Assessment of biological status, proposals for management measures and a new rebuilding plan.
This report highlights the status of Norwegian coastal cod north of 62˚N including stock structure, monitoring, a rebuilding plan and potential management measures. The report contains limited information with respect to biology, ecosystems and coastal environmental factors. Northeast Arctic cod (NEAC) and Norwegian coastal cod (NCC), cannot be separated visually at the landing site. Thus, there is no way to manage quotas for the two stocks separately in real time. Only when the catch year is over, and all sampling is analyzed and quality assured, can the catch of coastal cod be calculated accurately.
Genetic studies show a gradual change in the stock structure of coastal cod from north to south. This indicates that division into several management units can provide a basis for better protection of any "weaker components" throughout the distribution area. It must be noted, however, that such divisions will necessarily require increased scientific survey coverage and more intense fisheries sampling in order to provide data that would adequately represent any new management units.
In general, based on the current stock situation, there seems to be a need for stricter measures south of 67˚N than north of this latitude. The Institute of Marine Research recommends that the following proposals should be considered for commercial fisheries:
Reduced coastal fishing in the fall when there is less NEAC at the coast.
Close areas / months where a lot of coastal cod is usually caught.
Protect coastal cod spawning areas.
Preserve a broad age and size composition.
Consider revisions of fjord lines and more stringent fishery restrictions within fjord lines.
Monitor and measure the effects of regulations.
Further develop genetic methods and tools to operate on the "quayside", and hence quickly and efficiently capture dynamic changes in coastal cod catch.
Promote "local-regional regulations" in areas where there is a very good knowledge base.
The Institute of Marine Research also recommends special measures for the tourist- and recreational fishing.
1 - Bakgrunn
Denne rapporten setter søkelys på vurdering av status for kysttorsk nord for 62 o N, bestandsstruktur, overvåkning, gjenoppbyggingsplan og mulige forvaltningstiltak. Rapporten inneholder lite om biologi, alder, vekst og torskens diettsammensetning, og går heller ikke nevneverdig inn på naturlige dødsårsaker og økosystem aspekter. Rapporten beskriver heller ikke kystklimaet inkl. temperatur og saltholdighet, eller miljøstatus.
Fangst av kysttorsk avregnes mot en samlet kvote for kysttorsk og skrei. Begrensninger av fisket etter kysttorsk foretas derfor gjennom regler som styrer utøvelsen av fisket (områdebegrensninger, redskapsregulering, periodiseringer med mere). Erfaringene så langt har vist at disse reguleringene ikke har vært tilstrekkelig til å gi målbar vekst i bestanden.
Torsk fra de to bestandene lar seg ikke visuelt skille fra hverandre ved landing. Dermed foreligger ingen fortløpende oppdatering for gjenværende kvote. Først når fangståret er omme og alle stikkprøver er analysert og kvalitetssikret, blir siste års fangst av kysttorsk beregnet.
I tillegg er det betydelig usikkerhet i toktmålingene (særlig sør for Lofoten). Dette fører til stor grad av usikkerhet i bestandsvurderingen.
1.1 - MSC sertifisering
Det nordøstarktiske torskefisket har siden 2010 vært sertifisert etter MSC-standarden, og merkeordningen anses å være av stor betydning for markedsadgang og oppnådd pris på fisken. Situasjonen for kysttorsk har vært en flaskehals og en risikofaktor for sertifikatet helt siden det først kom på plass. Det var først etter at dagens gjeldende gjenoppbyggingsplan for kysttorsk kom på plass, at fisket innenfor 12 nm lot seg sertifisere, og da ble kysttorsk behandlet under en IPI (Inseparable or Practically Inseperable) provisjon i MSC-standarden. Gitt gjenoppbyggingsplanen, passerte da «bifangsten» av kysttorsk MSC-standarden, men ble fremdeles ansett som et sporbarhetsproblem.
I den opprinnelige sertifiseringen, ble det gitt 5 års frist til å enten bygge kysttorskbestanden opp på et sertifiserbart nivå som målart, eller finne en metode for å skille kysttorsk og nordøstarktisk torsk i verdikjedene. En ytterligere 5-årig forlengelse ble gitt ved resertifiseringen i 2015. Torsk og hyse er nå i full gang med sin tredje hovedsertifisering, og gjeldende sertifikat løper ut i oktober 2020. Etter den tid er det svært sannsynlig at store deler av det norske torskefisket vil være utestengt fra å bruke MSC-merket inntil de underliggende problemene med kysttorsken er løst.
1.2 - Definisjon av kysttorsk Nord
I kystområdene nord for 62 grader nord (Fiskeristatistikk-områdene 00, 03, 04, 05, 06 og 07) blir kysttorsk identifisert ut fra vekstmønsteret i øresteinene (otolittene, Rollefsen 1932). Dette blir gjort ved stikkprøvetaking, både i forskningstoktene og i prøvetakingen fra fiskeriene. Basert på denne prøvetakingen blir torskefangster per område og kvartal i ettertid (når fangståret er omme) splittet på kysttorsk og nordøst-arktisk torsk. For torsk yngre enn 2 år inneholder otolittene for lite informasjon til å foreta et pålitelig skille mellom kysttorsk og skrei. Disse aldersgruppene er bare sporadisk representert i yrkesfisket, men kan i noen områder inngå i fritids -og turistfiske. For 0-1 åringer er det bare genetiske analyser som kan avklare om det er kysttorsk eller nordøst-arktisk torsk.
1.3 - Skille mellom kysttorsk og skrei
Hylen et al (2008) oppsummerer hva man historisk og vitenskapelig har lagt til grunn for å skille kysttorsk og nordøstarktisk torsk (skrei). I flere hundre år har fiskere skilt mellom to hovedtyper av torsk i norske farvann; kysttorsken som lever nær kysten og i fjordene hele året, og de store torskemassene som kommer inn i kystfarvannet for å gyte, eller for å beite på lodde sent på vinteren og våren. De skilte mellom de to typene på grunnlag av formen og fargen på fisken. Det samme gjorde G.O. Sars da han tegnet dem (Sars 1879). Da Rollefsen studerte torskeotolittene (Rollefsen 1933, 1935) fant han at “ Otolittene til disse kysttorskene avviker betydelig på mange måter fra skrei (nordøst-arktisk torsk), ikke bare i relativ bredde på sonene og deres struktur, men til og med i den ytre formen ”. Resultater fra ganske nylige analyser støtter også bruken av otolittform for separasjon av nordøst-arktisk torsk og norsk kysttorsk (Stransky et al. 2007). Rollefsen fant også ut at de to torskegruppene viste forskjeller i vekst og alder ved første gyting; kysttorsken vokser raskere og når modning i en tidligere alder enn nordøst-arktisk torsk; observasjoner som siden har blitt bekreftet (f.eks Godø 1984). Studier gjort av britiske og russiske forskere på 1950-tallet avslørte forskjeller i otolittstruktur og sonemønster hos torsk fanget på forskjellige steder i Barentshavet (Trout 1955; Mankevich 1969).
Separasjonen i to hovedgrupper, kysttorsk og nordøst-arktisk torsk, ble støttet av de genetiske studiene til Møller (1968, 1969). Han foreslo at de to gruppene var genetisk distinkte bestander. Mork et al. (1984, 1985) fant at den genetiske avstanden mellom de forskjellige bestandene av atlantisk torsk var generelt lav, og de mente at dette sannsynligvis var et resultat av genstrømning mellom bestandene. Nyere undersøkelser som har sammenlignet resultatene fra genetiske studier, merkeforsøk og otolitt-mønstre, har ført til konklusjonen at de to gruppene skal betraktes som separate bestander (Jakobsen 1987; Dahle 1991, og senere publikasjoner referert i kapitlet om genetikk). Kysttorsk skiller seg også noe fra nordøst- arktisk torsk med hensyn til livshistorieparametere, som også viser forskjeller mellom områder (Berg og Albert 2003, og Berg og Pedersen 2001). Siden midten av 1990-tallet har en egen bestandsvurdering av kysttorsk blitt utført av ICES.
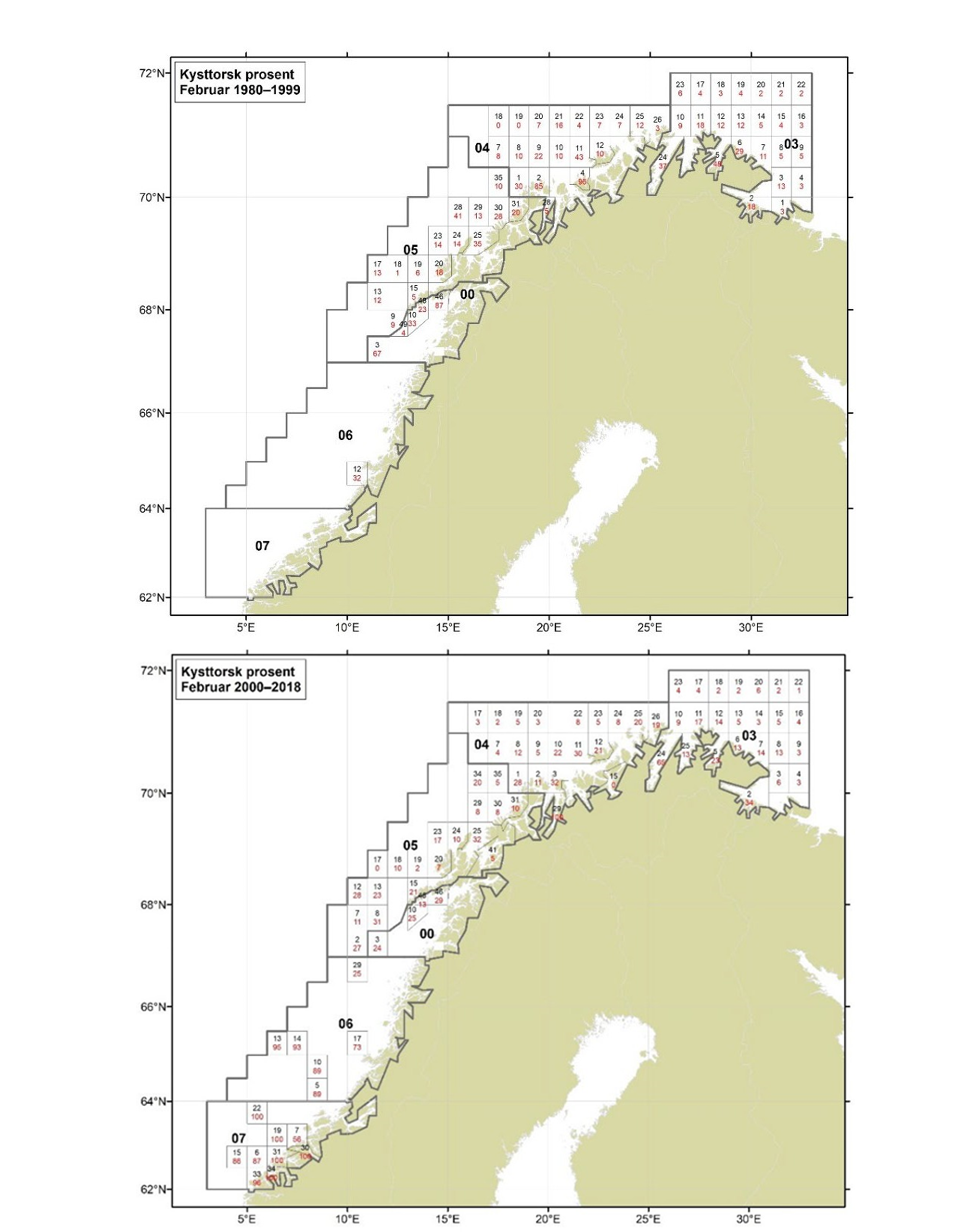
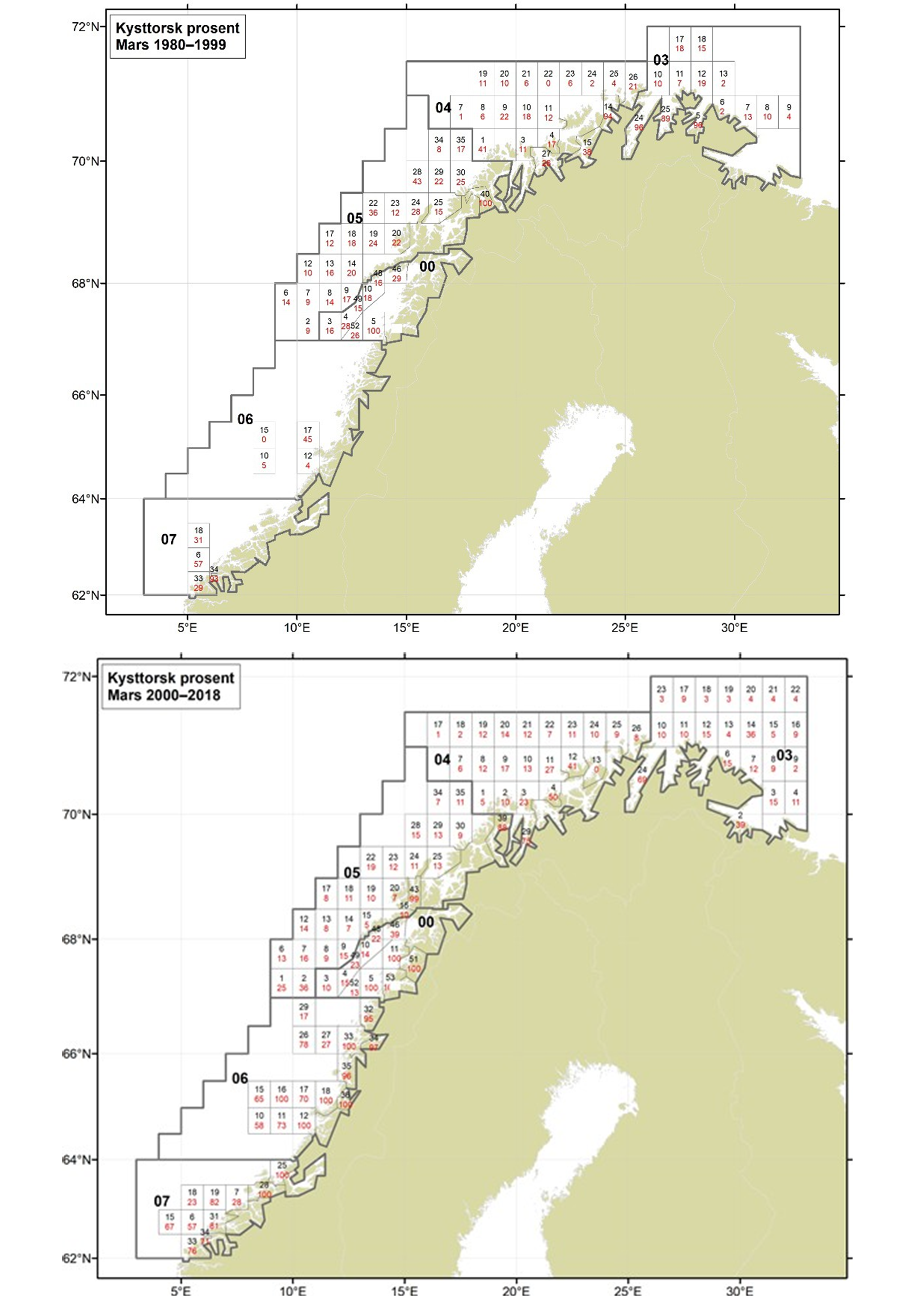
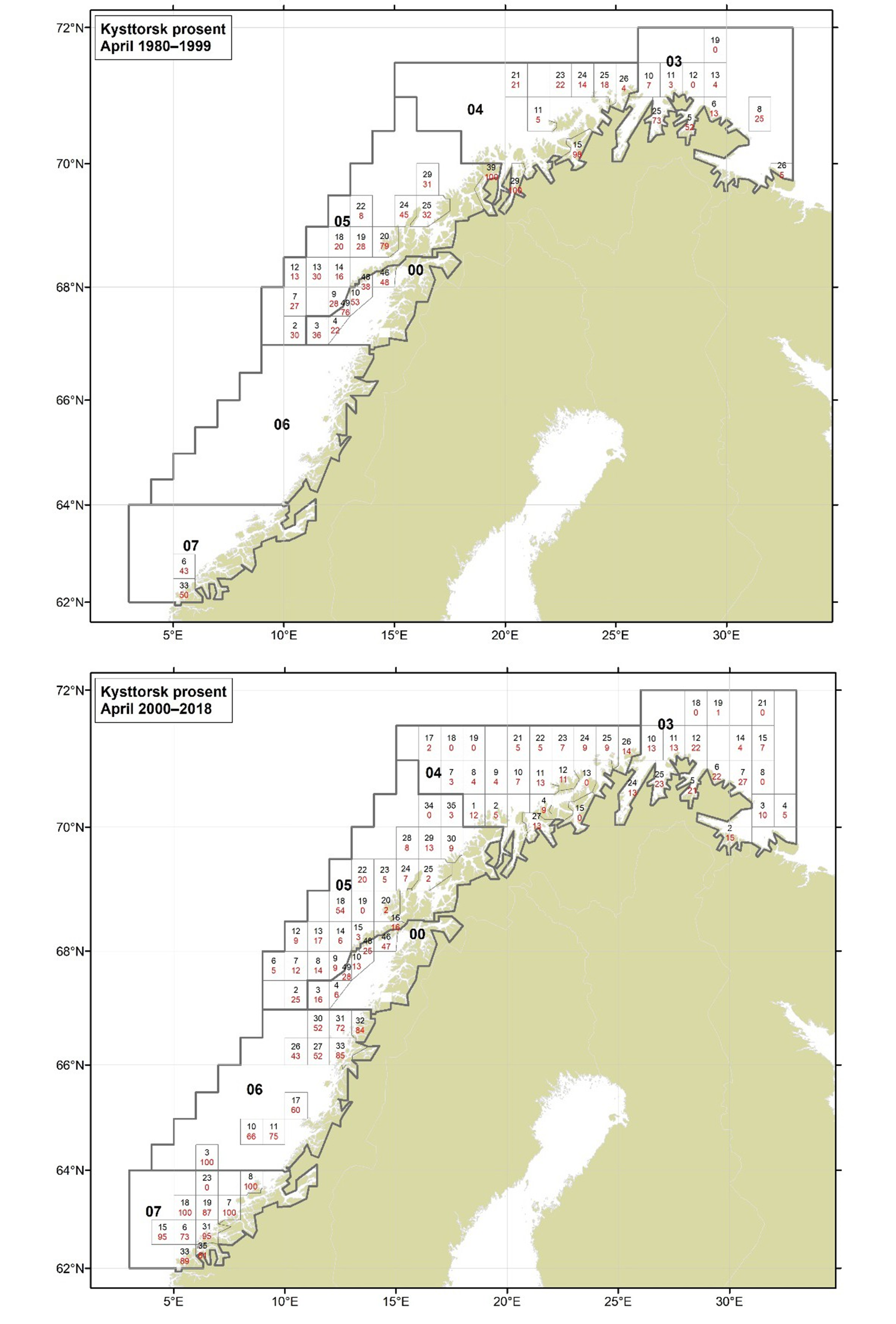
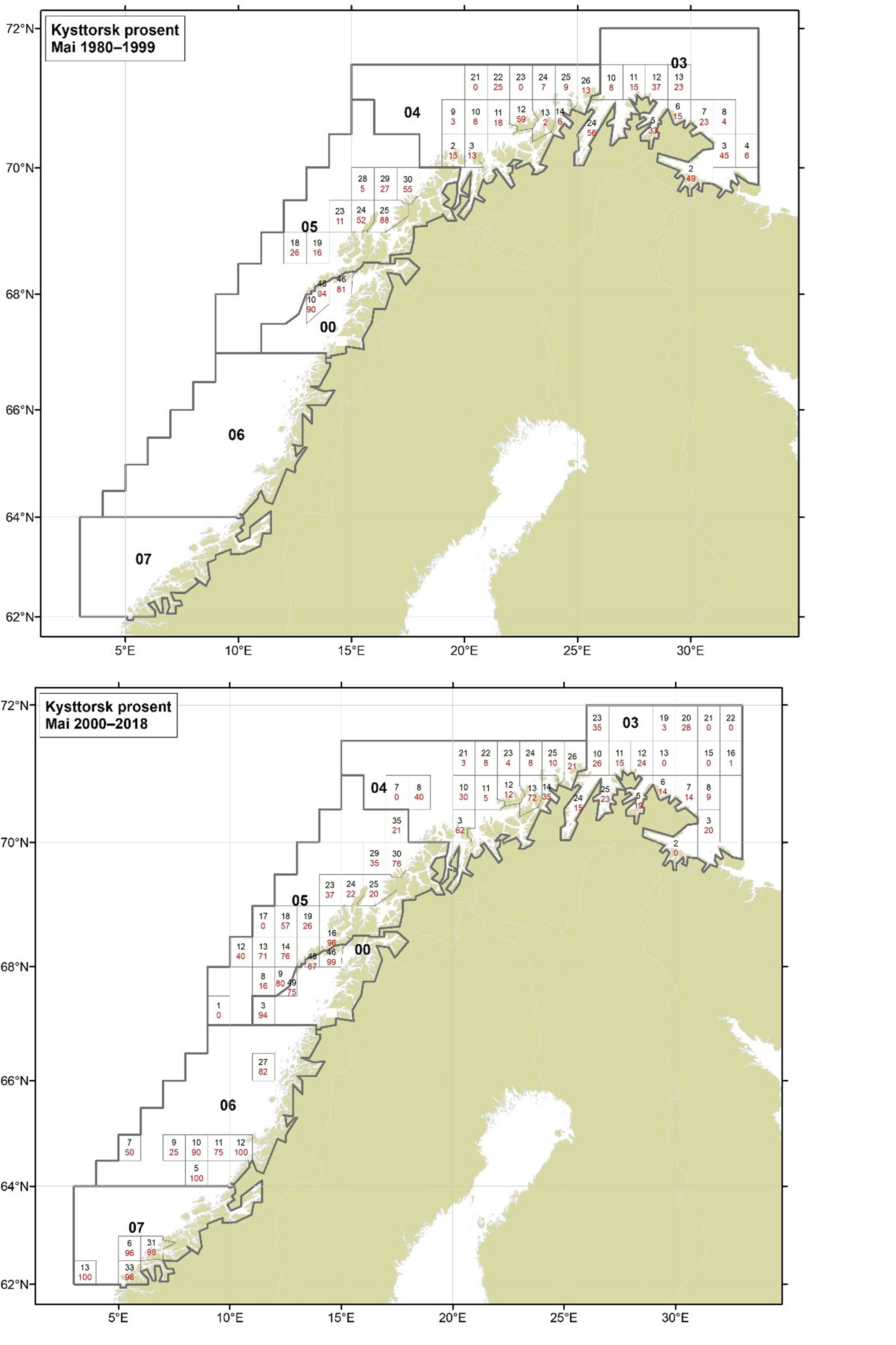
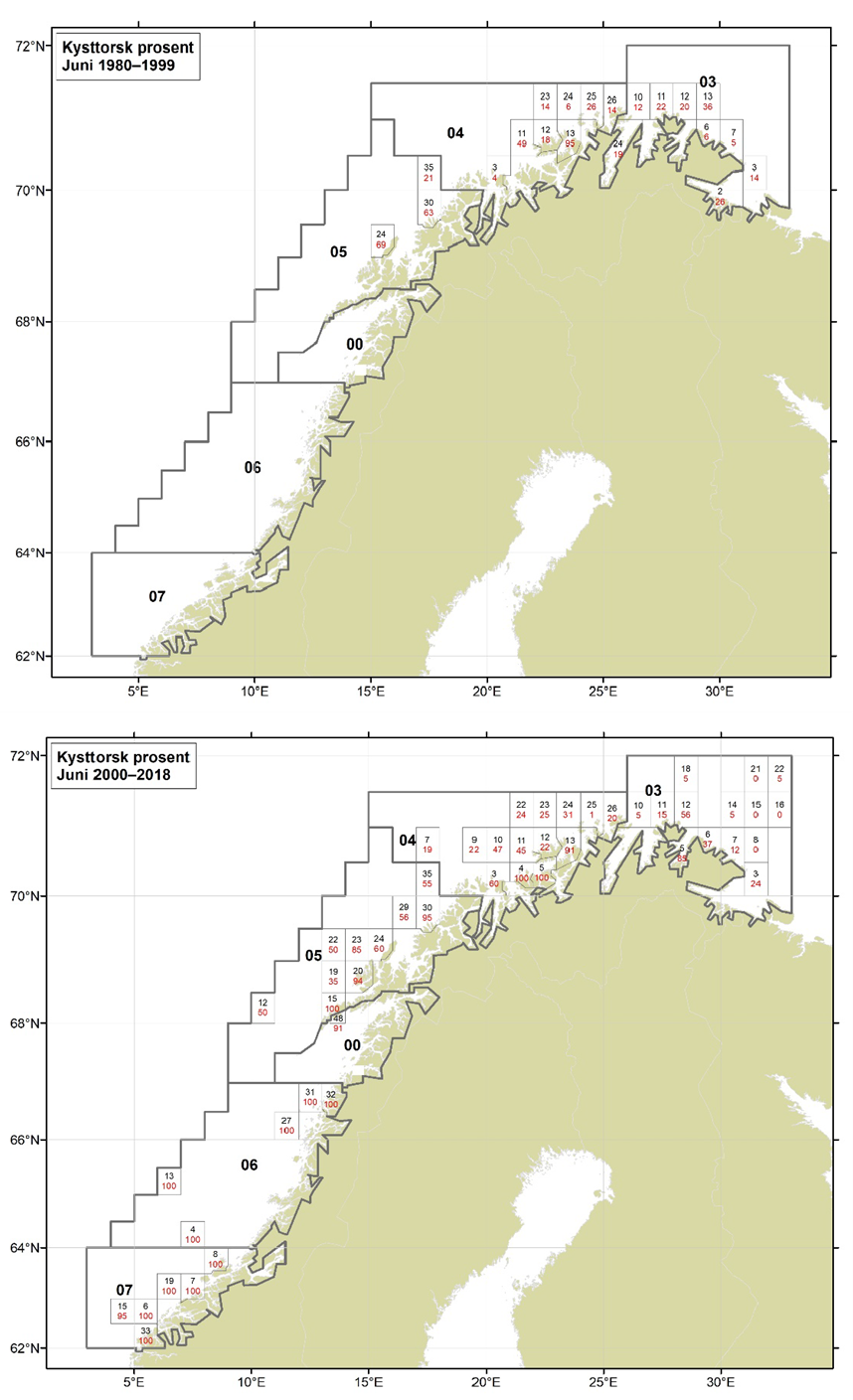
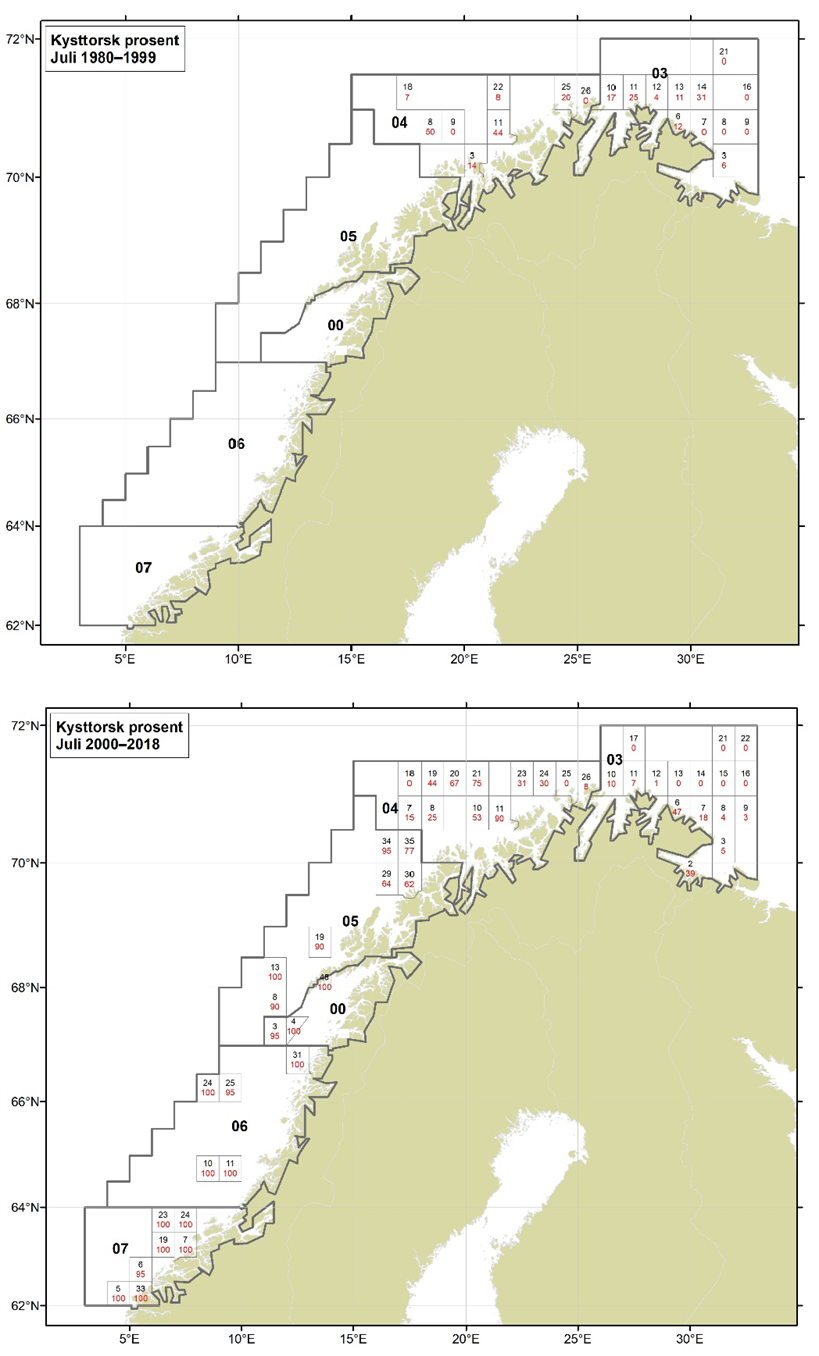
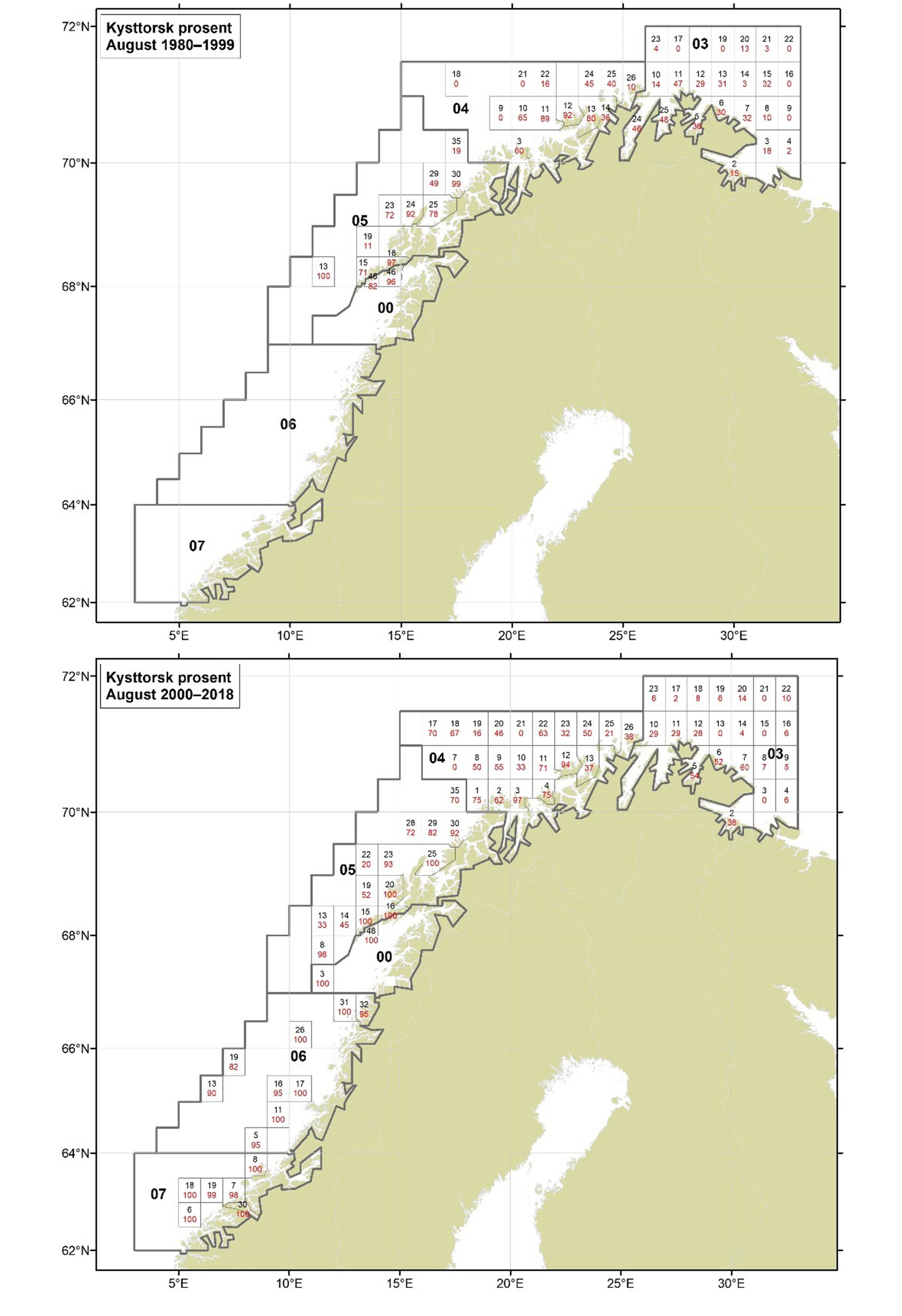
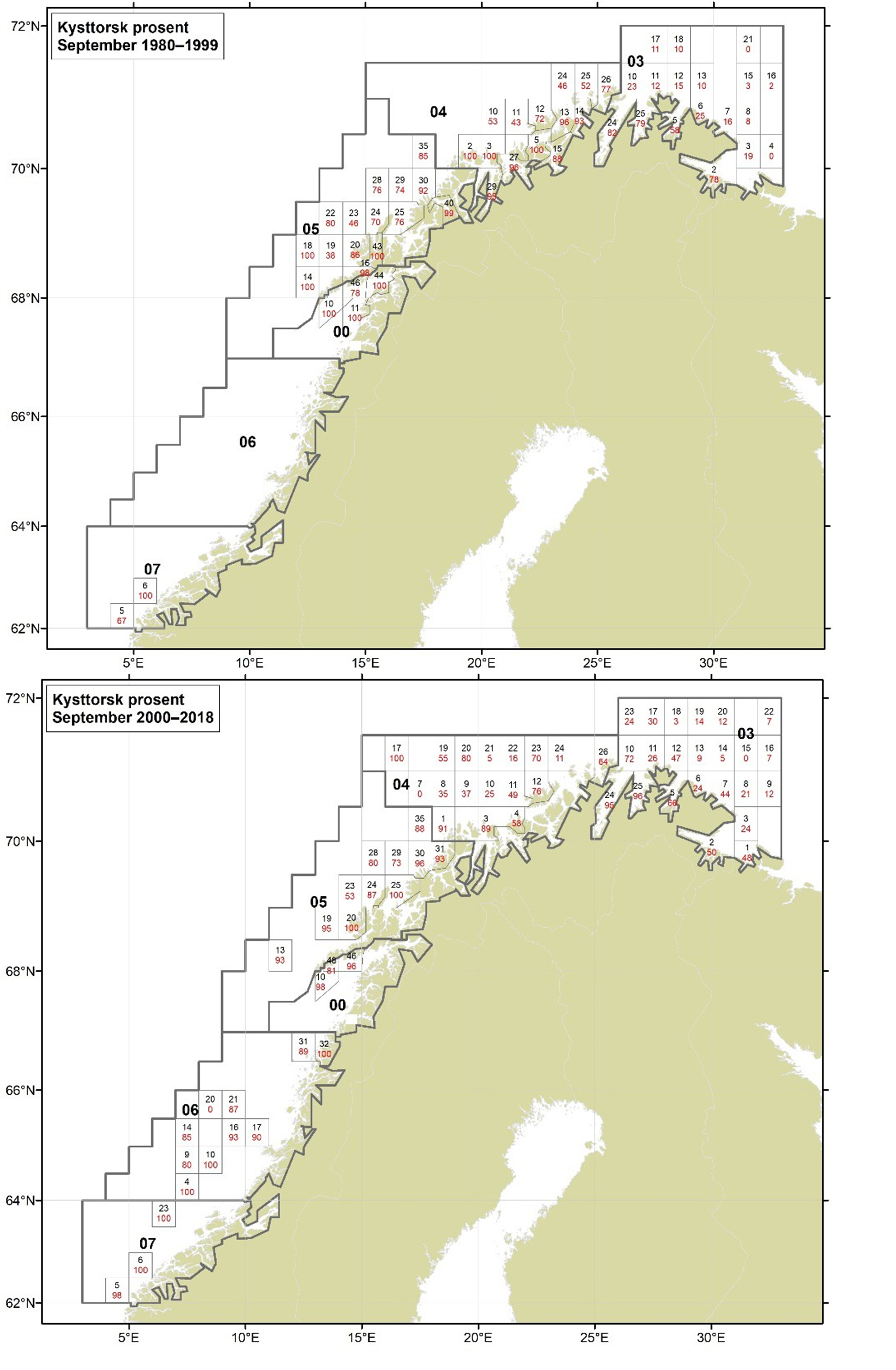
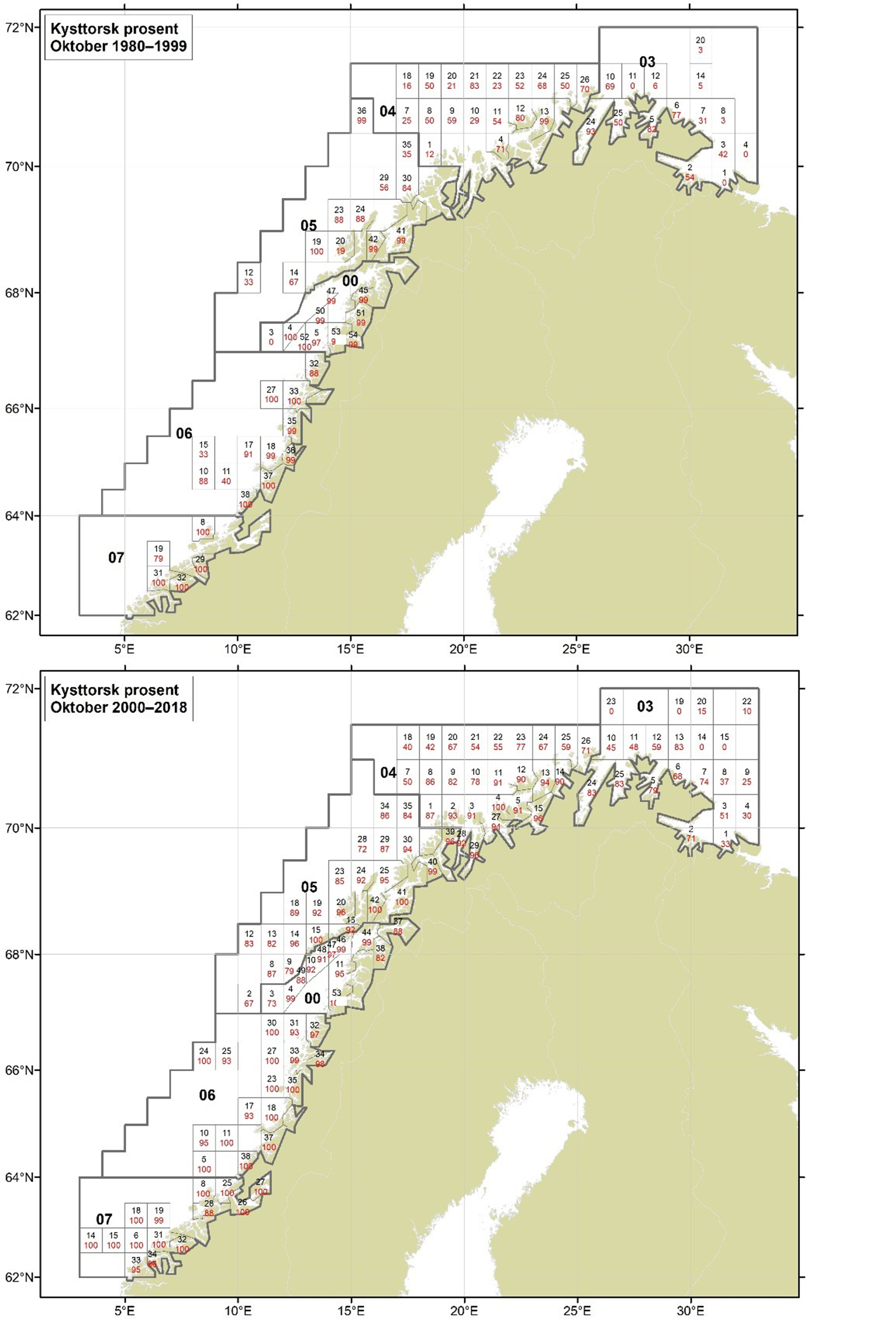
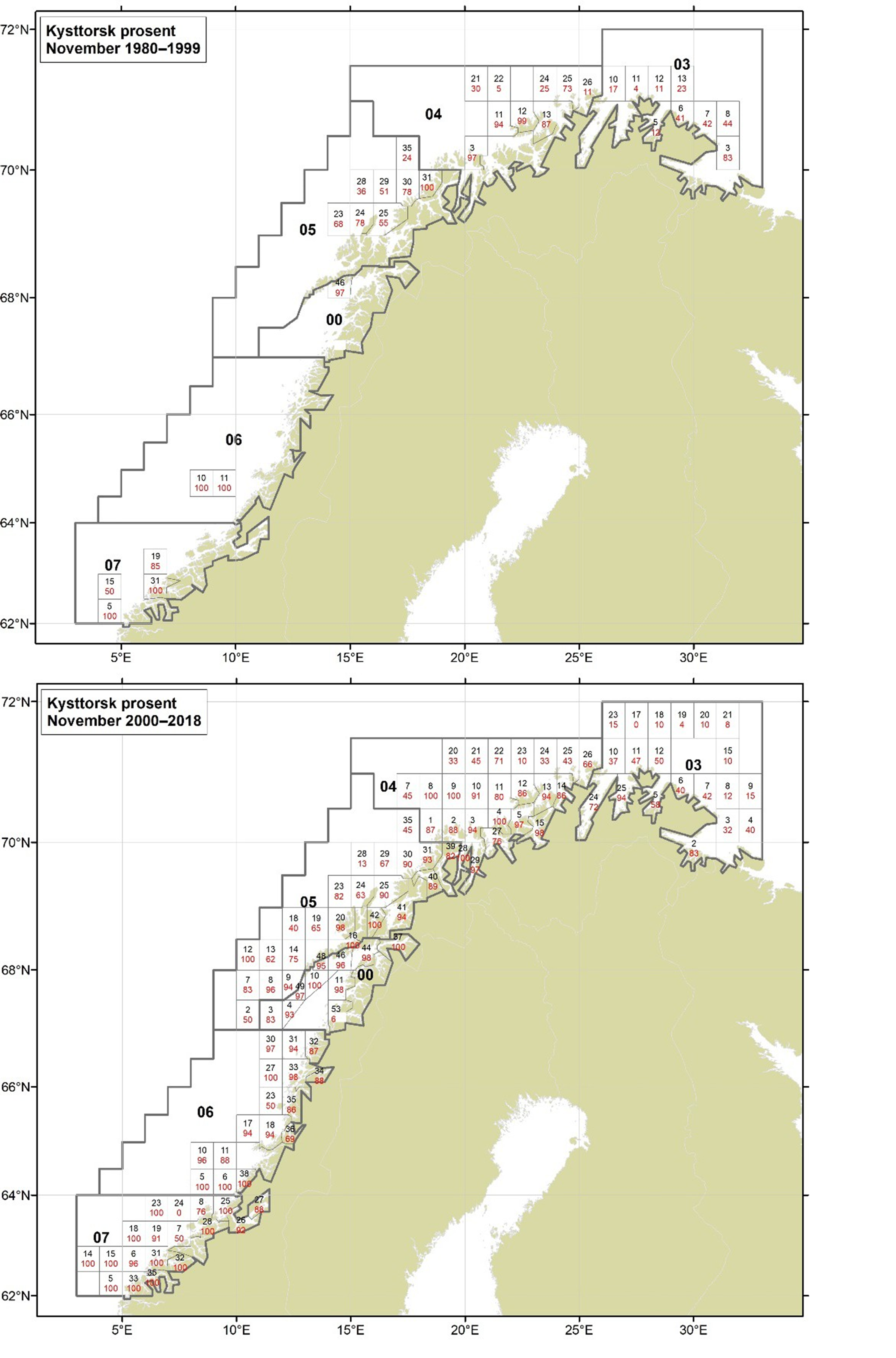
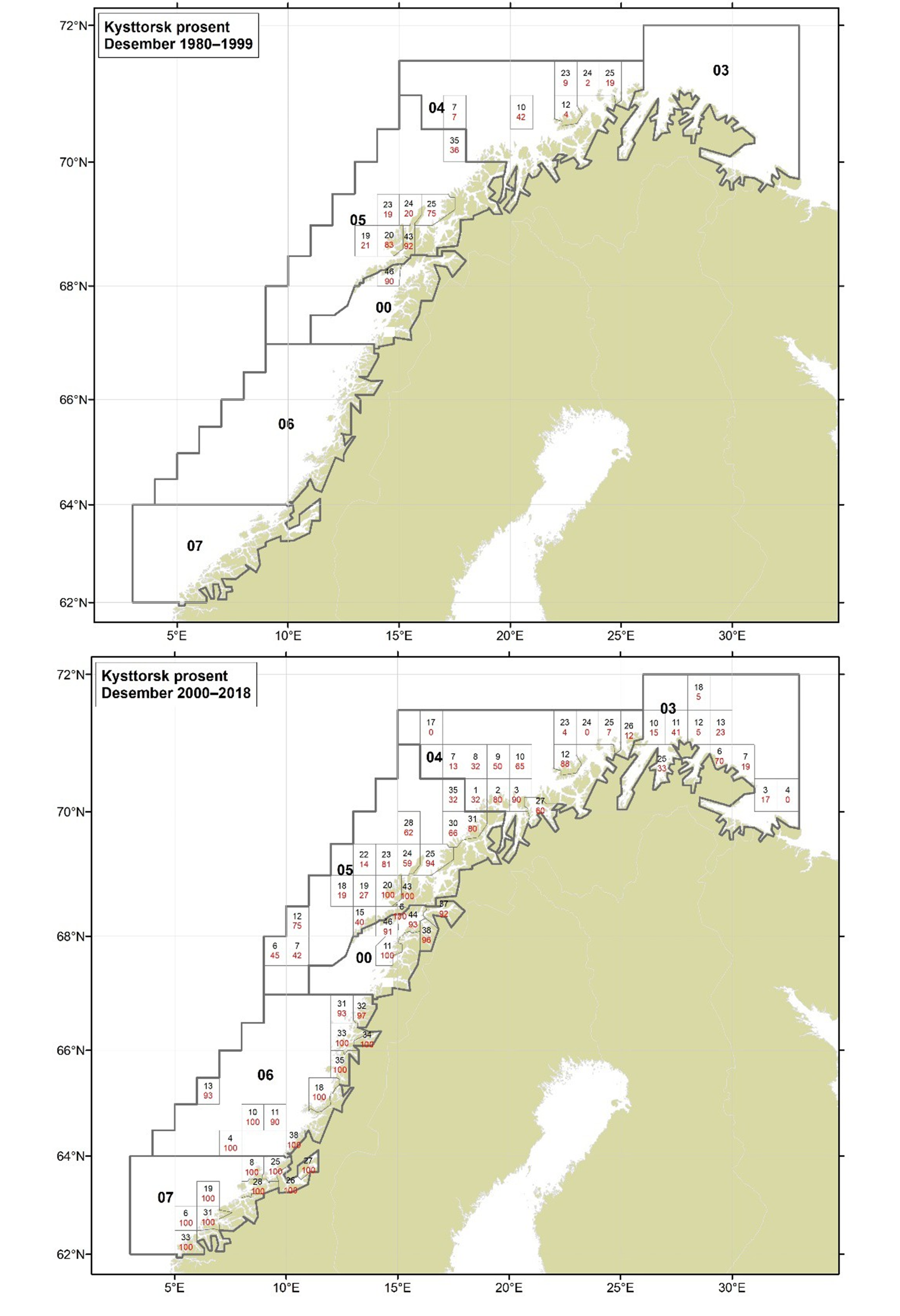
Figurer i Vedlegg 3 viser kart over prosent kysttorsk, bestemt utfra otolittmønster, per lokalitet innenfor alle kystområdene nord for 62N (03, 04, 05, 00, 06 og 07) hver måned i to tidsperioder 1980-1999 og 2000-2018. Kartene inkluderer prøvetaking både fra fiskeriene og fra forskningstoktene. Kartene viser at den geografiske prøvedekningen har blitt bedre i perioden 2000-2018 sammenlignet med 1980-1999. Vi ser også tydelige sesongmessige variasjoner. Det er disse data som benyttes ved estimering av kysttorskfangstene (se kapittel Landingsdata og forvaltningshistorikk).
1.4 - Landingsdata og forvaltningshistorikk
Tabeller og beretninger om Norges fiskerier finnes tilbake til 1866 (Anon. 1883; https://www.ssb.no/a/histstat/publikasjoner/histemne-06.html ). I eldre fiskeristatistikk «Norges Fiskerier» finnes offisiell norsk statistikk tilbake til 1914 på fangst av torsk pr landsdel/kommune i kategoriene: «Skrei» (gytetorsk), «Loddetorsk» (vårtorskefiske i Finnmark), «Banktorsk» og «Fjordtorsk». Slik er landingsstatistikken presentert for torsk frem til og med 1961 da «Banktorsk» og «Fjordtorsk» ble erstattet av «Annen torsk». For årene 1970-1976 presenteres landingsstatistikken av torsk bare som «Torsk». Betegnelsen «Skrei» inneholder trolig også noe kysttorsk på gytefeltene, og «Annen torsk» er nok i stor grad kysttorsk. Noe kysttorsk er sannsynligvis også inkludert i «Vårtorsk (loddetorsk)». Trolig ble betegnelsen «Skrei» i en del tilfeller også brukt for gytende kysttorsk sørpå (Opdal, 2009)
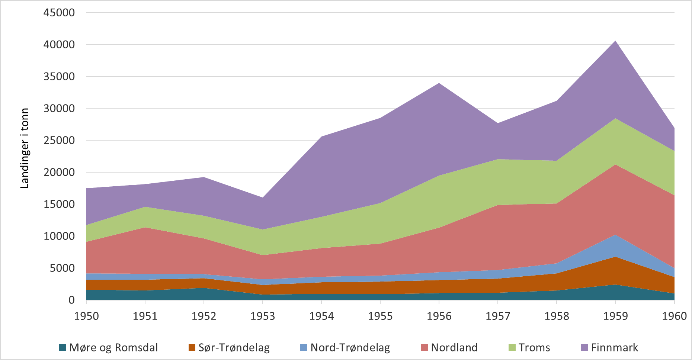
Figur 1 viser mengde fjordtorsk landet per fylke nord for Stad i perioden 1950-1960 ifølge Norges fiskerier (Anon. 1883). Slik statistikk finnes helt tilbake til 1914. Det finnes ingen presis definisjon av begrepet «fjordtorsk», men vi antar at det meste er kysttorsk. Det representerer trolig også det som ble fisket og levert innenfor det som nå defineres som fjordlinjer, og sammenligninger kan derfor til en viss grad gjøres mellom all torsk som i dag fiskes innenfor fjordlinjene og rapportert fjordtorsk i årene 1914-1960.
Figur 1. Mengde fjordtorsk (tonn) landet i perioden 1950-1960 (Norges fiskerier) per fylke nord for Stad (62 ˚ N).
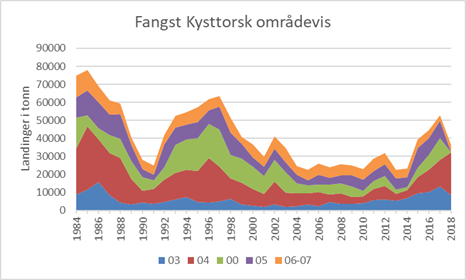
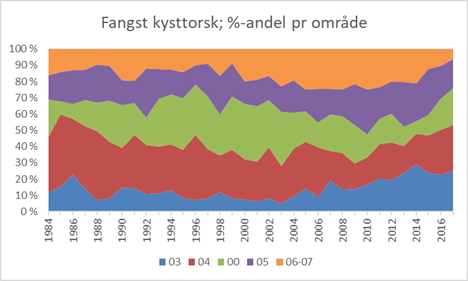
Ved Havforskningsinstituttet blir torsk sortert til kysttorsk og skrei basert på mønsteret i vekstsonene i ørestenene (otolittene) (Rollefsen 1933). Ut fra ørestensprøver er fangstkvantum av kysttorsk beregnet for årene 1984-2018 (Figur 2). Reviderte beregningsrutiner er årsak til at 2018 ikke er like godt fordelt på områder. Det bør utvikles rapporter i programvaren «Estimating Catch-at-age» (ECA) som kan presentere estimerte kysttorskfangster per område, kvartal og redskap.
Figur 2. Beregnet fangstkvantum av kysttorsk per statistisk område nord for Stad (62 ˚ N) i 1984-2017 (2018) basert på stikkprøver av øresteiner fra fangstene. Merk at fangstene i område 04, 05 og 00 er slått sammen i 2018 (rustrød farge).
1.5 - «Kysttorsk-kvoten»
Kysttorsk har inngått i den norsk-russiske kvoteavtalen siden 1977. I de årlige møtene i den Norsk-Russiske Fiskerikomiteen blir man først enig om en totalkvote for Nordøstarktisk torsk. Så avsettes et kvantum «Murmansktorsk» til Russland, så får Norge og Russland halvparten hver av resten. Til slutt legges det til et kvantum kysttorsk til den norske kvoten. Kvoten for «Murmansktorsk» og norsk kysttorsk blir satt likt, slik at begge land har lik samlet torskekvote. I perioden 1977-2003 var kysttorskkvoten 40 tusen tonn. I 2004 ble den redusert til 20 tusen tonn begrunnet med behovet for gjenoppbygging av kysttorskbestanden. I 2005 ble den økt til 21 tusen tonn og har siden vært uendret. Bakgrunnen for de 40 tusen tonn var basert på gjennomsnittlig årlig fangst i perioden 1960-1970 i statistikkområdene 06 og 07, pluss fangsten andre halvår i område 00 og 05 (ICES 1970, 1990).
Alt norsk torskefiske nord for 62-graden avregnes mot en samlet kvote for kysttorsk og den 5 til 10 ganger større kvoten for nordøst-arktisk torsk. Endringer i «kysttorskkvoten» har i seg selv liten effekt på uttaket av kysttorsk. Det har derfor værtet mål å innrette reguleringene slik at torskefisket i størst mulig grad rettes mot nordøstarktisk torsk . Dette har vist seg å være vanskelig, blant annet fordi det er mindre utgifter ved fiske nært kysten, og fordi små båter ikke kan fiske særlig langt utafor kysttorsk-områdene. Tiltak innført fra 2005 og senere har satt grenser for hvor nært kysten de ulike båtstørrelser har lov å drive direkte fiske på torsk. «Sildtorsken» har vært en spesiell utfordring: -Nvg sild samler seg på senhøsten/vinteren i spesielle kystområder og står der inntil den i slutten av januar starter på vandringen til gytefeltene. Disse sildeansamlingene tiltrekker seg torsk, og det er i perioder hardt fiske på kysttorsk i slike områder, spesielt i år når skreinnsiget lar vente på seg. Fra slutten av 1980-åra til 2003 skjedde dette i Vestfjord, Tysfjord og Ofotfjord, så dukket fisket opp i ytre Senja og Kvaløya fram til ca 2016, og har så flyttet seg til Skjervøy/ Kvænangen.
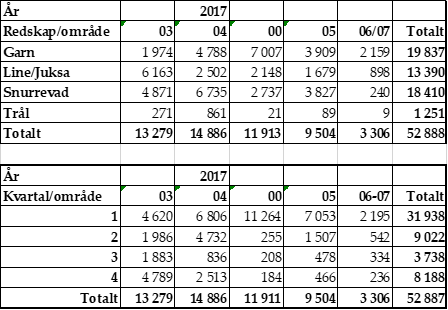
Dette tilsier at det er viktig med god prøvedekning fra fiskeriene fra Lofoten til Nord-Troms både under sildeinnsiget og skreiinnsiget (januar-april), og i områder med ferskfiskordning (Troms-Finnmark) i oktober-desember. Tabell 1 viser eksempel på kysttorskfangst pr område, kvartal og redskap og kvartal. Slike tabelloversikter har Havforskningsinstituttet for perioden 1984-2018 (se også Berg et al. 1998).
Tabell 1. Norsk fangst av kysttorsk nord for 62˚N fordelt på redskap, kvartal og områder i 2017 basert på splitting av den totale norske torskefangsten på kysttorsk og nordøst-arktisk torsk ved hjelp av prøvetaking fra fiskeriene og analyser av øresteinene (otolittene).
2 - Vurdering av status for kysttorsk nord for 62˚N
2.1 - Vurderinger og råd fra ICES; kort historikk om rammeverket for rådgivingen og den eksisterende gjenoppbyggingsplan
For bestander med en godkjent analytisk bestandsberegning gir ICES et råd om fiske som referer til diverse biologiske referansepunkt slik som Fmsy (den fiskedødelighet som teoretisk skal maksimere langtidsutbytte), eller Fpa (som skal sikre at bestanden ikke blir kritisk nedfisket). For bestander uten analytisk bestandsberegning, brukes ofte tidstrenden i toktindekser, eller tidstrender i fangst per enhet innsats, som grunnlag for å anbefale økning eller reduksjon i fisket. Når tilgjengelige data tilsier at bestanden er kritisk lav, kan ICES tilrå null fangst og innføring av en «hensiktsmessig» gjenoppbyggingsplan.
Kysttorsk nord for 62-graden har vært behandlet i ICES sin rådgivende komite siden 1987. I de første årene ble det ikke gitt spesifikke råd om fisket på denne bestanden, men i 2001 og 2003 var rådet å redusere fiskedødelighet betraktelig. I 2002 var rådet å redusere fisket i samme takt som reduksjonen for Nordøst-arktisk torsk. I alle årene 2004-2008 var rådet å innføre en gjenoppbyggingsplan. I årene 2009-2011 var rådet null fangst og innføring av gjenoppbyggingsplan. For alle årene 2012 til 2020 har rådet vært å følge den gjenoppbyggings-planen som ble innført i 2011.
Figur 3 (fra ICES 2019a). Årlige råd om reduksjon i fiskedødelighet i henhold til gjeldene gjenoppbyggings-plan. % reduksjon er relatert til beregnet F for fangståret 2009 (ca 0.3).
2010
2011
2012
2013
2014
2015
2016
2017
2018
2019
SSB survey compared to previous year
Rebuilding plan action step for the next year
-
Step 1
Step 2
Step 2
Step 2
Step 3
Step 3
Step 4
Step 4
Step 4
Management regulations for the next year*
-
Reduce F by ≥15%
Reduce F by ≥15%
Reduce F by ≥30%
Reduce F by ≥30%
Reduce F by ≥30%
Reduce F by ≥45%
Reduce F by ≥45%
Reduce F by ≥60%
Reduce F by ≥60%
I rådet fra ICES for 2019 og for 2020 ble det presisert at det bør innføres en ny plan, fordi den eksisterende planen ikke har ført til en klart målbar gjenoppbygging. Samtidig sier ICES at de vil gi råd i henhold til den eksisterende planen inntil en ny foreligger . Figur 3 viser hvordan gjenoppbyggingsplanen har ført til råd om større og større reduksjon i fiskedødelighet (F) for hvert år toktresultatet har gått ned. Det har vist seg vanskelig både å definere og å innføre de nødvendige tiltak som skulle føre til vesentlig reduksjon i F.
2.2 - Grunnlag for ICES-rådet gitt i 2019
Som i de 18 foregående år er det nyeste ICES rådet for kysttorsk (ICES 2019a) basert på beregninger og vurderinger gjort i ICES-arbeidsgruppen «ICES Working Group for Arctic Fisheries» (ICES-områdene I og IIa). Arbeidsgruppen i 2019 (ICES 2019b) påpeker at det fortsatt er betydelig usikkerhet i både toktdata og i beregnet fangst av kysttorsk. I arbeidsgruppen 2019 ble det tatt i bruk en revidert beregning av tidsserien for tonn fangst av kysttorsk (Figur 4), og antall kysttorsk ved alder i årlig fangst, splittet ut fra samlet kysttorsk og nordøstarktisk torsk. Denne er beregnet med programvaren «ECA» (Hirst et al 2012).
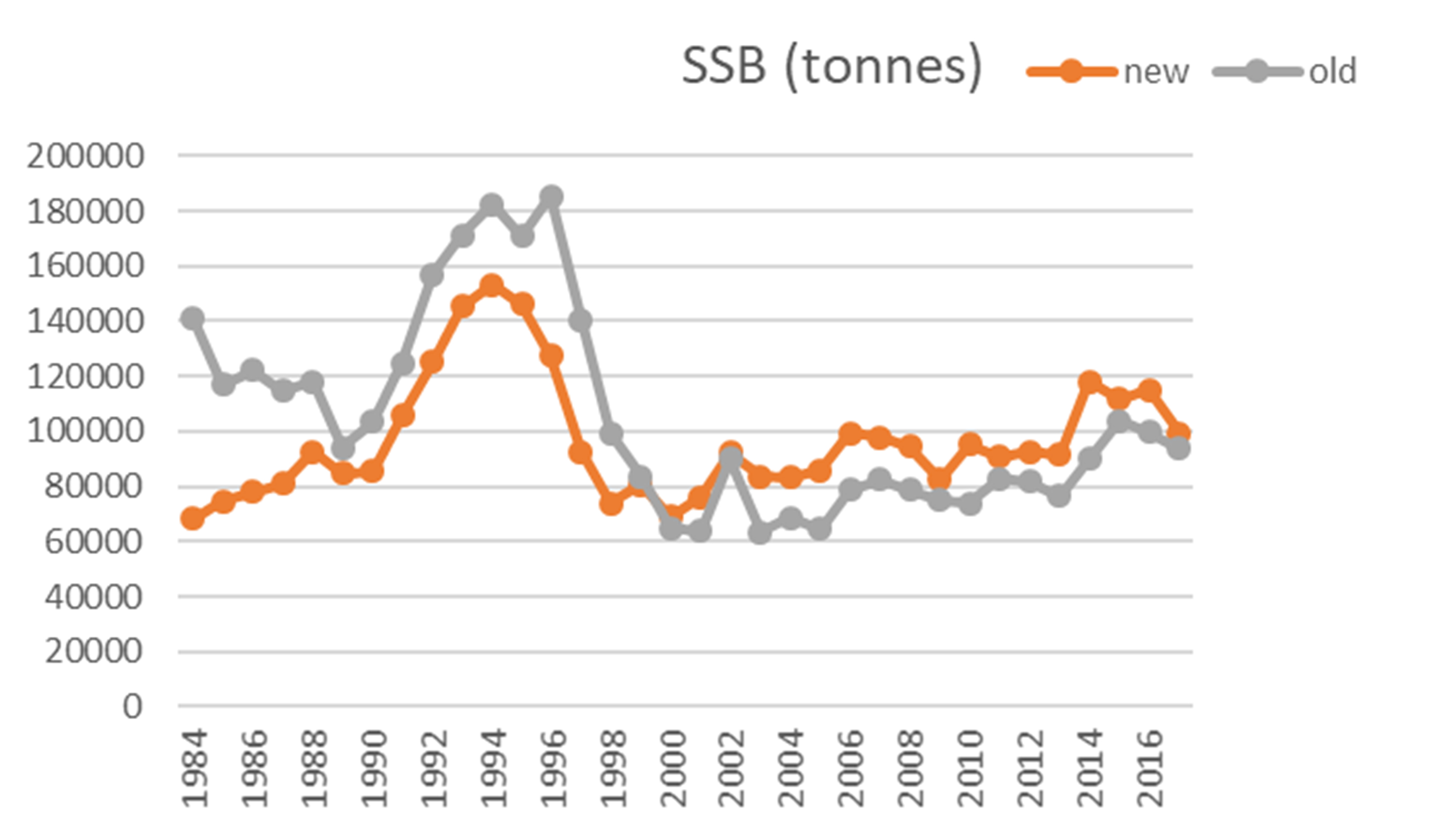
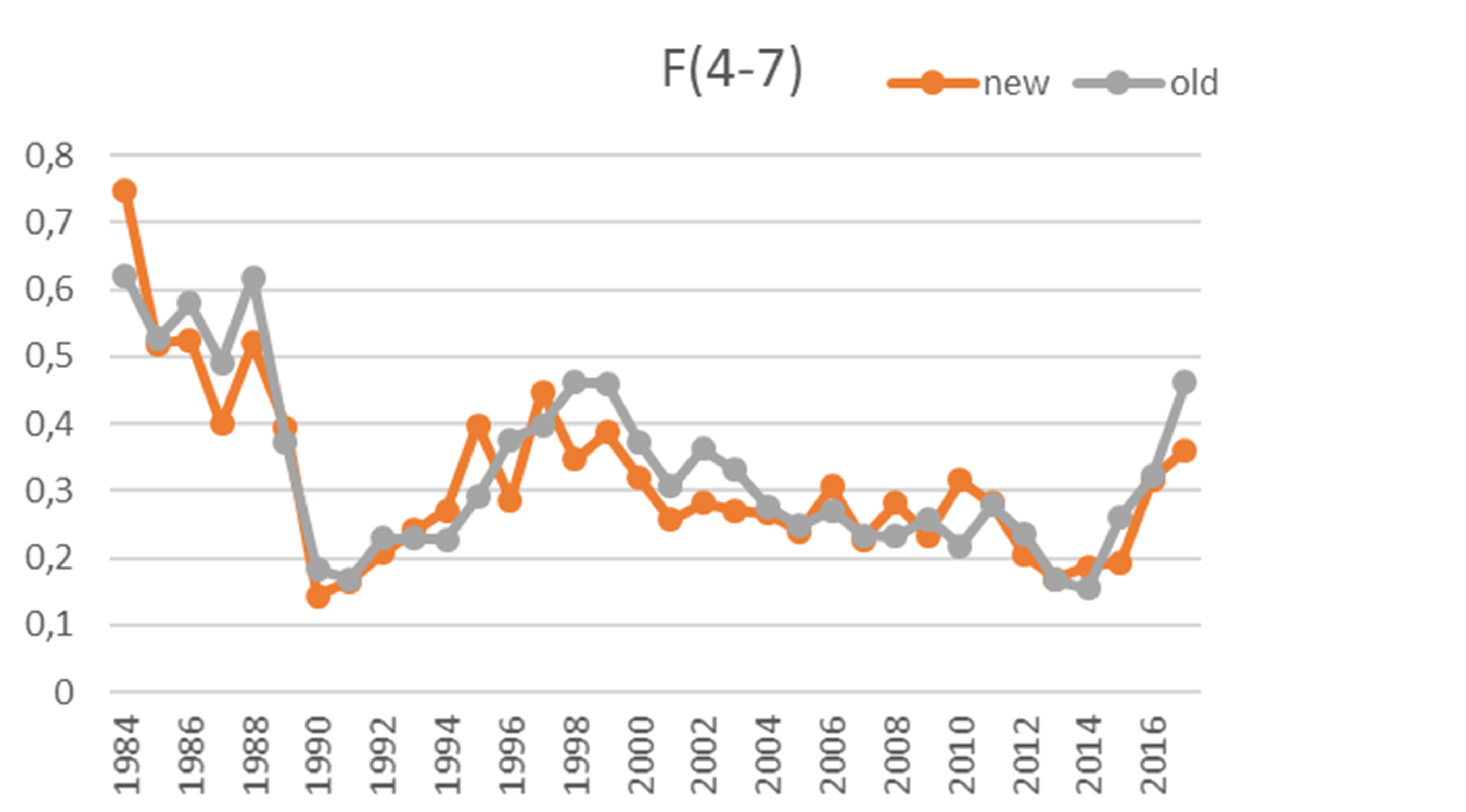
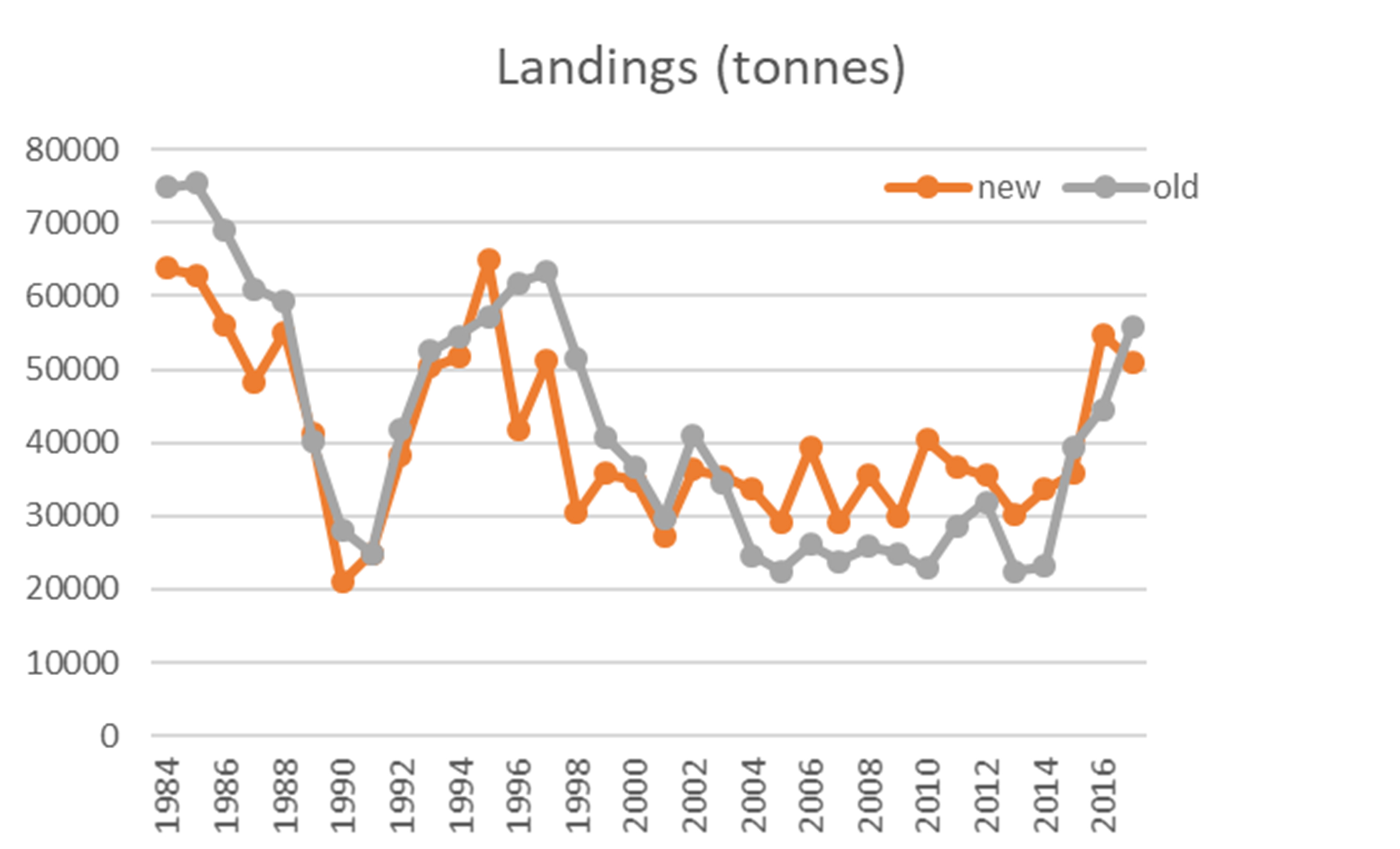
En bestandsberegning (vpa) basert på denne fangst-serien (Figur 5) tilsier at bestanden var på sitt laveste i de første 4 årene av tidsserien (1984-1987), mens beregningen med den gamle fangst-serien viser en bestand som i de 4 første årene var i overkant av langtids-gjennomsnittet. Den nye beregningen bekrefter at bestanden var på sitt høyeste rundt 1994, men forskjellen mellom dagens bestand og høyest observert bestand er litt mindre.
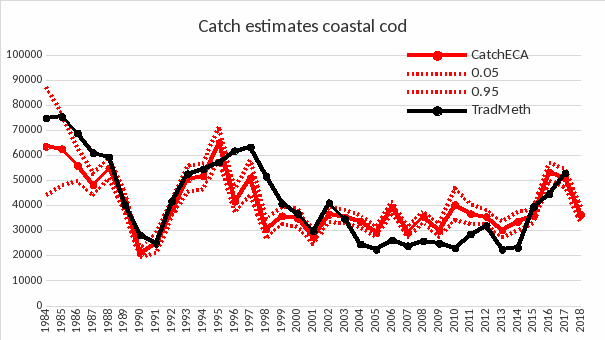
Figur 4. Beregnet fangst av kysttorsk med tradisjonell metode (sort linje) og ved bruk av ECA Rød linje for median-verdi, og stiplet rød linje for 5-persentilen og 95-persentilen.
Figur 5. Tentativ bestandsberegning basert på gamle fangstdata (grå linjer) og basert på nye fangstdata (oransje linjer). Øverst gytebestand (SSB), midterst Fiskedødelighet (F) som årlig gjennomsnitt for aldersgruppene 4-7 år (ICES 2019b), og nederst gamle og nye fangstdata (landingsdata).
2.3 - Tokt-målinger Kystressurstoktet
Tidsserie fra Kystressurs-toktet Varanger-Stad utført med to av Havforskningsinstituttets forskningsfartøy i perioden oktober-november hvert år (80 toktdøgn) (Figur 6).
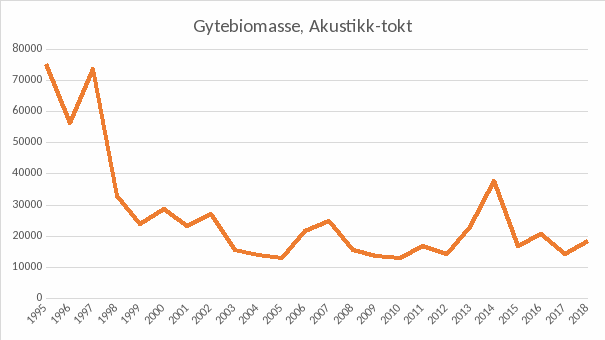
Figur 6. Beregnet gytebiomasse (tonn) kysttorsk nord for 62˚N. Basert på antakelse om den reelle fisketettheten per enhet akustikkverdi.
De eldste data anses mer usikre enn de nyere. Toktserien er basert på akustiske målinger, som før innføring av senkekjøl, var sterkt påvirket av værforholdene. I tillegg ble metodikken for kalibrering av ekkoloddene betydelig forbedret rundt 2000. De tre første årene ble det gjennomført mange pelagiske trålhal som i enkelte områder dokumenterte betydelig pelagiske forekomster av ungtorsk. I etterfølgende år ble det avsatt mindre tokt-tid, slik at få pelagiske hal har blitt prioritert. Senere erfaringer er at det er hyse som dominerer de pelagiske ekkolodd-registreringene. Det blir imidlertid diskutert om toktindeks-nivået etter 1995-1997 kan være noe underestimert i forhold til disse første årene i tidsserien.
Rapportering av landinger på statistiske lokaliteter har gradvis blitt bedre. I eldre data er det en tendens til at fangstene ble rapportert til den lokaliteten hvor fiskemottaket lå, selv om fangsten egentlig var tatt i en annen lokalitet. Innsamling av aldersprøver (otolitter) er avgjørende for presisjonen av både splitting av kysttorsk og skrei, og for beregning av antall fisk per aldersgruppe. Før 1984 er det for lite otolitt-prøver til å beregne fangst av kysttorsk. Denne prøvetakingen har økt, og er nå i større grad lagt opp i henhold til gode statistiske retningslinjer. Etablering av Kystreferanseflåten har bidratt betydelig til å bedre prøvedekning i tid og rom.
Da gjeldene gjenoppbyggingsplan ble foreslått, ble de akustiske beregningene fra kystressurstoktet valgt som referanse, ettersom den fangst-baserte bestandsberegningen var ansett som svært usikker (og ikke godkjent av ICES). Gjenoppbyggingsmålet i den gjeldende gjenoppbyggingsplanen ble definert som gjennomsnittet av toktmålingene av gytebestanden i de fire første årene i tidsserien (1995-1998). Toktet gjennomføres 3-4 måneder før gyting, og det er vanskelig å beregne andelen av fisk som skal gyte i kommende sesong. Beregnet gytebiomasse har derfor større usikkerhet enn totalbiomassen. Som nevnt over, er de første årene av toktserien (og fangstberegningen) mer usikre enn nyere målinger. Forhåpentligvis kan den nye fangstserien, kombinert med en bunntrålbasert toktindeks for 2 år og eldre fisk gi et bedre grunnlag for beregning av historisk bestandsutvikling og biologiske referansepunkter. Ambisjonene er nå å få etablert en bedre bestandsberegning, som også kan gi grunnlag for beregning av referansepunkter som svarer til maksimering av langtidsutbytte i fisket. Slike referansepunkter er nyttige for å spesifisere et fornuftig gjenoppbyggingsmål, men så lenge bestandsberegningen er usikker, vil også referansepunktene være usikre. Det er planlagt et nytt «ICES-Benchmark» (trolig ferdig i 2021) hvor målet er å bedre bestandsberegningen.
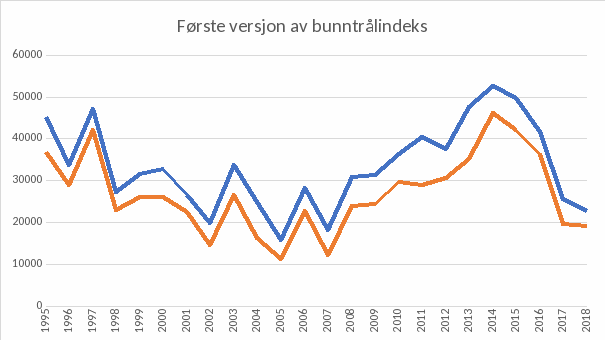
De faste bunntrålstasjonene i Kystressurstoktet er nå tatt i bruk for å beregne bunntrål-indekser. En første versjon av disse beregningene er vist i Figur 8. Denne viser en ganske forskjellig tidstrend sammenliknet med det akustikken viser. Disse forskjellene må undersøkes mer grundig.
Merk at figurene 5, 6 og 7 alle angir gytebiomasse i tonn, men det ligger ulike antakelser i beregningene, som gjør at de ikke er direkte sammenlignbare. Den fangstbaserte («VPA», Figur 5) har en antakelse om naturlig dødelighet lik 0.2, en antagelse om at fangbarheten i fisket og i toktene ikke er avhengig av fisketetthet, og den inneholder fangster fra hele området 03+04+05+00+06+07, som er noe større enn det toktene dekker. Akustikk-toktet(Figur 6) er basert på antakelser om den reelle fisketettheten per enhet akustikkverdi, og bunntrålindeksen (Figur 7) er basert på antakelse om den reelle fisketettheten per enhet bunntrålfangst.
Gjenoppbyggingsmålet på 60 000 tonn, i gjeldende gjenoppbyggingsplan, referer til gytebiomasse i akustikk-toktet. I slike analyser er ofte tidstrendene mer relevante enn absoluttverdiene.
Figur 7. Foreløpig beregning av bunntrålindeks (biomasse for 2år og eldre kysttorsk) basert på antakelse om den reelle fisketettheten per enhet bunntrålfangst (oransje: nord for 67 ˚ N, blå: hele området nord for 62 ˚ N). Data fra Kystressurstoktet.
2.4 - Tokt med trollgarn og åleruser i grunne områder mellom Steigen (68˚N) og Stad (62˚N).
I skjærgården langs Møre, Trøndelag og Helgeland finnes store gruntvannsområder hvor det er kjent at det finnes en del torsk som ikke lar seg kartlegge med store forskningsfartøy. Disse forekomstene er dermed lite representert i kystressurstoktet.
Basert på erfaringer med garn og rusetokt i Lofoten-Senja- området (Sundby et al 2013) er det gjennomført noen tokt med trollgarn og åleruser på grunt vann i området fra Steigen (68 ˚ N) og Vikna (65 ˚ N) i 2013, 2016 og 2018, og i gruntvannsområder mellom Vikna og Stad (62 ˚ N) i 2015, 2017 og 2019.
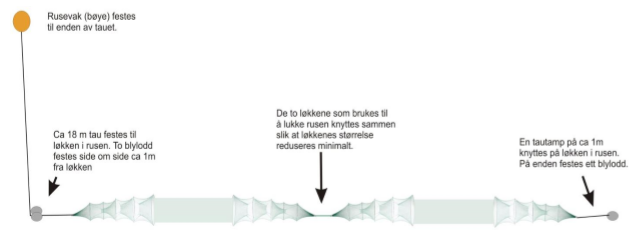
Daglig bruksmengde var 6 stk doble åleruser (Figur 8) og 2 trollgarnsett med 4 enkeltgarn i hvert sett (2 stk 36mm og 2 stk 45 mm stolpelengde).
Det er gjennomført både otolittyping og genetikk-analyser (pan 1) av all torsk fanget i disse toktene. Resultatene viser at alle individer er kysttorsk.
Figur 8. Dobbel åleruse: Fra venstre; ett rusekammer og 3 kalver, så et ledegarn og 3 kalver og et rusekammer. Andre halvdel er et speilbilde av første (fra van der Meeren, 2018).
3 - Resultat
3.1 - Tokt med trollgarn og åleruser i grunne områder mellom Steigen (68˚N) og Vikna (65˚N) i august 2013, 2016 og 2018
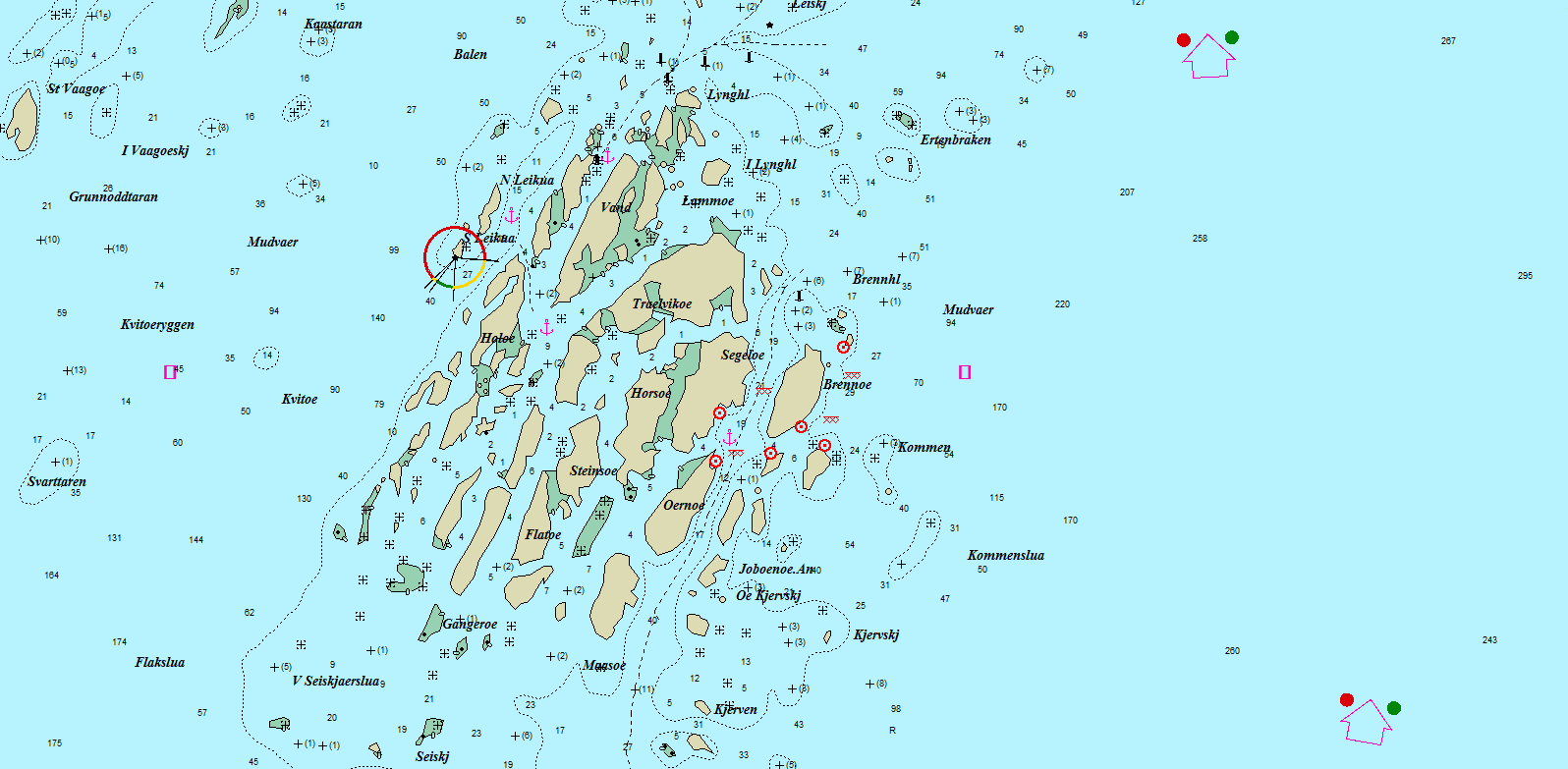
I Steigen-Vikna- området ble det definert 49 aktuelle gruntvannsområder, hver av dem høvelig for et døgns fiske. (Figur 9). Blandt disse ble, 12 fisket i 2013, 21 i 2016 og 20 i 2018. Blandt de 20 fisket i 2018; ble 11 fisket i alle de tre årene, 8 i to år og 4 ble fisket bare ett år. R/V “Fangst” ble benyttet I alle toktene. Kriteriene for valg av områder var et ønske om en rimelig spredning innenfor hele undersøkelses-området, med litt justering i forhold til værforhold og kjøredistansen mellom lokaliteter. Figur 10 viser et eksempel på plassering av redskapene innenfor en valgt lokalitet (Mudvær i Vega).
Figur 9. Forhånds-definerte fiskelokaliteter Steigen-Vikna. (Skala Max Sea; 1:1.5 mill).
Figure 10. Mudvær, Vega; Eksempel på fiskeområde. Røde sirkler er ruse-sett, og xxx er start -og sluttposisjon for trollgarnsettene. (Skala Max Sea; 1:25000).
3.2 - Status for kysttorsk, Steigen-Vikna
I alle de tre årene ble det fanget mest torsk i aldersgruppene 1, 2 og 3 (Tabell 2). Samlet for alle aldersgrupper er det små forskjeller mellom de tre årene i gjennomsnittlig antall torsk pr fiskedag (Tabell 2). Sammenliknet med 2013 og 2016 antyder 2018-resultatene litt økning for nullgruppen (Tabell 3). Variasjonskoeffisienten (CV) tilsier at små endringer fra år til år ikke er signifikante.
# fiske-
# torsk
CV for middelverdi
År
døgn
Pr dag
%
2013
12
42.8
17
2016
21
41.6
14
2018
20
47.5
12
Tabell 2. Gjennomsnittlig antall torsk pr fiskedøgn, Steigen-Vikna
Alder
0
1
2
3
4
5
6
7
8
9+
Total
2013
2.8
16.4
10.8
6.6
3.5
1.3
0.9
0.2
0.2
0.2
42.8
2016
2.9
11.3
12.0
7.0
5.6
1.0
1.2
0.4
0.3
0.1
41.6
2018
6.8
13.2
13.7
5.6
3.4
1.8
2.2
0.3
0.2
0.3
47.5
Tabell 3. Gjennomsnittlig antall torsk pr fiskedøgn fordelt på aldersgrupper
3.3 - Tokt med trollgarn og åleruser i grunne områder mellom Vikna (65˚N) og Stad (62˚N) i august 2015, 2017 og 2019
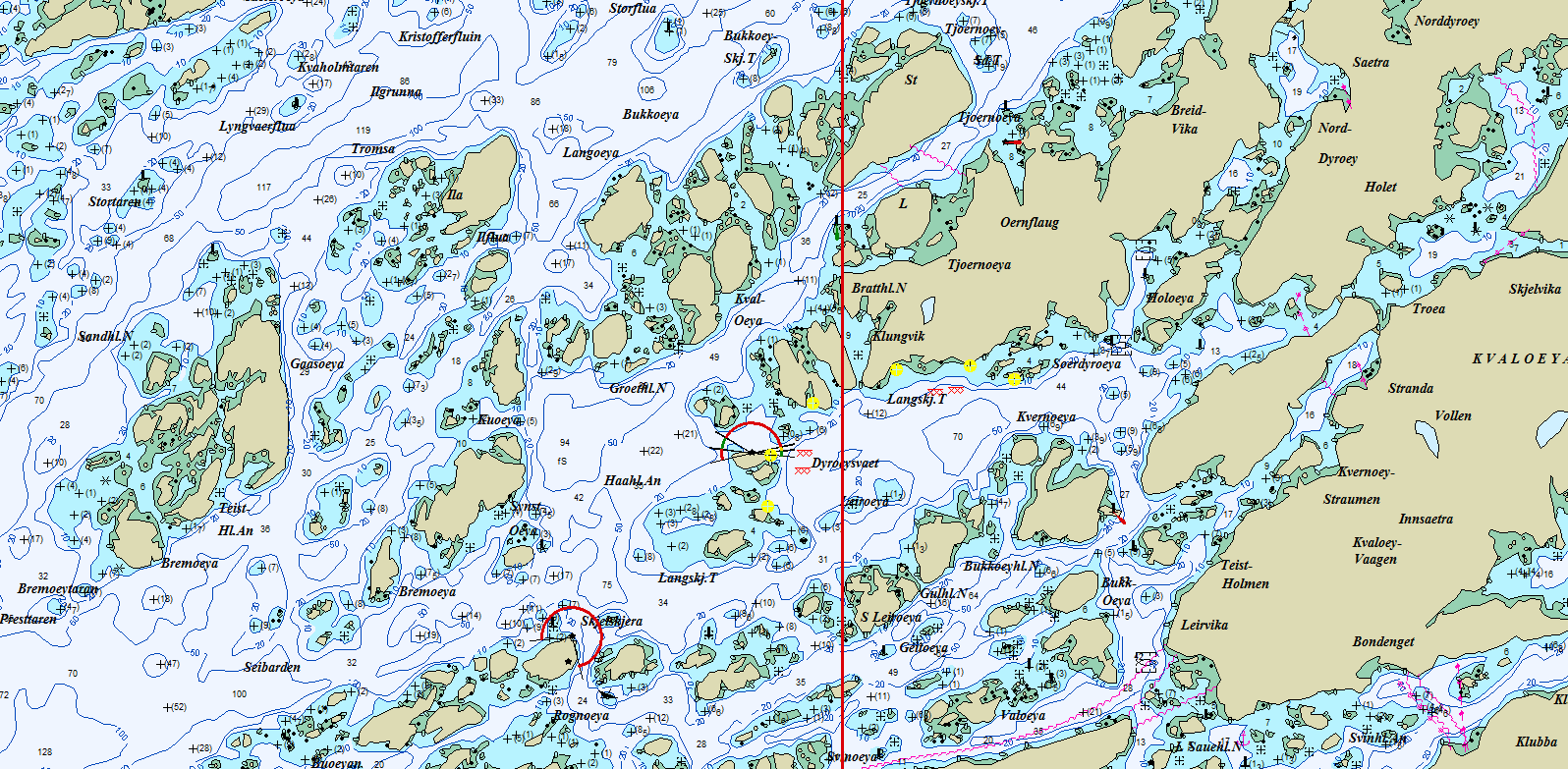
I Vikna-Stad- området ble det definert 46 aktuelle gruntvannsområder, hver av dem høvelig for et døgns fiske. (Figur 11). Blant disse ble, 23 fisket i 2015, 21 i 2017 og 22 in 2019. Blant de 22 fisket i 2019; har 20 blitt fisket enten i 2015 eller i 2017. To av lokalitetene i 2019 (Ona and Nordøyan) har ikke vært fisket tidligere år. F/F “Fangst” ble benyttet til alle toktene. Som i Steigen-Vikna var kriteriene for valg av områder et ønske om en rimelig spredning innenfor hele undersøkelses-området, med litt justering i forhold til værforhold og kjøredistansen mellom lokaliteter. Figur 12 viser et eksempel på plassering av redskapene innenfor en valgt lokalitet (Dyrøysvaet, Frøya).
Figur 11. Forhånds-definerte fiskelokaliteter Vikna-Stad. (Skala Max Sea; 1:1.5 mill).
Figur 12. Dyrøysvaet, Frøya; Eksempel på fiskeområde. Gule sirkler er ruse-sett, og xxx er start -og sluttposisjon for trollgarnsettene. (Skala Max Sea; 1:25000).
3.4 - Status for kysttorsk, Vikna-Stad
Samlet for alle aldersgrupper er det små forskjeller mellom de tre årene i gjennomsnittlig antall torsk pr fiskedag (Tabell 4). Sammenliknet med 2015 og 2017 ligger 2019-resultatene litt lavere for 4 år og eldre torsk (Tabell 5). Variasjonskoeffisienten (CV) tilsier at små endringer fra år til år ikke er signifikante.
# fiske-
Gj.sn ant
CV for middelverdi
År
døgn
torsk pr dag
%
2015
23
19.6
13
2017
21
22.1
11
2019
22
18.6
16
Tabell 4. Gjennomsnittlig antall torsk pr fiskedøgn Vikna-Stad
Ader
0
1
2
3
4
5
6
7
8
9+
Total
2015
0.10
6.57
5.76
3.34
1.31
1.31
0.71
0.20
0.20
0.00
19.6
2017
3.52
9.92
3.85
1.90
1.20
0.86
0.41
0.19
0.14
0.05
22.1
2019
1.09
9.28
4.12
2.31
0.77
0.76
0.08
0.09
0.09
0.00
18.6
Tabell 5. Gjennomsnittlig antall torsk pr fiskedøgn fordelt på aldersgrupper
Fangstratene for torsk i denne regionen (Tabell 4 og 5) var generelt lavere enn i området Steigen- Vikna (Tabell 2 og 3). I alle de tre årene ble det fanget mest torsk i aldersgruppene 1, 2 og 3.
3.5 - Fangst per garndøgn (CPUE) fra kystfiskeriene som indeks på bestandsendringer
Årlige indekser for størrelsen på fiskebestander basert på fangst- og innsatsdata er benyttet i mange fiskeribestandsvurderinger. Fangst per enhet innsats (CPUE) er et indirekte mål på forekomst av en art. Endringer i fangst per innsatsenhet indikerer endringer i artens virkelige størrelse. En synkende CPUE indikerer bestands-/populasjonsnedgang og overbeskatning, mens en stabil eller økende CPUE indikerer bærekraftig høsting. Det har blitt mer vanlig de siste årene å bruke avanserte statistiske metoder for å standardisere fangstrater mot forklaringsvariabler som et middel for å justere indekser for ulik prøvetaking over rom eller tid eller habitat/område. Vanlige brukte metoder inkluderer generelle lineære modeller (GLM), generaliserte lineære blandede modeller (GLMM), for ikke-normalfordelte data, delta - lognormale eller delta - gamma GLMMer og generaliserte additive modeller (GAM).
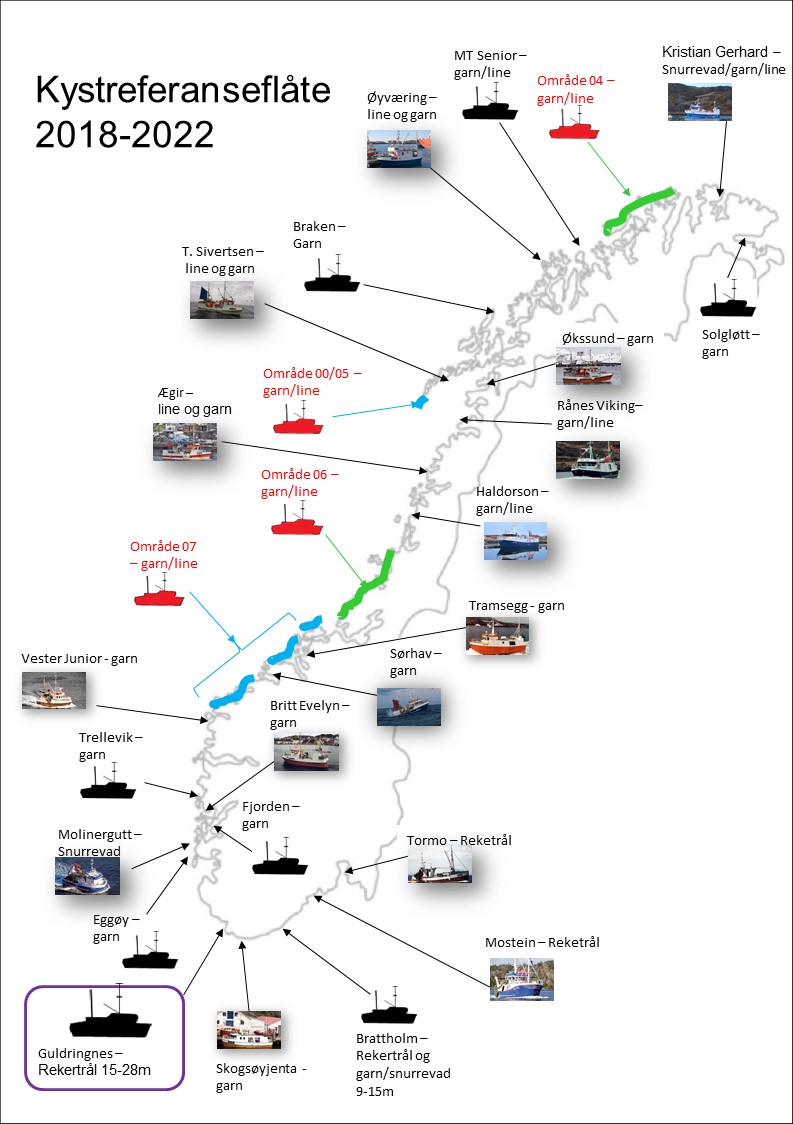
Siden slutten av 2005 har 10-13 garnbåter mellom 9 og 15 meter vært med i HIs kystreferanseflåte nord for 62 ˚ N (Figur 13). Kystreferanseflåten tar prøver av egen fangst og leverer data direkte til Havforskningsinstituttet, både av beholdt fangst og ikke-omsettelig fangst. Detaljert informasjon om redskap, fiskeinnsats og fangst blir gitt gjennom denne ordningen.
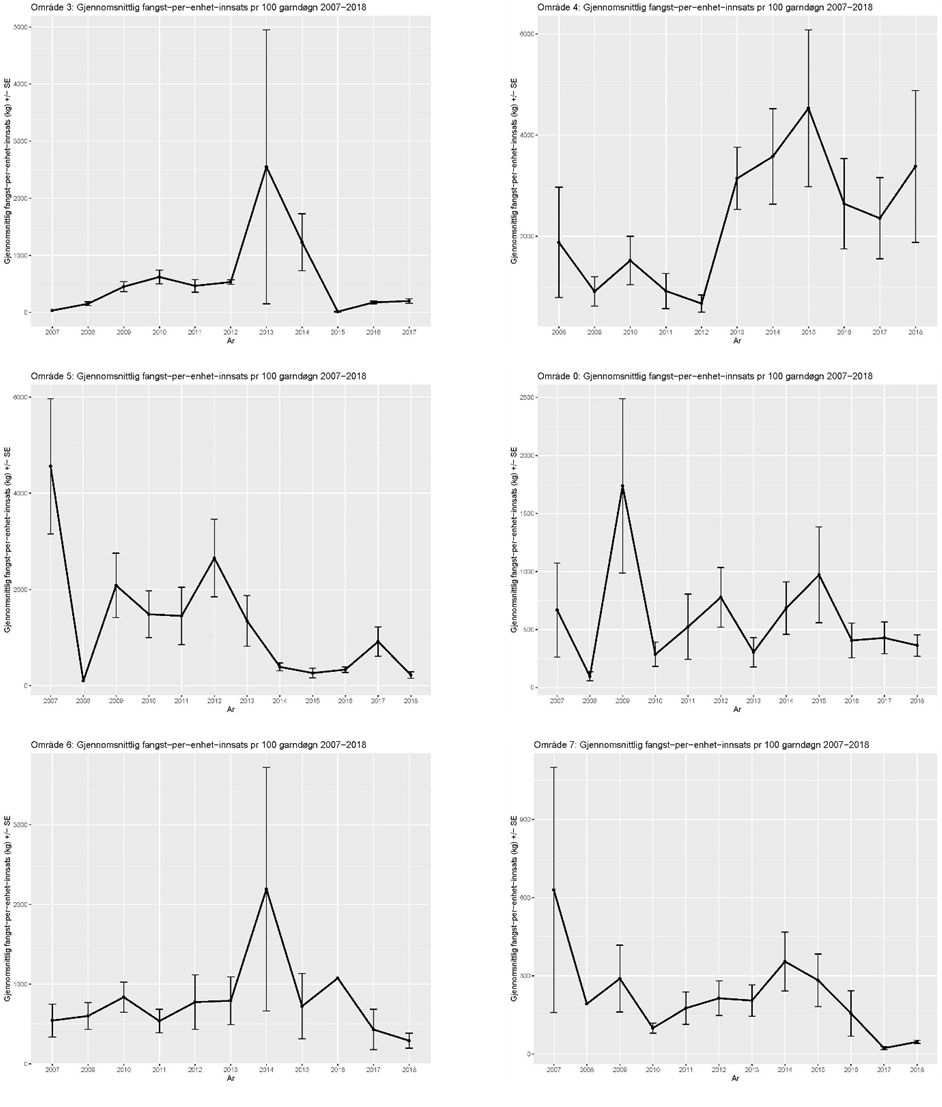
Figur 14. Gjennomsnittlig CPUE (kg kysttorsk per 100 garndøgn) per statistikkområde (03, 04, 05, 00, 06 og 07) for alle fartøyer i kystreferanseflåten som har fisket nord for 62 ˚ N med torskefisk-garn i perioden 2007-2018. Foreløpige data. Merk ulik skala på y-aksen. Vertikale linjer viser standardfeilen til middelverdien.
Datagrunnlaget for Figur 14 må sees på som foreløpig. Det pågår en kvalitetssikring av disse data. Noen topper i figurene har store standard-feil som må sjekkes nærmere. Etter 2013 er tendensen at de høyeste fangstratene for kysttorsk har vært i Område 4. Fangstratene indikerer en signifikant reduksjon i kysttorskforekomstene i noen områder (område 05, 06, og særlig 07). Dersom slike CPUE-serier skal brukes i overvåkningen av de ulike kysttorskpopulasjonene, så bør mer avanserte statistiske metoder benyttes for å standardisere fangstratene mot relevante forklaringsvariabler, bl.a. for å justere indekser for ulik prøvetaking over rom, tid og habitat/område.
3.6 - Kartlegging av gytefelt Nord for 62 grader
Gytefelt for kysttorsk nord for Stad er kartlagt på 18 tokt gjennomført i perioden 2008 til 2018 (Tabell 5). Tokt er gjennomført ved at det er gjort vertikale håvtrekk fra 50m og opp til overflaten. Fra alle prøver som ble samlet, ble alle egg artsbestemt så langt det var mulig, og torskeeggene ble bestemt til art og utviklingsstadium.
Tokt nr.
Område
Årstall
1
Møre og Romsdal – Stad til Ålesund
2016
2
Møre og Romsdal – Ålesund til Molde
2017
3
Møre og Romsdal – Molde til Sør av Hitra
2018
4
Sør-Trøndelag - Trondheimsleia
2008
5
Sør-Trøndelag – Ørland - Osen
2009
6
Nørd Trøndelag - indre deler
2010
7
Nord Trøndelag – Vikna (og Frøya/ Hitra)
2018-2019
8
Nordland – Bindal til Vega
2011
9
Nordland – Vega til Mo i Rana
2012
10
Nordland – Lurøy til Bodø
2014
11
Nordland – Saltfjorden - Steigen
2013
12
Nordland – Sagfjorden til Tysfjord
2012
13
Nordland/ Troms – Ofotfjorden til Senja
2011
14
Troms – Malangen, Balsfjord og Ullsfjord
2010
15
Troms - Storfjorden
2009
14
Troms - Kvænangen
2010
16
Finnmark - Altafjorden
2015
17
Finnmark – Sandlandsfjorden til Porsangerfjorden
2016
18
Finnmark – Laksefjorden til Varangerfjorden
2017
Tabell 5: Oversikt over tokt gjennomført for kartlegging av gytefelt for kysttorsk.
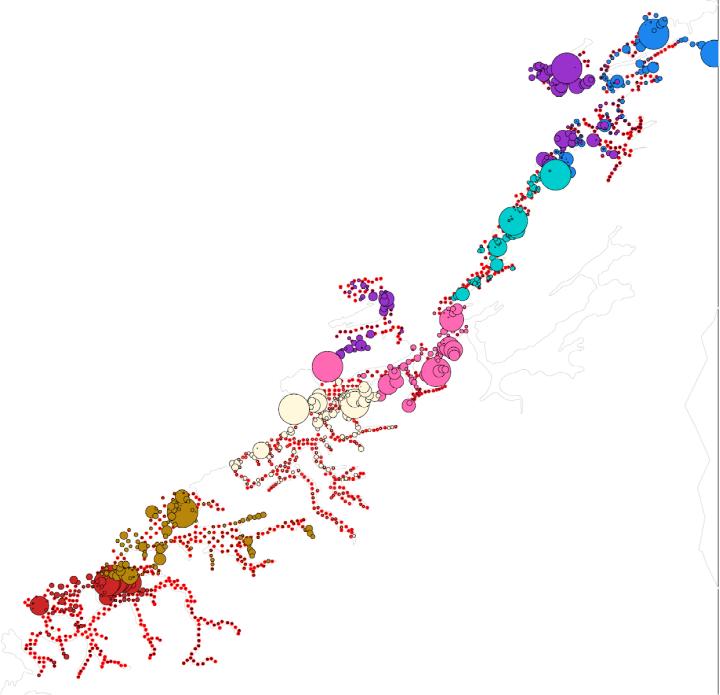
Figur 15: Fordeling av egg i område Møre og Romsdal, Sør- Trøndelag og Nord-Trøndelag. Røde prikker viser stasjoner som ble undersøkt. Sirkler er skalert i forhold til antall egg. Ulike farger på sirklene markerer ulike tokt. Sirklene er kun skalert i forhold til egg funnet på samme tokt, slik at stasjonene med flest egg hvert år vil ha like store sirkler selv om det absolutte nivået av egg er forskjellig.
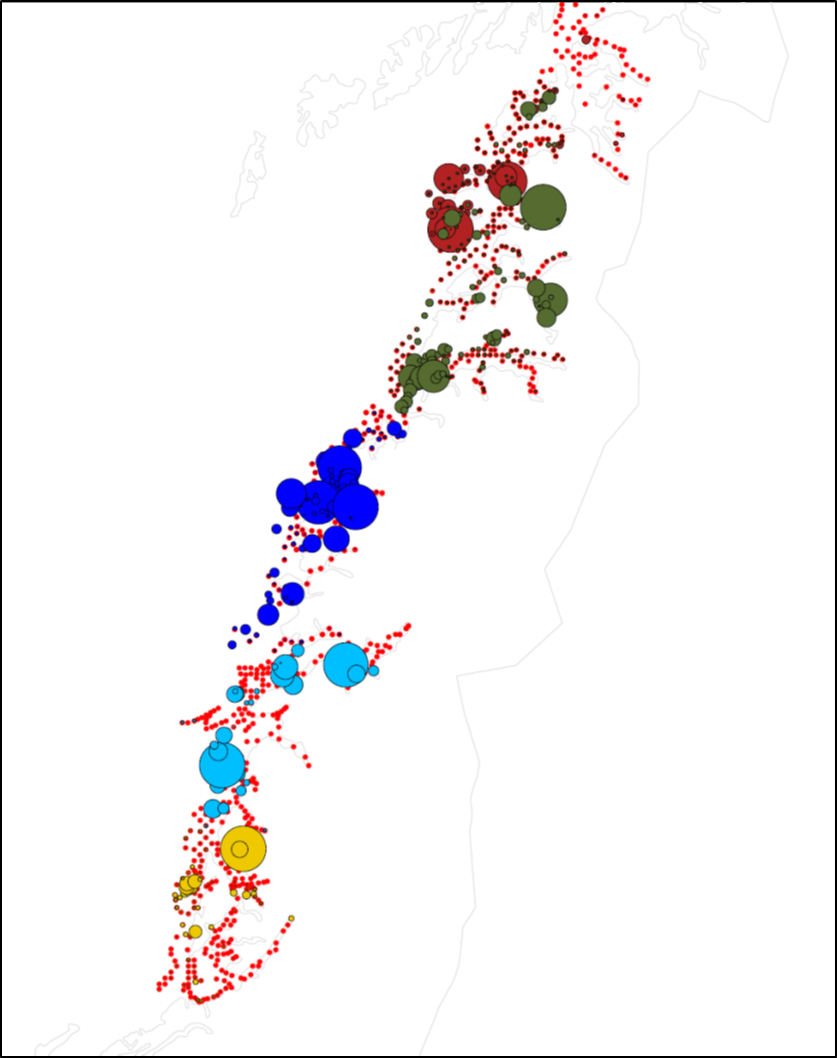
Figur 16: Fordeling av egg i Nordland. Røde prikker viser stasjoner som ble undersøkt. Sirkler er skalert i forhold til antall egg. Ulike farger på sirklene markerer ulike tokt. Sirklene er kun skalert i forhold til egg funnet på samme tokt, slik at stasjonene med flest egg hvert år vil ha like store sirkler selv om det absolutte nivået av egg er forskjellig.
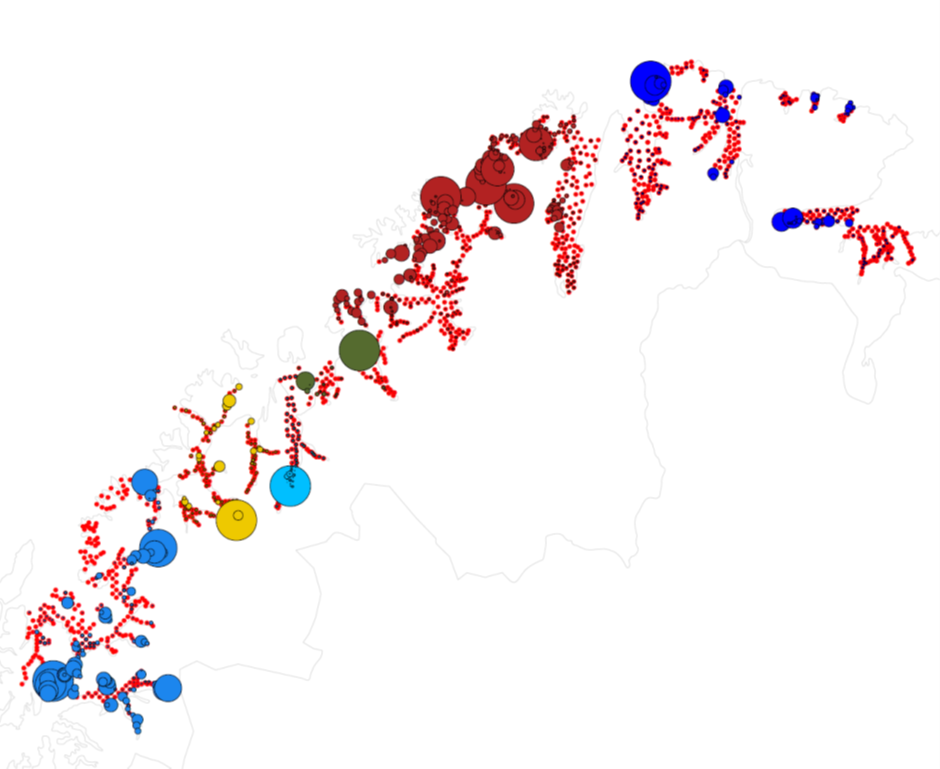
Figur 17: Fordeling av egg i Troms og Finnmark. Røde prikker viser stasjoner som ble undersøkt. Sirkler er skalert i forhold til antall egg. Ulike farger på sirklene markerer ulike tokt. Sirklene er kun skalert i forhold til egg funnet på samme tokt, slik at stasjonene med flest egg hvert år vil ha like store sirkler selv om det absolutte nivået av egg er forskjellig.
Figurene 15-17 viser fordeling av egg som ble artsbestemt for å være torsk. Det er likevel vanskelig å visuelt skille egg fra torsk og noen andre arter torskefisk. I enkelte områder vil det også kunne være store innslag av egg fra skrei som kan komme drivende inn på kysten.
For materialet som ble samlet de siste årene (2017 og 2018) er det blitt etablert en god protokoll for både artsidentifikasjon og for å skille kysttorsk og skrei. Dette viser at mye av de store konsentrasjonene av egg i Finnmark (mørkerøde og mørkeblå sirkler i Figur 17) er skreiegg som har drevet inn. Store konsentrasjoner av skreiegg vil i flere tilfeller da «overskygge» konsentrasjoner av kysttorskegg inne i fjordene. I noen tilfeller er det også store konsentrasjoner av kysttorskegg i ytre deler av kysten som også kan overskygge lave konsentrasjoner av egg i indre deler av kysten, men som likevel representerer viktig lokal gyting.
For å finne hvor det er viktige konsentrasjoner av egg, er det gjort en analyse hvor hver stasjon er sammenlignet mot et nabolag for å se om mengden egg er signifikant mer enn i det lokale nabolaget.
For hvert gytefelt er det gjort en modellering av drift for å undersøke hvor egg fra de forskjellige gytefeltene vil drive i en periode etter gyting. Siden kartleggingen har hatt et fokus på kartlegging av gytefelt for stasjonære bestander av kysttorsk, har bare gytefelt der det er stor grad av retensjon blitt registrert som gytefelt.
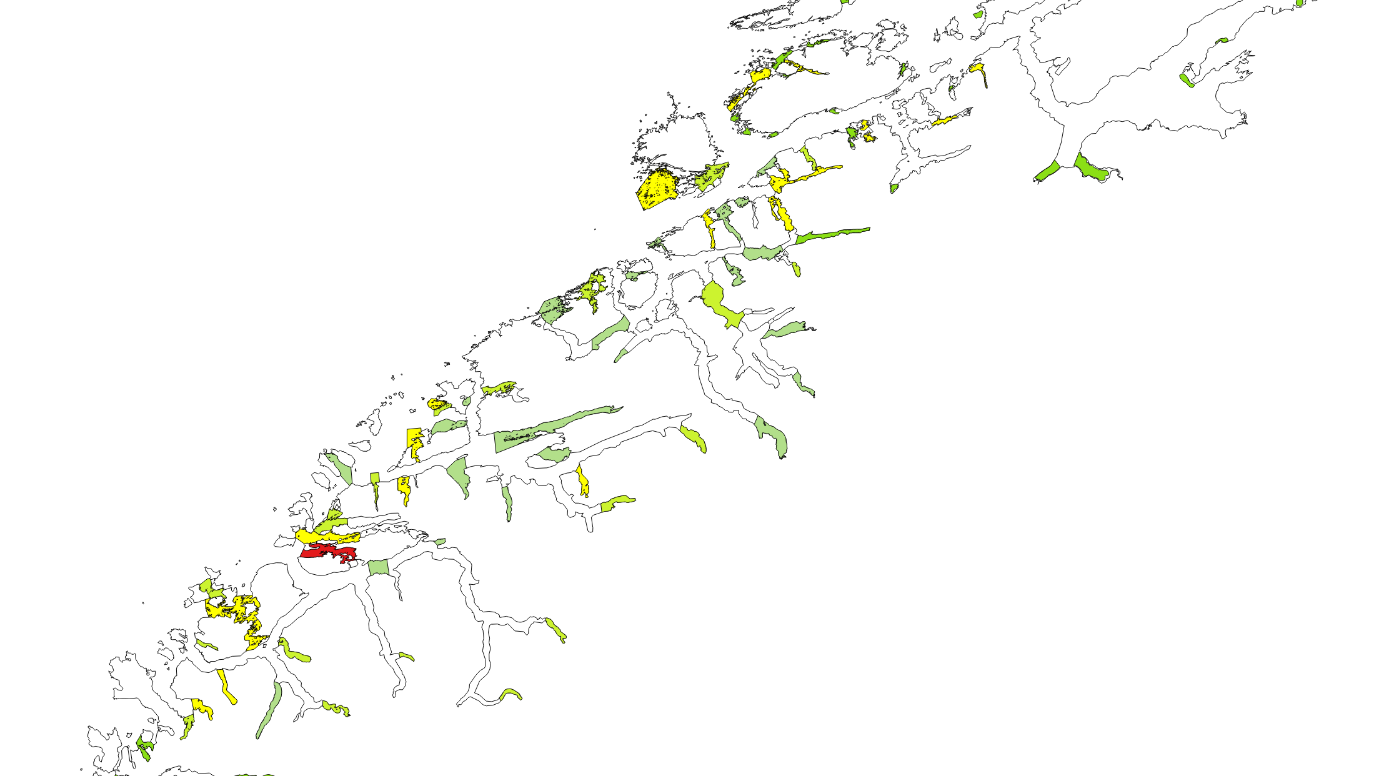
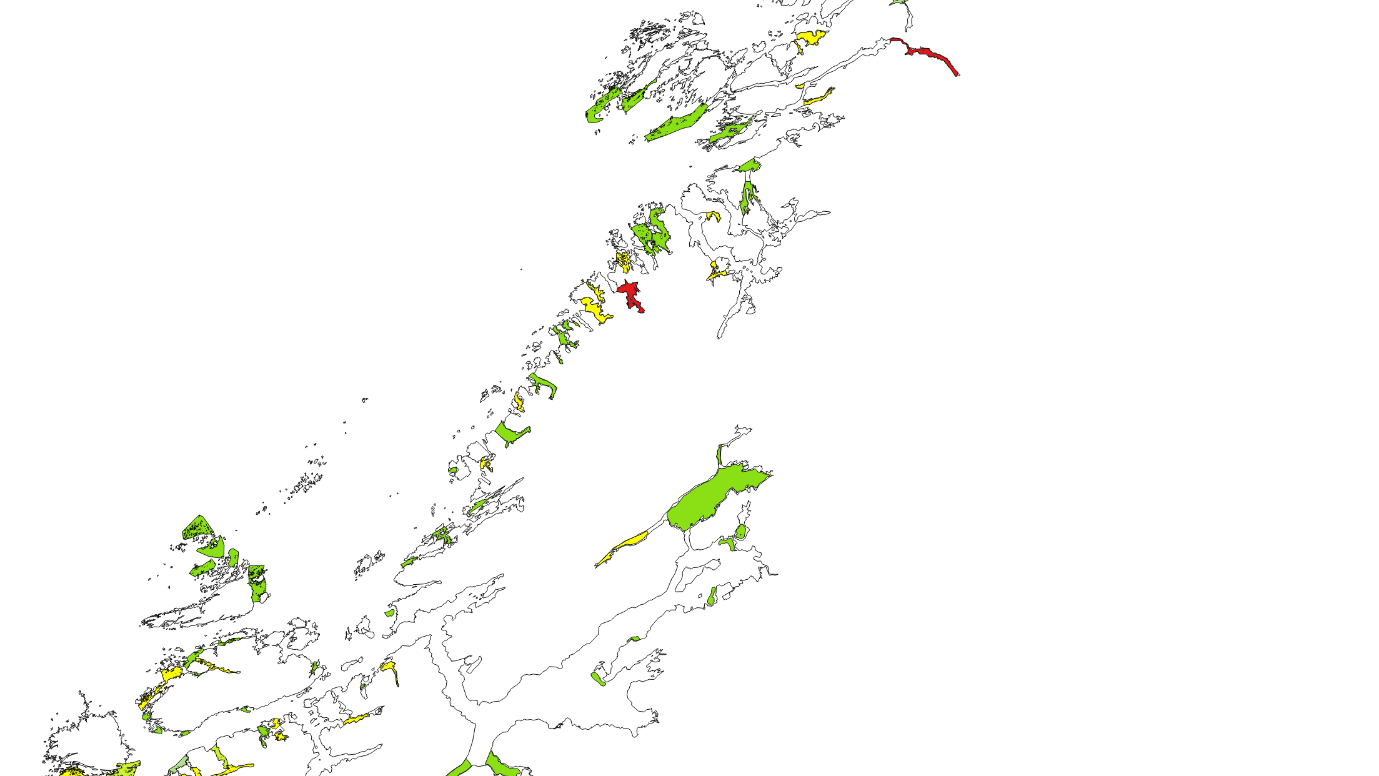
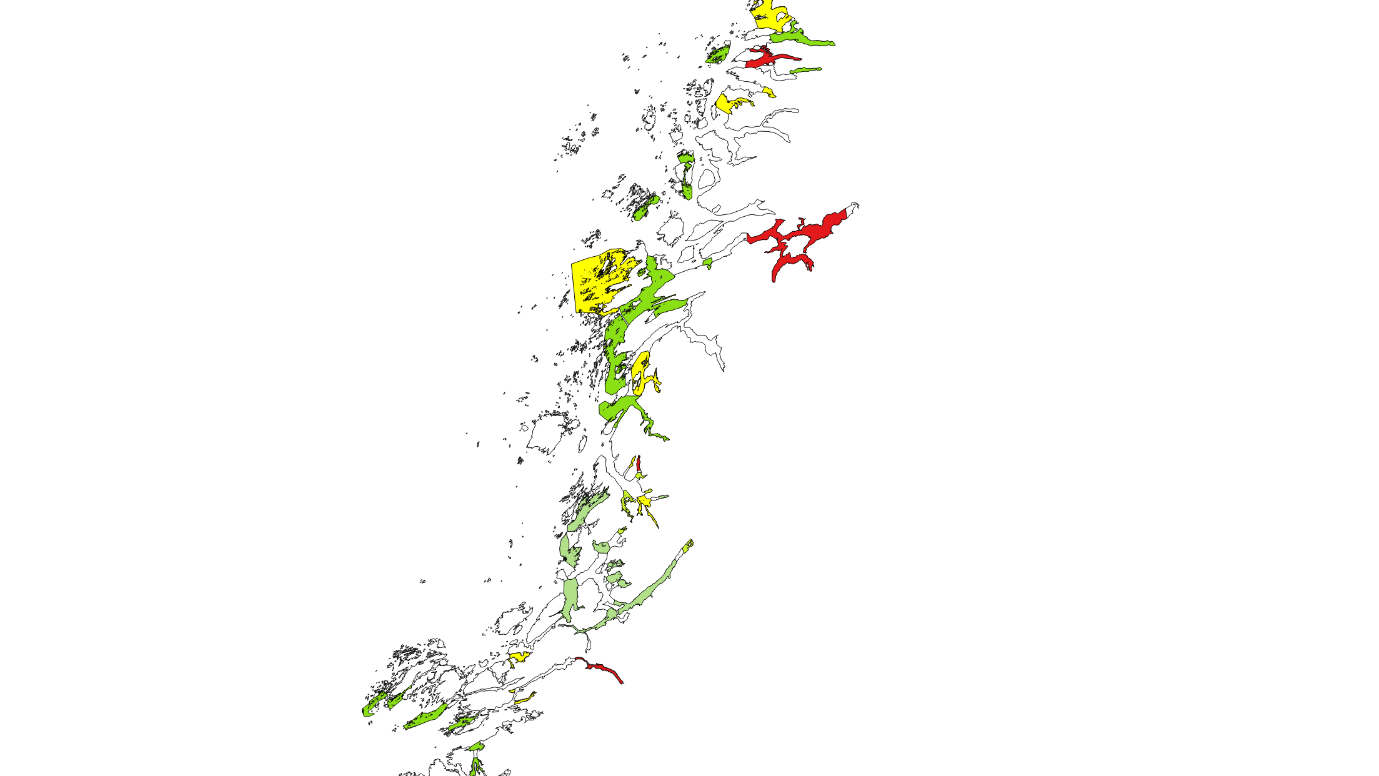
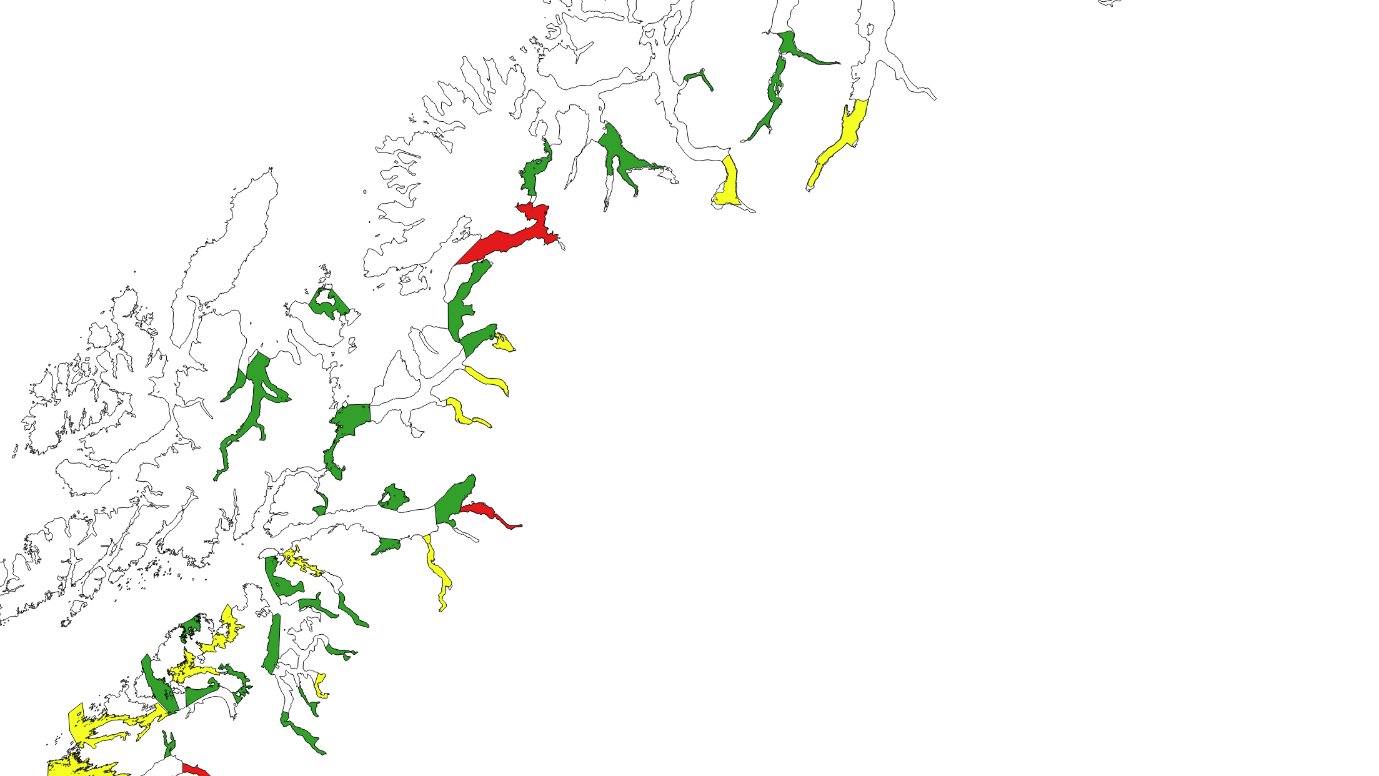
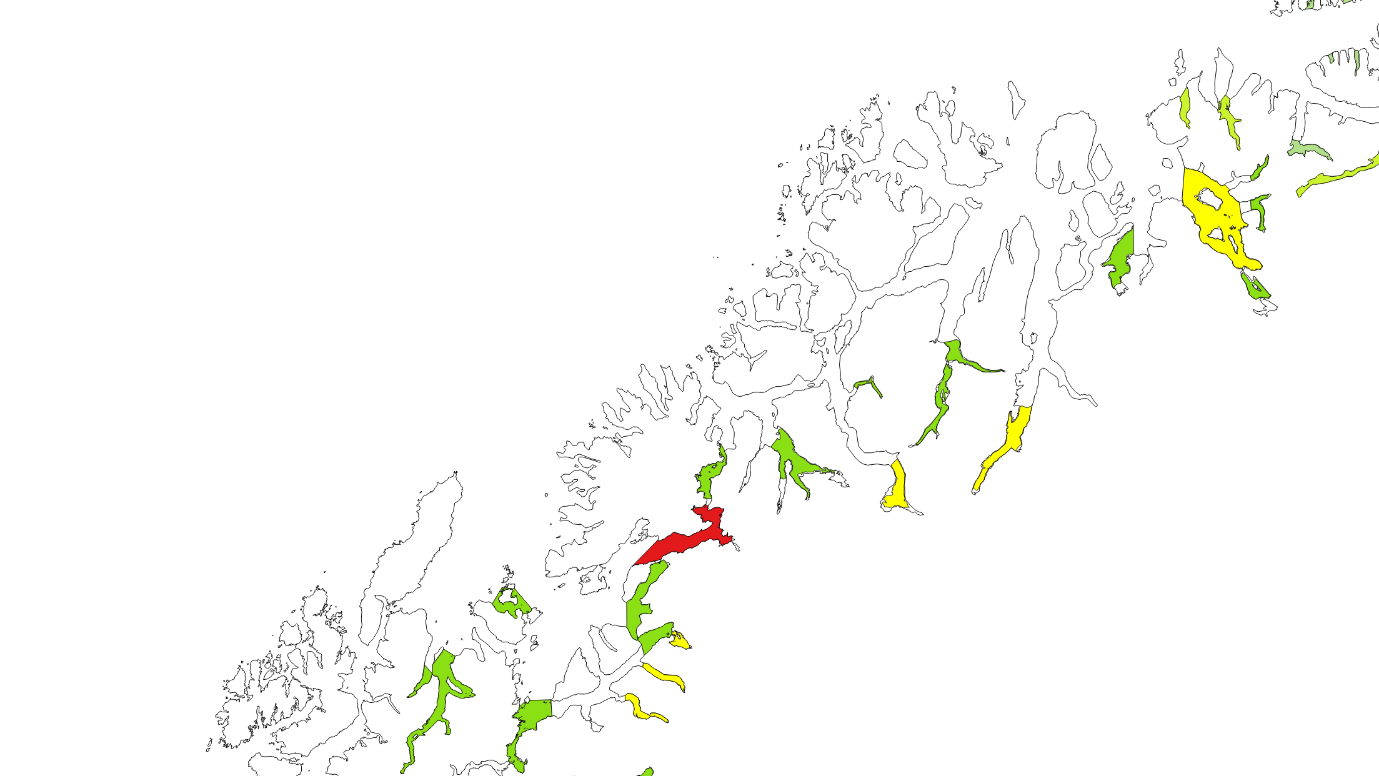
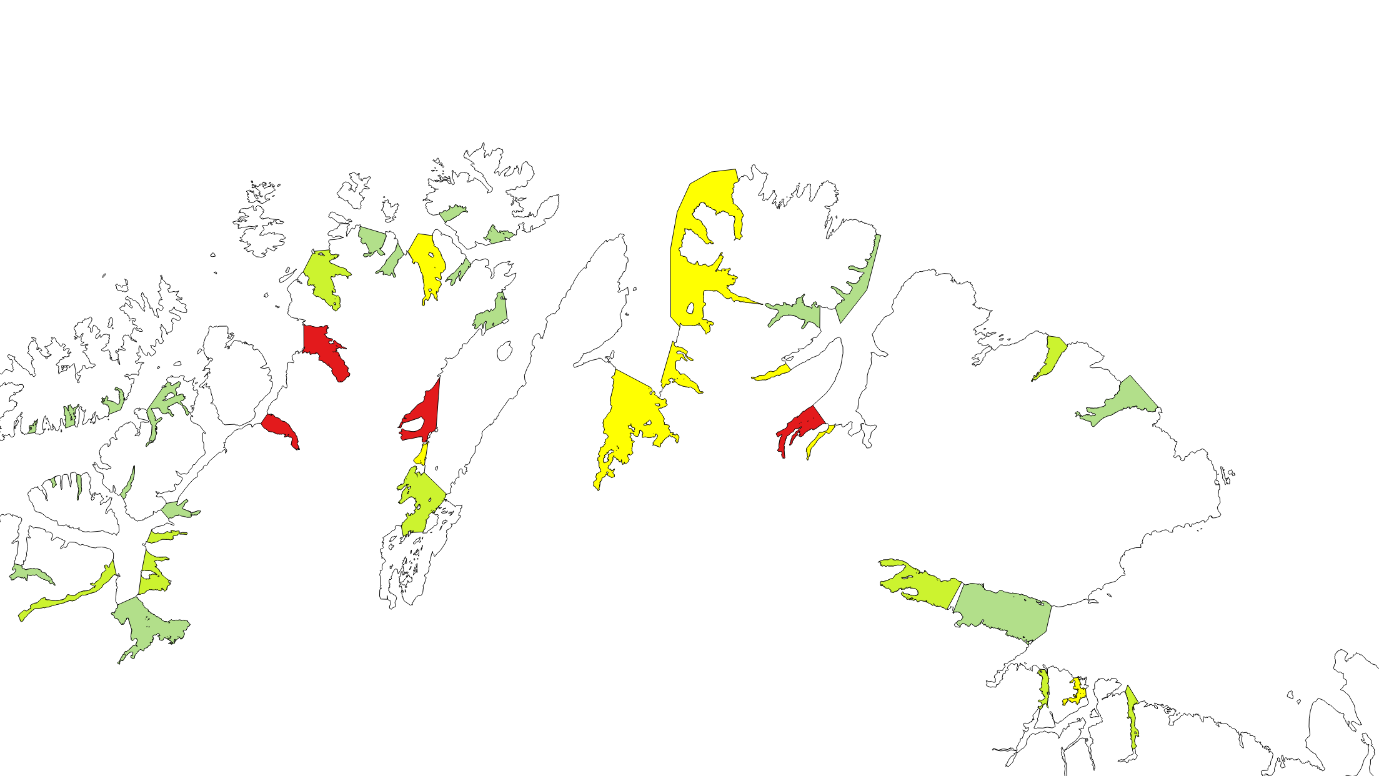
Kartlagte og verdisatte gytefelt for stasjonære bestander av kysttorsk ligger som «Gytefelt Torsk MB» på Fiskeridirektoratets kartside Yggdrasil (Figur 18-23).
Figur 18. Verdisatte gytefelt i Møre og Romsdal basert på kartleggingstokt. Gytefelt med kategori A er røde (nasjonalt viktig), B er gule (regionalt viktig), og de andre er grønne (lokalt viktig).
Figur 19. Verdisatte gytefelt Trøndelag. Gytefelt med kategori A er røde (nasjonalt viktig), B er gule (regionalt viktig), og de andre er grønne (lokalt viktig). Vikna-området er f.eks. et viktig gytefelt med masse egg, men har fått grønn farge pga liten retensjon (dvs eggene driver raskt videre bort fra gyteområdet).
Figur 20. Verdisatte gytefelt Nordland sør. Gytefelt med kategori A er røde (nasjonalt viktig), B er gule (regionalt viktig), og de andre er grønne (lokalt viktig).
Figur 21. Verdisatte gytefelt Nordland nord. Gytefelt med kategori A er røde (nasjonalt viktig), B er gule (regionalt viktig), og de andre er grønne (lokalt viktig).
Figur 22. Verdisatte gytefelt Troms. Gytefelt med kategori A er røde (nasjonalt viktig), B er gule (regionalt viktig), og de andre er grønne (lokalt viktig).
Figur 23. Verdisatte gytefelt Finnmark. Gytefelt med kategori A er røde (nasjonalt viktig), B er gule (regionalt viktig) og de andre er grønne (lokalt viktig).
3.7 - Vern av gytefelt – overvåkning og måling av reguleringseffekter
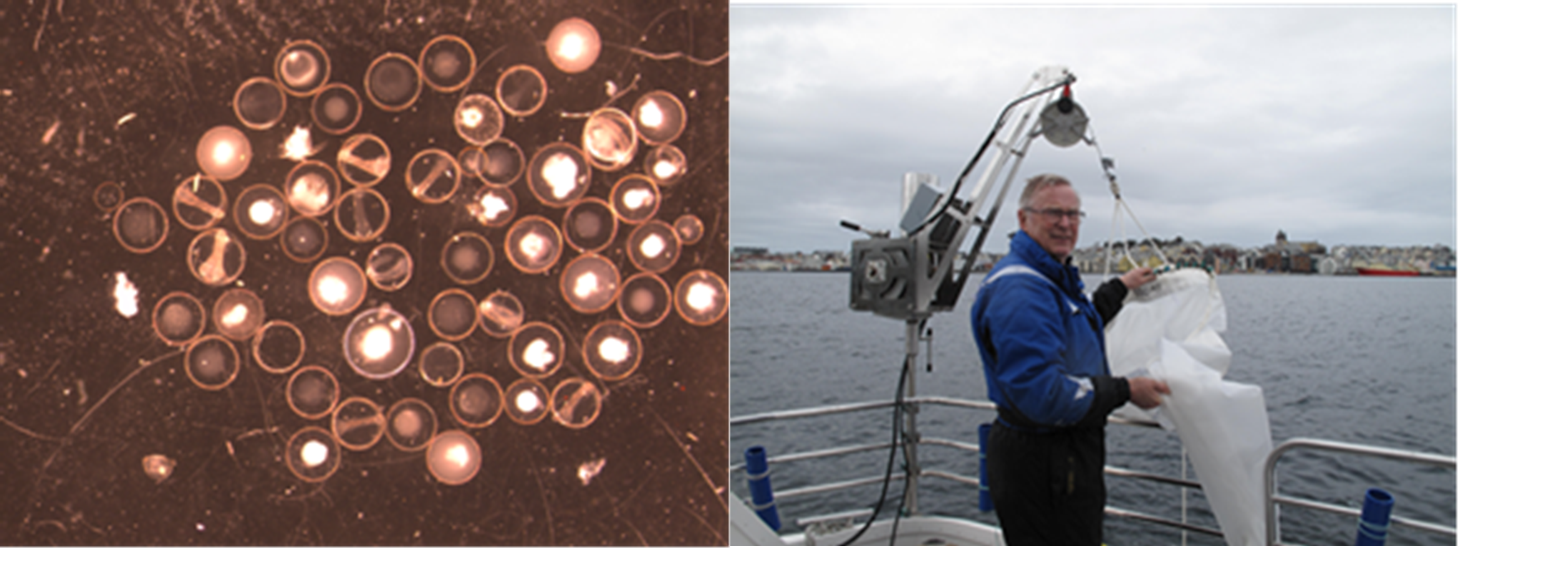
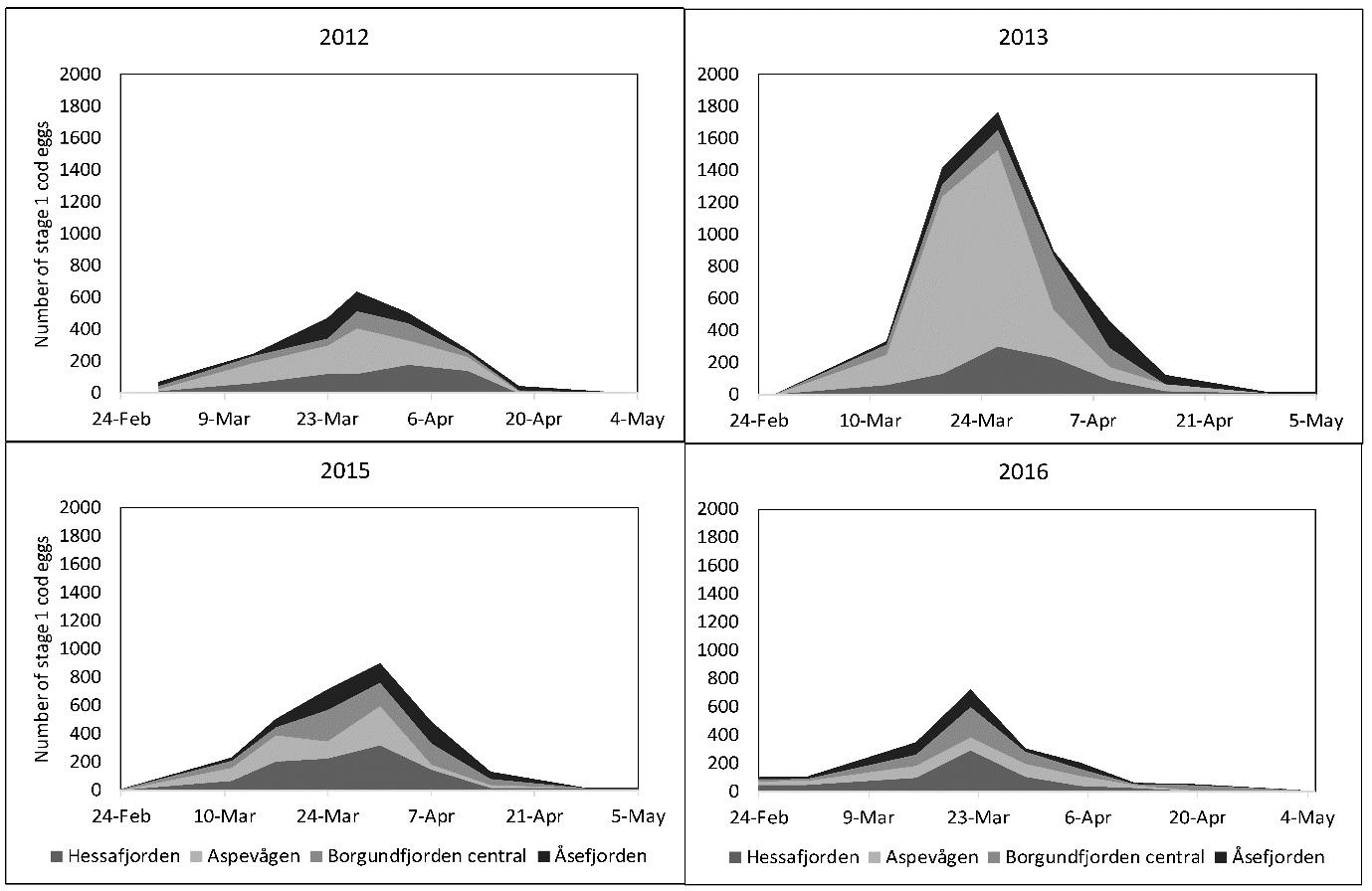
Ved å samle inn egg (og sortere de på art og utviklingsstadium) flere ganger gjennom hele gyteforløpet til torsk i et gyteområde i en avgrenset fjord, kan man kvantifisere gyteforløpet fra start til slutt (Figur 25). Basert på genetikkanalyser (panI) sorteres eggene på kysttorsk og nordøst-arktisk torsk. Har man i tillegg lengde, alder og kjønn av gytetorsken, kan fekunditet og dermed biomassen av torsk som har gytt på dette gytefeltet beregnes (Figur 26). I gytesesongene 2012-2019 har Havforskningsinstituttet, i samarbeid med Runde Miljøsenter (RMS), gjort slike biologiske undersøkelser på gytefeltene til torsk (nordøst-arktisk torsk og kysttorsk) i Hessafjorden, Aspevågen, Borgundfjorden og Åsefjorden på Sunnmøre med det formål å regne ut samlet antall torskeegg gytt i løpet av gytesesongen (Figur 24). Lignende undersøkelser er tidligere gjort i Lofoten (Sundby og Bratland 1986). Ved innføring av områdevernet i Borgundfjorden i 2009 var det nærmest en forutsetning at et slikt vern av gyteområde måtte følges opp med prøvetaking for å følge med på om vernet fører til økt gyting i fjorden, og på sikt dermed et større ressursgrunnlag å høste fra. Samlet mengde torskeegg gytt i fjorden i løpet av gytesesongen bør over tid gi oss en god indikasjon på om områdevernet virker som forventet, eller det må settes inn ytterligere tiltak. En slik egg-metode vil være en mer miljøvennlig metode enn å drive prøvefiske i det stengte området for å studere utvikling av fangstrater.
Figur 24. Fiskeegg frå Borgundfjorden i ulike utviklingsstadier (venstre). Skipper Arne Sævik med egghåven som blir benyttet (til høyre).
Figur 25. Gyteforløp til torsk i Borgundfjorden 2012-2016 per underområde kvantifisert ved hjelp av standard egghåving og stadiebestemmelse av torskeegg (Johansen et al. 2017).
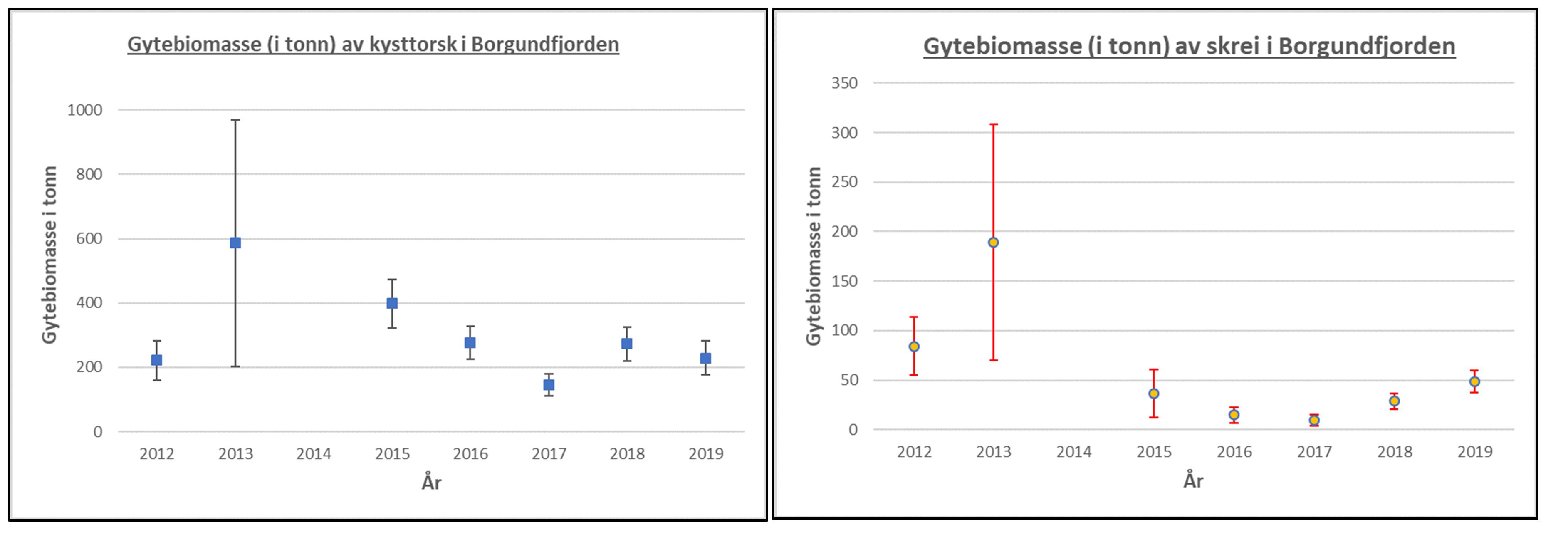
Figur 26. Foreløpige beregninger av gytebiomasse av kysttorsk og nordøst-arktisk torsk (skrei) i hele Borgundfjorden 2012-2019, basert på innsamlete egg-data, fekunditets-målinger (antall egg per gytefisk i området) og genetikk av eggene. (Masteroppgave William Aure Aannø, in prep.)
3.8 - Beskatning ved fritids -og turistfiske
På norskekysten har fritidsfiske etter torsk eksistert lenge. I de siste tiårene har det i tillegg vært betydelig turistfiske. I ICES sine bestandsvurderinger av kysttorsk har høyst usikre tall for årlig fritids -og turistfiske (ICES, 2010) vært addert til de rapporterte fangstene i yrkesfisket. Disse anslag for fritids -og turistfiske er i hovedsak basert på en undersøkelse fra 2003 (Hallenstvedt og Wulff, 2004), med små årlige skaleringer basert på Statistisk Sentralbyrå sine undersøkelser av fritidsbruk i den norske befolkning (hvert femte år).
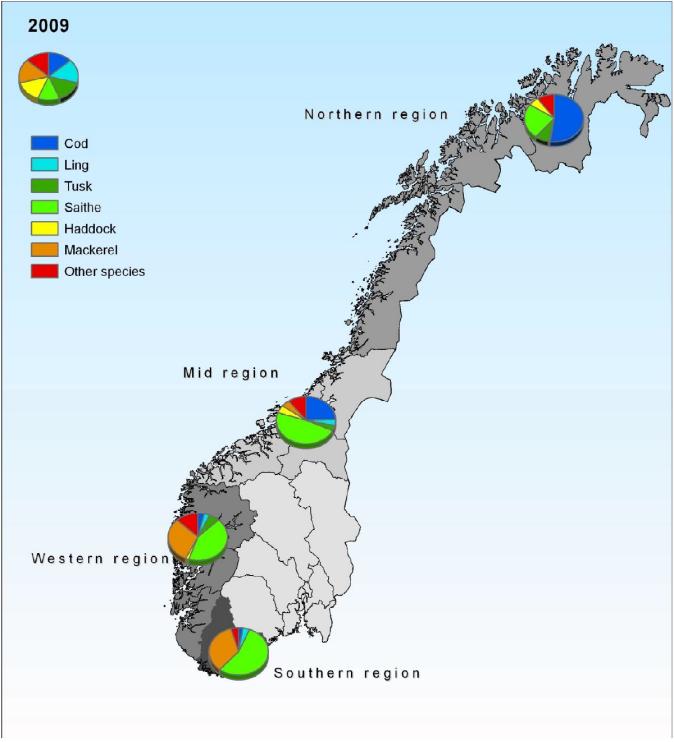
I 2008 og 2009 ble det gjennomført en undersøkelse av turistfisket knyttet til registrerte turistfiskeanlegg (Vølstad et al. 2009). Resultatene tilsier at antalls-andelen torsk i dette fisket er drøyt 50 % i Nord-Norge, ca 25% i Møre-Trøndelag, og mindre enn 10% i området fra Stad til Kristiansand (Figur 27). I områdene sør for Nordlandsgrensa var det sei det ble fanget mest av (om lag 50% av totalantall fisk). For de bedriftene som deltok i undersøkelsen nord for 62-graden i 2009 var total beregnet torskefangst (rundvekt) om lag 1600 tonn (konfidensintervall 903-2269).
Noen nye metodeprosjekter er satt i gang for å framskaffe bedre data som også inkluderer fritidsfisket. Dette er beskrevet i Vølstad et al. (2019), og inkluderer Troms, Hordaland og Oslofjorden. Prosjektene avsluttes i 2020.
Havforskningsinstituttet vil i løpet av 2020 i samarbeid med Fiskeridirektoratet oppsummere erfaringer med denne rapporteringen.
Figur 27. Antallsprosent fordeling på fiskearter i turistfisket i regionene Northern, Mid, Western og Southern (fra Vølstad et al. 2009).
3.9 - Bestandsstruktur basert på genetiske undersøkelser av kysttorsk nord for 62 ° N (Stad-Russegrensen)
Ved Havforskningsinstituttet har det over tid vært gjort et stort genetisk arbeid på torsk langs Norskekysten og i Barentshavet. Siden 2003 har det blitt gjennomført mer rettede studier av torsk og skrei langs Norskekysten, med hovedfokus på kysttorsk (NCC) nord og kysttorsk sør for 62°N og dens forhold til skrei (NEAC).
Det mest omfattende arbeidet med å definere kysttorskens populasjonsstruktur til dags dato er utført av Dahle et al. (2018). Dette arbeidet er basert på kysttorsk langs kysten fra Magerøysund i nord til Hvaler i sørøst, innsamlet under gytesesongene 2002-2007 (CODBIOBANK materialet). All fisk som kunne være skrei basert på otolitt-klassifisering ble fjernet fra analysen slik at vi satt igjen med >4000 kysttorsk fra totalt 55 ulike gyteområder. Kysttorsken ble genotypet med mikrosatellitter og genet Pan I som tidligere har vist seg å skille mellom kysttorsk og skrei (Westgaard & Fevolden 2007).
Resultatene viser i korte trekk at genetiske forskjeller mellom kysttorskprøvene øker med geografiske avstand mellom gyteområdene: jo større avstand fra hverandre, desto større genetisk differensiering uten at vi kan sette noen tydelig oppdeling i distinkte grupper. Videre viser dataene at denne differensiering langs kysten i hovedsak er drevet av kontakten med den store bestanden av skrei fra Barentshavet. Den nordligste kysten er mer påvirket enn kysttorsken lenger sør, og antall fisk som måtte fjernes basert på otolitt-typing viser noe av det samme mønsteret; med flere mulige skrei i de nordlige prøvene enn lenger sør. Den biologisk mest sannsynlige tolkningen av disse observasjonene er at kysttorsken består av et -kanskje stort- antall lokale gytebestander eller populasjoner som i bare begrenset omfang blandes med hverandre og med skrei. Dersom det ikke hadde vært noen begrensninger i utveksling mellom kysttorsk populasjonene ville vi forventet at resultatet av kontakten med skrei i nord ville spredd seg raskt nedover langs kysten og vi ville da ikke hatt den genetiske gradienten vi observerer.
3.9.1 - Pågående arbeider
Som en oppfølging av resultatene fra mikrosatellitter og Pan I summert ovenfor har Havforskningsinstituttet gjennomført en -fortsatt pågående- studie (SPACEFISH: 2016-2019) som benytter en annen type genetiske markører (kalt «snipper», SNP) enn tidligere og som har muliggjort mer detaljert klassifisering av kysttorsken på individnivå. Det har vist seg at skrei skiller seg fra kysttorsken i nord primært i 4 ulike områder i genomet, i kromosomene 1, 2, 7 og 12, og som inneholder bl.a. Pan I genet (Johansen et al. 2017). I en foreløpig analyse av ca 2000 kysttorsk fra 17 gyteområder nord for 60 ° N har vi valgt ut SNP gen-markører som dekker disse 4 kromosomene samt andre områder i torskegenomet (Johansen et al. 2017). Resultatene av disse bekrefter og utvider funnene fra Dahle et al. (2018) at kysttorsken ikke er en enkel, genetisk ensartet populasjon. Isteden observerer vi en genetisk strukturering som kjennetegnes ved: a) Økende genetiske forskjeller i geografisk avstand, mer eller mindre kontinuerlig uten noen åpenbar gruppering langs kysten. b) Genetiske forskjeller mellom år i enkelte områder og som kan antyde en vandrende bestandskomponent. c) Ulike deler av genomet viser forskjellig grad av genetisk divergens: grovt sett ligger forskjellene hos NCC i nettopp de kromosomene som skiller skrei og kysttorsk. Dette bekrefter konklusjonen at noe inn-kryssing av skrei i kysttorsken i nord har funnet sted og forklarer den observerte nord-sør gradienten. En nærmere beskrivelse og oppsummering av analysene er gjort i Vedlegg 1.
Havforskningens Strategiske Instituttsatsing ble startet opp for å studere genetisk struktur hos kysttorsk ved bruk av avanserte genetiske og genomiske metoder. Det overordna målet er å kartlegge genetisk struktur for å kunne bidra til en mer presis forvaltning av kysttorsk.
I gytesesongen våren 2019, ble det samlet inn prøver fra ca 3500 torsk, fra Varangerhalvøya i nord til Arnavågen i sør. Prøvene i denne perioden ble samlet inn ved en kombinasjon av eget prøvetakingsfartøy og personlig kontakt med fiskere og fiskemottak. En utfordring er at dette er også den perioden skreien kommer inn til kysten for å gyte, og spesielt i nord forventer vi en større innblanding av skrei. Det er tatt prøver til DNA analyser, otolitter (for å kunne bestemme alder og skille ut skreien) og lengde fra all torsken. I tillegg har det blitt samlet inn andre biologiske data hvor det har vært mulig (kjønn og vekt).
Prøvene fra vårfisket blir supplert med prøver tatt høsten 2019, siden det er på høsten vi estimerer mengde kysttorsk til rådgivningen vår. Vi forventer ut fra kunnskap om kysttorskens vandring at den torsken som står i fjordene og langs kysten på dette tidspunkt er noe stedegen og noe vandrende kysttorsk. Havforskningsinstituttet gjennomfører et eget tokt i det aktuelle området fra Varangerfjorden til Stad. Denne prøvetakingen vil supplere innsamlingen fra våren 2019. I tillegg vil vi ta prøver av torskehoder fra lokale fiskere og fiskemottak langs kysten og inn i fjordene også sør for Stad (62°N).
I vårt forrige prosjekt CODBIOBANK satset vi på å få en oversikt over nord-sør utbredelsen av kysttorsk i gytesesongen. I vår nye satsing vil vi gjenta denne prøvetakingen, men i tillegg få en bedre prøvetaking i «transekt» fra innerst i fjorden til ut på kysten, Dette fordi vi tror en del torsk ser ut til å vandre mellom fjord og kyst (Michaelsen et al. 2014) og i noen områder også gyte i fjordene (Johansen med flere 2017). Det vil derfor bli tatt prøver fra Varangerfjorden i nord til Bokn i sør, hvorav mange av prøvene utgjør «transekt» fra innerst i en fjord og helt ut til kyststrømmen. Vi har valgt ut noen fjorder, de fleste i Troms og Finnmark, for å studere eventuell genetisk struktur innen fjordene. Da er det også avgjørende å få samlet inn prøver fra ute på bankene for om mulig å få data på det mange anser er en «vandrende» kysttorsk som kan gyte inne i fjordene. De genetiske analysene vil starte opp våren 2020.
Planen er å gjennomføre ulike typer DNA analyser (full-genom og SNP) som vil gi oss mer detaljert informasjon om både den overordnede genetiske strukturen langs kysten, i tillegg til detaljert informasjon om eventuell struktur i noen av de viktigste fjordene våre – fra innerst i fjorden til ytterst på kysten.
3.10 - Merkeforsøk
Mange merkeforsøk er gjort på gytefelt i gytetiden. I disse forsøkene kommer de aller fleste gjenfangster fra samme gytefelt i samme og etterfølgende gytesesonger. Mange av utsettingene i Lofoten (og litt på Møre) inneholder mye skrei som typisk blir gjenfanget i Barentshavet.
Merkeforsøk gjennomført sør for Troms utenom gytetiden (utsatt om høsten) inneholder minimalt med skrei. Disse merkingene viser ganske små vandringer. (Se Vedlegg 2 «oversikt eldre merkeforsøk»).
Merkeforsøk utført i Finnmark (Altafjord, Revsbotn, Porsangerfjord, Laksefjord, Tanafjord og Varangerfjord) i 1980-1983 (Jakobsen 1987) viser at de fleste kysttorsk blir gjenfanget i samme fjord som de er merket, men at opptil 10% kan gjenfanges i nabofjorden.
4 - Forslag til ny gjenoppbyggingsplan for kysttorsk nord
4.1 - Gjenoppbyggingsmål for kysttorsk
Beregnet årlig fangst av kysttorsk (perioden 1984-2018) har en topp i 1984-1985 (64 og 63 tusen tonn), i 1995 (65 tusen tonn), og i 2016-2017 (55 og 51 tusen tonn). I eldre landingsstatistikk (1950-1960) for «fjordtorsk» (som ikke inkluderer all kysttorsk) er det en topp i 1959 på 35 tusen tonn. En noe usikker bestandsberegning for kysttorsk i perioden 1984-2018 (AFWG 2019) viser en topp i beregnet gytebestand i 1994, etterfulgt av betydelig reduksjon fram til 1998 og deretter en svakt økende tendens til noe i nærheten av langtidsgjennomsnittet (for hele perioden 1984-2018). Den gjeldende gjenoppbyggingsplanen referer til akustiske målinger i de fire første årene i Kystressurstoktet (1995-1998). Disse er de høyeste i hele tidsserien, og i ettertid framstår dette målet lite realistisk. Ingen senere målinger har vært på samme nivå. De akustiske målingene i kystressurstoktet viser en mer negativ utvikling enn det foreløpige bunntrål-indekser fra samme tokt viser. Det er uklart om det eksisterende målet som går på å gjenoppbygge kysttorskbestanden til nivået midt på 90-tallet er realistisk i dagens situasjon. Det akustiske toktet ble også gjennomført på en annerledes måte de tre første årene enn i den påfølgende perioden (Erik Berg pers. med.). Vi foreslår her noen grunnleggende endringer i bestandsrådgivningen og forvaltningen av kysttorsken.
4.2 - Oppdeling i flere forvaltningsområder
Både genetiske analyser (se Vedlegg 1) og merkeforsøkene (se Vedlegg 2) tilsier at det kan være hensiktsmessig å ha mer enn ett forvaltningsområde for kysttorsk i kystområdet fra Varanger til Stad. Kysttorsken kan best forstås som en såkalt metapopulasjon som består av forskjellige gytekomponenter langs kysten. Det generelle bildet er at gytekomponenter er mer beslektet med geografisk nære komponenter enn fjernere gytekomponenter.
Det er gjennomført en rekke genetiske analyser av torsk langs norskekysten i tillegg til merkeforsøk for å se på vandring av de ulike komponentene av torsk (NEAC og NCC). Dataene gir få klare grenser for genetisk differensiering, selv om de viser at det eksisterer en differensiering mellom populasjoner langs norskekysten (Dahle et al 2018). Det synes imidlertid å være en barriere ved Stadlandet (62 graden). I tillegg gir dataene antydninger om en grense ved Vesterålen. De populasjonene som er i området mellom Vestfjorden og Bodø, vil sannsynligvis ikke komme under noe større press. Av hensyn til geografisk dekning i toktene synes det mer hensiktsmessig å legge grensen ved 67 graden.
For kysttorsk nord for 67 graden vurderer vi at datagrunnlaget er godt nok til at vi kan utvikle en analytisk bestandsvurdering på tilsvarende måte som for nordøstarktisk torsk. Dette området gir rundt 80% av fangstene av kysttorsk. Som et element i en slik bestandsvurdering vil der være biologiske referansepunkter som angir grenseverdier for bestandsstørrelse og høstingsnivå. Det vil også bli utviklet en høstingsregel som vil virke slik at dersom denne følges så vil bestanden bevege seg mot bærekraftig bestandsnivå og høstingsnivå. Gjenoppbygningsplanen vil dermed bli å følge høstingsregelen.
Kystressurstoktet ser ut til å ha rimelig god dekning av kysttorsk sør til Vestfjorden (67-graden), men i kystområdene lengre sør er det betydelige grunne områder som ikke kan dekkes med store forskningsfartøy. (Denne problemstillingen er mindre relevant for seien, fordi de viktige aldersgruppene av sei finnes i stor grad på kystbankene, som dekkes godt av Kystressurstoktet). I områdene sør for Vestfjorden kan det være relevant å ha en rådgivning for kysttorsk basert på en datafattig tilnærming (ICES datafattig kategori 3- bestand), som kan gjøre nytte av garn-ruse toktene. Disse går annethvert år i området Steigen-Vikna, og annethvert år i området Vikna-Stad. For den sørlige kysttorsken vil vi også bruke fangstdata til å vurdere bestandsutviklingen. Vi vil også som en del av prosessen med å analysere disse toktdataene og fangstdataene og komme med forslag til gjenoppbyggingsplan for den sørlige bestanden.
Havforskningsinstituttet vil derfor faglig sett anbefale at en ny gjenoppbyggingsplan for kysttorsk nord for 62 ˚ N baseres på følgende:
Det etableres et skille for håndtering av kysttorsk ved 67-graden.
Nord for 67-graden (fjordene inkludert): her er det grunnlag for å utvikle en analytisk bestandsvurdering (basert på forvaltningsrelaterte referansepunkter. Det er behov for mer overvåkning av eggproduksjon og kartlegging av yngel/ungfisk i fjordene. Vern av gyte og oppvekstområder er relevant.
Sør for 67-graden: her anbefaler vi å basere rådgiving på overvåking av yngel/ungfisk med garn og ruser og overvåkning av viktige gyteområder. Fiskeribaserte innsatsdata og fangst per enhet innsats er nyttig grunnlag for å regulere innsatsen i fiskeriene.
Gjenoppbyggingsmålene blir satt for hvert av områdene basert på utledede biologiske referansepunkt.
4.3 - Lokal/regional forvaltning
Resultater fra eldre merkeforsøk (se vedlegg 2) tyder på begrenset vandring og lokal tilhørighet for en stor andel av merkede individer. I spesielle områder hvor det foreligger gode data og lokal kunnskap, kan det vurderes om det er grunnlag for å ha et mer lokalt fokusert forvaltningsregime etter mønster av «Aktiv forvaltning» (Espeland et al. 2014) og «Tvedestrandsprosjektet» (Espeland et al. 2016), eller som skissert i Strand (2019 https://www.hi.no/hi/nettrapporter/ porsangerfjorden-2.0-final.docx) og Langeland et al. (2010). Aktuelle områder kan for eksempel vurderes valgt ut på bakgrunn av verdisetting av gytefelt.
4.4 - Nytt ICES Benchmark assessment
En ny gjenoppbyggingsplan vil i stor grad bli basert på analyser gjennomført i ICES (AFWG), blant annet for å definere «sikre biologiske grenser» og andre biologiske referansepunkter. Noe av dette kan forberedes til det ordinære AFWG-møtet 16.-22. april 2020. Det formelle «ICES-Benchmark» starter opp med data-gjennomgang og eventuelle data-revisjoner, trolig i august 2020 og selve Benchmark-møtet blir trolig i februar 2021. Forbedret bestandsberegning og biologiske referansepunkter relevante for gjenoppbyggingsplanen, kan da være på plass til det ordinære AFWG-møtet i april 2021.
ICES vil gi råd i henhold til gjeldende plan inntil en ny plan er vedtatt og godkjent av ICES.
5 - Forslag til forvaltningstiltak
5.1 - Mulige tiltak for å fremme vekst i bestanden
Fangst av kysttorsk avregnes som nevnt mot en samlet kvote for kysttorsk og skrei. Begrensninger av fisket etter kysttorsk foretas derfor gjennom regler som styrer utøvelsen av fisket (områdebegrensninger, redskapsregulering, periodiseringer o.l.). Mulige forvaltningstiltak for å få gjenoppbyggingsplanene implementert vil være å innskrenke adgangen til å fiske innenfor fjordlinene til kun å gjelde fartøy <11 meter. Et annet tiltak er å etablere reell tids-stengning av fiskeområder der det fiskes mye kysttorsk. «Henningsværboksen» i Lofoten ble stengt for alt fiske unntatt fiske med håndsnøre og fiskestang i gytesesongen i 2005 på grunn av høy andel kysttorsk i fangstene. Boksen kunne åpnes hvis prøvetaking fra fisket like utenfor boksen viste mer enn 70% skrei. I årene 2005-2010 ga ingen prøver grunnlag for åpning, men fra og med 2011 har skreiandelen vært høy, og boksen har vært åpnet for fiske. En tilsvarende boks ble i 2009 etablert i Borgundfjorden ved Ålesund. Her har prøvene ikke gitt grunnlag for åpning. Prøvetakingen har vært betydelig redusert etter 2016. Slik operasjonell genetisk overvåking av kysttorskfiskeriene innebærer å daglig estimere andelen av kysttorsk som fiskes i viktige gytefelt. Vi foreslår å etablere operasjonell genetisk overvåking i flere sentrale kysttorskområder for den nordlige bestanden og etablere en reell tids-lukking av felter der det påføres høy fiskedødelighet på kysttorsk (funksjon av antall fisk fisket og andel kysttorsk av disse). HIs sitt fangstsamplingsfartøy (Falkungen) og Fiskeridirektoratets sjøtjeneste er aktuelle innsamlingsplattformer for en slik operasjonell genetisk overvåking.
Genetiske studier viser en gradvis endring i kysttorsken fra nord til sør. Det tilsier at oppdeling i flere forvaltningsenheter kan gi grunnlag for bedre vern av eventuelle «svakere komponenter», men samtidig vil oppdelinger øke kravet til omfang både av tokt-dekning og prøver fra fiskeriene.
Garn-ruse-toktene som ble startet opp i 2013 veksler annethvert år mellom området Steigen-Vikna og Vikna-Stad. Fangstene tyder på at lokale bestander på grunt vann i begge områder domineres av unge individer (1, 2 og 3 år gamle fisk, se tabell 3 og 5), som selvsagt kan være fordi de eldre individene på denne tiden av året lever dypere, men kan også være er en indikasjon på høy dødelighet. For hvert av disse områdene kan det være relevant å ha en «datafattig» rådgiving (ICES category3-stocks), basert på garn-ruse toktene, landingsdata og fangst-per-enhet-innsats i Referanseflåten. Dette vil kreve en utvidet prøvetaking fra fiskeriene, samt en tidsserie med relevante data for fiskeinnsats.
Både fiskeriene og toktene viser at det er mest kysttorsk nord for 67-graden (ca 75%), og at forekomstene avtar gradvis fra 67-graden til 62-graden. Det er grunn til å tro at i områder hvor tettheten av torsk er lav, kan bestandskomponentene være ekstra sårbare, slik at behovet for vern og gjenoppbygging er ekstra stort. Klimaendringene vil på kort sikt også kunne ha potensielt større effekt i sør enn i nord.
I NFDs bestilling anmodes Havforskningsinstituttet om å fremme forslag til tiltak som kan bidra til vekst i bestanden. Generelt synes det ut ifra den målte bestandssituasjonen å være behov for strengere tiltak sør for 67-graden enn nord for 67 -graden .
Havforskningsinstituttet anbefaler at følgende forslag bør vurderes i kommersielt fiskeri:
Innføring av bifangstregler, gjerne nyansert på område, tid, fartøystørrelse og redskap, bl.a. i forbindelse med tiltakene nedenfor.
Redusere kystfisket om høsten (i konflikt med dagens «ferskfiskordninger»).
Stenge områder/måneder der det vanligvis fanges mye kysttorsk (se Vedlegg 3; deriblant overvintringsområder for NVG-sild i november-januar).
I gytesesongen; -stenge utvalgte gytefelt som er dominert av kysttorsk.
Vurdere tiltak for bevaring av bred alders- og størrelsessammensetning
Vurdere revisjoner av fjordlinjer.
Strengere begrensninger innenfor fjordlinjer (redusere maks fartøylengde til 11 m, og begrensninger på bruksmengde).
Sør for 67-graden er innblandingen av skrei utenom gytetiden minimal. Her kan innsatsen i fisket reguleres for resten av året i forhold til fangst-per-enhet innsats (CPUE), og det som Havforskningsinstituttet vurderer som et bærekraftig uttak.
Overvåke og måle effekter av reguleringer.
Videreutvikle genetiske metoder og verktøy for å kunne operere på «kaikanten» og raskt og effektivt fange opp dynamiske endringer i fangst av kysttorsk.
Eventuelt fremme «lokale -regionale reguleringer» i områder hvor det er et svært godt kunnskapsgrunnlag (se Langeland et al. 2010). Jfr «Aktiv forvaltning» (Espeland et al 2014) og det som er skissert i (Strand, 2019): https://www.hi.no/hi/nettrapporter/porsangerfjorden-2.0-final.docx .
Havforskningsinstituttet anbefaler at følgende forslag bør vurderes i turist -og fritidsfisket:
Sørge for håndheving av registreringsplikt og rapporteringsplikt, og lage en plan for overvåking av disse fiskeriene.
Redusere tillatt bruksmengde.
Fang og slipp: formidle betydningen av beste praksis.
Maksmål kan vurderes som reguleringstiltak i tillegg til minstemål, særlig på Sør- og Vestlandskysten. Lenger nord blir dette vanskeligere på grunn av innblanding av nordøstarktisk torsk.
6 - TAKK
Takk til alle som har bidratt til denne rapporten! En særskilt takk til:
Erik Berg, Geir Dahle, Ove Djupevåg, Sigurd Heiberg Espeland, Karen Gjertsen, Kevin Glover, Sofie Gundersen, Jeff Hutchings (Canada), Torild Johansen, Per Erik Jorde, Halvor Knutsen, Trond Ottemo, Terje Svåsand og Jon Ivar Westgaard. Også en stor takk til alle feltarbeidere og alderslesere.
Berg, E., Eriksen, I. A. and Eliassen, J.-E. 1998. Catch-statistics for Norwegian Coastal cod 1984-1997, -data and methods. Fiskeriforskning, 10/1998.
Berg, E., and Pedersen, T. 2001. Variability in recruitment, growth and sexual maturity of coastal cod ( Gadus morhua L.) in a fjord system in northern Norway. Fisheries Research, 52: 179–189.
Berg, E., and Albert, O. T. 2003. Cod in fjords and coastal waters of North Norway: distribution and variation in length and maturity at age. – ICES Journal of Marine Science, 60: 787–797
Dahle, G. 1991. Gadus morhua L., populations identified by mitochondrial DNA. Journal of Fish Biology (1991) 38: 295–303.
Dahle G, M Quintela, T Johansen, J-I Westgaard, F Besnier, A Aglen, KE Jørstad, KA Glover 2018. Analysis of coastal cod ( Gadus morhua L.) sampled on spawning sites reveals a genetic gradient throughout Norway's coastline. BMC Genetics 19: 42.
Espeland, S. H., Kleiven A.R., Olsen E.M. and J.A. Knutsen, 2014. Aktiv forvaltning av marine ressurser – lokalt tilpasset forvaltning. Rapporter fra Havforskningen nr 34-2014.
Espeland S. H., Kleiven A.R., Moland, E., Kleiven, P. J. N., Halvorsen, K., Bodvin, T., Olsen, E.M., Knutsen, J. A. 2016. Aktiv forvaltning av marine ressurser – lokalt tilpasset forvaltning, Tvedestrand. Rapporter fra Havforskningen nr 40-2016.
Godø, O.R. 1984. Migration, mingling and homing of Northeast Arctic cod from two separated spawning grounds. In the proceedings of the Soviet-Norwegian symposium on Reproduction and Recruitment of Arctic Cod, Leningrad 26–30 September 1983, edited by O.R. Godø, S. Tilseth. Institute of Marine Research, Bergen, Norway, 1984.
Hallenstvedt, A. and Wulff, I., 2004. Recreational fishery in the sea 2003. (In Norwegian). Norwegian College for Fisheries/University of Tromsø, 2004.
Hirst, D., Storvik, G., Rognebakke, H., Aldrin, M., Aanes, S., and Vølstad, J. H. 2012. A Bayesian modelling framework for the estimation of catch-at-age of commercially harvested fish species. Can. J. Fish. Aquat. Sci. 69: 2064–2076.
Hylen, A., Nakken, O. and Nedreaas, K. 2008. Northeast Arctic cod: fisheries, life history, stock fluctuations and management. Pp 83-118 (chapter 5) In Nakken, O. (editor). Norwegian spring-spawning herring and northeast Arctic cod. 100 years of research and management. Tapir Academic Press, Trondheim. ISBN:978-82-519-2367-5. 177 pp.
ICES 1970. Report of the meeting of the North-East Arctic Fisheries Working Group Annex_1. International Council for the Exploration of the Sea, Copenhagen, 14th - 16th January 1970. 16 pp.
ICES 1990. Report of the Arctic Fisheries Working Group. International Council for the Exploration of the Sea, Copenhagen, 18-27 September 1990. CM 1991/Assess:3, 127 pp.
ICES 2010. Report of the Arctic Fisheries Working Group, Lisbon/Bergen, 22-28 April 2010. ICES C.M. 2010/ACOM:05, 664 pp.
ICES 2019a. Cod ( Gadus morhua ) in subareas 1 and 2 (Norwegian coastal waters cod). In Report of the ICES Advisory Committee, 2019. ICES Advice 2019, cod.27.1-2coast, https://doi.org/10.17895/ices.advice.4711
ICES 2019b. Arctic Fisheries Working Group (AFWG). ICES Scientific Reports.1:30. 930 pp. http:/doi.org/10.17895/ices.pub.5292
Jakobsen, T. 1987. Coastal Cod in Northern Norway. Fisheries Research 5: 223-234.
Johansen, T, Westgaard J-I, Seliussen BB, Nedreaas K, Dahle G, Glover KA, Kvalsund R, Aglen A, 2017. “Real-time” genetic monitoring of a commercial fishery on the doorstep of an MPA reveals unique insights into the interaction between coastal and migratory forms of the Atlantic cod. – ICES Journal of Marine Science, doi:10.1093/icesjms/fsx224.
Langeland, T., Olsen, E.M., Knutsen, H., Kleiven, A.R., Moland, E., Knutsen, J.A. (2010) Nye verktøy i forvaltningen av kystressursene. Pp. 62-66. I: Gjøsæter, H., Haug, T., Hauge, M., Karlsen, Ø., Knutsen, J.A., Røttingen, I., Skilbrei, O., Sunnset, B.H. (red.) 2010. Havforskningsrapporten 2010. Fisken og havet, særnr. 1-2010. Havforskningsinstituttet, Bergen.
Mankevich, E.M. 1969. Structure of the stock of Arcto-Norwegian cod in 1969 according to age samples obtained off the north western coast of Norway. Annales Biologiques, Vol. 26: 123–125.
Michalsen K, Johansen T, Subby S, Beck A, 2014. Linking tagging technology and molecular genetics to gain insight in the spatial dynamics of two stocks of cod in Northeast Atlantic waters. ICES Journal of Marine Science; doi:10.1093/icesjms/fsu083
Mork, J., Giskeødegård, R. and Sundnes, G. 1984. Population genetics studies in cod ( Gadus morhua L.) by means of the haemoglobin polymorphism; observations in a Norwegian coastal population. Fiskeridirektoratets Skrifter Serie Havundersøkelser 17, No. 12: 449-471.
Mork, J., Ryman, N., Ståhl, G., Utter, F., Sundnes, G. 1985. Genetic variation in Atlantic cod ( Gadus morhua L.): little divergence throughout the species range. Canadian Journal of Fisheries and Aquatic Sciences, 42: 1580–1587.
Møller, D. 1968. Genetic diversity in spawning cod along the Norwegian coast. Hereditas, 60: 1–32.
Møller, D. 1969. The relationship between Arctic and coastal cod in their immature stages illus-trated by frequencies of genetic charaters. Fiskeridirektoratets Skrifter Serie Havundersøkelser 15: 220-233.
Opdal, A.F. 2009. Fisheries change spawning ground distribution of Northeast Arctic cod. Biol. Lett (2010) 6, 261-264 doi:10.1098/rsbl.2009.0789
Rollefsen, G. 1933. The otoliths of the cod. Preliminary report. Fiskeridirektoratets Skrifter, Serie Havundersøkelser (Report of the Norwegian Fisheries and Marine Investigations), 4 (3): 3–14.
Rollefsen, G. 1935. The spawning zone in cod otoliths and prognosis of stock. Fiskeridirektoratets Skrifter, Serie Havundersøkelser (Report of the Norwegian Fisheries and Marine Investigations), 4: 3–10.
Sars, G.O. 1879. Inberetninger til Departementet for det Indre fra professor G.O. Sars om de af ham anstillede undersøgelser angaaende saltvandsfiskeriene. Berg og Ellefsens Bogtrykkeri, Christiana: 221 pp. (In Norwegian).
Stransky, C; Baumann, H; Fevolden, S-E; Harbitz, A; Høie, H; Nedreaas, KH; Salberg, A-B; Skarstein, TH. 2008. Separation of Norwegian coastal cod and Northeast Arctic cod by outer otolith shape analysis. Fisheries Research 90, 26-35.
Sundby, S. og Bratland, P. 1986. Kartlegging av gytefeltene for norskarktisk torsk i Nord-Norge og beregning av eggproduksjonen i årene 1983- 1985. fSpatial distribution and production of eggs from Northeast-arctic cad at the coast of Northern Norway 1983-1985]. Fisken Hav., 1987(1): 1-58.
Sundby, S, P. Fossum, A. Sandvik, F.B. Vikebø, A. Aglen, L. Buhl-Mortensen, A. Folkvord, K. Bakkeplass, P. Buhl-Mortensen, M. Johannessen, M.S. Jørgensen, T. Kristiansen, C. Landa, M.S. Myksvoll and R. Nash. 2013. KunnskapsInnhenting Barentshavet-Lofoten-Vesterålen (KILO). Fisken og havet nr. 3-2013, 186 pp (in Norwegian).
Trout, G.C. 1955. English Cod Tagging in 1954 and 1955. Annales Biologiques, 12: 138–140.
van der Meeren, T. 2018. Undersøkelser av gyte- og oppvekstområder for torsk i Smøla og Aure kommuner våren og høsten 2017. Rapport fra Havforskningen, Nr 6-2018
Vølstad, J.H., Korsbrekke, K.,Nedreaas, K., Nilsen, M., Nilsson, G.N., Pennington, M., Subbey, S. and Wienerroither. 2009. A probability based survey using self sampling to estimate catch and effort in Norway’s coastal tourist fishery. Coastal fish resources: the foundation for tourist fishing and related commerce (NFR project no. 173274/S40).
Vølstad, J. H., Christman, M., Ferter, K., Kleiven, A. R., Ottera˚, H., Aas, Ø., Arlinghaus, R. Borch, T. Colman, J., Hartill, B. Haugen, T. O., Hyder, K., Lyle, J. M., Ohldieck, M. J. Skov, C., Strehlow, H. V., van Voorhees, D., Weltersbach, M. S., and Weber, E. D. 2019. Field surveying of marine recreational fisheries in Norway using a novel spatial sampling frame reveals striking under-coverage of alternative sampling frames. – ICES Journal of Marine Science, doi:10.1093/icesjms/fsz108.
Westgaard J-I & S-E Fevolden 2007. Atlantic cod ( Gadus morhua L.) in inner and outer coastal zones of northern Norway display divergent genetic signature at non-neutral loci. Fisheries Research 85: 306-315.
Vedlegg 1. Oppsummering av genetiske undersøkelser av kysttorsk nord for 60°N (SPACEFISH)
Torild Johansen, Per Erik Jorde, Geir Dahle, Jon Ivar Westgaard og Kevin Glover
Som en oppfølging av resultatene fra mikrosatelitter og Pan I publisert av Dahle et al. (2018) har Havforskingsinstituttet gjennomført en --fortsatt pågående-- studie (SPACEFISH: 2016-2019) som benytter en annen type genetiske markører (kalt «snipper», SNPs) enn tidligere og som har muliggjort mer detaljert klassifisering av kysttorsken på individnivå. Hele torskens arvemasse eller genom er nylig blitt kartlagt (Star et al. 2011, Tørresen et al. 2017) og består av ca 650 millioner nukleotider fordelt på 23 kromosomer. Den overveiende andelen av nukleotidene er identiske hos alle torsk, men forskjeller mellom individer og bestander finnes i ca 1% av dem. Denne variasjonen er grunnlaget for å bruke genetikk som et verktøy til å separere torsk i ulike populasjoner og til å identifisere individer til den populasjonen de hører til. Det har vist seg at skrei (Nordøstarktisk torsk, NEAC) skiller seg fra kysttorsken i nord primært i 4 ulike områder i genomet, i kromosomene 1, 2, 7 og 12 (Kirubakaran et al. 2016, Berg et al. 2016). Disse kromosom-områdene er karakterisert ved en type strukturelle forandringer som kalles inversjoner, der en stor bit av kromosomet (ofte på flere millioner nukleotider) finnes i både "forlengs-" og "baklengs-" varianter. Individer som bærer på kromosomer av begge typene, dvs. som er heterozygote, vil bare produsere kjønnsceller der den ene eller andre av kromosom-typene er intakt, uten den sedvanlige rekombinasjonen mellom genene innen kromosomet. Resultatet er at alle genene som ligger inne i slike inversjoner nedarves sammen, som et stort "supergen". Vi formoder at de biologiske egenskapene som skiller skrei og kysttorsk har en genetisk basis som ligger i disse kromosomene, men hvilke gener som er involvert er ikke kjent.
I prosjektet SPACEFISH har vi for praktiske analyser av kysttorskens populasjonsstruktur valgt ut et antall SNP genmarkører i genomet og undersøker hvordan variasjonen i disse fordeler seg i tid og rom i utvalgte kysttorsk populasjoner fra CODBIOBANK. I en foreløpig analyse av ca 2000 kysttorsk fra 17 gyteområder har vi valgt ut SNP genmarkører som dekker alle 4 kromosom-inversjonene samt andre områder/kromosomer i torskegenomet (Johansen et al. 2017). For å teste hypotesen om at kysttorsken utgjør en enkel biologisk populasjon testet vi prøvene som er tatt på ulike gytelokaliteter og tester om disse er genetisk like, dvs. om forskjellene i forekomstene av ulike genvariantene er større enn det som kan forventes ved tilfeldige avvik under prøvetaking. Resultatet er entydige og bekrefter funnene fra Dahle et al. (2018) at kysttorsken ikke er en enkel, genetisk ensartet populasjon. Isteden observerer vi en romlig genetisk strukturering som kjennetegnes ved:
Økende genetiske forskjeller i geografisk avstand, mer eller mindre kontinuerlig uten noen åpenbar gruppering langs kysten.
Ulik grad av genetisk divergens i deler av genomet som ligger i og utenfor de 4 kromosom-inversjonene: grovt sett ligger forskjellene i de samme regionene som skiller skrei og kysttorsk, og kysttorsken blir genetisk mer lik skrei i nordområdene.
Det er tre spørsmål som må besvares før vi kan gi en forvaltningsmessig relevant tolkning av disse observasjonene:
1. Er det genetiske forskjeller blant "ekte" kysttorsk vi observerer, eller er forskjellene diktert av innslag av skrei i prøvene?
En mulig tilstedeværelse av skrei i kysttorskmaterialet, noe som er vanlig i gytesesongen, kan være problematisk siden det kan gi et feilaktig inntrykk av genetiske forskjeller hos kysttorsk. Mistanken om tilstedeværelse av skrei i kysttorskprøvene er det forhold at ialt 56 individer fra 2000 totalt har samme otolittype som skrei, og 54 individer har en genotype (BB) i PanI markøren som er karakteristisk for skrei (jfr. Westgaard & Fevolden 2007). Vi finner imidlertid ingen eksakt overensstemmelse mellom disse to karakteristika for skrei: bare 18 individer har begge.
Denne svake korrespondansen mellom Pan I og otolittype gjør det vanskelig med sikkerhet å skille skrei fra kysttorsk på individnivå. Videre er det uklart hva den relativt store andelen heterozygote (AB) individer har for populasjonstilhørighet. Dersom de er kysttorsk så forventer vi også en hel del BB homozygoter som følge av Mendelsk segregering, og da kan ikke alle de observerte homozygotene være skrei. Dermed er det også en mulighet for at ingen av dem er skrei men isteden representer variasjon innen gruppen kysttorsk (Michalsen et al. 2014).
2. Hvor stabil er den observerte romlige genetiske strukturen over tid?
I for eksempel kysttorsk tatt i 2004 og 2006 fra Eidsfjorden i Vesterålen viser tildels store, statistisk signifikante genetiske endringer imellom år (grad av genetisk differensiering, F ST = 0.0200). En mulig forklaring på de tilsynelatende store temporale genetiske forskjellene er derfor et varierende innslag av skrei i prøvene. Imidlertid støtter ikke de genetiske dataene en så enkel forklaring: skrei karakteriseres av høy grad av homozygositet for PanI-B markøren (Westgaard og Fevolden 2007, Michalsen et al . 2014), men vi ser tildels store innslag også av heterozygoter i disse kystsamplene, og de aller fleste av de heterozygote individene har otolitter av kysttorsk-typen.
Dersom årsaken til temporale endringer ikke er innslag av skrei kan det være at kysttorsken har en mer komplisert populasjonsstruktur, kanskje inkludert både lokale og mer mobile bestandskomponenter (Michalsen et al. 2014). Det skal heller ikke underslås at vi har begrenset erfaring med temporal stabilitet av kromosom-inversjonene, som trolig er under seleksjon, og genmarkører i disse regionene kan tenkes å gi misvisende informasjon om kysttorskens populasjonsstruktur (men se Jorde et al. 2018).
3. Hvor store genetiske forskjeller er "akseptable" innenfor en forvaltningsenhet?
Ideelt sett bør forvaltningen baseres på biologiske populasjoner som enheter, for å unngå overbeskatning av mindre, numerisk svakere populasjoner. I marine miljøer er det oftest uklart hvor den ene populasjonen slutter og den andre begynner og det er sannsynlig at en viss utveksling mellom populasjonene finner sted. Utveksling av individer er en effektiv mekanisme som reduserer genetiske forskjeller mellom populasjonene (f.eks. Waples og Gaggiotto 2006). Dette betyr at der vi faktisk finner genetiske forskjeller, som vi gjør hos kysttorsk nord for 62°N, så er det sannsynligvis flere enn én populasjon til stede.
Et vedvarende problem for videre analyser og tolkninger er temporale genetiske forskjeller og/eller genetisk strukturering på meget liten (kilometer) geografisk skala. I videre arbeid med SPACEFISH skal vi koble våre genetiske resultater med modellering for å teste om temporale innslag av skrei i kysttorskmaterialet kan forklare noe av den genetiske strukturen vi ser i dataene.
Som en foreløpig oppsummering av dagens kunnskap om populasjonsstruktur hos kysttorsk kan vi si at SPACEFISH analysene så langt bekrefter tidligere funn av Dahle et al. (2018), og viser at nåværende forvaltningsenhet for kysttorsk nord for Stad (62°N) må inneholde flere enn én enkelt kysttorskbestand. Denne bør trolig deles opp i mindre geografiske forvaltningsenheter, men resultatene så langt gir ikke sikre holdepunkter for hvor mange enheter og hvor disse bør ligge.
Referanser
Berg PR, B Star, C Pampoulie, M Sodeland, JMI Barth, H Knutsen, KS Jakobsen, S Jentoft 2016. Three chromosomal rearrangements promote genomic divergence between migratory and stationary ecotypes of Atlantic cod. Scientific Reports 6: 23246
Dahle G, M Quintela, T Johansen, J-I Westgaard, F Besnier, A Aglen, KE Jørstad, KA Glover 2018a. Analysis of coastal cod ( Gadus morhua L.) sampled on spawning sites reveals a genetic gradient throughout Norway's coastline. BMC Genetics 19: 42
Johansen, T, Westgaard J-I, Seliussen BB, Nedreaas K, Dahle G, Glover KA, Kvalsund R, Aglen A, 2017. “Real-time” genetic monitoring of a commercial fishery on the doorstep of an MPA reveals unique insights into the interaction between coastal and migratory forms of the Atlantic cod. – ICES Journal of Marine Science, doi:10.1093/icesjms/fsx224.
Jorde PE, A-E Synnes, SH Espeland, M Sodeland, H Knutsen 2018. Can we rely on selected genetic markers for population identification? Evidence from coastal Atlantic cod. Ecology and Evolution 8: 12547-12558
Michalsen K, Johansen T, Subby S, Beck A, 2014. Linking tagging technology and molecular genetics to gain insight in the spatial dynamics of two stocks of cod in Northeast Atlantic waters . ICES Journal of Marine Science ; doi:10.1093/icesjms/fsu083
Star B og 41 andre 2011. The genome sequence of Atlantic cod reveals a unique immune system. Nature 477: 207-210
Tørresen OK og 15 andre 2017. An improved genome assembly uncovers prolific tandem repeats in Atlantic cod. BMC Genomics 18: 95
Waples RS, O Gaggiotti 2007. What is a population? An empirical evaluation of some genetic methods for identifying the number of gene pools and their degree of connectivity. Molecular Ecology 15: 1419-1439
Westgaard J-I & S-E Fevolden 2007. Atlantic cod ( Gadus morhua L.) in inner and outer coastal zones of northern Norway display divergent genetic signature at non- neutral loci. Fisheries Research 85: 306-315
Vedlegg 2: Oversikt over eldre merkeforsøk
Eldre merke-gjenfangstdata av torsk relevant for kysttorsk i nord
Terje Svåsand, Sofie Gundersen, Ove Djupevåg og Asgeir Aglen
Historiske merke-gjenfangstforsøk med torsk er systematisert i en database på Havforskningsinstituttet. Et utvalg av disse dataene (Tabell 1) er analysert som del av arbeidet med å beskrive bestandsstruktur hos kysttorsk i nord. Da det ikke er tilgang til informasjon (otolitter/genetikk) om dette er kysttorsk eller nordøstarktisk torsk, er kun utsetting av merket torsk i perioden juni-oktober benyttet. I denne perioden er det forventet at det kun er kysttorsk langs kysten. Merkeforsøk i de kystnære fiskeriområdene 00, 05, 06, 07 er analysert (Tabell 1).
Utsettingssted (Fiskeriområde)
Periode
# Satt ut
Midd.lengde(cm) ved utsett
#Gjenfanget
% gjenfanget
Dager i sjø (median)
00
1964-1983
1247
51,8
492
39,5
170
05
1964-1981
2898
60,2
1079
37,2
130
06
1979-1983
2082
47,4
220
10,6
198
07
1958-1983
3620
56,7
1185
32,7
166
Alle
1958-1983
9847
56,5
2976
30,0
152
Tabell 1. Oversikt over merke-gjenfangstforsøk (se tekst).
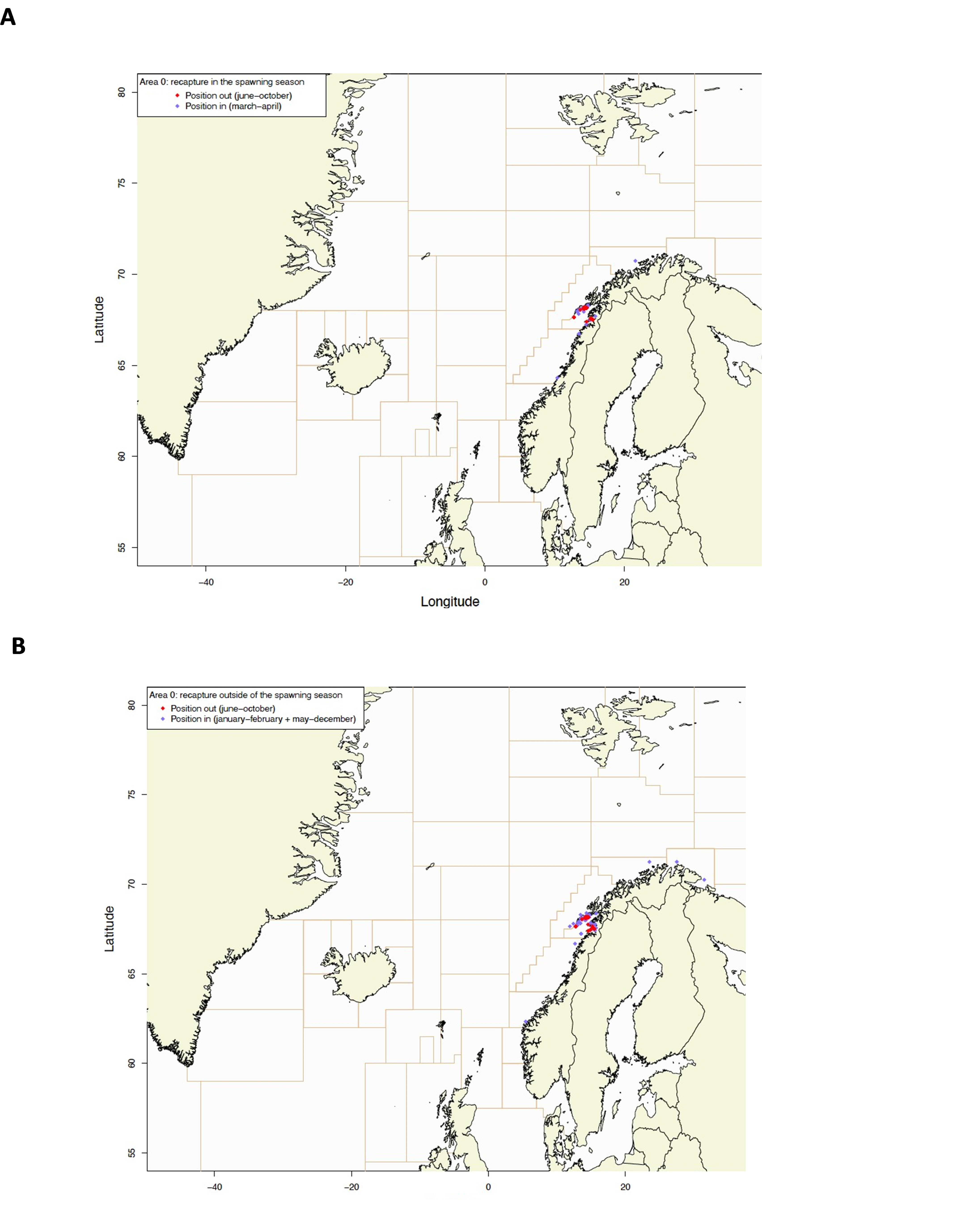
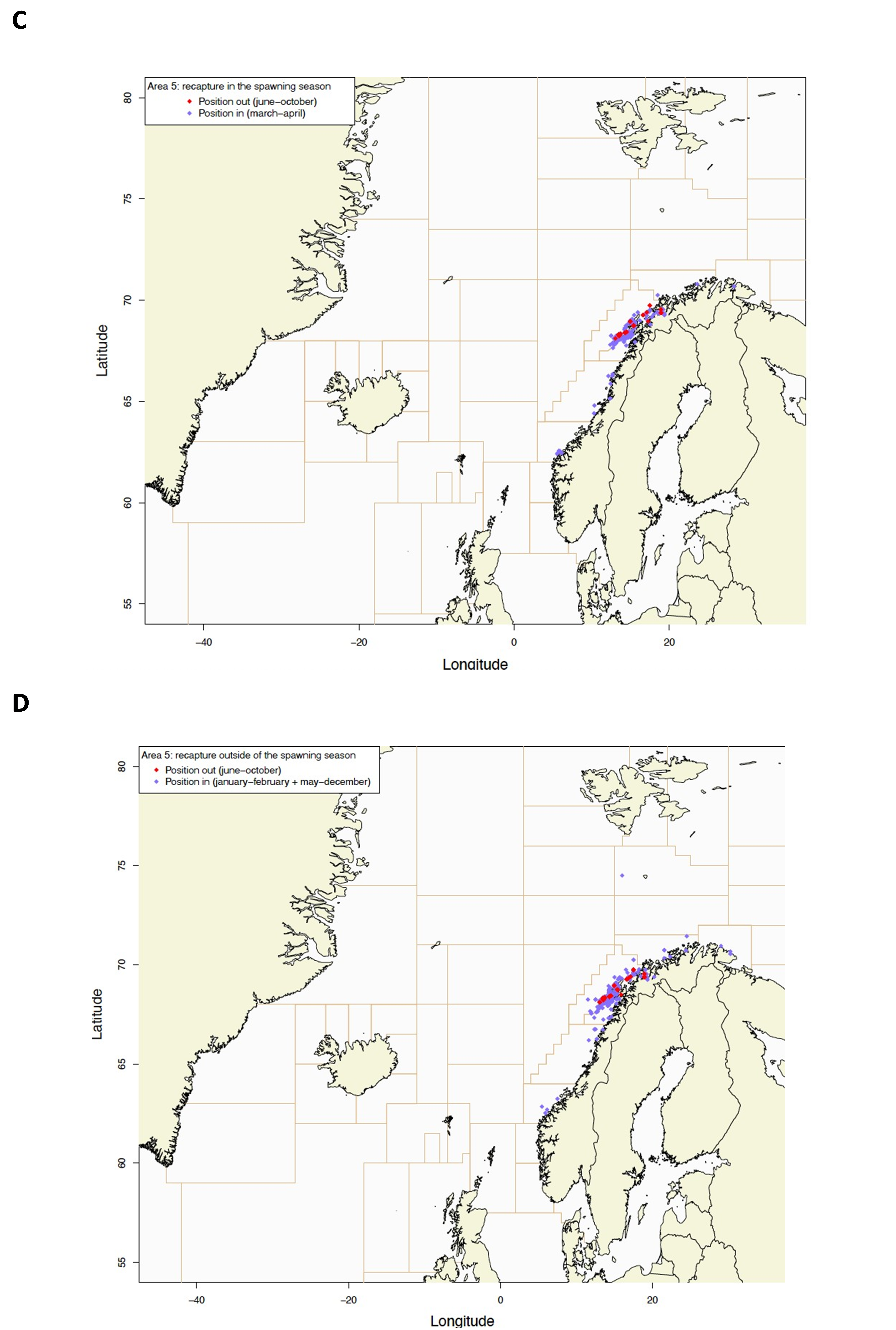
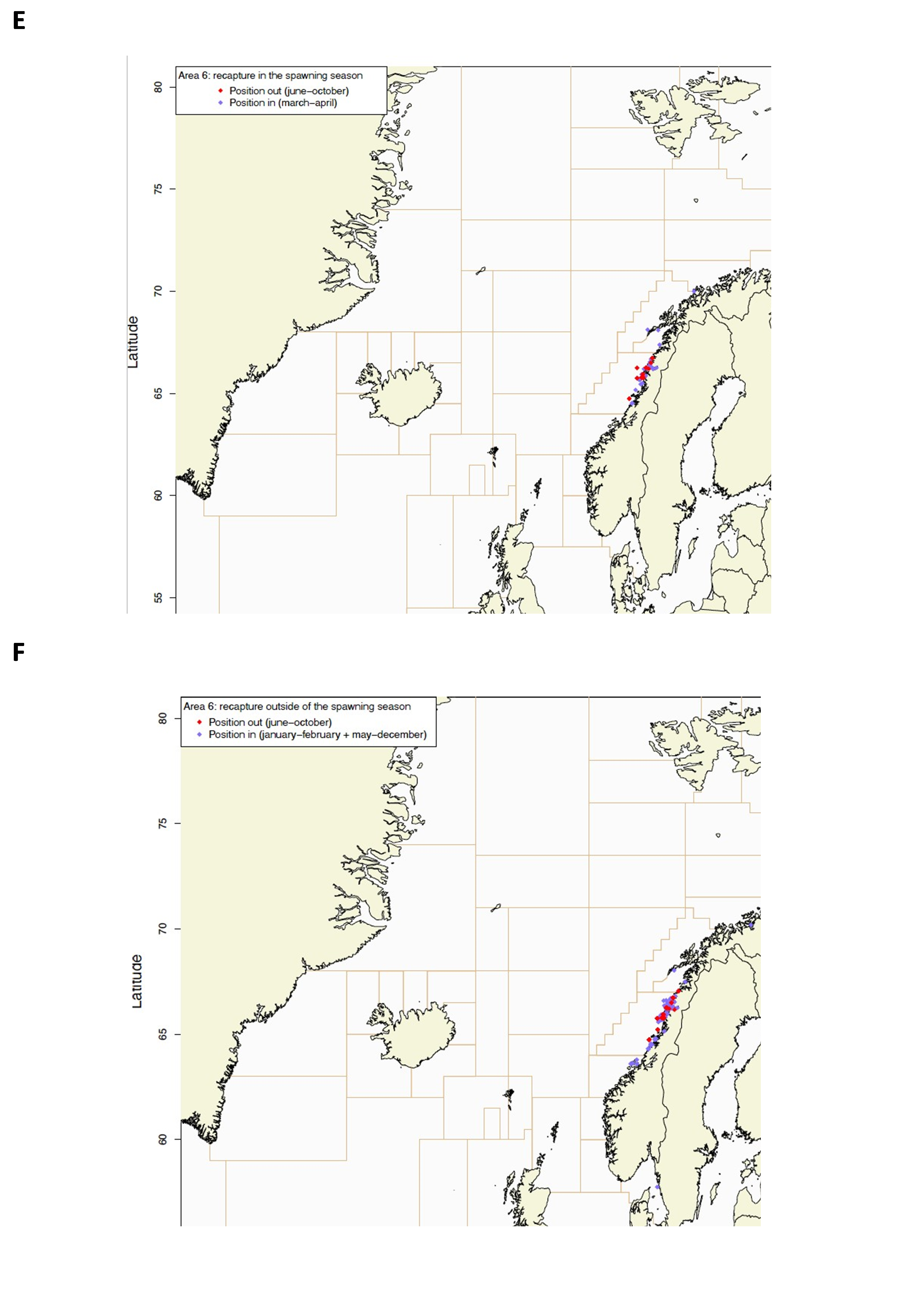
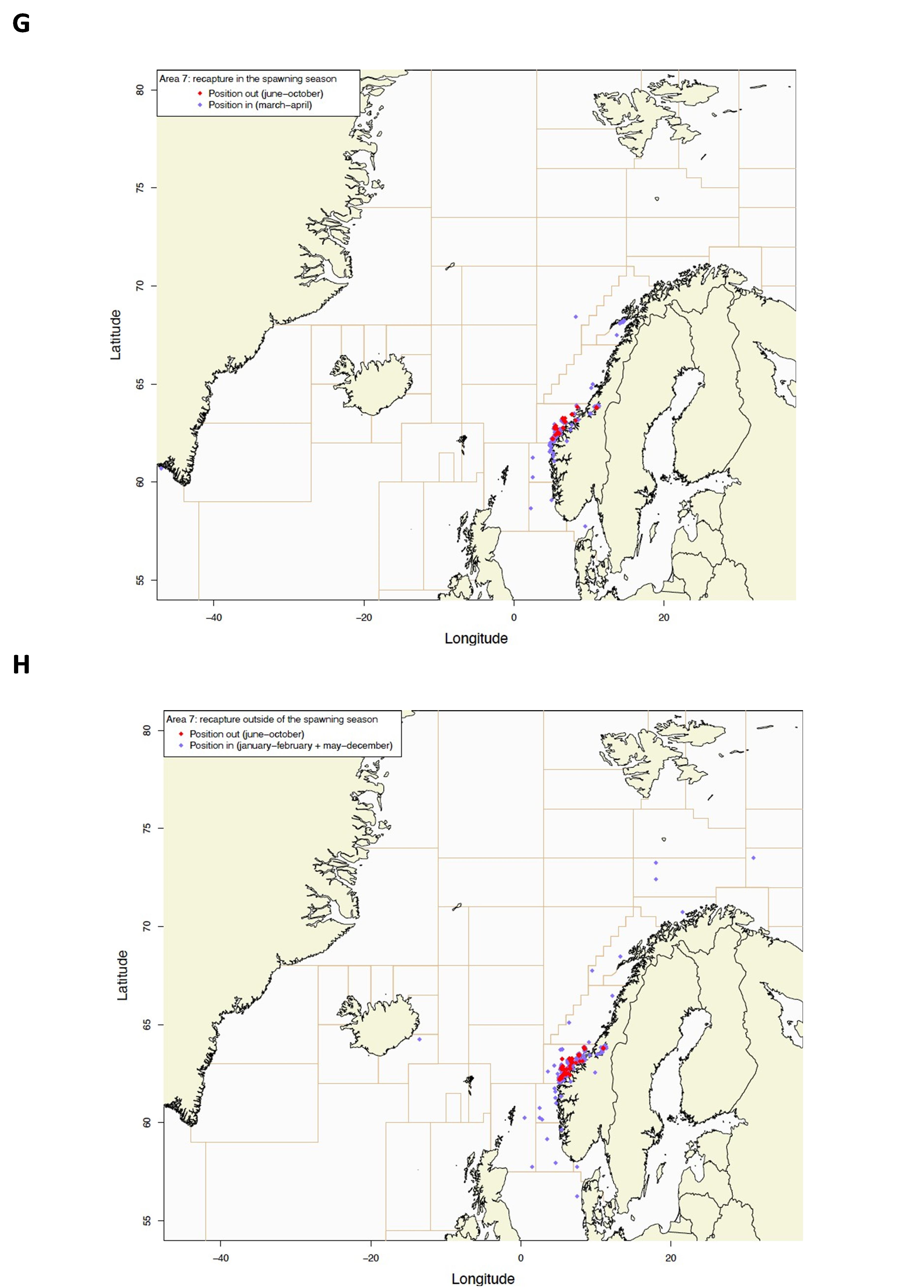
Gjenfangstprosent har variert mellom 10,6% og 39,5% med medianalder på den gjenfangede fisken som varierte fra 130-198 dager. Dette indikerer høy fiskedødelighet på de størrelsesgruppene som ble satt ut (middellengde 47,4-60,2 cm).
Utsettingslokalitet og gjenfangststed er plottet i Figur 2, fordelt på gjenfangster i gytesesong (mars-april) og utenom gytesesong (resten av året).
Tabell 2 og Figur 2 viser at hovedtrekket er at en stor del av gjenfangstene tas i nærhet av utsettingslokalitet (75% av gjenfangstene er tatt nærmere enn 27-35 km, og 90 % av gjenfangstene er tatt nærmere enn 40-75 km fra utsettingslokalitet). Noen enkeltindivider har vandret langt. Figur 1 viser at det er noe variasjon i avstand mellom utsettingslokalitet og gjenfangstlokalitet for torsk gjenfanget i ulike måneder i året.
Utsettingssted (fiskeriområde)
Min
25% kvartil
Median
Middel
75% kvartil
90% percentil
Max
0
0
0
7,4
22,5
27,0
40.0
752
5
0
5,4
16,6
37,6
29,6
57,1
799
6
0
7,8
15,5
42,3
30,6
63,0
893
7
0
4,9
17,0
51,2
35,0
74,8
2890
Alle
0
4,4
15,6
40,9
32,2
58,9
2890
Tabell 2. Avstand mellom utsettings- og gjenfangstlokalitet (km) for alle gjenfangster fra utsettinger i perioden juli-oktober i fiskeriområdene 0, 5, 6 og 7.
Figur 1. Boxplott av avstand mellom utsettingslokalitet og gjenfangststed forutsettinger i fiskeri-områdene 0 (a), 5 (b), 6 (c) og 7 (d). Outliers > 500 km er ikke tatt med.
Figur 2. Utsettingslokalitet og gjenfangststed for utsettinger i fiskeriområdene: 00 (A, B), 05 (C, D), 06 (E, F) og 07 (G, H). Utsetting i perioden juni-oktober i perioden 1958-1983 (Tabell 1), fordelt på gjenfangster i gyteperioden (A, C, E, G), og utenom den sentrale gyteperioden (B, D, F, H).
Vedlegg 3. Forekomst av kysttorsk per fangststatistikk-lokasjon
Asgeir Aglen, Karen Gjertsen, Sofie Gundersen og Kjell Nedreaas
Månedsvis prosent kysttorsk (basert på otolittmønster, angitt med røde tall) per fangststatistikk-lokasjon (angitt med svarte tall) per måned, for perioden 1980-1999, og for perioden 2000-2018. Inneholder data fra både forskningstokt og fra prøvetaking av fiskeriene.