Sammendrag
Siden det er praktisk umulig å få en fullstendig oversikt over luseinfeksjoner på laksefisk langs hele kysten ved hjelp av tradisjonelle feltobservasjoner, har Havforskningsinstituttet utviklet en lakselusmodell som utfyller observasjonene, både i tid og rom. Les mer om overvåking på https://www.hi.no/hi/temasider/arter/lakselus/overvaking-av-lakselus
Rapporten inneholder figurer som viser antall klekte nauplier per måned (aril - juli) i de ulike produksjonspmrådene. Videre beskrives og vises det figurer over hvordan tettheten av kopepoditter variere i tid og rom innen de ulike produksjonsområdene. Til slutt inneholder rapporten en forløpig diskusjon av betydningen av å gå fra 800m til 160m gitteroppløsning.
Rapporten er Appendiks IV til hovedrapporten om "Vurdering av lakseindusert dødelighet per produksjonsområde i 2019".
1 - Bakgrunn - hvorfor modellere?
Siden det er praktisk umulig å få en fullstendig oversikt over luseinfeksjoner på laksefisk langs hele kysten ved hjelp av tradisjonelle feltobservasjoner, har Havforskningsinstituttet utviklet en lakselusmodell som utfyller observasjonene, både i tid og rom. Les mer om overvåking på https://www.hi.no/hi/temasider/arter/lakselus/overvaking-av-lakselus
Vi vet at tetthet av lakseluskopepoditter er proporsjonal med sannsynligheten for smitte på villfisk (Myksvoll m.fl. 2018) og at lakselusa fordeler seg i vannmassene i fjord- og kystområdene på en svært variabel måte. Denne fordelingen varierer både over korte tidsperioder og små geografiske avstander (Asplin m.fl., 2014; Johnsen m.fl., 2014; Sandvik m.fl., 2016, Skarðhamar m.fl. 2018). Dette er en naturlig konsekvens av varierende strøm og lakselusenes vertikale posisjonering i varierende strømfelt (Johnsen m.fl., 2016). En rekke modellsimuleringer med likt utslipp av nauplier fra ulike anlegg har blitt utført (se for eksempel http://www.imr.no/hi/temasider/arter/lakselus/hvordan-spres-lakselusa og Asplin m.fl. 2014) og viser at en fullstendig beskrivelse av hvilken fordeling lakseluskopepoditter har langs kysten, bare kan gjøres gjennom å modellere spredning med bruk av informasjon om realistisk strøm, saltholdighet og temperatur.
Lakselusmodellen brukt ved Havforskningsinstituttet er en sammenkobling av flere ulike modeller (se https://github.com/bjornaa/ladim for en detaljert beskrivelse). Først beregnes utslipp (antall) av lakselus fra alle rapporteringspliktige oppdrettsanlegg i landet (Stien m.fl. 2005). Når luselarvene så er klekket ut i vannmassene opplever de temperatur, saltholdighet og strøm (representert ved den hydrodynamiske modellen) i den posisjonen de befinner seg. Basert på tilgjengelig kunnskap om biologi, atferd og dødelighet blir det beregnet hvordan larvene sprer seg med strømmen, først som ikke-smittsomme nauplier og videre som smittsomme kopepoditter (spredningsmodellen). Sluttproduktet er fordelingen av smittsomme lakseluslarver (kopepoditter) langs hele kysten time for time, noe som gir en god og detaljert oversikt over hvilke områder som har mye eller lite lus til enhver tid.
Modellsystemet baserer seg på eksisterende og veldokumenterte metoder. Den hydrodynamiske modellen (strøm) har mye til felles med værvarslings- og klimamodeller som har et omfattende vitenskapelig miljø for utvikling og validering, samt håndtering av usikkerhet. Vi kjenner heller ikke til eksisterer kunnskap om at lakselusa sin atferd eller dødelighet varierer mellom fjorder. Det er derfor rimelig å anta at modellsystemet for spredning av lakselus kan regnes som gyldig for hele kysten.
Modellen som blir benyttet i rådgivningen er satt opp på et gitter der hver rute er 800m x 800m. For 2019 har vi også satt opp en modell med et enda finere gitter (160m x 160m). Modellresultater fra 800m modellen blir hele tiden vurdert mot feltobservasjoner av lakseluspåslag på villfisk for å etablere så presise kvantitative sammenhenger som mulig.
Denne finere modellen er foreløpig ikke like godt testet mot observasjoner som det 800m modellen er, men de foreløpige testene som er utført tyder på at forskjellene først og fremst er lokale. Noen eksempler på resultat fra 160m modellen er beskrevet i avsnitt 8.
2 - Strømmodellsystem
Prinsippet for å skaffe informasjon om strøm er å inkludere en så fullstendig beskrivelse som mulig av de ulike fysiske prosessene og drivkrefter som vind, tidevann og ferskvannsavrenning. Dessuten trenger en like detaljert geografisk informasjon om bunndyp som modelloppløsningen. En hydrodynamisk modell beregner strøm, vannstand og hydrografi (saltinnhold og temperatur) prognostisk ved hjelp av numeriske teknikker med et tredimensjonalt beregningsgitter, og strømmodellen vi anvender er for tiden Regional Ocean Modeling System (ROMS, www.myroms.org; Albretsen m.fl., 2011). ROMS er satt opp for hele Norskekysten med gitteroppløsning 800 m, som er relativt høy romlig oppløsning for å dekke et så stort område. Det betyr at hele kysten og fjordene er delt inn i et rutenett der hver rute er på størrelse 800m ganger 800m horisontalt. Vertikalt er disse rutene delt i 35 lag som varierer tykkelsen fra noen dm nær overflaten til flere meter nedover i dypet. En horisontal gitteroppløsning på 800m representerer et kompromiss mellom tilgjengelige regnemaskiners kapasitet og behovet for å beskrive vannmassenes dynamikk og ikke minst bunntopografi og kystlinje. Atmosfæriske drivkrefter (vind og stråling) henter vi fra en separat vindmodell (WRF, www.wrf-model.org, eller operasjonelle atmosfæreprognoser fra Meteorologisk institutt) og åpne grenseverdier mot havet tas fra havmodellresultater med større geografisk utstrekning og verdier minst hver time. Modelloppsettet for Norskekysten inkludert systemet for realistiske inngangsdata og drivkrefter kalles NorKyst800 (Albretsen m.fl., 2011) og modellresultater er publisert i en rekke vitenskapelig publikasjoner (Asplin m.fl. (2014), Johnsen m.fl. (2014), Johnsen m.fl. (2016), Sandvik m.fl. (2016), Myksvoll m.fl. (2018), Skardhamar m.fl. (2018)), som viser god overensstemmelse med modellert og observert strøm og hydrografi.
Resultater fra strømmodellen brukt for spredningsberegninger for lus 2012-2019 er presentert i Appendiks III: Fysisk miljø.
I tillegg til 800m modellen, gjøres det en rekke modellsimuleringer der strømmodellen benytter 160m horisontal gitteroppløsning. Med likt vertikalt gitter er likevel disse simuleringene nesten 10 ganger så tidkrevende som simuleringene med 800m horisontal gitteroppløsning. Det er opplagt at 5 ganger finere gitter vil gjengi topografien og strømmene i trangere farvann og fjorder bedre enn med 800m gitteroppløsning.
3 - Spredningsmodell for lakselus
Som et ledd i arbeidet med å videreutvikle modellsystemet ble partikkelspredningsmodellen (LADIM) til HI omprogrammert til et mer fleksibelt system våren 2017. Den nye modellversjonen er fritt tilgjengelig på (https://github.com/bjornaa/ladim). Det har blitt utført en grundig sammenligning av resultatene fra ny og gammel versjon som viser at selv om resultatene er relativt like avviker de såpass mye at det var nødvendig med en rekalibrering av smittepresskartet (ROC) og den virtuelle smoltmodellen (Myksvoll m.fl. 2018a). I en nylig publisert artikkel (Crosbie m.fl. 2019) ble det i laboratorieeksperiment funnet at lakselus-kopepodittene i økende grad unnviker vann med saltholdighet fra 31 til 23. Dette ble implementert i Havforskningsinstituttet sin lakselusmodell, til erstatning fra den gamle saltholdighets unnvikelsen på 20 (Heuch 1995). Den nye metoden er validert mot smoltburdata fra 2012- 2107 og viste en betydelig forbedring av det estimerte smittepresset (Sandvik m.fl. 2019).
3.1 - Parametrisering av lakselusas utvikling og atferd
Vi har valgt å spre planktonisk lakselus som partikler og beregne spredningsprosessen, vekst, atferd og dødelighet basert på informasjon fra litteraturen og sensitivitetsstudier med modell (Asplin m.fl., 2011, Asplin m.fl., 2014, Johnsen m.fl., 2014, Johnsen m.fl., 2016). Horisontalt driver lakseluslarvene passivt med strømmen, men vertikalt er lakseluslarvene i stand til å posisjonere seg basert på miljøfaktorer som lys og saltholdighet. Lakselusene er i modellen gitt evnen til å plassere seg nær overflaten, når eksponert for lys, og å synke lenger ned i vannsøylen ved lave saltholdigheter. I tillegg vil den vertikale forflytningen inneholde småskala turbulent bevegelse. Dødelighet er inkludert i modellen og parameterisert som 17 % per døgn, basert på Stien m.fl. (2005).
Lakselusa vil vokse og utvikle seg som en funksjon av temperatur, og vi modellerer dette ved å bruke døgngrader som er tid multiplisert med omgivelsestemperatur. Grensene for når lakselusa er i de to første nauplie-stadiene og når de er i det smittsomme kopepodittstadiet angis i døgngrader og er basert på laboratoriestudier som har vist at det smittsomme kopepodittstadiet er mellom 40 og 170 døgngrader (Samsing m.fl., 2016). Det betyr at lakseluslarvene utvikler seg raskt til kopepoditter når det er varmt i vannet, men samtidig blir levetida kort (eks. 17 dager ved 10ºC). Ved lave vanntemperaturer tar utviklingen lengre tid, men lusene kan også leve lenger. Disse forskjellene i utvikling og levetid kan påvirke hvor langt lusene kan spres geografisk.
De modellerte verdiene for lakseluskonsentrasjon er avhengig av hvor mange nauplier som klekkes (kildeledd, hovedsakelig lus på oppdrettsfisk) og hvor stor dødeligheten er. Begge deler er i dag (og i overskuelig fremtid) beheftet med usikkerheter, og selv om vi benytter best tilgjengelig kunnskap per dags dato mener vi at det er et potensiale for å forbedringer.
Slike forbedringer av modellen pågår kontinuerlig etter hvert som ny kunnskap om biologien til lakselusa blir publisert vitenskapelig. Fordelingen av lakseluskopepoditter i rom og tid vil kvalitativt bare i mindre grad påvirkes av detaljer i kildeledd (se under) eller dødelighet. Manglende utslippskilder for lus, f.eks. fra slaktemerder, vil derimot kunne være av stor betydning.
3.2 - Kildeleddet
Kildeleddet i modellsystemet er antall egg som klekkes av lus i oppdrettsanleggene, altså antall luselarver som slippes ut fra hvert anlegg. Dette beregner vi basert på den rapporteringen industrien gir ukentlig for antall lus per fisk og månedlig for biomasse (omregnet til antall fisk) i merdene. Ideelt sett skulle denne informasjonen vært tilgjengelig hyppigere og med eksakt dato. En detaljert beskrivelse av hvordan vi beregner kildeleddet er beskrevet i Ådlandsvik et al (2017) .
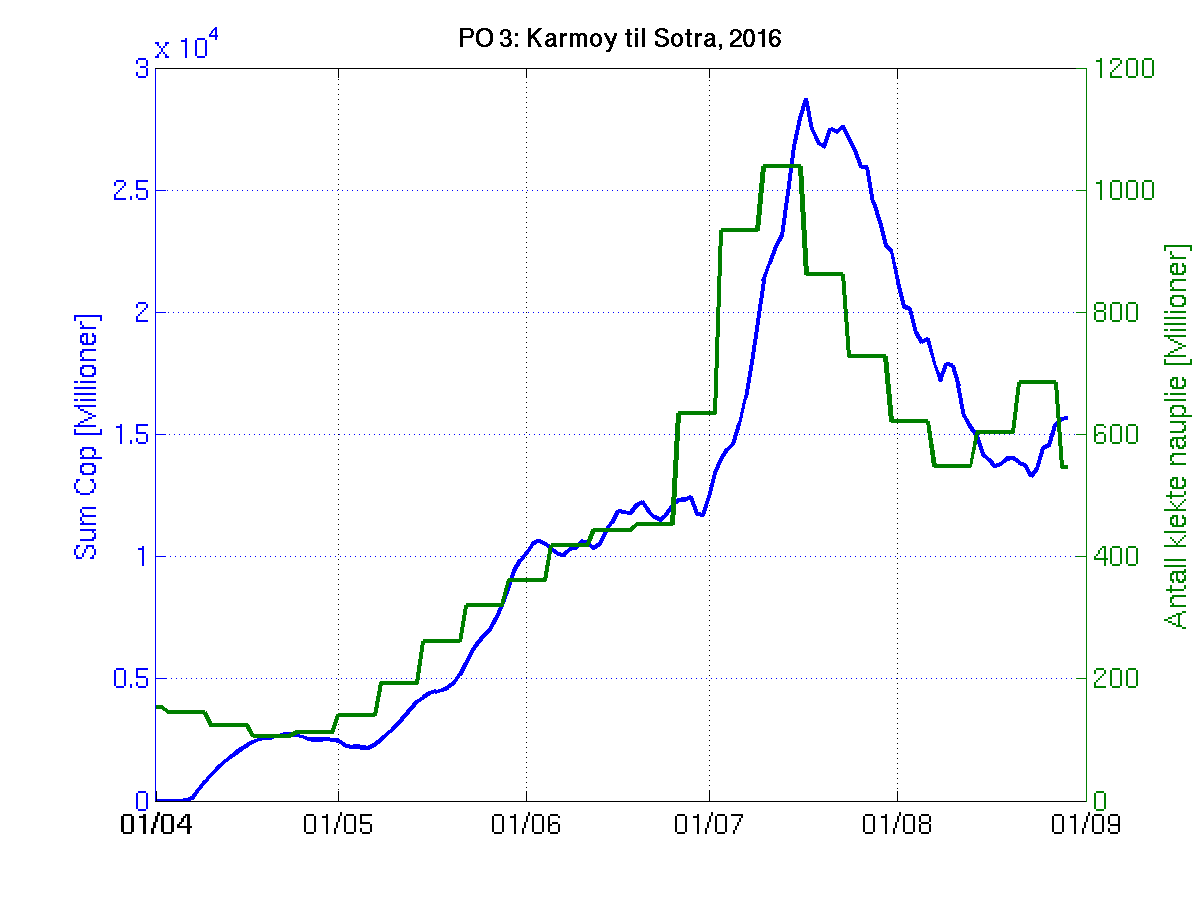
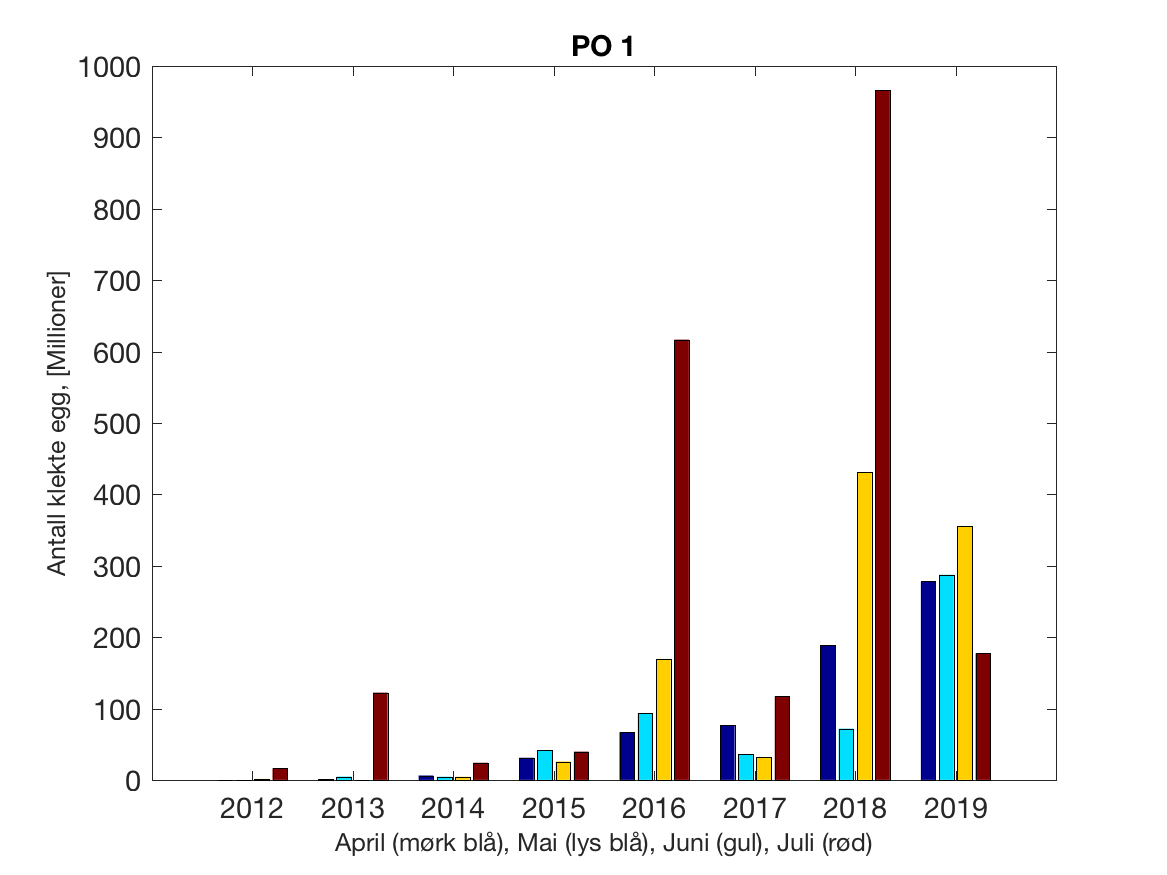
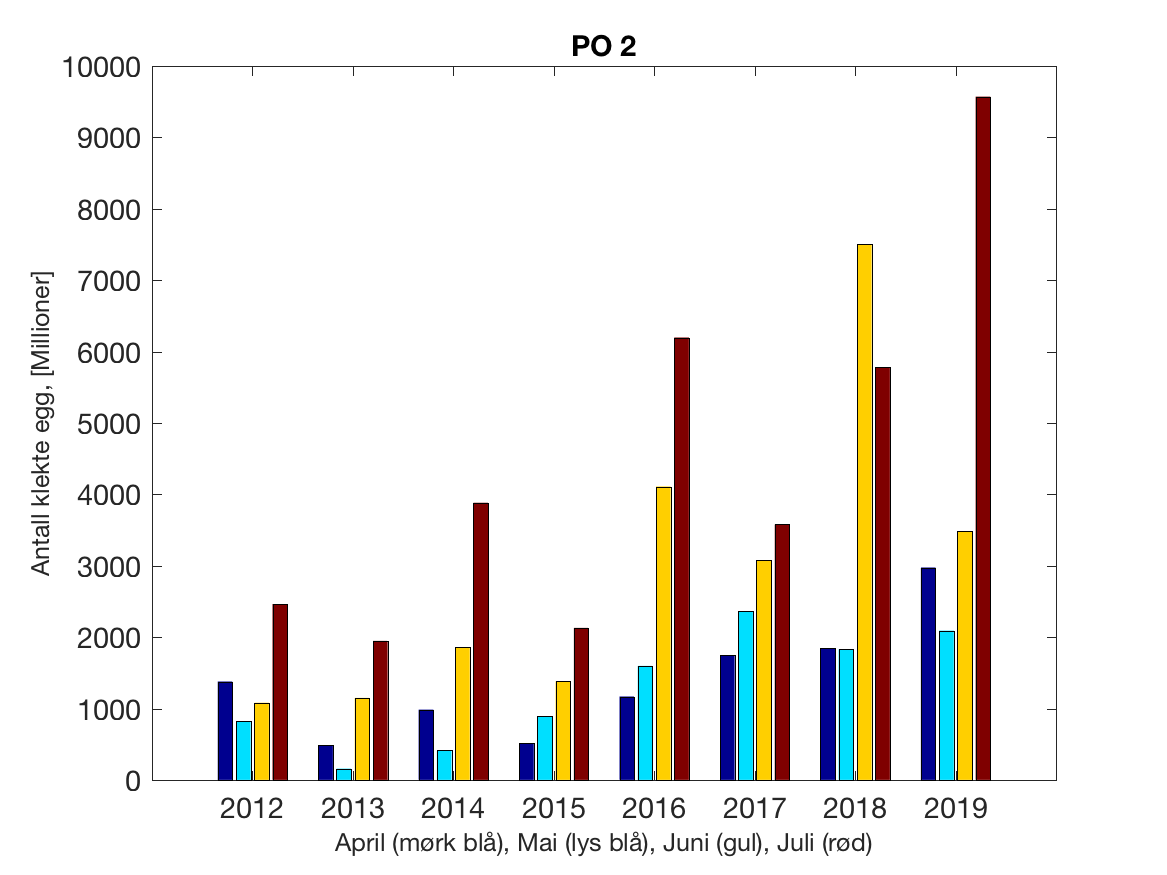
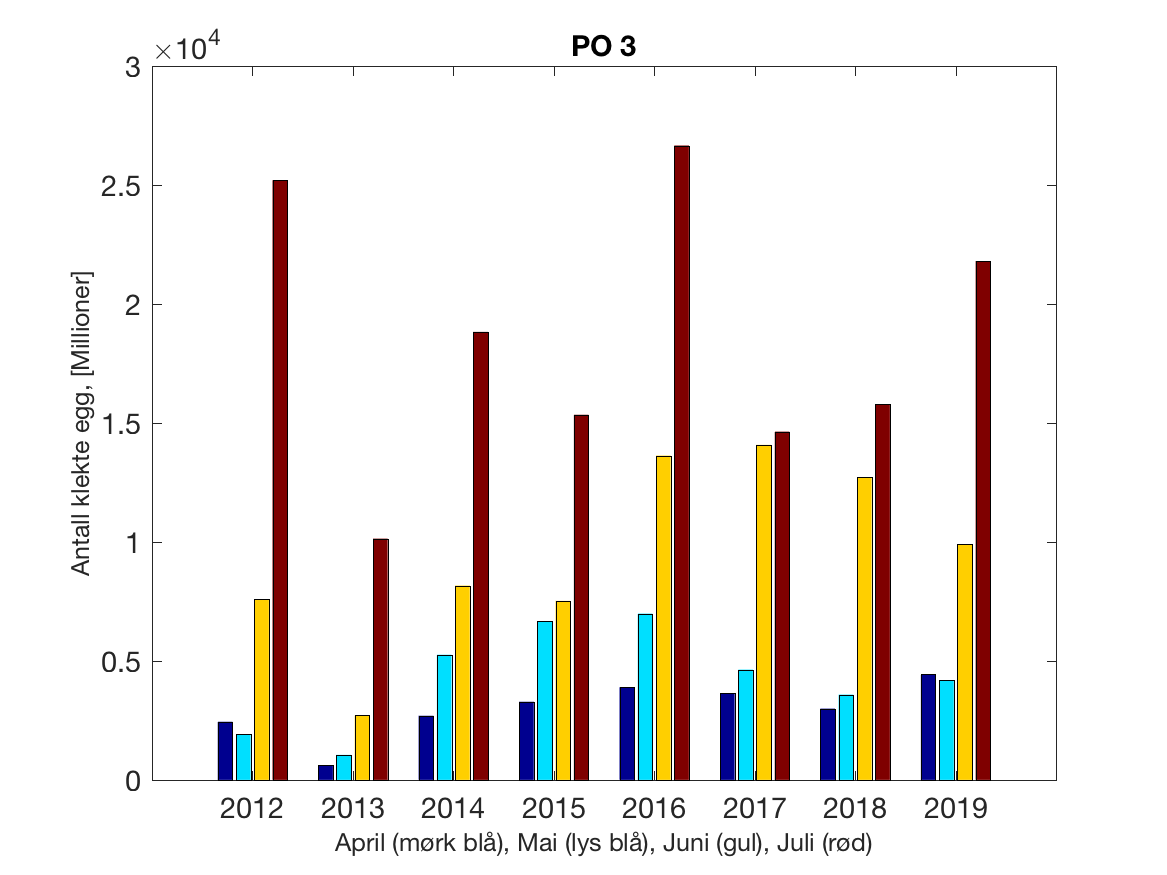
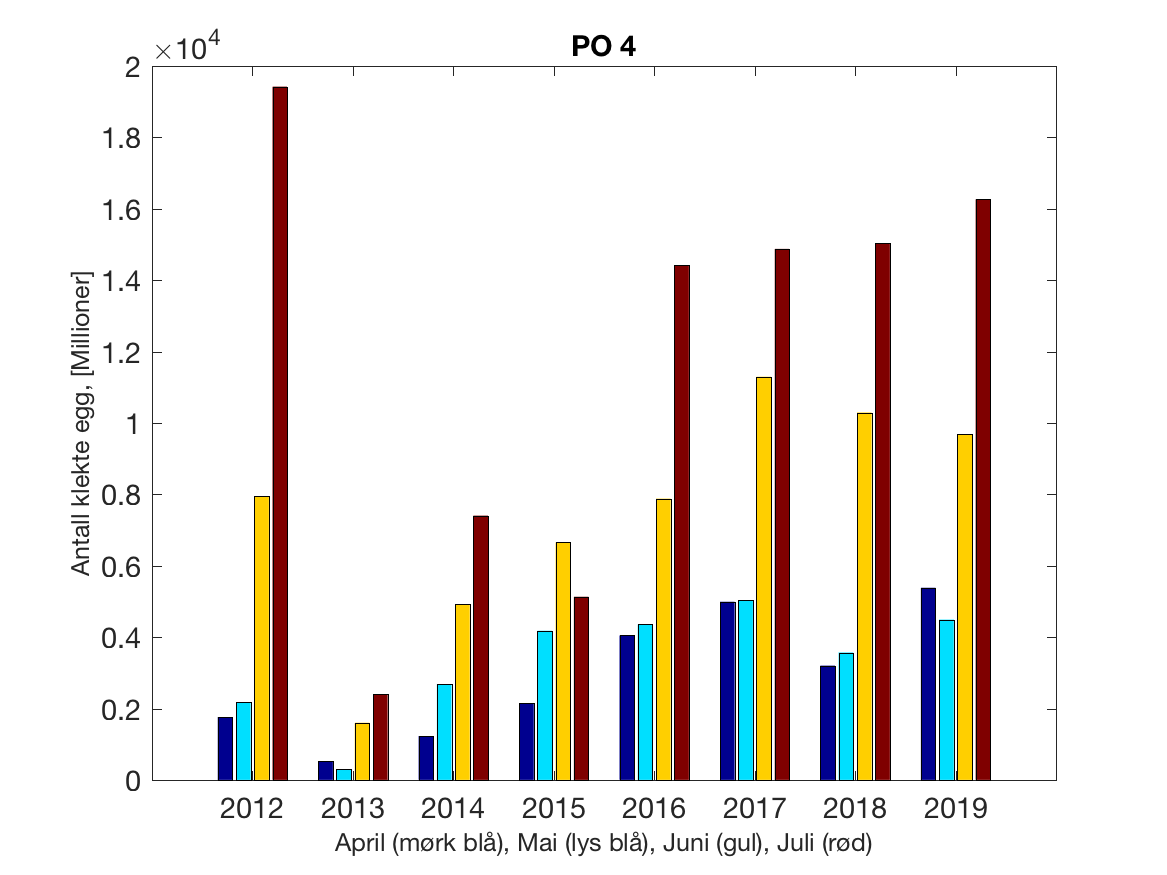
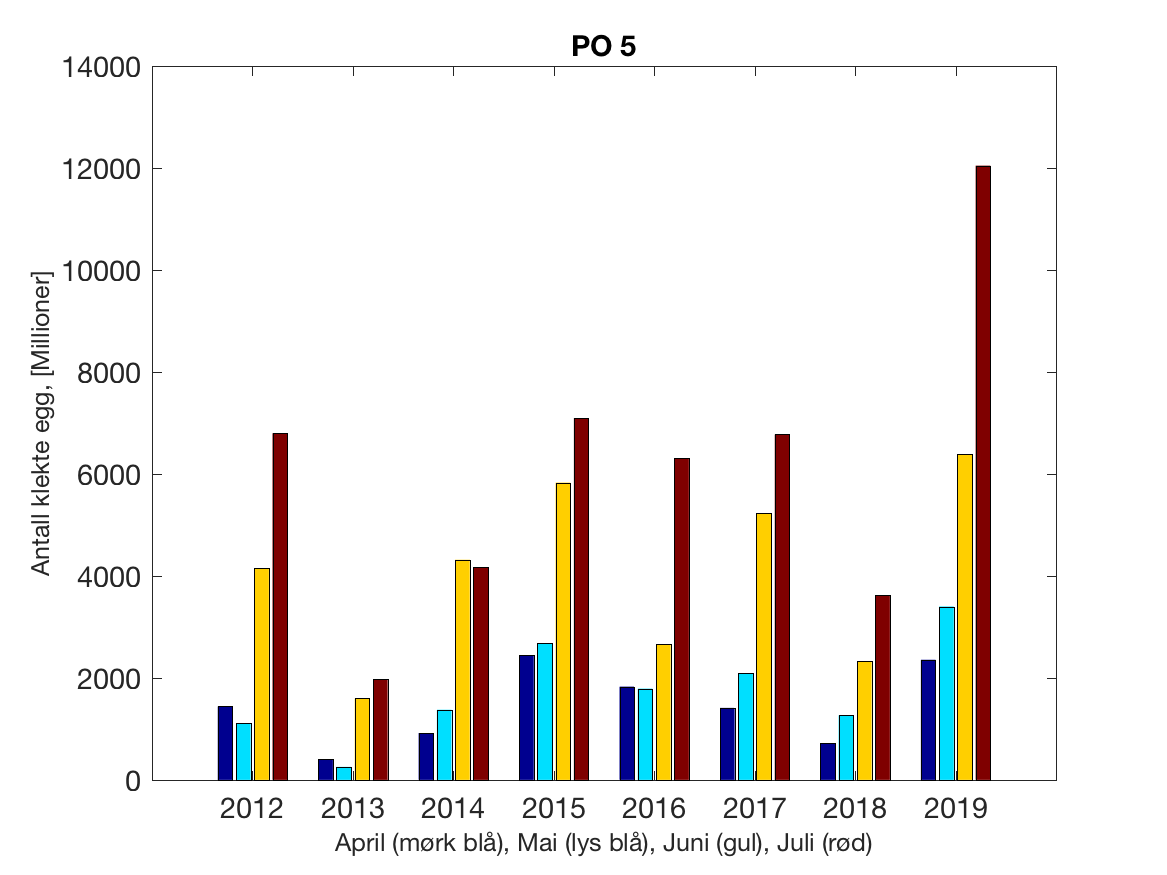
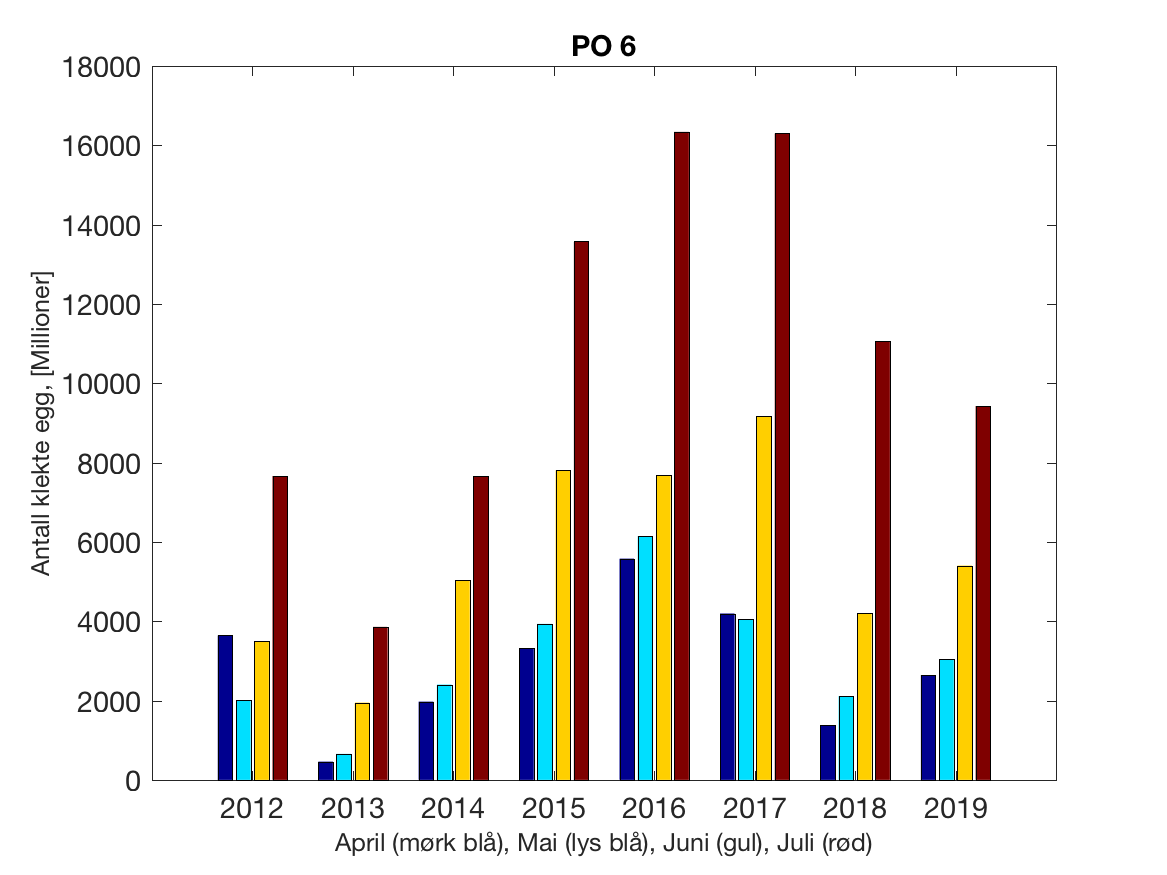
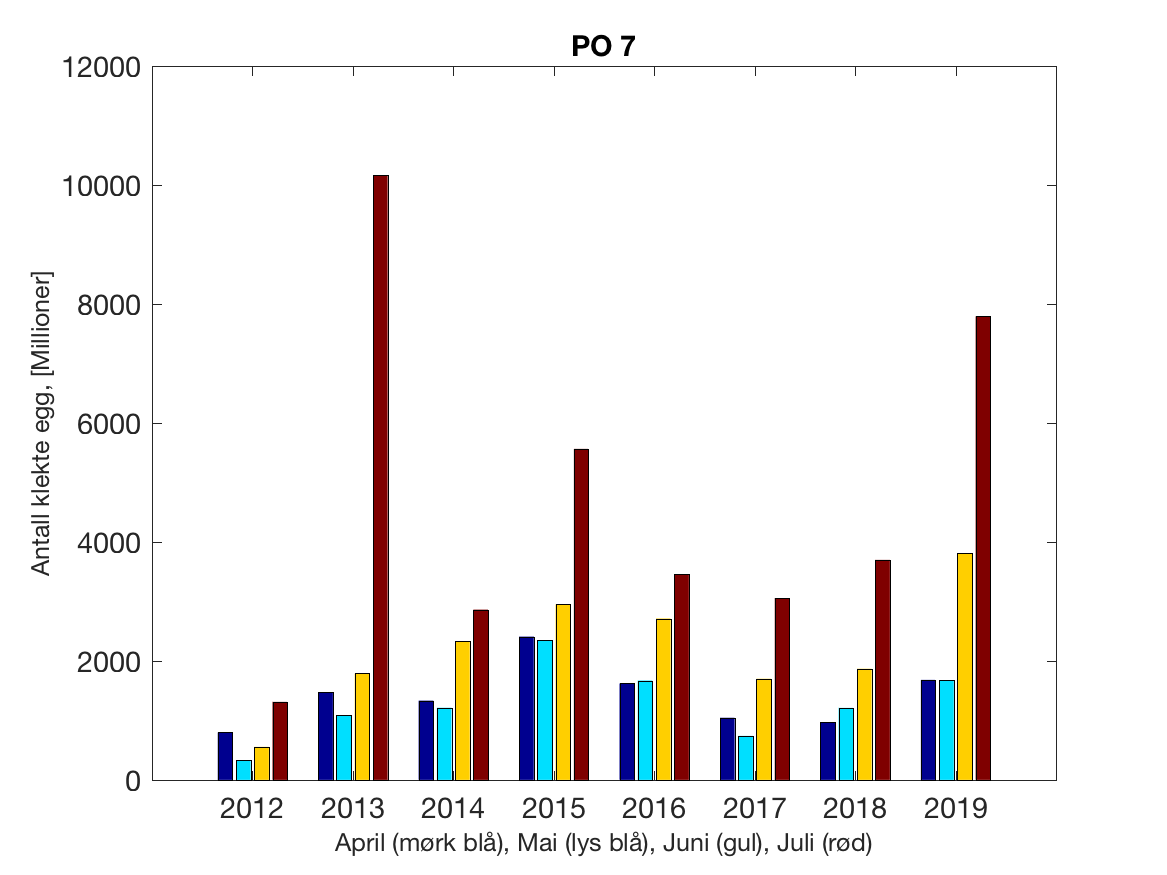
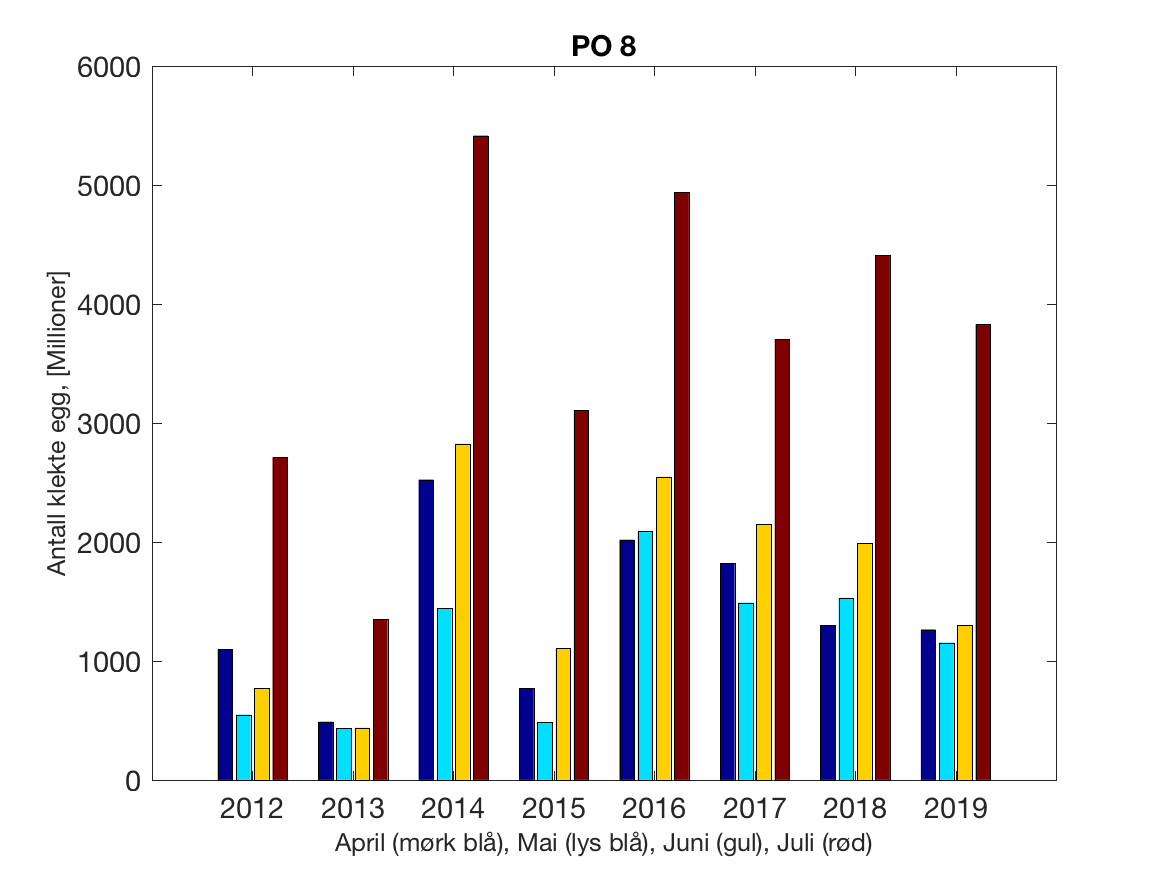
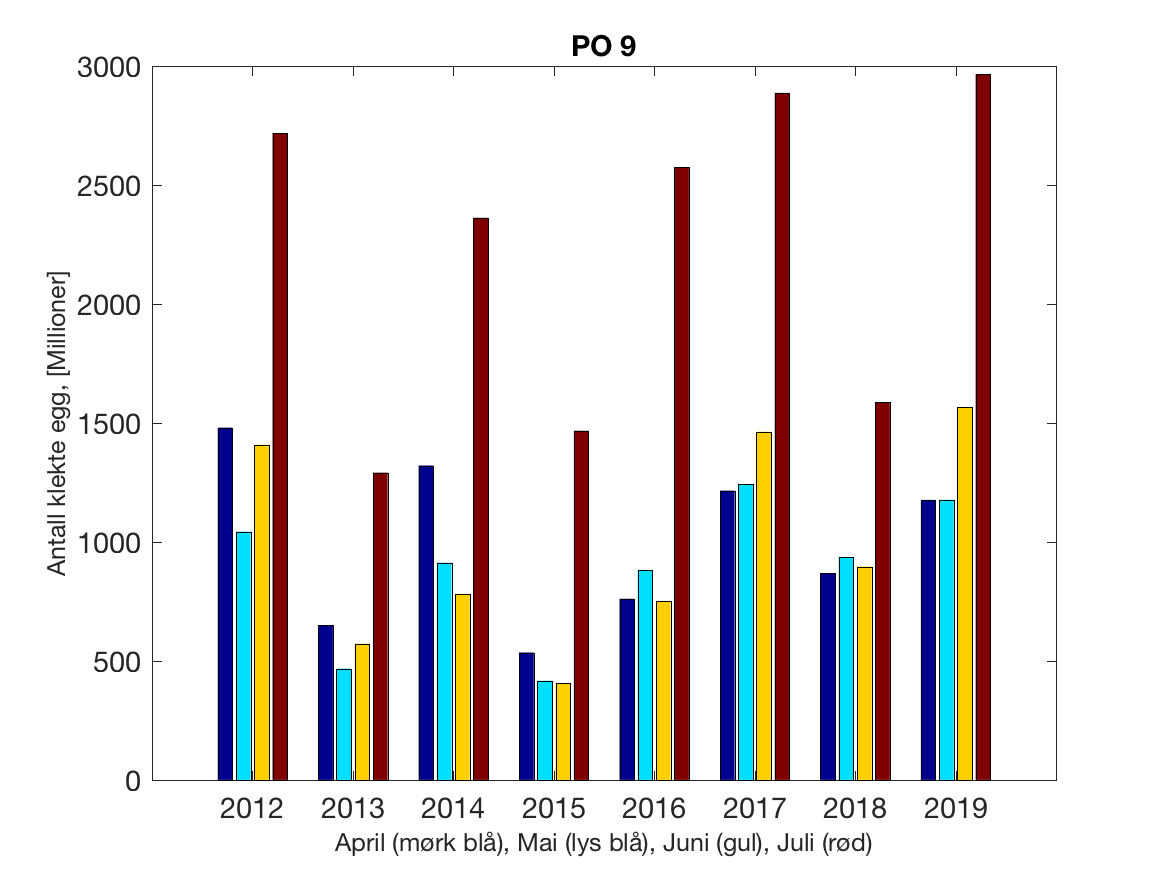
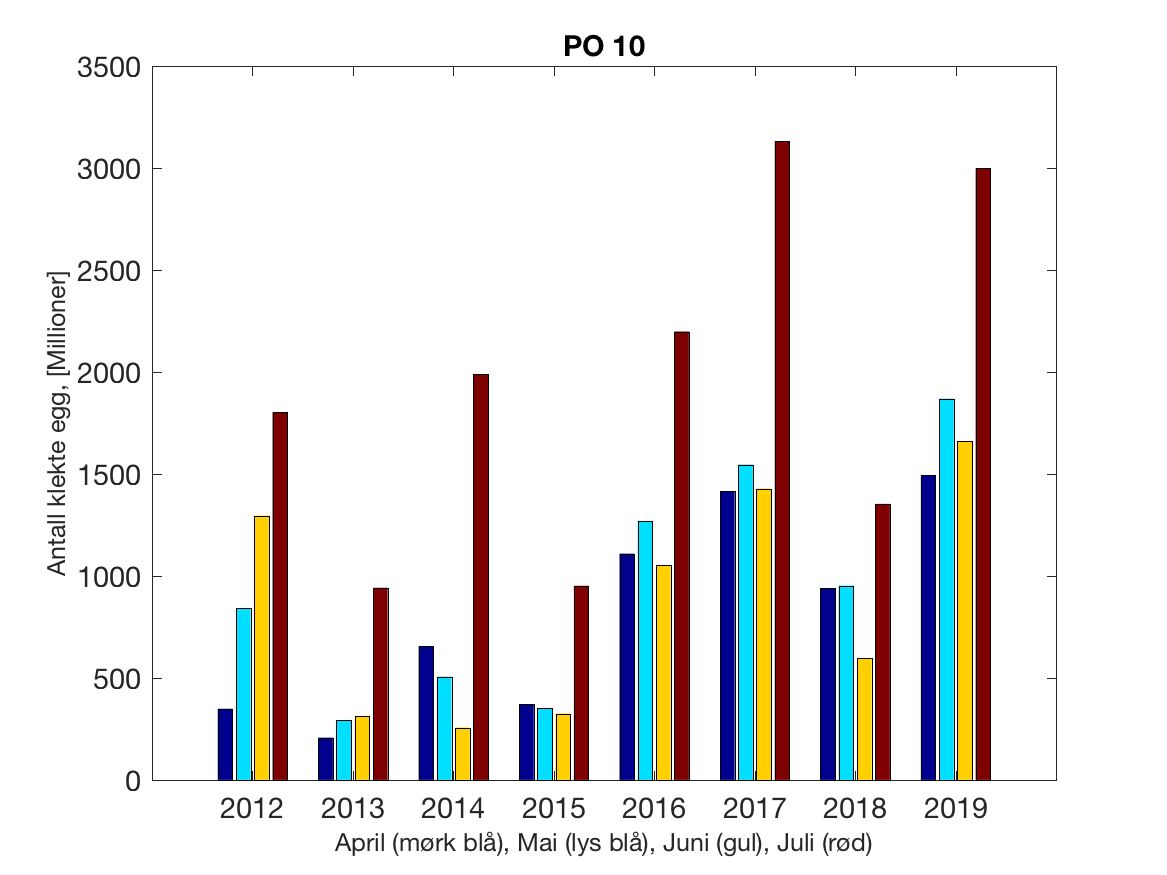
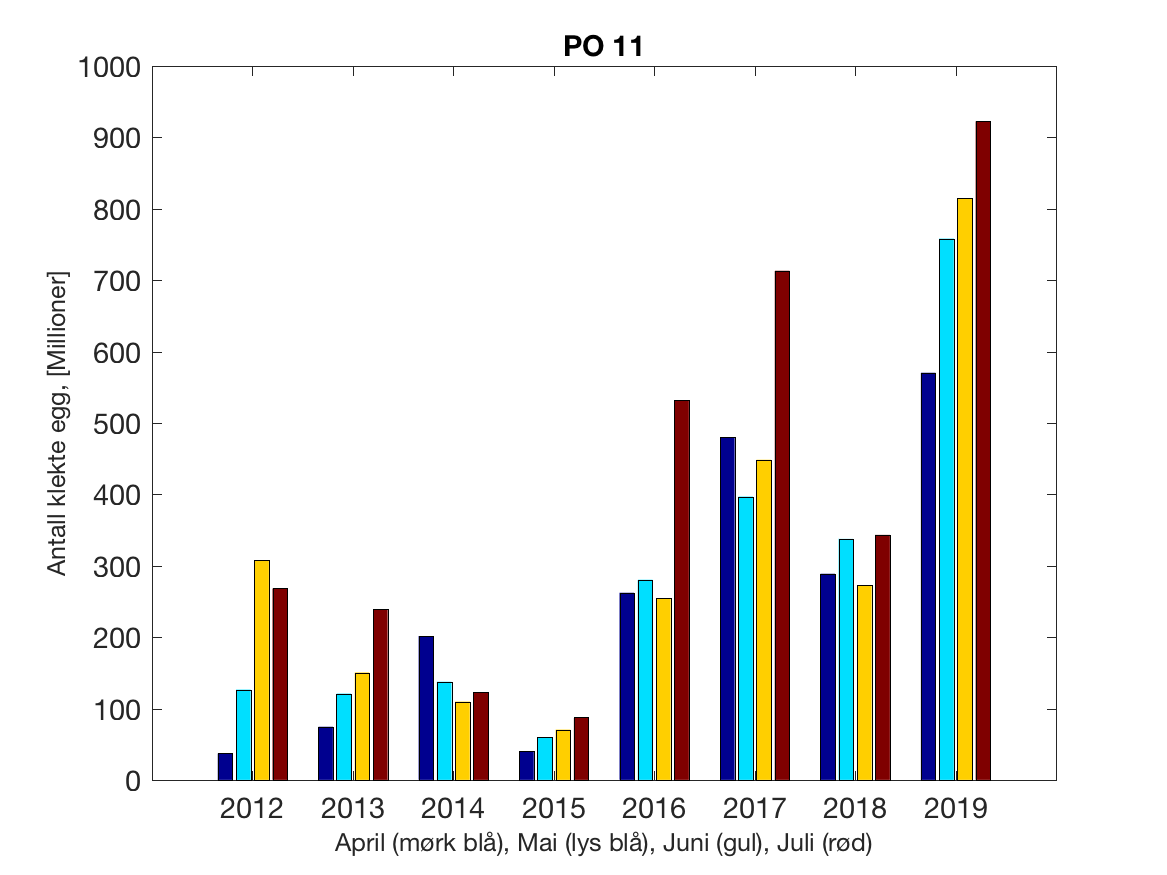
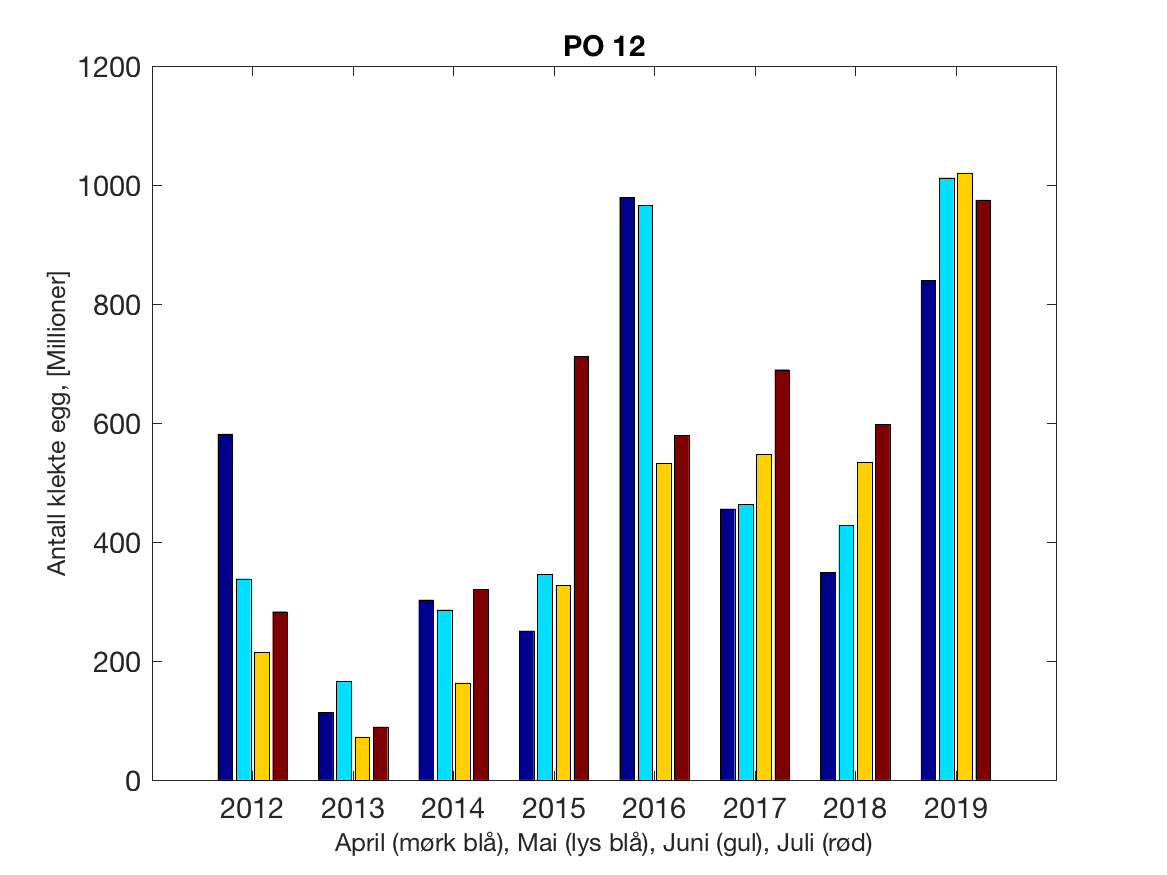
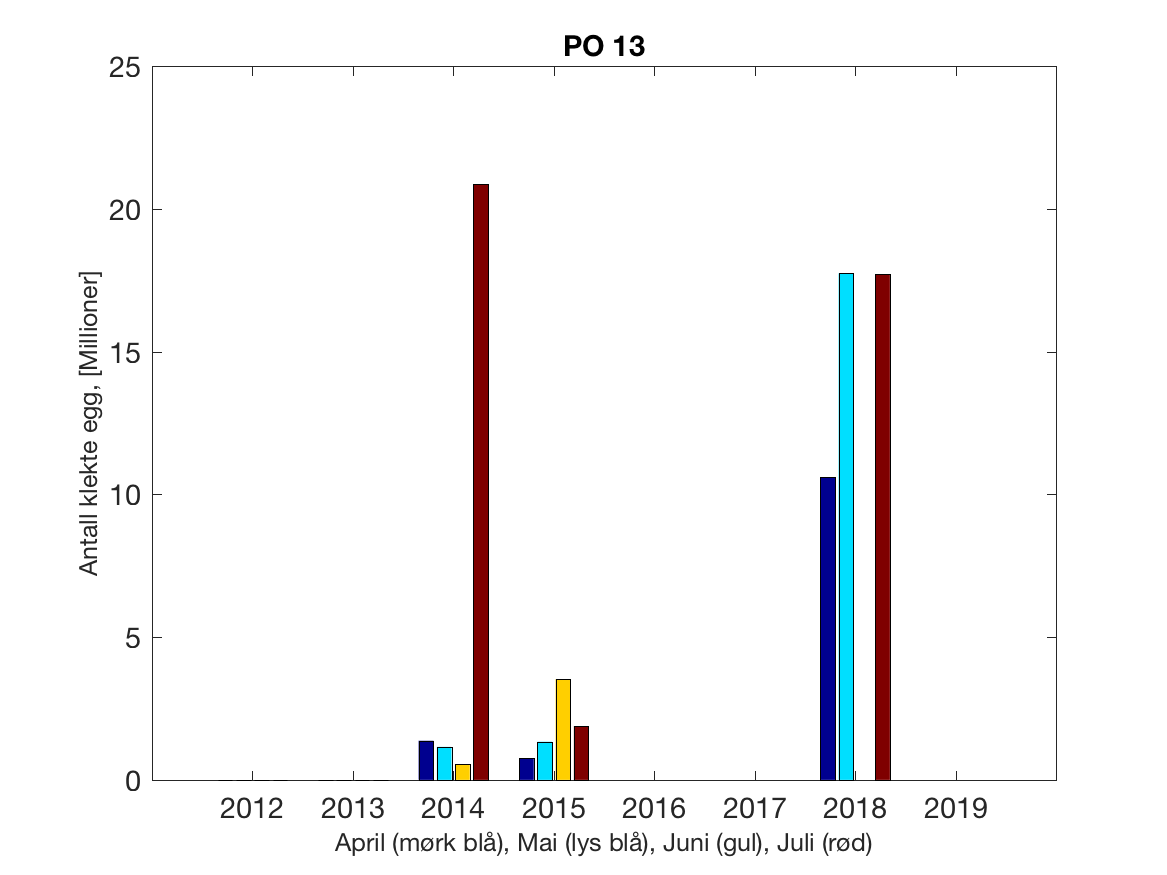
Siden grensene mellom produksjonsområdene er plassert slik at det skal være minimal utveksling av partikler mellom områdene vil summen av antall infektive kopepoditter følge klekkingen av nauplier (innen POet) med en tidsforsinkelse som er avhengig av vanntemperaturen, som vi kan se av eksempelet i Figur 1. I figur 2 er summen (per måned) av antall klekte nauplier per produksjonsområde plottet år for år. Merk at skalaen på y-aksen varierer mellom produksjonsområdene.
Kildeleddet har potensiale for forbedring i de kommende årene ved å få sikrere og mer detaljerte rapporteringstall fra oppdrettsanleggene (se f.eks. Skar ð hamar et al 2018).
Figur 1. Sum over antall nyklekte nauplier (grønn) og sum over antall smittsomme kopepoditter (blå) i produksjonsområde 3. Kurvene følger hverandre, men endringen av antall kopepoditter ligger litt etter endringen av nauplier.
Figur 2. Kildeleddet (antall klekte nauplier) integrert over produksjonsområde for månedene April-Juli. Merk at skalaen på y-aksen varierer mellom produksjonsområdene.
4 - Modellresultater
Sluttproduktet fra lakselusmodellen er en geografisk fordeling av planktonisk lakselus (stadiefordelt på nauplier og kopepoditter) time for time for en tidsperiode. Dette kan for eksempel presenteres som tetthet, antall lus per m², for en gitt periode og for en gitt del av vannsøylen
Grunnlagsdataene for de to kolonnene ”HI smittepress” og ”HI virtuell smolt” i hovedrapporten (”Vurdering av lakselusindusert villfiskdødelighet per produksjonsområde i 2019”) er døgnverdier av tetthet for den øvre delen av vannsøylen (summert fra overflaten og ned til 2m), hvor vi antar at den ville fisken hovedsakelig oppholder seg.
Denne typen resultater kan produseres for en hvilken som helst periode der vi har modellresultater for strøm, samt reelle estimat på hvor mange lakselus-nauplier som slippes ut (kildeleddet, beskrevet i 3.2). Konsistent informasjon om antall lus på fisk i anlegg finnes tilbake til 2012, og Havforskningsinstituttet har produsert et arkiv som inneholder geografisk fordeling av planktonisk lakselus time for time for perioden 1/3 – 1/9, 2012 til 2019. Dette arkivet oppdateres når for eksempel ny kunnskap om lakselusens biologi er tilgjengelig, slik at best mulige data kan benyttes i rådgivningen. I tillegg produseres det ukentlig 10-dagers gjennomsnittsverdier som publiseres på Havforskningsinstituttets webside www.lakselus.no .
5 - Validering av modellresultatene
Det er viktig å kunne bestemme i hvilken grad modellresultatene gir en realistisk beskrivelse av det smittepresset de ville laksefiskene opplever. Validering av denne type modellsystem er imidlertid ingen enkel øvelse:
For det første beregner modellen antall smittsomme lakseluskopepoditter (smittedose), mens de felt-observasjoner er fra tellinger av lakselus (alle stadier) som sitter på fisken (smitterespons). Vi trenger altså å etablere en sammenheng mellom antall kopepoditter i vannmassene og det antallet som fester seg på fisken. Det finnes noen studier som beskriver hvordan temperatur, saltholdighet og strøm påvirker smitteprosessen (Tucker m.fl., 2000; Browman m.fl., 2004; Genna m.fl., 2005; Frenzl, 2014). Hvordan man skal representere denne smitteprosessen som en funksjon av flere variable er imidlertid enda ikke beskrevet i litteraturen, og som en første ordens tilnærming antar vi en lineær sammenheng mellom smittedose og smitterespons for individuell fisk.
For det andre må vi finne en best mulig måte å sammenligne diskrete punktobservasjoner med grov tidsoppløsning, med modellresultater som har god oppløsning i både tid og rom. Vi vet at lakseluskopepodittene ikke er jevnt fordelt i vannmassene, men ofte som «flak» eller «flekker» med forhøya tetthet, og disse kan flyttes relativt raskt med strømmen i noen områder og perioder, og ha lang oppholdstid i andre områder/perioder. Sammenligning mellom de to datasettene, som har både forskjellig oppløsning og presisjon, må derfor gjøres med omhu. Dette kan gjøres på flere måter, og metoden vi har valgt er i beskrevet i Sandvik m. fl., 2016, basert på observasjoner fra smoltbur i Hardangerfjorden. Resultatene, som inkluderer fire år og mer enn 100 ulike datapunkt, viste at fordelingen av lakseluslarver estimert med modellen passet godt sammen med den skjeve (mange lave verdier og få høye) fordelingen som kom fram i det observasjons-datasettet, inndelt i infeksjonsklasser for gjennomsnittlig antall lus per fisk i burene. Infeksjonsklassene er definert som færre enn 1 lus i snitt per fisk (abundans), 1-5 lus, 5-10 lus og flere enn 10 lus per fisk. I 78 % av burobservasjonene var modellen i stand til å estimere riktig infeksjonsklasse, mens den i 18 % av burobservasjonene underestimerte med en luseklasse. I de resterende 4 % ble det over- eller underestimert med 2 luseklasser. Det ble også vist at den høye treffsikkerheten til modellen er langt over det vi kunne ha oppnådd ved en ren tilfeldighet.
Arbeidet i Sandvik m.fl. (2016) er videreført i Sandvik m.fl. (2019). Observasjonene er her utvidet til 6 år (2012-2017), og gjennom en stegvis prosess der en beregner både treff og falske alarmer er det utviklet en metode som enkelt kan kvantifisere kvaliteten på ulike modeller for lakselus. Dette er eksemplifisert ved å implementere en delvis saltunnvikelse basert på resultatene fra Crosbie m.fl. (2019), noe som gir en signifikant forbedring i forhold til resultatene fra Sandvik m.fl. (2016). Med den nye metoden er det også mulig å velge en vilkårlig grense for høy/lav slik en kan tegne «kart» med ulike grenser (f.eks. 2-6 og 1-10).
Myksvoll m.fl. (2018b) har sammenlignet modellert fordeling av lakseluskopepoditter med observert mengde lus på villfanget ørret. Kun fisk som ble fanget i den første perioden av overvåkingen i hvert produksjonsområde (Nilsen m.fl., 2017) er inkludert i sammenligningen, for å representere forventet utvandringstidspunkt for laksesmolt i de ulike områdene. Videre er studien avgrenset til fisk som er mindre enn 150 gram. Perioden strekker seg fra slutten av mai på Sørlandet til slutten av juni i Finnmark. Datagrunnlaget, etter disse avgrensningene, fra 2015 utgjør 1088 fisk fordelt på 21 stasjoner og fra 2016 var det 1733 fisk fordelt på 44 stasjoner. Fra observasjonene er summert antall tidlige lus (kopepoditter, chalimus 1+2) og beregnet totalt antall lus relativt til vekt (lus per gram kroppsvekt) i gjennomsnitt for hver stasjon. Fra modellen er antall kopepoditter beregnet innenfor ulike avstander fra observasjonspunktet (3x3, 5x5, 9x9 gridruter), midlet i rom og summert over en tre ukers periode som inkluderer de to ukene prøvefisket har foregått og en uke i forveien (Tabell 1, Figur 5).
Tabell 1: Spearman rank korrelasjon mellom observasjoner (tidlige stadier og relativt antall per gram) og modellresultater i ulike avstander omkring observasjonspunktet (3x3, 5x5, 9x9 gridpunkter).
| 2015/2016 | Tidlige stadier | Totalt antall (lus/gr) |
| 3x3 gridruter | 0,61/0,57 | 0,63/0,67 |
| 5x5 gridruter | 0,61/0,64 | 0,62/0,69 |
| 9x9 gridruter | 0,71/0,69 | 0,73/0,72 |
Når man slår sammen de to ulike årene til en lang serie blir korrelasjonen 0,675 for tilfellet med tidlige stadier og 9x9 gridpunkter (0,71/0,69). Ellers er tidsseriene holdt separat fordi antall observasjoner er cirka dobbelt så mange i 2016 sammenlignet med 2015 og stasjonene er ikke nødvendigvis de samme mellom år. 2016 var også det første året med observasjoner i alle produksjonsområder.
Sammenligningen viser at modellen viser høy korrelasjon mellom modellert og observert smittepress i hele landet. Kvaliteten på modellresultatene anses derfor som svært god og man kan dermed konkludere at modellen beregner realistisk smittepress i alle produksjonsområdene, både høye og lave verdier.
6 - Modellprodukter
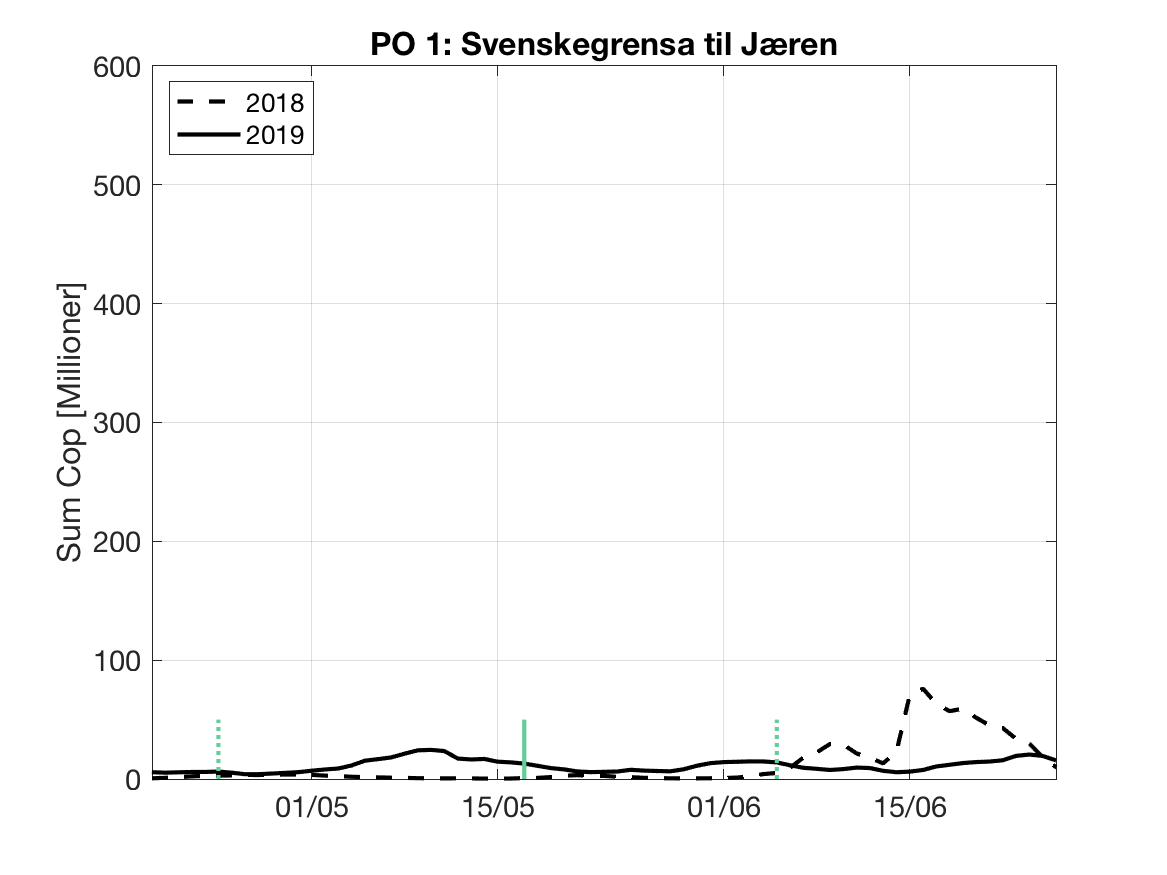
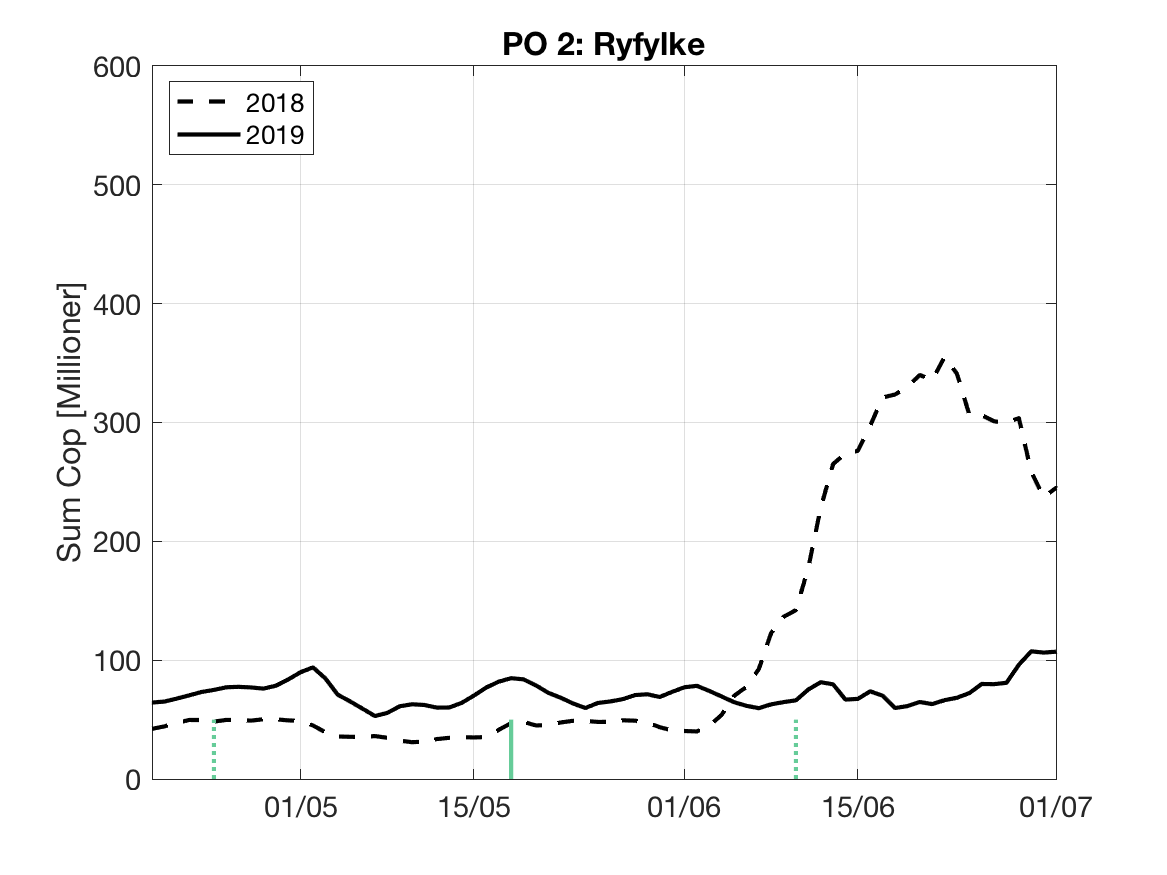
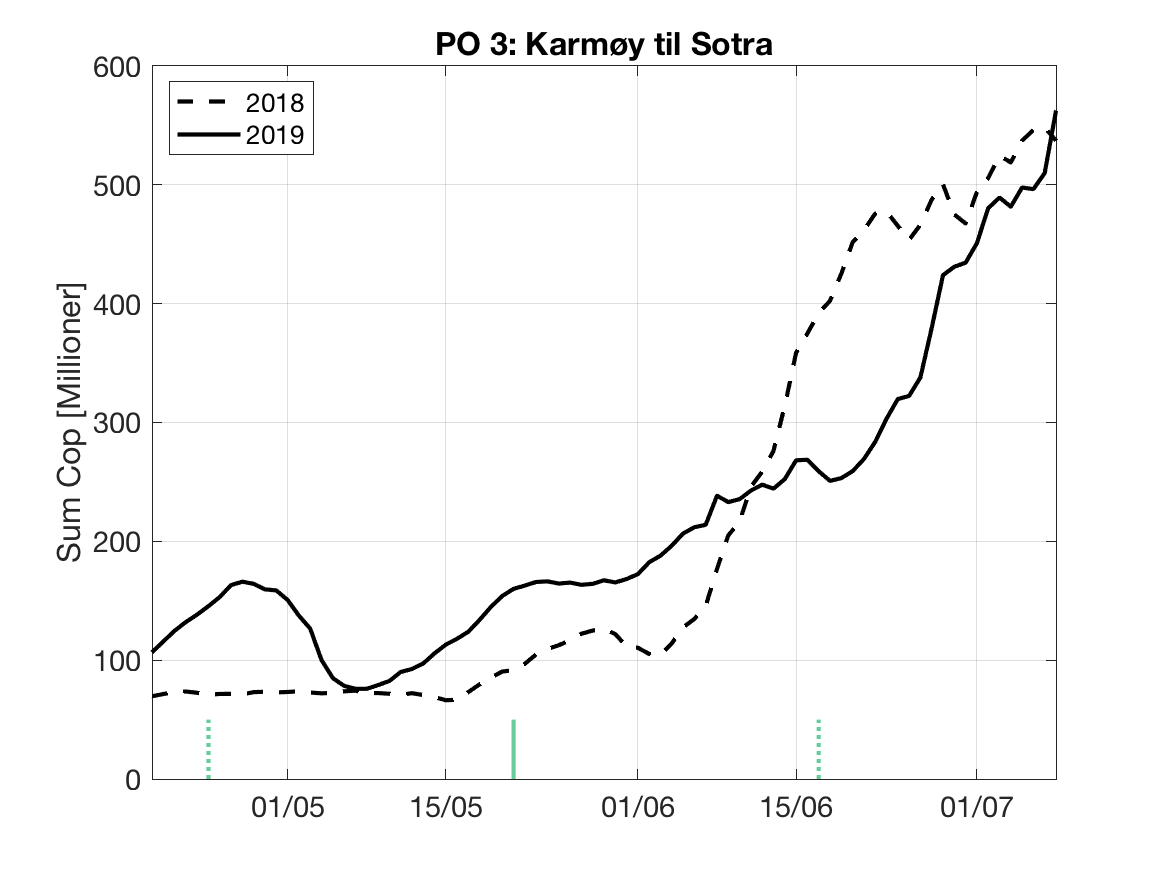
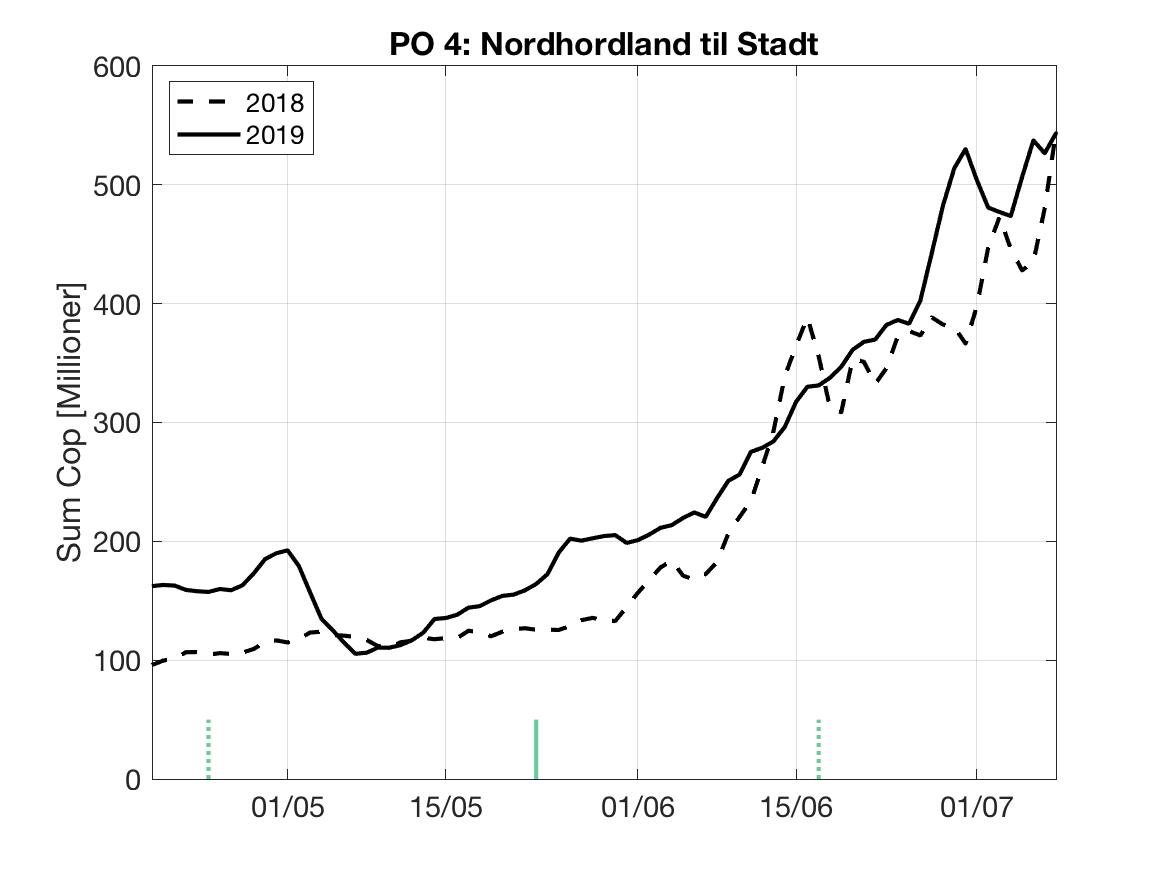
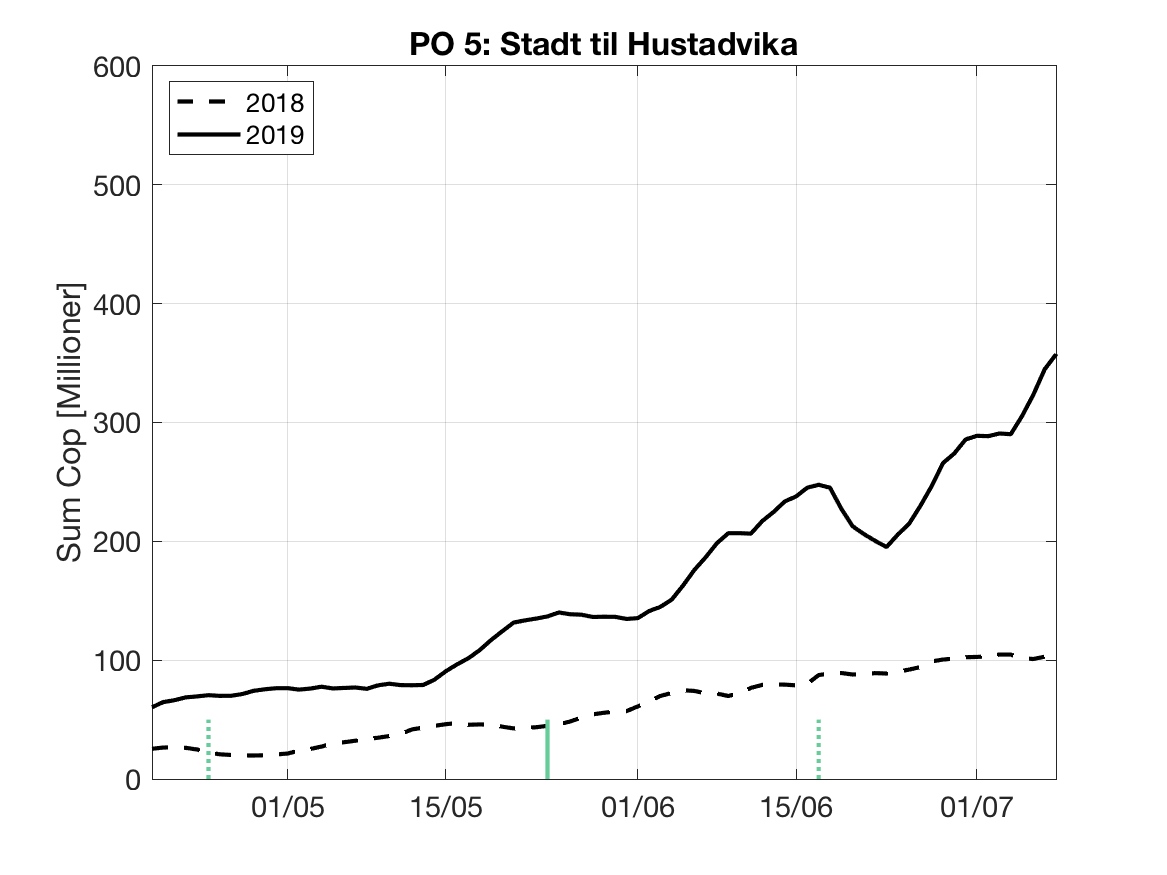
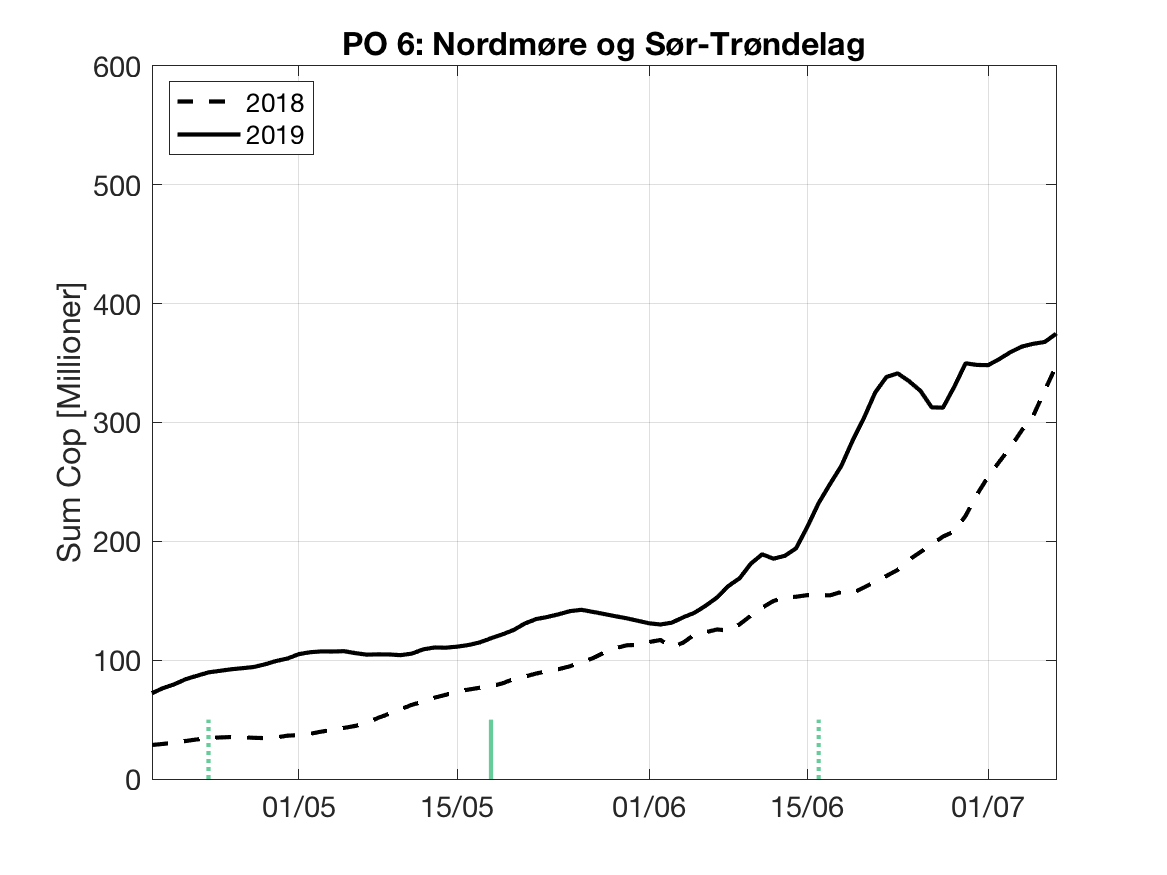
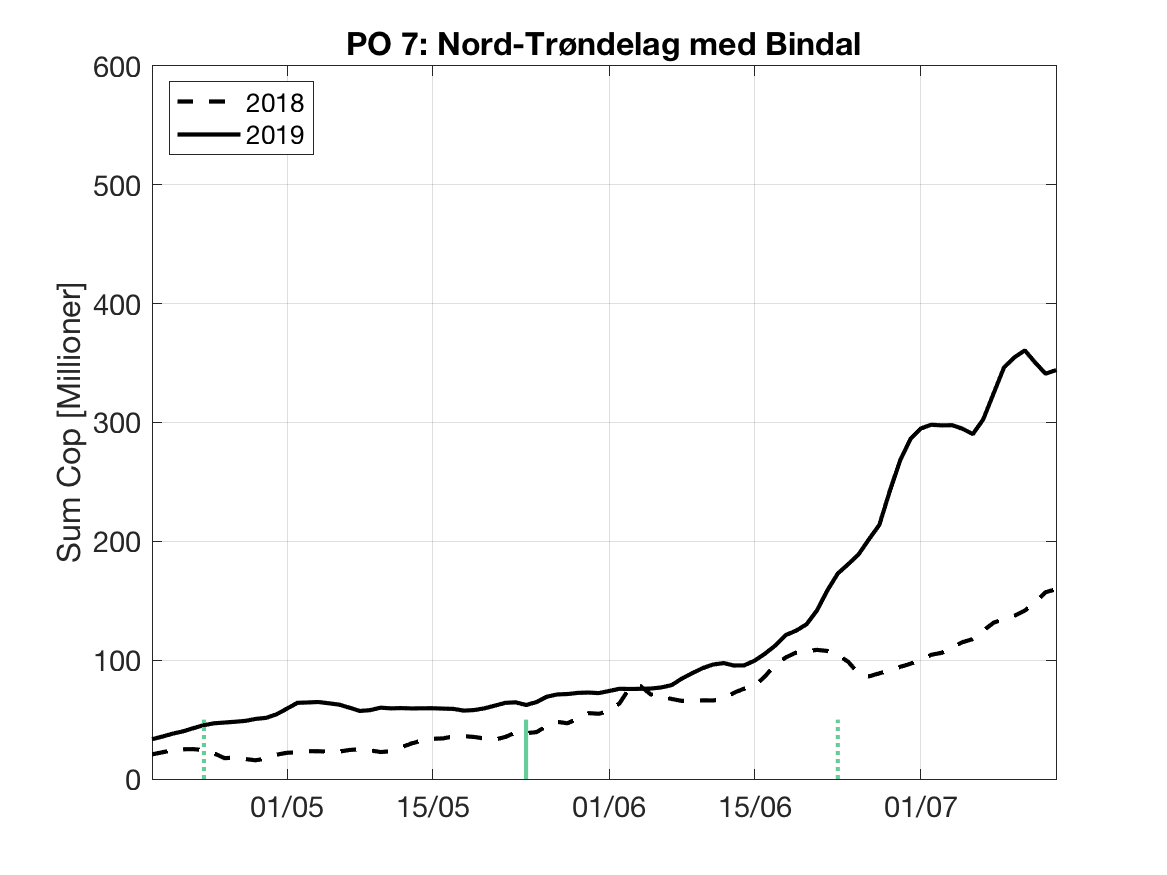
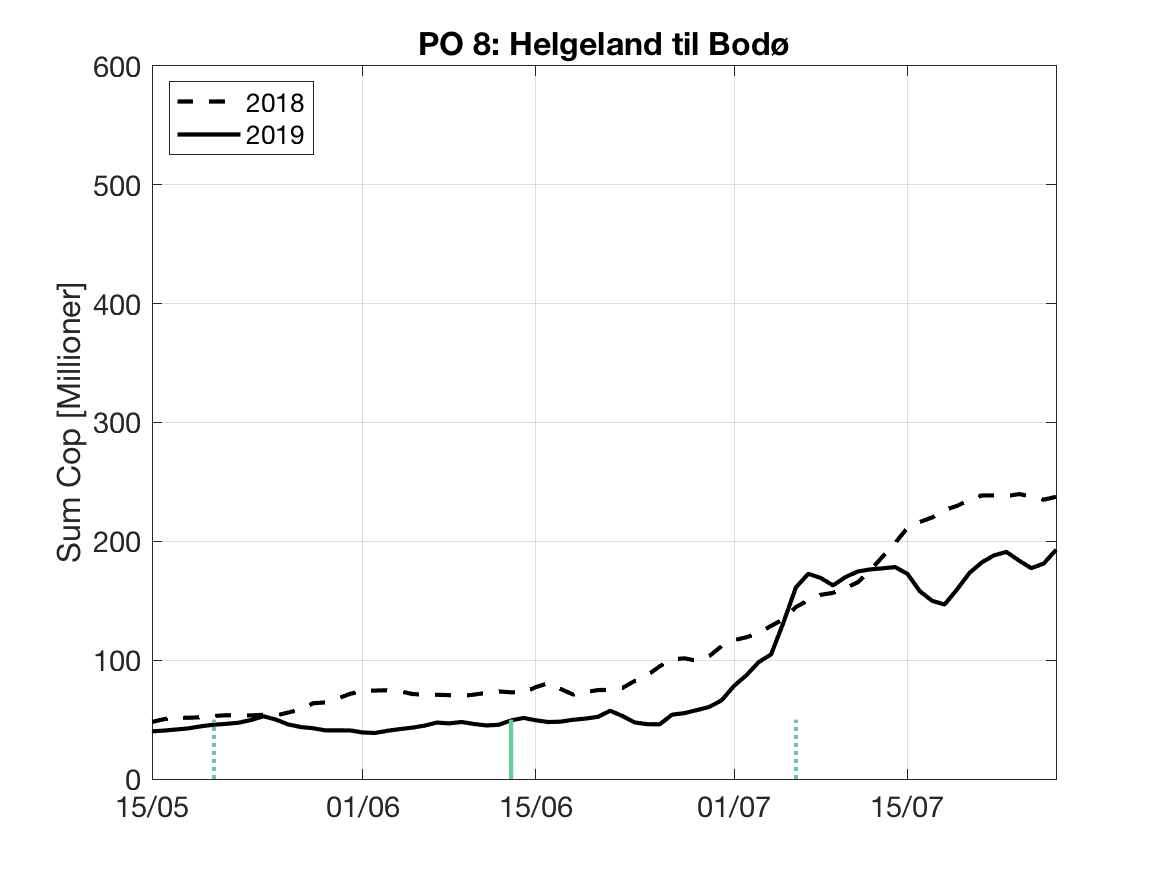
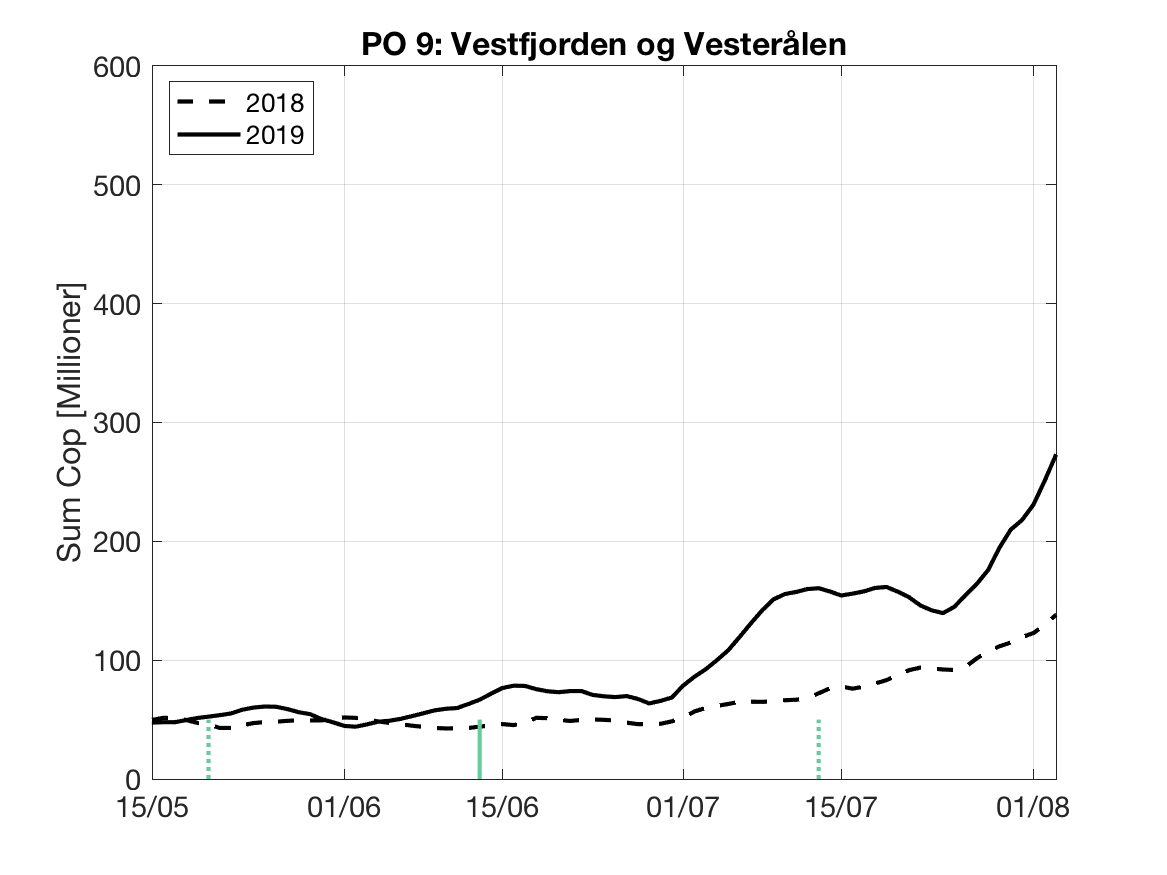
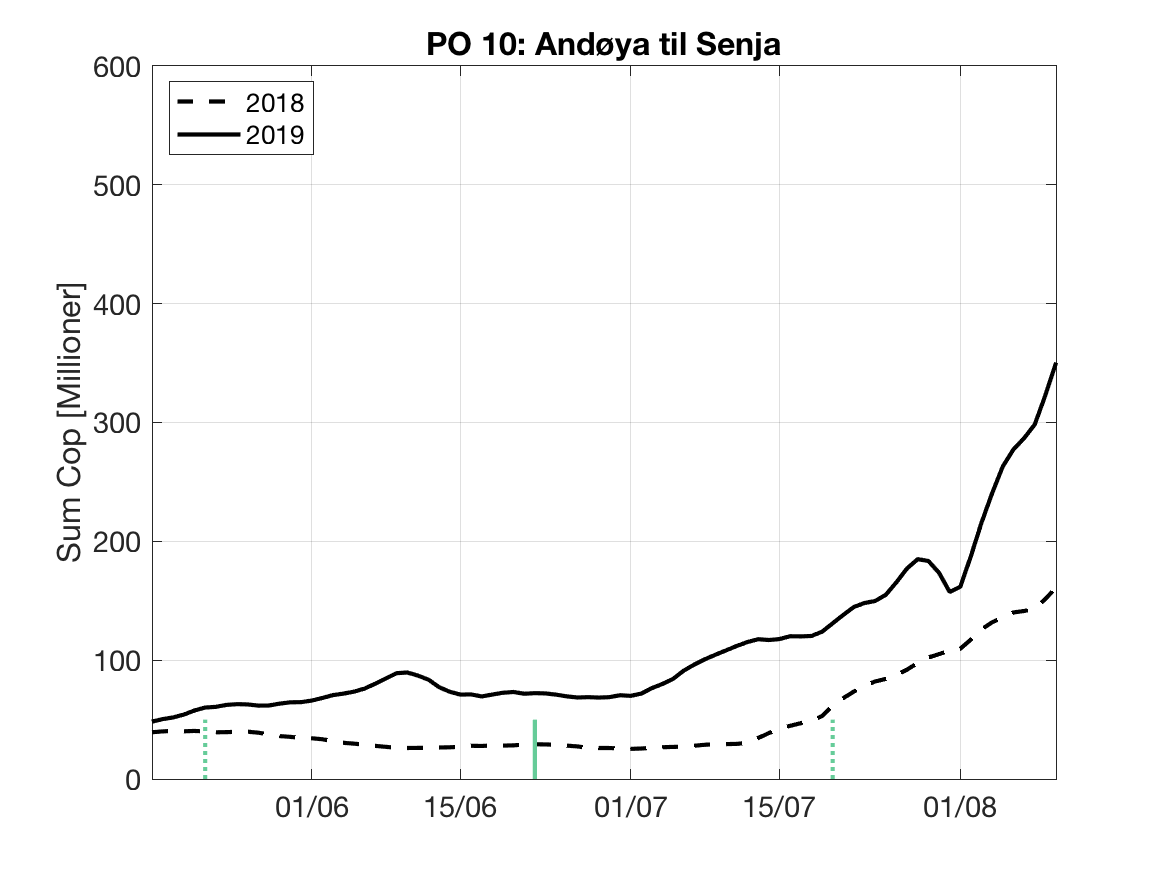
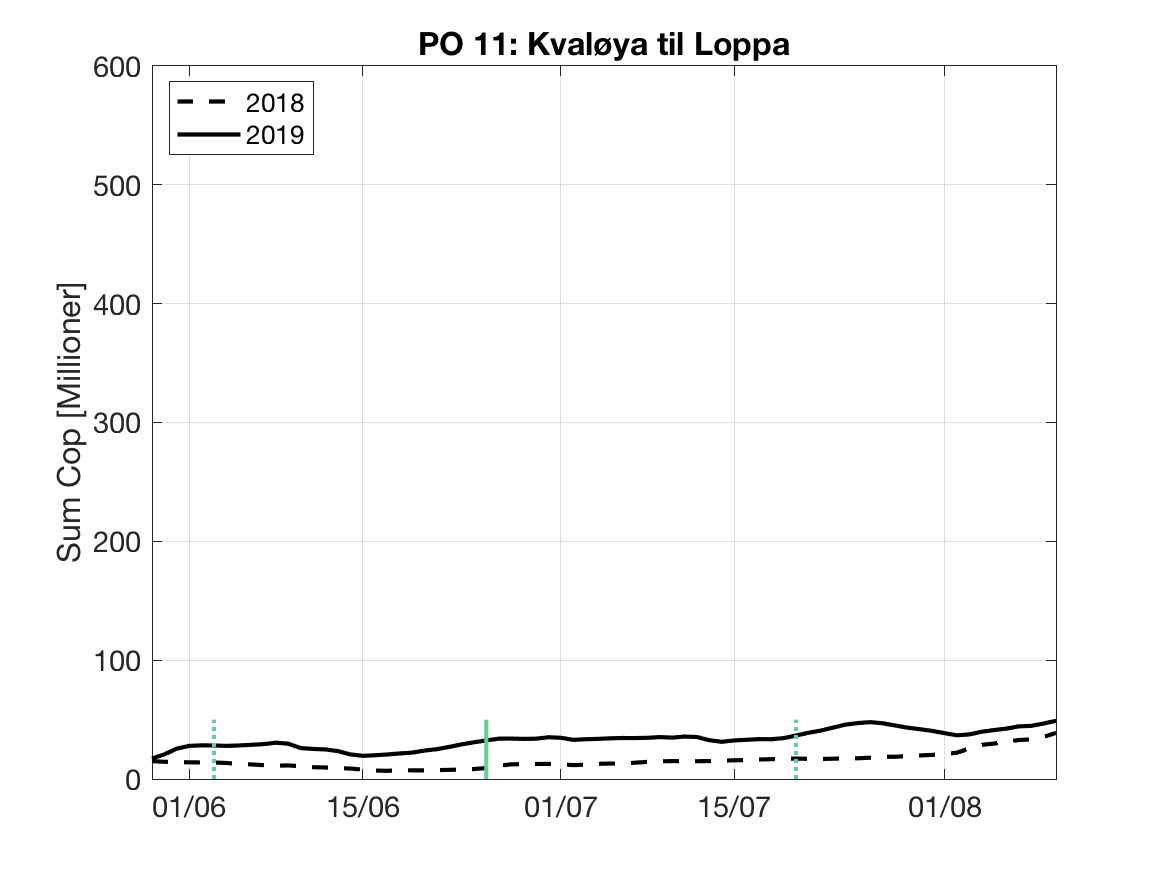
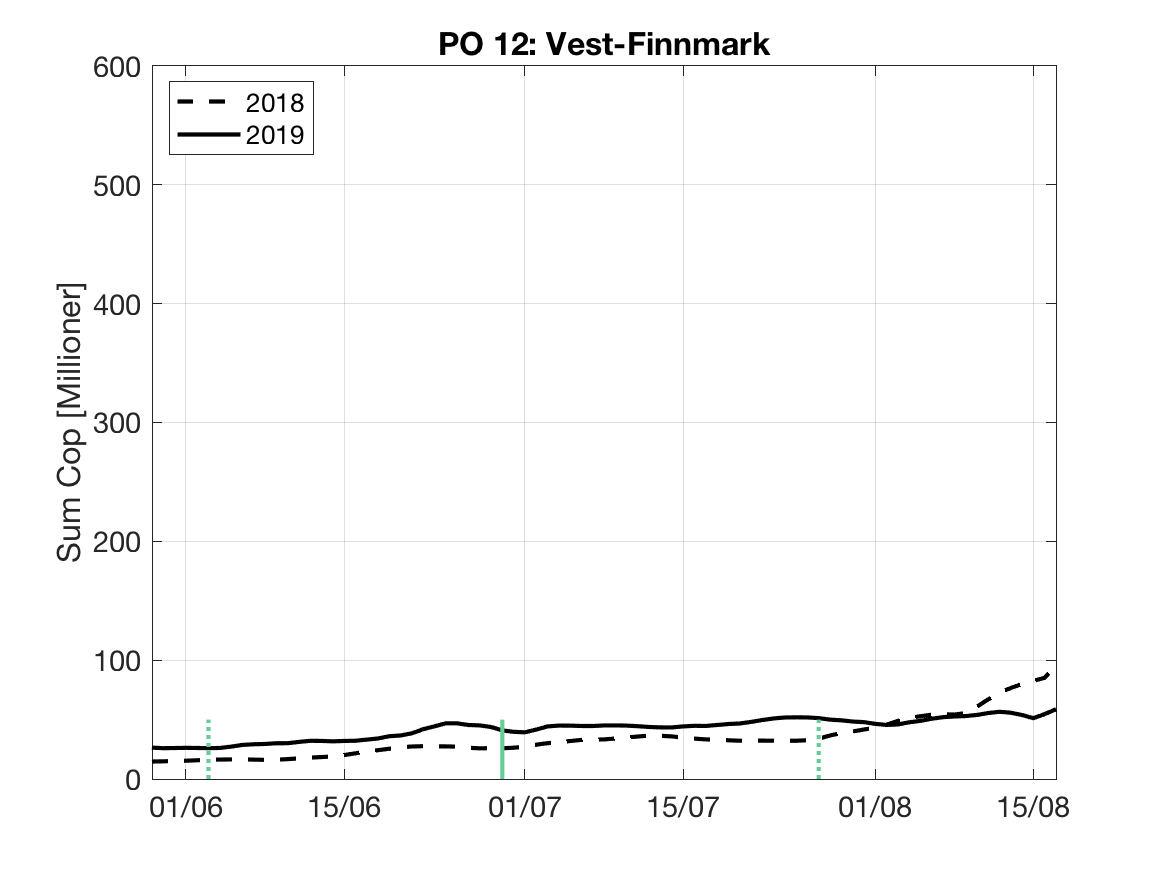
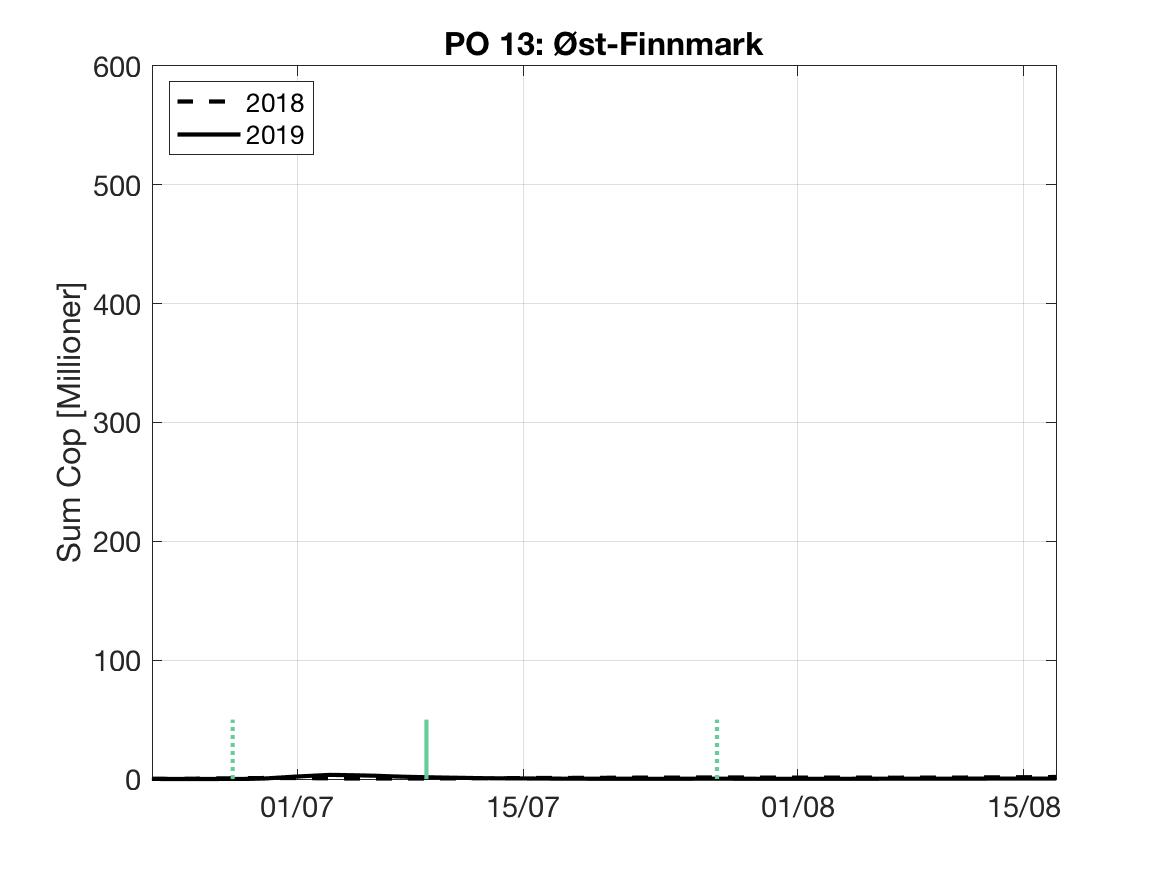
Sluttproduktet fra lakselusmodellen er antallet smittsomme kopepoditter i produksjonsområdene til enhver tid. Summen over hvert produksjonsområde er vist som tidsserier for 2018 og 2019 i figur 3 og som horisontale kart i figur 4.
Gitt at den modellerte fordelingen av smittsomme lakseluskopepoditter i rom og tid er en realistisk beskrivelse av det reelle smittepresset for villfisk, og gitt at sammenhengen mellom smittedose og smitterespons er tilnærmet lineær, kan vi estimere individuell smitte på den villfisken som befinner seg i modellområdet. Vi trenger da å kjenne hvor fisken er, og når den er der, for å sammenholde med den modellerte tettheten av kopepoditter. Med denne bakgrunnen har vi definert to ulike produkter som søker å relatere modellresultatene til dødelighet på villfisk i produksjonsområdene: Smittepresskart og Virtuell smoltutvandring. Disse produktene er beskrevet i Appendiks V og VI sammen med resultater for 2019.
Under følger en kort vurdering av bakgrunnsdataene for Havforskningsinstituttet sine modellprodukt til Trafikklysvurderingen 2019. Formålet er å vise når resultatene vil være mest følsomme for variasjoner i utvandringstidspunkt og forløp, samt gi et best mulig bilde av den horisontale variasjonen i tetthet av smittsomme lakseluslarver innen de ulike produksjonsområdene.
6.1 - Vurdering av tetthet av kopepoditter 2019
PO1: Lav konsentrasjon ved dato for første utvandring, og holder seg relativt stabil gjennom utvandringsperioden.
Tetthetskart: Lave verdier i hele området, bortsett fra en liten antydning til luselarver i området ved Flekkefjord.
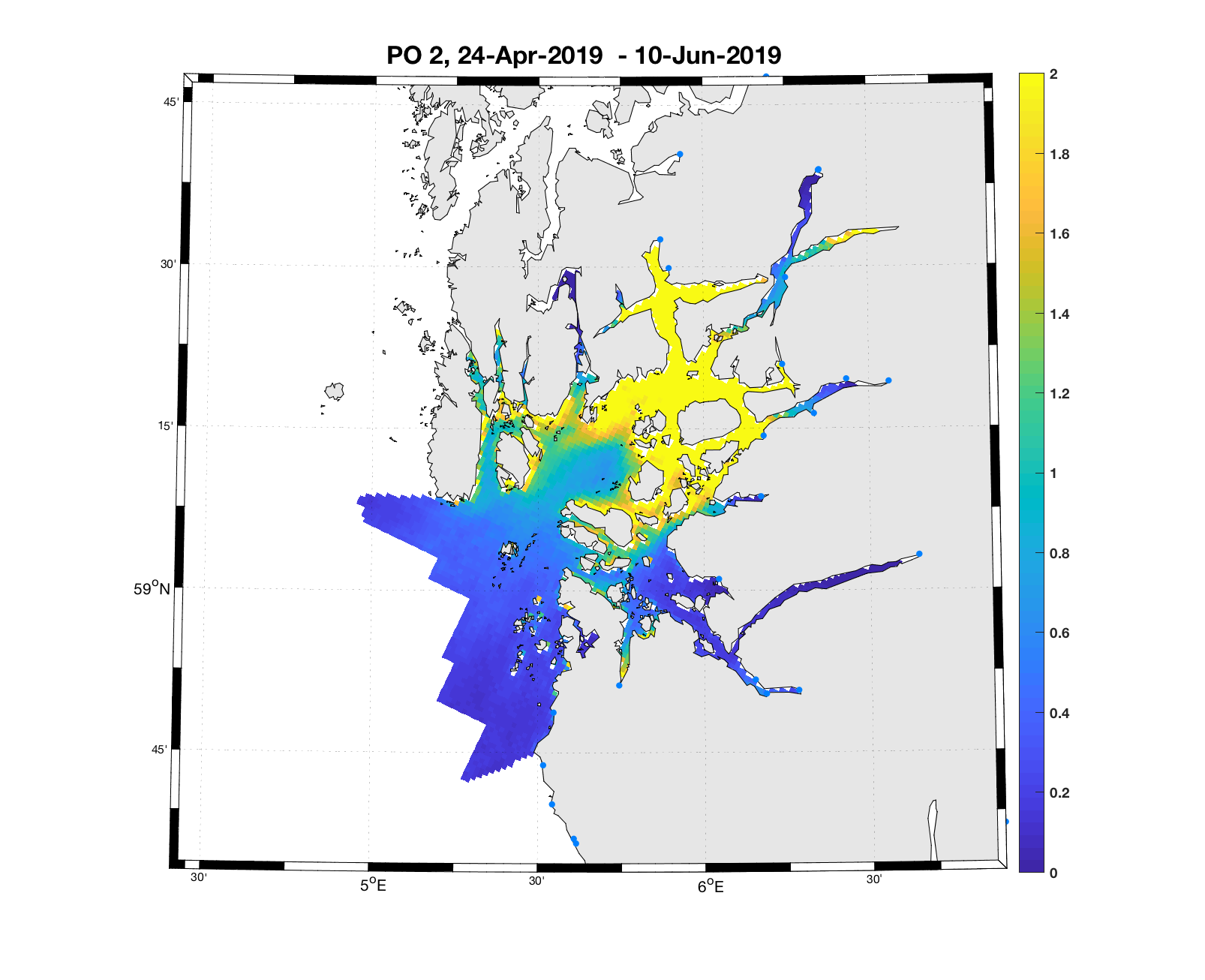
PO2:
Tidsutvikling: Relativ høy verdi ved dato for første utvandring, men holder seg relativt stabil gjennom utvandringsperioden.
Tetthetskart: Summert over den lange utvandringsperioden er det høye konsentrasjoner av lakseluslarver i nord og nordøst, ellers moderate og lave konsentrasjoner. Elvene i sørøst ( Dirdal, Espedal og Frafjord) vil ifølge dette kartet sannsynligvis få på seg små mengder lakselus, mens postsmolt fra elvene i nordøst ( Årdal, Vorma, Ulla og Suldalslågen) sannsynligvis vil få på seg relativt store mengder.
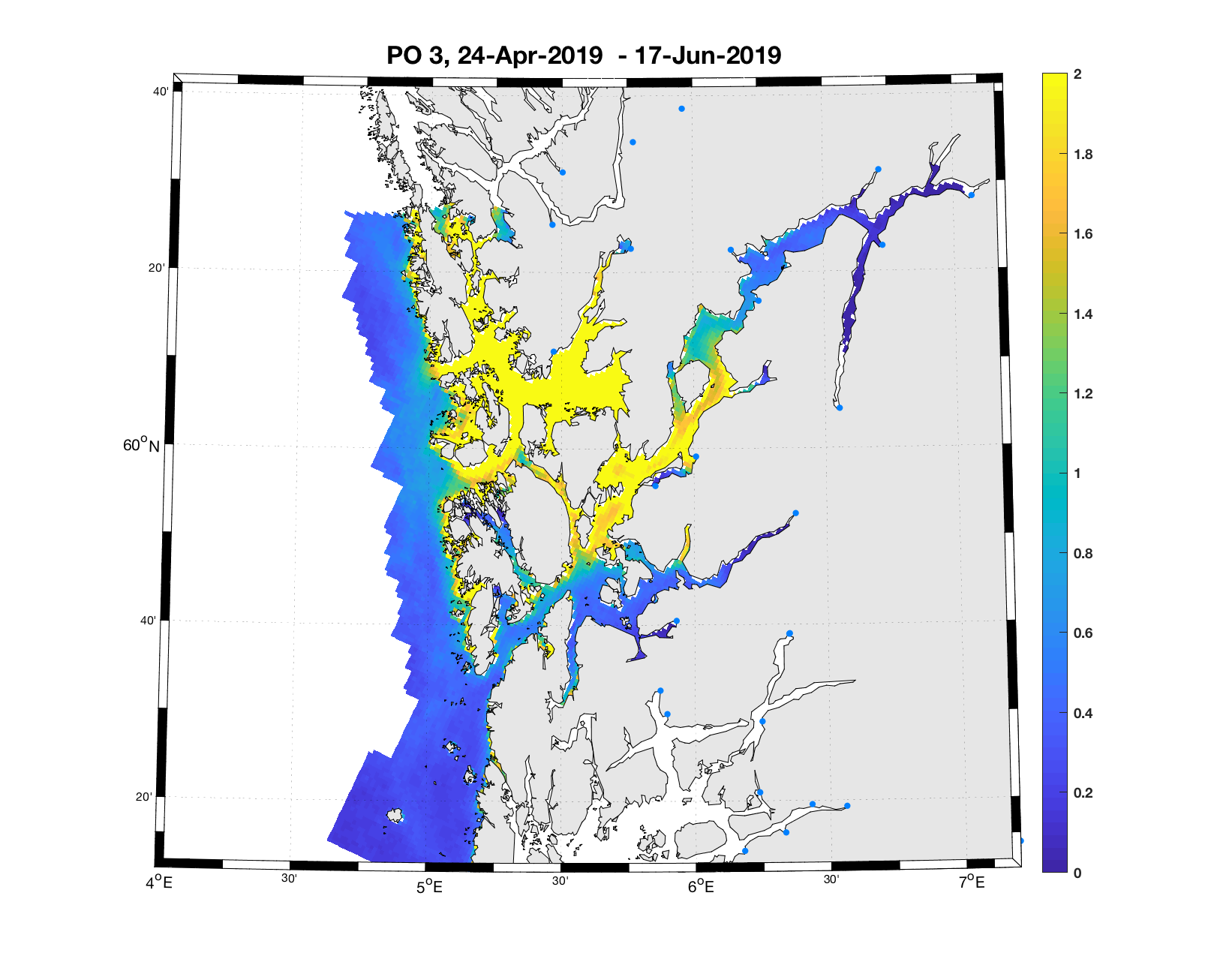
PO3: Tidsutvikling: Relativt høy verdi ved dato for første utvandring, holder seg stabil fram til dato for 50 % utvandring, øker deretter betraktelig.
Tetthetskart: Summert over den lange utvandringsperioden er det høye konsentrasjoner av lakseluslarver i nord (Bjørnafjorden) og i midtre Hardanger, ellers moderate og lave konsentrasjoner. Smolten som vandrer fra Etne vil oppleve svært lave konsentrasjoner i første del av vandringsruten, moderate mengder i andre halvdel, i sum moderate mengder. Postsmolt fra Oselva i Bjørnafjorden vil sannsynligvis få på seg store mengder lakseluslarver, det samme vil smolt fra elvene i indre Hardanger ( Opo, Kinso, Eidfjord og Granvin).
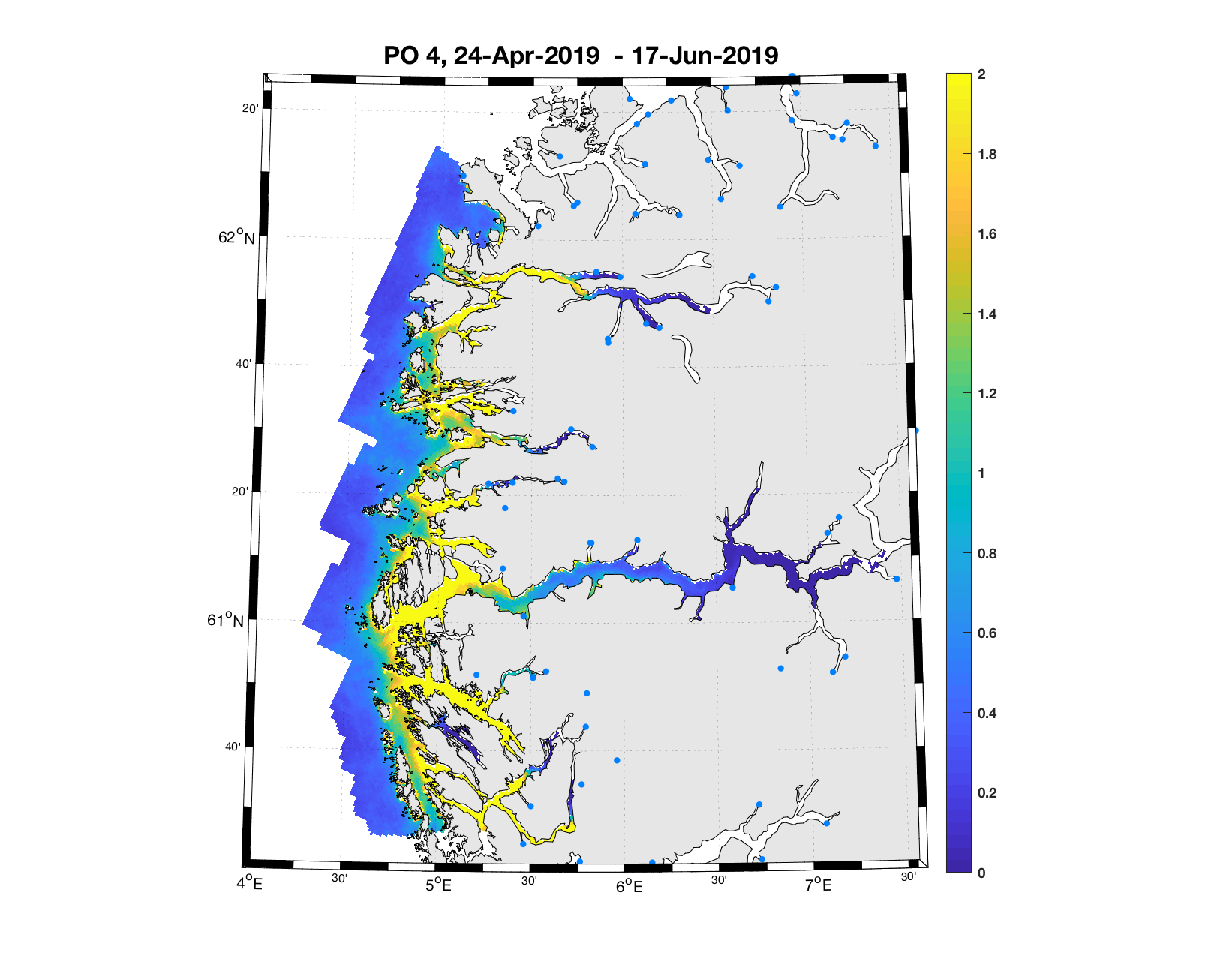
PO4: Tidsutvikling: Relativt høy verdi ved dato for første utvandring, holder seg stabil fram til dato for 50 % utvandring, øker deretter betraktelig.
Tetthetskart: Summert over den lange utvandringsperioden er der høye konsentrasjoner av lakseluslarver i alle fjordmunningene. Smolt fra de fleste elvene må passere gjennom et eller fler områder med høye tettheter av lakseluslarver og dermed sannsynligvis få på seg store mengder lakseluslarver.
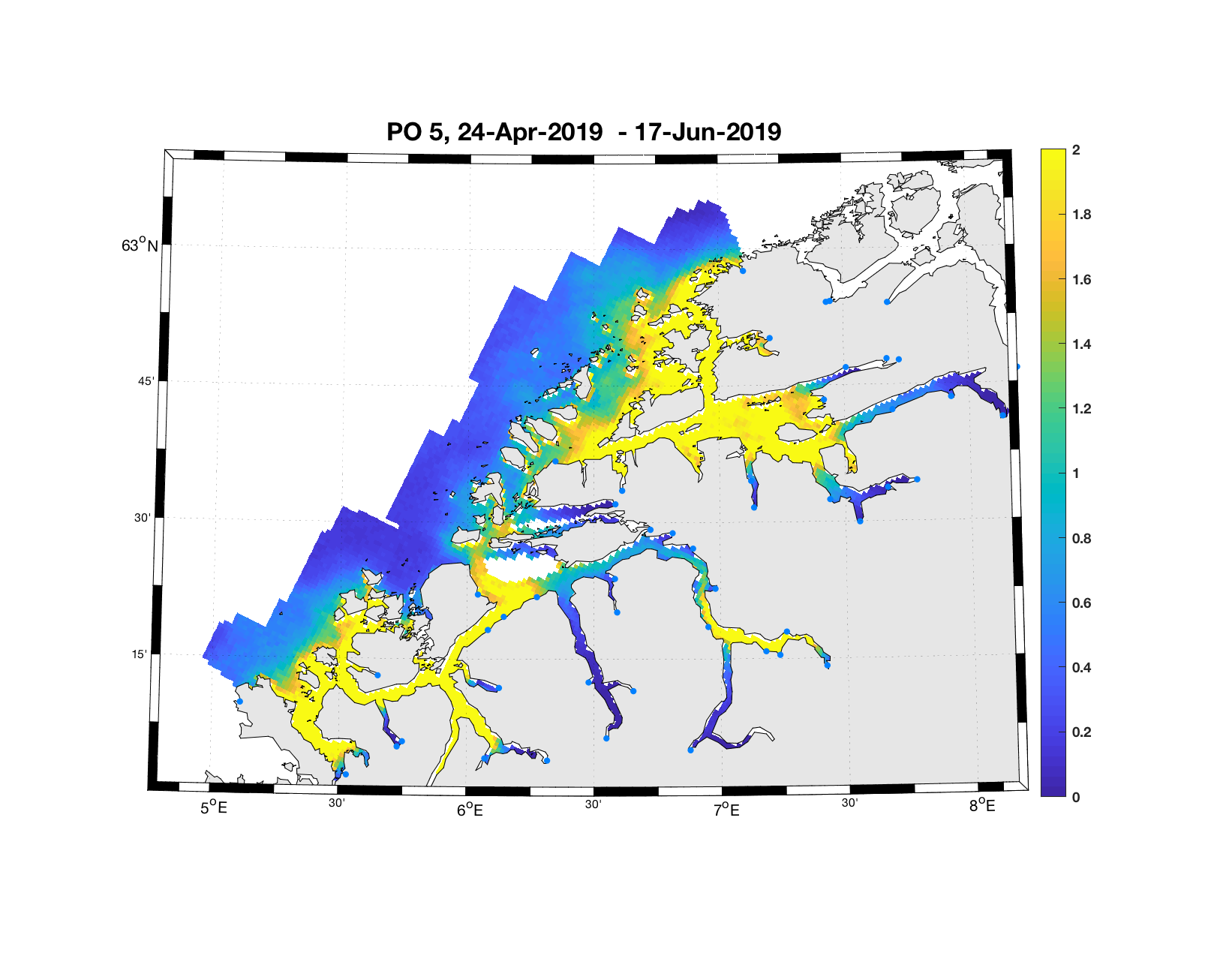
PO5: Tidsutvikling: Moderat verdi ved dato for første utvandring, øker kraftig fram til dato for 50 % utvandring og videre.
Tetthetskart: Summert over den lange utvandringsperioden er der høye konsentrasjoner av lakseluslarver i store deler av området og dermed vil smolt fra både indre Romsdal (Rauma og Eira) og kystnær-midtre (Ørsta) sannsynligvis få på seg store mengder lakseluslarver.
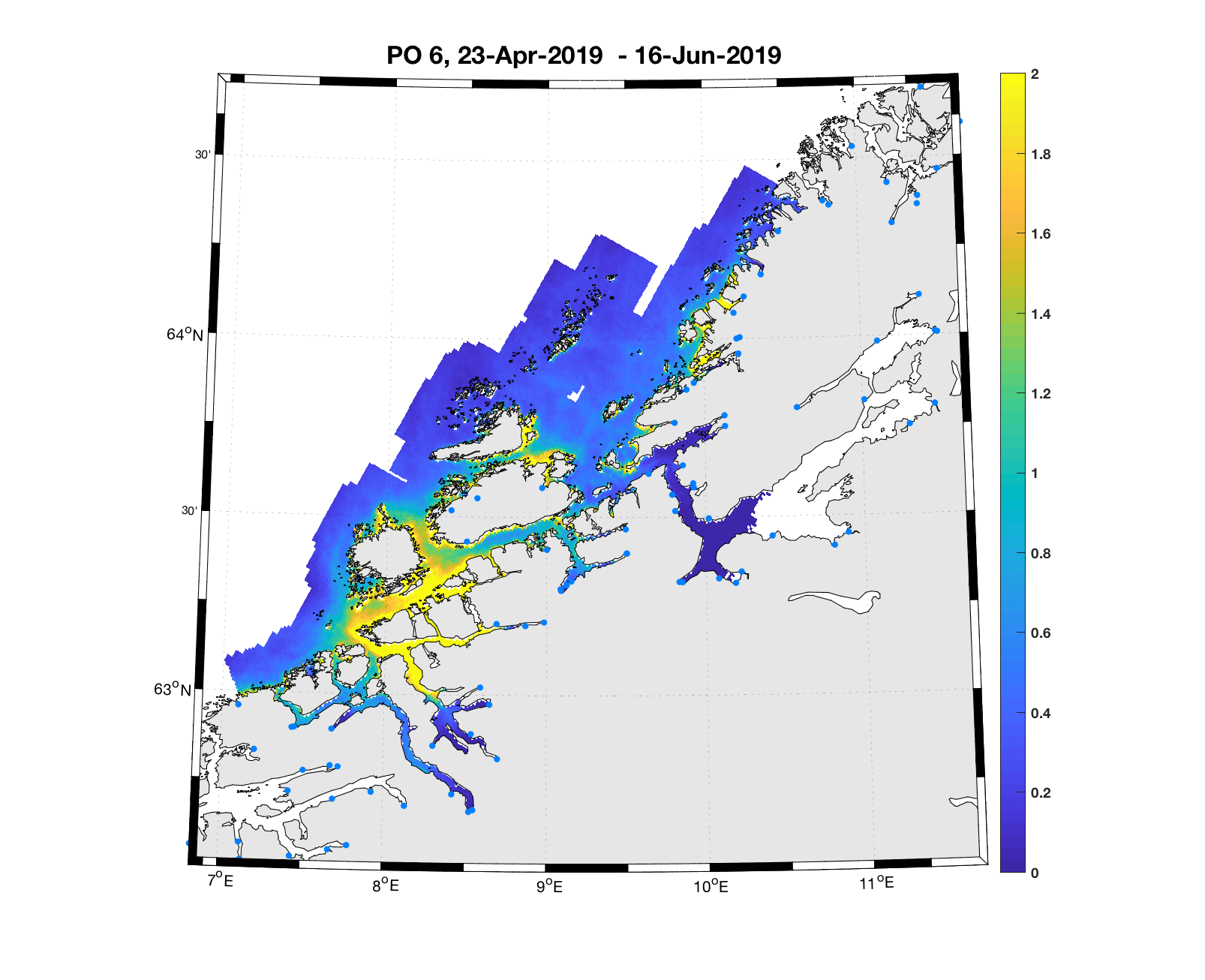
PO6: Tidsutvikling: Relativt høy verdi ved dato for første utvandring, øker moderat fram til dato for 50 % utvandring og videre kraftig.
Tetthetskart: Summert over den lange utvandringsperioden er det høye konsentrasjon av luselarver i sør, mens det er moderate konsentrasjoner i nord. Smolt fra elvene i Trondheimsfjorden ( Orkla, Gaula, Stjørdalselva og Verdalselva) vil sannsynligvis få på seg små mengder med lakseluslarver, mens smolt fra eleven lenger sør (Surna og Driva) sannsynligvis vil få på seg store mengder.
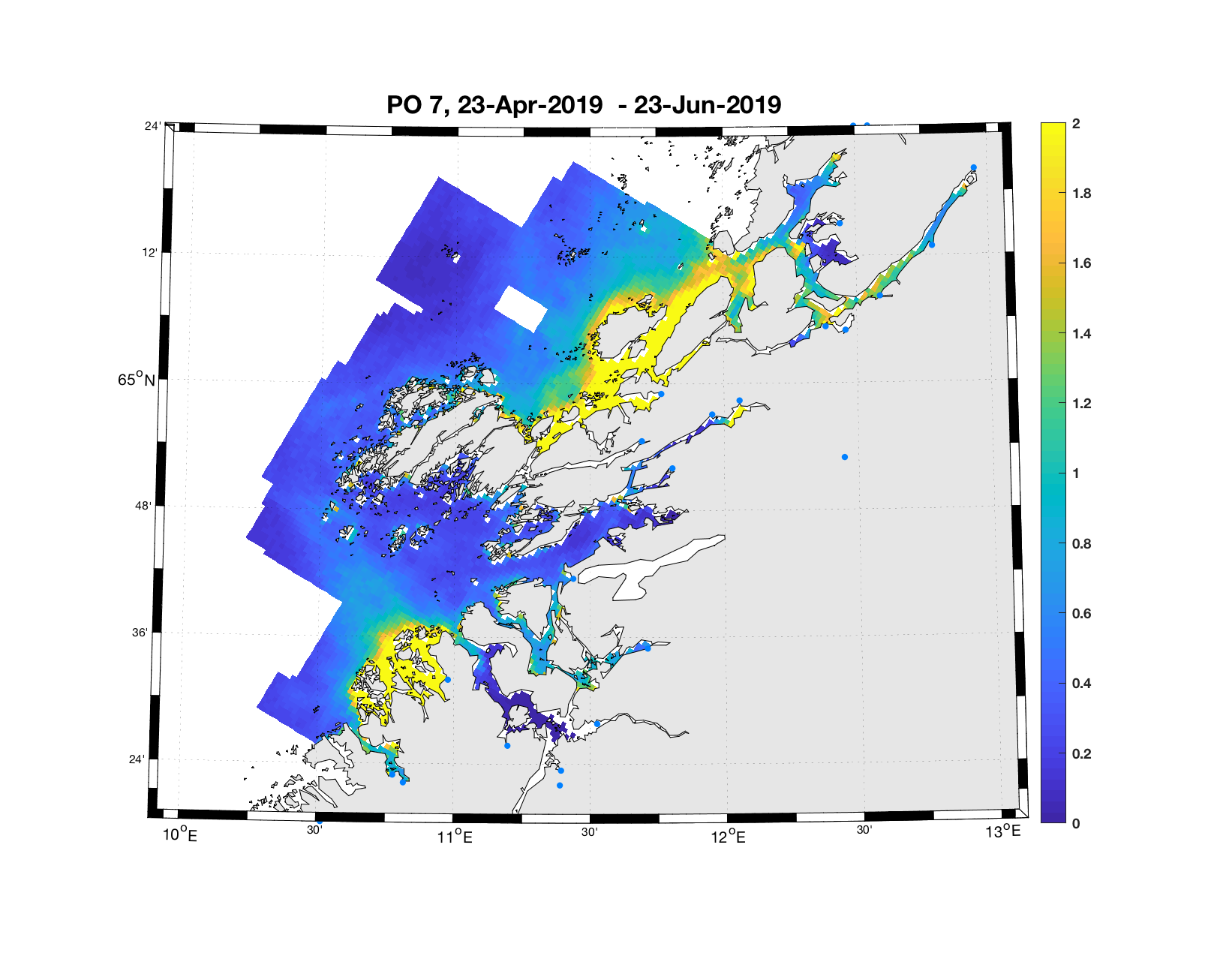
PO7: Tidsutvikling: Moderat verdi ved dato for første utvandring, øker moderat fram til dato for 50 % utvandring og videre kraftig.
Tetthetskart: Summert over den lange utvandringsperioden er der høye konsentrasjon av luselarver i sør og nord. Smolt fra Namsen blir sannsynligvis utsatt for en moderate mengde lakseluslarver.
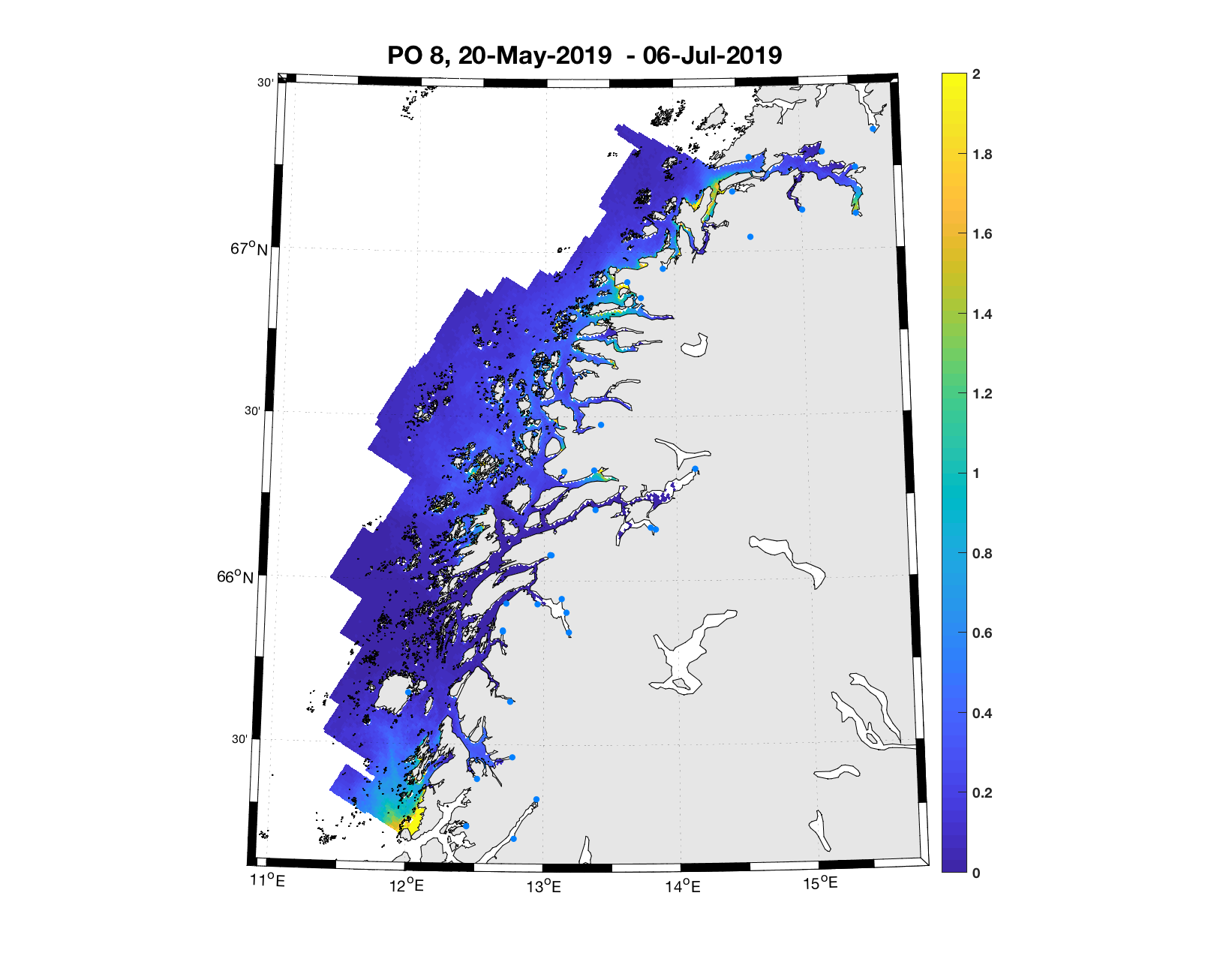
PO8: Tidsutvikling: Moderat verdi ved dato for første utvandring, holder seg stabil fram til dato for 50 % utvandring, øker deretter betraktelig.
Tetthetskart: Summert over den lange utvandringsperioden er der bare små områder med høye konsentrasjoner av lakseluslarver. I den antatte utvandringsruten fra Vefsna er der lave konsentrasjoner og postsmolten vil sannsynligvis få på seg små mengder lakseluslarver.
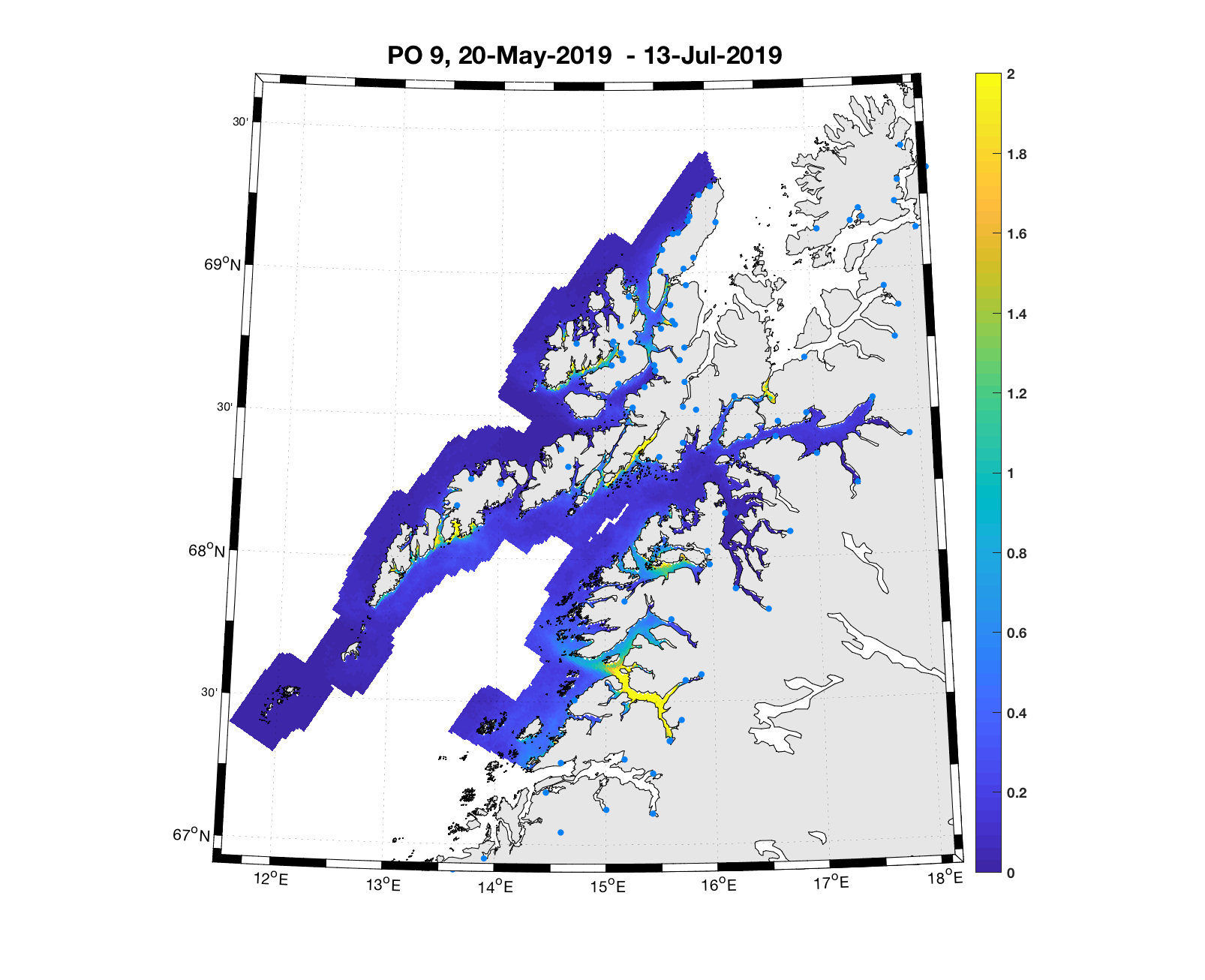
PO9: Tidsutvikling: Moderat verdi ved dato for første utvandring, øker moderat fram til dato for 50 % utvandring og videre kraftig.
Tetthetskart: Summert over den lange utvandringsperioden er der bare noen få områder med høye konsentrasjoner av lakseluslarver. Smolt fra elvene i Sør-Folda vil sannsynligvis få på seg store mengder lakseluslarver, smolt fra de andre eleven moderate eller små mengder.
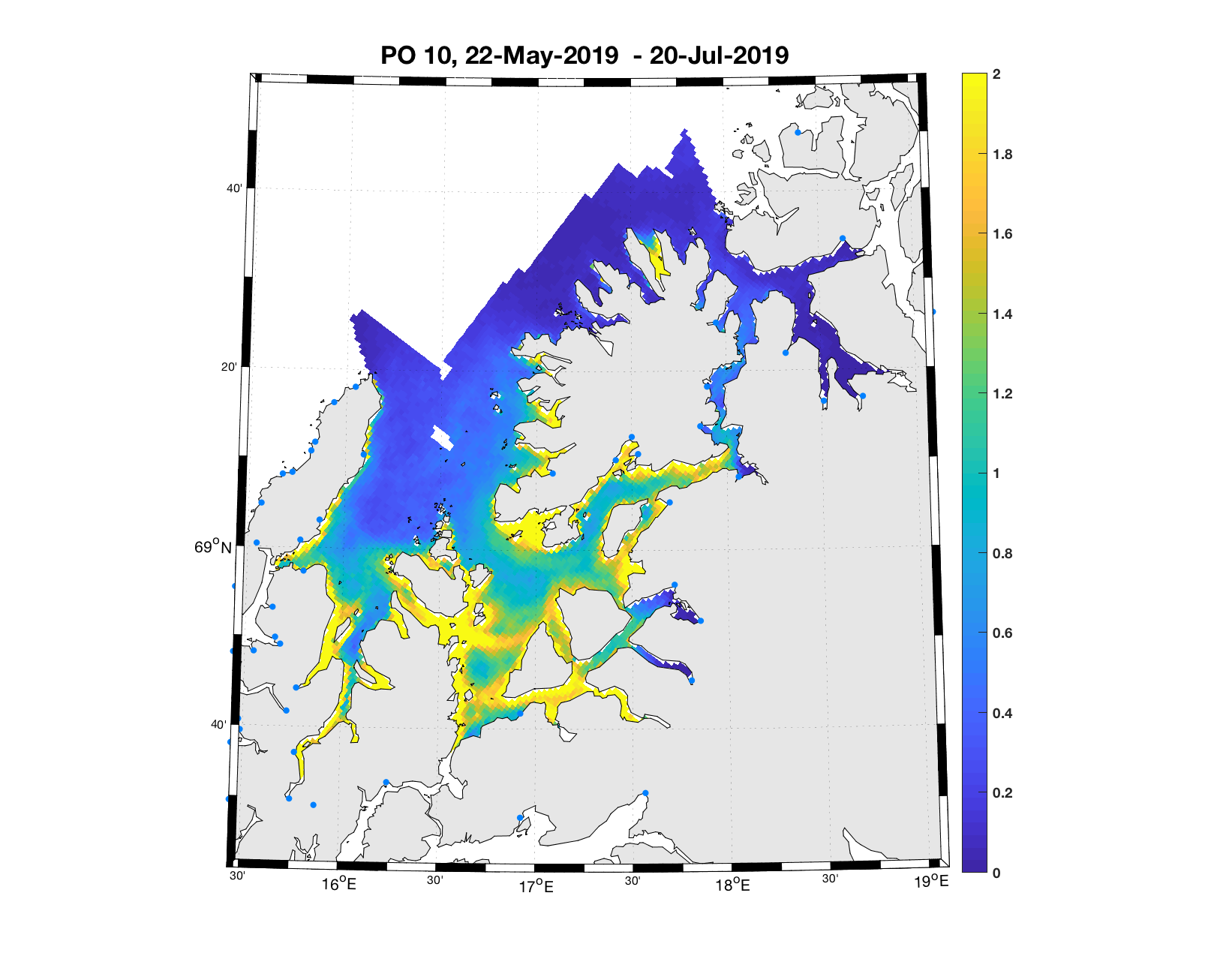
PO10: Tidsutvikling: Moderat verdi ved dato for første utvandring, øker moderat fram til dato for 50 % utvandring og videre kraftig.
Tetthetskart: Summert over den lange utvandringsperioden er der store områder med moderate til høye konsentrasjoner av lakseluslarver. Postsmolt fra de fleste elvene vil sannsynligvis få på seg moderate mengder lakseluslarver.
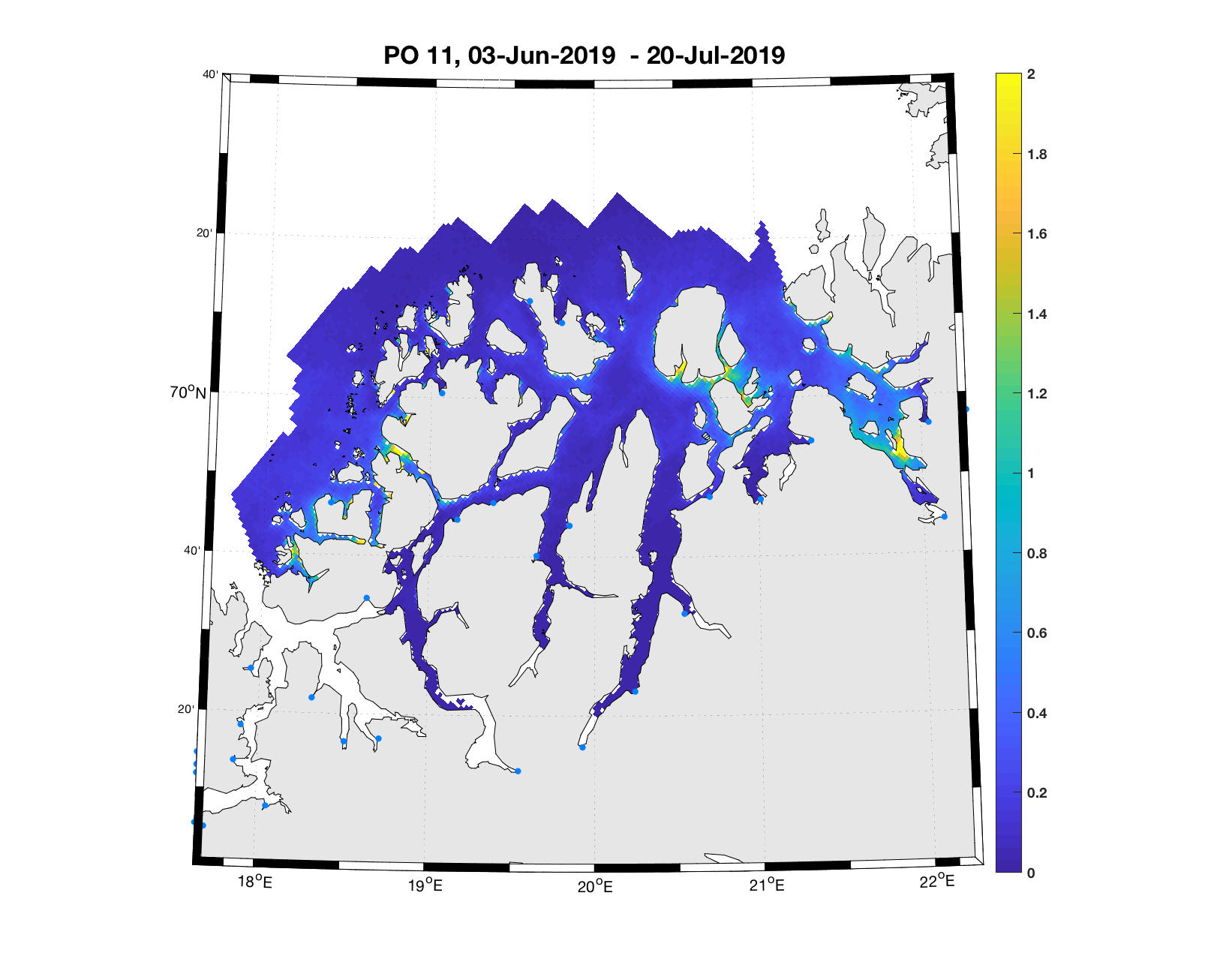
PO11: Tidsutvikling: Moderat til lav verdi ved dato for første utvandring, holder seg stabil fram til dato for 50 % utvandring, øker deretter moderat.
Tetthetskart: Summert over den lange utvandringsperioden er der bare noen få områder med høye konsentrasjoner av lakseluslarver. Postsmolt fra alle elvene vil sannsynligvis få på seg små mengder lakseluslarver.
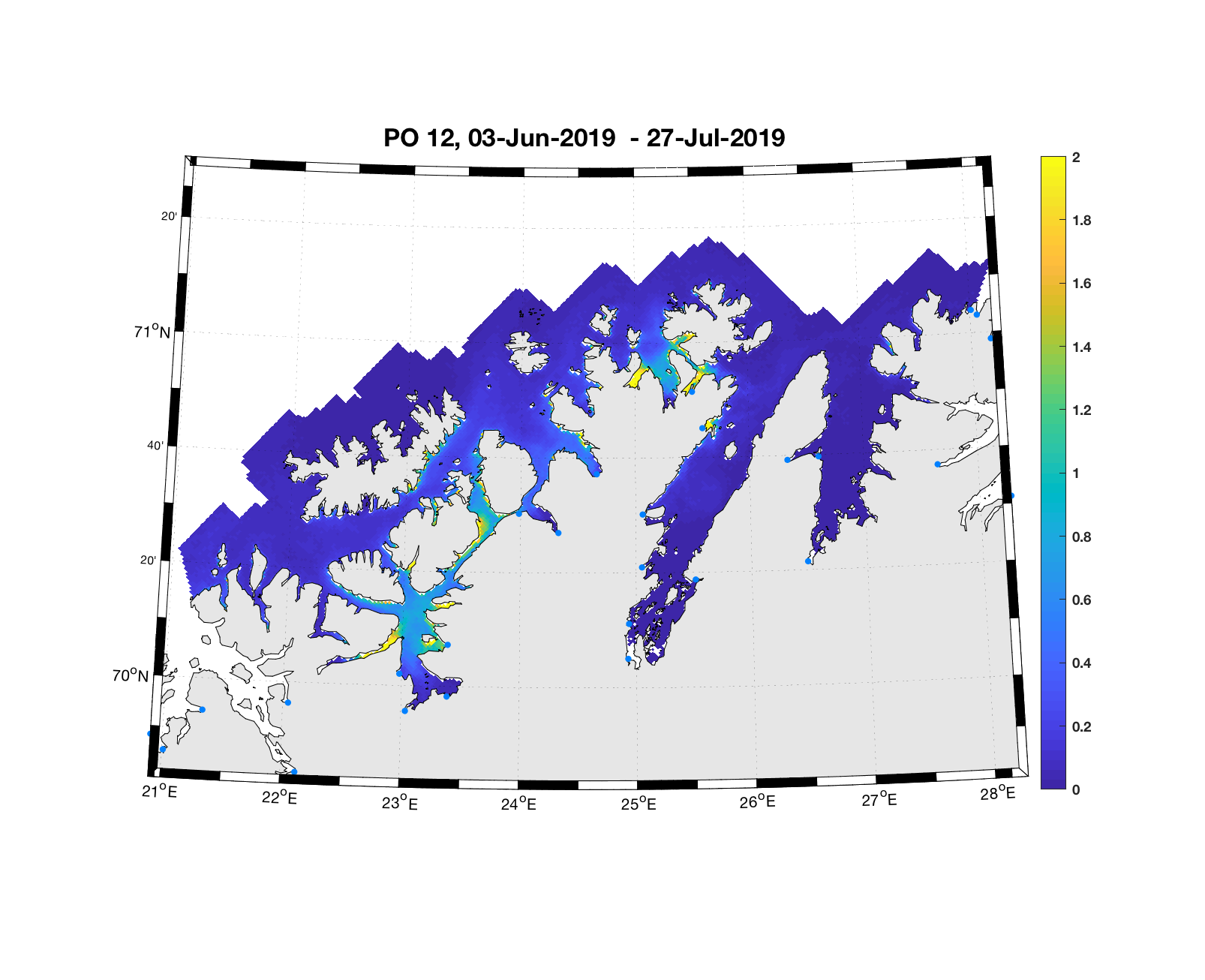
PO12: Tidsutvikling: Moderat til lav verdi ved dato for første utvandring, øker jevnt og betydelig fram til dato for 50 % utvandring og videre.
Tetthetskart: Summert over den lange utvandringsperioden er der bare noen områder i sør (Altafjorden) med moderate til høye konsentrasjoner av lakseluslarver. Smolt som vandre fra Altaelva vil sannsynligvis få på seg moderate mengder med lakseluslarver. Postsmolt fra elvene lenger øst vil sannsynligvis få på seg små mengder.
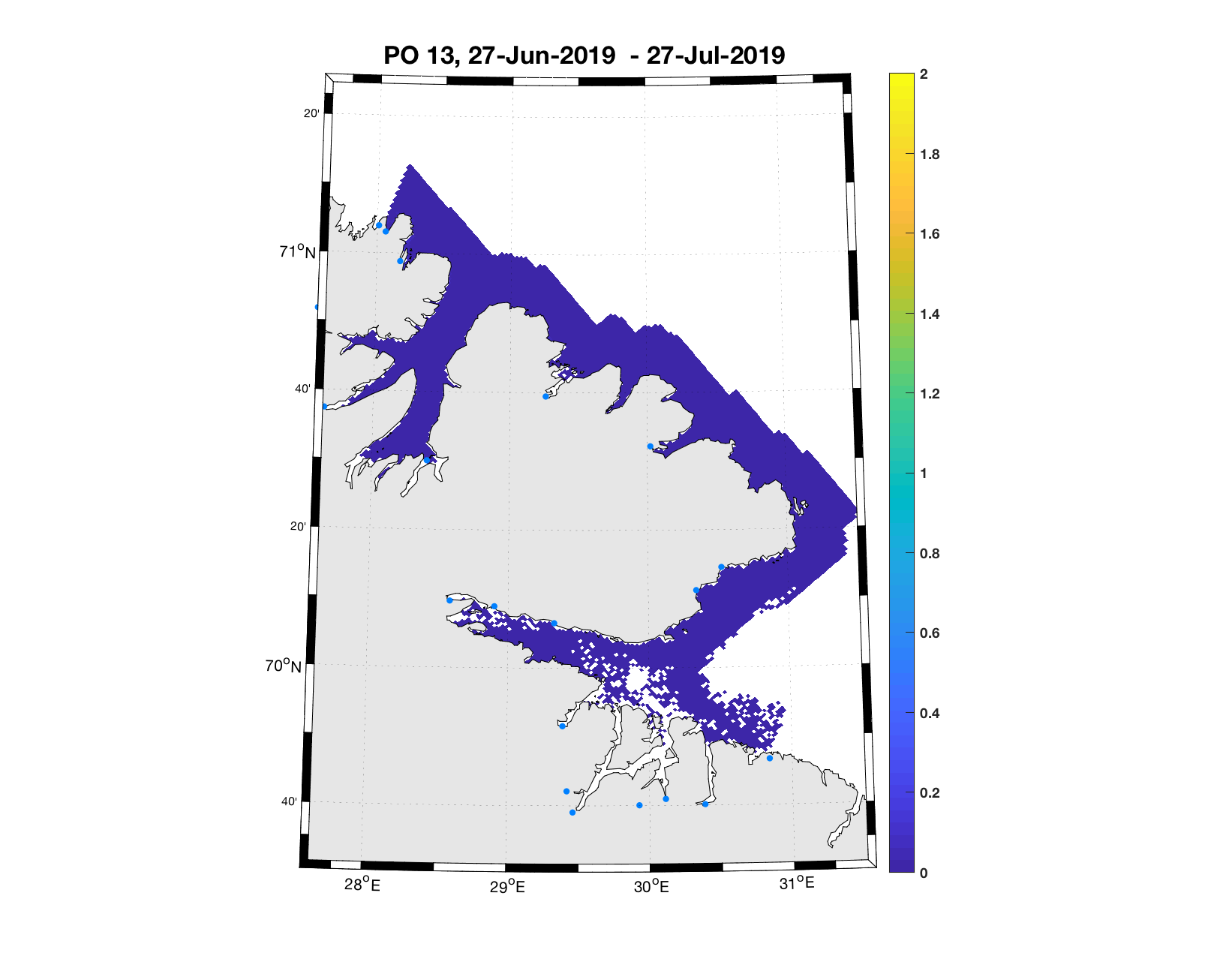
PO13: Tidsutvikling: Tetthetskart: Summert over den lange utvandringsperioden er der lave konsentrasjoner av lakseluslarver i hele området.
6.2 - Tidsutvikling av kopepoditter innen produksjonsområdene
Endringen i tetthet av kopepoditter med tiden er vist i figur 3. Tabell 1 gir en oppsummert der dato for første utvandring er benyttet som referansedato, og summen av smittsomme lakseluslarver ved 50 % utvandring på PO-nivå, siste dag for utvandring og 14 dager etter siste utvandring er gitt relativt til denne.
Figur 3. Tidsutviklingen av antall smittsomme kopepoditter i hvert produksjonsområde for 2018 og 2019, beregnet med den hydrodynamiske spredningsmodellen for lakselus.
| | T1 (Millioner) | T2/T1 | T3/T1 | T4/T1 |
| PO1 | 6.3 | 2.1 | 2.2 | 2.5 |
| PO2 | 75.0 | 1.1 | 0.9 | 1.4 |
| PO3 | 145.5 | 1.1 | 1.8 | 3.9 |
| PO4 | 157.4 | 1.0 | 2.1 | 3.5 |
| PO5 | 70.5 | 1.9 | 3.5 | 5.0 |
| PO6 | 90.0 | 1.3 | 2.6 | 4.2 |
| PO7 | 45.3 | 1.4 | 3.8 | 7.6 |
| PO8 | 45.7 | 1.1 | 3.5 | 4.2 |
| PO9 | 52.5 | 1.3 | 3.0 | 5.2 |
| PO10 | 60.1 | 1.2 | 2.2 | 5.9 |
| PO11 | 28.2 | 1.1 | 1.3 | 1.7 |
| PO12 | 26.0 | 1.6 | 2.0 | 2.3 |
| PO13 | - | - | - | - |
Tabell 1. Relative tall (relativt til referansedato, T1) for summen av kopepoditter innen hvert PO. T1 er antall kopepoditter ved dato for første utvandring i POet. T2 er antall kopepoditter ved midlere dato for 50 % utvandring på PO-nivå. T3 er antall kopepoditter ved dato for siste utvandring i POet. T4 er antall kopepoditter ved dato for siste utvandring i POet +14 dager. En fullstendig liste over utvandringstidspunkt kan finnes i Appendiks I.
6.3 - Geografisk fordeling av kopepoditter
Sluttproduktet fra lakselusmodellen er antallet smittsomme kopepoditter i produksjonsområdene til enhver tid. Med de variasjonene som er vist i figur 3 og tabell 1 er det egentlig umulig å lage kun ett kart med horisontal fordeling som er representativ for hele utvandringsperioden. Vi har tidligere vist kart der kopepodittene har blitt summert over 30 dager rundt den datoen det er estimert at 50 % av smolten vandrer fra elv til hav. For de produksjonsområdene der mengden kopepoditter endrer seg betydelig i løpet av utvandringsperioden blir dette et dårlig mål. Vi viser derfor i figur 4 summen over hele utvandringsperioden. Tiden det tar en postsmolt å vandre fra elven til havet vil variere mye mellom de ulike elvene. Vi har derfor i figur 4 valgt å utelate denne siste perioden (fra siste utvandringsdag til siste fisk har kommet seg til havet).
Figur 4. Tetthet av smittsomme kopepoditter mellom overflaten og 2m dyp, 2019. Figurene viser antall per kvadratmeter., Beregningen er gjort med den hydrodynamiske spredningsmodellen for lakselus. Skalaen går til 2, i de gule områdene kan verdiene være betydelig høyere.
7 - Betydning av gitteroppløsning
I tillegg til ukentlige simuleringer av lakselusspredning med 800m horisontal gitteroppløsning (NorKyst800), gjøres det en rekke modellsimuleringer der strømmodellen benytter 160m gitteroppløsning (NorFjords160). Med likt vertikalt gitter er likevel disse simuleringene nesten 10 ganger så tidkrevende som simuleringene med 800m gitteroppløsning. Det er opplagt at 5 ganger finere gitter vil beskrive topografien og strømforholdene i trangere farvann og fjorder bedre enn hva en kan med 800m gitteroppløsning.
Vi ønsker å finne ut om 800m gitteroppløsning for miljøinformasjon er tilstrekkelig til formålet som er å karakterisere smittepress fra lakselus på villfisk i de 13 produksjonsområdene langs kysten, eller om høyere romlig oppløsning av miljøforholdene er nødvendig i noen av, eller alle, områdene. I det følgende presenteres noen foreløpige resultater, men flere simuleringer og sammenligninger må gjøres for å konkludere.
Spredningssimuleringene av lakselus er gjennomført med identiske kildeledd (antall lakselusnauplier som slippes fra et oppdrettsanlegg) og lik metodikk for lakselusspredning, men med miljøinformasjon fra henholdsvis strømmodellene NorKyst800 og NorFjords160. Den eneste forskjellen mellom resultatene er altså detaljnivået på de fysiske feltene (strøm, saltholdighet, temperatur) pga. ulik horisontal gitteroppløsning. Vertikalgitteret er likt for både NorKyst800 og NorFjords160. Vi forventer en viss forskjell i fordelingen av kopepoditter i rom og tid, og at det er større ulikheter i områder der det er trangere farvann, med smale sund og fjordarmer.
Simuleringene starter 2. mars 2019 for begge modellene og avsluttes på dag 121 som er 30. juni. Resultatene er presentert som døgnmiddel av tetthet, uttrykt som antall kopepoditter pr. m² mellom overflaten og 2m dyp.
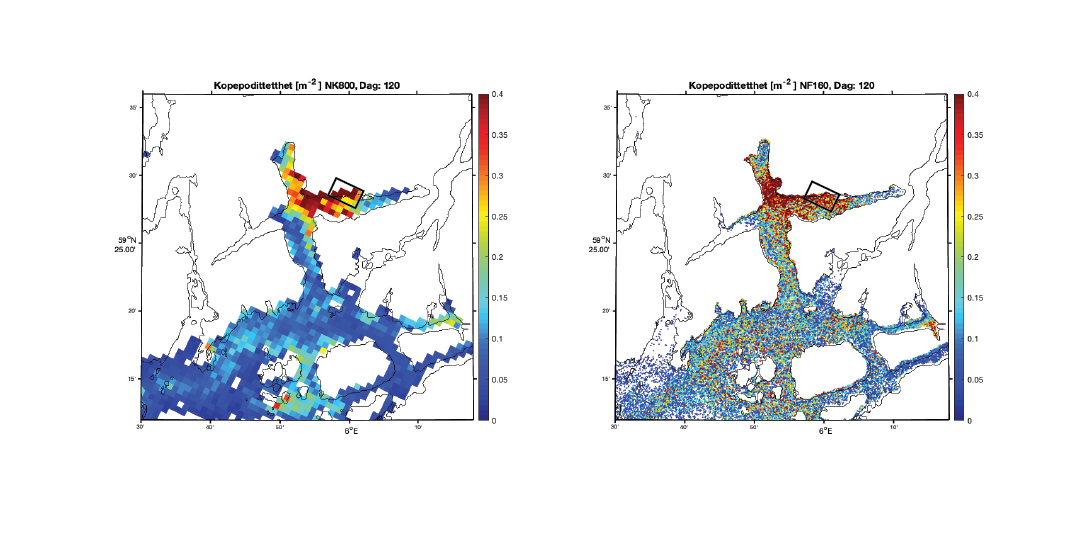
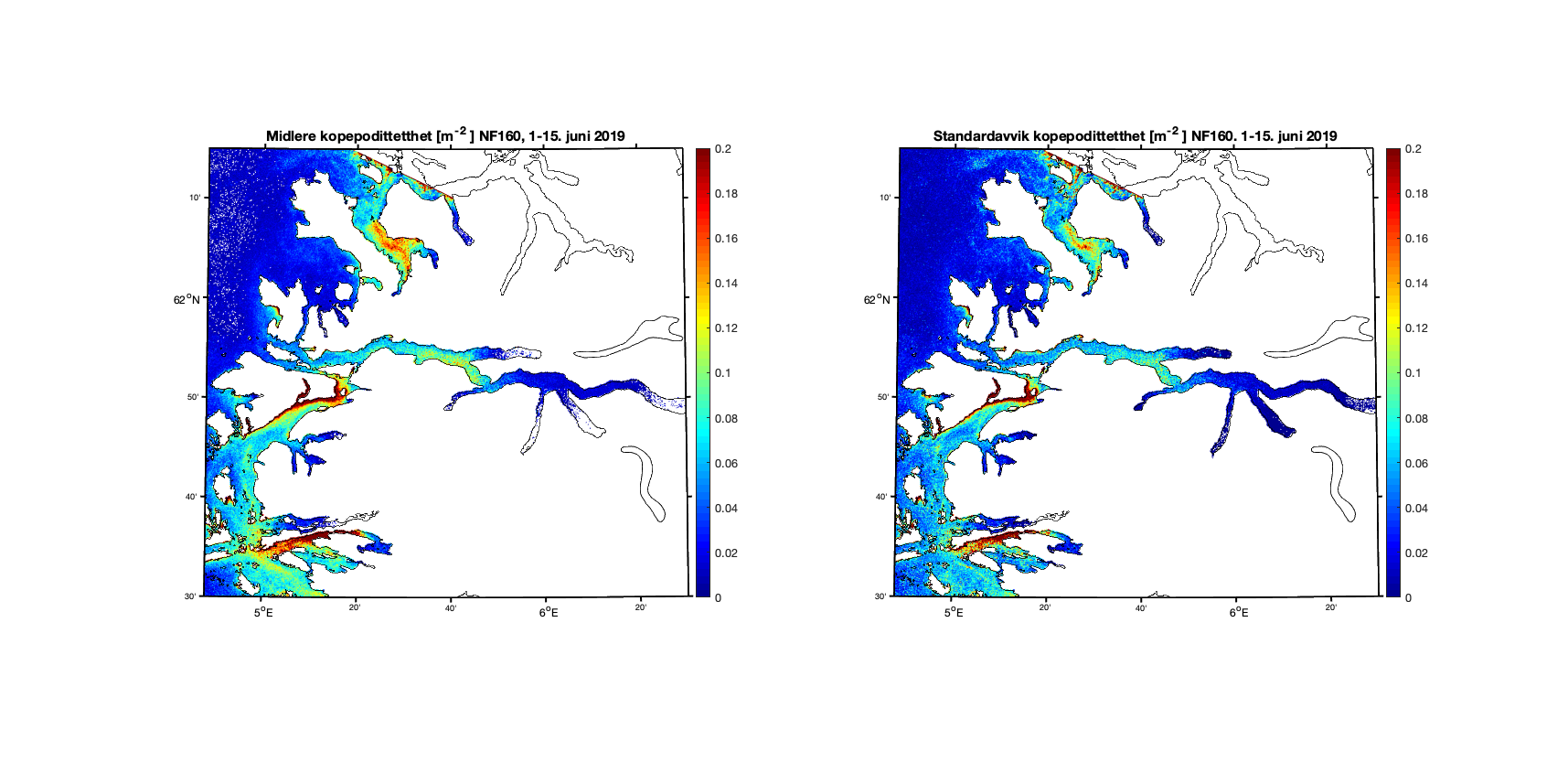
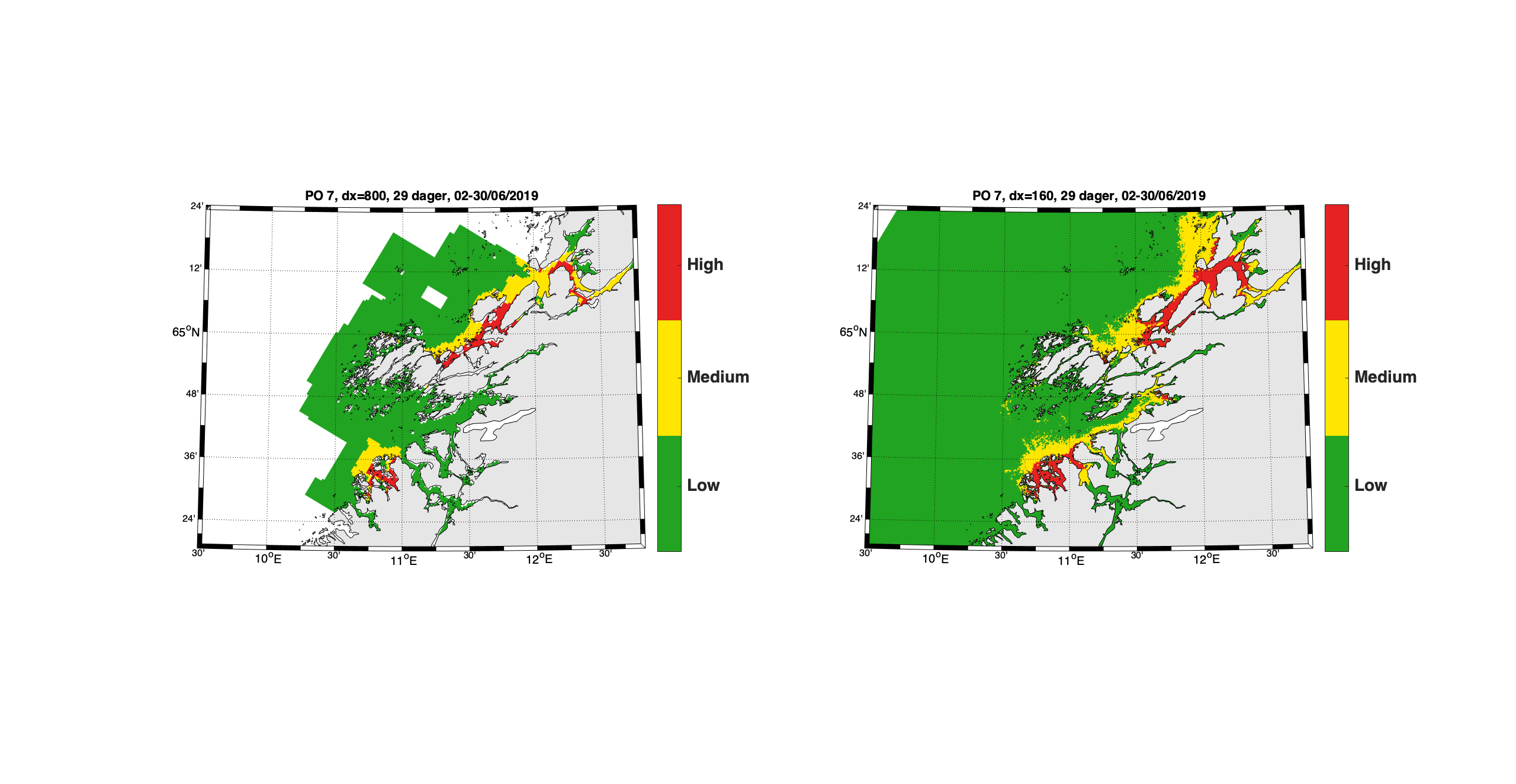
Et grunnleggende eksempel på de ulikhetene som finnes mellom resultatene, er fra Vindafjorden i Rogaland (Figur 5). Begge modellene indikerer en økt konsentrasjon langs nordsiden av fjorden med verdi mellom 0.5 og 1 kopepoditt pr. m² på dag 120 som er 29. juni 2019.
Figur 5. Konsentrasjon av kopepoditter den 29. juni 2019 beregnet med 800m-modellen (venstre) og 160m-modellen (høyre). Den svarte firkanten markerer et areal vi viser tidsserier fra i Figur 2 og 3 under.
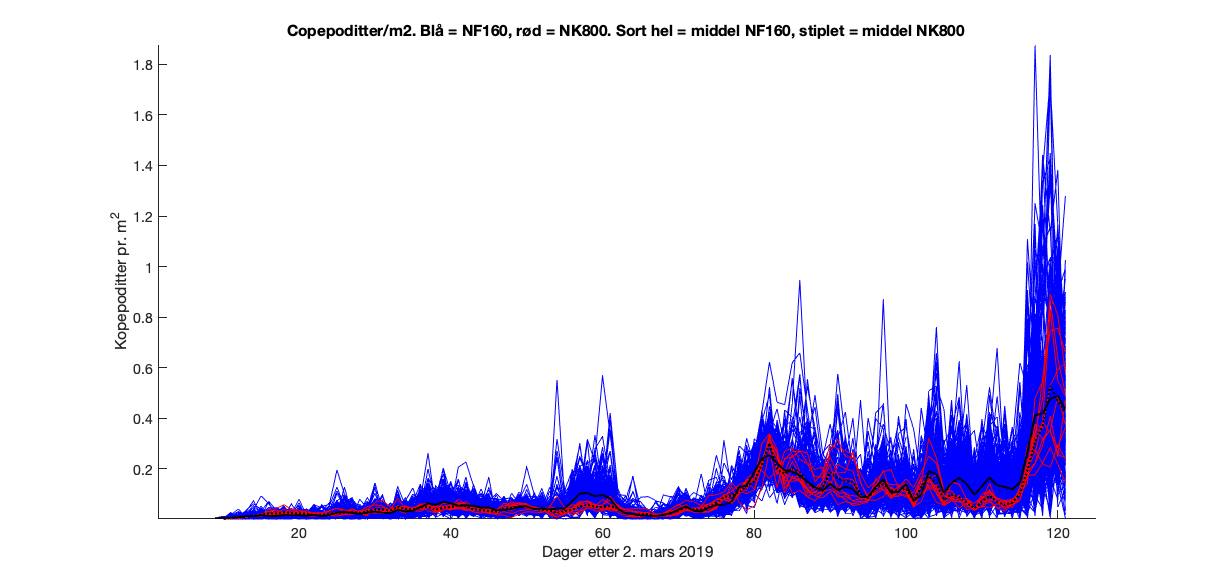
Ser vi nærmere på antall kopepoditter inne i den svarte firkanten markert i Figur 5, finner vi at det er en del variasjon mellom gitterrutene. I 800m-gitteret er det 11 ruter som ikke er landpunkt mens i 160m-gitteret er det 240 ruter. Tidsserier av verdiene i alle rutene viser at det er betydelig høyere verdier i enkelte av 160m-gitterrutene enn i 800m-rutene (Figur 6).
Figur 6. Tidsserie av kopepodittkonsentrasjon i gitterrutene for 800m-modellen (rød) og 160m-modellen (blå) i det markerte arealet i Vindafjorden.
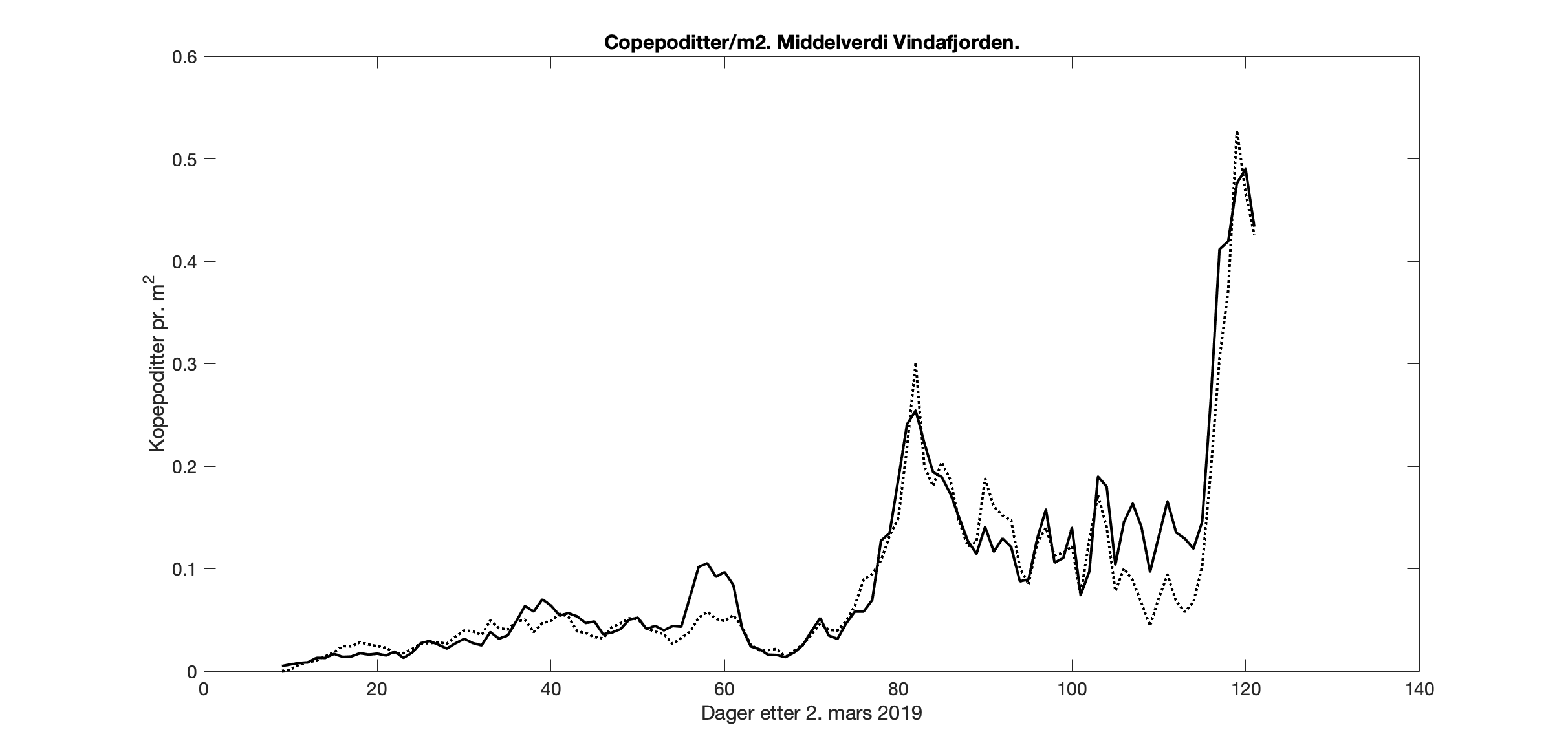
Beregner vi den romlige middelverdien er forskjellen mellom de to simuleringene liten, med bare kortere perioder med noe avvik (Figur 7).
Figur 7. Middelverdi av kopepodittetthet for arealet i Vindafjorden for 800m-modellen (prikket linje) og 160m-modellen (heltrukket linje).
NorKyst800 gir et landsdekkende bilde av situasjonen dag for dag, mens NorFjords160 er avgrenset til produksjonsområdene. Det vil derfor kunne være feil i resultatene for kopepodittkonsentrasjon med strøm fra NorFjords160 nær de åpne grensene for modellområdene.
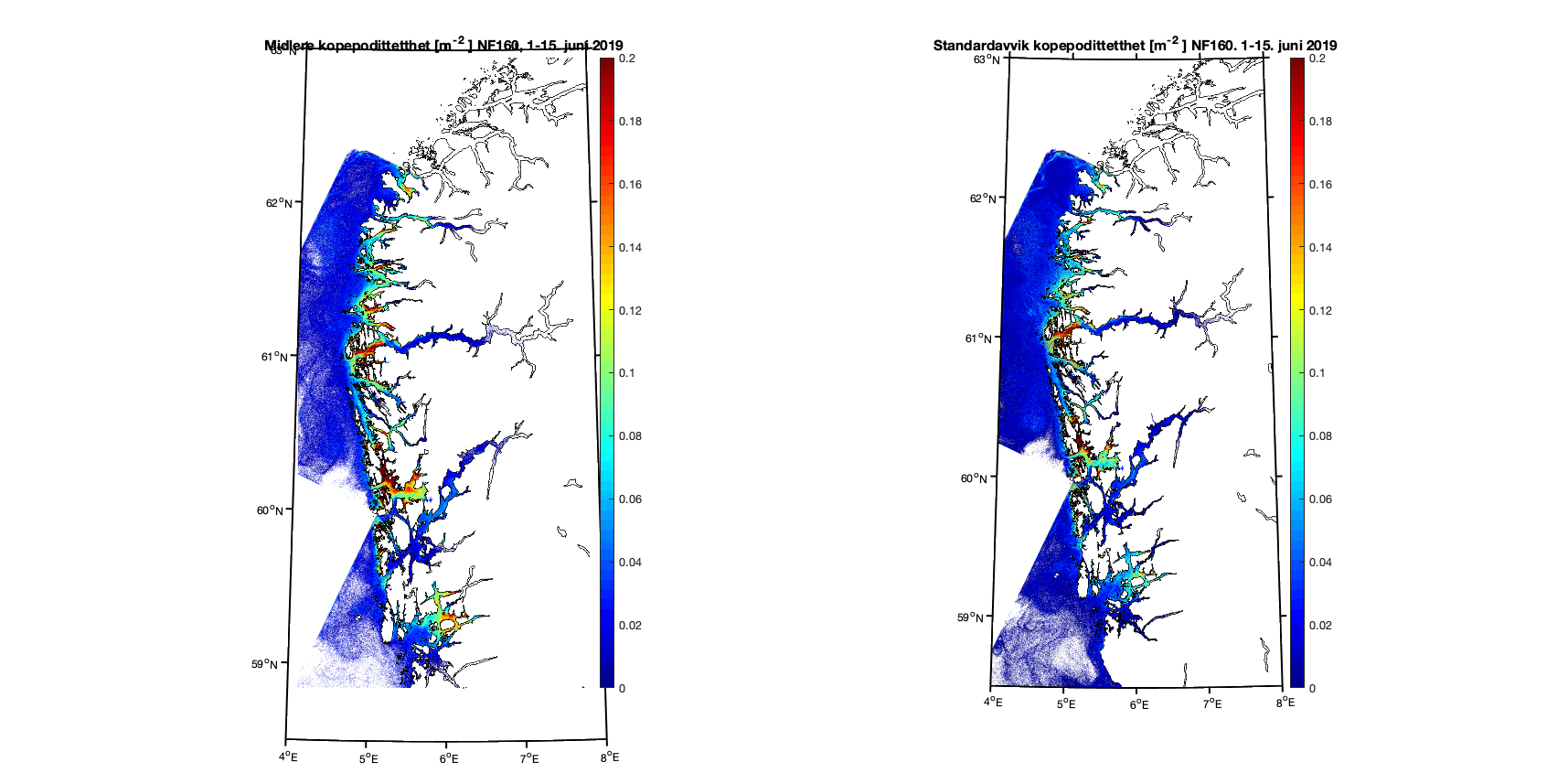
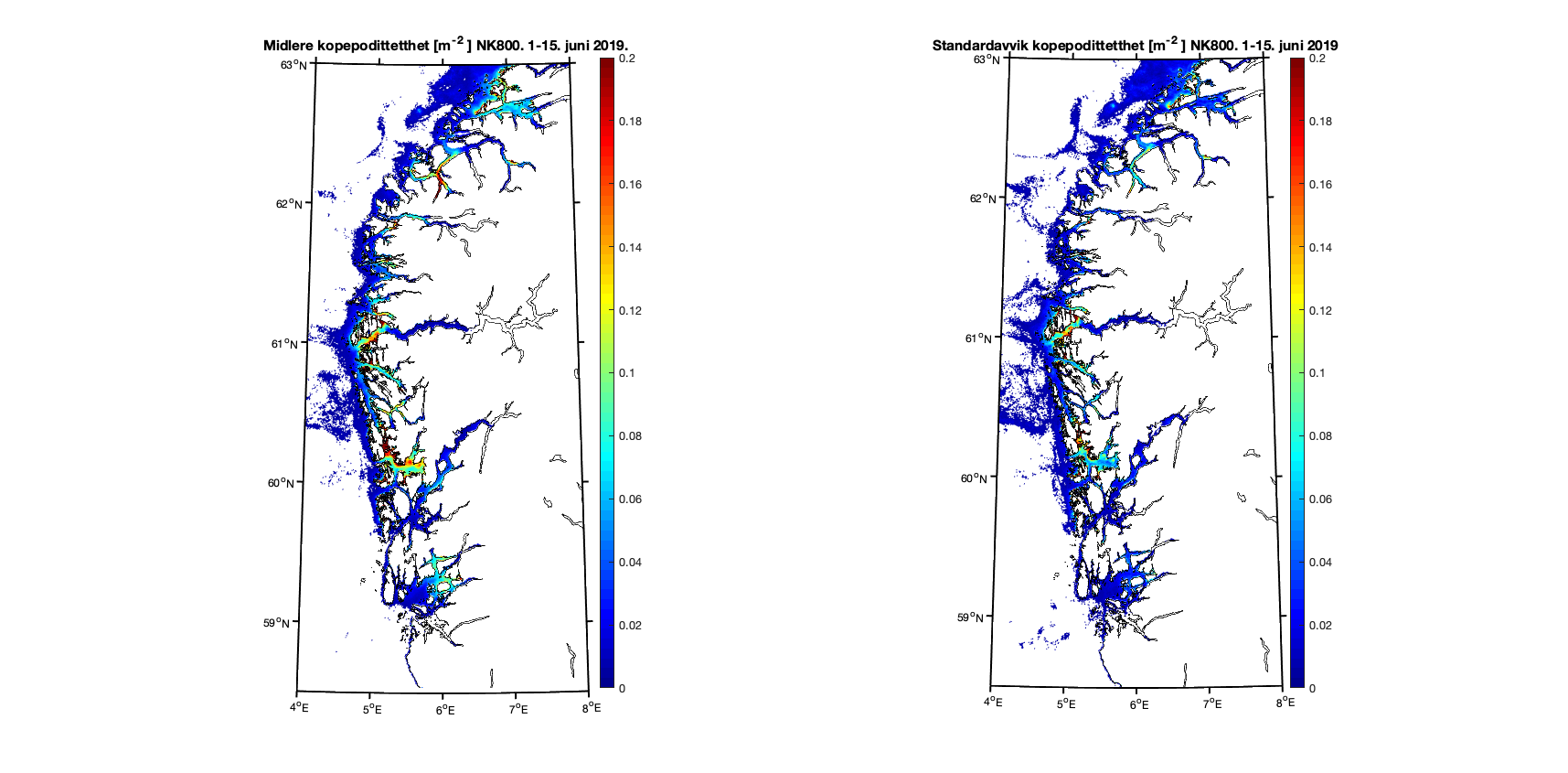
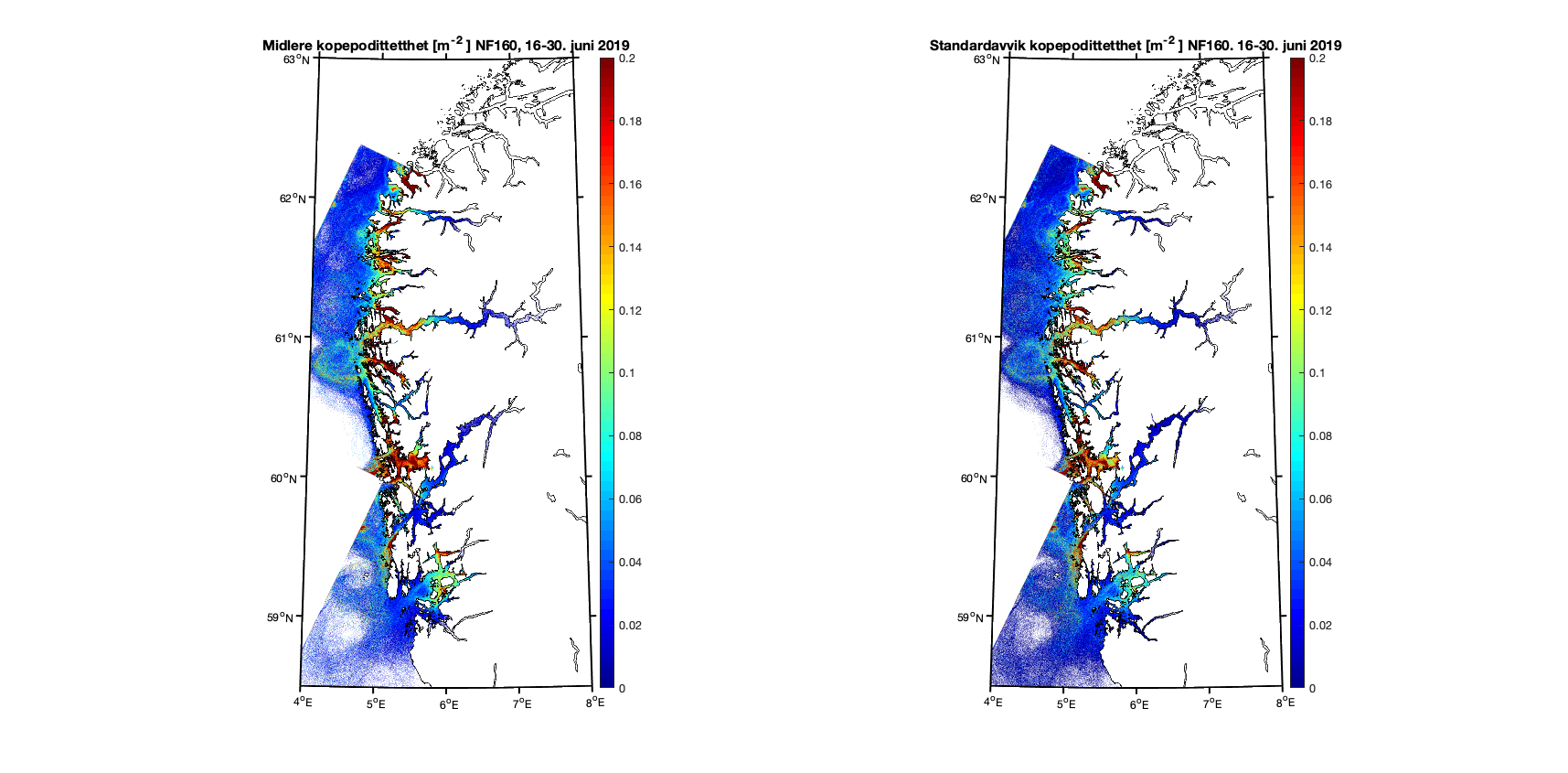
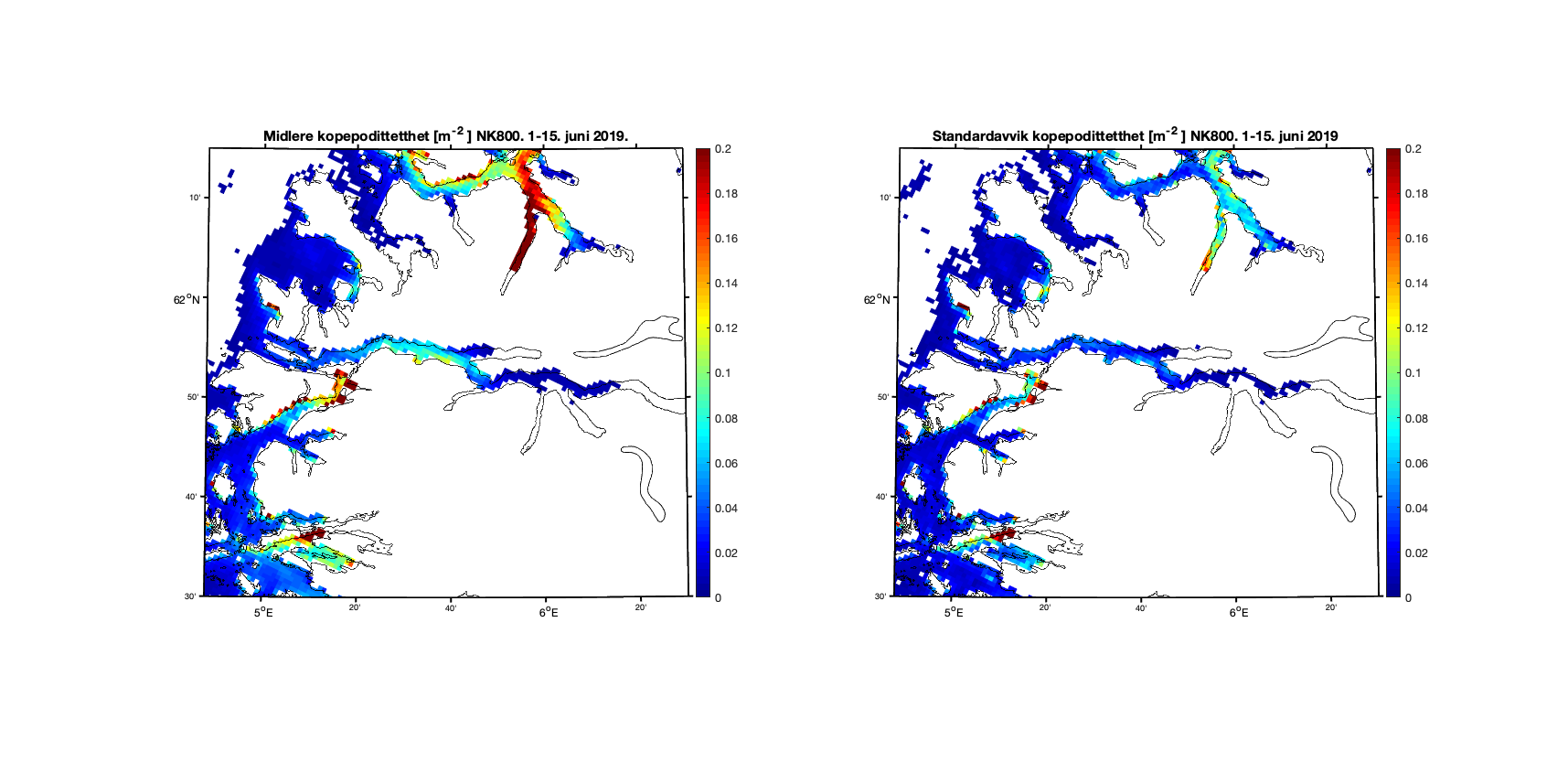
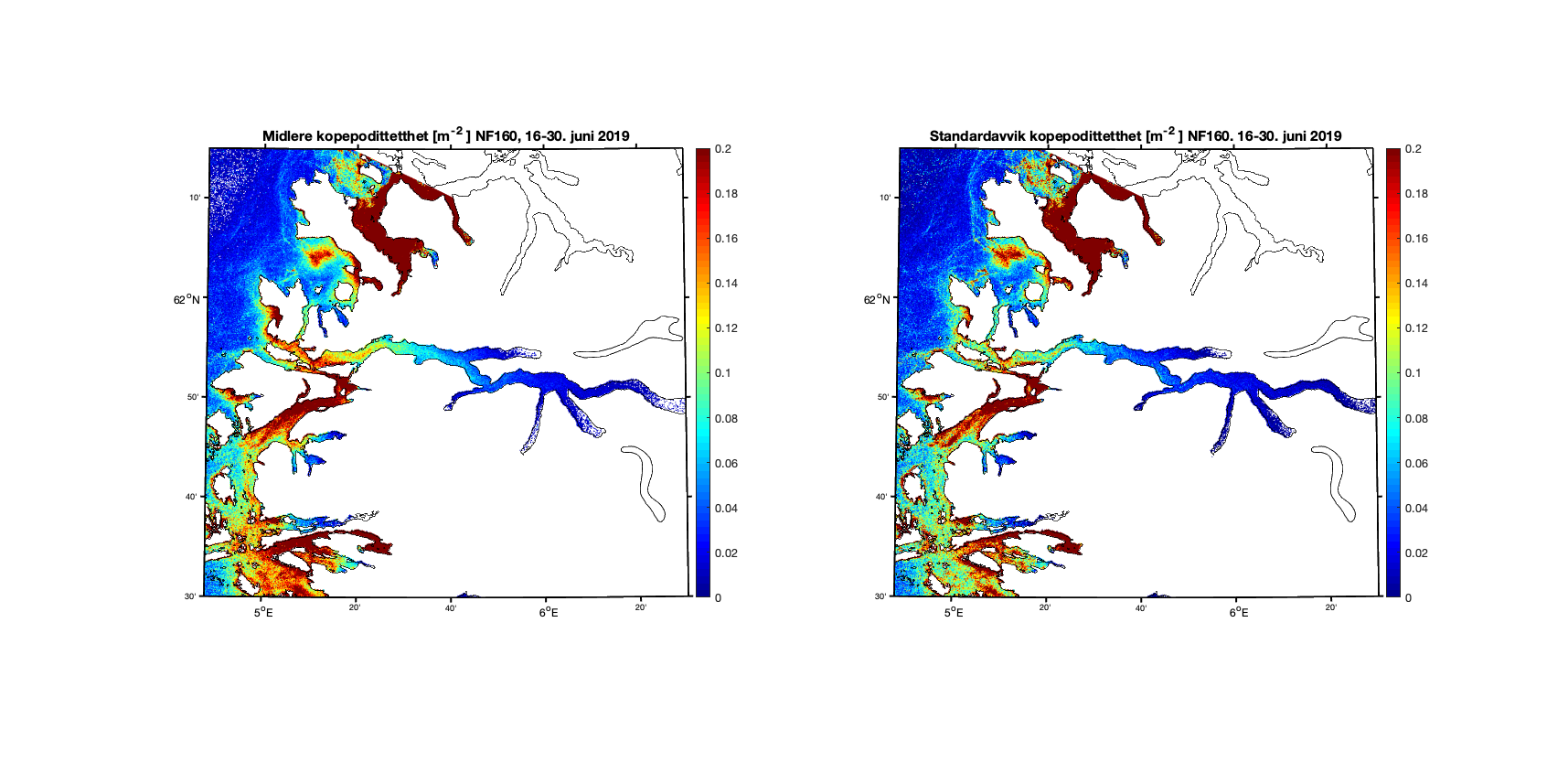
Aggregerer vi resultatene i toukersperioder finner vi for 1-15. juni 2019 at kopepodittfordelingen for begge de to strømmodellene er relativt like men med noen forskjeller på detaljnivå (Figur 8 og 9). Standardavviket (høyre kolonne i figurene) viser at det er mer variabilitet i resultatene med høyere oppløsning, omtrent som forventet.
Figur 8. Kopepodittetthet og standardavvik for NorKyst800 (nederst) og NorFjords160 (øverst) for perioden 1-15. juni 2019.
Figur 9. Kopepodittetthet og standardavvik for NorKyst800 (nederst) og NorFjords160 (øverst) for perioden 16-30. juni 2019.
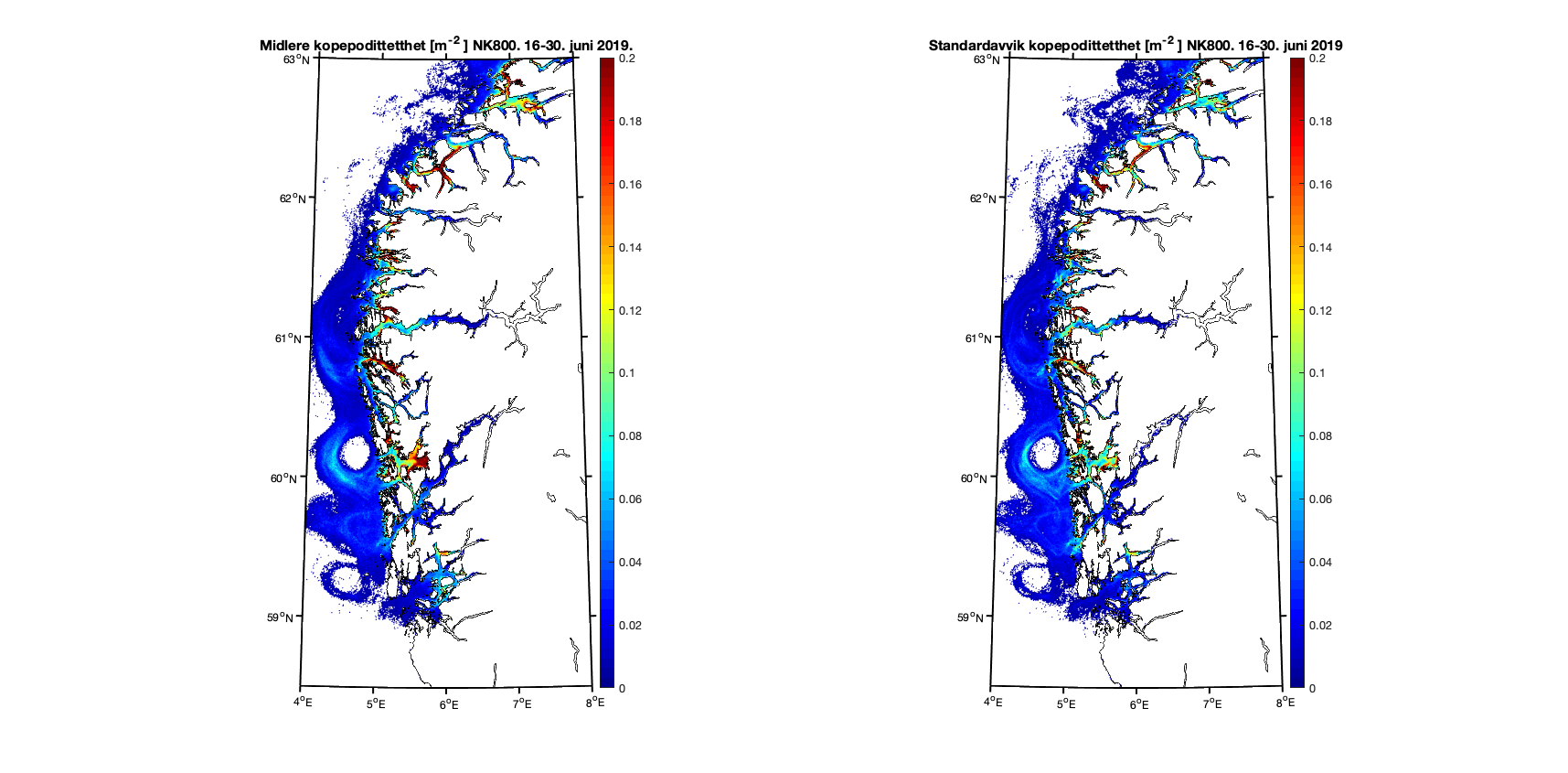
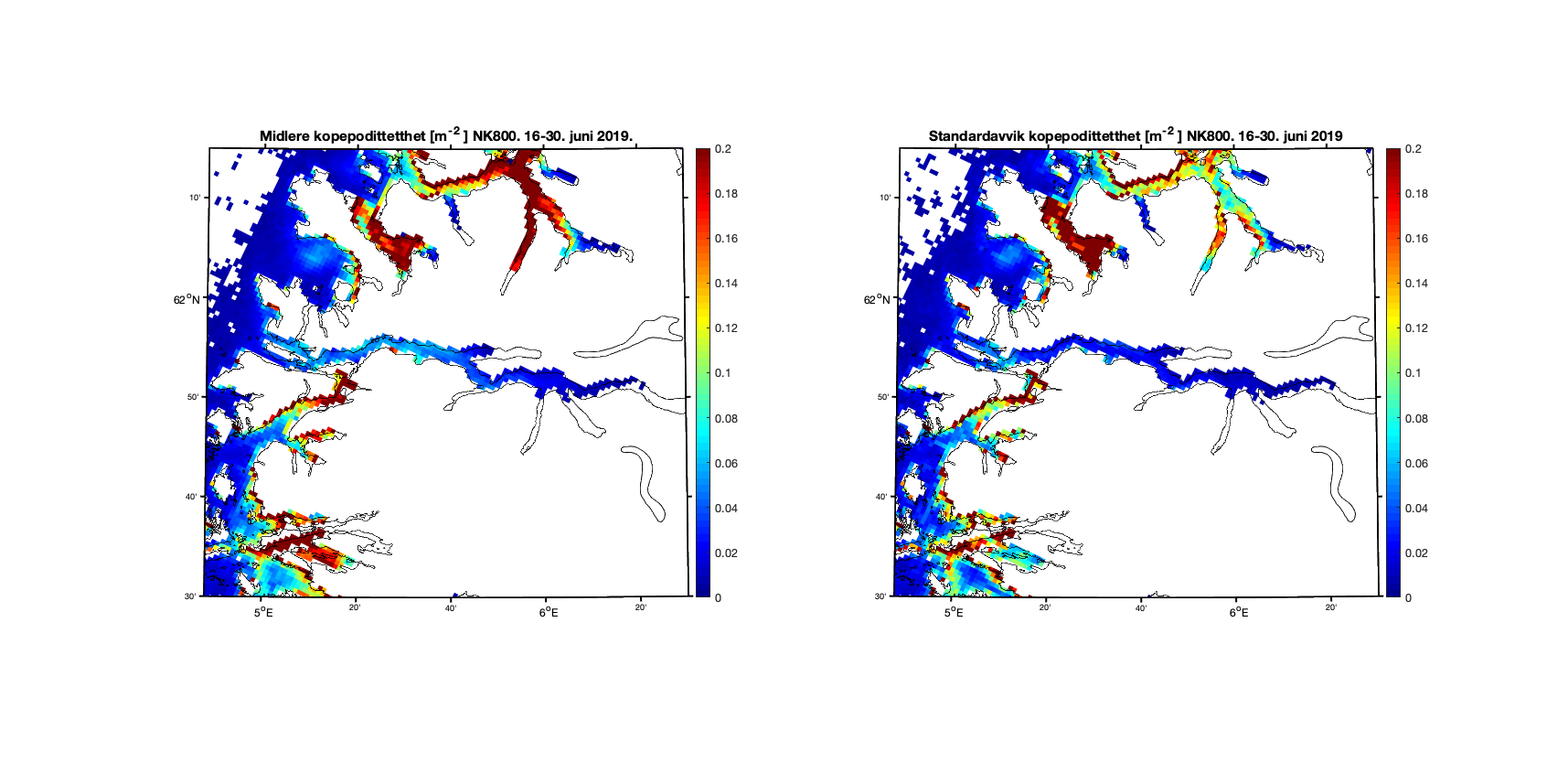
Forskjellene mellom modellresultatene viser seg klarest i områder der det er trange farvann og hvor 800m gitteroppløsning ikke inkluderer smale sund. Utløpet til Nordfjord er et eksempel på dette, og i første halvdel av juni er resultatene forholdsvis like (Figur 10) mens det i andre halvdel er det en god del kopepoditter i ytre del av Nordfjord i 160m-resultatene som ikke er der i 800m-resultatene (Figur 11).
Figur 10. Kopepodittetthet og standardavvik for NorKyst800 (nederst) og NorFjords160 (øverst) for perioden 1-15. juni 2019.
Figur 11. Kopepodittetthet og standardavvik for NorKyst800 (nederst) og NorFjords160 (øverst) for perioden 16-30. juni 2019.
Lenger nord i Vikna-området, er Nærøysundet forbi Rørvik bare ca. 400m bredt i store deler. Dette er så smalt at sundet ikke er åpnet i NorKyst800. Transport av lakselus nordover eller sørover dette sundet finner derfor bare sted i NorFjords160-modellen, og sundets påvirkning på sirkulasjonen på begge sider er ikke med i NorKyst800-resultatene. Simuleringene viser at det er en aggregering av kopepoditter rett nord for Nærøysundet i NorKyst800 som ikke er der i NorFjords160-resultatene (Figur 12).
Figur 12. Kopepodittetthet for NorKyst800 (venstre) og NorFjords160 (høyre) for 1-15. juni, 2019.
Smittepresskartet (ROC) viser at området nord for Vikna har en litt annen fordeling mellom rødt og gult fra NorKyst800 enn NorFjords160. Det samme gjelder sør for Vikna innover i Folda (Figur 13).
Figur 13. Smittepresskart basert på NorKyst800 (venstre) og NorFjords160 (høyre) for 1-15. juni, 2019.
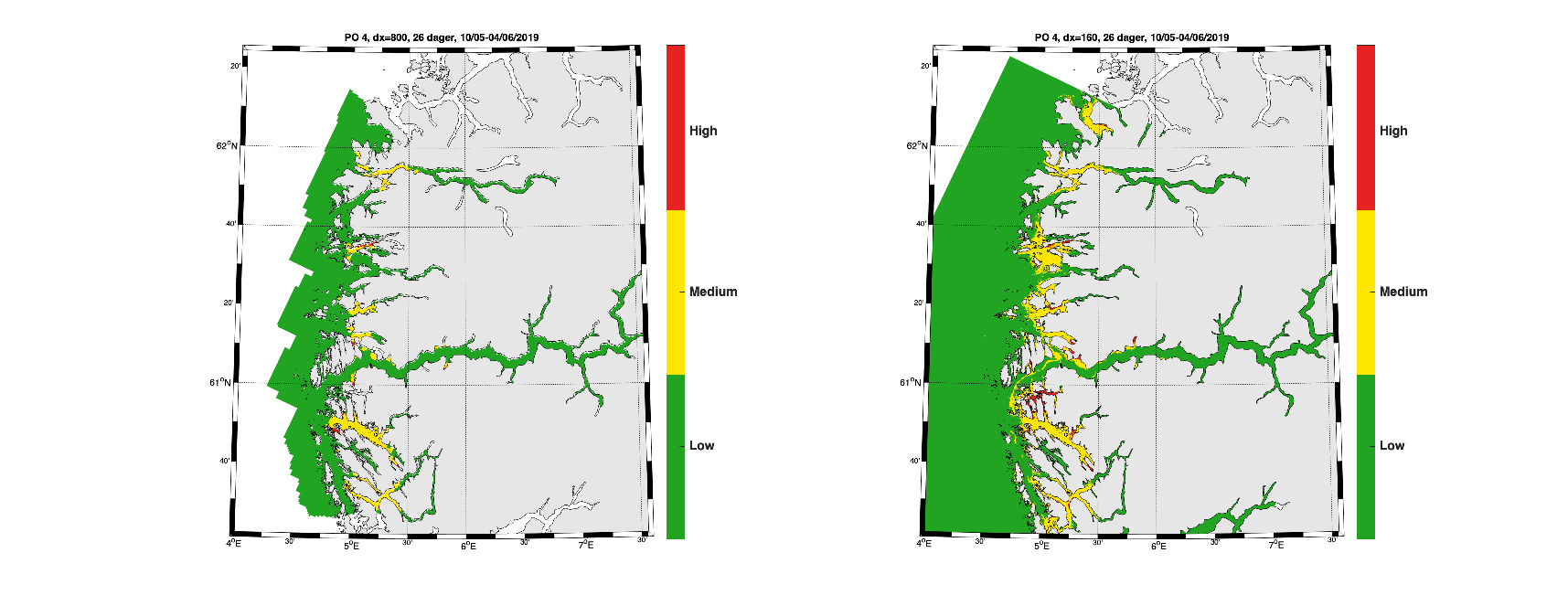
Smittepresskart fra Produksjonsområde 4 for mai viser at høyere oppløsning har en tendens til å gi litt større areal med gul og rød farge (Figur 14).
Figur 14. Smittepresskart basert på NorKyst800 (venstre) og NorFjords160 (høyre) for 10. mai til 4. juni, 2019, for Produksjonsområde 4.
Hvor stort utslag slike forskjeller kan ha på vurderingene for trafikklyssystemet er ikke undersøkt enda.
Våre foreløpige resultater av sammenligning mellom resultater mellom de to modellene med forskjellig oppløsning viser at:
- Vi finner ulikheter mellom spredningssimuleringsresultatene basert på strøm fra NorKyst800 og NorFjords160, men det er uklart om disse ulikhetene er signifikante i forhold til å vurdere smittesituasjonen i de ulike produksjonsområdene.
- NorFjords160-resultatene gir en god del mer variabilitet i rom og tid i avgrensede områder, men gjennomsnittsverdier ser ut til å være relativt sammenfallende.
- I trange områder der NorKyst800 ikke løser opp topografien vil det være noen forskjeller. Slike områder dominerer ikke modellområdene og totaleffekten på hele produksjonsområdet er ikke undersøkt enda.
- Det bør gjøres en grundigere studie for å tallfeste forskjellene, spesielt med fokus på resultatene av smittepresskart og populasjonsdødelighet til virtuell smolt. Dersom en slik studie påviser systematiske forskjeller, bør en bytte til modellen med høyere oppløsning for produksjon av resultater til Trafikklyssystemet. Enten for hele landet, eller for de produksjonsområdene der forskjellen er betydelige.
8 - Referanser
Albretsen, J., Sperrevik, A.K., Staalstrøm, A., Sandvik, A.D., Vikebø F. and Asplin, L., 2011. NorKyst-800 report no. 1: User manual and technical descriptions. Fisken og Havet nr. 2/2011, 51 pages.
Asplin, L., Boxaspen, K.K. & Sandvik, A.D. 2011. Modeling the distribution and abundance of planktonic larval stages of Lepeophtheirus salmonis in Norway. In S.R.M. Jones and R.J. Beamish (eds.) Salmon Lice: An integrated approach to understanding parasite abundance and distribution. Wiley-Blackwell, Hoboken, New Jersey, pp. 31-50.
Asplin, L., Johnsen, I.A., Sandvik, A.D., Albretsen, J., Sundfjord, V., Aure, J. & K.K. Boxaspen. 2014. Dispersion of salmon lice in the Hardangerfjord. Marine Biology Research, 10:3, 216-225, DOI:10.1080/17451000.2013.810755.
Browman, H., Boxaspen, K.K., Kuhn, P., 2004. The effect of light on the settlement of the salmon louse, Lepeophtheirus salmonis , on Atlantic salmon, Salmo salar L., Journal of Fish Disease, 27, 701-706.
Frenzl, B., 2014. Understanding key factors associated with the infection of farmed Atlantic salmon by the salmon louse Lepeophtheirus salmonis . PhD avhandling ved Institute of Aquaculture, University of Sterling, Skottland. 165 sider.
Genna, R.L., Mordue, W., Pike, A.W., Mordue (Luntz), A.J., 2005. Light intensity, salinity, and host velocity influence presettlement intensity and distribution on hosts by copepodids of sea lice, Lepeophtheirus salmonis , Canadian Journal of Fisheries and Aquatic Sciences, 62, 2675-2682.
Johnsen, I.A., Fiksen, Ø., Sandvik, A.D., Asplin, L., 2014. Vertical salmon lice behaviour as a response to environmental conditions and its influence on regional dispersion in a fjord system, Aquaculture Environment Interactions, 5, 127-141. DOI: 10.3354/aei00098.
Johnsen, IA., Asplin, L., Sandvik, AD., Serra-Llinares, RM., 2016. Salmon lice dispersion in a northern Norwegian fjord system and the impact of vertical movements. Aquaculture Environment Interactions, 8: 99-116, DOI: 10.3354/aei00162.
Nilsen, R. m.fl., 2017. Lakselusinfestasjon på vill laksefisk langs norskekysten i 2016, Rapport fra Havforskningen, Nr1-2017, 56 sider.
Sandvik, A.D., Bjørn, P.A., Ådlandsvik, B., Asplin, L., Skarðhamar, Johnsen, I.A., Myksvoll, M., Skogen, M.D., 2016. Toward a model-based prediction system for salmon lice infestation pressure. Aquaculture Environment Interactions, 8: 527-542, doi:10.3354/aei00193.
Sandvik, A.D, Johnsen, I.A., Myksvoll, M., Sævik P. N., Skogen, M.D., 2019. Prediction of the salmon lice infestation pressure in a Norwegian fjord. Innsendt til ICES Journal of Marine Science
Sandvik, A.D., Skagseth, Ø., Skogen M.D., 2016. Model validation: Issues regarding comparisons of point measurements and high-resolution modeling results, Ocean Modelling, 106, 68-73.
Skardhamar J, Albretsen J, Sandvik AD, Lien VS, Myksvoll MS, Johnsen IA, Asplin L, Ådlandsvik B, Halttunen E, Bjørn PA. Modelled salmon lice dispersion and infestation patterns in a sub-arctic fjord. ICES J Mar Sci. 2018; https://doi.org/10.1093/icesjms/fsx225
Ådlandsvik B, Morvik A, Sandvik AD 2017: « Salmon Lice Modelling at IMR - Source Term», https://docs.google.com/document/d/1zjAG6jxURC7UwVAVdZJseRl3ZDRRfZkxYc9nyXEb46A/edit?ts=5880a9e5#heading=h.bz2blmmrwuqn