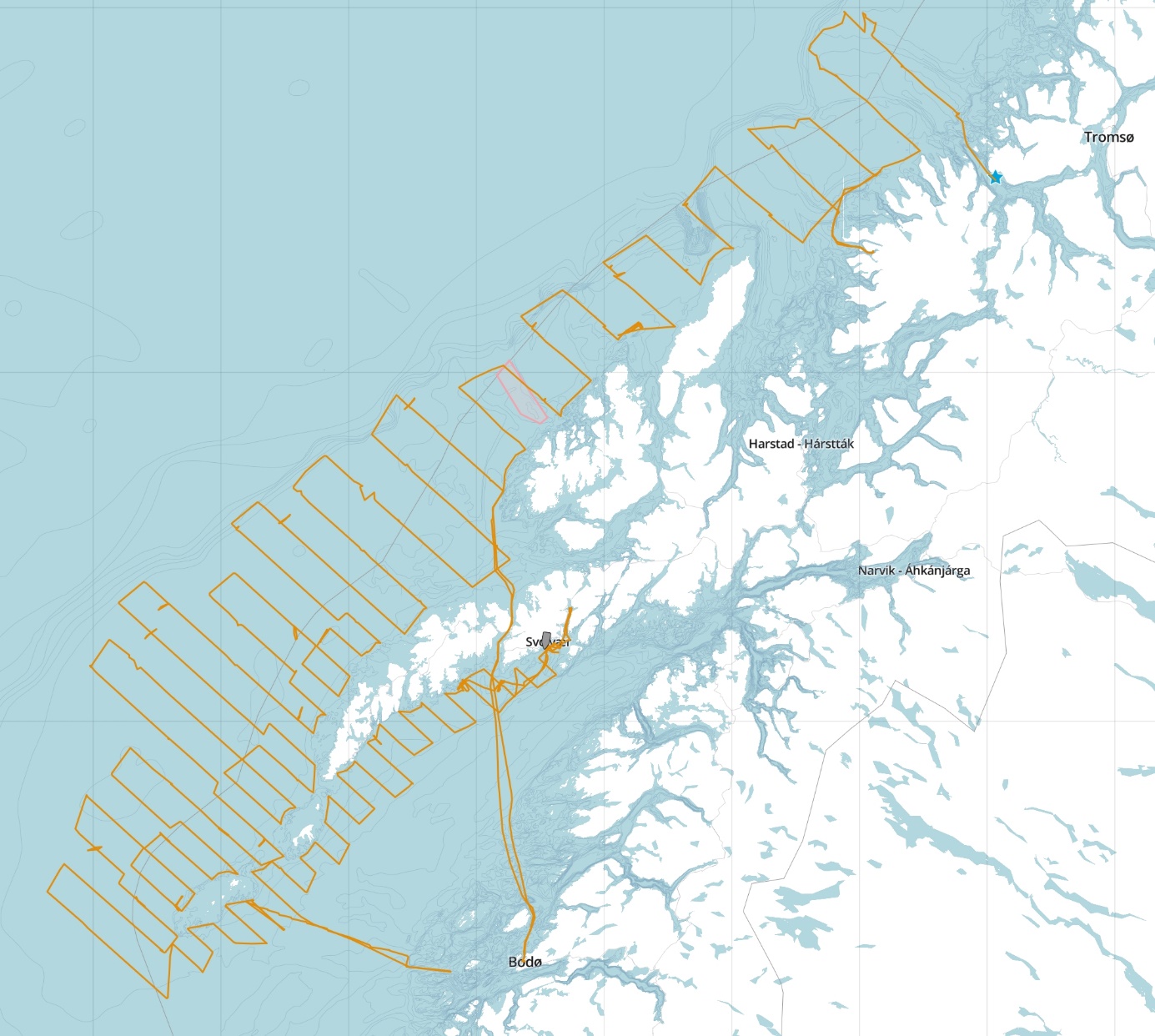
The 2025 skrei (North East Arctic Cod spawners) survey was largely carried out according to plan, but with significant cuts south of Røstbanken. Net hauls carried out downstream from the uncovered area indicated that spawning was not taking place there. The survey was carried out using standard methodology established for the time series.
The survey covered the coastal area from 70° north southwards to Røstbanken, as well as the Vestfjord, and yielded a quantity estimate of 32 thousand tonnes of mature skrei (age groups older than 5 years). This is dramatically and significantly lower than what could be expected from the stock assessment. The bulk of the spawning skrei were fish that were 9 years and older.
Typical skrei registrations were only found in a few limited areas and the observations corresponded well with the areas where fishing was taking place.
Observations of newly spawned cod eggs in the water column were mainly made in the same areas as where the largest concentrations of spawning fish were registered. The total quantities of cod eggs were very low.
Sammendrag
Skreitoktet 2025 ble i hovedsak gjennomført i henhold til plan, men med betydelige avkortninger sør på Røstbanken. Håvtrekk utført nedstrøms fra det udekkede området indikerte at det ikke foregikk gyting der. Toktet ble gjennomført med standard metodikk etablert for tidsserien.
Toktet dekket kystområdet fra 70° nord sørover til og med Røstbanken, samt Vestfjorden, og ga et mengdeanslag på 32 tusen tonn moden skrei (aldersgruppene eldre enn 5 år). Dette er dramatisk og betydelig lavere enn hva som kunne forventes ut ifra bestandsvurderingen. Hovedvekten av den gytende skreien var fisk som var 9 år og eldre.
Typiske skreiregistrering ble kun funnet på noen få avgrensede områder og observasjonene samsvarte godt med de områder hvor det foregikk fiske.
Observasjoner av nygytte torskeegg i vannsøylen ble i hovedsak gjort i de samme områdene som der de største konsentrasjonene av gytefisk ble registrert. De totale mengdene av torskeegg var svært lave.
1 - Gjennomføring
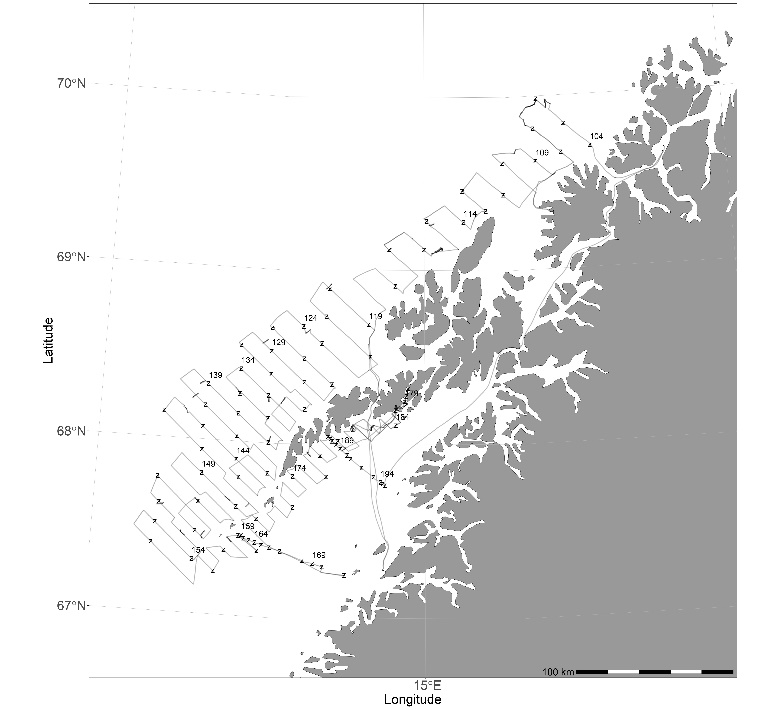
Toktet ble gjennomført med FF «Johan Hjort» i perioden 21. mars – 4. april ( A ). Toktet hadde noen dager værhindring og fikk ikke dekket hele det planlagte området. Håvtrekk etter egg nedstrøms fra det udekkede området indikerte at det ikke foregikk gyting der. Toktet består av parallelle kurser (transekter) og avstanden mellom kursene i de underområdene (strata) hvor det tradisjonelt er tettere registreringer er noe mindre. Informasjon fra trålstasjoner ble benyttet som støtte i vurdering av ekkogram og fordeling av ekkomengder på forskjellige arter.
Figur 1 Kursene fra årets tokt.
Det ble gjennomført to hydrografisk snitt (Røst-Tennholmen og Ballstad-Måløy/Skarholmen).
I tillegg til vanlig prøvetaking, samlet vi (som i fjor) genetiske prøver for verifisering av bestemmelse av bestandstilhørighet (kysttorsk/skrei) som rutinemessig blir bestemt fra undersøkelser av otolitt/ørestein. Disse prøvene ble samlet inn for et internt prosjekt ledet av Torild Johansen. Vi samlet også inn prøver av rogn fra torsk for å få mer nøyaktige modningsbeskrivelser ut fra mikroskopi metodikk.
I alt ble det samlet inn 1113 nautiske mil med tolkede akustiske observasjoner, 26 trålhal, håvtrekk fra 74 posisjoner, og CTD fra 93 posisjoner. Toktet ble gjennomført i henhold til toktmanual («Overvåkningstokt – Skreitokt», internt dokument), og biologisk prøvetaking i henhold til prøvetakingshåndboken («Håndbok for prøvetaking av fisk, krepsdyr og andre evertebrater», internt dokument). 373 torsk ble fullt opparbeidet med alle observasjoner og prøveinnsamling. Det er naturlig at det blir et begrenset antall slike enkeltfiskobservasjoner når innsiget er så svakt som i år. I et mer typisk gyteinnsig vil toktet samle inne detaljerte prøver av rundt 700 torsk.
2 - Metoder
2.1 - Akustikk
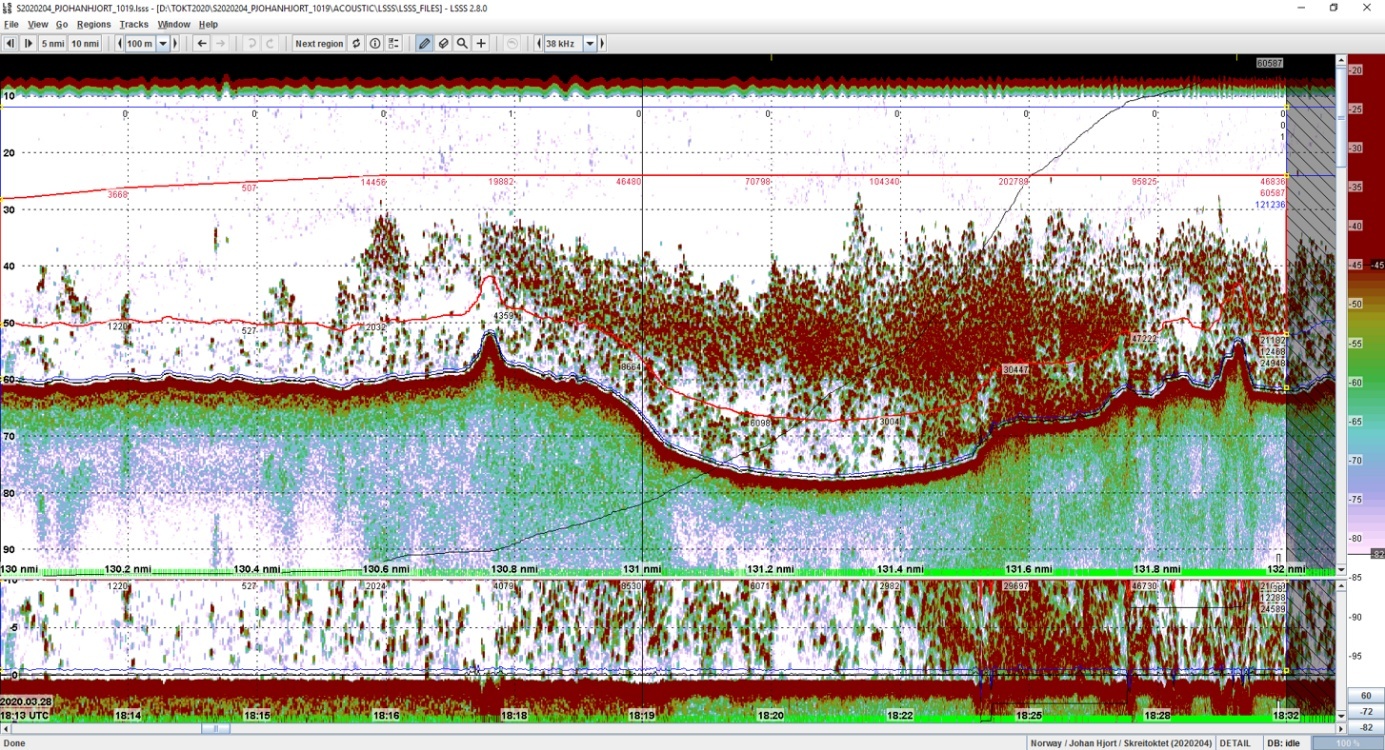
Skreitoktet blir utført som et akustisk tokt, hvor fiskemengde blir bestemt fra ekkoloddobservasjoner. Ekkoloddet observerer hele vannsøylen under båten, men gir ikke tilstrekkelig informasjon til å skille arter, og kan ikke observere fisk i «dødsonen», nær bunn. Siden torsken gyter pelagisk, er ekkoloddet likevel et godt verktøy for å mengdemåle gyteinnsiget av skrei.
Figur 2 Informasjon fra ekkolodd vises som "ekkogram". Dette eksemplet viser en strekning på 2 nautiske mil i 2020 med en rekordhøy registrering av gytende skrei i Henningsværstrømmen.
Vi er avhengig av trålprøver for å skille fisk med like ekkokarakteristikker (f.eks. torsk, hyse, og sei). Disse prøvene gir et litt selektivt utvalg av det vi observerer på ekkoloddet, og vi må regne tolkingen som sikrere når fiskeslagene ikke står blandet i sjøen. De siste år har vi sett at torsk og sei står mye blandet, og det introduserer noe mer usikkerhet enn normalt i tolkingen av akustiske data.
Akustiske observasjoner ble gjort med Simrad EK80 ekkolodd. Mengdebestemmelse ble gjort for registreringer fra 38 KHz svinger, mens andre frekvenser ble brukt som støtte til tolking (18 KHz, 120 KHz, 200 KHz og 333 KHz). LSSS (Large Scale Survey System) ble benyttet til fordeling av ekkomengder, og signalet ble tolket til de akustiske kategoriene «torsk», «hyse», «sei», «sild», «plankton» og «annen bunnfisk». Kategorien «annen bunnfisk» benyttes til all fisk som ikke passer i de andre kategoriene og kan også inneholde pelagisk fisk. Typiske for årets tokt bestod «annen bunnfisk» av øyepål, hvitting, vassild, mesopelagisk fisk og uerartene.
2.2 - Tråling
I tillegg til støtte for tolking av akustiske observasjoner, trenger vi trålprøver for å bestemme størrelsesammensetning, alderssammensetning, innblanding av kysttorsk, kjønn og grad av modning i skreiinnsiget. Som for artssammensetning må vi forholde oss til et redskap som fisker selektivt. I tillegg til må vi ofte tråle i utkanten av de største akustiske registreringene for å ta hensyn til bunnforhold og pågående fiskeaktivitet. Med tanke på gytende torsk, regner vi det som en viktig feilkilde ved bruk av trålinformasjon.
Figur 3 Kurskart med trålstasjoner.
2.3 - Håvtrekk
Eggprøvene ble tatt med en T-80 egghåv (Maskevidde 380 μm, diameter 80 cm). Håven ble senket til 100 M dyp (hvis grunnere, 5 m over bunnen) og trukket vertikalt opp med hastighet 0.5 m/s. I tillegg tok vi på hver stasjon også et trekk (samme dyp som T-80) med standard WP2 håv med 180 µm maskevidde. Denne prøven ble fiksert på sjøvann tilsatt borax (buffer) og 4 % formaldehyd (fikseringsmiddel), og ble satt på lager for senere planktonopparbeiding.
Eggprøven fra T-80 egghåven ble umiddelbart opparbeidet levende under stereomikroskop med hensyn på fiskeegg. Alle fiskeeggene ble sortert ut og deretter fotografert. Basert på fotografiene ble alle eggene størrelsesmålt og delt inn i fire kategorier, egg med utseende som torskegg, egg med fettdråpe, egg med stort perivitellint rom (vanligvis gapeflyndre), og andre egg. Alle torskelignende egg med diameter på mellom 1.2 og 1.6 mm ble som for tidligere tokt tolket som torskeegg. Selv om flere arter har egg som tilfredsstiller disse kriteriene indikerer tidligere resultater samt registreringer av fisk underveis at mesteparten av disse eggene faktisk var fra torsk. Genetiske undersøkelser av egg fra skreitoktet i 2022 viste liten forveksling med andre arter (Fuglebakk et al., 2024). En mindre innblanding av hyse kan det nok likevel ha vært. Etter fotografering ble eggene konservert på etanol slik at de senere kan artsidentifiseres ved hjelp av DNA teknikker.
I tillegg til inndeling i artstyper, ble eggene også stadiebestemt. Stadiene som ble brukt er i henhold til Fridgeirsson (1978). For torsk i toktområdet er den totale varigheten av eggstadiet typisk 2-3 uker, alt etter temperatur.
Når eggene var talt opp og kategorisert ble eggmengdene omregnet til antall/m2 overflate. Det kalkulerte eggtallet er bare helt riktig om man regner filtreringen for 100 % effektiv (dvs at håven ikke skyver vann foran seg) og at trekket er absolutt vertikalt. I virkeligheten er filtreringseffekten noe lavere og opptrekket aldri helt vertikalt. Disse feilkildene var nok likevel forholdsvis små under våre forhold; vi observerte aldri klogging av håven slik man kan oppleve med store planktonmengder, og visningen på wiren var beskjeden. Dette er ellers samme metodikk som har blitt brukt også på våre tidligere eggtokt i Lofoten og resultatene er derfor sammenlignbare.
Figur 4 Kurskart med håvtrekk.
2.4 - Hydrografi
Temperatur og saltholdighet (salinitet) kartlegges med en CTD-sonde (Conductivity, Temperature, Salinity) som føres vertikalt gjennom vannsøylen. Dette gir en beskrivelse av vannlag og gir oss muligheten til å overvåke temperaturen i dypet hvor torsken vandrer eller gyter og hvordan temperaturforholdene endrer seg mellom områder og mellom år.
Figur 5 Kurskart med CTD stasjoner.
3 - Beregninger
Beregningsmetodikken for fiskemengdeindekser er beskrevet i Korsbrekke (1997), og er kun kort oppsummert under, likelydende som i rapporten for 2023. De mest sentrale tabellene er gjengitt i vedlegg.
Fra de tolkede akustiske kursene utleder vi for hver nautisk mil en tilbakespredningskoeffisient som representerer gjennomsnittlig ekko fra torsk i arealet dekket av ekkoloddet. Disse summeres og multipliseres med avstand mellom kursene. Alternativt kan man se på det som å regne ut gjennomsnittlig ekko fra torsk per areal og multiplisere med arealet for området. Vektet lengdefordeling for utvalgte trålhal blir kombinert med informasjon om hvor mye ekko en fisk av en gitt lengde gir, slik at vi kan estimere totalt antall torsk i hver lengdegruppe og for hvert område.
Ettersom det oftest ikke lar seg gjøre å tråle i de tetteste registreringene, venter vi ikke at trålhalene gir et riktig bilde av fordelingen mellom kysttorsk og skrei. De tetteste konsentrasjonene er det rimelig å anta at er ren skrei, og en pragmatisk fremgangsmåte er etablert for å gi rimelig anslag på fordelingen mellom kysttorsk og skrei, og for å gi en rimelig vekting av tilhørende biologiske parametere (lengde, vekt, alder og modning). Denne er forklart i Korsbrekke og Thorsen (2020), og gir et estimat på totalt antall skrei i hver lengdegruppe og for hvert område, og eventuelle andre parametere innad i hver lengdegruppe (slik som alder, kjønn og modning).
Eggmengdeindeksen ble beregnet som summen av eggmengden fra tre ulike strata: Vestfjorden, Ytre Sør og Ytre Nord. Hvert av disse strataene har sin egen tetthet av hovstasjoner, som i hovedsak er den samme fra år til år. Innen hvert stratum ble gjennomsnittlig tetthet av egg per stasjon beregnet, og deretter multiplisert med stratumets areal. For å sikre sammenlignbarhet mellom år, ble eventuelt manglende stasjoner ekstrapolert basert på nabostasjonene. Eggmengden fra 2016 ble brukt som referanseverdi for eggindeksen, dvs 100 %, og alle andre eggindekser ble beregnet ut fra denne. For eggindeksene ble forekomsten av alle eggstadier inkludert.
4 - Resultater
4.1 - Observasjoner av skrei
4.1.1 - Årets tokt
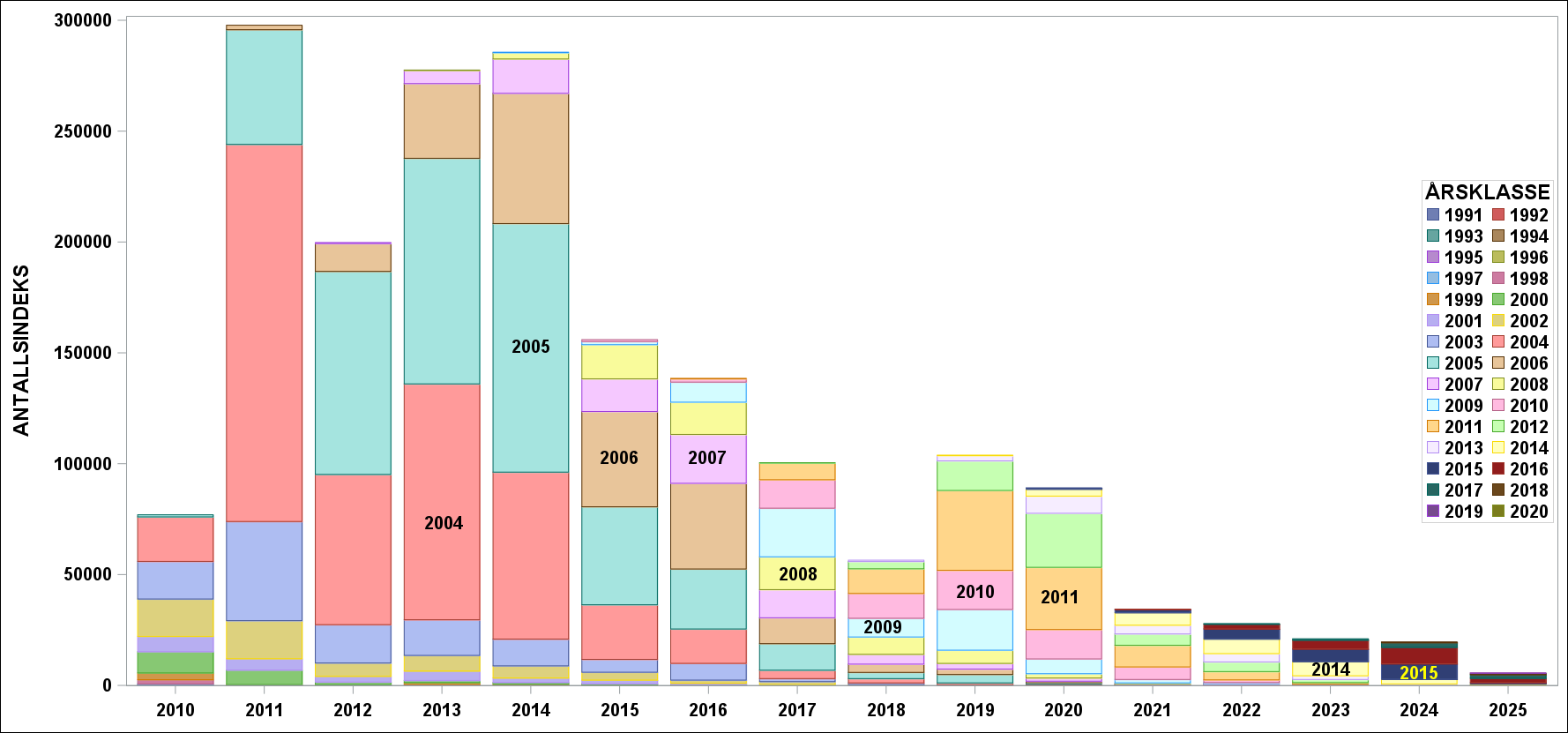
Det viktigste resultatet fra skreitoktet er det vi omtaler som antallsindekser på alder for skrei. Disse inngår i en tidsserie av toktresultater og er vist i vedlegg (Kapittel 7) til rapporten (2010-2025).
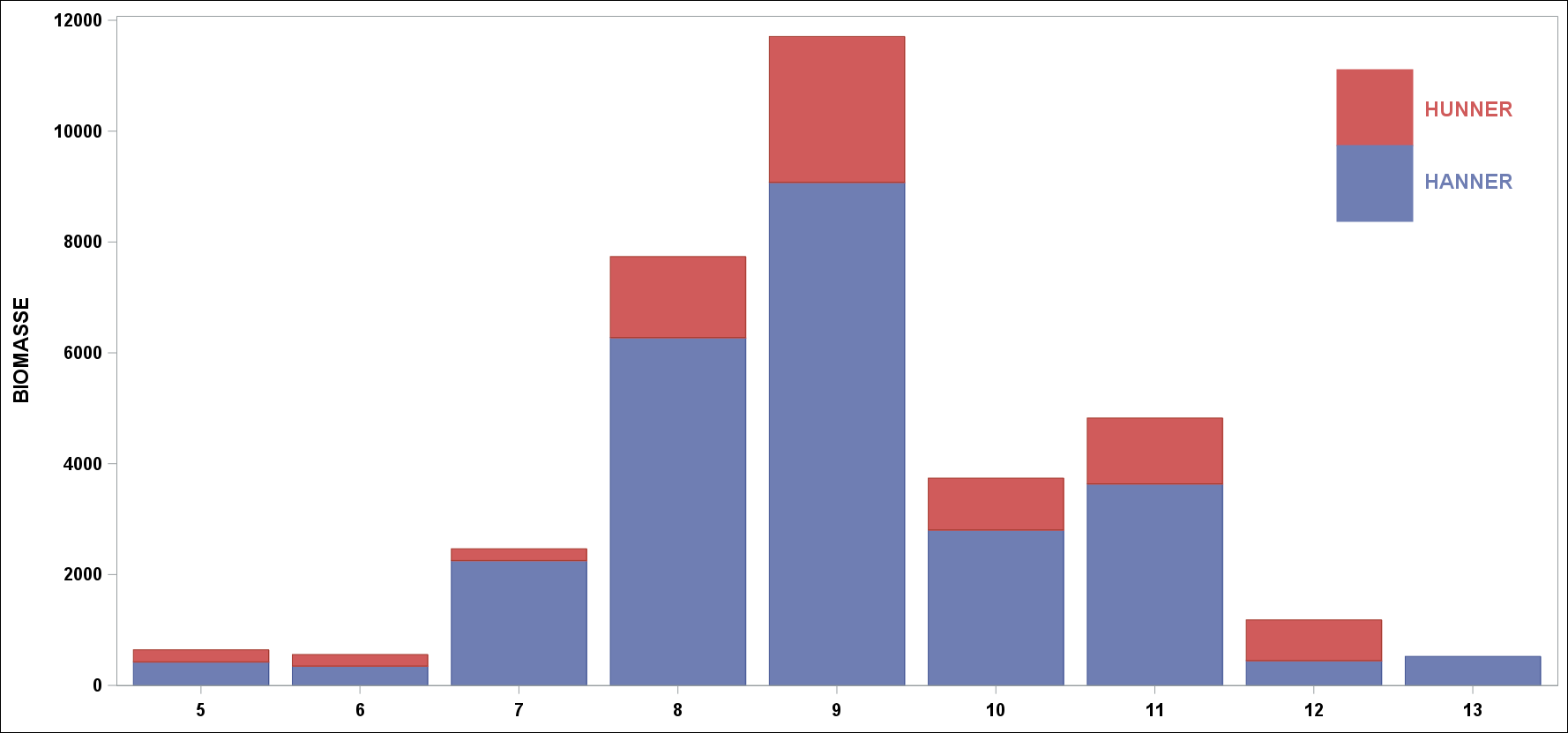
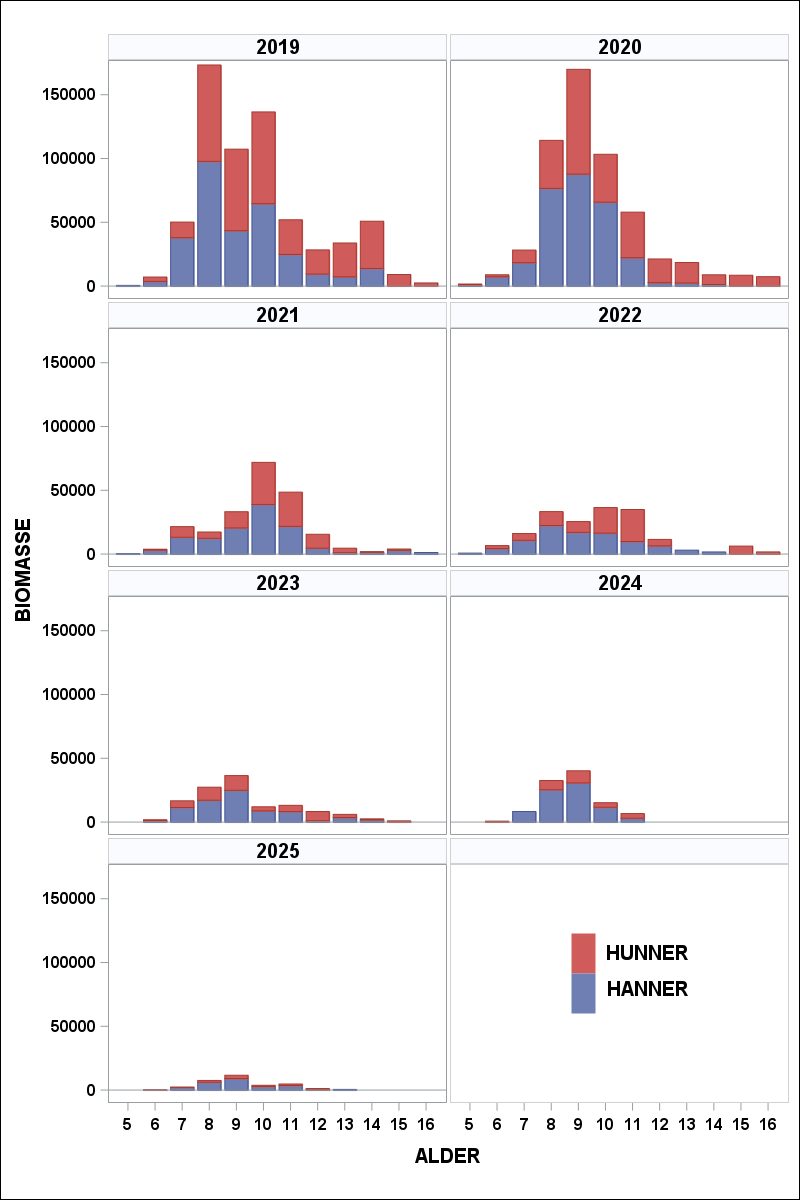
Toktresultatene inneholder også informasjon om størrelse på fisken, hvor langt den har kommet i gytingen og om fisken har gytt tidligere. Gjennomsnittlig vekt ved alder kan anvendes på antall fisk i en aldersgruppe og vi kan regne ut en totalvekt (biomasse). Dette gir en god pekepinn på hvilke årsklasser som bidrar mest i fisket. Følgende figur viser hvilke mengder som ble observert for hver aldersgruppe. Legg merke til at andelen hunnfisk er omlag 20% ( Figur 6 ). Lave andeler med hunnfisk er noe som vanligvis kan observeres etter perioder med forhøyet fiskepress.
Figur 6 Mengdeindekser (vekt) fordelt på alder og kjønn.
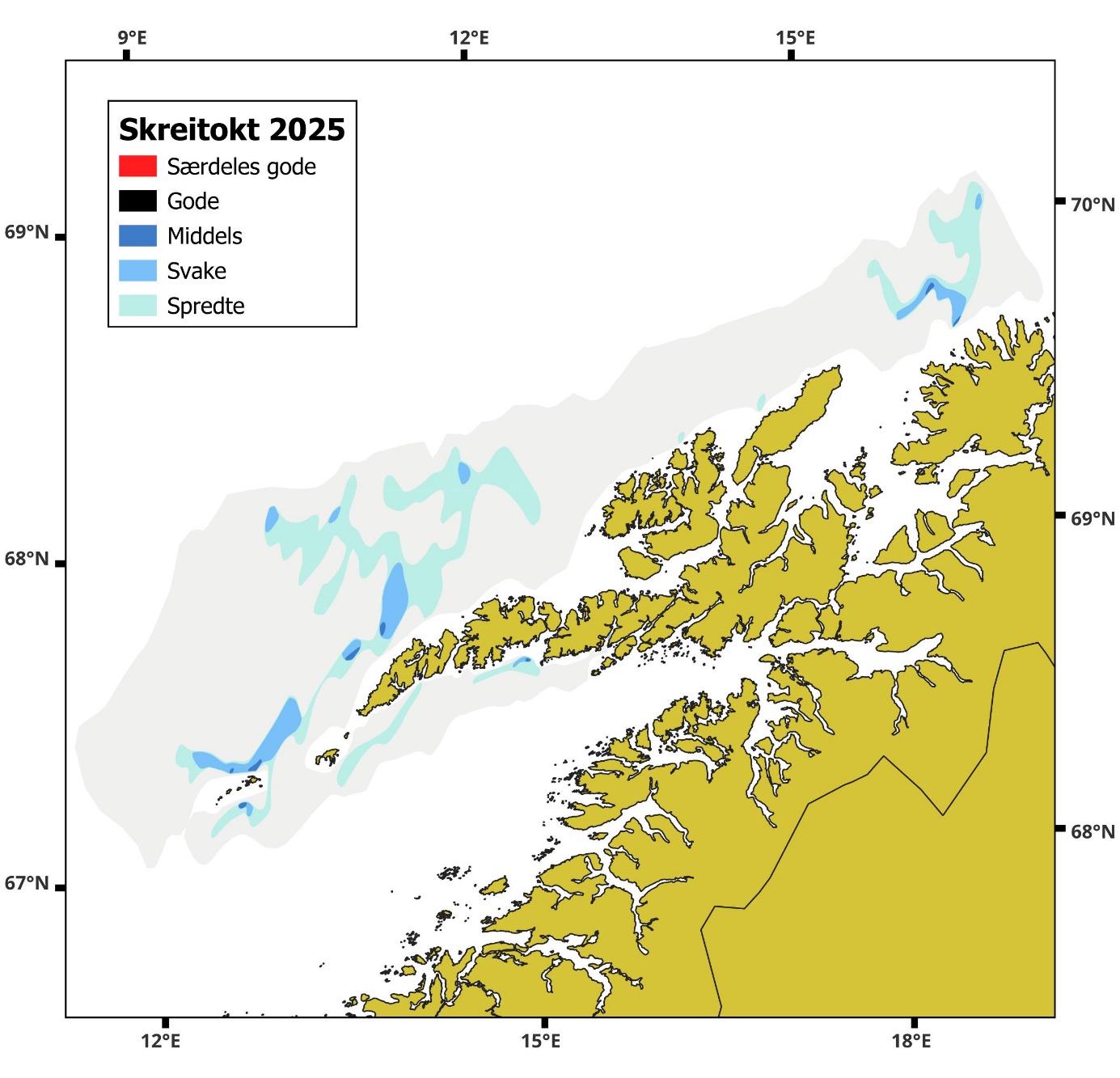
Områdene med registreringer av skrei var av svært begrenset omfang og bestod i all hovedsak av spredte og svake registreringer med noen få flekker med middels registrering. Mengden skrei i toktområdet var ikke mer enn 2,1% av rekordåret 2014.
Geografisk fordeling og tetthet i registreringene er vist i Figur 7 .
Figur 7 Fordeling av akustiske registreringer, skreikart.
Havforskningsinstituttet mottar noen henvendelser om toktdekning av mer nordlige gyteområder. Årets fiske var stort sett konsentrert i områdene nord for toktområdet og bestod av fisk fra lokale gytefelt, men ikke minst fra fisk som var på vandring sørover. Vandringsrutene til skrei varierer fra år til år og vi kjenner ikke de mekanismene som har innvirkning på valg av dyp for vandring mot gytefeltene. Havforskningsinstituttet dekket kjente gytefelt for skrei nord for toktområdet i 2023 med et ekstra fartøy (F/F Kristine Bonnevie). Dekningsområdet strekte seg forbi Nordkapp til Nordkinnhalvøya. Mengdene som ble registrert i 2023 representerte 13% av de samlede registreringene (begge toktene).
Skreitoktet vil utvide sitt dekningsområde fra 2026.
4.1.2 - Tidsserien av mengdeobservasjoner
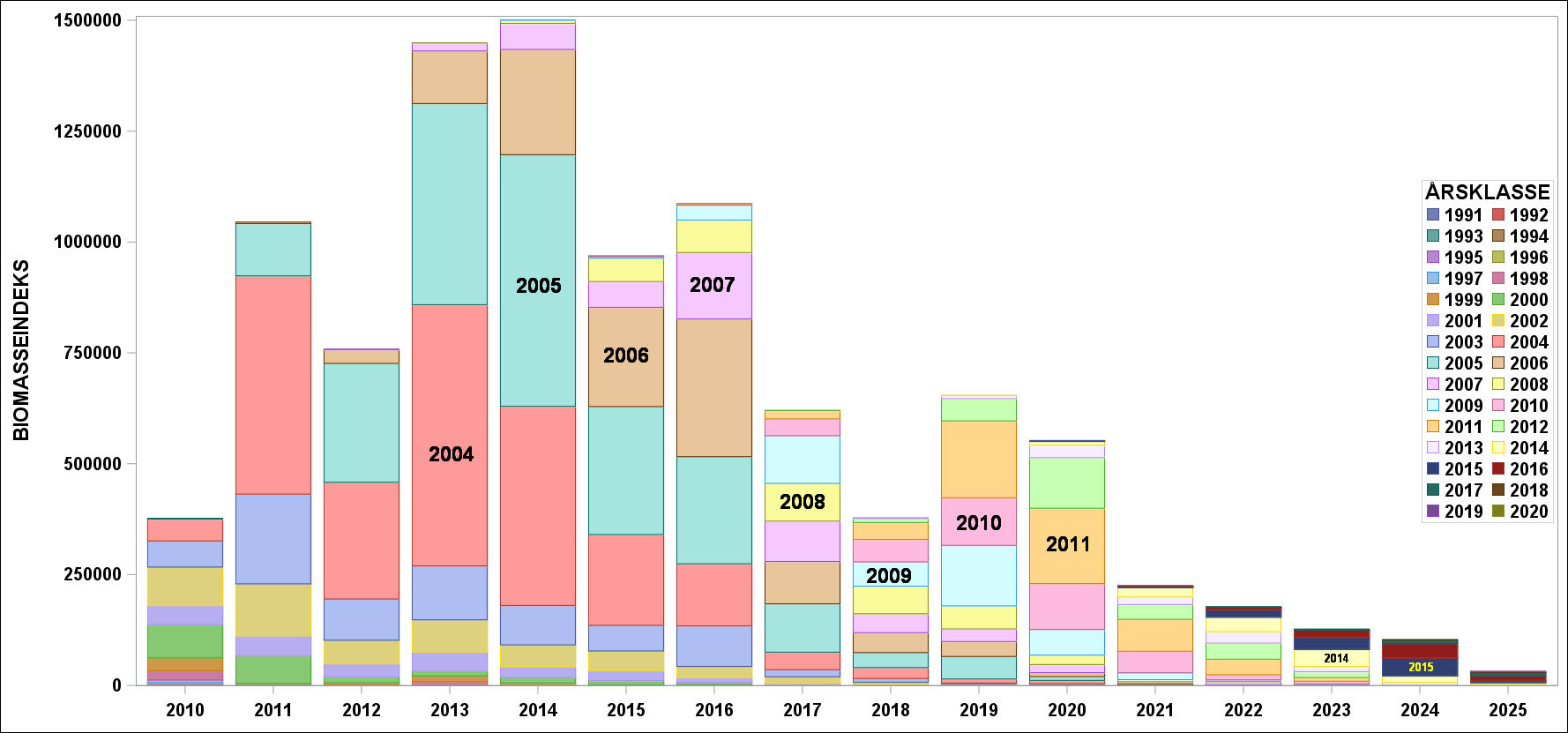
Årets innsig er det dårligste siden 1988 og det er 9 år gammel fisk som bidrar mest. Sett over tid er årets resultat overraskende svakt ( Figur 8 og Figur 9 ).
Figur 8 Biomasse (øverst) og antallsindekser (nederst)r indekser for perioden 2010-2025. Fargene representerer forskjellige årsklasser.
Figur 9 Aldersfordeling fra 2019 til 2025.
4.1.3 - Biologiske egenskaper over tid
Med biologiske egenskaper mener vi i denne sammenheng slikt som gjennomsnittlig lengde eller vekt ved alder, andeler førstegangsgytere, andeler hunnfisk og spesielle størrelser som gjennomsnittlig rognvekt som prosent av fiskens totalvekt og gjennomsnittlig levervekt gitt som en prosent.
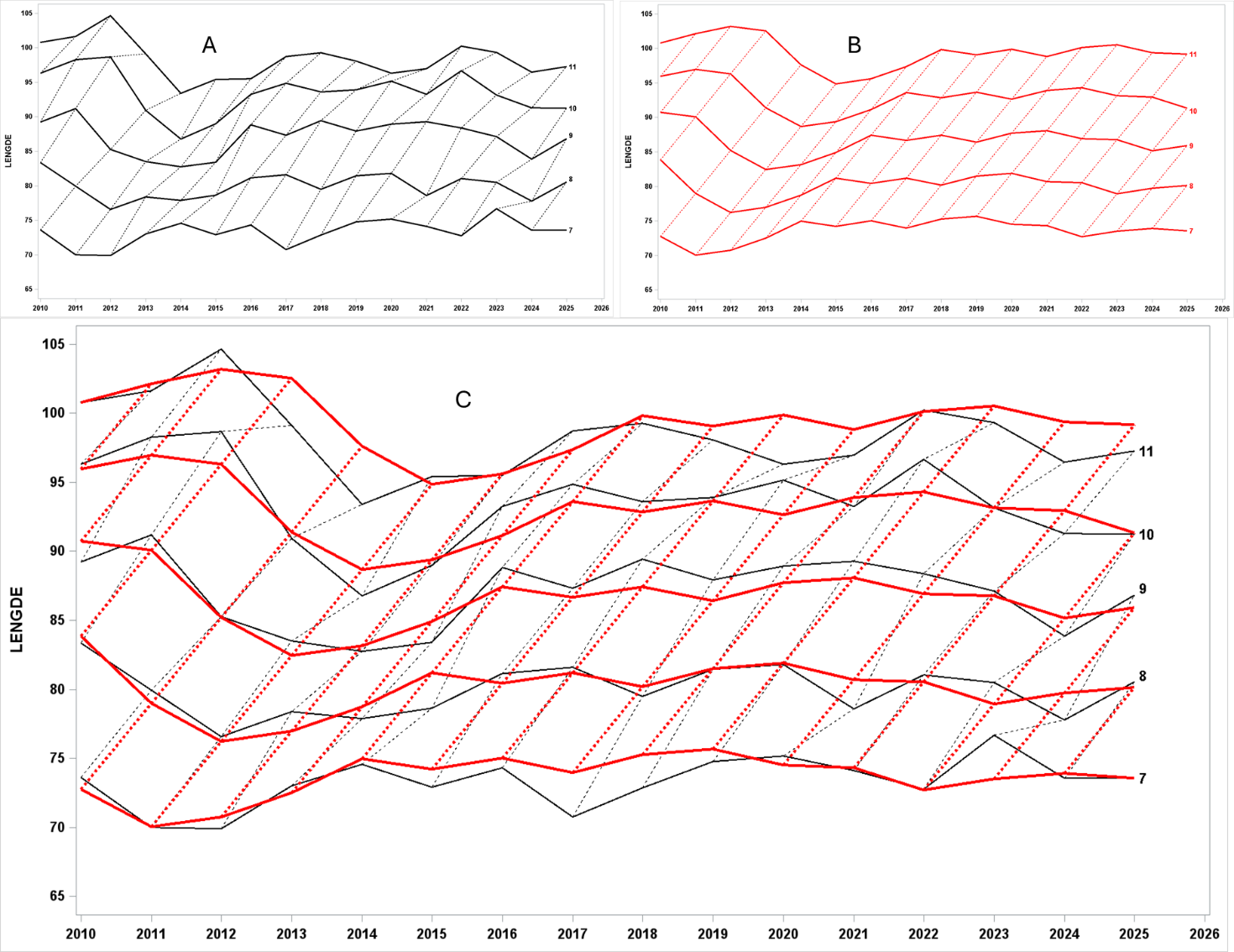
I Figur 10 vises gjennomsnittlig lengde ved alder i to utgaver. I svart (A) vises beregnet gjennomsnitt over tid, mens i rødt (B) vises en statistisk modell av det samme. Tokt er basert på stikkprøver og har derfor noe usikkerhet. Oppe til høyre (B) vises modellresultater og i denne modellen benyttes en antagelse om at veksten fra alder 7 til 11 er konstant og at all variasjon i vekst skjer før alder 7. Nederst (C) er figurene lagt oppå hverandre og man kan se hvor godt det passer. Tilpasningen blir for grov, men er «slettes ikke så verst». B viser oss ganske tydelig at det er årsklassene 2004 og 2005 som har hatt dårligst vekst i ung alder. Disse årsklassene er de sterkeste vi har observert i våre tokt og en slik reduksjon omtales ofte som en tetthetsavhengig reduksjon i vekst selv om flere faktorer kan spille inn. Fra plottene ser vi også at veksthastigheten for disse to årsklassene som modne var like god som for andre årsklasser.
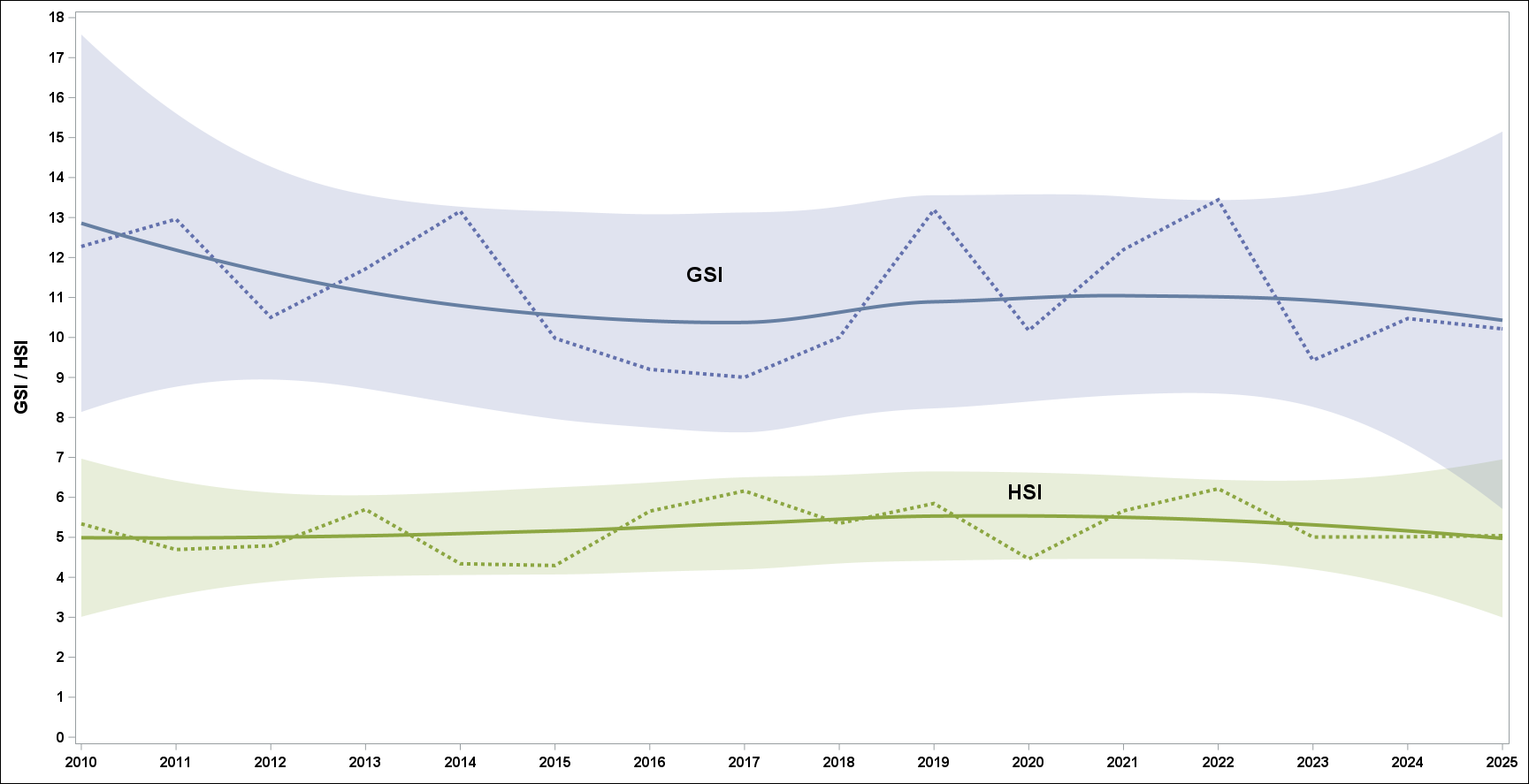
I Figur 11 vises variasjon i rognvekt og levervekt.
Figur 10 Her viser fiskens lengde ved alder. Heltrukken linje følger samme alder over tid, mens stiplede linjer følger den enkelte årsklasse. I figur A vises gjennomsnittlig lengde ved alder fra serien av skreitokt. Oppe til høyre (B) vises modellresultater i rødt. Nederst (C) er figurene lagt oppå hverandre og man kan se hvor godt det passer.
Figur 11 GSI står for Gonadosomatic Index, mens HSI er en forkortelse for Hepatosomatic Index. På norsk blir det rognindeks og leverindeks og regnes ut som vekt av rogn eller lever som en prosent av totalvekt. Her vises gjennomsnittlige indekser over år. Rogn og levermengde avhenger i stor grad av mattilbudet torsken har i Barentshavet. Mer mat medfører økt vekst og tidligere modning og mer rogn og større lever for fisk som allerede er modne. Store torsk er effektive predatorer og vil vanligvis ha en høyere rogn og leverindeks enn yngre fisk i gytebestanden. Det betyr at den variasjonen vi ser i figuren skyldes variasjon i mattilbud og av alderssammensetning i gytebestand.
4.2 - Observasjoner av egg
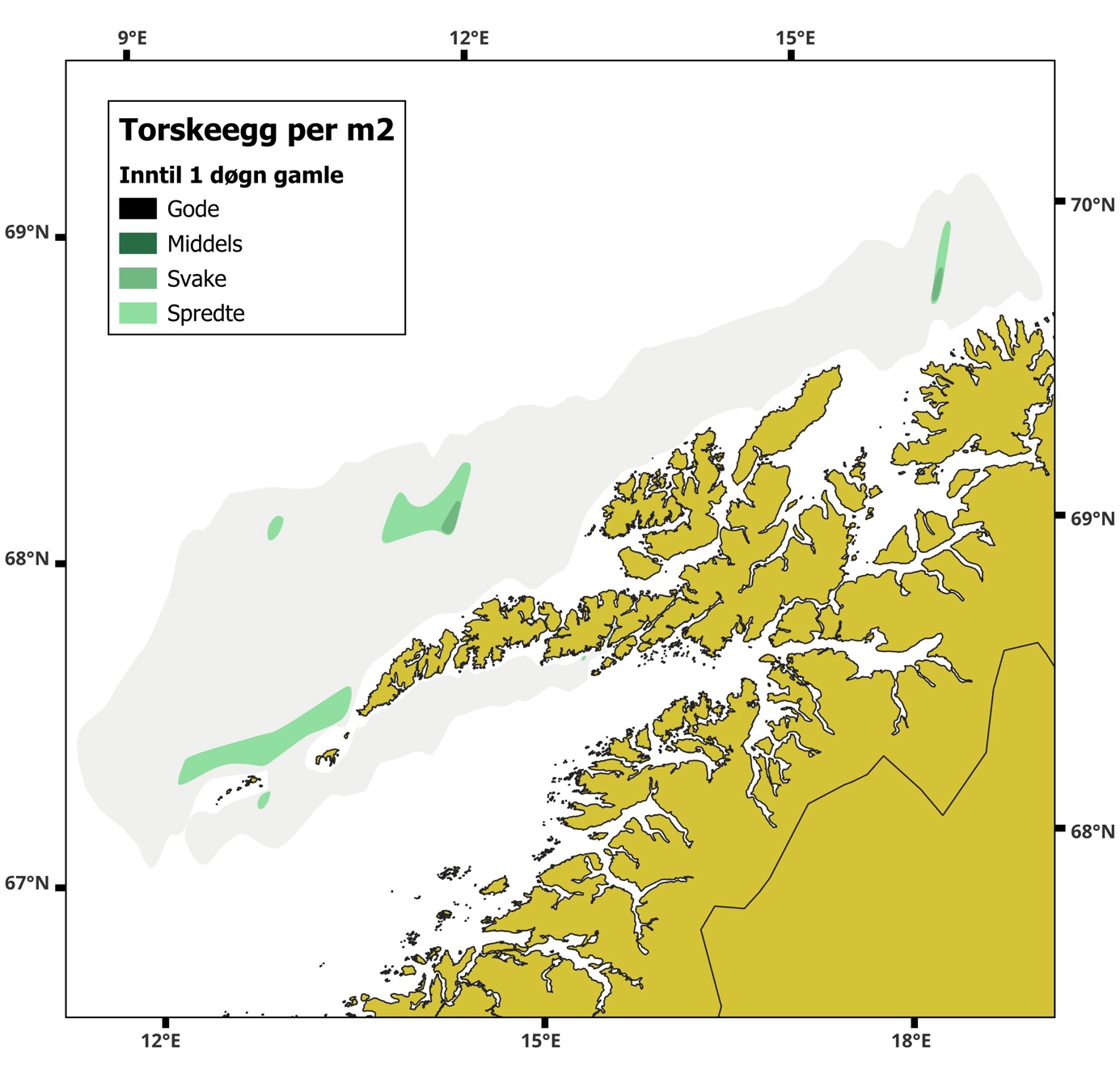
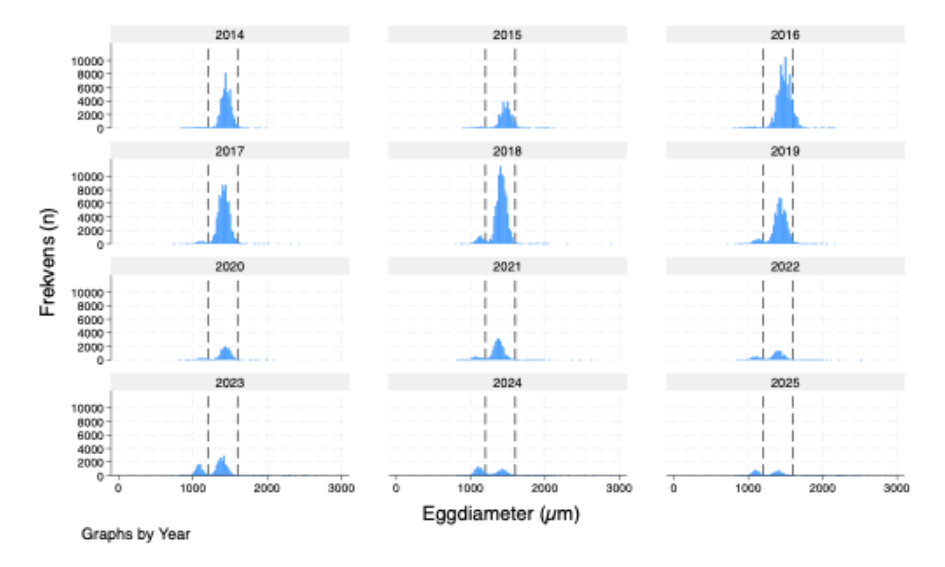
Figur 12 Utbredelse av torskeegg basert på hovtrekk. Egg inntil 1 dag gamle.
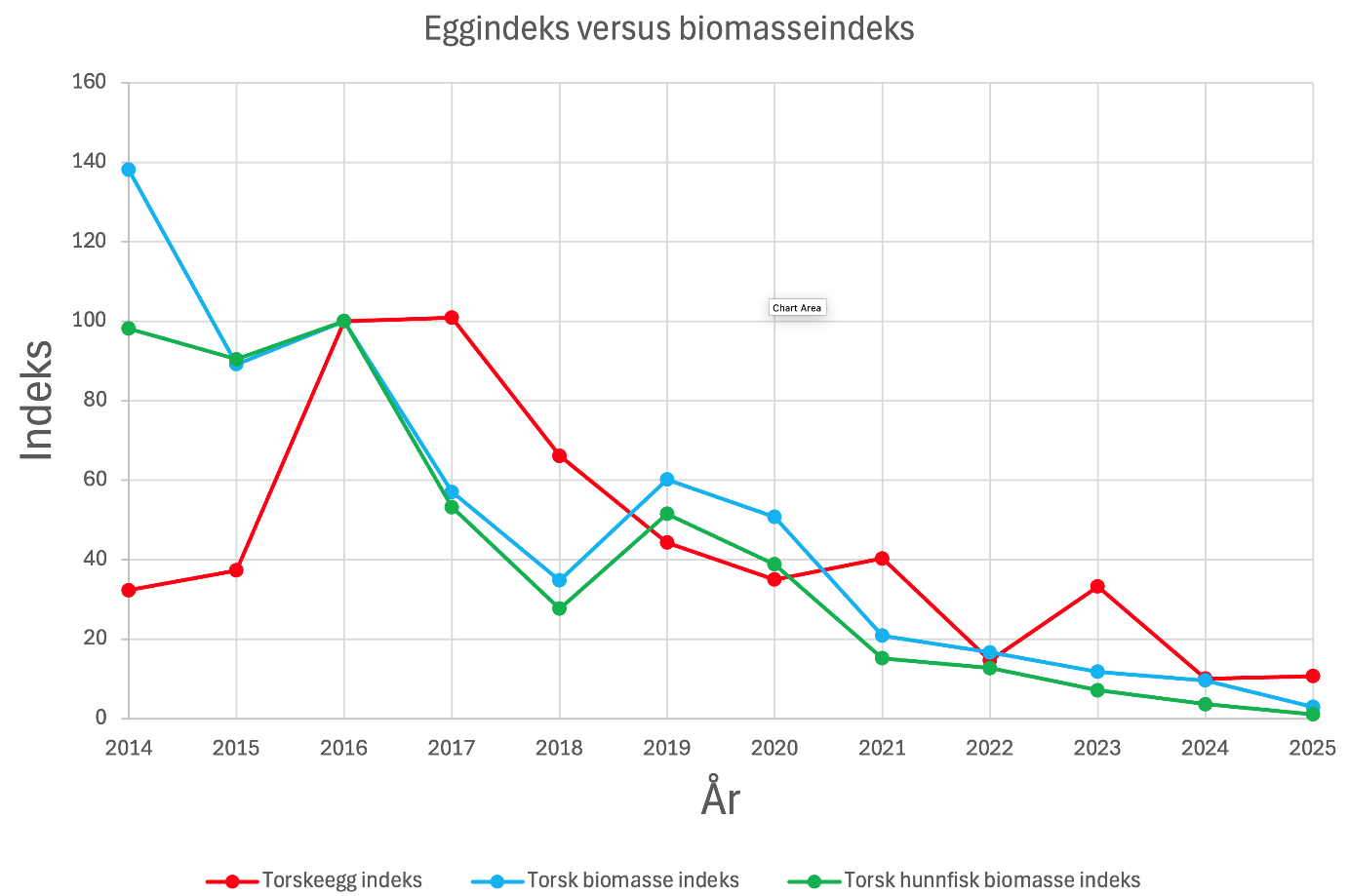
Skreikartene viser, som vanlig, at egg i tidlige utviklingsstadier ( Figur 12 ) hovedsakelig forekommer i rimelig nærhet til områder med høye konsentrasjoner av gytefisk. Områder med lite torsk viser tilsvarende lav tetthet av inntil ett døgn gamle egg ( Figur 12 ). Som i de senere årene følger også eggmengdeindeksen fiskemengdeindeksen over tid ( Figur 13 ).
Figur 13 Indeks for torskeegg, torsk biomasse, og torsk biomasse hunnfisk for årene 2014-2025.
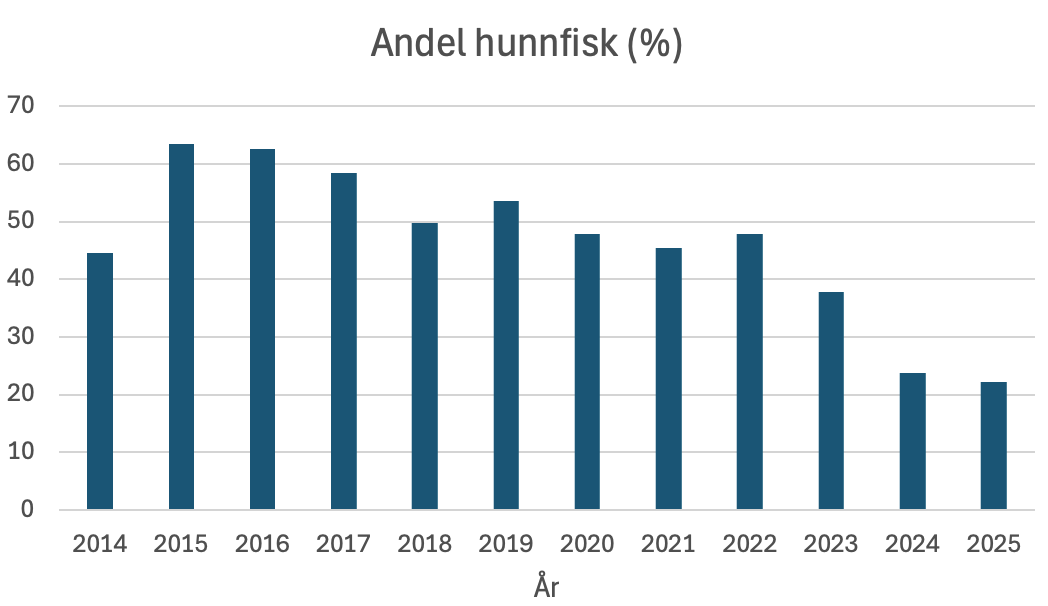
Figur 14 Andel hunnfisk (vektandel) i perioden 2014 til 2025 beregnet ut fra trålfangster.
Indeksene er her standardisert for å vise variasjon mellom år. Mengdene av fisk og egg i 2016 er brukt som referanseverdier (100 %), og for alle andre år er prosentindekser beregnet i forhold til 2016. Avvikene i 2014 og 2015 har vi ingen sikker forklaring på, men de kan tenkes å skyldes variasjon i kjønnsfordeling i skreiinnsiget, ulik grad av modning, fekunditet, samt forskjeller i tidspunkt for gytestart og gyteforløp i forhold til toktperioden. Vi anser det som lite sannsynlig at avvikene skyldes feil i artsbestemmelse av egg, ettersom genetiske analyser har vist godt samsvar med mikroskopisk identifisering (Fuglebakk et al. 2024). En eventuell innblanding av forvekslingsarter vil dessuten sannsynligvis bidra til en overestimering, snarere enn en underestimering, av eggtettheten.
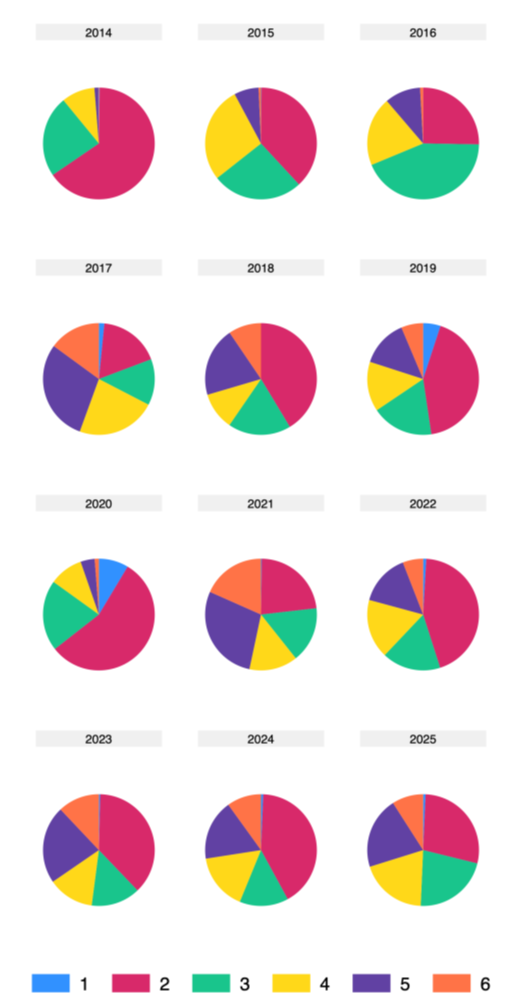
Figur 15 Fordeling av eggstadier (Fridgeirsson, 1978) i årene fra 2014 til 2025.
Endringer i gytestart og gyteforløp mellom år kan vurderes ut fra fordelingen av eggstadier i prøvene (Figur 15). Ved temperaturer på 3–6 °C, som er typiske for toktområdet, har torskeegg en utviklingstid på om lag 14–21 dager. Resultatene viser at andelen gamle eggstadier (stadium 5–6) var svært lav i 2014, mens andelen nygytte egg (stadium 1-2) var tilsvarende høy. Dette indikerer at gytingen nettopp hadde startet, noe som sannsynligvis påvirket eggmengdeindeksen for dette året, ettersom indeksen beregnes fra summen av alle utviklingsstadier. En lignende, men mindre uttalt, situasjon ble observert i 2015. I de øvrige årene varierer fordelingen av stadier noe, men bare i 2020 ser vi et mønster som minner om 2014–2015. Likevel avviker ikke eggindeksen for 2020 særlig fra fiskemengdeindeksen.
Andelen hunnfisk i fangstene varierte i perioden 2014–2022 mellom 45 % og 64 %. Fra og med 2023 har imidlertid andelen sunket betydelig, og i 2025 utgjorde hunnfisk bare 22 % av totalfangsten ( Figur 14 ). Det er likevel ingen tydelige indikasjoner på at variasjon i hunnfiskandel kan forklare avvikene mellom fiskeindeksen og eggindeksen ( Figur 13 ).
Tidspunktet for start av egginnsamlingen har variert noe mellom år, men har normalt vært omkring 22. mars. I 2014 og 2024 startet innsamlingen tidligere, 17. mars, mens den i 2020 ble forsinket til 28. mars på grunn av COVID-19-restriksjoner. Toktet i 2020 skilte seg også fra øvrige år ved at det ble gjennomført fra sør mot nord, i stedet for motsatt retning. Dette året ble Vestfjorden, som hadde mye gytende torsk, derfor dekket relativt tidlig, selv om toktstart var sen.
Sammenligning av eggstørrelsesfordeling mellom år ( Figur 16 ) viser at i år med mye torskeegg er fordelingen dominert av egg i typisk torskestørrelse (1,2–1,6 mm). I de senere årene har imidlertid mengden torskeegg blitt betydelig redusert, mens mengden små egg har økt – både relativt og delvis også i absolutte tall. De små eggene er sannsynligvis hovedsakelig fra øyepål, noe som ble bekreftet ved DNA-analyse i 2022. Øyepål er et vanlig byttedyr for torsk, og når mengden torsk går ned, er det derfor sannsynlig at bestanden av øyepål øker.
Figur 16 Egg størrelseshistogram for perioden 2014 til 2025. Stiplede vertikale linjer indikerer størrelsesområdet for torskeegg.
4.3 - Hydrografi
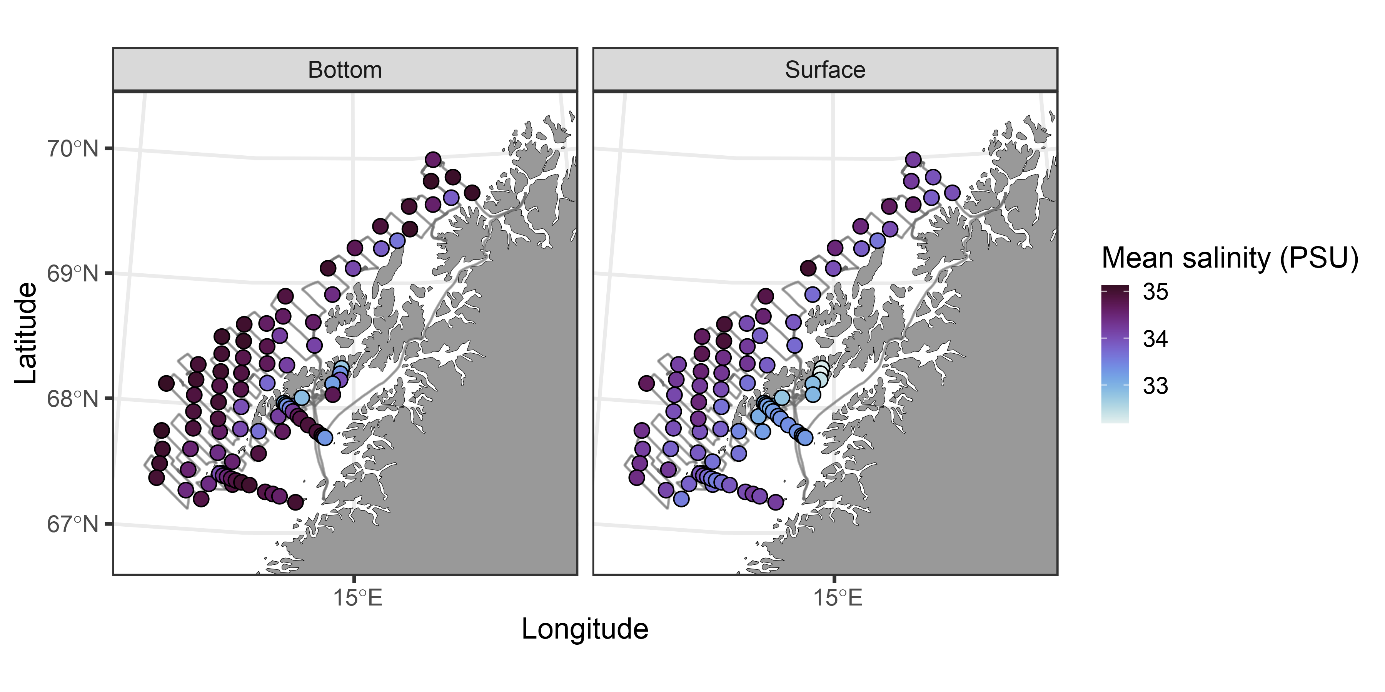
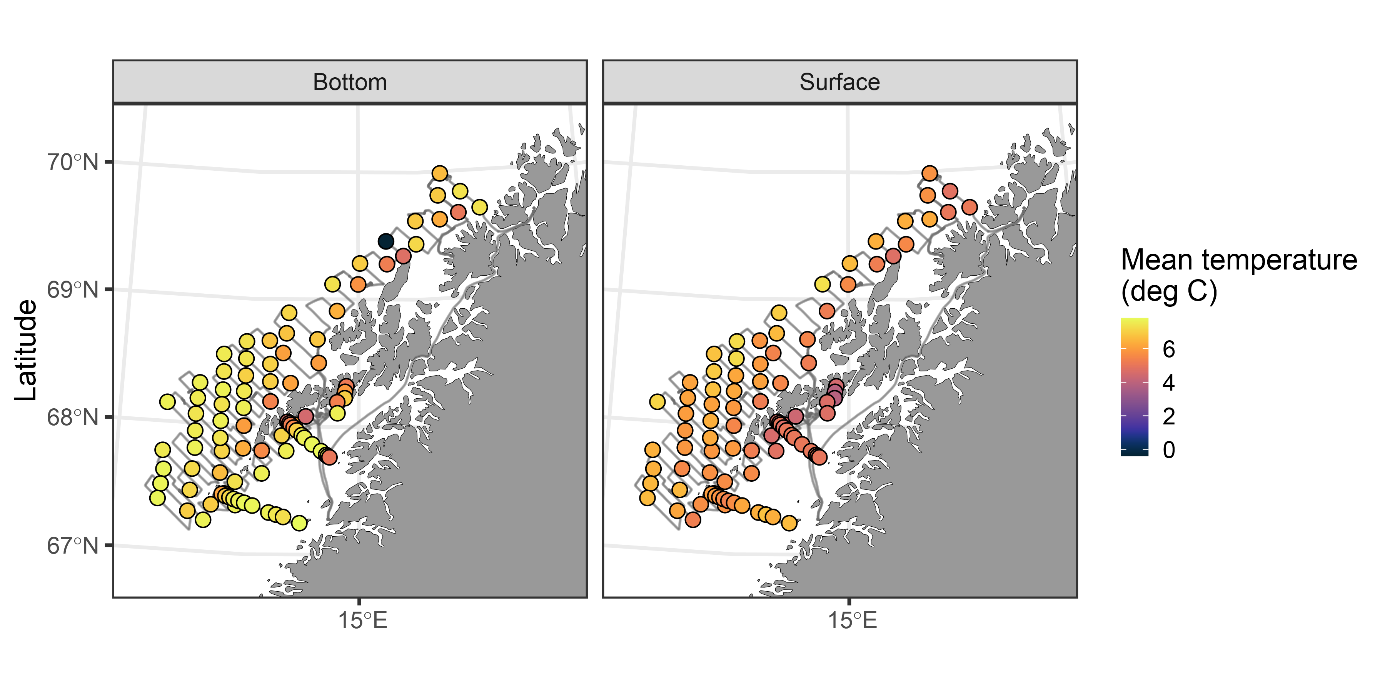
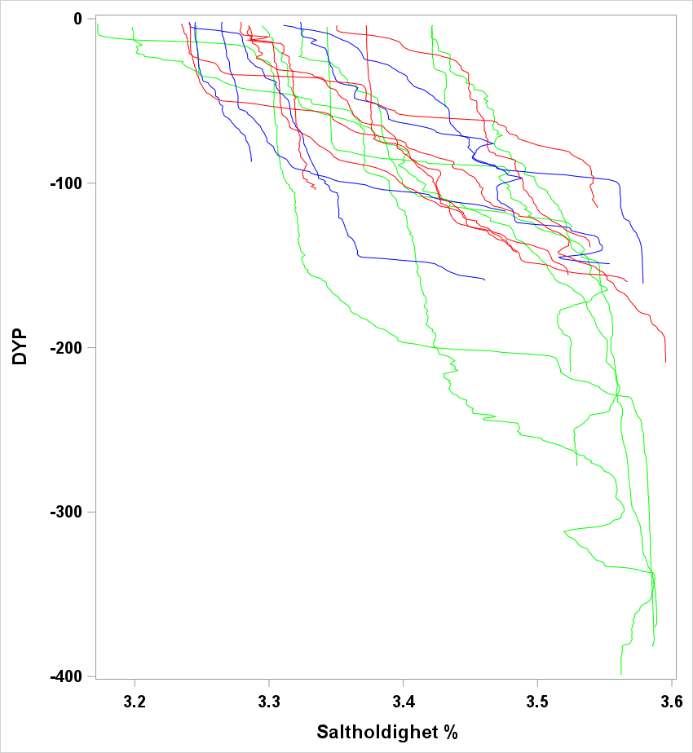
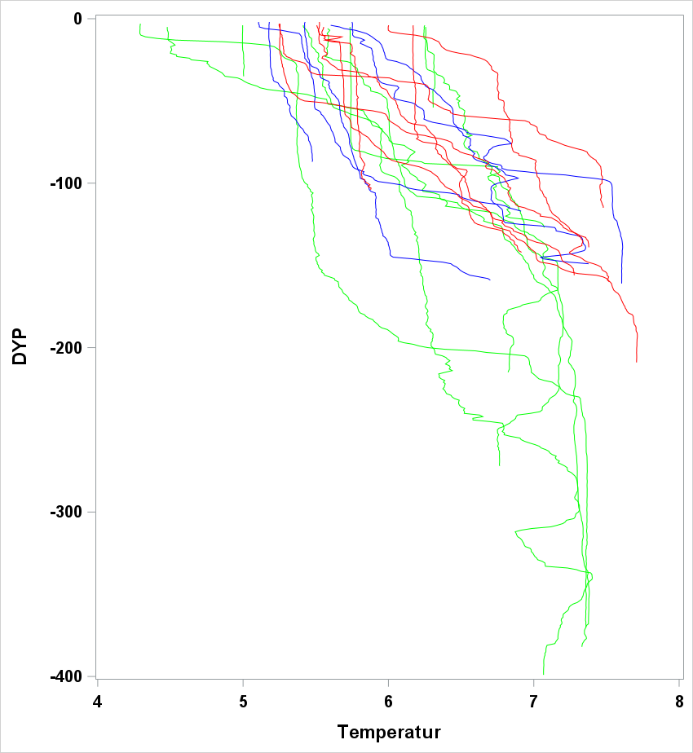
Figur 17 Saltholdighet og temperatur ved bunn og nær overflaten.
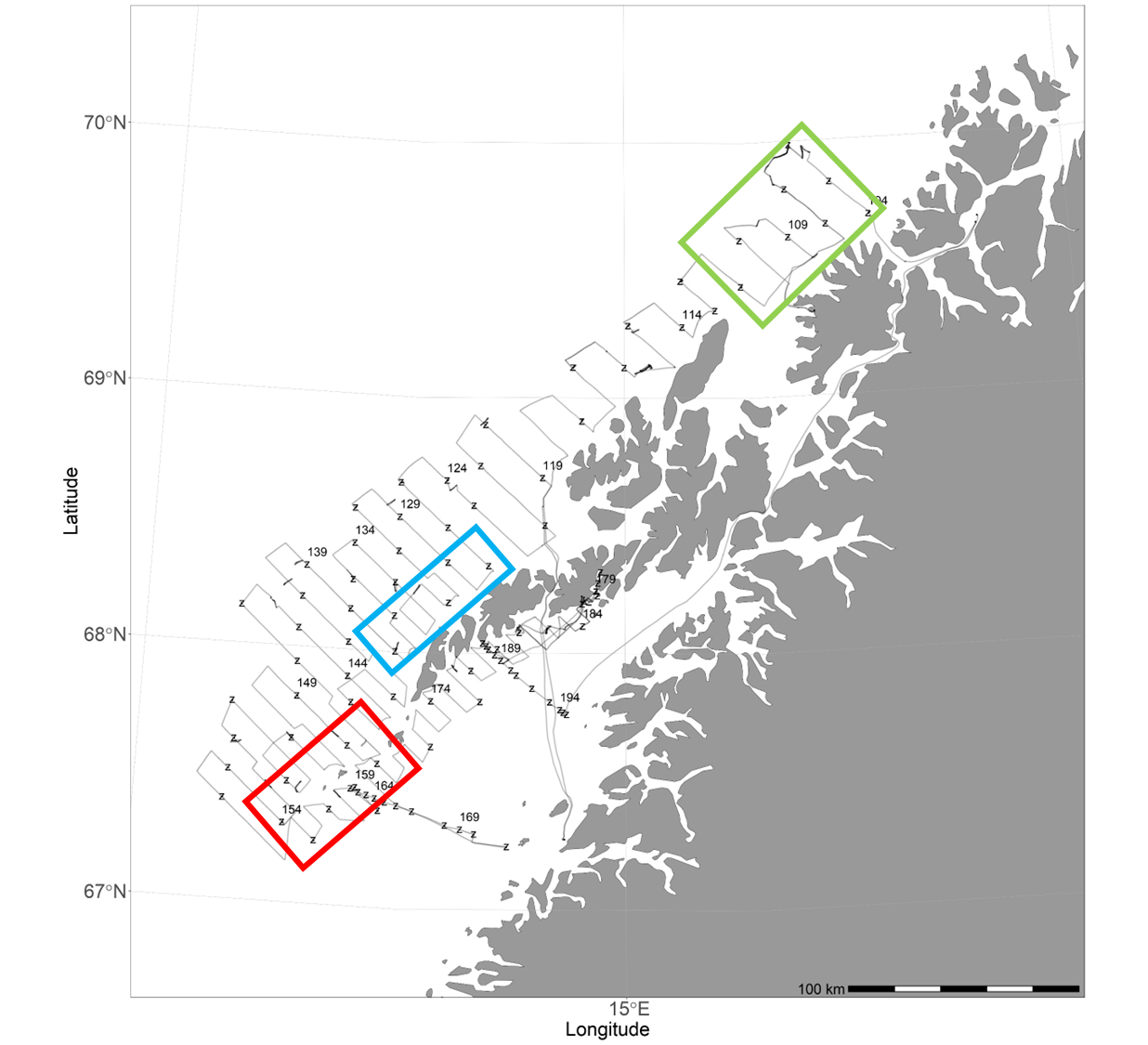
Figur 18 Kart som markerer de tre områdene blir vist i Figur 19 .
Observasjoner av saltholdighet og temperatur ved bunn og i overflaten er vist i Figur 17 . Temperaturene i overflaten er høye sammenlignet med historiske observasjoner. De hydrografiske forholdene ved bunn og i det meste av vannsøylen er typiske for dypere Atlanterhavsvann. Overgangen mellom det kaldere vannet nær overflaten (kyststrømmen) kalles overgangslaget og viser stor variasjon mellom områder og også innenfor områder. Ingen av kurvene viser et skarpt skille mellom lagene. Dette har vi illustrert i Figur 19 og fargekodene representerer de områdene som ble benyttet ( Figur 18 ).
Figur 19 Saltholdighet og temperaturprofil for noen utvalgte CTD stasjoner. De tre områdene som er valgt i Figur 18 er vist med tilsvarende strekfarger her.
5 - Oppsummering
Skreitoktet 2025 ble avkortet i forhold til plan. Dekningen har likevel vært tilfredsstillende. Vi begrunner det med at det ikke foregikk noe fiske i det udekkede området og eggprøvene nedstrøms viste at det var kun minimal gyting i dette området. Toktet fikk noe knapt med biologiske prøver, men den økte usikkerheten er håndterlig. Alderssammensetningen er som forventet, mens total mengde er betydelig lavere enn hva som var forventet ut ifra bestandsberegningene. De hydrografiske forholdene var preget av et mindre tydelig overgangslag (overgangslaget skiller det kalde overflatevannet i kyststrømmen fra det varmere og saltere atlanterhavsvannet dypere) og forholdene var gunstige for gyting på alle tradisjonelle gytefelt.
Toktresultater gitt som tidsserier er vist i eget Vedlegg . Der finnes også mer detaljerte resultater fra årets tokt.
6 - Referanser
Korsbrekke, Knut. 1997. Norwegian acoustic survey of North East Arctic Cod on the spawning grounds off Lofoten. ICES. C.M. 1997/Y:18 (https://imr.brage.unit.no/imr-xmlui/handle/11250/105785)
Korsbrekke, Knut og Thorsen Anders. 2021. Skreitokt 2020. Rapport fra havforskningen 2020-32 ISSN: 1893- 4536
Fra Havforskningsinstituttets Kvalitetsportal (intern): «Overvåkningstokt – Skreitokt» https://hi.dkhosting.no/docs/pub/dok06743.htm)
Fra Havforskningsinstituttets Kvalitetsportal (intern): «Håndbok for prøvetaking av fisk, krepsdyr og andre evertebrater»
(https://hi.dkhosting.no/docs/pub/DOK05957.pdf)
Fuglebakk, Edvin; Thorsen, Anders; Mellerud, Ida Kristin; Sodeland, Marte og Sannæs Hanne. Genetisk analyse av skreitoktets eggsurvey 2022. Rapport fra Havforskningen 2024-19 ISSN: 1893-4536 (https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-2024-19)
Fridgeirsson, E. (1978) Embryonic Development of Five Species of Gadoid Fishes in Icelandic Waters. Rit Fiskideildar 5 - Nr. 6: 1–68.
Korsbrekke, Knut. Skreitokt nord – Kartlegging av gytebestanden av skrei nord for Vesterålsbankene 2023. Toktrapport 2024-1 ISSN: 1503-6294 (https:// www.hi.no/hi/nettrapporter/toktrapport-2024-1)
7 - Vedlegg
År
a5
a6
a7
a8
a9
a10
a11
a12
a13
a14
a15
a16
a17
a18
a19
2010
956
20056
16977
16839
6886
9605
3046
1908
357
55
277
2011
2014
51730
170085
44722
17155
5115
6544
165
210
21
2012
458
12559
91582
67752
17304
5982
2585
1105
280
14
136
2013
220
5888
33691
101759
106390
16082
7047
4240
959
752
501
33
2014
250
2816
15493
58750
112101
75325
12065
5463
1976
737
640
2015
873
1401
15418
14734
42982
44198
24618
5792
3540
1587
684
143
2016
244
1459
9050
14533
22060
38650
27063
15497
7553
1356
545
340
109
45
2017
168
7507
12842
21937
14789
12699
11672
11987
3773
1366
1359
358
2018
612
3280
11106
11214
8439
7825
4415
3727
2806
1917
753
358
44
2019
250
2348
13339
36004
17676
18353
5965
2564
2435
3795
811
152
178
2020
578
3172
7752
24370
28045
13275
6659
1957
1426
632
564
496
121
99
2021
188
1387
5703
3943
5112
9754
5664
1531
390
158
461
63
81
2022
248
2158
4509
6635
3749
4353
3739
1224
460
135
547
141
2023
598
4007
5637
6453
1663
1401
695
394
146
49
2024
302
2095
7450
6976
2118
683
2025
57
668
1643
1980
532
565
125
64
Tabell 1 Antall ved alder (mengdeindeks).
År
a5
a6
a7
a8
a9
a10
a11
a12
a13
a14
a15
a16
a17
a18
a19
2010
1.90
2.46
3.47
5.13
6.26
7.83
9.59
10.77
18.31
20.84
17.23
2011
1.66
2.28
2.89
4.52
6.82
8.82
9.55
9.08
13.38
10.70
2012
3.07
2.47
2.93
3.89
5.37
8.79
11.53
12.28
15.04
5.41
12.98
2013
2.49
3.05
3.52
4.46
5.54
7.56
10.26
10.23
11.49
16.61
16.80
11.95
2014
1.90
2.52
3.80
4.04
5.06
5.96
7.36
9.01
12.20
16.95
8.85
2015
2.16
2.62
3.42
3.95
5.21
6.53
8.32
9.95
12.45
14.21
13.90
11.69
2016
2.53
2.31
3.72
5.05
6.79
8.03
8.93
9.02
12.12
18.46
20.78
13.08
12.90
15.25
2017
2.01
2.52
2.94
4.91
5.75
7.16
8.18
9.10
10.49
11.59
10.92
13.25
2018
3.25
2.77
3.41
4.53
6.51
7.94
9.65
12.05
12.04
12.85
11.05
18.80
13.86
2019
2.12
3.02
3.76
4.81
6.07
7.44
8.71
11.06
13.86
13.40
11.17
16.10
16.69
2020
2.75
2.79
3.64
4.69
6.06
7.78
8.70
10.86
12.93
13.95
15.04
14.79
18.51
18.26
2021
2.28
2.73
3.77
4.41
6.49
7.37
8.57
10.13
11.82
12.00
8.56
19.20
16.75
2022
2.98
3.07
3.56
5.01
7.14
8.37
9.34
9.38
9.44
12.28
11.42
11.85
2023
3.03
4.15
4.85
5.99
7.19
9.36
11.88
15.31
17.22
19.07
2024
2.38
3.95
4.37
5.76
7.16
9.72
2025
2.43
3.46
4.58
5.87
7.02
8.41
9.44
8.17
Tabell 2 Vekt ved alder (kg).
Type: Skrei
Gytehistorie
Totalt
Umoden
Førstegangsgyter
Flergangsgyter
Antall
Biomasse
Antall
Biomasse
Antall
Biomasse
Antall
Biomasse
Alder
4
6.3
4
6.3
5
6
75
190.5
57
139.6
132
330.1
7
12
37.5
582
2076.6
86
236.5
680
2350.6
8
1121
5182.0
522
2352.2
1643
7534.2
9
941
5224.4
1039
6391.3
1980
11615.6
10
236
1614.4
297
2125.5
532
3739.9
11
85
659.9
480
4094.1
565
4754.1
12
125
1181.9
125
1181.9
13
64
523.2
64
523.2
Sum
91
234.3
3022
14896.9
2614
16904.6
5727
32035.9
Tabell 3 Antalls- og biomasseindekser fordelt på alder og gytehistorie. Umoden fisk benyttes ikke i tidsserien av indekser for moden fisk.
Type: Skrei
Gytehistorie
Totalt
Umoden
Førstegangsgyter
Flergangsgyter
Lengde
Vekt
Lengde
Vekt
Lengde
Vekt
Lengde
Vekt
Alder
59.0
1.440
59.0
1.440
5
6
67.4
2.555
61.9
2.432
65.0
2.502
7
69.0
3.040
74.3
3.568
68.7
2.748
73.5
3.455
8
80.9
4.622
79.7
4.504
80.6
4.585
9
86.4
5.555
87.3
6.150
86.8
5.867
10
91.2
6.845
91.3
7.167
91.2
7.024
11
95.6
7.759
97.5
8.523
97.2
8.408
12
99.8
9.442
99.8
9.442
13
97.7
8.171
97.7
8.171
Gj. snitt
67.2
2.567
82.2
4.929
88.3
6.468
84.8
5.594
Tabell 4 Gjennomsnittlig lengde og vekt fordelt på alder og gytehistorie.