67°N som følge av torskeoppdrett i åpne merder i sjø ved en produksjonsintensitet på 13 000 tonn». Det vurderes å være lav sannsynlighet for negative endringer i fysiologi og adferd, økt sykdomsforekomst og genetisk påvirkning på kysttorskbestandene som følge av torskeoppdrett nord for 67°N. Det vektlegges at de lokale kysttorskbestandene vurderes å være i god forfatning og dermed robuste for å motstå en negativ påvirkning og dermed også en eventuell genetisk innkrysning fra oppdrettstorsk. I tillegg er det lav produksjon av oppdrettstorsk og lokalitetene er spredt innenfor forvaltningsområdet. Totalt sett vurderes derfor sannsynligheten for «negative effekter på kysttorskbestander nord for 67°N» som lav.
Det mangler kunnskap om de fleste av de underliggende risikokildene og hendelsene og kunnskapsstyrken vurderes derfor totalt sett som moderat.
Lav sannsynlighet kombinert med moderat kunnskapsstyrke gir noe usikkerhet grunnet manglende kunnskap. På tross av begrenset kunnskap om noen av risikokildene er det forventet svært lav produksjon av oppdrettstorsk i området de neste fem årene, og kysttorskbestandene er i god forfatning. Risikoen for ««Negative effekter på kysttorskbestander nord for 67°N som følge av torskeoppdrett i åpne merder i sjø ved en produksjonsintensitet på 13 000» tonn vurderes derfor som lav.
1.5 - Konklusjon
Risikoen for «Negative effekter på kysttorskbestander som følge av påvirkning fra torskeoppdrett i åpne merder i sjø» vurderes som moderat mellom 62°N til 67°N og lav nord for 67°N. I begge områdene er risikoen justert ned sammenlignet med vurderingen som ble gjort i 2022. Grunnet veldig lav produksjon og få lokaliteter sør for 62°N, er det ikke gjort en egen vurdering for dette forvaltningsområdet. En risikovurdering av så store geografiske områder som brukt her, gir ikke et dekkende bilde av risikoen for kysttorskebestandene på lokal skala. For eksempel, nord for 67 °N vil det være store områder uten torsk i oppdrett, men vi kan likevel ikke utelukke lokal påvirkning i umiddelbar nærhet av de få anleggene som er i drift. Også mellom 62°N og 67°N vil det være områder uten torskeoppdrett, men også områder hvor flere anlegg ligger nær hverandre og påvirkningen fra oppdrett vil være større.
En av utfordringene i beregningen av produksjonsvolum frem i tid er gitte tillatelser som ikke er i bruk. Disse utgjør en stor andel av den mulige totale produksjonen. Om, og eventuelt når, disse vil tas i bruk er uvisst. Det er likevel ikke mulig å utelate disse fra en risikovurdering, da vi anser at alle tillatelser teoretisk sett kan realiseres innen kort tid hvis forholdene ligger til rette for det. I motsetning til vurderingen som ble gjort i 2022, der vi tok høyde for 100% kapasitetsutnyttelse, har vi i denne vurderingen tatt utgangpunkt i 40 % kapasitetsutnyttelse.
Som ved forrige risikovurdering vurderes tilstanden til kysttorskbestanden i forvaltningsområde mellom 62°N og 67°N som svak. For området nord for 67°N, der kysttorskbestanden i 2022 ble vurdert som moderat, vurderes bestanden nå som sterk. Da produksjonsvolum, antall lokaliteter og tilstanden til kysttorskbestandene er styrende for effekten av risikokildene, hendelsene og konsekvensene i risikovurderingen, reduseres risikoen for «Negative effekter på kysttorskbestander som følge av påvirkning fra torskeoppdrett i åpne merder i sjø» i begge forvaltningsområdene vi har inkludert.
Vi anser fortsatt gyting i merd og rømming som de største utfordringene i torskeoppdrett. Dette er problemstillinger næringen jobber med gjennom optimalisering av lysstyring og forbedring av oppdrettsnøter og oppdrettsteknologi, inkludert nedsenkbar merdteknologi og lukkede anlegg. Status på kjønnsmodning viser at det er en betydelig andel som når stadiet for gyting i merd. Overlevelse fra egg gytt i merd til kjønnsmoden torsk er påvist, men kunnskapen om dette er svak og fragmentert. Hvis slik overlevelse er av en viss størrelsesorden kan gyting i merd være betydelig mer alvorlig enn rømming i et regionalt perspektiv.
Det er nå 6.-9. generasjons oppdrettstorsk som benyttes, og det er påvist en reduksjon i genetisk variasjon på nær 50 % i en av oppdrettslinjene. Genetiske interaksjoner mellom oppdrettsfisk og villfisk er godt dokumentert for andre arter, og vil kunne føre til redusert genetisk mangfold og tilpasningsevne (f.eks. med hensyn til klima- og miljøendringer). Det mangler kunnskap om i hvilken grad dette gjelder også for torsk. Erfaringer fra lakseoppdrett og interaksjoner med villaks, overført til torsk, antyder at sterke bestander av villtorsk vil være mer motstandsdyktige mot påvirkning fra torskeoppdrett på grunn av uttynningseffekten. Men små bestander med lokale gytefelt og oppvekstområder i indre fjordområder med høy grad av retensjon og lokal bunnslåing av larver, vil være ekstra utsatt. Hva konsekvensene av innkrysning av oppdrettstorsk vil være for kysttorskbestandene vet vi ikke, men det er et uttrykt faglig og politisk mål å stoppe tap av biologisk mangfold, inkludert genetisk variasjon innen arten.
Utvikling av gode metoder for å overvåke genetisk påvirkning, gjennom for eksempel genetiske markører for oppdrettstorsk, vil derfor være viktig verktøy for forvaltningen. Siden det er utfordrende å overvåke større geografiske områder vil trolig genetiske metoder være mest aktuelle å ta i bruk nær lokaliteter med observert rømming og/eller gyting i merd.
Med en kapasitetsutnyttelse på 40% av total kapasitet (total produksjon på 67 000 tonn) vurderer vi at det er lav sannsynlighet for at torskeoppdrett vil føre til negative endringer i adferd og fysiologi hos kysttorsken og at det også vil være lav sannsynlighet for økt sykdomsforekomst i villfisken. Vi vil likevel påpeke at forebyggende tiltak for å begrense sykdom og smittespredning, inkludert tilgang på gode og beskyttende vaksiner, vil være essensielt i den videre utviklingen av næringen. Det finnes i dag liten tilgang på kommersielt godkjente vaksiner, og oppdrettstorsk vaksineres i stor grad med autogene vaksiner. Selv om det per i dag er få sykdomsproblemer relatert til dødelighet hos oppdrettstorsk, tilsier all erfaring fra andre arter at med en oppskalering vil sykdomssituasjonen endre seg. Man må derfor anta at også oppdrettstorsken vil møte på utfordringer med sykdom i takt med at produksjonen øker. Det er heller ikke gitt at det vil være de samme kjente sykdommene som vil gjøre seg gjeldene i framtida. Man må være forberedt på at både ukjente og nye varianter av kjente patogener (skadelige virus, bakterier og parasitter) vil skape utfordringer. Nye patogener og sykdommer blir derfor i vår vurdering sett på som potensielle overraskelser, symbolisert med svarte svaner i risikokartene. Noen patogener er lite artsspesifikke og forårsaker sykdommer hos mange ulike arter. Et eksempel er bakterier som forårsaker atypisk furunkulose som er kjent fra både rensefisk og torsk. Det er derfor viktig å tenke på biosikkerhet og potensiell smittespredning, både mellom arter i oppdrett og til eller fra villfisk.
Fôrspill fra torskeoppdrett forventes å tiltrekke seg villfisk inkludert torsk. Selv om oppdrettstorsk fôres med pellets som er bedre tilpasset marin fisk enn laksefisk, vil torsk som beiter på spillfôr forventes å få endringer i fysiologi på grunn av innblandet plantemateriale, og særlig planteoljer. Det er derimot usikkert i hvilken grad dette vil påvirke overlevelse, vekst og rekruttering. Stasjonær fjordtorsk ser ut til å ha et relativt begrenset hjemmeområde og vil i perioder sannsynligvis beite rundt torskeoppdrettsanlegg og vandre mellom disse og nærliggende gytefelt. Det er foreløpig begrenset kunnskap om dette vil kunne påvirke de ville torskebestandene.
Som ved forrige risikovurdering mangler det kunnskap for en rekke av risikokildene og hendelsene som er identifisert i denne risikovurderingen. Mangel på kunnskap skaper usikkerhet og resultater som bygger på svak kunnskap, kan skjule deler av risikobildet gjennom eksempelvis feilaktige antakelser. Vi har fått mer kunnskap om modning og gyting i merd gjennom målrettet forskning, men fortsatt er kunnskapen svak i forhold til hvorvidt egg fra gyting i merd vil ende opp i overlevelse frem til gytende individer som igjen vil kunne krysse seg inn i de lokale kysttorskbestandene. «Bruk av kunstig belysning for å utsette kjønnsmodning» er en ny risikokilde vi har valgt å ta med i årets vurdering. Vi antar at kunstig belysning kan bidra til at kysttorsk tiltrekkes oppdrettsanlegg og endrer adferd, men det er foreløpig lite kunnskap om dette. Å bygge opp denne type kunnskap tar tid og det vil være nødvendig å identifisere og prioritere de viktigste kunnskapshullene for en videre utvikling av næringen.
Begrenset tilgang til areal og nye lokaliteter, i hovedsak på grunn av usikkerhet knyttet til rømming, gyting i merd og påvirkning på gytefelt, ser derfor på kort og mellomlang sikt ut til å være en flaskehals for ytterligere vekst av torskeoppdrett i åpne merder.
Risikovurderingen konkluderer med at økt kontroll på gyting i merd og rømming er de viktigste tiltakene for å redusere risiko for «Negative effekter på kysttorskbestander som følge av påvirkning fra torskeoppdrett i åpne merder i sjø».
2 - Kunnskapsstatus
Denne kunnskapsstatusen er en oppdatert versjon av den som ble utgitt i 2022. Den presenterer en utvidet gjennomgang av faktorene knyttet til selve risikovurderingen; genetisk påvirkning på kysttorskbestander (tilstanden på kysttorskbestandene, rømming av oppdrettstorsk og gyting i merd med overlevelse av avkom til gytende individer), økt sykdomsforekomst hos kysttorskbestander (sykdomsutbrudd og smittespredning i merd), endring i adferd og i fysiologien hos kysttorsk (bruk av kunstig belysning for å utsette kjønnsmodning, spillfôr).
2.1 - Torskebestander og torskeoppdrett i Norge
Torsk ( Gadus morhua ) er en torskefisk i familien Gadidae som er utbredt langs hele norskekysten og i havområdene rundt. Torsken har vært, og er, en av de viktigste kommersielle artene i Norge. I norske farvann finnes det ulike bestander av torsk; nordøst arktisk torsk, nordsjøtorsk og kysttorsk, sistnevnte kan deles inn i mange underbestander. Skrei brukes om gytemodne individer av den vandrende nordøstarktiske torsken, som for det meste holder til i Barentshavet og vandrer inn mot kysten om våren (i mars /april) for å gyte.
2.1.1 - Generelt om kysttorskbestander – tilstand og utbredelse
Kysttorsk er utbredt langs hele norskekysten fra lukkede fjordområder til mer åpne bankområder. På disse bankområdene grenser kysttorsk mot, og delvis blander seg med, de større oseaniske bestandene i Barentshavet i nord og Nordsjøen i sør. Genetiske undersøkelser har vist at kysttorsk ofte skiller seg fra de oseaniske bestandene, og at kysttorsk ikke er en ensartet bestand, men viser økende grad av forskjell med geografisk avstand (Dahle mfl. 2018; Johansen mfl. 2020; Jorde mfl. 2021). Men det er også kysttorskkomponenter langs ytre skjærgård i sør som genetisk er lik, og kanskje er, Nordsjøtorsk (Sodeland mfl. 2016; Knutsen mfl. 2018). Det finnes sannsynligvis en rekke torskebestander langs kysten, men det er vanskelig å avgrense utbredelsen til hver enkelt av dem. Selv om enkelte bestander ser ut til være tilknyttet begrensede arealer, er det i større eller mindre grad egnede torskehabitater langs hele kysten.
Modellberegninger av pelagisk egg- og larvedrift indikerer at fjord- og kystbestander har spesifikke gytefelt (Espeland mfl. 2015, Myksvoll mfl. 2011; 2014). Avkommets spredning viser lokal retensjon (tilbakeholdelse), og kysttorsken har gjerne tidlig bunnslåing i kontrast til skrei der avkommet hovedsakelig transporteres fra ytre kystområder til oppvekstområder i Barentshavet, med bunnslåing etter ca. 5 måneders drift i de frie vannmassene. Det finnes imidlertid også skrei som bunnslår hovedsakelig på dypt vann i nordnorske fjorder (Fevolden mfl. 2012). Nyere forskning har imidlertid vist at skrei kan oppholde seg året rundt i fjorder i Vest-Finnmark (Strøm mfl. 2023). Noe kysttorsk finnes innblandet på de samme gytefelt som benyttes av skreien ved Lofoten og tilsvarende finnes noe skrei innblandet i gytefelt som hovedsakelig brukes av kysttorsk, for eksempel i Borgundfjorden (Johansen mfl. 2018). Det er ukjent i hvilken grad torsk fra de to hovedgruppene gyter sammen og krysser seg med hverandre (hybridiserer) (Nordeide & Båmstedt 1998; Nordeide & Folstad 2000), men genetiske studier viser økende grad av skreigener i kysttorskbestandene nordover fra Stad (Dahle mfl. 2018; Johansen mfl. 2020; Jorde mfl. 2021; Breistein mfl. 2022; Helmerson mfl. 2025). Tidligere var man av den oppfatning at slik hybridisering forekommer relativt sjeldent, da omfattende hybridisering vil bryte ned de genetiske forskjellene mellom økotypene. Nyere studier viser dog at mengden hybridisering er betydelig nok til å påvirke den genetiske strukturen hos kysttorsk, primært nord for 67°N, og ytterst ute i fjordene (Jorde mfl. 2021; Breistein mfl. 2022), og at hyppigheten på hybridiseringen har vært økende de siste tiårene (Helmerson mfl. 2025). Vi kjenner ikke til hvilke egenskaper kysttorsk-skrei hybrider vil ha: om de vil være stasjonære og vil blande seg med lokal kysttorsk, eller om de vil vandre ut fra kysten. Det er fremdeles uklart hvordan og hvorvidt pelagisk avkom fra de to komponentene, når de forekommer på samme gyteplass, har ulike sprednings- og bunnslåingsmønstre. I Barentshavet bunnslår torskeyngel på bunn med dyp fra 150-300 m, mens i kyst- fjordhabitater bunnslår torskeyngel betraktelig grunnere og delvis i littoralsonen (Godø & Sunnanå 1984; Jørstad mfl. 2014).
Genetiske forskjeller mellom skrei og kysttorsk ligger hovedsakelig i de delene av kromosomene som er karakterisert ved store såkalte «inversjoner». Inversjoner er intakte genetiske «pakker» som nedarves nesten uten å rekombineres, og dette antas å være en viktig mekanisme for å opprettholde en separasjon av de to sameksisterende økotypene (Berg mfl. 2016; Kirubakaran mfl. 2016; Matschiner mfl. 2022; Sodeland mfl. 2022). Slike inversjoner ser også ut til å være viktige for å skille vandrende og stasjonære økotyper av torsk på et mer generelt grunnlag (Johansen mfl. 2020).
Torsk som fanges i fiskeriene, inneholder ofte fangster av blandede bestander, og dette er en generell utfordring ved fiskeriene som har vist seg vanskelig å eliminere. I nord gjelder dette spesielt bifangst av kysttorsk under skreifisket, og Havforskningsinstituttet klassifiserer rutinemessig fangster fra fiskeriene og forskningstokt til skrei og kysttorsk, basert på øresteiner og genetiske metoder. Fenomenet med sameksisterende bestander eller økotyper av torsk i fjordene og langs kysten har vi også i øvrige deler av landet, men datagrunnlaget er best i nord (kysttorsk og skrei) og helt i sør (Nordsjø- og fjordtorsk på Skagerrakkysten). Tidligere merkeforsøk har vist at f.eks. Balsfjordtorsk og Porsangertorsk har en lokal utbredelse i større grad enn andre fjordbestander i regionen nord for 67°N (Jakobsen 1987).
2.1.2 - Status kysttorsk sør for 62°N
Det finnes ikke en formell bestandsvurdering for kysttorsk sør for 62°N, men Havforskningsinstituttet har nylig publisert en rapport med oppdatert status for torsk i dette forvaltningsområdet (Stock mfl. 2025). Alle tilgjengelige data viser at bestanden er i svært dårlig forfatning. For eksempel viser fangst-per-enhet-innsats data fra referanseflaten en nedgang på 75 – 80 % i Skagerrak og på 50 % på Vestlandet siden 2006 (figur 2.1). Den negative utviklingen i Skagerrak tydeliggjøres ytterligere med data fra Havforskningsinstituttet garntokt, hvor det i Øst-Skagerrak ble observert en markant nedgang på 1990-tallet. Til sammenligning var torskebiomassen i Vest-Skagerrak relativ stabil før den gikk kraftig ned fra 2017 (figur 2.1). I dag er derfor biomassen av torsk antatt å være på historisk lave nivåer i begge områdene.
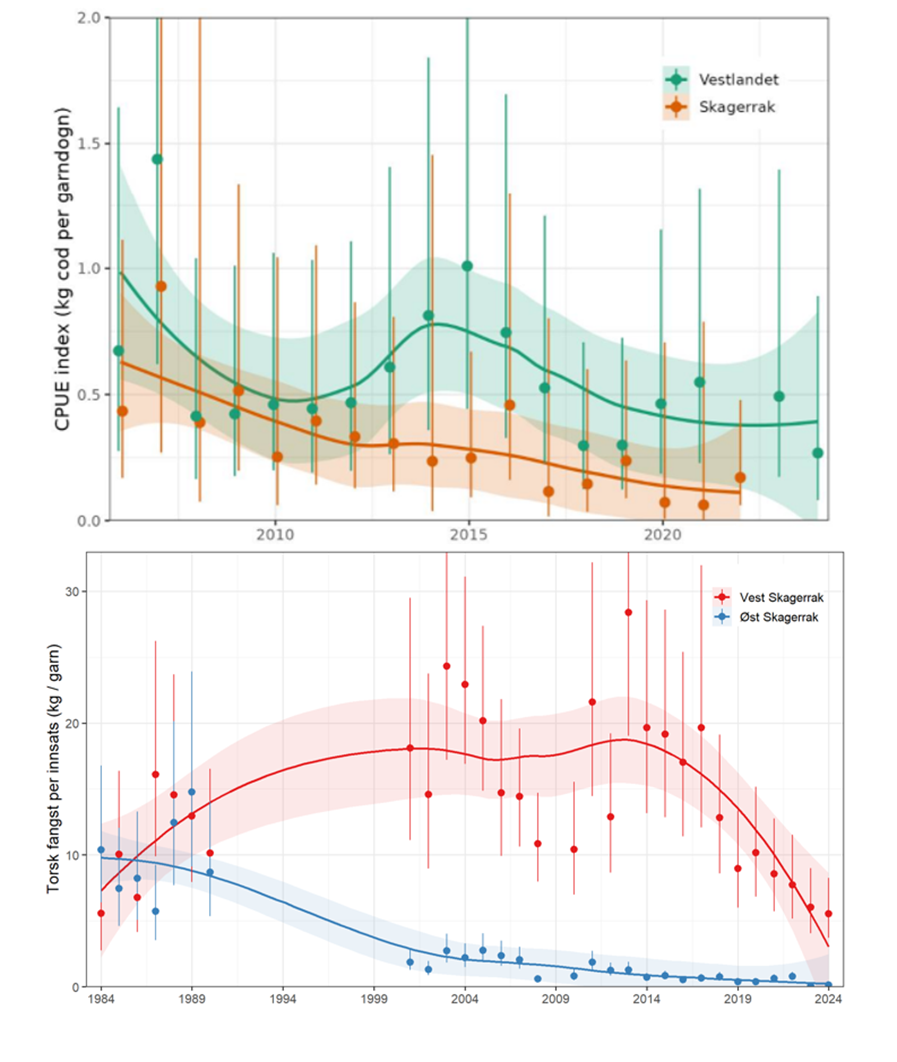
Videre viser de kommersielle fangstene av torsk sør for 62°N en nedgang på ca. 90 % sammenlignet med historiske fangster på 2000 – 4000 tonn fra 1920- tallet til rundt år 2000. Det er lite som tyder på at vekstforholdene (individvekst) begrenser torskebestandene sør for 62°N, ettersom vekt ved alder har økt i alle regioner for de fleste aldre. Merke-gjenfangst analyser har vist at fiskedødelighet (F) overgår naturlig dødelighet både i Skagerrak og på Vestlandet. Av mulige påvirkningsfaktorer i tillegg til klimaendringer (i.e., vekst, rekruttering, naturlig dødelighet og fiskedødelighet), ansees fiskedødelighet en viktig årsak til den observerte nedgangen i torskebestanden sør for 62°N, og da spesielt dødelighet fra fritidsfiske ettersom denne antas å være høyere enn fra kommersielt fiskeri (Stock mfl. 2025).
2.1.3 - Status kysttorsk 62°N – 67°N
Kysttorsk 62-67°N er definert som kategori 2-bestand av ICES. Det betyr at bestandsvurderingen er godkjent med hensyn til trender, men ikke absolutte estimater av bestandens biomasse (B). Bestandsvurderingen gjøres ved en produksjonsmodell (Pedersen & Berg 2017). I en slik modell er relativ fiskedødelighet og den relative biomassen to sentrale elementer. Relativ fiskedødelighet gis ved å dele fiskedødelighet (F) på den fiskedødeligheten som gir det maksimale langtidsutbyttet (Fmsy), det vil si det fisket som er bærekraftig over tid. Ideelt sett bør dette forholdet (F/Fmsy) være mindre eller lik 1 og det anses som kritisk hvis den relative fiskedødeligheten overskrider 1,7. Tilsvarende er den relative biomassen av en bestand den faktiske biomassen (B) delt på biomassen som resulterer fra fiske ved Fmsy (Bmsy). Dette forholdet (B/Bmsy) bør være større eller lik 0,5 og 0,3 anses som kritisk nedre grense. Når bestanden når kritisk grense betyr det at gytebiomassen er så lav at bestandens rekruttering påvirkes negativt. Det er viktig å presisere at i denne type modeller refererer både relativ fiskedødelighet og biomasse til den fiskbare delen av bestanden.
Bestandsmodellen inkluderer fangstdata fra kommersielt fiske, turist/fritidsfiske, samt biomasseindekser fra Havforskningsinstituttets kysttokt (akustikkdata og trålfangster) og garn-ruse tokt. Både kommersielle fangster og tetthetsindikatorer fra Havforskningsinstituttets overvåkningstokt har vist en tydelig nedgang det siste tiåret. Den relative fiskedødeligheten (F/Fmsy) økte fra midten av 1990-tallet og frem til 2008 hvor den flatet noe ut. For 2024 er denne estimert til å være 1,80 med 95 % konfidensintervall på 1,21 – 2,67, det vil si over bærekraftig nivå (figur 2.2) (ICES 2025b). Dette gjenspeiler seg i den relative biomassen (B/Bmsy) som er mer enn halvert siden starten av 1990-tallet og for 2024 er estimert til 0,39 med 95% konfidensintervall på 0,25 – 0,66 (figur 2.2) (ICES 2025b). For 2025 forventes en total fangst på 6 834 tonn (ICES 2025c), noe som er over tre ganger høyere enn ICES sitt samlede råd for kommersielt-, turist- og fritidsfiske på 1 926 tonn for bærekraftig høsting for 2025 (ICES 2025b).
Det største problemet med denne bestandsvurderingen er den høye usikkerheten rundt fangstene i turist- og fritidsfiske, da dette fisket sannsynligvis utgjør mer enn halvparten (ca. 60 %) av de totale fangstene. Usikkerheten kan ikke kvantifiseres og er ikke inkludert i modellen. Fangstdata er kritisk for modellen da den i stor grad bestemmer nivået på bestanden, mens trendene påvirkes av toktindeks. Når i tillegg sensitivitetstester av bestandsmodellen viser at absolutte estimater av bestandsstørrelse og fiskeridødelighet (F) er mye mer følsomme enn når disse utrykkes relativt Bmsy/Fmsy, og disse referansepunktene oppdateres hver gang modellen kjøres, se også ICES 2025b), er det åpenbart lav presisjon i bestandsestimatet for dette området. For å kunne vurdere risiko fra torskeoppdrett for bestanden er del likevel behov for et anslag av biomassen. Fra modellen får vi et estimat på 30 000 tonn (95% konfidensintervall: 12 900 – 69 800) for 2024. Dette vil være et maksimalt estimat av gytebiomassen, da modellen definerer høstbar biomasse som fisk over lengde 50 cm, som omtrent motsvarer lengden der 50 % er kjønnsmodne. Historiske toktdata viser at omtrent 75 % av kysttorsken i de to bestandsområdene befinner seg nord for 67°N (Aglen mfl. 2020). I senere tid har bestandsutviklingen vært mer positiv for kysttorsken i dette området sammenlignet med området 62-67°N der trenden er synkende, og andelen kysttorsk nord for 67°N har derfor økt (ICES 2025c). Ser vi på de totale fangstene (kommersielle + rekreasjonsfangster) i de to forvaltningsområdene så ligger området 62-67°N på ca. 20 % av nord for 67°N. Gitt proporsjonalitet mellom bestandsstørrelse og fangster vil dette tilsi en gytebestandsstørrelse i dette forvaltningsområdet på i underkant av 20 000 tonn.
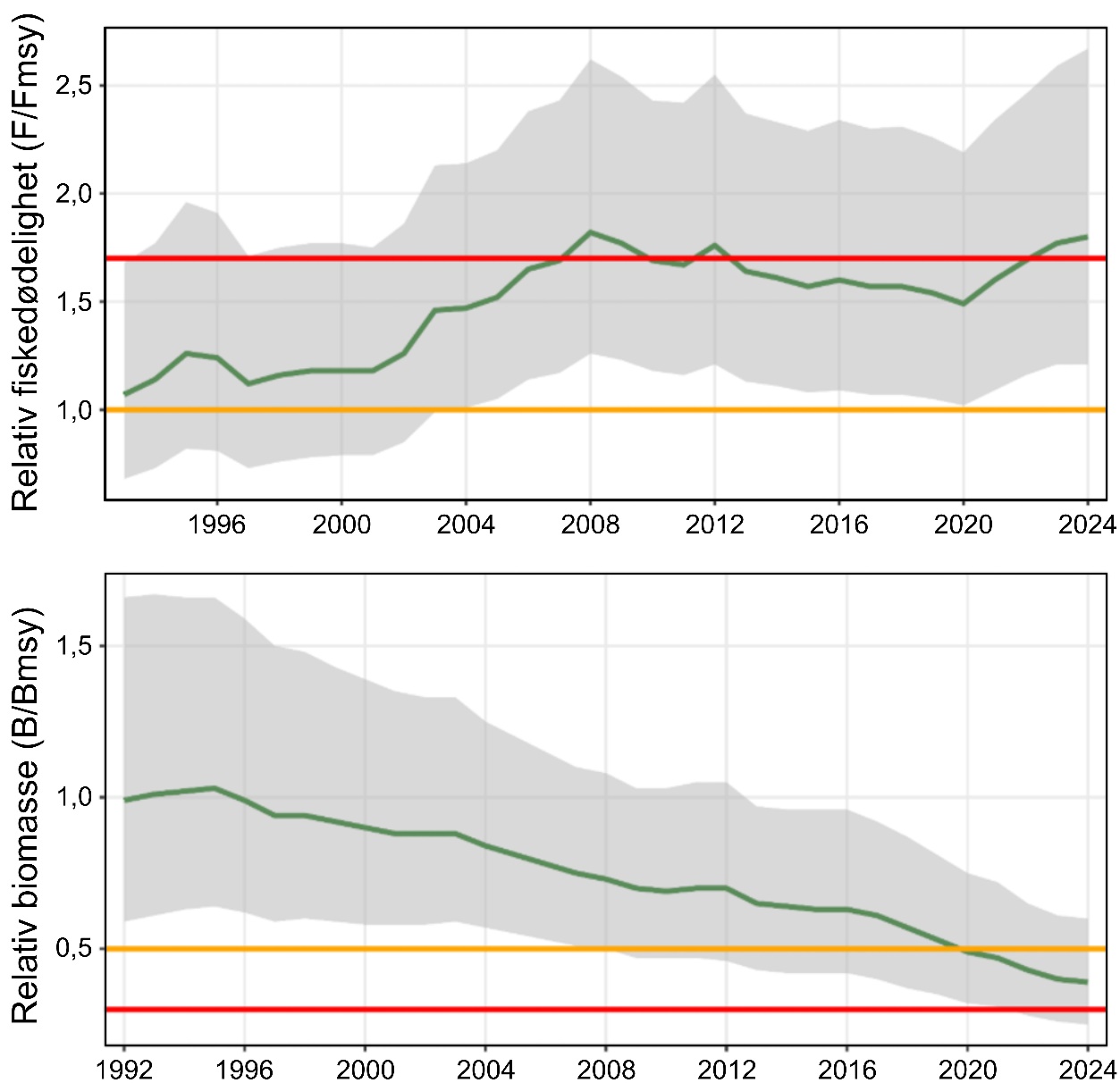
Videre har både kommersielle fangster og tetthetsindikatorer fra Havforskningsinstituttets overvåkningstokt vist en tydelig nedgang det siste tiåret. Det største problemet med den nåværende vurderingen av bestandsstatus for kysttorsk 62°N – 67°N er den høye usikkerheten rundt fangstene i turist- og fritidsfiske, da dette fisket sannsynligvis utgjør mer enn halvparten (ca. 60 %) av de totale fangstene. For 2025 estimeres den totale fangsten å bli 6 834 tonn (ICES 2025c), noe som er over tre ganger høyere enn ICES sitt samlede råd for kommersielt-, turist- og fritidsfiske på 1 926 tonn for bærekraftig høsting for 2025 (ICES 2025b).
2.1.4 - Status kysttorsk nord for 67°N
Datagrunnlaget for bestandsvurdering av kysttorsk nord for 67°N er bedre enn for kysttorsk i området 62°N – 67°N, og ICES gir sin bestandsvurdering og kvoteråd basert på en aldersstrukturert bestandsmodell (ICES 2021). Denne modellen har påvist en svak sammenheng mellom størrelse på gytebestanden og rekruttering, noe som tyder på varierende trender i rekruttering til lokale populasjoner innenfor bestandsområdet (ICES, 2022a). Som følge av dette er det ikke fastsatt en kritisk grense for biomasse i gytebestanden (Blim), et referansepunkt som normalt brukes som en del av høstingsregelen. En alternativ høstingsregel ble derfor foreslått og testet, der det ble beregnet en fiskedødelighet som var «føre-var» i hele spennet av observerte gytebestandsstørrelser og varierte lite mellom ulike antakelser om blant annet nivået på Blim (ICES, 2022b). Denne høstingsregelen, som ofte blir referert til som Den norske forvaltningsplanen, ble vedtatt av Norge ved Nærings- og fiskeridepartementet og brukt for første gang i kvoterådet for 2023.
Siden midten av 1990-tallet har fiskedødeligheten (4-8 åringer) for kysttorsk nord for 67°N for det meste vært langt over anbefalt nivå. Etter en periode med lavere fiskedødelighet i 2011-2015, som følge av lavere fangster og gode årsklasser på slutten av 90-tallet, med påfølgende vekst i bestanden, har fiskedødeligheten igjen økt det siste tiåret (figur 2.3). Fiskedødelighet for yngre årsklasser (dvs. 3 – 6 åringer) var spesielt høy i 2023 før den i 2024 returnerte til et nivå i bedre samsvar med de siste års gjennomsnitt (ICES 2025a). Denne reduksjonen i dødelighet for ungfisk sammenfalt med en økning i minstemålet fra 44 til 55 cm, som tredde i kraft 1. januar 2024. En sterk 2020-årsklasse kombinert med økt vekst og sannsynlighet for kjønnsmodning hos yngre fisk (3 – 6 åringer) bidro til den relativt markante økningen i gytebiomasse fra 2023 til 2024 (ICES 2025a). De kommersielle fangstene av eldre fisk (7+) var mye lavere i 2024, noe som sannsynligvis har sammenheng med særdeles lav biomasse av disse aldersklassene i bestanden. For 2024 var gytebiomassen estimert til 85 000 tonn, mens den totale biomassen var anslått å være 276 000 tonn (ICES 2025c). I 2024 ble totale fangsten estimert til 35 684 tonn (ICES 2025a). Dette tilsvarte en 32 % reduksjon sammenlignet med 2023, men var fortsatt en betydelig overskridelse (28 %) av den anbefalte kvoten for samme år. Det er uvisst hvordan den reduserte torskekvoten, som reflekterer nedgangen i skreibestanden, vil slå ut på kysttorskfangstene i tid og rom. For 2026 har ICES anbefalt at fangsten av kysttorsk nord for 67°N ikke bør overskride 27 887 tonn (ICES 2025a).
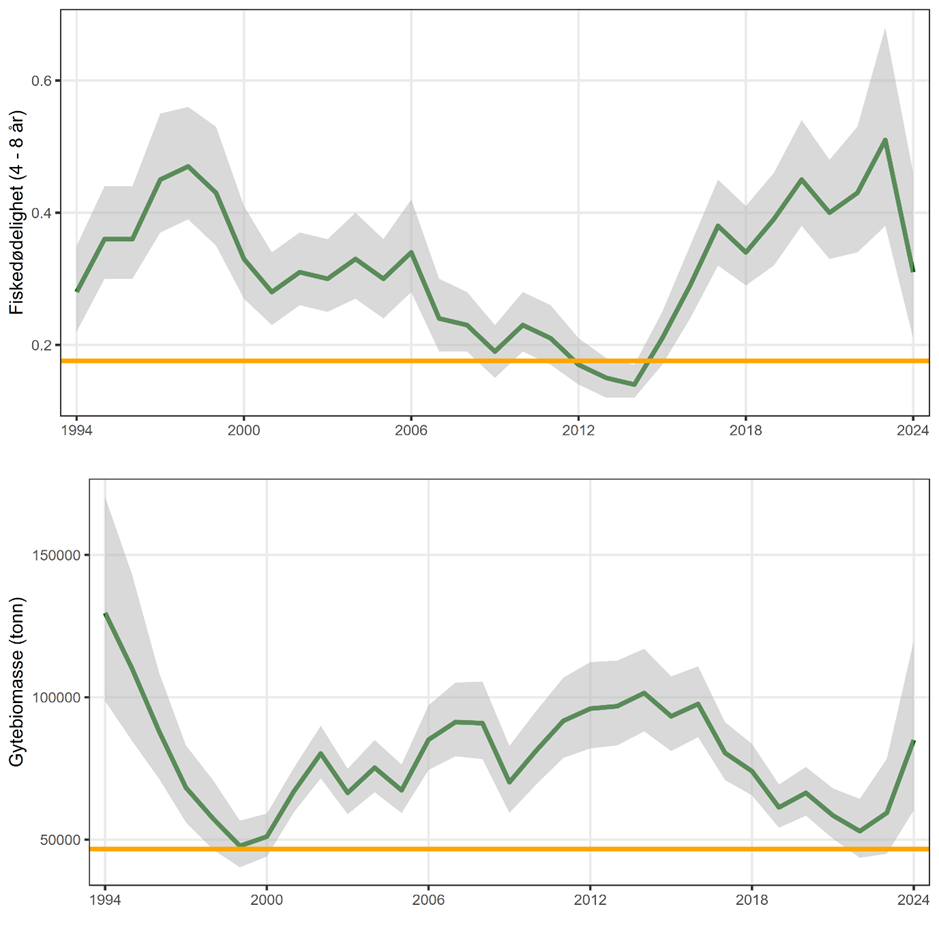
En oppsummering av status for kysttorskbestandene innen de tre forvaltningsområdene sees i figur 2.4.

2.1.5 - Torsk i oppdrett
2.1.5.1 - Bakgrunn og historikk
I Norge ble de første forsøkene med atlantisk torsk utført som gjenoppbyggingsprogrammer på slutten av 1880-tallet med store utsett av plommesekklarver (Rognerud 1887). Krav om bevis på at disse var levedyktige førte til startfôring i et sjøvannsbasseng ved den biologiske stasjonen i Flødevigen, der ca. 5000 torskeyngel ble drettet opp på naturlig dyreplankton. Først 100 år senere ble den samme metoden benyttet i Hyltropollen i Austevoll for å skaffe torskeyngel til akvakultur (Kvenseth & Øiestad 1984; Øiestad mfl. 1985). Dette førte til den første «torskeboomen» der yngel ble forsøkt produsert i poller eller poseanlegg knyttet til poller (van der Meeren & Naas 1997). Begrensning i yngeltilgang, optimalt vekstfôr, tidlig kjønnsmodning og endringer i markedssituasjonen førte imidlertid til at interessen for torsk i oppdrett falt sammen på begynnelsen av 90-tallet (Svåsand mfl. 2004; Engelsen mfl. 2004). Imidlertid beholdt Havforskningsinstituttet pollen «Parisvatnet» i Øygarden (Blom mfl. 1991) som forsynte noen få oppdrettere med yngel.
Inspirert av veksten for andre marine arter i akvakultur i Sør-Europa ble forsøk med intensiv produksjon av torskyngel basert på produksjon av levendefôr som hjuldyr (rotatorier) og saltkreps (Artemia sp.), utført i løpet av 1990-1993 av PB Ernæring ARC og Salar AS på Bessaker nord for Trondheim, sammen med flere norske forskningsinstitusjoner (Svåsand mfl. 2004). Til tross for et høyt antall yngel produsert i disse storskalaforsøkene, ble det observert høy dødelighet på grunn av kannibalisme forårsaket av suboptimale oppdretts- og fôrforhold, og forsøkene ble avsluttet. Torskeyngel drettet opp på naturlig plankton (hoppekreps) har vist både bedre vekst og lavere andel av deformiteter enn torsk startfôret på rotatorier og Artemia (Fjelldal mfl. 2009; Busch mfl. 2010; Karlsen mfl. 2015). Ernæring i tidlige livsstadier ser derfor ut til å være viktig for yngelkvaliteten. Videre utvikling av ernæringsmessig kvalitet gjennom anrikning av rotatorier og Artemia mot slutten av 90-tallet, sammen med oppløftende forsøk med intensiv startfôring av torsk i Canada og økende priser på grunn av nedgang i ville bestander, førte til fornyet interesse for torskeoppdrett ved årtusenskiftet både i Norge og i utlandet (Nardi mfl. 2021). Forsøk i Austevoll viste at yngelproduksjon av torsk kunne gjøres uavhengig av årstid ved hjelp av lysstyring (Norberg mfl. 2004; van der Meeren & Ivannikov 2006), og dette initierte den andre «torskeboomen» basert på intensiv startfôring og yngelproduksjon. Det ble etablert et Nasjonalt avlsprogram for torsk i Tromsø, og tillatelser for etablering av torskeklekkerier og matfiskanlegg økte raskt fra 94 i 2000 til 560 ved utgangen av 2004. Videreføring av tillatelsene var imidlertid avhengig av at det ble satt fisk i anleggene innen få år, og kapasiteten med hensyn til produksjon av yngel av god kvalitet klarte ikke å følge opp etterspørselen. Yngel med dårlig vekstevne og misdannelser i skjelettet ble satt ut i merder, og påfølgende rømminger resulterte i negative presseoppslag der betegnelsen «monstertorsk» ble brukt. Torsk ble observert i å aktivt bite hull i notveggene (Damsgård mfl. 2012), og rømt torsk ble funnet igjen på lokale gytefelt i betydelig avstand fra matfiskanlegg (Jørstad mfl. 2014). Dårlig vekst, rømminger og dårlig kvalitet reduserte de økonomiske marginene hos oppdretterne.
I motsetning til laksefisk er torsk en marin fisk som enkelt kan gyte i merdene (Jørstad mfl. 2008; van der Meeren mfl. 2012). Ettersom kjønnsmodning bare delvis kunne kontrolleres ved lysstyring i merder (Taranger mfl. 2006; Skulstad mfl. 2013) og torskeyngel ble flyttet langs hele norskekysten, ble dette betraktet som en mulig genetisk trussel mot lokale tilpasninger hos torskestammer som viser stor genetisk diversitet fra syd til nord (Johansen mfl. 2020). Kjønnsmodningen representerte i tillegg et tap i biomasse som oppdretterne ikke fikk dekning for. Kjønnsmodningen førte også til stor dødelighet i enkelte matfiskanlegg fordi torsken fikk problemer med å slippe eggene, såkalt «gytespreng» (Árnason & Björnson 2012).
Under den andre «torskeboomen» ble torskens genom kartlagt (Johansen mfl. 2009), og nye molekylære metoder for undersøkelser av gener og genaktivering dukket opp og har ekspandert raskt de siste 15 årene. Dette har ført til bedre verktøy for å forstå biokjemiske og fysiologiske prosesser hos torsken, og hvordan disse påvirkes av ernæring og miljø.
I 2004 brøt bakteriesykdommen, francisellose (Francisella noatunensis subsp. noatunensis) ut i norsk torskeoppdrett og i årene som fulgte forårsaket sykdommen store utfordringer med fiskevelferd og dødelighet. Francisellose er inkludert i den den nasjonale sykdomslisten og Mattilsynet krevde at all fisk ble slaktet dersom francisellose ble påvist i et matfiskanlegg. Francisellose, sammen med rømminger, yngelkvalitet, problemer knyttet til kjønnsmodning og lav vekst, ga den andre «torskeboomen» dårlige odds. Når dette i tillegg falt sammen med den økonomiske krisen i 2008 og rask fremvekst av en annen og billig og konkurrerende hvitfiskkart fra Vietnam, pangasius (Pangasianodon hypophthalmus), kollapset produksjonen av oppdrettstorsk i perioden fra 2011 til 2014. Det er likevel ikke mulig å peke på en enkelt faktor for sammenbruddet. Mange av yngelanleggene gikk over til oppdrett av rensefisk grunnet behovet for et alternativ til kjemisk lusebehandling da resistens mot lusemidler oppstod hos lakselus på den tiden (Børretzen Fjørtoft mfl. 2017), og er derfor intakte i dag.
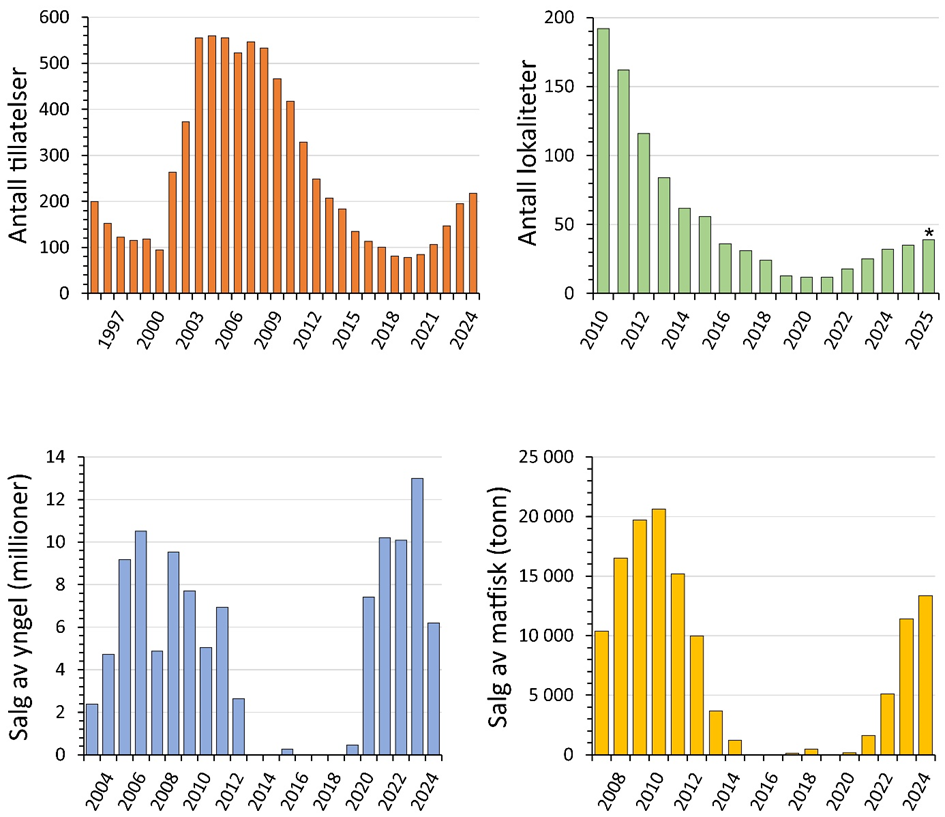
Etter sammenbruddet i torskeoppdrett for ti år siden har en av de private aktørene, Havlandet AS i Florø, beholdt en stamfisklinje for torsk, i tillegg til Nasjonalt avlsprogram for torsk ved Nofima i Tromsø. Disse to har systematisk avlet torsk nå i 6-9 generasjoner, og det hevdes at man har fått frem en mer domestisert torsk bedre tilpasset en oppdrettssituasjon og med betydelig fremgang i veksthastighet (Henriksen mfl. 2018; Puvanendran mfl. 2021). Yngelkvaliteten er forbedret og det er en stabil produksjon av yngel/settefisk. Sammenslutningen CodCluster, som består av blant annet 15 torskeoppdrettere, har en visjon om en samlet produksjon på 75 000 tonn i 2030, men det er enighet i næringen om at de ikke vil nå dette målet grunnet lav tilgang på nye lokaliteter. Det finnes 39 godkjente lokaliteter for oppdrett av torsk der 12 anlegg har torsk i sjøen i oktober 2025 (figur 2.5 og 2.6). Søknader om ytterligere 18 lokaliteter er under behandling.

Fremtiden for torskeoppdrett vil være avhengig av flere faktorer hvor avl vil spille en vesentlig rolle (Nardi mfl. 2021). Den fornyede interessen for torsken i oppdrett har derfor sin bakgrunn i avlsprogrammene i Tromsø og Florø som nå har foregått over henholdsvis 6 og 9 generasjoner. Ut fra erfaringene med forrige runde av torskeoppdrett vil forbedring av faktorer som vekst og yngelkvalitet, økt motstand mot sykdommer (særlig Francisellose), en roligere og mindre rømmingsvillig torsk samt kontroll med kjønnsmodning, være nøkkelfaktorer for fremtidig suksess. Det hevdes for eksempel at optimalisert bruk av lys i åpne merder vil kunne hindre kjønnsmodning før slakting.
Dokumentasjon av forbedringene gjennom avl er imidlertid mangelfull, og det foreligger ikke data publisert i internasjonale vitenskapelige tidsskrifter eller i offentlig tilgjengelige rapporter fra næringsaktører eller nasjonale forskningsinstitusjoner. Økt vekstrate er den faktoren som er best dokumentert, mens andre av de nevnte faktorene i stor grad mangler dokumentasjon. At disse kunnskapshullene lukkes, vil være viktig både for investorene, næringen og forvaltningen.
Videre vil torskeoppdrett konkurrere med oppdrett av laks og ørret med hensyn til egnede lokaliteter. Miljøproblemene som oppdrett av laksefisk sliter med, sammen med at befolkningen langs kysten utnytter en minkende kysttorskbestand til både næring og fritidsfiske, har ført til motstand mot torskeoppdrett. I den sammenheng er det nå planlagt både anlegg på land og lukkede anlegg i sjø. Kjønnsmodning kan se ut til å være mulig å kontrollere i anlegg som hindrer tilførsel av naturlig lys (Karlsen mfl. 2000; Almeida mfl. 2009), men organiske utslipp må fremdeles kontrolleres. Økte kostnader ved et mer teknologiintensivt oppdrett må heller ikke undervurderes. Til syvende og sist vil det være produktkvalitet og betalingsvillighet i markedene som vil være utslagsgivende for om oppdrettstorsken vil lykkes. Miljøsertifisering kan også bli en viktig faktor i denne sammenheng.
2.1.5.2 - Status og produksjon av oppdrettstorsk
Kysttorskbestandene forvaltes etter tre ulike forvaltningsområder. Disse forvaltningsområde er; sør for 62°N, mellom 62°N til 67°N, og nord for 67°N. Norskekysten er delt inn i 13 ulike produksjonsområder (PO) for akvakultur. Forvaltningsområdene for kysttorsk overlapper delvis med produksjonsområdene; henholdsvis PO 1-4, PO 5-8, og PO 9-13 (figur 2.7). Produksjon av torsk og søknader om torskeoppdrett er ikke jevnt fordelt mellom de ulike produksjonsområdene. Ved utgangen av august 2025 foreligger det ingen godkjente torskelokaliteter eller søknader om torskeoppdrett i PO1-3, 11 og 13. I PO7, 9 og 12 er det per i dag ingen aktive lokaliteter med fisk i sjø (produksjon av torsk), men det foreligger godkjente lokaliteter i tillegg til at det er nye søknader til behandling (tabell 2.1.a, b). I 2024 og 2025 har det vært 11-12 aktive sjølokaliteter hvor det produseres oppdrettstorsk. Disse har en samlet total kapasitet på 39 156 tonn. Ved utgangen av august 2025 var det gitt tillatelse til en økning i produksjon med drøyt 12.000 tonn fordelt på 4 lokaliteter, hvorav én er en utvidelse av allerede eksisterende lokalitet. Per 9. oktober 2025, foreligger det søknader om 18 nye lokaliteter med åpne merder i sjø til behandling. Disse har en samlet omsøkt biomasseøkning på 56 915 tonn. Sammen med nåværende godkjente kapasitet på 108 291 tonn, vil dette utgjøre en total kapasitet på 165 206 tonn, hvis alle søknadene godkjennes (tabell 2.1.a, b).
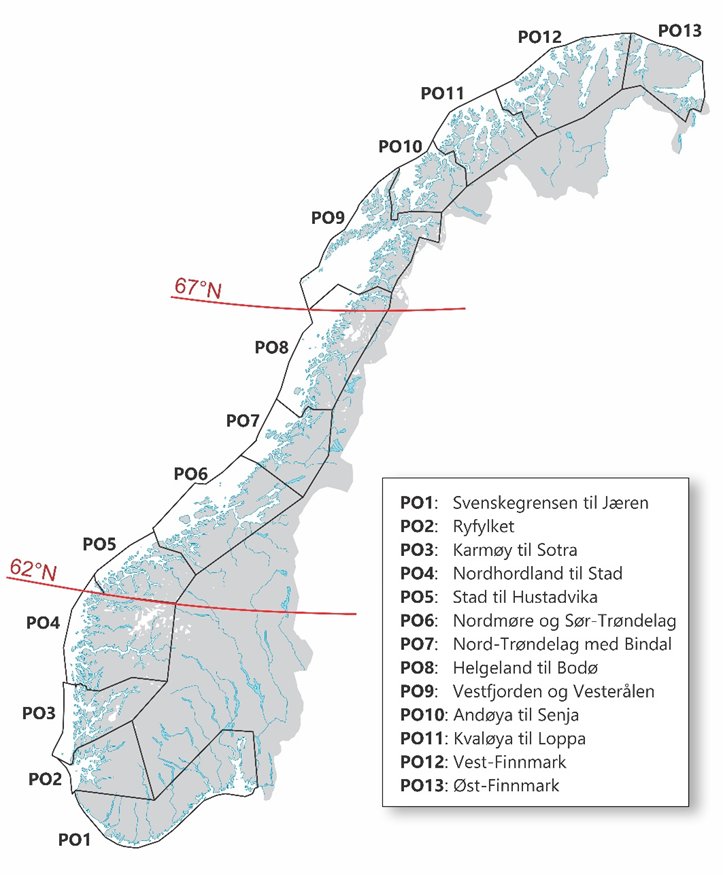
| Produksjons-område (PO) | KKB for godkjente lokaliteter per 09.10.2025 ** | KKB for lokaliteter i drift per 09.10.2025 | Biomasse i søknader per 25.8.2025* | Samlet biomassekapasitet for lokaliteter i drift og søknader |
| PO3 | 780 | 780 | ||
| PO4 | 7 340 | 5 780 | 3 599 | 10 939 |
| PO5 | 29 897 | 15 477 | 8 101 | 37 998 |
| PO6 | 20 059 | 11 879 | 14 099 | 34 158 |
| PO7 | 12 480 | 3 500 | 15 980 | |
| PO8 | 21 597 | 3 599 | 12 179 | 33 776 |
| PO9 | 3 210 | 9 197 | 12 317 | |
| PO10 | 10 199 | 4 600 | 1 860 | 12 059 |
| PO11 | ||||
| PO12 | 3 599 | 3 600 | 7 199 | |
| SUM: | 108291 | 41 335 | 56 915 | 165 206 |
| < 62°N | 7 340 | 5 780 | 4 379 | 11 719 |
| 62-67°N | 84 033 | 30 955 | 37 879 | 121 912 |
| > 67°N | 16 918 | 4 600 | 14 657 | 31 575 |
| SUM: | 108291 | 41 335 | 56 915 | 165 206 |
| Område | Totalt antall godkjente lokaliteter per 09.10.2025 | Lokaliteter i drift pr. 09.10.2025 | Søknader om nye lokaliteter * | Samlet antall søknader og godkjente lokaliteter |
| PO3 | 1 | 1 | ||
| PO4 | 2 | 1 | 1 | 3 |
| PO5 | 9 | 4 | 2 | 11 |
| PO6 | 7 | 4 | 4 | 11 |
| PO7 | 7 | 1 | 8 | |
| PO8 | 6 | 1 | 5 | 11 |
| PO9 | 3 | 3 | 6 | |
| PO10 | 4 | 2 | 4 | |
| PO11 | ||||
| PO12 | 1 | 1 | 2 | |
| SUM: | 39 | 12 | 18 | 57 |
| < 62°N | 2 | 1 | 2 | 4 |
| 62-67°N | 29 | 9 | 12 | 41 |
| > 67°N | 8 | 2 | 4 | 12 |
| SUM: | 39 | 12 | 18 | 57 |
2.2 - Kartlegging av gytefelt og oppvekstområder
Tradisjonelt har begrepet gytefelt blitt brukt for å omtale områder der fisken samles for å gyte. Aktive gytefelt er verdifulle for den enkelte art, og bør ikke forstyrres i gytetiden. Mange marine fisk har pelagiske egg som slippes og flyter fritt i vannmassene på gytefeltene (f.eks. torsk), mens hos noen arter limes eggene fast til vegetasjon eller stein og grus på bunnen (f.eks. sild). Ofte er de kystnære gytefeltene lokalisert til terskler og grunner i nærheten av større vannvolum som gir gode og stabile forhold for egg og larver i første fase av livet (Ciannelli mfl. 2010). Aktive gytefelt er et resultat av artens evolusjonære historie på den måten at her har gytingen med påfølgende overlevelse av egg, larver og yngel vært vellykket.
Et gytefelt kan ha en rekke egenskaper slik som spesiell bunntopografi, bassenger og terskler. Noen arter liker å gyte nær bestemte bunntyper eller i bestemte habitater. Også spesielle strømforhold kan være viktig. Fisk som har gytt på steder der avkommet er tatt med strømmene til ufordelaktige steder, har ikke overlevd i særlig grad. Gytefelt kan derimot naturlig ligge på steder der de pelagiske eggene og larvene driver til områder det vil være fordelaktig for avkommet å vokse opp. Mange arter i kystsonen gyter også på steder der egg og larver blir holdt tilbake av strømmene slik at avkommet vokser opp i nærheten av gytefeltene. Slike områder kaller vi retensjonsområder. Gytefelt der avkom ikke blir blandet med avkom fra andre gytefelt, kan bidra til å etablere genetiske forskjeller og gi grunnlag for lokale bestander av fisk. Dette er påvist for blant annet sild og torsk. Ofte vil fisk oppholde seg i et større område utenom gytesesongen, for så å svømme tilbake til gytefeltene i gytetiden. Da er det også viktig at den gytemodne fisken kan svømme til gytefeltet uten å møte hindringer eller blir skremt. Her kan du lese mer om gytefelt og gytetider.
Gytefelt har som regel vært definert som områder der man fanger fisk med rennende rogn og melke. Kartlegging av gytefelt etter denne tilnærmingen er problematisk fordi den er avhengig av fangbarhet og innsats. Gytefelt for kommersielt attraktive bestander vil være mer kjent enn gytefelt der fisken enten ikke er kommersielt like interessant eller er vanskeligere å fange. Noen ganger kan det være lettere å fange fisk på vei til et gytefelt enn på selve gytefeltet. Fisk som er rennende når de trekkes opp i båten, trenger ikke nødvendigvis gyte akkurat på stedet de er fanget.
Havforskningsinstituttet har kartlagt gytefelt langs hele kysten gjennom innsamling av nygytte egg i vannsøylen i tidsrommet februar til mai (Espeland mfl. 2007; Knutsen mfl. 2007; Espeland mfl. 2013; 2015). Denne metoden vil være påvirket av at innsamlingen gjøres på et tidspunkt det foregår gyting på gytefeltene. Det er vist at blant annet sjøtemperaturen kan forskyve gytetidspunktet (van der Meeren mfl. 2012), men også genetikk kan påvirke torskens gytetidspunkt (Otterå mfl. 2012). For å kunne vurdere om det er nok egg i et område til å kategorisere det som gytefelt, er det viktig også å ha dekket områder som ikke er gytefelt (kontrollområder). Biologisk kan små gytefelt være viktige dersom egg og avkom holdes tilbake på gytefelt som følge av strømforholdene i området (retensjon) og i liten grad blander seg med avkom fra andre gytefelt. Dette kan gi opphav til lokale bestander, med mulige unike tilpasninger (Ciannelli mfl. 2010; Jorde mfl. 2007; Olsen mfl. 2004).
Kartlegging av gytefelt for kysttorsk har benyttet data fra både feltinnsamling av egg, Fiskeridirektoratets intervjuundersøkelser og modellering av oseanografi for å vurdere hvilke områder som kan være de viktigste gytefeltene for stasjonære bestander av kysttorsk. Ettersom kartleggingen av gytefelter gjennomført av Havforskningsinstituttet har inkludert områder der egg oppholder seg i vannmassene i perioden umiddelbart etter gyting, vil ofte arealene for disse gytefeltene være større enn de intervjubaserte gytegytefeltene. I mange tilfeller er det god overlapp mellom Havforskningsinstituttets kartlagte gytefelt og de intervjubaserte gytefeltene, men de sistnevnte vil enkelte ganger reflektere gode fiskeplasser for gytemoden fisk som ikke nødvendigvis er gytefelt. I områder der det ikke er gjennomført feltundersøkelser vil de intervjubaserte gytefeltene representere best tilgjengelige kunnskap.
Kartleggingen av gytefelt med feltundersøkelser ble ferdigstilt, for alle fjorder i Norge, i 2019 gjennom prosjektet “Kartlegging av marine naturtyper”. Videre kartlegging og overvåkning av gytefelter er fra 2020 videreført med i prosjektet «Kartlegging av gytefelt og oppvekstområder for kommersielt viktige arter i kystsonen». Dette prosjektet vil overvåke gytefelt gjennom å besøke alle tidligere undersøkte stasjoner på en rullerende basis der hele kysten vil dekkes i løpet av 6-8 års perioder.
Gjennom prosjektet samles det inn egg fra vårgytende arter ved hjelp av vertikale håvtrekk. Stasjoner som undersøkes er primært stasjoner i fjordene og indre deler av kysten (eksempel i figur 2.8). En del områder i ytre kyststrøk, deriblant Lofoten og Vesterålen, er derfor ikke undersøkt (figur 2.9). Alle egg artsbestemmes, fotograferes og lagres på etanol (Espeland & Sannæs 2018). Det er lagt ned et stort arbeid i å etablere en genetisk metode for artsbestemmelse av egg. Teknologien som benyttes, tillater samkjøring av sekvensering av DNA fragmenter fra et høyt antall individuelle prøver. Det benyttes mitokondrielle DNA kjent for sin egnethet til å skille mellom arter hvor det er høy innbyrdes grad av slektskap, som for eksempel mellom forskjellige arter av torskefisk eller flyndrefisk.

For alle områder som undersøkes, gjennomføres det strømmodellering med tilhørende simulering av drift av egg (Espeland mfl. 2015). På denne måten kan driftsmønsteret tolkes slik at det er mulig å skille gytefelt der egg og larver holdes tilbake på gytefeltet og gytefelt som har større spredning av egg og larver og på den måten er knyttet til andre gytefelt gjennom utveksling av egg og larver (figur 2.10).
Den endelige registreringen av gytefelt baserer seg både på genetisk artsidentifiserte egg funnet i felt, strømmodellering og intervjuinformasjon. Alle gytefelt blir kartlagt og gjøres tilgjengelig gjennom Fiskeridirektoratets kartverk Yggdrasil, som kartlaget “Gytefelt Torsk MB”. Intervjubaserte gytefelt samlet inn av Fiskeridirektoratet ligger som kartlaget «Gyteområder». Det er viktig å bemerke at for ytre områder der det ikke er gjort eggundersøkelser vil intervjubaserte gytefelt representere «best tilgjengelige kunnskap».
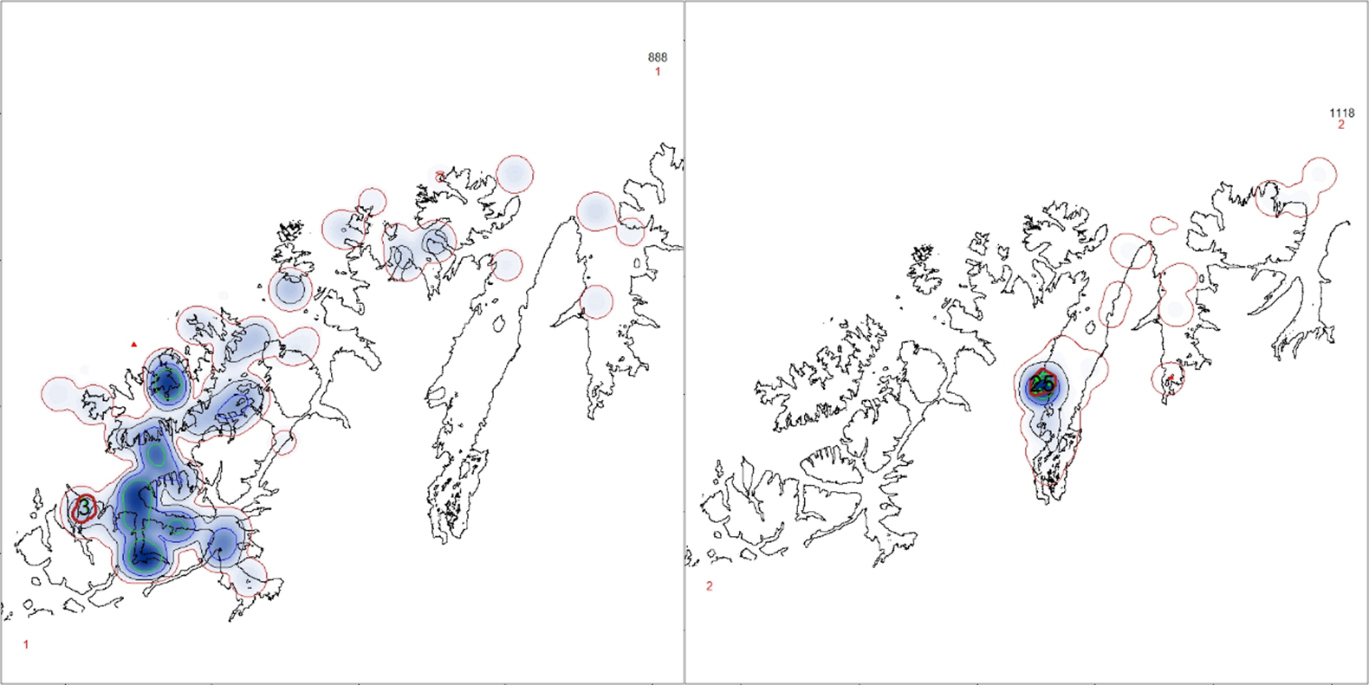
I 2025 har Havforskningsinstituttet også samlet inn data om gytemoden torsk utenom det som regnes som gytesesong om våren. Her ble det tatt i bruk folkeforskning gjennom nettsiden «Dugnad for Havet», og det ble registrert gytemoden og gytende torsk basert på bilder av gonadene i perioden fra juni til og med juli. Alle observasjonene ble gjort i Troms fylke (figur 2.11), men å kartfeste utbredelsen av gytefelt hvor denne sommergytingen finner sted er ikke mulig ut fra de få observasjonene som er gjort.

«Nasjonalt program for kartlegging av marine naturtyper» har ikke bare kartlagt gytefelt, men også ålegressenger, tareskog og andre viktige habitater som kan fungere som oppvekstområder i kystsonen. Gjennom prosjektet «Kartlegging av gytefelt og oppvekstområder for kommersielt viktige arter i kystsonen» gjøres også undersøkelser av betydningen av ulike habitater som oppvekstområde på utvalgte steder. Foreløpige resultater fra disse undersøkelsene tyder på at flere ulike habitater kan være viktige som oppvekstområder for kysttorsk. Mens ålegrasenger og tareskog i noen grad er kartfestet som oppvekstområder, er dette i liten grad tilfelle for andre typer oppvekstområder. Yngel av kysttorsk ser ut til å bunnslå på grunt vann (Godø & Sunnanå 1984; Jørstad mfl. 2014), og det er funnet gode forekomster av torskeyngel på grunn sand- og grusbunn med rødalger (van der Meeren mfl. 2021, Dunlop mfl. 2022). Dette habitatet er ikke kartfestet. Generelt vil de fleste grunne områder med påvekst av ålegras, tare eller rødalger kunne være viktige oppvekstområder for yngel av torsk og andre fiskearter.
2.3 - Risikokilder og hendelser inkludert i risikovurderingen
2.3.1 - Genetisk påvirkning på kysttorskbestandene
Oppdrettstorsk er avlet for et liv i merd under andre forhold enn hva villtorsken er tilpasset, og skiller seg fra denne i genetisk sammensetting. Ved innkrysning fra oppdrett introduseres ufordelaktige genvarianter og genkombinasjoner inn i de ville bestandene. Ufordelaktige gener er slike som favoriseres i oppdrett, fordi de forbedrer produksjonsegenskapene, men har lavere fitness (overlevelse og/eller reproduktiv verdi) i det naturlige miljøet. Så lenge slike genvarianter fortsetter å introduseres i kysttorskbestandene vil det være en naturlig seleksjon mot dem. Seleksjonen vil redusere forekomsten av ufordelaktige gener og på sikt kanskje eliminere dem dersom videre innkrysning fra oppdrett skulle opphøre på et tidspunkt (Glover mfl. 2025a). Denne seleksjonen har imidlertid en kostnad i form av økt dødelighet og redusert reproduktiv evne og -spesielt gitt de svake bestandene i dette området i utgangspunktet- vil kunne medføre negative effekter på kysttorskbestandene.
Genetiske interaksjoner mellom domestisert oppdrettsfisk og ville bestander er godt dokumentert og representerer en av de største utfordringene for et bærekraftig havbruk (Bekkevold mfl. 2006; Taranger mfl. 2015; Forseth mfl. 2017; Glover mfl. 2017).
Ser man på Atlantisk laks der denne problemstillingen er både mest utbredt (Diserud mfl. 2019; Glover mfl. 2019; 2020) og uten tvil har vært best studert, ser man at genetisk innblanding av rømt oppdrettsfisk forandrer livshistorie-trekkene hos de ville bestandene, og vil kunne føre til redusert produktivitet og tilpasning. Kort fortalt – mindre fisk og svakere bestander. En metastudie om økologiske effekter (13 fiskearter) og genetiske effekter (15 fiskearter) ved utsetting fra klekkerier viser at reduksjon av reproduktiv suksess og genetisk variasjon snarere er regelen enn unntaket for fiskearter i akvakultur (Araki & Schmid 2010).
Vi vet at oppdrettslaks er genetisk forskjellig fra villaks (se punktene nedenfor som også gjelder for torsk), og det er svært godt dokumentert at deres avkom viser redusert overlevelse i naturen (McGinnity mfl. 1997; 2003; Fleming mfl. 2000; Skaala mfl. 2012; 2019). Genetisk innblanding av oppdrettsfisk vil derfor, dersom den er høy nok, kunne føre til forandringer i livshistorie-egenskaper hos de ville bestandene, i for eksempel alder ved kjønnsmodning, størrelse ved alder, og i forandret timing av kritiske livshistorietrekk som tidspunkt for smoltutvandringen (Bolstad mfl. 2017; Skaala mfl. 2019). Slike forandringer kan føre til redusert produktivitet i ville bestander, og i det lange løp redusere bestandenes muligheter til å tilpasse seg naturens variasjoner og fremtidige endringer (f.eks. klimaendringer). Det bør derfor ta lærdom fra lakseoppdrett i denne tidlige fasen av torskeoppdrett.
Oppdrettsfisk, også oppdrettstorsk, er genetisk annerledes enn vill fisk på grunn av følgende faktorer:
-
Målrettet seleksjon for produksjons-egenskaper. Dette kan omfatte f.eks. hurtig vekst, sykdomsresistens, utsatt kjønnsmodning og reduksjon i aggressiv eller annen uønsket adferd.
-
Redusert naturlig seleksjon under oppdrettsforhold. Oppdrettsfisk er i liten grad utsatt for predasjon eller andre viktige naturlige prosesser (jakt på mat, migrasjon), men er i stedet oppdrettet i et trygt miljø med mye mat og høy overlevelse.
-
Founder-effekter. Oppdrettsfisk kan stamme fra andre geografiske områder/bestander og derfor tilhøre en annen genetisk gruppe enn bestandene de eventuelt kan blande seg med.
-
Innavl og tilfeldige genetisk forandringer er knyttet til begrenset antall gytefisk i avl og produksjon og forsterkes av at oppdrettsbestanden ikke mottar gener utenfra, noe som på sikt medfører redusert mengde genetisk variasjon og økt grad av innavl i bestanden.
Disse prosessene, spesielt de to første punktene, gjør fisken mer produktiv og velfungerende i et oppdrettsmiljø, men gjør dem samtidig dårligere tilpasset et liv i naturen. Innavl og tap av genetisk variasjon (pkt. 4) er uheldige bivirkninger i oppdrett som er vanskelige å eliminere, siden de følger av at et begrenset utvalg foreldre brukes i avlen, og at avlslinjene er lukket for genetisk påvirkning fra vill fisk. Vill torsk, som laks, utgjør ikke én enkel genetisk homogen bestand, men består av mange lokale bestander med ulike genetiske, adferdsmessige (f.eks. stasjonære/vandrende) og økologiske egenskaper (pkt. 3) (Otterå mfl. 2012; Dahle mfl. 2018; Johansen mfl. 2020). I nord er det spesielt store genetiske forskjeller mellom bestander av vandrende (skrei) og mer stasjonære (kysttorsk) økotyper (Berg mfl. 2016; Kirubakaran mfl. 2016). Tilsvarende i sør har vi Nordsjøtorsk og fjord-torsk som begge opptrer langs Skagerrakkysten (Barth mfl. 2017; Knutsen mfl. 2018) og på Vestlandet (Breistein mfl. 2022). De vandrende økotypene synes å stå sterkest på ytre kyst, og skreien minker også i en gradient fra nord mot sør (Breistein mfl. 2022). Innblanding av rømt oppdrettstorsk kan som følge av alle disse genetiske faktorene ødelegge naturlige tilpasninger hos lokale bestander. De eksakte konsekvensene av slik genetisk innblanding vil variere i tid og rom, være avhengig av mange faktorer og kan derfor vanskelig forutsies med presisjon.
Det er relativt lite publiserte data på genetiske egenskaper hos domestisert torsk (Otterå mfl. 2018), men det vises til økt veksthastighet ved allerede etter tre generasjoner med avl (Henriksen mfl. 2018; Puvanendran mfl. 2021). På laks har avl resultert i doblet veksthastighet i fangenskap i løpet av de fem første generasjonene (Gjedrem 2000; 2010). Allerede etter så kort tid har dette ført til en målbar reduksjon i overlevelsen i et naturlig miljø (McGinnity mfl. 1997; Fleming mfl. 2000; McGinnity mfl. 2003). På torsk er det lengst komne avlsprogram nå ca. 6-9 generasjoner og derfor antakelig godt på vei i domestiseringsprosessen. Ved 2-3 generasjoner var det dokumentert at avkom av oppdrettstorsk har høyere vekst enn avkom av vill torsk under identiske forhold (Otterå mfl. 2018; Henriksen mfl. 2018). Det betyr at det allerede da var målbare genetiske forskjeller i egenskaper mellom oppdrettet og vill torsk. Forskjellene er antakelig enda større i dag siden det nå er gått flere generasjoner i de to eksisterende avlsprogrammene. Selv om det ikke er studert ennå, kan avkom av en avlet oppdrettstorsk, basert på erfaring fra andre arter, få en redusert overlevelse i naturen (Glover mfl. 2017). Jo flere fisk som rømmer i forhold til den lokale bestandens størrelse, jo større er sannsynligheten for genetisk innblanding. Hos laks er det en god korrelasjon mellom andel rømt fisk observert i en bestand over tid, og graden av genetisk innblanding (Glover mfl. 2013; Heino mfl. 2015; Karlsson mfl. 2016). Økt genetisk innblanding gir økt konsekvens for den ville bestanden (Hindar mfl. 2006; Bolstad mfl. 2017; Castellani mfl. 2018). De samme prinsipper og mekanismer vil også gjelde for torsk. Dette betyr at små lokale bestander antakelig er mer utsatt for genetisk innblanding enn de større og mer robuste bestandene. For eksempel, kan lokale bestander i fjorder lett bli oversvømt genetisk av oppdrettsfisk fra et lokalt anlegg dersom det er mye gyting i merd, eller ved en rømmingshendelse.
I motsetning til laks som etter rømming må vandre opp i elva til gyteplassene for å kunne blande seg med vill fisk, vil rømt oppdrettstorsk i en del tilfeller allerede være ved gyteplassen og har derfor stort potensiale til å kunne gyte med vill torsk. Derfor er plassering av anlegg i henhold til naturlige gyteplasser en viktig risikofaktor. Når først rømming har skjedd vil adferdsmessige faktorer også kunne være bestemmende for om gyting med vill torsk kan skje. Studier har vist at hunnfiskens valg ser ut til å være av betydning, og mens oppdrettshanner har dårlige odds til å få gyte med ville hunner, kan ville hanner ha bedre muligheter hos oppdrettshunner (Meager mfl. 2009; 2010; Skjæraasen mfl. 2010).
Det vil antakelig være vanskeligere å spore genetisk innblanding av rømt oppdrettstorsk i ville bestander enn for laks, men dette vil være avhengig av hva slags torskemateriale avlsprogrammene har brukt som utgangspunkt (Glover mfl. 2011). Genetiske markører for sporing av rømt oppdrettslaks til sitt opprinnelsesanlegg (ved urapporterte rømminger) vil kunne fungere også for oppdrettstorsk (Glover 2010; Glover mfl. 2010). Dette fordi det er genetiske forskjeller mellom grupper med torsk i et anlegg, antakelig fordi hver merd består av et relativt begrenset antall familier (Glover mfl. 2010; Glover mfl. 2011). Empiriske data som ble samlet inn i forbindelse med en større rømmingshendelse i Meløy i januar 2023 bekrefter at rømt oppdrettstorsk kan skilles fra ville artsfrender med samme metodikk som for laks (Jorde mfl. 2024). Men, dersom rømt torsk krysser seg med vill fisk vil det på sikt bli vanskelig å måle hvor mye innkrysning som har skjedd fra oppdrettstorsk til vill torsk med nåværende metodikk.
For laks konkluderes det med at antall rømmingshendelser må reduseres for å redusere risikoen for genetisk innblanding med påfølgende negative konsekvenser for ville bestander. Det bør også vurderes å ta i bruk steril fisk slik at de ikke kan formerer seg med vill fisk. Disse to tiltakene vil være å anbefale også for torskeoppdrett.
2.3.1.1 - Rømming av oppdrettstorsk
På samme måte som rømming av oppdrettslaks har blitt et betydelig problem for laksenæringen og forvaltningen av ville bestander av laks, er det sannsynlig at rømming av oppdrettstorsk vil bli et tilsvarende problem for torskenæringen, ville bestander av torsk og forvaltningen av torsk. Dette understrekes av flere større rømmingshendelser med oppdrettstorsk de siste årene (2022 i Voldsfjorden, Møre og Romsdal; 2023 i Meløy og 2024 i Nesna, begge i Nordland) som til sammen inkluderer over 100 000 fisk, i to av tilfellene gytemoden, rømt oppdrettstorsk (figur 2.12).
I Voldsfjorden var det ca. 87 000 torsk med ca. 600 g snittvekt som rømte. Prøvetaking av torsk ble tatt i fjordområdene rundt oppdrettsanlegget vintrene 2022/2023 og 2023/2024, og egg ble samlet inn fra vannmassene i mars 2024 da første kjønnsmodning hos eventuell overlevende rømt torsk var ventet. Ved DNA analyser ble det verifisert rømt torsk opp til 20 km fra anlegget, men kun ett av i alt 233 analyserte egg var fra oppdrettstorsk (Glover mfl. 2025b). Den rømte torsken må lære seg å fange byttedyr, og det er derfor usikkerhet knyttet til overlevelse og tid som trengs for å nå kjønnsmodning. Muligheten er likevel til stede for at individer fra denne rømmingen kan dukke opp seinere som gytere.
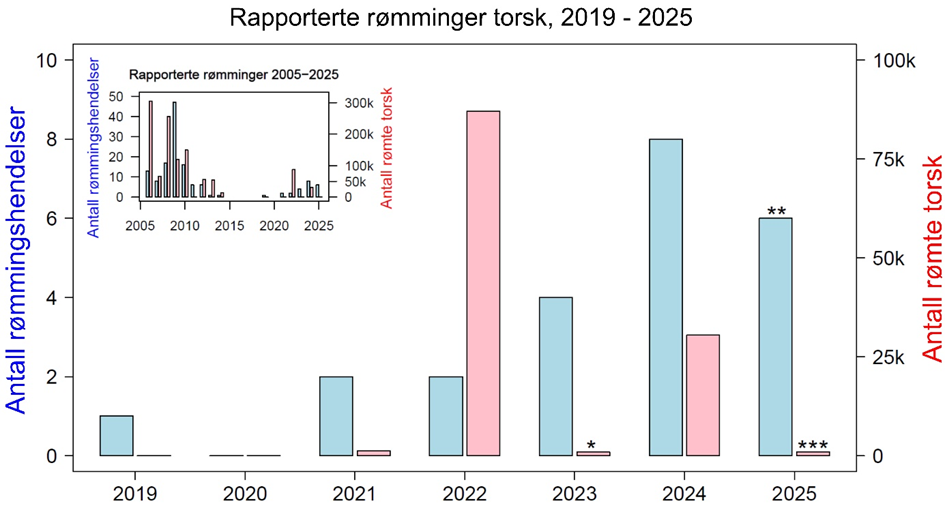
Det er derfor viktig å ta med seg den kunnskapen om rømming av oppdrettstorsk som ble opparbeidet parallelt med den forrige bølgen av torskeoppdrett, blant annet i regi av Havforskningsinstituttet, Norges Forskningsråd og EU, for å forstå “hvorfor, hvordan og hvor” torsken rømmer (f.eks. Hansen mfl. 2009; Uglem mfl. 2008; Dahle mfl. 2011; Jørstad mfl. 2014). Hensikten med disse prosjektene var å undersøke adferdsmekanismene bak rømming og å kartlegge spredningen av rømt torsk under naturlige forhold. Prosjektene bestod av kontrollerte lab- og merdeksperimenter, samt storskala fjordeksperimenter, og de spesifikke målsettingene var å studere grunnleggende adferdsmekanismer som er involvert når torsk rømmer. Dette inkluderte å identifisere typiske karaktertrekk bak “rømmeadferd”, undersøke adferd og bevegelser hos torsk etter rømming og foreslå forbedrete metoder for overvåkningsfiske og gjenfangst. I tillegg var det et mål å framskaffe kunnskap som legger til rette for å evaluere mulige negative miljøeffekter av rømt oppdrettstorsk.
Hensikten med lab- og merdeksperimenter var å undersøke om faktorer som størrelse, fôringsstatus og genetisk opprinnelse påvirker motivasjonen til å svømme gjennom en åpning i en notvegg (Hansen mfl. 2009). Torsken som ble brukt, kom fra første generasjons stamfisk fra torskeavlsprogrammet til Nofima. Kort tid etter forsøkstart begynte torsken å svømme gjennom hullet i notveggen, og rømmefrekvensen var høyest like etter at hullet var laget. Torskens størrelse hadde ingen innvirkning på motivasjonen til å rømme, og var heller ikke betydelig påvirket av fiskens fôrstatus, men ni dagers sulting førte til flere torsk på “rømmesiden” av merden. Det var også en høyere andel rømt kysttorsk enn skrei, og gjentatte forsøk viste en betydelig andel gjengangere på “rømningssiden”. Oppsummert viste disse studiene at torsken ofte raskt svømmer ut av et hull som oppstår i en notvegg, at rømming er et individuelt valg og at det ikke er tilfeldig hvilke individer som rømmer (Hansen mfl. 2009). Påfølgende studier viste at faktorer som legger til rette for denne typen undersøkende adferd hos torsk, kunne knyttes til både fysiske faktorer i notveggen og tilgang på fôr (Zimmermann mfl. 2012; Damsgård mfl. 2012). Både lengre perioder uten fôr og kortere perioder med begrenset tilførsel av fôr økte slik undersøkende rømmeadferd – det samme gjorde også skader eller reparasjoner i notvegg. Torsk var i tillegg i stand til å øke omfanget av notskader gjennom aktiv biting på notlinen (Moe mfl. 2007; Damsgård mfl. 2012).
Spredning av oppdrettstorsk etter rømning har blant annet blitt undersøkt ved simulert rømming fra oppdrettsanlegg i fullskala fjordstudier der torsk ble merket med akustiske sendere slik at vandringer i fjorden kunne undersøkes (Uglem mfl. 2008; Uglem mfl. 2010). Resultatene viste at rømt oppdrettstorsk forlater området like rundt anlegget raskt etter en simulert rømning. Bare noen få timer etter rømningen var over halvparten av oppdrettstorsken forsvunnet fra anlegget, og i løpet av fire dager var nesten all fisken vekk. De rømte torskene spredte seg raskt utover relativt store områder av fjorden. Noen av oppdrettstorskene vandret utover mens andre vandret innover og oppsøkte hyppig kjente gyte- og fiskegrunner i indre deler av fjorden, spesielt i løpet av gyteperioden for den lokale fjordtorsken. Imidlertid ble omtrent 30 – 50 % av fisken også regelmessig observert i anleggets nærområde etter rømning (innenfor 3 km av anlegget). Fjordfiskernes gjenfangst var også forbausende høy, og hele 44 % av all merket oppdrettstorsk ble gjenfanget uten ekstraordinær fiskeinnsats (Uglem mfl. 2008). Tilsvarende forsøk med stor kysttorsk og stor skrei (første generasjon fra Nofima sitt avlsprogram), viste ingen større forskjeller mellom kysttorsk og skrei. Også disse forlot anleggene raskt, dykket ned til dyp > 20 meter, beveget seg mye langs land og til dels over (større områder, oppsøkte gytefelt, men returnere også regelmessig til oppdrettsanlegget. Gjenfangsten var høy også i denne studien (31 %) (Uglem mfl. 2010).
Påfølgende forsøk med torsk av mindre størrelse i et annet fjordsystem i Nord-Norge viste at rømt småtorsk var utsatt for en betydelig størrelsesrelatert predasjon fra vill torsk og sei rundt oppdrettsanlegget (Serra-Llinares mfl. 2013). Den rømte småtorsken forlot anlegget raskt og søkte tilflukt på grunt vann opp mot fjæresonen hvor de oppholdt seg relativt nært anlegget i flere måneder. Gjenfangsten av den mindre fisken var imidlertid lav (< 5 %) på tross av aktivt gjenfangstfiske med flere redskaper (Serra-Llinares mfl. 2013). I en parallell studie i samme fjordsystem ble det merket 20 tidligere rømte oppdrettstorsk som ble gjenfanget like ved oppdrettsanlegget (Hedger mfl. 2017). Disse viste høy tilknytning til anlegget også etter utsett, spesielt gjennom mørketiden. Få individer besøkte andre anlegg eller forlot fjorden, og dødeligheten var høy (> 40%) i løpet av de fire månedene forsøket varte.
Tilsvarende observasjoner har blitt gjort i et merke-gjenfangstforsøk i Trondheimsfjorden (Varne 2016) like i etterkant av to større rømminger fra et torskeoppdrettsanlegg. Gjenfangsten var høyest de to første månedene etter rømming, spredningen i fjorden var tilsynelatende tilfeldig og relativt begrenset, og få ble gjenfanget etter mer enn 6 måneder, noe som kan indikere lav overlevelse og begrenset tilpasningssuksess. Som en del av avhandlingen til Varne (2016), ble det utviklet metodikker for å skille rømt oppdrettstorsk fra villtorsk basert på morfologi og skjell (Uglem mfl. 2011). Mikrostrukturen i øresteinene (otolittene) er også foreslått som metode til å kunne skille oppdrettstorsk fra villtorsk (Dahle mfl. 2011).
I forsøk gjennomført av Havforskningsinstituttet ble ett år gammel merket oppdrettstorsk av skrei og lokal kysttorsk (24-27 cm lengde) satt ut i et eksponert kystlandskap i Øygarden nordvest for Bergen. Gjenfangsten skjedde hovedsakelig innen et år og var lav (< 8,6 %) men størst for kysttorsken, trolig på grunn av forskjeller i dødelighet mellom de to stammene (Otterå mfl. 1999). Alle gjenfangster skjedde innen 15 km fra utslippsstedet, og ingen forskjeller ble funnet mellom skrei og kysttorsk når det gjaldt distanse fra utslippssted. Disse resultatene støttes også fra en større havbeitesatsing i årene fra 1985-2000, hvor nærmere 1 mill. kultivert torsk (8-34 cm) med opprinnelse fra oppdrett i poll, ble merket og satt ut i kyst- og fjordområder langs Norskekysten (Svåsand mfl. 2000). Det generelle trekket viser at de fleste gjenfangstene ble tatt nært utsettingsstedet, men også at det er en økende spredning fra sør til nord for hovedutsettingsområdene.
I et stort merke/gjenfangst-forsøk i Ullsfjorden i Troms var bevegelsesmønsteret for liten torsk (< 27 cm ved merking) likt for vill og oppdrettet torsk, mens hos større oppdrettstorsk hadde raskere og større spredning enn tilsvarende størrelse vill torsk (Pedersen mfl. 2008). Gjenfangstraten totalt i dette forsøket var 14,7 % (2877 fisk) for vill torsk og 8 % (2073 fisk) for oppdrettstorsk. Imidlertid ble kun 2,2 % av gjenfangstene (107 fisk) gjort utenfor Ullsfjorden, der halvparten av disse be fanget 50-100 km fra utslippspunktet. Maksimal gjenfangstdistanse var 726 og 780 km for henholdsvis oppdrettet og vill torsk. I en canadisk studie fra Newfoundland var tilsvarende gjenfangstdistanse 256 km for vill torsk og 157 km for oppdrettet torsk (Zimmermann mfl. 2013).
Observasjoner i Norddalsfjorden ved Florø og Masfjordområdet av rømt oppdrettstorsk, vurdert ut fra både en genetisk markør og morfologiske avvik, har vist at en betydelig andel holdt seg i nærheten av anlegget der rømmingen skjedde (Dahle mfl. 2011). Her ble det observert vedvarende episoder med rømming (Jørstad mfl. 2014), men når driften opphørte forsvant den rømte oppdrettstorsken i stor grad. Tre mulige forklaringer har blitt lansert for dette (Dahle mfl. 2011): a) rømt torsk finner mat ved anlegget, b) høyt fiskepress kan føre til at andelen av rømt torsk fiskes ut relativt raskt når ikke ny tilførsel kan skje gjennom nye rømminger, og c) rømt torsk overlever ikke særlig lenge utenfor merden grunnet problemer med å omstille kostholdet til «levende mat». Angående det siste punktet ble det observert lav kondisjon på en del rømt torsk samt individer med magen full av blåskjell (Dahle mfl. 2011), men vi har generelt lite data på næringsvalg til domestisert oppdrettstorsk, inkludert eventuell predasjon på yngre torsk. Jørstad mfl. (2014) viste at rømt torsk overlevde 3 år etter rømming, og at de utgjorde en betydelig andel av den tilsvarende årsklassen hos vill torsk. Tilpasning, eller mangel på tilpasning til et liv i det fri vil sannsynligvis avhenge av tid i oppdrett og grad av domestisering. Problemstillingen ble grundig studert som del av havbeiteforsøkene, og her ble det dokumentert at polloppdrettet torsk forholdsvis raskt tok til seg føde og skilte seg lite fra villtorsk kort tid etter utsetting som 0-gruppe (Svåsand mfl. 2000). Tilsvarende studier har ikke vært gjennomført med domestisert oppdrettstorsk.
Den genetiske markøren hos pollprodusert oppdrettstorsk gjorde det mulig å vurdere vandring etter rømming i et større område. Her ble det funnet torsk med genmarkøren på gytefelt opp til 21 km unna i samme fjordsystem som oppdrettsanlegget, og i tillegg 32 km inn i nabofjorden. Tendens til økt frekvens av den genetiske markøren ble observert hos larver og juvenil torsk, men ikke statistisk signifikant til å konkludere med at oppdrettstorsk krysset seg med villtorsk (Jørstad mfl. 2014).
Oppsummert tyder resultatene fra alle disse studiene på at rømt oppdrettstorsk har motivasjon for å finne eller til og med lage hull i notveggen, at det ikke er tilfeldig hvilken fisk som rømmer og at dette også påvirkes av egenskaper ved notvegg og ernæringsmessig status. Etter rømming dykker større torsk raskt ned til ca. 20 meters dyp, vandrer inn mot land, og kan raskt spres over større områder, selv om en del rømt torsk også vil holde seg i nærheten av anlegget de har rømt fra. Gjenfangst av rømt torsk ved anlegget bør skje umiddelbart etter rømning og nært land, men en høy total gjenfangst antyder også at rømt oppdrettstorsk kan være lettere å gjenfange enn rømt laks i et mer langsiktig gjenfangstfiske i et større område. Et problem kan imidlertid være at et intensivt gjenfangstfiske også kan gå ut over lokal villtorsk, og risikoforebyggende tiltak i anlegg er derfor viktig. Yngre torsk ser ut til å bevege seg i mindre grad enn større torsk etter rømming, og bruk av redskaper som fisker effektivt på grunt vann (f.eks. åleruser) kan være en strategi for gjenfangst av ung rømt torsk. Ruser vil også skåne andre arter i et eventuelt gjenfangstfiske.
Videre tyder spredningsmønsteret hos rømt oppdrettstorsk på at det er et klart potensial for interaksjon mellom rømt fisk og villfisk, også i gytetiden, selv om studiene fra Trondheimsfjorden og Norddalsfjorden ikke klarte å finne klare indikasjoner på innkryssing av oppdrettsgener hos den lokale villtorsken. Den nye interessen for torskeoppdrett er knyttet til en domestisert oppdrettstorsk. Det mangler imidlertid dokumentasjon på adferd hos denne torsken som kan belyse om den har større eller mindre trang til å rømme fra merdene, og hvordan den eventuelt vil fordele og klare seg i et fjordsystem hvis rømming skulle skje. Slik informasjon vil være vesentlig for valg av strategier med hensyn til å både å forebygge og redusere effektene av rømminger.
2.3.1.2 - Gyting i merd
I motsetning til laksen vil torsken kunne gyte i merdene. Gyteadferd hos torsk involverer trolig partnervalg med kurtise (Karlsen & van der Meeren 2013), og det er observert at torsk kan gyte i små vannvolum (van der Meeren & Ivannikov 2006). Fra et forsøk i Heimarkspollen i Austevoll der oppdrettstorsk med en genetisk markør fikk gyte i en merd, ble det påvist at befruktede egg vil kunne unnslippe merdene og overleve til kjønnsmoden torsk som igjen vil kunne bidra til gyting på lokale gytefelt (Jørstad mfl. 2008; van der Meeren mfl. 2012). Det ble funnet en betydelig økning av den genetiske markøren, med forekomst hos opp til 2,4 % av torskelarvene i neste generasjon 5 år etter at første forsøket med gyting i merd ble gjennomført. Dette er opp fra 0,02-0,04 % som er naturlig forekommende for denne markøren (Jørstad mfl. 1991). Resultatene viste et tydelig signal på overlevelse av egg gytt i merd til kjønnsmoden fisk, men overlevelsen kunne ikke kvantifiseres.
En studie med genetisk merket torsk i et kommersielt oppdrettsanlegg i Norddalsfjorden ved Florø påviste larver og yngel av torsk med den genetiske markøren i og ved gytefelt opp til 19 km unna anlegget, men grunnet rømmingsepisoder fra anlegget var det ikke mulig å si om dette var avkom fra gyting i merd eller fra rømt torsk (Jørstad mfl. 2014). I forsøk med genetisk merket torsk i merder i Heimarkspollen i Austevoll ble det observert at 12 % av torskelarver samlet inn 8 km unna merdene hadde sin opprinnelse fra gyting i merd inne i pollen, mens tilsvarende andel inne i pollen var 36 %. Gener fra oppdrettstorsken vil derfor enkelt kunne spres uten at oppdrettstorsken faktisk rømmer fra merdene. En studie i Trondheimsfjorden viste at betydelige mengder med egg fra gyting i merd hovedsakelig drev i overflaten utover fjorden, og det ble derfor ikke funnet torskelarver fra denne gytingen eller noe genetisk signal for juvenil torsk i indre fjordbasseng (Beistadfjorden) tre år etter observert gyting (Varne mfl. 2015). Det er påvist at torsk med opphav fra gyting i merd, gyter med hverandre, men det er ikke sikkert påvist at slik torsk faktisk krysser seg med villtorsk (van der Meeren mfl. 2012). Data fra Jørstad mfl. (2014) indikerte at krysninger mellom oppdrettstorsk og villtorsk kan ha funnet sted, men at dette ikke kunne påvises sikkert statistisk sett. Også Jorde mfl. (2024) indikerer at slik krysning kan ha funnet sted. Adferdsstudier i tankforsøk og merkeforsøk på gytefelt med oppdrettstorsk og villtorsk har imidlertid vist at det er sannsynlig at kryssing kan skje mellom vill- og oppdrettstorsk (Meager mfl. 2009, 2010; Skjæraasen mfl. 2010).
I forbindelse med en større rømmingshendelse i januar 2023 i Meløy (Nordland) ble det konstatert gytemoden, rømt oppdrettstorsk og at oppdrettstorsken i det lokale anlegget på Frosvika var i ferd med å utvikles til gytemoden fisk. Basert på vevsprøver samlet inn av Fiskeridirektoratet på anlegget og fra mistenkt rømt torsk levert av lokale fiskere på mottak, ble det etter genetiske analyser konstatert egg av oppdrettstorsk på lokale gytefelt som mest sannsynlig stammer fra gyting i merd (Jorde mfl. 2024). Av i alt 47 egg med oppdrettsbakgrunn hadde ett egg en genetisk profil som er konsistent med krysning mellom oppdrettet og vill torsk. Det gjenstår å undersøke i hvilken grad slik gyting mellom vill og oppdrettet torsk forekommer, og hvilke langsiktige konsekvenser det har for ville bestander.
Biomassen i et matfiskanlegg for torsk kan trolig være betydelig sammenlignet med biomassen av villtorsk på lokale gytefelt, men data for dette mangler fordi bestandene av vill torsk knyttet til gytefeltene i de enkelte fjordsystemene ikke er kjent. I forsøkene med gyting i merd i Heimarkspollen i Austevoll førte gyting hos 2 til 3 tonn oppdrettstorsk til en andel av larver fra gyting i merd på opp mot 36 % av alle torskelarver i pollen (Jørstad mfl. 2008). Videre vil oppdrettstorsken ha betydelig flere egg (fekunditet) enn villtorsk (Wroblewski mfl. 1999). Teoretisk vil en fullført gytesesong fra et matfiskanlegg på 500 tonn med lik kjønnsfordeling kunne produsere mellom 150 og 200 milliarder egg hvis all torsken blir kjønnsmoden, og 50 til 65 milliarder egg om kun en tredjedel av torsken kjønnsmodner. Det forventes imidlertid at kvaliteten på egg fra fremavlet oppdrettstorsk er dårligere enn egg fra villfisk, og at overlevelsen til ulike stadier frem mot voksne individer også vil være redusert.
En forutsetning for å kontrollere gyting i merd er kontroll på kjønnsmodning. Kjønnsmodning hos torsk er vanskeligere å hindre ved bruk av lys enn det som er tilfelle for oppdrettslaks. Ved bruk av lys kan gyting i merd derfor forekomme hos en viss andel av bestanden, men modningen vil forsinkes noe (Taranger mfl. 2006; Korsøen mfl. 2013) slik at gyting kan skje utenfor den naturlige gytesesongen før slakting.
Det finnes ikke publisert informasjon om en domestisert torsk reagerer annerledes på lysstyring enn det som har vært observert tidligere hos torsk som ikke har vært gjennom et avlsprogram. Et pågående prosjekt ledet av HI (MOTOR, Modningsfri torsk i merd, FHF, prosjektnr 901815) setter søkelys på å utvikle en innovativ og bærekraftig metode for å produsere torsk i merder frem til slaktestørrelse, utsette kjønnsmodning og forebygge spredning av egg fra gyting i merd ved bruk av LED-lys og demping ved nedsenkning og bruk av skyggenett. De foreløpige resultatene viser at fisk under lysstyring ikke viste avanserte stadier av kjønnsmodning før de nådde slaktestørrelse (Havforskningsinstituttet, upubliserte data fra MOTOR-prosjektet). Det er verd å merke seg at disse resultatene ble oppnådd under lysbehandling i merdene i deler av produksjonssyklusen for torsk, og de bør derfor tolkes med forsiktighet. Funnene bør verifiseres gjennom studier som omfatter hele produksjonssyklusen i sjømerder. Oppdrettstorsk i naturlig lys var i stand til å gyte levedyktige egg, selv om befruktningsratene og utviklingssuksessen til embryoene var lavere enn hos referansegruppen av villtorsk. Villtorskelarvene så imidlertid ut til å være mindre enn larvene fra oppdrettsgruppene (Alix mfl. 2025).
Ved å benytte lys i merder kan kjønnsmodning utsettes og derved inntreffe i temperaturer som er ugunstig for gonadeutvikling før gyting. Det er også observert at eggmodning skjer raskere ved økt temperatur (Kjesbu mfl. 2010). En temperatur på over ca. 10°C vil føre til redusert befruktning og feilutvikling hos en betydelig andel av eggene (van der Meeren & Ivannikov 2006). Etter befruktning er det observert økt eggdødelighet over 9°C (Dahlke mfl. 2018, Skjærven mfl. 2024), mens nyklekte torskelarver har en øvre temperaturtoleranse på 10-12°C (Iversen & Danielsen 1984). Torskelarver utvider imidlertid sin øvre toleransegrense gjennom utviklingen, og larver med lengde mellom 25 og 40 mm har vært observert å kunne vokse med opptil 37 % i vekt pr. døgn ved ca. 17°C (Finn mfl. 2002). Det er vist at optimal temperatur for vekst kan endres over kun en generasjon hos fisk ut fra hvilken temperatur foreldregenerasjonen holdes under (Salinas & Munch 2012). Om slike mekanismer kan endre temperaturtoleranse hos torsk som har vært avlet over flere generasjoner, er imidlertid ikke kjent. Torskelarvers mulighet for overlevelse utenom normal gytesesong om våren er diskutert i van der Meeren mfl. (2024), hvor det pekes på at temperaturforhold og tilgjengelighet av byttedyr er til stede for perioder av året og deler av kysten der overlevelse er mulig. I denne sammenheng er det også verd å nevne de nye observasjonene av sommergytende villtorsk, som kan indikere overlevelse av egg og larver utenom normal gytesesong om våren i de nordlige landsdeler (figur. 6.11).
Bedret vekstrate hos en domestisert torsk kan påvirke tidspunkt for modning og derav tidspunkt for utslakting. Otterå mfl. (2018) fant variabel effekt i vekst hos 2. generasjon oppdrettstorsk fra to avlsprogram i Norge (Torskeavlsprogrammet i Tromsø og Marine Breed), sammenlignet med avkom fra villtorsk. Fra avlsprogrammet i Tromsø er det imidlertid rapportert om 10 % økning i vekst for hver generasjon (Henriksen mfl. 2018).
2.3.1.3 - Vurdering av modningsgrad hos oppdrettstorsk 2023-2025
I en rapport fra Havforskningsinstituttet er det publisert en protokoll for å vurdere modningsgrad hos oppdrettstorsk (Alix mfl. 2024). Denne rapporten presenterer en visuell metode for overvåkning med hensyn til modningsgrad hos oppdrettstorsk, samt data fra fire oppdrettsanlegg andre vinteren (2023–2024) i sjø. Altså hvor fisken snart ville være slakteklar. Andelen fisk identifisert som gytende eller utgytt i merdene var mellom 11,2 og 43,3 % fra mai til juli, mens tilsvarende andel for torsken i samme tidsrom ved slakteriet var mellom 19,4 og 28,4 %. Andelen umoden torsk i merdene sank fra 11,5 % i januar til 1,8 % i juni, mens i juli ble det ikke funnet umoden torsk.. Det ble registrert en økning i forekomsten av gytende/utgytte individer frem mot juni, både i merdene og i slakteri. Denne utviklingen kan trolig knyttes til bruken av kontinuerlig lys i merdene, som benyttes for å hindre eller forsinke kjønnsmodning (Taranger mfl. 2006; Korsøen mfl. 2013).
I slutten av 2024 sendte Fiskeridirektoratet ut skriftlig vedtak til de to torskeoppdretterne som hadde torsk i sin andre vinter i sjø. Oppdretterne ble pålagt å overvåke og rapportere modningsgrad i henhold til en egen prosedyre, som var vedlagt sammen med protokollutkastet fra Havforskningsinstituttet (Haugland & Jensen 2025). Det ble rapportert modningsstadier og fiskedata i både første og andre vinter i sjø, modningsgrad ved slakteri samt bilder av gonadene (rognsekk og testes). Innrapporteringen til Fiskeridirektoratet med rådataene fra oppdretterne viste en andel gytende/utgytt torsk på mellom 0 og 20 % i merder og 0 og 33 % ved slakteri i perioden januar til juli i 2025 (Haugland & Jensen 2025).
Disse dataene ble også analysert og kontrollert av Havforskningsinstituttet (upubliserte data). Totalt ble 4284 bilder av gonader gjennomgått av Havforskningsinstituttet i perioden desember 2024 til juli 2025. Fire lokaliteter inngikk i denne overvåkningen. Henholdsvis ble totalt 2221 hunntorsk (blant alle fiskene tilsvarer det 52% i merd og 53% ved slakteri) og 2063 hanner (blant alle fiskene tilsvarer det 48% i merd og 47% ved slakteri) identifisert i sin andre vinter i sjøen.
Andre vinter i sjø varierte andelen umoden fisk mellom 5 og 33 %. Andelen gytende eller utgytt torsk i merd varierte mellom 1 og 35 %, og mellom 2 og 40 % ved slakteri (Tabell 2.2). Andre vinter i sjø viste vurderingene i gjennomsnitt at 16 % av torsken var gytende eller utgytt (21 % hunner blant alle gytende/utgytt fisk) i merd, og 30 % (34 % hunner blant alle gytende/utgytt fisk) ved slakteri i perioden desember 2024 til juli 2025.
| Måned | Antall prøver | Prosent umoden | Prosent modnende | Prosent gytende/utgytt |
| Desember merd i sjø | 735 | 27 | 71 | 1 |
| Desember slakteri | 100 | 7 | 81 | 12 |
| Januar merd i sjø | 894 | 26 | 59 | 14 |
| Januar slakteri | 150 | 5 | 60 | 33 |
| Februar merd i sjø | 1026 | 18 | 57 | 24 |
| Februar slakteri | 42 | 24 | 74 | 2 |
| Mars merd i sjø | 607 | 16 | 60 | 23 |
| Mars slakteri | 150 | 10 | 56 | 34 |
| April merd i sjø | 270 | 19 | 63 | 18 |
| April slakteri | 150 | 15 | 47 | 37 |
| Mai merd i sjø | 40 | 33 | 33 | 35 |
| Mai slakteri | 50 | 6 | 60 | 34 |
| Juni merd i sjø | 20 | 15 | 70 | 15 |
| Juni slakteri | 0 | IK | IK | IK |
| Juli merd i sjø | 0 | IK | IK | IK |
| Juli slakteri | 50 | 16 | 44 | 40 |
| Totalt | 4284 | 20 | 61 | 19 |
| Totalt for merd i sjø | 3592 | 22 | 61 | 16 |
| Totalt for slakteri | 692 | 11 | 59 | 30 |
| a. Merd 2023-2025 | |||||||||
| Måned | Antall prøver F (N) | Umoden F | Modnende F | Gytende/ Utgytt F | Modnende + gytende/utgytt F | ||||
| % | N | % | N | % | N | % | N | ||
| Desember i sjø | 373 | 34 | 126 | 66 | 245 | 1 | 2 | 66 | 247 |
| Januar merd i sjø | 529 | 33 | 177 | 61 | 321 | 6 | 31 | 67 | 352 |
| Februar merd i sjø | 655 | 22 | 143 | 67 | 438 | 11 | 74 | 78 | 512 |
| Mars merd i sjø | 506 | 19 | 96 | 74 | 372 | 8 | 38 | 81 | 410 |
| April merd i sjø | 411 | 13 | 54 | 81 | 331 | 6 | 26 | 87 | 357 |
| Mai merd i sjø | 148 | 11 | 16 | 78 | 116 | 11 | 16 | 89 | 132 |
| Juni merd i sjø | 124 | 5 | 6 | 71 | 88 | 24 | 30 | 95 | 118 |
| Juli merd i sjø | 12 | 0 | 0 | 83 | 10 | 17 | 2 | 100 | 12 |
| SUM: | 2758 | 618 | 1921 | 219 | 2140 | ||||
| Andel av total (F): | 22 | 70 | 8 | 78 | |||||
| b. Slakteri 2023-2025 | |||||||||
| Måned | Antall prøver F (N) | Umoden F | Modnende F | Gytende/ Utgytt F | Modnende + gytende/utgytt F | ||||
| % | N | % | N | % | N | % | N | ||
| Desember slakteri | 66 | 9 | 6 | 89 | 59 | 2 | 1 | 91 | 60 |
| Januar slakteri | 187 | 18 | 33 | 66 | 124 | 16 | 30 | 82 | 154 |
| Februar slakteri | 110 | 11 | 12 | 69 | 76 | 20 | 22 | 89 | 98 |
| Mars slakteri | 187 | 9 | 16 | 68 | 128 | 23 | 43 | 91 | 171 |
| April slakteri | 137 | 15 | 21 | 64 | 88 | 20 | 28 | 85 | 116 |
| Mai slakteri | 232 | 3 | 8 | 79 | 183 | 18 | 41 | 97 | 224 |
| Juni slakteri | 40 | 10 | 4 | 63 | 25 | 28 | 11 | 90 | 36 |
| Juli slakteri | 52 | 42 | 22 | 42 | 22 | 15 | 8 | 58 | 30 |
| SUM: | 1011 | 122 | 681 | 184 | 889 | ||||
| Andel av total (F): | 12 | 70 | 18 | 88 | |||||
Det er usikkerhet knyttet til andel torsk som modner og gyter i oppdrettsanleggene. Data i tabell 2.3 omfatter perioden desember til juli fra i alt 2758 hunnfisk fra merder og 1011 hunnfisk tatt ved slakting både i 2023-2024 (4 oppdrettere og 4 lokaliteter) og 2024-2025 (2 oppdrettere og 4 lokaliteter) (Tabell 2.3a, b). Det er en lavere andel torsk som er gytende/utgytt i merdene, og data ved slakting representerer et endepunkt for oppdrettstorsken. Gytende og utgytt torsk har allerede bidratt til gyting i merd når de når dette endepunktet. Den samlende andelen modnende og gytende/utgytt hunntorsk for disse to årene var 78 og 88 % for henholdsvis merder og slakteri (Tabell 2.3a, b), der gytende/utgytt av dette utgjorde 10 % i merder og 21 % ved slakteri. Det var liten forskjell mellom 2024 og 2025, men med utgangspunkt i innføring av forvaltningstiltak er det knyttet usikkerhet til om andelen modnende og gytende/utgytt de to siste årene vil vedvare fremover.
2.3.1.4 - Overlevelse av avkom gytt i merd til gytende individer
Egg og larvedrift fra gyting hos torsk i merd vil mest sannsynlig være sammenlignbart med drift av gyteprodukter fra vill torsk ved lignende miljøforhold. Lengden på eggstadiet vil variere med temperaturen, fra ca. 14 dager på Vestlandet og opp til 3 uker i mars i Nord-Norge på grunn av lave temperaturer (Myksvoll mfl. 2014; Westernhagen 1970). Gyting utenfor sesong i den varme årstiden, for eksempel ved bruk av lys for å utsette kjønnsmodning, kan redusere utviklingstiden på eggene. Liten variasjon i eggenes flyteevne i interaksjon med variasjonen i saltholdighet i vann gjør at det er hovedsakelig det lokale miljøet som bestemmer vertikalfordelingen av egg (Myksvoll mfl. 2011). Egg kan også ha relativt stor variasjon i spesifikk oppdrift og derved ha en spredning i vertikalfordeling (Jung mfl. 2012). Kysttorsken har ofte egg som ligger under overflatelaget og derfor spres mindre (Ciannelli mfl. 2010; Espeland mfl. 2015).
Lokalisering av anlegg inne i en fjord er av stor betydning for spredning av egg (Uglem mfl. 2012). Mange av gytefeltene til kysttorsk ligger innerst i fjordarmene, der er det ofte litt lavere saltholdighet, og eggene er fordelt litt dypere i vannsøylen. Den dypere fordelingen og de lokale strømforholdene gjør at eggene kan få høy retensjon i området de er gytt, altså at potensialet for spredning er begrenset (Myksvoll mfl. 2011). Spredningsmodellering har vist at torskeegg har høyest retensjon i fjordområder (> 90 %), moderat retensjon i kystområder innenfor grunnlinjen (rundt 50 %) og ingen retensjon utenfor grunnlinjen (Myksvoll mfl. 2014). Plasseringen av anlegg på strømsterke lokaliteter, som for lakseoppdrett, vil medføre en betydelig større spredning enn om anlegget plasseres i skjermede fjordarmer, tilsvarende der hvor gytefelt for kysttorsk ofte finnes. Gytefelt som ligger ute på kysten har større potensiale for spredning fordi eggene kan flyte helt i overflaten der strømmen er sterkest på grunn av vindpådraget. Der kontinentalsokkelen er smal, kommer kyststrømmen tett på kysten og bidrar til betydelig transport nordover.
Torskelarver lever fritt i vannmassene og vil også være påvirket av strømforholdene. Larvene har imidlertid egenbevegelse, både vertikalt og horisontalt, og egenbevegelsen øker med larvenes alder. Som for torskeegg vil torskelarver også befinne seg i de øvre vannlagene (Jørstad mfl. 2008). Den pelagiske fasen frem til det tidlige yngelstadiet varer til om lag 40-50 mm størrelse da yngelen vil søke mot bunnen på grunt vann (Jørstad mfl. 2014). Tiden det tar for å nå denne størrelsen vil variere med temperatur og matforhold, med det kan ta opp mot to måneder etter klekking. Det er vist at hyselarver og sildelarver kan ha en retningsbestemt bevegelse (Cresci mfl. 2019; 2020), men det er ukjent om torskelarver har samme egenskap. Torskelarver har så langt vist seg å tiltrekkes lavfrekvent lyd (Cresci mfl. 2023). Egg og larver spres på samme måte som lakselus de første ukene før egenbevegelsen gjør seg gjeldende, og kan derfor spres flere titalls kilometer også innenfor en fjord (Asplin mfl. 2014).
Simulering av spredning er nå allment tilgjengelig gjennom «Strømkatalogen». Dette on-line verktøyet simulerer spredning av partikler i en oppløsning med ruter på 800 x 800 m. Denne oppløsningen kan være noe grov for trange fjorder, men den vil kunne gi en pekepinn på hvor omfattende spredningen kan være. For en bedre oppløsning kan for eksempel en modell tilsvarende NorKyst-modellen med 160 x 160 m rutenett benyttes, som også brukes for å simulere spredning av lakselus.
En annen faktor som gir usikkerhet og kan påvirke overlevelse av avkom fra gyting i merd, er plassering av oppdrettsanlegg med hensyn til gunstige eller ugunstige forhold for avkommet (habitatkvalitet, mat- og predatorforhold, drift gjennom egg- og larvefasen til oppvekstområder for yngelen, osv.). Det samme gjelder for gyting og klekking utenom normal gytetid grunnet bruk av lys for å utsette kjønnsmodning. I 2025 er det verifisert at vill torsk i Troms kan gyte om sommeren, faktisk så seint som i slutten av juli (prosjekt «Sommer- og høstgyting hos torsk», Dugnad for havet). Dette, sammen med en vurdering av byttedyrforhold ved gyting utenom normal gytetid om våren, indikerer at overlevelse av larver til yngel kan finne sted (van der Meeren mfl. 2024). Usikkerheten ligger imidlertid i å kvantifisere denne overlevelsen.
Videre overlevelse i yngelstadiet forutsetter at torskelarvene spres til gunstige oppvekstområder. Som beskrevet i tidligere avsnitt er disse områdene mangelfullt kartlagt. Grunne områder med bunnvegetasjon antas å være særlig egnede oppvekstområder, men større forekomster av torskeyngel er også funnet på mer «naken» grunn bunn bestående av sand, grus eller stein, gjerne med forekomst av ulike rødalger (Dunlop mfl. 2022). Ved gyting i merd i indre fjordområder er det større sannsynlighet for at larver spres til lokale oppvekstområder inne i fjordene, sammenlignet med gyting i ytre fjordområder. Dersom gyting i merd skjer i nærheten av naturlige gytefelt, er det stor sannsynlighet for at larver av villtorsk og oppdrettstorsk spres til samme sted.
Relativ forskjell i overlevelse av egg fra oppdrettet og vill torsk har også usikkerhet knyttet til seg og vil være viktig i vurdering av risiko ved gyting i merd. Forskjeller i befruktning, feilutvikling av embryoene i eggene og klekkesuksess kan ha ulike årsaker, blant annet temperatur under utviklingen av rogna i morfisken og om morfisken er førstegangsgyter (van der Meeren & Ivannikov 2006; Kjesbu mfl. 1996). Også morfiskens ernæringsmessige status vil spille en rolle for eggkvalitet hos torsk (Pickova mfl. 1997; Salze mfl. 2005; Røjbek mfl. 2014; Norberg mfl. 2017). Torsk som modner i merder, vil være førstegangsgytere men vil også produsere flere egg (ha høyere fekunditet) enn vill torsk (van der Meeren & Ivannikov 2006; Thorsen mfl. 2010; upublisert data fra Havforskningsinstituttet for kysttorsk fra Andenes-Myre serien mars 1986-2025). Hos vill torsk bidrar flergangsgytere i større grad, og flergangsgytere kan ha større suksess med hensyn til overlevelse av avkom (Kjesbu mfl. 1996). Egg samlet inn under gytesesongen i Lofoten viser imidlertid at egg fra gyting hos vill torsk kan ha en betydelig grad av feilutvikling (Makhotin mfl. 2001). I et gyteforsøk med vill og oppdrettet torsk er det vist at forskjeller i befruktning og feilutvikling i gjennomsnitt ga 29 % overlevelse av egg til klekking hos oppdrettstorsk og 76 % overlevelse hos egg av vill torsk (Havforskningsinstituttet, upubliserte data fra MOTOR-prosjektet). I et nylig startfôringsforsøk ble det ikke funnet forskjeller i overlevelse til yngelstadiet mellom larver fra oppdrettstorsk og vill torsk som begge ble startfôret på naturlig plankton (Havforskningsinstituttet, upubliserte data fra MOTOR-prosjektet). Dette utelukker ikke at forskjeller i overlevelse knyttet til genetikk eller atferd kan eksistere når oppdrettsfisk settes ut i naturen.
Hva er kjent om overlevelse av torsk i tidlige livsstadier som settes ut i sjøen? I et forsøk med utsetting av en større mengde genetisk merkede plommesekklarver i Heimarkspollen i Austevoll ble effekten av utsettingen frem til ett år gammel yngel vurdert å være liten (Kristiansen mfl. 1997). I PUSH-programmet på 1990-tallet ble det observert betydelige forskjeller i overlevelse mellom utsatt pollprodusert yngel av torsk og vill torsk av samme størrelse, selv om dietten etter få uker endret seg til den samme som hos vill torsk (Svåsand mfl. 2000). I disse forsøkene ble stamfisken ikke vurdert å være mye forskjellig fra vill torsk i områdene for utsetting. Overlevelsen av utsatt torsk i PUSH-programmet varierte mye, med gjenfangster fra 0 til 30 % avhengig av område, tidspunkt og fiskestørrelse ved utsetting. Forskjellene i gjenfangst mellom utsettingsområdene ble forklart med forskjeller i naturlig dødelighet, størrelses-selektivt fiske og fiskeintensitet. Overlevelsen kan ha vært bedre enn gjenfangstdata tilsier, da manglende inkludering av ikke-rapportert gjenfangst og merketap kan ha underestimert gjenfangsten med 1,5-2 ganger (Svåsand mfl. 2000). Overlevelse av oppdrettstorsk i alle livsstadier fra larve til voksen kjønnsmoden fisk ble også undersøkt under forrige runde med torskeoppdrett (2006-2012) i et forsøk med gyting i merd i Heimarkspollen i Austevoll. Som presentert i kapitlet ovenfor om gyting i merd, ble det her funnet et tydelig signal på overlevelse frem til kjønnsmoden fisk med påfølgende gyting (van der Meeren mfl. 2012).
2.3.2 - Endring i adferd hos kysttorsk
Oppdrettsanlegg fungerer som innretninger for samling av fisk ved at de fysiske strukturene tilbyr ly og habitat for ulike organismer, og ved økt tilgjengelighet av mat, hovedsakelig gjennom spillfôr (Sanchez-Jerez mfl. 2011; Uglem mfl. 2014; Callier mfl. 2018; Skjæraasen mfl. 2022; Meier mfl. 2023). Hvis et betydelig antall fisk fra den lokale bestanden samler seg rundt oppdrettsanlegg, kan det fungere som en økologisk felle. Økologiske feller oppstår når et kunstig habitat introduseres i det naturlige miljøet, tiltrekker seg fisk, og effekten av dette leder til negative økologiske konsekvenser for fisken (Dempster mfl. 2011). Fisk kan foretrekke et kunstig habitat fremfor et naturlig om dette habitatet fremstår attraktivt for fisken, men som i realiteten er et dårligere habitat med lav kvalitet som kan lede til dårligere reproduksjon eller overlevelse (Battin 2004).
I en metaanalyse om miljøpåvirkningene fra havbruk viste Barret mfl. (2019) økt tetthet og mangfold av dyreliv rundt oppdrettsanlegg, hovedsakelig drevet at økte forekomster av fisk. I Norge er det vist at fisk samler seg rundt oppdrettsanlegg. Dette gjelder spesielt for sei, men også andre arter som torsk, hyse og makrell er påvist å samle seg rundt laksemerder (Dempster mfl. 2009; 2010; Dunlop mfl. 2024). Vill fisk, inkludert torsk og sei, kan oppholde seg svært nært oppdrettsanlegg over forholdsvis lange perioder og vandre raskt og ofte mellom flere anlegg, samt til andre nærliggende habitater som for eksempel gytefelt eller tradisjonelle fiskefelt (Uglem mfl. 2008; 2009; Dempster mfl. 2010; Otterå & Skilbrei 2014; Sanchez mfl. 2011; Skjæraasen mfl. 2022). Videoundersøkelser har vist at oppdrettsanlegg for laks kan ha mengder av vill fisk som tilsvarer 1 til 3 størrelsesordener mer enn i kontrollområder uten anlegg (Dempster mfl. 2009), og et oppdrettsanlegg kan derfor betraktes som en «fiskeaggregeringsenhet» (FAD) (Sanchez-Jerez mfl. 2011). Tettheten av fisk som samler seg rundt oppdrettsanlegg kan henge sammen med fôrintensitet (Ballester-Moltó mfl. 2015) eller fôringstider (Uglem mfl. 2009). Torsk beiter på organismer og annen mindre fisk som samler seg rundt oppdrettsanlegg (Dempster mfl. 2011; Sæther mfl. 2012; Serra-Llinares mfl. 2013). Videre er det vist at fisk som oppholdt seg ved oppdrettsanlegg for laks, var både større og tyngre enn fisk fra områder uten oppdrett (Barett mfl. 2019). Torsk som spiser laksefôr, vil blant annet få en større lever som følge av overskudd av fett (Meier mfl. 2023). Endring i mangfold og mengde av arter kan ha konsekvenser for økosystemets funksjon (Gamfeldt mfl. 2015), men det er usikkert om dette kan skje ved etablering og drift av oppdrettsanlegg.
Habitatvalg hos torsk i fjorder med torskeoppdrett («torskehoteller») har også blitt undersøkt, og det er bekreftet at det er en forhøyet tetthet av torsk rundt slike oppdrettsanlegg, og at enkelte individer er svært stasjonære rundt anleggene (Bjørn mfl. 2007; Serra Llinnares mfl. 2013; Hedger mfl. 2017). I en studie fra Balsfjorden ble vill torsk fanget ved et oppdrettsanlegg merket med akustiske sendere både høst og vinter og gjenutsatt (Uglem mfl. 2008). Adferden til disse ble deretter studert både før, under og etter gytesesongen. Resultatene viste at en høy andel (ca. 50%) av villtorsken forble ved anlegget utover høsten, og de benyttet nærområdet til anlegget mer enn simulert rømt oppdrettstorsk som ble satt ut samtidig med villtorsken. Om vinteren var den ville torsken mer på gytefeltene, spesielt det indre gytefeltet i fjorden, og dette kan indikere en stasjonær torsk «hjemmehørende» i indre fjord, og som benyttet anlegget som en ressurs gjennom året. Den «rømte» oppdrettstorsken spredte seg over store områder, men ble også funnet på lokale gytefelt (Uglem mfl. 2008).
Kunstig belysning brukes i både i torske- og lakseoppdrett for å hemme kjønnsmodning og for å øke somatisk vekst om våren (Hansen mfl. 2017). Flere virvelløse dyr tiltrekkes av kunstig belysning, og det er observert forhøyede mengder virvelløse dyr rundt oppdrettsanlegg (Fernandez-Jover mfl. 2016; McConnell mfl. 2010). Krill er et viktig byttedyr for torsk, og krill vil svømme mot lys (Humborstad mfl. 2018). Ansamlinger av krill kan derfor tiltrekke torsk til oppdrettsanlegg som benytter kunstig lys for å forhindre kjønnsmodning. Det har blitt funnet krill i mageprøver av både makrell og hyse ved lakseanlegg (Svåsand mfl. 2017).
Tidligere har det blitt beskrevet av fiskere at torsk unngår områder med aktive oppdrettsanlegg (Maurstad mfl. 2007). Det er også vist i laboratorieeksperimenter at villfanget torsk unngikk vanntanker med vann fra lakseoppdrettsanlegg, men ettersom en lignende respons også var til stede når tankene ble tilsatt vann fra tanker med domestisert torsk, er det det sannsynlig at denne adferden er en respons på høye tettheter av fisk og ikke lakseoppdrett (Sæther mfl. 2007). Videre har eksperimentelle adferdsundersøkelser vist at villtorsk med blokkert luktesans har lik adferd og områdebruk som torsk med upåvirket luktesans i områder med mye lakseoppdrett (Bjørn mfl. 2009). Som følge av dette vurderes det som lite sannsynlig at kysttorsk konsekvent unngår områder med akvakulturvirksomhet, noe som settes som et premiss for vårt valg av de antatt viktigste risikofaktorene, hendelsene og konsekvensene knyttet til negative effekter av lakseoppdrett på kysttorsk som presenteres her.
Maskiner og rutinemessige operasjoner i lakseoppdrett som fartøy, fôringsutstyr, høytrykksspylere, aggregater, m.m., lager støy som kan forstyrre livet for dyr i miljøet rundt. Adferd fra marine dyr som en respons på menneskeskapte støy inkluderer fluktreaksjoner, unngåelse eller endringer i svømmeadferd (Olesiuk mfl. 2010). Andre effekter av miljøstøy er rapportert, som lydskader (Casper mfl. 2013) og en signifikant økning av stressrelaterte blodparametere (Buscaino mfl. 2010). Torsk kommuniserer med "grynt" under parring, aggresjon eller flukt (Brawn, 1961). Hunntorsk foretrekker hanner med større kropper og lange finner, men gytesuksess er også korrelert med volum på «gryntene» fra hannfisken (Rowe and Hutchings, 2008). Karforsøk med torsk utsatt for langvarig støy har blitt rapportert å ha en lavere befruktningsgrad, opptil 40% reduksjon, enn ikke-eksponert fisk. Fisk utsatt for kortvarig støy har også vist en økning i blodkortisol (Sierra-Flores mfl. 2015). Resultatene av denne studien viser at støygenererende havbruksaktiviteter kan påvirke reproduksjon, både når det gjelder forstyrrelse av naturlig kommunikasjon og de negative effektene av stress. En studie fra oppdrettsintensive og oppdrettsekstensive områder på Vestlandet fant imidlertid små forskjeller i faktorer knyttet til reproduksjon hos vill torsk (Barrett mfl. 2018).
Internasjonalt er det relativt liten kunnskap om unnvikelse av fisk fra oppdrettsanlegg, eller eventuell påvirkning på vandringsmønsteret (Callier mfl. 2018; Barrett mfl. 2019), og det har vært lite eller ingen ny forskning på dette de siste 10-15 årene. I Norge har fiskere imidlertid lenge hevdet at «innsigstorsk» på gytevandring, ikke vandret inn til gytefelt i oppdrettsintensive fjorder. Fiskernes tradisjonelle økologiske kunnskap er studert gjennom intervjuundersøkelser og samfunnsvitenskapelige metoder (Maurstad mfl. 2007). Konklusjonen er at fiskerne har gjort pålitelige observasjoner av adferd til torsk i oppdrettsintensive områder, både unnvikelse og tiltrekning, men at det trengs naturvitenskaplige studier for å konkludere med større sikkerhet, samt foreslå mekanismer bak eventuelle endringer (Maurstad mfl. 2007).
Gjentatte eksperimentelle studier i laboratoriet, hvor vill kysttorsk gis et valg mellom å oppholde seg i eller unngå kar iblandet vann fra et kar med oppdrettslaks, bekrefter at villfanget antatt oppdrettsnaiv kysttorsk unngår å oppholde seg i kar som tilføres en svært liten (2,5 ‰) andel vann fra et kar med laks (Sæther mfl. 2007). Det er luktstoffer i vannet torsken reagerer på, ikke endringer i vannkvalitet som sådan, da fisk med blokkert luktesans ikke viste denne adferden. Luktesansen er svært viktig for fiskens evne til å orientere seg i miljøet, blant annet for å finne mat, orientere seg om gode/dårlige habitater, unngå predatorer, kommunikasjon med artsfrender etc. (f.eks. Hara 1994; Sorensen & Caprio 1998). Oppfølgende eksperimenter viser at fisk fanget nær eller under oppdrettsanlegg, som antas å være stasjonær kysttorsk, også unngår vann fra kar med laks, men i mindre grad enn antatt oppdrettsnaiv vandrende kysttorsk.
Eksponering for lukt fra annen torsk gav tilsvarende adferdsendring hos villtorsken; den tilbrakte mindre tid i karet hvor lukten ble tilsatt. Det kan dermed synes som om de observerte adferdsendringene ikke er knyttet til lukten av noen spesiell art, men snarere generelle komponenter som akkumuleres i vannet når store tettheter av fisk oppholder seg der. Første generasjon oppdrettstorsk endrer imidlertid ikke adferd når den eksponeres for vann fra laksekaret, noe som kan skyldes at disse er tilvent høye konsentrasjoner av komponenten(e) som villtorsken reagerer på (Sæther mfl. 2007). Det eksperimentelle designet tillater imidlertid ikke at man konkluderer med at lukt av annen fisk i en fjord kan føre til endret adferd hos torsk som har naturlig tilhørighet der, eksempelvis ved at vandrende kysttorsk på gytevandring stanser og snur før de kommer inn til gytefeltet, fordi kompleksiteten i problemstillingen er alt for stor til at dette kan gjenskapes i laboratoriet, og det kan også tenkes at oppsettet er «for sensitivt» ved at fisken ikke måtte velge bort noe (f.eks. beskyttelse eller mat) dersom den unngikk kar med lukt fra laks. Resultatene fra stasjonær torsk kan tyde på dette, ettersom denne fisken var fanget ved oppdrettsanlegg, og som frittlevende valgte å oppholde seg der, men likevel unngikk kar med lukt av laks i eksperimentet.
Vandringsmønsteret til innvandrende kysttorsk («innsigsfisk») under naturlige forhold har derfor også blitt undersøkt i en storskala telemetristudie (Bjørn mfl. 2009). Torsken ble fanget i midtveis inn i yte del av Øksfjorden, en oppdrettsintensiv fjord med et tradisjonelt gytefelt (Maurstad mfl. 2007; Fiskeridirektoratet.no) innerst i fjorden. Halvparten av fisken ble luktblokkert, og satt ut på fangststedet mens resten ble transportert inn til gytefeltet og i oppdrettsintensive områder. All fisk, foruten en, som ble fanget og gjenutsatt i ytre del av Øksfjorden, snur og forlater fjorden innen en uke etter utsett. Når de derimot fysisk flyttes inn til de oppdrettsintensive indre områdene av fjorden oppholder majoriteten av torsken seg der inne i ca. 3 uker, de synes ikke avskrekket av anlegg, og det kan heller ikke utelukkes at de gyter i fjorden. Det var ingen sammenheng mellom luktesans og vandringsadferd, ettersom luktblokkert fisk viste samme adferd som fisken med intakt luktesans (Bjørn mfl. 2009). Feltstudien frambrakte med andre ord ingen bevis på at vandrende torsk unngår oppdrettsintensive områder som en følge av luktstoffer, men resultatene strider heller ikke imot fiskernes observasjoner om at torsken ikke lenger går inn til gytefeltet i fjorden. I en studie fra Smøla ble det heller ikke påvist at oppdrettsanlegg for laks hindret torsk i å bruke nærliggende gytefelt (Skjæraasen mfl. 2021).
Våre resultater viser dermed at laboratorie- og feltstudier av den typen som hittil er gjennomført, ikke vil være tilstrekkelig for å kunne vise om intensiv oppdrettsaktivitet (først og fremst lakseoppdrett, men det kan også gjelde torskeoppdrett) faktisk fører til at vill innsigstorsk skyr tradisjonelle gyteplasser. Dersom vi antar at laboratorieresultatene er valide (jamfør sensibilitet), viser disse at antatt oppdrettsnaiv vandrende kysttorsk responderer klart negativt på lukt fra høye tettheter av fisk. Denne responsen er svakere (men fortsatt signifikant) hos antatt stasjonær kysttorsk fra områder med oppdrett, og helt fraværende hos oppdrettstorsk (Sæther mfl. 2007).
Samtidig viser telemetristudien (Bjørn mfl. 2009) og de andre feltresultatene (Bjørn mfl. 2007) at det i all hovedsak er stasjonær fjordtorsk som aggregeres rundt oppdrettsanlegg. Det kan derfor tenkes at det kan være to helt forskjellige responser hos torsk på samme stimuli; noen kan tilvennes og tiltrekkes (stasjonær fjordtorsk) mens andre kan frastøtes (vandrende kysttorsk på gytevandring). Dette er også i overensstemmelse med fiskernes observasjoner om at det er «innsigsfisken» som unngår fjorder med oppdrett (Maurstad mfl. 2007). Samlet sett kan det derfor fortsatt ikke utelukkes at tilstedeværelse av annen fisk kan bidra til å forklare endringene i kysttorskens tilstedeværelse som fiskerne observerer. Lyd, lys eller andre faktorer fra industrielt oppdrett kan også påvirke adferd, men det kan heller ikke utelukkes at torsk ikke reagerer negativt på intensiv oppdrettsaktivitet. Skjæraasen mfl. (2022) fant at noen individer av torsk ser ut til å ha lang oppholdstid ved oppdrettsanlegg (måneder), mens andre individer har kort oppholdstid (dager). I hvilken grad adferd på individnivå (dvs. torskens “personlighet”) er bestemmende for tiltrekking eller unnvikelse av oppdrettsanlegg, er uklart, men slike forskjeller i individuell adferd kan tenkes å forekomme både blant vandrende og stasjonær kysttorsk. Derfor vil langsiktige feltstudier med individmerking og genetisk karakterisering (før og etter etablering av oppdrett og i fjorder med og uten oppdrett, slik som Icod og SalCod prosjektene, antakeligvis være en mer treffende metodikk for å forsøke å dokumentere om intensivt fiskeoppdrett kan påvirke adferd, gyting og populasjoner hos vill torsk.
2.3.3 - Endringer i fysiologi hos kysttorsk
Torsk er en generalist som endrer diett gjennom livsløpet. Generelt sett består dietten til torsk under 20 cm på grunt vann i kystsonen hovedsakelig av ulike krepsdyr og små fisk (Arnott & Pihl 2000) Tilsvarende størrelse torsk hos de havgående bestandene spiser krill og andre små pelagiske byttedyr, og med større innslag av bunndyr og større byttefisk med økende størrelse (Link & Garrison 2002; Holt mfl. 2019). Videre varierer også torskens diett som en funksjon av byttedyrtetthet (Link & Garrison 2002). Denne fleksibiliteten kommer tydelig frem i diettundersøkelser i relasjon til nærhet til oppdrettsanlegg. Torsk tilknyttet laksemerder spiser mer spillfôr, fisk og flerbørstemark enn torsk fanget lengre bort fra anleggene (Dempster mfl. 2011; Fernandez-Jover mfl. 2011). Studier av dietten til torsk fanget i nærheten av 9 oppdrettsanlegg i viste at 20 % av torsken hadde pellet i magesekken, mens torsk fanget på kontrollokaliteter ikke hadde pellet i magesekken (Dempster mfl. 2011). Sæther mfl. (2012) viste også at torsk av ulik størrelse varierte med hensyn til hvor mange som hadde pellets i mageinnholdet. Andelen av torsk som var mindre (n=34) eller større (n=46) enn 60 cm som hadde spist pellets, var henholdsvis 32 % og 11 % (Sæther mfl. 2012). Lignende resultater er også vist av Keeley mfl. (2025), hvor forfatterne fant at 69 % av torsk fanget nært (<100 m) laksemerdene i Langfjorden i Vest-Finnmark hadde en fettsyresammensetning i leveren som viser at dietten er dominert av laksefôr. Mageundersøkelser fant i tillegg betydelig mengder av opportunistiske sediment-levende bunndyr som flerbørstemark, slangestjerner og muslinger som forbindes med typisk fauna under oppdrettsanlegg. Dette studiet fant at det kun var torsk som ble fanget helt nær oppdrettsanleggene, som hadde spesialisert seg til å beite på slike byttedyr, så det tyder på at noen fisk tilvenner seg mens andre ikke gjør det. Sanchez-Jeres mfl. (2025) viste at både kysttorsk og skrei har sterke signaler av spillfôr fra lakseoppdrett i sin fettsyresammensetning, men at de potensielt negative fysiologiske effektene av endringen vil kreve nye og mer detaljerte studier. Meier mfl. (2023) fant at henholdsvis 13 % (2018) og 20 % (2019) av gytemoden fisk som var fanget på gytefeltene rundt Smøla på Nordmøre, hadde spesialisert seg på beiting på oppdrettsfôr. Sammensetningen av fôret som brukes i norsk akvakultur av laksefisk, har endret seg markant over tid. Andelen protein har sunket og fettinnholdet har økt, andelen marine ingredienser har sunket fra 90 til 25 % i perioden fra 1990 til 2016 og samtidig har andelen plantebaserte ingredienser økt fra nesten 0 til 71 % (Ytrestøyl mfl. 2015; Aas mfl. 2019). Fiskeoljer har primært blitt erstattet av rapsolje (Brassica napus var. napus) som inneholder meget høye nivåer (67 % av total mengde fettsyrer) av de terrestriske fettsyrene oljesyre (18:1n-9), linolsyre (18:2n-6) og α-linolensyre (18:3n-3) (Bell mfl. 2003). Laksefôr inneholder i dag typisk 24 % fett, og 56 % av dette består av terrestriske fettsyrer, mens nivåene av langkjedede flerumettede fettsyrene (LC-PUFA) er betydelig lavere enn marine oljer. Typiske verdier i laksefôr er 0,2 % arakidonsyre (20:4n-6) (ARA), 4 % eikosapentaensyre (20:5n-3, EPA) og 5 % dokosaheksaensyre (22:6n-3, DHA) (Meier mfl. 2023).
Fôret som brukes i torskeoppdrett, har et større innslag av marine ingredienser, og dermed er nivåene av terrestriske fettsyrer lavere (23 %) og marine fettsyrer høyere (0,5 % ARA, 8 % EPA og 11 % DHA) (tabell 2.4). De negative effekter man (kanskje) kan få hos vill torsk som beiter på spillfôr fra fiskeoppdrett, vil derfor kunne forekomme i større grad hos villfisk som beiter på laksefôr enn hos villfisk som beiter på marint-fôr fra torskeoppdrett.
| Laksefôr | Marint fôr | |
| Fettsyremengde (mg/100 mg tørrvekt) | 24 | 21 |
| Fettsyreprofil (% av total fettsyremengde) | ||
| 14:0 | 2,3 | 6,2 |
| 16:0 | 10,9 | 15,8 |
| 18:0 | 2,3 | 2,1 |
| ∑SFA | 16,7 | 26,2 |
| 16:1n-7 | 2,6 | 4,9 |
| 18:1n-9 | 37,2 | 14,0 |
| 18:1n-7 | 2,8 | 2,1 |
| 20:1n-9 | 2,0 | 4,6 |
| 22:1n-11 | 1,6 | 6,3 |
| ∑MUFA | 48,4 | 36,1 |
| 18:2n-6 | 16,5 | 6,8 |
| 20:4n-6 (ARA) | 0,2 | 0,5 |
| 18:3n-3 | 6,2 | 2,0 |
| 18:4n-3 | 3,3 | 3,3 |
| 20:5n-3 (EPA) | 3,9 | 8,3 |
| 22:5n-3 | 0,7 | 1,0 |
| 22:6n-3 (DHA) | 4,8 | 11,2 |
| ∑PUFA | 34,9 | 37,3 |
| ∑PUFA (n-6) | 17,2 | 8,2 |
| ∑PUFA (n-3) | 17,2 | 27,6 |
| PUFA n-6/n-3 ratio | 1,0 | 0,3 |
| ∑terrestriske fettsyrer (%) | 56 | 23 |
SFA = mettede fettsyrer, MUFA = monoumettede fettsyrer, PUFA = polyumettede fettsyrer. ∑terrestriske fettsyrer er summen av18:1n-9, 18:2n-6 og 18:3n-3. Tabellen viser de 15 dominerende fettsyrer av 70 analyserte fettsyrer.
Det at torsk kan samle seg rundt oppdrettsanlegg for å spise spillfôr og byttedyr forbundet med oppdrett, kan forventes å føre til en næringsendring der dietten går fra å bestå av naturlige byttedyr med mye omega-3-fettsyrer til en diett som inkluderer et høyere innhold av terrestriske omega-6-fettsyrer. Flere feltundersøkelser har vist at torsk fanget nært oppdrettsanlegg har høyere nivåer av terrestriske fettsyrer i muskel og lever enn fisk fanget lengre fra laskemerdene (Fernandez-Jover mfl. 2011; Keeley mfl. 2025; Meier mfl. 2023; Sanchez-Jerez mfl. 2024; 2025). I en feltundersøkelse gjort på gytende torsk rundt Smøla ble det også påvist en betydelig endring i fettsyresammensetningen hos gytebestanden. Her utgjorde terrestriske fettsyrer over 50 % av lipidene i leveren for 17 % av gytefisken, mens de resterende individene hadde lipidsammensetninger dominert av naturlige byttedyr (Meier mfl. 2023). Samme studie viste videre at de terrestriske fettsyrene også ble overført til muskel og gonader (rogn og melke), eksemplifisert med en 20 % økning av terrestriske fettsyrer i rogn hos hunnfisk, fra 7 til 27 % (Meier mfl. 2023).
Et pågående prosjekt ledet av HI (MOTOR, Modningsfri torsk i merd, FHF, prosjekt 901815) undersøker om oppdrettstorsk er i stand til å få levedyktig avkom. MOTOR-prosjektet gjennomføres med eksperimentelle studier som skal kartlegge hvordan forhøyede mengder terrestriske fettsyrer fra kommersielt marint fôr påvirker gonademodning, fekunditet og larveutvikling hos torsk, og denne kunnskapen vil være klar i løpet av 2026.
Fettsyreanalyser av leverprøver viser at oppdrettstorsk har forhøyede nivåer av terrestriske fettsyrer (32%) sammenlignet med vill fisk (19 %) som beiter på bunndyr, men dette er betydelig lavere enn det man finner hos vill torsk som har beitet på laksefôr (tabell 2.5). Det gjenstår å analysere fettsyresammensetning hos gonader og gyteprodukter (egg/larver) fra MOTOR-prosjektet, men gyteforsøkene fra MOTOR viste at egg fra oppdrettstorsk er levedyktige. Det er et stort behov for tilsvarende eksperimentelle studier for å undersøke hvordan vill torsk sin beiting på laksefôr (som har en mer ekstrem fettsyre sammensetning) påvirker torsken reproduksjon.
| Vill torsk som beiter på laksefôr | Vill torsk som beiter på bunndyr | Oppdrettstorsk | |
| Fettsyremengde (mg/100 mg lever vekt) | 54 | 22 | 55 |
| Fettsyreprofil (% av total fettsyremengde) | |||
| 14:0 | 1,9 | 2,3 | 2,3 |
| 16:0 | 9,3 | 14,9 | 12,7 |
| 18:0 | 2,3 | 3,5 | 4,2 |
| ∑SFA | 16,7 | 23,3 | 20,8 |
| 16:1n-7 | 3,3 | 5,9 | 4,5 |
| 18:1n-9 | 36,4 | 16,4 | 23,4 |
| 18:1n-7 | 3,9 | 6,4 | 4,1 |
| 20:1n-9 | 3,1 | 2,2 | 4,0 |
| 22:1n-11 | 1,5 | 1,1 | 2,7 |
| ∑MUFA | 51,3 | 37,9 | 43,4 |
| 18:2n-6 | 12,9 | 1,7 | 7,3 |
| 20:4n-6 (ARA) | 0,5 | 3,0 | 0,7 |
| 18:3n-3 | 5,0 | 1,0 | 1,4 |
| 18:4n-3 | 0,8 | 1,3 | 1,3 |
| 20:5n-3 (EPA) | 4,8 | 8,4 | 7,3 |
| 22:5n-3 | 1,1 | 2,7 | 1,3 |
| 22:6n-3 (DHA) | 6,2 | 16,6 | 12,2 |
| ∑PUFA | 34,2 | 38,8 | 35,4 |
| ∑PUFA (n-6) | 14,6 | 7,0 | 9,5 |
| ∑PUFA (n-3) | 18,9 | 38,8 | 24,7 |
| PUFA n-6/n-3 ratio | 0,7 | 0,2 | 0,4 |
| ∑terrestriske fettsyrer (%) | 54 | 19 | 32 |
Per nå er det lite kunnskap om konsekvensene av en slik feilernæring, og de langsiktige effektene av beiting på spillfôr er identifisert som det viktigste kunnskapshullet for hvordan lakseoppdrett kan påvirke ville torskebestander (Bøhn mfl. 2024). Undesøkelser gjort i laboratorier har vist at torsk tolererer en plantebasert diett og kan opprettholde vekst så lenge fisken får nok marine omega-3 fettsyrer (Hansen & Hemre 2013). Dette støttes av feltstudier som har vist at torsk som beiter ved oppdrettsanlegg, kan ha bedre kondisjon og et vesentlig større energireservoar i form av leverstørrelse enn annen torsk (Dempster mfl. 2011; Fernandez-Jover mfl. 2011; Meier mfl. 2023). Til tross for dette representerer store mengder mat av avvikende kvalitet og fettsyresammensetning en mulig risiko, ettersom det er svært lite kunnskap om hva langtidseffektene av en slik endring vil være, både på individ- og bestandsnivå. Av spesiell interesse er hvordan en overgang til en diett dominert av terrestriske fettsyrer påvirker reproduksjon. Selv om økt mattilgang og kondisjon isolert sett er positivt for gonadevekst hos torsk (Lambert & Dutil, 1997, 2000; Marshall mfl. 1999; Wroblewski mfl. 1999; Ouellet mfl. 2001; Skjæraasen mfl. 2010), har flere studier vist at både fekunditet og larveoverlevelse avhenger av tilgang til enkelte langkjedede marine fettsyrer (Pickova mfl. 1997; Izquierdo mfl. 2001; Salze mfl. 2005; Røjbek mfl. 2014; Norberg mfl. 2017).
2.3.4 - Bruk av kunstig belysning for å utsette kjønnsmodning
Lysstyring brukes for å kontrollere og å utsette kjønnsmodning hos oppdrettsfisk. I oppdrett av torsk benyttes en sterkere lyskilde enn i produksjonen av laks. Typen lys (lyskilde) og antall varierer med om det benyttes i overflaten eller som del av nedsenkbar teknologi. For best mulig effekt er det viktig med så jevnt lys i merden som mulig for å sikre jevnt og tilstrekkelig med lys all fisken.
Lys har en effektiv tiltrekningskraft på mange dyr i havet og benyttes ofte i forbindelse med fiskeriaktivitet. Det er derfor nærliggende å tenke seg at ulike organismer vil tiltrekkes av oppdrettsanlegg som følge av lysbruken. Dette kan være ulike plankton, inkludert krill, og fisk av ulik størrelse som igjen tiltrekker seg større predatorer som for eksempel sel. Økt tilstedeværelse av villfisk rundt oppdrettsmerder, kan føre til økt interagering mellom oppdretts- og villfisken. Det vil være særlig i den mørke årstiden at kunstig belysning vil ha størst effekt på livet utenfor merden.
Det er store variasjoner i hvilke lysstyringsregimer som benyttes i oppdrettsnæringen generelt, og også innenfor torskeoppdrett. Her følger beskrivelse av et eksempel på lysstyring som benyttes av Ode AS (pers komm. Falk D. Øveraas): Det benyttes kontinuerlig lys der lyskildene typisk er satt ut på 12 og 25m dyp for å sikre at lyset fordeles så jevnt som mulig i hele vannsøylen. Etter montering av lysoppheng gjennomføres lux-målinger i not på flere dyp og radiale posisjoner for å verifisere at det ikke finnes «mørke» soner hvor fisk kan samle seg, og for å kunne justere plassering og effekt ved behov. Et vanlig oppsett i en 120 m merd vil være bruk av undervanslamper med enten 28,4 eller 24,2 kW pr. merd. Lyskilden er et oppsett med ledlyslamper med effekt på 600 eller 2500W, og oppsettet tar utgangspunkt i en studie utført av Dahl & Bjørkevoll (2023). Det benyttes grønt lys da dette har god, dokumentert gjennomtrengningsevne i sjøvann (30-50 m) (Apel 1988). Lyskildene plasseres i en avstand fra notveggen som minimerer sjansen for kontakt mellom not og lyskilde. Dette gjør også at lysintensiteten er svakere nær notveggene (ytterkanten) av merden enn i senter. Dybden under en merd er stort sett så dyp at lyset ikke trenger gjennom ned til bunnen, og det antas at bunnfauna ikke påvirkes i særlig grad av dette lyset.
Nedsenkede merder til 20 m dyp i kombinasjon med kontinuerlig lys vil utsette kjønnsmodningen med 3 til 6 måneder, og det er mulig at kjønnsmodning kan utsettes helt ved nedsenking til større dyp og i kombinasjon med skyggenett (Korsøen mfl. 2013). «Taket» på en typisk nedsenkbar merd til torsk vil være på ca. 36 m dyp, mens bunnen vil være på ca. 65 m dybde.
2.3.5 - Økt sykdomsforekomst hos kysttorsk som følge av smitte fra oppdrettstorsk
Smittespredning vil påvirkes av mange ulike miljøforhold (f.eks. temperatur, saltholdighet og strøm), og med type smitte (agens, smittestoff). Antallet og størrelsen på utbrudd (mengde syk fisk som skiller ut smittestoff over tid) vil direkte påvirke mengden smittestoff som slippes ut i miljøet, og dermed smittepresset mottakelig fisk kan være utsatt for. Smittespredning med vannmassene vil blant annet være avhengig av strømforhold, overlevelsen til agens i miljøet, og avstand til kilden (fortynningseffekt). En annen viktig forutsetning for smittespredning er overlapp i tid og rom mellom en mottakelig vert, altså vill torsk, og høy nok smittedose. Et eksempel kan være smitte til torskeyngel som er stasjonære i de lokale oppvekstområdene over tid. Torsk og annen marin fisk tiltrekkes av oppdrettsanlegg der de spiser spillfôr, eller andre organismer som beiter på spillfôr (evertebrater). Slike ansamlinger av fisk nært oppdrettsanlegg øker sannsynligheten for smittespredning fra oppdrettstorsk til villtorsk. Det er viktig å merke seg at forskjellige agens vil kunne ha ulik påvirkning på ulike livsstadier hos villtorsk. Det vil også være nærliggende å se for seg at forskjellige sykdommer vil være mer fremtredende på gitte tider av året (for eksempel temperaturavhengige). Overlevelse av et agens i miljøet er en av nøkkelfaktorene for spredningspotensialet, og dette varierer mellom ulike agens. Smitte fra oppdrettet torsk til villtorsk kan skje direkte, men da det er andre mottagelige arter (for eksempel andre torskefisk) i miljøet, er det mulig at noe smittepåvirkning også kan skje indirekte via disse artene. Økt smittepress til miljøet vil også kunne bidra til oppbygging av reservoar. Erfaringsmessig vil det også komme ukjente sykdommer (hittil nye og uoppdagede) med oppskalering og/eller oppdrett av nye arter.
Når fisk smittes er det i hovedsak tre ulike scenarier som kan oppstå. 1. Fisken bekjemper infeksjonen og forblir frisk. 2. Fisken blir smittet og utvikler sykdom, men bekjemper sykdommen og blir frisk. I slike tilfeller kan fisken også fortsette som smittebærer. 3. Fisken blir smittet, utvikler sykdom og dør eller blir spist (økt dødelighet som følge av predasjon). Smittespredning til villfisk kan føre til økt dødelighet ut over den naturlige dødeligheten i populasjonen. Samtidig vil sårbarheten til en lokal torskebestand være større om den er liten.
Rømming av smittet fisk vil også kunne spre smitte til og i miljøet, både horisontalt (mellom individer) og vertikalt (til avkom, ved gyting). Torsk er kjent som en kannibalistisk art. Enkelte agens kan også overføres oralt, og rømt oppdrettstorsk med smitte vil kunne smitte villtorsk med enkelte agens, som f.eks. Francisella noatunensis, om den spises.
Eliminasjon eller begrensing av sykdomstilfeller er viktig for å hindre smittespredning fra torskeoppdrett. God forebygging gjennom biosikkerhetstiltak i alle ledd av produksjonen og beskyttende vaksinering er tiltak som hindrer infeksjoner i oppdrettsfisken og dermed kan redusere risiko for smitte fra anlegg til omgivelsene og villfisken (Sitja-Bobadilla mfl. 2017).
Det er sannsynlig at det spres patogener til miljøet fra smittet og syk torsk i oppdrett. Slik smittespredning kan deles opp i følgende scenarier:
-
Smitte til villfisk, både torsk og andre mottakelige arter
-
Smitte innad i anlegg og mellom anlegg (torsk – torsk)
-
Smitte til lakseanlegg både laks og rensefisk (leppefisk og rognkjeks).
Det er per i dag en lav oppdrettsintensitet av torsk og det meldes om få og sporadiske sykdomstilfeller på enkeltlokaliteter av vibriose (Vibrio anguillarum, O2 β, Vibrio anguillarum), Aliivibrioser (Aliivibrio logei), sår (Moritella viscosa og Tenacibaculum finnmarkense) og gjelleproblemer (epiteliocyster, Trichodina sp.) (Moldal mfl. 2025). I dagens torskeoppdrett oppgir næringen selv at de største utfordringer er av ikke-infeksiøs årsak: tarmslyng / tarmproblemer, tapersyndrom, gytespreng, hjerteforandringer og rygg- /kjevedeformiteter. Tapersyndrom er en generell betegnelse på svekkede individer, og en uavklart diagnose, som ikke kan utelukkes at skyldes et smittsomt agens.
Hvordan sykdomsutviklingen vil arte og utvikle seg etter hvert som torskenæringen vokser i volum, er usikkert. Det har i de siste 10-12 årene vært gjennomført svært lite sykdomsforskning på torsk, og det er generelt lite kunnskap om sykdom i torsk, inkludert for kjente agens og deres sykdommer.
Vi vet i dag lite om hvor stor smittefaren er eller konsekvensene av en eventuell smittespredning fra oppdrettstorsk til gyte- og oppvekstområder for torsk. Kartlegging, overvåking og forskning vil være nødvendig for å forstå dette bedre. Forekomst av relevante parasitter, virus og bakterier bør derfor kartlegges og overvåkes systematisk i utvalgte bestander både før og etter at torskeoppdrett har økt i omfang. Denne typen overvåking vil også øke sjansen for å identifisere både nye agens, eller nye varianter av allerede kjente agens. Økt kunnskap om dette vil være viktig for å øke biosikkerheten, minske spredning av smitte fra oppdrett til villfisk og også hvilke konsekvenser eventuell smitte vil kunne ha på villtorsk. I tillegg vil slik kunnskap også være viktig for raskt å kunne iverksette tiltak når nye sykdomsproblemer oppstår.
De viktigste kjente parasitter, bakterier og virus fra torsk og torskeoppdrett er her gitt en kort omtale. Alle disse agensene er i dag naturlig hjemmehørende i villtorskpopulasjoner, som utgjør naturlige reservoarer. Men høyt smittepress over tid vil kunne føre til økt sykdomsforekomst hos villtorsk. Over tid kan en også få virulensutvikling. Hvis dette skjer i yngelproduksjon kan en få omfattende sykdomsutbrudd, men også spredning med frisk settefisk som kan være bærere.
2.3.5.1 - Parasitter
Vi kjenner over hundre forskjellige parasittarter fra torsk, og noen av disse kan forårsake sykdomsproblemer i torskeoppdrett. Generelt vil parasitter med direkte smittevei, dvs. uten mellomverter, være de som gir størst problemer. Slike parasitter kan få gode livsbetingelser i et system med høy tetthet av verter – som er normalt i oppdrett. Av de mange parasittene som vi vet infiserer torsk, har minst 22 arter slik direkte smitte (encellede parasitter, Gyrodactylus spp. og copepoder), og de finnes hos torsk i Norge. Noen av disse har allerede forårsaket problemer i torskeoppdrett (Trichodina spp., Gyrodactylus spp., Ichthyobodo spp., Caligus elongatus), og dette har medført behov for behandling med kjemikalier (Nygaard 2005; Karlsbakk mfl. 2009; Khan 2009; Heuch mfl. 2011).
Det har de siste årene vært problemer med systemiske infeksjoner med den encellede parasitten Gadixcellia gadi. Denne parasitten er kjent å forårsake såkalte pseudobrankie-svulster hos villtorsk (f.eks. Lange & Johannessen 1977, Egidius mfl.1981). Dette er ikke kreftsvulster som en tidligere trodde, men nå vist å være forårsaket av en encellet parasitt (Freeman mfl. 2011, 2017). En kaller derfor disse svulstaktige parasittansamlingene for xenoma. Der er to pseudobrankier i ganetaket på fisk, og det er blitt vist at ensidige slike xenoma er lite skadelige for torsken, mens tosidige kan være dødelige. En annen slik x-celleparasitt, Salmoxcellia vastator, forekommer i oppdrettede laksefisk, og den danner vanligvis ikke slike celleklumper eller xenoma, men spres med blodet (Karlsbakk mfl. 2021). Den nye torskesykdommen (systemisk gadixcelliose) arter seg likedan, med celler i blodet, men disse kan også danne aggregater og blokkeringer i blodårer i gjeller, hjerte, muskel og andre organ (f.eks. Kvalvik 2025). Der er dødelighet. Tilstanden er påvist fra Nordfjord til Finnmark, i noen anlegg et særfunn, i andre med mye syk fisk. Det er gjort smitteforsøk med G. gadi, uten at en har klart å smitte torsken. Derfor har en spekulert på om en ytterligere vert er involvert (Eydal mfl. 2010, Freeman mfl. 2011, 2017). Per nå, er kunnskapen om denne og beslektede parasitter så begrenset at det ikke er mulig å vurdere en smitterisiko. G. gadi er kun forekommende i torsk.
Eksempler på andre agens som kan forårsake sykdom hos oppdrettstorsk og dermed føre til risiko for smittespredning er mikrosporidien Loma morhuae (Bricknell mfl. 2006; Khan 2009). Denne parasitten forårsaker sykdom i nordvest-Atlanteren, mens infeksjoner i oppdrettstorsk i norske farvann er uvanlige og begrensede (harmløse).
Villtorsk parasitteres av to arter lus, torskelus (Caligus curtus) og skottelus (Caligus elongatus). Begge kan forekomme på oppdrettstorsk i merd. Disse er slektninger av lakselus, som de på mange måter kan sammenlignes med. Torskelus finner man på torsk og noen andre torskefisk som hyse, lyr, sei, lange og brosme. Den er derfor spesifikk for torskefisk, og ikke bare for torsk. Skottelusa derimot er en generalist (ikke spesifikk) som man har funnet på mange ubeslektede fiskearter i hele landet. Skottelus vokser ofte opp på småfisk, men også på større fisk som rognkjeks og laks. Der er to varianter (genotyper) av lusen, trolig to arter. Disse ser ut til å ha forskjellig forekomst gjennom året (Øines & Heuch 2007) og de synes ha forskjellige vertspreferanser (Øines mfl. 2006; Øines & Heuch 2007; Persson 2021). I Sør-Norge er ikke skottelus fra voksen torsk (type 2) den samme som er vanlig på laks og rognkjeks (type 1), særlig sommer høst. En kan finne skotteluselarver på små torskeyngel i naturen, men sjelden på den større fisken en holder i merdanlegg Karlsbakk mfl. 2008, 2009; Heuch mfl. 2011).
Det er forskjeller mellom Nord-Norge og Sør-Norge når det gjelder påslag på torsk i merd av disse luseartene. I Sør-Norge forekommer torskelusa på vandrende torsk i gytetiden, men er svært uvanlig på gruntlevende lokal villtorsk og på oppdrettstorsk. I sør er det derfor ikke påslag med torskelus. I Nord-Norge er det derimot registrert påslag av torskelus på torsk i merd (Karlsbakk mfl. 2009; Heuch mfl. 2011). Denne lusen har larver som vokser opp på torsken, og en kan derfor tenke seg at torskelusen kan forårsake lignende problemer som lakselus på laks hvis oppdrettsvolumene blir store. En har så langt ikke måttet behandle mot torskelus. Villtorsk og andre torskefisk kan ha kraftige infeksjoner og utgjør et betydelig reservoar av smitte. Derfor, per i dag er smitteveien hovedsakelig fra villfisk til oppdrettsfisk, og kun i nord.
Skottelusa forekommer som nevnt i to genetiske varianter, med forskjellig vertstilfang og geografisk utbredelse. Det kan være snakk om to arter (Øines & Schram 2008; Persson 2021), og begge forekommer på torsk. Skottelus er omtrent like vanlig nord og sør i Norge. Denne lusa kan forårsake omfattende skottelusepåslag på torsk i merd. Dette er da med voksne lus som har vokst opp på andre fiskearter (villfisk) og som er frittsvømmende i vannet, og ikke lus som har vokst opp på merdtorsken (Karlsbakk mfl. 2009; Paulsen 2018; Persson 2021). Påslagene av skottelus kan derfor være plutselige, og beiteskader har medført at en har måttet behandle fisken (Nygaard 2005). Denne lusens biologi er derfor annerledes på torsk enn lakselusens på laks. Likevel kan kraftige luseinfeksjoner på torsken tenkes å forårsake økt smittepress på villfisk i omgivelsene. Voksne skottelus på oppdrettstorsk kan frigjøre luselarver som kan smitte villfisk. Det er lite trolig at de vil smitte større torsk, fordi disse er lite mottagelige og er nesten aldri infisert med skottelus chalimi (Heuch mfl. 2011; King 2023). Småyngel kan smittes, men da uten alvorlige konsekvenser da de voksne lusene vil forlate yngelen (Karlsbakk mfl. 2001). Oppdrettspåvirkning på villtorsk må derfor trolig skje via andre fisk der lusene vokser opp, før de eventuelt smitter som voksne til større villtorsk. Da skottelus som regel forårsaker lite skade på torsk, forventes det ikke påviselige negative effekter før næringen er betydelig større enn nå. Forekomst, økologi og adferd hos torskelus og de to skottelusvariantene er lite undersøkt, og det trengs derfor forskning på begge disse Caligus-artene i årene fremover.
2.3.5.2 - Bakterier
En forutsetning for å kontrollere bakterielle sykdommer i oppdrett er gode og funksjonelle vaksiner tilpasset den aktuelle oppdrettsarten. Bakteriesykdommer det tidligere var utfordringer med var vibriose og kaldtsvannsvibriose, forårsaket av ulike Vibrio og Aliivibrio-bakterier, samt furunkulose, som forårsakes av ulike varianter av bakterien Aeromonas salmonicida. Før vi fikk gode beskyttende vaksiner var disse sykdommene et stort problem i lakseoppdrett.
Mange bakterier som har forårsaket sykdom hos oppdrettsfisk har vært såkalte opportunister: de er lite artsspesifikke og invaderer eller oppformeres voldsomt når det blir mulig. Slike bakterier utgjør et kjent problem for både torsk og rensefisk i oppdrett. De finnes ofte i miljøet og kan gjerne overleve uavhengig av verter, eller de utgjør en del av vertenes normale mikroflora. De kan forårsake sykdom når verten svekkes. De kan også utvikle mer fisketilpassede skadelige stammer som sirkulerer i oppdrett. Slike bakterier, som for eksempel Aliivibrio spp., ulike arter og serotyper av Vibrio og atypiske furunkulosebakterier (Aeromonas salmonicida sspp.), er kjent fra mange fiskearter. Sist torskeoppdrett var stort i Norge, sto disse bakterietypene for en stor del av sykdomsutbruddene. I tillegg kommer utfordringer med sårbakterier som for eksempel Moritella viscosa (Hellberg mfl. 2011), Flavobacterium frigidarium (data fra Havforskningsinstituttet 2010, upublisert) og Tenacibaculum spp. De vaksiner, i all hovedsak autogene vaksiner, som benyttes i dagens torskeoppdrett meldes å gi grei beskyttelse og det rapporteres om få bakterielle problemer og utbrudd.
En sykdom det ennå ikke finnes en kommersiell vaksine mot er francisellose hos torsk som er forårsaket av den parasittiske bakterien Francisella noatunensis subsp. noatunensis. Bakterien er intracellulær (lever inni fiskens celler) og forårsaker systemisk granulomatøs inflammasjon (francisellose) som ikke kan behandles med antibiotika (Nylund mfl. 2006; Furevik mfl. 2011). På tidlig 2000-tallet ble det observert utbrudd fra Boknafjorden til Gildeskål like sør for Bodø, med hovedvekt på Vestlandet og Møre. Dette indikerer at temperatur kan spille en viktig rolle i smitte og sykdomsutbrudd (Bakkemo mfl. 2016). Francisellose er kategori F listeført sykdom i Norge. Den var sist påvist i akvakulturanlegg i Norge i 2014 (Sommerseth mfl. 2022) inntil en enkeltstående påvisning i februar 2025. Andre arter enn torsk er ikke kjent å være mottagelige.
Smittet torsk kan ved høyere sjøtemperatur utvikle betennelsesknuter i indre organer, særlig i milt og nyre som kan bli svært oppsvulmet (Karlsbakk mfl. 2008; 2010; Colquhoun og Duodu 2011; Ottem mfl. 2008; Omdal 2009). Francisellose utvikler seg under normale norske miljøforhold langsomt, og fisken kan være smittebærer over lengre tid uten at det oppdages. Det gjør at både behandling og forebygging av sykdommen kan være vanskelig. Oppblomstring av bakterien og smittespredning er særlig knyttet til høyere temperaturer (> ca. 12°C), og en har derfor tidligere hatt mest problemer med francisellose i Sør-Norge. Smitte skjer horisontalt ved at fisk med francisellose skiller ut bakterien i vannet, og oralt ved kannibalisme. Det er blitt registrert økt forekomst av francisellose i villtorsk i etterkant av sykdomsutbrudd i oppdrett. Det er uavklart om overlevelsen til affisert villtorsk er redusert ved normale sjøtemperaturer, men ved høye temperaturer (>18°C) kan oppdrettstorsk dø (normalt er flere andre agens involvert samtidig).
Mye tyder på at bakterien er vanlig forekommende hos vill torsk i sørlige deler av Norge. Det har vært epidemier av sykdommen i villtorsk i Skagerrak/Kattegat i varme somre. Med varmere klima og høyere sjøtemperaturer kan en derfor ikke utelukke at denne sykdommen vil kunne øke i omfang, både i vill og oppdrettet torsk. Villtorsk i Sør-Norge er relativt ofte (7-11 %) bærere av bakterien. Villtorsk med francisellose er observert på Sørlandet, langs den svenske vestkysten og sørover i Nordsjøen, fra før en begynte med torskeoppdrett. Francisellose vanskeliggjorde tidligere torskeoppdrett i Rogaland, men bakterien er uvanlig i villtorsk i Nord-Norge og oppdrettstorsk der vil normalt ikke kunne utvikle sykdommen på grunn av lavere sjøtemperatur. Flytting av settefisk fra sør har tidligere ført til smittepåvisninger i nord. Dette er et scenario som kan unngås med regional yngelproduksjon, smittefri stamfisk, intensiv yngelproduksjon og påvekst til settefisk i lukkede systemer. I sør vil forekomst av bakterien i villtorsk kunne føre til at oppdrettsfisk i åpne merder smittes. En kan i så fall få utbrudd og smittespredning på sommer eller høst. Smitte fra mordyr via egg til yngel (vertikal smitte) er usannsynlig, men ikke umulig. I dag kontrolleres all stamfisk for tilstedeværelse av Francisella, så smittespredning fra settefisk er svært usannsynlig.
Bakterier av arten Aeromonas salmonicida er velkjent fra både oppdretts- og villfisk og forårsaker sykdommen furunkulose. Bakterien forårsaket store problemer for laksenæringen i Norge på slutten av 80 og begynnelsen av 90-tallet etter å ha blitt importert til landet på midten av 80-tallet med laksesmolt fra Skottland (Egidius 1987). Man regner bakterien for å ha et reservoar i naturen, både salt- og ferskvann. Utvikling av vaksiner førte til god beskyttelse mot sykdommen og det har siden vært få utbrudd i lakseoppdrett. Navnet på sykdommen kommer av symptomet med byller (furunkler) i muskulaturen hos syk fisk. Det skilles mellom typisk eller klassisk og atypisk furunkulose avhengig av hvilken type (underart) av bakterien som forårsaker sykdommen. Klassisk eller typisk furunkulose forårsakes av Aeromonas salmonicida subsp. salmonicida og er kjent fra laks og laksefisk. Furunkulose hos vill laksefisk opptrer fra tid til annen ved høye temperaturer og gjerne ved lav vannføring i elv (Sommerseth mfl. 2022). Hverken atypisk eller typisk furunkulose ble påvist på vill eller oppdrettet laks i 2024 (Garseth mfl. 2025). Furunkulose forårsaket av andre subarter som: achromogenes, masoucida, pectinolytica og smithia kalles atypisk furunkulose. Atypisk furunkulose var, sammen med vibriose og francisellose, en vanlig bakteriell sykdom sist torskeoppdrett var på sitt høyeste (Hellberg mfl. 2011). Atypisk furunkulose forårsaker fortsatt dødelighet hos oppdrettstorsk i 2024 (Moldal mfl. 2025). Hos torsk i oppdrett var det historisk sett subarten achromogenes som dominerte (oppsummert i Puvanendran mfl. 2021). Denne subarten er kjent for å forårsake sykdom og dødelighet også i andre marine oppdrettsarter som piggvar og sjøtunge (Torres-Corral mfl. 2022).
Med bruken av rensefisk har også forekomsten av furunkuloseutbrudd i oppdrett økt, og atypisk furunkulose er en av de hyppigste påviste bakterielle sykdommene hos både leppefisk og rognkjeks. Tilbake i 2020 meldte Veterinærinstituttet om økt bekymring for klassisk furunkulose i matfiskproduksjon av laks med fem påvisninger i 2020 og 2021. Da var ikke bakterien påvist i laks siden 2014. I tillegg er også denne varianten av bakterien nå påvist hos syk rognkjeks brukt som rensefisk (Sommerseth mfl. 2021). Påvisning av furunkulose hos laks har i de siste årene gått ned og i 2023 og 2024 var det ingen slike påvisninger (Moldal mfl. 2025).
Klassisk furunkulose (Aeromonas salmonicida subsp. salmonicida) er en listeført sykdom i Norge (kategori F) og påvisning fører til opprettelse av et kontrollområde med en bekjempelsessone og observasjonssone. Bakterien smitter horisontalt, og utbrudd av furunkulose forekommer ofte i perioder på året med vanntemperaturer over 10 grader. Aeromonas-bakterier har et tykt ytterlag bestående av protein. Dette ytterlaget, kalt A-laget, er en viktig virulensfaktor og gir bakterien en sterk hydrofob overflate som gjør at de klumper seg sammen. Bakteriens hydrofobe egenskaper gjør at den vil finnes i overflatefilm rundt anlegg med utbrudd. Bakterien danner også biofilm (Desbios mfl. 2020) og vil derfor kunne trives godt i biofilm på alle overflater i tilknytning til oppdrett (som kar, merder, rørsystemer o.l.) I forsøk er det også vist at tilstedeværelse av sediment øker overlevelsen til A. salmonicida i miljøet, noe som peker på at det her kan finnes et reservoar. Det er også indikasjoner på økt overlevelsen i brakkvann (6,4 +/- 0,5 ‰) i forhold til i saltvann (30 ‰) (Wiklund 1995). Vibriose forårsakes av ulike varianter og serotyper av Vibrio-bakterier og sykdomsutbrudd er kjent fra både oppdretts- og villfisk som torsk og sei. Bakterien smitter horisontalt og man regner med at Vibrio-bakterier har et marint reservoar i villfisk (oppsummert i Puvanendran mfl. 2021). Hos torsk i oppdrett har historisk sett vibrioseutbrudd vært dominert Vibrio anguillarum serotypene O2β og O2α (Hellberg mfl. 2011; oppsummert i Puvanendran mfl. 2021). Det bør også nevnes at det fra syk oppdrettstorsk ble isolert V. anguillarum-varianter som var ulike de vanlige beskrevne og kjente serotypene (Mikkelsen mfl. 2007). Det er viktig å ha kunnskap om de variantene som forårsaker sykdomsutbrudd for å sikre best mulige og beskyttende vaksiner.
Vibrio-bakterier er opportunister og sykdomsutbrudd kommer gjerne når fisk har svekket immunforsvar som følge av stress eller endring i miljøforhold (f. eks svingninger i temperatur, gjerne høye temperaturer). Vanlige sykdomstegn for vibriose er blødninger i hoderegionen, ved finnebasis, finneråte og sår i hud. Vibriose er også en utfordring hos annen marin fisk i oppdrett, som leppefisk og rognkjeks. Men bør derfor være oppmerksom på smitte mellom alle arter av marin fisk og laksefisk i oppdrett.
2.3.5.3 - Virus
Betanodavirus varianten av Nodavirus som infiserer torsk i Norge (GMNNV, en type BFNNV) er utbredt i villtorsk i hele landet (Nylund mfl. 2008). Viruset angriper nervesystemet, og kan forårsake nervevevsdød - sykdommen viral nervøs nekrose (VNN, også kalt viral encephalopati og retinopati VER). VNN er vanligst hos yngel og er kun kjent fra oppdrettssituasjoner. Vi vet i dag altfor lite om hvor stor faren for vannbåren smitte med GMNNV er, fra smittet oppdrettstorsk til forskjellige livsstadier av villtorsk og for andre villfisk.
Det har forekommet sykdomsutbrudd tidligere, da det var mye torskeoppdrett, men disse var få og sporadiske. Det begynte med tre utbrudd i 2006, toppet seg med seks utbrudd i 2007, men en har ikke hatt utbrudd etter 2009. Utbruddene var både blant yngel og større torsk. Det har altså ikke vært mange VNN tilfeller, gitt det relativt store antallet anlegg i drift. GMNNV forekomst i villtorsk ble tidlig kartlagt (2005-2009), og funnet vanlig forekommende (10-20% er infisert). De infiserte villfiskene er friske, men har viruset i seg (de er "bærere"). Slike fisk vil normalt ikke spre smitte, men viruset kan blomstre opp under visse forhold, inkludert ved kjønnsmodning, og da kan fisken bli smittespredende. GMNNV vil normalt ikke utgjøre en stor fare for villtorsken (som jo ofte er smittet). Derimot vil import av settefisk fra andre land representere en trussel, da de kan bære på andre NNV-typer som kan være mer skadelige for norsk torsk. Import av levende oppdrettsfisk medfører vesentlig risiko, og bør ikke tillates.
Et problem med denne typen virus er at de i tillegg til å smitte horisontalt kan smitte vertikalt, altså mordyr-egg-yngel (Breuil mfl. 2002). Avkom av smittet stamfisk kan altså bli bærere av viruset, og senere være opphav til NNV utbrudd. Oppdrettstorsk kan modnes og gyte i merd og det kan dermed skje omfattende smittespredning hvis GMNNV infisert fisk gyter i merd. Siden en relativt høy andel av villfisken også er smittet naturlig utgjør dette fenomenet primært en trussel hvis:
-
Det introduseres eksotiske skadelige virusvarianter (import av fisk)
-
Det utvikles særlig skadelige (virulente) varianter av viruset i oppdrett
Det er derfor viktig å sikre smittefri stamfisk, og unngå kjønnsmodning. Det bør ikke tillates import av settefisk fra andre land. Det finnes vaksiner, men så langt ikke for torskevarianten og torsk.
Et annet marint fiskevirus som kan skape problemer, er rabdoviruset viral haemorragisk septikemi virus (VHSV) som forårsaker den alvorlige sykdommen viral haemorragisk septikemi (VHS). VHSV er ifølge OIE kjent fra over 80 arter av både saltvanns og ferskvannfisk inkl torsk og salmonider (Anonym 2014). En lite skadelig variant av VHSV forekommer naturlig i sild (Sandlund mfl. 2014; Johansen mfl. 2013) og i villtorsk (Karlsbakk mfl. 2008) i Norge. Smitteforsøk tyder på at torsk er forholdsvis resistent mot VHS (Sandlund mfl. 2021; Snow mfl. 2000; Snow mfl. 2005), men kan utvikle sykdommen. VHS-virusets overlevelse i vann er rapportert som dager (Hawley & Garver 2008) og uker (Brun & Lillehaug 2010). Smitte via føde er en kjent smittevei for VHSV (Ahne 1980; Schönherz mfl., 2012). Viruset kan også overleve i frossen fisk og vil kunne smitte videre om frossen fisk benytte som fôr (Gudding & Lillehaug 2018).
Infeksiøs pankreas nekrose virus (IPNV) som forårsaker infeksiøs pankreas nekrose (IPN), har et vidt vertsspekter (Munang’andu mfl. 2016). Det er viktig hos laks, og har også vært en viktig dødsårsak i oppdrett av kveite (Bloch mfl. 1991, Johnsen mfl. 2004). Viruset er isolert fra norsk villtorsk (Jensen mfl. 2009), og IPNV har forårsaket IPN hos torskeyngel på Førøyene og i Norge (ett tilfelle). Viruset er også påvist å kunne forårsake dødelighet under eksperimentelle betingelser (Urquhart mfl. 2009). I de senere år er også IPNV isolert fra oppdrettstorsk i Norge. Viruset har hittil ikke forårsaket store sykdomsutfordringer eller dødelighet i torskeoppdrett. I et nylig gjennomført smitteforsøk (kohabitantsmitte) ble det vist at viruset kan smitte videre fra ip-injisert torsk til både torsk, rognkjeks, kveite og laks (Weli mfl. 2025). Denne typen forsøk viser potensialet for smitte mellom arter i oppdrett og til villfisk.
2.3.5.4 - Nylige påviste sykdommer hos oppdrettstorsk
Det er påvist flere nye agens på torsk, og nye sykdommer. Systemisk gadixcellliose er nevnt over. Poxvirus er kjent å forårsake gjellesykdom hos laks i oppdrett. Nylig er også denne typen virus beskrevet hos torsk i oppdrett med alvorlig gjellesykdom. Varianten av viruset som er beskrevet fra torsk er genetisk forskjellig fra laksevarianten og omtales som «cod gill pox virus» (CGPV) (Gjessing mfl. 2024).
Et annet gjellepatogen, kjent fra laks, er nå også påvist hos torsk, Candidatus Branchiomonas cysticola (Gjessing mfl. 2024). Signifikansen er uklar.
2.4 - Takk
Takk til Monica F Solberg, Kevin Glover, Ørjan Karlsen, Bjarte Bogstad og Åge Høines for diskusjoner og kvalitetssikring av tekst. Spesiell takk til Johanna Fall og Brian Stock for bidrag til tekst om kysttorskbestandene i kunnskapsstatus.
2.5 - Referanser
Aas, T.S., Ytrestøyl, T. & Åsgård, T. (2019). Utilization of feed resources in the production of Atlantic salmon ( Salmo salar ) in Norway: An update for 2016. Aquaculture Reports 15, 100216. https://doi.org/10.1016/j.aqrep.2022.101316
Ahne, W. (1980). Experimental infection of pike ( Esox lucius ) with Egtvedvirus. Tierarztliche Umschau 35, 225-229.
Aglen, A., Nedreaas, K., Knutsen, J.A. & Huse, G. (2020). Kysttorsk nord for 62-grader nord. Vurdering av status og forslag til forvaltningstiltak og ny gjenoppbyggingsplan. Fisken og Havet nr. 2020-2. 64 pp. https://www.hi.no/hi/nettrapporter/fisken-og-havet-2020-2
Alix, M., Mihaljevic, M. & Norberg, B. (2024). Videreutviklet protokoll for vurdering av modningsgrad hos oppdrettstorsk. Rapport fra havforskningen 2024-58, 61 pp. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-2024-58
Alix, M., Yadav Ahir S., Geitung L., Perrichon P., Fraser T., Fjelldal PG, van der Meeren T., Harboe T., Utskot S.O., Nola V., Oppedal F., Dempster T., Norberg B. (2025). Reproductive biology and offspring development success in farmed cod are affected by nutrition, light and submergence. AQUA 2025, 22-25 September, Valencia, Spania.
Almeida, F.F.L., Taranger, G.L., Norberg, B., Karlsen, K., Bogerd, J. & Schulz, R.W. (2009). Photoperiod-Modulated Testis Maturation in Atlantic Cod ( Gadus morhua , L.). Biology of Reproduction 80, 631-640. https://doi.org/10.1095/biolreprod.108.071340
André, C., Svedäng, H., Knutsen, H., Dahle, G., Jonsson, P., Ring, A.-K., Sköld, M. & Jorde, P.E. (2016). Population structure in Atlantic cod in the eastern North Sea-Skagerrak-Kattegat: early life stage dispersal and adult migration. BMC Res. Notes 9, 63. https://doi.org/10.1186/s13104-016-1878-9
Andersen, L.B., Grefsrud, E.S., Svåsand, T. & Sandlund, N. (2022). Risk understanding and risk acknowledgement: a new approach to environmental risk assessment in marine aquaculture. ICES Journal of Marine Science 79, 987-996. https://doi.org/10.1093/icesjms/fsac028
Apel, J.R. (1988). Chapter Nine Optics of the Sea. In: Apel, J.R., (Ed.), International Geophysics, Academic Press Volume 38, 511-605.
Araki, H. & Schmid, C. (2010). Is hatchery stocking a help or harm?: Evidence, limitations and future directions in ecological and genetic surveys. Aquaculture 308, Supplement 1, S2-S11. https://doi.org/10.1016/j.aquaculture.2010.05.036
Árnason, T. & Björnsson, B. (2012). Spawning-related mortality in captive Atlantic cod ( Gadus morhua L.). Aquaculture Research 43, 292-296. https://doi.org/10.1111/j.1365-2109.2011.02829.x
Arnott, S.A. & Pihl, L. (2000). Selection of prey size and prey species by 1-group cod Gadus morhua : effects of satiation level and prey handling times. Marine Ecology Progress Series 198, 225-238. https://doi.org/10.3354/meps198225
Asplin, L., Johnsen, I.A., Sandvik, A., Albretsen, J., Sundfjord, V., Aure, J. & Boxaspen, K.K. (2014). Dispersion of salmon lice in the Hardangerfjord. Marine Biology Research 10, 216-225. https://doi.org/10.1080/17451000.2013.810755
Aven, T. (2014). Risk, Surprises and Black Swans: Fundamental Ideas and Concepts in Risk Assessment and risk management. Routledge, London. 276 s.
Bakkemo, K.R., Mikkelsen, H., Johansen, A., Robertsen, B. & Seppola, M. (2016). Francisella noatunensis subsp. noatunensis invades, survives and replicates in Atlantic cod cells. Diseases of Aquatic Organisms 121, 149-159. https://doi.org/10.3354/dao03043
Barrett, L.T., Swearer, S.E., Harboe, T., Karlsen, Ø., Meier, S. & Dempster, T. (2018). Limited evidence for differential reproductive fitness of wild Atlantic cod in areas of high and low salmon farming density. Aquaculture Environment Interactions 10, 369-383. https://doi.org/10.3354/aei00275
Barrett, L.T., Swearer, S.E. & Dempster, T. (2019). Impacts of marine and freshwater aquaculture on wildlife: a global meta-analysis. Reviews in Aquaculture 11, 1022–1044. https://doi.org/10.1111/raq.12277
Barth, J.M.I., Berg, P.R., Jonsson, P.R., Bonanomi, S., Corell, H., Hemmer-Hansen, J., Jakobsen, K.S., Johannesson, K., Jorde, P.E., Knutsen, H., Moksnes, P.O., Star, B., Stenseth, N.C., Svedang, H., Jentoft S. & Andre, C. (2017). Genome architecture enables local adaptation of Atlantic cod despite high connectivity. Molecular Ecology 26, 4452-4466. https://doi.org/10.1111/mec.14207
Battin, J. (2004). When good animals love bad habitats: ecological traps and the conservation of animal populations. Conservation Biology 18, 1482–1491. https://doi.org/10.1111/j.1523-1739.2004.00417.x
Bekkevold, D., Hansen, M.M. & Nielsen, E.E. (2006). Genetic impact of gadoid culture on wild fish populations: predictions, lessons from salmonids, and possibilities for minimizing adverse effects. ICES Journal of Marine Science 63, 198-208. https://doi.org/10.1016/j.icesjms.2005.11.003
Bell, J. G.,McGhee, F.,Campbell, P. J. & Sargent, J.R. (2003). Rapeseed oil as an alternative to marine fish oil in diets of post-smolt Atlantic salmon ( Salmo salar ): changes in flesh fatty acid composition and effectiveness of subsequent fish oil “wash out”. Aquaculture 218, 515–528. https://doi.org/10.1016/S0044-8486(02)00462-3
Berg, P.R., Star, B., Pampoulie, C., Sodeland, M., Barth, J.M.I., Knutsen, H., Jakobsen, K.S. & Jentoft, S. (2016). Three chromosomal rearrangements promote genomic divergence between migratory and stationary ecotypes of Atlantic cod. Scientific Reports 6, 23246. https://www.nature.com/articles/srep2324
Berg, E. & Albert, O.T. (2003). Cod in fjords and coastal waters of North Norway: distribution and variation in length and maturity at age. ICES Journal of Marine Science 60, 787–797. https://doi.org/10.1016/S1054-3139(03)00037-7
Bjørn, P.A., Uglem, I., Sæther, B.S., Dale, T., Kerwath, S., Økland, F., Nilsen, R., Aas, K. & Tobiassen, T. (2007). Videreføring av prosjektet «Behavioural responses in wild coastal cod exposed to salmon farms: possible effects of salmon holding water – a field and experimental study. Rapport 6/2007, Fiskeriforskning. 38 pp. www.fhf.no/prosjekter/prosjektbasen/242021/?fileurl=https://fhfno.sharepoint.com/sites/pdb/Dokumenter/Import1/242021.pdf&filename=sluttrapport
Bjørn, P.A., Uglem, I., Kerwath, S., Sæther, B.S., Nilsen, R. (2009). Spatiotemporal distribution of Atlantic cod ( Gadus morhua L.) with intact and blocked olfactory sense during the spawning season in a Norwegian fjord with intensive salmon farming. Aquaculture 286, 36-44. https://doi.org/10.1016/j.aquaculture.2008.08.038
Bloch, B., Gravningen K. & Larsen J. L. (1991). "Encephalomyelitis among turbot associated with a Picornavirus-like agent." Diseases of Aquatic Organisms 10, 65-70. https://doi.org/10.3354/dao010065
Blom, G., Otterå, H., Svåsand, T., Kristiansen, T.S. & Serigstad, B. (1991). The relationship between feeding conditions and production of cod fry ( Gadus morhua L.) in a semi-enclosed marine ecosystem in western Norway, illustrated by use of a consumption model. ICES Marine Science Symposia 192, 176-189.
Bolstad G.H., Hindar K., Robertsen G., Jonsson B., Saegrov H., Diserud O.H., Fiske P., Jensen A.J., Urdal K., Naesje T.F., Barlaup B.T., Florø-Larsen B., Lo H., Niemela E. & Karlsson S. (2017). Gene flow from domesticated escapees alters the life history of wild Atlantic salmon. Nature Ecology & Evolution 1, 0124. https://doi.org/10.1038/s41559-017-0124
Brawn, V.M. (1961). Sound Production By the Cod ( Gadus callarias L.). Behaviour 18, 239–255. https://doi.org/10.1163/156853961X00150
Breistein, B., Dahle, G., Johansen, T., Besnier, F., Quintela, M., Jorde, P.E., Knutsen, H., Westgaard, J.-I., Nedreaas, K., Farestveit, E. & Glover, K.A. (2022). Geographic variation in gene flow from a genetically distinct migratory ecotype drives population genetic structure of coastal Atlantic cod ( Gadus morhua L.). Evol. Appl. 15, 1162-1176 https://onlinelibrary.wiley.com/doi/10.1111/eva.13422
Breuil, G., Pépin, J.F.P., Boscher, S. and Thiéry, R. (2002). Experimental vertical transmission of nodavirus from broodfish to eggs and larvae of the sea bass, Dicentrarchus labrax (L.). Journal of Fish Diseases, 25, 697-702. https://doi.org/10.1046/j.1365-2761.2002.00406.x
Bricknell, I.R., Bron, J.E. & Bowden, T.J. (2006). Diseases of gadoid fish in cultivation: a review. ICES J. Mar. Sci. 63, 253-266, https://doi.org/10.1016/j.icesjms.2005.10.009
Brun, E. & Lillehaug, A. (2010). Risikoprofil for sykdommer i norsk fiskeoppdrett. Norge, Veterinærinstituttets rapportserie nr. 09, 46 pp. https://www.vetinst.no/rapporter-og-publikasjoner/rapporter/2010/risikoprofil-for-sykdommer-i-norsk-fiskeoppdrett
Buscaino, G., Filiciotto, F., Buffa, G., Bellante, A., Stefano, V.D., Assenza, A., Fazio, F., Caola, G., Mazzola, S., (2010). Impact of an acoustic stimulus on the motility and blood parameters of European sea bass ( Dicentrarchus labrax L.) and gilthead sea bream (Sparus aurata L.). Marine Environmental Research 69, 136–142. https://doi.org/10.1016/j.marenvres.2009.09.004
Busch, K.E.T., Falk-Petersen, I.-B., Peruzzi, S., Rist, N.A. & Hamre, K. (2010). Natural zooplankton as larval feed in intensive rearing systems for juvenile production of Atlantic cod ( Gadus morhua L.). Aquaculture Research 41, 1727-1740 https://doi.org/10.1111/j.1365-2109.2009.02450.x
Bøhn, T., Strøm, J.F., Sanchez-Jerez, P., Keeley, N.B., Johansen, T., Gjelland, K.Ø., Sandlund, N., Sæther, B.-S., Sætra, I., Olsen, E.M., Skjæraasen, J.E., Meier, S., van der Meeren, T. & Bjørn, P.A. (2024). Ecological interactions between farmed Atlantic salmon and wild Atlantic cod populations in Norway – a review of risk sources and knowledge gaps. Reviews in Aquaculture 202, 1-18. https://doi.org/10.1111/raq.12899
Børretzen Fjørtoft, H., Besnier, F., Stene, A., Nilsen, F., Bjørn, P.A., Tveten, A.-K., Finstad, B., Aspehaug, V. & Glover, K.A. (2017). The Phe362Tyr mutation conveying resistance to organophosphates occurs in high frequencies in salmon lice collected from wild salmon and trout. Scientific Reports 7, 14258 https://doi.org/10.1038/s41598-017-14681-6
Callier, M.D., Byron, C.J., Bengtson, D.A., Cranford, P.J., Cross, S.F., Focken, U., Jansen, H.M., Kamermans, P., Kiessling, A., Landry, T., O’Beirn, F., Petersson, E., Rheault, R.B., Strand, Ø., Sundell, K., Svåsand, T., Wikfors, G.H. & McKindsey, C.W. (2018). Attraction and repulsion of mobile wild organisms to finfish and shellfish aquaculture: a review. Reviews in Aquaculture 10, 924–949. https://doi.org/10.1111/raq.12208
Casper, B.M., Smith, M.E., Halvorsen, M.B., Sun, H., Carlson, T.J. & Popper, A.N. (2013). Effects of exposure to pile driving sounds on fish inner ear tissues. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 166, 352–360. https://doi.org/10.1016/j.cbpa.2013.07.008
Castellani M., Heino M., Gilbey J., Araki H., Svåsand T. & Glover K.A. (2018). Modelling fitness changes in wild Atlantic salmon populations faced by spawning intrusion of domesticated escapees. Evolutionary Applications 11, 1010-1025. https://doi.org/10.1111/eva.12615
Ciannelli L, Knutsen, H, Olsen EM, Espeland SH, Asplin L, Jelmert A, Knutsen JA, Stenseth NC (2010). Small-scale genetic structure in a marine population in relation to water circulation and egg characteristics. Ecology 91, 2918-2930. https://doi.org/10.1890/09-1548.1
Colquhoun, D.J. & Duodu, S., (2011). Francisella infections in farmed and wild aquatic organisms. Veterinary Research 42, 47. https://doi.org/10.1186/1297-9716-42-47
Cresci, A. Paris, C.B., Foretich, M.A., Durif, C.M., Shema, S., O’Brien, C.J.E., Vikebø, F.B., Skiftesvik, A.B. & Browman, H.I. (2019). Atlantic haddock ( Melanogrammus aeglefinus ) larvae have a magnetic compass that guides their orientation. iScience 19, 1173–1178. https://doi.org/10.1016/j.isci.2019.09.001
Cresci, A., Allan, B. J. M., Shema, D.D., Skiftesvik, A.B. & Browman, H.I. (2020). Orientation behavior and swimming speed of Atlantic herring larvae ( Clupea harengus ) in situ and in laboratory exposures to rotated artificial magnetic fields. Journal of Experimental Marine Biology and Ecology 526, 151358. https://doi.org/10.1016/j.jembe.2020.151358
Cresci, A., Zhang, G., Durif, C.M.F., Larsen, T., Shema, S., Skiftesvik, A.B. &Browman, H.I. (2023). Atlantic cod ( Gadus morhua ) larvae are attracted by low-frequency noise simulating that of operating offshore wind farms. Communications Biology 6, 353. https://doi.org/10.1038/s42003-023-04728-y
Dahl, T.H. & Bjørkevoll, I. (2023). Lysstyrt kjønnsmodning hos oppdrettstorsk. Studium av oppdrettstorsk fra lokaliteten Svartekari i perioden 2022-2023. Møreforskning, Rapport nr. 2303, 17 pp. https://www.moreforsk.no/download.aspx?object_id=upload_images/7EC4F89875AD4A4C92E8C11873A8E7F1.pdf
Dahle G., Quintela M., Johansen T., Westgaard J.I., Besnier F., Aglen A., Jorstad K.E. & Glover K.A. (2018). Analysis of coastal cod ( Gadus morhua L.) sampled on spawning sites reveals a genetic gradient throughout Norway's coastline. Bmc Genetics 19, 42. https://doi.org/10.1186/s12863-018-0625-8
Dahle, G. Jørstad, K.E., van der Meeren, T. & Svåsand, T. (2011). Oppdrettet torsk sin innflytelse på vill torsk og mulige løsninger for overvåkning. Rapport fra Havforskningen 26-2011. 19 pp. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen/2011/hi-rapp_26-2011_til_web
Dahlke, F. T., Butzin, M., Nahrgang, J., Puvanendran, V., Mortensen, A., Pörtner, H.-O., & Storch, D. (2018). Northern cod species face spawning habitat losses if global warming exceeds 1.5°C. Science Advances, 4(11), eaas8821. https://doi.org/10.1126/sciadv.aas8821
Desbois, A.P., Cook, K.J. & Buba, E. (2020). Antibiotics modulate biofilm formation in fish pathogenic isolates of atypical Aeromonas salmonicida . Journal of Fish Diseases 43, 1373-1379. https://doi.org/10.1111/jfd.13232
Damsgård, B., Høy, E., Uglem, I., Hedger, R.D., Izquierdo-Gomez, D. & Bjørn, P.A. (2012). Net-biting and escape behaviour in farmed Atlantic cod Gadus morhua : effects of feed stimulants and net traits. Aquaculture Environmental Interactions 3, 1-9. https://doi.org/10.3354/aei00047
Dempster, T., Sanchez-Jerez, P., Fernandez-Jover, D., Bayle-Sempere, J., Nilsen, R., Bjørn, P.-A. & Uglem, I. (2011). Proxy Measures of Fitness Suggest Coastal Fish Farms Can Act as Population Sources and Not Ecological Traps for Wild Gadoid Fish. PLOS ONE 6, e15646. https://doi.org/10.1371/journal.pone.0015646
Dempster, T., Sanchez-Jerez, P., Uglem, I. & Bjørn, P.-A. (2010). Species-specific patterns of aggregation of wild fish around fish farms. Estuarine, Coastal and Shelf Science 86, 271–275. https://doi.org/10.1016/j.ecss.2009.11.007
Dempster, T., Uglem, I., Sanchez-Jerez, P., Fernandez-Jover, D., Bayle-Sempere, J., Nilsen, R. & Bjørn, P.A., (2009). Coastal salmon farms attract large and persistent aggregations of wild fish: An ecosystem effect. Marine Ecology Progress Series 385, 1–14. https://doi.org/10.3354/meps08050
Diserud O., Fiske P., Sægrov H., Urdal K., Aronsen T., Lo H., Barlaup B.T., Niemela E., Orell P., Erkinaro J., Lund R.A., Økland F., Østborg G.M., Hansen L.P. & Hindar K. (2019). Escaped farmed Atlantic salmon in Norwegian rivers during 1989–2013. ICES Journal of Marine Science 76, 1140-50. https://doi.org/10.1093/icesjms/fsy202
Dunlop, K., Staby, A., van der Meeren, T., Keeley, N., Olsen, E.M., Bannister, R. & Skjæraasen, J.E. (2022). Habitat associations of juvenile Atlantic cod ( Gadus morhua L.) and sympatric demersal fish communities within shallow inshore nursery grounds. Estuarine, Coastal and Shelf Science 279, 108111. https://doi.org/10.1016/j.ecss.2022.108111
Dunlop, K., Strammer, I. & Keeley, N. (2024). "Attraction and avoidance of wild demersal fish and crustaceans to open-net aquaculture pens resolved by baited and towed underwater camera surveys." Frontiers in Marine Science 11 - 2024. https://doi.org/10.3389/fmars.2024.1400195
Egidius, E., Johannessen, J.V., Lange, E., (1981). Pseudobranchial tumours in Atlantic cod, Gadus morhua L., from the Barents Sea. J. Fish Diseases. 4, 527-532. https://doi.org/10.1111/j.1365-2761.1981.tb01166.x
Egidius, E. (1987). Import of furunculosis to Norway with Atlantic salmon smolts from Scotland. ICES CM Documents;1987/F:8. (in mimeo). https://imr.brage.unit.no/imr-xmlui/bitstream/handle/11250/104304/CM_1987_F_8.pdf?sequence=1&isAllowed=y
Engelsen, R., Asche, F., Skjennum, F. & Adoff, G. (2004). New species in aquaculture: some basic economic aspects. In: Moksnes, E., Kjørsvik, E. & Olsen, Y. (Eds). Culture of cold-water marine fish. Blackwell Publishing, Oxford, UK.
Espeland, S.H., Gundersen, A.F. Olsen, E.M. Knutsen, H. Gjøsæter, J. & Stenseth. N.C. (2007). Home range and elevated egg densities within an inshore spawning ground of coastal cod. ICES Journal of Marine Science 64, 920–928. https://doi.org/10.1093/icesjms/fsm028
Espeland, S.H., Olsen, E.M., Knutsen, H., Gjøsæter, J., Danielssen, D. & Stenseth, N.C. (2008). New perspectives on fish movement: kernel and GAM smoothers applied to a century of tagging data on coastal Atlantic cod. Mar. Ecol. Prog. Ser. 372, 231–241. https://doi.org/10.3354/meps07721
Espeland, S.H., Albretsen, J., Nedreaas, K., Sannæs, H., Bodvin, T. & Moy, F. (2013). Kartlegging av gytefelt. Gytefelt for kysttorsk. Fisken og havet nr. 1-2013. 43 pp. https://www.hi.no/resources/publikasjoner/fisken-og-havet/2013/fh_1-2013.pdf
Espeland, S.H., Albretsen, J, Olsen, E.B. & Bodvin, T. (2015). Modelling drift of pelagic offspring: the importance of egg surveys in providing a realistic model initialization. ICES Journal of Marine Science 72, 2578-2589. https://doi.org/10.1093/icesjms/fsv134
Espeland, S., & Sannæs, H. (2018). Estimating cod egg developmental stage based on DNA concentration. ICES Journal of Marine Science 75: 825-830, https://doi.org/10.1093/icesjms/fsx172
Eydal, M., Kristmundsson, Á., Bambir S.H. (2010) Pseudobranchial X-cell pseudotumors in young wild and farmed Atlantic cod Gadus morhua in Iceland. DAO 91, 83–88. https://doi.org/10.3354/dao02248
Fernandez-Jover, D., Martinez-Rubio, L., Sanchez-Jerez, P., Bayle-Sempere, J.T., Lopez Jimenez, J. A., Martinez Lopez, F.J., Bjørn, P.-A., Uglem, I. & Dempster, T. (2011). Waste feed from coastal fish farms: A trophic subsidy with compositional side-effects for wild gadoids. Estuarine, Coastal and Shelf Science 91, 559-568. https://doi.org/10.1016/j.ecss.2010.12.009
Fernandez-Jover, D., Toledo-Guedes, K., Valero-Rodríguez, J.M., Fernandez-Gonzalez, V. & Sanchez-Jerez, P. (2016). Potential retention effect at fish farms boosts zooplankton abundance. Estuarine, Coastal and Shelf Science 181, 144–152. https://doi.org/10.1016/j.ecss.2016.08.015
Fevolden, S.E., Westgaard, J.I., Pedersen, T. & Præbel, K. (2012). Settling-depth vs. genotype and size vs. genotype correlations at the Pan I locus in 0-group Atlantic cod Gadus morhua . Marine Ecology Progress Series 468, 267-278. https://doi.org/10.3354/meps09990
Finn, R.N., Rønnestad, I., van der Meeren, T., & Fyhn, H.J. (2002). Fuel and metabolic scaling during the early life stages of Atlantic cod Gadus morhua . Marine Ecology Progress Series 243, 217-234. https://doi.org/10.3354/meps243217
Fjelldal, P.G., van der Meeren, T., Jørstad, K.E. & Hansen, T.J. (2009). A radiological study on vertebral deformities in cultured and wild Atlantic cod ( Gadus morhua , L.). Aquaculture 289, 6-12. https://doi.org/10.1016/j.aquaculture.2008.12.025
Fleming I.A., Hindar K., Mjolnerod I.B., Jonsson B., Balstad T. & Lamberg A. (2000). Lifetime success and interactions of farm salmon invading a native population. Proceedings of the Royal Society of London Series B-Biological Sciences 267, 1517-1523. https://doi.org/10.1098/rspb.2000.1173
Forseth, T., Barlaup, B.T., Finstad, B., Fiske, P., Gjøsæter H., Falkegard, M., Hindar, A., Mo, T.A., Rikardsen, A.H., Thorstad, E.B., Vollestad, L.A. & Wennevik V. (2017). The major threats to Atlantic salmon in Norway. ICES Journal of Marine Science 74, 1496-513. https://doi.org/10.1093/icesjms/fsx020
Freeman, M.A., Eydal, M., Yoshimizu, M., Watanabe, K., Shinn, A.P., Miura, K., Ogawa, K., (2011). Molecular identification and transmission studies of X-cell parasites from Atlantic cod Gadus morhua (Gadiformes: Gadidae) and the northern black flounder Pseudopleuronectes obscurus (Pleuronectiformes: Pleuronectidae). Parasites & Vectors. 4, 13 p. https://doi.org/10.1186/1756-3305-4-15
Freeman, M., Fuss, J., Kristmundsson, A., Bjorbækmo, M.F.M., Mangot, J.-F., del Campo, J., Keeling, P.J., Shalchian-Tabrizi, K., Bass, D., (2017). X-Cells Are Globally Distributed, Genetically Divergent Fish Parasites Related to Perkinsids and Dinoflagellates. Current Biology. 27, 1645–1651. http://dx.doi.org/10.1016/j.cub.2017.04.045
Furevik, A., Pettersen, E.F., Colquhoun, D. & Wergeland, H.I. (2011). The intracellular lifestyle of Francisella noatunensis in Atlantic cod ( Gadus morhua L.) leucocytes. Fish & Shellfish Immunology 30: 488-494. https://doi.org/10.1016/j.fsi.2010.11.019
Gamfeldt, L., Lefcheck, J.S., Byrnes, J.E.K., Cardinale, B.J., Duffy, J.E. & Griffin, J.N. (2015). Marine biodiversity and ecosystem functioning: what’s known and what’s next? Oikos 124, 252–265. https://doi.org/10.1111/oik.01549
Garseth, Å.H., Furnesvik, L., Hansen, H., Haukland, H., Kielland, Ø.K. & Strand, D.A. (2025). Villfiskrapporten 2024. Veterinærinsituttets rapportserie 10/2025, 123 pp. . https://www.vetinst.no/rapporter-og-publikasjoner/rapporter/2025/villfiskrapporten-2024/_/attachment/inline/eeec2c12-bc26-4039-9788-ddd2136ec8de:d5148f17f1c15c36117124d6f6268fa3f7fc2e01/Villfiskrapporten%202024.pdf
Gjedrem, T. (2000). Genetic improvement of cold-water fish species. Aquaculture Research 31, 25-33. https://doi.org/10.1046/j.1365-2109.2000.00389.x
Gjedrem, T. (2010). The first family-based breeding program in aquaculture. Reviews in Aquaculture 2, 2-15. https://doi.org/10.1111/j.1753-5131.2010.01011.x
Gjessing, M.C., Tengs, T., Nilsen, H., Mohammad, S. & Weli, S.C. (2024). The first report of cod gill poxvirus in gills of Atlantic cod ( Gadus morhua L.) suffering from cardiorespiratory disease. J. Fish Dis. e14078, https://doi.org/10.1111/jfd.14078
Glover, K.A. (2010). Forensic identification of fish farm escapees: the Norwegian experience. Aquaculture Environment Interactions 1, 1-10. https://doi.org/10.3354/aei00002
Glover, K.A., Dahle, G., Westgaard, J.I., Johansen, T., Knutsen, H. & Jørstad, K.E. (2010). Genetic diversity within and among Atlantic cod ( Gadus morhua ) farmed in marine cages: a proof-of-concept study for the identification of escapees. Animal Genetics 41, 515-22. https://doi.org/10.1111/j.1365-2052.2010.02025.x
Glover, K.A., Dahle, G. & Jørstad, K.E. (2011). Genetic identification of farmed and wild Atlantic cod, Gadus morhua , in coastal Norway. ICES Journal of Marine Science 68, 901-10. https://doi.org/10.1093/icesjms/fsr048
Glover K.A., Pertoldi C., Besnier F., Wennevik V., Kent M. & Skaala Ø. (2013). Atlantic salmon populations invaded by farmed escapees: quantifying genetic introgression with a Bayesian approach and SNPs. BMC Genetics 14, 74. https://doi.org/10.1186/1471-2156-14-74
Glover K.A., Solberg M.F., McGinnity P., Hindar K., Verspoor E., Coulson M.W., Hansen M.M., Araki H., Skaala Ø. & Svåsand T. (2017). Half a century of genetic interaction between farmed and wild Atlantic salmon: status of knowledge and unanswered questions. Fish and Fisheries 18, 890-927. https://doi.org/10.1111/faf.12214
Glover K.A., Urdal K., Næsje T., Skoglund H., Florø-Larsen B., Otterå H., Fiske P., Heino M., Aronsen T., Sægrov H., Diserud O., Barlaup B.T., Hindar K., Bakke G., Solberg I., Lo H., Karlsson S., Skaala Ø., Lamberg A., Kanstad-Hanssen Ø., Muladal R., Skilbrei O.T. & Wennevik V. (2019). Domesticated escapees on the run: the second-generation monitoring program reports the numbers and proportions of farmed Atlantic salmon in >200 rivers annually. ICES Journal of Marine Science 76, 1151-1161. https://doi.org/10.1093/icesjms/fsy207
Glover K.A., Wennevik V., Hindar K., Skaala O., Fiske P., Solberg M.F., Diserud O.H., Svåsand T., Karlsson S., Andersen L.B. & Grefsrud E.S. (2020). The future looks like the past: Introgression of domesticated Atlantic salmon escapees in a risk assessment framework. Fish and Fisheries 21, 1077-1091. https://doi.org/10.1111/faf.12478
Glover, K.A., Castelliani, M., Heino, M. & Besnier, F. (2025a). Modelling the consequences of domestication-introgression in wild populations using genetic markers under varying degrees of selection. Evolutionary Applications 18, e70140. https://doi.org/10.1111/eva.70140
Glover, K., Quintela Sanchez, M., van der Meeren, T., Mateos Rivera, A., Eide Sørvik, A.G. & Stöger, E. (2025b). Genetisk identifikasjon av torsk og torskeegg etter en rømmingshendelse i Voldsfjorden i 2022. Rapport fra Havforskningen nr. 2025-24. 13 pp. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-2025-24
Godø, O.R. & Sunnanå, K. (1984). Spawning area and distribution of 0-group cod ( Gadus morhua L.), on the Møre coast. In (Eds) Dahl, E., Danielssen, D. S., Moksness, E., & Solemdal, P., The Propagation of Cod ( Gadus morhua L.). Flødevigen rapportser. 1, 519-526.
Godø, O.R. (1995). Transplantation-tagging-experiments in preliminary studies of migration of cod off Norway. ICES Journal of Marine Science 52, 955–962. https://doi.org/10.1006/jmsc.1995.0091
Gudding, R. & Lillehaug, A. (red.) (2018). Smitte mellom oppdrettsfisk og villfisk: Kunnskapsstatus og risikovurdering. Veterinærinstituttet Rapport 12, 107 pp. https://www.vetinst.no/rapporter-og-publikasjoner/rapporter/2018/smitte-mellom-oppdrettsfisk-og-villfisk-kunnskapsstatus-og-risikovurdering
Hansen, J.Ø., Berge, G.M., Hillestad, M., Krogdahl, Å., Galloway, T.F., Holm, H., Holm, J. & Ruyter, B. (2008). Apparent digestion and apparent retention of lipid and fatty acids in Atlantic cod ( Gadus morhua ) fed increasing dietary lipid levels. Aquaculture 284, 159–166. https://doi.org/10.1016/j.aquaculture.2008.07.043
Hansen, L.A., Dale, T., Damsgård, B. Uglem, I., Aas, K. & Bjørn, P.‐A. (2009). Escape‐related behaviour of Atlantic cod, Gadus morhua L., in a simulated farm situation. Aquaculture Research 40, 26-34. https://doi.org/10.1111/j.1365-2109.2008.02057.x
Hansen, T.J., Fjelldal, P.G., Folkedal, O., Vågseth, T. & Oppedal, F. (2017). Effects of light source and intensity on sexual maturation, growth and swimming behavior of Atlantic salmon in sea cages. Aquacult Environ Interact 9, 193-204. https://doi.org/10.3354/aei00224
Hansen, A.C. & Hemre, G.I. (2013). "Effects of replacing fish meal and oil with plant resources in on-growing diets for Atlantic cod Gadus morhua L." Aquaculture Nutrition 19, 641-650. https://doi.org/10.1111/anu.12078
Hara T.J. (1994). The diversity of chemical stimulation in fish olfaction and gustation. Rev. Fish. Biol. Fish. 4, 1-35. https://doi.org/10.1007/BF00043259
Haugland, M. & Jensen, S.-K. (2025). Evaluering av tilsynssatsing rettet mot modning og gyting i merd hos oppdrettstorsk sesongen 2024-2025. Evalueringsrapport, 18 pp. https://www.fiskeridir.no/nyheter/fortsatt-risiko-for-gyting-hos-oppdrettstorsk-i-merd
Hawley, L.M. & Garver, K.A. (2008). Stability of viral hemorrhagic septicemia virus (VHSV) in freshwater and seawater at various temperatures. Diseases of Aquatic Organisms 82, 171-178. https://doi.org/10.3354/dao01998
Hedger, R.D., Serra-Llinares, R.M., Arechavala-Lopez, P., Nilsen, R., Bjørn, P.A. & Uglem, I. (2017). Tracking escaped Atlantic cod ( Gadus morhua L.) aggregated around Norwegian farms: Considerations for management strategies. Fish. Manag. Ecol. 24, 265-273. https://doi.org/10.1111/fme.12219
Heino M., Svåsand T., Wennevik V. & Glover K.A. (2015). Genetic introgression of farmed salmon in native populations: quantifying the relative influence of population size and frequency of escapees. Aquaculture Environment Interactions 6, 185-190. https://doi.org/10.3354/aei00126
Hellberg, H., Colquhoun, D., Nilsen, H. & Jensen, B.B. (2011). Helsesituasjonen hos marin fisk 2010. In: Hellberg, H. (red.) Fiskehelserapporten 2010, Veterinærinstituttet, s 26-34. https://www.vetinst.no/rapporter-og-publikasjoner/rapporter/2011/fiskehelserapporten-2010
Helmerson, C., Durant, J.M., Hoff, S.N.K., Brieuc, M. S.O., Berg, P.R., Jorde, P.E., Jentoft, S. (2025). A century of anthropogenic perturbations impact genomic signatures of the iconic migratory Atlantic cod. Science Advances, 11(31), eadp3342. https://doi.org/10.1126/sciadv.adp3342
Henriksen, E., Heide, M., Hansen, Ø.J. & Mortensen, A. (2018). Kunnskaps- og erfaringsgrunnlag for torskeoppdrett. Nofima Rapport 23/2018. 127 pp. https://nofima.no/wp-content/uploads/2018/10/Rapport-23-2018-Kunnskaps-og-erfaringsgrunnlag-for-torskeoppdrett.pdf
Heuch, P.A., Jansen, P.A., Hansen, H., Sterud, E., Mackenzie, K., Haugen, P. & Hemmingsen, W. (2011). Parasite faunas of farmed cod and adjacent wild cod populations in Norway: a comparison. Aquaculture Environment Interactions, 2, 1-13. https://doi.org/10.3354/aei00027
Hindar K., Fleming I.A., McGinnity P. & Diserud A. (2006). Genetic and ecological effects of salmon farming on wild salmon: modelling from experimental results. ICES Journal of Marine Science 63, 1234-1247. https://doi.org/10.1016/j.icesjms.2006.04.025
Holt, R.E., Bogstad, B., Durant, J.M., Dolgov, A.V. & Ottersen, G. (2019). Barents Sea cod ( Gadus morhua ) diet composition: long-term interannual, seasonal, and ontogenetic patterns. ICES Journal of Marine Science 76, 1641–1652. https://doi.org/10.1093/icesjms/fsz082
Humborstad, O.-B., Utne-Palm, A.C., Breen, M., Løkkeborg, S., (2018). Artificial light in baited pots substantially increases the catch of cod ( Gadus morhua ) by attracting active bait, krill ( Thysanoessa inermis ). ICES J. Mar. Sci. 75, 2257–2264. https://doi.org/10.1093/icesjms/fsy099
ICES (2016) Technical Guidelines - Definitions of stock status. ICES Technical Guidelines. ICES Technical Guidelines 2016. https://ices-library.figshare.com/ndownloader/files/34123176
ICES (2021). Benchmark Workshop for Barents Sea and Faroese Stocks (WKBARFAR 2021). ICES Scientific Reports 3, 205 pp. https://ices-library.figshare.com/ndownloader/files/33397481
ICES (2022a). Norwegian request for evaluation of management of Norwegian coastal cod north of 67° N. I Report of the ICES Advisory Committee, 2022. ICES Advice 2022, sr.2022.08, 5 pp. https://ices-library.figshare.com/ndownloader/files/35770520
ICES (2022b). Workshop on the evaluation of northern Norwegian coastal cod harvest control rules (WKNCCHCR). ICES Scientific Reports 4, 115 pp. https://ices-library.figshare.com/ndownloader/files/35775509
ICES (2025a). Cod ( Gadus morhua ) in subareas 1 and 2 north of 67°N (Norwegian Sea and Barents Sea), northern Norwegian coastal cod. In Report of the ICES Advisory Committee, 2025. ICES Advice 2025, cod.27.1-2coastN, https://doi.org/10.17895/ices.advice.27202557
ICES (2025b). Cod ( Gadus morhua ) in Subarea 2 between 62°N and 67°N (Norwegian Sea), southern Norwegian coastal cod. In Report of the ICES Advisory Committee, 2025. ICES Advice 2025, cod.27.2.coastS. 7 pp. https://doi.org/10.17895/ices.advice.28194458
ICES (2025c). Arctic fisheries Working Group (AFWG). ICES Scientific Reports. 7 (66). 339 pp. https://doi.org/10.17895/ices.pub.29234186
Iversen, S.A. & Danielsen, D.S. (1984). Development and mortality of cod ( Gadus morhua L.), eggs and larvae in different temperatures. In: (Eds) Dahl, E., Danielssen, D.S., Moksness, E., & Solemdal, P., The Propagation of Cod ( Gadus morhua L.). Flødevigen rapportser. 1, 49-65.
Izquierdo, M.S., Fernández-Palacios, H. & Tacon, A.G.J. (2001). Effect of broodstock nutrition on reproductive performance of fish. Aquaculture 197, 25-42. https://doi.org/10.1016/S0044-8486(01)00581-6
Jakobsen, T. (1987). Coastal cod in northern Norway. Fish. Res. 5, 223–234. https://doi.org/10.1016/0165-7836(87)90042-7
Jensen, I., Seppola, M., Steiro, K., Sandaker, E., Mennen, S. & Sommer, A.I. (2009). Susceptibility of Atlantic cod Gadus morhua juveniles to different routes of experimental challenge with infectious pancreatic necrosis virus (IPNV). Diseases of Aquatic Organisms, 85, 105-113. https://doi.org/10.3354/dao02066
Johansen, R., Bergh, Ø., Modahl, I., Dahle, G., Gjerset, B., Holst, J.C. & Sandlund, N. (2013). High prevalence of viral haemorrhagic septicaemia virus (VHSV) in Norwegian spring-spawning herring. Marine Ecology Progress Series, 478, 223-230. https://doi.org/10.3354/meps10208
Johansen, S.D., Coucheron, D.H., Andreassen, M., Karlsen, B.O., Furmanek, T., Jørgensen, T.E., Emblem, Å., Breines, R., Nordeide, J.T., Moum, T., Nederbragt, A.J., Stenseth, N.C. & Jakobsen, K.S. (2009). Large-scale sequence analyses of Atlantic cod. New Biotechnology 25, 263-271. https://doi.org/10.1016/j.nbt.2009.03.014
Johansen, T., Besnier, F., Quintela, M., Jorde, P.E., Glover, K.A. Westgaard, J.-I., Dahle, G., Lien, S. & Kent, M.P. (2020). Genomic analysis reveals neutral and adaptive patterns that challenge the current management regime for East Atlantic cod Gadus morhua L. Evolutionary Applications 13, 2673-2688. https://doi.org/10.1111/eva.13070
Johansen, T., Westgaard, J.-I., Seliussen, B.B., Nedreaas, K., Dahle, G., Glover, K.A., Kvalsund, R., Aglen, A. (2018). “Real-time” genetic monitoring of a commercial fi shery on the doorstep of an MPA reveals unique insights into the interaction between coastal and migratory forms of the Atlantic cod. ICES Journal of Marine Science 75, 1093-1104, https://doi.org/10.1093/icesjms/fsx224
Jorde, P.E., Knutsen, H., Espeland, S.H., Stenseth, N.C. (2007). Spatial scale of genetic structuring in coastal cod Gadus morhua and geographic extent of local populations. Marine Ecology Progress Series 343, 229-237. https://doi.org/10.3354/meps06922
Jorde, P.E., van der Meeren, T., Quintela, M., Dahle, G., Mateos-Rivera, A., Aase, M., Norberg, B., Næverlid Sævik, P., Bjørn, P.A. & Glover, K.A. (2024). Genetic analyses verify sexually mature escaped farmed Atlantic cod and farmed cod eggs in the natural environment. Evolutionary Applications 17, e13688 https://onlinelibrary.wiley.com/doi/full/10.1111/eva.13688
Jorde, P. E., Synnes, A.-E., Espeland, S. H., Sodeland, M. & Knutsen, H. (2018). Can we rely on selected genetic markers for population identification? Evidence from coastal Atlantic cod. Ecology & Evolution 8, 12547–12558. https://doi.org/10.1002/ece3.4648
Jorde, P.E., Huserbråten, M.B.O., Seliussen, B.B., Myksvoll, M.S., Vikebø, F.B., Dahle, G., Aglen, A. & Johansen, T. (2021). The making of a genetic cline: introgression of oceanic genes into coastal cod populations in the Northeast Atlantic. Canadian Journal of Fisheries and Aquatic Sciences 78, 958-968. https://cdnsciencepub.com/doi/abs/10.1139/cjfas-2020-0380
Jung, K.-M., Folkvord, A., Kjesbu, O.S., Agnalt, A.L., Thorsen, A. & Sundby, S. (2012). Egg buoyancy variability in local populations of Atlantic cod ( Gadus morhua ). Marine Biology 159, 1969-1980. https://doi.org/10.1007/s00227-012-1984-8
Jørstad, K.E., Skaala, Ø. & Dahle, G. (1991). The development of biochemical and visible genetic markers and their potential use in evaluating interactions between cultured and wild fish populations. ICES Mar. Sci. Symp. 192, 200-205. https://ices-library.figshare.com/ndownloader/files/55980086
Jørstad, K.E., Otterå, H., van der Meeren, T., Dahle, G., Paulsen, Bakke, G. & Svåsand, T. (2014). Genetic marking of farmed Atlantic cod ( Gadus morhua L.) and detection of escapes from a commercial cod farm. ICES Journal of Marine Science 71, 574-584. https://doi.org/10.1093/icesjms/fst171
Jørstad, K.E., van der Meeren, T., Paulsen, O.I., Thomsen, T., Thorsen, A. & Svåsand, T. (2008). “Escapes” of eggs from farmed cod spawning in net pens: recruitment to wild stocks. Reviews in Fisheries Science 16, 285–295. https://doi.org/10.1080/10641260701678017
Karlsbakk, E., Otterlei E., Høie, H. & Nylund, A. (2001). Parasites of cultured cod ( Gadus morhua ) postlarvae fed natural zooplankton. Bull. Eur. Ass. Fish Pathol. 21, 63-70. https://eafp.org/download/2001-Volume21/Issue%202/21_063.pdf
Karlsbakk, E., Isaksen, T.E., Ottem, K.F., Nylund, A., Korsnes, K., Nerland A. mfl. (2008). 1.10 Viktige patogener i kystsonen. Fisken & Havet (Kyst og havbruk), 48-51. https://hdl.handle.net/11250/114125
Karlsbakk, E., Isaksen, T.E., Nylund, A., Devold, M., Korsnes, K., Nylund, S., Ottem, K.F., Brevik, Ø., Andersen, L., Karlsen, M., Jørstad, K.E., Sleire, L., Johansen, L., Ranasinghe, M.P.N., Handeland, S.O., (2008). CodPath prosjektet: kartlegging av sykdomsagens hos villtorsk - en kort presentasjon. Fiskehelse 10, 10-18.
Karlsbakk, E., Isaksen, T.E. & Hamre, L.A. (2009). 1.10 Hva vet vi om parasitter og oppdrett av torsk? Fisken og Havet (Kyst og havbruk 2009), 50-53. https://imr.brage.unit.no/imr-xmlui/handle/11250/113972
Karlsbakk, E., mfl. (2010). Smittespredning ved francisellose hos torsk. Fisken & Havet 2010 (Havforskningsrapporten), 101-102. https://imr.brage.unit.no/imr-xmlui/handle/11250/113966
Karlsbakk, E., Nystøyl, C.F., Plarre, H., Nylund, A. (2021). A novel protist parasite, Salmoxcellia vastator n. gen., n. sp. (Xcelliidae, Perkinsozoa), infecting farmed salmonids in Norway. Parasites Vectors (2021) 14:431 https://doi.org/10.1186/s13071-021-04886-0
Karlsen, Ø. & van der Meeren, T. (2013). Kunnskapsstatus - plassering av oppdrettsanlegg og mulige interaksjoner med gytefelt og oppvekstområder for marin fisk og vandringsruter for laks. Fisken og Havet 6-2013, 47 pp. https://www.hi.no/hi/nettrapporter/fisken-og-havet/2013/fh_6-2013_plassering_oppdrettsanlegg_til_web
Karlsen, Ø., van der Meeren, T., Rønnestad, I., Mangor-Jensen, A., Galloway, T.F., Kjørsvik, E. & Hamre, K. (2015). Copepods enhance nutritional status, growth and development in Atlantic cod ( Gadus morhua L.) larvae – can we identify the underlying factors? PeerJ 3, e902. https://doi.org/10.7717/peerj.902
Karlsen, Ø., Taranger, G.L., Dahle, R., & Norberg, B. (2000). Effect of exercise and continuous light on early sexual maturation in cod. In Proceedings of the 6th International Symposium on the Reproductive Physiology of Fish Bergen, July 4e9, 1999, pp. 328-330. (Eds) Norberg, B. Kjesbu, O. S., Taranger, G. L., Andersson, E., & Stefansson. S. O. John Grieg, Bergen. 499 pp.
Karlsson S., Diserud O.H., Fiske P. & Hindar K. (2016). Widespread genetic introgression of escaped farmed Atlantic salmon in wild salmon populations. ICES Journal of Marine Science 73, 2488-2498. https://doi.org/10.1093/icesjms/fsw121
Kennedy, D.A., Kurath, G., Brito, I.L., Purcell, M.K., Read, A.F., Winton, J.R., & Wargo, A.R. (2016). Potential drivers of virulence evolution in aquaculture. Evolutionary Applications, 9, 344-354. https://doi.org/10.1111/eva.12342
Khan, R.A. (2009). Parasites causing disease in wild and cultured fish in Newfoundland. Icelandic Agricultural Sciences, 29-35. http://ias.is/wp-content/uploads/Icelandic_Agricultural_Sciences_22_2009/Parasites-causing-disease-in-wild-and-cultured-fish.pdf
King, A.M (2023). The sea louse Caligus elongatus (Caligidae). Hosts susceptibility to the two genotypes. MSc oppgave i fiskehelse, Univ. Bergen. 48 sider. https://hdl.handle.net/11250/2761777
Kirubakaran, T.G., Grove, H., Kent, M.P., Sandve, S.R., Baranski, M., Nome, T., De Rosa, M.C., Righino, B., Johansen, T., Otterå, H., Sonesson, A., Lien, S. & Andersen, Ø. (2016). Two adjacent inversions maintain genomic differentiation between migratory and stationary ecotypes of Atlantic cod. Mol. Ecol. 25, 2130–2143. https://doi.org/10.1111/mec.13592
Kjesbu, O.S., Solemdal, P., Bratland, P. & Fonn, M. (1996). Variation in annual egg production in individual captive Atlantic cod ( Gadus morhua ). Canadian Journal of Fisheries and Aquatic Sciences, 53, 610-620. https://doi.org/10.1139/cjfas-53-3-610
Kjesbu, O.S., Righton, D., Krüger-Johnsen, M. Thorsen, A., Michalsen, K., Fonn, M., & Witthames, P.R. (2010). Thermal dynamics of ovarian maturation in Atlantic cod ( Gadus morhua ). Canadian Journal of Fisheries and Aquatic Sciences 67, 605-625. https://doi.org/10.1139/F10-011https://doi.org/10.1111/eva.12640
Knutsen, H. Olsen, E.M., Ciannelli, L., Espeland, S.H., Knutsen, J.A., Simonsen, J.H., Skreslet, S. & Stenseth, N.C. (2007). Egg distribution, bottom topography and small-scale cod population structure in a coastal marine system. Mar. Ecol. Prog. Ser. 333, 249–255. https://doi.org/10.3354/meps333249
Knutsen, H., Olsen, E.M., Jorde, P.E., Espeland, S.H., André, C. & Stenseth, N.C. (2011). Are low but statistically significant levels of genetic differentiation in marine fishes ‘biologically meaningful’? A case study of coastal Atlantic cod. Molecular Ecololgy 20, 768–783. https://doi.org/10.1111/j.1365-294X.2010.04979.x
Knutsen, H., Jorde, P.E., Hutchings, J.A., Hemmer-Hansen, J., Grønkjær, P., Jørgensen, K.-E.M., André, C., Sodeland, M., Albretsen, J. & Olsen, E.M. (2018). Stable coexistence of genetically divergent Atlantic cod ecotypes at multiple spatial scales. Evolutionary Applications 11, 1527–1539. https://doi.org/10.1111/eva.12640
Korsøen, Ø.J., Dempster, T., Fosseidengen, J.E., Karlsen, Ø., Oppedal, F., Stien, L.H. & Kristiansen, T.S. (2013). Towards cod without spawning: artificial continuous light in submerged sea-cages maintains growth and delays sexual maturation for farmed Atlantic cod Gadus morhua . Aquaculture Environmental Interactions 3, 245-255. https://doi.org/10.3354/aei00067
Kristiansen, T.S., Jørstad, K.E., Otterå, H., Paulsen, O.I. & Svåsand. T. (1997). Estimates of larval survival of cod by releases of genetically marked yolk-sac larvae. Journal of Fish Biology 51, 264-283. https://doi.org/10.1111/j.1095-8649.1997.tb06103.x
Kvalvik, R. (2025) Histologisk studie av tarm og hjerte hos oppdrettstorsk Gadus morhua med tarmlidelser. MSc oppgave I fiskehelse, Univ. Bergen https://hdl.handle.net/11250/3215468
Kvenseth, P.G. & Øiestad, V. (1984). Large-scale rearing of cod fry on the natural food production in an enclosed pond. In: Dahl, E., Danielssen, D. S., Moksness, E., & Solemdal, P. (Eds). The propagation of cod Gadus morhua L. Flødevigen rapportser. 1, 645-656.
Lambert, Y. & Dutil, J.D. (2000). Energetic consequences of reproduction in Atlantic cod ( Gadus morhua ) in relation to spawning level of somatic energy reserves. Canadian Journal of Fisheries and Aquatic Sciences 57, 815-825. https://doi.org/10.1139/f00-022
Lambert, Y. & Dutil, J.-D. (1997). Can simple condition indices be used to monitor and quantify seasonal changes in the energy reserves of Atlantic cod ( Gadus morhua )? Canadian Journal of Fisheries and Aquatic Sciences 54, 104-112. https://doi.org/10.1139/f96-149
Lange, E., Johannessen, J.V., (1977). Histochemical and ultrastructural studies of chemodectoma-like tumors in the cod ( Gadus morhua L.). Laboratory Investigation. 37, 96-104.
Link, J.S. & Garrison, L.P. (2002). Trophic ecology of Atlantic cod Gadus morhua on the northeast US continental shelf. Marine Ecology Progress Series 227, 109-123. https://doi.org/10.3354/meps227109
Makhotin, V., Solemdal, P., Korsbrekke, K. & Salthaug, A. (2001). Types and frequency of malformations and mortality in eggs of Arcto-Norwegian cod: A field study. ICES CM 2001/N:12, 17 pp. https://files.core.ac.uk/download/pdf/225944019.pdf
Marshall, C.T., Yaragina, N.A., Lambert, Y. & Kjesbu, O.S. (1999). Total lipid energy as a proxy for total egg production by fish stocks. Nature 402, 288-290. https://doi.org/10.1038/46272
Matschiner, M, Barth, J.M.I., Tørresen, O.K., Star, B., Baalsrud, H.T., Brieuc, M.S.O., Pampoulie, C., Bradbury, I., Jakobsen, K.S. & Jentoft, S. (2022). Super gene origin and maintenance in Atlantic cod. Nature Ecology and Evolution 6, 469–481. https://doi.org/10.1038/s41559-022-01661-x
Maurstad, A., Dale, T., & Bjørn, P.A. (2007). You wouldn’t spawn in a septic tank, would you? Hum. Ecol. 35, 601–610. https://doi.org/10.1007/s10745-007-9126-5
McConnell, A., Routledge, R., Connors, B.M. (2010). Effect of artificial light on marine invertebrate and fish abundance in an area of salmon farming. Marine Ecology Progress Series 419, 147–156. https://doi.org/10.3354/meps08822
McGinnity, P., Prodohl, P., Ferguson, K., Hynes, R., O'Maoileidigh, N., Baker, N., Cotter, D., O'Hea, B., Cooke, D., Rogan, G., Taggart, J. & Cross, T. (2003). Fitness reduction and potential extinction of wild populations of Atlantic salmon, Salmo salar , as a result of interactions with escaped farm salmon. Proceedings of the Royal Society of London Series B-Biological Sciences 270, 2443-2450. https://doi.org/10.1098/rspb.2003.2520
McGinnity, P., Stone, C., Taggart, J.B., Cooke, D., Cotter, D., Hynes, R., McCamley, C., Cross, T. & Ferguson, A. (1997). Genetic impact of escaped farmed Atlantic salmon ( Salmo salar L.) on native populations: use of DNA profiling to assess freshwater performance of wild, farmed, and hybrid progeny in a natural river environment. ICES Journal of Marine Science 54, 998-1008. https://doi.org/10.1016/S1054-3139(97)80004-5
Meager, J.J., Skjæraasen, J.E., Fernö, A., Karlsen, Ø., Løkkeborg, S., Michalsen, K. & Utskot, S.O. (2009). Vertical dynamics and reproductive behaviour of farmed and wild cod Gadus morhua . Marine Ecology Progress Series 389, 233-243. https://doi.org/10.3354/meps08156
Meager, J.J., Skjæraasen, J.E., Fernö, A. & Løkkeborg, S. (2010). Reproductive interactions between fugitive farmed and wild cod ( Gadus morhua ) in the field. Canadian Journal of Fisheries and Aquatic Sciences 67, 1221-1231. https://doi.org/10.3354/meps08670
Meier, S., van der Meeren, T., Skjæraasen, J.E., Bannister, R.J., Rasinger, J.D. & Karlsen, Ø. (2023). Terrestrial fatty acids from feed oil in feed for farmed salmonids are transferred to the liver, gonads, and muscle of wild Atlantic cod (Gadus morhua). ICES Journal of Marine Science 80, 1143-1154. https://doi.org/10.1093/icesjms/fsad051
Michalsen, K., Johansen, T., Subbey, S., Beck, A. (2014). Linking tagging technology and molecular genetics to gain insight in the spatial dynamics of two stocks of cod in Northeast Atlantic waters. ICES Journal of Marine Science 71, 1417-1432. https://doi.org/10.1093/icesjms/fsu083
Mikkelsen, H., Lund, V., Martinsen, L.C., Gravningen, K. & Schrøder, M.B. (2007). Variability among Vibrio anguillarum O2 isolates from Atlantic cod ( Gadus morhua L.): Characterisation and vaccination studies. Aquaculture 266, 16-25. https://doi.org/10.1016/j.aquaculture.2007.02.041
Moe, H., Dempster, T., Sunde, L.M., Winther, U. & Fredheim, A. (2007). Technological solutions and operational measures to prevent escapes of Atlantic cod ( Gadus morhua ) from sea cages. Aquaculture Research 38, 91-99. https://doi.org/10.1111/j.1365-2109.2006.01638.x
Moldal, T., Wiik-Nielsen, J., Oliveira, V.H.S., Svendsen, J.C. & Sommerset, I. (2025). Fiskehelserapporten 2024. Veterinærinstituttets rapportserie nr. 1a/2025, 234 pp. https://www.vetinst.no/rapporter-og-publikasjoner/rapporter/2025/fiskehelserapporten-2024
Munang’andu, H.M., Mutoloki, S. & Evensen, Ø. (2016). Birnaviruses of Aquatic Organisms. Aquaculture Virology, 237-250. https://doi.org/10.1016/B978-0-12-801573-5.00015-2
Myksvoll, M.S., Jung, K.M., Albretsen, J. & Sundby, S. (2014). Modelling dispersal of eggs and quantifying connectivity among Norwegian coastal cod subpopulations. ICES Journal of Marine Science 71, 957-969. https://doi.org/10.1093/icesjms/fst022
Myksvoll, M.S., Sundby, S., Ådlandsvik, B. & Vikebø F.B. (2011). Retention of coastal cod eggs in a fjord caused by interactions between egg buoyancy and circulation pattern. Marine and Coastal Fisheries 3, 279-294. https://doi.org/10.1080/19425120.2011.595258
Nardi, G., Prickett, R., van der Meeren, T., Boyce, D. & Moir, J. (2021). Atlantic cod aquaculture: Boom, bust, and rebirth? Journal of World Aquaculture Society 52, 672-690. https://www.doi.org/10.1111/jwas.12811
Nedreaas, K. (2006). Kysttorsk nord for Stad. Artsdatabankens faktaark nr. 87. https://www2.artsdatabanken.no/faktaark/Faktaark87.pdf
Neuenfeldt, S. Righton, D., Neat, F., Wright, P. J., Svedäng, H., Michalsen, K., Subbey, S., Steingrund, P., Thorsteinsson, V., Pampoulie, C., Andersen, K.H., Pedersen, M.W. & Metcalfe J. (2013). Analysing migrations of Atlantic cod Gadus morhua in the north‐east Atlantic Ocean: then, now and the future. J. Fish Biol. 82, 741–763. https://doi.org/10.1111/jfb.12043
Norberg, B., Brown, C.L., Halldorsson, O., Stensland, K. & Björnsson, B.T. (2004). Photoperiod regulates the timing of sexual maturation, spawning, sex steroid and thyroid hormone profiles in the Atlantic cod ( Gadus morhua ). Aquaculture 229, 451-467. https://doi.org/10.1016/S0044-8486(03)00393-4
Norberg, B., Kleppe, L., Andersson, E., Thorsen, A., Rosenlund, G. & Hamre, K. (2017). Effects of dietary arachidonic acid on the reproductive physiology of female Atlantic cod ( Gadus morhua L.). General and Comparative Endocrinology 250, 21-35. https://doi.org/10.1016/j.ygcen.2017.05.020
Nordeide, J.T. & Båmstedt, U. (1998). Coastal cod and north-east Arctic cod - do they mingle at the spawning grounds in Lofoten? Sarsia 83, 373–379. https://doi.org/10.1080/00364827.1998.10413696
Nordeide, J.T. & Folstad, I. (2000). Is cod lekking or a promiscuous group spawner? Fish and Fisheries 1, 90–93. https://doi.org/10.1046/j.1467-2979.2000.00005.x
Nygaard, S.M.R. (2005). Skader av lus på torsk i merd. Norsk Veterinærtidsskrift 117, 34.
Nylund, A., Karlsbakk, E., Nylund, S. Isaksen, T.E., Karlsen, M., Korsnes, K., Handeland, S., Martinsen, R., Mork Pedersen T. & Ottem K.F. (2008). New clade of betanodaviruses detected in wild and farmed cod ( Gadus morhua ) in Norway. Arch. Virol. 153, 541–547. https://doi.org/10.1007/s00705-007-0015-4
Nylund, A., Ottem, K.F., Watanabe, K., Karlsbakk, E. & Krossøy, B. (2006). Francisella sp. (Family Francisellaceae) causing mortality in Norwegian cod ( Gadus morhua ) farming. Archives of Microbiology 185, 383-392. https://doi.org/10.1007/s00203-006-0109-5
Olesiuk, P.F., Lawson, J.W. & Trippel, E.A. (2010). Pathways of effects of noise associated with aquaculture on natural marine ecosystems in Canada. Canadian Science Advisory Secretariat Research Document 2010/017, 70. https://waves-vagues.dfo-mpo.gc.ca/Library/345630.pdf
Olsen, E.M., Knutsen, H., Gjøsæter, J., Jorde, P.E., Knutsen, J.A. and Stenseth, N.C. (2004). Life‐history variation among local populations of Atlantic cod from the Norwegian Skagerrak coast. Journal of Fish Biology 64, 1725-1730. https://doi.org/10.1111/j.0022-1112.2004.00402.x
Omdal, L.M. (2009). Effekter av temperatur og alder ved badsmitte med Francisella noatunensis (isolat GM 2212) på tidlige stadier av torsk, University of Bergen. Bergen. 83 pp.
Ottem, K.F., Nylund, A., Isaksen, T.E., Karlsbakk, E. & Bergh, Ø. (2008). Occurrence of Francisella piscicida in farmed and wild Atlantic cod, Gadus morhua L., in Norway. J. Fish Dis. 31, 525-534. https://doi.org/10.1111/j.1365-2761.2008.00930.x
Otterå H., Agnalt A.L., Thorsen A., Kjesbu O.S., Dahle G. & Jørstad K. (2012). Is spawning time of marine fish imprinted in the genes? A two-generation experiment on local Atlantic cod ( Gadus morhua L.) populations from different geographical regions. ICES Journal of Marine Science 69, 1722-8. https://doi.org/10.1093/icesjms/fss135
Otterå, H., Heino, M., Sorvik, A. G.E., Svåsand, T., Karlsen, O., Thorsen, A., & Glover, K.A. (2018). Growth of wild and domesticated Atlantic cod Gadus morhua reared under semi-commercial conditions. Aquaculture Environment Interactions 10, 187-200. https://doi.org/10.3354/aei00262
Otterå, H., Jørstad, K.E., Svåsand, T. & Kristiansen, T.S. (1999). Migration patterns and recapture rates of North‐east Arctic and Norwegian coastal cod reared and released under similar conditions. Journal of Fish Biology 54, 213–217. https://doi.org/10.1111/j.1095-8649.1999.tb00625.x
Otterå, H. & Skilbrei, O.T. (2014). Possible influence of salmon farming on long-term resident behaviour of wild saithe ( Pollachius virens L.). ICES Journal of Marine Science 71, 2484–2493. https://doi.org/10.1093/icesjms/fsu096
Ouellet, P., Lambert, Y. & Bérubé, I. (2001). Cod egg characteristics and viability in relation to low temperature and maternal nutritional condition. ICES Journal of Marine Science 58, 672-686. https://doi.org/10.1006/jmsc.2001.1065
Paulsen, N. (2018). Skottelus (Caligus elongatus). En litteraturgjennomgang. MSc thesis, Dept. Bioscienses, Univ. Bergen. 53 pp.
Pedersen, M.W. & Berg, C.W. (2017). A stochastic surplus production model in continuous time. Fish and Fisheries 18, 226-243. https://doi.org/10.1111/faf.12174
Pedersen, T., Cuveliers, E.L. & Berg, E. (2008). Spatial scales of movement in Northeast Atlantic coastal cod. Reviews in Fisheries Science 16, 348–356. https://doi.org/10.1080/10641260701678462
Persson, H.L. (2021). The sea louse Caligus elongatus (Caligidae). Genetic variation and host use by its two genotypes. MSc thesis, Dept. Bioscienses, Univ. Bergen. 71 pp.
Pickova, J., Dutta, P.C., Larsson, P.-O. & Kiessling, A. (1997). Early embryonic cleavage pattern, hatching success, and egg-lipid fatty acid composition: comparision between two cod ( Gadus morhua ) stocks. Canadian Journal of Fisheries and Aquatic Sciences 54, 2410-2416. https://doi.org/10.1139/f97-148
Puvanendran, V., Mortensen, A., Johansen, L.-H., Kettunen, A., Hansen, Ø.J. Henriksen, E. & Heide, M. (2021). Development of cod farming in Norway: Past and current biological and market status and future prospects and directions. Reviews in Aquaculture 14, 308-342. https://doi.org/10.1111/raq.12599
Rogers, L.A., Olsen, E.M., Knutsen, H. & Stenseth, N.C. (2014). Habitat effects on population connectivity in a coastal seascape. Mar. Ecol. Prog. Ser. 511, 153–163. https://doi.org/10.3354/meps10944
Rognerud, C. (1887). Hatching cod in Norway. Bulletin of the United States Fish Commission 7(8): 113-119. https://spo.nmfs.noaa.gov/sites/default/files/pdf-content/fish-bull/fb7.38.pdf
Rowe, S., Hutchings, J.A., (2008). A link between sound producing musculature and mating success in Atlantic cod. Journal of Fish Biology 72, 500–511. https://doi.org/10.1111/j.1095-8649.2007.01713.x
Røjbek, M.C., Støttrup, J.G., Jacobsen, C., Tomkiewicz, J., Nielsen, A. & Trippel, E.A. (2014). Effects of dietary fatty acids on the production and quality of eggs and larvae of Atlantic cod ( Gadus morhua L.). Aquaculture Nutrition 20, 654-666. https://doi.org/10.1111/anu.12124
Salinas, S. & Munch, S.B. (2012). Thermal legacies: transgenerational effects of temperature on growth in a vertebrate. Ecology Letters 15, 159-163. https://doi.org/10.1111/j.1461-0248.2011.01721.x
Salze, G., Tocher, D.R., Roy, W.J. & Robertson, D.A. (2005). Egg quality determinants in cod ( Gadus morhua L.): egg performance and lipids in eggs from farmed and wild broodstock. Aquaculture Research 36, 1488-1499. https://doi.org/10.1111/j.1365-2109.2005.01367.x
Sanchez-Jerez, P., Fernandez-Jover, D., Uglem, I., Arechavala-Lopez, P., Dempster, T., Bayle-Sempere, J., Pérez, C., Izquierdo-Gomez, D., Bjørn, P.-A. & Nilsen, R., (2011). Coastal Fish Farms as Fish Aggregation Devices (FADs), In Bortone, S.A., Pereira, B.F., Fabi, G. & Otake, S. (Eds.), Artificial Reefs in Fisheries Management. CRC Press, Boca Raton, FL, pp. 187–208.
Sanchez-Jerez, P., Atalah, J., Sætra, I.M., Bøhn, T., Sæther, B.-S., Johansen, T., Keeley, N., van der Meeren, T. & Bjørn, P.A. (2025). Effects of salmon farming on Atlantic cod: insights from liver fatty acid composition in relationship with biological and genetic variables. Aquaculture Environment Interactions 17, 149-160. https://doi.org/10.3354/aei00501
Sanchez-Jerez, P., Atalah, J., Sætra, I.M., Bøhn, T., Saether, B.-S., Johansen, T., Keeley, N., van der Meeren, T. & Bjørn, P.A. (2024). Comparative study of fatty acid composition of muscles of Atlantic cod ( Gadus morhua Linnaeus, 1758) with natural diet and feeding near salmon farms. Aquaculture Journal 4, 246-254. https://doi.org/10.3390/aquacj4040018
Sandlund, N., Gjerset, B., Bergh, Ø., Modahl, I., Olesen, N. J. & Johansen, R. (2014). Screening for viral hemorrhagic septicemia virus in marine fish along the Norwegian coastal line. PLoS ONE, 9(9), e108529. https://doi.org/10.1371/journal.pone.0108529
Sandlund, N., Johansen, R., Fiksdal, I.U., Einen, A.C.B., Modahl, I., Gjerset, B., & Bergh, Ø. (2021). Susceptibility and pathology in juvenile Atlantic cod Gadus morhua to a marine viral haemorrhagic septicaemia virus isolated from diseased rainbow trout Oncorhynchus mykiss. Animals 11, 3523. https://doi.org/10.3390/ani11123523
https://doi.org/10.1016/S0044-8486(99)00191-XSerra-Llinares, R.M., Nilsen, R., Uglem, I., Arechavala-Lopez, P., Bjørn, P.A. & Noble, C. (2013). Post-escape dispersal of juvenile Atlantic cod Gadus morhua from Norwegian fish farms and their potential for recapture. Aquaculture Environmental Interactions 3, 107−116. https://doi.org/10.3354/aei00051
Schönherz, A.A., Hansen, M.H.H., Jorgensen, H.B.H., Berg, P., Lorenzen, N. & Einer-Jensen, K. (2012). Oral transmission as a route of infection for viral haemorrhagic septicaemia virus in rainbow trout, Oncorhynchus mykiss (Walbaum). Journal of Fish Diseases, 35, 395-406, https://doi.org/10.1111/j.1365-2761.2012.01358.x
Sierra-Flores, R., Atack, T., Migaud, H., Davie, A., (2015). Stress response to anthropogenic noise in Atlantic cod Gadus morhua L. Aquacultural Engineering 67, 67–76. https://doi.org/10.1016/j.aquaeng.2015.06.003
Sitjà-Bobadilla, A. & Oidtmann B. (2017). Integrated Pathogen Management Strategies in Fish Farming. Fish Diseases, 119-144. https://doi.org/10.1016/B978-0-12-804564-0.00005-3
Skaala O., Besnier F., Borgstrom R., Barlaup B., Sorvik A.G., Normann E., Ostebo B.I., Hansen M.M. & Glover K.A. (2019). An extensive common-garden study with domesticated and wild Atlantic salmon in the wild reveals impact on smolt production and shifts in fitness traits. Evolutionary Applications 12, 1001-1016. https://doi.org/10.1111/eva.12777
Skaala Ø., Glover K.A., Barlaup B.T., Svåsand T., Besnier F., Hansen M.M. & Borgstrøm R. (2012). Performance of farmed, hybrid, and wild Atlantic salmon ( Salmo salar ) families in a natural river environment. Canadian Journal of Fisheries and Aquatic Sciences 69, 1994-2006. https://doi.org/10.1139/f2012-118
Skjæraasen, J.E., Meager, J.J., Karlsen, Ø., Mayer, I., Dahle, G., Rudolfsen, G. & Fernö, A. (2010). Mating competition between farmed and wild cod Gadus morhua . Marine Ecology Progress Series 412, 247-258. https://doi.org/10.3354/meps08156
Skjæraasen, J.E., Karlsen, Ø., Langangen, Ø., van der Meeren, T., Keeley, N.R., Myksvoll, M.S., Dahle, G., Moland, E., Nilsen, R., Elvik Schrøder, K.M., Bannister, R.J. & Olsen, E.M. (2021). Impact of salmon farming on Atlantic cod spatio-temporal reproductive dynamics. Aquaculture Environment Interactions 13: 399-412. https://doi.org/10.3354/aei00415
Skjæraasen, J.E., Karlsen, Ø., Langangen, Ø., Meier, S., Dunlop, K.M., van der Meeren, T., Keeley, N.B., Myksvoll, M.S., Dahle, G., Moland, E., Nilsen, R., Elvik, K.M.S., Bannister, R.J. & Olsen E.M. (2022). The attraction of cod Gadus morhua from local coastal spawning grounds to salmon farms. Aquaculture Environment Interactions 14, 229-242. https://doi.org/10.3354/aei00440
Skjærven, K. H., Alix, M., Kleppe, L., Fernandes, J. M. O., Whatmore, P., Nedoluzhko, A., Andersson, E., & Kjesbu, O. S. (2024). Ocean warming shapes embryonic developmental prospects of the next generation in Atlantic cod. ICES Journal of Marine Science, 81(4), 733–747. https://doi.org/10.1093/icesjms/fsae025
Skulstad, O.F., Taylor, J., Davie, A., Migaud, H., Kristiansen, T., Mayer, I., Taranger, G.L., Olsen, R.E. & Karlsen, Ø. (2013). Effects of light regime on diurnal plasma melatonin levels and vertical distribution in farmed Atlantic cod ( Gadus morhua L.). Aquaculture 414-415, 280-287. http://dx.doi.org/10.1016/j.aquaculture.2013.08.007
Snow, M., Cunningham, C.O., & Bricknell, I.R. (2000). Susceptibility of juvenile Atlantic cod Gadus morhua to viral haemorrhagic septicaemia virus isolated from wild-caught Atlantic cod. Diseases of Aquatic Organisms, 41, 225-229. https://doi.org/10.3354/dao041225
Snow, M., King, J. A., Garden, A. & Raynard, R.S. (2005). Experimental susceptibility of Atlantic cod, Gadus morhua (L.), and Atlantic halibut, Hippoglossus hippoglossus (L.), to different genotypes of viral haemorrhagic septicaemia virus. Journal of Fish Diseases 28, 737-742. https://doi.org/10.1111/j.1365-2761.2005.00682.x
Sodeland, M., Jentoft, S., Jorde, P.E., Mattingsdal, M., Albretsen, J., Kleiven, A.R., Synnes, A.-E., Espeland, S.H., Olsen, E.M., Andrè, C., Stenseth, N.C. & Knutsen, H. (2022). Stabilizing selection on Atlantic cod supergenes through a millennium of extensive exploitation. Proc. Natl. Acad. Sci., USA 119, e2114904119. https://www.pnas.org/doi/full/10.1073/pnas.2114904119
Sodeland, M., Jorde, P.E., Lien, S., Jentoft, S., Berg, P.R., Grove, H., Kent, M.P., Arnyasi, M., Olsen, E.M., & Knutsen, H. (2016). “Islands of Divergence” in the Atlantic cod genome represent polymorphic chromosomal rearrangements. Genome Biol. Evol. 8, 1012–1022. https://doi.org/10.1093/gbe/evw057
Sorensen, P.W. & Caprio, J. (1998). Chemoreception. In D.H. Evans (Ed), The Physiology of Fishes. 2nd ed. Marine Science Series. CRC Press, Boca Raton, Florida. 519 pp.
Stock, B., Knutsen, H., Nedreaas, K., Otterå, H., Fall, J., Denechaud, C., Moland, E., Jorde, P.E. & Kleiven, A.R. (2025). Oppdatert status om torsk langs Vestlandet- og Skagerrakkysten: potensial for bestandsvurdering. Rapport fra Havforskningen Nr. 2025-42, 64 pp. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-en-2025-42
Strøm, J.F., Bøhn, T., Skjæraasen, J.E., Gjelland, K.Ø. Karlsen, Ø., Johansen, T., Hanebrekke, T., Bjørn, P.A. & Olsen, E.M. (2023). Movement diversity and partial sympatry of coastal and Northeast Arctic cod ecotypes at high latitudes. Journal of Animal Ecology 92, 1966-1978. https://doi.org/10.1111/1365-2656.13989
Svåsand, T., Kristiansen, T.S., Pedersen, T., Salvanes, A.G.V., Engelsen, R., Nævdal, G. & Nødtvedt, M. (2000). The enhancement of cod stocks. Fish and Fisheries 1, 173-205. https://doi.org/10.1046/j.1467-2979.2000.00017.x
Svåsand, T., Grefsrud, E.S., Karlsen, Ø., Kvamme, B.O., Glover, K.S., Husa, V. & Kristiansen, T.S., (2017). Risikorapport norsk fiskeoppdrett 2017 Fisken og havet, Særnr. 2-2017. 179 pp. https://www.hi.no/resources/publikasjoner/risikorapport-norsk-fiskeoppdrett/2017/risikorapport_2017.pdf
Svåsand, T., Otterå, H. & Taranger, G.L., Litvak, M., Skiftesvik, A.B., Bjelland, R.M., Pavlov, D.A., Holm, J.Chr. Harboe, T., Mangor-Jensen, A., Norberg, B. & Howell, B. (2004). The status and perspective for the species. In Moksnes, E., Kjørsvik, E. & Olsen, Y. (Eds). Culture of cold-water marine fish. Blackwell Publishing, Oxford, UK, p 433-474.
Sæther, B.S., Bjørn, P.A. & Dale, T. (2007). Behavioural responses in wild cod ( Gadus morhua L.) exposed to fish holding water. Aquaculture 262, 260-267. https://doi.org/10.1016/j.aquaculture.2006.11.020
Sæther, B.-S., Løkkeborg, S., Humborstad, O.-B., Tobiassen, T., Hermansen, Ø. & Midling, K.Ø., (2012). Fangst og mellomlagring av villfisk ved oppdrettsanlegg. Nofima rapport nr. 8/2012, 53 pp. https://nofima.brage.unit.no/nofima-xmlui/handle/11250/2504608
Taranger, G.L., Karlsen, O., Bannister, R.J., Glover, K.A., Husa, V., Karlsbakk, E., Kvamme, B.O., Boxaspen, K.K., Bjørn, P.A., Finstad, B., Madhun, A.S., Morton, H.C. & Svåsand T. (2015). Risk assessment of the environmental impact of Norwegian Atlantic salmon farming. ICES Journal of Marine Science 72, 997-1021. https://doi.org/10.1093/icesjms/fsu132
Taranger, G.L., Aardal, L., Hansen, T. & Kjesbu, O.S. (2006). Continuous light delays sexual maturation and increases growth of Atlantic cod ( Gadus morhua L.) in sea cages. ICES Journal of Marine Science 63, 365-375. https://doi.org/10.1016/j.icesjms.2005.10.014
Torres-Corral, Y., González-Barreiro, O., Riaza, A.; Santos, Y. (2022). Establishment of different challenge models for Aeromonas salmonicida subsp. achromogenes in turbot and sole. Aquaculture, 555, 738261, https://doi.org/10.1016/j.aquaculture.2022.738261
Uglem, I., Karlsen, Ø., Sanches-Jerez, P. & Sæther, B.S. (2014). Impacts of wild fishes attracted to open-cage salmonids in Norway. Aquacult. Environ. Interact. 6, 91-103. https://doi.org/10.3354/aei00112
Uglem, I., Berg, M., Varne, R., Nilsen, R., Mork, J. & Bjørn, P.A. (2011). Discrimination of wild and farmed Atlantic cod ( Gadus morhua ) based on morphology and scale-circuli pattern. ICES Journal of Marine Science 68, 1928-1936. https://doi.org/10.1093/icesjms/fsr120
Uglem, I., Bjørn, P.A., Dale, T., Kerwath, S., Økland, F., Nilsen, R., Aas, K., Fleming, I. & McKinley, R.S. (2008). Movements and spatiotemporal distribution of escaped famed and local wild Atlantic cod ( Gadus morhua L.) in a Norwegian fjord. Aquaculture Research 39, 158–170. https://doi.org/10.1111/j.1365-2109.2007.01872.x
Uglem, I., Bjørn, P.A., Mitamura, H. & Nilsen, R. (2010). Spatiotemporal distribution of coastal and oceanic Atlantic cod Gadus morhua sub-groups after escape from a farm. Aquaculture Environment Interactions 1, 11-19. https://doi.org/10.3354/aei00003
Uglem, I., Dempster, T., Bjørn, P.-A., Sanchez-Jerez, P. & Økland, F. (2009). High connectivity of salmon farms revealed by aggregation, residence and repeated movements of wild fish among farms. Marine Ecology Progress Series 384, 251–260. https://doi.org/10.3354/meps08001
Uglem, I., Karlsen, Ø., Sanches-Jerez, P. & Sæther, B.S. (2014). Impacts of wild fishes attracted to open-cage salmonids in Norway. Aquacult. Environ. Interact. 6, 91-103. https://doi.org/10.3354/aei00112
Uglem, I., Knutsen, Ø., Kjesbu, O.S., Hansen, Ø.J., Mork, J., Bjørn, P.A., Varne, R., Nilsen, R., Ellingsen, I. & Dempster, T. (2012). Extent and ecological importance of escape through spawning in sea-cages for Atlantic cod. Aquaculture Environment Interactions 3, 33-49. https://doi.org/10.3354/aei00049
Urquhart, K., Bowden, T.J., Buckett, B.-E., Garcia, J., Fryer, R.J. & Ellis, A.E. (2009). Experimental study of the susceptibility of Atlantic cod, Gadus morhua (L.), to infection with an IPNV strain pathogenic for Atlantic salmon, Salmo salar L. Journal of fish diseases. 32, 447-456. https://doi.org/10.1111/j.1365-2761.2009.01036.x
van der Meeren, T. & Ivannikov, V.P. (2006). Seasonal shift in spawning of Atlantic cod ( Gadus morhua L.) by photoperiod manipulation: Egg quality in relation to temperature, and intensive larval rearing. Aquaculture Research 37: 898-913. https://doi.org/10.1111/j.1365-2109.2006.01510.x
van der Meeren, T. & Naas, K.E. (1997). Development of rearing techniques using large enclosed ecosystems in the mass production of marine fish fry. Reviews in Fisheries Science 5, 367-390. https://doi.org/10.1080/10641269709388606
van der Meeren, T., Jørstad, K.E., Paulsen, O.I., & Dahle G. (2012). Offspring from farmed cod ( Gadus morhua spawning in net pens: documentation of larval survival, recruitment to spawning stock, and successful reproduction. ICES CM 2012/P:11. (in mimeo). https://www.ices.dk/sites/pub/CM%20Doccuments/CM-2012/P/P1112.pdf
van der Meeren, T., Meier, S., Myksvoll, M.S., Dahle, G., Karlsen, Ø., Staby, A., Mjanger, H., Engevik, A., Dunlop, K.M., Bannister, R. & Skjæraasen, J.E. (2021). Sluttrapport fra ICOD-prosjektet. Arbeidspakke 2: Egg- og yngelundersøkelser på gyte- og oppvekstområder på Smøla og i Aure i forbindelse med etablering av oppdrettsanlegg nær lokale gyteområder for kysttorsk. Rapport fra Havforskningen Nr. 2021-32. 88 pp. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-2021-32
van der Meeren, T., Alix, M., Asplin, L. & Falkenhaug, T. (2024). Risiko knyttet til gyting i merd hos oppdrettstorsk ulike deler av året. Rapport fra havforskningen Nr. 2024-60. 20 pp. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-2024-60 https://doi.org/10.13140/RG.2.2.11795.80168
Varne, R. (2016). Tracing the fate of escaped farmed cod ( Gadus morhua L.) in a Norwegian fjord system. Norwegian University of Science and Technology, Doctoral theses at NTNU, 2016:116, 169 pp. https://ntnuopen.ntnu.no/ntnu-xmlui/handle/11250/2390705
Varne, R., Kunz, K.L., Johansen, T., Westgaard, J.-I., Uglem, I. & Mork, J. (2015). Farmed cod escapees and net-pen spawning left no clear genetic footprint in the local wild cod population. Aquaculture Environment Interactions 7, 253-266. https://doi.org/10.3354/aei00153
Weli, S.C., Gjessing, M., Patel, S., Olsen, A.B. & Sandlund, N. (2025). Infectious pancreatic necrosis virus can be transmitted through cohabitation from Atlantic cod ( Gadus morhua ) to Atlantic cod, Atlantic salmon ( Salmo salar ), Atlantic halibut ( Hippoglossus hippiglossus ) and lumpfish ( Cyclopterus lumpus ). Journal of Fish Diseases, e70042. https://doi.org/10.1111/jfd.70042
Westernhagen, H. (1970). Erbrütung der eier von Dorsch (Gadus morhua), Flunder (Pleuronectes flesus) und Scholle (Pleuronectes platessa) unter kombinierten temperature- und salzgehaltsbedingungen. Helgoländer Wissenschaftliche Meeresuntersuchen 21: 21–102. https://doi.org/10.1007/BF01630518
Wiklund, T. (1995). Survival of atypical Aeromonas salmonicida in water and sediment microcosms of different salinities and temperatures. Diseases of Aquatic Organisms 21, 137-143. https://doi.org/10.3354/dao021137
Wroblewski, J.S., Hiscock, H.W. & Bradbury, I.R. (1999). Fecundity of Atlantic cod ( Gadus morhua ) farmed for stock enhancement in Newfoundland bays. Aquaculture 171, 163–180. https://doi.org/10.1016/S0044-8486(98)00407-4
https://doi.org/10.1016/j.aquaculture.2010.11.009Zimmermann, E.W., Purchase, C.F. & Fleming, I.A. (2012). Reducing the incidence of net cage biting and the expression of escape-related behaviors in Atlantic cod ( Gadus morhua ) with feeding and cage enrichment. Applied Animal Behaviour Science 141, 71-78. https://doi.org/10.1016/j.applanim.2012.07.009
Zimmermann, E.W., Purchase, C.F., Fleming, I.A. & Brattey, J. (2013). Dispersal of wild and escapee farmed Atlantic cod ( Gadus morhua ) in Newfoundland. Canadian Journal of Fisheries and Aquatic Sciences 70, 747–755. https://doi.org/10.1139/cjfas-2012-042
Øiestad, V., Kvenseth, P. G., & Folkvord, A. (1985). Mass production of Atlantic cod juveniles Gadus morhua in a Norwegian saltwater pond. Transactions of the American Fisheries Society 114: 590-595. https://doi.org/10.1577/1548-8659(1985)114<590:MPOACJ>2.0.CO;2
Øines, Ø., Simonsen, J.H., Knutsen, J.A., Heuch P.A. (2006) Host preference of adult Caligus elongatus Nordmann in the laboratory and its implications for Atlantic cod aquaculture. Journal of Fish Diseases 2006, 29, 167–174 https://doi.org/10.1111/j.1365-2761.2006.00702.x
Øines, Ø., Heuch, P.A (2007). Caligus elongatus Nordmann genotypes on wild and farmed fish. Journal of Fish Diseases 2007, 30, 81–91 https://doi.org/10.1111/j.1365-2761.2007.00783.x
Øines, Ø. og T. Schram (2008). Intra- or inter-specific difference in genotypes of Caligus elongatus Nordmann 1832? Acta Parasitologica 53, 93-105. https://doi.org/10.2478/s11686-008-0002-2