The Institute of Marine Research have monitored kelp (Laminaria hyperborea) communities in southern parts (65° 13’ N - 65° 33’ N) of Nordland County in Norway in 2020-24. The monitoring was performed by underwater camera and acoustic observations of seabed vegetation along transects and stations included both harvesting fields and non-harvested reference areas. Kelp samplings for studies of age, morphology, associated epiphytes and sea urchin recruitment were performed at a subset of the stations. This part of Nordland County was opened for commercial kelp harvesting in 2022 after promising results from previous kelp harvesting trial studies. New observations at the fields previously subjected to harvesting trials indicate that kelp canopy size and biomass density were reestablished five years after harvesting, whereas kelp population age structure and epiphytic communities were not. Eight years after harvesting the age- and epiphytic structure of the kelp populations appeared reestablished and not different from that in unharvested reference populations.
Before the upstart of commercial kelp harvesting in 2022 video- and acoustic transects were established at all the harvesting fields. New observations along the same transect in 2023 and 2024 showed reduced kelp vegetation after harvesting. The recruitment of kelp has been good at the fields harvested in 2022, but lower at some fields harvested in 2023 due to increased coverage of furbelows (Saccorhiza polyschides) one year after harvest. The abundance of sea urchins was generally low and the recovery of the kelp vegetation after harvest appeared unaffected by sea urchin grazing.
To estimate the available kelp resources a biomass model was developed to predict the standing biomass of kelp at each harvesting field. The modelled estimates indicated large variation in standing kelp biomass between fields with a total sum of 516 000 tons kelp for all harvesting areas and an outtake of 23 % for the areas available for harvesting in the period 2022-24.
Sammendrag
Havforskningsinstituttet har i perioden 2020-24 gjennomført undersøkelser av taresamfunn på kyststrekningen Bindal (65° 13’ N) – Vega (65° 33’ N) i Nordland. Deler av denne kyststrekningen ble åpnet for kommersiell høsting av stortare (Laminaria hyperborea) i 2022, etter en prøvehøstingsperiode i 2013-17. Effekter av den tidligere prøvehøstingen på taresamfunn og kråkebolleforekomster ble undersøkt ved hjelp av undervannsvideo langs transekter og innsamlinger av tareplanter på utvalgte stasjoner for studier av morfologi, alder, assosiert biomangfold (epifyttsamfunn) og kråkebollerekruttering. Resultatene fra disse undersøkelsene indikerer at stortarevegetasjonens størrelse og biomasse er reetablert 5 år etter høsting, mens begroingssamfunnene (epifyttene) knyttet til tarestilkene er reetablert 8 år etter høsting og da lik tilsvarende observert i uhøstede referanseområder. Resultatene tyder på at forekomstene av kråkeboller er på et moderat nivå og at reetableringen av tare på høstefeltene ikke har vært hemmet av kråkebollebeiting.
Det ble i forkant av oppstarten av kommersiell tarehøsting i 2022 etablert overvåkingsstasjoner (transekter) for video og akustikk i samtlige høstefelt i Nordland der tilstanden sammenlignes med tilsvarende på stasjoner i uhøstede referanseområder. Akustikk er et verktøy som er under utvikling for å estimere biomassetetthet av stortare. En sammenligning av stasjoner som ble undersøkt før høsteoppstarten i 2022 og på ny i 2023 og 2024 viste redusert stortarevegetasjon etter høsting. Restitusjonen av stortare har imidlertid vært god på feltene som ble høstet i 2022, men stedvis noe redusert på enkelte felt høstet i 2023 pga høyere innslag av draugtare (Saccorhiza polyschides) året etter høsting.
I 2021 og 2022 ble det samlet inn data for etablering og validering av en biomassemodell for stortare i denne delen av Nordland for å få en mer helhetlig oversikt over ressursgrunnlaget sammenlignet med uttaket av stortare på de enkelte høstefeltene. Resultatene fra modellkjøringene indikerte stor variasjon i stortarebiomasse mellom felt med et totalt biomasseestimat på 516 000 tonn for alle høstearealene tilsammen og en uttaksgrad på 23 % totalt for arealene som har vært åpne for høsting i perioden 2022-24.
1 - Innledning
Den ytre del av kystrekningen i Nordland som strekker seg fra Trøndelagsgrensen i sør til Vega (sør for breddegrad 65° 33’ N) i nord ble åpnet for kommersiell høsting av stortare (Laminaria hyperborea) sommeren 2022 (Fiskeridirektoratet J-131-2022). Denne regionen har tidligere vært preget av kråkebollebeiting, men observasjoner av tilbakegang i kråkebollebestandene og gjenvekst av tare (Norderhaug & Christie 2009, Fagerli et al. 2013, 2015) har gjort dette området attraktivt for tarehøsting. Kunnskapsgrunnlaget for igangsetting av kommersiell høsting av stortare i denne regionen er basert på erfaringer og oppfølgende undersøkelser gjennomført av Havforskningsinstituttet i forbindelse med prøvehøstinger i perioden 2013-17 (Steen et al. 2018ab, 2020). Tarehøstingen i Nordland foregår, som i øvrige områder langs norskekysten der denne aktiviteten er tillatt, ved bruk av tindetrål.
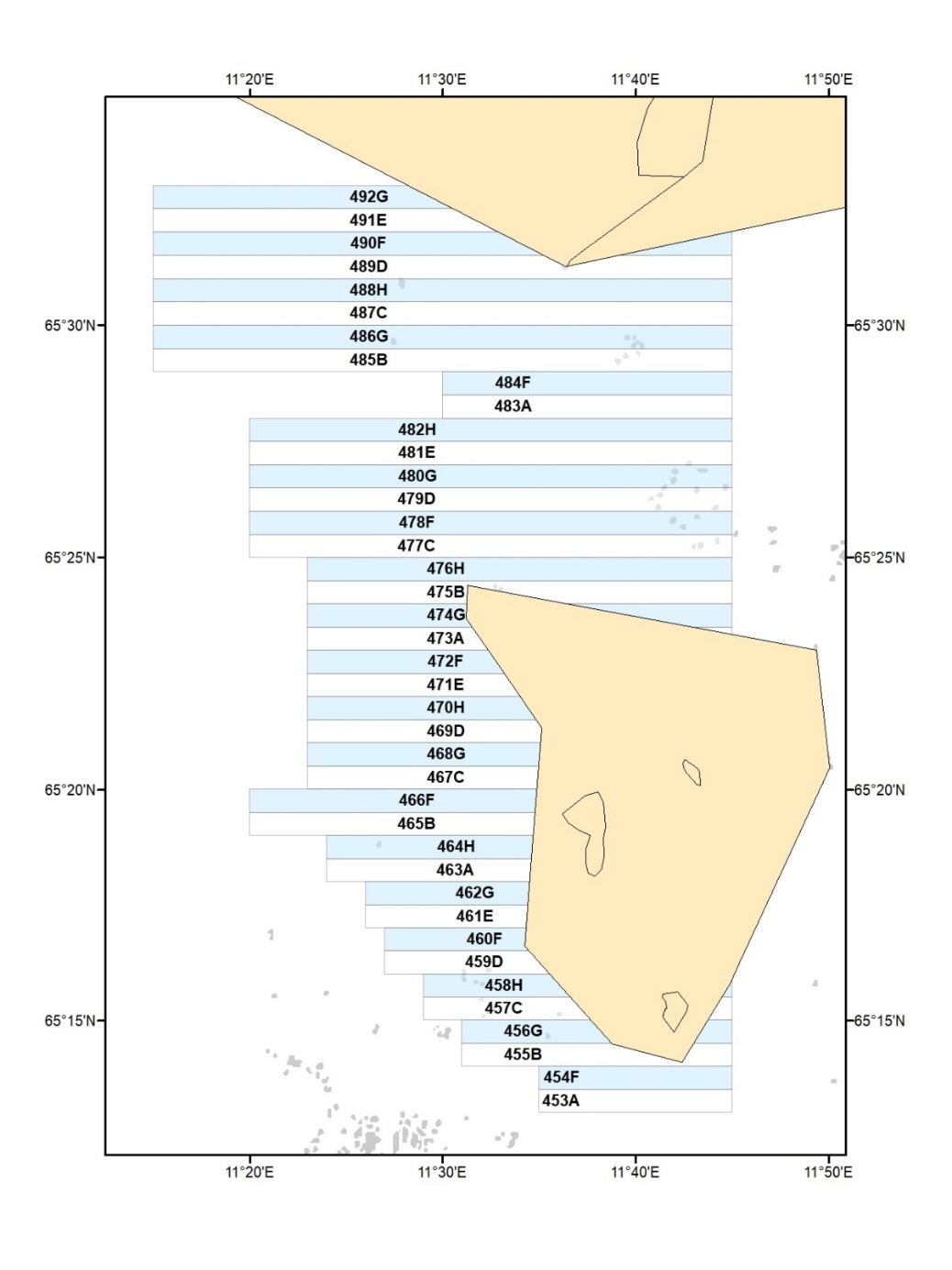
Høsteområdet for stortare i Nordland er inndelt i 40 felt, der den ene halvparten av feltene høstes med 5 års syklus og resterende halvpart høstes med 10 års syklus (Fig. 1). En slik differensiert høstesyklus byr blant annet på muligheter for å teste i hvilken grad lengden på hvileperioden mellom høstingene vil påvirke taresamfunnenes produksjon, restitusjonsevne og økologiske funksjonalitet over tid. Høstesesongen i Nordland er begrenset til perioden juni – august, mens høstingen i øvrige regioner langs norskekysten foregår året rundt. Feltene i Nordland er angitt med et nummer (453 – 492) og en bokstavkode (A - H), der sistnevnte refererer til hvilke år feltene er åpne for høsting (Tab. 1).
Tabell 1. Bokstavkoder og høsteår for tarehøstefelt i Nordland.
Feltkode
1 høsteår
2 høsteår
A
2022
2027
B
2023
2028
C
2024
2029
D
2025
2030
E
2026
2031
F
2022
2032
G
2025
2035
H
2028
2038
Havforskningsinstituttet gjennomførte i 2020 og 2022 oppfølgende videoobservasjoner på feltene som også ble undersøkt før den siste prøvehøstingen i 2017 (Steen et al. 2018a) for å undersøke utviklingen av tarevegetasjon og kråkebolleforekomster. I perioden 2022-24 er det også samlet inn tareprøver på et utvalg av feltene som ble prøvehøstet henholdsvis 5 år og 8 år tidligere for å sammenligne tarevegetasjonens morfologi, alder, veksthistorikk og påvekstorganismer (epifyttsamfunn) her med tilsvarende hos tareplanter samlet inn i uhøstede referanseområder. Disse undersøkelsene vil bidra med utfyllende kunnskap om taresamfunnenes reetableringsevne og restitusjonsperiodens varighet i denne regionen.
I forkant av oppstart av den kommersielle tarehøstingen etablerte Havforskningsinstituttet i 2022 overvåkingstransekter som ble undersøkt ved hjelp av undervannsvideo og akustikk i samtlige høstefelt i Nordland. Sammenlignbare undersøkelser ble også gjennomført langs transekter i uhøstede referanseområder. Akustikk er ny kostnadseffektiv metode for estimering av tarebiomasse som tidligere har vist lovende resultater (Kartveit et al. 2022), og som planlegges å tas mer i bruk i tareovervåkingen langs kysten. Det ble i 2023 og 2024 gjennomført oppfølgende undersøkelser med tilsvarende metodikk som i 2022 langs samtlige transekter, for å undersøke utviklingen på nylig høstede felt sammenlignet med andre felt og referanseområder.
For å få mer helhetlig kunnskap om forekomstene av tareressurser på høstefeltene i Nordland ble det i 2021-22 samlet inn data fra randomiserte stasjoner (videoobservasjoner) for utvikling av en prediksjonsmodell for stortarebiomasse i dette området. En sammenligning av biomasseestimatene fra modellen med høstekvantumet vil gi informasjon om aktivitetens omfang og uttaksgraden av stortare på høstefeltene, noe som vil være nyttig kunnskap for tareforvaltningen dersom man i framtiden vurderer å innføre feltspesifikke kvantumsgrenser på uttaket av stortare (Norderhaug et al. 2021).
Figur 1. Kart over høstefelt for stortare i Nordland. Hvite felt er åpne for høsting hvert 5 år,
og lyseblå felt er åpne for høsting hvert 10 år. Beige polygoner representerer
referanseområder (Horsvær naturreservat i sør og Vega verdensarvområde i nord).
2 - Metodikk
2.1 - Videoundersøkelser
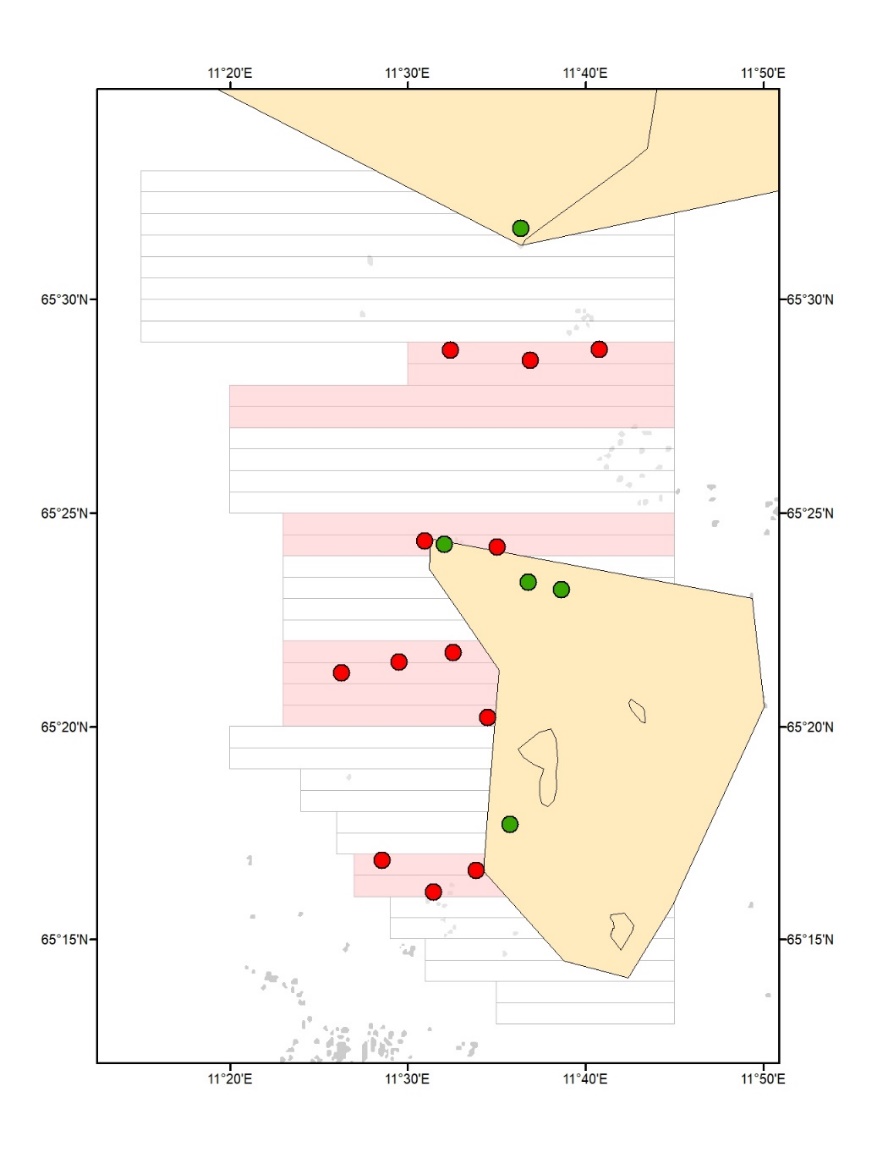
Havforskningsinstituttet har i 2020 og 2022 gjennomført oppfølgende videoundersøkelser på stasjoner som også ble undersøkt i forkant av den siste prøvehøstingen som ble gjennomført i Nordland, i 2017 (Steen et al. 2018a), for å sammenligne tarevegetasjonens utvikling på prøvehøstefeltene med tilstanden i uhøstede referanseområder (Fig. 2). Disse stasjonene vil også inngå i stasjonsnettet som benyttes for overvåking av den kommersielle tarehøstingen fra 2022 (Fig. 3). Alle undersøkelsene i Nordland er gjennomført i samme årstid (juni) for å sikre at resultatene ikke påvirkes av sesongvariasjoner.
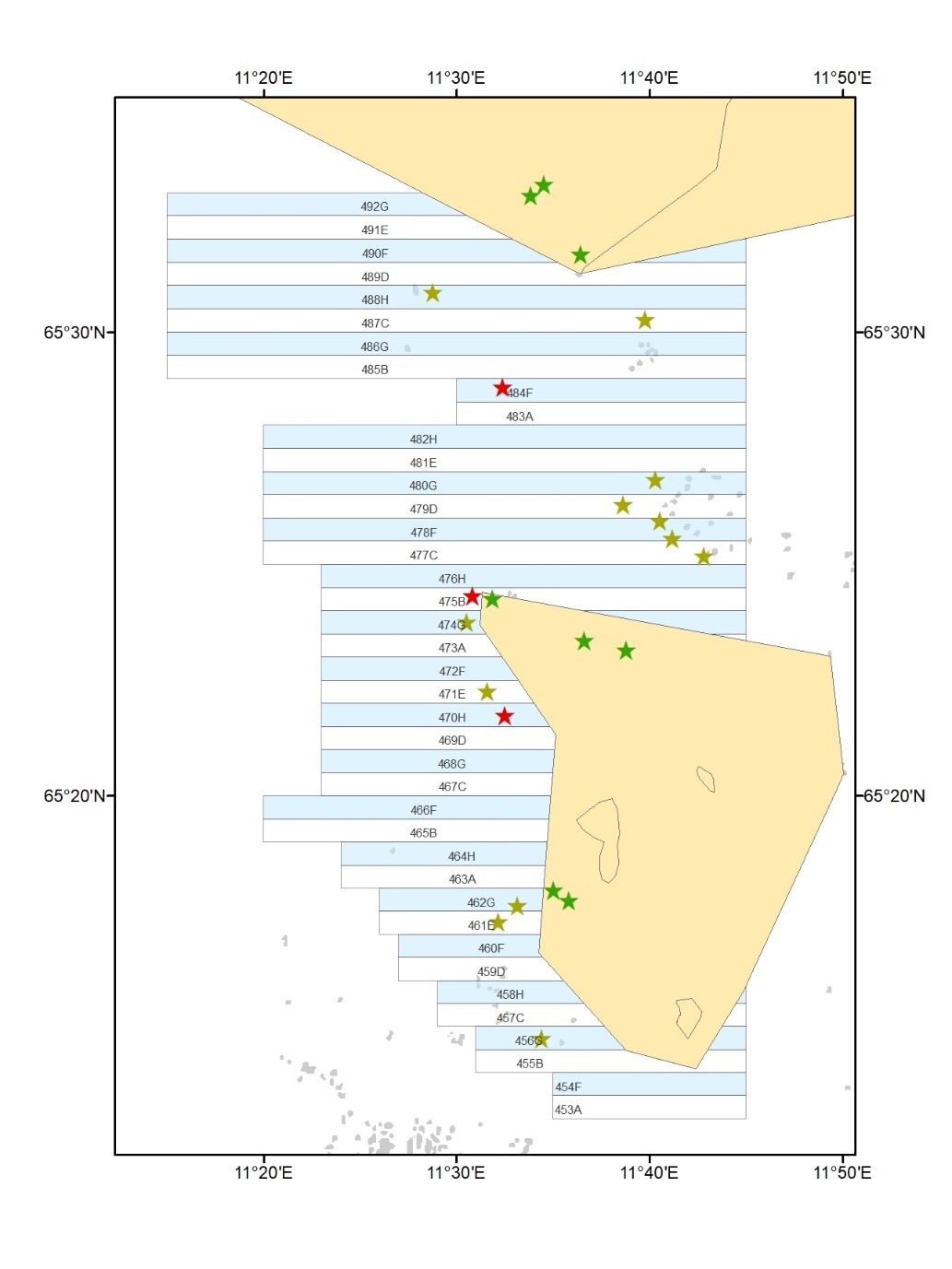
Figur 2. Videostasjoner som ble undersøkt i forkant av prøvehøstingen i 2017 og
på ny i 2020 og 2022.Stasjoner i prøvehøstefelt (røde symboler) og uhøstede
referanseområder (grønne symboler). Felt som ble prøvehøstet i 2017 (røde polygon).
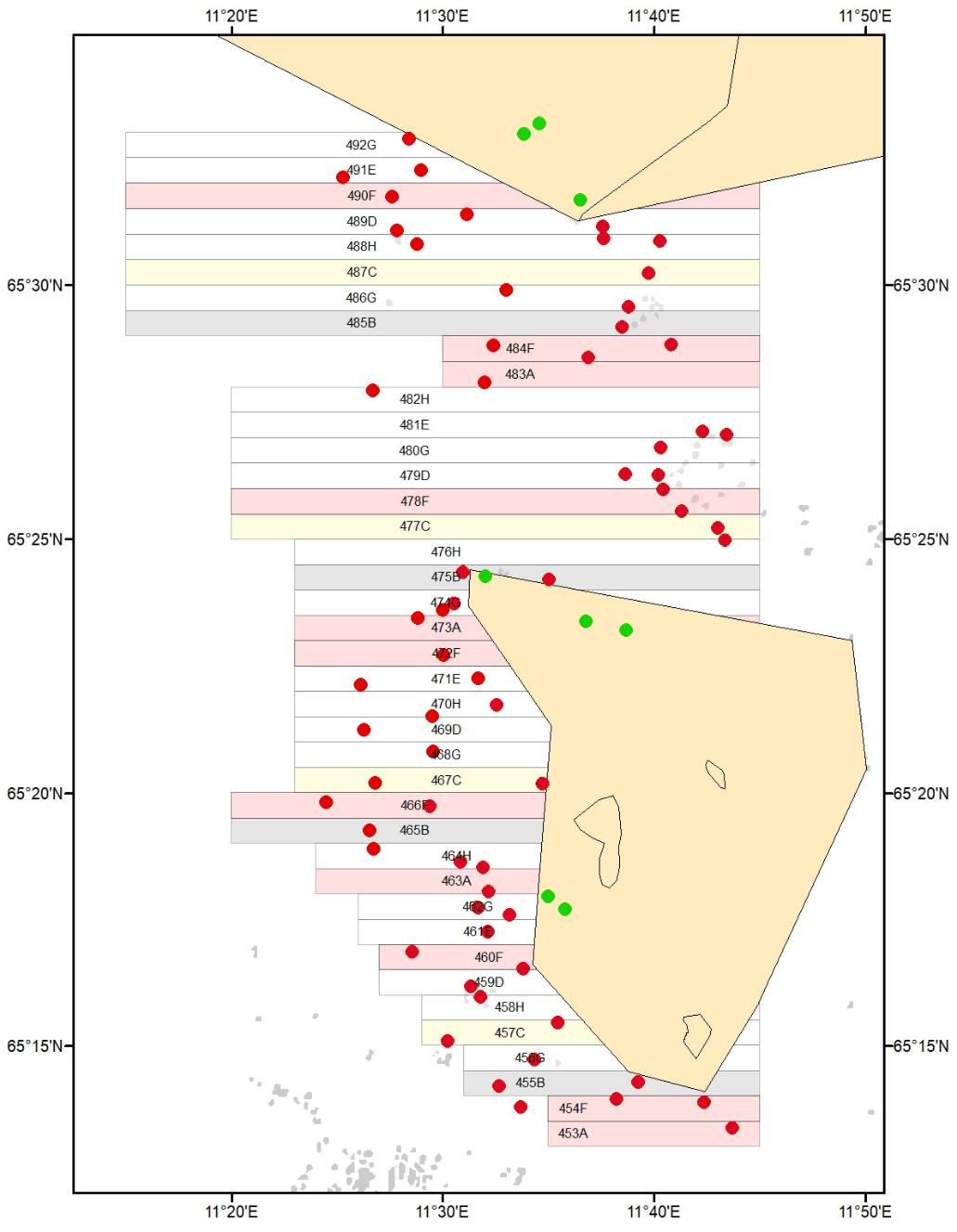
Før oppstart av kommersiell tarehøsting i Nordland etablerte Havforskningsinstituttet, i juni 2022, overvåkingsstasjoner for undervannsvideo og akustikk i samtlige felt som inngår i høsteplanen og i nærliggende referanseområder som er stengt for tarehøsting (Fig. 3, Appendix 1). Overvåkingsstasjonene ble plukket ut på basis av dybde- og bunnforhold, samt innrapporterte sporingsdata etter tidligere høsteaktivitet i feltene der denne type informasjon foreligger. Totalt 65 stasjoner ble undersøkt i høstefeltene og 8 stasjoner i referanseområdene. I juni 2023 og 2024 ble det gjennomført oppfølgende undersøkelser med tilsvarende metodikk på samtlige av disse stasjonene (Fig. 3, Appendix 2, 3) for å undersøke tilstanden på de nylig høstede feltene sammenlignet med tilstanden i uhøstede områder.
Bølgeeksponering er en faktor som påvirker utbredelse og vekst av stortare (Norderhaug et al. 2011, 2012, Pedersen et al. 2012, Bekkby et al. 2014, Steen et al. 2018b). Bølgeeksponering basert på strøklengde og topografi med 50m x 50m oppløsning ble beregnet i form av midlere signifikante bølgehøyde (MSWH) for overflatebølger i meter (beskrevet i Steen et al. 2018b) på samtlige stasjoner. Dette for å undersøke i hvilken grad denne faktoren varierer mellom referanse- og høstefeltene og påvirker de ulike målvariablene.
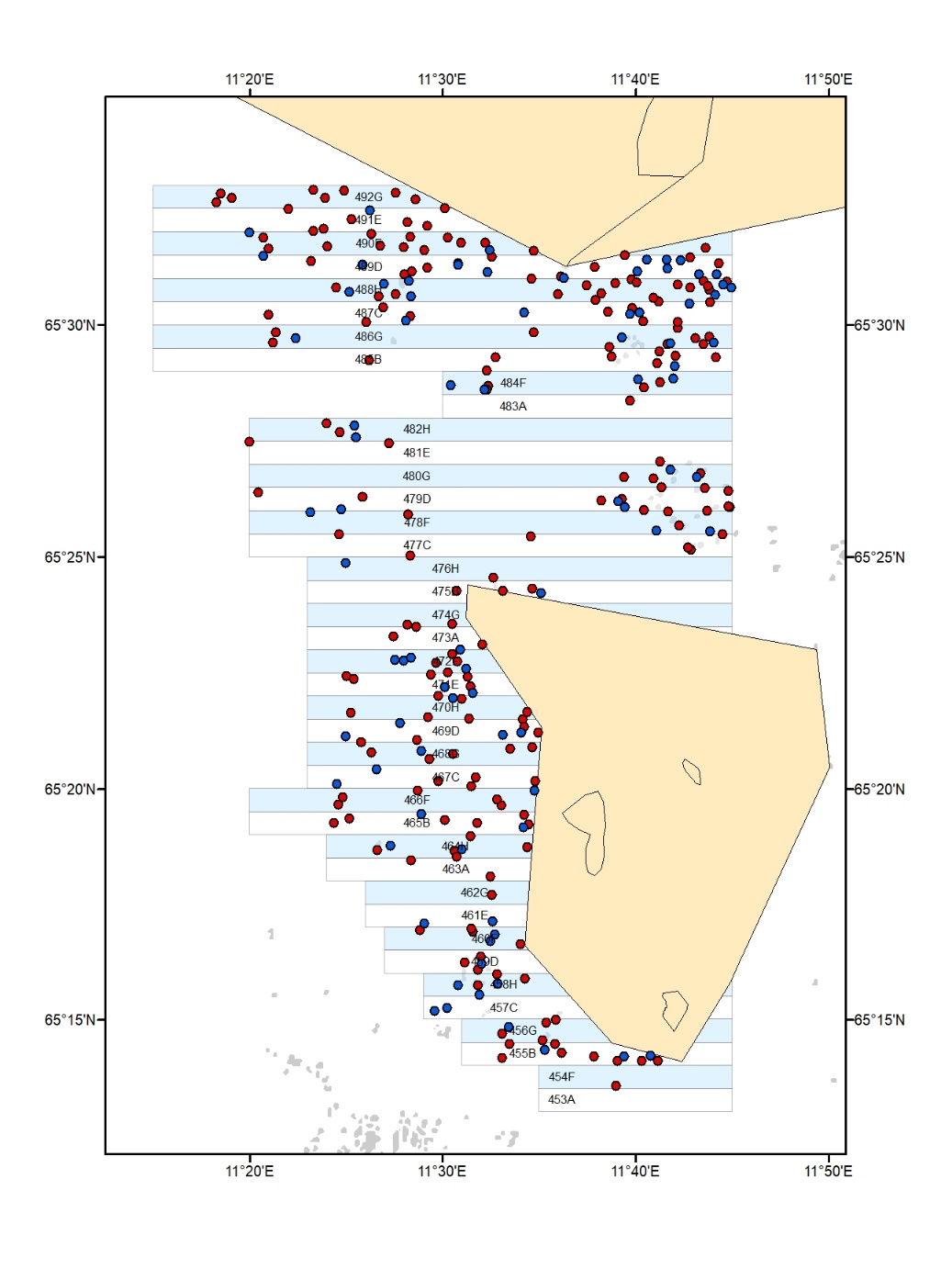
Figur 3. Video- og akustikkstasjoner som ble undersøkt i høstefelt (røde punkt)
og uhøstede referanseområder (grønne punkt) i 2022-24. Felt åpne for høsting
i 2022 (lyserøde rektangler), felt åpne for høsting i 2023 (lysegrå rektangler),
felt åpne for høsting i 2024 (lysegule rektangler).
Videoundersøkelsene ble gjennomført og analysert etter samme metode som benyttet ved tidligere tarerelaterte undersøkelser og overvåking (Steen 2019, 2020, 2021, 2022, Steen et al. 2018ab, 2020, 2023, Steen & Kartveit 2024). Videoobservasjonene på de enkelte stasjonene ble gjennomført med nedsenkbart undervannskamera (UVS 5080), med innebygd dybdesensor, langs transekter fra båt (FF Fangst) med kartplotter og ekkolodd, med en gjennomsnittshastighet på ca 0,5 - 0,8 knop. I tillegg ble det montert et Paralenz dive camera med dybdesensor på siden av UVS 5080-kameraet for å få høyoppløselige bilder (Figur 4). Undervannskameraene ble vekselvis ført rett over og gjennom tarevegetasjonen over en strekning (transekt) på ca 100-200m, der kamerapiloten justerer høyden i forhold til tarevegetasjonen og bunnen ved hjelp av en monitor. Filmene fra UVS 5080 kameraet ble fortløpende konvertert til PC-format (mpg) vha en Pinnacle movie box og lagret på eksterne harddisker, mens filmene fra Paralenz-kameraet ble lagret på interne minnebrikker og daglig overført til eksterne harddisker.
Figur 4. Undervannskamerasystemer benyttet i Nordland.
Før analyse ble videotransektene splittet opp i mindre avsnitt (for hvert minutt film) der gjennomsnittlig dyp, bunntype, tarevegetasjonens dekningsgrad (definert som andel bunnflate dekket av tarevegetasjon), tetthet og høyde ble anslått. Høyden av tareplantene måles fra tareplantenes festeorgan (dvs bunnen) til tarebladet vha kameraets innebygde dybdesensor. Det ble også gjennomført tellinger av kråkeboller og forekomstene ble angitt som antall registreringer per meter videotransekt.
Det ble også gjort anslag av gjennomsnittlig biomassetetthet av stortare på hver stasjon ved å multiplisere gjennomsnittlig plantevekt med den gjennomsnittlige plantettheten observert langs videotransektene. Stortareplantenes vekt ble beregnet ut ifra plantehøyden som observeres langs videotransektene ved å benytte en relasjon mellom plantehøyde og vekt utviklet gjennom tidligere målinger av innsamlede stortareplanter (Steen et al. 2020, 2023, Van Son et al. 2020).
For å teste effektene av tarehøsting på målvariablene ble det benyttet lineære sammensatte modeller («linear mixed-effect models») tilpasset med “lmer” funksjonen i “Lme4” pakken i R (Bates et al. 2015). Fordeling (grafiske plot) og varianshomogenitet (Levene’s test) av residualer ble kontrollert før kjøring av testene. Korrelasjoner mellom variablene ble estimert vha Pearsons produkt-moment korrelasjonskoeffisient (r).
2.2 - Akustikkundersøkelser
Langs samtlige transekter som ble undersøkt i Nordland i perioden 2022-24 (Fig. 3) ble det gjort akustiske registreringer for estimering av stortareforekomstenes biomassetetthet. Det ble benyttet Simrad EK60/80 ekkolodd med split beam transducer ES38B og ES120-7C for innsamling av 38 og 120 kHz data (Kartveit et al. 2022, Steen & Kartveit 2024). Dataene ble prosessert med programvaren KORONA for, blant annet, å fjerne støy og detektere bunn, og tolket i Large Scale Survey System (LSSS). Den manuelle tolkningen i LSSS består i hovedsak av å beskjære korrumperte datasett og tegne integrasjonslinjer rundt tareskogen. På denne måten kan man beregne den akustiske mengden (nautical area backscatter coefficient - NASC) i integrasjonsarealet (tareskogen). NASC-verdiene lagres så i en database og kan hentes derfra for å brukes videre i statistisk analyse og for å estimere biomassetetthet av stortare for hvert transekt. De akustiske målingene ble filtrert ved hjelp av interquartile range (IQR-metoden), og uteliggere er gitt ekstremalverdi. Dette er gjort fordi den lineære modellen kan gi estimater langt over det som er realistisk – for eksempel i tilfeller der luftbobler fra sjøgang og fartøy har penetrert tarevegetasjonen og gitt ekstremt høye NASC-verdier (og dermed også biomasseestimat). Akustikkdata ble samlet inn samtidig med videoundersøkelsene da fartøyet beveger seg sidelengs, og i tillegg ble det samlet inn et sett med akustiske data fra der fartøyet beveger seg rettvendt med en høyere hastighet (ca. 5-6 knop) langs den samme transektlinjen. Dette ble gjort for å undersøke hvordan datakvalitet og tolkningsresultater påvirkes av ulike innsamlingsforhold.
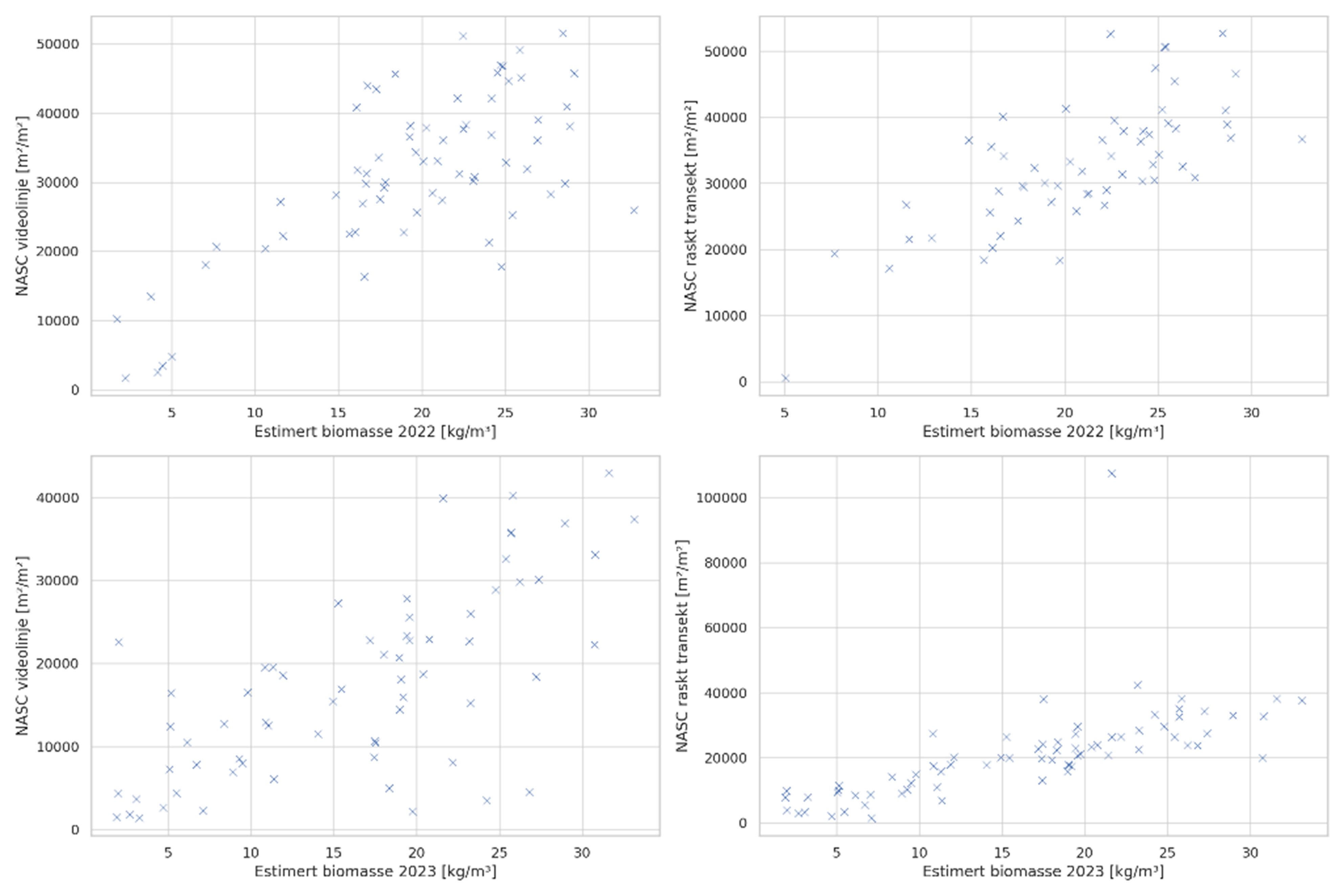
For å estimere biomassen basert på NASC-verdiene ble det brukt en modell der historiske akustikkmålinger er plottet mot biomasse estimert fra videoanalysene fra de samme transektene (Fig. 5), før det ble utført lineær regresjon på hvert datasett samt alle datapunkter samlet. Figur 5 viser en langt bedre samling av akustikkdata innhentet med høyere hastighet (heretter kalt «akustiske transekter») enn datasettet som ble samlet inn simultant med video. Ekkogrammene fra de akustiske transektene viser også tydelig mindre støy enn for ekkogrammene basert på data innhentet samtidig med videoinnsamlingen, og på bakgrunn av dette er det derfor de akustiske transektene som er benyttet som grunnlag for biomasseestimering. I de tilfellene der de akustiske transektene er korrumperte eller mangler (som tilfelle i 2022) har akustikkdata samlet inn samtidig med videokjøringene blitt brukt som substitutt. Data fra stasjon 455Be var korrumperte i 2022 og dette transektet er derfor ikke inkludert i datasettet. Vi har derfor et datasett med 72 transekter fra 2022 (hvorav 12 er innhentet samtidig med videokjøringene), og 73 akustiske transekter fra 2023 og 2024.
Figur 5: Sammenligning av NASC-verdier og estimert biomasse fra videoanalyser langs de samme transektlinjene
fra 2022-2023. Plot til venstre er fra de langsomme videotransektkjøringene, plot til høyre er fra de raske
akustiske transektkjøringene. Data fra samtlige akustiske transekter er inkludert i disse plottene.
2.3 - Tareinnsamlinger
For undersøkelser av tarevegetasjonens morfologi, alder, veksthistorikk, epifyttsammensetning og forekomst av kråkebollerekrutter, er det i perioden 2022-24 samlet inn tareplanter ved bruk av samme metode som er benyttet ved tidligere undersøkelser (Steen 2021, 2022, Steen et al. 2 018b, 2020, 2023, Steen & Kartveit 2024). 10 tareplanter ble samlet inn fra hver av 23 ulike stasjoner ved hjelp av tareskrape (Fig. 7, Appendix 4). For å sammenligne taresamfunnenes utvikling etter tidligere høstinger med ulik hvileperiode ble innsamlingsstasjonene fordelt med 3 stasjoner på felt som ble prøvehøstet 5 år tidligere, 12 stasjoner i felt som ble prøvehøstet 8 år tidligere og 8 stasjoner i uhøstede referanseområder (Fig. 7). Tillatelse til innsamlinger av stortare i Horsvær naturreservat ble gitt av Statsforvalteren i Nordland og Miljødirektoratet.
Figur 6. Tareskrape i stål (ca 60 cm bred), benyttet til innsamling
av stortareplanter i Nordland
Figur 7. Innsamlingsstasjoner for stortare i juni 2022-24. Stasjoner i felt som ble høstet
5 år tidligere (røde stjerner), stasjoner i felt som ble høstet 8 år tidligere (gule stjerner)
og stasjoner i referanseområder (grønne stjerner).
De innsamlede stortareplantene ble analysert vha samme metode som benyttet ved tidligere prøvetakinger (Steen 2021, 2022, Steen et al. 2018b, 2020, 2023). Tarestilkenes lengde ble målt fra overgang festeorgan/stilk til overgang stilk/blad, mens bladlengden ble målt fra overgang blad/stilk til enden av blad. Påvekstorganismene (epifyttene) ble skrapt av tarestilkene og veid (våt vekt), og de dominerende arter/grupper ble rangert etter forekomst for hver enkelt tarestilk. Tareplantenes epifytter har stor betydning for tareskogens totale biomangfold og produksjon (Christie et al. 1998, 2003, 2007, Norderhaug et al. 2003, Pedersen et al. 2014, Teagle et al. 2017) og benyttes her som en indikator på taresamfunnenes økologiske funksjonalitet.
Tareplantenes blad (lamina), stilkdel (stipes) og festeorgan (hapter) ble separert og veid (våt vekt) hver for seg. Forekomster av juvenile kråkeboller (rekrutter) i tareplantenes festeorgan ble registrert. Tarestilkenes diameter ble målt i tverrsnitt tatt ca 1 cm over festeorganet, der stilkene er på sitt tykkeste. Alderen på de innsamlede tareplantene ble anslått ved å telle vekstsoner i tverrsnittene og samtlige tverrsnitt ble avfotografert for senere målinger av vekstsonearealene og rekonstruksjon av tareplantenes veksthistorikk. Tarestilkene har sin beste vekstsesong i vinterhalvåret og størrelsen (arealet) av hver enkelt vekstsone vil gi informasjon om tareplantenes vekst i en gitt sesong, der den indre ringen representerer veksten i plantenes første leveår (0-1 år), den nest innerste ringen veksten i andre leveår (1-2 år), etc, etc (Fig. 8). Arealet på en vekstsone (a) ble beregnet gjennom målinger av bredden fra tverrsnittets sentrum til indre (i) og ytre (y) vektsonegrense langs to akser med ca 90 graders vinkel og beregnet utifra formelen a = π y 2 – π i 2 .
Variasjoner knyttet til hvileperiodens lengde ble testet vha. multiple sammenligner (post-hoc Tukey tester) av faktornivåer (uhøstet vs. 5 år etter høsting vs. 8 år etter høsting), mens variasjoner mellom to enkelte faktornivåer ble undersøkt vha. t-tester.
Figur 8. Aldersberegninger gjøres ved telling av vekstsoner i tarestilkenes tverrsnitt og
årlig tykkelsestilvekst estimeres utifra størrelsen (arealet) av hver enkelt vekstsone.
2.4 - Modellering av stortarebiomasse
2.4.1 - Datagrunnlaget
Det ble i juni 2021 gjennomført videoundersøkelser på 200 tilfeldig utvalgte (randomiserte) stasjoner (punkt) for utvikling av en biomassemodell for stortare for høstefeltene i Nordland (Fig. 9). I juni 2022 ble det gjort tilsvarende observasjoner på ytterligere 80 randomiserte stasjoner for å validere modellens presisjonsnivå. På hver stasjon ble det gjort anslag av andel hardbunn, samt tetthet og høyde av stortarevegetasjonen per kvadratmeter. Vegetasjonsmålingene ble deretter omregnet til biomassetetthet av stortare per kvadratmeter ved å multiplisere den observerte plantetettheten med gjennomsnittlig vekt per plante etter samme metode som er benyttet ved tidligere biomassemodellstudier (Van Son et al. 2020, Norderhaug et al. 2021). Her beregnes stortareplantenes vekt ut fra plantehøyden som observeres på videoene ved å benytte en relasjon mellom plantehøyde og -vekt utviklet gjennom målinger av innsamlede stortareplanter.
Figur 9. Lokaliteter (punkt) undersøkt for utvikling av biomassemodell for stortare
i 2021 (røde symboler) og for å validere modellen i 2022 (blå symboler).
2.4.2 - Variabler
Fysiske variabler knyttet til terrenget i høstingsområdet ble valgt ut basert på kunnskap om relasjon mellom tarebiomasse og terrengkarakteristikker (Norderhaug et al. 2021). Vi vurderte dybde (m), substrat (hardbunnprosent), helning, krumning, topografisk posisjonsindeks (TPI) og terrengets ujevnhetsindeks (TRI). I tillegg vurderte vi effekten av bølgedynamikk på tarebiomassen (van Son et al. 2020) og inkluderte bølgehøyde og bølgeeksponering mot sjøbunn i valget av forklaringsvariabler. Før analysen tilpasset vi utvalget av variabler til et sett med kovariater som viste lav kollinearitet, ved først å velge variabler med lav korrelasjon ( Pearson’s R < 0,65) og deretter variabler som viste lav variasjonsinflasjonsfaktor (VIF < 3). Det endelige utvalget inkluderte: dybde, bølgehøyde, hardbunnsprosent, krumning og TRI. Bølgehøyde og bølgeeksponering mot sjøbunn hadde en høy korrelasjonskoeffisient, noe som indikerer høy kollinearitet mellom disse variablene. I tidligere biomassemodelleringsstudier har bølgeeksponering mot sjøbunn vært foretrukket fremfor bølgehøyde (van Son et al. 2020, Norderhaug et al. 2021), ettersom denne inkluderer både bølgehøyde, bølgelengde og bunndyp. For Nordland ble imidlertid bølgehøyde foretrukket framfor bølgeeksponering mot sjøbunn på grunn av bedre modellytelse målt ved Akaike Information Criterion (Akaike 1974) og mer homogen fordeling av residualer.
2.4.3 - Modellbygging
Basert på tidligere kunnskap (van Son et al. 2020, Norderhaug et al. 2021) startet vi analysen med en generalisert additiv modell (GAM) med en tweedie-fordeling som tilpasser seg responsvariabelens verdiintervall. Tweedie-fordelingen gjør det også mulig for modellen å håndtere observasjoner av biomassefravær uten å ty til nullinflasjonsmodellering. Den fullstendige modellformelen var som følger:
Der a er modellens skjæringspunkt, fi betegner de spesifikke utjevningsfunksjonene (penalized regression splines) for den i -te forklaringsvariabelen (dybde, substrat (hardbunnprosent), kurvatur, TRI (terrengets ujevnhetsindeks) og bølgehøyde). x og y er koordinatene til stasjonene (Northings og Eastings) og ε er den normalfordelte tilfeldige feilen (residual). Modellen er knyttet til den observerte biomassetettheten gjennom en logaritmisk koblingsfunksjon.
Vi implementerte først GAM-modellen med pakken «mgcv» (Wood et al. 2016). Tweedie-distribusjonsmodellens residualer ble kontrollert via randomiserte kvantilresidualer (Dunn & Smyth 2005). Vi brukte DHARMa-pakken (Hartig 2022) til å generere randomiserte kvantilresidualer og kontrollerte disse for mønstre, fordeling, spredning, forekomst av ekstremverdier, nullinflasjon og romlig autokorrelasjon. Kontrollen indikerte romlig autokorrelasjon i residualene (Morans I-test for avstandsbasert autokorrelasjon, p-verdi < 0,001). Tilstedeværelsen av romlig autokorrelasjon betyr at modellen bryter med forutsetningen om uavhengige feil, og må tas hensyn til i modellen for å kunne vurdere standardfeilene til modellparametrene korrekt (Zuur et al. 2017).
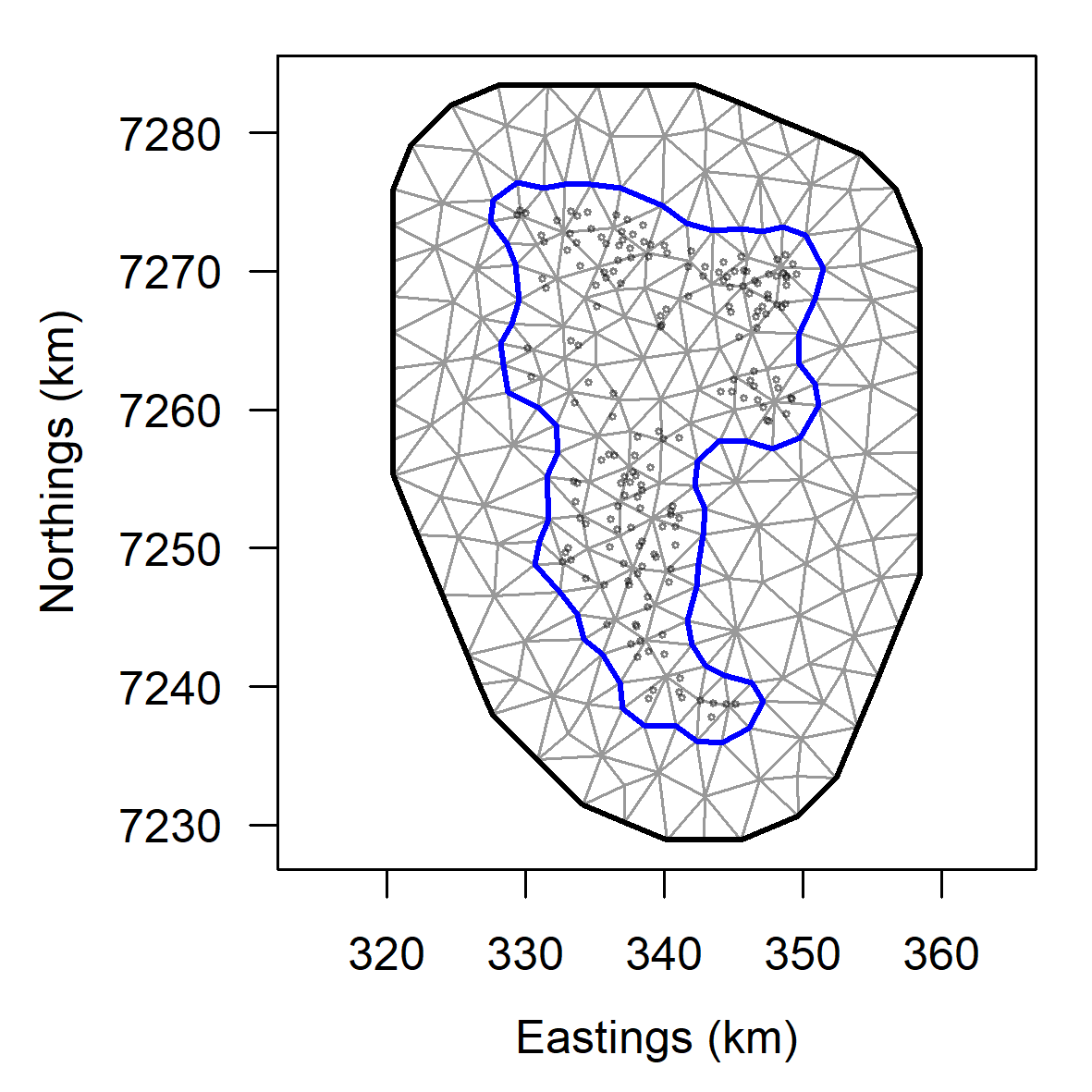
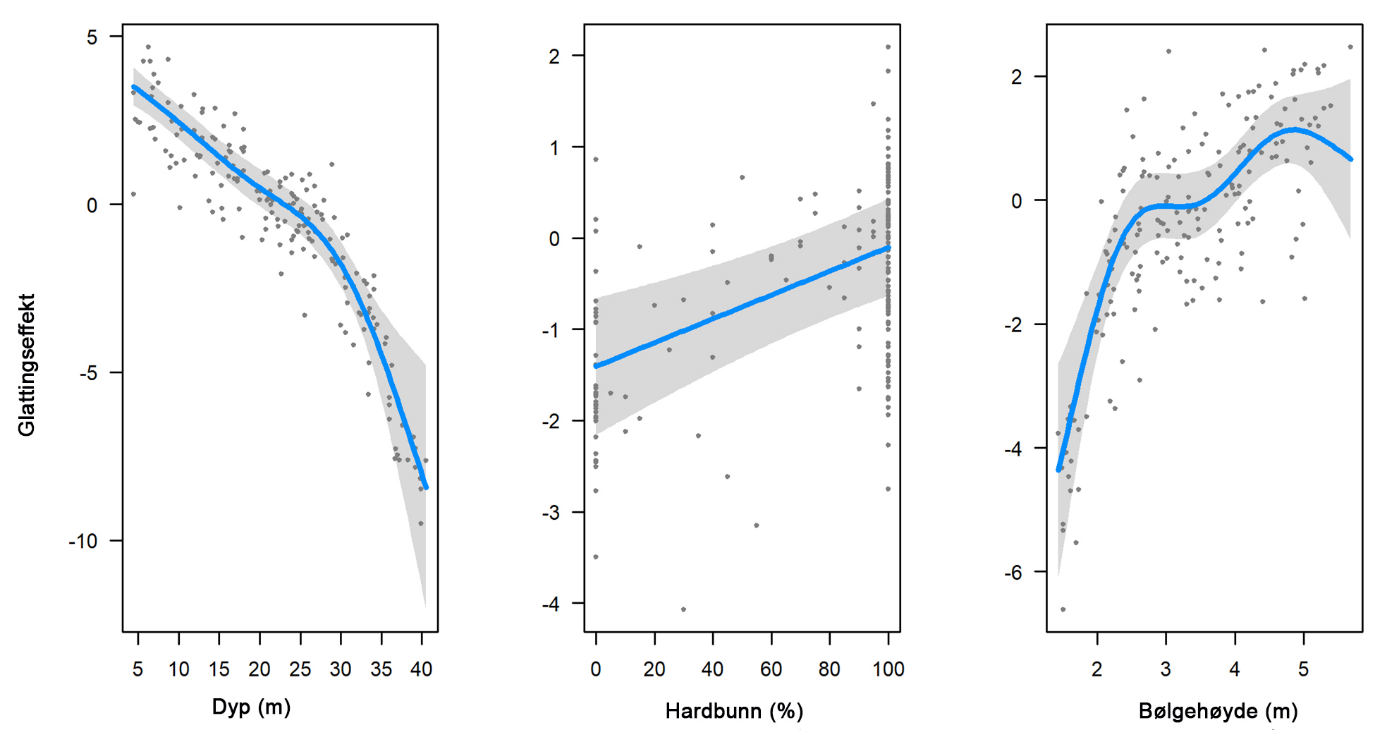
For å ta hensyn til romlig autokorrelasjon benyttet vi en blandet modell (general additive mixed model GAMM) og statistikkpakken sdmTMB (Anderson et al. 2024). Pakken sdmTMB implementerer et tilfeldig romlig felt basert på Matèrn-korrelasjonsfunksjonen og tar hensyn til romlig basert kovarians i residualene. Først definerte vi et nett der det romlige tilfeldige feltet (random spatial field) evalueres gjennom tesseleringer over prøvetakingsområdet (Fig. 10). Deretter kjørte vi en GAM-modell uten det tilfeldige romlige feltet og sammenlignet den med en modell med et tilfeldig romlig felt ved hjelp av AIC. Modellen med det tilfeldige romlige feltet hadde en lavere AIC-verdi og fravær av romlig autokorrelasjon i residualene. I et andre trinn valgte vi ut de mest relevante forklaringsvariablene gjennom en 7 gangers kryssvalidering (7-fold cross-validation). Resultatene viste en lav sum av sannsynlighetsfunksjonen (log-likelihood) for alle 7 valideringsrepetisjoner for modellvarianten som inkluderte dybde, substrat (hardbunnprosent) og bølgehøyde. Denne ble derfor valgt ut som beste modell for prediksjon av biomassetetthet. sdmTMB-pakken evaluerer ikke sannsynlighetsverdier knyttet til glattingsfunksjoner (smoothers), men forfatterne anbefaler en visuell analyse av disse med tilhørende konfidensintervall. Glattingsfunksjonene viste tydelig en signifikant reduksjon av biomassetetthet med økende dybde, mens biomassetettheten økte signifikant med økende bølgehøyde og hardbunnprosent (Fig. 11).
Figur 10. Todimensjonalt nett som definerer lokalitetene for estimering av tilfeldig
romlig felt (random spatial field) som skjæringspunktene mellom tesseleringstrianglene .
Figur 11: Glattingsfunksjoner som indikerer effekt på biomassetetthet av stortare (på logaritmisk skala) for de utvalgte miljøvariablene (dyp, hardbunnprosent, bølgehøyde). Grå areal markerer 95% konfidensintervall, mens punktene markerer observasjoner.
2.4.4 - Biomasseberegninger basert på beste utvalgt modell
Vi valgte ut den beste modellen som inkluderte dyp, hardbunnprosent og bølgehøyde for å predikere biomassetetthet av stortare i høsteområdet i Nordland på et rutenett (grid) med 50 meters oppløsning. Dybdedata (fra NGU-databasen) med en oppløsning på 50 m ble benyttet til biomassemodelleringen. Bølgehøyde (i form av 90% prosentil) ble estimert med kystbølgemodellen SWAN (https://swanmodel.sourceforge.io/home.htm) på et rutenett som hadde 200 m x 200 m oppløsning. Dette vil antakelig overestimere bølgehøyden i de områdene der mindre holmer, skjær og grunner egentlig vil dempe bølgeaktiviteten. I tillegg er bølgehøyden overestimert i forhold til helårsverdiene ettersom simuleringsperioden kun omfatter vintersesongen. Bølgedataene fra SWAN (200 m rutenett) ble bilineært interpolert til 50 m rutenett som ble brukt i biomassemodelleringen. Prosentandelen av hardbunn var ikke tilgjengelig for alle arealer i rutenettet, og ble beregnet ved hjelp av en lineær modell med en beta-fordeling med hardbunnprosent som responsvariabel og dybde og bølgehøyde som forklaringsvariabler (beste modell valgt basert på AIC-kriteriet).
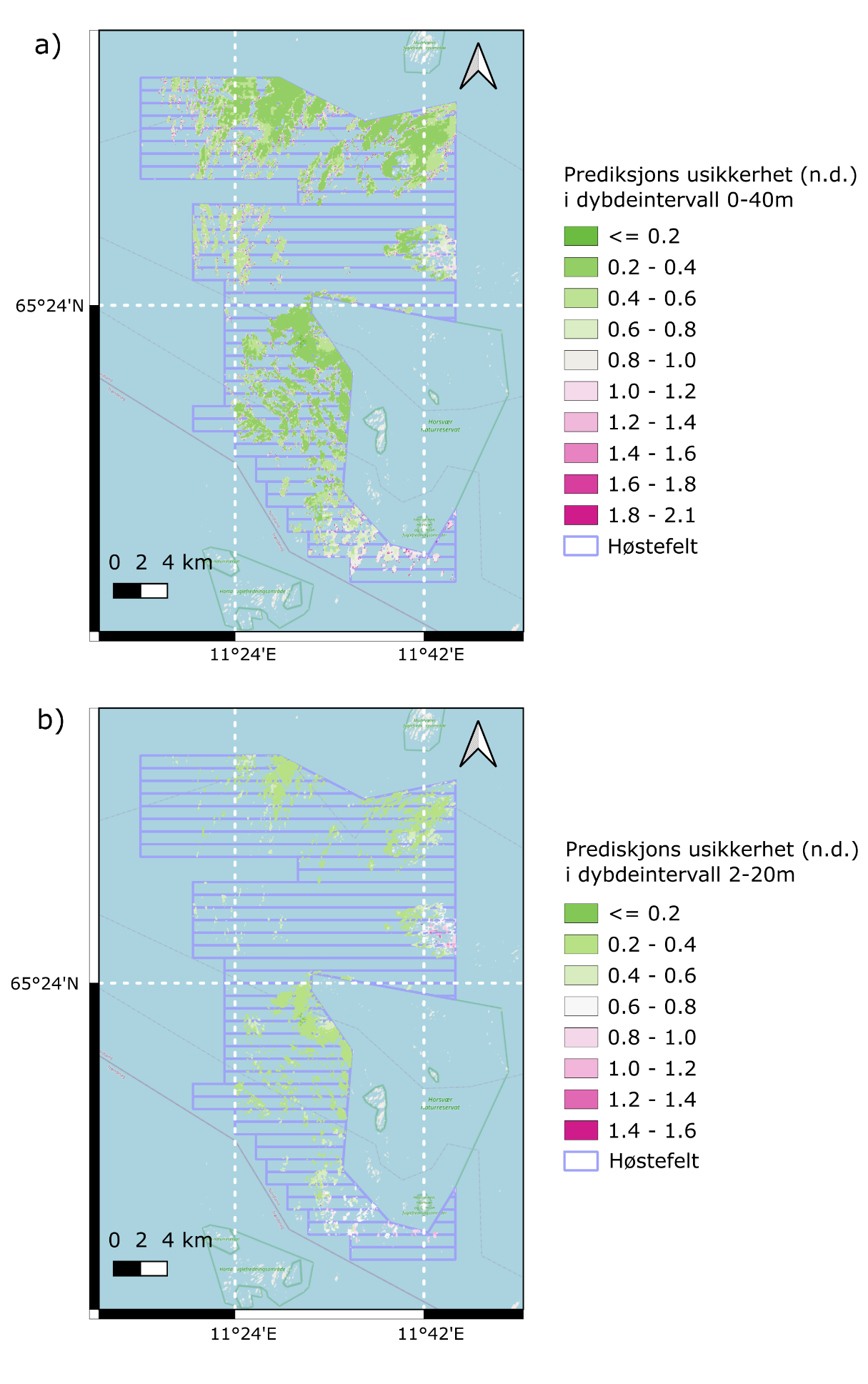
For å beregne usikkerheten av prediksjonene i hver av rutene ble det kjørt 500 repetisjonspredikeringer i koblingsrommet for presisjonsmatrisen som er knyttet til de ulike modellparameterne der usikkerheten måles i form av standardavviket til fordelingen i koblingsrommet på logaritmisk skala. Prediksjonene ble gjort for to ulike dybdeintervall: 0-40 m dyp som dekker hele utbredelsesdypet for stortare og 2-20 m dyp som dekker dybdeintervallet som er tilgjengelig for tarehøsting (Lovdata 2004). Prediksjonene viste høyest grad av usikkerhet i den sørligste del av modelleringsområdet, samt den østlige delen rett nord for Horsvær verneområde (Fig. 12).
Vi estimerte deretter den totale biomassen av stortare i hvert høstefelt ved å summere medianverdiene for den predikerte biomasstettheten på hver rutenettlokasjon og konvertere disse fra kg m -2 til tonn biomasse ved å bruke de respektive høstingsfeltenes areal som sammenleggingsfaktor. For å konstruere 95% konfidensintervall ble tilsvarende summeringer gjort for henholdsvis nedre 2,5 % og øvre 97,5 % prosentiler av ruteprediksjonene.
Figur 12. Romlig usikkerhet (variasjonskoeffisient) for prediksjoner av biomassetetthet av
stortare i a) 0-40 m dyp og b) 2-20 m dyp.
3 - Resultater
3.1 - Effekter av tidligere prøvehøstinger av stortare
3.1.1 - Videoundersøkelser
Videoobservasjoner etter prøvehøstingen i Nordland i 2017 viste at kronesjikthøyde og biomassetetthet av stortare var reetablert 5 år, men ikke 3 år, etter høsting. Kråkebolleforekomstene har ikke vært påvirket av tarehøstingen og kråkebollebeiting har ikke påvirket reetableringen av tare på høstefeltene. Forekomstene av rød kråkebolle var i 2020 og 2022 lavere enn det som ble observert før prøvehøstingen i 2017. Det ble ikke observert forekomster av grønn kråkebolle på noen av de undersøkte stasjonene i perioden 2017-22.
Den siste prøvehøstingen av stortare i Nordland ble gjennomført i 2017, da totalt 12.837 tonn ble høstet fordelt på 4 områder. I alt 12 videotransekter i prøvehøstefeltene og 5 videotransekter i uhøstede referanseområder (Fig. 3) har blitt fulgt opp med etterundersøkelser i 2020 og 2022 for å studere gjenvekst av stortarevegetasjon og utvikling av kråkebolleforekomster.
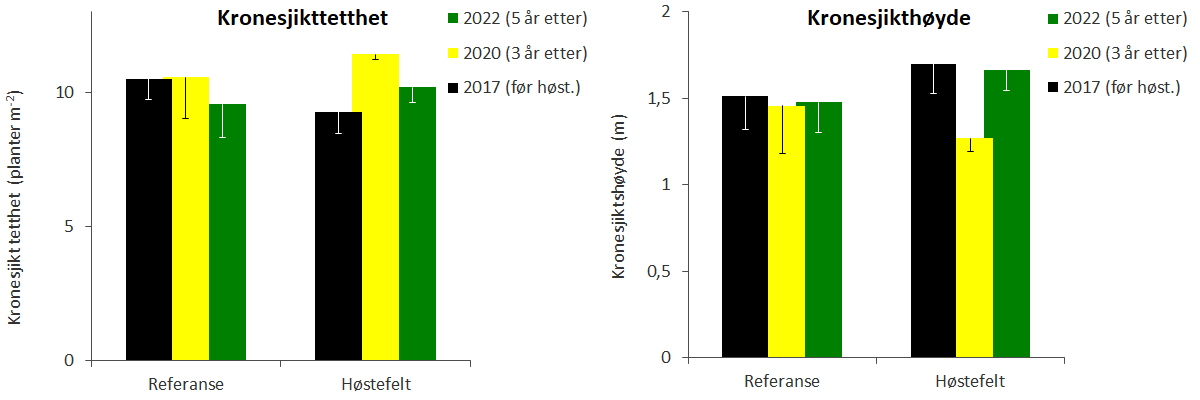
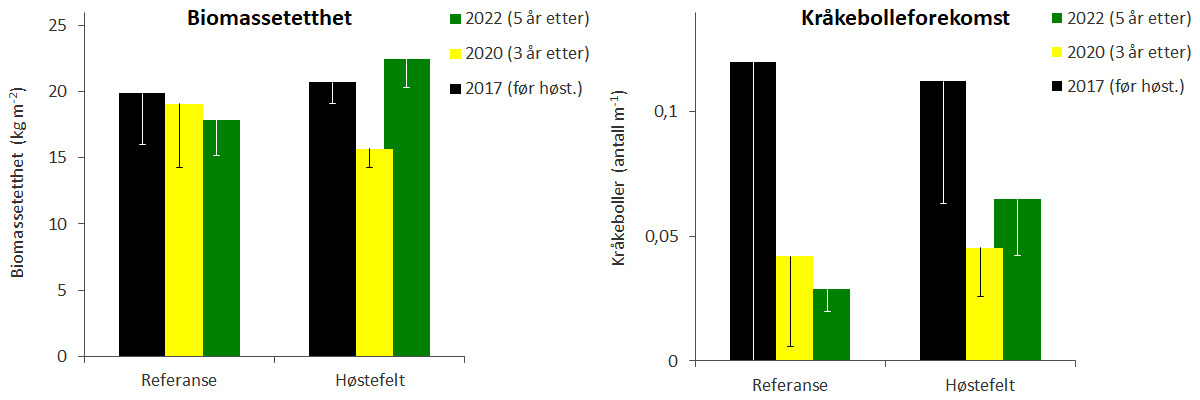
Observasjonene gjennomført i 2020 indikerer høyere tetthet av stortareplanter, men lavere kronesjikthøyde og biomassetetthet på høstefeltene enn det som ble observert før prøvehøstingen tre år tidligere (Fig. 13-15, Tab. 2). Observasjonene fra 2022 viste at stortarevegetasjonens kronesjikthøyde og biomassetetthet var reetablert 5 år etter prøvehøstingen. Registreringer av kråkeboller langs videotransektene indikerer at forekomstene i 2020 og 2022 var lavere enn i 2017 (Fig. 15, Tab. 2). Det ble ikke registrert signifikante forskjeller i utviklingen av kråkebolleforekomster mellom stasjoner i høstefelt og referanseområder, og kråkebollebeiting har ikke påvirket reetableringen av tare på høstefeltene. Det ble kun observert individer av rød kråkebolle (Echinus esculentus), mens grønn kråkebolle (Strongylocentrotus droebachiensis) ikke ble observert langs noen av de undersøkte videotransektene i perioden 2017-22.
Figur 13. Gjennomsnittlig tetthet (venstre panel) og høyde (høyre panel) av stortarevegetasjonens kronesjikt observert før høsting i 2017 (sorte søyler), 3 år etter høsting i 2020 (gule søyler) og 5 år etter høsting i 2022 (grønne søyler) på stasjoner i høstefelt og referanseområder i Nordland. Nedre grense for 95 % konfidensintervall er tegnet inn.
Figur 14. Eksempler på stortarevegetasjon fra felt 484F før prøvehøstingen i 2017 (øvre bilde),
3 år etter prøvehøsting i 2020 (midtre bilde) og 5 år etter prøvehøsting i 2022 (nedre bilde).
Figur 15. Gjennomsnittlig biomassetetthet av stortare (venstre panel) og registreringsfrekvens av kråkeboller (høyre panel) før høsting i 2017 (sorte søyler), 3 år etter høsting i 2020 (gule søyler) og 5 år etter høsting i 2022 (grønne søyler) på stasjoner i høstefelt og referanseområder i Nordland. Nedre grense for 95 % konfidensintervall er tegnet inn.
Tabell 2. Lineære sammensatte modeller («linear mixed-effect models») for videodata med felttype (høstefelt vs referanse) krysset med periode (før høsting (2017) vs etter 3 år etter høsting (2020) vs 5 år etter høsting (2022) som uavhengige variabler. Modellen ble kjørt for responsvariablene (i kursiv): Kronesjikttetthet, kronesjikthøyde, tarebiomasse og kråkebolleforekomst. Signifikante sannsynlighetsverdier (p < 0,05) for interaksjonsleddene (hvis signifikans indikerer effekt av høsteaktiviteten) er uthevet.
Responsvariabel
Effekt
Estimat
t verdi
p verdi
Kronesjikttetthet (per m-2)
(Skjæringspunkt)
10,27
20,62
<0,001
Periode (2017 vs 2020)
-0,214
-0,421
0,677
Periode (2017 vs 2022)
-1,230
-2,413
0,022
Felttype (referanse vs høstefelt)
-0,839
-1,414
0,167
Interaksjon (2020 x høstefelt)
2,200
3,628
0,001
Interaksjon (2022 x høstefelt)
1,939
3,197
0,003
Kronesjikthøyde (m)
(Skjæringspunkt)
1,478
14,37
<0,001
Periode (2017 vs 2020)
-0,019
-0,217
0,830
Periode (2017 vs 2022)
0,009
0,105
0,917
Felttype (referanse vs høstefelt)
0,192
1,567
0,130
Interaksjon (2020 x høstefelt)
-0,389
-3,818
0,015
Interaksjon (2022 x høstefelt)
-0,034
-0,331
0,743
Tarebiomasse (kg m-2)
(Skjæringspunkt)
18,99
8,823
<0,001
Periode (2017 vs 2020)
-0,493
-0,337
0,738
Periode (2017 vs 2022)
-1,717
-1,176
0,249
Felttype (referanse vs høstefelt)
1,585
0,619
0,543
Interaksjon (2020 x høstefelt)
-4,546
-2,615
0,014
Interaksjon (2022 x høstefelt)
3,185
1,832
0,077
Kråkebolleforekomst (per m-1)
(Skjæringspunkt)
0,23
5,074
<0,001
Periode (2017 vs 2020)
-0,120
-2,180
0,037
Periode (2017 vs 2022)
-0,162
-2,943
0,006
Felttype (referanse vs høstefelt)
-0,096
-1,776
0,083
Interaksjon (2020 x høstefelt)
0,033
0,496
0,623
Interaksjon (2022 x høstefelt)
0,095
1,455
0,160
3.1.2 - Tareinnsamlinger
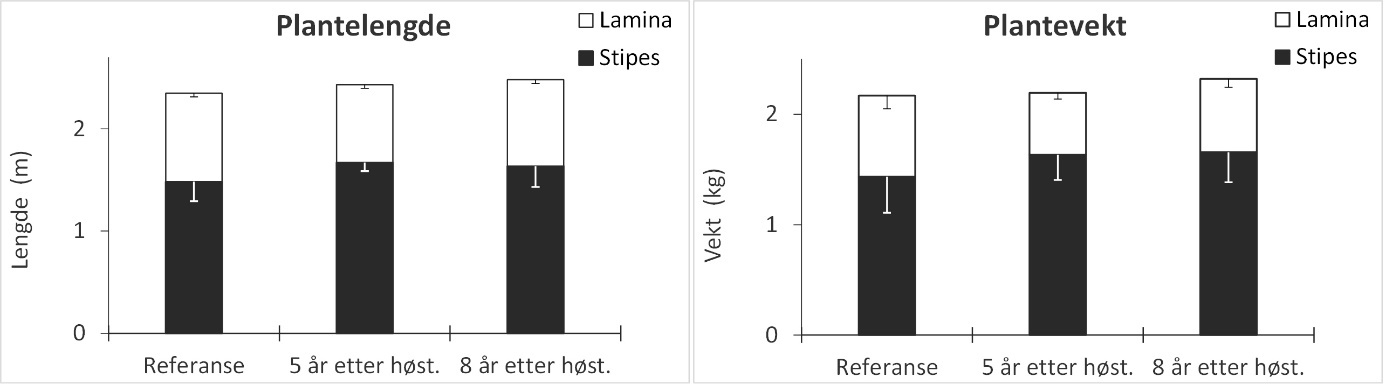
Stortarepopulasjonene samlet inn på stasjoner høstet 5 år tidligere var dominert av yngre aldersgrupper enn på uhøstede referansestasjoner. Det ble ikke registrert signifikante forskjeller i størrelse og vekt mellom stortareplanter fra referanseområdene og felt høstet 5 år tidligere. Vekstsoneanalyser viste at den gjennomsnittlige årlige stilktilveksten hos stortare har vært høyere på stasjonene høstet 5 år tidligere enn på referansestasjonene. Sammenlignet med referansestasjonene var den gjennomsnittlige epifyttbiomassen på tarestilkene ca 50 % og epifyttdiversiteten signifikant lavere 5 år etter høsting. Epifyttforekomstene økte med tareplantenes alder og hadde kraftigst utvikling mellom 6 år og 8 år. Stortareplanter samlet inn på stasjoner høstet 8 år tidligere viste ingen signifikante forskjeller fra uhøstede referansestasjoner mhp alder, størrelse og epifyttsammensetning.
Det ble i perioden 2022-24 samlet inn stortare på 3 stasjoner prøvehøstet 5 år tidligere, 12 stasjoner prøvehøstefelt 8 år tidligere og 8 stasjoner i uhøstede referanseområder for sammenligninger av aldersstruktur, morfologi, vekst og epifyttforekomster mellom områder med ulik høstehistorikk. I tillegg ble forekomst av juvenile kråkeboller i tareplantenes festeorgan (hapterkråkeboller) registrert som en indikator på kråkebollerekruttering.
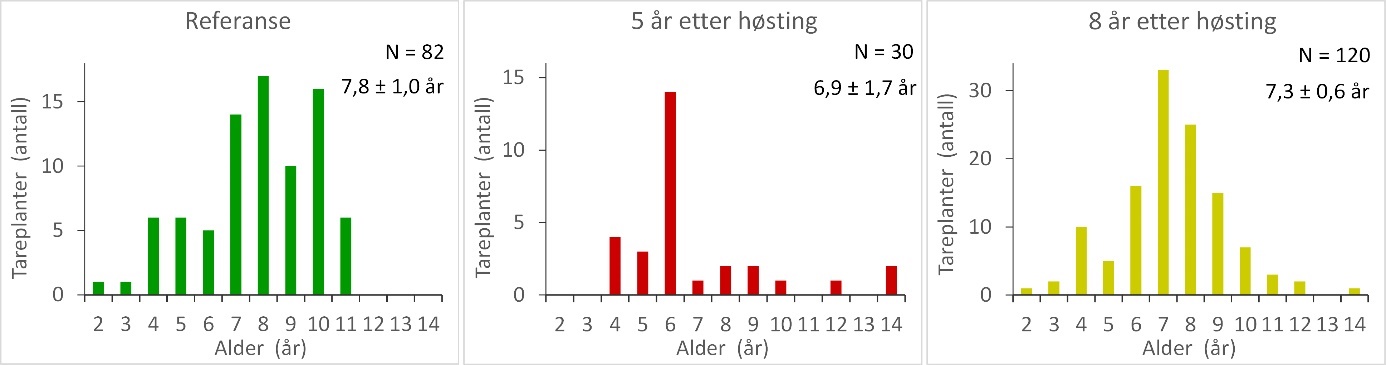
Innsamlingsdypet varierte fra 5 – 8 m og bølgeeksponeringen (MSWH) fra 0,5 – 1,1 m mellom stasjonene. Det var ingen signifikante forskjeller mellom stasjoner med ulik høstehistorikk og referansestasjoner med hensyn til innsamlingsdyp og bølgeeksponering (Tukey tester i Appendix 5). Det ble ikke registrert signifikante forskjeller i gjennomsnittlig alder for tareplantene samlet inn i de tidligere høstede områdene og referanseområdene (Appendix 5). Likevel viser aldersfordelingsdiagrammene at stortarepopulasjonene på stasjonene som ble høstet 5 år tidligere var dominert av yngre aldersgrupper enn stortarepopulasjonene på stasjonene høstet 8 år tidligere og i referanseområdene (Fig. 16). På stasjonene som ble høstet 5 år tidligere var f.eks 70 % av tareplantene yngre enn 7 år, mens kun 28 % av tareplantene på stasjonene som ble høstet 8 år tidligere og 23 % av tareplantene på referansestasjonene var yngre enn 7 år. Det ble ikke registrert signifikante forskjeller i stortareplantenes størrelse og vekt mellom de tidligere høstede stasjonene og referansestasjonene (Fig. 17, Appendix 5).
Figur 16. Aldersfordeling av stortarepopulasjoner samlet inn på stasjoner i uhøstede referanseområder (venstre panel), felt som ble høstet 5 år tidligere (midtre panel) og felt som ble høstet 8 år tidligere (høyre panel). Gjennomsnittlig alder ± 95 % konfidensintervall og totalt antall innsamlede planter (N) er vist i hvert diagram.
Figur 17. Gjennomsnittlig stilk (stipes) og blad (lamina) lengder (venstre panel) og gjennomsnittlig stilk- og bladvekt (høyre panel) for stortareplanter samlet inn på stasjoner i uhøstede referanseområder, felt som ble høstet 5 år tidligere og felt som ble høstet 8 år tidligere. Nedre grense for 95 % konfidensintervall er tegnet inn.
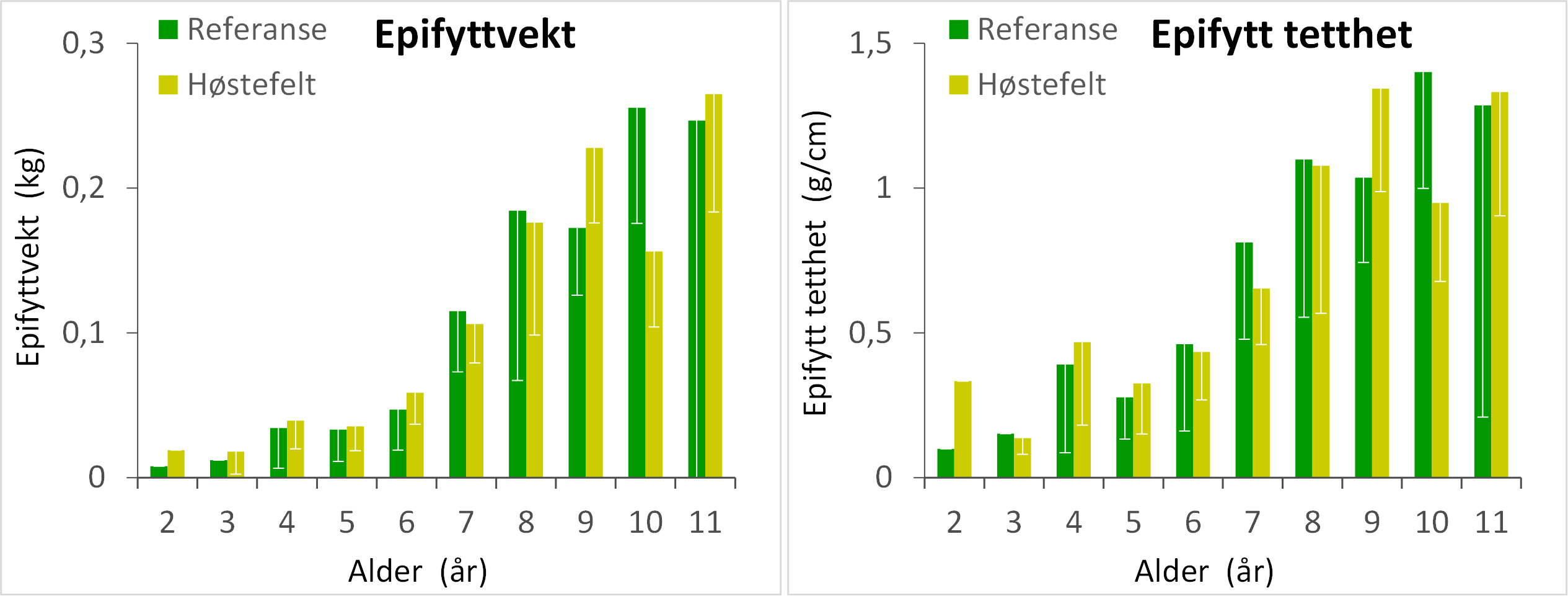
Den gjennomsnittlige epifyttbiomassen på tarestilkene samlet inn på stasjoner høstet 5 år tidligere (82 ± 65 g) var ca halvparten av den gjennomsnittlige epifyttbiomassen på referansestasjonene (157 ± 85 g), men likevel ikke signifikant forskjellig pga stor variasjon i epifyttbiomasse mellom enkeltstasjoner (Appendix 5). For stortareplanter samlet inn på stasjonene høstet 8 år tidligere var den gjennomsnittlige epifyttbiomassen 129 ± 35 g og ikke signifikant forskjellig fra referansestasjonene.
Det var en signifikant positiv korrelasjon mellom epifyttvekt og alder (r = 0,46, p < 0,001) og mellom epifytt tetthet (i gram epifytter per cm tarestilk) og alder (r = 0,39, p < 0,001). Sammenhengen mellom tareplantenes alder og epifytt tetthet indikerer at epifyttutviklingen også er avhengig av andre aldersrelaterte faktorer enn tarestilkenes størrelsesutvikling. Den kraftigste utviklingen finner sted mellom 6 og 8 års alder da epifyttvekten tredobles og epifytt tettheten mer enn dobles (Fig. 18). Det var ingen signifikante forskjeller i epifyttvekt og epifytt tetthet mellom tareplanter samlet inn i feltene høstet 8 år tidligere og referanseområdene for de sammenlignbare aldersgruppene. Tareprøvene samlet inn på felt høstet 5 år tidligere inneholdt få planter som var eldre enn 6 år og er derfor ikke tatt med i Fig.18. Det var imidlertid ingen signifikante forskjeller i epifyttbiomasse mellom tareplanter samlet inn i feltene høstet 5 år tidligere og i referanseområder for aldersgruppene 4 år (t-test, p = 0,229), 5 år (t-test, p = 0,333) eller 6 år (t-test, p = 0,619).
Figur 18. Gjennomsnittlig epifyttvekt (venstre panel) og epifytt tetthet i form av gram epifytter per cm tarestilk (høyre panel) for ulike aldersgrupper av stortareplanter samlet inn på stasjoner i felt høstet 8 år tidligere og i uhøstede referanseområder. Nedre grense for 95 % konfidensintervall er tegnet inn.
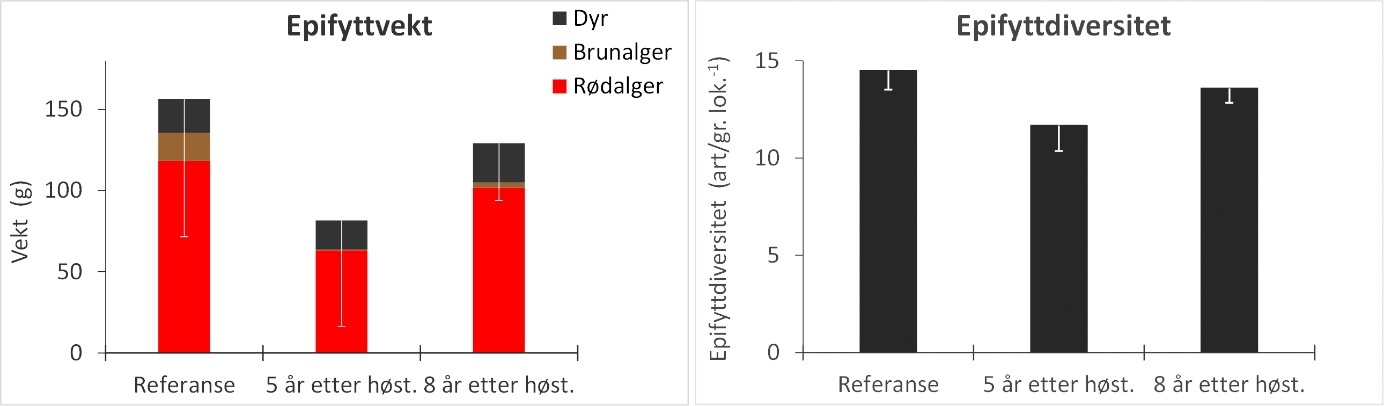
Epifytter på innsamlede stortarestilker ble bestemt til art/slekt/gruppe og rangert etter forekomst. Epifyttsamfunnene på de fleste innsamlede tarestilkene domineres av tråd- og bladformede rødalger som i gjennomsnitt utgjør ca 75 % av epifyttbiomassen (Fig. 19). Den gjennomsnittlige epifyttdiversiteten på stasjonene var signifikant lavere på feltene som ble høstet 5 år tidligere enn i referanseområdene, mens det ikke ble registrert forskjeller i epifyttdiversitet mellom stasjoner i felt som ble høstet 8 år tidligere og referanseområder (Fig. 19, Appendix 4). Eksempler på epifyttstruktur på tarestilker fra stasjoner i et referanseområde og felt høstet henholdsvis 5 og 8 år tidligere er vist i Fig. 20.
Figur 19. Gjennomsnittlig vekt av epifytter (venstre panel) og epifyttdiversitet per lokalitet (høyre panel) for stortareplanter samlet inn på stasjoner i uhøstede referanseområder, felt som ble høstet 5 år tidligere og felt som ble høstet 8 år tidligere. Den totale høyden av søylene i venstre panel angir total epifyttvekt, mens det relative bidraget fra henholdsvis rødalger, brunalger og dyr til epifyttbiomassen indikeres av høyden på de fargede søylesegmentene. Nedre grense for 95 % konfidensintervall for total epifyttvekt og diversitet er tegnet inn.
Figur 20. Eksempler på epifyttstruktur på stortarevegetasjonen i et referanseområde (Horsvær)
og felt som ble høstet henholdsvis 5 år (475B) og 8 år (477C) tidligere.
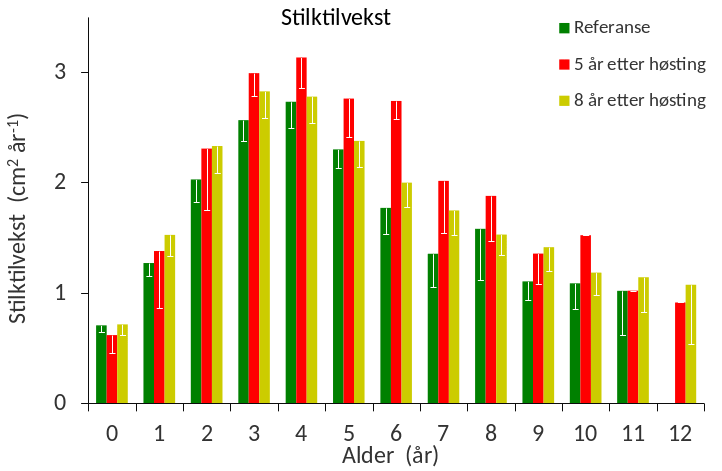
Den årlige stilktilveksten beregnet ut fra arealet av åringene i stilktverrsnittene indikerer at stortareplantene har sin mest intensive vekst ved ca 3-4 års alder (Fig. 21). Vekstsoneanalysene viste også at stilktilveksten for de fleste sammenlignbare årsklasser har vært jevnt over noe høyere på de tidligere høstede stasjonene enn på referansestasjonene.
Juvenile individer av grønn kråkebolle (S. droebachiensis) ble registrert i tareplantenes festeorgan (hapter) på 21 av 23 innsamlingsstasjoner i 2022-24. Den gjennomsnittlige registreringsfrekvensen lå på 0,4 ± 0,1 kråkeboller per tarehapter og det ble ikke registrert signifikante forskjeller mellom referansestasjoner og tidligere høstede stasjoner (Appendix 4).
Figur 21 Gjennomsnittlig årlig stilktilvekst (i form av vekstsoneareal) for ulike aldersgrupper
av stortareplanter samlet inn på referansestasjoner (grønne søyler), stasjoner høstet 5 år
tidligere (røde søyler) og stasjoner høstet 8 år tidligere (gule søyler). Nedre grense for 95 %
konfidensintervall er tegnet inn.
3.2 - Undersøkelser før og etter oppstart av kommersiell stortarehøsting
3.2.1 - Videoundersøkelser
Video- og akustikkstasjoner ble etablert i samtlige høstefelt i Nordland i forkant av oppstart av kommersiell tarehøsting i 2022 og fulgt opp med nye undersøkelser i 2023 og 2024. Med unntak av de fire sørligste feltene ble det registrert tette forekomster av høyvokst stortarevegetasjon på de fleste stasjoner i 2022. Rekoloniseringen og tilveksten av stortare på feltene som ble høstet i 2022 har vært god, mens feltene som ble høstet i 2023 har hatt mer variabel tilvekst av stortare og økning i forekomster av brunalgen draugtare året etter høsting. Registreringsfrekvensen av rød kråkebolle indikerer at forekomstene er på et moderat nivå og beiting fra kråkeboller har hatt liten effekt på reetableringen av tare på høsteflatene. Grønn kråkebolle er ikke observert på noen av videostasjonene i perioden 2022-24.
I forkant av oppstarten av kommersiell tarehøsting i Nordland i 2022 ble det etablert 1-3 video- og akustikkstasjoner (transekter) i hvert av de 40 høstefeltene, samt i uhøstede referanseområder (Fig. 3). Ettersom tarevegetasjonen i 2022 var reetablert etter tidligere prøvehøstinger (Fig. 15, Tab. 2), vil disse observasjonene kunne benyttes som sammenligningsgrunnlag for å vurdere effektene av den framtidige kommersielle tarehøstingsaktiviteten i dette området. Bølgeeksponeringen (MSWH) varierte fra 0,4 – 1,3 m mellom transektene, og det ble ikke registrert signifikante forskjeller i bølgeeksponering mellom transekter i høstefelt og referanseområder (t-test, p = 0,903).
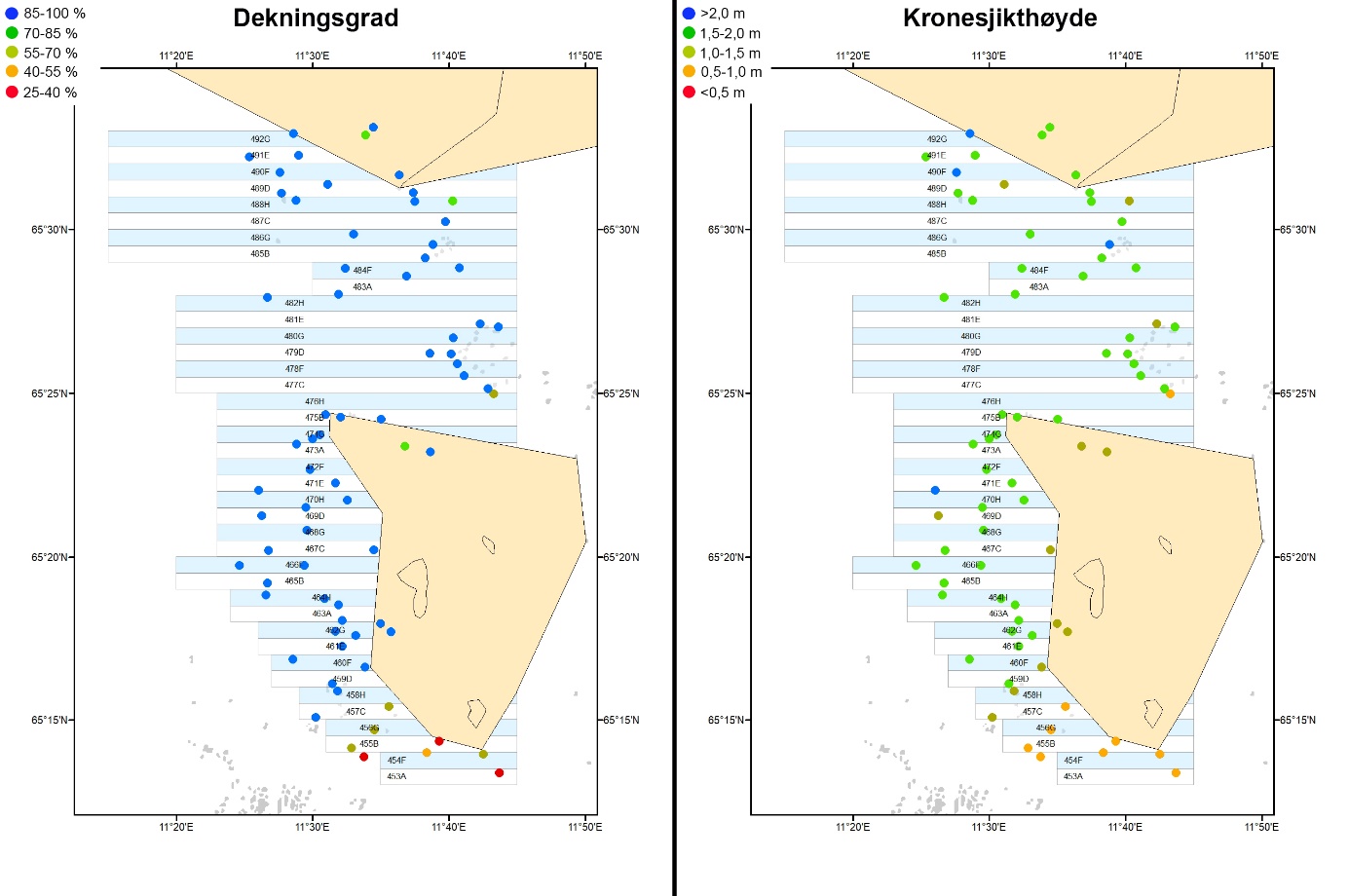
Videoundersøkelsene gjennomført i 2022 viste at stortare dominerte bunnvegetasjonen med en gjennomsnittlig dekningsgrad på >85 % og kronesjikthøyde på >1,5 m på de fleste stasjoner (Fig. 22). Unntaket er stasjonene i de fire sørligste høstefeltene og felt 476H rett nord for Horsvær der den gjennomsnittlige dekningsgraden av stortare var <70 % og kronesjikthøyden <1 m. På de sørligste feltene dominerte stedvis andre arter som butare, sukkertare og draugtare bunnvegetasjonen langs transektene (Fig. 23). Det ble også registrert en signifikant positiv korrelasjon mellom grad av bølgeeksponering og stortarevegetasjonens dekningsgrad (r = 0,29, p = 0,014) og kronesjikthøyde (r = 0,39, p > 0,001).
Figur 22. Gjennomsnittlig kategorisert dekningsgrad (venstre kart) og kronesjikthøyde (høyre kart) av stortare på stasjoner undersøkt ved hjelp av undervannsvideo i 2022. Dekningsgrad for stortare er estimert som prosentandel dekket bunnflate og kronesjikthøyden estimert i meter. Intervallinndeling for kategorier er listet øverst til venstre i kartene.
Figur 23. Eksempler på områder med innslag av butare (Alaria esculenta) på felt
454F (øvre bilde), draugtare (Saccorhiza polyschides) på felt 455B (midtre bilde)
og sukkertare (Saccharina latissima) på felt 456G (nedre bilde).
I 2022 ble høstefeltene i kategori A og F åpnet for tarehøsting og det ble til sammen høstet 28.531 tonn stortare. Uttaket varierte mellom de enkelte feltene fra 0 på de to sørligste feltene (453A, 454F) til 9.216 tonn på 473A-feltet (Appendix 6). Det ble i juni 2023 gjennomført oppfølgende video- og akustikkundersøkelser på samtlige stasjoner som også ble undersøkt før høstingen i 2022. Undersøkelsene i 2023 foregikk samtidig med den pågående høstingen av B-feltene (som var åpne for høsting i perioden juni-august 2023) som ikke var sluttført på undersøkelsestidspunktet. Selv om det ble observert ferske trålspor på flere av stasjonene i B-feltene, gir ikke observasjonene i 2023 noe komplett bilde av høstepåvirkningen i disse feltene.
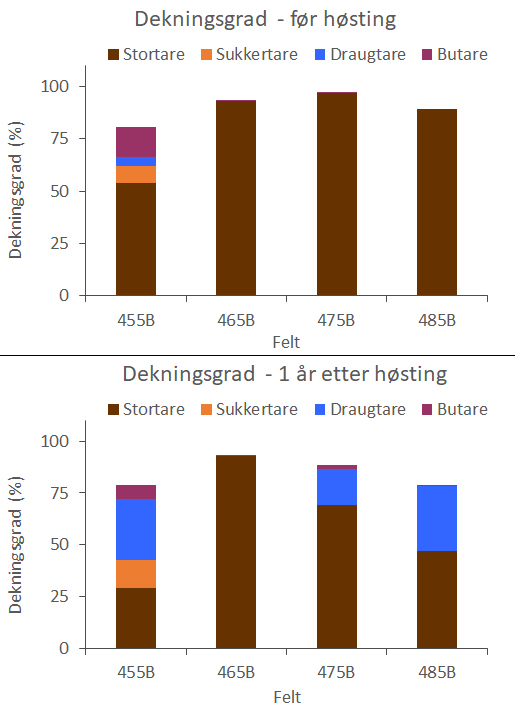
Det ble observert trålspor i samtlige høstede A- og F-felt i 2023, og dekningsgraden av stortare i de ett år gamle trålsporene er betydelig høyere enn i trålsporene som ble observert på de nylig høstede B-feltene (Fig. 24). Selv om den reetablerende stortarevegetasjonen i de årsgamle trålsporene fortsatt var kortvokst (ca 0,3 m høy) var den gjennomsnittlige dekningsgraden av stortare på disse feltene > 80 %.
Figur 24. Eksempler på tilstand i trålspor observert langs videotransekter i juni 2023 på nylig høstede felt (B felt til venstre) og felt høstet ett år tidligere (A og F felt til høyre).
Det ble i juni 2024 gjennomført oppfølgende video- og akustikkobservasjoner langs de samme transektlinjene som ble undersøkt i 2022 og 2023. Det innrapporterte høstekvantumet for B feltene i 2023 var totalt på 11.699 tonn stortare og varierte fra 1.048 tonn på felt 455B til 4.479 tonn på felt 485B (Appendix 6). Det ble observert trålspor i samtlige fire B-felt, og året etter høstingen var den gjennomsnittlige dekningsgraden av stortare på disse feltene ca 60 % og signifikant lavere (t-test, p = 0,019) enn året etter høstingen av A- og F-feltene. Dette har sammenheng med en økning i forekomster av draugtare (Saccorhiza polyschides) på tre av B-feltene i 2024 (Fig. 25, 26). På B-feltene som ble høstet i 2023 har gjennomsnittlig dekningsgrad av draugtare økt signifikant (t-test, p = 0,010) fra 1 % før høsting til 20 % året etter høsting. På feltene som ble høstet i 2022 (A og F) er det ikke registrert økninger i forekomst av draugtare fra før høsting (2022) til etter høsting (verken i 2023 eller 2024) og den gjennomsnittlige dekningsgraden av draugtare på disse feltene har vært <1 % i hele perioden. To år etter høstingen av A- og F-feltene har stortarevegetasjonen en dekningsgrad på >90 % og vegetasjonshøyden langs trålsporene er doblet fra året før (Fig. 27). Undersøkelsene i 2024 foregikk samtidig med den pågående høstingen av C-feltene, og det ble observert ferske trålspor på samtlige av disse allerede i løpet av de to første ukene etter at feltene var åpnet for høsting (Fig. 28).
Figur 25. Dekningsgrad av stortare, sukkertare, draugtare og butare på B-felt,
henholdsvis før høsting (øvre panel) og et år etter høsting (nedre panel).
Feltene er listet fra sør (venstre) mot nord (høyre).
Figur 26. Eksempler på tilstand i trålspor observert langs videotransekter i juni 2024 et år etter høsting (B-felt til venstre) og to år etter høsting (A- og F-felt til høyre). Bildene er tatt langs samme transekter som i Fig. 21a. Tre av B-feltene (455B, 475B og 485B) høstet i 2023 domineres året etter stedvis av draugtare (Saccorhiza polyschides).
Figur 27. Eksempler på tilstand i trålspor observert langs videotransekter i juni 2024, to år etter høstingen av felt 472F og 473A. Høstekvantumet av stortare på disse feltene i 2022 var henholdsvis 5.833 tonn (472F) og 9.216 tonn (473A).
Figur 28. Eksempler på trålspor observert langs videotransekter på nylig høstede C-felt i juni 2024.
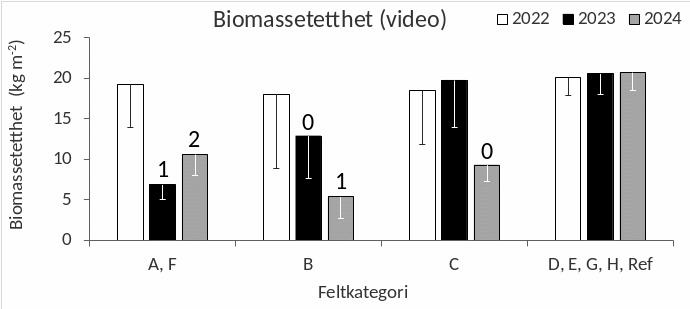
Den gjennomsnittlige biomassetettheten av stortare beregnet fra videoobservasjonene lå før høsteoppstart på 18-20 kg m -2 (Fig. 29). Året etter høsting var den gjennomsnittlige biomassetettheten 7 kg m -2 for A- og F-feltene og 5 kg m -2 for B-feltene, noe som tilsvarer en reduksjon på 64 % for A- og F-feltene og 70 % for B-feltene i forhold til førhøstenivået. To år etter høstingen av A- og F-feltene hadde den gjennomsnittlige biomassetettheten av stortare økt til 11 kg m -2 og reduksjonen i forhold til førhøstenivået var da 45 %. Selv om det ble observert reduksjoner i biomassetetthet i forbindelse med den pågående høstingen av B-feltene (i 2023) og C-feltene (i 2024) gir ikke disse estimatene noe komplett bilde av biomassereduksjonene her da høstingen av disse feltene ikke var sluttført på undersøkelsestidspunktet. Det ble ikke observert endringer i biomassetetthet av stortare i perioden 2022-24 for uhøstede felt og referanseområder (Fig. 29).
Figur 29. Gjennomsnittlig biomassetetthet av stortare ( Laminaria hyperborea ) langs videotransekter i årene 2022, 2023, 2024 i felt som ble høstet sommeren 2022 (A, F), sommeren 2023 (B), sommeren 2024 (C) og uhøstede felt (D, E, G, H, Referanse). Tallene over kolonnene indikerer tidsrom mellom høsting og observasjon (0: Pågående høsting, 1: Et år etter høsting, 2: To år etter høsting). Nedre grense for 95 % konfidensintervall er tegnet inn.
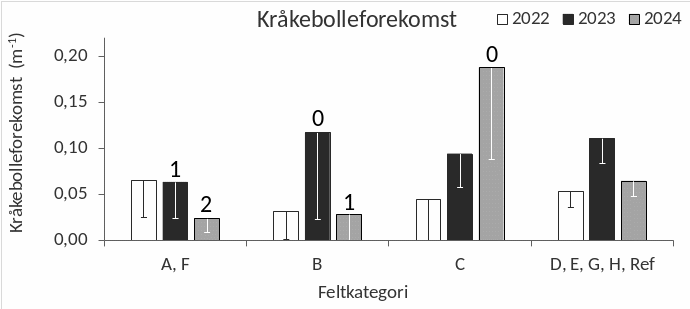
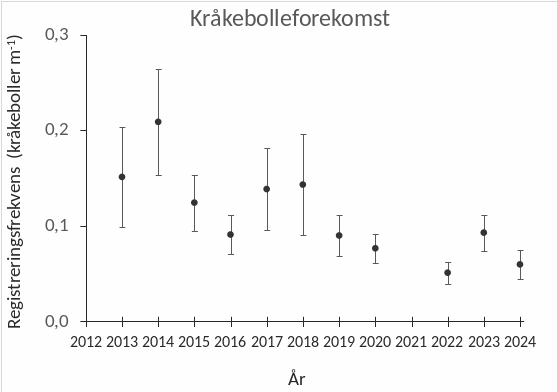
Kråkeboller registreres gjennom visuelle observasjoner der registreringsfrekvensen av ulike arter per transektmeter benyttes som et mål på forekomst. Det ble ikke observert individer av grønn kråkebolle (Strongylocentrotus droebachiensis) langs noen av videotransektene undersøkt i perioden 2022-24, mens rød kråkebolle (Echinus esculentus) observeres i de fleste områder (Fig. 30). Det ble registrert høyere forekomster av rød kråkebolle i 2023, sammenlignet med registreringene i 2022 (t-test, p = 0,001) og 2024 (t-test, p = 0,049). Det var også en tendens til økning i registreringsfrekvensen av kråkeboller på feltene som var under høsting på observasjonstidspunktene i 2023 (B-felt) og 2024 (C-felt), mens registreringsfrekvensen året etter høsting var på samme nivå som før høsting (Fig. 31). En sammenligning mellom videoobservasjoner gjennomført i sørlig del av Nordland i perioden 2013-24 indikerer at kråkebolleforekomstene i denne regionen har vist en nedadgående trend i denne perioden (Fig. 32).
Figur 30. Eksempler på rød kråkebolle (Echinus esculentus) beitende på epifytter på stortarestilker.
Figur 31. Gjennomsnittlig registreringsfrekvens av rød kråkebolle (Echinus esculentus) langs videotransekter i årene 2022, 2023, 2024 i felt som ble høstet sommeren 2022 (A, F), sommeren 2023 (B), sommeren 2024 (C) og uhøstede felt (D, E, G, H, Referanse). Tallene over kolonnene indikerer tidsrom mellom høsting og observasjon (0: Pågående høsting, 1: Et år etter høsting, 2: To år etter høsting). Nedre grense for 95 % konfidensintervall er tegnet inn.
Figur 32. Gjennomsnittlig registreringsfrekvens av rød kråkebolle langs videotransekt som
er undersøkt i sørlig del av Nordland i perioden 2013-24. Stasjonsantallet var lavere i perioden
2013-20 enn i perioden 2022-24. De vertikale linjene representerer 95 % konfidensintervall.
3.2.2 - Akustikkundersøkelser
Akustikkmålinger (EK60/80) ble gjennomført langs samme transektlinjer som videomålingene, der tarens akustiske styrke (NASC) brukes til å beregne tarebiomasse, som deretter sammenlignes med de videoestimerte biomassetetthetene. Regresjonsanalyser viser godt samsvar mellom de akustiske- og videobaserte målingene, og et lineært forhold mellom de to målingene basert på historiske data benyttes for akustisk biomasseestimering. Resultatene fra de akustikkbaserte biomasseanslagene viste en klar nedgang i gjennomsnittlig biomassetetthet fra året før til året etter høsting. Akustikkmålingene gjennomført på felt høstet to år tidligere ga imidlertid en overestimering av biomassetetthet i forhold til videoestimatene, noe som sannsynligvis skyldes at mesteparten av den reflekterte energien fra tarevegetasjonen kommer fra bladverket og i mindre grad påvirkes av vegetasjonshøyden. Slik vil kortvokst, men tett, vegetasjon bli overestimert av den eksisterende modellen. Vi arbeider nå med akustiske eksperiment i et kontrollert miljø slik at vi får en bedre forståelse for den akustiske responsen av tarevegetasjon i ulike høydesegment og kan utvikle en modell som inkluderer vegetasjonshøyde i de akustiske biomassemålingene.
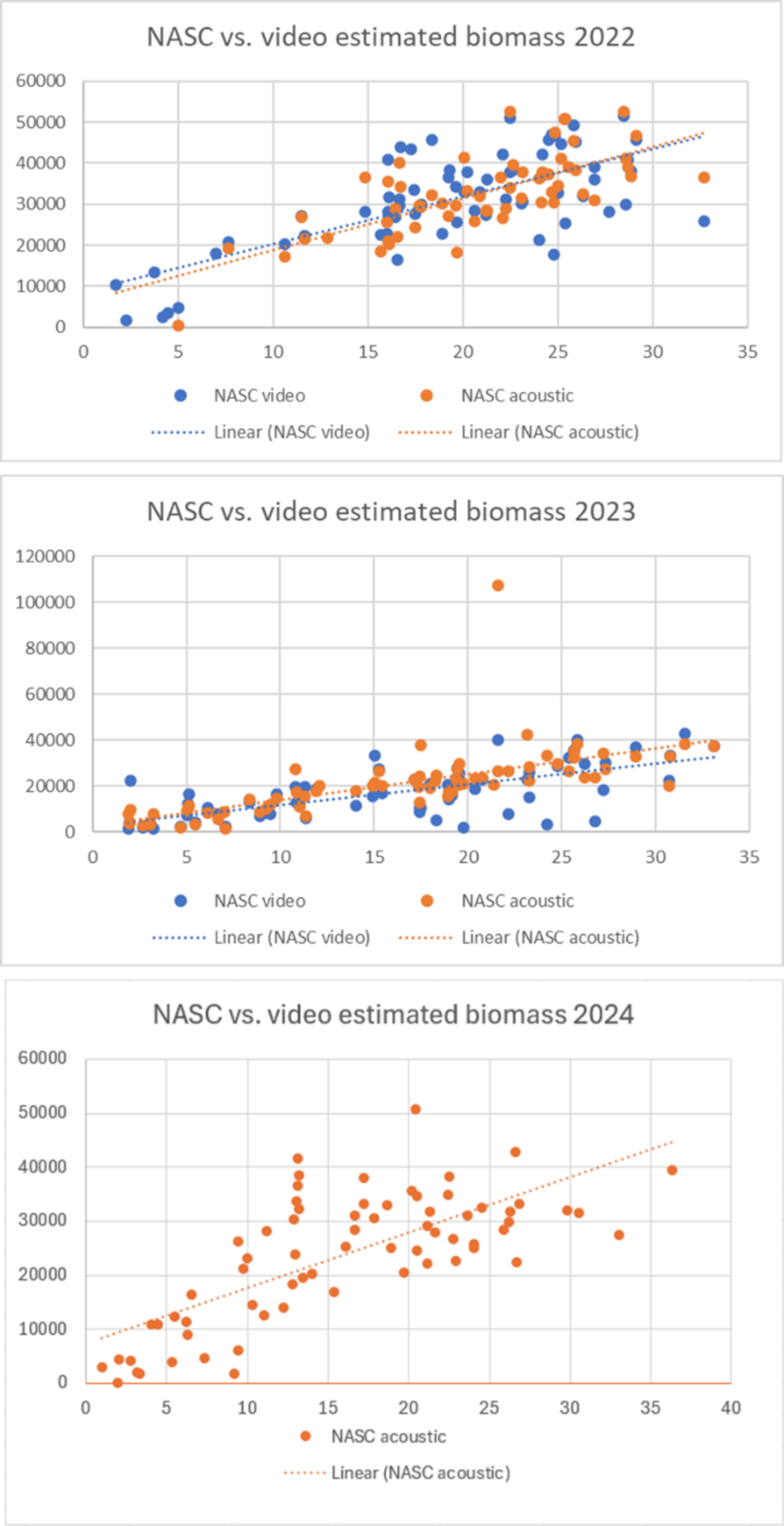
Plottene av NASC og videoestimert biomasse (Fig. 33) viser en til dels stor forskjell i NASC for enkelte stasjoner mellom datasettene som er innsamlet samtidig med videokjøringene og de akustiske transektene. Til tross for dette er det et godt samsvar mellom regresjonslinjene for de to innsamlingsmetodene (Fig. 33). Det er få åpenbare uteliggere, og IQR-metoden luker kun ut en stasjon; stasjon 491Ea i det akustiske datasettet fra 2023 (Fig. 33, nederst). Datasettet fra 2022 (r 2 = 0,634) viser også betydelig større spredning enn de innsamlede dataene fra 2023 (r 2 = 0,730). De akustiske datasettene, som danner grunnlaget for biomasseestimatet, viser en gjennomsnittlig nedgang i akustisk mengde fra 2022 til 2023 (Fig. 33, 34).
Figur 33: NASC fra alle transekter plottet mot estimert biomasse fra videoanalyse.
2022 (øverst), 2023 (midt), 2024 (nederst). «NASC video» er akustisk mengde fra
ekkoloddata samlet inn samtidig med videoopptakene (for 2024 er ikke «NASC video»
målingene inkludert), «NASC acoustic» er akustisk mengde fra ekkoloddata samlet inn
angs den samme transektlinjen i høyere hastighet. Lineære trendlinjer er stiplet.
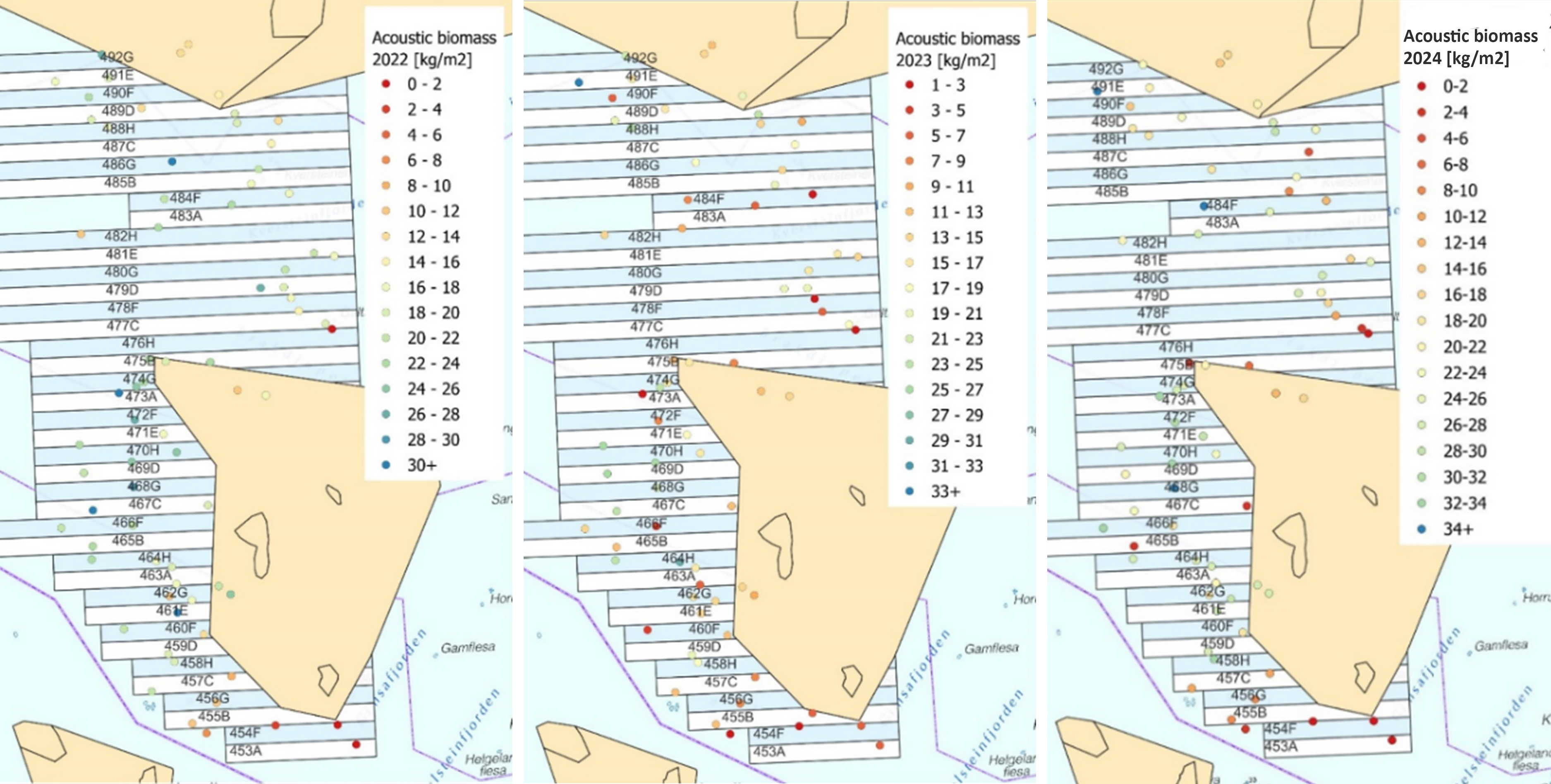
Biomassetetthet estimert på bakgrunn av den akustiske mengden (NASC) langs transektene fra 2022-24 er vist i Fig. 34. Disse viser stor variasjon i biomasse mellom stasjoner, der de høyeste verdiene generelt sett sees på stasjonene vest for Horsvær. Akustikkmålingene i 2022 var også generelt mer preget av støy enn 2023-målingene, noe som sannsynligvis har gitt overestimat av tarebiomasse på enkelte stasjoner.
Figur 34. Akustisk estimering av biomassetetthet av tare i 2022 (til venstre), 2023 (i midten) og 2024 (til høyre)
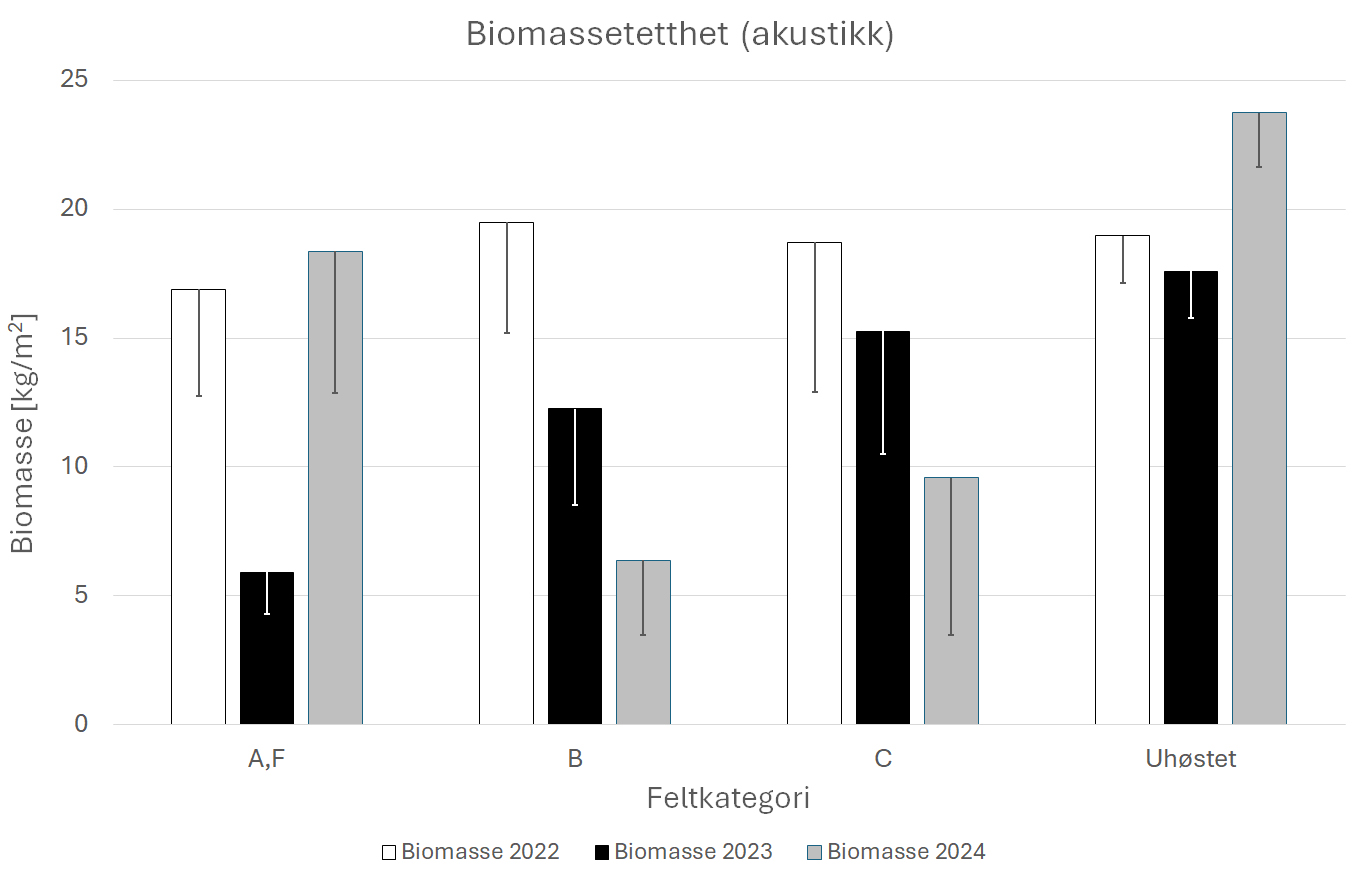
Gjennomsnittlig biomassetetthet av stortare fra akustikkmålingene langs transektene i A - og F - feltene i 2023 og B-feltene i 2024 var ca 65 % lavere året etter høsting enn før høsting i 2022 (Fig. 35). To år etter høsting av A- og F-feltene ga de akustiske signalene imidlertid et estimat av biomassetetthet som lå over estimatene fra før høsting. Den ekstreme økningen på A- og F-feltene to år etter høsting skyldes sannsynligvis akustisk overestimering av biomassen i tett, lav, tareskog. Dette kommer av at ca. 90% av den reflekterte energien fra tareskogen kommer fra bladverket og i liten grad inkorporerer tarestilkenes biomassebidrag. I de uhøstede feltene viser den akustiske biomasseestimeringen en betydelig økning i biomasse i 2024 som vi per i dag ikke kan forklare til det fulle. Den akustiske biomassemodellen er en dynamisk modell som utvikler seg etter hvert som flere datapunkter blir tilgjengelig. En plausibel feilkilde er at et datasett som er implementert i modellen etter at 2023-estimatene ble gjort har ført til en overestimering av 2024-dataene. Dette vil i så fall påvirke alle biomasseestimat i 2024 og følgelig resultere i en overestimering tilsynelatende i størrelsesordenen 10-20%. Dette vil undersøkes når arbeidet med en ny og forbedret modell som også vekter inn tarevegetasjonens høyde er fullført.
Figur 35. Gjennomsnittlig akustikkestimert biomassetetthet av stortare (Laminaria hyperborea) i 2022-24
i felt som ble høstet sommeren 2022 (A, F), sommeren 2023 (B), sommeren 2024 (C) og uhøstede
felt (D, E, G, H, Referanse). Nedre grense for 95 % konfidensintervall er tegnet inn.
3.3 - Modellering av stortarebiomasse
3.3.1 - Validering av modellen
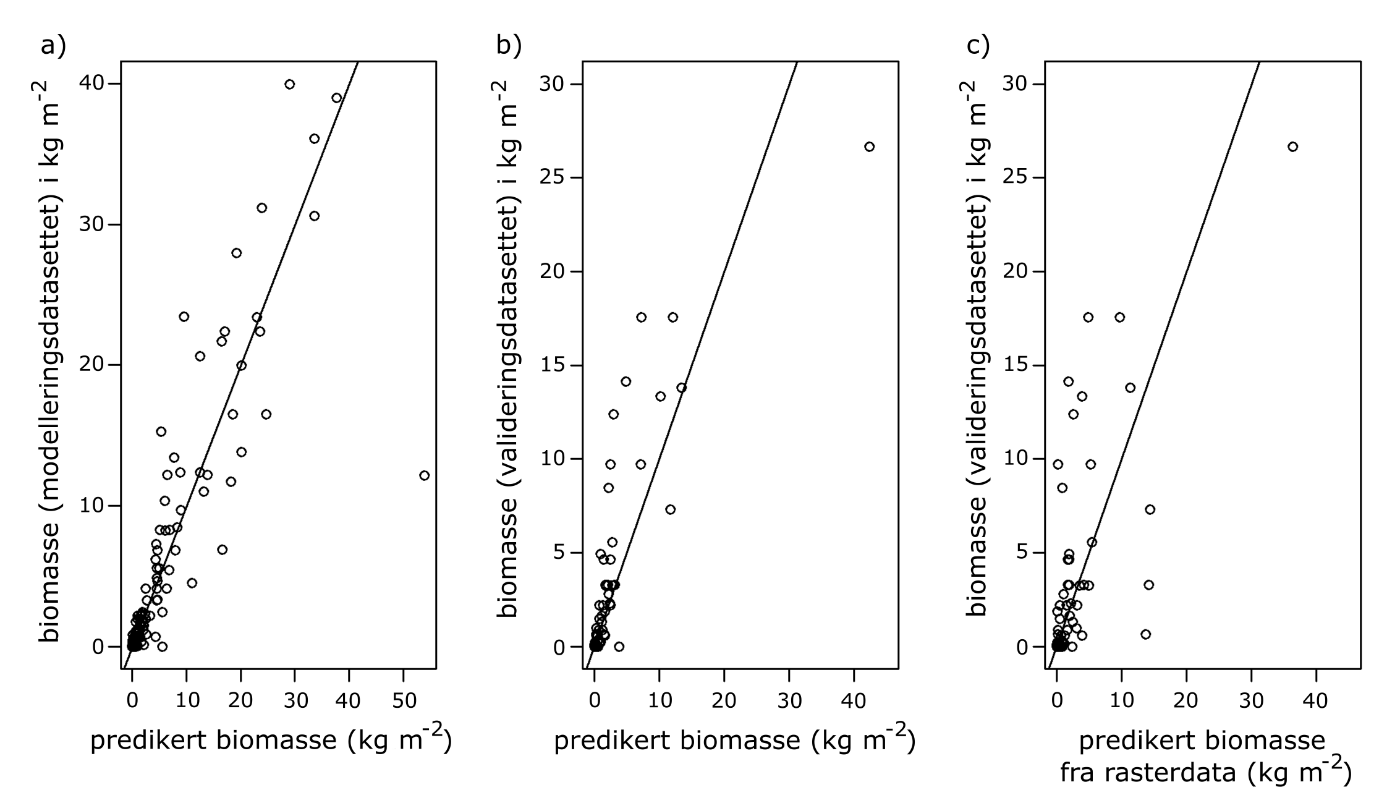
Biomassemodellen for stortare ble validert i tre-trinn og resultatene viste signifikant korrelasjon mellom predikert og observert biomassetetthet av stortare, hvilket indikerer at modellens presisjonsnivå er tilfredsstillende.
Modellen ble først validert med treningsdatasettet samlet inn i 2021 (datasettet som ble brukt til å bygge modellen) ved hjelp av begrenset maksimal sannsynlighetsanalyse (restricted maximum likelihood). Denne valideringen viste en sterk korrelasjon mellom predikert og observert biomassetetthet av stortare på den logaritmiske skalaen (Pea rson’s R = 0,91, p < 0,001 ). I tillegg validerte vi modellen ved å predikere biomassetetthet ved hjelp av de lokalt observerte forklaringsvariablene fra det uavhengige romlig randomiserte datasettet som ble samlet inn før høsteoppstarten i 2022 (Fig. 9). Denne valideringen viste god korrelasjon mellom predikert og observert biomassetetthet (Pea rson’s R = 0,83, p < 0,001 ) på logaritmisk skala. Ved bruk av romlig oppløste forklaringsvariabler i modellen ble imidlertid korrelasjonen redusert, men var fortsatt signifikant (Pea rson’s R = 0,69, p < 0,001 ). Dette utgjør den strengeste formen for validering av modellen (van Son et al. 2020, Norderhaug et al. 2021). Samlet sett tyder likevel resultatene på at den valgte modellen fungerer tilfredsstillende når man tar alle tre valideringene i betraktning (Fig. 36).
Figur 36. Modellpredikert biomassetetthet versus observert biomassetetthet av stortare med a) predikert biomassetetthet fra observerte forklaringsvariabler for treningsdatasettet, b) predikert biomassetetthet fra uavhengige observerte forklaringsvariabler fra valideringsdatasettet, c) predikert biomassetetthet fra romlig oppløste forklaringsvariabler. Den svarte linjen representerer likhet mellom observasjoner og prediksjoner.
3.3.2 - Romlig biomassefordeling
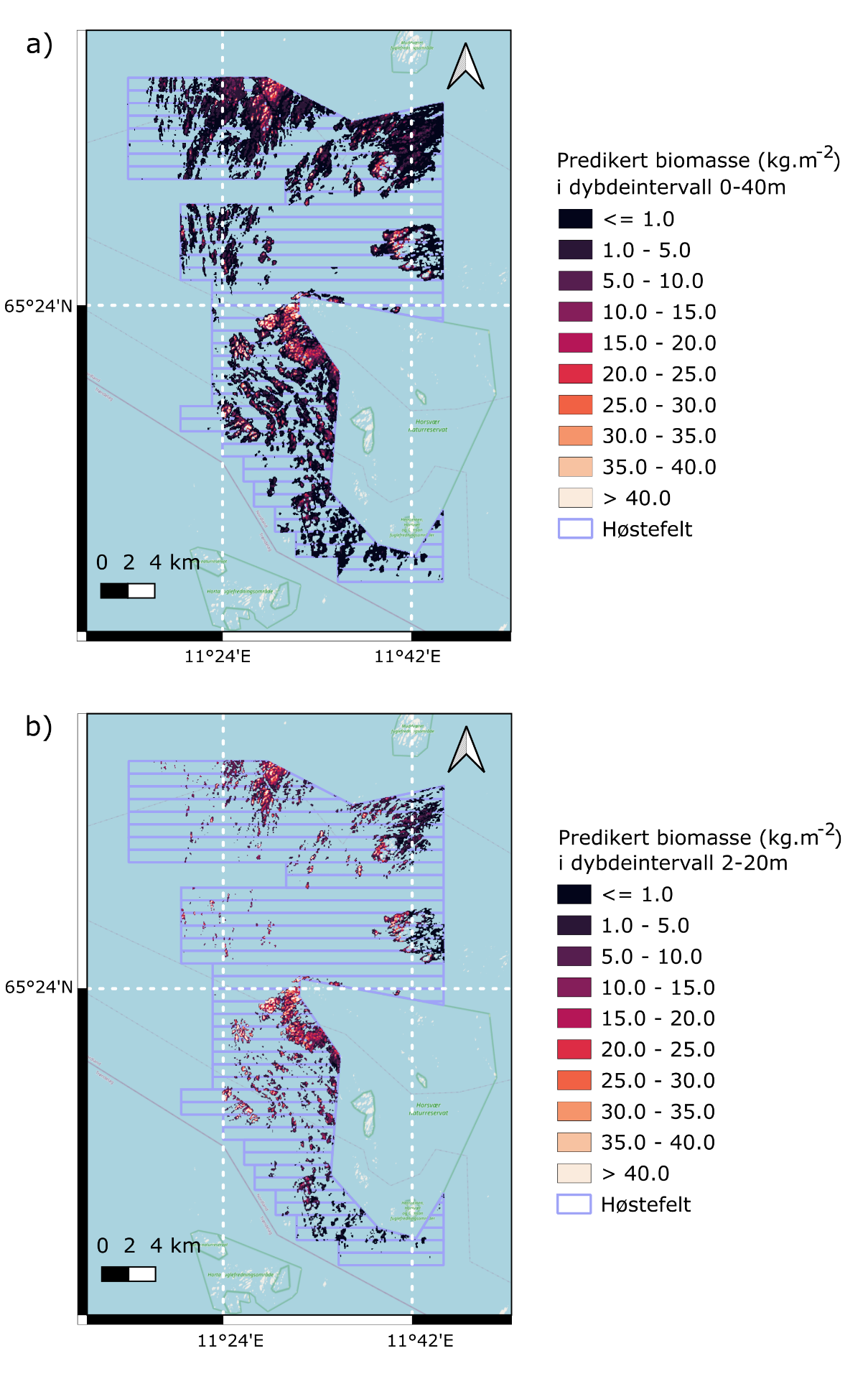
Modelleringen viste store geografiske variasjoner i den romlige fordelingen av stortarebiomasse for tarehøstingsområdet i Nordland, med de høyeste forekomstene i sentrale deler vest av Horsvær naturreservat og i nordlige deler sørvest av Vega verdensarvområde, samt lavest forekomst i sørlig del av modellområdet.
Prediksjonene avslørte store geografiske variasjoner i stortarebiomasse innenfor det modellerte området (Fig. 37). Prediksjonene indikerer også at arealene utenfor dybdeintervallet 2-20 meter har gjennomgående lav biomassetetthet av stortare (Fig. 37). De høyeste biomasseforekomstene ble predikert i sentrale deler av høstingsområdet, vest av Horsvær naturreservat, og i nordlige deler, sørvest av Vega verdensarvområde. Disse to områdene er adskilt av et område, nord av Horsvær, med lavere biomasse grunnet færre gruntvannsarealer egnet for stortarevekst her. Lav biomasse av stortare ble også predikert for de sørlige delene av modellområdet, sannsynligvis knyttet til lav bølgeeksponering i disse områdene.
Figur 37. Romlig fordeling av predikert biomassetetthet (kg m-2 ) av stortare i
tarehøstingsområdene i Nordland i a) 0-40 m dyp og b) 2-20 m dyp.
3.3.3 - Omfang av tarehøsting
Den totale modellerte biomassen av stortare i høstefeltene i Nordland ble summert til 579 000 tonn hvorav 516 000 tonn (89 %) av biomassen står i dybdeintervallet 2-20 m som er tilgjengelig for tarehøsting. En sammenligning mellom høstekvantum og modellert stortarebiomasse for arealene som har vært tilgjengelig for tarehøsting i perioden 2022-24 indikerte en uttaksgrad på 23%.
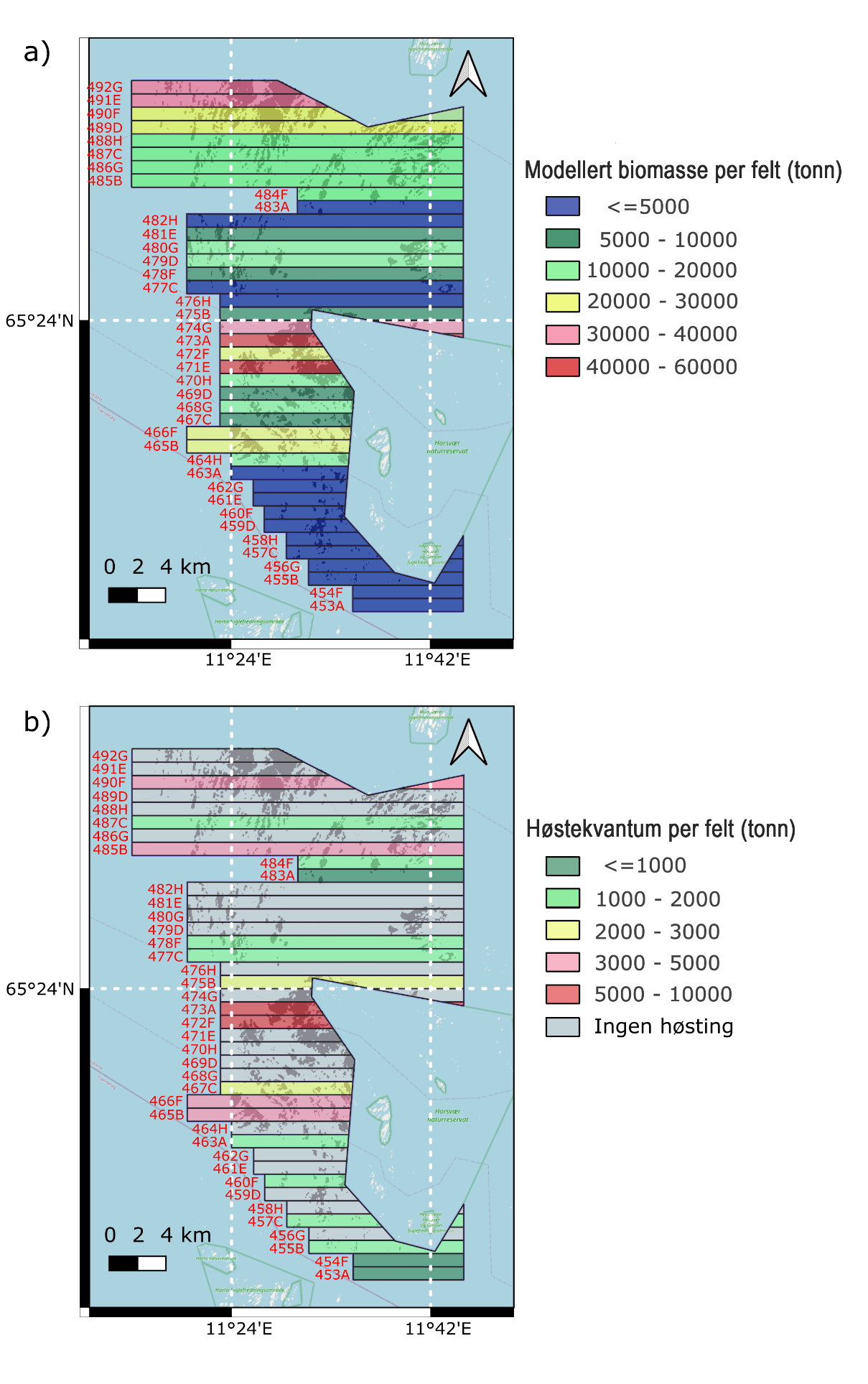
Den totale modellerte biomassen av stortare summert over alle høstefeltene i Nordland var 579 000 tonn og varierte fra et minimum på 2 tonn på felt 453A til et maksimum på 57 800 tonn i felt 471E (Fig. 38). Generelt varierte usikkerheten rundt estimatene med en faktor på ca 2 og usikkerhetsintervallet var relativt sett større for lave enn for høye biomasseestimat, spesielt for de sørlige feltene som alle hadde lave biomasseestimat. For eksempel var forholdet mellom 97,5 % prosentilratio og medianverdi for den estimerte biomassen i felt 453A (2 tonn) og felt 473A (45 400 tonn) henholdsvis ~5 og ~1,75. Tarehøsting i Norge er begrenset til dybdeintervallet 2-20 m (LOVDATA 2004) og estimat over hvor stor andel av tarebiomassen som høstes i dette dybdeintervallet vil derfor gi den mest presise kunnskapen om uttaksgraden innenfor de arealene som faktisk er tilgjengelig for høsting. Den totale modellerte biomassen av stortare innenfor dybdeintervallet 2-20 m ble summert til 516 000 tonn (89 % av den totale biomassen).
Det samlede høstekvantumet av stortare for feltene som har vært åpne for høsting i perioden 2022-24 i Nordland er 47 398 tonn. For disse feltene var den totale modellerte biomassen (0-40 m dyp) 230 000 tonn, med 205 000 tonn modellert innenfor dybdeintervallet 2-20 m (Appendix 6). Når vi sammenligner den modellerte tarebiomassen for dybdeintervallet 2-20 m med høstekvantumet for feltene som har vært åpne for høsting i perioden 2022-24, var den totale prosentandelen av høstet biomasse ift modellert biomasse (uttaksgraden) 23 % med et 95 % konfidensintervall mellom 12 % og 45 %. Det var stor variasjon i uttaksgrad mellom feltene som har vært høstet til nå (fra 14 % på felt 487C til 185 % på felt 455B), og uttaksgraden var høyest for feltene i den sørlige delen der de modellerte biomasseestimatene var gjennomgående lave (Fig. 38). For to av de sørligste feltene (455B og 457C) var høstekvantumet høyere enn den modellerte tarebiomassen, og uttaksgraden følgelig over 100%, men likevel innenfor 95 % konfidensintervallene. Denne underestimeringen i biomassemodelleringene er sannsynligvis knyttet til større usikkerhet rundt prediksjonene ved lave bølgeeksponeringsverdier, samt at en del av arealene i de sørlige feltene manglet dybdeverdier og derfor ikke inkluderes i biomassesummene.
Figur 38: a) Modellert stortarebiomasse per felt (tonn) i dybdeintervallet 2-20 m, b) Innrapportert
høstekvantum per felt (tonn) i Nordland per 2024.
4 - Diskusjon
De ytre deler av kyststrekningen sør i Nordland fylke fra Trøndelagsgrensen til Vega ble åpnet for kommersiell høsting av stortare sommeren 2022 (Fiskeridirektoratet J-131-2022), på bakgrunn av resultater fra tidligere prøvehøstinger i denne regionen. For å få mer kunnskap om taresamfunnenes reetableringsevne og restitusjonsperiodens varighet i denne regionen har Havforskningsinstituttet i perioden 2020-24 gjennomført oppfølgende studier av tilstanden i tidligere prøvehøstede områder sammenlignet med nærliggende referanseområder.
Videoundersøkelsene viste som ved tidligere undersøkelser (Steen et al. 2018b, 2020) at tarevegetasjonens kronesjikthøyde og biomassetetthet var reetablert i løpet av 5 år, og at restitusjonen etter prøvehøstingene ikke har vært påvirket av kråkebollebeiting. Innsamlede stortareplanter fra felt høstet 5 år tidligere var av samme størrelse, men ble dominert av yngre aldersgrupper og hadde lavere diversitet i epifyttsamfunnene enn stortareplanter, samlet inn i referanseområdene. Vekstsoneanalyser av årringene i tarestilkene viste også at den årlige stilktilveksten har vært høyere i feltene som ble høstet 5 år tidligere enn i referanseområdene. Dette samsvarer med resultater fra tidligere undersøkelser i denne regionen og indikerer at en 5 års høstesyklus kan være gunstig med henblikk på effektiv ressursutnyttelse av tarebiomassen, men ikke gir tilstrekkelig hviletid for restitusjon av aldersstruktur og epifyttsamfunn (Steen et al. 2020).
Det ble derimot ikke observert forskjeller i aldersstruktur og epifyttsammensetning mellom stortareplanter samlet inn i referanseområder og felt høstet 8 år tidligere. Dette tyder på at effektene av tarehøstingen er reversible og at 8 år vil være tilstrekkelig hviletid for restitusjon av tarepopulasjonenes aldersstruktur og assosierte epifyttsamfunn i denne regionen. En restitusjonsperiode på 8 år for epifyttsamfunnene understøttes også av relasjonen mellom epifyttforekomster og stortareplantenes alder, der den kraftigste epifyttutviklingen finner sted når plantene er mellom 6 år og 8 år gamle. En 10 års høstesyklus vil dermed gi taresamfunnene tid til å reetablere klimakstilstand og økosystemfunksjonalitet.
Prøvehøstingene i Nordland gir kun informasjon om taresamfunnenes restitusjon etter en enkel høsterunde med stedvis kvantumsbegrensninger på høstevolumene (Steen et al. 2020). Gjentagende høstinger vil kunne påvirke taresamfunnenes sårbarhet og restitusjonsevne (Buschmann et al. 2014, Mac Monagail et al. 2017, Carranza et al. 2024), og høsteaktivitetens bærekraftighet bør derfor undersøkes over tid langs denne kyststrekningen.
For å undersøke taresamfunnenes tilstand og overvåke videre utvikling etablerte Havforskningsinstituttet i forkant av oppstarten av kommersiell tarehøsting i Nordland i 2022, overvåkingsstasjoner (transekter) for video og akustikk i samtlige høstefelt og i nærliggende referanseområder. Disse observasjonene vil gi et sammenligningsgrunnlag for å måle effekter av den kommersielle tarehøstingsaktiviteten i denne delen av Nordland, der den differensierte høstesyklusen også byr på muligheter for å teste betydningen av hvileperiodens varighet på taresamfunnenes restitusjonsevne over tid. Observasjonene gjennomført i Nordland før høsteoppstarten i 2022 viste, med unntak av de sørligste feltene, gode forekomster av høyvokst stortarevegetasjon i de fleste felt. De sørligste feltene i denne regionen har i liten grad vært gjenstand for tidligere prøvehøstinger og vegetasjonen langs de undersøkte transektene domineres her stedvis av andre arter enn stortare. Årsakene til variasjoner i forekomst og størrelse av stortare har sammenheng med variasjoner i graden av bølgeeksponering, men manglende reetablering av stortarevegetasjon etter tidligere nedbeitingsepisoder i deler av området kan heller ikke utelukkes.
I 2023 og 2024 ble det gjennomført oppfølgende undersøkelser langs de de samme transektene som ble etablert i 2022, som viste små endringer av tarevegetasjonen i uhøstede felt og referanseområder, men forekomst av tydelige trålspor i de fleste felt som har vært åpne for kommersiell tarehøsting i perioden 2022-24. Til tross for redusert stortarevegetasjon på feltene som ble høstet i 2022 (A og F felt), har rekoloniseringen her vært god og på nivå med det som er observert etter tidligere prøvehøstinger i dette området (Steen et al. 2015ab, 2016, 2018ab, 2020).
For feltene som ble høstet i 2023 (B-felt) var tilstanden året etter høsting mer variabel og rekoloniseringen av stortare redusert i forhold til utviklingen året etter høstingen av A- og F-feltene. Den reduserte gjenveksten av stortare var korrelert med en økning i forekomst av draugtare (Saccorhiza polyschides) på tre av de fire B-feltene i løpet av det første året etter høstingen. Draugtare er en varmekjær brunalge som har hatt en sørvestlig utbredelse i Norge (Brattegard & Holte 1997, Artsdatabanken 2024) og økte sjøtemperaturer kan føre til at arten får bedre vekstvilkår og brer seg nordover langs kysten. I områder med redusert stortarevegetasjon, som for eksempel på trålflater, vil sannsynligvis draugtare også lettere kunne få fotfeste sammenlignet med områder med mer heldekkende stortarevegetasjon. Draugtare er en hurtigvoksende, ettårig, art som det er vanskelig å forutsi utviklingen til fra år til år, men oppblomstringen av denne vil sannsynligvis være et forbigående stadium før den flerårige stortarevegetasjonen igjen overtar dominansen. Liknende fenomen ble også observert, dog i noe mindre omfang, etter prøvehøstingene i 2017, der draugtare og den trådformede brunalgen mykt kjerringhår (Desmarestia viridis) økte i forekomst på enkelte trålflater året etter høsting (Steen et al. 2018b) før stortarevegetasjonen senere reetablerte dominansen (Steen et al. 2020, Fig: 39). Det blir derfor viktig å følge vegetasjonsutviklingen på høstefeltene videre for å undersøke om oppblomstringen av draugtare etter høstingen i 2023 er et forbigående stadium og i hvilken grad restitusjonen av stortare påvirkes av denne oppblomstringen. Hvorvidt draugtare også vil øke i forekomst på feltene som ble høstet i 2024 (C-felt) er det foreløpig ikke grunnlag for å si noe om, men vegetasjonsutviklingen bør også her følges videre.
Figur 39. Eksempler på tilstand observert i trålspor langs videotransekter, henholdsvis et år (øvre bilder)
og to år (nedre bilder) etter prøvehøstingene av stortare i 2017. Stedvis dominans av mykt kjerringhår
(Desmarestia viridis) og draugtare (Saccorhiza polyschides) året etter prøvehøstingen (øvre bilder) og
dominans av stortare (Laminaria hyperborea) to år etter prøvehøstingen (nedre bilder). Tallene nederst
til høyre i bildene refererer til høstefeltenes nåværende nummerering.
Akustikkmålinger vil kunne påvirkes av støy, blant annet luftbobler fra propeller og sjøgang på innsamlingstidspunktet. Generelt sett var akustikkmålingene fra 2022 preget av støy, noe som sannsynligvis har gitt overestimat av biomasse på enkelte stasjoner. Datasettet fra 2022 manglet i tillegg kjøringer av raske akustiske transekter for 12 av stasjonene, som ble erstattet med akustikkdata samlet inn under videotransektkjøringene. Selv om disse dataene er samlet inn langs de samme transektlinjene vil de ikke være helt kompatible og derfor tilføre en viss usikkerhet til datasettet. De akustiske datasettene fra videotransektkjøringene der fartøyet beveger seg sideveis er ofte betydelig mer påvirket av boblestøy fra sidepropeller enn de raske akustiske transektene som tas opp når fartøyet beveger seg rettvendt. Dette kan både føre til over- og underestimering av biomasse. På grunne stasjoner der luftbobler penetrerer tareskogen vil disse føre til en stor økning i akustisk mengde i integrasjonsområdet, noe som gir urealistisk høyt biomasseestimat. På dypere stasjoner vil luftboblene danne et akustisk lokk over tareskogen og dempe de akustiske signalene, som gjør at mindre av den akustiske energien når ned til tareskogen og videre til at mindre akustisk energi reflekteres. Dette vil igjen føre til et urealistisk lavt biomasseestimat. Datasettene inneholdt bare en uteligger som ble luket ut av IQR-metoden, det vestlige transektet i 491E feltet i 2023. Den gjennomsnittlige akustiske mengden i dette transektet er den høyeste som til dags dato er innsamlet, og ansees som korrumpert data. Vi har ikke klart å forstå bakgrunnen til dette avviket til det fulle: Ved visuell inspeksjon av dataene ser de ikke ut til å være spesielt påvirket av bølgestøy, men det virker til å være en del punktstøy i vannmassene. På videoen fra transektet kan det sees en veldig stor tetthet av maneter, og en mulig forklaring kan være at dette påvirker den akustiske mengden noe. Det virker dog urealistisk at dette alene vil kunne påvirke de akustiske dataene i stor nok grad til å ødelegge dem. Uavhengig av underliggende grunn så ble altså dette datapunktet flagget som en uteligger og korrigert til ekstremalverdien i regresjonsmodellen i det endelige biomasseestimatet.
Et annet overraskende resultat kan sees på referansestasjonene sørvest i naturvernområdet i Horsvær der den akustikkestimerte biomassetettheten, spesielt for den østligste stasjonen, går markant ned fra 2022 til 2023 for så å øke igjen i 2024, mens den videoestimerte biomassetettheten viste kun små år til år variasjoner. Dette skyldes sannsynligvis at tidevannsforskjeller og posisjonsavvik mellom innsamlingsperiodene har påvirket de akustiske biomasseestimatene. I 2023 måtte f.eks den grunneste, mest vegetasjonsrike delen av det østligste transektet maskeres fordi vannsøylen over tarevegetasjonen var for liten grunnet lav vannstand, i tillegg til at akustikkmålingene ble startet for tidlig og inneholdt mer dype, vegetasjonsfattige, områder sammenlignet med 2022 og 2024 målingene. I tillegg til variasjoner i støymengde kan avvik også til en viss grad skyldes at akustikk- og videotransektene ikke overlapper fullstendig geografisk, da det er ca. 5-6 meter mellom kameraet og ekkoloddsvingerne. Det er imidlertid lite sannsynlig at disse usikkerhetene har påvirket biomassestimatene fra høstede felt i mindre eller større grad enn estimatene fra uhøstede felt.
Til tross for disse innførte usikkerhetene gir de akustiske undersøkelsene generelt sett et godt bilde av den stående biomassen som samsvarer med estimatene fra videoanalysene, selv om de absolutte verdiene avviker noe på enkelte stasjoner. Dette gjelder spesielt for målingene gjennomført på felt høstet to år tidligere der de akustiske biomasseestimatene overestimeres i forhold til videoestimatene. Dette skyldes sannsynligvis at mesteparten av den reflekterte energien fra tarevegetasjonen kommer fra bladverket og i mindre grad inkluderer vegetasjonshøyden og biomassebidraget fra tarestilkene. En kortvokst, men tett, tarevegetasjonstype blir derfor sannsynligvis overestimert av den eksisterende metoden som kun er basert på NASC-signalene. For å utvikle akustikkmetoden og inkorporere vegetasjonshøyde i biomasseestimatene arbeider vi nå med akustiske eksperiment i et kontrollert miljø slik at vi får en bedre forståelse for den akustiske responsen av tarevegetasjon i ulike høydesegment.
Kråkebollebeiting er den viktigste årsaken til tap av tareskog i de nordlige kystområdene i Norge, og oppblomstringene av kråkeboller som ble observert på begynnelsen av 1970-tallet (Sivertsen & Bjørge 1980, Sivertsen 1982, 1997, 2006, Sakshaug & Sjøtun 2002) er senere satt i sammenheng med utfiskingen av kråkebollespisende fiskeslag (Norderhaug et al. 2020). Omfattende nedbeitingsepisoder av taresamfunn har også vært rapportert fra andre kystområder i flere verdensdeler (Filbee-Dexter et al. 2014, 2017, Ling et al. 2015). Det er grønn kråkebolle (Strongylocentrotus droebachiensis) som utgjør den største trusselen og som har forårsaket nedbeitingene av tareskogene i Nord-Norge, men det er også observert at høye tettheter av rød kråkebolle (Echinus esculentus) kan redusere tarevegetasjonen i enkelte kystområder (Sjøtun et al. 2006, Steen 2016, 2022).
Med tanke på kråkebollenes herjinger og nedbeitingshistorikken til tareskogene i nordområdene kan kystområdene i Nordland være mer utsatt for nye kråkebolleoppblomstringer enn områdene lenger sør langs norskekysten. Bestandsutviklingen av kråkeboller bør derfor overvåkes regelmessig i denne regionen. Kråkebolleforekomstene ble registrert ved å telle individene som visuelt observeres langs videotransektene og viste en økning i registreringsfrekvens av rød kråkebolle på felt som var under høsting på observasjonstidspunktene. Denne økningen er sannsynligvis relatert til observasjonsmetoden da kråkeboller vil være mer synlige og lettere registreres i nytrålte områder, enn i områder med mer heldekkende tarevegetasjon. Forekomstene av kråkeboller som observeres er likevel på et moderat nivå og det er heller ingen indikasjoner på at beiting fra kråkeboller har hemmet vegetasjonsutviklingen etter tarehøstingene som til nå er gjennomført i Nordland. Sammenligninger av videoobservasjoner gjennomført i denne regionen i perioden 2013-24 (Steen et al. 2014, 2015ab, 2016b, 2018ab, 2020) tyder på at forekomstene i perioden 2022-24 ikke er høyere enn tidligere og at det sågar har vært en svak nedadgående trend gjennom hele denne perioden.
Forekomster av juvenile individer av grønn kråkebolle i tareplantenes festeorgan (hapterkråkeboller) vil være en indikator på denne artens rekrutteringspotensial i et område. Tidligere innsamlinger av stortare på strekningen Flekkefjord – Bodø viste en økning i registreringsfrekvens av hapterkråkeboller fra sør til nord, med høyere forekomster på kyststrekningen fra Vega til Bodø, enn i områdene sør for Vega i Nordland (Steen et al. 2020). For tidligere innsamlinger av tareplanter i sørlig del av Nordland (Trøndelagsgrensen - Vega) i perioden 2013-19 har den årlige gjennomsnittlige registreringsfrekvensen variert fra 0,2 – 1,1 hapterkråkeboller per tareplante (Steen et al. 2014, 2015ab, 2016b, 2018ab, 2020), mens den i 2022-24 var 0,4 hapterkråkeboller per tareplante. Dette tyder på at rekrutteringspotensialet for grønn kråkebolle ikke har vært økende i sørlig del av Nordland i de senere år, men at arten fortsatt har en viss rekruttering og kan være en latent trussel for regionens tareskoger.
Effektene av tarehøsting og dens bærekraftighet vil avhenge av inngrepets omfang og taresamfunnenes restitusjonsevne (Norderhaug et al. 2021). Selv om resultatene fra Havforskningsinstituttets gjenvekststudier tilsier at tarehøsting er et reversibelt inngrep vil likevel tarevegetasjonen og dens økologiske funksjonalitet i et høsteområde være redusert gjennom restitusjonsperioden. I tillegg til taresamfunnenes restitusjonsevne vil kunnskap om tarehøstingens omfang i forhold til ressursgrunnlaget være vesentlig for å vurdere den samlede påvirkningen av høsteaktiviteten i et område.
For å utrede ressursgrunnlaget på høstefeltene i Nordland har Havforskningsinstituttet utviklet en biomassemodell for stortare. Denne er basert på biologiske data samlet inn i forkant av høsteoppstarten og deres relasjon med ulike miljøvariable. Uttaksgraden av stortare beregnes deretter ved å sammenligne innrapporterte høstedata med de modellerte biomasseestimatene for en kvantitativ beregning av inngrepets omfang i forhold til ressursgrunnlaget på høstefeltene.
Den totale modellerte biomassen av stortare i høstefeltene i Nordland ble beregnet til 579 000 tonn, hvorav 516 000 tonn står i dybdeintervallet 2-20 m der tarehøsting er tillatt (LOVDATA 2004). Dette betyr at 89 % av feltenes stortareforekomster i prinsippet vil kunne påvirkes av høsteaktiviteten, mens 11 % av forekomstene står i dybdeintervall som er skjermet for tarehøsting. En sammenligning mellom høstekvantum og modellert biomasse av stortare (i 2-20 m dyp) for feltene som har vært høstet i perioden 2022-24 indikerer en uttaksgrad på 23 % med et 95 % konfidensintervall mellom 12 % og 45 %. Selv om det er en betydelig grad av usikkerhet knyttet til de modellerte biomasseanslagene ligger uttaksgraden på et klart lavere nivå enn reduksjonene på 65-70 % i tarebiomasse som observeres fra året før til året etter høsting langs overvåkingstransektene. Dette kan blant annet skyldes at overvåkingstransektene er lokalisert i grunne partier spesielt egnet for tarehøsting, mens biomassemodellen er basert på tilfeldig lokaliserte (randomiserte) data som også inkluderer arealer med tareforekomster som kan være mindre tilgjengelige for trålredskapen som benyttes. Det er sannsynligvis også et visst biomassesvinn knyttet til trålfelling av tareplanter som blir liggende igjen på bunnen og følgelig ikke inngår i uttaksgradberegningene da høstekvantum kun er basert på landet biomasse (Fig. 40).
Figur 40. Eksempler på felling av tareplanter som blir liggende tilbake på bunnen etter tråling, observert langs videotransekter i juni 2023 på nylig høstede B-felt (475B) til venstre og i juni 2024 på nylig høstede C-felt (467C) til høyre.
Det er tidligere utviklet biomassemodeller for stortare langs andre deler av norskekysten, blant annet for Vikna-området i Trøndelag (Norderhaug et al. 2021) som har en geografisk nærhet til høsteområdene i Nordland. I Vikna omfattet modellområdet syv høstefelt, hvorav fire (414E, 416B, 417C, 419E) er høstet i perioden 2021-24. For disse fire feltene ble den totale biomassen av stortare modellert til 99 000 tonn (Norderhaug et al. 2021), mens det samlede høstekvantumet i siste treårsperiode har vært 25 000 tonn (Kilde: Fiskeridirektoratet), noe som gir en uttaksgrad på 25 %. I Vikna ble uttaksgraden beregnet i forhold til modellanslagene for den samlede stortarebiomassen i alle dyp og hvis vi sammenligner med tilsvarende modell i Nordland gir dette her en uttaksgrad på 21 % av den totale stående biomassen (0-40 m dyp). Den noe høyere uttaksgraden i Vikna kan bero på forskjeller i modellvariablenes presisjonsnivå, men også skyldes variasjoner i ressurstilgjengelighet og høstetrykk da feltene i Trøndelag har fem måneder lengre høstesesong enn i Nordland.
Hvis vi gjør en enkel prognoseberegning basert på de modellerte biomassesummene i 2-20 m dyp for alle tarehøstingsfeltene i Nordland og antar en gjennomsnittlig uttaksgrad på 23% for samtlige felt vil det totale forventede høstekvantumet, etter at alle feltene er høstet en gang, være ca 120 000 tonn. Til sammenligning så er det høstet ca 693 000 tonn på kyststrekningen Rogaland-Trøndelag ila siste femårsperiode (Kilde: Fiskeridirektoratet) da de aller fleste av høstefeltene på denne kyststrekningen har vært åpne for en enkel høsterunde. Hvis prognoseberegningene holder stikk så innebærer dette at høstefeltene i Nordland kan holde ca 15 % av tareressursene som for tiden utnyttes langs norskekysten til tross for at dette området ikke utgjør mer enn ca 5 % av den totale kystlinjen som er tilgjengelig for kommersiell tarehøsting i Norge.
5 - Konklusjoner
Havforskningsinstituttet gjennomførte i perioden 2020-24 oppfølgende tilstandsundersøkelser av taresamfunnene på tidligere prøvehøstede felt og i uhøstede referanseområder i sørlig del av Nordland. Resultatene fra disse undersøkelsene viser, i tråd med tidligere studier, at stortarevegetasjonens størrelsesstruktur og biomassetetthet er reetablert 5 år etter høsting. Undersøkelsene viste også at tarevegetasjonens aldersstruktur og epifyttsamfunn ikke reetableres i løpet av 5 år, men var reetablert 8 år etter høsting og da ikke forskjellig fra uhøstede referanseområder. Dette indikerer at den differensierte høstesyklusen som er innført for tare i Nordland vil gi en kombinasjon av felt der tareresursene utnyttes effektivt gjennom en 5 års høstesyklus og felt der taresamfunnene gis tid til å reetablere klimakstilstand av en viss varighet gjennom en 10 års høstesyklus. Den differensierte høstesyklusen vil også gi muligheter til å teste betydningen av hvileperiodens varighet på taresamfunnenes produksjon og økosystemfunksjonalitet, over tid. Vi anbefaler derfor at denne høstesyklusen videreføres for sørlig del av Nordland og at restitusjon av taresamfunnene overvåkes regelmessig med evaluering av resultater og høsteplan etter fullført syklus.
For å følge den videre utviklingen etablerte Havforskningsinstituttet, i forkant av oppstarten av den kommersielle tarehøstingen i Nordland i 2022, stasjoner i samtlige høstefelt og i uhøstede referanseområder for video- og akustikkbasert overvåking av tareforekomstene. Oppfølgende undersøkelser på disse stasjonene i 2023 og 2024 viste klare spor etter tråling og betydelig reduksjon av tarebiomasse på de høstede feltene, men også at rekoloniseringen og tilveksten av stortare har vært god de første årene etter høstingen på de fleste felt. På flere av feltene som ble høstet i 2023 ble det imidlertid året etter høstingen observert økte forekomster av den hurtigvoksende brunalgen draugtare og vegetasjonsutviklingen bør her følges videre i årene framover.
Tareforekomstene i Nordland har tidligere vært preget av kråkebollebeiting og utviklingen av kråkebolleforekomster og deres eventuelle påvirkning på reetableringen etter tarehøstinger bør derfor overvåkes spesielt i denne regionen. Resultatene fra de senere års undersøkelser tyder på at kråkebolleforekomstene i denne delen av Nordland for tiden er på et moderat nivå som ikke har påvirket tarevegetasjonens restitusjon etter høstingene som har vært gjennomført til nå.
For å få mer helhetlig kunnskap om forekomstene av tareressurser på høstefeltene i Nordland ble det etablert en prediksjonsmodell for stortarebiomasse i dette området. Kvantitativ kunnskap om hvor store tareressurser som finnes på de enkelte høstefeltene og omfanget av høsteaktiviteten vil være viktig dersom man i framtiden vurderer å innføre feltspesifikke kvantumsgrenser på uttaket. Modellberegningene indikerte en total stående biomasse av stortare på høstefeltene i Nordland på til sammen 579 000 tonn, med 516 000 tonn stående innenfor dybdeintervallet (2-20 m) som er tilgjengelig for tarehøsting. En sammenligning av mellom høstekvantum og den modellberegnede stortarebiomassen for arealene som har vært åpne for høsting i perioden 2022-24 indikerer en uttaksgrad på til sammen 23 % av stortarebiomassen. Videre detaljert rapportering av høstekvantum og høsteinnsats (tidsbruk per høstet kvantum) vil være viktig for beregninger av uttaksgrad og høsteffektivitet, og også bidra med informasjon om eventuelle endringer i tareressursgrunnlaget over tid.
6 - Referanser
Akaike H. 1974. A new look at the statistical model identification. IEEE Transactions on Automatic Control 19 (6): 716-723. DOI: 10.1109/TAC.1974.1100705.
Anderson SC, Ward EJ, English PA, Barnett LAK, Thorson JT. 2024. “sdmTMB: an R package for fast, flexible, and user-friendly generalized linear mixed effects models with spatial and spatiotemporal random fields”. bioRxiv. *2022.03.24.485545*. https://doi.org/10.1101/2022.03.24.485545.
Bates DM, Maechler M, Bolker B, Walker S. 2015. Fitting Linear Mixed-Effects Models Using lme4. Journal of Statistical Software, 67 (1), 1-48. <doi:10.18637/jss.v067.i01>.
Bekkby T, Rinde E, Gundersen G, Norderhaug KM, Gitmark JK, Christie H. 2014. Length, strength and water flow: relative importance of wave and current exposure on morphology in kelp Laminaria hyperborea . Marine Ecology Progress Series. 506: 61–70.
Brattegard T, Holthe T (eds). 1997. Distribution of marine, benthic macro-organisms in Norway. A tabulated catalogue. Preliminary edition. Research Report for DN 1997-1. Directorate for Nature Management. 409 pp.
Buschmann AH, Prescott S, Potin P, Faugeron S, Vasquez JA, Camus C, Infante J, Hernandez-Gonzalez MC, Gutierrez A, Varela DA. 2014. The status of kelp exploitation and marine agronomy, with emphasis on Macrocystis pyrifera , in Chile. Advances in Botanical Research. 71: 161–188.
Carranza DM, Wieters EA, Vásquez JA, Stotz BS. 2024. Exploring the consequences of kelp removal: a review shows we are missing a broader perspective. Biodiversity and Conservation (2024). https://doi.org/10.1007/s10531-023-02769-7.
Christie H, Fredriksen S, Rinde E. 1998. Regrowth of kelp and colonization of epiphyte and fauna community after kelp trawling at the coast of Norway. Hydrobiologia 375 (376): 49–58.
Christie H, Jørgensen, NM, Norderhaug KM, Waage-Nielsen E. 2003. Species distribution and habitat exploitation of fauna associated with kelp ( Laminaria hyperborea ) along the Norwegian coast. Journal of the Marine Biological Association of the United Kingdom 83: 687–699.
Christie H, Jørgensen NM, Norderhaug KM. 2007. Bushy or smooth, high or low; importance of habitat architecture and vertical position for distribution of fauna on kelp. Journal of Sea Research 58: 198–208.
Dunn PK, Smyth GK. 2005. Series evaluation of Tweedie exponential dispersion model densities. Statistics and Computing 15: 267–280. https://doi.org/10.1007/s11222-005-4070-y.
Fagerli CW, Norderhaug KM, Christie H. 2013. Lack of sea urchin settlement may explain kelp forest recovery in overgrazed areas in Norway. Marine Ecology Progress Series. 488: 119–132.
Fagerli CW, Stadniczeñko SG, Pedersen MF, Christie H, Fredriksen S, Norderhaug KM. 2015. Population dynamics of Strongylocentrotus droebachiensis in kelp forests and barren grounds in Norway. Marine Biology. 162: 1215–1226.
Filbee-Dexter K, Scheibling RE. 2014. Sea urchin barrens as alternative stable states of collapsed kelp ecosystems. Marine Ecology Progress Series. 495: 1–25.
Filbee-Dexter K, Scheibling RE. 2017. The present is the key to the past: linking regime shifts in kelp beds to the distribution of deep-living sea urchins. Ecology. 98(1): 253–264.
Hartig F. 2022. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level / Mixed) Regression Models. R package version 0.4.6. https://CRAN.R-project.org/package=DHARMa.
Kartveit KH, Filbee-Dexter K, Steen H, Christensen L, Norderhaug KM. 2022. Efficient spatial kelp biomass estimations using acoustic methods. Frontiers in Marine Science 9: 1065914
Ling SD, Scheibling RE, Rassweiler A, Johnson CR, Shears N, Connell SD, Salomon AK, Norderhaug KM, Pérez-Matus A, Hernández JC, Clemente S, Blamey LK, Hereu B, Ballesteros E, Sala E, Garrabou J, Cebrian E, Zabala1 M, Fujita D, Johnson LE. 2015. Global regime shift dynamics of catastrophic sea urchin overgrazing. Philosophical Transactions Royal Society B. 370: 20130269.
Mac Monagail M, Cornish L, Morrison L, Araújo R, Critchley AT. 2017. Sustainable harvesting of wild seaweed resources. European Journal of Phycology. 52: 371-390.
Norderhaug KM, Fredriksen S, Nygaard K. 2003. Trophic importance of Laminaria hyperborea to kelp forest consumers and the importance of bacterial degradation to food quality. Marine Ecology Progress Series 255: 135–144.
Norderhaug KM., Christie H. 2009. Sea urchin grazing and kelp re-vegetation in the NE Atlantic. Marine Biology Research 5: 515-528.
Norderhaug KM., Christie H. 2011. Secondary production in a Laminaria hyperborea kelp forest and variation according to wave exposure. Estuarine, Coastal and Shelf Science 95: 135-144.
Norderhaug KM, Christie H, Andersen GS, Bekkby T. 2012. Does the diversity of kelp forest macrofauna increase with wave exposure? Journal of Sea Research 69: 36–42.
Norderhaug KM, Nedreaas KH, Huserbråten M, Moland E. 2020. Depletion of coastal predatory fish sub-stocks coincided with the largest sea urchin grazing event observed in the NE Atlantic. Ambio, 50: 163-171. doi: 10.1007/s13280-020-01362-4.
Norderhaug KM, Freitas Brandt C, Heiberg Espeland S, Albretsen J, Christensen-Dalsgaard S, Ohldieck MJ, van Son TC, Knutsen JA, Moy FE, Steen H. 2021. Bærekraftig taretråling - Vurdering av bærekraftskriterier ved Vikna. Rapport fra Havforskningen 2021-46.
Pedersen MF, Nejrup LB, Fredriksen S, Christie H, Norderhaug KM. 2012. Effects of wave exposure on population structure, demography, biomass and productivity of the kelp Laminaria hyperborea . Marine Ecology Progress Series. 451: 45–60.
Pedersen MF, Nejrup LB, Pedersen TM, Fredriksen S. 2014. Sub-canopy light conditions only allow low annual net productivity of epiphytic algae on kelp Laminaria hyperborea . Marine Ecology Progress Series. 516: 163-176.
Sakshaug E, Sjøtun K. 2002. Nedbeiting av tareskog i Norge. Rapport fra arbeidsgruppe nedsatt av Fiskeridepartementet og Miljøverndepartementet. 1-47.
Sivertsen K, Bjørge A. 1980. Reduksjon av tareskogen på Helgelandskysten. Fisken og Havet 4: 1-9.
Sivertsen K. 1982. Utbredelse og variasjon i kråkebollebeiting av tareskogen på vestkysten av Norge. - NF-Rapport nr. 7/82. Nordlandsforskning. 1-31.
Sivertsen K. 1997. Geographic and environmental factors affecting the distribution of kelp beds and barren grounds and changes in biota associated with kelp reduction at sites along the Norwegian coast. Canadian Journal of Fisheries and Aquatic Sciences. 54: 2872–2887.
Sivertsen K. 2006. Overgrazing of kelp beds along the coast of Norway. Journal of Applied Phycology 18: 599-610.
Sjøtun K, Christie H, Fosså JH. 2006. Effects of kelp recruitment and sea urchin grazing on stability in kelp forest ( Laminaria hyperborea ). Marine Biology Research 2:24-32.
Steen H. 2016. Undersøkelser av A-høstefelt for tare i Sør-Trøndelag og Nord-Trøndelag i 2016. Rapport fra Havforskningen. Nr. 31-2016.
Steen H. 2019. Tilstandsvurdering av C-felt for tarehøsting i Rogaland og Sogn og Fjordane i 2019. Rapport fra Havforskningen Nr. 32-2019.
Steen H. 2020. Tilstandsvurdering av høstefelt for stortare i Møre og Romsdal og Trøndelag i 2020. Rapport fra Havforskningen Nr. 31-2020.
Steen H. 2021. Tilstandsvurdering av Høstefelt for Stortare i Rogaland og Vestland i 2021. Rapport fra Havforskningen 2021-34.
Steen H. 2022. Tilstandsvurdering av høstefelt for stortare i Møre og Romsdal og Trøndelag i 2022. Nr. 28-2022.
Steen H, Moy FE, Bodvin T. 2014a. Undersøkelser av stortarehøsting i Nord-Trøndelag og Nordland i 2013. Fisken og Havet 2014-3.
Steen H, Husa V, Moy FE, Bodvin T, Øverbø Hansen H, Sannæs H, Bosgraaf S. 2015a. Undersøkelser av stortarehøsting i Nordland i 2014. Rapport fra Havforskningen Nr. 1-2015.
Steen H, Bodvin T, Moy FE, Sannes H, Øverbø Hansen H. 2015b. Undersøkelser av stortarehøsting i Nordland i 2015. Rapport fra Havforskningen Nr. 26-2015.
Steen H, Moy FE, Bodvin T, Husa V. 2016a. Regrowth after kelp harvesting in Nord-Trøndelag, Norway. ICES Journal of Marine Science. 73(10): 2708–2720.
Steen H, Bodvin T, Moy FE, Gustad E, Øverbø Hansen H, Jelmert A, Baardsen P. 2016b. Effekter av stortarehøsting i Nordland i 2016. Rapport fra Havforskningen Nr. 38-2016.
Steen H, Norderhaug KM, Moy FE. 2018a. Tareundersøkelser i Nordland i 2017. Rapport fra Havforskningen Nr. 9-2018.
Steen H, Norderhaug KM, Moy FE. 2018b. Tareundersøkelser i Nordland i 2018. Rapport fra Havforskningen Nr. 44-2018.
Steen H, Norderhaug KM, Moy FE. 2020. Tareundersøkelser i Nordland i 2019. Rapport fra Havforskningen Nr. 9-2020.
Steen H, Christensen L, Norderhaug KM. 2023. Tilstandsvurdering av høstefelt for stortare i Rogaland og Vestland i 2023. Rapport fra Havforskningen Nr. 38-2023.
Steen H, Kartveit KH. 2024. Tilstandsvurdering av høstefelt for stortare i Møre og Romsdal og Trøndelag i 2024. Rapport fra Havforskningen Nr. 40-2024.
Teagle H, Hawkins SJ, Moore PJ, Smale DA. 2017. The role of kelp species as biogenic habitat formers in coastal marine ecosystems. Journal of Experimental Marine Biology and Ecology. 492: 81-98.
van Son TC, Nikolioudakis N, Steen H, Albretsen J, Furevik BR, Elvenes S, Moy F, Norderhaug KM. 2020. Achieving Reliable Estimates of the Spatial Distribution of Kelp Biomass. Frontiers in Marine Science. 7:107. doi: 10.3389/fmars.2022.
Wood SN, Pya N, Saefken B. 2016. Smoothing parameter and model selection for general smooth models (with discussion). Journal of the American Statistical Association 111: 1548-1575.
Zuur AF, Leno EN, Saveliev AA. 2017. Recognizing statistical dependency. Beginner’s Guide to Spatial, Temporal, and Spatial-Temporal Ecological Data Analysis with R-INLA. Newburgh, Highland Statistics Ltd. 1: 5-32.
7 - Appendix
Appendix 1. Felt, stasjon, siste høsteaktivitet, undersøkelsestidspunkt, start-, stopposisjoner (WGS84 i grader,desimaler), gjennomsnittlig dyp og midlere signifikante bølgehøyde for video- og akustikktransekter undersøkt på kyststrekningen fra Trøndelagsgrensen til Vega i Nordland i juni 2022.
Felt
St.
Høstet (år)
Dato
Kl.
Lat. start
Lon. start
Lat. end.
Lon. end.
Dyp (m)
Bølgeh. (m)
453A
f
uhøstet
15.06.2022
13:19
65,2227⁰N
11,7283⁰Ø
65,2218⁰N
11,7284⁰Ø
12
0,56
454F
c
uhøstet
15.06.2022
14:35
65,2332⁰N
11,6396⁰Ø
65,2324⁰N
11,6372⁰Ø
9
0,56
454F
d
uhøstet
15.06.2022
15:19
65,2308⁰N
11,5628⁰Ø
65,2296⁰N
11,5613⁰Ø
10
0,71
454F
e
uhøstet
15.06.2022
13:38
65,2322⁰N
11,7085⁰Ø
65,2313⁰N
11,7063⁰Ø
10
0,54
455B
b
2015
15.06.2022
15:43
65,2356⁰N
11,5474⁰Ø
65,2364⁰N
11,5441⁰Ø
9
0,70
455B
e
2015
15.06.2022
14:19
65,2390⁰N
11,6550⁰Ø
65,2380⁰N
11,6543⁰Ø
8
0,68
456G
c
2015
15.06.2022
16:19
65,2449⁰N
11,5753⁰Ø
65,2455⁰N
11,5721⁰Ø
7
0,59
457C
b
2016
15.06.2022
18:47
65,2512⁰N
11,5038⁰Ø
65,2524⁰N
11,5015⁰Ø
9
0,73
457C
d
2016
15.06.2022
16:45
65,2568⁰N
11,5931⁰Ø
65,2575⁰N
11,5908⁰Ø
7
0,73
458H
e
2016
15.06.2022
17:27
65,2647⁰N
11,5303⁰Ø
65,2660⁰N
11,5299⁰Ø
6
0,73
459D
b
2017
15.06.2022
17:49
65,2683⁰N
11,5239⁰Ø
65,2695⁰N
11,5219⁰Ø
8
0,75
460F
a
2017
16.06.2022
12:05
65,2768⁰N
11,5641⁰Ø
65,2752⁰N
11,5635⁰Ø
11
0,72
460F
c
2017
16.06.2022
09:36
65,2808⁰N
11,4756⁰Ø
65,2799⁰N
11,4774⁰Ø
11
0,82
461E
g
2015
16.06.2022
11:11
65,2875⁰N
11,5359⁰Ø
65,2874⁰N
11,5398⁰Ø
9
0,70
462G
f
2015
16.06.2022
10:48
65,2930⁰N
11,5524⁰Ø
65,2940⁰N
11,5530⁰Ø
9
0,69
462G
h
2015
16.06.2022
10:03
65,2953⁰N
11,5279⁰Ø
65,2962⁰N
11,5284⁰Ø
11
0,68
463A
h
uhøstet
16.06.2022
10:29
65,3008⁰N
11,5360⁰Ø
65,3013⁰N
11,5380⁰Ø
10
0,73
464H
e
2016
17.06.2022
09:42
65,3138⁰N
11,4428⁰Ø
65,3149⁰N
11,4451⁰Ø
8
0,99
464H
f
uhøstet
16.06.2022
17:09
65,3119⁰N
11,5147⁰Ø
65,3108⁰N
11,5141⁰Ø
10
0,75
464H
g
uhøstet
16.06.2022
17:32
65,3089⁰N
11,5317⁰Ø
65,3099⁰N
11,5339⁰Ø
8
0,79
465B
b
2016
17.06.2022
10:02
65,3199⁰N
11,4445⁰Ø
65,3210⁰N
11,4419⁰Ø
8
1,05
466F
a
2016
17.06.2022
10:42
65,3290⁰N
11,4104⁰Ø
65,3301⁰N
11,4077⁰Ø
9
1,22
466F
d
2016
16.06.2022
16:34
65,3289⁰N
11,4898⁰Ø
65,3278⁰N
11,4922⁰Ø
10
0,90
467C
a
2017
16.06.2022
13:53
65,3369⁰N
11,5747⁰Ø
65,3363⁰N
11,5786⁰Ø
9
0,84
467C
f
2017
17.06.2022
11:20
65,3366⁰N
11,4463⁰Ø
65,3354⁰N
11,4480⁰Ø
10
1,09
468G
e
2017
16.06.2022
16:05
65,3468⁰N
11,4927⁰Ø
65,3462⁰N
11,4954⁰Ø
10
0,93
469D
c
2017
16.06.2022
15:38
65,3585⁰N
11,4916⁰Ø
65,3569⁰N
11,4922⁰Ø
9
0,95
469D
d
2017
17.06.2022
12:24
65,3542⁰N
11,4379⁰Ø
65,3552⁰N
11,4404⁰Ø
11
1,13
470H
b
2017
16.06.2022
14:50
65,3621⁰N
11,5422⁰Ø
65,3602⁰N
11,5422⁰Ø
9
0,90
471E
g
2015
17.06.2022
15:57
65,3708⁰N
11,5281⁰Ø
65,3704⁰N
11,5258⁰Ø
8
0,95
471E
i
2015
17.06.2022
13:06
65,3673⁰N
11,4342⁰Ø
65,3690⁰N
11,4351⁰Ø
10
1,15
472F
h
2015
17.06.2022
14:10
65,3779⁰N
11,4970⁰Ø
65,3787⁰N
11,5006⁰Ø
9
0,97
473A
d
2016
17.06.2022
16:12
65,3907⁰N
11,4804⁰Ø
65,3907⁰N
11,4763⁰Ø
9
1,00
474G
a
2016
17.06.2022
16:55
65,3954⁰N
11,5085⁰Ø
65,3952⁰N
11,5046⁰Ø
10
0,90
474G
b
2016
17.06.2022
16:38
65,3934⁰N
11,5002⁰Ø
65,3935⁰N
11,4968⁰Ø
10
0,99
475B
a
2017
17.06.2022
17:19
65,4059⁰N
11,5161⁰Ø
65,4044⁰N
11,5125⁰Ø
9
0,90
475B
b
2017
17.06.2022
18:36
65,4034⁰N
11,5838⁰Ø
65,4021⁰N
11,5813⁰Ø
9
0,80
476H
c
2017
20.06.2022
13:49
65,4164⁰N
11,7213⁰Ø
65,4165⁰N
11,7225⁰Ø
11
0,60
477C
f
2014
20.06.2022
13:14
65,4190⁰N
11,7142⁰Ø
65,4202⁰N
11,7168⁰Ø
7
0,57
478F
g
2014
20.06.2022
12:10
65,4255⁰N
11,6850⁰Ø
65,4261⁰N
11,6889⁰Ø
9
0,57
478F
h
2014
20.06.2022
11:39
65,4317⁰N
11,6770⁰Ø
65,4329⁰N
11,6737⁰Ø
8
0,66
479D
d
2015
20.06.2022
10:34
65,4370⁰N
11,6435⁰Ø
65,4381⁰N
11,6439⁰Ø
8
0,72
479D
e
2015
20.06.2022
10:52
65,4368⁰N
11,6694⁰Ø
65,4380⁰N
11,6699⁰Ø
8
0,66
480G
c
2015
20.06.2022
10:06
65,4449⁰N
11,6717⁰Ø
65,4467⁰N
11,6717⁰Ø
9
0,50
481E
d
2017
20.06.2022
09:24
65,4521⁰N
11,7048⁰Ø
65,4513⁰N
11,7019⁰Ø
8
0,39
481E
e
2017
20.06.2022
09:03
65,4503⁰N
11,7274⁰Ø
65,4510⁰N
11,7241⁰Ø
9
0,40
482H
f
uhøstet
25.06.2022
10:04
65,4654⁰N
11,4450⁰Ø
65,4645⁰N
11,4453⁰Ø
13
1,20
483A
d
2017
22.06.2022
16:10
65,4670⁰N
11,5318⁰Ø
65,4679⁰N
11,5332⁰Ø
10
0,93
484F
a
2017
22.06.2022
08:50
65,4804⁰N
11,6798⁰Ø
65,4814⁰N
11,6819⁰Ø
8
0,49
484F
b
2017
22.06.2022
16:41
65,4762⁰N
11,6148⁰Ø
65,4754⁰N
11,6177⁰Ø
9
0,73
484F
c
2017
22.06.2022
15:07
65,4802⁰N
11,5398⁰Ø
65,4792⁰N
11,5413⁰Ø
9
0,93
485B
d
2014
22.06.2022
09:25
65,4855⁰N
11,6378⁰Ø
65,4864⁰N
11,6412⁰Ø
11
0,70
486G
e
2014
22.06.2022
09:53
65,4922⁰N
11,6468⁰Ø
65,4939⁰N
11,6473⁰Ø
7
0,61
486G
f
2014
22.06.2022
14:34
65,4975⁰N
11,5503⁰Ø
65,4983⁰N
11,5499⁰Ø
7
0,92
487C
g
2015
22.06.2022
10:51
65,5039⁰N
11,6622⁰Ø
65,5043⁰N
11,6634⁰Ø
9
0,51
488H
a
2015
22.06.2022
11:12
65,5144⁰N
11,6712⁰Ø
65,5142⁰N
11,6745⁰Ø
9
0,59
488H
c
2015
25.06.2022
14:33
65,5147⁰N
11,4795⁰Ø
65,5133⁰N
11,4793⁰Ø
10
1,06
488H
f
uhøstet
22.06.2022
12:38
65,5142⁰N
11,6248⁰Ø
65,5151⁰N
11,6266⁰Ø
7
0,71
489D
b
2014
25.06.2022
15:06
65,5184⁰N
11,4616⁰Ø
65,5178⁰N
11,4640⁰Ø
11
1,22
489D
c
2014
24.06.2022
15:57
65,5229⁰N
11,5183⁰Ø
65,5226⁰N
11,5209⁰Ø
11
1,03
489D
d
2014
22.06.2022
12:58
65,5186⁰N
11,6227⁰Ø
65,5191⁰N
11,6262⁰Ø
7
0,60
490F
a
2014
25.06.2022
13:19
65,5290⁰N
11,4597⁰Ø
65,5298⁰N
11,4580⁰Ø
10
1,08
491E
a
2013
25.06.2022
11:42
65,5368⁰N
11,4227⁰Ø
65,5352⁰N
11,4215⁰Ø
11
1,31
491E
c
2013
25.06.2022
12:53
65,5375⁰N
11,4826⁰Ø
65,5393⁰N
11,4799⁰Ø
11
1,18
492G
b
2013
25.06.2022
12:32
65,5485⁰N
11,4762⁰Ø
65,5499⁰N
11,4751⁰Ø
10
1,19
HorsværVest
1b
uhøstet
16.06.2022
12:41
65,2950⁰N
11,5958⁰Ø
65,2955⁰N
11,5978⁰Ø
8
0,69
HorsværVest
1c
uhøstet
16.06.2022
13:00
65,2991⁰N
11,5829⁰Ø
65,2985⁰N
11,5853⁰Ø
10
0,76
HorsværNord
5a
uhøstet
17.06.2022
17:43
65,4046⁰N
11,5339⁰Ø
65,4037⁰N
11,5308⁰Ø
8
0,85
HorsværNord
5c
uhøstet
19.06.2022
10:39
65,3898⁰N
11,6129⁰Ø
65,3883⁰N
11,6096⁰Ø
9
0,84
HorsværNord
5d
uhøstet
19.06.2022
11:27
65,3869⁰N
11,6443⁰Ø
65,3860⁰N
11,6474⁰Ø
7
0,74
VegaVerd.arv
7a
uhøstet
22.06.2022
13:35
65,5276⁰N
11,6056⁰Ø
65,5277⁰N
11,6083⁰Ø
7
0,72
VegaVerd.arv
7c
uhøstet
24.06.2022
17:20
65,5516⁰N
11,5739⁰Ø
65,5528⁰N
11,5759⁰Ø
7
0,96
VegaVerd.arv
7d
2013
24.06.2022
16:55
65,5478⁰N
11,5647⁰Ø
65,5492⁰N
11,5641⁰Ø
10
0,99
Appendix 2. Felt, stasjon, siste høsteaktivitet, undersøkelsestidspunkt, start-, stopposisjoner (WGS84 i grader,desimaler), gjennomsnittlig dyp og midlere signifikante bølgehøyde for video- og akustikktransekter undersøkt på kyststrekningen fra Trøndelagsgrensen til Vega i Nordland i juni 2023.
Felt
St.
Høstet (år)
Dato
Kl.
Lat. start
Lon. start
Lat. end.
Lon. end.
Dyp (m)
Bølgeh. (m)
453A
f
uhøstet
02.06.2023
10:21
65,2227⁰N
11,7283⁰Ø
65,2218⁰N
11,7284⁰Ø
12
0,56
454F
c
uhøstet
02.06.2023
11:38
65,2332⁰N
11,6396⁰Ø
65,2324⁰N
11,6372⁰Ø
9
0,56
454F
d
uhøstet
02.06.2023
12:17
65,2308⁰N
11,5628⁰Ø
65,2296⁰N
11,5613⁰Ø
11
0,71
454F
e
uhøstet
02.06.2023
10:47
65,2322⁰N
11,7085⁰Ø
65,2313⁰N
11,7063⁰Ø
10
0,54
455B
b
2023
02.06.2023
12:38
65,2356⁰N
11,5474⁰Ø
65,2364⁰N
11,5441⁰Ø
9
0,70
455B
e
2023
02.06.2023
11:17
65,2390⁰N
11,6550⁰Ø
65,2380⁰N
11,6543⁰Ø
8
0,68
456G
c
2015
02.06.2023
13:53
65,2449⁰N
11,5753⁰Ø
65,2455⁰N
11,5721⁰Ø
7
0,59
457C
b
2016
02.06.2023
13:16
65,2512⁰N
11,5038⁰Ø
65,2524⁰N
11,5015⁰Ø
10
0,73
457C
d
2016
02.06.2023
14:24
65,2568⁰N
11,5931⁰Ø
65,2575⁰N
11,5908⁰Ø
8
0,73
458H
e
2016
02.06.2023
14:56
65,2647⁰N
11,5303⁰Ø
65,2660⁰N
11,5299⁰Ø
6
0,73
459D
b
2017
02.06.2023
15:17
65,2683⁰N
11,5239⁰Ø
65,2695⁰N
11,5219⁰Ø
8
0,75
460F
a
2022
02.06.2023
17:42
65,2768⁰N
11,5641⁰Ø
65,2752⁰N
11,5635⁰Ø
10
0,72
460F
c
2022
02.06.2023
15:50
65,2808⁰N
11,4756⁰Ø
65,2799⁰N
11,4774⁰Ø
10
0,82
461E
g
2015
02.06.2023
16:29
65,2875⁰N
11,5359⁰Ø
65,2874⁰N
11,5398⁰Ø
8
0,70
462G
f
2015
02.06.2023
17:11
65,2930⁰N
11,5524⁰Ø
65,2940⁰N
11,5530⁰Ø
9
0,69
462G
h
2015
06.06.2023
09:42
65,2953⁰N
11,5279⁰Ø
65,2962⁰N
11,5284⁰Ø
10
0,68
463A
h
2022
06.06.2023
09:59
65,3008⁰N
11,5360⁰Ø
65,3013⁰N
11,5380⁰Ø
9
0,73
464H
e
2016
06.06.2023
11:18
65,3138⁰N
11,4428⁰Ø
65,3149⁰N
11,4451⁰Ø
10
0,99
464H
f
uhøstet
06.06.2023
10:40
65,3119⁰N
11,5147⁰Ø
65,3108⁰N
11,5141⁰Ø
10
0,75
464H
g
uhøstet
06.06.2023
10:18
65,3089⁰N
11,5317⁰Ø
65,3099⁰N
11,5339⁰Ø
9
0,79
465B
b
2023
06.06.2023
11:39
65,3199⁰N
11,4445⁰Ø
65,3210⁰N
11,4419⁰Ø
9
1,05
466F
a
2022
06.06.2023
12:59
65,3290⁰N
11,4104⁰Ø
65,3301⁰N
11,4077⁰Ø
10
1,22
466F
d
2022
06.06.2023
12:09
65,3289⁰N
11,4898⁰Ø
65,3278⁰N
11,4922⁰Ø
11
0,90
467C
a
2017
06.06.2023
08:56
65,3369⁰N
11,5747⁰Ø
65,3363⁰N
11,5786⁰Ø
7
0,84
467C
f
2017
06.06.2023
12:34
65,3366⁰N
11,4463⁰Ø
65,3354⁰N
11,4480⁰Ø
11
1,09
468G
e
2017
06.06.2023
14:16
65,3468⁰N
11,4927⁰Ø
65,3462⁰N
11,4954⁰Ø
10
0,93
469D
c
2017
06.06.2023
15:17
65,3585⁰N
11,4916⁰Ø
65,3569⁰N
11,4922⁰Ø
9
0,95
469D
d
2017
06.06.2023
13:43
65,3542⁰N
11,4379⁰Ø
65,3552⁰N
11,4404⁰Ø
12
1,13
470H
b
2017
06.06.2023
14:48
65,3621⁰N
11,5422⁰Ø
65,3602⁰N
11,5422⁰Ø
10
0,90
471E
g
2015
06.06.2023
16:51
65,3708⁰N
11,5281⁰Ø
65,3704⁰N
11,5258⁰Ø
7
0,95
471E
i
2015
06.06.2023
15:47
65,3673⁰N
11,4342⁰Ø
65,3690⁰N
11,4351⁰Ø
10
1,15
472F
h
2022
06.06.2023
16:22
65,3779⁰N
11,497⁰Ø
65,3787⁰N
11,5006⁰Ø
9
0,97
473A
d
2022
09.06.2023
10:05
65,3907⁰N
11,4804⁰Ø
65,3907⁰N
11,4763⁰Ø
8
1,00
474G
a
2016
09.06.2023
09:30
65,3954⁰N
11,5085⁰Ø
65,3952⁰N
11,5046⁰Ø
8
0,90
474G
b
2016
09.06.2023
09:47
65,3934⁰N
11,5002⁰Ø
65,3935⁰N
11,4968⁰Ø
8
0,99
475B
a
2023
09.06.2023
09:01
65,4059⁰N
11,5161⁰Ø
65,4044⁰N
11,5125⁰Ø
8
0,90
475B
b
2023
08.06.2023
19:40
65,4034⁰N
11,5838⁰Ø
65,4021⁰N
11,5813⁰Ø
9
0,80
476H
c
2017
08.06.2023
18:52
65,4165⁰N
11,7226⁰Ø
65,4164⁰N
11,7210⁰Ø
12
0,60
477C
f
2014
08.06.2023
18:31
65,4203⁰N
11,7167⁰Ø
65,4190⁰N
11,7141⁰Ø
8
0,57
478F
g
2022
08.06.2023
18:06
65,4260⁰N
11,6887⁰Ø
65,4255⁰N
11,6851⁰Ø
10
0,57
478F
h
2022
08.06.2023
17:41
65,4330⁰N
11,6736⁰Ø
65,4317⁰N
11,6769⁰Ø
9
0,66
479D
d
2015
06.06.2023
18:03
65,4381⁰N
11,6440⁰Ø
65,4369⁰N
11,6435⁰Ø
8
0,72
479D
e
2015
08.06.2023
17:27
65,4378⁰N
11,6700⁰Ø
65,4368⁰N
11,6696⁰Ø
9
0,66
480G
c
2015
06.06.2023
18:32
65,4467⁰N
11,6719⁰Ø
65,4449⁰N
11,6717⁰Ø
9
0,50
481E
d
2017
08.06.2023
16:28
65,4521⁰N
11,7048⁰Ø
65,4513⁰N
11,702⁰Ø
9
0,39
481E
e
2017
08.06.2023
16:52
65,4510⁰N
11,7241⁰Ø
65,4504⁰N
11,7272⁰Ø
9
0,40
482H
f
uhøstet
09.06.2023
11:06
65,4654⁰N
11,4450⁰Ø
65,4645⁰N
11,4453⁰Ø
12
1,20
483A
d
2022
09.06.2023
11:40
65,4680⁰N
11,5332⁰Ø
65,4669⁰N
11,5316⁰Ø
10
0,93
484F
a
2022
04.06.2023
10:13
65,4804⁰N
11,6799⁰Ø
65,4814⁰N
11,6819⁰Ø
8
0,49
484F
b
2022
09.06.2023
12:51
65,4763⁰N
11,6147⁰Ø
65,4754⁰N
11,6180⁰Ø
8
0,73
484F
c
2022
09.06.2023
12:01
65,4802⁰N
11,5399⁰Ø
65,4793⁰N
11,5415⁰Ø
9
0,93
485B
d
2023
09.06.2023
13:14
65,4864⁰N
11,6414⁰Ø
65,4855⁰N
11,6377⁰Ø
9
0,70
486G
e
2014
04.06.2023
12:46
65,4927⁰N
11,6467⁰Ø
65,4942⁰N
11,6473⁰Ø
10
0,61
486G
f
2014
09.06.2023
12:20
65,4983⁰N
11,5500⁰Ø
65,4974⁰N
11,5505⁰Ø
7
0,92
487C
g
2015
04.06.2023
10:45
65,5038⁰N
11,6621⁰Ø
65,5043⁰N
11,6634⁰Ø
10
0,51
488H
a
2015
09.06.2023
13:47
65,5143⁰N
11,6715⁰Ø
65,5141⁰N
11,6748⁰Ø
9
0,59
488H
c
2015
10.06.2023
10:54
65,5132⁰N
11,4793⁰Ø
65,5147⁰N
11,4796⁰Ø
9
1,06
488H
f
uhøstet
09.06.2023
14:09
65,5151⁰N
11,6267⁰Ø
65,5141⁰N
11,6249⁰Ø
8
0,71
489D
b
2014
10.06.2023
11:34
65,5178⁰N
11,4641⁰Ø
65,5184⁰N
11,4616⁰Ø
11
1,22
489D
c
2014
04.06.2023
11:49
65,5230⁰N
11,5188⁰Ø
65,5228⁰N
11,5211⁰Ø
12
1,03
489D
d
2014
09.06.2023
14:27
65,5190⁰N
11,6263⁰Ø
65,5185⁰N
11,6226⁰Ø
8
0,60
490F
a
2022
10.06.2023
10:13
65,5288⁰N
11,4599⁰Ø
65,5298⁰N
11,4580⁰Ø
9
1,08
491E
a
2013
10.06.2023
09:45
65,5351⁰N
11,4215⁰Ø
65,5367⁰N
11,4228⁰Ø
10
1,31
491E
c
2013
10.06.2023
11:26
65,5374⁰N
11,4827⁰Ø
65,5395⁰N
11,4796⁰Ø
9
1,18
492G
b
2013
10.06.2023
12:06
65,5477⁰N
11,4732⁰Ø
65,5481⁰N
11,4760⁰Ø
9
1,19
HorsværVest
1b
uhøstet
02.06.2023
18:12
65,2950⁰N
11,5963⁰Ø
65,2955⁰N
11,5982⁰Ø
6
0,69
HorsværVest
1c
uhøstet
02.06.2023
18:37
65,2992⁰N
11,5829⁰Ø
65,2985⁰N
11,5852⁰Ø
8
0,76
HorsværNord
5a
uhøstet
09.06.2023
08:41
65,4045⁰N
11,5339⁰Ø
65,4036⁰N
11,5307⁰Ø
7
0,85
HorsværNord
5c
uhøstet
08.06.2023
20:04
65,3898⁰N
11,6130⁰Ø
65,3884⁰N
11,6097⁰Ø
10
0,84
HorsværNord
5d
uhøstet
08.06.2023
20:29
65,3868⁰N
11,6444⁰Ø
65,3860⁰N
11,6474⁰Ø
8
0,74
VegaVerd.arv
7a
uhøstet
09.06.2023
14:59
65,5278⁰N
11,6086⁰Ø
65,5275⁰N
11,6056⁰Ø
8
0,72
VegaVerd.arv
7c
uhøstet
09.06.2023
15:32
65,5529⁰N
11,5759⁰Ø
65,5516⁰N
11,5737⁰Ø
10
0,96
VegaVerd.arv
7d
2013
09.06.2023
15:53
65,5493⁰N
11,5640⁰Ø
65,5478⁰N
11,5648⁰Ø
11
0,99
Appendix 3. Felt, stasjon, siste høsteaktivitet, undersøkelsestidspunkt, start-, stopposisjoner (WGS84 i grader,desimaler), gjennomsnittlig dyp og midlere signifikante bølgehøyde for video- og akustikktransekter undersøkt på kyststrekningen fra Trøndelagsgrensen til Vega i Nordland i juni 2024.
Felt
St.
Høstet (år)
Dato
Kl.
Lat. start
Lon. start
Lat. end.
Lon. end.
Dyp (m)
Bølgeh. (m)
453A
f
uhøstet
08.06.2024
11:33
65,2218⁰N
11,7283⁰Ø
65,2228⁰N
11,7281⁰Ø
12
0,56
454F
c
uhøstet
08.06.2024
12:50
65,2331⁰N
11,6395⁰Ø
65,2324⁰N
11,6372⁰Ø
9
0,56
454F
d
uhøstet
08.06.2024
13:24
65,2309⁰N
11,5627⁰Ø
65,2296⁰N
11,5613⁰Ø
10
0,71
454F
e
uhøstet
08.06.2024
11:55
65,2312⁰N
11,7063⁰Ø
65,2321⁰N
11,7084⁰Ø
9
0,54
455B
b
2023
08.06.2024
13:56
65,2356⁰N
11,5475⁰Ø
65,2365⁰N
11,5441⁰Ø
10
0,70
455B
e
2023
08.06.2024
12:21
65,2379⁰N
11,6542⁰Ø
65,2391⁰N
11,6549⁰Ø
7
0,68
456G
c
2015
08.06.2024
14:20
65,2449⁰N
11,5752⁰Ø
65,2455⁰N
11,5722⁰Ø
8
0,59
457C
b
2024
08.06.2024
14:55
65,2513⁰N
11,5039⁰Ø
65,2524⁰N
11,5016⁰Ø
10
0,73
457C
d
2024
09.06.2024
09:35
65,2568⁰N
11,5931⁰Ø
65,2575⁰N
11,5907⁰Ø
7
0,73
458H
e
2016
08.06.2024
15:22
65,2658⁰N
11,5301⁰Ø
65,2647⁰N
11,5304⁰Ø
8
0,73
459D
b
2017
08.06.2024
15:35
65,2682⁰N
11,5239⁰Ø
65,2694⁰N
11,5219⁰Ø
10
0,75
460F
a
2022
09.06.2024
10:02
65,2752⁰N
11,5637⁰Ø
65,2768⁰N
11,5642⁰Ø
10
0,72
460F
c
2022
08.06.2024
16:04
65,2799⁰N
11,4774⁰Ø
65,2808⁰N
11,4755⁰Ø
11
0,82
461E
g
2015
09.06.2024
10:25
65,2874⁰N
11,5399⁰Ø
65,2876⁰N
11,5356⁰Ø
8
0,70
462G
f
2015
09.06.2024
10:49
65,2941⁰N
11,5530⁰Ø
65,2930⁰N
11,5523⁰Ø
8
0,69
462G
h
2015
08.06.2024
18:55
65,2955⁰N
11,5277⁰Ø
65,2962⁰N
11,5614⁰Ø
11
0,68
463A
h
2022
08.06.2024
18:37
65,3014⁰N
11,5381⁰Ø
65,3009⁰N
11,5359⁰Ø
11
0,73
464H
e
2016
08.06.2024
17:10
65,3149⁰N
11,4450⁰Ø
65,3138⁰N
11,4427⁰Ø
10
0,99
464H
f
uhøstet
08.06.2024
17:49
65,3118⁰N
11,5148⁰Ø
65,3107⁰N
11,5142⁰Ø
9
0,75
464H
g
uhøstet
08.06.2024
18:09
65,3101⁰N
11,5339⁰Ø
65,3089⁰N
11,5316⁰Ø
9
0,79
465B
b
2023
08.06.2024
16:51
65,3197⁰N
11,4448⁰Ø
65,3210⁰N
11,4418⁰Ø
10
1,05
466F
a
2022
09.06.2024
13:25
65,3301⁰N
11,4079⁰Ø
65,3289⁰N
11,4109⁰Ø
10
1,22
466F
d
2022
09.06.2024
12:47
65,3288⁰N
11,4896⁰Ø
65,3276⁰N
11,4923⁰Ø
11
0,90
467C
a
2024
09.06.2024
12:09
65,3368⁰N
11,5749⁰Ø
65,3362⁰N
11,5711⁰Ø
8
0,84
467C
f
2024
09.06.2024
13:53
65,3366⁰N
11,4464⁰Ø
65,3355⁰N
11,4481⁰Ø
11
1,09
468G
e
2017
09.06.2024
14:19
65,3468⁰N
11,4925⁰Ø
65,3461⁰N
11,4958⁰Ø
10
0,93
469D
c
2017
09.06.2024
15:17
65,3583⁰N
11,4917⁰Ø
65,3568⁰N
11,4923⁰Ø
9
0,95
469D
d
2017
09.06.2024
15:45
65,3551⁰N
11,4403⁰Ø
65,3541⁰N
11,4378⁰Ø
12
1,13
470H
b
2017
09.06.2024
14:48
65,3622⁰N
11,5421⁰Ø
65,3602⁰N
11,5421⁰Ø
10
0,90
471E
g
2015
09.06.2024
17:09
65,3708⁰N
11,5279⁰Ø
65,3704⁰N
11,5255⁰Ø
7
0,95
471E
i
2015
09.06.2024
16:07
65,3689⁰N
11,4352⁰Ø
65,3673⁰N
11,4343⁰Ø
10
1,15
472F
h
2022
09.06.2024
16:41
65,3788⁰N
11,5010⁰Ø
65,3779⁰N
11,4967⁰Ø
9
0,97
473A
d
2022
13.06.2024
12:24
65,3907⁰N
11,4804⁰Ø
65,3906⁰N
11,4762⁰Ø
8
1,00
474G
a
2016
09.06.2024
17:36
65,3955⁰N
11,5089⁰Ø
65,3952⁰N
11,5044⁰Ø
9
0,90
474G
b
2016
13.06.2024
12:45
65,3934⁰N
11,5001⁰Ø
65,3942⁰N
11,4968⁰Ø
8
0,99
475B
a
2023
13.06.2024
11:50
65,4059⁰N
11,5161⁰Ø
65,4044⁰N
11,5125⁰Ø
8
0,90
475B
b
2023
13.06.2024
11:01
65,4034⁰N
11,5837⁰Ø
65,4021⁰N
11,5814⁰Ø
9
0,80
476H
c
2017
12.06.2024
18:13
65,4164⁰N
11,7210⁰Ø
65,4165⁰N
11,7226⁰Ø
11
0,60
477C
f
2024
12.06.2024
17:56
65,4191⁰N
11,7143⁰Ø
65,4203⁰N
11,7168⁰Ø
8
0,57
478F
g
2022
12.06.2024
17:43
65,4255⁰N
11,6849⁰Ø
65,4260⁰N
11,6886⁰Ø
10
0,57
478F
h
2022
12.06.2024
17:13
65,4317⁰N
11,6770⁰Ø
65,4330⁰N
11,6736⁰Ø
9
0,66
479D
d
2015
12.06.2024
16:41
65,4369⁰N
11,6436⁰Ø
65,4381⁰N
11,6440⁰Ø
10
0,72
479D
e
2015
12.06.2024
16:59
65,4368⁰N
11,6695⁰Ø
65,4379⁰N
11,6700⁰Ø
9
0,66
480G
c
2015
12.06.2024
16:15
65,4448⁰N
11,6718⁰Ø
65,4467⁰N
11,6718⁰Ø
10
0,50
481E
d
2017
12.06.2024
15:47
65,4513⁰N
11,7019⁰Ø
65,4521⁰N
11,7049⁰Ø
9
0,39
481E
e
2017
12.06.2024
14:29
65,4510⁰N
11,7243⁰Ø
65,4504⁰N
11,7272⁰Ø
9
0,40
482H
f
uhøstet
12.06.2024
14:17
65,4654⁰N
11,4450⁰Ø
65,4644⁰N
11,4454⁰Ø
14
1,20
483A
d
2022
10.06.2024
11:48
65,4669⁰N
11,5317⁰Ø
65,4680⁰N
11,5332⁰Ø
11
0,93
484F
a
2022
10.06.2024
10:38
65,4804⁰N
11,6799⁰Ø
65,4814⁰N
11,6820⁰Ø
7
0,49
484F
b
2022
10.06.2024
11:08
65,4763⁰N
11,6146⁰Ø
65,4753⁰N
11,6183⁰Ø
9
0,73
484F
c
2022
10.06.2024
12:09
65,4802⁰N
11,5399⁰Ø
65,4792⁰N
11,5416⁰Ø
10
0,93
485B
d
2023
10.06.2024
13:03
65,4855⁰N
11,6378⁰Ø
65,4864⁰N
11,6415⁰Ø
10
0,70
486G
e
2014
10.06.2024
13:33
65,4928⁰N
11,6469⁰Ø
65,4941⁰N
11,6474⁰Ø
8
0,61
486G
f
2014
10.06.2024
12:30
65,4983⁰N
11,5499⁰Ø
65,4974⁰N
11,5505⁰Ø
9
0,92
487C
g
2024
10.06.2024
13:56
65,5038⁰N
11,6622⁰Ø
65,5043⁰N
11,6635⁰Ø
10
0,51
488H
a
2015
10.06.2024
14:11
65,5143⁰N
11,6714⁰Ø
65,5141⁰N
11,6747⁰Ø
10
0,59
488H
c
2015
12.06.2024
13:07
65,5148⁰N
11,4796⁰Ø
65,5133⁰N
11,4794⁰Ø
10
1,06
488H
f
uhøstet
10.06.2024
14:36
65,5141⁰N
11,6248⁰Ø
65,5151⁰N
11,6268⁰Ø
8
0,71
489D
b
2014
12.06.2024
12:48
65,5184⁰N
11,4615⁰Ø
65,5178⁰N
11,4641⁰Ø
11
1,22
489D
c
2014
12.06.2024
13:32
65,5230⁰N
11,5188⁰Ø
65,5228⁰N
11,5210⁰Ø
11
1,03
489D
d
2014
10.06.2024
14:52
65,5185⁰N
11,6224⁰Ø
65,5191⁰N
11,6262⁰Ø
8
0,60
490F
a
2022
12.06.2024
12:26
65,5298⁰N
11,4580⁰Ø
65,5289⁰N
11,4599⁰Ø
9
1,08
491E
a
2013
12.06.2024
12:02
65,5351⁰N
11,4215⁰Ø
65,5367⁰N
11,4227⁰Ø
10
1,31
491E
c
2013
12.06.2024
11:22
65,5395⁰N
11,4797⁰Ø
65,5375⁰N
11,4827⁰Ø
10
1,18
492G
b
2013
12.06.2024
11:02
65,5477⁰N
11,4731⁰Ø
65,5481⁰N
11,4761⁰Ø
9
1,19
HorsværVest
1b
uhøstet
09.06.2024
11:17
65,2950⁰N
11,5961⁰Ø
65,2955⁰N
11,5981⁰Ø
7
0,69
HorsværVest
1c
uhøstet
09.06.2024
11:30
65,2985⁰N
11,5851⁰Ø
65,2992⁰N
11,5829⁰Ø
9
0,76
HorsværNord
5a
uhøstet
13.06.2024
11:31
65,4036⁰N
11,5307⁰Ø
65,4045⁰N
11,5338⁰Ø
7
0,85
HorsværNord
5c
uhøstet
13.06.2024
10:32
65,3899⁰N
11,6131⁰Ø
65,3884⁰N
11,6097⁰Ø
10
0,84
HorsværNord
5d
uhøstet
13.06.2024
10:10
65,3861⁰N
11,6473⁰Ø
65,3868⁰N
11,6444⁰Ø
8
0,74
VegaVerd.arv
7a
uhøstet
10.06.2024
15:15
65,5276⁰N
11,6056⁰Ø
65,5278⁰N
11,6086⁰Ø
9
0,72
VegaVerd.arv
7c
uhøstet
10.06.2024
15:43
65,5517⁰N
11,5738⁰Ø
65,5528⁰N
11,5759⁰Ø
9
0,96
VegaVerd.arv
7d
2013
10.06.2024
16:00
65,5478⁰N
11,5649⁰Ø
65,5493⁰N
11,5639⁰Ø
11
0,99
Appendix 4. Feltnavn, stasjonsnavn, høstestatus, tidspunkt, posisjon, innsamlingsdyp, midlere signifikant bølgehøyde (MSWH) og antall planter undersøkt på innsamlingsstasjoner for stortare på strekningen fra Trøndelagsgrensen til Vega i Nordland i 2022-24.
Felt
St.
Høstestatus
Dato
Kl.
Latitud
Longitud
Dyp (m)
Bølgeh. (m)
Pl.
HorsværVest
1b
Referanse
02.06.2023
18:25
65,2953⁰N
11,5970⁰Ø
5
0,69
10
HorsværVest
1c
Referanse
02.06.2023
18:50
65,2989⁰N
11,5837⁰Ø
6
0,76
11
HorsværNord
5a
Referanse
17.06.2022
18:15
65,4039⁰N
11,5311⁰Ø
6
0,87
10
HorsværNord
5c
Referanse
19.06.2022
11:00
65,3889⁰N
11,6101⁰Ø
7
0,83
10
HorsværNord
5d
Referanse
19.06.2022
12:00
65,3854⁰N
11,6463⁰Ø
8
0,74
10
VegaVerd.arv
7a
Referanse
22.06.2022
13:55
65,5276⁰N
11,6068⁰Ø
8
0,75
10
VegaVerd.arv
7c
Referanse
24.06.2022
17:45
65,5524⁰N
11,5751⁰Ø
7
0,96
11
VegaVerd.arv
7d
Referanse
10.06.2024
16:25
65,5486⁰N
11,5642⁰Ø
6
0,99
10
470H
b
5 år etter høst.
16.06.2022
15:15
65,3618⁰N
11,5416⁰Ø
5
0,9
10
475B
a
5 år etter høst.
17.06.2022
18:00
65,4051⁰N
11,5137⁰Ø
8
0,9
10
484F
c
5 år etter høst.
22.06.2022
15:25
65,4799⁰N
11,5399⁰Ø
6
0,93
10
477C
f
8 år etter høst.
20.06.2022
14:40
65,4193⁰N
11,7140⁰Ø
6
0,66
10
478F
g
8 år etter høst.
20.06.2022
14:55
65,4257⁰N
11,6867⁰Ø
7
0,57
10
478F
h
8 år etter høst.
20.06.2022
15:15
65,4320⁰N
11,6760⁰Ø
6
0,66
10
456G
c
8 år etter høst.
02.06.2023
14:05
65,2452⁰N
11,5735⁰Ø
5
0,6
10
461E
g
8 år etter høst.
02.06.2023
16:50
65,2875⁰N
11,5362⁰Ø
7
0,69
10
462G
f
8 år etter høst.
02.06.2023
17:25
65,2933⁰N
11,5524⁰Ø
8
0,67
10
471E
g
8 år etter høst.
06.06.2023
17:15
65,3706⁰N
11,5267⁰Ø
6
0,94
10
474G
a
8 år etter høst.
09.06.2024
17:55
65,3956⁰N
11,5087⁰Ø
7
0,89
10
479D
d
8 år etter høst.
06.06.2023
18:20
65,4376⁰N
11,6438⁰Ø
6
0,72
10
480G
c
8 år etter høst.
06.06.2023
19:00
65,4466⁰N
11,6720⁰Ø
8
0,48
10
487C
g
8 år etter høst.
04.06.2023
11:05
65,5040⁰N
11,6628⁰Ø
5
0,51
10
488H
c
8 år etter høst.
10.06.2023
11:10
65,5139⁰N
11,4795⁰Ø
8
1,13
10
Appendix 5. Tukey test på forskjeller mellom innsamlingsstasjoner for stortare i henholdsvis referanseområder, felt høstet 5 år tidligere og felt høstet 8 år tidligere. Signifikante p-verdier (<0,05) er understreket.
Responsvariabel
Sammenligning
Differanse
95 % konf. int.
p verdi
Midlere signifikant bølgehøyde (m)
Referanse vs 5 år etter høst.
-0,086
(-0,35 - 0,18)
0,692
Referanse vs 8 år etter høst.
0,114
(-0,06 - 0,29)
0,263
8 år vs 5 år etter høst.
-0,200
(-0,45 - 0,05)
0,136
Innsamlingsdyp (m)
Referanse vs 5 år etter høst.
0,292
(-1,66 - 2,24)
0,924
Referanse vs 8 år etter høst.
0,183
(-1,13 - 1,50)
0,934
8 år vs 5 år etter høst.
0,108
(-1,75 - 1,97)
0,990
Alder (år)
Referanse vs 5 år etter høst.
0,933
(-1,26 - 3,13)
0,539
Referanse vs 8 år etter høst.
0,500
(-0,98 - 1,98)
0,674
8 år vs 5 år etter høst.
0,433
(-1,66 - 2,53)
0,861
Stipeslengde (cm)
Referanse vs 5 år etter høst.
-18,72
(-72,4 - 34,9)
0,657
Referanse vs 8 år etter høst.
-15,08
(-51,2 - 21,1)
0,552
8 år vs 5 år etter høst.
-3,633
(-54,8 - 47,5)
0,982
Stipesvekt (g)
Referanse vs 5 år etter høst.
-143,0
(-752 - 466)
0,825
Referanse vs 8 år etter høst.
-146,4
(-557 - 264)
0,645
8 år vs 5 år etter høst.
3,533
(-577 - 584)
1,000
Laminavekt (g)
Referanse vs 5 år etter høst.
172,6
(-74,3 - 420)
0,206
Referanse vs 8 år etter høst.
69,42
(-97,1 - 236)
0,552
8 år vs 5 år etter høst.
103,2
(-132 - 339)
0,520
Kråkeboller (antall hapter-1)
Referanse vs 5 år etter høst.
0,215
(-0,40 - 0,83)
0,653
Referanse vs 8 år etter høst.
0,090
(-0,32 - 0,50)
0,847
8 år vs 5 år etter høst.
0,125
(-0,46 - 0,71)
0,851
Epifyttvekt (g)
Referanse vs 5 år etter høst.
75,04
(-75,8 - 226)
0,434
Referanse vs 8 år etter høst.
27,40
(-74,2 - 129)
0,777
8 år vs 5 år etter høst.
47,63
(-96,2 - 191)
0,684
Epifyttdiversitet (grupper lok.-1)
Referanse vs 5 år etter høst.
2,833
(0,55 - 5,12)
0,014
Referanse vs 8 år etter høst.
0,917
(-0,62 - 2,46)
0,310
8 år vs 5 år etter høst.
1,917
(-0,26 - 4,09)
0,091
Stilktilvekst (cm2 år-1)
Referanse vs 5 år etter høst.
-0,483
(-0,90 - -0,07)
0,021
Referanse vs 8 år etter høst.
-0,230
(-0,51 - 0,05)
0,119
8 år vs 5 år etter høst.
-0,253
(-0,65 - 0,14)
0,259
Appendix 6. Felt, høsteår og høstekvantum i perioden 2022-24 (Kilde: Fiskeridirektoratet). Sum av modellert stortarebiomasse og gjennomsnittlig uttaksgrad totalt i dybdeintervallet 2-20 m (dybdeintervall for tarehøsting), med grenser for 2,5 % og 97,5 % konfidensintervall per felt. Felt som har vært åpne for høsting i perioden 2022-24 er markert i rødt.
Felt
Høsteår
Høste-kvant. (tonn)
Biomasse 2-20 m (tonn)
Biomasse 2-20 m (2,5 % konf. grense) (tonn)
Biomasse 2-20m (97,5 % konf. grense) (tonn)
Uttak av biomasse 2-20 m (%)
Uttak av biomasse 2-20 m (2,5 % konf. grense) (%)
Uttak av biomasse 2-20 m (97,5 % konf. grense) (%)