Dette kapitlet rapporterer de første stegene i utviklingen av en rettsmedisinsk verktøykasse for å avgjøre om atlantisk laks ble korrekt bedøvd og bløgget i henhold til norske forskrifter, eller om fisken allerede var død før prosessering. Å oppdage selvdød fisk er avgjørende for matsikkerhet og for å forebygge svindel.
To eksperimenter undersøkte postmortale endringer hos laks lagret i sjøvann ved 5 °C, 10 °C og 15 °C i opptil 72 timer. Eksperiment 1 undersøkte kroppens helhetlige tilstand, indre organer, filetkvalitet, muskel-pH, farge og mikrobiell vekst. Eksperiment 2 fokuserte på ytre trekk ved hodet, inkludert gjeller, øyne og skin.
Resultatene viser at forsinket bløgging raskt mister effekt, med restblod som blir igjen i nyrene og i blodkarene i bukhulen. Høyere temperaturer fremskyndet postmortale endringer, inkludert misfarging av hud og hale, opphopning av slim, tap av skjell, bleking av gjeller og oppmykning av muskulaturen. Lavere temperaturer bremset disse prosessene og bevarte kroppens integritet.
De indre organene reflekterte også tidspunktet etter død: ubløgget fisk beholdt mørkere farge i lever og tarm, og vev viste autolyse og økt lukt ved høyere temperaturer. Muskel-pH sank raskere ved forhøyede temperaturer, mens filetens rødfarge forble relativt stabil, men lyshet og tekstur endret seg over tid. Mikrobiell vekst økte med temperaturen, særlig ved 15 °C.
Samlet sett påvirker temperatur og tid postmortale endringer i stor grad. Kombinerte indikatorer – bløggeeffektivitet, blodretensjon, utseende på lever og tarm, tilstand i gjeller og øyne, slimdannelse, nedbrytning av halen, muskel-pH og bakteriebelastning – gir et robust grunnlag for å skille korrekt slaktet laks fra selvdød fisk, og kan dermed støtte myndighetenes kontroll og avdekking av svindel.
Til slutt er det viktig å understreke at denne studien ble gjennomført under kontrollerte forhold ved bruk av gjennomstrømmende UV-behandlet sjøvann for å sikre temperaturkontroll og standardiserte miljøforhold. Fremtidige studier bør ta sikte på å gjenta disse forsøkene i kommersielle merdmiljøer, hvor nedbrytning kan skje raskere som følge av høyere biologisk aktivitet.
Summary
This chapter reports the initial steps in developing a forensic toolbox to determine whether Atlantic salmon were properly stunned and bled according to Norwegian regulations or had been dead before processing. Detecting self-dead fish is critical for food safety and fraud prevention.
Two experiments assessed postmortem changes in salmon stored in seawater at 5 °C, 10 °C, and 15 °C for up to 72 hours. Experiment 1 examined whole-body condition, internal organs, fillet quality, muscle pH, colour, and microbial growth. Experiment 2 focused on external head features, including gills, eyes, and skin.
Results show that delayed bleeding rapidly loses effectiveness, with residual blood retained in the kidney and peritoneal vessels. Higher temperatures accelerated postmortem changes, including skin and tail discoloration, mucus accumulation, scale loss, gill bleaching and muscle softening . Lower temperatures slowed these processes.
Internal organs also reflected postmortem timing: unbled fish retained darker liver and gut coloration, and tissues showed autolysis and increased odor at higher temperatures. Muscle pH declined faster at elevated temperatures, while fillet redness remained stable but brightness and texture changed over time. Microbial growth increased with temperature, especially at 15 °C.
Overall, temperature and time strongly affect postmortem changes. Combined indicators—bleeding efficiency, blood retention, liver and gut appearance, gill and eye condition, mucus, tail degradation, muscle pH, and bacterial load—provide a robust basis for distinguishing properly slaughtered salmon from self-dead fish, supporting regulatory enforcement and fraud detection.
Finally, it is important to acknowledge that this study was conducted under controlled conditions, using flow-through UV-treated seawater, to ensure temperature control and environmental standardisation. Future studies should aim to replicate these experiments in commercial sea-cage environments, where spoilage may occur more rapidly due to higher biological activity.
1 - Introduction
The Norwegian aquaculture industry annually harvests more than 1.6 million tonnes of salmon and rainbow trout, at a value of more than one hundred billion Norwegian kroner (The Norwegian Fisheries Directorate, Aquaculture statistics). Norway has strict regulations to ensure fish welfare during harvest (Dyrevelferdsloven, LOV-2009-06-19-97), good hygiene and food safety (Matloven, LOV-2003-12-19-124). Farmed salmon and rainbow trout must be killed at an approved harvest plant/boat (§ 16, FOR-2008-06-17-822). They must be stunned prior to killing in such a way that they remain unconscious until death (§ 34, FOR-2008-06-17-822). Upon effective stunning, typically either achieved through electrical or percussive methods, the unconscious fish must be bled immediately to achieve mortal blood loss to the brain (§ 34, FOR-2008-06-17-822).
Fish that have been lying dead in the water for a while can contain dangerous bacteria and should not enter the processing line (Regulation (EC) 852/2004, Attachment I, part A). Any dead fish should therefore be removed before the fish are pumped up into the processing line. However, breaches in routines, or other special circumstances, may result in suspicion of that newly dead fish have been processes as live fish. Depending on temperature and how long the fish had been dead before it was processed it can be difficult to assert this for certain when inspecting the salmon afterwards. There is therefore a need for developing a toolbox with indicators that can be used to determine if a salmon carcass have undergone a slaughtering process according to the regulations or have been self-dead for a limited time before being bled.
It is strictly forbidden to market fish that are self-dead for human consumption (§ 14, FOR-2023-02-27-266). There is, however, suspicion that criminals may try to sell discarded self-dead fish for human consumption. A recent TV documentary, for example, alleged the export of self-dead salmon to Kazakhstan (NRK Brennpunkt 2025). Such practice poses a serious threat to public health and to industry’s reputation. Consequently, the industry and Norwegian authorities urgently need reliable tools to detect and prevent such fraud.
We have therefore started a series of experiments with the ultimate aim to develop a forensic toolbox for determining if a salmon carcass has been stunned and bled according to the regulations, or if it had already been dead for a while before it was bled. Here we report results from the two first experiments from this series. The experiments were designed to monitor self-dead fish at different temperatures. Experiment 1 monitored changes over the whole body including internal organs and measures of fillet quality, while Experiment 2 specifically focused on external changes during the decay of the head, including changes in eye and gill tissue.
One possible indicator of that a salmon have been bled correctly is the amount of visible blood in the fillet. Bleeding gives the best effect if done immediately postmortem, and that there is no longer any effect if the fish are bled more than an hour postmortem (Roth et al., 2005). In addition to visible blood in the fillet we also investigate indicators as blood in water if bled, fish outer appearance (discoloration), surface mucus, appearance of head, eyes and gills (redness), tail discoloration, appearance of internal organs (liver), odor, fillet appearance, gaping, fillet pH, fillet colouration and microbial load.
2 - Material and Methods
Two experiments were designed to follow the decay of fish subjected to delayed bleeding after death over a period of 72 hours at different temperatures. Experiment 1 monitored changes over the whole body including internal organs and measures of fillet quality, while Experiment 2 specifically focused on external changes during the decay of the head, including changes in eye and gill tissue.
2.1 - Experiment 1
The experiment was conducted at the Institute of Marine Research (IMR), Matre, Norway. Atlantic salmon (Salmo salar, Aquagen strain) were maintained in an outdoor tank (7 m in diameter; volume 57,000 L) at a temperature of approximately 9°C and a salinity of 28 ppt. At the start of the experiment, 130 fish (length: 60.4 ± 3.99 cm; weight: 2,595.95 ± 468.57 g) were netted and euthanized with an overdose of Finquel Vet (MS-222; 500 mg L⁻¹). Of those fish, 120 were then transported to the experimental facility and distributed into 12 square tanks (400 L each) containing seawater at 28 ppt salinity. Each tank contained 10 fish, with four replicate tanks assigned to each of the three temperature treatments: 5, 10, and 15°C. A net was fitted inside the tank to lift and support the dead fish, preventing them from lying directly on the bottom. This setup was designed to mimic how carcasses settle at the bottom of a sea cage (Figure 1). For each temperature-treatment group, 10 fish were bled for at least 30 minutes and then sampled approximately 4, 24, 48 and 72 hours after death. The bleeding procedure for all groups consisted of cutting the right gill arches and placing the fish in a transport tank filled with seawater at 10°C and 28 ppt salinity. Continuous water movement was maintained in the tank through an inlet pipe to ensure proper flow during bleeding. The excess water was allowed to overflow from the tanks. The remaining 10 fish were used as a control group and were euthanized in the same way as the other fish, but bled immediately after they reached anaesthesia and sampled 30 minutes later.
Figure1 Atlantic salmon carcasses lying on a net suspended inside a tank to mimic how self-dead salmon lie on the bottom of a sea cage
2.1.1 - Sampling procedure
Each fish was tagged with an identification number attached to the tail to enable individual tracking. The control group was sampled first, while the remaining groups were sampled starting four hours postmortem. To minimize potential sampling bias, the order of sampling among the temperature-treatment groups was varied each day in a randomized manner.
For each fish, body weight and fork length were recorded. Photographs were taken of the whole fish, the left gills after removal of the operculum, the viscera, the peritoneal area, and muscle fillet.
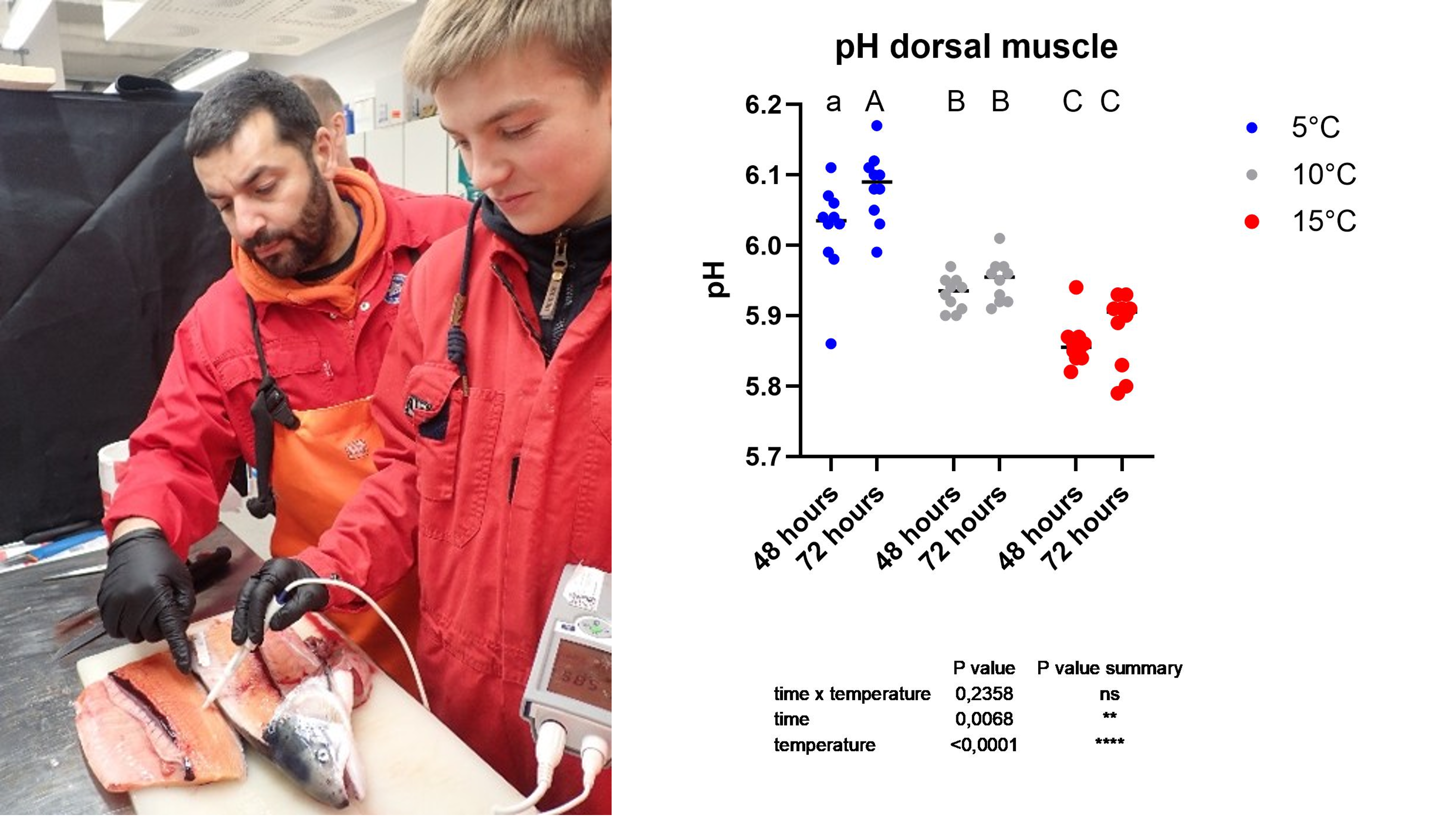
The Norwegian quality cut (NQC) of both sides was collected. The left side was parted in slice of 0.5 cm sections and stored at -20°C for later analyses. From these, two aliquots of samples were used for total aerobic bacterial plate count (TPC) and hydrogen sulfide–producing bacteria analyses. The right side was used to measure muscle colour using the Salmofan (Digital SalmoFan™).
In addition, at the 48 and 72h sampling, muscle pH was measured in all groups on the dorsal fillet, between the dorsal fin and the region just posterior to the head.
2.2 - Experiment 2
Nine salmon from the same stock tank, as these used in Experiment 1, were euthanized with an overdose of Finquel Vet (500 mg L⁻¹) but were not bled at any point during the trial. The operculum on the left side was thereafter cut to expose the gill for easier observation on pictures. Three individuals were placed in each of three transparent flow through plastic boxes supplied with seawater of the same temperatures as in Experiment 1, i.e., 5°C, 10°C and 15°C (Figure 2). In one fish the gill tissue was accidentally damaged when the operculum was cut, resulting in heavy bleeding. This fish was therefore removed, leaving only 2 fish in the 5 °C group. Photographs of the fish were taken at regular intervals up to 73 (15 °C), 96 (10 °C) or 120 (5 °C) hours.
Figure 2 Experimental set up (left and right) that focused on the observation of the fish head decay at different time point (up to 5 days) under 3 different temperature condition (5, 10 and 15 °C).
3 - Observations and discussions
3.1 - Bleeding fail after fish are long dead
Fish that are stunned and bled immediately release blood rapidly into the water. In contrast, delayed bleeding and possible coagulation of blood within the body after death can reduce or even completely prevent bleeding (Tavaris-Dias & Oliveira 2009). Consequently, in the present study, we observed a decrease in the amount of blood released into the water following a cut to the right gill arch, the longer the fish remained unbled.
Figure 3 shows fish 48 hours postmortem, kept at 5 °C (left) and 15 °C (right), and then bled for 30 minutes after cutting their right gills. In both cases, no blood was observed in the water, and the fish were clearly visible in the tanks. The only noticeable difference between the two temperature groups is the cloudiness of the water, caused by the release of mucus from fish kept at higher temperature. The mucus developed along the body and the gills (Figure 6) and was associated with a strong odor, possibly having a negative effect on water quality.
Figure 3 Bleeding bath 48 hours after Atlantic salmon were overdosed with anesthesia and left in tanks equipped with nets and supplied with seawater at 5 °C (left) or 15 °C (right). In all fish, the right gills arch was cut. Note that almost no blood is visible in the running water, whereas water cloudiness is influenced by mucus released from the fish
3.2 - Whole fish assessment
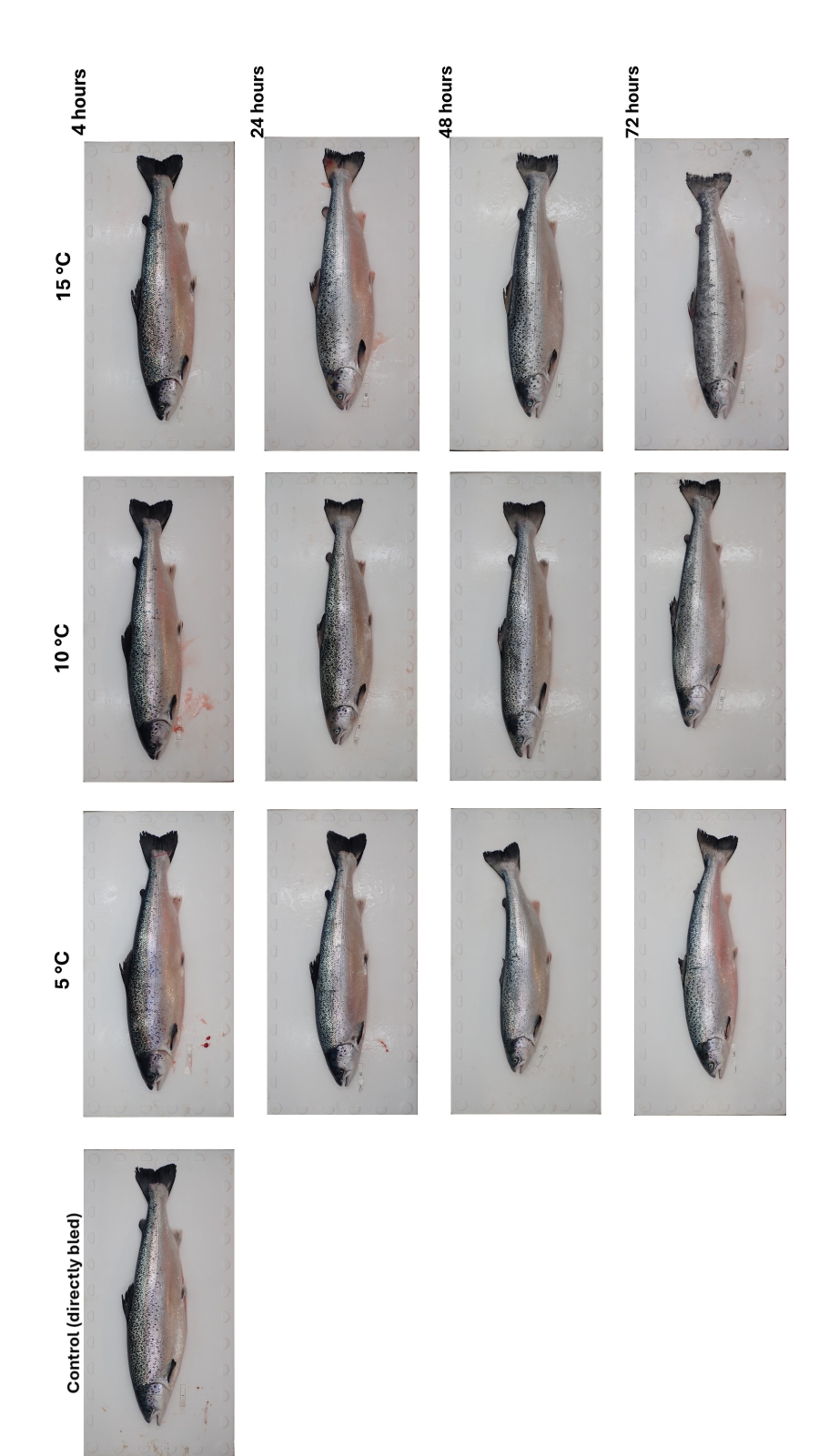
After the fish carcasses were placed in nets, the bodies quickly began to lose their iridescent brightness (Figure 4). After 4 hours, the appearance of the fish was similar across all temperature conditions, with no noticeable differences among individuals. At 10 °C and 15 °C, discolouration, including depigmentation of dark skin areas, progressed rapidly and was clearly apparent within 24 hours postmortem. In contrast, carcasses stored at 5 °C retained their natural skin and scale colouration more effectively. Overall, the degree of discolouration increased with both temperature and storage time, becoming pronounced after 72 hours. Furthermore, fish stored at higher temperatures exhibited noticeable scale detachment upon handling, particularly evident 72 hours postmortem at 15°C (Figure 5).
When handled, fish turgidity differed noticeably among the three temperature treatment groups. Fish kept at 5 °C remained in rigor mortis 48 hours after death. In contrast, carcasses stored at 10 °C and 15 °C showed reduced firmness at the same time point, with fish held at 15 °C becoming noticeably soft after 72 hours. Surface mucus formation was evident on the entire body of fish within 24 hours postmortem, primarily in fish stored at 15 °C. Mucus accumulation became apparent in the other temperature groups after 48 hours, although it was consistently less pronounced in fish held at 5 °C.
Figure 4 Photographs of Atlantic salmon, which were euthanized with an overdose of anesthesia and subsequently bled either immediately after death or at 4, 24, 48, and 72 hours postmortem, under three different temperature conditions.
Figure 5: Atlantic salmon lying dead in water at 5 °C, 10 °C, and 15 °C for 72 hours show progressive quality deterioration. At 5°C, scales and fins remain largely intact, skin retains bright silvery coloration, body firmness is preserved, and eyes appear clearer than in higher-temperature treatments. At 10°C, fish exhibit moderate scale loosening, fin fraying, dulling of skin and tail color, slight softening of tissue, and mild eye cloudiness. At 15°C, salmon display extensive scale loss, advanced fin degradation, pronounced discoloration, tissue breakdown with a flattened body profile, and marked eye cloudiness, reflecting accelerated spoilage.
3.3 - Head eye and gills
A closer observation of the fish head (Figure 6) reveals three main factors that change with time and storage temperature:
i) Gill redness decreased over time, with the discoloration becoming more pronounced at higher temperatures. This effect is most clearly observed 72 hours postmortem but also clearly visible after 48 hours at 15 C.
ii) Mucus accumulation on the gills increased over time but decreased in areas where gill bleaching (loss of redness) occurred.
iii) The skin of the head progressively lost color, noticeable as early as 24 hours postmortem. This can be observed particularly on the dorsal side of the head, which is not covered by scales.
Furthermore, although not visible it in the photographs, several fish exhibited corneal dehydration and sunken eyes.
It should be noted that the relationship between discoloration and time and temperature is less clearly depicted in Figure 6. The extent and pattern of discoloration varied among individuals and may have been influenced by factors such as position in the tank or contact with other fish or equipment. This is exemplified in Figure 7, where a net imprint on the head indicates that the fish had been lying dead at the bottom of the net.
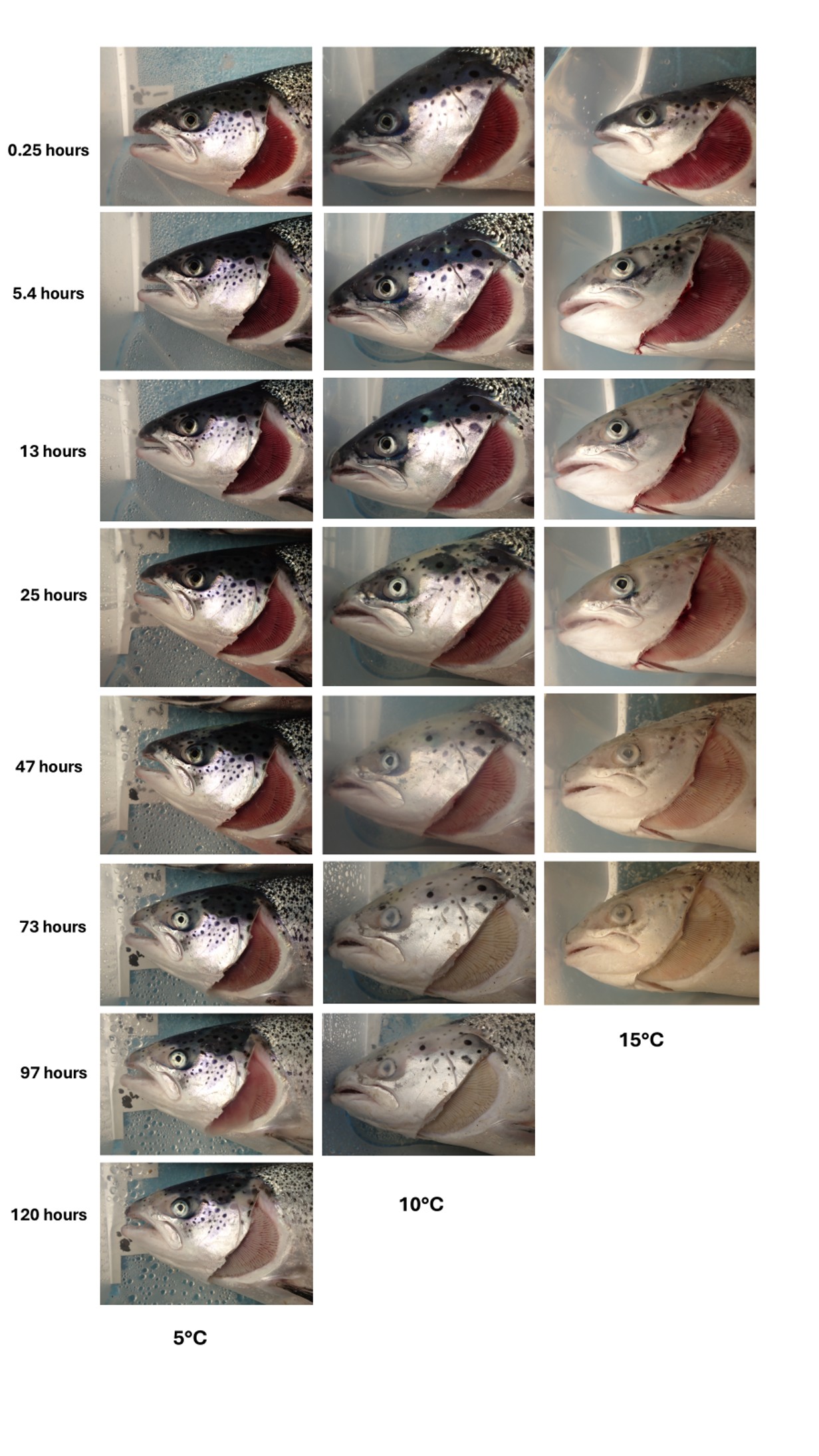
In Figure 6, it is also evident that the brightness of the fish eyes decreases over time. However, a proper comparison between time points is not possible due to the lack of baseline measurements for each fish at the beginning of the trial and the potential presence of pre-existing eye conditions, such as cataracts. To more clearly evaluate the influence of time and temperature on head-related features, Experiment 2 was conducted to monitor postmortem changes over time. This experiment focused on eye condition, gill appearance, and overall head characteristics, as shown in Figure 8.
Figure 6 Atlantic salmon sampled immediately after bleeding (control) and at 4, 24, 48, and 72 hours postmortem after euthanasia by anesthetic overdose, while kept in water tanks at 5, 10, and 15 °C
Figure 7 Close-up of skin discoloration in the head of a fish held at 10 °C for 72 hours postmortem. The observed cross‑pattern corresponds to the points of contact between the fish head and the net.
Figure 8 illustrates more clearly how the decay process in fish is accelerated at higher temperatures. The time-lapse sequence highlights a progressive discolouration of the head skin, which began as early as 13 hours postmortem at 15 °C and within 25 hours at 10 °C. The skin discolouration appeared to occur in parallel with the loss of the dark area surrounding the eye. Within 47 hours, corneal opacification was evident in both the 10 °C and 15 °C groups, whereas this process was not observed in the 5 °C group. Similarly, at 73 hours, the gills of fish held at 10 °C and 15 °C appeared completely white, while those maintained at 5 °C still exhibited a pale pink colouration even 120 hours postmortem.
Figure 8 Atlantic salmon placed in three rectangular boxes supplied with seawater through an overflow system and divided into three temperature groups, at 5 °C, 10 °C, and 15 °C. Each column represents fish maintained at a specific temperature, and each row corresponds to increasing postmortem intervals. Fish remained unbled in the boxes for up to 120 hours.
3.4 - Tail discolouration
A visual inspection of the caudal area (Figure 9) clearly shows that postmortem degradation progresses faster at higher temperatures. The tail region pictures of fish stored at 10 and 15 °C displays early discolouration and skin dullness within the first 24 hours, followed by scale detachment and tissue softening after 48–72 hours. In contrast, fish stored at 5°C retain their natural brightness and scale integrity for a longer period, indicating a slower rate of decay. These observations suggest that the tail region could serve as a promising external indicator of postmortem changes and overall freshness in Atlantic salmon
Figure 9 Progressive postmortem changes in the tail region of Atlantic salmon lying in seawater at 5 °C, 10 °C, and 15 °C over time. Each row represents fish maintained at a specific temperature, and each column corresponds to increasing postmortem intervals. The images illustrate a gradual loss of skin brightness and iridescence, increasing opacity, and scale loosening with higher temperatures and longer postmortem time. At 15 °C, the onset of discolouration and tissue degradation occurs noticeably earlier compared to 10 °C and 5 °C.
3.5 - Muscle and organs appearance
After stunning, fish should be bled immediately and thoroughly gutted, as blood residues and remaining tissues (e.g. kidney) in the abdominal cavity have been shown to negatively affect fillet quality, its firmness and increase gaping (Jacobsen et al., 2017). In the present study, however, all fish were handled identically and left ungutted postmortem, except the control fish that were gutted after 30 min bleeding. This allows us to focus specifically on the effects of temperature and postmortem time on the muscle and internal organs, and to identify potential differences compared with fish that are bled immediately after death. Figures 10 and 11 illustrate the fillet, internal organs, and body cavity, from which several major observations can be made:
The liver is a sensitive indicator of proper bleeding and early spoilage. In fish that are bled shortly after death (control fish), the liver colour changes from dark red/purple to a clearer dark orange or mustard-like colour. In unbled fish, the liver tends to retain blood, resulting in a darker appearance. In some cases, puncturing the liver during filleting can cause localized bleeding, leading to discolouration around the puncture site (e.g., Figure 11, fish bled 4 hours postmortem at 10 °C). Around 72 hours postmortem, at 15 °C, liver decay (autolysis) becomes more evident, with the tissue appearing gelatinous and shiny. At this stage, the liver can begin to affect the fillet, leaving marks on the parietal peritoneum (Figure 10, fish number 129). Fish stored at 5 °C maintained fillet brightness, red muscle definition, and liver integrity for much longer, confirming the protective effect of lower storage temperatures.
Figure 10 Postmortem changes in fillet appearance and internal organs of Atlantic salmon lying in seawater at 5 °C, 10 °C, and 15 °C over time. Each column represents at a specific temperature treatment, and each row corresponds to increasing postmortem time intervals. The images aimed to show progressive changes in fillet colour, gaping, blood clearing, and liver appearance throughout the storage period.
The fish gut also appears to be a reliable indicator of whether bleeding occurred soon after death. In the control group and in fish bled 4 hours postmortem and stored in water at 5 °C and 10 °C, the visceral fat exhibited a clearer, pale colour. In contrast, in fish bled later after death and those stored at higher temperature, the areas around the pyloric caeca and the surrounding fat became redder. By 48 and 72 hours postmortem, the gut of fish stored at 10 °C and 15 °C showed signs of autolytic degradation.
The blood that is not released into the water during bleeding (see Figure 3) is retained within the body, primarily in the vena cava, which lies between the fish’s spine and kidney. Although not immediately noticeable, careful examination of Figures 10 and 11 reveals that after 24 hours, for fish stored at 15 °C, the kidney line appears thicker due to the retention of coagulated blood. In addition, although not immediately noticeable, compared with the control and fish bled 4 hours postmortem, unbled fish later show more blood in the vessels around the parietal peritoneum, over the ribs (Figure 11), and in the vessels surrounding the backbone in the dorsal area of the fillet.
Figure 11 Zoom on postmortem changes on the internal organs of Atlantic salmon lying in seawater at 5 °C, 10 °C, and 15 °C over time. Each column represents at a specific temperature treatment, and each row corresponds to increasing postmortem time intervals. The images aimed to show progressive changes in blood clearing, and liver appearance throughout the storage period.
Sensorially, the odor intensity of fish gut samples stored at 10 °C and 15 °C increased markedly, becoming strongly pronounced by after 72 hour, particularly at 15 °C. Fillet firmness was also described as low. When carefully filleted, the fillets did not exhibit any visible gaps; however, after 72 hours kept in water at 15 °C, the fillets became fragile and prone to tearing when handled (Figure 12). In addition, the skin was easily detached from the underlying muscle.
Figure 12 gaping on salmon filet, 72 hours postmortem and resting in water at 15 °C
3.6 - The muscle fillet pH
As postmortem time progresses, anaerobic ATP production also ceases. This occurs primarily due to the depletion of glycogen substrates and the progressive inhibition of glycolytic enzymes, the latter largely driven by the decline in muscle pH associated with lactic acid buildup. Together, these processes contribute to the biochemical environment characteristic of early postmortem muscle and set the stage for subsequent quality changes observed during storage. Furthermore, the hydrolysis of ATP within the muscle contributes additional protons (H+) to the system, thereby intensifying the postmortem decline in pH (reviewed by Daskalova 2019).
In this trial, the muscle pH of fish stored at 5 °C for 48 and 72 hours was similar to values previously reported for Atlantic salmon (Skjold et al., 2020), Atlantic cod (Gadus morhua, Kristoffersen et al., 2006) and rainbow trout (Oncorhynchus mykiss, Germond et al., 2023) about 40 hours after slaughter. However, it is important to note that in those studies fish were gutted and bled shortly after slaughter, while in the current trials pH was measured on unbled and ungutted fish (Figure 13, left).
In the present experiment, fish maintained at 5 °C exhibited higher muscle pH values (6.02 ± 0.02 at 48 hours and 6.08 ± 0.02 at 72 hours poststress(Figure 13, right), indicating a slower rate of postmortem metabolism. In contrast, elevated water temperatures appeared to accelerate biochemical processes and, consequently, lower muscle pH. This pattern was reflected in the measurements obtained at 10 °C (5.93 ± 0.01 at 48 hours and 5.95 ± 0.01 at 72 h) and at 15 °C (5.86 ± 0.01 at 48 hour and 5.88 ± 0.01 at 72 h), respectively.
In theory, during spoilage, fish pH levels can increase over time due to the production of alkaline bacterial metabolites, which coincides with rising levels of total volatile basic nitrogen (TVBN; not measured here; Abbas et al., 2008). However, this increase was only partially observed at 5 °C (between 48–72 hours) and was not observed in the present study for the higher temperature groups, even after three days of exposure to elevated water temperature. It is important to note that most knowledge on fish spoilage is based on studies of fish shelf life and storage out of seawater, whereas fish in seawater may be subjected to different processes than those occurring in fish stored on ice or under controlled-atmosphere conditions.
Figure 13 pH of dorsal muscle (right) in fish stored at 5 °C (blue), 10 °C (gray), and 15 °C (red) measured after 48 h and 72 hours postmortem. Values represent individual measurements with mean ± SEM. Different uppercase letters (A–C) indicate significant differences between group temperatures (p < 0.05), while lowercase letters (a, b) indicate significant differences between times within the same temperature groups.
3.7 - Muscle coloration
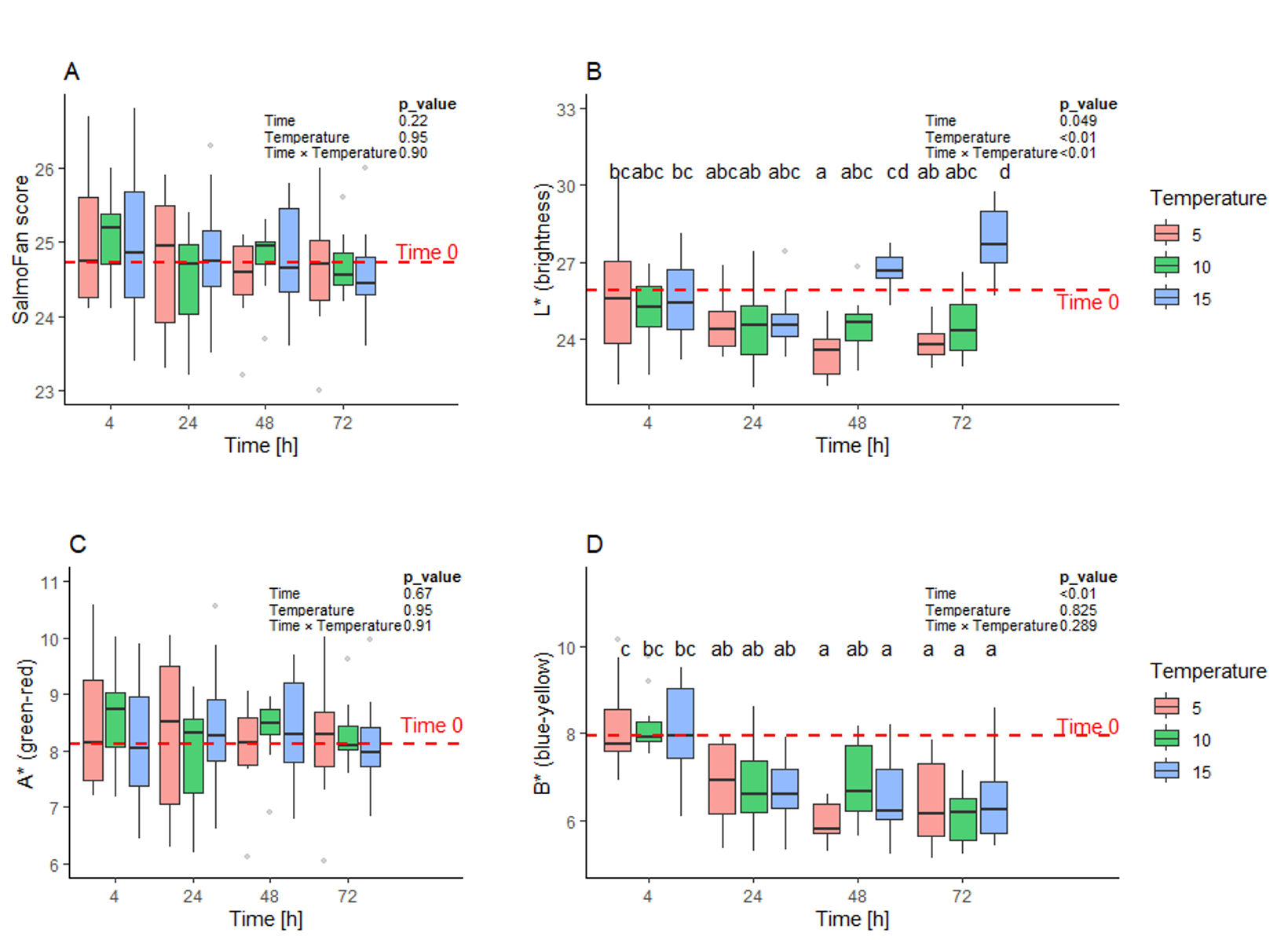
Fish colour remained largely stable over 72 hours, with SalmoFan general scores (Figure 14 A) and A* (red–green, Figure 14 C) values showing no clear temporal or temperature trends. In contrast, brightness (L*), and blue-yellow (B*) values changed significantly over time (Figure 14 B and D respectively), while only the brightness was influenced by temperature. Brightness generally decreased from 4 to 48 hours, particularly at 5 °C and 10 °C, but increased between 48 and 72 hours in the 15 °C group. B* values declined over time, reflecting a shift toward the blue-end of the spectrum. These results indicate that, while overall colour and redness remain stable, both brightness and blue–yellow balance are sensitive to the pre-bleeding time duration and temperature.
Fillet colour and pH are influenced not only by storage and handling conditions but also by the fish’s physiological state prior to slaughter. Exposure to varying levels of stress in commercial operations—such as during transport, pumping, crowding, or stunning—can negatively affect fillet quality and reduce shelf life. Erikson and Misimi (2008) reported that premortem stress in Atlantic salmon resulted in darker fillets, as indicated by lower L* values. However, the impact of stress on fish meat colour remains somewhat inconsistent, likely due to the influence of additional factors such as temperature. Flesh colour changes are also associated with storage conditions (reviewed in Daskalova, 2019). Colour variations may arise from rapid ATP depletion and accompanying changes in muscle pH. Interestingly, consistent with the findings of the present study, previous research has identified a relationship between fillet texture and colour, with fillets exhibiting higher L* values being softer than those with lower L* values (Kiessling et al., 2004).
Elevated temperatures appear to sustain or even enhance L* values over extended pre-bleeding period. Comparable trends have been observed in land farmed animals. In broiler breast meat (reviewed in Daskalova, 2019), reduced muscle pH is linked to lighter meat colour and increased light reflectance. These effects are largely driven by alterations in sarcoplasmic protein solubility and pigment interactions (Swatland, 2008; Van Laack et al., 2000; Joo et al., 1999). In support of this, Van Laack et al. (2000) reported a strong positive association between muscle pH and sarcoplasmic protein solubility, alongside an inverse relationship between these factors and L* (lightness) values. Additionally, a rapid postmortem decline in pH and lower ultimate pH are widely recognized as key contributors to paler appearance in pork and poultry. For example, chicken breast meat with a lower pH (around 5.91) demonstrates greater surface reflectance and a lighter appearance, whereas meat with a higher pH (approximately 6.36) permits deeper light penetration, producing a darker visual impression (Swatland, 2008).
Figure 14 SalmoFan score and color values of Atlantic salmon fillets from fish lying dead in water at three different temperatures (5°C, 10°C, and 15°C) and sampled at four time points (4, 24, 48, and 72 h postmortem). Statistical analysis (n = 10 fish per treatment) was performed using two-way ANOVA followed by Tukey’s HSD post hoc test.
3.8 - Microbial spoilage
A homogenized mixture of salmon fillet and skin was embedded into agar culture Petri dishes for the enumeration of total aerobic bacteria and hydrogen sulphide–producing microorganisms.
The total amount of bacteria can be a general indicator for hygiene and freshness of food, and specific bacteria are often associated with spoilage. The main spoilage bacteria in fresh or chilled seafood are characterized by their ability to produce hydrogen sulfide and reduce TMAO (trimethylamine oxide). In seafood, these include for instance Shewanella spp., Aeromonas spp., Enterobacteriaceae Photobacterium spp. and Vibrio spp (Fogarty et al., 2019; Gram et al., 1987; Nian et al., 2022). Most of these typical “spoilage” bacteria such as Shewanella spp., Pseudomonas spp. and Photobacterium spp. are naturally present in seawater, and can in fisheries products stored at low temperatures increase over time. As such, their amount is an indicator of the quality and “freshness” of fish-products (Fogarty et al., 2019; Gram, 1992). The focus here has been on total counts of aerobic bacteria and H2S-producing spoilage bacteria.
Three analytical methods were initially evaluated to determine the most suitable approach for this sample type: IMR method 111, which is based on NMKL method 184 (NMKL, 2006); a modified version of IMR method 111 (m111); and IMR method 384 for aerobic microorganisms using Petrifilm™ (Neogen).
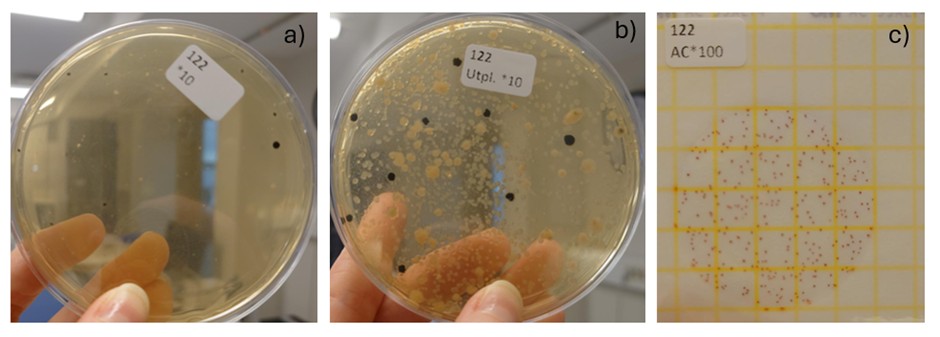
The modified method 111 (m111; a summary of the method is displayed in box1) was chosen for all samples because it consistently yielded higher aerobic counts than the original method 111 and allowed detection of H2S-producing bacteria through black-centred colonies. The lower counts in the original method may be due to thermal stress when embedding samples in molten agar at ~45 °C, which could reduce survival of heat-sensitive bacteria. Method 384 (Petri film) was not used, as it quantifies only total aerobic bacteria and cannot distinguish spoilage organisms. Extending incubation to 96 hours in m111 further improved visibility of black colonies, ensuring reliable assessment of both total aerobes and H2S-producing spoilage bacteria. Example of the three different culture can be observed in Figure 15.
Figure 15: Examples of bacterial growth on the three methods tested in this study. A) Method 111, b) modified method 111, and c) Methods 384.
Figure 15: Examples of bacterial growth on the three methods tested in this study. A) Method 111, b) modified method 111, and c) Methods 384.
Box 1: Brief description of modified method 111
Based on method 111 – Aerobe counts and hydrogen sulphide producing bacteria
NMKL method 184, 2006
1. Weigh 20 g of sample into a stomacher bag with filter
2. Dilute the sample 1:10 with dilution water via dilutor
3. Homogenize for 30 sec
4. Set up a 10-fold dilution series (1ml+9ml) according to the expected amount of bacteria in the sample
5. Transfer 1 ml of the dilution to an Iron-agar petri dish containing L-cysteine
6. Distribute sample material with an L-rod, and air dry in a sterile bench
7. The petri-dishes are incubated at 25 °C for 72-96 hours
8. The petri-dishes are counted both after 73 hours and 96 hours
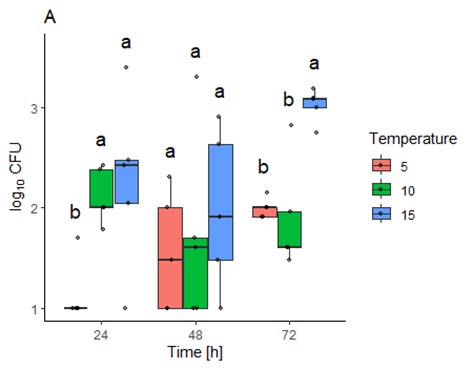
Figure 16. Bacteria abundance as log10CFU (colony forming units) as effect of time. As data were not normally distributed, were log-transformed (since the bacterial growth is exponential and 10-fold differences in CFU reflect this exponential growth). Likelihood ratio tests were used to identify main effects and interactions between temperature and time (survival, version 3.5-7). Values <10 were treated as left centered. A sidak-test (emmeans, version 1.10.3) was used for pairwise comparison between groups.
Bacterial cultures showed though that at 24 hours post-bleeding the colony forming bacteria units (CFU, bacteria abundance) were significantly higher in fish kept at 10 or 15 ˚C compared to those kept at 5 ˚C (Figure 16). After 48 hours no significant differences could be detected between the groups. However, after 72h the CFU was significantly higher in fish kept at 15 ˚C compared to those kept at 5 or 10 ˚C.
Growth of H2S producing bacteria was overall poor, and too few to be subjected to statistical analyses. Few colonies grew, with the highest numbers observed being at 48 hours in samples from fish stored at 10 and 15 ˚C. However, as the number of colonies counted in all samples were below 20 (O’Toole, 2016), the results are unreliable. Most likely there were few H2S producing bacteria present. Another possibility is that there were more H2S-producers present, but they did not handle freezing very well, and that this analyses could have been more successful if fresh samples were analysed, the chosen method is generally used for fresh samples (NMKL, 2006). However, previous studies have indicated that freezing for up to 5 weeks should have little effect on bacterial counts (Magnússon & Martinsdóttir, 1995).
It is important to note that this pilot trial was conducted in flow-through experimental tanks under controlled conditions, where seawater was filtered and treated with UV before entering the tanks. While such controlled conditions allow the effect of water temperature on postmortem processes to be carefully regulated, they may not fully reflect the complexity of commercial or natural environments. In sea-cage settings, fish may exhibit signs of spoilage and postmortem deterioration in a shorter time than observed in this study. These processes could be further accelerated by the presence of saprophagous organisms other than bacteria, including invertebrates or other fish. This hypothesis is worthy of further investigation to better understand how environmental conditions and biological interactions influence spoilage dynamics.
Fresh fish harbor different initial microbiota associated with environmental factors such as temperature and salinity, and marine fish is often dominated by Firmicutes, Bacteroidetes, and Proteobacteria (Chi et al., 2025). After harvest and during storage the community shifts, and psychrotrophic, cold‑tolerant bacteria such as Pseudomonas spp. and Shewanella spp. can grow well at cold temperatures and often dominate spoilage of chilled fish (Fasolato & Andreani, 2025; Vogel et al., 2005). Unpreserved fish can also be spoiled by gram‑negative fermentative bacteria such as Vibrios (Gram & Dalgaard, 2002). Maldi-tof identification of twelve randomly selected colonies identified eight bacteria, whereas four could not be identified (Table 1). Of the identified bacteria, two were Shewanella baltica, which are well known spoilage bacteria (Gram & Dalgaard, 2002; Vogel et al., 2005). The other five isolates included at least three that have the potential to contribute to spoilage, Vibrio, Aeromonas and Pseudomonas (Table 1). The specific strains identified here have not been associated specifically with spoilage, but closely related strains have (Chi et al., 2025; Gram & Dalgaard, 2002). It is possible that a more in-depth analyses of microbial community changes over time using next-generation sequencing (NGS) could give more details of microbial community shifts after fish-death, and should be tested in future studies.
Table 1: Identification of isolates using Maldi-tof. Best match indicates best match in the Maldi-tof database. ID score indicates how certain it is that the result is correct, where good means very certain, and medium means there might not be a very good match in the database, or the isolate was not pure.
Fish pathogen, other Aeromonas spp. Are known to cause spoilage
medium beige
Pseudomonas libanensis
Good
Associated with food spoilage
medium dark beige
Glutamicibacter bergerei
Good
Uncertain, little information available
pink-beige
Psychrobacter namhaensis
Medium
Little information on P. namhaensis, but other Psychrobacter spp. are common spoilage bacteria
4 - References
§ 14, FOR-2013-06-28-844 (2013) Nærings- og fiskeridepartementet (akvakulturfordriften).
§ 14, FOR-2023-02-27-266 Nærings- og fiskeridepartementet
§ 16, FOR-2008-06-17-822 (2008). Forskrift om drift av akvakulturanlegg (akvakulturfordriften). Nærings- og fiskeridepartemente
§ 34, FOR-2008-06-17-822 (2008). Forskrift om drift av akvakulturanlegg (akvakulturfordriften). Nærings- og fiskeridepartementet.
Abbas, K. A., Mohamed, A., Jamilah, B., & Ebrahimian, M. (2008). A review on correlations between fish freshness and pH during cold storage. American journal of biochemistry and biotechnology, 4(4), 416-421.
Chi, Y., Luo, M., & Ding, C. (2025). The role of microbiota in fish spoilage: biochemical mechanisms and innovative preservation strategies. Antonie van Leeuwenhoek 2025 118:7, 118(7), 89-. https://doi.org/10.1007/S10482-025-02101-Z
Daskalova, A. (2019). Farmed fish welfare: stress, post-mortem muscle metabolism, and stress-related meat quality changes. International aquatic research, 11(2), 113-124.
Dyrevelferdsloven (LOV-2009-06-19-97) The Norwegian Animal Welfare Act. Dyrevelferdsloven LOV-2009-06-19-97. Norwegian Ministry of Agriculture and Food. [Cited 1 November 2019.] Available from URL: www.lovdata.no
Erikson, U., & Misimi, E. (2008). Atlantic salmon skin and fillet color changes effected by perimortem handling stress, rigor mortis, and ice storage. Journal of food science, 73(2), C50-C59.
Fasolato, L., & Andreani, N. A. (2025). The genus Pseudomonas. The Microbiological Quality of Food: Foodborne Spoilers, Second Edition, 23–55. https://doi.org/10.1016/B978-0-323-91160-3.00007-6
Fogarty, C., Whyte, P., Brunton, N., Lyng, J., Smyth, C., Fagan, J., & Bolton, D. (2019). Spoilage indicator bacteria in farmed Atlantic salmon (Salmo salar) stored on ice for 10 days. Food Microbiology, 77, 38–42. https://doi.org/10.1016/J.FM.2018.08.001
Germond, A., Vénien, A., Ravel, C., Castulovich, B., Rouel, J., Hutin, M., ... & Astruc, T. (2023). The effects of postmortem time on muscle trout biochemical composition and structure. Foods, 12(10), 1957.
Ghaly, A. E., Dave, D., Budge, S., & Brooks, M. S. (2010). Fish spoilage mechanisms and preservation techniques. American journal of applied sciences, 7(7), 859.
Gram, L. (1992). Evaluation of the bacteriological quality of seafood. International Journal of Food Microbiology (Vol. 16, 25-39).
Gram, L., & Dalgaard, P. (2002). Fish spoilage bacteria – problems and solutions. Current Opinion in Biotechnology, 13(3), 262–266. https://doi.org/10.1016/S0958-1669(02)00309-9
Gram, L., Trolle, G., & Huss, H. H. (1987). Detection of specific spoilage bacteria from fish stored at low (0 o C) and high (20 o C) temperatures. In International Journal of Food Microbiology (Vol. 4).
Jacobsen, A., Joensen, H., & Eysturskarð, J. (2017). Gaping and loss of fillet firmness in farmed salmon (Salmo salar L.) closely correlated with post‐slaughter cleaning of the abdominal cavity. Aquaculture Research, 48(1), 321-331.
Joo, S. T., Kauffman, R. G., Kim, B. C., & Park, G. B. (1999). The relationship of sarcoplasmic and myofibrillar protein solubility to colour and water-holding capacity in porcine longissimus muscle. Meat science, 52(3), 291-297.
Kiessling A, Espe M, Ruohonen K, Mørkøre T (2004) Texture, gaping and colour of fresh and frozen Atlantic salmon flesh as affected by pre-slaughter iso-eugenol or CO2 anaesthesia. Aquaculture 236(1–4):645–657. https://doi.org/10.1016/j. aquaculture.2004.02.030
Kristoffersen, S., Tobiassen, T., Steinsund, V., & Olsen, RL (2006). Slaughter stress, postmortem muscle pH and rigor development in farmed Atlantic cod (Gadus morhua L.). International Journal of Food Science and Technology , 41 (7), 861-864.
Magnússon, H., & Martinsdóttir, E. (1995). Storage Quality of Fresh and Frozen-thawed Fish in Ice. Journal of Food Science, 60:2.
Matloven (LOV-2003-12-19-124) The Norwegian Food Act. Matloven LOV-2003-12-19-124. Norwegian Ministry of Health and Care Services. [Cited 1 November 2019.] Available from URL: www.lovdata.no
Nian, L., Wang, M., Pan, M., Cheng, S., Zhang, W., & Cao, C. (2022). A potential spoilage bacteria inactivation approach on frozen fish. Food Chemistry: X, 14, 100335. https://doi.org/10.1016/J.FOCHX.2022.100335
NMKL. (2006). NMKL 184. Aerobic count and specific spoilage organisms in fish and fish products. - NMKL. https://www.nmkl.org/nmkl-184-aerobic-count-and-specific-spoilage-organisms-in-fish-and-fish-products/
O’Toole, G. A. (2016). Classic Spotlight: Plate Counting You Can Count On. Journal of Bacteriology, 198(23), 3127. https://doi.org/10.1128/JB.00711-16
Olsen, S. H., Sorensen, N. K., Stormo, S. K., & Elvevoll, E. O. (2006). Effect of slaughter methods on blood spotting and residual blood in fillets of Atlantic salmon (Salmo salar). Aquaculture, 258(1-4), 462-469.
Regulation (EC) No 852/2004 of the European Parliament and of the Council of 29 April 2004 on the hygiene of foodstuffs
Roth, B., Torrissen, O. J., & Slinde, E. (2005). The effect of slaughtering procedures on blood spotting in rainbow trout (Oncorhynchus mykiss) and Atlantic salmon (Salmo salar). Aquaculture, 250(3-4), 796-803[AM1] .
Skjold, V., Joensen, J. K., Esaiassen, M., & Olsen, R. L. (2020). Determination of pH in pre rigor fish muscle–method matters. Journal of aquatic food product technology, 29(5), 480-485.
Swatland, H. J. (2008). How pH causes paleness or darkness in chicken breast meat. Meat Science, 80(2), 396-400.
Tavares-Dias, M., & Oliveira S.R., (2009) . "A review of the blood coagulation system of fish." Revista Brasileira de Biociências 7(2):205-224
Van Laack, R. L. J. M., Liu, C. H., Smith, M. O., & Loveday, H. D. (2000). Characteristics of pale, soft, exudative broiler breast meat. Poultry Science, 79(7), 1057-1061.