This report presents preliminary results from the mapping of biotopes and vulnerable habitats in the North Sea and Skagerrak conducted by the MAREANO programme. The results are based on field observations collected during seven research cruises carried out between 2022 and 2025 using the research vessels G.O. Sars and Kronprins Haakon. The mapping covers priority areas related to offshore wind planning, assessment of proposed Particularly Valuable and Vulnerable Areas (SVO) in the Norwegian Trench, and follow-up of commitments to conserve Vulnerable Marine Ecosystems (VME). A total of 552 video stations were investigated at depths between 40 and 700 m. The analyses include 523 stations located within the mapped areas. Video data from approximately 200 m long transects were analysed using Detrended Correspondence Analysis (DCA) and TWINSPAN classification, resulting in twelve biotope groups reflecting gradients in depth, substrate composition and the occurrence of habitat-forming species. The most species-rich biotopes occur in coastal and intermediate-depth areas with complex substrates, whereas deeper parts of the Skagerrak and the Norwegian Trench are dominated by muddy sediments and sea pen communities. In total, 472 taxa were recorded, with the highest species richness between 100 and 250 m depth. VMEs were recorded at 30% of the stations. Sea pen communities were the most widespread habitat type, particularly in deep sedimentary areas of the Skagerrak and the Norwegian Trench, dominated by Funiculina quadrangularis and Kophobelemnon stelliferum. Sponge communities constituted the second most important group of vulnerable habitats and included both sponge grounds on hard substrates and sponge spicule beds in more muddy environments at intermediate depths, particularly in the Utsira coastal belt. Soft-bottom coral gardens of Isidella lofotensis, cup coral communities dominated by Flabellum macandrewi, and horse mussel beds of Modiolus modiolus were recorded at relatively few localities and appear to have a more fragmented distribution than sea pen and sponge communities. Several of the biotopes identified in the analyses are characterised by indicator species of vulnerable habitats, but the relationship between biotopes and VME types is not one-to-one. Some VME types occur in several biotopes, while some biotopes contain multiple VME elements. This reflects that biotopes describe entire faunal assemblages along environmental gradients, whereas VME classification is based on the occurrence and density of particular habitat-forming species. It should be emphasised that the MAREANO mapping so far covers only parts of the North Sea and Skagerrak. When combined with previous observations of sea pens and Isidella in adjacent areas, the results suggest that additional occurrences of such VMEs likely exist in areas that have not yet been mapped. The results demonstrate a clear depth- and substrate-related distribution of vulnerable habitats. At the same time, many indicator species occur at low densities that do not always meet operational threshold values for habitat classification, illustrating methodological challenges associated with the use of fixed criteria. Human impact was documented through observations of trawl marks, marine litter and mechanically damaged colonies of habitat-forming species. Trawl marks were most frequent in the Norwegian Trench and the outer Oslofjord, and the overlap with damaged sea pens indicates physical disturbance of vulnerable habitats in some areas, particularly in the outer Oslofjord. The results provide an updated knowledge base for marine spatial planning and management in the North Sea and Skagerrak. However, they are based on preliminary field annotations, and further analyses will provide a more detailed understanding of species diversity and community structure.
Kartlegging av biotoper og sårbare habitater i Nordsjøen og Skagerrak
— Foreløpige resultater fra syv tokt i Mareano
Rapportserie:
Rapport fra havforskningen 2026-16
ISSN: 1893-4536
Publisert: 24.03.2026
Prosjektnr: 16155
Forskningsgruppe(r):
Bunnsamfunn
Program:
Nordsjøen,
Marine prosesser og menneskelig påvirkning
Forskningsgruppeleder(e):
Sigurd Heiberg Espeland (Bunnsamfunn)
Godkjent av:
Forskningsdirektør(er):
Geir Lasse Taranger
Programleder(e):
Frode Vikebø
English summary
Sammendrag
Rapporten presenterer foreløpige resultater fra Mareanos kartlegging av biotoper og sårbare habitater i Nordsjøen og Skagerrak, basert på feltregistreringer fra syv tokt i perioden 2022–2025 med FF G.O. Sars og FF Kronprins Haakon. Kartleggingen dekker prioriterte områder knyttet til havvindplanlegging, vurdering av foreslåtte SVO-områder (svært verdifulle og sårbare områder) i Norskerenna og oppfølging av forpliktelser om bevaring av sårbare marine økosystemer (VME). Totalt ble 552 videostasjoner undersøkt på 40–700 m dyp. Analysene omfatter 523 stasjoner innenfor Mareanos kartleggingsområder. Videodata fra ca. 200 m lange transekter ble analysert ved hjelp av DCA og TWINSPAN, og resulterte i tolv biotopgrupper som reflekterer gradienter i dyp, substrat og forekomst av habitatdannende arter. De mest artsrike biotopene forekommer i kystnære og mellomdype områder med komplekst substrat, mens dypere deler av Skagerrak og Norskerenna domineres av slamrike sedimenter og sjøfjærsamfunn. Totalt ble 472 taxa registrert, med høyest artsmangfold mellom 100 og 250 m. Sårbare habitater ble registrert på 30 % av stasjonene. Sjøfjærsamfunn var den mest utbredte habitattypen og forekom særlig i dype sedimentområder i Skagerrak og Norskerenna, dominert av Funiculina quadrangularis og Kophobelemnon stelliferum. Svampsamfunn utgjorde den nest viktigste gruppen av sårbare habitater og omfattet både svampskog i hardbunnsområder og svampspikelbunn i mer slamrike miljøer i mellomdyp sone, særlig i Kystbelte Utsira. Bløtbunnskorallskog (Isidella lofotensis), begerkorallsamfunn (Flabellum macandrewi) og O-skjellbanker (Modiolus modiolus) ble registrert på få lokaliteter og fremstår som mer fragmentert utbredt enn sjøfjær- og svampsamfunn. Flere av biotopene identifisert i analysene er karakterisert av indikatorarter for sårbare habitater, men sammenhengen mellom biotoper og VME-typer er ikke entydig. Enkelte VME-typer forekommer i flere biotoper, og enkelte biotoper inneholder flere ulike VME-elementer. Dette reflekterer at biotopene beskriver helhetlige faunasamfunn langs miljøgradienter, mens VME-klassifiseringen er basert på forekomst og tetthet av bestemte strukturdannende arter. Det må imidlertid understrekes at Mareanos kartlegging så langt kun omfatter deler av Nordsjøen og Skagerrak. Sammenstilt med tidligere undersøkelser av sjøfjær og Isidella i tilgrensende områder, er det grunn til å anta at flere forekomster av sårbare habitater finnes i områder som ennå ikke er kartlagt. Resultatene viser en tydelig dybde- og substratavhengig fordeling av sårbare habitater. Samtidig opptrer mange indikatorarter i lave tettheter som ikke alltid tilfredsstiller operasjonelle terskelverdier for habitatklassifisering, noe som illustrerer metodiske utfordringer ved bruk av faste kriterier. Menneskelig påvirkning ble dokumentert gjennom registrering av trålspor, marint avfall og mekanisk skadede kolonier av habitatdannende arter. Trålspor var hyppigst i Norskerenna og Ytre Oslofjord, og overlapp med skadde sjøfjær indikerer fysisk påvirkning på sårbare habitater i enkelte områder, spesielt i Ytre Oslofjord. Resultatene gir et oppdatert kunnskapsgrunnlag for marin arealforvaltning i Nordsjøen og Skagerrak, men er basert på foreløpige feltannoteringer. Videre analyser vil gi et mer presist bilde av artsmangfold og samfunnsstruktur.
1 - Innledning
Mareano-programmet ble etablert i 2005 for å kartlegge dybde, bunnforhold, biologisk mangfold og forurensning av havbunnen i norske havområder. Programmet er et samarbeid mellom Havforskningsinstituttet, Norges geologiske undersøkelse og Kartverket, og leverer kunnskap som er sentral for økosystembasert forvaltning av havområdene (Mareano.no; Buhl-Mortensen et al. 2015a; 2015b; Gonzalez-Mirelis et al. 2015; Marques et al 2024). Data fra Mareano brukes blant annet i arbeidet med de helhetlige forvaltningsplanene for norske havområder, og i vurderinger av særlig verdifulle og sårbare områder (SVO) (Eriksen et al. 2021).
Kartleggingen av Nordsjøen og Skagerrak har siden 2022 vært prioritert på bakgrunn av flere aktuelle forvaltningsbehov. Det er behov for basiskunnskap om havbunnen i områder som er aktuelle for utbygging av havvind, der både fysiske og biologiske forhold må inngå i konsekvensutredninger og arealplanlegging. Videre er det behov for mer detaljert kunnskap om foreslåtte SVO-områder, særlig i Norskerenna, som er et kjerneområde for dypvannsfauna og økologiske nøkkelfunksjoner (Eriksen et al. 2021). Tidligere undersøkelser har dokumentert omfattende forekomster av sjøfjærsamfunn og bambuskorallskog i Skagerrak og Norskerenna (Buhl-Mortensen et al. 2023). I tillegg har områdene i Nordsjøen en sentral rolle for flere marine ressurser (Eriksen et al. 2021). Deler av Nordsjøen og Skagerrak er viktige gytefelt for tobis (Ammodytes marinus), en nøkkelart i økosystemet og en viktig ressurs for sjøfugl og andre fiskebestander. Områdene er også viktige for gyte- og oppvekstområder for kommersielt viktige bestander, inkludert torsk (Gadus morhua), hyse (Melanogrammus aeglefinus) og sild (Clupea harengus). Kunnskap om habitatforholdene i disse områdene er derfor nødvendig både for å sikre bevaring av økologiske funksjoner og for en bærekraftig forvaltning av fiskeressursene med stor økonomisk verdi.
Sentrale deler av kartleggingen omfatter identifisering og avgrensning av sårbare marine økosystemer (Vulnerable Marine Ecosystems, VME). Begrepet VME er utviklet og operasjonalisert gjennom internasjonale rammeverk, særlig av Food and Agriculture Organization (FAO 2009) og OSPAR Commission (OSPAR 2008). Selv om definisjonene varierer noe i form og formål, har de betydelig overlapp når det gjelder hvilke artsgrupper som anses som indikatorer på sårbarhet. Felles kriterier omfatter forekomst av habitatdannende og langsomt voksende arter, høy strukturell kompleksitet, lav motstandsdyktighet mot fysisk påvirkning og begrenset reetableringsevne. Typiske VME-indikatorer inkluderer kaldtvannskoraller (Scleractinia og Gorgonacea), svampsamfunn, sjøfjær (Pennatulacea), og O-skjellbanker. Fordelingen av flere av disse artene i Nord-Atlanteren er nært knyttet til batymetri og hydrografiske forhold (Buhl-Mortensen et al. 2015). I denne rapporten benyttes en operasjonell tilnærming til VME-begrepet der overordnede habitatklasser (for eksempel korallskog og svampsamfunn) deles inn i undertyper basert på dominerende indikatorarter og substratforhold. Denne inndelingen gjør det mulig å skille mellom økologisk ulike samfunn innenfor samme hovedkategori, samtidig som klassifiseringen er forankret i internasjonale kriterier.
VME-rammeverket utvikles kontinuerlig i takt med ny kunnskap. Nye arter og organismegrupper vurderes jevnlig som potensielle indikatorer på sårbare økosystemer. Et eksempel er sylindersjøroser (Cerianthidae), som er foreslått som VME-indikatorer i NAFO-området basert på kriterier om tetthet, habitatdannende funksjon og sårbarhet for fysisk forstyrrelse (Murillo et al. 2011). I denne rapporten er forekomster av sylindersjøroser vurdert i lys av disse kriteriene og inkludert som en egen undertype der tetthetskrav er oppfylt.
Denne rapporten presenterer foreløpige resultater fra syv tokt gjennomført i perioden 2022–2025 med forskningsfartøyene G.O. Sars og Kronprins Haakon. Materialet består av videodata fra 552 lokaliteter på dyp mellom 40 og 700 meter i Nordsjøen og Skagerrak. Observasjonene er analysert med ordinasjons- og klassifikasjonsmetoder for å identifisere biotoper og kartfeste forekomster av sårbare habitater. Resultatene gir et første regionalt overblikk over biotopmønstre og VME-forekomster, men er basert på foreløpige feltannoteringer og videodata alene. Videre bearbeiding, inkludert mer detaljert artsidentifikasjon og integrering av prøvetakingsdata, vil gi et mer presist bilde av samfunnsstruktur, habitatutbredelse og økologiske sammenhenger. Rapporten må derfor sees som et første kunnskapsgrunnlag som vil bli videreutviklet i kommende analyser og publikasjoner.
2 - Materiale og metode
2.1 - Undersøkelsesområder
Kartleggingen omfatter utvalgte deler av Nordsjøen og Skagerrak, med hovedvekt på områder identifisert som aktuelle for utbygging av havvind (Nordsjøplatået og nordlig del av Norskerenna) samt områder foreslått som SVO (Norskerenna, Skagerrak–Ytre Oslofjord/NS4 og kystbelter i Skagerrak og Nordsjøen). Områdene representerer ulike naturgeografiske og oseanografiske forhold og dekker både grunne, vindpåvirkede platåområder og dype bassenger med mer stabile miljøforhold. Samlet utgjør de viktige leveområder for en rekke bentiske samfunn. Skagerrak er karakterisert av store dybdeforskjeller, der Norskerenna danner et dypt basseng med relativt stabile hydrografiske forhold sammenlignet med de grunnere områdene mot Danmark og i Kattegat. I de sørlige og grunne delene av Nordsjøen (<50 m) er sirkulasjonen i større grad vinddrevet, med en dominerende transport mot nord og øst.
Vannmassene i regionen består av en dynamisk blanding av atlantisk vann, kystvann og ferskvannspåvirkede vannmasser fra Østersjøen og store skandinaviske elver. Atlantisk vann strømmer inn i Nordsjøen fra nord og vest og følger i stor grad Norskerenna inn i Skagerrak på dypere nivåer (>100 m). I Skagerrak møter dette vannet lettere og mindre saltholdige overflatevannmasser fra Kattegat og Østersjøen, tilført ferskvann fra blant annet Glomma, Göta älv og Drammenselva. Tetthetsforskjeller fører til en lagdeling der brakkere vann ligger over det saltere atlantiske vannet. Samspillet mellom atlantisk vann og ferskvannstilførsel danner Den norske kyststrømmen, som transporterer ferskvannspåvirket kystvann nordover langs norskekysten. Kystvannet har lavere og mer variabel saltholdighet og større sesongmessige temperatursvingninger enn det underliggende atlantiske vannet. Denne kombinasjonen av topografi, vannmasseutveksling og ferskvannstilførsel skaper tydelige miljøgradienter i temperatur og saltholdighet, som er sentrale for fordelingen av arter og habitater i undersøkelsesområdet.
Nordsjøplatået Fra dette området dekker Mareanos kartlegging områdene Sørvest F, NSJ-1 og NSJ-2 (Figur 1). Området består i hovedsak av grunne områder (<200 m) dominert av sand- og grusbunn, avbrutt av partier med bløtbunn og enkelte topografiske strukturer som sandbølger og banker. Store deler av platået var eksponert over havflaten under tidligere istider, og er formet av en kombinasjon av marin sedimentasjon, erosjon og istidsrelaterte prosesser. Dette har skapt en variert bunnmorfologi som fungerer som viktige leveområder for en rekke arter, inkludert fiskebestander med stor økologisk og økonomisk betydning.
Norskerenna omfatter i denne rapporten delområdene kalt Norskerenna, Utsira Nord og Vestavind B (Figur 1). Norskerenna er en markert, dyp renne som strekker seg fra Skagerrak, langs norskekysten, og nordover til Norskehavet. Den ble i stor grad dannet gjennom erosjon fra ismassene under istidene, og har dyp ned mot 700 m. Området kjennetegnes av et karakteristisk dypvannsfauna-samfunn, og tidligere undersøkelser har dokumentert forekomster av sårbare habitater, inkludert sjøfjærsamfunn og bambuskorallskog.
Skagerrak omfatter i denne rapporten delområdene kalt Skagerrak og Ytre Oslofjord (NS4). Skagerrak dekker også de østlige og dypeste deler av Norskerenna med dyp ned til 725 m. Ytre Oslofjord dekker et stort dybdespenn fra kystnære områder med blandede bunnforhold til dypere deler hvor Norskerenna møter den ytterste delen av Oslofjorden, med kupert bunn ofte med eksponert fjell.
Kystbelte-områder Flere kystnære områder med kystbelte Skagerrak (KB Skagerrak) i øst, kystbelte Jæren (KB Jæren) fra Farsund til Egersund, og kystbelte Utsira fra vest av Stavanger til vest av Utsira (Figur1) dekker store dybdegradienter og variert bunnforhold.
2.2 - Feltarbeid og innsamlingsmetoder
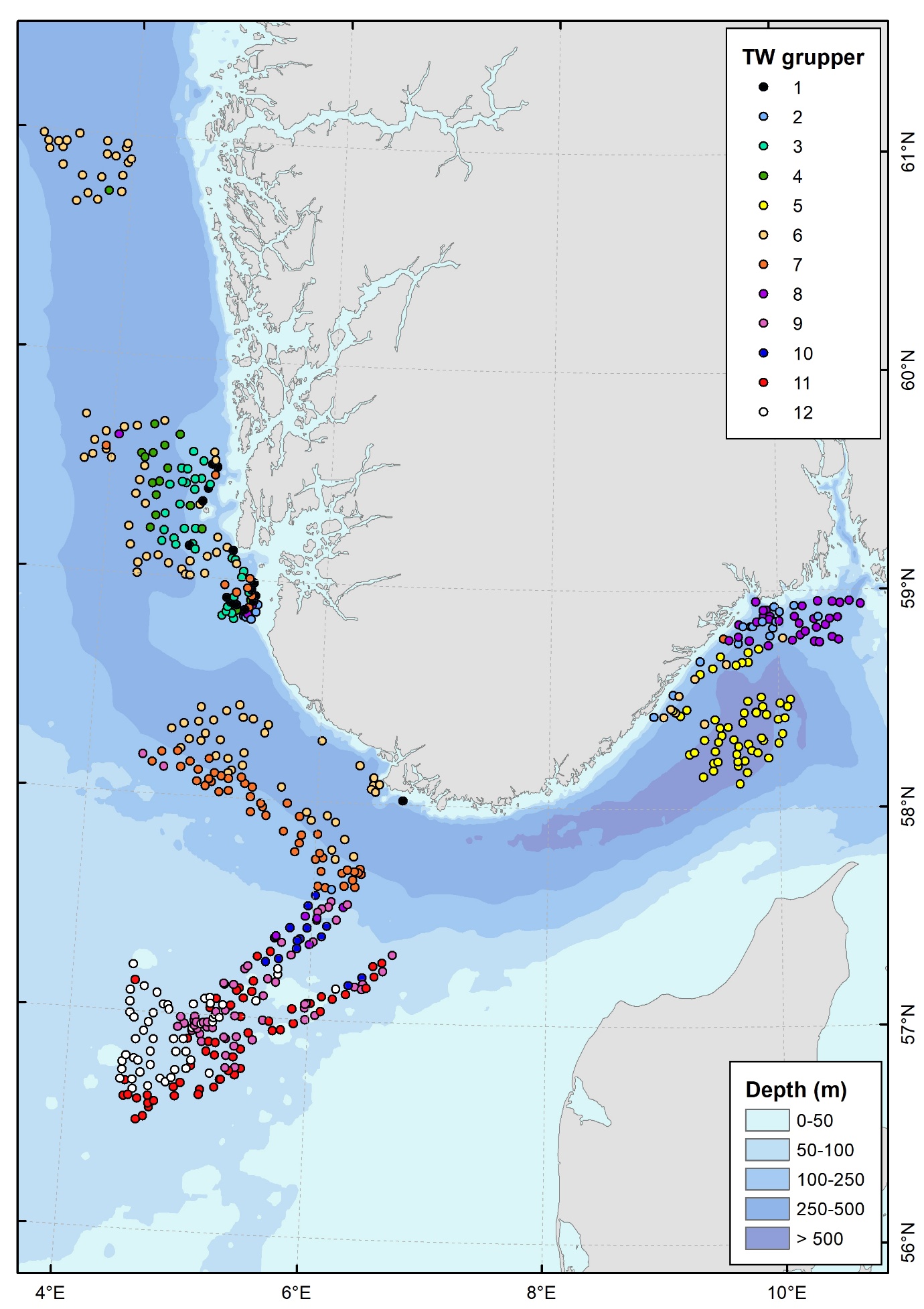
Datainnsamlingen ble gjennomført på syv tokt med forskningsfartøyene G.O. Sars og Kronprins Haakon i perioden 2022 til 2025. Totalt ble 552 lokaliteter undersøkt, på dyp mellom 40 og 700 m (Figur 1). Under perioder med dårlig vær i åpne havområder ble tiden benyttet til undersøkelser i mer beskyttede fjordområder (bl.a. Boknafjorden, Fedafjorden, Hardangerfjorden og Oslofjorden). Disse stasjonene ligger utenfor Mareanos prioriterte kartleggingsområder og er derfor ekskludert fra videre analyser av biologiske samfunn og biotoper i denne rapporten. Kartet nedenfor (Figur 1) gir en oversikt over hvilke delområder som inngår i rapporten og danner grunnlaget for den regionale tolkningen av biotopmønstre.
På en tiendedel av stasjonene ble det i tillegg til videofilming også tatt bunnprøver med ulike redskaper. Disse stasjonene klassifiseres som fullstasjoner (FS). Resten av stasjonene er rene videostasjoner (VL), hvor bare visuell dokumentasjon av bunnen ble gjennomført.
Prøvetaking på fullstasjonene omfattet bruk av:
-
Grabb
-
Boxcorer
-
Multicorer
-
Bomtrål
-
RP-slede
De syv toktene til Nordsjøen/Skagerrak utgjør til sammen 80 toktdøgn hvor 552 videolinjer (VL) ble inspisert og filmet og 57 lokaliteter (FS = fullstasjoner) ble prøvetatt med ulike redskaper (Tabell 1).
| Toktnr. | Fartøy | Toktleder | Start dato | Slutt dato | Ant VL | Ant FS |
| 2022118 | G.O. Sars | Genoveva Gonzalez-Mirelis | 14.10.22 | 29.10.22 | 78 | 8 |
| 2023001005 | G.O. Sars | Rebecca Ross | 30.03.23 | 12.04.23 | 113 | 9 |
| 2023001009 | G.O. Sars | Pål Buhl-Mortensen | 03.07.23 | 14.07.23 | 64 | 13 |
| 2023001014 | G.O. Sars | Kyrre Heldal Kartveit | 08.10.23 | 14.10.23 | 21 | 2 |
| 202400703 | Kronprins Haakon | Kyrre Heldal Kartveit | 08.03.24 | 19.03.24 | 84 | 6 |
| 2024001021 | G.O. Sars | Hilde Elise Heldal | 22.11.24 | 03.12.24 | 51 | 10 |
| 2025001009 | G.O. Sars | Genoveva Gonzalez-Mirelis | 20.06.25 | 03.07.25 | 141 | 9 |
| Sum | 552 | 57 |
En oversikt over hvilke stasjoner som ble prøvetatt, og hvilke redskaper som ble benyttet på hver enkelt stasjon, er gitt i Appendiks 1 . Denne rapporten beskriver ikke innholdet i prøvene, men viser til andre Mareano-publikasjoner for detaljert behandling av fullstasjonsdata (Gonzalez-Mirelis G 2022; 2026, Kartveit et al. 2023, Ross et al. 2023, Buhl-Mortensen et al. 2024, Kartveit et al. 2024). Analysene i denne rapporten baserer seg utelukkende på resultatene fra videoregistreringene.
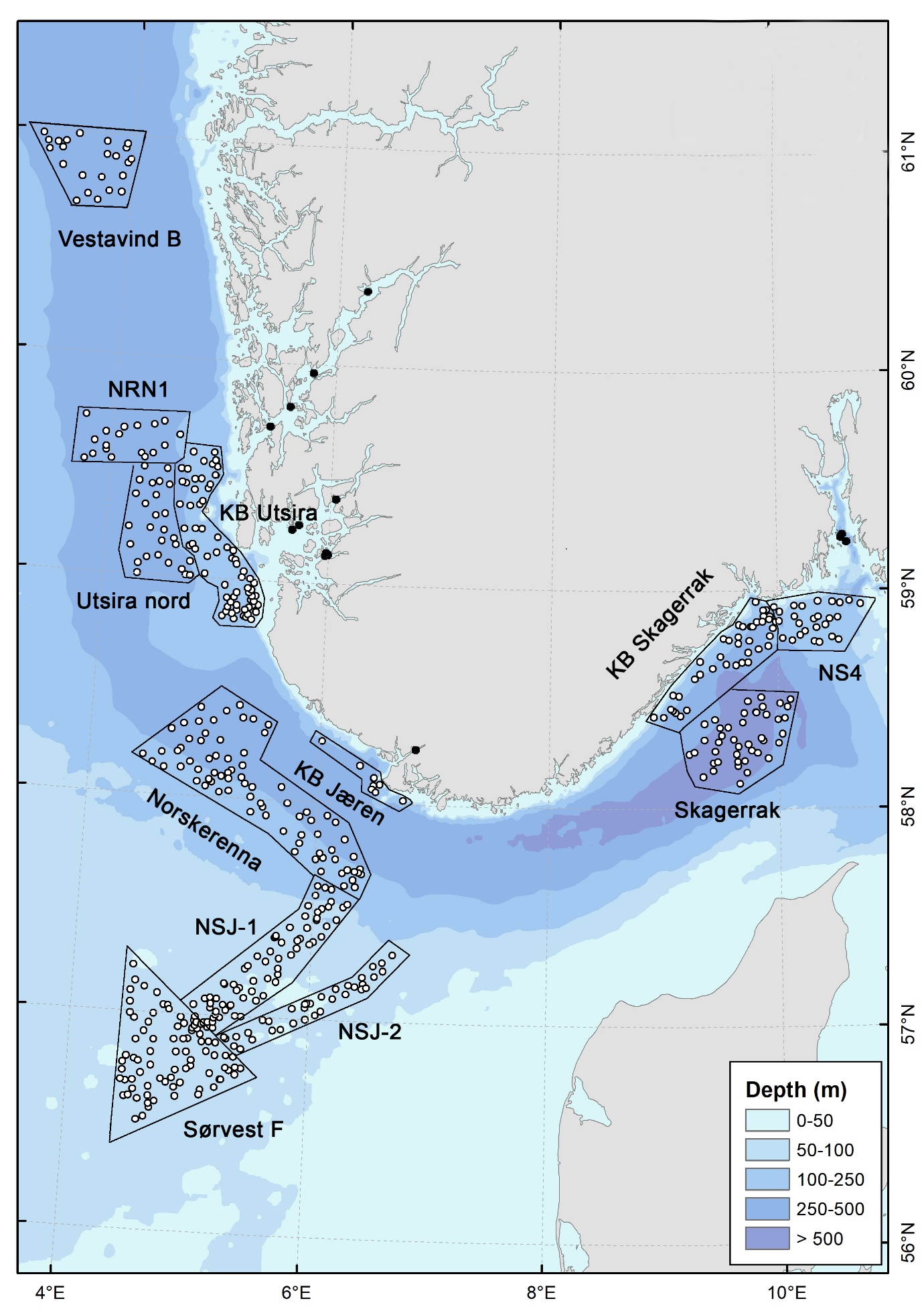 Figur 1. Kart over Nordsjøen og Skagerrak som viser inndelingen i geografiske områder brukt i rapporten og plasseringen av videostasjoner. Sorte punkt er stasjoner utenfor Mareanos kartleggingsområde, som ikke er tatt med i ordinasjonsanalyser.
Figur 1. Kart over Nordsjøen og Skagerrak som viser inndelingen i geografiske områder brukt i rapporten og plasseringen av videostasjoner. Sorte punkt er stasjoner utenfor Mareanos kartleggingsområde, som ikke er tatt med i ordinasjonsanalyser.
Havbunnen ble filmet ved hjelp av videoriggen Chimaera og ROVene Freya og Ægir 6000. Disse videoplattformene er utstyrt med høyoppløselige videokamera, lys og posisjoneringssystemer som muliggjør presis dokumentasjon av bunntyper og bunnfauna. Videoopptakene utgjør grunnlaget for identifikasjon av biotoper og registrering av sårbare habitater (VME-er).
2.3 - Annotering av havbunnsobservasjoner i felt
Hver videostasjon består av et ca. 200 m langt videotransekt, filmet med jevn fart og en høyde på omtrent 1,5 meter over havbunnen. Feltobservasjonene ble annotert (registrert med en rekke metadata koblet til hver observasjon) i sanntid ved hjelp av programvarene Campodlogger og Seabed FieldObserver (SFO). Disse verktøyene kobler annoteringer av arter, habitattyper og substrat direkte til metadata fra GPS, ekkolodd og undervannsposisjonering.
Bunntypene ble registrert i felt av geologer fra NGU, etter et klassifikasjonssystem med 19 bunntyper (Tabell 2). Den relative forekomsten av hver bunntype ble beregnet som prosentandel av observasjonene langs transektene.
| Norsk navn | Engelsk navn |
| Slam | Mud |
| Sandig slam | Sandy mud |
| Slamholdig sand | Muddy sand |
| Grus- og sandholdig slam | Gravelly sandy mud |
| Grus- og slamholdig sand | Gravelly muddy sand |
| Slam og sand med grus | Mud and sand with gravel |
| Slam- og sandholdig grus | Muddy sandy gravel |
| Slam og sand med grus, stein og blokk | Mud and sand with gravel, cobbles and boulders |
| Fin sand | Fine sand |
| Sand | Sand |
| Grusholdig sand | Gravelly sand |
| Sandholdig grus | Sandy gravel |
| Sand, grus og stein | Sand, gravel and cobbles |
| Sand, grus, stein og blokk | Sand, gravel, cobbles and boulders |
| Sand og blokk | Sand and boulders |
| Grus | Gravel |
| Grus og stein | Gravel and cobbles |
| Grus, stein og blokk | Gravel, cobbles and boulders |
| Bart fjell | Exposed bedrock |
Annoteringene i felt blir gjort på et mer overordnet nivå (forekomst av bunntyper og relativ mengde av hovedgrupper av arter og lett gjenkjennelige arter) enn det som er mulig etter tokt hvor videoopptakene kan settes på pause og man har bedre tid til å vurdere hver observasjon. Mer detaljerte analyser, inkludert artsidentifikasjon og kvantitative beregninger, tar lengre tid og blir rapportert senere.
Forekomst av sårbare habitater (VME) ble registrert basert på frekvens-kriterier, der tiden mellom observasjoner av indikatorarter brukes som mål på tetthet. Metoden er spesielt egnet for registrering i felt hvor direkte kvantitative tellinger ikke er praktisk mulig. Frekvensgrensene som benyttes i denne rapporten (Tabell 3) er basert på tidligere analyser av tettheter fra detaljerte videoannoteringer. Forekomsten av hver VME-type ble beregnet som andelen av transektets lengde der habitatet ble observert, etter samme prinsipp som for bunntyper.
| Habitatnavn | Indikator-arter (en eller flere av disse) | Hyppighet (tid mellom hver observasjon) |
| Svamspikelbunn | Geodia, Stelletta, Stryphnus, Aplysilla | < 5 sekunder mellom hver observasjon |
| Svampskog | Axinella, Phakellia, Clathria, Porifera branched | < 5 sekunder mellom hver observasjon |
| Kaldtvanns-svampsamfunn | Caulophacus, Cladorhiza, Hexactinellida | < 10 sekunder mellom hver observasjon |
| Hardbunns korallskog | Paragorgia, Primnoa, Paramuricea, Hornkoraller | < 10 sekunder mellom hver observasjon |
| Bløtbunns korallskog | Radicipes, Isidella | < 5 sekunder mellom hver observasjon |
| Blomkålkorallskog | Duva, Drifa, Gersemia, Nephtheidae | < 5 sekunder mellom hver observasjon |
| Sjøfjærsamfunn | Funiculina, Virgularia, Kophobelemnon, Halipteris, Pennatula, Pennatulacea | < 5 sekunder mellom hver observasjon |
| Sylindersjørosebunn | Cerianthus, Pachycerianthus, Ceriantharia | < 5 sekunder mellom hver observasjon |
| Umbellula-samfunn | Umbellula | < 30 sekunder mellom hver observasjon |
| Begerkorallsamfunn | Flabellum, Fungiacyathus, Scleractinia solitary | < 5 sekunder mellom hver observasjon |
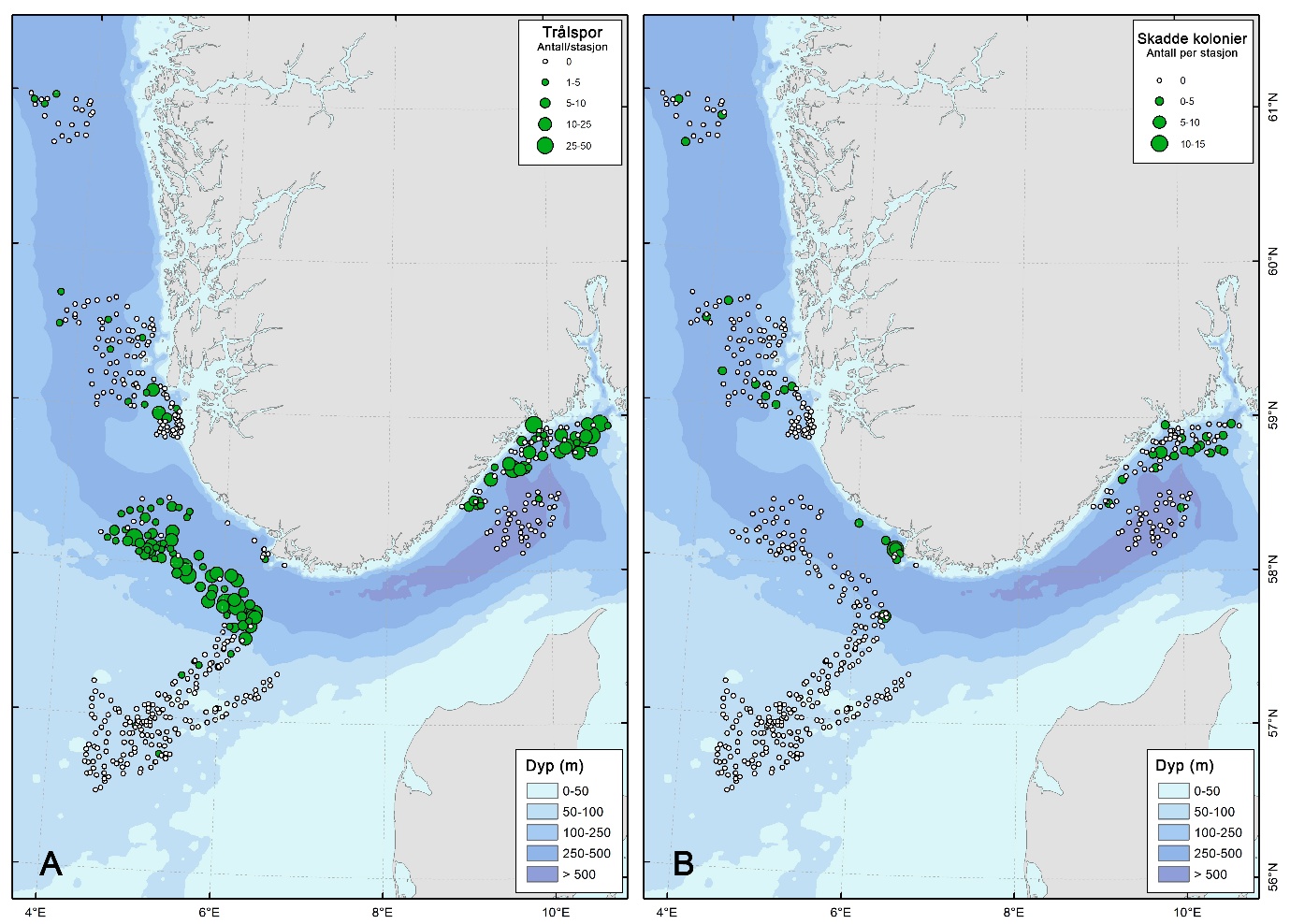
I tillegg til registrering av organismer og bunntyper, ble synlige tegn på menneskelig påvirkning dokumentert under feltannoteringen. Dette omfattet både fysisk skade på habitatdannende arter og direkte spor etter menneskelig aktivitet på havbunnen. Kolonier av sjøfjær (Pennatulacea) og hornkoraller (Gorgonacea) som fremstod som brukket, veltet eller tydelig mekanisk skadet, ble telt separat. Antall skadde kolonier ble registrert per transekt og benyttet som en indikator på mulig fysisk påvirkning, særlig fra bunnredskap.
Videre ble trålspor registrert og kvantifisert som antall observerte spor per videotransekt. Forekomst av marint søppel ble også registrert, inkludert type og antall objekter der dette var mulig å identifisere. Registreringene av trålspor, skadde kolonier og marint søppel inngår samlet som indikatorer på menneskelig påvirkning.
Metoder for feltannotering, etterarbeid, analyser og habitatmodellering er nærmere beskrevet i tidligere publikasjoner og rapporter (se Buhl-Mortensen et al. 2020, 2024a, 2024b)
2.4 - Databehandling og analyser
Etter feltarbeidet ble alle videoannotasjoner gjennomgått, kvalitetssikret og strukturert for videre analyser. Dataene ble delt inn i to matriser: en artsmatrise og en miljømatrise. Stasjoner utenfor Mareanos kartleggingsområder ble ekskludert. I artsmatrisen ble kun arter som forekom på mer enn fire stasjoner inkludert, og kun stasjoner med flere enn fire arter ble inkludert. Dette gav en endelig artsmatrise med 523 stasjoner og 222 arter/taxa. Miljømatrisen inneholdt 51 variabler: fire kategoriske variabler (toktnummer, geografisk område, dominerende bunntype, og VME-type). Miljømatrisen ble brukt som støttevariabler i DCA- og TWINSPAN-analyser, samt som grunnlag for å tolke gradientstrukturer og gruppedifferensiering.
Resultatene fra feltannoteringene ble analysert for å identifisere mønstre i arts- og habitatfordeling. To komplementære multivariate metoder ble benyttet:
-
DCA (Detrended Correspondence Analysis) (Hill & Gauch 1980), som gir en oversikt over gradienter i artsdata.
-
TWINSPAN (Two-Way Indicator Species Analysis) (Hill 1979), som benyttes til å klassifisere stasjoner i grupper basert på forekomst av indikatorarter og habitattyper.
TWINSPAN er en hierarkisk, divisjonsbasert metode som kombinerer ordinasjon og indikatorartsanalyse. Analysen starter med en ordinering som rangerer stasjonene langs viktigste artsgradient. Basert på denne deles datasettet i to grupper (første divisjon). Hver divisjon identifiserer indikatorarter som karakteriserer gruppene, og prosessen gjentas over flere nivåer. Resultatet er en hierarkisk gruppering av stasjoner som representerer ulike samfunnstyper og habitatmønstre.
Metoden er spesielt nyttig når målet er å skille ut økologiske samfunn, identifisere habitatgradienter og støtte biotopklassifisering. Den er godt egnet for store videobaserte datasett fordi den gir en robust og reproduserbar gruppering selv når artsdataene er ufullstendige eller kategoriske.
3 - Resultater
3.1 - Biomangfold
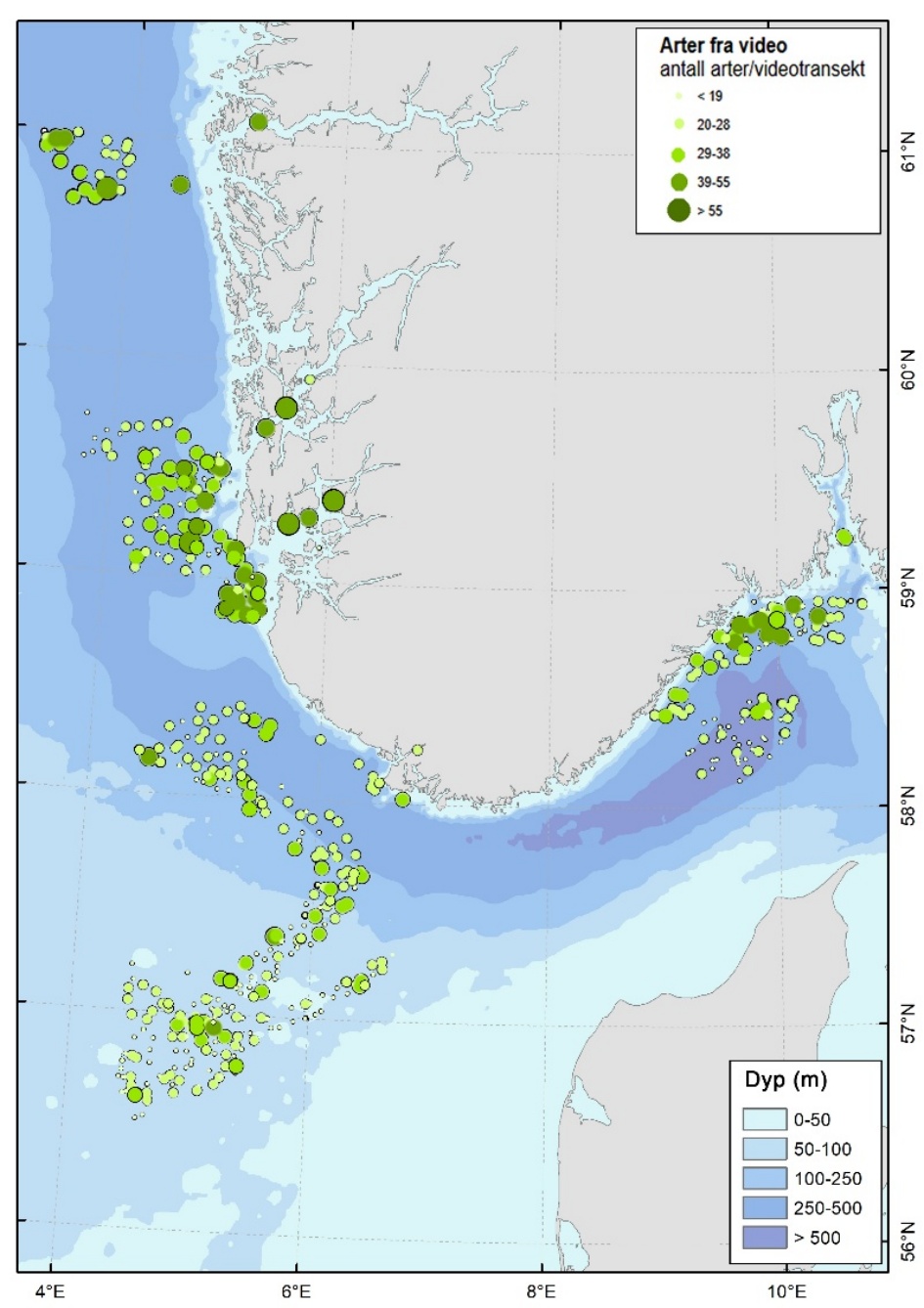
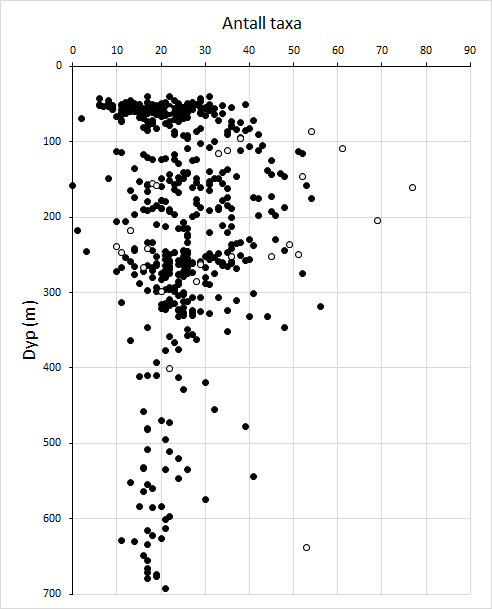
Totalt ble 472 taxa (arter og høyere taksonomiske nivåer) registrert i videomaterialet fra de syv toktene. Artsmangfoldet viste tydelig variasjon med både dyp og geografisk område. Antall observerte taxa var høyest mellom 100 og 250 m dyp i områder som representerer skråningen ned mot Norskerenna (Figur 2 & 3).
De høyeste artsantallene per stasjon ble registrert i fjordområdene Boknafjorden og Hardangerfjorden, som ligger utenfor Mareanos prioriterte kartleggingsområde, med opptil 77 taxa per stasjon på lokaliteter med korallrev og komplekse hardbunnshabitater.
Innenfor Mareanos kartleggingsområde ble det høyeste artsmangfoldet funnet i kystnære områder som Vestavind B, kystbeltet utenfor Utsira og i deler av Skagerrak, med opptil 56 taxa per stasjon.
I Nordsjøens grunne platåområder, dominert av sand og sandig slam, var artsmangfoldet lavere og mer homogent, med gjennomsnittlige verdier på 19–30 taxa per stasjon. Lokalt, i områder med stabile bunnhabitater som grus og stein på sandbunn, ble det ofte observert høye tettheter av svampskog og Alcyonium-samfunn med høyere artsmangfold. De dypere områdene i Skagerrak og Norskerenna har færre taxa i gjennomsnitt, men dekker viktige sårbare habitater på større dyp. Innenfor disse områdene ble det dokumentert omfattende forekomster av sjøfjærsamfunn, og lokale høye tettheter av begerkoraller, bambuskorallskog og andre sårbare habitater.
De mest dominerende hovedgruppene av bunndyr, observert på mer enn halvparten av stasjonene, var:
-
Asteroidea (sjøstjerner)
-
Pandalidae (reker)
-
Paguridae (eremittkreps)
-
Pleuronectiformes (flyndrer)
-
Munida sp. (trollhummer)
-
Ophiuroidea (slangestjerner)
3.2 - Biotopinndeling basert på ordinasjons- og klassifikasjonsanalyser
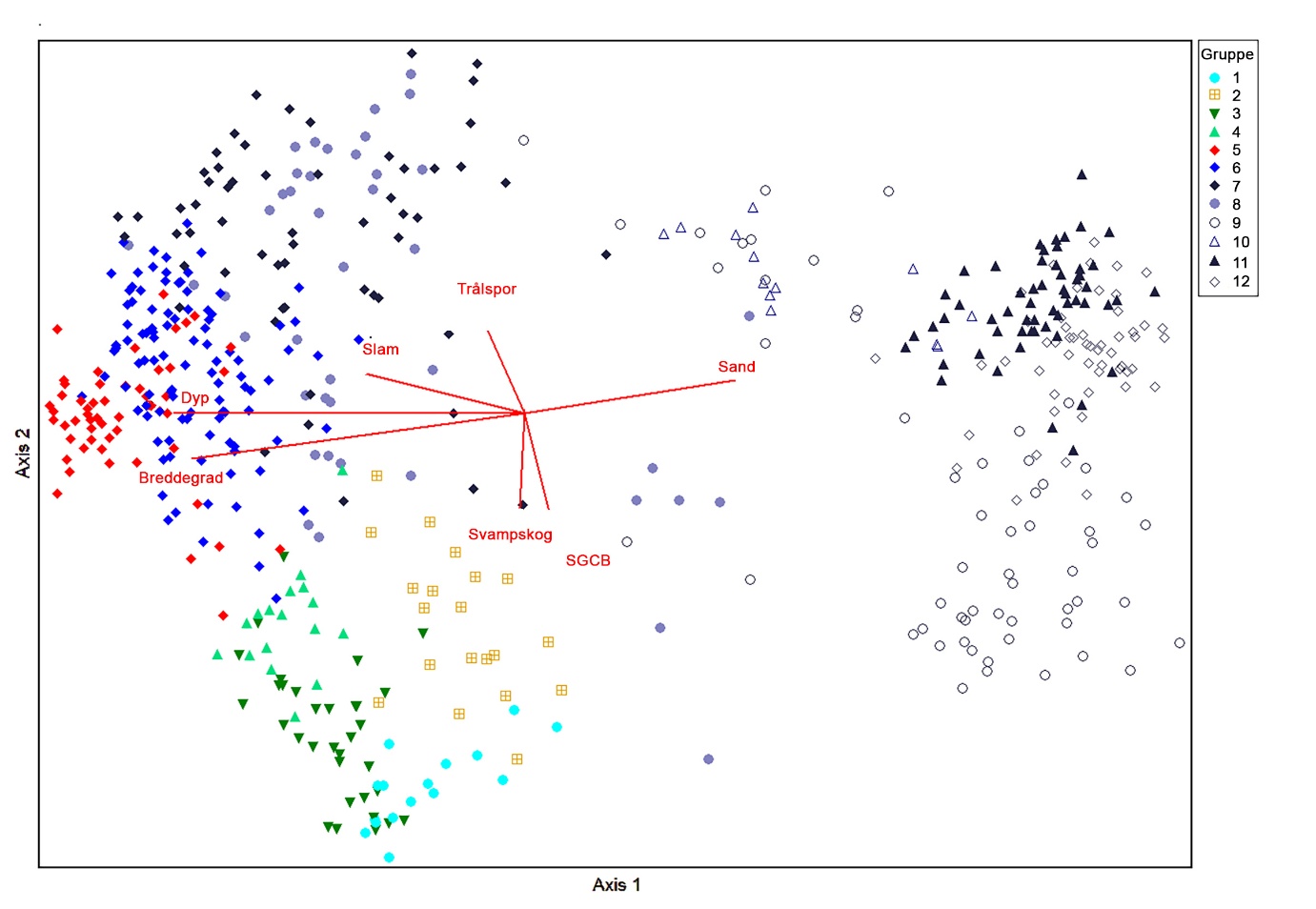
Ordinasjonsanalysene (DCA) og den hierarkiske klassifikasjonen (TWINSPAN) av 523 videostasjoner og 222 taxa (etter filtrering) identifiserte tydelige mønstre i sammensetningen av bunndyrsamfunn. TWINSPAN-analysen delte datasettet i tolv økologiske grupper (4 grupper ved tredje delingsnivå og 8 ved fjerde delingsnivå). Disse omtales videre som biotoper og presenteres i DCA-plot (Figur 4), kart (Figur 5), og som en sammenfattende tabell med kjennetegn (Tabell 4). Faunamønstrene i ordinasjonsplottet var sterkest korrelert med dyp, breddegrad og relativ mengde sand og slam (r² ≤ –0,54) langs førsteaksen (Figur 4). Dette indikerer at hovedgradienten i datasettet i stor grad reflekterer en kombinert dybde- og geografisk gradient, der de sørlige, grunnere og mer sanddominerte områdene skilles fra de dypere og mer finmaterialedominerte områdene lenger nord. Andreaksen var sterkest korrelert med antall trålspor, relativ mengde sandig grus med stein og blokk (SGCB), samt andel av videotransektene med svampskog-habitat (r² ≥ 0,38). Denne aksen kan tolkes som en gradient knyttet til substrattype og fysisk påvirkning. Hardbunnsområder (høy andel SGCB) og forekomst av svampskog var negativt korrelert med trålspor, noe som indikerer at tråling i hovedsak foregår i bløtbunnsområder og i mindre grad i områder med hardbunn og utviklede svampsamfunn. Samlet viser korrelasjonene at førsteaksen primært representerer en storskala miljøgradient (dyp–geografi–sedimenttype), mens andreaksen i større grad reflekterer variasjon i substrat og menneskelig påvirkning.
3.3 - Beskrivelse av biotopgruppene (TWINSPAN-grupper)
De mest artsrike gruppene (gr. 1–4) forekom hovedsakelig i kystnære og mellomdype områder rundt Utsira, Vestavind og Skagerrak, med gjennomsnittlige artsantall på 32–42 taxa. Disse gruppene er preget av sammensatte bunnsubstrater bestående av sand, grus, stein og blokk, samt stedvis innslag av slam. Flere av disse gruppene (gruppe 1 – 3) karakteriseres av svampdominerte samfunn (svampskog) , med typiske arter som Antho dichotoma, Axinella infundibuliformis, Mycale lingua og Phakellia spp. Gruppe 3 skiller seg ut ved å kombinere svamphage med bløtbunnsfauna med innslag av sjøfjær (spesielt Funiculina quadrangularis) og domineres av Bonnelida (skjeormer) og sjøpølsen Mesothuria intestinalis. Gruppe 4, med stasjoner hovedsakelig fra Vestavind F, Utsira nord, og KB-Utsira karakteriseres av bunn med sandig slam dominert av svampene Oceanapia robusta, Thenea sp., og Craniella zetlandica typiske for blandete slike sedimenter, samt sekkedyret Styela sp., og sjøpølsen Mesothuria intestinalis.
Grupper fra større dyp (grupper 5–7), hovedsakelig fra Skagerrak og Norskerenna, på dyp fra ca. 200 til over 500 m, er knyttet til slam- og sandig slam-dominerte sedimenter og kjennetegnes av forekomster av sjøfjær, begerkoraller, bambuskorall (Isidella lofotensis) og svampen Thenea. Typiske indikatorarter inkluderer Funiculina quadrangularis, Asteronyx loveni, Kophobelemnon stelliferum, Atlantopandalus propinqvus og uidentifiserte Pennatulacea (sjøfjær). Gruppe 6 viser særlig høy dominans av sjøfjærarter og representerer et tydelig sjøfjærsamfunn i Norskerenna. Gruppe 8 representerer blandete slamdominerte sedimenter i NS4-området (ytre Oslofjord), med arter som Nephrops norvegicus (sjøkreps) og Echinoidea irregularia (irregulære kråkeboller). Blant stasjonene representert i gruppe 5, 6 og 8 ble det observert områder med høye tettheter av sjøfjær (se kapittel 3.4).
De grunneste samfunnene (gr. 9–12), hovedsakelig fra NSJ-1, NSJ-2 og Sørvest-F, forekommer på dybder mellom 40 og 150 meter og er preget av sand- og grusdominerte substrater . Flere av disse gruppene (gr. 9 og 11) er assosiert med Alcyonium-hager , der dødmannshånd (Alcyonium digitatum ) og annen fastsittende fauna (bl.a. begerkorallen Caryophyllia smithii og skorpedannende rødalger) dominerer. De grunneste gruppene har generelt lavere artsmangfold (15–24 taxa) og karakteriseres også av bevegelige predatorer som sjøstjerner (Asterias rubens, Luidia sarsii), flerbørstemark (Oxydromus flexuosus) og av tobis (Ammodytidae).
| Gruppe | Område | Dyp (intervall) | Antall taxa (snitt) | VME/Habitat | Substrat | Karakteristiske arter/taxa | |
| 1 | KB-Utsira | 125 | (63-193) | 42 | Svampskog | Sand, grus, stein og blokk | Antho dichotoma, Axinella infundibuliformis, Mycale lingua, Phakellia sp. |
| 2 | KB-Skagerrak, NS4 | 140 | (39-347) | 37 | Svampskog | Sand, grus, stein og blokk | Echinoidea regular, Phakellia ventilabrum, Porifera forgrenet, Stichastrella rosea |
| 3 | KB-Utsira | 222 | (137-275) | 35 | Svampskog og svampspikelbunn | Slam og sand m. grus, stein og blokk | Geodia sp., Hymedesmia paupertas, Bonellida , Mesothuria intestinalis |
| 4 | Vestavind F, Utsira nord, KB-Utsira | 245 | (117-318) | 32 | Thenea | Sandig slam | Oceanapia robusta, Styela sp., Mesothuria intestinalis, Thenea sp., Craniella zetlandica |
| 5 | Skagerrak | 528 | (215-693) | 21 | Sjøfjær | Slam | Craniella sp., Atlantopandalus propinqvus, Munida sarsi, Virgulariidae red, Kophobelemnon stelliferum, Cerianthus lloydii |
| 6 | Norskerenna | 285 | (164-410) | 24 | Sjøfjær, Flabellum, og Isidella | Sandig slam | Funiculina quadrangularis, Kophobelemnon stelliferum, Asteronyx loveni, Pennatula phosphorea, Flabellum macandrewi, Isidella lofotensis |
| 7 | Norskerenna | 204 | (73-295) | 22 | Sandig slam | Pandalidae , Spirontocaris sp., Gadiculus argenteus, Myxine glutinosa, Nephrops norvegicus, | |
| 8 | NS4 | 170 | (71-412) | 22 | Sjøfjær | Blandet finkornet | Balticina sp., Nephrops norvegicus, Echinoidea irregular, Trisopterus sp. |
| 9 | NSJ-1, NSJ-2, Sørvest-F | 63 | (40-153) | 24 | Alcyonium | Sand, grus, stein og blokk | Caryophyllia smithii, Alcyonium digitatum, Rhodophyta encrusting calcareous, Modiolus modiolus |
| 10 | NSJ-1 | 75 | (53-125) | 20 | Sand | Cottidae , Hormathiidae, Ammodytidae, Spatangus sp., Melanogrammus aeglefinus | |
| 11 | NSJ-2, Sørvest-F | 57 | (45-73) | 15 | Sand | Aphroditidae, Pleuronectiformes, Asteriidae , Luidia sarsii | |
| 12 | Sørvest-F | 57 | (48-63) | 19 | Sand | Astropecten irregularis, Asterias rubens, Oxydromus flexuosus, Limanda limanda | |
3.4 - Forekomster av sårbare habitater (VME)
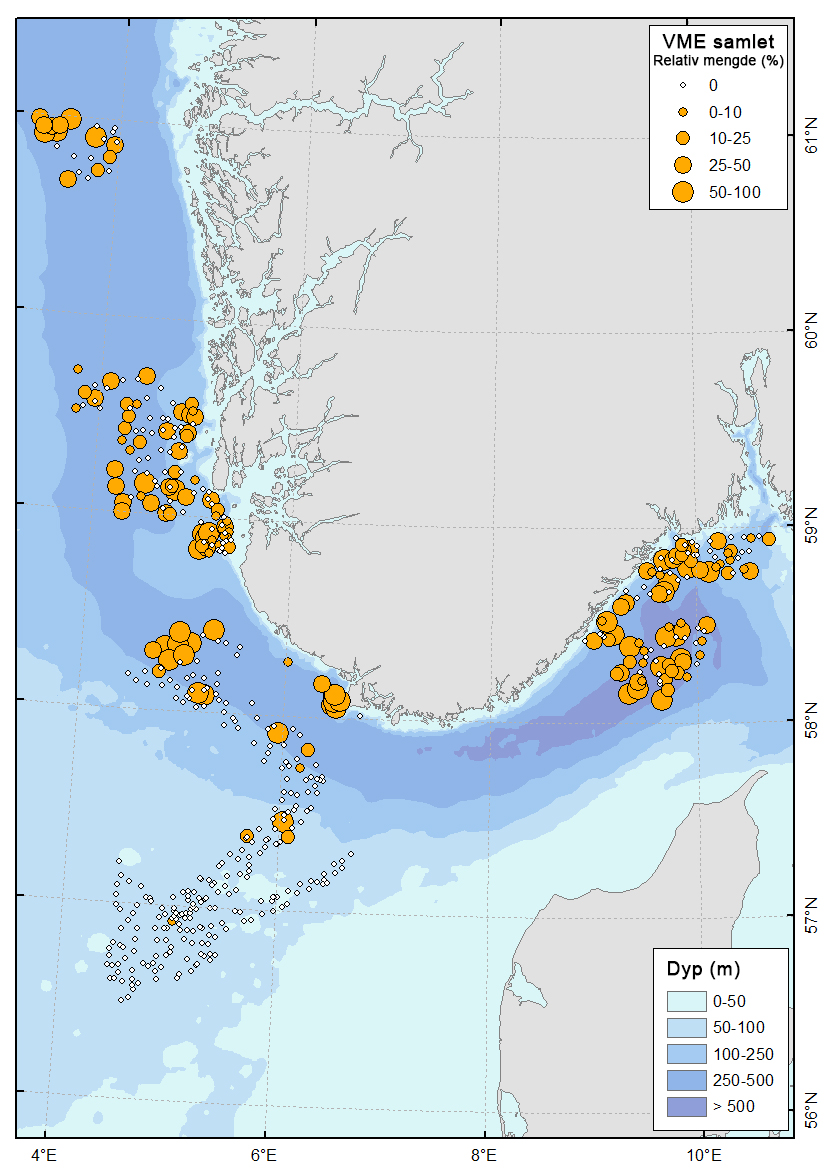
En oversikt over observasjoner av indikator-taksa for sårbare habitater er gitt i tabell 5. Totalt ble slike habitater (jfr tabell 3) registrert på 30 % av stasjonene (158 av 523). Forekomsten viser tydelige regionale og dybderelaterte forskjeller.
Sårbare habitater forekom hovedsakelig på stasjoner dypere enn 350 m (Figur 6 & 7), mens de grunne områdene på Nordsjøplatået i mindre grad oppfylte kriteriene. Andelen stasjoner med VME var høyest i Kystbelte Skagerrak (89 %), Kystbelte Jæren (89 %), Utsira Nord (78 %) og i Skagerrak (70 %). Lavest andel ble registrert i NSJ-områdene og Sørvest-F, hvor under 30 % av stasjonene oppfylte kriteriene. Utenfor Mareanos kartleggingsområde hadde Boknafjorden og Hardangerfjorden en relativt høy andel av lokaliteter med sårbare habitater (ca. 50–60 %).
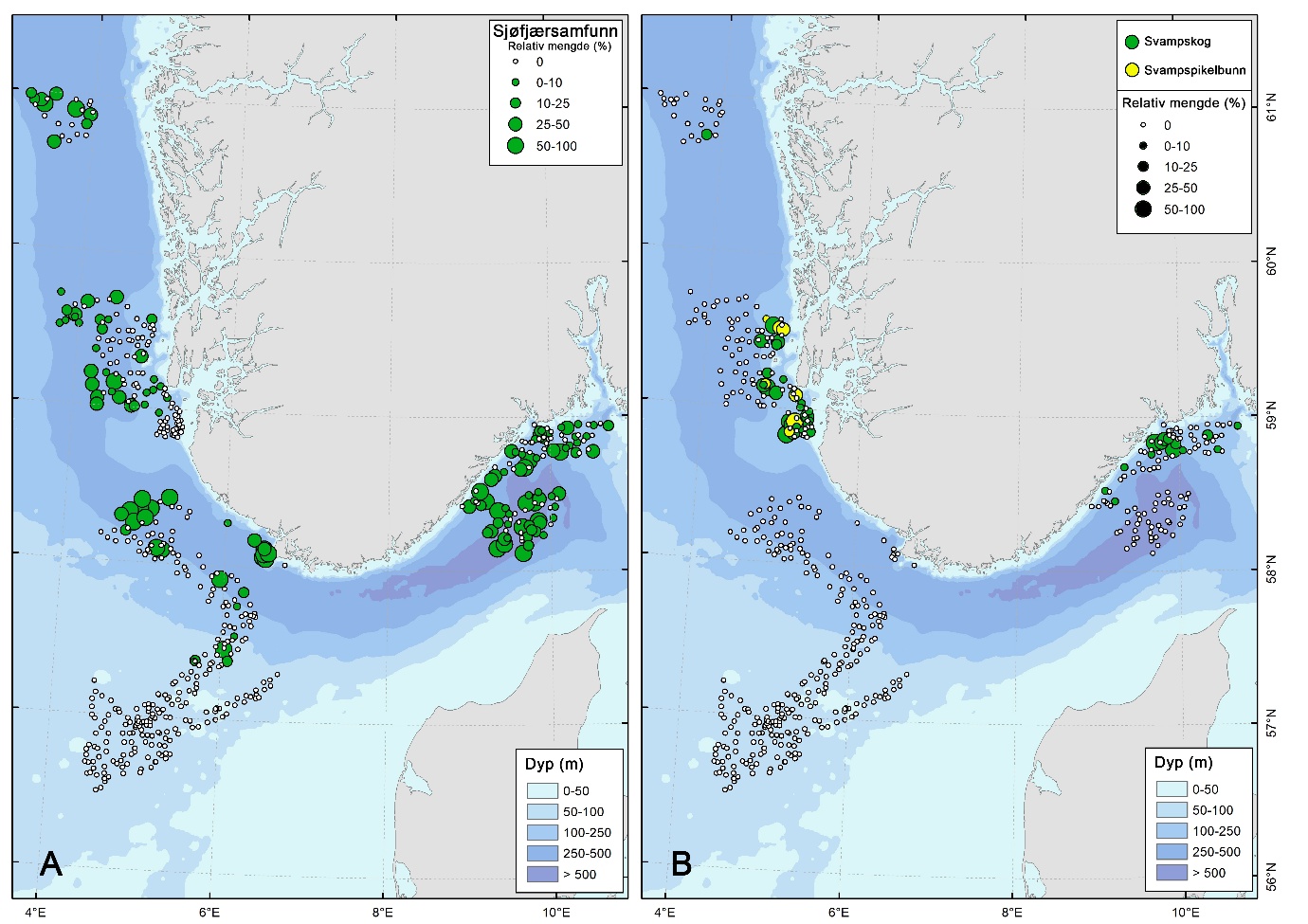
Sjøfjærsamfunn var den klart dominerende VME-typen og preget særlig de dype sedimentområdene (Figur 6 & 9). Gjennomsnittlig forekomst av sjøfjær var høyest i Norskerenna (87 observasjoner per stasjon), Skagerrak (47), Vestavind B (42) og Kystbelte Jæren (36) (Tabell 5). Forekomstene besto hovedsakelig av Funiculina quadrangularis og Kophobelemnon stelliferum . I kontrast var sjøfjær sjeldne i de grunne delene av Nordsjøplatået og i Sørvest-F.
Svampsamfunn viste et annet romlig mønster og omfattet både svampskog i hardbunnsområder og svampspikelbunn i mer slamrike miljøer i mellomdyp sone (særlig i Kystbelte Utsira). Svampskog var særlig konsentrert i hardbunnsområdene i Kystbelte Utsira (27 observasjoner per stasjon), Kystbelte Skagerrak (11) og NS4 (7). Forekomsten av svamp var lav i de dype renneområdene.
Øvrige VME-indikatorarter forekom mer lokalt og i lavere tettheter. Bløtbunnskorallskoger var representert ved Isidella lofotensis (særlig i Kystbelte Jæren, 11 per stasjon) og begerkorallsamfunn med Flabellum macandrewi (særlig i Vestavind B, 12 per stasjon). Sylindersjøroser hadde høyest forekomst i Kystbelte Skagerrak (6) og O-skjellbanker ( Modiolus modiolus ) ble kun observert i grunne NSJ-områder og ble registrert på to lokaliteter.
Samlet viser resultatene en tydelig dybde- og substratavhengig fordeling av VME-indikatorarter: sjøfjær dominerer i dype sedimentmiljøer, svampsamfunn i hardbunnsområder, mens øvrige habitattyper forekommer mer fragmentert og regionavhengig.
| Vestavind B | KB Utsira | Vestavind F | Utsira nord | Norskerenna | KB Jæren | NSJ-1 | NSJ-2 | Sørvest-F | Skagerrak | KB Skagerrak | NS4 | Utenfor Mareano | |
| Antall st | 23 | 78 | 17 | 18 | 79 | 9 | 68 | 37 | 84 | 43 | 32 | 35 | 28 |
| Tot ant arter | 120 | 250 | 98 | 117 | 144 | 102 | 179 | 109 | 149 | 124 | 146 | 140 | 207 |
| Snitt arter per st | 30 | 32 | 23 | 27 | 23 | 26 | 22 | 18 | 20 | 20 | 27 | 25 | 30 |
| Snitt dyp | 322 | 189 | 267 | 258 | 243 | 290 | 63 | 57 | 58 | 570 | 254 | 169 | 209 |
| Min dyp | 266 | 37 | 227 | 236 | 123 | 71 | 40 | 36 | 42 | 386 | 80 | 38 | 34 |
| Maks dyp | 367 | 276 | 277 | 272 | 333 | 361 | 144 | 81 | 65 | 695 | 462 | 355 | 847 |
| Bløtkoraller | |||||||||||||
| Alcyonium digitatum | 0.01 | 0.11 | 14.13 | 11.81 | 9.63 | ||||||||
| Anthomastus sp. | 0.57 | ||||||||||||
| Sum bløtkoraller | 0.06 | 0.11 | 14.13 | 11.81 | 9.63 | 0.64 | |||||||
| Hornkoraller | |||||||||||||
| Anthothela grandiflora | 0.01 | 0.14 | |||||||||||
| Clavularia borealis | 0.01 | 0.18 | |||||||||||
| Paragorgia arborea | 0.06 | 0.36 | |||||||||||
| Paramuricea placomus | 0.01 | 1.25 | |||||||||||
| Primnoa resedaeformis | 0.09 | 0.02 | 0.03 | ||||||||||
| Isidella lofotensis | 0.65 | 0.09 | 11.44 | 0.04 | |||||||||
| Sum hornkoraller | 0.65 | 0.27 | 11.44 | 0.01 | 0.02 | 0.03 | 1.96 | ||||||
| Steinkoraller | |||||||||||||
| Flabellum macandrewi | 11.83 | 0.04 | 0.03 | 0.13 | |||||||||
| Lophelia pertusa | 1.43 | ||||||||||||
| Sum steinkoraller | 12.83 | 0.04 | 0.24 | 0.14 | 1.43 | ||||||||
| Sylindersjøroser | 0.74 | 0.13 | 0.12 | 0.11 | 0.77 | 2.00 | 0.63 | 0.46 | 0.01 | 1.72 | 6.13 | 1.29 | 8.46 |
| Svampskog | |||||||||||||
| Antho dichotoma | 0.09 | 7.18 | 0.61 | 0.04 | 0.11 | 0.28 | 0.04 | 0.31 | 0.17 | 4.93 | |||
| Axinella infundibuliformis | 2.44 | 0.01 | 0.33 | 0.16 | 0.33 | 0.97 | 0.89 | 0.71 | |||||
| Axinellidae | 1.19 | 0.17 | 0.01 | 0.11 | 0.01 | 0.01 | 0.05 | 2.88 | 1.20 | 2.25 | |||
| Phakellia sp. | 0.43 | 4.58 | 0.28 | 0.22 | 0.15 | 0.28 | 2.84 | 1.11 | 0.54 | ||||
| Phakellia ventilabrum | 3.37 | 0.28 | 0.22 | 0.07 | 0.09 | 1.00 | 0.37 | 4.75 | |||||
| Porifera branched dense | 0.86 | 0.22 | 2.38 | 1.91 | 0.04 | ||||||||
| Mycale lingua | 0.87 | 7.18 | 0.24 | 0.72 | 0.01 | 0.01 | 0.01 | 0.88 | 1.71 | 2.61 | |||
| Sum svampskog | 1.39 | 26.79 | 0.24 | 2.06 | 0.08 | 1.22 | 0.69 | 0.00 | 0.06 | 0.74 | 11.25 | 7.37 | 15.82 |
| Svampspikelbunn | |||||||||||||
| Geodia atlantica | 0.65 | 0.11 | 0.02 | 1.61 | |||||||||
| Geodia barretti | 0.04 | 1.10 | 0.67 | 1.05 | 0.34 | 0.93 | |||||||
| Geodia macandrewii | 0.29 | 0.06 | 0.44 | 0.54 | |||||||||
| Geodia phlegraei | 0.10 | 0.06 | 0.05 | 0.36 | |||||||||
| Geodia sp. | 0.56 | 0.41 | 0.11 | 0.21 | 0.22 | 0.03 | 0.89 | ||||||
| Hexadella dedritifera | 0.04 | 2.00 | 0.11 | 0.25 | 0.14 | 0.06 | 0.40 | 1.39 | |||||
| Stryphnus ponderosus | 0.55 | 0.05 | 0.43 | ||||||||||
| Sum svampspikelbunn | 0.09 | 5.27 | 0.47 | 1.50 | 0.25 | 1.51 | 0.63 | 0.43 | 6.14 | ||||
| Sjøfjær | |||||||||||||
| Balticina finmarchica | 0.56 | ||||||||||||
| Balticina sp. | 0.78 | 0.05 | 0.33 | 5.94 | |||||||||
| Funiculina quadrangularis | 17.57 | 6.47 | 4.29 | 9.17 | 22.66 | 6.22 | 0.01 | 2.26 | 15.75 | 5.00 | 5.29 | ||
| Kophobelemnon stelliferum | 16.04 | 4.63 | 8.35 | 12.28 | 18.97 | 17.00 | 23.93 | 7.31 | 5.71 | 1.79 | |||
| Pennatula phosphorea | 0.04 | 0.13 | 1.41 | 4.61 | 17.22 | 0.89 | 0.07 | 0.75 | 1.63 | 0.04 | |||
| Pennatulacea | 7.43 | 1.55 | 0.59 | 0.39 | 1.94 | 0.22 | 0.09 | 0.01 | 5.07 | 1.19 | 2.63 | 1.68 | |
| Virgularia sp. | 0.54 | 3.71 | 0.56 | 2.80 | 2.78 | 0.18 | 0.51 | 5.91 | 0.59 | 1.69 | 1.29 | ||
| Virgulariidae | 0.04 | 0.90 | 1.61 | 13.13 | 5.44 | 3.28 | 0.76 | 9.58 | 3.25 | 0.26 | 3.25 | ||
| Sum sjøfjær | 41.96 | 15.04 | 18.35 | 30.00 | 86.99 | 36.33 | 6.91 | 2.00 | 0.01 | 46.91 | 29.00 | 22.86 | 15.75 |
| Modiolus modiolus | 6.10 | 0.24 | 1.15 |
Sjøfjærsamfunn
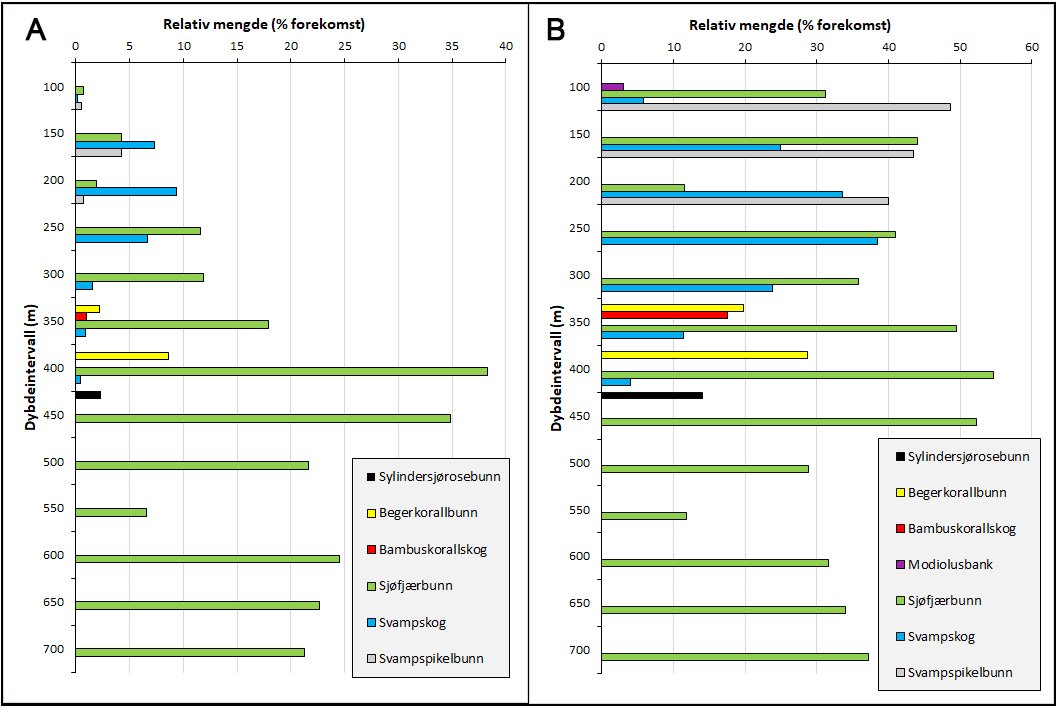
Sjøfjærsamfunn var det mest utbredte sårbare habitatet og ble registrert på 21 % av stasjonene (112 av 523). Habitatet forekom primært dypere enn 400 m (Figur 6 & 9). De største forekomstene ble registrert i dybdeintervallet 400–500 m, hvor habitatet i gjennomsnitt dekket 35–38 % av videolinjene. Beregnet kun for stasjoner hvor habitatet forekom, var gjennomsnittlig dekning 52–54 % av videolinjene i dette dybdeintervallet.
Indikatorartene Funiculina quadrangularis (stor piperenser) og Kophobelemnon stelliferum (hanefot) ble registrert på henholdsvis 237 og 327 stasjoner. De fleste observasjonene var imidlertid enkeltindivider eller lave tettheter som ikke tilfredsstilte kriteriene for sjøfjærsamfunn (jf. Tabell 3). Pennatula phosphorea (Figur 8) ble observert på 56 stasjoner, med høy tetthet på seks lokaliteter mellom 250 og 320 m dyp.

Svampskog
Svampskog ble påvist i flere områder, særlig i kystnære lokaliteter i dybdeintervallet 150–250 m, knyttet til hardbunn dominert av grus, stein og blokk. I dette intervallet dekket habitatet i gjennomsnitt 7–9 % av videolinjene. Beregnet kun for stasjoner hvor habitatet forekom, var gjennomsnittlig dekning 34-39 % av videolinjene i dybdeintervallet 200-250 m (Figur 7 & 9).
Svampspikelbunn
Svampspikelbunn ble registrert på kun syv stasjoner (3 % av stasjonene) i Kystbelte Utsira, i dybdeintervallet 138–236 m. Habitatet var knyttet til områder med større innslag av slam, og forekom delvis overlappende med lokaliteter der svampskog ble registrert. På stasjoner hvor habitatet ble identifisert dekket det 40-49 % av videolinjene (Figur 6).
Indikatorartene omfattet de store svampene Geodia atlantica, G. barretti, G. macandrewii, G. phlegraei, Hexadella detrifera, Stryphnus fortis, S. ponderosus og Stelletta sp. Det ble også registrert innslag av indikatorarter for svampskog på disse lokalitetene, noe som indikerer en gradient eller mosaikk mellom disse to svampdominerte habitattypene.
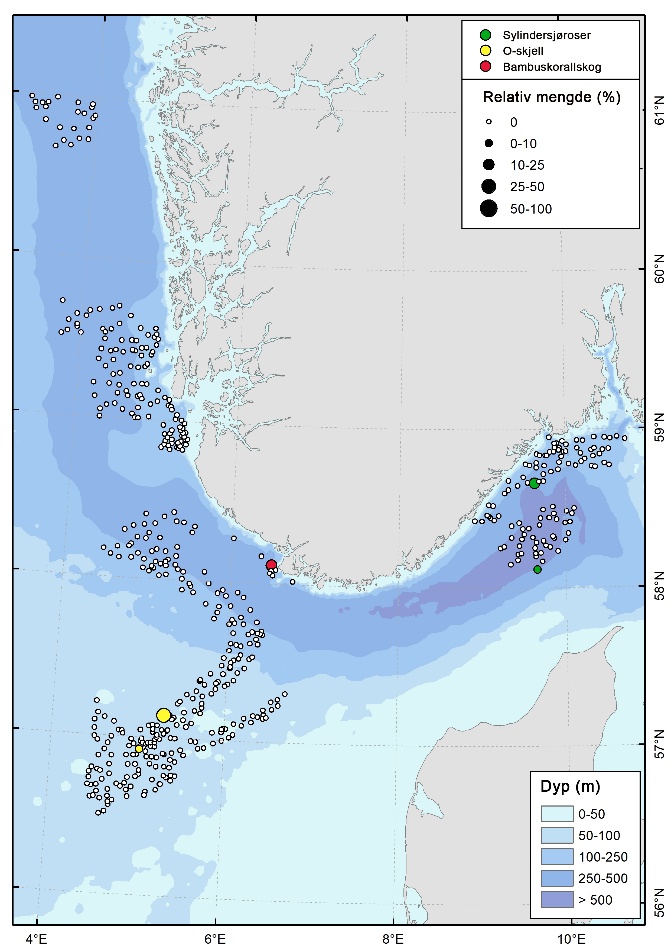
Bløtbunnskorallskog
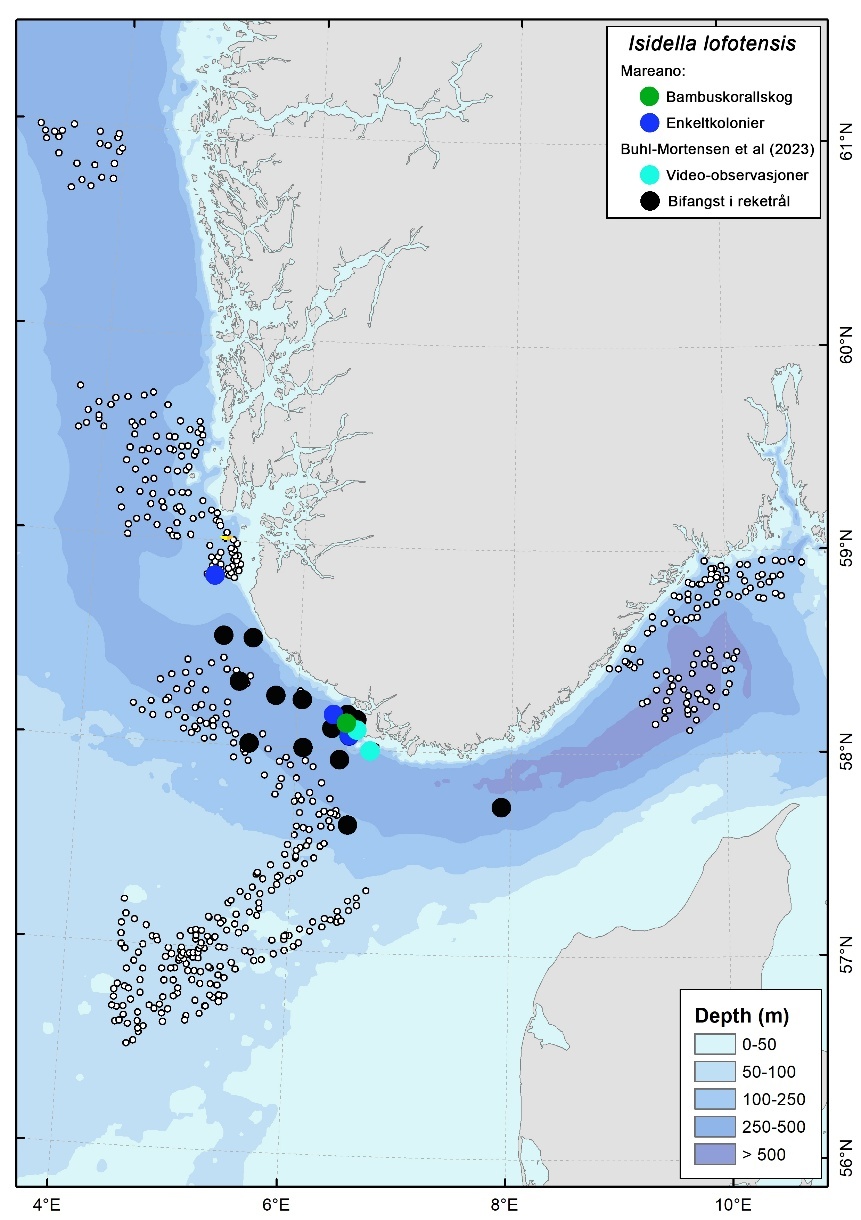
Bambuskorallskog dominert av Isidella lofotensis ble registrert på to lokaliteter på 300-350 m dyp, i kystbeltet Jæren (Figur 6, 10 & 12). Her dekket habitatet rundt 18 % av videolinjene. Begerkorallsamfunn dominert av Flabellum macandrewi ble dokumentert på syv lokaliteter i Vestavind B (Figur 11), i dybdeintervallet 350–450 m, med en dekning på 20-29 % av videolinjene.

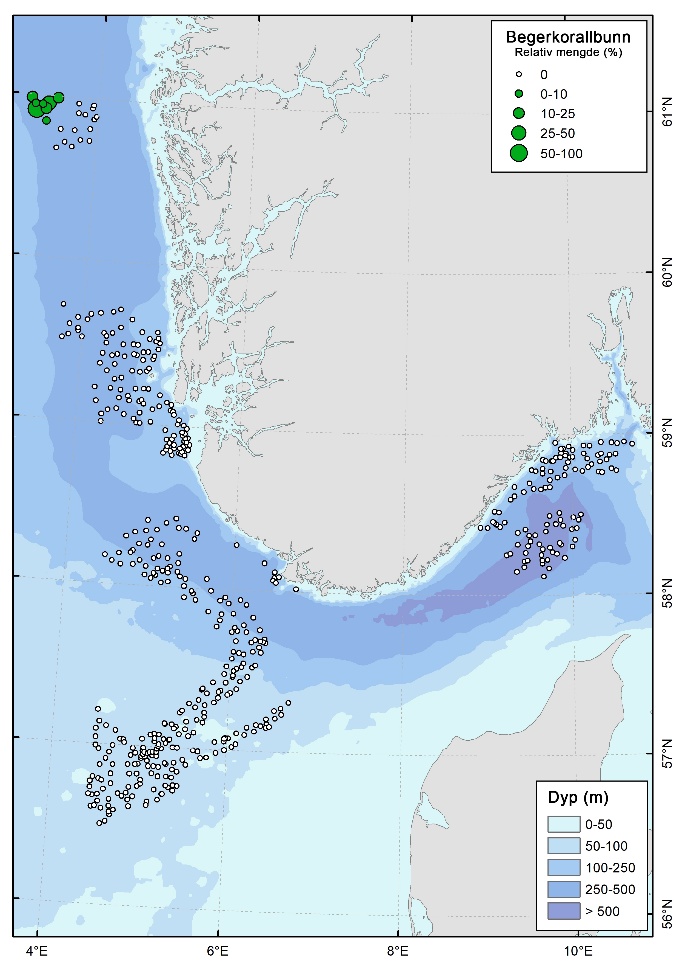
Sylindersjørosebunn
Sylindersjørosebunn ble registrert med høye tettheter på to lokaliteter i ytterkanten av den flate, dype delen (dybdeintervallet 450 – 500 m) av Skagerrak i nordlige deler av kystbeltet ved Risør og i sørlige deler av Skagerrak (Figur 6 & 9). Her forekom sylindersjøroser med høye tettheter, med en dekning på rundt 14 % av videolinjene.
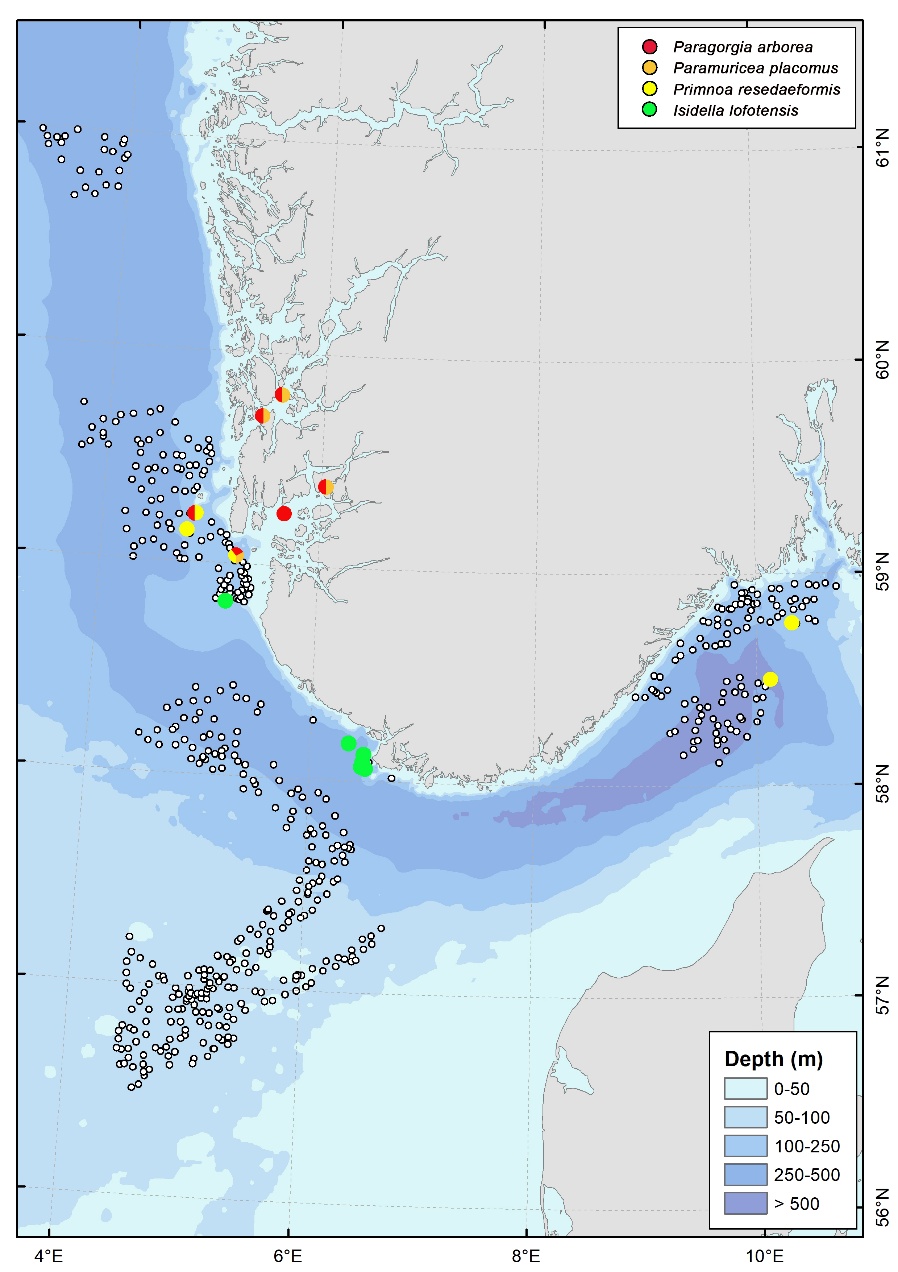
Enkeltforekomster av hornkoraller
Utover bambuskoraller ble det ikke registrert tettheter av hornkoraller som tilfredsstilte kriteriene for korallskog. Enkeltkolonier ble observert i kystbelte Utsira (Paragorgia arborea, Paramuricea placomus og Primnoa resedaeformis) og på to lokaliteter i Skagerrak (P. resedaeformis). Utenfor Mareanos kartleggingsområder ble P. arborea og P. placomus observert i Hardangerfjorden og Boknafjorden (Figur 13).
Andre strukturdannende arter
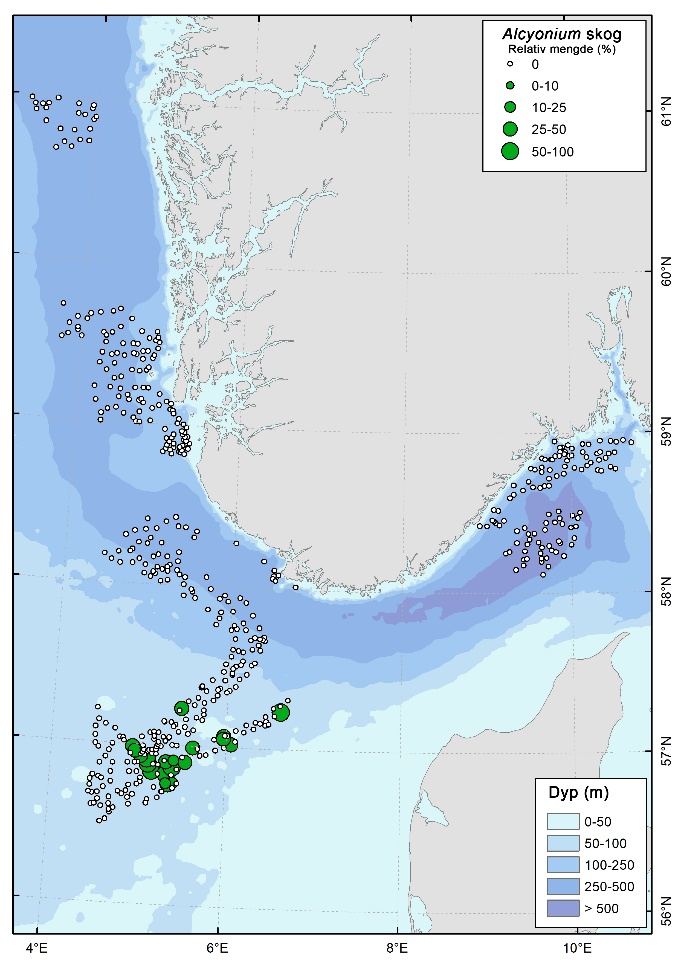
Høye tettheter av bløtkorallen dødmannshånd (Alcyonium digitatum) ble registrert i flere områder med grus og stein i Sørvest F og i sørlige, grunne deler av NSJ-områdene (Figur 14). Disse forekomstene er imidlertid ikke klassifisert som sårbare habitater av OSPAR eller FAO.

3.5 - Menneskelig påvirkning
3.5.1 - Trålspor og skadde kolonier
Trålspor ble observert i flere deler av undersøkelsesområdet, men forekomsten var klart høyest i skråningen opp fra Norskerenna til Nordsjøplatået, nordlig del av kystbelte Skagerrak og i Ytre Oslofjord (NS4) (Figur 17). I disse områdene var det gjennomsnittlige antallet trålspor per videostasjon henholdsvis 10 og 6, med et maksimum på 38 og 50 spor per stasjon. I nordlig del av kystbelte Skagerrak og Ytre Oslofjord var det et overlapp mellom trålspor og skadde kolonier av sjøfjær, som indikerer fysisk påvirkning av bunntråling. Trålspor var sjeldne eller fraværende i de fleste fjordområdene og på de grunne, sanddominerte områdene på Nordsjøplatået.
Samtidig viste videomaterialet betydelige interne forskjeller innen Norskerenna. Flere av de dypeste lokalitetene fremsto som lite eller ikke påvirket av bunntråling og var preget av tette bestander av sjøfjær. Dette samsvarer med at de dypere deler av Norskerenna inngår i et dumpingsområde for militære stridsmidler som er stengt for tråling, og det ser ut til at fiskeriaktiviteten i stor grad også unngår nærliggende områder. Videre er de dypeste delene av Norskerenna generelt mindre attraktive for bunntråling, og den mest intensive reketrålingen foregår i grunnere deler av skråningen opp fra renna.

3.5.2 - Søppel på havbunnen
Forekomsten av marint søppel var generelt lav i undersøkelsesområdet, men søppel ble registrert i et flertall av de geografiske delområdene. Totalt ble marint søppel observert på 23 % av videostasjonene. De høyeste gjennomsnittlige forekomstene ble registrert i Skagerrak (1,16 objekter per stasjon) og i Norskerenna (0,57 objekter per stasjon), med enkelte stasjoner der opptil 18 søppelobjekter ble registrert. I de øvrige områdene var den gjennomsnittlige forekomsten lav, med verdier på 0,3–0,4 objekter per stasjon.
Det var en tydelig forskjell i både forekomst og type søppel mellom de dype stasjonene i Norskerenna og de grunnere stasjonene i kystbeltet. I kystbeltet ble søppel registrert på 21 % av stasjonene, mens søppel ble observert på 60 % av stasjonene i Norskerenna. De største mengdene søppel ble dermed funnet i Norskerenna, der plast og uidentifisert søppel dominerte funnene. I kystbeltet ble det ikke registrert plastbasert søppel.
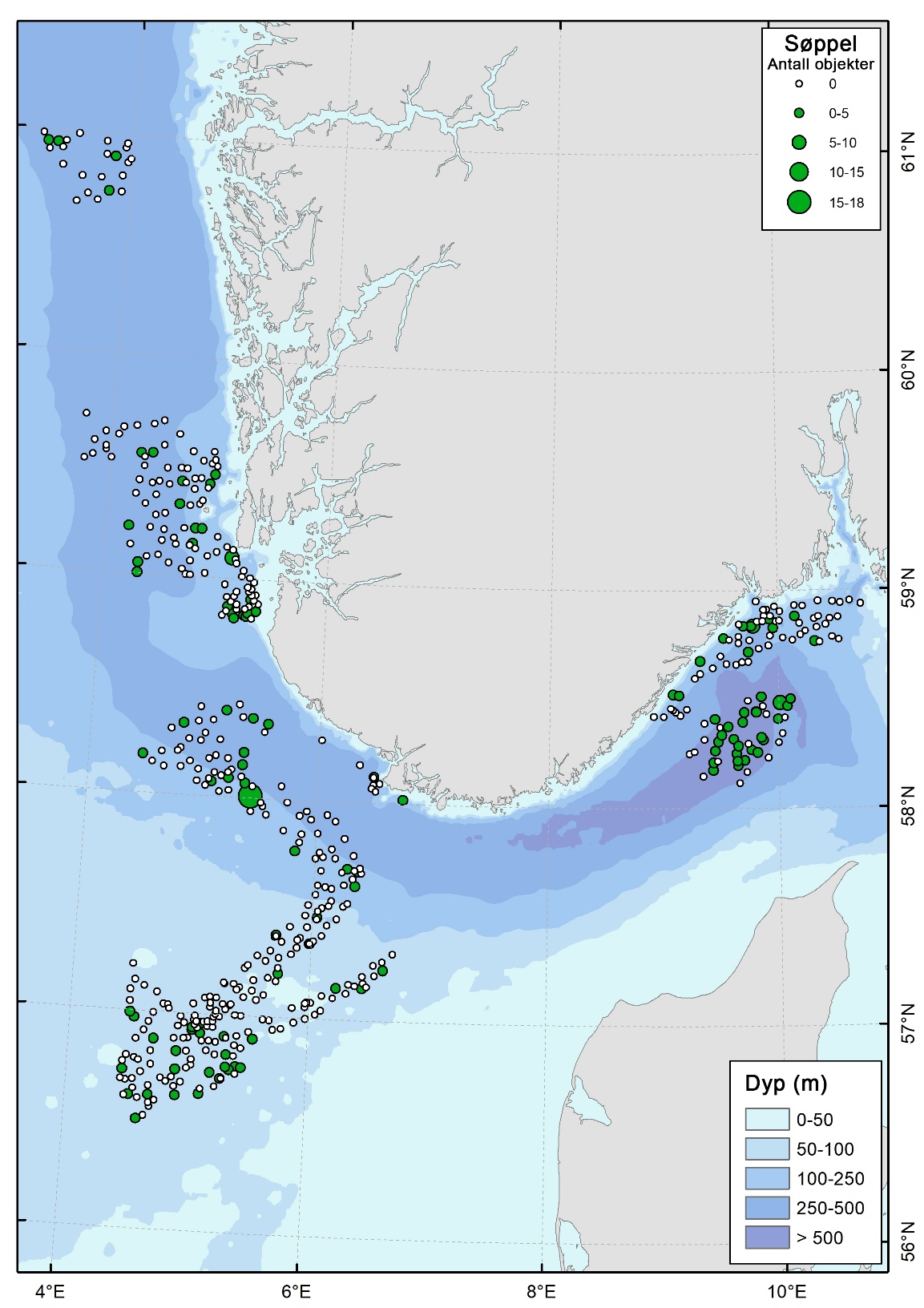
Plast har lav egenvekt og transporteres effektivt med havstrømmer. Norskerenna fungerer derfor trolig som et akkumuleringområde for plast (Figur 19) og annet lett søppel som kan ha sin opprinnelse utenfor selve kartleggingsområdet. Kategorien «uidentifisert søppel» består i stor grad av objekter som antas å være knyttet til dumping av stridsmidler etter andre verdenskrig. Det ble imidlertid ikke observert større ammunisjon eller vrak innenfor kartleggingsområdet i Norskerenna. Ett skipsvrak ble derimot registrert i kystbeltet i Skagerrak, øst for Tromøy.

4 - Diskusjon
4.1 - Overordnede mønstre i biotoper og biodiversitet
Resultatene viser at utbredelsen av biotoper og variasjonen i artsmangfold i Nordsjøen og Skagerrak i hovedsak reflekterer landskapstyper og gradienter i dyp, substrat og hydrografiske forhold. Disse faktorene virker sammen og skaper tydelige økologiske gradienter som fanges opp både i DCA-ordinasjonen og TWINSPAN-klassifikasjonen.
Det høyeste artsmangfoldet ble observert i kystnære og mellomdype områder (omtrent 100–250 m), særlig i kystbeltet ved Utsira, Vestavind og deler av Skagerrak. Disse områdene er preget av komplekse bunnsubstrater med innslag av grus, stein og blokk, som gir økt habitatheterogenitet og tredimensjonal struktur. Slike forhold er kjent for å fremme høyere biodiversitet, og samsvarer godt med tidligere funn fra Mareano og andre regionale studier (Buhl-Mortensen & Buhl-Mortensen 2014).
I kontrast viser de grunne, sanddominerte områdene på Nordsjøplatået et lavere og mer homogent artsmangfold. Disse områdene dekker imidlertid store arealer og har høy økologisk betydning som funksjonelle habitater for mobile arter, inkludert fisk og krepsdyr, og som gyte- og oppvekstområder for flere kommersielt viktige bestander.
De dypere områdene i Skagerrak og Norskerenna har i gjennomsnitt færre taxa per stasjon, men rommer flere sårbare og habitatdannende arter, og representerer dermed økologiske nøkkelområder til tross for lavere lokal artsrikdom. Dette illustrerer at høy biodiversitet og høy strukturell eller økologisk betydning ikke nødvendigvis sammenfaller.
4.2 - Biotopinndeling og økologisk tolkning av Twinspan-gruppene
Biotopinndelingen identifisert med TWINSPAN følger i stor grad kjente geomorfologiske og oseanografiske strukturer i regionen. De artsrike gruppene i kystnære og mellomdype områder (grupper 1–4) er kjennetegnet av hardbunn eller blandet substrat og domineres av habitatdannende svamper og bløtkoraller. Slike samfunn skaper kompleks tredimensjonal struktur og fungerer som viktige leveområder for et bredt spekter av assosierte arter, inkludert både fastsittende og mobile taxa.
De dypere biotopene i Skagerrak og Norskerenna (grupper 5–7) er knyttet til finere sedimenter og er dominert av sjøfjærsamfunn, bambuskorallskog og forekomster av begerkoraller. Disse samfunnene er karakteristiske for stabile dypvannsmiljøer med lav fysisk forstyrrelse og representerer sårbare marine økosystemer (VME) i henhold til internasjonale kriterier. Den tydelige fremtredenen av disse gruppene i analysene understreker Norskerenna sin rolle som et kjerneområde for dypvannsfauna i Nordsjø–Skagerrak-regionen.
De grunneste biotopene (gruppe 9–12), dominert av sand og grus, har generelt lavere artsrikdom, men viser tydelige økologiske funksjoner. Forekomsten av Alcyonium-hager og mobile predatorer indikerer aktive bentiske samfunn med sterke koblinger til pelagiske næringsnett. Flere av disse områdene overlapper også med kjente gyte- og oppvekstområder for fisk, noe som gir dem høy funksjonell betydning til tross for begrenset strukturell kompleksitet
4.3 - Forekomster av sårbare habitater (VME)
Kartleggingen dokumenterer at sårbare habitater er vidt utbredt i Nordsjøen og Skagerrak, men med betydelig regional variasjon i forekomst, tetthet og fragmentering. Sjøfjærsamfunn er det klart mest utbredte sårbare habitatet i materialet, særlig i Skagerrak og Norskerenna. Den brede forekomsten av indikatorartene Funiculina quadrangularis og Kophobelemnon stelliferum viser at disse artene inngår som en viktig komponent i store deler av det dypere bentiske økosystemet. Samtidig viser analysene at tetthetene ofte er lave og fragmenterte, slik at kun en del av forekomstene tilfredsstiller de operasjonelle kriteriene for å bli klassifisert som sjøfjærsamfunn. Selv om sjøfjærsamfunn generelt har lavere artsrikdom av direkte assosiert fauna enn større hornkoraller, fungerer de likevel som viktige biogeniske habitater. De Clippele et al. (2015) viser at sjøfjær bidrar med romlig kompleksitet i ellers sedimentdominerte miljøer og gir skjul, oppvekst- og næringsmuligheter for mobile bunndyr.
Den assosierte faunaen er mindre artsrik enn hos hornkoraller, men består særlig av krepsdyr (bl.a. reker) og slangestjerner som bruker koloniene som tilflukt eller som et gunstig sted for fødeopptak. Enkelte arter viser tydelig vertsspesifisitet; eksempelvis ble slangestjernen Asteronyx loveni kun observert på Funiculina quadrangularis , der den sannsynligvis utnytter høyden over bunnen for effektiv partikkelfangst. Dette understreker at selv relativt spredte sjøfjærforekomster kan ha økologisk betydning utover det som reflekteres i tetthetskriteriene alene. Svampskog forekommer hovedsakelig i hardbunnsområder i mellomdype kystsoner. Her er substrattilgjengelighet den avgjørende faktoren. De fleste svamper krever fast feste, og forekomsten reflekterer direkte tilgjengelighet av grus, stein og blokk. I motsetning til sjøfjær er svampskog mindre knyttet til dyp alene og mer til substrat og vannbevegelse. Dette gir en tydelig romlig adskillelse mellom sedimentdominerte VME-er og hardbunnsbaserte VME-er.
Bløtbunnskorallskog ( Isidella lofotensis ), begerkorallsamfunn ( Flabellum macandrewi ) og O-skjellbanker ( Modiolus modiolus ) forekommer mer fragmentert og regionalt. Indikatorarter for hardbunnskorallskog ble kun observert med spredte forekomster. Det må understrekes at kun deler av Nordsjøen og Skagerrak er kartlagt og at man ikke kan utelukke større utbredelse av sjeldnere sårbare naturtyper eller at det kan finnes hardbunnskorallskog, spesielt i hardbunnsområder i skråningen ned mot Norskerenna. Sammenstilt med observasjoner fra andre videoinspeksjoner og bifangst i reketrål (Figur 20) indikerer Mareanos observasjon av Isidella at bambuskorallskog sannsynligvis er mer utbredt enn Mareano-resultater indikerer alene. Spesielt er sannsynligheten stor i skråningen ned i Norskerenna i vestlig del.
Påvisning av sårbare habitater i denne studien er basert på operasjonelle tetthetskriterier (jf. Tabell 3), hvor frekvensen av observasjoner av indikatorarter langs videolinjene brukes som en proxy for habitatdannelse. Denne tilnærmingen er metodisk robust og nødvendig for standardisert kartlegging, men man kan reise spørsmål om hvorvidt faste terskelverdier for klassifisering fullt ut fanger opp økologisk relevans i alle områder. Naturlige bakgrunnstettheter av habitatdannende arter varierer betydelig mellom havområder. For eksempel viser både tidligere studier og foreliggende materiale at tetthetene av sjøfjær generelt er høyere i Nordsjøen og Skagerrak enn i Barentshavet og på midtnorsk sokkel. Et tetthetsnivå som representerer et strukturdannende samfunn i ett havområde, kan derfor være uvanlig høyt – eller motsatt, relativt lavt – i et annet. Dette innebærer at absolutte terskelverdier for klassifisering av sårbare habitater ikke nødvendigvis er direkte overførbare mellom regioner. Dersom tersklene settes for høyt i områder med naturlig lav bakgrunnstetthet, kan viktige habitatforekomster bli underestimert. Omvendt kan lave terskler i områder med høy naturlig tetthet føre til at svært store arealer klassifiseres som VME, noe som kan redusere forvaltningsmessig presisjon.
Resultatene fra Nordsjøen og Skagerrak illustrerer denne problemstillingen tydelig. Sjøfjær forekommer på en stor andel av stasjonene, men kun en mindre del av disse tilfredsstiller kriteriene for sjøfjærsamfunn. Dette kan indikere at dagens terskler fanger opp de mest strukturelt utviklede forekomstene, men samtidig ikke fullt ut reflekterer den økologiske betydningen av mer spredte forekomster.
Resultatene viser også at forholdet mellom de identifiserte biotopene og forekomsten av sårbare habitater ikke er entydig. Omtrent halvparten av biotopene identifisert med TWINSPAN er karakterisert av indikatorarter for sårbare habitater (Tabell 4), men sammenhengen er ikke én-til-én. Enkelte VME-typer forekommer i flere biotoper, og enkelte biotoper inneholder flere ulike VME-elementer. Dette reflekterer at biotopene primært beskriver helhetlige faunasamfunn, mens VME-klassifiseringen er basert på forekomst og tetthet av bestemte strukturdannende arter. Innenfor enkelte habitattyper kan ulike indikatorarter dessuten dominere i ulike biotoper. For sjøfjær er for eksempel Kophobelemnon stelliferum karakteristisk for biotop 5 i de dype slamdominerte områdene i Skagerrak, mens Funiculina quadrangularis er mer fremtredende i biotop 6 i Norskerenna. I biotop 8 i NS4-området forekommer derimot Balticina sp. hyppigere. Slike forskjeller kan reflektere variasjoner i sedimentegenskaper, som kornstørrelse, organisk innhold eller sedimentstruktur, faktorer som ikke er undersøkt i detalj i denne studien. En mer finmasket analyse av sammensetningen av indikatorarter kunne i prinsippet gi en videre oppdeling av enkelte VME-typer i undergrupper. En slik inndeling vil imidlertid i begrenset grad gi tilleggsverdi i en forvaltningsmessig sammenheng, ettersom de overordnede habitattypene allerede fanger opp de viktigste økologiske og strukturelle forskjellene mellom områdene.
4.4 - Sammenheng mellom biologiske mønstre og menneskelig påvirkning
Registreringene av trålspor og marint søppel viser at menneskelig påvirkning er utbredt i deler av undersøkelsesområdet, særlig i Norskerenna og Ytre Oslofjord. Samtidig er det i disse områdene flere sårbare habitater forekommer, inkludert sjøfjærsamfunn og andre VME-typer. Det romlige sammenfallet mellom påvirkningsindikatorer og sårbare biotoper peker på en potensiell konflikt mellom bevaring av bentiske økosystemer og pågående eller historisk bruk av havbunnen.
Trålspor dokumenterer fysisk forstyrrelse av både sediment og hardbunn. Dette kan ha betydelige konsekvenser for langsomt voksende og strukturdannende arter som sjøfjær, svamper og koraller, som ofte har lav restitusjonsevne og begrenset rekruttering. Gjentatt forstyrrelse kan derfor hindre utvikling og opprettholdelse av velstrukturerte samfunn, selv der de abiotiske forholdene ellers er gunstige. Det tydeligste romlige overlappet mellom trålspor og mekanisk skadde sjøfjærkolonier ble registrert i Kystbelte Skagerrak og NS4. Dette styrker tolkningen av at fysisk bunnpåvirkning bidrar til direkte skade på strukturdannende arter i enkelte områder. Trålspor ble også hyppig observert i den sørlige delen av Norskerenna, særlig i skråningen opp mot Nordsjøplatået. I disse områdene ble det imidlertid ikke registrert skadde sjøfjær eller andre habitatdannende arter. Dette samsvarer med at området i liten grad fremstår som egnet habitat for slike sårbare samfunn, og illustrerer at effekten av bunnpåvirkning må vurderes i lys av den underliggende habitatfordelingen.
Forekomsten av marint søppel, særlig plast, i Norskerenna indikerer at området fungerer som et akkumulasjonsområde for avfall transportert med havstrømmer og topografisk styrt sedimentasjon. Dette representerer en annen type påvirkning enn tråling, og kan påvirke både habitatkvalitet og organismer direkte. Mens effektene på bentiske samfunn i denne studien ikke er kvantifisert, kan plast og annet avfall ha negative konsekvenser for både bunndyr, fisk og marine pattedyr gjennom fysisk skade, tildekking og inntak.
Selv om rapporten ikke analyserer årsak–virkning eksplisitt, viser resultatene at enkelte biotoper og VME-forekomster er eksponert for flere samtidige påvirkningsfaktorer. Dette peker mot behovet for å vurdere kumulative effekter og samlet belastning i videre forvaltnings- og arealplanlegging, der både naturlige miljøgradienter og menneskeskapte påvirkninger inngår i en helhetlig vurdering.
4.5 - Metodiske styrker og begrensninger
En sentral styrke ved denne studien er det store antallet videostasjoner og den brede geografiske dekningen, som gir et solid grunnlag for regional biotopklassifisering. Bruken av standardiserte feltannoteringer kombinert med robuste multivariate metoder (DCA og TWINSPAN) gjør det mulig å identifisere overordnede mønstre i arts- og habitatfordeling, selv når dataene er basert på overordnede taksonomiske nivåer. Samtidig er det viktig å understreke at resultatene er foreløpige. Artsidentifikasjonene er i stor grad gjort på høyt taksonomisk nivå, og kvantitative estimater av tetthet og biomasse mangler. Analysene baserer seg utelukkende på videodata, uten direkte integrering av bunnprøvedata fra fullstasjonene, noe som kan føre til underrepresentasjon av mindre, kryptiske eller infauna-arter. Videre bearbeiding av materialet, inkludert mer detaljert artsbestemmelse og kobling mot prøvetakingsdata, vil kunne gi en mer presis og økologisk detaljert beskrivelse av samfunnsstruktur og prosesser.
5 - Litteratur
Buhl-Mortensen P, Buhl-Mortensen L (2014). Diverse and vulnerable deep-water biotopes in the Hardangerfjord. – Marine Biology Research 10: 253-267, http://dx.doi.org/10.1080/17451000.2013.810759
Buhl-Mortensen L, Buhl-Mortensen P, Dolan MFJ, Holte B (2015a). The MAREANO programme – A full coverage mapping of the Norwegian off-shore benthic environment and fauna. Marine Biology Research 11(1): 4-17. http://www.tandfonline.com/doi/full/10.1080/17451000.2014.952312#.VL5h6EfF9oM
Buhl-Mortensen L, Buhl-Mortensen P, Dolan M, Gonzales-Mirelis G (2015b). Habitat mapping as a tool for conservation and sustainable use of marine resources: some perspectives from the MAREANO programme, Norway. – Journal of Sea Research, Gonzalez-Mirelis G (2022). MAREANO North Sea/Frisk Oslofjord cruise (2022118) — Utsira, SVO “North Sea & Skagerrak #4” and Ytre Hvaler. RV "G.O. Sars" 14.10–30.10 2022. TOKTRAPPORT Nr.13 2022.
Buhl-Mortensen L, Olafsdottir SH, Buhl-Mortensen P, Burgos JM, Ragnarsson SA (2015). Distribution of nine cold-water coral species (Scleractinia and Gorgonacea) in the cold temperate North Atlantic in light of bathymetry and hydrography. Hydrobiologia, 759:39-61.
Buhl-Mortensen L, Burgos JM, Steingrund P, Buhl-Mortensen P, Ólafsdóttir SH, Ragnarsson SÁ (2019) Vulnerable marine ecosystems (VME): Coral and sponge VMEs in Arctic and sub-Arctic waters - Distribution and threats. TemaNord 2019:519, ISSN 0908-6692
Buhl-Mortensen L, Thangstad TH, Søvik G, Wehde H (2023) Sea pens and bamboo corals in Skagerrak and the Norwegian trench, Marine Biology Research, 19:2-3, 191-206, DOI: 10.1080/17451000.2023.2224967
Buhl-Mortensen P, Dolan MFJ, Ross RE, Gonzalez-Mirelis G, Buhl-Mortensen L, Bjarnadóttir LR and Albretsen J (2020). Classification and Mapping of Benthic Biotopes in Arctic and Sub-Arctic Norwegian Waters. Front. Mar. Sci. 7:271. doi: 10.3389/fmars.2020.00271
Buhl-Mortensen P, Bellec V, Strømme ML, Olssøn R, Wiberg DH, Sveistrup AK (2024a). MAREANOTOKT 2023 LEG 2 – Toktrapport 2023001009. TOKTRAPPORT Nr. 2 2024
Buhl-Mortensen P, Buhl-Mortensen L, Bakkeplass K. (2024b). Plastsøppel på havbunnen kartlagt av Mareano. Rapport fra Havforskningen 2024-12, ISSN: 1893-4536
Clippele L. H. De, Buhl-Mortensen P, Buhl-Mortensen L. 2015. Fauna associated with cold water gorgonians and sea pens. Continental Shelf Research, 105:67-78.
Eriksen E, Meeren GI, Nilsen BM, Quillfeldt CH, Johnsen H (2021). Særlig verdifulle og sårbare områder (SVO) i norske havområder - Miljøverdi – En gjennomgang av miljøverdier og grenser i eksisterende SVO og forslag til nye områder. Rapport fra havforskningen År - Nr. 2021-26, ISSN:1893-4536, 359 sider.
FAO (2009). International guidelines for the management of deep-sea fisheries in the high seas. Rome, Italy: Food and Agriculture Organization. Available at https:// www.fao.org/3/i0816t/I0816T.pdf. Accessed September 20, 2024.
Gonzalez-Mirelis G, Buhl-Mortensen P. 2015. Modelling benthic habitats and biotopes off the coast of Norway to support spatial management. Ecological Informatics 30: 284–292, http://dx.doi.org/10.1016/j.ecoinf.2015.06.005
Gonzalez-Mirelis G (2026). MAREANO - NORTH SEA LEG 2 – Cruise report 2025001009 - Sørvest F and Norskerenna FF. TOKTRAPPORT No.1 2026.
Hill MO (1979). TWINSPAN a Fortran Program for Arranging Multivariate Data in an Ordered Two-Way Table by Classification of the Individuals and Attributes. Ecology and Systematic (Ithaca, NY: Cornell University).
Hill MO, and Gauch HG (1980). Detrended correspondence analysis: An improved ordination technique. Vegetatio 42, 47–58. doi: 10.1007/ BF00048870
Kartveit KH, Bellec V, Josefina Johansson J, Piechaud N (2023). Mareano leg 3 2023. TOKTRAPPORT Nr. 9 2024.
Kartveit KH, Bøe R, Strømme ML, Skjefstad J, Fuhrmann MM, Gabrielsen H, Saint-André C, Bienfait AM (2024). MAREANO 2024 LEG1 – Nordsjøtokt 2024007003. TOKTRAPPORT Nr. 11 2024.
Marquez JF, Meeren G, Meeren T, Albretsen J, Ross R, et al. (2024). Liste over forvaltningsrelevante naturenheter. Rapport fra Havforskningen 2024-53. ISSN: 1893-4536. 74pp.
Murillo FJ, Kenchington E, Sacau M, Piper DJW, Wareham V, Muñoz A (2011). New VME indicator species (excluding corals and sponges) and some potential VME elements of the NAFO Regulatory Area. NAFO SCR Doc. 11/73.
OSPAR (2008). Descriptions of habitats on the OSPAR list of threatened and/or declining species and habitats. Reference number 2008-7.
Ross R, Thorsnes T, Meek TJ, Johansson J (2023). MAREANO LEG 1 2023 EASTERCRUISE REPORT – Cruise Report 2023001005. TOKTRAPPORT Nr. 2 2024.
6 - Appendiks
| Art/Taxon | Ant st | Ant obs | Maks forekomst |
| Bacteria | |||
| bacterial mat | 7 | 21 | 11 |
| Rhodophyta encrusting calcareous | 39 | 996 | 109 |
| Protozoa | |||
| Pelosina arborescens | 89 | 291 | 16 |
| Porifera | |||
| Antho dichotoma | 70 | 615 | 33 |
| Craniella sp. | 16 | 67 | 15 |
| Craniella zetlandica | 24 | 107 | 38 |
| Asbestopluma pennatula | 16 | 45 | 10 |
| Axinellidae | 55 | 236 | 29 |
| Axinella infundibuliformis | 71 | 281 | 15 |
| Geodia atlantica | 17 | 54 | 14 |
| Geodia barretti | 46 | 155 | 20 |
| Geodia macandrewii | 13 | 32 | 12 |
| Geodia phlegraei | 6 | 11 | 4 |
| Geodia sp. | 29 | 70 | 7 |
| Haliclona sp. | 6 | 9 | 2 |
| Hexadella dedritifera | 47 | 198 | 21 |
| Hamacantha bowerbanki | 5 | 32 | 22 |
| Hymedesmia crux | 20 | 134 | 36 |
| Hymedesmia jecusculum | 12 | 52 | 13 |
| Hymedesmia paupertas | 63 | 395 | 22 |
| Mycale lingua | 75 | 688 | 52 |
| Oceanapia robusta | 41 | 355 | 65 |
| Phakellia sp. | 81 | 533 | 38 |
| Phakellia ventilabrum | 44 | 324 | 41 |
| Polymastiidae | 34 | 111 | 14 |
| Polymastia sp. | 36 | 85 | 14 |
| Porifera branched dense | 44 | 212 | 24 |
| Quasillina brevis | 37 | 155 | 23 |
| Spinularia spinularia | 4 | 9 | 3 |
| Stryphnus ponderosus | 14 | 45 | 9 |
| Stylocordyla borealis | 19 | 89 | 17 |
| Tentorium semisuberites | 17 | 50 | 8 |
| Thenea sp. | 78 | 828 | 67 |
| Cnidaria | |||
| Actinostola callosa | 9 | 13 | 3 |
| Alcyonium digitatum | 121 | 2209 | 138 |
| Balticina sp. | 23 | 238 | 38 |
| Bolocera tuediae | 166 | 1356 | 67 |
| Caryophyllia smithii | 5 | 15 | 7 |
| Cerianthus lloydii | 19 | 232 | 62 |
| Art/Taxon | Ant st | Ant obs | Maks forekomst |
| Cerianthidae | 105 | 254 | 21 |
| Corymorpha sp. | 6 | 7 | 2 |
| Edwardsiidae | 21 | 110 | 20 |
| Epizoanthus sp. | 15 | 48 | 17 |
| Flabellum macandrewi | 21 | 288 | 57 |
| Flustridae | 116 | 1004 | 132 |
| Funiculina quadrangularis | 223 | 3770 | 245 |
| Halcampoides sp. | 4 | 4 | 1 |
| Hormathia sp. | 9 | 26 | 7 |
| Hormathiidae | 50 | 151 | 24 |
| Hydractiniidae | 23 | 39 | 4 |
| Hydrozoa bush | 23 | 42 | 8 |
| Hydrozoa feather | 17 | 21 | 2 |
| Isidella lofotensis | 15 | 125 | 39 |
| Kophobelemnon stelliferum | 219 | 4208 | 206 |
| Nephtheidae | 4 | 7 | 4 |
| Pachycerianthus multiplicatus | 27 | 96 | 13 |
| Pennatulacea | 158 | 819 | 71 |
| Pennatula phosphorea | 55 | 1572 | 548 |
| Primnoa resedaeformis | 5 | 9 | 4 |
| Ptychodactis patula | 5 | 7 | 3 |
| Sertulariidae | 21 | 48 | 9 |
| Virgularia mirabilis | 24 | 232 | 21 |
| Virgulariidae red | 93 | 776 | 78 |
| Virgularia sp. | 84 | 492 | 44 |
| Virgulariidae | 97 | 1186 | 108 |
| Nemertea | |||
| Nemertea | 17 | 25 | 5 |
| Polychaeta | |||
| Nothria sp. | 10 | 411 | 200 |
| Oxydromus flexuosus | 34 | 1105 | 288 |
| Polychaeta tube | 77 | 260 | 110 |
| Sabellidae | 132 | 358 | 30 |
| Aphroditidae | 51 | 172 | 9 |
| Aphrodita aculeata | 15 | 40 | 7 |
| Ditrupa arietina | 22 | 531 | 265 |
| Filograna implexa | 86 | 353 | 23 |
| Pycnogonida | |||
| Nymphon sp. | 4 | 7 | 4 |
| Pycnogonida | 24 | 34 | 5 |
| Crustacea | |||
| Atlantopandalus propinqvus | 89 | 579 | 33 |
| Balanidae | 18 | 37 | 9 |
| Calocaris macandreae | 6 | 13 | 5 |
| Art/Taxon | Ant st | Ant obs | Maks forekomst |
| Cancer pagurus | 16 | 37 | 6 |
| Caridea | 109 | 366 | 29 |
| Crangonidae | 98 | 540 | 113 |
| Eurycope sp. | 7 | 7 | 1 |
| Galatheoidea | 26 | 43 | 7 |
| Geryon trispinosus | 9 | 11 | 2 |
| Hyas sp. | 5 | 5 | 1 |
| Munida sp. | 284 | 4348 | 175 |
| Neohela sp. | 35 | 49 | 4 |
| Nephrops norvegicus | 105 | 277 | 27 |
| Nudibranchia | 14 | 15 | 2 |
| Pagurus bernhardus | 14 | 24 | 3 |
| Pagurus pubescens | 5 | 5 | 1 |
| Paguridae | 308 | 1728 | 48 |
| Pandalidae | 307 | 9509 | 867 |
| Pontophilus sp. | 25 | 43 | 9 |
| Sabinea sp. | 6 | 7 | 2 |
| Scalpellidae | 11 | 14 | 2 |
| Spirontocaris sp. | 42 | 267 | 35 |
| Mollusca | |||
| Acesta excavata | 5 | 13 | 6 |
| Aporrhais sp. | 6 | 9 | 3 |
| Astarte sp. | 11 | 20 | 8 |
| Buccinidae | 62 | 161 | 14 |
| Buccinoidea | 137 | 621 | 26 |
| Buccinum undatum | 24 | 37 | 6 |
| Cephalopoda benthic | 9 | 10 | 2 |
| Colus sp. | 32 | 50 | 11 |
| Modiolus modiolus | 28 | 521 | 273 |
| Mytilidae | 4 | 18 | 10 |
| Naticidae | 4 | 4 | 1 |
| Neptunea antiqua | 14 | 29 | 5 |
| Neptunea sp. | 31 | 35 | 3 |
| Pectinidae | 31 | 58 | 19 |
| Prosobranchia | 106 | 225 | 9 |
| Pseudamussium peslutrae | 19 | 159 | 62 |
| Echinodermata | |||
| Antedonoidea | 19 | 39 | 7 |
| Asteroidea | 328 | 2572 | 187 |
| Asteriidae | 59 | 294 | 24 |
| Astropecten irregularis | 147 | 2978 | 198 |
| Asteronyx loveni | 158 | 1687 | 83 |
| Asterias rubens | 156 | 1313 | 91 |
| Bathyplotes natans | 6 | 10 | 4 |
| Art/Taxon | Ant st | Ant obs | Maks forekomst |
| Ceramaster granularis | 46 | 158 | 10 |
| Echinus esculentus | 24 | 130 | 24 |
| Echinoidea irregular | 44 | 194 | 45 |
| Echinoidea regular | 39 | 153 | 31 |
| Echinus sp. | 4 | 5 | 2 |
| Gorgonocephalus sp. | 5 | 8 | 3 |
| Gracilechinus acutus | 193 | 5506 | 668 |
| Hippasteria phrygiana | 16 | 18 | 2 |
| Holothuroidea | 9 | 11 | 3 |
| Henricia sp. | 151 | 1026 | 30 |
| Leptasterias muelleri | 7 | 14 | 5 |
| Leptasterias sp. | 31 | 240 | 35 |
| Mesothuria intestinalis | 64 | 287 | 14 |
| Ophioscolecidae | 19 | 133 | 20 |
| Ophiuroidea | 235 | 2288 | 676 |
| Ophiopholis aculeata | 8 | 14 | 7 |
| Ophiura sarsii | 7 | 14 | 4 |
| Ophiura sp. | 4 | 7 | 4 |
| Parastichopus tremulus | 191 | 1159 | 39 |
| Pontaster tenuispinus | 5 | 9 | 4 |
| Porania pulvillus | 40 | 163 | 20 |
| Poraniomorpha sp. | 15 | 22 | 6 |
| Poraniidae | 5 | 6 | 2 |
| Psilaster andromeda | 81 | 441 | 42 |
| Pteraster militaris | 7 | 15 | 6 |
| Pteraster sp. | 16 | 18 | 2 |
| Rhizocrinus lofotensis | 10 | 21 | 7 |
| Solaster sp. | 25 | 48 | 6 |
| Spatangoida | 18 | 95 | 30 |
| Spatangus sp. | 39 | 377 | 45 |
| Stichastrella rosea | 78 | 514 | 68 |
| Echiura | |||
| Bonelliidae | 46 | 503 | 71 |
| Maxmuelleria faex | 30 | 118 | 22 |
| Bryozoa | |||
| Brosme brosme | 9 | 12 | 2 |
| Bryozoa branched | 12 | 18 | 4 |
| Bryozoa calcareous branched | 29 | 58 | 7 |
| Bryozoa encrusting | 39 | 103 | 14 |
| Horneridae | 30 | 47 | 5 |
| Reteporella beaniana | 59 | 151 | 17 |
| Brachiopoda | |||
| Brachiopoda | 35 | 46 | 3 |
| Novocrania anomala | 27 | 73 | 13 |
| Art/Taxon | Ant st | Ant obs | Maks forekomst |
| Ascidiacea | |||
| Ascidia conchilega | 4 | 17 | 8 |
| Ascidiacea colonial | 27 | 61 | 10 |
| Ascidiacea solitary | 56 | 259 | 24 |
| Ascidia sp. | 7 | 15 | 5 |
| Ascidia virginea | 19 | 65 | 30 |
| Ascidiacea | 22 | 134 | 101 |
| Didemnidae | 29 | 57 | 9 |
| Styela rustica | 4 | 25 | 9 |
| Styela sp. | 17 | 93 | 15 |
| Teleostei | |||
| Amblyraja radiata | 4 | 4 | 1 |
| Ammodytidae | 22 | 83 | 12 |
| Anarhichas sp. | 5 | 7 | 2 |
| Chimaera monstrosa | 72 | 106 | 7 |
| Corystes cassivelaunus | 21 | 59 | 10 |
| Coryphaenoides rupestris | 34 | 126 | 13 |
| Cottidae | 18 | 40 | 8 |
| Gadiculus argenteus | 84 | 457 | 25 |
| Gadus morhua | 73 | 183 | 12 |
| Gadidae | 150 | 331 | 20 |
| Galeus melastomus | 26 | 32 | 3 |
| Glyptocephalus cynoglossus | 35 | 45 | 3 |
| Helicolenus dactylopterus | 17 | 34 | 7 |
| Etmopterus spinax | 50 | 90 | 11 |
| Lepidorhombus boscii | 5 | 7 | 3 |
| Limanda limanda | 45 | 167 | 15 |
| Lophius piscatorius | 4 | 4 | 1 |
| Lophius sp. | 17 | 19 | 2 |
| Lithodes maja | 64 | 109 | 7 |
| Lithothamnion sp. | 4 | 9 | 5 |
| Luidia sarsii | 71 | 488 | 41 |
| Luidia sp. | 4 | 11 | 7 |
| Lumpeninae | 20 | 38 | 7 |
| Lycodonus sp. | 7 | 8 | 2 |
| Lycenchelys sarsii | 6 | 7 | 2 |
| Lycenchelys sp. | 19 | 53 | 8 |
| Lycodes sp. | 37 | 40 | 3 |
| Macrourus berglax | 7 | 30 | 9 |
| Macrouridae | 16 | 107 | 20 |
| Maurolicus muelleri | 6 | 33 | 19 |
| Melanogrammus aeglefinus | 157 | 660 | 36 |
| Merlangius merlangus | 34 | 59 | 9 |
| Merluccius merluccius | 7 | 13 | 4 |
| Art/Taxon | Ant st | Ant obs | Maks forekomst |
| Molva dipterygia | 7 | 8 | 2 |
| Molva molva | 23 | 51 | 11 |
| Myxine glutinosa | 191 | 1901 | 144 |
| Pollachius pollachius | 5 | 12 | 7 |
| Pollachius virens | 125 | 547 | 43 |
| Pollachius sp. | 5 | 10 | 3 |
| Rajella fyllae | 6 | 6 | 1 |
| Phycis blennoides | 4 | 5 | 2 |
| Pleuronectiformes | 302 | 1933 | 31 |
| Pleuronectes platessa | 38 | 58 | 4 |
| Rajiformes | 54 | 70 | 4 |
| Sebastidae | 6 | 8 | 2 |
| Sebastes sp. | 45 | 70 | 8 |
| Selachii | 10 | 16 | 3 |
| Triglidae | 74 | 159 | 10 |
| Trisopterus esmarkii | 29 | 132 | 14 |
| Trisopterus minutus | 4 | 10 | 5 |
| Trisopterus sp. | 109 | 561 | 34 |
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| Toktnr 2022118, FF G.O. Sars, Toktleder Genoveva Gonzalez-Mirelis | |||||||||||||
| 3110 | 14.10.22 | Utsira kontroll | 59°42.784633 | 3°44.82871 | 272 | 1 | |||||||
| 3111 | 14.10.22 | Utsira kontroll | 59°38.31623 | 3°56.47601 | 275 | 1 | |||||||
| 3112 | 15.10.22 | Utsira kontroll | 59°37.75956 | 4°03.33029 | 274 | 1 | |||||||
| 3113 | 15.10.22 | Utsira kontroll | 59°39.9092 | 4°06.01301 | 267 | 1 | |||||||
| 3114 | 15.10.22 | Utsira kontroll | 59°40.414439 | 4°13.086054 | 265 | 1 | |||||||
| 3115 | 15.10.22 | Utsira kontroll | 59°41.20876 | 4°22.28581 | 273 | 1 | |||||||
| 3116 | 15.10.22 | Utsira kontroll | 59°42.221933 | 4°27.623028 | 279 | 1 | |||||||
| 3117 | 15.10.22 | Utsira kontroll | 59°38.76056 | 4°36.36223 | 267 | 1 | |||||||
| 3118 | 15.10.22 | Utsira kontroll | 59°35.54311 | 4°28.45487 | 261 | 1 | |||||||
| 3119 | 15.10.22 | Utsira kontroll | 59°33.25531 | 4°22.62183 | 260 | 1 | |||||||
| 3120 | 15.10.22 | Utsira kontroll | 59°33.03692 | 4°16.336712 | 266 | 1 | 5 | 1 | 2 | 1 | 1 | ||
| 3121 | 16.10.22 | Utsira kontroll | 59°32.015028 | 4°18.305789 | 265 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3122 | 16.10.22 | Utsira kontroll | 59°31.226673 | 4°00.393665 | 279 | 1 | |||||||
| 3123 | 16.10.22 | Utsira kontroll | 59°33.44232 | 3°57.000615 | 275 | 1 | |||||||
| 3124 | 16.10.22 | Utsira kontroll | 59°34.44942 | 3°56.924498 | 275 | 1 | |||||||
| 3125 | 16.10.22 | Utsira kontroll | 59°35.78222 | 3°50.48996 | 268 | 1 | |||||||
| 3126 | 16.10.22 | Utsira kontroll | 59°31.8857 | 3°50.04573 | 265 | 1 | |||||||
| 3127 | 16.10.22 | KB-Utsira | 59°29.69164 | 4°38.51581 | 257 | 1 | |||||||
| 3128 | 16.10.22 | KB-Utsira | 59°29.553798 | 4°41.710181 | 254 | 1 | |||||||
| 3129 | 17.10.22 | KB-Utsira | 59°25.674089 | 4°41.47088 | 261 | 1 | |||||||
| 3130 | 17.10.22 | KB-Utsira | 59°25.918684 | 4°39.447333 | 265 | 1 | |||||||
| 3131 | 17.10.22 | KB-Utsira | 59°17.012324 | 4°31.384914 | 269 | 1 | 10 | 1 | 1 | 1 | 1 | 1 | |
| 3132 | 17.10.22 | KB-Utsira | 59°19.451244 | 4°44.394037 | 257 | 1 | |||||||
| 3133 | 18.10.22 | KB-Utsira | 59°19.70717 | 4°38.82358 | 269 | 1 | |||||||
| 3134 | 18.10.22 | KB-Utsira | 59°24.858936 | 4°32.705751 | 270 | 1 | |||||||
| 3135 | 18.10.22 | KB-Utsira | 59°12.783345 | 4°23.983704 | 256 | 1 | |||||||
| 3136 | 18.10.22 | KB-Utsira | 59°12.873089 | 4°12.266428 | 278 | 1 | 9 | 3 | 1 | 1 | 1 | 1 | |
| 3137 | 18.10.22 | KB-Utsira | 59°12.363896 | 4°31.415589 | 256 | 1 | |||||||
| 3138 | 18.10.22 | KB-Utsira | 59°13.09992 | 4°42.02716 | 276 | 1 | |||||||
| 3139 | 19.10.22 | KB-Utsira | 59°10.286573 | 4°36.74787 | 266 | 1 | |||||||
| 3140 | 19.10.22 | KB-Utsira | 59°08.52771 | 4°38.033924 | 268 | 1 | |||||||
| 3141 | 29.10.22 | KB-Utsira | 59°08.489821 | 4°45.28181 | 176 | 2 | |||||||
| 3142 | 20.10.22 | NS4 | 58°46.823499 | 9°31.262005 | 113 | 1 | |||||||
| 3143 | 20.10.22 | NS4 | 58°46.42757 | 9°34.34922 | 176 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3144 | 20.10.22 | NS4 | 58°45.514904 | 9°39.020053 | 234 | 1 | |||||||
| 3145 | 20.10.22 | NS4 | 58°47.2842 | 9°39.04292 | 179 | 1 | |||||||
| 3146 | 20.10.22 | NS4 | 58°50.658714 | 9°39.36449 | 119 | 1 | |||||||
| 3147 | 20.10.22 | NS4 | 58°46.21494 | 9°44.91533 | 413 | 1 | |||||||
| 3148 | 21.10.22 | NS4 | 58°50.176621 | 9°45.920062 | 146 | 1 | |||||||
| 3149 | 21.10.22 | NS4 | 58°52.959748 | 9°52.767075 | 276 | 1 | 8 | 1 | 1 | 1 | 1 | 1 | |
| 3150 | 21.10.22 | NS4 | 58°54.685131 | 9°54.045698 | 147 | 1 | |||||||
| 3151 | 21.10.22 | NS4 | 58°52.88618 | 9°55.98393 | 129 | 1 | |||||||
| 3152 | 21.10.22 | NS4 | 58°52.10741 | 9°55.51989 | 146 | 1 | |||||||
| 3153 | 21.10.22 | NS4 | 58°47.443633 | 9°55.883511 | 184 | 1 | |||||||
| 3154 | 21.10.22 | NS4 | 58°49.644364 | 9°57.33695 | 179 | 1 | |||||||
| 3155 | 21.10.22 | NS4 | 58°51.457569 | 9°59.069896 | 161 | 1 | |||||||
| 3156 | 21.10.22 | NS4 | 58°55.481863 | 9°58.322553 | 38 | 1 | |||||||
| 3157 | 21.10.22 | NS4 | 58°54.23437 | 10°01.115667 | 94 | 1 | |||||||
| 3158 | 22.10.22 | NS4 | 58°51.643956 | 10°00.836708 | 215 | 1 | |||||||
| 3159 | 22.10.22 | NS4 | 58°46.900302 | 10°02.762999 | 310 | 1 | |||||||
| 3160 | 22.10.22 | NS4 | 58°46.274045 | 10°07.715414 | 239 | 1 | |||||||
| 3161 | 22.10.22 | NS4 | 58°52.794379 | 10°08.881553 | 178 | 1 | |||||||
| 3162 | 22.10.22 | NS4 | 58°55.862089 | 10°08.8548 | 124 | 1 | |||||||
| 3163 | 22.10.22 | NS4 | 58°50.97983 | 10°11.41198 | 193 | 1 | |||||||
| 3164 | 22.10.22 | NS4 | 58°47.77751 | 10°11.92741 | 193 | 1 | |||||||
| 3165 | 22.10.22 | NS4 | 58°48.76679 | 10°14.82401 | 170 | 1 | 5 | 3 | 1 | 3 | 1 | 1 | |
| 3166 | 22.10.22 | NS4 | 58°55.64285 | 10°13.46133 | 121 | 1 | |||||||
| 3167 | 23.10.22 | NS4 | 58°52.108892 | 10°19.386501 | 182 | 1 | |||||||
| 3168 | 23.10.22 | NS4 | 58°45.8941 | 10°19.47947 | 165 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3169 | 23.10.22 | NS4 | 58°49.73065 | 10°20.72391 | 178 | 1 | |||||||
| 3170 | 23.10.22 | NS4 | 58°52.78211 | 10°21.45786 | 189 | 1 | |||||||
| 3171 | 23.10.22 | NS4 | 58°57.060423 | 10°21.583491 | 121 | 1 | |||||||
| 3172 | 23.10.22 | NS4 | 58°50.41764 | 10°25.351967 | 157 | 1 | |||||||
| 3173 | 23.10.22 | NS4 | 58°47.014971 | 10°28.99181 | 120 | 1 | |||||||
| 3174 | 23.10.22 | NS4 | 58°52.10831 | 10°27.72462 | 144 | 1 | |||||||
| 3175 | 23.10.22 | NS4 | 58°56.747492 | 10°29.598282 | 157 | 1 | 7 | 2 | 1 | 2 | 1 | 1 | |
| 3176 | 24.10.22 | NS4 | 58°56.41599 | 10°33.32268 | 247 | 1 | |||||||
| 3177 | 24.10.22 | NS4 | 58°52.5302 | 10°32.16253 | 147 | 1 | |||||||
| 3178 | 24.10.22 | NS4 | 58°57.03291 | 10°38.23988 | 343 | 1 | |||||||
| 3179 | 24.10.22 | NS4 | 58°56.05391 | 10°44.285051 | 76 | 1 | |||||||
| 3180 | 27.10.22 | NS4 | 58°46.38168 | 10°32.3814 | 109 | 1 | |||||||
| 3181 | 27.10.22 | NS4 | 58°45.70876 | 10°21.872 | 158 | 1 | |||||||
| 3182 | 27.10.22 | NS4 | 58°44.835791 | 9°54.889012 | 321 | 1 | |||||||
| 3183 | 27.10.22 | NS4 | 58°44.404226 | 9°50.798254 | 412 | 1 | 7 | 2 | 1 | 3 | 1 | 1 | |
| 3184 | 29.10.22 | Utsira Nord | 59°01.886976 | 4°41.933986 | 277 | 1 | 6 | 2 | 1 | 2 | 1 | 1 | |
| 3185 | 29.10.22 | Utsira Nord | 59°04.27921 | 4°46.03876 | 254 | 1 | |||||||
| 3186 | 29.10.22 | Utsira Nord | 59°09.097424 | 4°46.976736 | 235 | 1 | |||||||
| 3187 | 29.10.22 | Utsira Nord | 59°30.72987 | 3°45.56254 | 258 | 1 | |||||||
| Toktnr 2023001005, FF G.O. Sars, Toktleder: Rebecca Ross | 0 | ||||||||||||
| 3188 | 30.03.23 | KB-Utsira | 59°34.56609 | 4°55.817505 | 196 | 1 | 5 | 1 | 1 | 2 | 1 | 1 | |
| 3189 | 31.03.23 | KB-Utsira | 59°34.37611 | 4°44.30687 | 237 | 1 | |||||||
| 3190 | 31.03.23 | KB-Utsira | 59°19.93798 | 4°49.59401 | 211 | 1 | 1 | 1 | |||||
| 3191 | 31.03.23 | KB-Utsira | 59°20.934271 | 4°51.000815 | 100 | 1 | |||||||
| 3192 | 31.03.23 | KB-Utsira | 59°23.93948 | 4°46.16764 | 245 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3193 | 31.03.23 | KB-Utsira | 59°24.571154 | 4°53.563071 | 120 | 1 | |||||||
| 3194 | 31.03.23 | KB-Utsira | 59°25.635909 | 4°54.59557 | 140 | 1 | |||||||
| 3195 | 31.03.23 | KB-Utsira | 59°27.130891 | 4°49.789053 | 203 | 1 | |||||||
| 3196 | 01.04.23 | KB-Utsira | 59°07.124093 | 5°00.272747 | 236 | 1 | 7 | 1 | 1 | 2 | 1 | 1 | |
| 3197 | 01.04.23 | KB-Utsira | 59°13.22751 | 4°48.07637 | 246 | 1 | |||||||
| 3198 | 01.04.23 | KB-Utsira | 59°07.61949 | 4°48.505326 | 243 | 1 | |||||||
| 3199 | 01.04.23 | KB-Utsira | 59°05.76639 | 4°55.26922 | 243 | 1 | |||||||
| 3200 | 02.04.23 | KB-Utsira | 58°58.351267 | 5°05.525245 | 247 | 1 | 6 | 1 | 1 | 2 | 1 | 1 | |
| 3201 | 02.04.23 | NSJ-1 | 57°04.571121 | 5°13.354361 | 55 | 1 | 6 | 2 | 1 | 2 | 1 | ||
| 3202 | 02.04.23 | NSJ-1 | 57°00.335611 | 4°55.69569 | 57 | 1 | 1 | ||||||
| 3203 | 02.04.23 | NSJ-1 | 57°08.634552 | 5°17.115004 | 45 | 1 | |||||||
| 3204 | 02.04.23 | NSJ-1 | 57°07.78256 | 5°24.58659 | 50 | 1 | |||||||
| 3205 | 02.04.23 | NSJ-1 | 57°07.169568 | 5°23.227406 | 48 | 1 | |||||||
| 3206 | 02.04.23 | NSJ-1 | 57°05.037649 | 5°26.882956 | 55 | 1 | |||||||
| 3207 | 03.04.23 | NSJ-1 | 56°59.89755 | 5°25.95882 | 49 | 1 | |||||||
| 3208 | 04.04.23 | NSJ-1 | 57°07.026601 | 5°31.101901 | 48 | 1 | 6 | 1 | 1 | 2 | 1 | ||
| 3209 | 03.04.23 | NSJ-1 | 57°09.96978 | 5°31.63242 | 50 | 1 | |||||||
| 3210 | 03.04.23 | NSJ-1 | 57°13.11389 | 5°26.78889 | 51 | 1 | |||||||
| 3211 | 03.04.23 | NSJ-1 | 57°02.47011 | 5°20.023234 | 49 | 1 | 1 | ||||||
| 3212 | 03.04.23 | NSJ-1 | 57°01.56273 | 5°18.04187 | 57 | 1 | |||||||
| 3213 | 03.04.23 | NSJ-1 | 57°02.93267 | 5°16.66043 | 58 | 1 | |||||||
| 3214 | 03.04.23 | NSJ-1 | 57°02.971146 | 5°20.618977 | 55 | 1 | |||||||
| 3215 | 03.04.23 | NSJ-1 | 57°22.062895 | 5°41.187608 | 73 | 1 | |||||||
| 3216 | 03.04.23 | NSJ-1 | 57°22.355964 | 5°41.875129 | 74 | 1 | |||||||
| 3217 | 03.04.23 | NSJ-1 | 57°25.08208 | 5°48.83085 | 77 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3218 | 03.04.23 | NSJ-1 | 57°25.150455 | 5°57.549694 | 75 | 1 | |||||||
| 3219 | 03.04.23 | NSJ-1 | 57°22.049571 | 5°54.077299 | 76 | 1 | |||||||
| 3220 | 03.04.23 | NSJ-1 | 57°21.552627 | 5°52.729494 | 78 | 1 | |||||||
| 3221 | 04.04.23 | NSJ-1 | 57°20.862025 | 5°45.00829 | 74 | 1 | |||||||
| 3222 | 05.04.23 | NSJ-1 | 57°17.5546 | 5°50.027335 | 71 | 2 | |||||||
| 3223 | 04.04.23 | NSJ-1 | 57°13.206064 | 5°42.41038 | 56 | 1 | 1 | ||||||
| 3224 | 04.04.23 | NSJ-1 | 57°36.850482 | 6°02.044474 | 143 | 1 | 5 | 2 | 1 | 2 | 1 | 1 | |
| 3225 | 05.04.23 | NSJ-1 | 57°22.654533 | 5°41.770684 | 77 | 1 | 1 | ||||||
| 3226 | 05.04.23 | NSJ-1 | 57°18.190169 | 5°39.464465 | 68 | 1 | |||||||
| 3227 | 05.04.23 | NSJ-1 | 57°15.359132 | 5°37.309215 | 56 | 1 | |||||||
| 3228 | 05.04.23 | NSJ-1 | 57°16.716119 | 5°32.899583 | 58 | 1 | 1 | ||||||
| 3229 | 05.04.23 | NSJ-1 | 57°13.870288 | 5°28.264293 | 53 | 1 | |||||||
| 3230 | 05.04.23 | NSJ-1 | 57°16.41007 | 5°43.66968 | 67 | 1 | |||||||
| 3231 | 05.04.23 | NSJ-1 | 57°19.4708 | 5°52.663 | 74 | 1 | |||||||
| 3232 | 06.04.23 | NSJ-1 | 57°21.390961 | 6°00.790847 | 77 | 1 | |||||||
| 3233 | 06.04.23 | NSJ-1 | 57°22.928856 | 6°05.103277 | 79 | 1 | |||||||
| 3234 | 06.04.23 | NSJ-1 | 57°25.834748 | 6°07.474915 | 72 | 1 | |||||||
| 3235 | 06.04.23 | NSJ-1 | 57°27.65818 | 6°12.41761 | 95 | 1 | |||||||
| 3236 | 06.04.23 | NSJ-1 | 57°31.273249 | 6°15.395719 | 90 | 1 | |||||||
| 3237 | 06.04.23 | NSJ-1 | 57°31.998325 | 6°17.694836 | 104 | 1 | |||||||
| 3238 | 06.04.23 | NSJ-1 | 57°32.655024 | 6°09.339086 | 94 | 1 | |||||||
| 3239 | 06.04.23 | NSJ-1 | 57°31.202038 | 6°08.233018 | 95 | 1 | |||||||
| 3240 | 06.04.23 | NSJ-1 | 57°30.62354 | 6°04.64164 | 68 | 1 | |||||||
| 3241 | 06.04.23 | NSJ-1 | 57°29.594341 | 6°02.619701 | 83 | 1 | |||||||
| 3242 | 06.04.23 | NSJ-1 | 57°27.39881 | 6°02.30041 | 88 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3243 | 06.04.23 | NSJ-1 | 57°28.360931 | 5°56.307874 | 88 | 1 | |||||||
| 3244 | 06.04.23 | NSJ-1 | 57°31.265793 | 5°57.437749 | 86 | 1 | |||||||
| 3245 | 06.04.23 | NSJ-1 | 57°34.24541 | 6°01.12473 | 125 | 1 | |||||||
| 3246 | 06.04.23 | NSJ-1 | 57°36.428221 | 6°05.920326 | 145 | 1 | |||||||
| 3247 | 06.04.23 | NSJ-1 | 57°36.064554 | 6°09.271194 | 143 | 1 | |||||||
| 3248 | 07.04.23 | NSJ-2 | 57°12.016977 | 6°26.4163 | 65 | 1 | |||||||
| 3249 | 07.04.23 | NSJ-2 | 57°10.244637 | 6°26.917361 | 67 | 1 | |||||||
| 3250 | 07.04.23 | NSJ-2 | 57°09.434591 | 6°22.841469 | 66 | 1 | |||||||
| 3251 | 07.04.23 | NSJ-2 | 57°09.765662 | 6°19.717549 | 67 | 1 | |||||||
| 3252 | 07.04.23 | NSJ-2 | 57°08.662609 | 6°13.291586 | 64 | 1 | |||||||
| 3253 | 07.04.23 | NSJ-2 | 57°04.36257 | 6°00.70523 | 46 | 1 | |||||||
| 3254 | 07.04.23 | NSJ-2 | 57°04.425761 | 5°58.867373 | 44 | 1 | |||||||
| 3255 | 07.04.23 | KB-Utsira | 58°49.872496 | 5°04.895803 | 203 | 1 | |||||||
| 3256 | 08.04.23 | KB-Utsira | 58°49.120697 | 5°11.148557 | 193 | 1 | |||||||
| 3257 | 08.04.23 | KB-Utsira | 58°50.02843 | 5°16.42541 | 121 | 1 | |||||||
| 3258 | 08.04.23 | KB-Utsira | 58°49.691813 | 5°17.642872 | 85 | 1 | |||||||
| 3259 | 08.04.23 | KB-Utsira | 58°50.664598 | 5°18.641798 | 75 | 1 | |||||||
| 3260 | 08.04.23 | KB-Utsira | 58°51.359827 | 5°22.854051 | 102 | 1 | |||||||
| 3261 | 08.04.23 | KB-Utsira | 58°53.36448 | 5°23.68163 | 92 | 1 | |||||||
| 3262 | 08.04.23 | KB-Utsira | 58°53.085995 | 5°20.840681 | 116 | 1 | 6 | 1 | 1 | 2 | 1 | ||
| 3263 | 08.04.23 | KB-Utsira | 58°52.38648 | 5°18.84078 | 114 | 1 | |||||||
| 3264 | 08.04.23 | KB-Utsira | 58°51.754842 | 5°16.841847 | 114 | 1 | |||||||
| 3265 | 08.04.23 | KB-Utsira | 58°51.523007 | 5°12.56747 | 163 | 1 | |||||||
| 3266 | 08.04.23 | KB-Utsira | 58°50.622104 | 5°10.088271 | 186 | 1 | |||||||
| 3267 | 08.04.23 | KB-Utsira | 58°51.05578 | 5°07.09188 | 198 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3268 | 08.04.23 | KB-Utsira | 58°52.332308 | 5°07.699241 | 198 | 1 | |||||||
| 3269 | 08.04.23 | KB-Utsira | 58°53.710882 | 5°09.125954 | 173 | 1 | |||||||
| 3270 | 08.04.23 | KB-Utsira | 58°52.970031 | 5°12.407928 | 145 | 1 | |||||||
| 3271 | 08.04.23 | KB-Utsira | 58°54.49593 | 5°19.82454 | 117 | 1 | |||||||
| 3272 | 08.04.23 | KB-Utsira | 58°54.228353 | 5°21.387097 | 115 | 1 | |||||||
| 3273 | 08.04.23 | KB-Utsira | 58°55.672574 | 5°22.276164 | 58 | 1 | |||||||
| 3274 | 09.04.23 | KB-Utsira | 58°55.545876 | 5°20.102381 | 126 | 1 | |||||||
| 3275 | 09.04.23 | KB-Utsira | 58°55.073671 | 5°10.798098 | 159 | 1 | |||||||
| 3276 | 09.04.23 | KB-Utsira | 58°54.864179 | 5°07.044039 | 190 | 1 | |||||||
| 3277 | 09.04.23 | KB-Utsira | 58°56.42815 | 5°11.86571 | 251 | 1 | |||||||
| 3278 | 09.04.23 | KB-Utsira | 58°56.64589 | 5°18.48621 | 151 | 1 | |||||||
| 3279 | 09.04.23 | KB-Utsira | 58°57.229213 | 5°19.773017 | 121 | 1 | |||||||
| 3280 | 09.04.23 | KB-Utsira | 58°57.70006 | 5°17.84614 | 254 | 1 | |||||||
| 3281 | 09.04.23 | KB-Utsira | 59°01.085608 | 4°54.547784 | 235 | 1 | |||||||
| 3282 | 09.04.23 | KB-Utsira | 59°08.675217 | 5°05.614099 | 166 | 1 | |||||||
| 3283 | 09.04.23 | KB-Utsira | 59°07.85983 | 5°08.889798 | 124 | 1 | |||||||
| 3284 | 09.04.23 | KB-Utsira | 59°05.73934 | 5°08.53838 | 165 | 2 | 1 | ||||||
| 3285 | 09.04.23 | KB-Utsira | 59°05.395985 | 5°10.352258 | 161 | 1 | |||||||
| 3286 | 09.04.23 | KB-Utsira | 59°04.235114 | 5°11.080321 | 182 | 1 | |||||||
| 3287 | 10.04.23 | KB-Utsira | 59°02.30928 | 5°15.26556 | 261 | 1 | |||||||
| 3288 | 10.04.23 | Boknafjorden | 59°07.96931 | 5°58.290456 | 260 | 1 | 1 | ||||||
| 3289 | 10.04.23 | Boknafjorden | 59°07.925376 | 5°58.906575 | 251 | 1 | |||||||
| 3290 | 10.04.23 | Boknafjorden | 59°07.852897 | 5°58.035062 | 289 | 1 | |||||||
| 3291 | 11.04.23 | KB-Utsira | 58°59.2372 | 5°21.033061 | 88 | 1 | |||||||
| 3292 | 11.04.23 | KB-Utsira | 59°00.292046 | 5°18.718235 | 277 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3293 | 11.04.23 | KB-Utsira | 59°00.65874 | 5°14.25178 | 230 | 1 | |||||||
| 3294 | 11.04.23 | KB-Utsira | 59°11.204231 | 5°00.325182 | 182 | 1 | |||||||
| 3295 | 11.04.23 | KB-Utsira | 59°13.29444 | 4°51.609252 | 217 | 1 | |||||||
| 3296 | 11.04.23 | KB-Utsira | 59°26.916144 | 4°46.075657 | 237 | 1 | |||||||
| 3297 | 11.04.23 | KB-Utsira | 59°28.291018 | 4°56.963667 | 178 | 1 | |||||||
| 3298 | 11.04.23 | KB-Utsira | 59°30.623575 | 4°57.960214 | 144 | 1 | |||||||
| 3299 | 11.04.23 | KB-Utsira | 59°31.145427 | 4°55.161648 | 147 | 1 | |||||||
| 3300 | 11.04.23 | KB-Utsira | 59°32.364076 | 4°56.726464 | 171 | 1 | |||||||
| 3301 | 11.04.23 | KB-Utsira | 59°31.988269 | 4°50.220154 | 195 | 1 | |||||||
| Toktnr 2023001009, FF G.O. Sars, Toktleder: Pål Buhl-Mortensen | 0 | ||||||||||||
| 3183 | 12.07.23 | KB-Skagerrak | 58°44.040056 | 9°49.80229 | 394 | 1 | 7 | 2 | 1 | 3 | 1 | 1 | |
| 3188 | 03.07.23 | KB-Utsira | 59°34.511578 | 4°55.864912 | 194 | 1 | 5 | 1 | 1 | 2 | 1 | 1 | |
| 3190 | 04.07.23 | KB-Utsira | 59°19.92759 | 4°49.73522 | 205 | 1 | 1 | 1 | |||||
| 3196 | 04.07.23 | KB-Utsira | 59°07.11861 | 5°00.121784 | 237 | 1 | 7 | 1 | 1 | 2 | 1 | 1 | |
| 3200 | 04.07.23 | KB-Utsira | 58°58.3688 | 5°05.623504 | 248 | 1 | 6 | 1 | 1 | 2 | 1 | 1 | |
| 3201 | 05.07.23 | NSJ-1 | 57°04.592803 | 5°13.46598 | 57 | 1 | 6 | 2 | 1 | 2 | 1 | ||
| 3208 | 05.07.23 | NSJ-1 | 57°06.987918 | 5°31.150091 | 50 | 1 | 6 | 1 | 1 | 2 | 1 | ||
| 3224 | 06.07.23 | NSJ-1 | 57°36.796448 | 6°02.011973 | 145 | 1 | 5 | 2 | 1 | 2 | 1 | 1 | |
| 3262 | 05.07.23 | KB-Utsira | 58°53.085092 | 5°20.726362 | 118 | 1 | 6 | 1 | 1 | 2 | 1 | ||
| 3302 | 04.07.23 | KB-Utsira | 58°49.168005 | 5°20.499999 | 81 | 1 | |||||||
| 3303 | 06.07.23 | Skagerrak | 58°10.35179 | 9°25.58006 | 634 | 1 | 3 | 3 | 1 | 2 | 1 | 1 | |
| 3304 | 07.07.23 | Skagerrak | 58°08.83739 | 9°20.17345 | 627 | 1 | |||||||
| 3305 | 07.07.23 | Skagerrak | 58°14.9894 | 9°13.13094 | 484 | 1 | |||||||
| 3306 | 07.07.23 | Skagerrak | 58°15.49299 | 9°15.60666 | 631 | 1 | |||||||
| 3307 | 07.07.23 | Skagerrak | 58°18.89767 | 9°20.90007 | 585 | 1 | |||||||
| 3308 | 07.07.23 | Skagerrak | 58°23.41199 | 9°20.92952 | 409 | 1 | |||||||
| 3309 | 07.07.23 | Skagerrak | 58°24.532242 | 9°26.420425 | 515 | 1 | |||||||
| 3310 | 07.07.23 | Skagerrak | 58°22.170029 | 9°29.124068 | 589 | 1 | 2 | 1 | 1 | 2 | 1 | 1 | |
| 3311 | 07.07.23 | Skagerrak | 58°22.479686 | 9°33.369808 | 632 | 1 | |||||||
| 3312 | 07.07.23 | Skagerrak | 58°20.251902 | 9°30.057201 | 605 | 1 | |||||||
| 3313 | 08.07.23 | Skagerrak | 58°19.023727 | 9°36.409919 | 676 | 1 | |||||||
| 3314 | 08.07.23 | Skagerrak | 58°18.335919 | 9°28.385569 | 621 | 1 | |||||||
| 3315 | 08.07.23 | Skagerrak | 58°15.875413 | 9°26.511132 | 681 | 1 | |||||||
| 3316 | 08.07.23 | Skagerrak | 58°12.62526 | 9°25.686133 | 653 | 1 | |||||||
| 3317 | 08.07.23 | Skagerrak | 58°12.977754 | 9°27.873513 | 650 | 1 | |||||||
| 3318 | 08.07.23 | Skagerrak | 58°06.857311 | 9°39.245022 | 453 | 1 | |||||||
| 3319 | 08.07.23 | Skagerrak | 58°09.99994 | 9°43.17854 | 512 | 1 | |||||||
| 3320 | 08.07.23 | Skagerrak | 58°11.921971 | 9°38.612784 | 564 | 1 | |||||||
| 3321 | 08.07.23 | Skagerrak | 58°13.184619 | 9°42.004518 | 536 | 1 | |||||||
| 3322 | 09.07.23 | Skagerrak | 58°13.059518 | 9°38.450813 | 557 | 1 | |||||||
| 3323 | 09.07.23 | Skagerrak | 58°14.978199 | 9°37.944649 | 607 | 1 | |||||||
| 3324 | 09.07.23 | Skagerrak | 58°17.13081 | 9°38.712829 | 684 | 1 | |||||||
| 3325 | 09.07.23 | Skagerrak | 58°17.753328 | 9°43.854333 | 618 | 1 | |||||||
| 3326 | 09.07.23 | Skagerrak | 58°16.085843 | 9°45.62962 | 589 | 1 | |||||||
| 3327 | 09.07.23 | Skagerrak | 58°15.42668 | 9°48.7257 | 540 | 1 | |||||||
| 3328 | 09.07.23 | Skagerrak | 58°13.811588 | 9°54.224196 | 486 | 1 | 6 | 1 | 1 | 2 | 1 | 1 | |
| 3329 | 09.07.23 | Skagerrak | 58°18.16003 | 9°59.92819 | 481 | 1 | |||||||
| 3330 | 09.07.23 | Skagerrak | 58°18.78336 | 9°51.75012 | 555 | 1 | |||||||
| 3331 | 09.07.23 | Skagerrak | 58°19.47574 | 9°50.67573 | 564 | 1 | |||||||
| 3332 | 10.07.23 | Skagerrak | 58°23.692143 | 9°41.033516 | 677 | 1 | |||||||
| 3333 | 10.07.23 | Skagerrak | 58°26.424356 | 9°41.981323 | 697 | 1 | 3 | 1 | 1 | 2 | 1 | 1 | |
| 3334 | 11.07.23 | Skagerrak | 58°29.637485 | 9°43.782695 | 668 | 1 | |||||||
| 3335 | 11.07.23 | Skagerrak | 58°30.708558 | 9°50.828272 | 601 | 1 | |||||||
| 3336 | 11.07.23 | Skagerrak | 58°28.13451 | 9°51.462858 | 462 | 1 | |||||||
| 3337 | 11.07.23 | Skagerrak | 58°26.614528 | 9°47.862584 | 577 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3338 | 11.07.23 | Skagerrak | 58°26.55112 | 9°48.055867 | 540 | 1 | |||||||
| 3339 | 11.07.23 | Skagerrak | 58°25.94144 | 9°53.40633 | 470 | 1 | |||||||
| 3340 | 11.07.23 | Skagerrak | 58°24.52777 | 9°59.73072 | 533 | 1 | |||||||
| 3341 | 11.07.23 | Skagerrak | 58°20.634418 | 10°01.912713 | 488 | 1 | |||||||
| 3342 | 11.07.23 | Skagerrak | 58°24.92321 | 10°03.08338 | 515 | 1 | |||||||
| 3343 | 11.07.23 | Skagerrak | 58°29.10352 | 10°00.74711 | 546 | 1 | |||||||
| 3344 | 11.07.23 | Skagerrak | 58°30.034987 | 10°06.307699 | 494 | 1 | |||||||
| 3345 | 11.07.23 | Skagerrak | 58°28.028094 | 10°04.932561 | 537 | 1 | 1 | ||||||
| 3346 | 12.07.23 | KB-Skagerrak | 58°43.063085 | 9°44.258266 | 435 | 1 | |||||||
| 3347 | 12.07.23 | KB-Skagerrak | 58°40.34299 | 9°44.18271 | 430 | 1 | |||||||
| 3348 | 12.07.23 | KB-Skagerrak | 58°40.301646 | 9°41.143094 | 378 | 1 | |||||||
| 3349 | 12.07.23 | KB-Skagerrak | 58°39.63285 | 9°37.92247 | 403 | 1 | |||||||
| 3350 | 12.07.23 | KB-Skagerrak | 58°39.7641 | 9°32.58665 | 321 | 1 | |||||||
| 3351 | 12.07.23 | KB-Skagerrak | 58°41.886185 | 9°29.596325 | 250 | 1 | |||||||
| 3352 | 12.07.23 | KB-Skagerrak | 58°38.556445 | 9°25.411592 | 304 | 1 | |||||||
| 3353 | 12.07.23 | KB-Skagerrak | 58°40.572585 | 9°18.664026 | 101 | 1 | |||||||
| 3354 | 12.07.23 | KB-Skagerrak | 58°37.13153 | 9°18.81634 | 213 | 1 | |||||||
| 3355 | 12.07.23 | KB-Skagerrak | 58°35.75693 | 9°15.8684 | 204 | 1 | |||||||
| 3356 | 13.07.23 | KB-Skagerrak | 58°31.362788 | 9°04.471306 | 91 | 1 | |||||||
| 3357 | 13.07.23 | KB-Skagerrak | 58°31.116459 | 9°07.57078 | 189 | 1 | |||||||
| 3358 | 13.07.23 | KB-Skagerrak | 58°27.244015 | 9°11.864771 | 376 | 1 | |||||||
| 3359 | 13.07.23 | KB-Skagerrak | 58°25.595998 | 9°08.603032 | 427 | 1 | |||||||
| 3360 | 13.07.23 | KB-Skagerrak | 58°26.550391 | 9°05.950174 | 314 | 1 | |||||||
| 3361 | 13.07.23 | KB-Skagerrak | 58°26.929107 | 9°04.558511 | 354 | 1 | |||||||
| 3362 | 13.07.23 | KB-Skagerrak | 58°27.408589 | 9°03.354421 | 223 | 1 | |||||||
| 3363 | 13.07.23 | KB-Skagerrak | 58°25.360451 | 8°54.366172 | 87 | 1 | |||||||
| 3364 | 13.07.23 | KB-Skagerrak | 58°25.280037 | 8°59.655911 | 217 | 1 | |||||||
| 3365 | 14.07.23 | KB-Skagerrak | 58°22.62689 | 9°01.031184 | 315 | 9 | 2 | 1 | 2 | 1 | 1 | ||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| Toktnr 2023001014, FF G.O. Sars, Toktleder: Kyrre Heldal Kartveit | 0 | ||||||||||||
| 3284 | 08.10.23 | KB-Utsira | 59°05.736706 | 5°08.540245 | 169 | 2 | 1 | ||||||
| 3366 | 09.10.23 | NS-SK | 58°50.10611 | 9°41.552 | 138 | 1 | |||||||
| 3367 | 09.10.23 | NS-SK | 58°50.09027 | 9°46.69346 | 211 | 1 | |||||||
| 3368 | 10.10.23 | NS-SK | 58°51.345779 | 9°49.011186 | 257 | 1 | |||||||
| 3369 | 10.10.23 | NS-SK | 58°51.514426 | 9°51.783278 | 372 | 1 | |||||||
| 3370 | 11.10.23 | NS-SK | 59°15.113391 | 10°34.09767 | 167 | 4 | |||||||
| 3371 | 10.10.23 | NS-SK | 58°47.893754 | 10°12.30776 | 192 | 6 | 1 | 2 | 1 | ||||
| 3372 | 10.10.23 | NS-SK | 58°54.56722 | 9°52.46394 | 248 | 1 | |||||||
| 3373 | 10.10.23 | NS-SK | 58°56.91979 | 9°48.40499 | 240 | 1 | |||||||
| 3374 | 11.10.23 | NS-SK | 59°14.749583 | 10°33.893925 | 133 | 2 | |||||||
| 3375 | 11.10.23 | NS-SK | 59°14.577286 | 10°34.655381 | 139 | 1 | |||||||
| 3376 | 12.10.23 | NS-SK | 59°14.05902 | 10°34.97374 | 138 | 1 | |||||||
| 3377 | 13.10.23 | Utenfor Lista | 58°07.23843 | 6°28.8977 | 298 | 2 | |||||||
| 3378 | 14.10.23 | Hardanger | 59°42.076769 | 5°25.629575 | 172 | 1 | |||||||
| 3379 | 14.10.23 | Hardanger | 59°48.47785 | 5°35.54818 | 165 | 1 | |||||||
| 3380 | 14.10.23 | Hardanger | 59°56.96592 | 5°47.15349 | 414 | 1 | |||||||
| 3381 | 14.10.23 | Hardanger | 60°20.0598 | 6°15.75456 | 846 | 1 | |||||||
| Toktnr 202400703, FF Kronprins Haakon, Toktleder: Kyrre Heldal Kartveit | 0 | ||||||||||||
| 3382 | 08.03.24 | VestavindB | 60°52.423785 | 3°57.143619 | 326 | 1 | 1 | ||||||
| 3383 | 09.03.24 | VestavindB | 60°53.350612 | 3°58.643311 | 326 | 1 | |||||||
| 3384 | 09.03.24 | VestavindB | 60°56.312829 | 3°55.521695 | 335 | 1 | |||||||
| 3385 | 09.03.24 | VestavindB | 60°57.540835 | 3°56.214697 | 336 | 1 | |||||||
| 3386 | 09.03.24 | VestavindB | 60°57.876715 | 3°44.724012 | 342 | 1 | |||||||
| 3387 | 09.03.24 | VestavindB | 60°53.812345 | 3°49.948928 | 339 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3388 | 09.03.24 | VestavindB | 60°54.146621 | 3°44.979863 | 342 | 1 | 7 | 1 | 2 | 1 | 1 | ||
| 3389 | 10.03.24 | VestavindB | 60°48.569084 | 3°54.681848 | 328 | 1 | |||||||
| 3390 | 10.03.24 | VestavindB | 60°47.859152 | 3°42.94124 | 335 | 1 | |||||||
| 3391 | 10.03.24 | VestavindB | 60°47.876483 | 3°31.886598 | 329 | 1 | |||||||
| 3392 | 10.03.24 | VestavindB | 60°50.502846 | 3°20.667798 | 341 | 1 | |||||||
| 3393 | 10.03.24 | VestavindB | 60°55.151422 | 3°19.88734 | 345 | 1 | 5 | 1 | 1 | 2 | 1 | 1 | |
| 3394 | 10.03.24 | VestavindB | 60°59.407927 | 3°28.37223 | 338 | 1 | |||||||
| 3395 | 11.03.24 | VestavindB | 60°58.872972 | 3°08.469554 | 360 | 1 | |||||||
| 3396 | 11.03.24 | VestavindB | 60°56.709745 | 3°11.507793 | 365 | 1 | |||||||
| 3397 | 11.03.24 | VestavindB | 60°54.581673 | 3°12.528017 | 352 | 1 | |||||||
| 3398 | 11.03.24 | VestavindB | 60°56.736919 | 3°16.986208 | 349 | 1 | |||||||
| 3399 | 11.03.24 | VestavindB | 60°57.102824 | 3°21.605766 | 348 | 1 | |||||||
| 3400 | 11.03.24 | VestavindB | 60°40.797175 | 3°29.682297 | 318 | 1 | |||||||
| 3401 | 11.03.24 | VestavindB | 60°43.249314 | 3°35.839237 | 314 | 1 | |||||||
| 3402 | 11.03.24 | VestavindB | 60°54.581688 | 3°12.52782 | 346 | 1 | |||||||
| 3403 | 11.03.24 | VestavindB | 60°44.222926 | 3°47.664576 | 332 | 1 | |||||||
| 3404 | 11.03.24 | VestavindB | 60°44.186633 | 3°54.850995 | 319 | 1 | 7 | 1 | 2 | 1 | 1 | ||
| 3405 | 13.03.24 | NSJ-2 | 56°30.144641 | 4°36.392866 | 64 | 1 | 1 | ||||||
| 3406 | 13.03.24 | NSJ-2 | 56°31.264705 | 4°39.91178 | 63 | 1 | |||||||
| 3407 | 13.03.24 | NSJ-2 | 56°33.666487 | 4°42.713108 | 64 | 1 | |||||||
| 3408 | 13.03.24 | NSJ-2 | 56°35.872658 | 4°36.546395 | 63 | 1 | |||||||
| 3409 | 13.03.24 | NSJ-2 | 56°34.805474 | 4°42.236769 | 63 | 1 | |||||||
| 3410 | 13.03.24 | NSJ-2 | 56°36.93947 | 4°41.7044 | 62 | 1 | |||||||
| 3411 | 14.03.24 | NSJ-2 | 56°35.503895 | 4°45.087102 | 63 | 1 | |||||||
| 3412 | 14.03.24 | NSJ-2 | 56°39.624673 | 4°54.193576 | 63 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3413 | 14.03.24 | NSJ-2 | 56°40.859749 | 4°57.379758 | 63 | 1 | |||||||
| 3414 | 15.03.24 | NSJ-2 | 56°37.026906 | 4°54.630751 | 64 | 1 | 5 | 4 | 1 | 2 | 1 | ||
| 3415 | 14.03.24 | NSJ-2 | 56°39.308982 | 5°07.553393 | 63 | 1 | |||||||
| 3416 | 14.03.24 | NSJ-2 | 56°37.949777 | 5°06.811422 | 64 | 1 | |||||||
| 3417 | 14.03.24 | NSJ-2 | 56°40.199975 | 5°14.581121 | 61 | 1 | |||||||
| 3418 | 14.03.24 | NSJ-2 | 56°42.371089 | 5°17.142056 | 60 | 1 | |||||||
| 3419 | 14.03.24 | NSJ-2 | 56°42.440704 | 5°17.756362 | 60 | 1 | |||||||
| 3420 | 14.03.24 | NSJ-2 | 56°44.008177 | 5°24.493638 | 57 | 1 | |||||||
| 3421 | 15.03.24 | NSJ-2 | 56°45.697963 | 5°27.314717 | 55 | 1 | |||||||
| 3422 | 15.03.24 | NSJ-2 | 56°45.977881 | 5°24.495069 | 54 | 1 | |||||||
| 3423 | 15.03.24 | NSJ-2 | 56°44.932366 | 5°21.224065 | 56 | 1 | |||||||
| 3424 | 15.03.24 | NSJ-2 | 56°45.605614 | 5°19.476206 | 60 | 1 | |||||||
| 3425 | 15.03.24 | NSJ-2 | 56°44.008716 | 5°11.984753 | 61 | 1 | 5 | 1 | 1 | 2 | 1 | 1 | |
| 3426 | 15.03.24 | NSJ-2 | 56°49.047103 | 5°13.955578 | 58 | 1 | |||||||
| 3427 | 15.03.24 | NSJ-2 | 56°49.025359 | 5°19.581646 | 57 | 1 | |||||||
| 3428 | 16.03.24 | NSJ-2 | 56°48.783793 | 5°23.268918 | 54 | 1 | |||||||
| 3429 | 16.03.24 | NSJ-2 | 56°51.011427 | 5°26.714002 | 52 | 1 | |||||||
| 3430 | 16.03.24 | NSJ-2 | 56°53.785226 | 5°32.406736 | 50 | 1 | |||||||
| 3431 | 16.03.24 | NSJ-2 | 56°56.885714 | 5°46.497189 | 53 | 1 | |||||||
| 3432 | 16.03.24 | NSJ-2 | 56°58.865514 | 5°52.641569 | 57 | 1 | |||||||
| 3433 | 16.03.24 | NSJ-2 | 56°59.940497 | 5°58.334262 | 51 | 1 | |||||||
| 3434 | 16.03.24 | NSJ-2 | 57°01.213361 | 6°03.256772 | 20 | 1 | |||||||
| 3435 | 17.03.24 | NSJ-2 | 56°56.507343 | 5°42.435547 | 54 | 1 | 6 | 1 | 1 | 2 | 1 | ||
| 3436 | 17.03.24 | NSJ-2 | 56°53.616489 | 5°19.014269 | 54 | 1 | |||||||
| 3437 | 17.03.24 | NSJ-2 | 56°54.084452 | 5°18.089622 | 55 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3438 | 17.03.24 | NSJ-2 | 56°54.615683 | 5°05.967131 | 59 | 1 | |||||||
| 3439 | 17.03.24 | NSJ-2 | 56°55.472814 | 5°01.805005 | 61 | 1 | |||||||
| 3440 | 17.03.24 | NSJ-2 | 56°57.338373 | 4°57.707383 | 61 | 1 | |||||||
| 3441 | 17.03.24 | NSJ-2 | 56°59.049541 | 4°55.931157 | 61 | 1 | 6 | 1 | |||||
| 3442 | 17.03.24 | NSJ-2 | 56°53.159184 | 4°59.706615 | 64 | 1 | |||||||
| 3443 | 17.03.24 | NSJ-2 | 56°52.535335 | 5°07.033735 | 59 | 1 | |||||||
| 3444 | 18.03.24 | NSJ-2 | 56°49.955579 | 5°09.644712 | 62 | 1 | |||||||
| 3445 | 18.03.24 | NSJ-2 | 56°52.741388 | 5°10.606777 | 59 | 1 | |||||||
| 3446 | 18.03.24 | NSJ-2 | 56°52.409559 | 5°13.991813 | 56 | 1 | |||||||
| 3447 | 18.03.24 | NSJ-2 | 56°51.762304 | 5°23.570035 | 53 | 1 | |||||||
| 3448 | 18.03.24 | NSJ-2 | 56°54.384106 | 5°24.348117 | 52 | 1 | |||||||
| 3449 | 18.03.24 | NSJ-2 | 56°55.67551 | 5°30.272499 | 52 | 1 | |||||||
| 3450 | 18.03.24 | NSJ-2 | 56°59.316479 | 5°37.324202 | 53 | 1 | |||||||
| 3451 | 18.03.24 | NSJ-2 | 56°59.072368 | 5°40.415524 | 55 | 1 | |||||||
| 3452 | 18.03.24 | NSJ-2 | 57°02.763235 | 5°51.742503 | 52 | 1 | |||||||
| 3453 | 18.03.24 | NSJ-2 | 57°03.175026 | 5°58.124452 | 51 | 1 | |||||||
| 3454 | 18.03.24 | NSJ-2 | 57°04.236418 | 5°58.074601 | 42 | 1 | |||||||
| 3455 | 18.03.24 | NSJ-2 | 57°02.311072 | 6°06.12372 | 50 | 1 | |||||||
| 3456 | 18.03.24 | NSJ-2 | 57°06.409453 | 6°06.429411 | 49 | 1 | |||||||
| 3457 | 18.03.24 | NSJ-2 | 57°05.853332 | 6°11.86867 | 50 | 1 | |||||||
| 3458 | 18.03.24 | NSJ-2 | 57°07.374348 | 6°18.5352 | 58 | 1 | |||||||
| 3459 | 18.03.24 | NSJ-2 | 57°08.927587 | 6°26.505522 | 66 | 1 | |||||||
| 3460 | 19.03.24 | NSJ-2 | 57°09.234384 | 6°28.757502 | 124 | 1 | |||||||
| 3461 | 19.03.24 | NSJ-2 | 57°12.459197 | 6°32.644633 | 68 | 1 | |||||||
| 3462 | 19.03.24 | NSJ-2 | 57°14.101144 | 6°36.902901 | 70 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3463 | 19.03.24 | NSJ-2 | 57°15.233039 | 6°31.867434 | 71 | 1 | |||||||
| 3464 | 19.03.24 | NSJ-2 | 57°16.295096 | 6°36.468692 | 74 | 1 | |||||||
| 3465 | 19.03.24 | NSJ-2 | 57°18.529665 | 6°41.486631 | 81 | 1 | |||||||
| Toktnr 2024001021, FF G.O. Sars, Toktleder: Hilde Elise Heldal | 0 | ||||||||||||
| 3425 | 01.12.24 | NSJ-2 | 56°43.92507 | 5°11.88571 | 64 | 1 | 5 | 1 | 1 | 2 | 1 | 1 | |
| 3555 | 22.11.24 | Vestavind-F | 59°29.419944 | 4°30.996271 | 266 | 1 | |||||||
| 3556 | 22.11.24 | Vestavind-F | 59°29.605781 | 4°18.388366 | 267 | 1 | |||||||
| 3557 | 22.11.24 | Vestavind-F | 59°25.69029 | 4°16.28671 | 260 | 1 | |||||||
| 3558 | 22.11.24 | Vestavind-F | 59°24.974125 | 4°23.215686 | 261 | 1 | |||||||
| 3559 | 22.11.24 | Vestavind-F | 59°25.606434 | 4°27.067908 | 261 | 1 | |||||||
| 3560 | 22.11.24 | Vestavind-F | 59°21.788068 | 4°14.538856 | 272 | 1 | 5 | 1 | 1 | 2 | 1 | 1 | |
| 3561 | 23.11.24 | Vestavind-F | 59°19.164007 | 4°20.242192 | 266 | 1 | |||||||
| 3562 | 23.11.24 | Vestavind-F | 59°21.748854 | 4°25.799291 | 263 | 1 | |||||||
| 3563 | 23.11.24 | Vestavind-F | 59°16.180515 | 4°26.191201 | 260 | 1 | |||||||
| 3564 | 23.11.24 | Vestavind-F | 59°09.40123 | 4°30.50598 | 249 | 1 | |||||||
| 3565 | 23.11.24 | Vestavind-F | 59°05.417142 | 4°28.885376 | 257 | 1 | 6 | 1 | 1 | 1 | 1 | ||
| 3566 | 25.11.24 | Boknafjorden | 59°15.503879 | 5°41.184034 | 265 | 1 | |||||||
| 3567 | 25.11.24 | Boknafjorden | 59°23.121324 | 6°02.385884 | 62 | 1 | 1 | ||||||
| 3568 | 26.11.24 | Boknafjorden | 59°16.97363 | 5°50.04657 | 130 | 3 | |||||||
| 3569 | 26.11.24 | Boknafjorden | 59°15.048712 | 5°39.531297 | 244 | 1 | |||||||
| 3570 | 26.11.24 | Boknafjorden | 59°15.618912 | 5°40.915956 | 235 | 1 | |||||||
| 3571 | 27.11.24 | Vestavind-F | 59°07.729427 | 4°13.597587 | 279 | 1 | |||||||
| 3572 | 27.11.24 | Vestavind-F | 59°03.01932 | 4°18.38398 | 261 | 1 | |||||||
| 3573 | 27.11.24 | Vestavind-F | 59°00.119175 | 4°18.235404 | 270 | 1 | |||||||
| 3574 | 27.11.24 | Vestavind-F | 59°04.696602 | 4°22.794213 | 259 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3575 | 27.11.24 | Vestavind-F | 59°03.35135 | 4°34.90869 | 255 | 1 | |||||||
| 3576 | 27.11.24 | Vestavind-F | 59°00.548669 | 4°44.055421 | 271 | 1 | |||||||
| 3577 | 27.11.24 | Vestavind-F | 59°00.437713 | 4°46.431825 | 274 | 1 | |||||||
| 3578 | 28.11.24 | Sørvest-F | 57°12.715586 | 4°30.229773 | 63 | 1 | 1 | ||||||
| 3579 | 28.11.24 | Sørvest-F | 57°08.46851 | 4°31.76798 | 58 | 1 | 8 | 1 | 2 | 1 | 1 | ||
| 3580 | 28.11.24 | Sørvest-F | 57°05.315512 | 4°43.131777 | 63 | 1 | 1 | ||||||
| 3581 | 28.11.24 | Sørvest-F | 57°06.89339 | 4°36.407125 | 63 | 1 | 1 | ||||||
| 3582 | 28.11.24 | Sørvest-F | 57°05.640072 | 4°29.5385 | 62 | 1 | 1 | ||||||
| 3583 | 28.11.24 | Sørvest-F | 57°02.474657 | 4°29.875754 | 66 | 1 | 1 | ||||||
| 3584 | 29.11.24 | Sørvest-F | 56°59.415908 | 4°30.32477 | 65 | 1 | 1 | ||||||
| 3585 | 29.11.24 | Sørvest-F | 56°58.085986 | 4°32.4272 | 65 | 1 | 1 | ||||||
| 3586 | 29.11.24 | Sørvest-F | 56°57.117989 | 4°39.99021 | 66 | 1 | |||||||
| 3587 | 29.11.24 | Sørvest-F | 56°59.678537 | 4°42.13237 | 66 | 1 | 1 | ||||||
| 3588 | 29.11.24 | Sørvest-F | 57°02.115415 | 4°46.960015 | 65 | 1 | 1 | ||||||
| 3589 | 29.11.24 | Sørvest-F | 56°52.68573 | 4°53.130981 | 66 | 1 | 6 | 1 | 2 | 2 | 1 | ||
| 3590 | 29.11.24 | Sørvest-F | 56°45.414816 | 4°41.94344 | 61 | 1 | 7 | 1 | 2 | 2 | 1 | ||
| 3591 | 30.11.24 | Sørvest-F | 56°47.615683 | 4°29.614054 | 60 | 1 | 6 | 1 | 2 | 2 | 1 | ||
| 3592 | 30.11.24 | Sørvest-F | 56°47.042719 | 4°33.728253 | 61 | 1 | 1 | ||||||
| 3593 | 30.11.24 | Sørvest-F | 56°52.145394 | 4°33.887811 | 64 | 1 | 1 | ||||||
| 3594 | 30.11.24 | Sørvest-F | 56°52.447318 | 4°42.95973 | 65 | 1 | |||||||
| 3595 | 01.12.24 | Sørvest-F | 56°43.032386 | 4°37.139097 | 60 | 1 | 1 | ||||||
| 3596 | 01.12.24 | Sørvest-F | 56°38.56386 | 4°33.82458 | 63 | 1 | 1 | ||||||
| 3597 | 01.12.24 | Sørvest-F | 56°44.41794 | 4°54.46375 | 61 | 1 | 1 | ||||||
| 3598 | 01.12.24 | Sørvest-F | 56°45.710137 | 5°02.256471 | 62 | 1 | 1 | ||||||
| 3599 | 01.12.24 | NSJ-1 | 56°59.559151 | 5°15.420209 | 56 | 1 | 5 | 1 | 1 | 2 | 2 | 1 | |
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3600 | 01.12.24 | Sørvest-F | 57°50.647998 | 5°52.637683 | 224 | 1 | 6 | 3 | 1 | 2 | 1 | 1 | |
| 3601 | 03.12.24 | KB Jæren | 58°04.976295 | 6°28.718862 | 368 | 1 | 8 | 1 | 2 | 1 | 1 | ||
| 3602 | 02.12.24 | KB Jæren | 58°04.03598 | 6°27.69674 | 359 | 1 | |||||||
| 3603 | 02.12.24 | KB Jæren | 58°03.221626 | 6°29.899901 | 367 | 1 | |||||||
| 3604 | 02.12.24 | KB Jæren | 58°01.037121 | 6°44.118045 | 104 | 1 | |||||||
| 3605 | 02.12.24 | Fedafjorden | 58°15.48966 | 6°50.22603 | 65 | 1 | 1 | ||||||
| 3606 | 03.12.24 | KB Jæren | 58°10.449743 | 6°21.114221 | 325 | 1 | |||||||
| 3607 | 03.12.24 | KB Jæren | 58°05.442384 | 6°31.949649 | 327 | 1 | |||||||
| Toktnr 2025001009, FF G.O. Sars, Toktleder: Genoveva Gonzalez-Mirelis | 0 | ||||||||||||
| 3589 | 21.06.25 | Sørvest-F | 56°52.897876 | 4°52.785883 | 65 | 1 | 6 | 1 | 2 | 2 | 1 | ||
| 3591 | 21.06.25 | Sørvest-F | 56°47.638284 | 4°29.598947 | 59 | 1 | 6 | 1 | 2 | 2 | 1 | ||
| 3599 | 23.06.25 | Sørvest-F | 56°59.472293 | 5°15.359009 | 55 | 1 | 5 | 1 | 1 | 2 | 2 | 1 | |
| 3600 | 26.06.25 | NS3-B1 | 57°50.678858 | 5°52.438442 | 223 | 1 | 6 | 3 | 1 | 2 | 1 | 1 | |
| 3608 | 20.06.25 | NS_Gas flare extra station | 57°11.584571 | 4°57.570339 | 62 | 2 | |||||||
| 3609 | 20.06.25 | Sørvest-F | 56°36.510681 | 4°29.561387 | 64 | 1 | |||||||
| 3610 | 20.06.25 | Sørvest-F | 56°36.71328 | 4°31.75853 | 60 | 1 | |||||||
| 3611 | 20.06.25 | Sørvest-F | 56°40.71162 | 4°29.78596 | 52 | 1 | |||||||
| 3612 | 20.06.25 | Sørvest-F | 56°41.04648 | 4°27.384 | 48 | 1 | |||||||
| 3613 | 20.06.25 | Sørvest-F | 56°43.63312 | 4°28.02693 | 50 | 1 | |||||||
| 3614 | 21.06.25 | Sørvest-F | 56°45.86928 | 4°28.02578 | 53 | 1 | |||||||
| 3615 | 21.06.25 | Sørvest-F | 56°41.009622 | 4°33.249397 | 59 | 1 | |||||||
| 3616 | 21.06.25 | Sørvest-F | 56°46.419238 | 4°35.684676 | 59 | 1 | |||||||
| 3617 | 21.06.25 | Sørvest-F | 56°39.475406 | 4°41.413189 | 60 | 1 | |||||||
| 3618 | 21.06.25 | Sørvest-F | 56°54.67373 | 4°37.55152 | 60 | 1 | |||||||
| 3619 | 21.06.25 | Sørvest-F | 56°49.091193 | 4°41.818894 | 62 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3620 | 21.06.25 | Sørvest-F | 56°41.766705 | 4°47.419045 | 60 | 1 | |||||||
| 3621 | 21.06.25 | Sørvest-F | 56°40.892685 | 4°50.122939 | 60 | 1 | |||||||
| 3622 | 21.06.25 | Sørvest-F | 56°42.186755 | 4°52.885838 | 60 | 1 | |||||||
| 3623 | 21.06.25 | Sørvest-F | 56°44.74701 | 5°00.03641 | 57 | 1 | |||||||
| 3624 | 21.06.25 | Sørvest-F | 56°49.31857 | 4°54.41952 | 59 | 1 | |||||||
| 3625 | 21.06.25 | Sørvest-F | 57°02.033927 | 4°49.111809 | 64 | 1 | |||||||
| 3626 | 22.06.25 | Sørvest-F | 57°00.985362 | 4°49.844194 | 63 | 1 | |||||||
| 3627 | 22.06.25 | Sørvest-F | 56°56.273346 | 4°54.58905 | 66 | 1 | |||||||
| 3628 | 22.06.25 | Sørvest-F | 56°53.08938 | 4°57.81326 | 61 | 1 | |||||||
| 3629 | 22.06.25 | Sørvest-F | 56°47.18603 | 5°02.16432 | 58 | 1 | |||||||
| 3630 | 22.06.25 | Sørvest-F | 56°49.470246 | 4°59.506659 | 62 | 1 | |||||||
| 3631 | 22.06.25 | Sørvest-F | 56°56.186723 | 5°02.001123 | 67 | 1 | |||||||
| 3632 | 22.06.25 | Sørvest-F | 56°56.019853 | 5°04.29399 | 60 | 1 | |||||||
| 3633 | 22.06.25 | Sørvest-F | 56°56.945681 | 5°03.066243 | 60 | 1 | |||||||
| 3634 | 22.06.25 | NSJ-1 | 56°58.3802 | 5°02.606622 | 61 | 1 | |||||||
| 3635 | 22.06.25 | NSJ-1 | 57°02.724148 | 5°01.865136 | 56 | 1 | |||||||
| 3636 | 22.06.25 | NSJ-1 | 56°59.31466 | 5°02.92687 | 55 | 1 | |||||||
| 3637 | 22.06.25 | NSJ-1 | 56°57.796797 | 5°03.784571 | 60 | 1 | |||||||
| 3638 | 22.06.25 | NSJ-1 | 56°57.324193 | 5°06.293945 | 61 | 1 | |||||||
| 3639 | 22.06.25 | NSJ-1 | 56°57.779317 | 5°07.344685 | 59 | 1 | |||||||
| 3640 | 22.06.25 | NSJ-1 | 56°56.556303 | 5°09.450429 | 56 | 1 | |||||||
| 3641 | 22.06.25 | NSJ-1 | 56°57.983667 | 5°09.445779 | 57 | 1 | |||||||
| 3642 | 22.06.25 | NSJ-1 | 56°56.479112 | 5°12.365597 | 55 | 1 | |||||||
| 3643 | 22.06.25 | NSJ-1 | 56°58.079966 | 5°12.027288 | 59 | 1 | |||||||
| 3644 | 23.06.25 | NSJ-1 | 57°01.84858 | 5°10.384313 | 54 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3645 | 23.06.25 | NSJ-1 | 57°04.473502 | 5°07.405909 | 61 | 1 | |||||||
| 3646 | 23.06.25 | Sørvest-F | 57°04.74094 | 5°09.92806 | 59 | 1 | |||||||
| 3647 | 23.06.25 | Sørvest-F | 57°03.141245 | 5°10.545032 | 57 | 1 | |||||||
| 3648 | 23.06.25 | Sørvest-F | 56°59.185893 | 5°12.236651 | 59 | 1 | |||||||
| 3649 | 23.06.25 | Sørvest-F | 57°00.091963 | 5°13.194679 | 58 | 1 | |||||||
| 3650 | 23.06.25 | Sørvest-F | 57°01.041129 | 5°13.654155 | 58 | 1 | |||||||
| 3651 | 23.06.25 | NSJ-1 | 57°08.903464 | 5°16.846734 | 54 | 1 | |||||||
| 3652 | 23.06.25 | NSJ-1 | 57°08.633211 | 5°21.530078 | 55 | 1 | |||||||
| 3653 | 23.06.25 | NSJ-1 | 56°59.991638 | 5°22.957895 | 53 | 1 | |||||||
| 3654 | 23.06.25 | NSJ-1 | 57°04.577953 | 5°33.410782 | 52 | 1 | |||||||
| 3655 | 23.06.25 | NSJ-1 | 57°06.026345 | 5°36.911226 | 47 | 1 | |||||||
| 3656 | 23.06.25 | NSJ-1 | 57°10.752066 | 5°38.925398 | 0 | 1 | |||||||
| 3657 | 24.06.25 | NSJ-1 | 57°09.922854 | 5°42.921986 | 53 | 1 | |||||||
| 3658 | 24.06.25 | NSJ-1 | 57°12.040754 | 5°43.927348 | 56 | 1 | |||||||
| 3659 | 24.06.25 | NSJ-1 | 57°13.664146 | 5°43.794098 | 59 | 1 | |||||||
| 3660 | 24.06.25 | NSJ-1 | 57°20.575069 | 5°59.038356 | 77 | 2 | |||||||
| 3661 | 24.06.25 | NSJ-1 | 57°28.076228 | 6°02.369604 | 84 | 1 | |||||||
| 3662 | 24.06.25 | NS3-B1 | 57°36.910128 | 6°21.085097 | 153 | 2 | |||||||
| 3663 | 24.06.25 | NS3-B1 | 57°45.338582 | 6°19.825203 | 281 | 1 | 7 | 1 | 1 | 2 | 1 | 1 | |
| 3664 | 25.06.25 | NS3-B1 | 57°55.764078 | 5°51.027069 | 257 | 1 | 5 | 4 | 1 | 2 | 1 | 1 | |
| 3665 | 25.06.25 | NS3-B1 | 58°09.052377 | 5°20.187543 | 291 | 1 | 5 | 1 | 1 | 2 | 1 | 1 | |
| 3666 | 25.06.25 | NS3-B1 | 58°15.419037 | 4°51.540492 | 303 | 1 | 7 | 3 | 1 | 2 | 1 | 1 | |
| 3667 | 26.06.25 | NS3-B1 | 58°19.818593 | 4°48.394939 | 294 | 1 | 7 | 1 | 1 | 2 | 1 | 1 | |
| 3668 | 27.06.25 | NS3-B1 | 58°16.82713 | 6°01.17341 | 276 | 1 | |||||||
| 3669 | 27.06.25 | NS3-B1 | 57°40.495068 | 6°24.106151 | 199 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3670 | 28.06.25 | NS3-B1 | 57°41.9528 | 6°23.38579 | 299 | 1 | |||||||
| 3671 | 28.06.25 | NS3-B1 | 57°40.945812 | 6°22.427166 | 231 | 1 | |||||||
| 3672 | 28.06.25 | NS3-B1 | 57°41.058982 | 6°20.552317 | 248 | 2 | |||||||
| 3673 | 28.06.25 | NS3-B1 | 57°38.79344 | 6°19.68282 | 151 | 1 | |||||||
| 3674 | 28.06.25 | NS3-B1 | 57°37.13909 | 6°15.51676 | 151 | 1 | |||||||
| 3675 | 28.06.25 | NS3-B1 | 57°40.62742 | 6°14.2587 | 193 | 1 | |||||||
| 3676 | 28.06.25 | NS3-B1 | 57°41.627725 | 6°16.730053 | 268 | 1 | |||||||
| 3677 | 28.06.25 | NS3-B1 | 57°49.993256 | 6°14.772332 | 288 | 1 | |||||||
| 3678 | 28.06.25 | NS3-B1 | 57°44.43015 | 6°10.63612 | 261 | 1 | |||||||
| 3679 | 28.06.25 | NS3-B1 | 57°46.983186 | 6°08.402359 | 261 | 1 | |||||||
| 3680 | 28.06.25 | NS3-B1 | 57°44.683509 | 6°04.187538 | 218 | 1 | |||||||
| 3681 | 28.06.25 | NS3-B1 | 57°40.91721 | 6°03.60099 | 150 | 1 | |||||||
| 3682 | 28.06.25 | NS3-B1 | 57°44.229388 | 6°00.228242 | 182 | 1 | |||||||
| 3683 | 28.06.25 | NS3-B1 | 57°45.883533 | 6°02.219375 | 252 | 1 | |||||||
| 3684 | 29.06.25 | NS3-B1 | 57°51.108901 | 6°00.838474 | 257 | 1 | |||||||
| 3685 | 29.06.25 | NS3-B1 | 57°54.53734 | 6°09.96364 | 295 | 1 | |||||||
| 3686 | 29.06.25 | NS3-B1 | 57°56.327399 | 6°05.661047 | 300 | 1 | |||||||
| 3687 | 29.06.25 | NS3-B1 | 57°57.04975 | 5°54.65953 | 273 | 1 | |||||||
| 3688 | 29.06.25 | NS3-B1 | 57°54.85436 | 5°57.03916 | 264 | 1 | |||||||
| 3689 | 29.06.25 | NS3-B1 | 57°48.456778 | 5°51.431323 | 211 | 1 | |||||||
| 3690 | 29.06.25 | NS3-B1 | 57°46.03378 | 5°49.50183 | 150 | 1 | |||||||
| 3691 | 29.06.25 | NS3-B1 | 57°51.450856 | 5°43.089697 | 193 | 1 | |||||||
| 3692 | 30.06.25 | NS3-B1 | 57°59.26018 | 5°44.25597 | 261 | 1 | |||||||
| 3693 | 30.06.25 | NS3-B1 | 58°03.67564 | 5°40.90591 | 282 | 1 | |||||||
| 3694 | 30.06.25 | NS3-B1 | 57°55.71262 | 5°33.65274 | 189 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3695 | 30.06.25 | NS3-B1 | 57°57.978135 | 5°31.592925 | 213 | 1 | |||||||
| 3696 | 30.06.25 | NS3-B1 | 57°58.87913 | 5°30.76364 | 215 | 1 | |||||||
| 3697 | 30.06.25 | NS3-B1 | 57°56.31363 | 5°25.43413 | 162 | 1 | |||||||
| 3698 | 30.06.25 | NS3-B1 | 58°00.492954 | 5°25.098806 | 217 | 1 | |||||||
| 3699 | 30.06.25 | NS3-B1 | 58°01.07011 | 5°24.0891 | 218 | 1 | |||||||
| 3700 | 30.06.25 | NS3-B1 | 58°03.22857 | 5°25.92527 | 263 | 1 | |||||||
| 3701 | 30.06.25 | NS3-B1 | 58°04.021198 | 5°21.709849 | 263 | 1 | |||||||
| 3702 | 30.06.25 | NS3-B1 | 58°05.7381 | 5°19.88766 | 276 | 1 | |||||||
| 3703 | 30.06.25 | NS3-B1 | 58°07.553958 | 5°14.560251 | 284 | 1 | |||||||
| 3704 | 30.06.25 | NS3-B1 | 58°06.845878 | 5°12.875663 | 272 | 1 | |||||||
| 3705 | 30.06.25 | NS3-B1 | 58°05.123224 | 5°13.099315 | 248 | 1 | |||||||
| 3706 | 30.06.25 | NS3-B1 | 58°01.80529 | 5°13.6931 | 186 | 1 | |||||||
| 3707 | 30.06.25 | NS3-B1 | 58°01.367464 | 5°08.552845 | 161 | 1 | |||||||
| 3708 | 30.06.25 | NS3-B1 | 58°05.811029 | 5°09.285527 | 249 | 1 | |||||||
| 3709 | 30.06.25 | NS3-B1 | 58°10.73274 | 5°12.46762 | 300 | 1 | |||||||
| 3710 | 01.07.25 | NS3-B1 | 58°11.29166 | 5°06.088769 | 298 | 1 | |||||||
| 3711 | 01.07.25 | NS3-B1 | 58°06.62261 | 5°05.86613 | 248 | 1 | |||||||
| 3712 | 01.07.25 | NS3-B1 | 58°04.069187 | 5°04.382033 | 193 | 1 | |||||||
| 3713 | 01.07.25 | NS3-B1 | 58°02.86497 | 5°01.37486 | 153 | 1 | |||||||
| 3714 | 01.07.25 | NS3-B1 | 58°04.717111 | 5°03.236201 | 201 | 2 | |||||||
| 3715 | 01.07.25 | NS3-B1 | 58°10.220052 | 5°02.705792 | 279 | 1 | |||||||
| 3716 | 01.07.25 | NS3-B1 | 58°07.176218 | 4°57.387676 | 228 | 1 | |||||||
| 3717 | 01.07.25 | NS3-B1 | 58°03.94657 | 4°56.72764 | 155 | 1 | |||||||
| 3718 | 01.07.25 | NS3-B1 | 58°07.66681 | 4°48.06411 | 195 | 1 | |||||||
| 3719 | 01.07.25 | NS3-B1 | 58°09.26853 | 4°52.74893 | 248 | 1 | |||||||
| St nr | Dato | Område | Latitude | Longitude | Dyp | Video | Grabb | Boxcorer | Bomtrål | RP-slede | Multicorer | CTD | Gr corer |
| 3720 | 02.07.25 | NS3-B1 | 58°12.35897 | 5°20.43271 | 299 | 1 | |||||||
| 3721 | 02.07.25 | NS3-B1 | 58°15.532896 | 5°07.985426 | 308 | 1 | |||||||
| 3722 | 02.07.25 | NS3-B1 | 58°17.296993 | 4°59.98915 | 304 | 1 | |||||||
| 3723 | 02.07.25 | NS3-B1 | 58°15.040854 | 4°57.856422 | 304 | 1 | |||||||
| 3724 | 02.07.25 | NS3-B1 | 58°18.153332 | 4°42.118149 | 297 | 1 | |||||||
| 3725 | 02.07.25 | NS3-B1 | 58°12.764682 | 4°47.417662 | 285 | 1 | |||||||
| 3726 | 02.07.25 | NS3-B1 | 58°11.878238 | 4°45.941206 | 269 | 1 | |||||||
| 3727 | 02.07.25 | NS3-B1 | 58°11.515424 | 4°37.922045 | 230 | 1 | |||||||
| 3728 | 02.07.25 | NS3-B1 | 58°07.54421 | 4°39.26886 | 156 | 1 | |||||||
| 3729 | 02.07.25 | NS3-B1 | 58°08.660365 | 4°33.260653 | 151 | 1 | |||||||
| 3730 | 02.07.25 | NS3-B1 | 58°10.650291 | 4°27.978532 | 150 | 1 | |||||||
| 3731 | 02.07.25 | NS3-B1 | 58°20.414435 | 4°56.157422 | 298 | 1 | |||||||
| 3732 | 02.07.25 | NS3-B1 | 58°24.443736 | 4°57.020081 | 294 | 1 | |||||||
| 3733 | 02.07.25 | NS3-B1 | 58°20.955505 | 5°03.877848 | 308 | 1 | |||||||
| 3734 | 02.07.25 | NS3-B1 | 58°23.692092 | 5°10.545498 | 314 | 1 | |||||||
| 3735 | 02.07.25 | NS3-B1 | 58°25.661271 | 5°16.983504 | 316 | 1 | |||||||
| 3736 | 03.07.25 | NS3-B1 | 58°22.242866 | 5°19.264648 | 326 | 1 | |||||||
| 3737 | 03.07.25 | NS3-B1 | 58°21.794409 | 5°24.516729 | 330 | 1 | |||||||
| 3738 | 03.07.25 | NS3-B1 | 58°20.576473 | 5°32.782692 | 336 | 1 | |||||||
| 3739 | 03.07.25 | NS3-B1 | 58°18.284956 | 5°30.92951 | 329 | 1 | |||||||