Rapportserie:
Rapport fra havforskningen 2025-65ISSN: 1893-4536Publisert: 28.10.2025Prosjektnr: 15954Oppdragsgiver(e): Fiskeri og Havbruksnærings forskningsfond (FHF)Referanse: 901831 Godkjent av:
Forskningsdirektør(er):
Geir Lasse Taranger
Infection trials with Spironucleus salmonicida in lumpfish and Atlantic salmon
The report concerns infection trials with Spironucleus salmonicida in lumpfish and Atlantic salmon in spring 2024. The background for the work is the severe outbreak of spironucleosis in northern Norway in 2022–23. The outbreak led to high mortality, major economic losses and reduced fish welfare. An unexpected finding from the outbreak was the detection of systemic infection with S. salmonicida in lumpfish cohabiting with S. salmonicida infected salmon. This indicated that the parasite has a broader host range than previously assumed, and that transmission may occur between species that normally do not have close contact in nature.
The main aim of the infection trials was to test whether S. salmonicida can infect lumpfish through waterborne transmission (bath challenge) or by cohabitation with infected salmon (cohabitant challenge), and to investigate the consequences of such infection. Two separate infection trials were carried out.
Trial 1 included both bath challenge and cohabitation. The results showed very low transmission success. Only a few individuals tested positive, and there were no clinical signs or mortality during the eight weeks of the experiment. The lumpfish showed growth and apparently good health. There were indications that infected salmon can transmit infection to lumpfish when kept in the same tank, but the results were weak.
Trial 2 used a modified infection model, in which both the washing procedure, the infection dose and the salinity in the challenge tank were adjusted. Here, high transmission efficiency was achieved. Three weeks after infection the majority of lumpfish (83%) and salmon (78%) were infected with S. salmonicida. The parasite load in blood and intestine of infected lumpfish was high. Lumpfish proved to be highly susceptible to S. salmonicida. Shortly after the planned mid-sampling point, three weeks post-infection, acute mortality occurred among the lumpfish. Mortality in lumpfish started around day 23 post-infection and increased rapidly, and the experiment was therefore terminated four weeks after infection.
The results indicate that lumpfish may be as susceptible to S. salmonicida as salmon. The experiment also suggest that the consequences of infection are more severe and develop more rapidly in lumpfish than in salmon. There were few clinical signs, but varying degrees of exophthalmia were common, and some diseased fish had pale gills. Examination of moribund and dead fish showed pale hearts, watery blood, and necrotic foci in the liver. Large numbers of flagellates were observed in blood, peritoneal fluid, and extensively in stomach and intestine. Dead fish had feed in the gastrointestinal tract, supporting the impression that the fish were in good condition until sudden death. For salmon, there were no clinical signs or mortality during the experimental period, even though they had bloodstream infection. The course in lumpfish resembled acute spironucleosis in salmon, and as in salmon, a brain infection may be suspected as the cause since flagellates were observed in the brains of lumpfish in this trial.
Conclusions
Lumpfish are susceptible to S. salmonicida free in water.
Infected salmon can transmit S. salmonicida to lumpfish.
Lumpfish develop severe disease and may die as a consequence of infection.
Lumpfish should not be used at sites with S. salmonicida infections.
Sammendrag
Rapporten omhandlar smitteforsøk med Spironucleus salmonicida på rognkjeks og laks våren 2024. Bakgrunnen for arbeidet er det alvorlege utbrotet av spironukleose i Nord-Noreg i 2022-23. Utbrotet førte til høg dødelegheit, store økonomiske tap og redusert fiskevelferd. Eit uventa funn frå utbrotet var påvising av systemisk infeksjon med S. salmonicida hos rognkjeks som gjekk saman med S. salmonicida infisert laks. Dette indikerte at parasitten har breiare vertsspekter enn tidlegare antatt, samt at det kan forekomme smitte mellom artar som normalt ikkje har tett kontakt i naturen.
Hovudmålet med smitteforsøka var å teste om S. salmonicida kan smitte rognkjeks gjennom vatn (badsmitte) eller ved kohabitasjon med infisert laks (kohabitantsmitte), samt undersøke kva følgjer ein slik infeksjonen vil ha. To separate smitteforsøk vart gjennomført.
Forsøk 1 inkluderte både badsmitte og kohabitasjon. Resultata viste svært låg smittesuksess. Berre eit fåtal individ vart påvist positive, og det var ingen sjukdomsteikn eller dødelegheit i løpet av dei åtte vekene forsøket varte. Rognkjeksa hadde vekst og tilsynelatande god helse. Det var indikasjonar på at infisert laks kan overføre smitte til rognkjeks når desse er haldt i same kar, men resultata var svake.
Forsøk 2 nytta ein justert smittemodell, der både vaskeprosedyre, smittedose og salinitet i smittekaret vart endra. Her vart det oppnådd høg smitteeffektivitet. Tre veker etter smitte var størsteparten av rognkjeks (83 %) og laks (78 %) infisert med S. salmonicida. Mengda parasitt i blod og tarm på smitta rognkjeks var høg. Rognkjeks viste seg svært mottakeleg for S. salmonicida. Kort tid etter det planlagde midtuttaket, 3 veker etter smitte, oppstod det akutt dødelegheit mellom rognkjeksene. Dødelegheita i rognkjeks starta rundt dag 23 etter smitte og auka raskt, og forsøket vart derfor avslutta 4 veker etter smitte.
Resultata indikerar at rognkjeks kan vera like mottakeleg for S. salmonicida som laks. Forsøket tyder og på at konsekvensane av infeksjon er alvorlegare og oppstår raskare hos rognkjeks enn laks. Det var lite klinikk, men varierande grad av eksoftalmi var vanleg og nokre sjuke fisk hadde bleike gjeller. Undersøking av døyande og død fisk viste bleike hjarte, vassent blod, innslag av nekroser (flekkar) på lever. Det var store mengder flagellatar i blod, bokholevæske, og mykje i mage og tarm. Dødfisk hadde för i mage-tarm, og dette underbygger inntrykket av at fisken var i godt hald fram til dei brått døydde. For laks var det ikkje sjukdomsteikn eller dødelegheit i forsøksperioden, sjølv om dei hadde blodinfeksjon. Forløpet i rognkjeks liknar akutt spironucleose hjå laks, og som for laks kan ein mistenke hjerneinfeksjon som årsak sidan det vart sett flagellater i hjerna hos rognkjeksa i dette forsøket.
Konklusjonar
Rognkjeks er mottakeleg for S. salmonicida fritt i vatn.
Infisert laks kan smitte rognkjeks.
Rognkjeks blir alvorleg sjuk og kan døy som følgje av infeksjon.
Rognkjeks bør ikkje nyttast på lokalitetar med S. salmonicida infeksjonar
1 - Introduksjon
Fiskeri og Havbruksnæringens forskingsfinansiering har med bakgrunn i utbrota av spironukleose i Nord-Noreg bevilga midler til gjennomføring av prosjektet «Spironucleus salmonicida infeksjoner og spironucleose i norsk akvakultur: smitteveier, vertsspekter og førebygging (SpiroFri)» (FHF prosjektnummer 901831). SpiroFri-prosjektet er leia av Havforskingsinstituttet, og gjennomførast i samarbeid med Nofima Tromsø, Veterinærinstituttet, Pure salmon Kaldnes v/ Erik Sterud, og Grieg Seafood ASA. Formålet med prosjektet er å få frem kunnskap for førebygging og bekjemping av S. salmonicida, både med tanke på laksefisk, men òg rensefisk og villfisk.
S. salmonicida er ein diplomonad organisme. Diplomonadar er små anaerobe (mikroaerofile) flagellatar (Adam, 2017) som er frittlevande eller symbiontar i fordøyingssystemet til dyr. Nokre av desse flagellatane kan vera skadelege parasittar og forårsake sjukdom. Mest kjend er nok Giardia intestinalis, som kan gje plagsom magesjuke hos menneske. Spironucleus artane lever særleg i tarmen hjå kaldblodige dyr som fisk, amfibiar og skilpadder, men der er og nokre artar i tarmen på fugl og pattedyr som er klassifisert i denne slekta. Fleire av artane forårsakar sjukdom. I fisk inkluderar dette S. vortens, S. salmonis, og S. salmonicida (Fard et al., 2007; Jorgensen & Sterud, 2006; Paull & Matthews, 2001; Poynton et al., 2004; Uldal & Buchmann, 1996).
I Nord-Noreg har S. salmonicida gjentatte gongar forårsaka sjukdomsutbrot i oppdrett av laks. Arten var først skildra og navngjeven i 2006, og flagellaten vart tidlegare identifisert som Hexamita sp., Hexamita salmonis og som Spironucleus barkhanus. S. salmonicida (då som Hexamita sp.) vart først påvist i 1989, i samband med eit utbrot av spironukleose i oppdrettslaks i Nord-Noreg (Mo et al., 1990; Poppe et al., 1992). Etter 1989, der 6 lakselokalitetar vart affisert, er det påvist utbrot på ein lokalitet med oppdrettslaks i 2001 (Sterud et al., 2002), nokre lakselokalitetar i 2007-08 (Johansen et al., 2009; Skjelstad et al., 2008), samt ved ein oppdrettslokalitet for røyr i 2002 (Sterud et al., 2003). Det hittil siste, og største, utbrotet av spironukleose i norsk oppdrett var i 2022-2023 der 10-12 matfisklokalitetar vart råka (Lie et al., 2022; Sommerset et al., 2024; Sommerset et al., 2023). Utbrota av spironukleose har vore alvorlege med store konsekvensar for affiserte anlegg, med tilfelle av destruksjon, høg dødelegheit, redusert produksjon, samt nedklassifisering av fisken (Grieg Seafood ASA, 2023; Mo et al., 1990; Sterud et al., 2002). 2022-utbrotet er det hittil største, med estimerte tap for oppdrettar opp mot 900 millionar norske kroner (Grieg Seafood ASA, 2023).
Kunnskapsstatus for S. salmonicida er oppsummert i Karlsbakk mfl. 2025 (Karlsbakk, Hansen, et al., 2025). Det er framleis lite kunnskap om livssyklus, smittevegar og infeksjons- og sjukdomsdynamikk for S. salmonicida. Det er kjent fire ulike studier med smitteforsøk med S. salmonicida i laks i ferskvatn. Etter eit utbrot av spironukleose på oppdretta kongelaks (chinook salmon, Oncorhynchus tshawytscha) i sjøvatn i 1991 gjennomførte Kent mfl. (1992) smitteforsøk med kongelaks og atlantisk laks (Kent et al., 1992). Laks vart intubert med infektivt materiale, men i løpet av dei ca fem vekene intubasjonssmitteforsøket pågjekk, vart atlantisk laks ikkje påviseleg smitta. Kongelaks vart derimot smitta med fleire ulike smittemodellar i dette forsøket, og det var høg dødelegheit over kort tid (1.5-4.5 veker). Parasitten på kongelaks i British Columbia er S. salmonicida, men er nok ein annan variant enn me har i Noreg. Som oppfølging av spironukleoseutbrotet i Noreg i 1989-1991 gjorde Mo mfl. (1993) kohabitasjonssmitteforsøk med laks, aure, røyr og regnbogeaure i brakkvatn (25 ‰) og ferskvatn (Mo et al., 1993). Smittekjelda i forsøket var stor laks med spironukleose. Etter 26 dagar var kun ein laks smitta i brakkvatn. For å undersøke smittevegar og infeksjonsdynamikk gjennomførte Guo og Woo (2004) smitteforsøk med S. salmonicida på laks i ferskvatn (Guo & Woo, 2004). Laks nytta som smittespreiarar («sheddarar») vart smitta med injeksjon i bukhola (intraperitonal injeksjon; i.p.), og kohabitantar tilsett. All injisert fisk vart infisert, og 50 % av kohabitasjonsfisken var smitta etter 4-6 veker. I løpet av forsøksperioden på inntil 21 veker etter smitte utvikla fisken byllar og sår, og det var høg dødelegheit. I 2018 gjorde Alfjorden smitteforsøk med laks i ferskvatn med to ulike dosar parasitt pipettert direkte ned i magen (oral intubasjon) eller injisert intraperitonalt i fisken (Alfjorden, 2018; Alfjorden et al., 2019). Vekentlege blodprøvar viste aukande prevalens til 100 % i dei 10 vekene forsøket gjekk, men det var lite lesjonar og ingen fisk døydde.
I 2022-23 utbrotet i Nord-Noreg vart det overraskande funne systemisk infeksjon med S. salmonicida i rognkjeks frå lokalitet med spironukleoseutbrot (Sommerset et al., 2024). Før denne påvisinga var det ikkje kjent at S. salmonicida kunne smitte andre artar enn laksefisk. Funnet indikerte at rognkjeks som går saman med infisert laks kan bli smitta av S. salmonicida. Men verken smittevegen frå laks til rognkjeks eller konsekvensane av smitten var kjent.
Funnet av systemisk S. salmonicida-infeksjon i rognkjeks er elles ei påminning om at smitte til nye artar kan skje når fleire artar vert helde tett saman, spesielt når økologien til artane tilseier at dei i naturen ikkje har mykje kontakt. Over tid kan dette få utilsikta konsekvensar knytt til smitte mellom artar og etablering av nye agens i oppdrett av fisk eller i ville populasjonar.
Som følgje av manglande kunnskap om interaksjonen mellom laks, rognkjeks og S. salmonicida, og konsekvensane av denne, vart ei av målsetjingane i SpiroFri-prosjektet å undersøke mottakelegheit, smittevegar og konsekvensar av infeksjon med S. salmonicida hos rognkjeks. Det vart gjennomført eit hovudforsøk med badsmitte og kohabitasjonssmitte (forsøk 1), og eit mindre forsøk med badsmitte (forsøk 2). I begge forsøka vart det teke blod- og tarmprøvar til påvising og kvantitering av S. salmonicida, samt histologiske prøvar. Ved sluttuttaka vart fisken òg undersøkt for indre og ytre teikn til spironukleose.
2 - Metodar
2.1 - Dyrking av S. salmonicida
Isolat SS1-1 (12/12/2023) isolert frå halebyll av laks i smitteforsøket med laks og aure vart nytta som smittemateriale (Karlsbakk, Nordbø, Austgulen, et al., 2025).
S. salmonicida vart dyrka i henhald til standard dyrkingsbetingar. Modifisert Keister’s medie (ATCC 2695 Medium; Keisters modifiserte Giardia TYI-S-33) utan glukose, og med MEM vitaminløysing (Sigma Aldrich, M6895) i staden for Diamonds vitamintilsetjing (Merck, 58980C) tilsatt, vart nytta som dyrkingsmedie (Keister, 1983). Keisters basemedie bestående av trypton, gjærekstrakt, NaCl, K2HPO4, KH2PO4 og jernammonium citrat vart laga, justert til pH 7.4 og autoklavert. Før bruk vart basemediet tilsatt oksegalle, cystein, vitaminer og antibiotika (antimycotic). Parasitten vart dyrka aseptisk i celledyrkingsflasker med 25, 75 eller 175 cm2 overflate (Nunc EasyFlask Cell Culture Flasks; Thermo Scientific, henholdsvis art. nr 156340, 156472, 159920) ved 6-10 °C utan risting. Kulturane vart splitta med 3-6 dagars mellomrom avhengig av vekst. Ved dyrking i 75 cm2 celledyrkingsflasker vart 10 ml kultur overført til 30 ml ferskt medium ved splitting. For å halde kulturane oksygenfrie vart mediet «lufta» med nitrogengass kvar gong flasker eller røyr vart opna og utsett for oksygen (luft). Rein nitrogengass vart bobla ned i mediet i ca 10 sekund og deretter vart flasker/røyr raskt lukka. Kulturar vart overvaka med omvendt mikroskop ved 10-20x, for vekst og vitalitet.
2.1.1 - Smittemateriale - Forsøk 1
Før smitte vart S. salmonicida kultur vaska i PBS for å fjerne vekstmedium og avfallstoff frå flagellatane. Totalt 2240 mL S. salmonicida-kultur i 40 mL alikvotar vart sentrifugert i 50 mL røyr ved 2000 x G i 10 min ved 4 °C. Pellet vart vaska ved å fjerne 35 mL supernatant, tilsette ca 5 mL PBS og løyse opp pellet. Deretter vart to og to røyr slått saman, volumet justert til 40 mL med PBS og deretter sentrifugert på nytt ved 2000 x G, 10 min, 4 °C. Dette vart gjort totalt to gongar. Etter siste vask vart supernatanten fjerna og pellet løyst opp i 25 mL PBS før alle røyr vart slått saman og antall S. salmonicida i smitteløysinga talt. For kvantifisering vart flagellatar immobilisert med glyserol i PBS (Glyserol:PBS 1:3) og talt med hemocytometer (Kova Glasstic, 87144E) i mikroskop i henhold til produsentens skildring. Kvar prøve vart talt tre gongar. Smitteløysinga stod på is under arbeidet. Det vart estimert totalt ca 560 Mill S. salmonicida i smitteløysinga.
2.1.2 - Smittemateriale - Forsøk 2
Etter resultata i forsøk 1, samt frå overlevelsesforsøk (Helsø, 2024), var vaskeprosedyra for preparering av parasitten til smitte ei mistenkt årsak til låg smittesuksess i forsøk 1. I forsøk 2 vart derfor vaskeprosedyra justert, blant anna basert på overlevelsesdata frå Helsø (2024). S. salmonicida kultur (40 mL) vart fordelt i 50 mL sentrifugerøyr, og sentrifugert ved 1000 x G i 10 min ved 4 °C. Supernatant, 35 mL, vart fjerna og pellet løyst i ca 45 mL 10 ‰ sjøvatn før alle røyr vart slått saman. Antal parasittar vart funnet som skildra for forsøk 1. Det vart estimert totalt ca 1 400 Mill S. salmonicida i smitteløysinga.
2.2 - Fisk
Laks post-smolt (AquaGen; 232.9 g) (Tabell 1) til forsøket vart produsert og PIT-merka ved Havforskningsinstituttet sin stasjon i Matre. Fisken vart transportert frå Matre med bil til Havforskingsinstituttets smittelaboratorium (Nordnesgaten 50, Bergen) 26. januar 2024.
Kommersielt produsert vaksinert og uvaksinert rognkjeks (hhv 29,2 ±6,1 g og 22,5 ±5,1 g) (Tabell 1) vart kjøpt frå Vest Aqua Base. Rognkjeksa vart transportert frå produksjonsanlegget (Leirpollvegen 56, 5419 Fitjar) med bil til Havforskingsinstituttets smittelaboratorie 09. januar 2024. Rognkjeksa vart PIT-merka ved Havforskingsinstituttet.
Etter mottak vart laks og rognkjeks holdt i individuelle 600 L kar med 350 L 24 ±2.3 ‰ saltvatn time-1, ca 9 °C og 12:12 L:D lysregime fram til smittedagen. Etter smitte vart fisk fordelt i kar à 250 L (Tabell 2), med same vassparametre som før smitten. Gjennom heile forsøksperioden vart fisken forá med Nutra RC3, 3 mm fôr (Skretting AS). Fisken vart røkta minimum to gongar/dag, og temperatur, salthald og oksygen vart målt dagleg.
Fisk i forsøk 2 var overskotsfisk og kontrollfisk frå forsøk 1. Forsøket vart gjennomført i 250 L kara nytta i forsøk 1 med laks og rognkjeks i to individuelle kar (Tabell 3). Fisken vart elles behandla som skildra for forsøk 1.
Tabell 1. Vekt av fisk før start av forsøk 1
Forsøksgruppe
N
Kar
Snittvekt (g)
SD (g)
Laks Kontroll
50
VK1-2
232,91
na
Laks Kohabitant
32
VK6-9
Laks Badsmitta
40
VK6-9
Rognkjeks Uvaksinert Kontroll
55
VK3
22,52
5.1
Rognkjeks Uvaksinert Kohabitant
160
VK6-9
Rognkjeks Uvaksinert Badsmitta
100
VK4-5
Rognkjeks Vaksinert Kontroll
55
VK3
29,23
6.1
Rognkjeks Vaksinert Badsmitta
100
VK4-5
1 Snitt av bulk vekter (2) med henholdsvis 10 og 11 laks vegd 25. januar
2 Snitt av 321 uvaksinerte rognkjeks individuelt vegd 29. januar
3 Snitt av 198 vaksinerte rognkjeks individuelt vegd 26. januar
Tabell 2. Oversikt over fordeling av fisk i forsøk 1.
Fiskegrupper/Kar
Kar
Laks
(bad)
Laks
(naiv)
Rognkjeks
(bad)
Rognkjeks
(naiv)
Antall
Kontroll Laks
VK1
25
25
Kontroll Laks
VK2
25
25
Kontroll Kjeks (uvaks+vaks)
VK3
110
110
Badsmitta Kjeks (uvaks+vaks)
VK4
100
100
VK5
101
101
Smitta Laks
Naiv Laks
Kjeks uvaks
VK6
10
8
40
58
VK7
10
8
40
58
VK8
10
8
40
58
VK9
10
8
40
58
Antall
40
82
201
270
593
Tabell 3. Vekt av fisk før start av forsøk 2
Forsøksgruppe
N
Snittvekt (g)
SD (g)
Laks Badsmitta
10
225,6 1
27,6
Rognkjeks Badsmitta
30
71,9 2
11,6
1 Snittvekt Kontroll Laks (n=38)frå forsøk 1 vegd 5. april
2 Snittvekt Kontroll Rognkjeks (n=80) frå forsøk 1 vegd 5. april
2.3 - Smitteforsøk
2.3.1 - Forsøk 1
Før smitte 7. februar var all fisk som skulle badsmittast (40 laks og 201 rognkjeks, Tabell 2) håva inn i smittekaret (1 m2, vassvolum 560 L). Ved start av badsmitte vart vatnet til karet stengt av, og vassvolumet redusert til ca 100 L. Gjennom heile perioden utan vasstilførsel vart det tilsett luft med to luftesteinar. Vaska S. salmonicida (ca 560 Mill) vart miksa inn i vatnet. Smittedosen tilsvarte ca 5-6 Mill S. salmonicida L-1. Fisk og oksygennivå vart observert kontinuerleg gjennom smitteperioden. Etter 60 minuttar vart vasstilførselen (700 L time-1) satt i gang. Temperatur og salthald i smitteperioden var henholdsvis ca 7,8 °C og ca 22,9 ‰. Fisken vart halden i smittekaret over natt før den vart fordelt til forsøkskar (vassvolum ca 135 L) (Tabell 2). I forsøksperioden fekk kvart forsøkskar ca 350 L time-1 vatn, og temperatur og salinitet var henholdsvis 8,0 ± 0,2 °C og ca 25 ‰, og med 12:12 lysregime.
2.3.2 - Forsøk 2
Før smitte 18. april vart fisken som skulle badsmittast (10 laks og 30 rognkjeks) håva inn i smittekaret (vassvolum ca 135 L). Ved start av badsmitte vart vatnet til karet stengt av, og vassvolumet redusert til ca 50 L. Gjennom heile perioden utan vasstilførsel vart det tilsett luft til karet. Vaska S. salmonicida (ca 1 400 Mill) vart miksa inn i vatnet. Smittedosen tilsvarte ca 28 Mill S. salmonicida L-1. Fisk og oksygennivå vart observert kontinuerleg gjennom smitteperioden. Etter 30 minuttar vart vasstilførselen (350 L time-1) satt i gang. Temperatur og salthald i smitteperioden var henholdsvis ca 8,0 °C og ca 10,5 ‰. All fisken vart holdt i smittekaret over natt før laks og rognkjeks vart fordelt på to like forsøkskar (vassvolum ca 135 L). I forsøksperioden var vassflow i kvart kar ca 350 L time-1, temperatur og salinitet var henholdsvis ca 8,8 ± 0,4 °C og ca 25 ‰, med 12:12 lysregime.
Forsøket var godkjent gjennom Forsøksdyrforvaltningens tilsyns- og søknadssystem (FOTS-ID: 30519) før oppstart.
2.4 - Prøvetaking
I forsøk 1 vart det teke blodprøve og tarmprøve av alle fisk 4 (midtuttak; 4 veker etter smitte) og 8 (sluttuttak; 8 veker etter smitte) veker etter smitte. Ved sluttuttaket vart fisk i tillegg undersøkt for ytre og indre sjukdomsteikn som tidlegare skildra (Kvamme et al., 2025). Ved moglege lesjonar vart det mikroskopert på nativpreparat for påvising av S. salmonicida, og tatt prøvar for histologi. Det vart tatt prøvar til qPCR analyse og histologi frå kvar forsøksgruppe. Etter prøveuttak og undersøking vart all fisk frosne enkeltvis i posar (-20 °C) for eventuell seinare prøvetaking.
I forsøk 2 vart det teke blodprøve og tarmprøve av all fisk 3 veker etter smitte («midtuttak»), og frå gjennværande fisk, 10 laks og 19 rognkjeks, 4 veker etter smitte (sluttuttaket). Det vart då tatt prøvar til histologi og qPCR (Tabell 4) frå alle fiskane.
For både forsøk 1 og 2 vart blodprøvar, tarmsvaber, og prøvar til qPCR og histologi tekne på same måte.
Blodprøvar vart tekne med 1 mL sprøyter og 0.6G kanylespissar, frå dei caudale blodkara. Volumet var 100 µL, tatt forsiktig for å ikkje gje interne skader på fisken, og deretter overført til prøverøyr tilsett antikoagulant (10 µl 20 mM EDTA). Prøvane vart haldne på is inntil dei vart frosne på -20 °C.
Tarmsvaber vart tekne med med mjuk svaber (PurFlock Ultra, MWE Ltd, #MW814). Svaber vart ført forsiktig inn gjennom endetarmsåpningen og bevega forsiktig fram og attende nokre gongar i bakre del av tarm. Svabertupp vart deretter klipt av og ned i individuelle eppendorfrøyr, og halden på is inntil den vart frose ved -20 °C.
Prøvar til histologi varte tekne så raskt som mogleg etter avliving og fysisk undersøkjing. Det vart tatt prøvar av ni vev (Tabell 4), samt venstre auge, frå kvar prøvetatt fisk. Alle vev frå kvar fisk vart fiksert i 10 % forsfatbufra formalin (ca. 4 % formaldehydløysing) (Cellstor Pot 60 (10 % NBF), Histolab #02165) ved romtemperatur. Etter 24 timar vart formalin helt av, prøvane vaska i 70 % etanol, og deretter lagra ved romtemperatur i ca 50 mL 70 % etanol inntil opparbeiding.
Prøvar til qPCR (molekylærbiologiske analyser) vart tekne samstundes med histologiprøvane. Prøvar av veva, utanom auge, (Tabell 4) for kvar fisk vart lagt på individuelle eppendorfrøyr som stod i tørris for frysing. Prøvane vart lagra ved -20 °C inntil opparbeiding.
Tabell 4. Oversikt over vev prøvetatt til histologi og qPCR
Vev
Merknad
Gjelle
Midt på første venstre gjellebue
Kaudalmuskel
Muskel ved venstre halefinnerot
Hjerte
Ventrikkel
Lever
Fremre del, venstre side
Milt
Tilfeldig bit
Nyre
Framre, bakre
Pylurusblindsekkar
Tarm
Bak siste caeca
CNS
Hovedsakelig metencephalon.
Venstre auge
Kun histologi
2.5 - Rensing Genomisk DNA
Rensing av DNA frå blodprøvar vart gjort med Genefinder V3 kit (Beckman Coulter) på ein Biomek i5 Automated Workstation (Beckman Coulter) i 96-brønnars brett. Blodprøvane vart tint og deretter homogenisert ved hjelp av risting med stålkuler. Frå den homogeniserte prøven vart 5 µl overført til 195 µl PBS-buffer og DNA rensa i henhold til produsentens anbefalingar. DNA vart eluert i 50 µl elueringsbuffer (EBB-buffer) og lagra på -20 °C inntil analysert. Genefinder protokollen vart i all hovudsak fulgt, med nokre tilpasningar av det automatiske programmet for arbeidsstasjonen.
Rensing av DNA frå tarmsvaber vart gjort med DNAdvance kit (Beckman Coulter) på ein Biomek i5 Automated Workstation (Beckman Coulter) i 96-brønnars brett. I kortheit vart svaber overført frå eppendorfrøyr til 96-brønners deepwell-plate (DW-plate), tilsett 200 µl lysis buffer med Proteinase K (ca 1,4 mg/mL) og blanda godt. Deretter vart DW-platene sentrifugert og inkubert på ein Corning benchtop shaking incubator over natt med 130 rpm ved 55 °C. Neste dag vart platene vorteksa og sentrifugert ved 1000 x G i 1 min. Deretter vart 200 µl lysat overført til ny isoleringsplate og plassert i arbeidsstasjonen og reinsa. DNA vart eluert i 50µl elueringsbuffer (EBA-buffer) og lagra på -20 °C inntil analysert.
2.6 - qPCR-analyser
Kvantifisering av S. salmonicida DNA i blod og tarm (svaberprøvar) vart gjort med kvantitativ real-time PCR (qPCR) med EagleTag 2x Universal Master Mix (ROX) (Merck, art.nr. 7260296190) etter produsentens anbefalingar. Rensa DNA frå tarmvæske- og blodprøve vart nytta som templat, enten ufortynna eller fortynna 1:10 med dH2O. Det vart nytta 384 brønnars brett og totalvolum 8 µl reaksjon-1; 4 µl 2X qPCR Master Mix, 0,4 µl av kvar primer (10 µM), 0,4 µL probe (4 µM), 0.8 µl vatn og 2 µl templat. Primerar og probar er gjevne i Tabell 5. Effektiviteten for S. salmonicida qPCR assayet (Tabell 5) vart berekna til mellom 0,87 (Blod) og 1,00 (PBS) (R2 hhv. 0.99 og 0.98).
All qPCR vart utført i Quantstudio TM 5 Real-Time PCR instrument (384-block) (Applied Biosystems, by Thermo Fisher Scientific) med følgande temperaturvilkår: 10 min ved 95 °C, 45 syklar: 15 sek ved 95 °C, 60 sek ved 60 °C, 1 sek ved 72 °C, deretter 30 sek ved 40 °C, og til sist ∞ ved 4 °C.
Ct-verdiar vart bestemt ved hjelp av QuantStudio TM Design & Analysis Software v1.4.2 (Applied Biosystems), med grenseverdi for berekning av Ct satt til 0,2.
Tabell 5. Primerar og probar nytta for qPCR-analysar. F=forward, R=reverse, P=probe.
Gen
Assay navn
qPCR assay (5’-3’)
Referanse
S. salmonicida 18S (AY677182)
Spiro
F: GCGAGATTTGTTGCATTTAAAAAG
R: CGAACTTTTTAACTGCAGCAACA
P: 6FAM-ACACGGAGAGTATTCT-MGB
(Miller et al., 2016)
S. salar GAPDH§
(XM_014152708.2)
GAPDH
F: GCCACCCAGAAGACTGTTGA
R: GCAGGGATGATGTTCTGGCT
P: 6FAM-GTGGAGGGATGGACGTGGCG-QSY
Skaftnesmo i (Kvamme et al., 2025)
§ Glyseraldehyd 3-fosfat dehydrogenase
3 - Resultat
3.1 - Fisk
Tabellane 6 og 7, og Figur 2, 3 og 4 oppsummerar vektdata for forsøk 1 og 2.
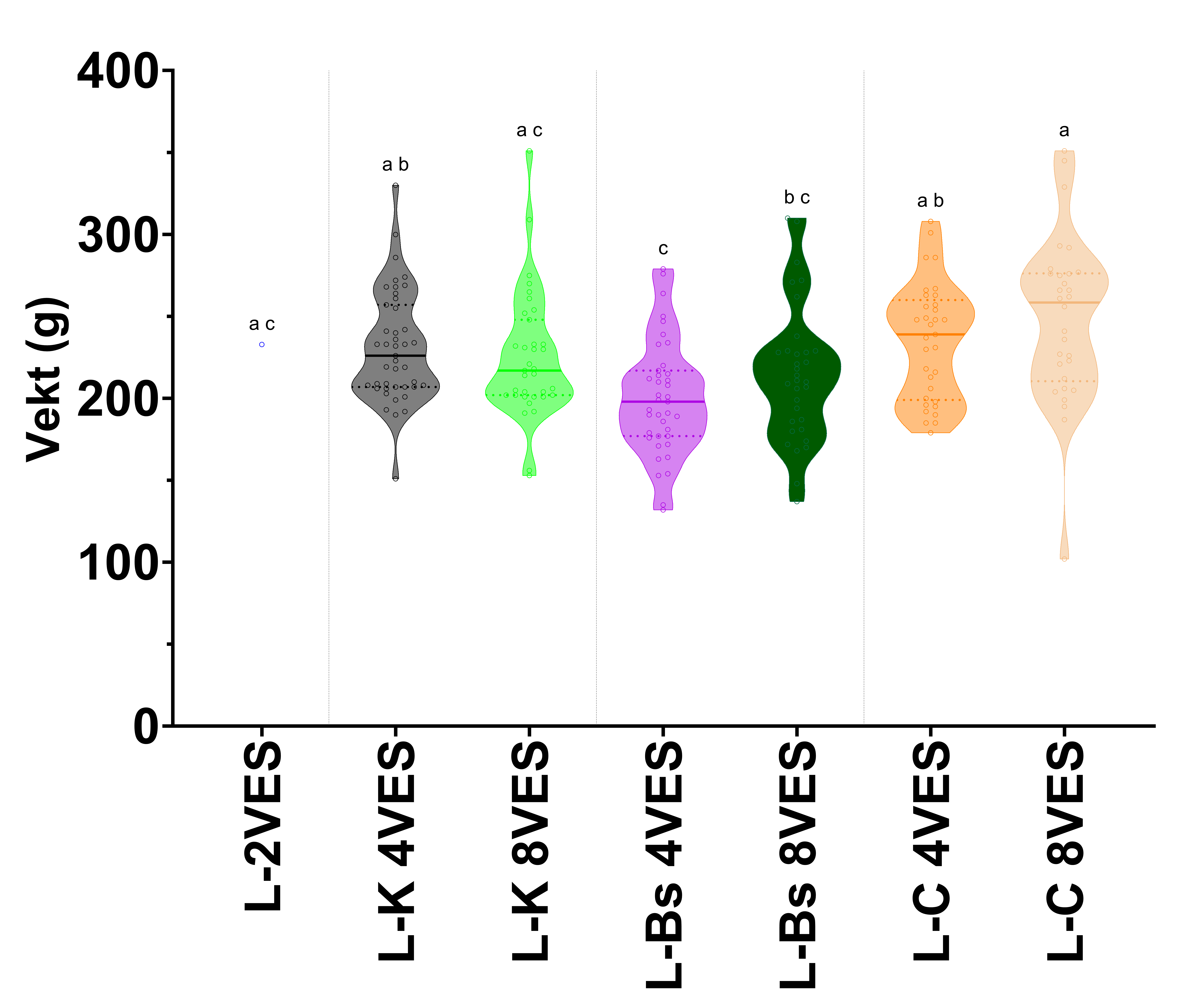
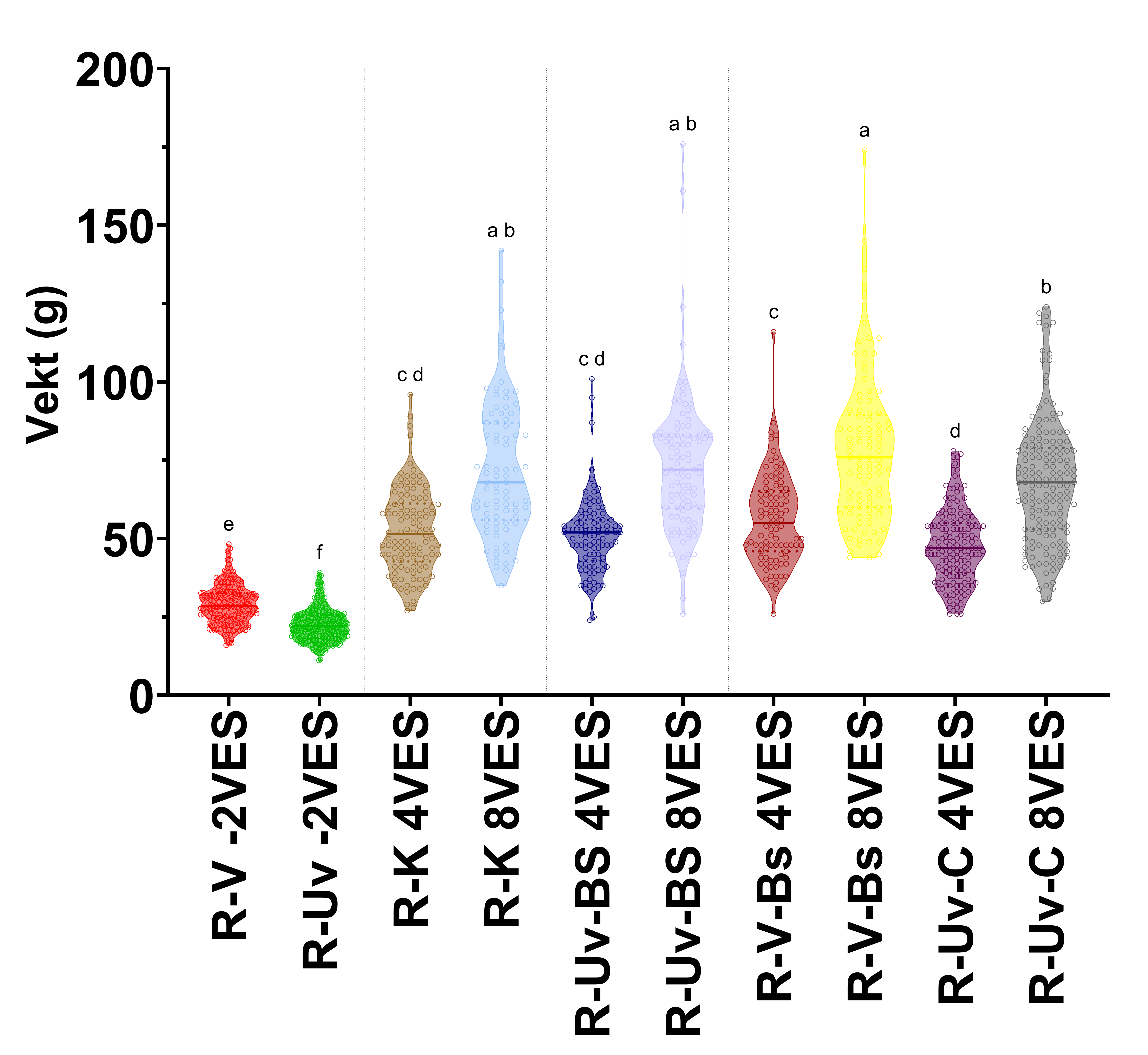
I forsøk 1 var det ingen vektauke for laks (Tabell 6 og Figur 2), inkludert i kontrollgruppa, for heile forsøksperioden. Rognkjeksa vaks derimot i alle grupper (Tabell 6 og Figur 3), med signifikant forskjell i vekt før forsøkstart til forsøksslutt for alle forsøksgrupper inkludert kontroll. Ved forsøksslutt, 8 veker etter smitte, var ingen av rognkjeksgruppene signifikant forskjellig frå kontrollgruppa. Kun uvaksinerte kohabiterande rognkjeks hadde lågare vekt 8 veker etter smitte enn vaksinert badsmitta rognkjeks (Figur 3).
Tabell 6. Vektdata for forsøk 1.
Gruppe
VES
DES
Snittvekt (g)
SD (g)
n
SGR (70d)
Laks Kontroll
-2
-14
232,9 1
na
2 (10+11)
-0,05
4
28
229,5
33,5
50
8
56
225,6
37,9
38
Laks Kohabitant
-2
-14
232,9 1
na
2 (10+11)
0,09
4
28
235,0
35,5
34
8
56
248,6
51,1
30
Laks Badsmitta
-2
-14
232,9 1
na
2 (10+11)
-0,11
2
14
196,6
34,78
40
4
28
200,1
35,1
40
8
56
215,1
41,7
40
Rognkjeks Uvaksinert Kontroll
-2
-14
22,5 2
5,1
321
1,83
4
28
57,1
12,7
54
8
56
80,8
20,7
36
Rognkjeks Uvaksinert Kohabitant
-2
-14
22,5 2
5,1
321
1,58
4
28
47,6
11,6
163
8
56
68,1
19,8
156
Rognkjeks Uvaksinert Badsmitta
-2
-14
22,5 2
5,1
321
1,68
4
28
51,1
12,2
99
8
56
73,1
22,1
98
Rognkjeks Vaksinert Kontroll
-2
-14
29,2 3
6,1
198
1,08
4
28
47,3
11,9
51
8
56
62,1
18,4
40
Rognkjeks Vaksinert Badsmitta
-2
-14
29,2 3
6.1
198
1,40
4
28
56,2
14,3
94
8
56
77,8
24,2
91
1 Snitt av to bulk vekter med henholdsvis 10 og 11 fisk vegd 25. januar
2 Snitt av 321 uvaksinerte Rognkjeks individuelt vegd 29. januar
3 Snitt av 198 vaksinerte Rognkjeks individuelt vegd 26. januar
Figur 2. Trunkert fiolinplot for vekt av laks i forsøk 1 før forsøksstart, og 4 og 8 veker etter badsmitte. Grupper er adskilt med vertikale liner. L-Laks, K-Kontroll, Bs-Badsmitte, C-Kohabitant, VES-veker etter smitte. Bokstavene over søylene indikerar statistiske grupper basert på Tukeys multiple samanlikningstest med kompakt bokstavvisning (p < 0.05). Grupper med ulike bokstaver er signifikant forskjellige.
Figur 3. Trunkert fiolinplot for vekt av rognkjeks i forsøk 1 før forsøksstart, og 4 og 8 veker etter badsmitte. Grupper er adskilt med vertikale liner. R-Rognkjeks, V-Vaksinert, Uv-Uvaksinert, K-Kontroll, Bs-Badsmitte, C-Kohabitant, VES-veker etter smitte. Bokstavene over søylene indikerar statistiske grupper basert på Tukeys multiple samanlikningstest med kompakt bokstavvisning (p ≤ 0,05). Grupper med ulike bokstaver er signifikant forskjellige.
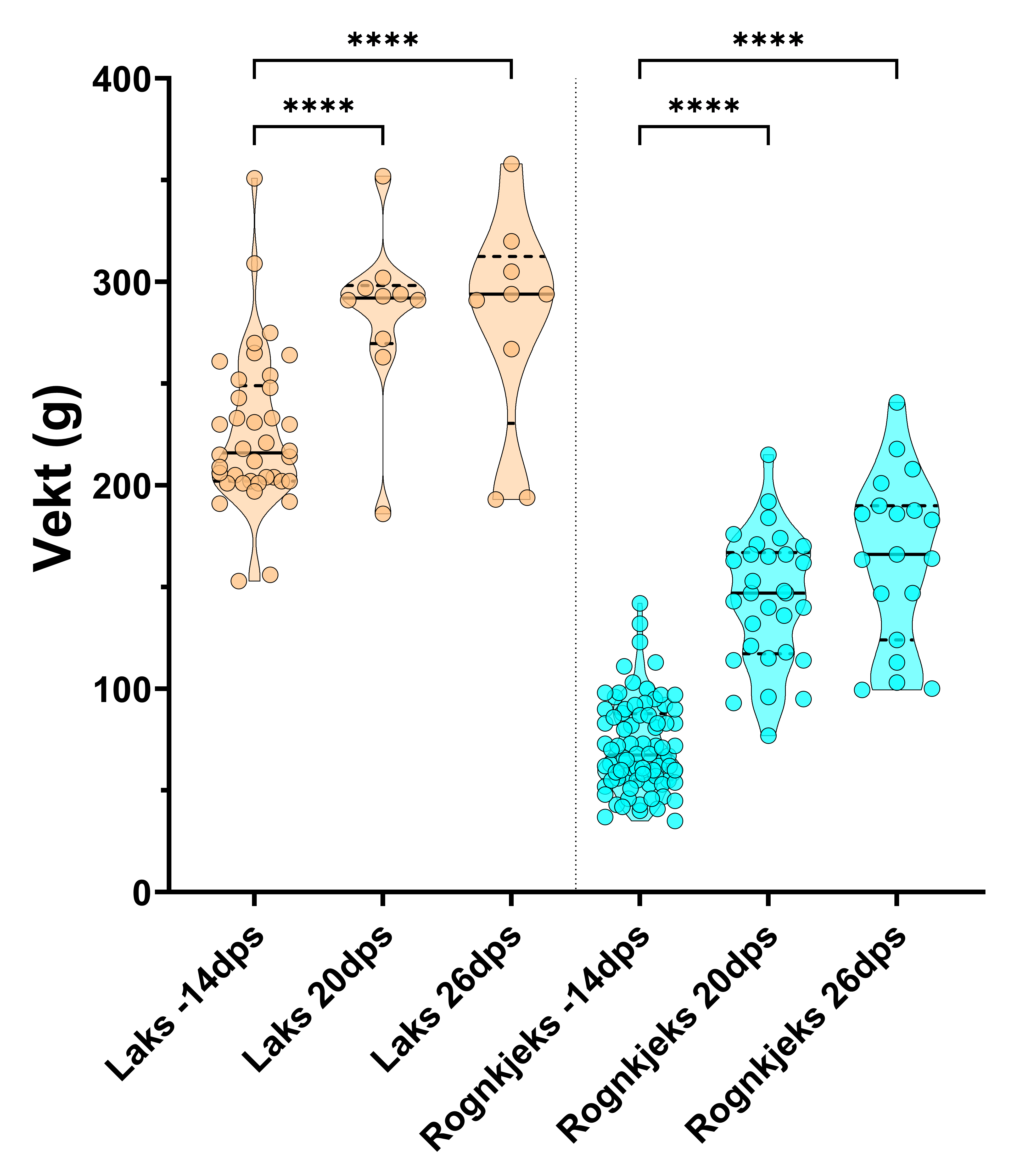
I forsøk 2 auka vekta (p<0,001, Anova med Tukey multiple test) til både laks og rognkjeks gjennom forsøksperioden (Tabell 7 og Figur 4). Det var i tillegg ein liten forskjell (p<0,5, einsidig t-test) vektauke for rognkjeks dei siste 6 dagane i forsøket (20 DES til 26 DES).
Tabell 7. Vektdata for forsøk 2
Gruppe
VES
DES
Snittvekt (g)
SD (g)
n
SGR (40d)
Laks Badsmitta
-2
-14
225,6 *
27,6
40
0,36
3
20
284,1
41,6
10
4
26
290,7
44,4
9
Rognkjeks Badsmitta
-2
-14
71,9 **
11,6
80
1,18
3
20
144,4
32,4
30
4
26
164,6
41,8
19
* Snittvekt Kontroll Laks (n=38) frå forsøk 1 vegd 5. april
** Snittvekt Kontroll Rognkjeks (n=80) forsøk 1 vegd 5. april
Figur 4. Trunkert fiolinplot for vekt av laks og rognkjeks i forsøk 2 før forsøksstart, og 3 og 4 veker etter badsmitte. Det var forskjell i vekt frå før forsøksstart til 3 og 4 veker etter smitte (One-Way Anova, Šídák's multiple comparisons test, **** = p < 0,0001). Det var og skilnad i vekt for rognkjeks mellom 20 og 26 dps (one tailed t-test, p = 0,03; ikkje vist). Gruppene er adskilt med vertikale liner. Dps – dagar etter smitte.
3.2 - Prevalens og Mengde S. salmonicida i blod og tarmvæske (qPCR)
3.2.1 - Forsøk 1
I forsøk 1 var kun eit fåtal fisk positive for S. salmonicida i blod eller tarmvæske (Tabell 8). For å bekrefte eigne analyser vart hjarteprøvar frå 31 badsmitta laksesheddarar tatt 8 VES sendt til Pharmaq Analytiq. Analysen bekrefta den låge prevalensen, med tre prøvar (Ct 18,4, 30,3 og 31,7) positive for S. salmonicida.
Tabell 8: Prevalens i blod og tarm 8 veker etter smitte for laks og rognkjeks i forsøk 1.
Antall Positive / antall analysert 1
Tarm
Blod
Gruppe
VES
DES
GAPDH 3
Spiro
GAPDH
Spiro
Laks Kontroll
8
56
23/38
0/23
21/30
0/21
Laks Kohabitant
8
56
13/30
0/13
22/25
0/22
Laks Badsmitta
8
56
9/40
0/9
29/33
2/29 (3/31 2)
Rognkjeks Kontroll
(uvaksinert og vaksinert)
8
56
Ingen Data
0/79
Ingen Data
0/78
Rognkjeks Uvaksinert Kohabitant
8
56
Ingen Data
0/158
Ingen Data
1/149
Rognkjeks Uvaksinert Badsmitta
8
56
Ingen Data
0/101
Ingen Data
2/197
Rognkjeks Vaksinert Badsmitta
8
56
Ingen Data
0/97
Ingen Data
0/92
1 Analyse av blod og tarm frå all fisk 8 veker etter smitte (Havforskingsinstituttet) og shedderfisk (Pharmaq Analytiq) viste svært låg prevalens. Andre prøvar vart derfor ikkje analysert.
2 31 prøvar analysert av Pharmaq Analytic
3 GAPDH-assayet (Tabell 5) er for laks, og ikkje påliteleg som amplifikasjonskontroll for prøvar frå rognkjeks
3.2.2 - Forsøk 2
I forsøk 2 var smittemodellen justert, med mildare vaskebetingingar, høgre smittedose, og salinitet redusert til 10 ‰. Rett etter uttaket 3 veker etter smitte oppsto det dødelegheit hos rognkjeks. Det vart påvist massive infeksjonar med S. salmonicida, og forsøket vart terminert i henhald til humant endepunkt og all fisk prøvetatt.
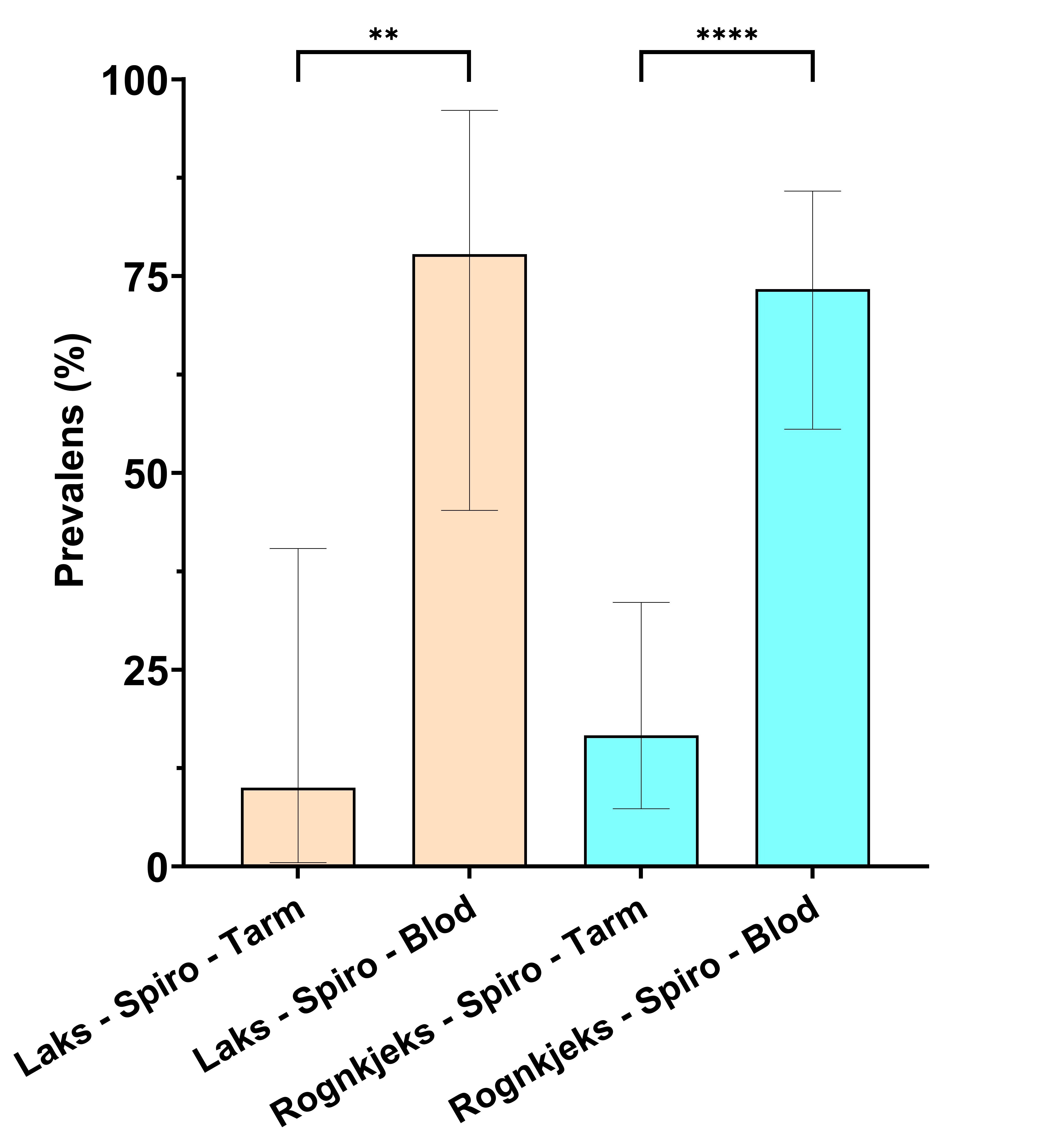
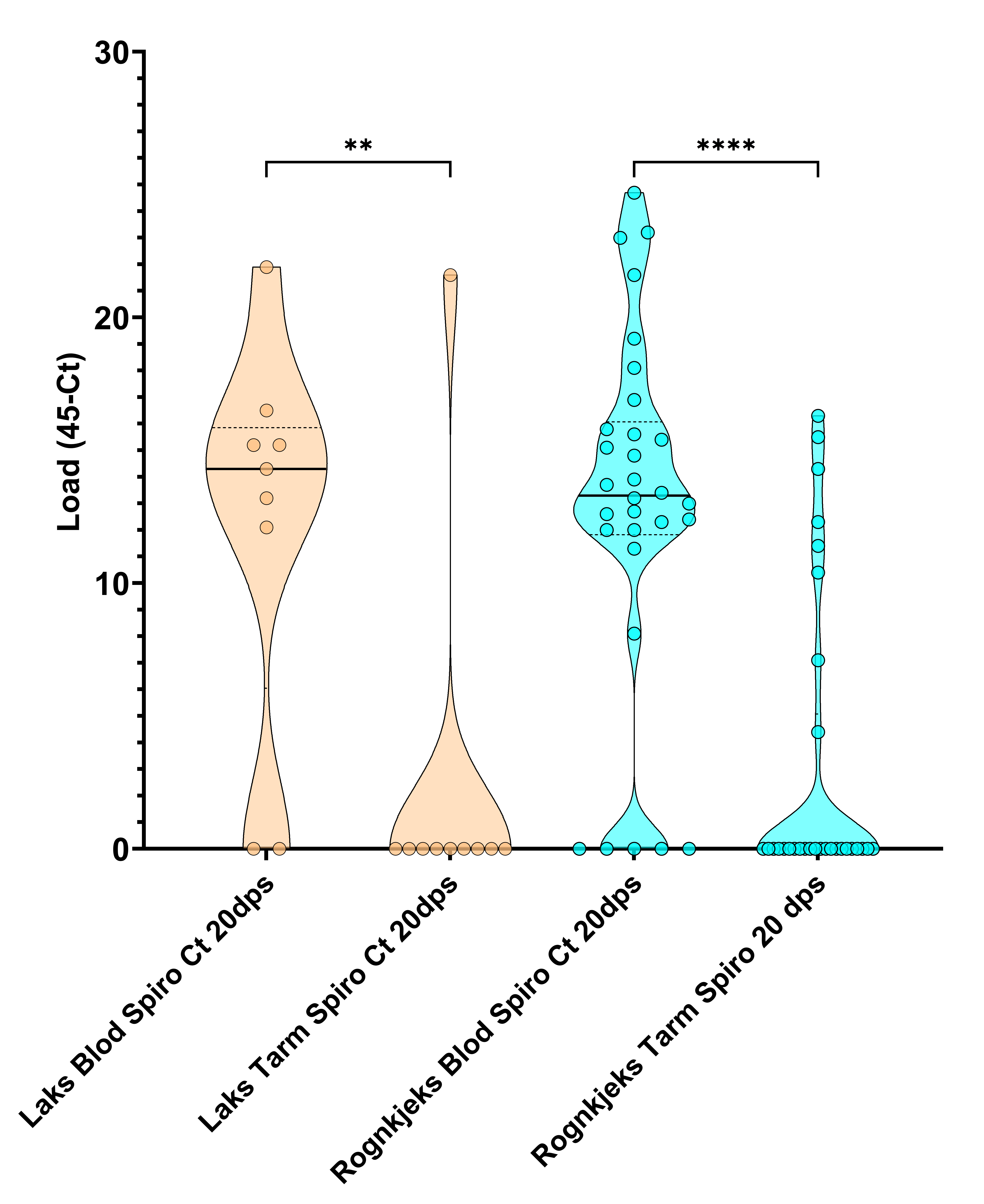
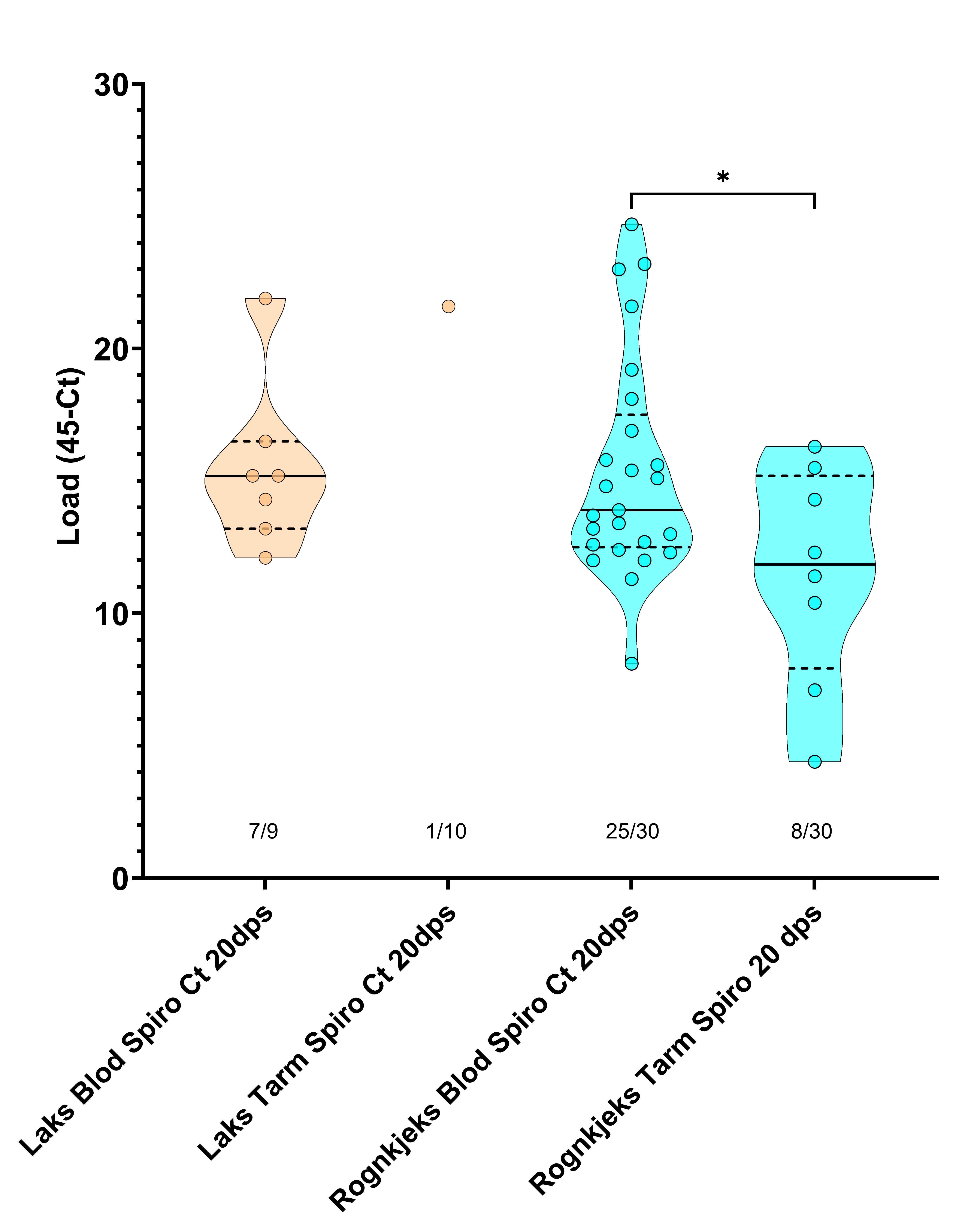
Både laks og rognkjeks var sterkt infisert av S. salmonicida i blod (Tabell 9, Figur 5, 6, 7) tre veker etter smitte. Tilnærma all laks (78 %) og rognkjeks (83 %) var smitta med S. salmonicida (Tabell 9). qPCR analysene indikerte store mengder S. salmonicida i blod og tarm i infisert fisk, med Load (45-Ct) frå 12,1-21,9 (laks) og 8,1-24,7 (rognkjeks) i blod (Figur 7).
Tabell 9. Prevalens i blod og tarm tre veker etter smitte for laks og rognkjeks i forsøk 2.
Antall Positive / antall analysert
Tarm
Blod
Gruppe
VES
DES
GAPDH
Spiro
GAPDH
Spiro
Laks Badsmitta
3
20
9/10
1/10
7/9
7/9
Rognkjeks Badsmitta
3
20
Ingen Data 1
8/30
Ingen Data
25/30
1 GAPDH-assayet (Tabell 5) er designa for laks, og er ikkje påliteleg som amplifikasjonskontroll for prøvar frå rognkjeks
Figur 5. Prevalens av S. salmonicida i tarmsvaber- og blodprøvar hos laks og rognkjeks tre veker etter smitte i forsøk 2.
Figur 6. Trunkert fiolinplot for mengde S. salmonicida i laks og rognkjeks i forsøk 2 tre veker etter badsmitte. Alle fiskar analysert er vist. Det var forskjell i mengde S. salmonicida mellom tarm og blod. Dps – dagar etter smitte.
Figur 7. Trunkert fiolinplot for mengde (Load (45-Ct)) S. salmonicida i laks og rognkjeks i forsøk 2 tre veker etter badsmitte. Kun fisk positive for S. salmonicida er vist, og her var det kun forskjell (p < 0,05) i mengde S. salmonicida mellom tarm og blod for rognkjeks. Dps – dagar etter smitte.
3.3 - Kliniske teikn og Histologi
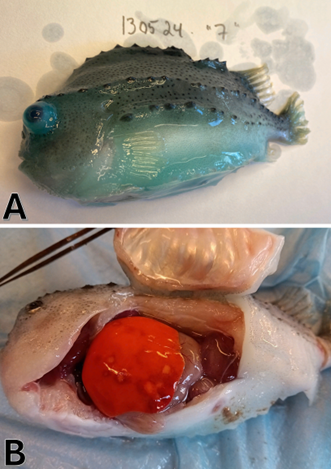
Generelt vart det få makroskopiske teikn på og i rognkjeksene som vart undersøkt. Det vart ikkje gjort makroskopiske funn på laks. Ved uttaket 3 veker etter smitte vart det notert enkelte rognkjeks med eksoftalmi, oftast einsidig. Ved sluttuttaket 4 veker etter smitte vart det funne fleire rognkjeks med eksoftalmi, nokre med til dels betydeleg, på eit eller begge auga. Det vart og funnet nokre fisk med lesjonar på lever (Figur 8) ved autopsien 4 veker etter smitte. Her vart det og observert at rognkjeksene hadde mat i magen ved uttak, og hadde spist fram til dei vart avliva.
Histologisk materiale av rognkjeks frå forsøk 2 er opparbeida i mastergradsarbeidet til Astrid Lofnes (2024-25), og nærare skildring av materialet og funn kan ein finne der (Lofnes, 2025).
Figur 8. A: Rogkjeksindivid med kraftig eksoftalmi (blod Ct = 31,7; Foto: Sandlund, N.). B. Rogkjeksindivid med fleire nekrotiske områder på lever (blod Ct = 28,1; Foto: Karlsbakk, E.)
3.4 - Dødelegheit
Det var inga dødelegheit i forsøk 1
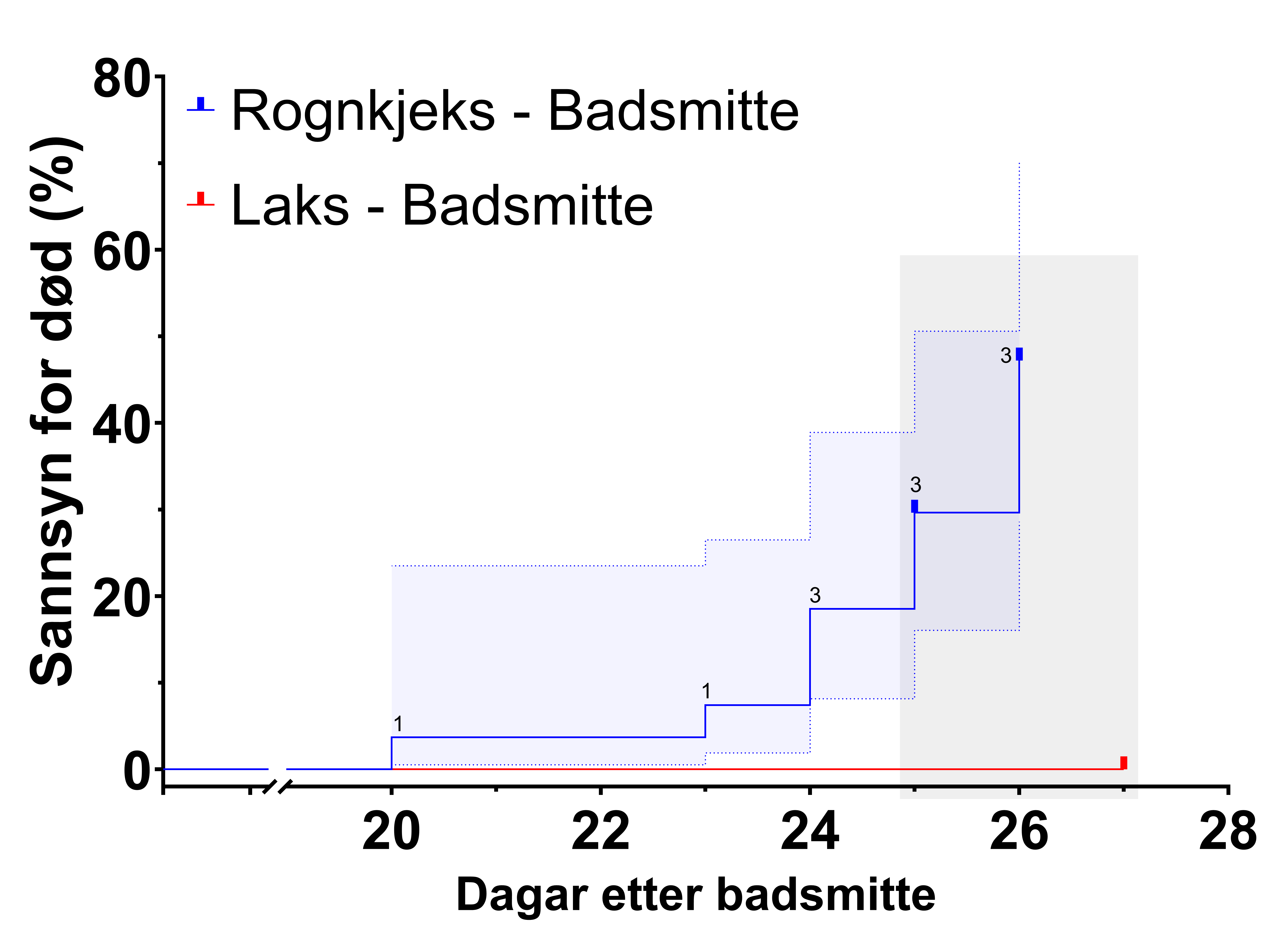
I forsøk 2 var det inga dødelegheit mellom dei badsmitta laksane, men det var vedvarande dødelegheit for rognkjeks frå 23 dagar etter smitte (Figur 9). Forskjellen i dødelegheit mellom laks og rognkjeks var signifikant (p<0.05, log-rank test).
Ved mitdtuttaket «3 veker» etter smitte, 20 dagar etter badsmitte, vart det funnet ei daud rognkjeks ved start av uttaket. Denne var lytefri, med unnatak av svak eksoftalmi på eit auge. Tre dagar etter midtuttaket vart det funnet ei daud rognkjeks og det vart mistanke om sjukdom. Søndag, 24 dagar etter smitte, vart ytterlegare 3 daude rognkjeks funnet og undersøkt makroskopisk og mikroskopisk, med påvising av store mengder S. salmonicida i blod og vevsimprint. Fisken hadde pellet i magen, so den døydde akutt.
Med bakgrunn i dødelegheita vart forsøk 2 avslutta med uttak av prøvar frå all fisk 25-27 dagar etter smitte. Det var 3 svimarar/daude rognkjeks før uttak både dag 25 og 26 etter smitte, alle inkludert som daude i analyse av dødelegheit. All rognkjeks var avliva og ferdig prøvetatt dag 26 etter smitte. Laksen vart avliva og prøvetatt dag 27 etter smitte.
Figur 9. Dødelegheitskurve for rognkjeks og laks i forsøk 2. Det var inga dødelegheit før dag 20 etter smitte og x-aksa er avkorta. Ingen laks døydde i forsøket. Antal svimar/daude rognkjeks er indikert for kvar dag det vart funnet daudfisk. Blått felt indikerar 95 % c.i., og grått felt indikerar tidsrom for sluttuttak. Log-rank test viste signifikant (p<0,05) forskjell i dødelegheit mellom laks og rognkjeks. All rognkjeks og laks avliva i sluttuttaket vart sensurert i analysa.
4 - Diskusjon
Målsetjinga med forsøket var å teste om S. salmonicida kan smitte rognkjeks gjennom vatn åleine (badsmitte) eller etter utskillelse frå infisert laks (kohabiteringssmitte). Det var og eit mål å få kunnskap om eventuelle konsekvensar av slik smitte i rognkjeks. Det var svært låg smittesuksess i forsøk 1. Resultata for smitte frå laks til rognkjeks var derfor svake, og vidare arbeid på dette er naudsynt, men den eine infiserte rognkjeksa kohabitert med infisert laks kan indikere at infisert laks kan smitte rognkjeks. Resultata frå forsøk 2 var derimot gode. Forsøket viste S. salmonicida fritt i vatn infiserar rognkjeks, at rognkjeksa er svært mottakeleg for smitte, og at konsekvensane av S. salmonicida infeksjonar hos rognkjeks kan være alvorlege.
I forsøk 1, hovudforsøket, var det svært avgrensa smittesuksess, og kun eit fåtal badsmitta laks og rognkjeks vart smitta. Etter enderingar i førebuinga av smittemateriale og gjennomføring av badsmitte fekk vi derimot svært godt tilslag på badsmitten med S. salmonicida i forsøk 2. Her var prevalensen 3 veker etter smitte høg både for laks (78 %) og rognkjeks (83 %).
Moglege årsaker til forskjellane er blant anna endring i førebuing av smittematerialet, dosen, og/eller vasskvaliteten ved smitte. I forsøket der aure vart smitta med S. salmonicida (Karlsbakk, Nordbø, Austgulen, et al., 2025) var der medierestar att i smittematerialet, som førte til skumming i karet. Det var då vanskelegare å observere fisken, og verknaden av synkande oksygen. For å unngå dette vart parasitten «vaska» betre enn tidlegare for å fjerne meir vekstmedium i smittematerialet. Vaskeprosedyra innebar fleire sentrifugeringar, skifting av PBS og tok tid. PBS har osmolaritet (ca 300 mOsm/L) omtrent tilsvarande fysiologisk saltvatn (9 ‰; 308 mOsm/L). Salthaldet i smittekaret var i forsøk 1 den same som vasskvaliteten nytta under sjølve forsøket - ca 25 ‰. Overlevinga til S. salmonicida er best ved 10 ‰ (Helsø 2024), og miljøet i smittekaret vil derfor kunna ha påverka smitteeffektiviteten negativt. I pilot og aure forsøka var det ca 25 ‰ promille ved smitte, og god smittesuksess. Mindre gunstige forhold utan næring, handtering og tidsbruken kan alle være faktorar som reduserte vitalitet og infektivitet til parasitten slik at smitte ikkje kunne etablerast effektivt. Årsaka til den låge smittesuksessen i dette forsøket er derfor ikkje kjent, og det er behov for ytterlegare metodeutvikling.
Då hovedmålet med forsøket var å eksponere rognkjeks for eit smittepress som smittar laks, vart smittedosen i forsøk 2 auka betydeleg, vaskeprosedyre justert til mildare betingingar og raskare gjennomføring, og saliniteten brukt i smittekaret satt til ca 10 ‰. Dette ga god smitteeffektivitet, og betingingane kan være utgangspunkt for kommande badsmitteforsøk. Det er ikkje kjent om betringa i smitteeffektiviteten skuldast endring i ein av enkeltfaktorane eller kombinasjonen av endringane. Vi hadde og godt tilslag på smitten i forsøket med aure, der ein annan vaskeprotokoll vart nytta i førebuing av smittematerialet (Karlsbakk, Nordbø, Austgulen, et al., 2025). Sidan det framleis er begrensa erfaring med modellen, og at både dyrking og handtering av parasitten er utfordrande, er det naudsynt med optimalisering av både produksjon av smittemateriale og smittemodellen. Etableringa av robuste protokollar for dyrking og smitte er derfor noko som krev vidare arbeid.
Som i pilotstudiet (Kvamme et al., 2025) var det stor forskjell på prevalens mellom blod og tarm i dette studiet. Dette gjaldt for både laks og for rognkjeks. Resultatet er ikkje som forventa ut frå tidlegare studier, samt at S. salmonicida truleg er ein tarmparasitt i dei naturlege vertane. Ei hypotese har vore at den valgte samplingsprotokollen og/eller analysemetoden ikkje er optimal for påvising av parasitten. Truleg er flagellatane vanlegast i framtarmen, og det vart og sett i mikroskopiering gjort. Men det synest lite truleg at ein infeksjon i framtarmen ikkje er påviseleg bakerst i tarmen med qPCR, med mindre tarminnhaldet inneheld inhibitorar for PCR. Det er mogleg at metoden for analyse av tarmprøvar for S. salmonicida kan optimaliserast ut over det vi har hatt anledning til i dette prosjektet. Men, analyser av forekomst av S. salmonicida i ulike vev frå bærarfisk frå ulike artar viste tilsvarande store ulikeheiter mellom ulike vevsprøvar, som det vi har funnet mellom prøvar tatt med tarmsvaber og blodprøvar (Lofnes, 2025). Lofnes (2025) konkluderte at påvising (post mortem) av S. salmonicida var best gjort i hodenyre hos laks, men fortarm og gjelle i røyr. For begge desse artane kom analyse av vevsprøvar frå tarm, tatt rett bak siste caecum, ut med låg påvisningsgrad samanlikna med dei fleste andre veva testa. Dette kan indikere at prøvar av tarm ikkje er godt eigna for å påvise infeksjonar av S. salmonicida.
I forsøk 1 var det ikkje vekst på laksen i forsøket verken for kontroll, badsmitta eller kohabitantar. Årsaka til dette er ikkje kjent. Rognkjeksa vaks derimot gjennom heile forsøksperioden. Men, som forventa ut frå at det ikkje var smitte i nokon grupper, og at forholda elles var like, var det ikkje forskjell i vekt mellom dei ulike forsøksgruppene 8 veker etter smitte.
I forsøk 2 vaks både laksen og rognkjeksa i forsøksperioden. Rognkjeksa vaks godt også i siste del av forsøket då dødelegheita starta. Det er interessant at rognkjeksa, med høge mengder parasitt i blodet hos dei fleste individa, og etter kvart akutt dødelegheit, hadde ete nok til å ha ein auke (one-tailed t-test, p=0,03) i vekt dei 5-6 dagane mellom uttaket 3 veker etter smitte til avslutning av forsøket. Det vart og funnet mat i magen til rognkjeksa som døydde og i individer ved sluttuttaket. Appetitt og vekst påverkast ofte av sjukdom. Funna tyder på at S. salmonicida infeksjonen ikkje utgjorde ei høg belastning for rognkjeksa, og dødelegheita var akutt.
Til tross for den låge smitteeffektiviteten i forsøk 1 er nokre forsiktige tolkningar av resultata mogleg. Det vart påvist smitte hos ei kohabiterande rognkjeks som gjekk saman med badsmitta laks. Dette indikerar at smitte frå laks til rognkjeks kan skje. Det vart kun påvist S. salmonicida infeksjon i 2-3 badsmitta laks, og ingen kohabiterande laks vart påvist infisert 8 veker etter smitte. Smittepresset i karet må derfor ha vore lågt i heile forsøksperioden. Saman med resultata frå forsøk 2, indikerar dette at rognkjeksa er meir mottakeleg for smitte med S. salmonicida enn laks. Sidan resultata frå forsøk 1 er svært svake bør dette undersøkast nærare. Det er og ei rekkje andre kunnskapshol. Smittevegen ved kohabitering er ukjent, og kan vere fritt gjennom vatn, via fysisk kontakt, frå napping eller at rognkjeksa et faeces. Det er heller ikkje kjent om smitte kan skje frå rognkjeks til laks, eller om andre rensefisk eller andre marine fisk kan bli infisert med S. salmonicida. Det er derfor tilråeleg å følgje opp desse resultata.
I forsøk 2 fekk vi derimot svært godt tilslag på badsmitte med S. salmonicida. Prevalensen 3 veker etter smitte var høg både for laks (78 %) og rognkjeks (83 %). All infisert fisk hadde til dels høge mengder parasitt i blodet. Det var då ikkje forskjell i mengda parasitt i blod hos laks og rognkjeks som var infisert. Resultata viste utvetydig at rognkjeks er mottakeleg for S. salmonicida fritt i vatnet, og er i samsvar med funna gjort for rognkjeks frå anlegg med spironukleose hjå laks.
Det var ikkje adferdsmessige eller makroskopiske teikn til sjukdom på verken laks eller rognkjeks 3 veker etter smitte. Som diskutert i rapporten frå pilotforsøket er dette som forventa for laks (Kvamme et al., 2025), og i tråd med tidlegare gjennomførte forsøk i SpiroFri-prosjektet (Karlsbakk, Nordbø, Austgulen, et al., 2025; Karlsbakk, Nordbø, Skår, et al., 2025; Kvamme et al., 2025), og andre studier (Alfjorden, 2018; Alfjorden et al., 2019; Guo & Woo, 2004; Kent et al., 1992; Mo et al., 1993). Det samsvarar og med erfaring frå utbrota i næringa.
For rognkjeks finnest det ikkje data for S. salmonicida infeksjonar og sjukdomsforløp. Som for laks vart det ikkje funnet teikn på sjukdom ved uttaket 3 veker (21 dagar) etter smitte, og fisken var stort sett lytefri. Likevel oppstod det uventa akutt dødelegheit 23 dagar etter smitte hos rognkjeks.
I laks er spironukleose ofte ein langsomt utviklande sjukdom med førekomst av lesjonar først etter 6-8 veker etter smitte og mortalitet, om nokon, etter dette (Alfjorden, 2018; Alfjorden et al., 2019; Guo & Woo, 2004; Karlsbakk, Hansen, et al., 2025; Kent et al., 1992; Kvamme et al., 2025; Mo et al., 1993). Det vart og kun funne moderate lesjonar og lite histopatologi i rognkjeks. Dette indikerar at mekanismene for akutt dødelegheit hos rognkjeks som følgje av S. salmonicida infeksjon ikkje skuldast akkumulerande vevsskader. Den faktiske årsaka til den akutte dødelegheita som S. salmonicida forårsaka hos rognkjeks er ikkje kjend.
5 - Konklusjonar
Rognkjeks er mottakeleg for S. salmonicida fritt i vatn.
Infisert laks kan smitte rognkjeks.
Rognkjeks blir alvorleg sjuk og kan døy som følgje av infeksjon.
Rognkjeks bør ikkje nyttast på lokalitetar med S. salmonicida infeksjonar
6 - Referansar
Adam, R. D. (2017). Diplomonadida. In J. M. Archibald, A. G. B. Simpson, C. H. Slamovits, L. Margulis, M. Melkonian, D. J. Chapman, & J. O. Corliss (Eds.), Handbook of the Protists (pp. 1–28). Springer International Publishing. https://doi.org/10.1007/978-3-319-32669-6_1-1
Alfjorden, A. (2018). Experimental Spironucleus infections in Atlantic salmon (Salmo salar) - Hidden secrets of the life cycle of Spironucleus salmonicida revealed Uppsala University. MSc oppgåve.Uppsala, Sweden.
Alfjorden, A., Astvaldsson, A., Jansson, E., & Svärd, S. (2019). Experimental challenge of Atlantic salmon (Salmo salar) with the diplomonad parasite Spironucleus salmonicida to characterize the infection cycle.
Fard, M. R. S., Jorgensen, A., Sterud, E., Bleiss, W., & Poynton, S. L. (2007). Ultrastructure and molecular diagnosis of Spironucleus salmonis (Diplomonadida) from rainbow trout Oncorhynchus mykiss in Germany. Diseases of Aquatic Organisms, 75(1), 37–50. https://doi.org/10.3354/dao075037
Grieg Seafood ASA. (2023). Annual Report 2023.
Guo, F. C., & Woo, P. T. (2004). Experimental infections of Atlantic salmon Salmo salar with Spironucleus barkhanus. Dis Aquat Organ, 61(1-2), 59–66. https://doi.org/10.3354/dao061059
Helsø, M. (2024). Miljøoverlevelse hos Spironuclues salmonicida - parasitten som forårsaker systemisk spironukleose hos oppdrettslaks. MSc oppgåve. Universitetet i Bergen.
Johansen, R., Kongtorp, R. T., Bornø, G., Skjelstad, H. R., Olsen, A. B., Flesjå, K., Colquhoun, D., Ørpetveit, I., Hansen, H., Garseth, Å. H., & Hjeltnes, B. (2009). Fiskehelserapporten 2008.
Jorgensen, A., & Sterud, E. (2006). The marine pathogenic genotype of Spironucleus barkhanus from farmed salmonids redescribed as Spironucleus salmonicida n. sp. Journal of Eukaryotic Microbiology, 53(6), 531–541. https://doi.org/10.1111/j.1550-7408.2006.00144.x
Karlsbakk, E., Hansen, H., Sterud, E., Tjessem, A., & Heggebø, R. (2025). Spironucleus-infeksjoner og Spironucleose hos laks – En kunnskapsoppsummering (2025). Havforskningsinstituttet.
Karlsbakk, E., Nordbø, J., Austgulen, M. H., Berg-Rolness, H., Mæhle, S., Ghebretnsae, D. B., Røttingen, T. B., & Kvamme, B. O. (2025). Er sjøaure mottagelig for Spironucleus salmonicida – parasitten som kan forårsake systemisk spironucleose i oppdrettslaks? (2025). Havforskningsinstituttets rapportserie. Havforskningsinstituttet.
Karlsbakk, E., Nordbø, J., Skår, C. K., Berg-Rolness, H., Mæhle, S., Ghebretnsae, D. B., Aga, J., Sengee, A., Lofnes, A., Røttingen, T. B., & Kvamme, B. O. (2025). Hvor mottagelig er sjørøyr for den laksepatogene flagellaten Spironucleus salmonicida? (2025). Havforskningsinstituttets rapportserie.
Keister, D. B. (1983). Axenic culture of Giardia lamblia in TYI-S-33 medium supplemented with bile. Transactions of the Royal Society of Tropical Medicine and Hygiene, 77(4), 487–488. https://doi.org/10.1016/0035-9203(83)90120-7
Kent, M. L., Ellis, J., Fournie, J. W., Dawe, S. C., Bagshaw, J. W., & Whitaker, D. J. (1992). Systemic hexamitid (Protozoa, Diplomonadida) infection in seawater pen-reared chinook salmon Oncorhynchus tshawytscha. Diseases of Aquatic Organisms, 14(2), 81–89. https://doi.org/10.3354/dao014081
Kvamme, B. O., Mæhle, S., Berg-Rolness, H., austgulen, M. H., Sengee, A., Ghebretnsae, D. B., Nordbø, J., Skår, C. K., Penaranda, M. M. D., Skaftnesmo, K. O., & Karlsbakk, E. (2025). Utprøving av fem ulike Spironucleus salmonicida smittemodellar for postsmolt laks i sjøvatn - SpiroFri Pilotsmitteforsøk Våren 2023 (2025-01). Havforskningsinstituttet. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-2025-25
Lie, K.-I., Wisløff, H., Østevik, L., Alacorn, M., & Nylund, S. (2022). Spironukleose hos atlantisk laks. Norsk Veterinaer Tidsskrift, 134(06-2022), 400–401.
Lofnes, A. (2025). Spironucleus salmonicida – histopatologi og vevstropisme i ulike vertsarter. MSc-oppgåve. Universitetet i Bergen.
Miller, K. M., Gardner, I. A., Vanderstichel, R., Burnley, T., Schulze, A. D., Li, S., Tabata, A., Kaukinen, K. H., Ming, T. J., & Ginther, N. G. (2016). Report on the Performance Evaluation of the Fluidigm BioMark Platform for High-Throughput Microbe Monitoring in Salmon. D. C. S. A. Sec.
Mo, T. A., Poppe, T., & Appleby, C. (1993). Kohabiteringsforsøk med fire ulike arter laksefisk og en encellet parasitt i slekten "Hexamita". Norsk Veterinærtidsskrift, 105(3).
Mo, T. A., Poppe, T. T., & Iversen, L. (1990). Systemic hexamitosis in salt-water reared Atlantic salmon (Salmo salar L.). Bull. eur. ass. Fish Pathol, 10(3).
Paull, G. C., & Matthews, R. A. (2001). Spironucleus vortens, a possible cause of hole-in-the-head disease in cichlids. Diseases of Aquatic Organisms, 45(3), 197–202. https://doi.org/10.3354/dao045197
Poppe, T. T., Mo, T. A., & Iversen, L. (1992). Disseminated hexamitosis in sea-caged atlantic salmon Salmo salar. Diseases of Aquatic Organisms, 14(2), 91–97. https://doi.org/10.3354/dao014091
Poynton, S. L., Fard, M. R. S., Jenkins, J., & Ferguson, H. W. (2004). Ultrastructure of Spironucleus salmonis n. comb. (formerly Octomitus salmonis sensu Moore 1922, Davis 1926, and Hexamita salmonis sensu Ferguson 1979), with a guide to Spironucleus species. Diseases of Aquatic Organisms, 60(1), 49–64. https://doi.org/10.3354/dao060049
Skjelstad, H. R., Bornø, G., Flesjå, K., Hansen, H., Nilsen, H., Wasmuth, M. A., & Hjeltnes, B. (2008). Fiskehelserapporten 2007.
Sommerset, I., Wiik-Nielsen, J., Moldal, T., Oliveira, V. H. S., Svendsen, J. C., Haukaas, A., & Brun, E. (2024). Fiskehelserapporten 2023.
Sommerset, I., Wiik-Nielsen, J., Oliveira, V. H. S., Moldal, T., Bornø, G., Haukaas, A., & Brun, E. (2023). Fiskehelserapporten 2022.
Sterud, E., Poppe, T., & Borno, G. (2003). Intracellular infection with Spironucleus barkhanus (Diplomonadida : Hexamitidae) in farmed Arctic char Salvelinus alpinus. Diseases of Aquatic Organisms, 56(2), 155–161. https://doi.org/DOI 10.3354/dao056155
Sterud, E., Poppe, T. T., Iversen, L., & Larsen, T. (2002). Men Spironucleus var ikke død. kyst.no/arkiv/men-spironucleus-var-ikke-dod/276649
Uldal, A., & Buchmann, K. (1996). Parasite host relations: Hexamita salmonis in rainbow trout Oncorhynchus mykiss. Diseases of Aquatic Organisms, 25(3), 229–231. https://doi.org/10.3354/dao025229