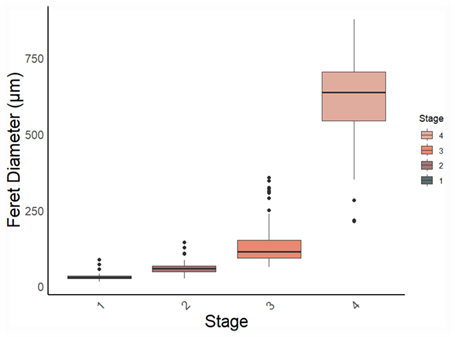
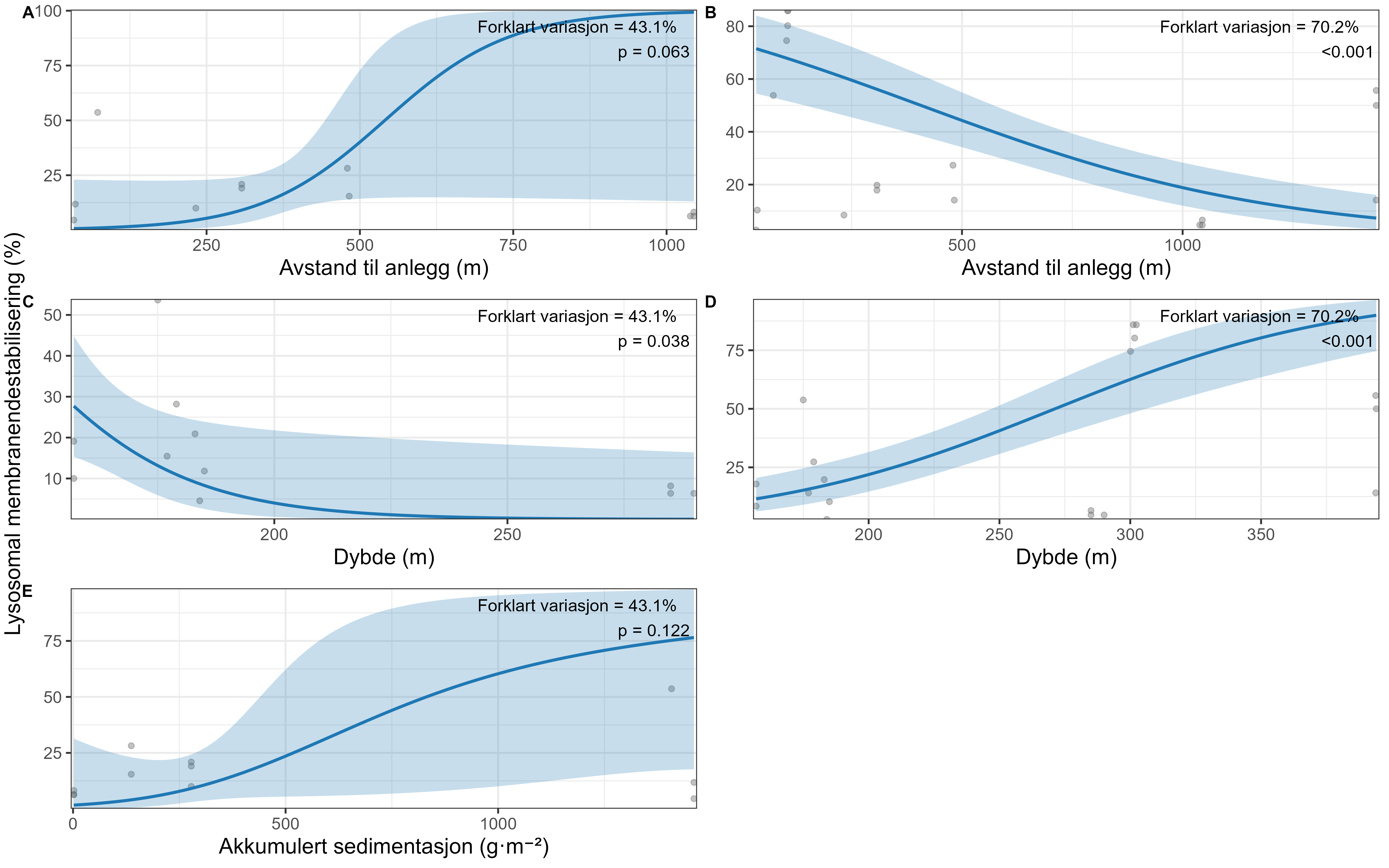
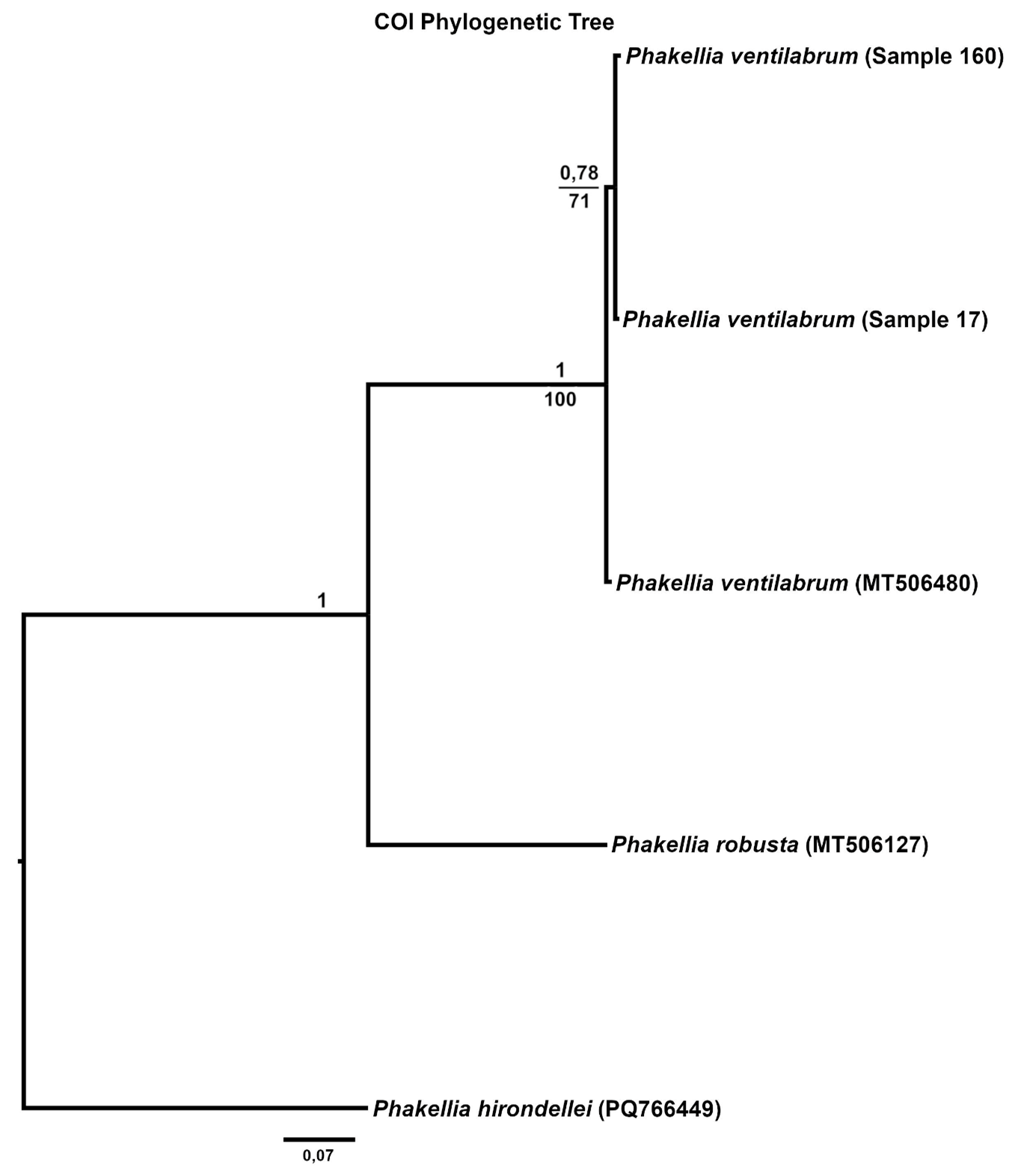
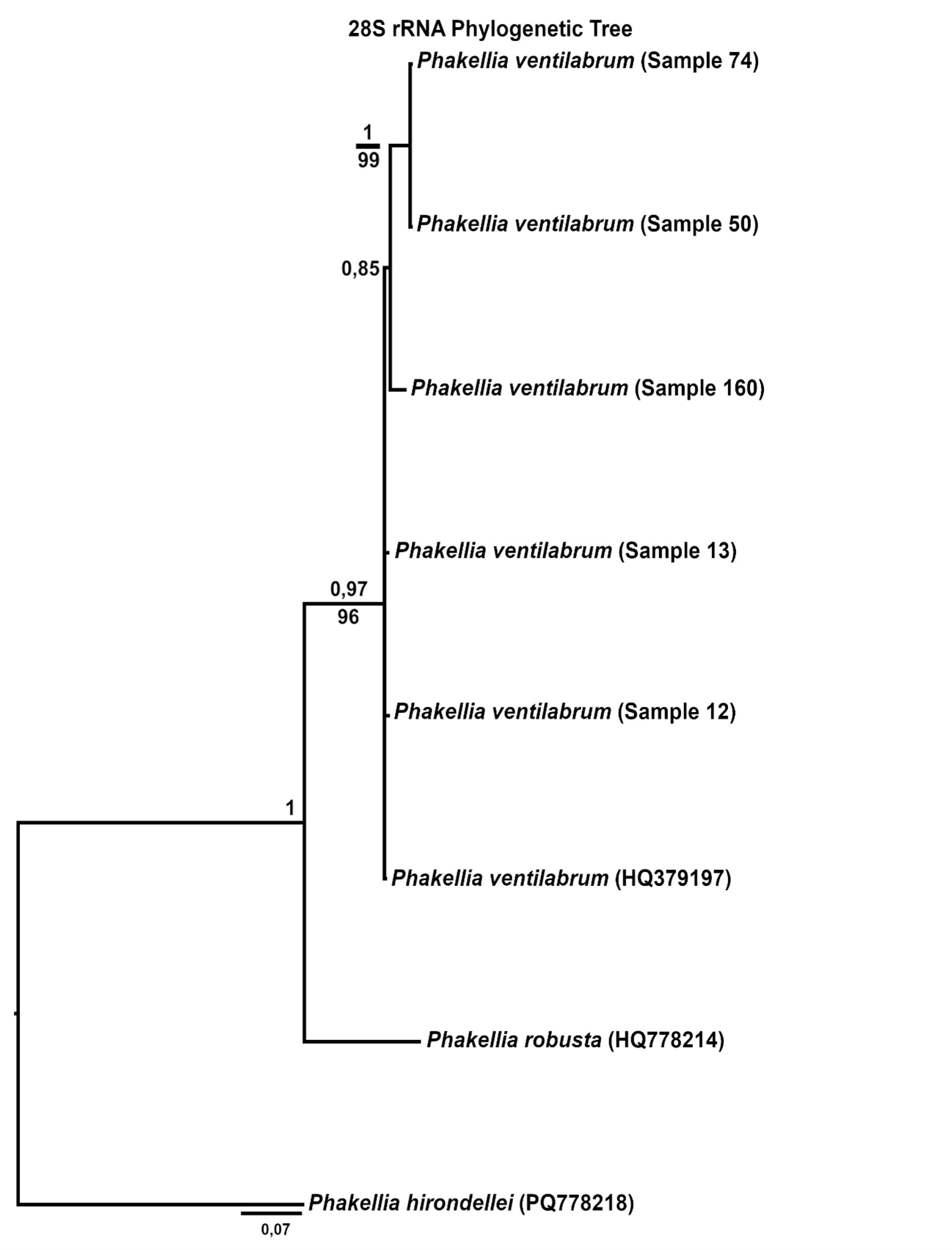
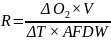The effects and tolerances of selected management-relevant species to organic fish farm waste and salmon lice medicines were investigated using newly developed field methods for assessing organism health. Focus species included the gorgonians Paragorgia arborea, Paramuricea placomus, Primnoa resedaeformis, the reef-forming stony coral Desmophyllum pertusum, the sponges Geodia barretti and Phakellia spp. and the bivalve Acesta excavata.
Samples from the selected species were collected by ROV from within the waste influence area of seven fish farms and from reference sites more than 750 m away along the coasts of Nordland and Møre og Romsdal. Modelled organic sedimentation (accumulated over two years prior to the collection of fauna samples) was compared with the Bacterial Metabarcoding Biotic Index (b-MBI), an indicator of organic enrichment based on the presence of bacterial indicator taxa. b-MBI values were positively correlated with modelled sedimentation and depth, reflecting moderate to high enrichment near farms and highlighting the method’s potential as an early-warning indicator of aquaculture impacts.
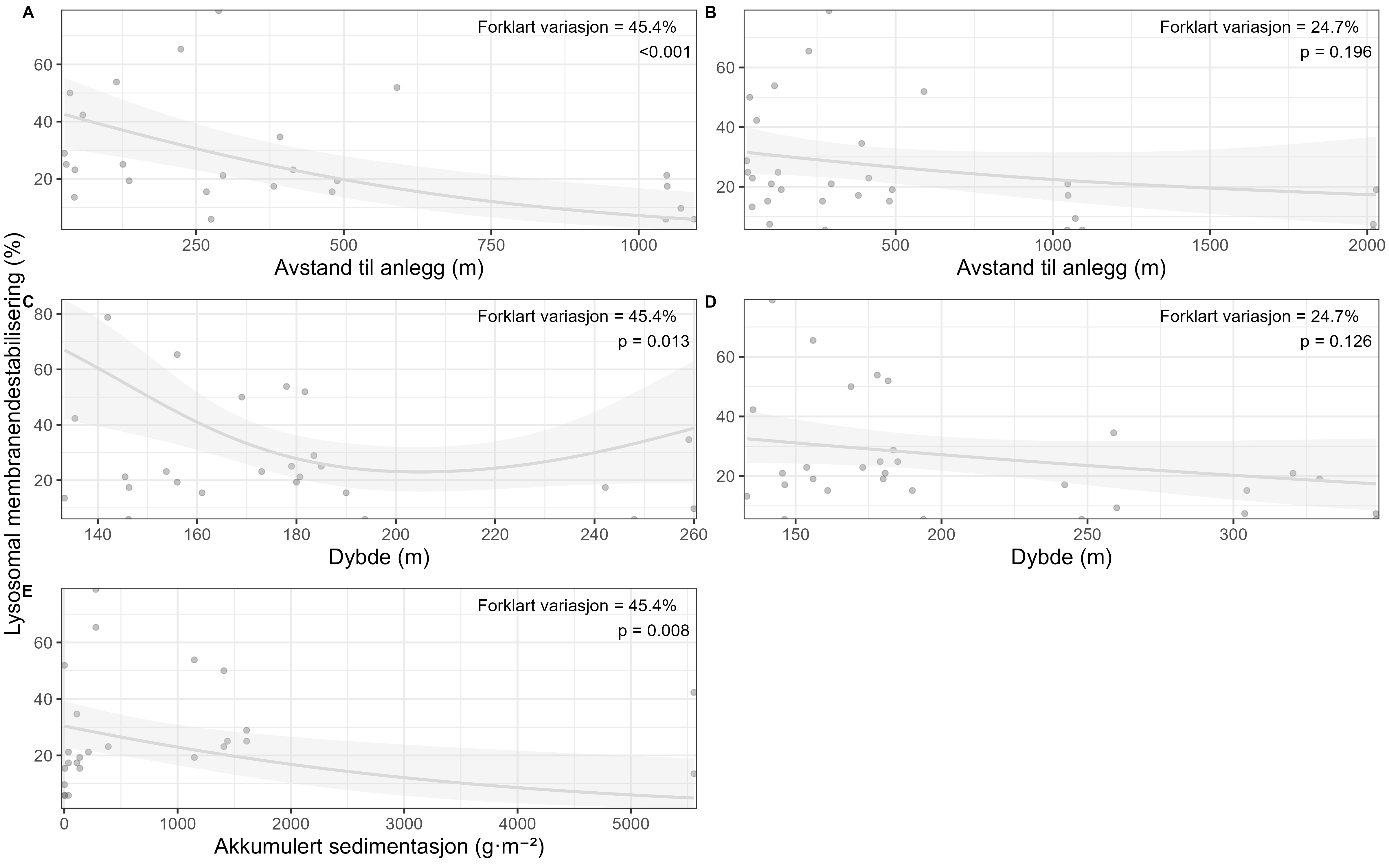
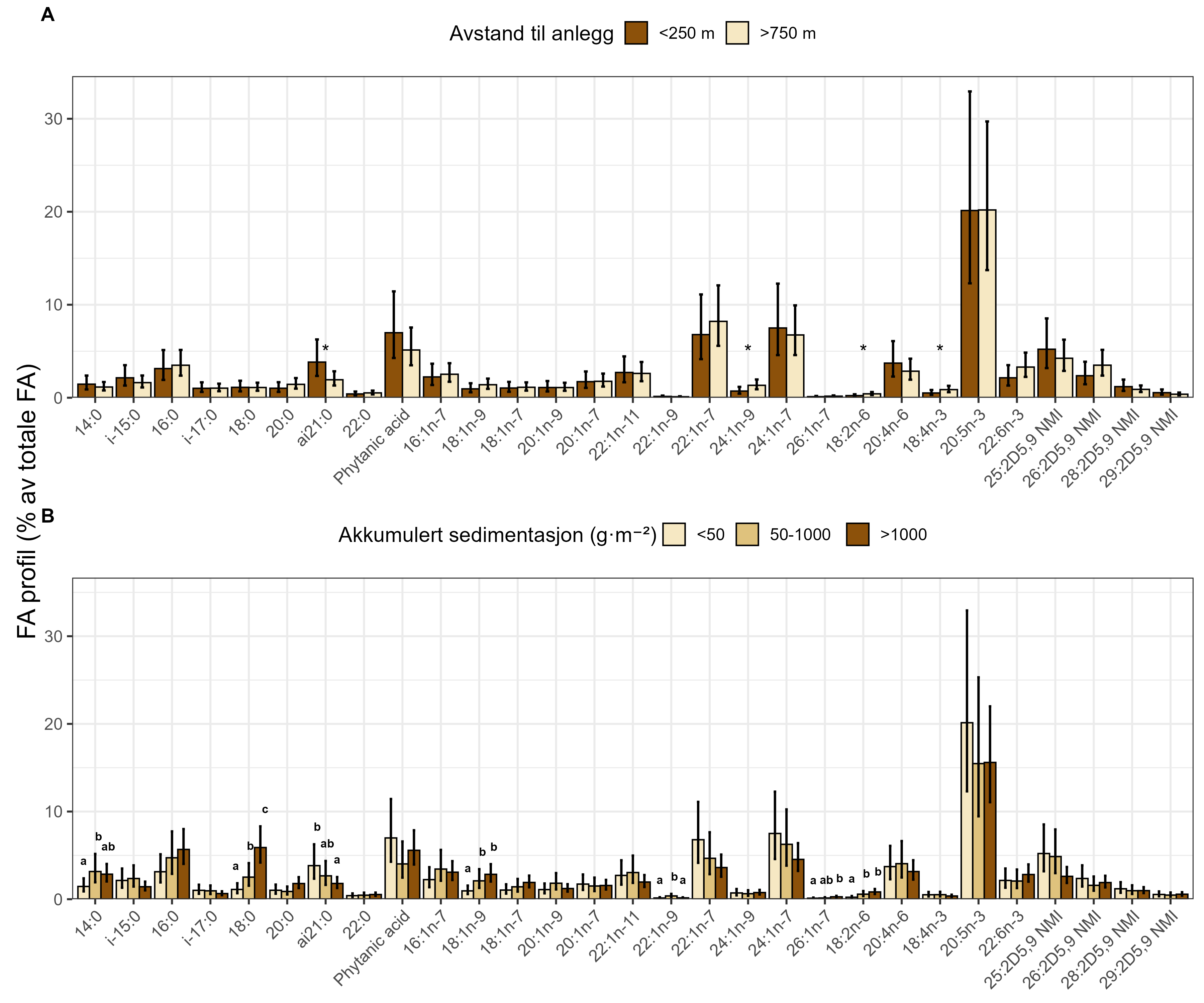
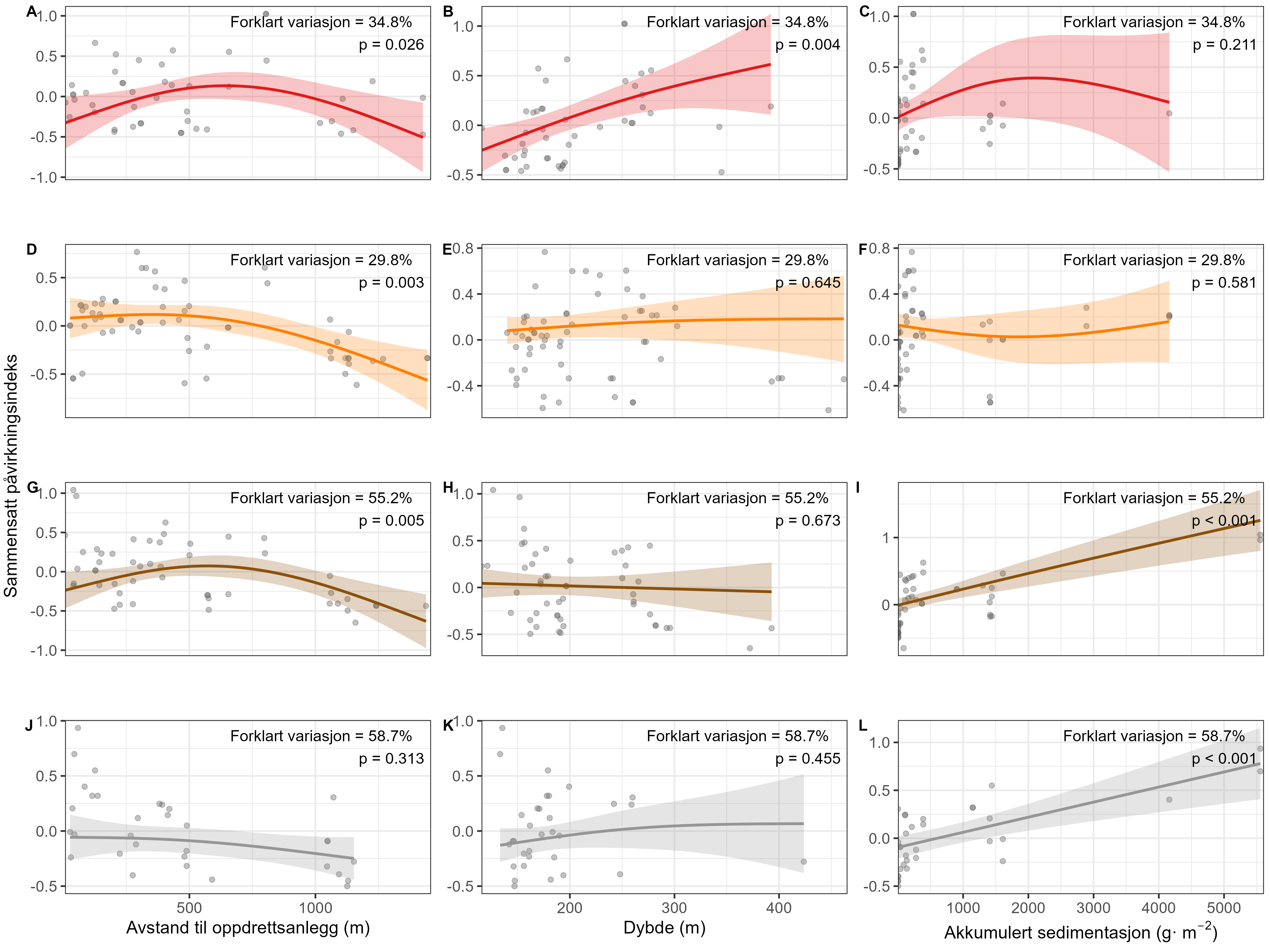
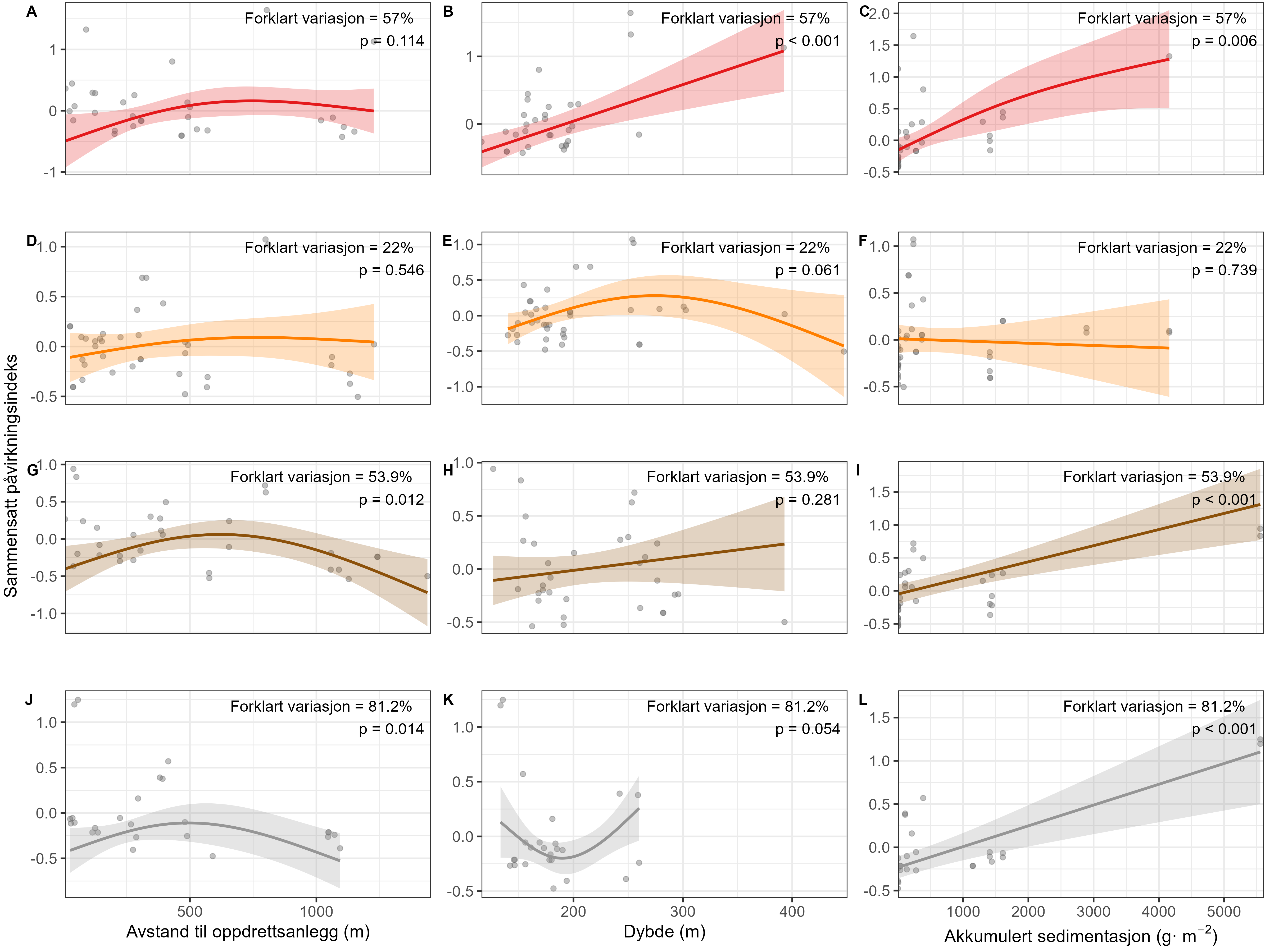
Visual index of negative health condition were positively correlated with modeled sedimentation for P. resedaeformis and Phakellia spp., showing reduced visual condition at higher sedimentation levels. The visual condition of G. barretti was also correlated with sedimentation although showing that some individuals remained unaffected. P. placomus showed reduced visual condition closer to the farm. Health criteria for P. arborea was more strongly related to depth than to distance from farms. Lysosomal membrane stability (LMS) analysis revealed a higher proportion of destabilized cells in the two sponges and A. excavata close to fish farms, suggesting sensitivity to aquaculture waste. The LMS method proved to be a rapid and cost-effective biomarker for field assessment of physiological stress.
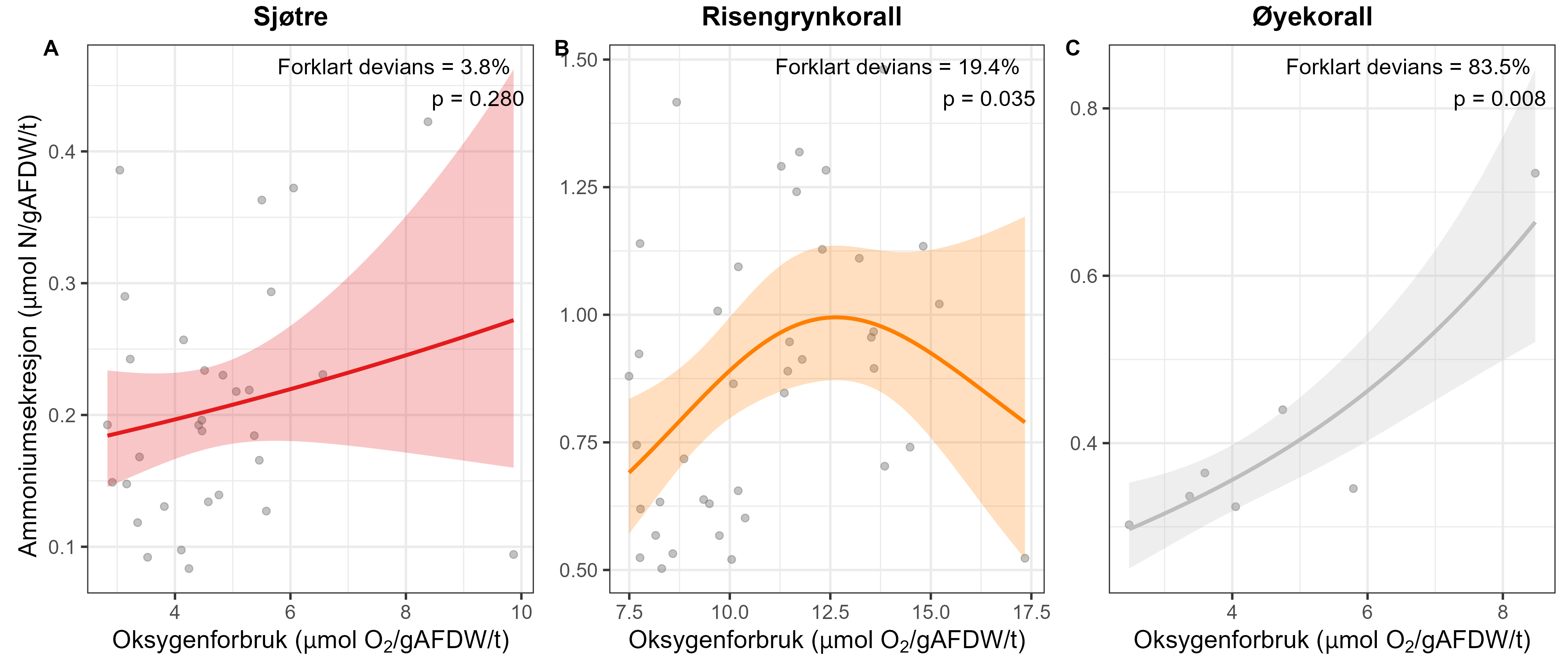
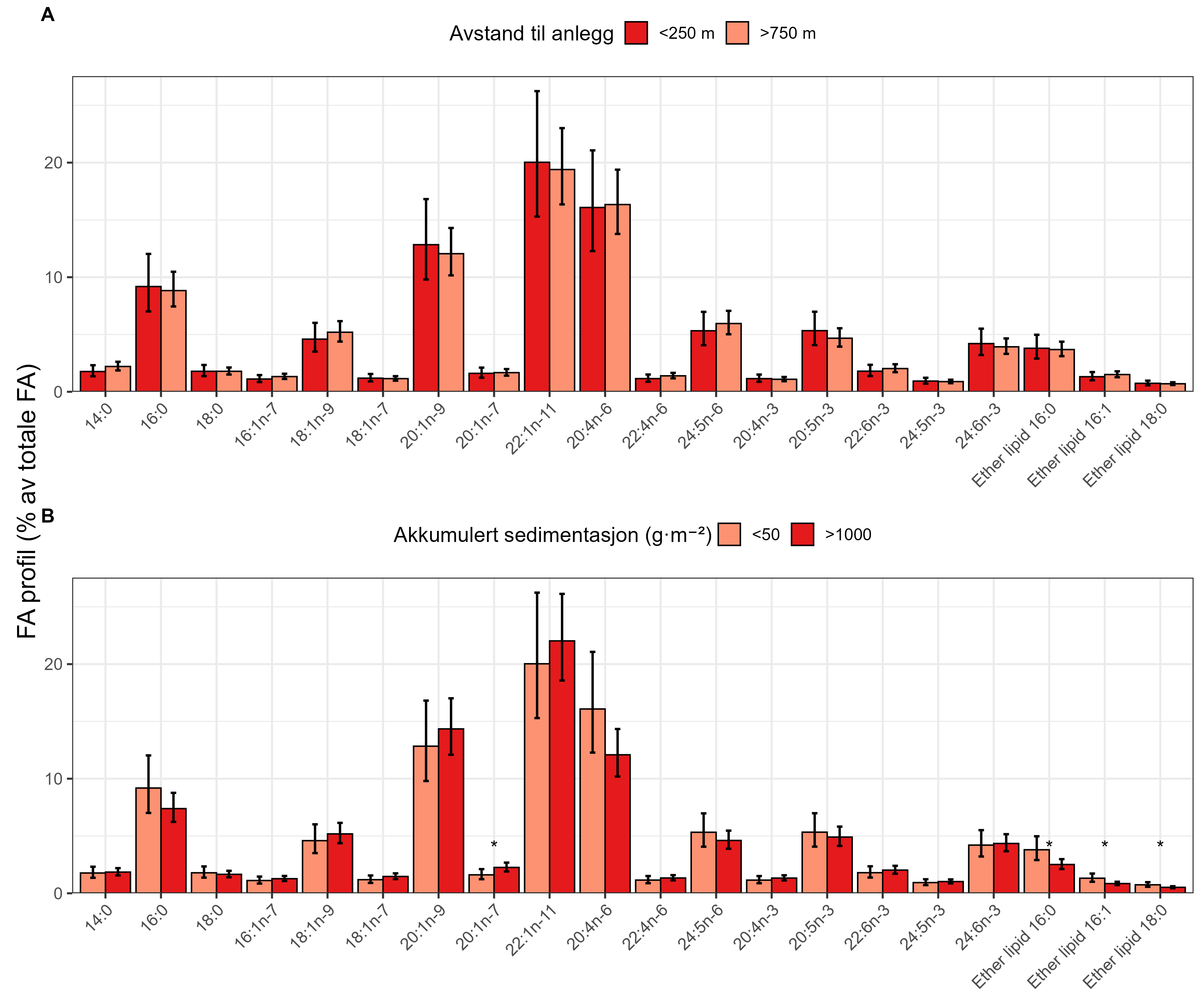
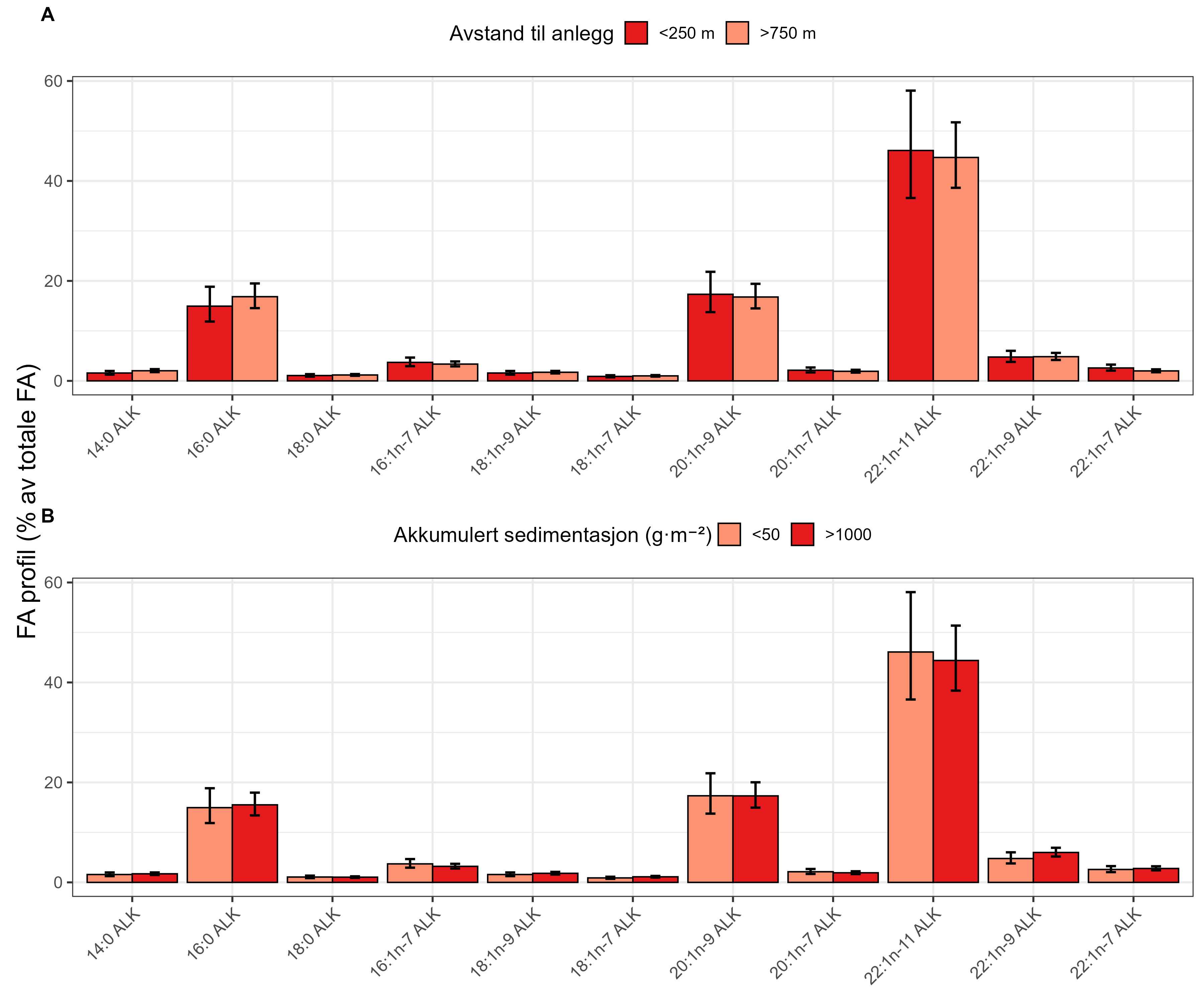
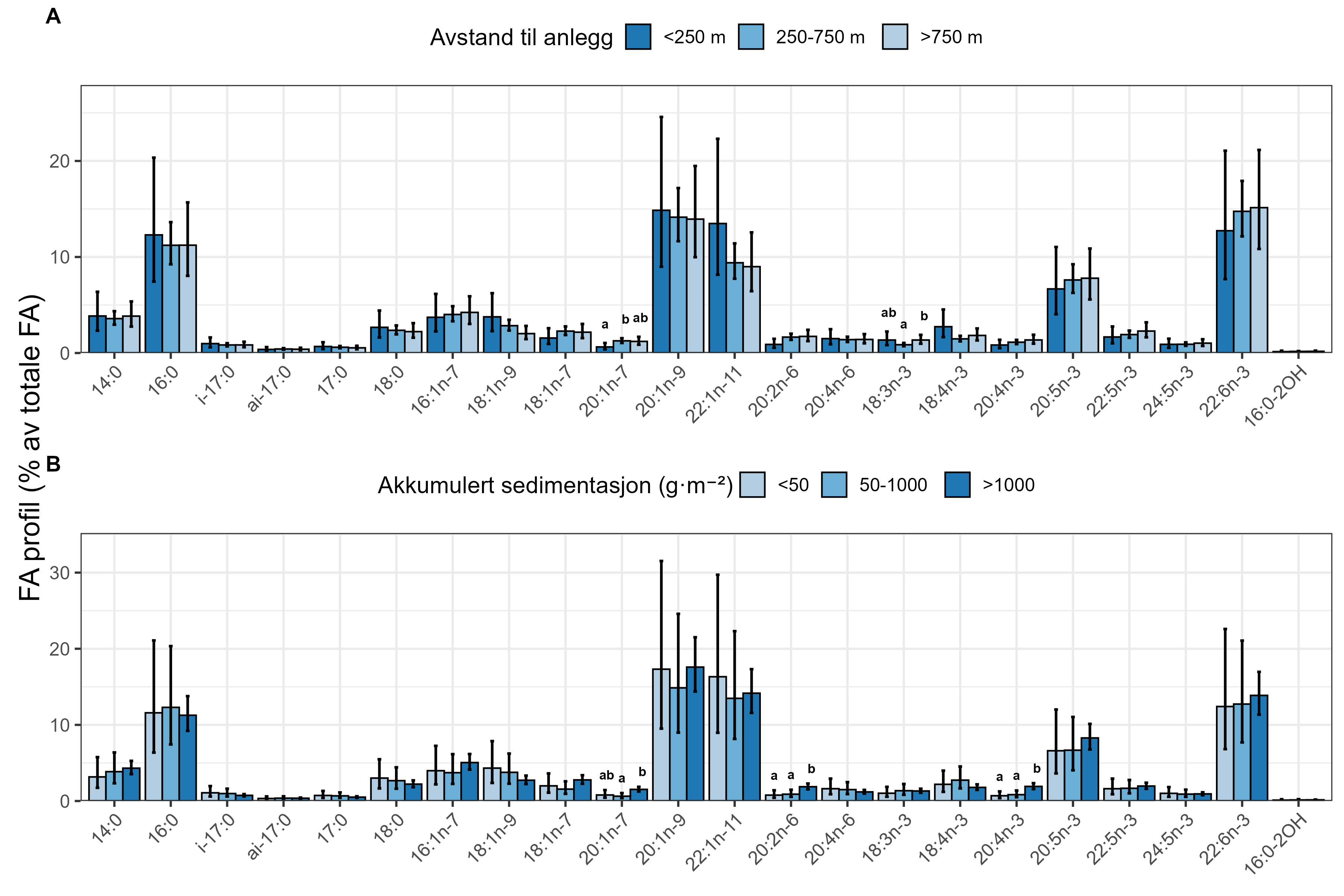
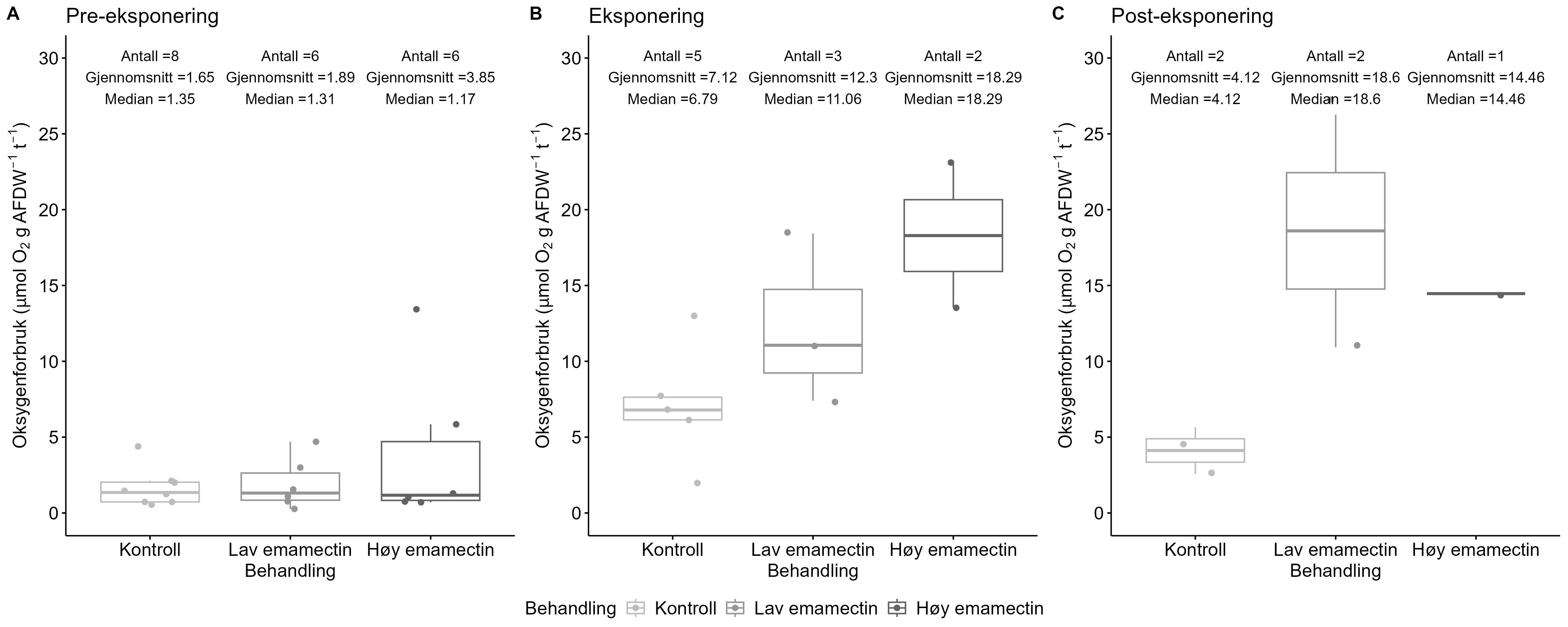
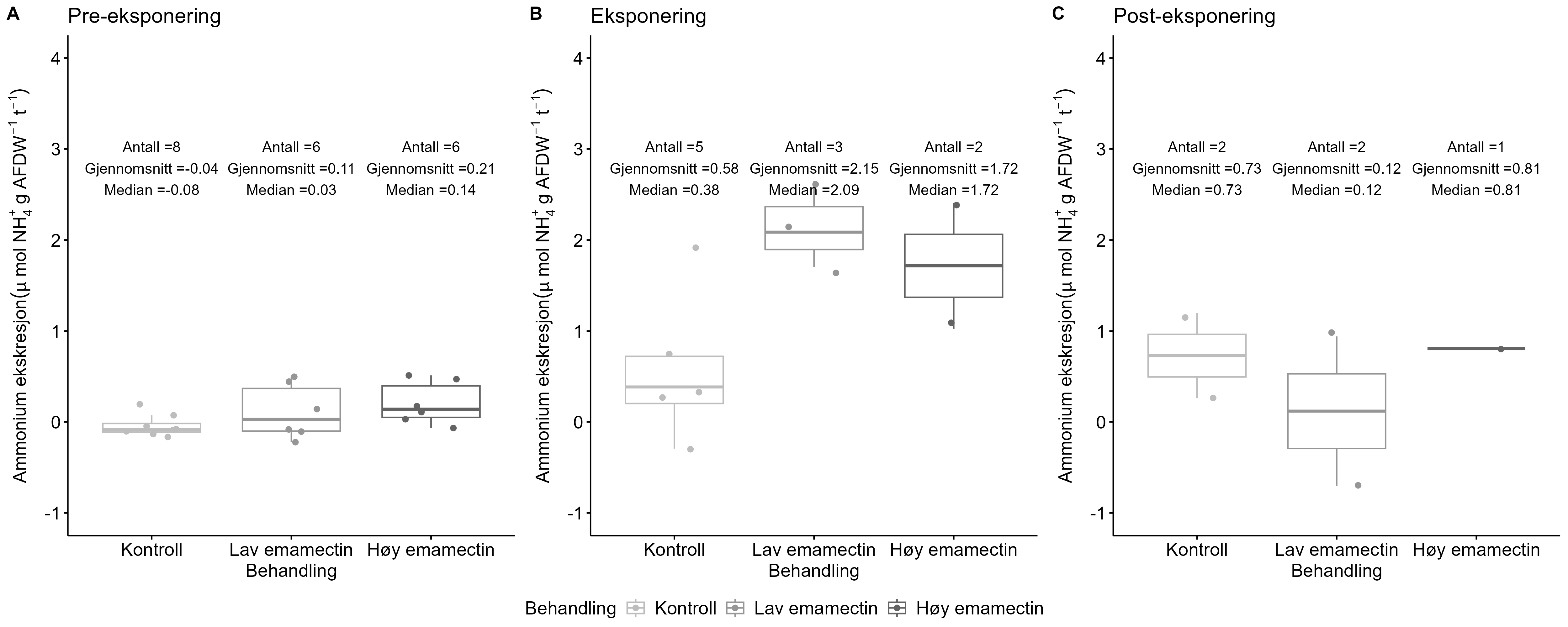
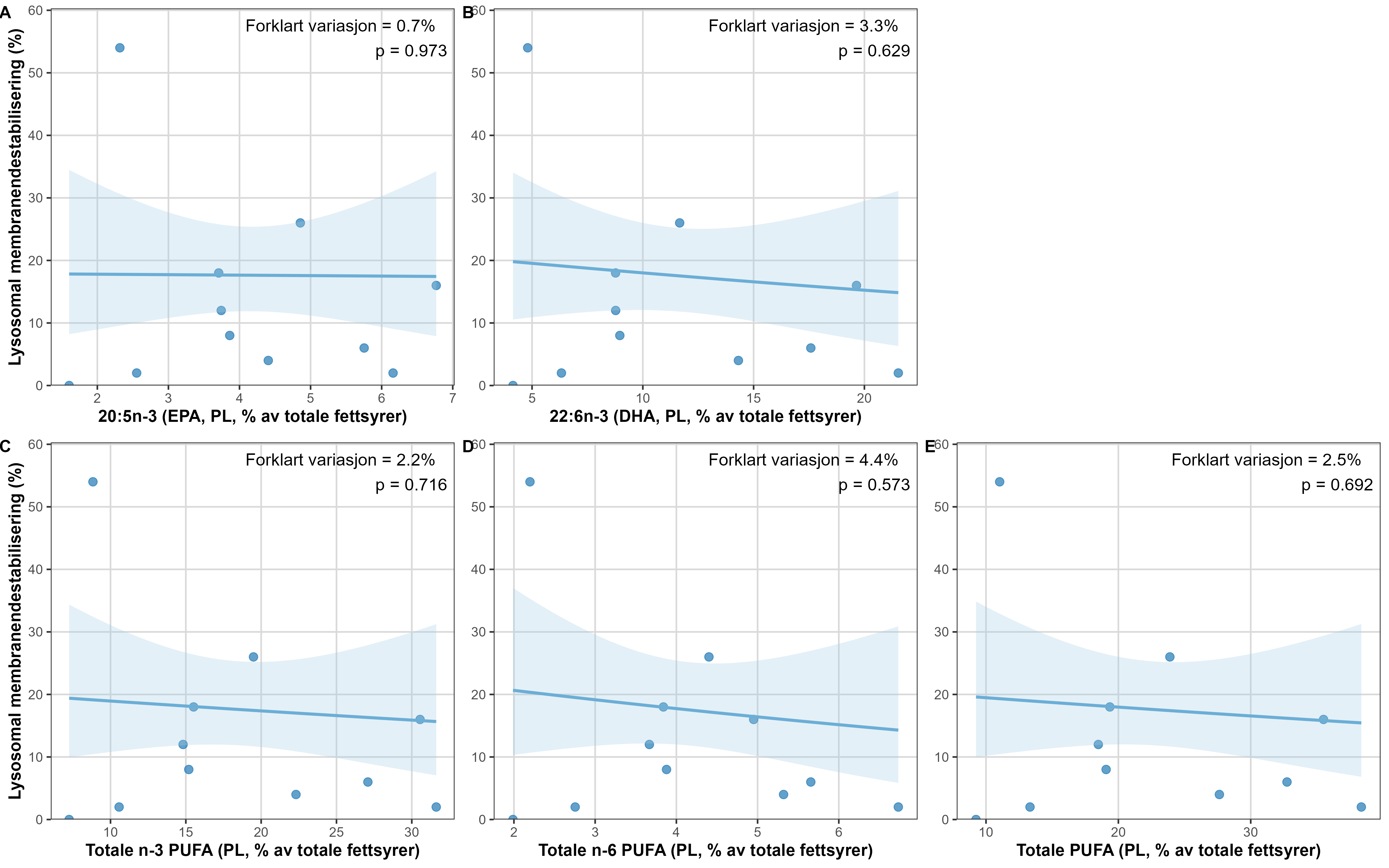
Measurements of oxygen consumption and ammonium excretion in corals showed limited direct metabolic effects of fish farm proximity, although P. arborea displayed a slight increase with increased sedimentation. Terrestrial fatty acids were detected at low levels across all species. A relative increase in total lipid content was observed with higher levels of predicted accumulated sedimentation for P. arborea and A. excavata, but overall results suggested limited uptake of farm-related material. Fatty acid profiles were largely stable and species-specific: P. arborea was unaffected by farm proximity, Phakellia spp. showed minor shifts related to organic sediment content, and A. excavata and G. barretti exhibited minimal responses.
Microbiome (16S rRNA) analyses showed low overlap between the microbial communities associated with the host species (P. arborea, Phakellia spp., G. barretti, and A. excavata) and those in surrounding sediments, indicating host-specific microbiomes. Predicted accumulated sedimentation and depth were the main environmental drivers of microbiome composition, while distance from farms had limited influence. Phakellia spp. displayed the strongest microbial response to sedimentation, whereas G. barretti showed few changes, highlighting species-specific host–microbiome interactions. Additionally, RNA-seq analyses revealed distinct transcriptional responses among species: A. excavata upregulated genes related to cellular protection and metabolic modulation; Phakellia spp. upregulated protein synthesis pathways; P. arborea showed gene regulation linked to symbiont and viral interactions; and G. barretti downregulated energy signaling and intracellular transport. Only intracellular transport genes were commonly affected across species, reflecting highly individualized stress-response strategies.
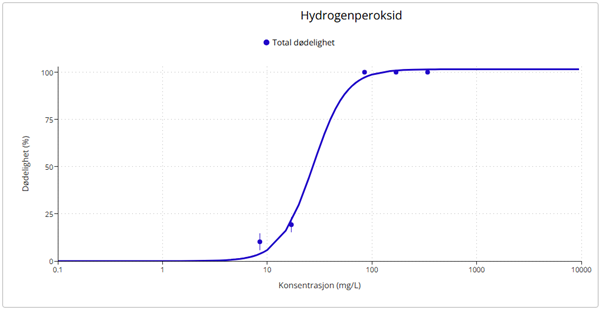
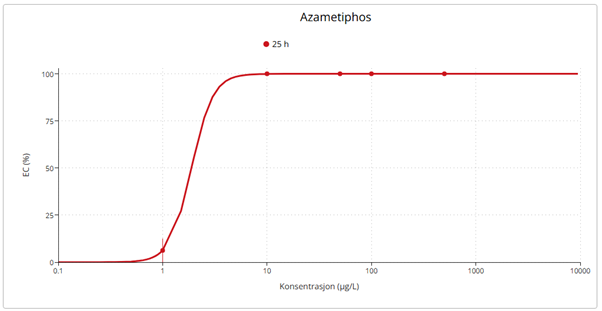
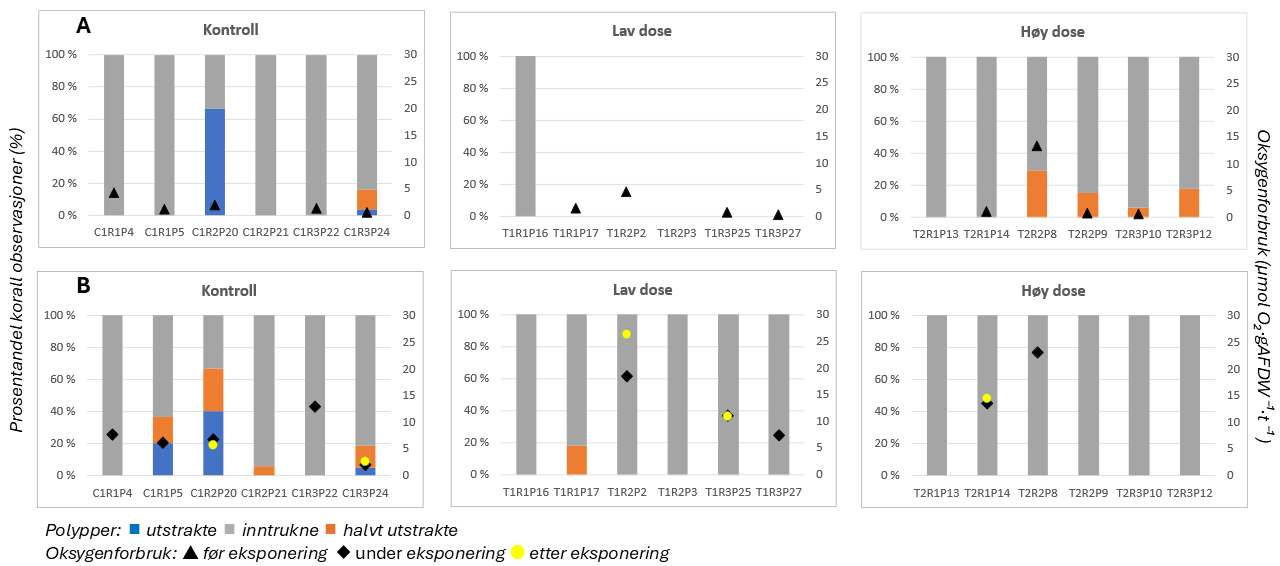
Results from field samples were complemented with laboratory exposure experiments to examine the effects of salmon lice chemotherapeutants on D. pertusum adults and larvae and particulate organic material on P. arborea, P. placomus, P. resedaeformis and D. pertusum. D. pertusum larvae were highly sensitive to hydrogen peroxide (LC50 = 27.20 mg·L⁻¹), similar to copepods but lower than lobster larvae, and especially to azametiphos (EC50 = 1.89 µg·L⁻¹). Adults exposed to emamectin benzoate (0.129 and 0.596 mg·L⁻¹) accumulated the compound in their tissues and showed metabolic and behavioral changes, including increased oxygen consumption, ammonium release and mucus production, and polyp mortality, particularly at high doses. P. resedaeformis was most sensitive to organic material, followed by P. arborea. The daily rythmn of polyp activity for both species was disturbed at the lowest exposure concentation.
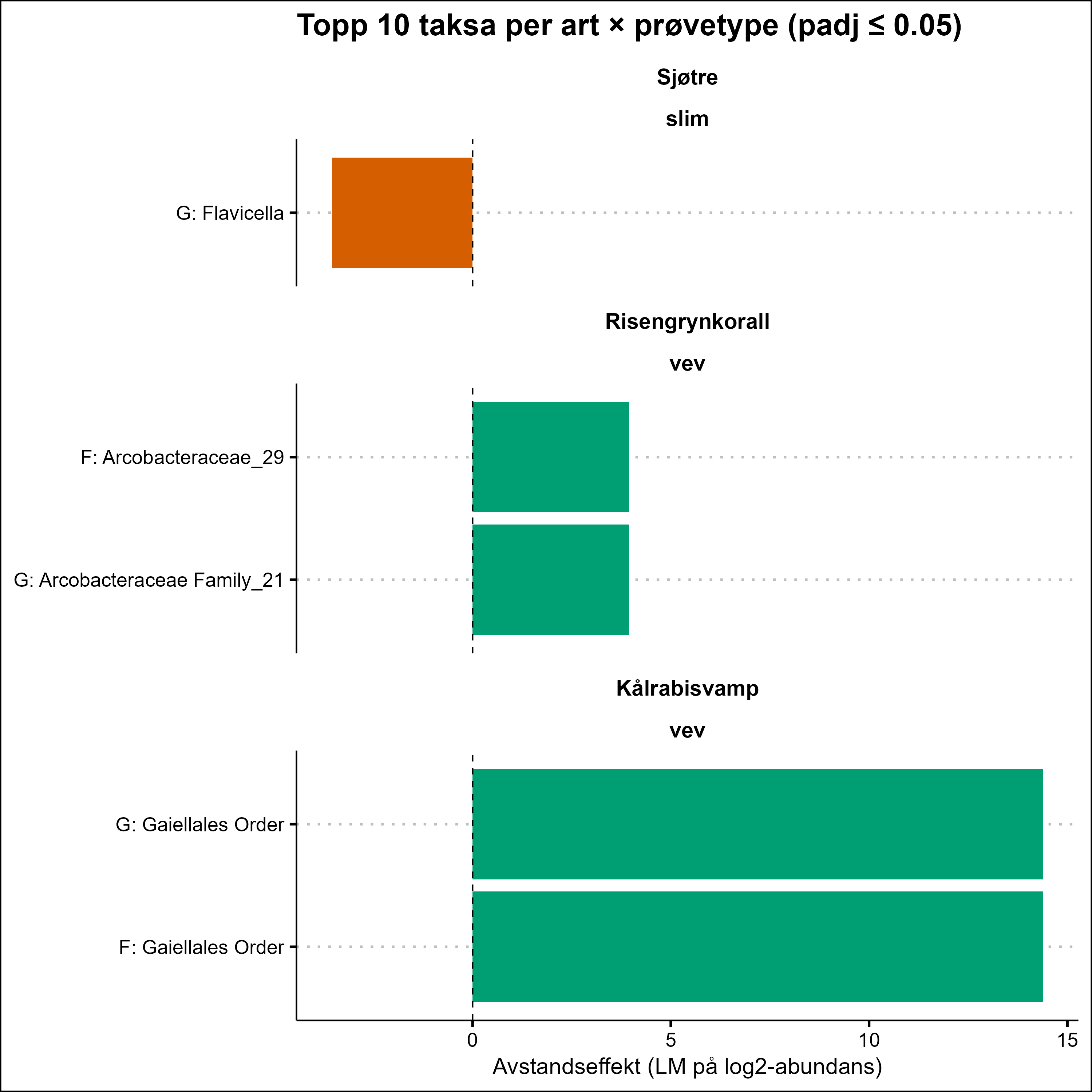
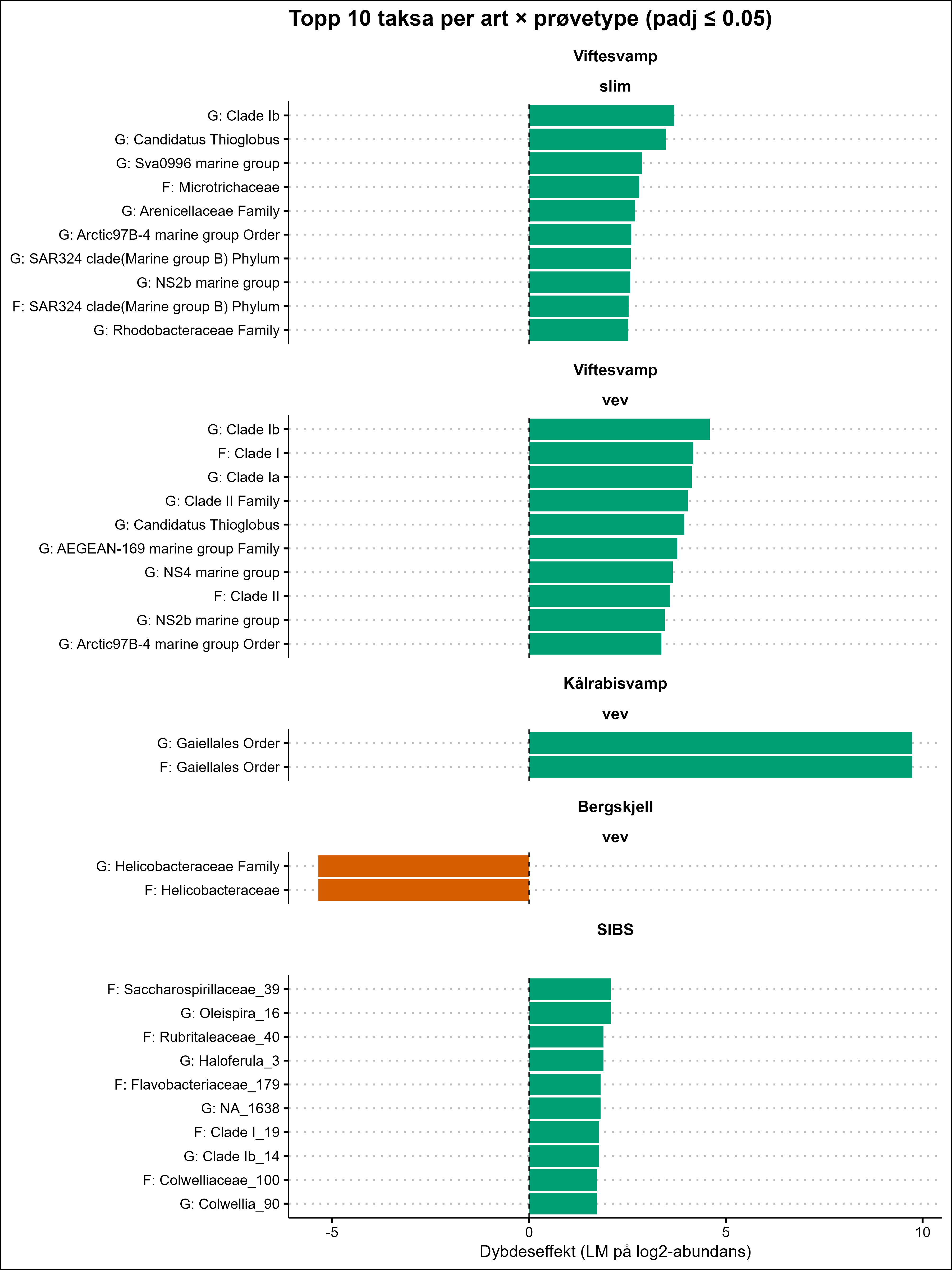
The VDWS Transition project provides an integrated assessment of sublethal effects of aquaculture waste on management-relevant benthic species in the field, combining visual, physiological, biochemical, microbial, and transcriptomic indicators. Based on a summary of results from field work and exposure experiments, Phakellia spp. emerges as the most sensitive species to aquaculture stress, with high confidence. P. resedaeformis is assessed to have moderate to high sensitivity, but with moderate confidence due to contradictory signals between methods (high visual sensitivity but absent metabolic response) and limited methodological coverage compared to other species. G. barretti and P. arborea appear to have high and moderate to high sensitivity respectively, both with high confidence. A. excavata was more robust, while the sensitivity of P. placomus and D. pertusum could not be assessed due to low sample size in the study. The combination of SIBS and b-MBI sampling, physiological biomarkers, and molecular (omics) analyses offers a scalable, minimally invasive approach for monitoring aquaculture impacts in complex habitats. Current knowledge of the sensitivity of the study species to aquaculture waste is summarized, and these field-based results complement laboratory studies by providing new insights into species-specific responses. A summary of the methods used to assess aquaculture impacts is also provided, highlighting their development, limitations, and potential.
Sammendrag
Effektene og toleransene til utvalgte forvaltningsrelevante arter overfor organisk utslipp fra oppdrettsanlegg og legemidler mot lakselus ble undersøkt ved bruk av nyutviklede feltmetoder for vurdering av organismers helsetilstand. Fokusartene inkluderte hornkoraller (sjøtre, sjøbusk, risengrynkorall), den revbyggende steinkorallen øyekorall, svampene kålrabisvamp og viftesvamp, og muslingen bergskjell.
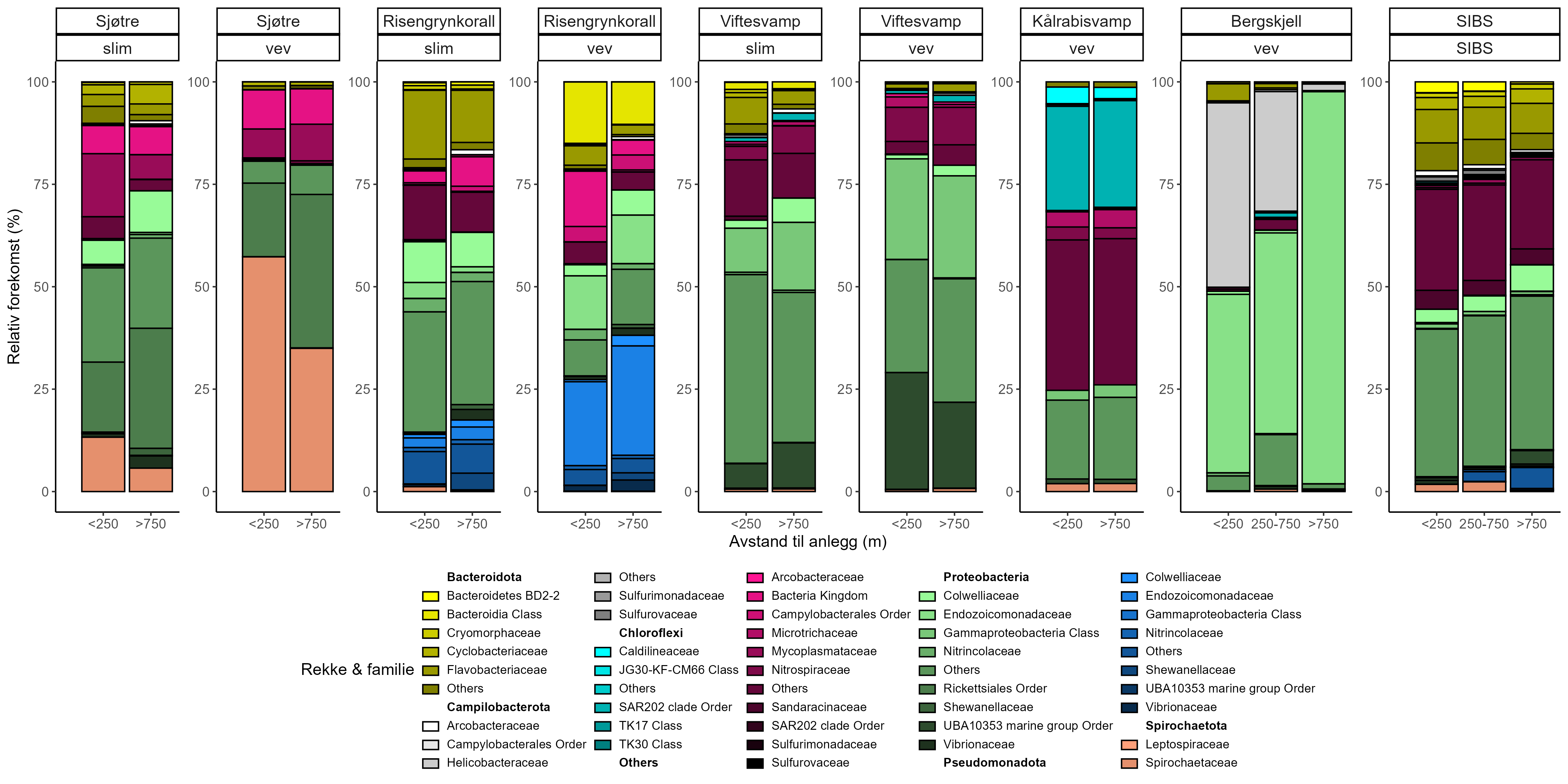
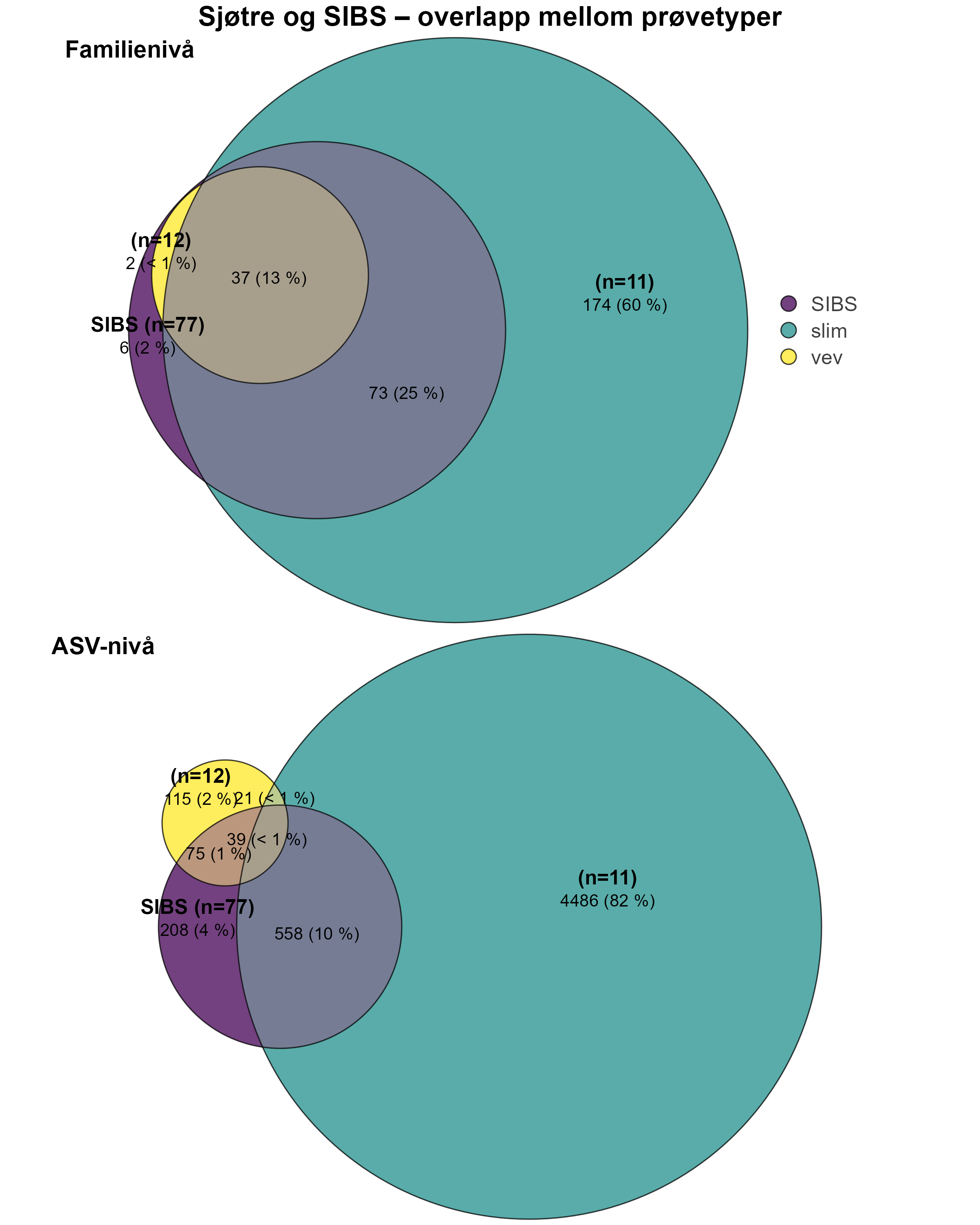
Prøver fra de utvalgte artene ble samlet inn med ROV fra områder innenfor influensområdet rundt sju oppdrettsanlegg og fra referanseområder mer enn 750 m unna langs kysten av Nordland og Møre og Romsdal. Modellert organisk sedimentasjon (akkumulert over to år før prøvetaking) ble sammenlignet med Bacterial Metabarcoding Biotic Index (b-MBI), en indikator på organisk anrikelse basert på tilstedeværelse av bakterielle indikatortaxa. b-MBI-verdier viste en positiv korrelasjon med modellert sedimentasjon og dybde, og reflekterte moderat til høy anrikelse nær anleggene, noe som fremhever metodens potensial som tidlig-varslingsindikator for oppdrettspåvirkning.
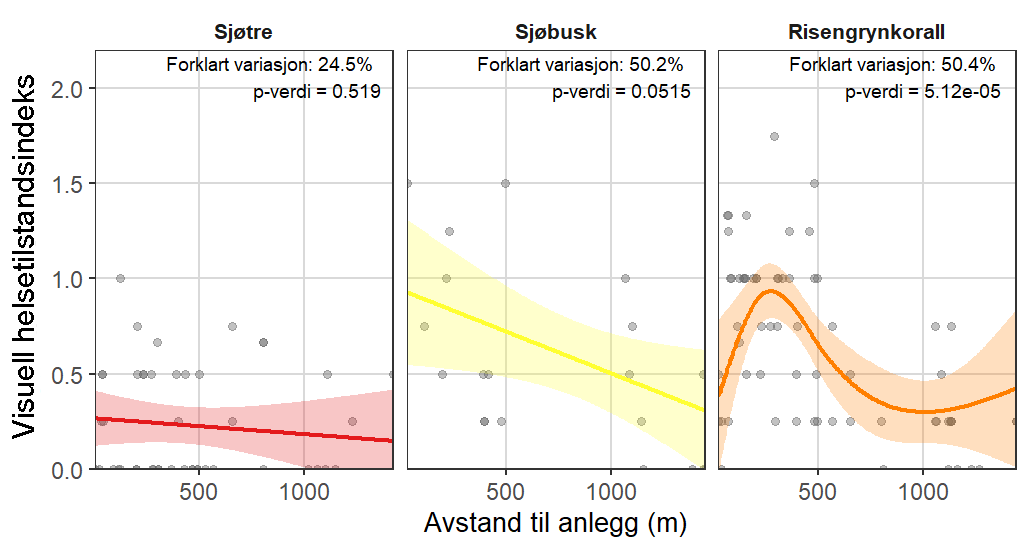
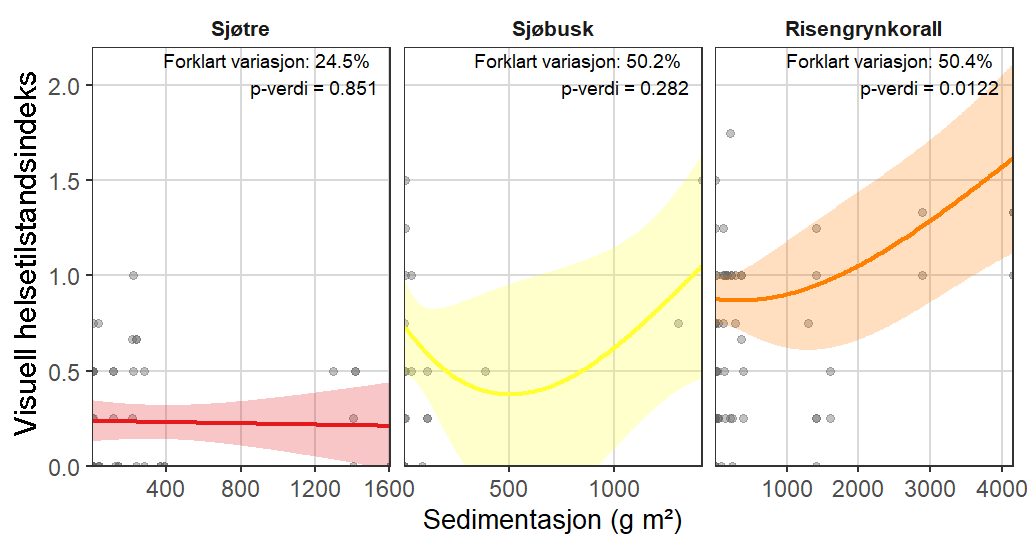
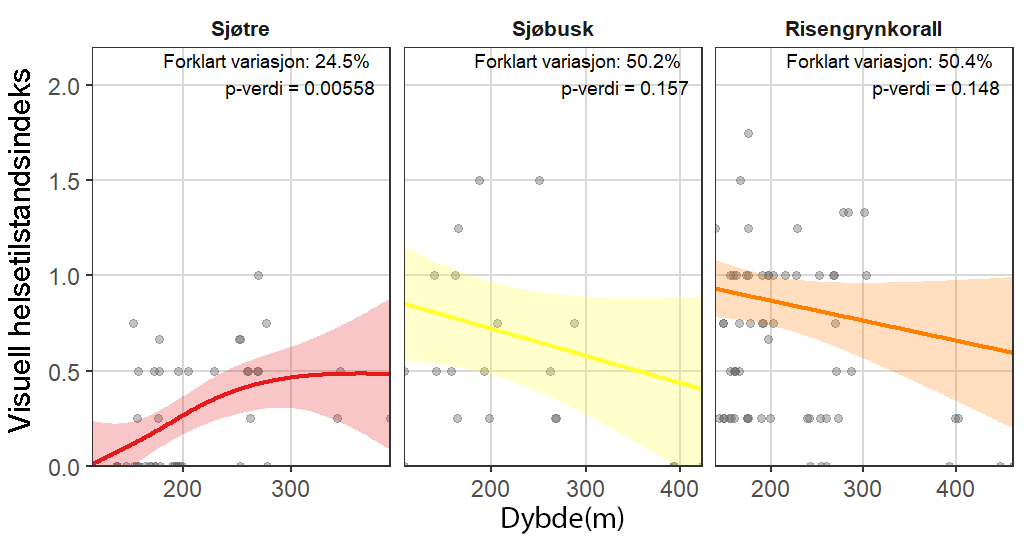
Visuell indeks på dårlig helsetilstand korrelerte med modellert sedimentasjon for risengrynkorall og viftesvamp, som viste redusert visuell tilstand ved høyere sedimentasjonsnivåer. Den visuelle tilstanden til kålrabisvamp var også korrelert med sedimentasjon, men viste at enkelte individer forble upåvirket. Sjøbusk viste redusert visuell tilstand nærmere oppdrettsanlegget. Helsekriterier for sjøtre var sterkere relatert til dybde enn til avstand fra oppdrettsanlegg. Analyse av lysosomal membranstabilitet (LMS) viste en høyere andel destabiliserte celler hos de to svampene og bergskjell nær oppdrettsanlegg, noe som indikerer følsomhet for oppdrettsutslipp. LMS-metoden viste seg å være en rask og kostnadseffektiv biomarkør for vurdering av fysiologisk stress i felt.
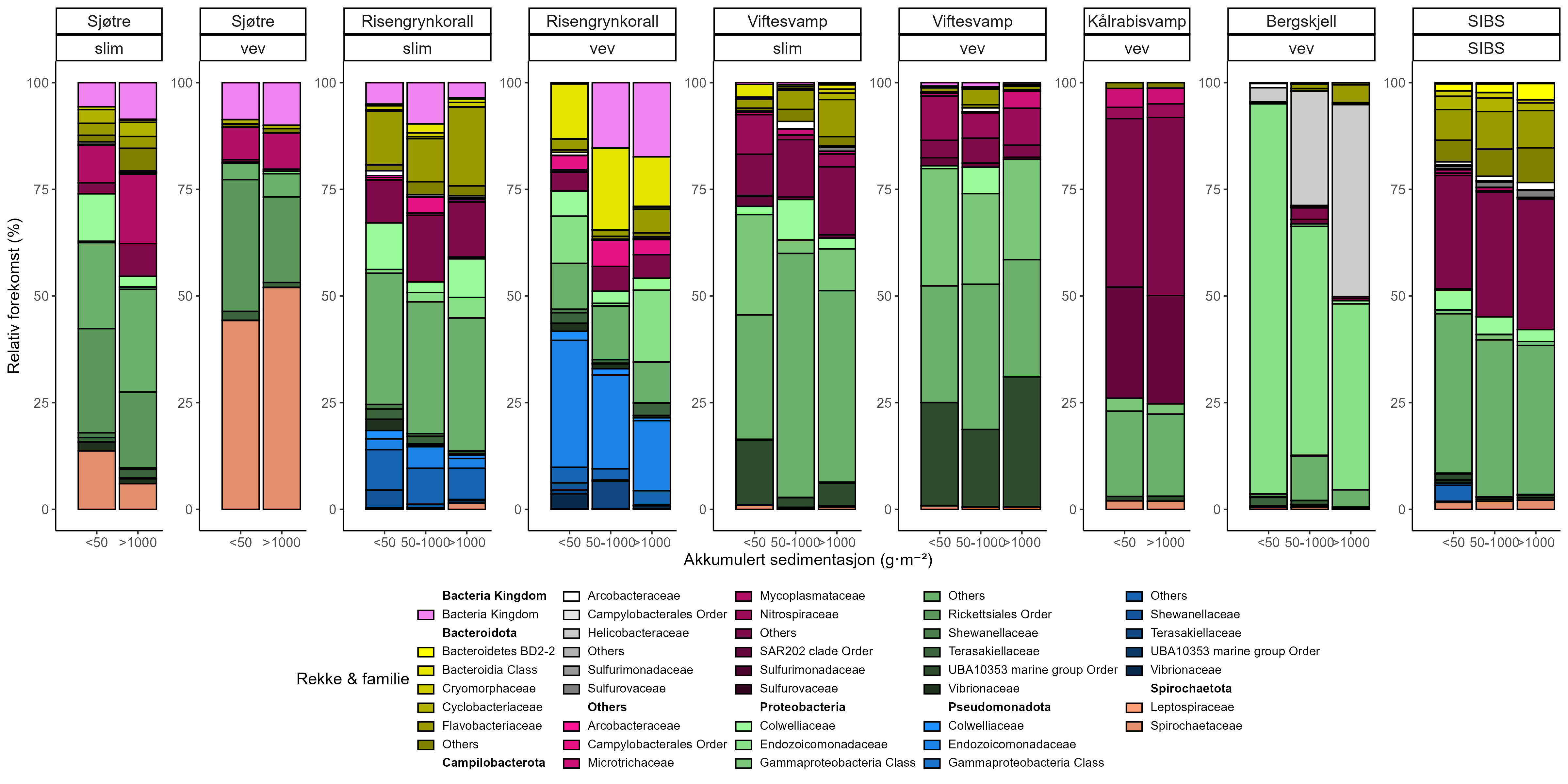
Målinger av oksygenforbruk og ammoniakkutskillelse hos koraller viste begrensede metabolske effekter av nærhet fra oppdrettsanlegg, med en svak økning hos sjøtre ved økt sedimentasjon. Terrestriske fettsyrer ble påvist i lave nivåer hos alle arter. En relativ økning i totalt lipidinnhold ble observert ved høyere nivåer av predikert akkumulert sedimentasjon for sjøtre og bergskjell, men totalt sett antydet resultatene begrenset opptak av oppdrettsrelatert materiale. Fettsyreprofilene var stort sett stabile og artsspesifikke: fettsyreprofilene til sjøtre ble ikke påvirket av nærhet til anlegg, viftesvamp viste derimot små endringer relatert til organisk sedimentinnhold, mens bergskjell og kålrabisvamp hadde minimale responser.
Mikrobiom-analyser (16S rRNA) viste lite overlapp mellom mikrobielle samfunn assosiert med vertene (sjøtre, viftesvamp, kålrabisvamp og bergskjell) og omgivende sedimenter, noe som indikerer vertsspesifikke mikrobiomer. Predikert akkumulert sedimentasjon og dybde var de viktigste miljødriverne for mikrobiomsammensetning, mens avstand fra oppdrettsanlegg hadde begrenset effekt. Viftesvamp viste sterkest mikrobiell respons på sedimentasjon, mens kålrabisvamp viste få endringer, noe som fremhever arts-spesifikke vert–mikrobiom-interaksjoner. RNA-sekvens-analyser viste også distinkte transkripsjonelle responser mellom arter: bergskjell oppregulerte gener knyttet til cellulær beskyttelse og metabolsk modulering; viftesvamp oppregulerte gener relatert til proteinsyntese; sjøtre viste genregulering knyttet til symbiont- og virusinteraksjoner; og kålrabisvamp nedregulerte energisignalisering og intracellulær transport. Kun gener knyttet til intracellulær transport var påvirket på tvers av arter, noe som reflekterer høyt individualiserte stress-responsstrategier.
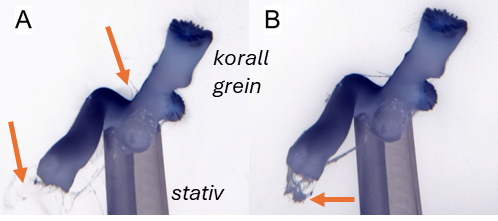
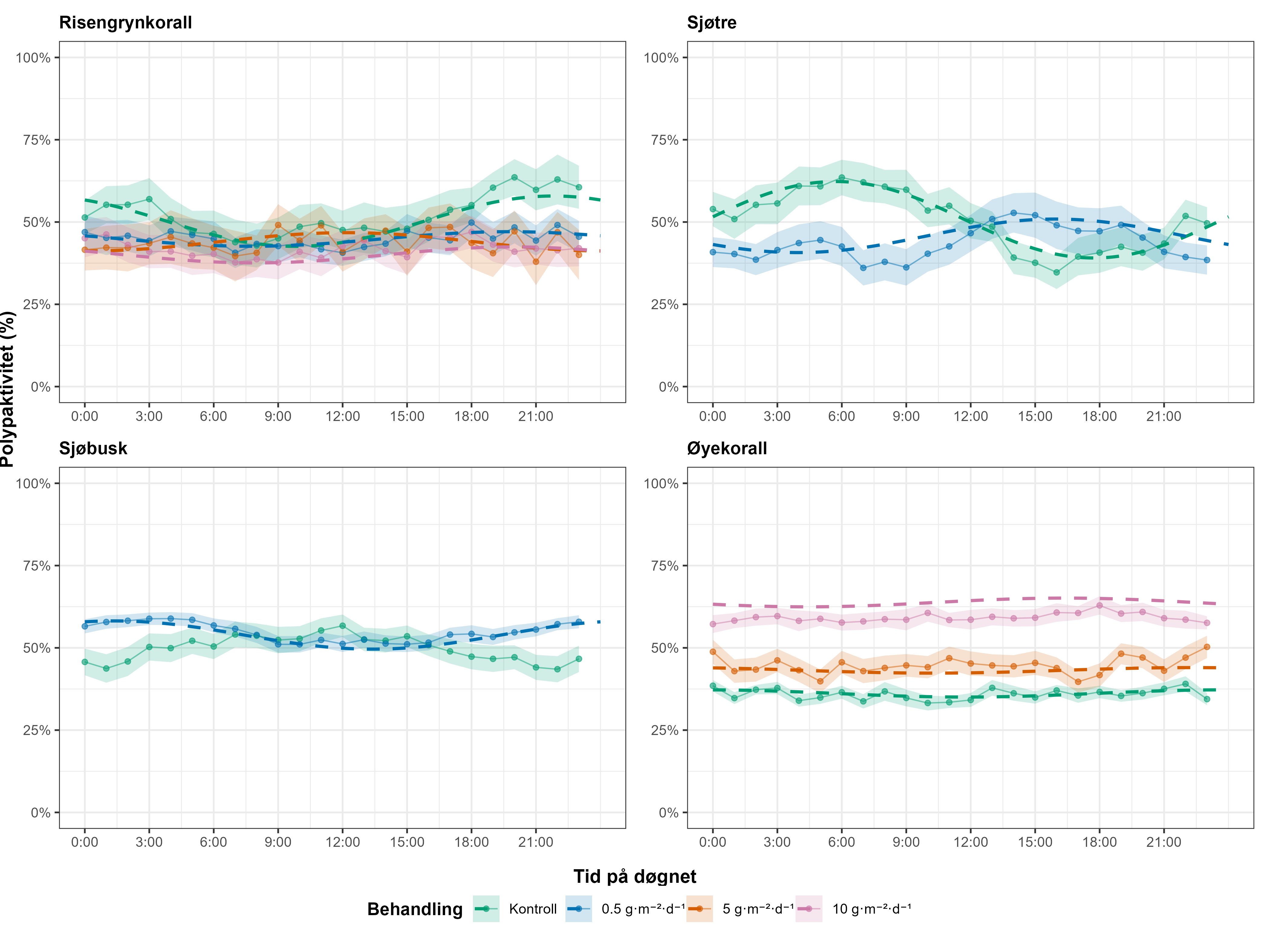
Resultater fra feltprøver ble supplert med laboratorieeksponeringer for å undersøke effektene av avlusningsmidler på voksne øyekorall og larver, samt effektene av partikulært organisk materiale på sjøtre, sjøbusk, risengrynkorall og øyekorall. Øyekorallarver var svært følsomme for hydrogenperoksid (LC50 = 27, 20 mg·L⁻¹) og spesielt følsomme for azametiphos (EC50 = 1,89 µg·L⁻¹). Voksne øyekoraller eksponert for emamektin benzoat (0,129 og 0,596 mg·L⁻¹) akkumulerte forbindelsen i vevet og viste metabolske og atferdsmessige endringer, inkludert økt oksygenforbruk, ammoniakkutskillelse, slimproduksjon og polyppdød, særlig ved høye doser. Risengrynkorall var den arten som i disse studiene var mest følsom for organisk materiale, etterfulgt av sjøtre. Døgnrytmen i polyppaktiviteten for begge arter ble også forstyrret allerede ved laveste eksponeringsnivå.
VDWS -prosjektet gir en integrert vurdering av subletale effekter av akvakulturutslipp på forvaltningsrelevante bunnarter i felt, ved å kombinere visuelle, fysiologiske, biokjemiske, mikrobielle og transkriptomiske indikatorer. Basert på en oppsummering av resultatene fra feltarbeid og eksponeringsforsøk, fremstår viftesvamp som den mest følsomme arten for stress fra organiske akvakulturutslipp, med høy grad av sikkerhet. Risengrynkorall vurderes å ha moderat til høy følsomhet, men med moderat sikkerhet på grunn av motstridende signaler mellom metodene (høy visuell sensitivitet, men fraværende metabolsk respons) og begrenset metodisk dekning sammenlignet med andre arter. Kålrabisvamp og sjøtre ser ut til å ha henholdsvis høy og moderat til høy følsomhet, begge med en høy grad av sikkerhet. Bergskjell var mer robust, mens følsomheten til sjøbusk og øyekorall kunne ikke vurderes på grunn av lavt utvalg i studien. Kombinasjonen av SIBS og b-MBI-prøvetaking, fysiologiske biomarkører og molekylære (omics)-analyser tilbyr en skalerbar og minimalt invasiv metode for å påvise effekter fra oppdrett i komplekse habitater. Nåværende kunnskap om følsomheten til bunndyr inkludert i denne studien for oppdrettsutslipp oppsummeres, og disse feltbaserte resultatene utfyller laboratoriestudier ved å gi nye innsikt i arts-spesifikke responser. En oppsummering av metodene brukt for å vurdere påvirkning av akvakulturutslipp på helsetilstand er også gitt, med vekt på videre utvikling, begrensninger og potensial.
Oppdatering:
09.04.2026: Kapittel 1.1 og 1.8 er oppdatert. I tillegg er figur 11 og 14 byttet ut som følg av skrivefeil, og det er lagt til en ekstra setning i kapittel 3.11.
1 - Anbefalinger
Utslipp fra fiskeoppdrettsanlegg i sjø kommer i form av løste næringssalter, organiske partikler, fremmedstoff og legemidler. Vi gir her en oppsummering av sju utvalgte forvaltningsrelevante bunnarters (viftesvamp, risengrynkorall, sjøbusk, bergskjell, kålrabisvamp, sjøtre og øyekorall) følsomhet for utslipp fra akvakultur, hvilke metoder som er benyttet for å dokumentere endringer i deres helsetilstand, samt anbefalinger for videre metodeutvikling. Analysen av resultater fra feltprøvene fokuserte primært på effekten av en anrikningsgradient fra organiske partikler, men bunndyr samlet inn i felt kunne også bli påvirket av fremmedstoffer og legemidler. Laboratorieforsøk ble brukt til å studere effekten av både avlusningsmidler og organisk partikulært materiale. Anbefalingene er basert på resultater fra prosjektet VDWS (Fra føre-var-prinsippet til en kunnskapsbasert forvaltning av sårbar natur nær matfiskanlegg), et eksponerings forsøk finansiert av Havforkningsinstituttets program for akvakultur, marint miljø og teknologi og eksisterende vitenskapelig kunnskap. Basert på ny kunnskap fra disse prosjektene foreslås det tiltak som kan iverksettes for å redusere påvirkningen av akvakulturutslipp på disse artene.
1.1 - Viftesvamp
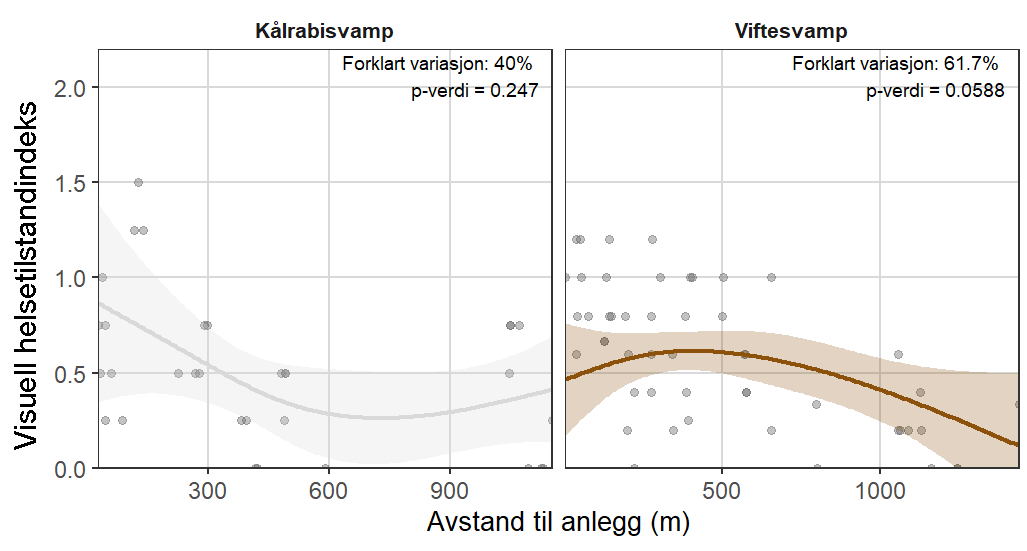
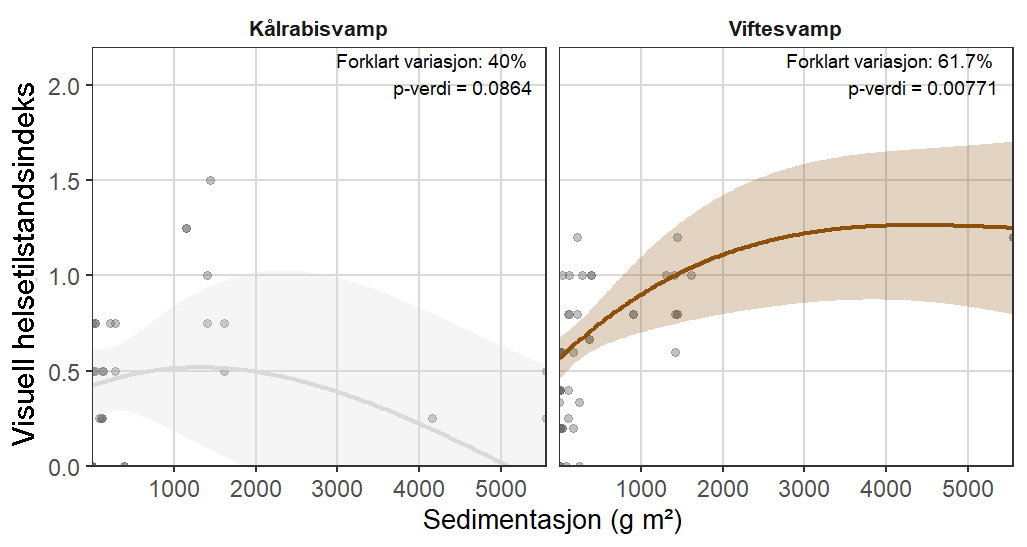
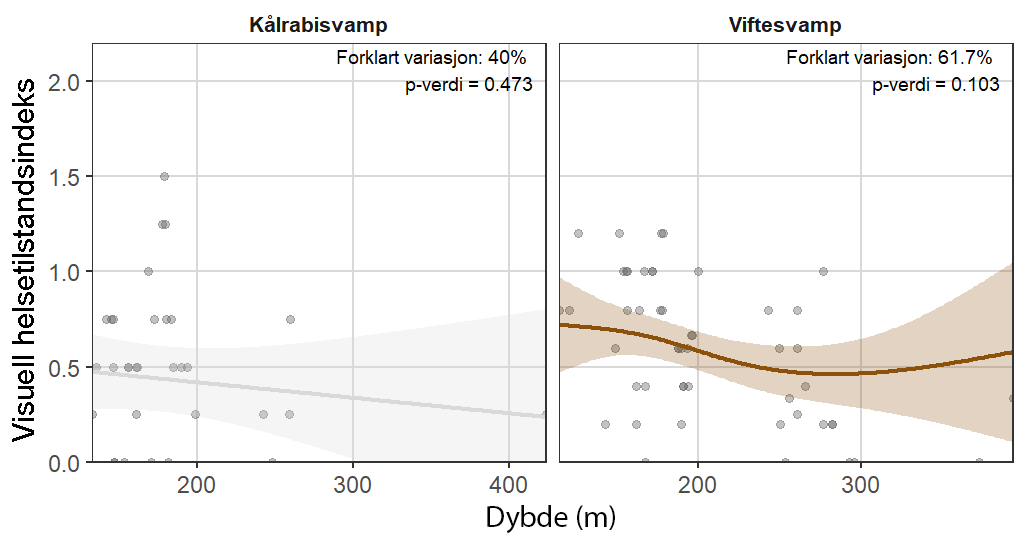
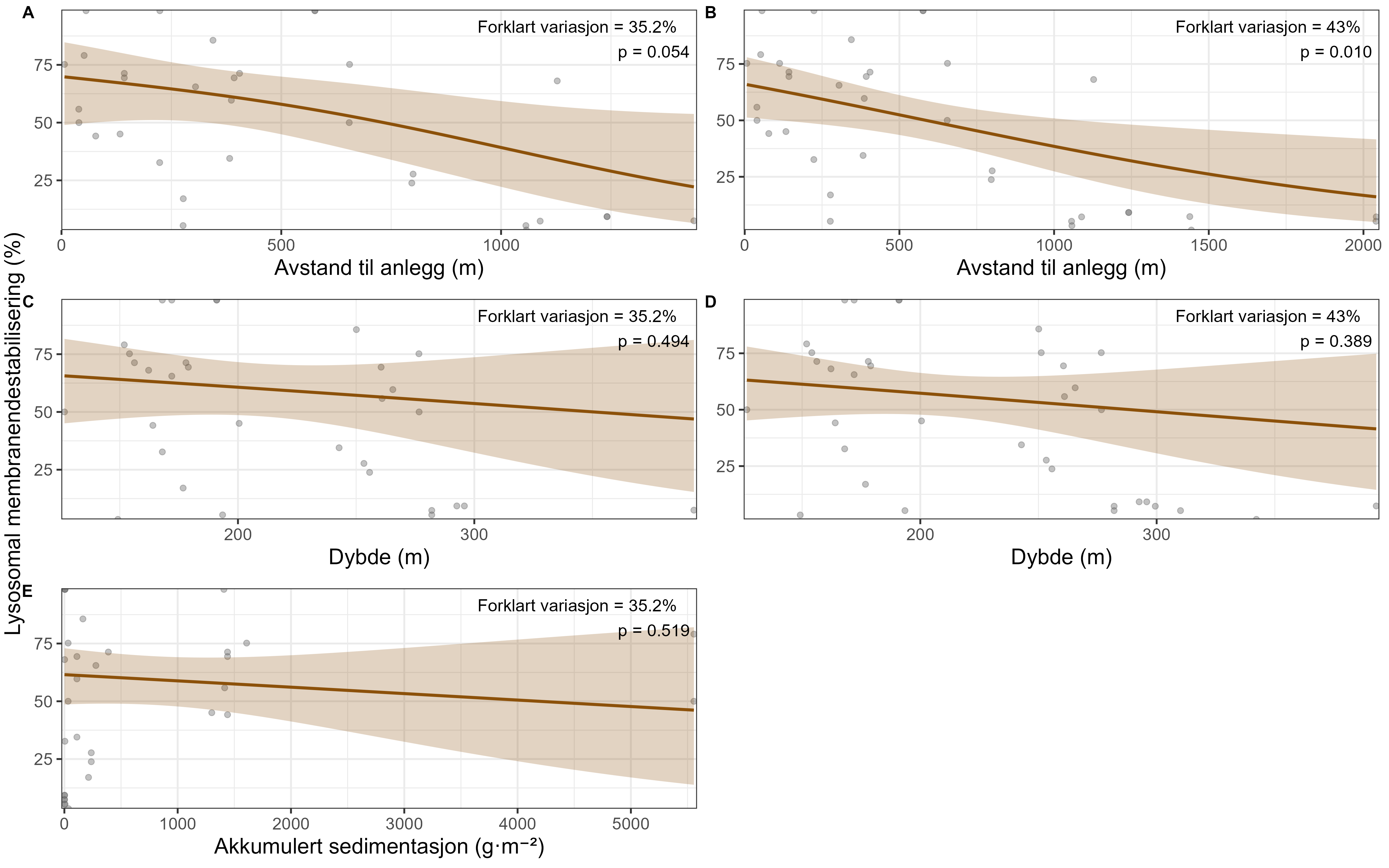
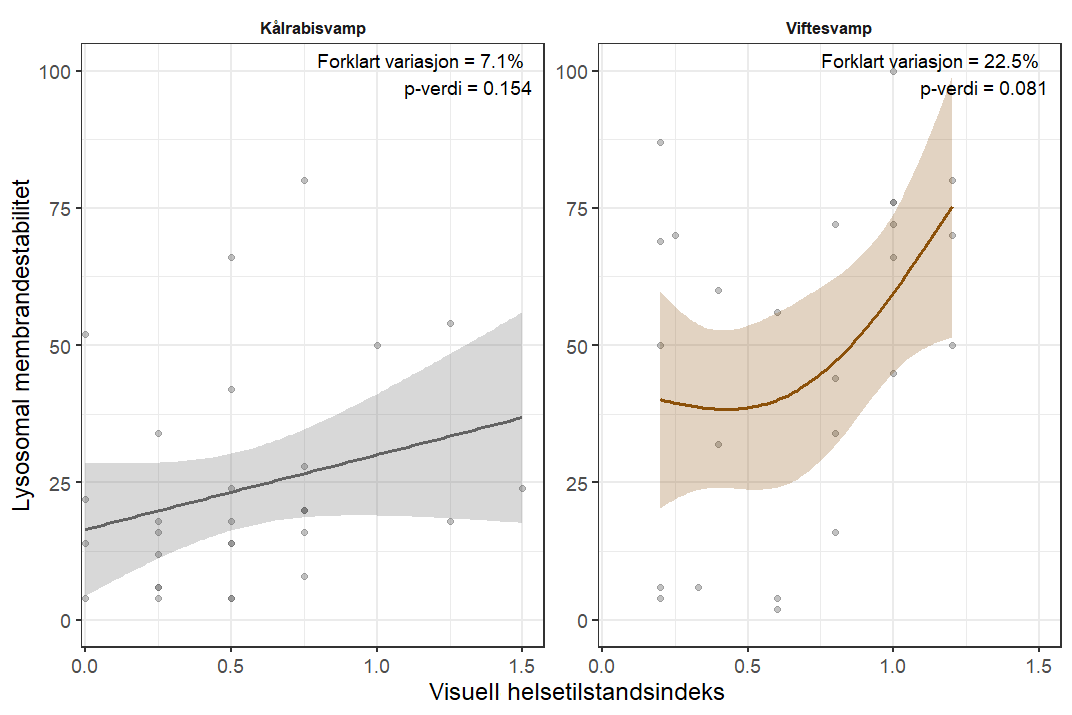
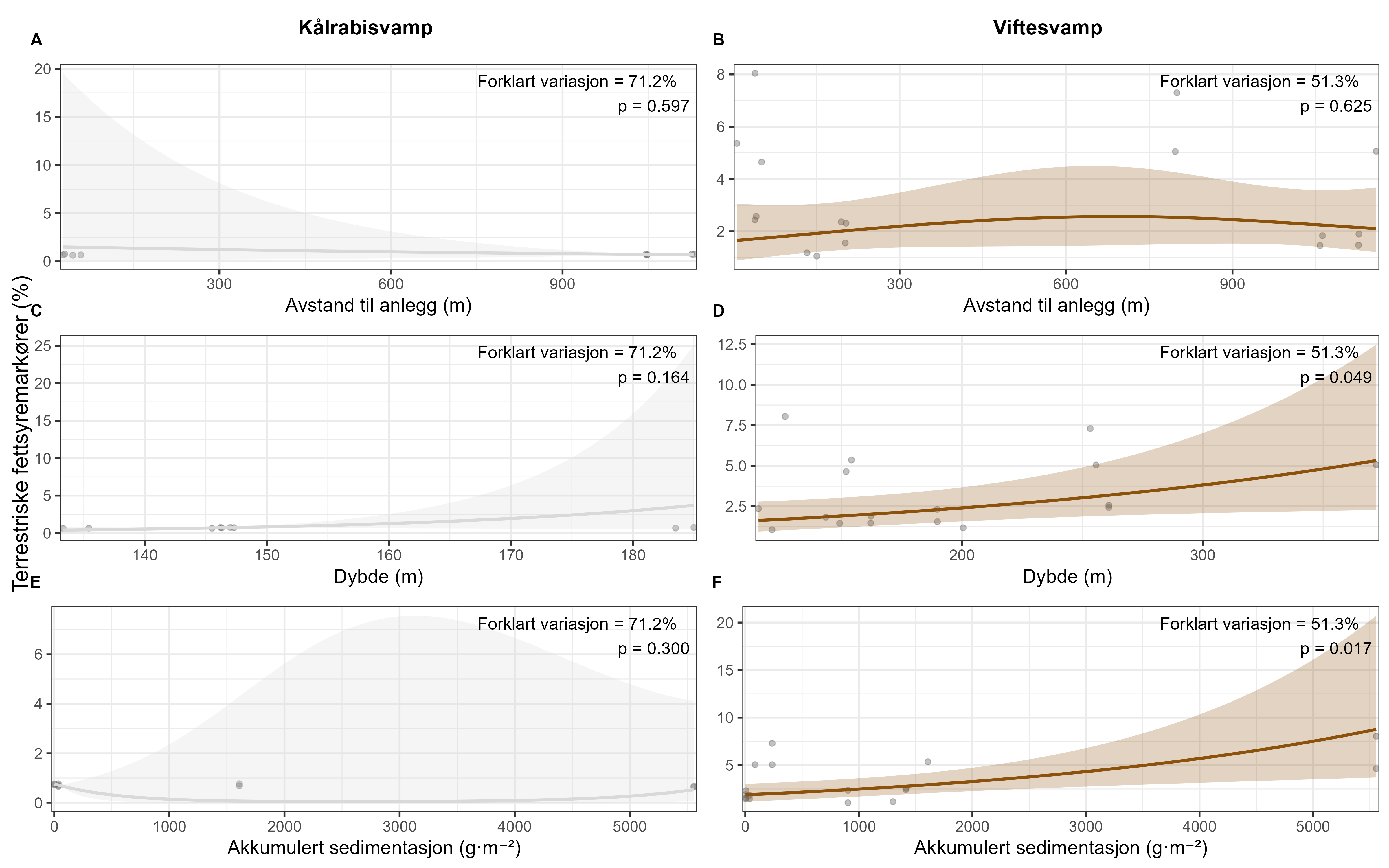
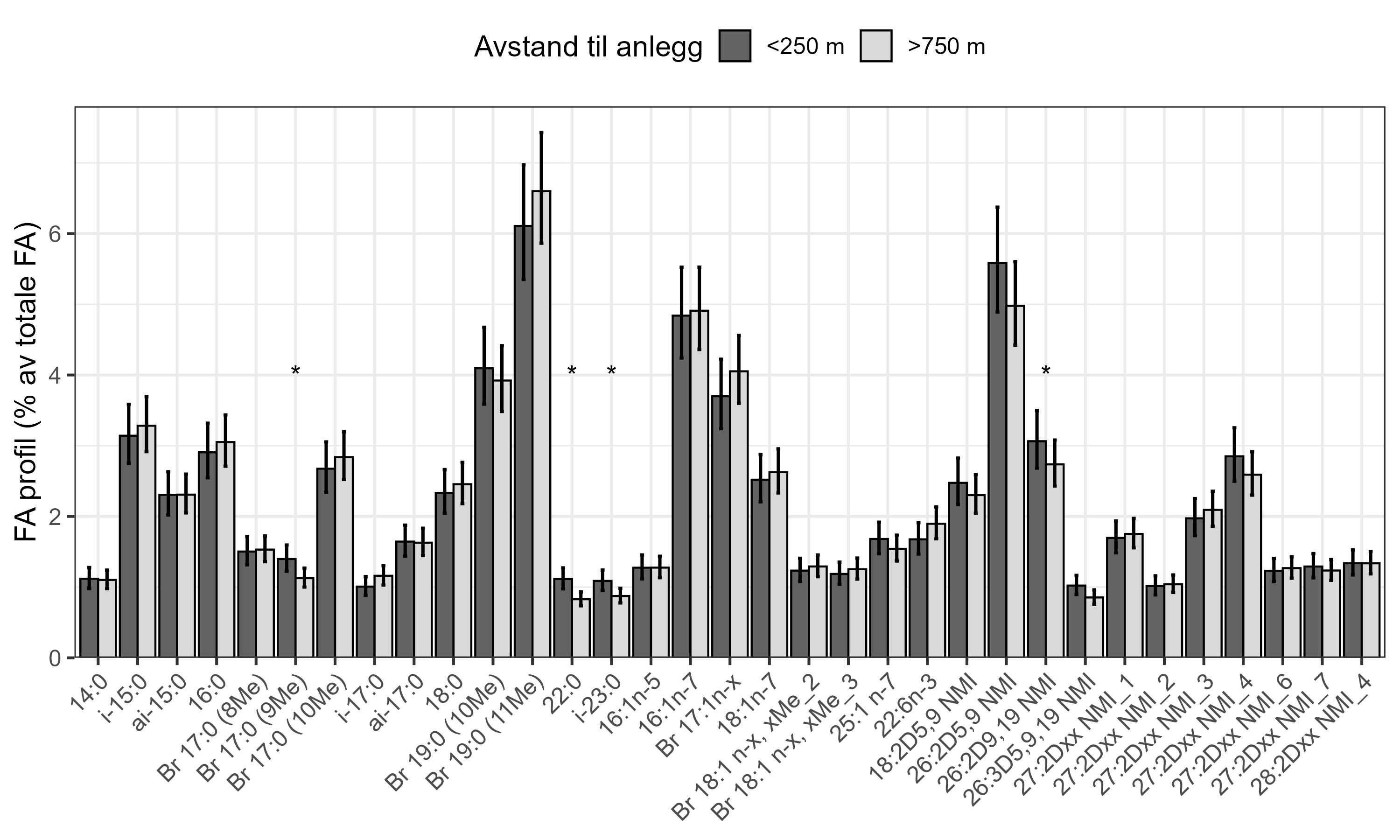
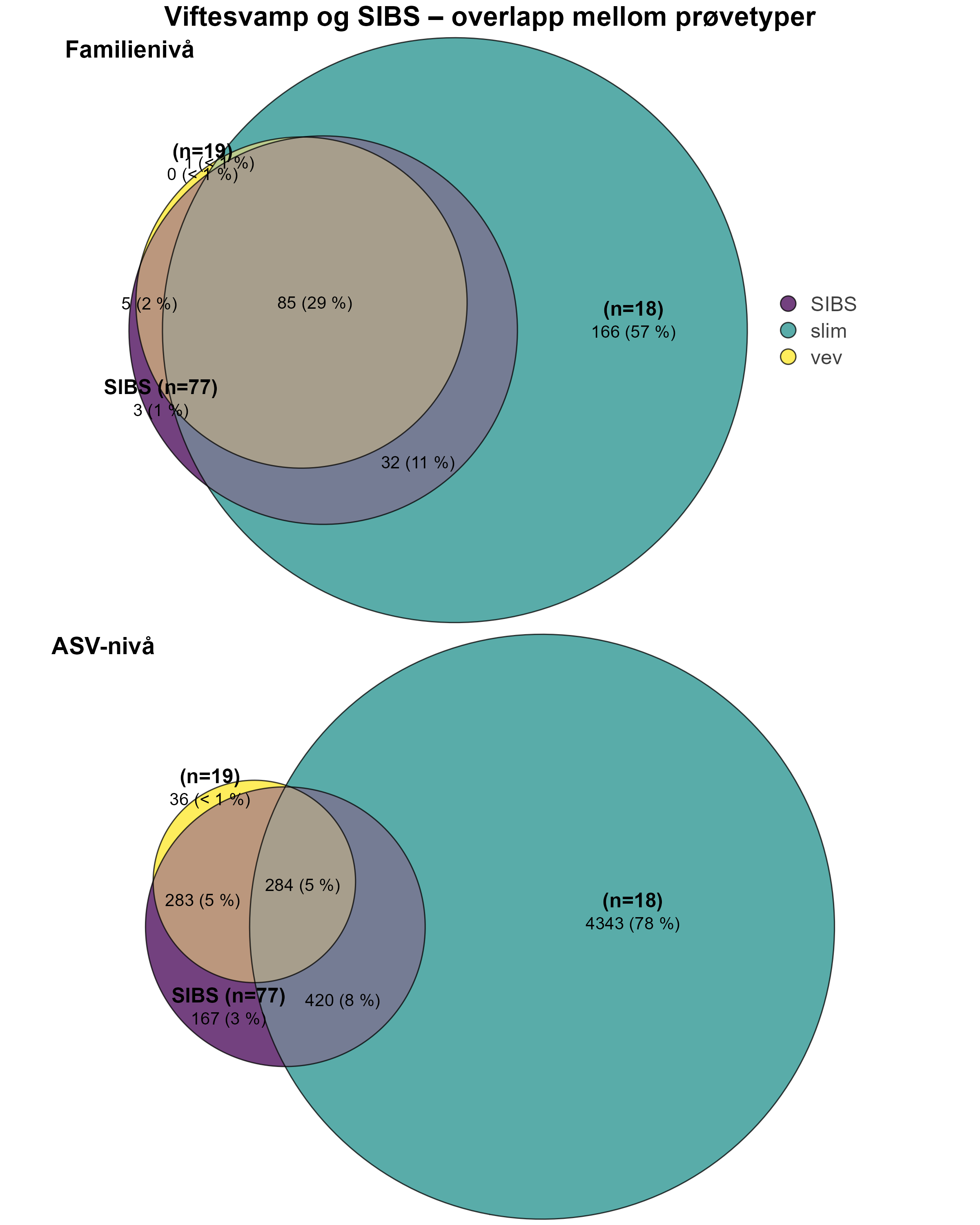
Følsomheten til viftesvamp for utslipp fra akvakultur ble vurdert på prøver samlet inn langs en utslippsgradient til sju oppdrettsanlegg. Dette ble gjort ved bruk av visuell helsevurdering, lysosomal membranstabilitet, lipid- og fettsyreprofiler og 16S- og RNA-sekvensering. Resultatene indikerte at viftesvamp er sensitiv for påvirkning fra organiske akvakulturutslipp. Det ble observert synlig vevsdegradering, redusert lysosomal membranstabilitet, et svak indikasjon på reduserte lipidreserver og endret fettsyreprofil som er respons på høy sedimentasjon av organisk utslipp. Det ble også observert moderate til sterke transkriptomiske responser på sedimentasjon. Disse subletale effektener forekom 10–600 meter fra oppdrettsanleggene og fraværet av viftesvamper innenfor 250 m fra enkelte lokaliteter tyder på at effektene noen ganger også kan være letale (dødelige) nært anleggene, selv om fravær også kan gjenspeile historiske forstyrrelser, substratforhold eller begrensninger i kartleggingsdekning. Ettersom det var et stort samsvar mellom de forskjellige metodene som ble brukt for å måle stress, kan man med stor grad av sikkerhet si at viftesvamp er svært følsom for organisk utslipp fra akvakulturanlegg.
Metoder for å dokumentere endringer i helsetilstand på grunn av akvakulturutslipp
En ikke-inngripende visuell helsetilstandsindeks ble brukt og vurdert som godt egnet til å dokumentere helserelaterte endringer hos viftesvamp innenfor influensområdet for spredning av organisk akvakulturutslipp. Metoden virket særlig robust når den ble brukt sammen med Bacterial Metabarcoding Biotic Index (b-MBI), en indikator på organisk anrikning basert på tilstedeværelse av bakterielle indikatortaksa tatt fra ved siden av svampprøvene. Det er derfor ikke nødvendig å inkludere fysisk prøvetaking og bruke inngripende metoder som lysosomal membranstabilitet, hvis man ikke trenger å ytterligere bekrefte tilstandsvurderingen. Mer presis kunnskap om effekter av utslipp på viftesvamp og biologiske og fysiologiske påvirkningsmekanismer kan oppnås ved bruk av analyser av fettsyreprofiler, 16S-samfunnsstruktur og RNA-sekvensering.
1.2 - Risengrynkorall
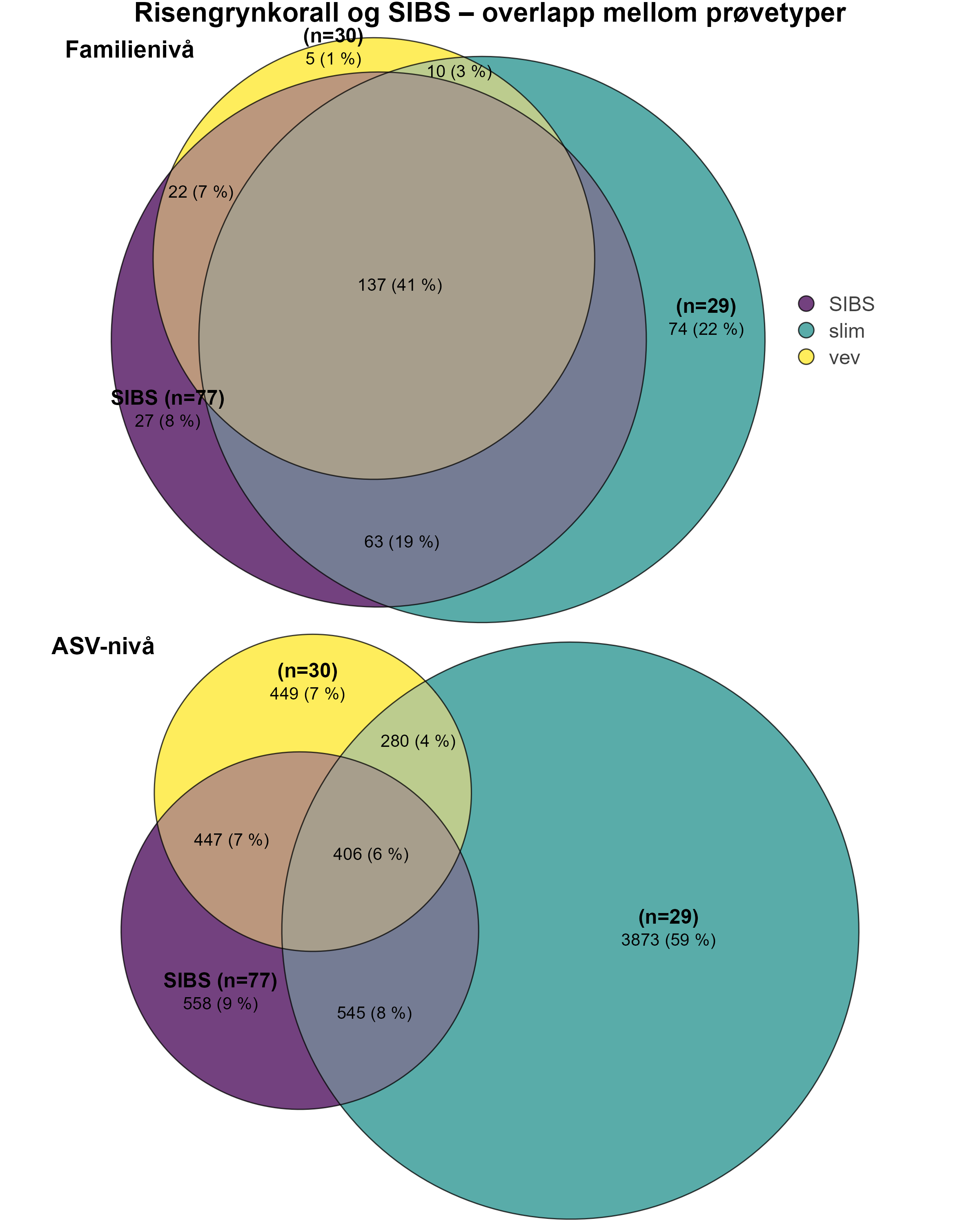
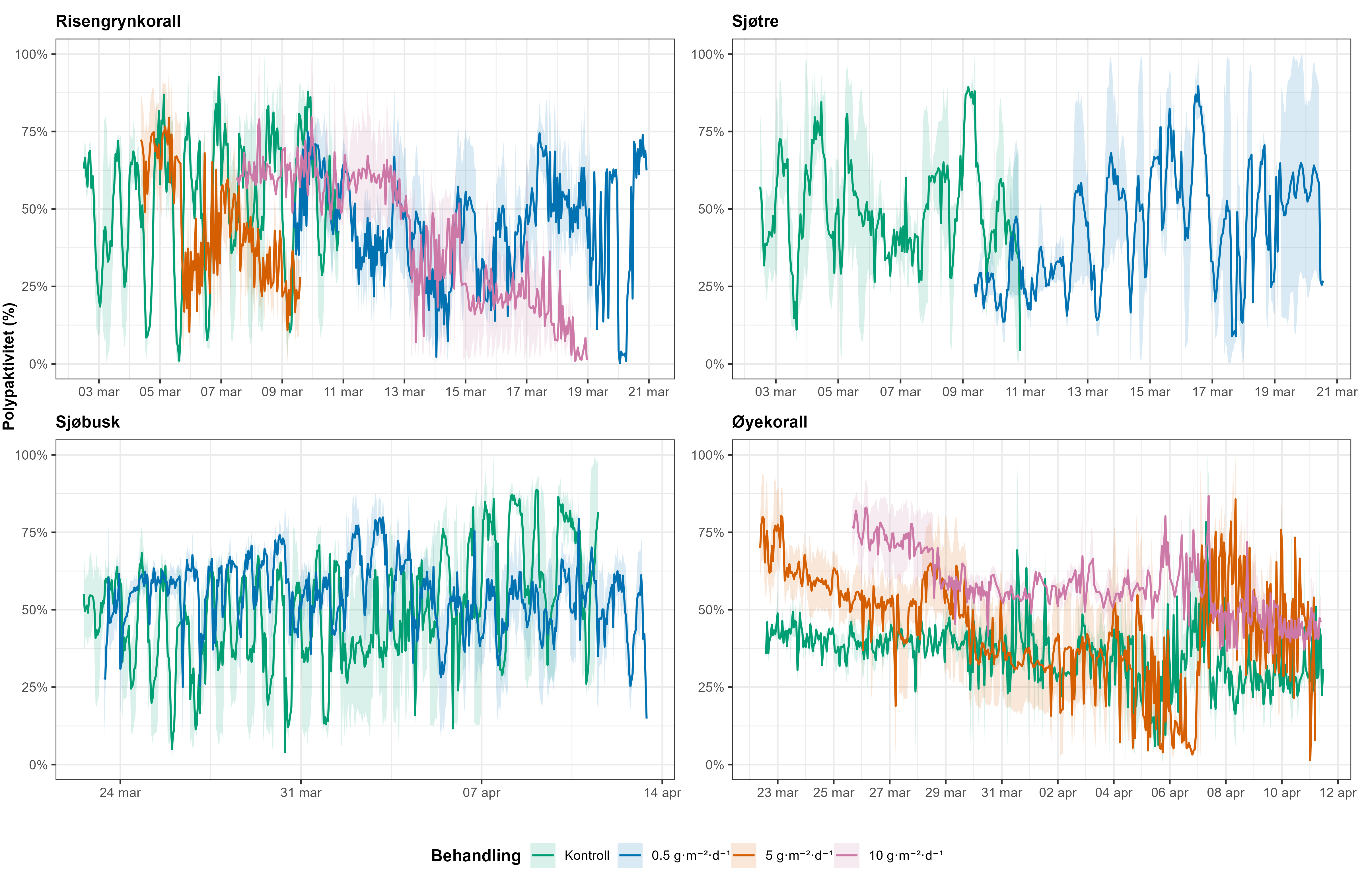
Følsomheten til risengrynkorall for utslipp fra akvakulturanlegg ble vurdert på prøver samlet inn langs organisk utslippsgradienter til sju oppdrettsanlegg. Dette ble gjort ved bruk av visuell helsevurdering, O₂-forbruk, ammoniumsekresjon, 16S-sekvensering og reproduksjonsevne. Synlig vevsdegradering og reduksjon av den visuelle helsetilstanden forekom ved høye sedimentasjonsnivåer av organisk material (> 2000 g·m⁻²) og innenfor 10–500 m fra oppdrettsanleggene. Det ble det også observert endringer i de mikrobielle samfunnene i vev og slim hos korallene. Oksygenforbruk, ammoniumsekresjon eller fekunditet ble ikke påvirket av utslipp. Redusert polyppaktivitet, vevsnedbrytning og økt dødelighet ble observert hos individer eksponert for 5–10 g·m⁻² organisk materiale per dag i laboratorieforsøk. Døgnrytmen i polyppaktivitet ble forstyrret hos risengrynkorall eksponert for konsentrasjoner over 0,5 g·m⁻²·dag⁻¹. Dette tilsvarer sedimentasjonsnivåer som man kan finne 300 m fra et oppdrettsanlegg. Basert på resultater fra felt og forsøk, vurderes risengrynkorall til å ha moderat til høy følsomhet for organisk utslipp fra oppdrettsanlegg, med moderat grad av sikkerhet. På denne arten ble det imidlertid benyttet et begrenset sett med molekylære metoder. Lysosomal membranstabilitet, lipid- og fettsyreprofiler, RNA-sekvensering eller DNA-metylering ble ikke analysert. Det kan derfor være at enkelte fysiologiske endringer ikke har blitt oppdaget.
Metoder for å dokumentere endringer i helsetilstand på grunn av akvakulturutslipp
En ikke-inngripende visuell helsetilstandsindeks ble brukt og vurdert godt egnet til å dokumentere helserelaterte endringer hos risengrynkorall innenfor 10–500 meter fra oppdrettsanlegg. De få molekylære metodene som ble testet på arten, viste begrenset sammenheng mellom fysiologiske endringer og organisk utslippsgradienten. 16S-sekvensering kan påvise endringer i mikrobielle samfunn knyttet til utslippsgradienten, men er en kostnadskrevende metode. Den sammensatte påvirkningsindeksen som ble brukt, en kombinasjon av visuell helsetilstandsindeks og b-MBI, beskrev bare delvis helseeffekten av utslippene. Forholdet var enda svakere når man inkluderte oksygenforbruk og ammoniakkutskillelse, noe som tyder på at vi trenger data fra andre metoder, som for eksempel analyser av lysosomal membranstabilitet.
1.3 - Kålrabisvamp
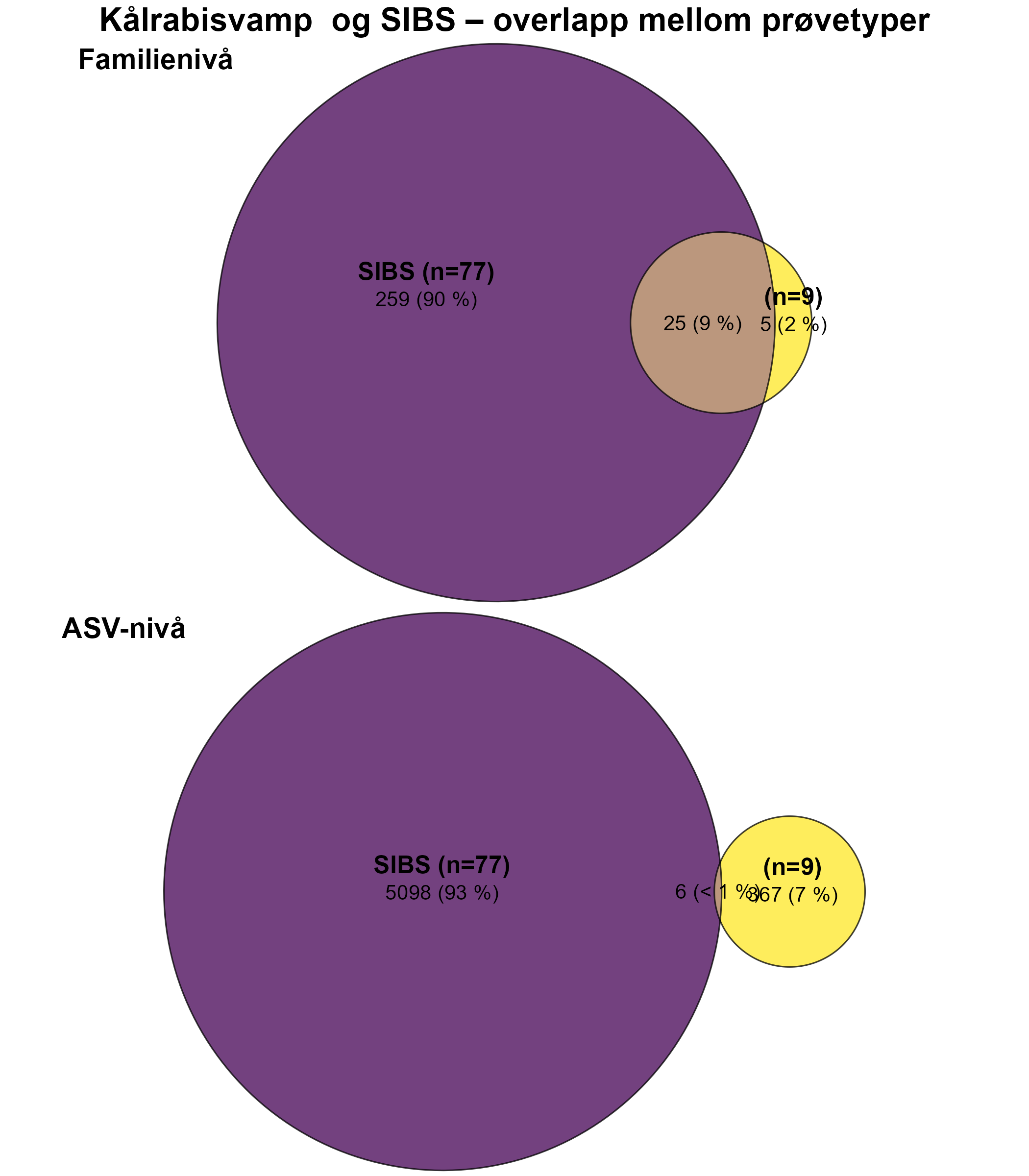
Følsomheten til kålrabisvamp for organisk utslipp fra akvakulturanlegg ble vurdert på prøver samlet inn langs utslippsgradienter til sju oppdrettsanlegg. Dette ble gjort ved bruk av visuell helsevurdering, lysosomal membranstabilitet, lipid- og fettsyreprofiler, 16S - og RNA-sekvensering. Resultatene viste en betydelig cellulær stressrespons ved økt organisk anrikning, og noe effekter på den visuelle helsetilstandsindeksen og på lipidlagre ble observert innenfor 300 m fra anleggene og ved sedimentasjon over 1000 g·m⁻². Til tross at noen individer ser ut til å være mer tolerante for påvirkning, er kålrabisvamp vurdert å ha en høy følsomhet for organisk utslipp fra oppdrettsanlegg, med høy grad av sikkerhet.
Metoder for å dokumentere endringer i helsetilstand på grunn av akvakulturutslipp
En artsspesifikk sammensatt påvirkningsindeks, som kombinerer visuell helsetilstandsindeks og b-MBI, vurderes som en effektiv metode for å måle stressrespons hos kålrabisvamp, men det vil styrke indeksen å inkludere analyse av lysosomal membranstabilitet for å oppnå høyere sikkerhet i vurderingen. Mer presis kunnskap om effekter av utslipp på kålrabisvamp og biologiske og fysiologiske påvirkningsmekanismer, kan oppnås ved bruk av flere molekylære metoder (lipid- og fettsyreprofiler, 16S-samfunnsstruktur, RNA-sekvensering).
1.4 - Sjøtre
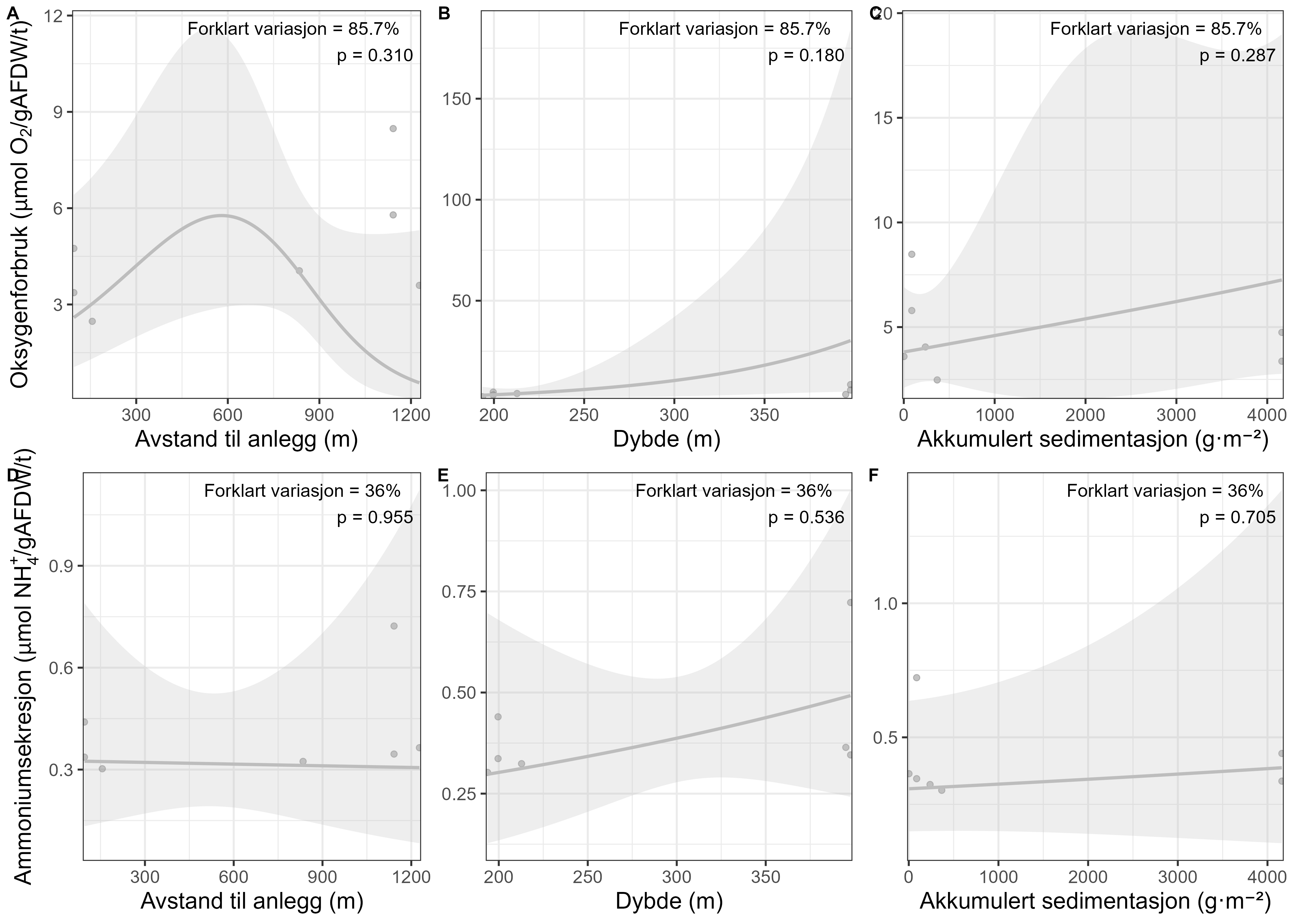
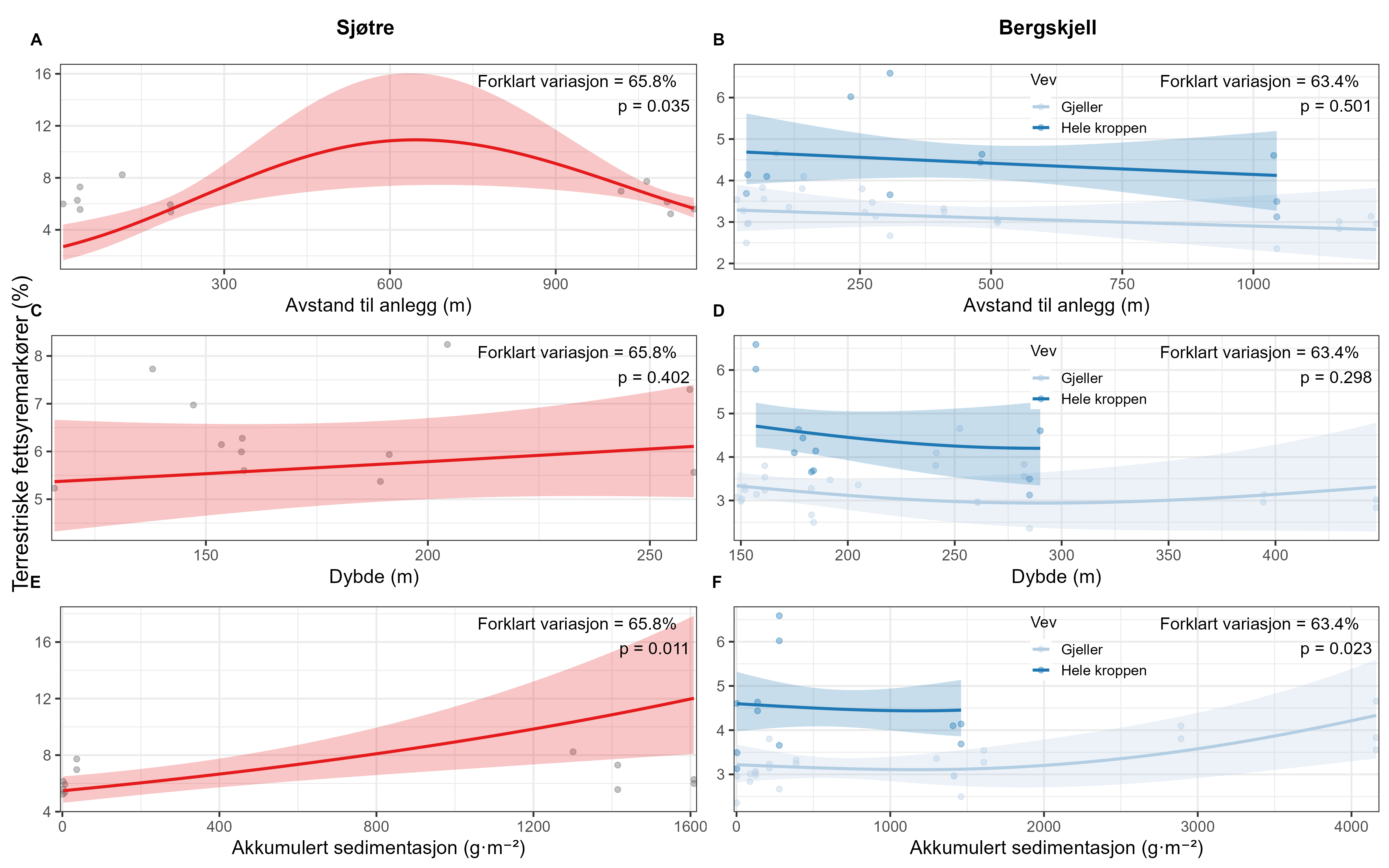
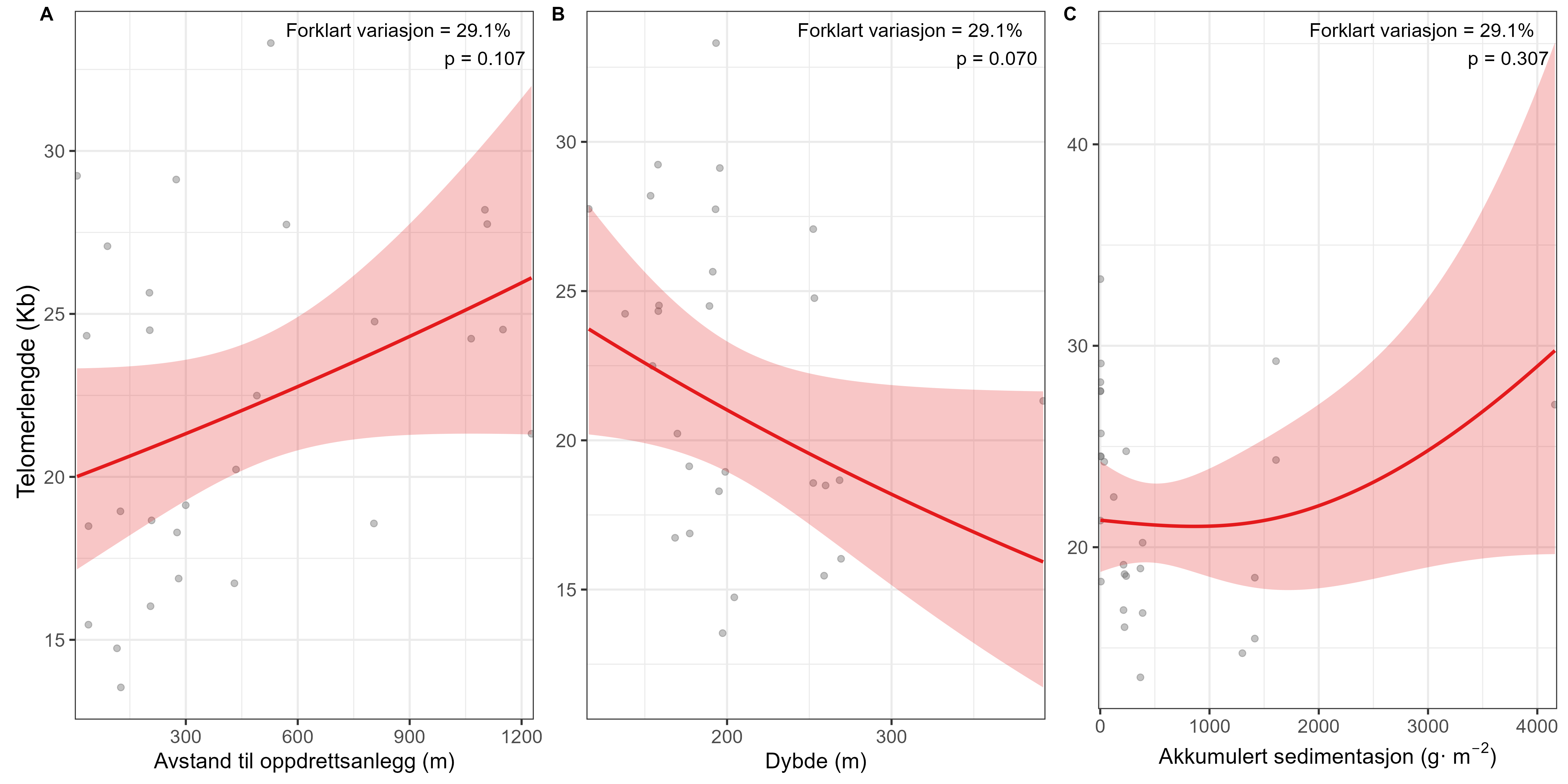
Følsomheten til sjøtre for organisk utslipp fra akvakultanlegg ble vurdert ved bruk av visuell helsevurdering, O₂-forbruk, ammoniumsekresjon, lipid- og fettsyreprofiler, 16S- og RNA-sekvensering og telomeranalyser. Endringer i helsetilstand hos sjøtre var vanskelige å observere visuelt, men metabolske og molekylære responser knyttet til sedimentasjon over 1000 g·m⁻² ble imidlertid observert innenfor en avstand på 200 m fra anleggene. Resultatene antyder at oksygenforbruket hos sjøtre i noen grad kan påvirkes av miljøfaktorer knyttet til oppdrettsaktivitet og sedimentasjonsnivå, mens ammoniumsekresjon ikke ble påvirket. Sjøtre viste noe forhøyede nivåer av terrestriske fettsyrer nær anleggene, men påvirkningen på fettsyresammensetningen var begrenset. Økt sedimentasjon førte til mindre endringer i fettsyresammensetningen. De transkriptomiske responsene var derimot mer uttalte og kan tyde på stressrelaterte endringer i vert–mikrobe-interaksjoner. Økt akkumulering av sediment på vevet, vevsnedbrytning og slimproduksjon ble observert hos individer eksponert for 5–10 g·m⁻² organisk materiale per dag i laboratorieforsøk. Døgnrytmen i polyppaktivitet ble forstyrret hos sjøtre eksponert for en konsentrasjon på 0,5 g·m⁻²·dag⁻¹. Dette tilsvarer sedimentasjonsnivåer som man kan finne 300 m fra et oppdrettsanlegg. Basert på resultater fra felt og forsøk, vurderes sjøtre til å ha moderat til høy følsomhet for organisk utslipp fra oppdrettsanlegg, med høy grad av sikkerhet. Både den visuelle helsetilstandsindeksen og telomerlengden viser en viss nedgang i helsetilstand med økende dybde.
Metoder for å dokumentere endringer i helsetilstand på grunn av akvakulturutslipp
Basert på de utviklede helsekriteriene vi fant få sjøtre få negative effekter av oppdrettsutslipp. Dette tyder på at disse kriteriene ikke er godt egnet til å påvise negative effekter av sedimentering for arten. Derfor bør den sammensatte påvirkningsindeksen, som kombinerer visuell helsetilstandsindeks og b-MBI, styrkes med data fra andre metoder knyttet til fysiologiske effekter på korallen. Metoder som måling av oksygenforbruk, lipid- og fettsyreprofiler samt 16S-sekvensering kan brukes til å karakterisere endringer i sjøtres helsetilstand som følge av utslipp fra oppdrettsanlegg, og kan videreutvikles for å styrke den sammensatte påvirkningsindeksen. Disse metodene er imidlertid kostbare og tidkrevende, og krever invasiv prøvetaking, så videre arbeid bør sette søkelys på videre utvikling og uttesting av den visuelle helsetilstandsindeksen in situ .
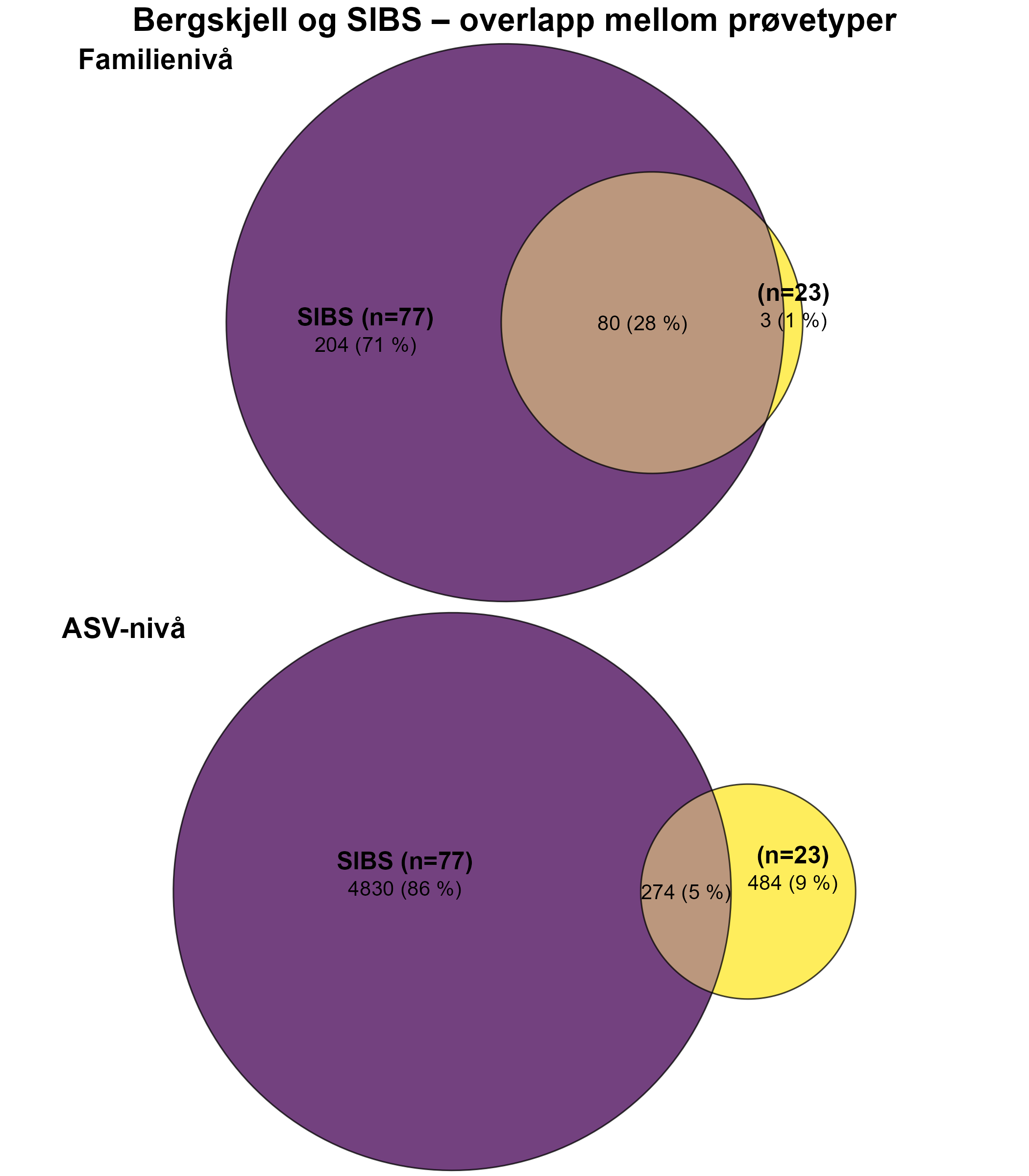
1.5 - Bergskjell
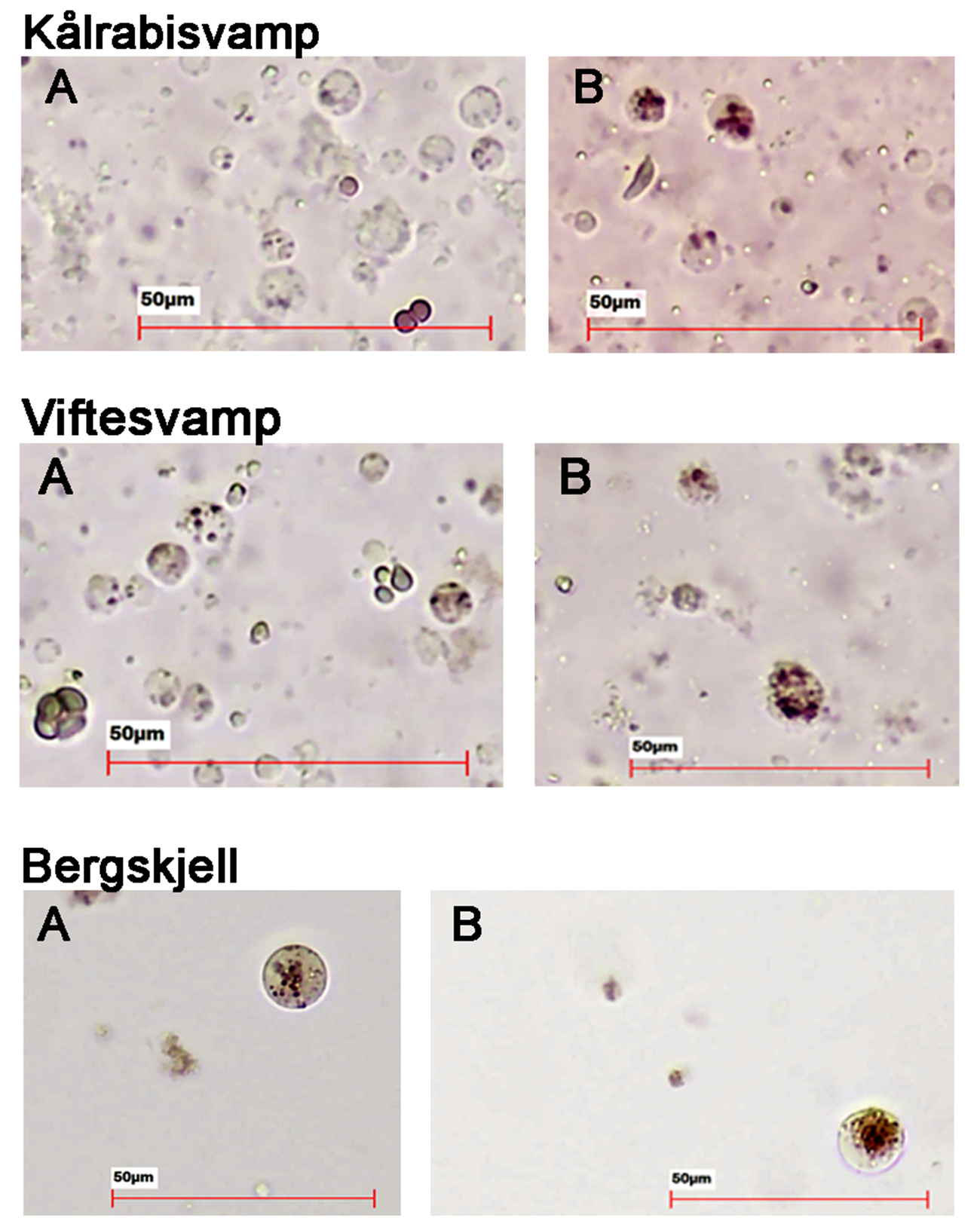
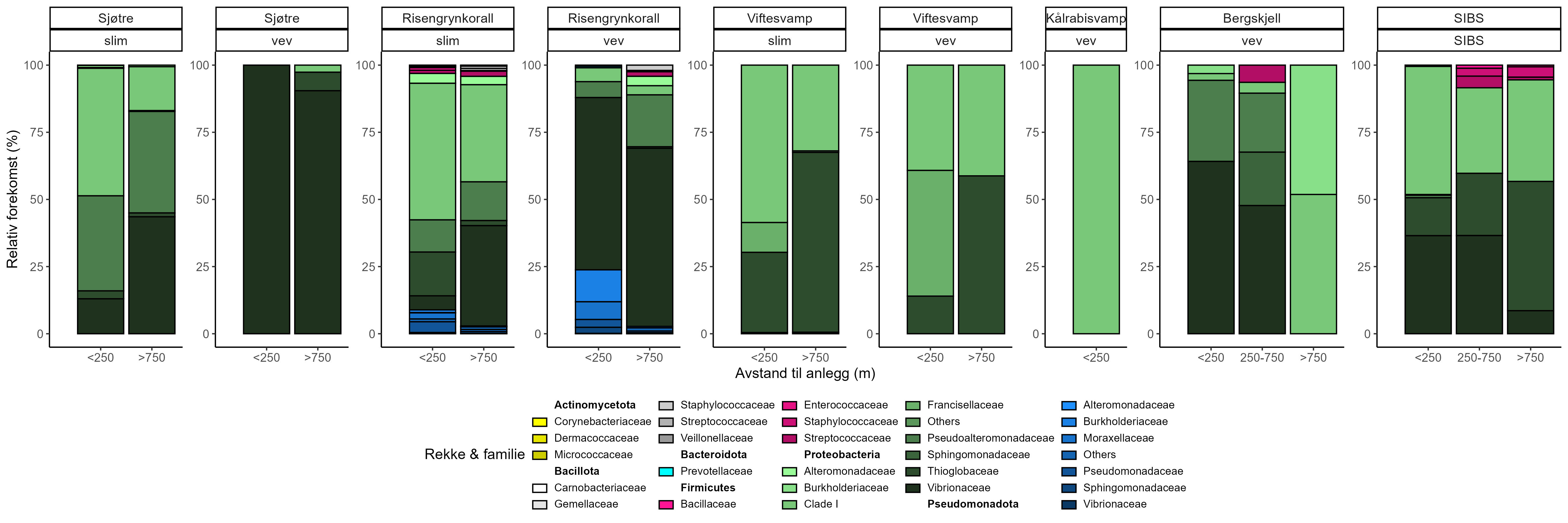
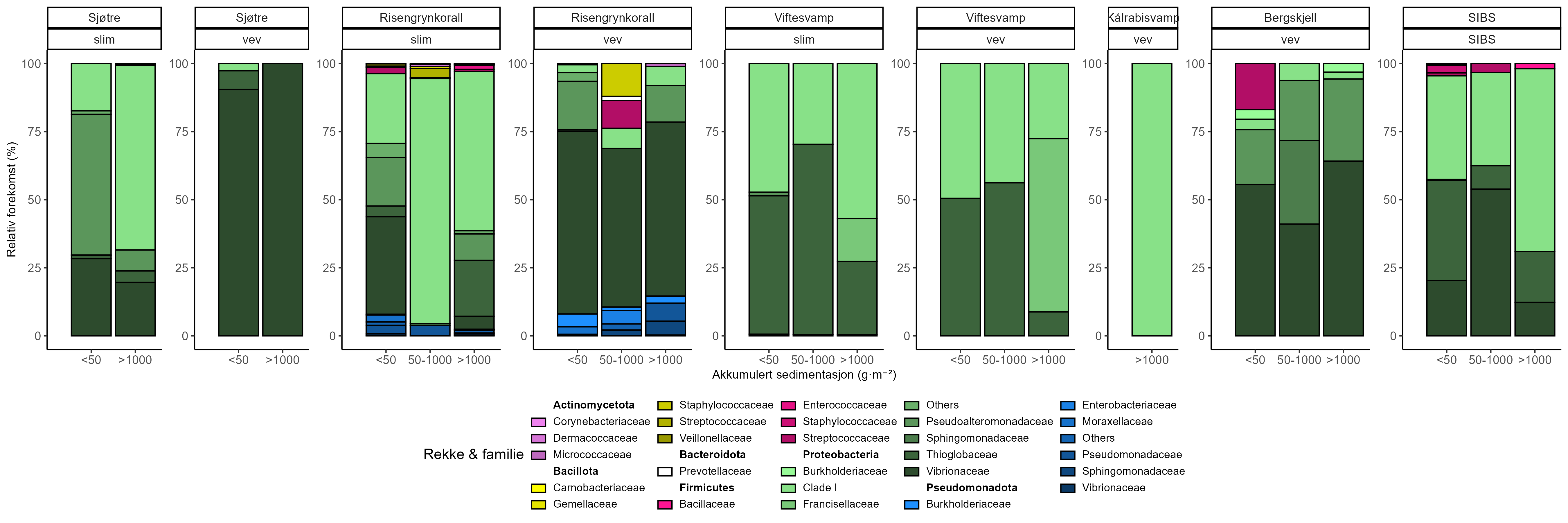
Følsomhet til bergskjell for organisk utslipp fra akvakultur ble vurdert ved bruk av lysosomal membranstabilitet, 16S- og RNA-sekvensering. Bergskjell viste noe cellulært stress basert på lysosomal membranstabilitet innenfor omtrent 100 m fra oppdrettsanleggene, samt endringer i mikrobielle samfunn på gjellevev i områder med høy sedimentasjon nær anleggene (over 1000 g·m⁻²). RNA-prøvene viste påvirkning på metabolske og enzymatiske prosesser knyttet til cellulært stress, immunrespons og reparasjonsmekanismer. Resultatene indikerer derfor at bergskjell er sensitive kun for høye nivåer av sedimentasjon nær merdene. Det vurderes derfor at bergskjell har lav følsomhet for utslipp fra oppdrettsanlegg med en moderat grad av sikkerhet.
Metoder for å dokumentere endringer i helsetilstand på grunn av akvakulturutslipp
Det var få tydelige visuelle kriterier knyttet til negativ påvirkning på bergskjellhelse som kunne brukes til å utvikle en robust helseindeks for denne arten. Forståelsen av artens respons på utslipp fra akvakultur var derfor basert på fysiske prøver. Lysosomal membranstabilitet ble ansett som en effektiv metode for å påvise og måle subletale effekter hos bergskjell, som avdekket cellulære stressresponser hos individer nær merdene.
1.6 - Sjøbusk
Følsomheten til sjøbusk ble vurdert kun ved bruk av visuell helsetilstandsvurdering, og et lite utvalg individer ble samlet inn, noe som begrenset konklusjonene om artens toleranse og følsomhetsterskler. Likevel indikerte den visuelle helsetilstandsindeksen en viss følsomhet for organiske utslipp nært oppdrettsanlegg. Sjøbusk eksponert for høye konsentrasjoner av fiskefekalier under eksponeringsforsøk viste høye nivåer av akkumulering av sediment på korallene, men et begrenset tidsseriedatasett for arten gjorde det ikke mulig å vurdere effekter på polyppaktivitet. Det vurderes derfor at til tross for noen indikasjoner på følsomhet for akvakulturutslipp, er sjøbusk faktiske følsomhet foreløpig ukjent.
Metoder for å dokumentere endringer i helsetilstand på grunn av akvakulturutslipp
Det ble funnet relativt få sjøbuskindivider rundt de syv oppdrettsanleggene som ble undersøkt, og derfor ble andre arter med høyere tetthet valgt for utvikling av fysiologiske, biokjemiske og molekylære metoder. Basert på indikasjonene fra den visuelle helsetilstandsindeksen om at arten kan være sensitiv for organiske akvakulturutslipp, vil sjøbusk være i fokus for utvikling av metoder som kan dokumentere endringer i helsetilstand.
1.7 - Øyekorall
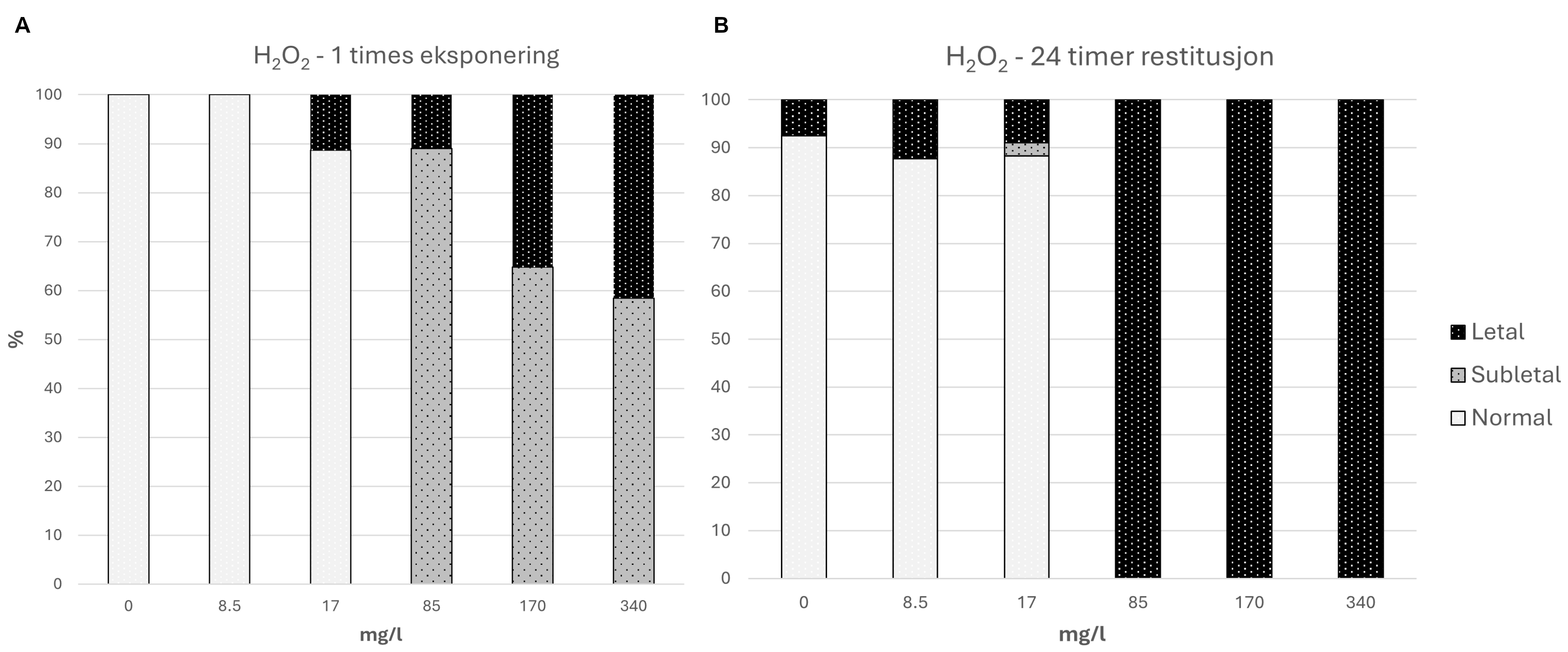
Følsomhet til øyekorall for utslipp av organiske materiale ble vurdert kun ved bruk av visuell helsetilstandsvurdering, oksygenforbruk og ammoniumsekresjon, men ingen effekt av utslipp fra oppdrettsanlegg ble observert med disse metodene. Antallet individ som ble undersøkt var imidlertid lavt, noe som gjør det vanskelig å trekke konklusjoner fra disse resultatene, og følsomhet forblir foreløpig ukjent for øyekorall basert på feltprøver i denne studien. Øyekorall var imidlertid den minst følsomme korallarten ved eksponering for fiskefekalier under eksponeringsforsøk, og tentakkelutstrekking økte ved høye konsentrasjoner, noe som indikerer at korallene må øke tentakkelaktiviteten for å rense seg for partikler. Eksperimentelle laboratoriestudier har vist at legemiddelet hydrogenperoksid gir letale effekter på øyekorall-larver opptil en kilometer fra anlegg, mens azametifos gir subletale effekter opptil om lag 200 m fra anleggene, noe som kan redusere artens reproduksjonspotensial.
Metoder for å dokumentere endringer i helsetilstand på grunn av akvakulturutslipp
Som følge av det lille antallet av øyekorall som ble samlet inn i felt i dette prosjektet, ble det lagt mindre vekt på utvikling av metoder for denne arten, m en vi fokuserte på denne arten i flere laboratorieforsøk. Endringer i polyppaktiviteten hos øyekorall kan brukes til å vurdere artens respons på eksponering for partikulært organisk materiale og avlusningsmidler under laboratorieforhold. Visuelle timelapse-observasjoner av polyppaktivitet fra tentakkelutstrekning anbefales derfor også å brukes i feltbaserte studier, og automatiske bildeanalysemetoder bør videreutvikles for mer effektiv datautvinning fra store bildedatasett. Arten bør derfor være i fokus for fremtidig forskning, spesielt på grunn av dens høye forvaltningsmessige relevans.
1.8 - Metoder for å vurdere anrikningsgradient rundt oppdrettanlegg
Substrate Independent Benthic Sampler (SIBS) ble med suksess tilpasset for ROV ( Remotely Operated Vehicles )-rigging på dypt vann (< 500 m), og muliggjorde direkte prøvetaking av overflatesediment og tilhørende mikrobielle samfunn fra bratte vegger og hardbunn. Denne metoden gir et pålitelig, ikke- inngripende alternativ til tradisjonell grabb-prøvetaking i hardbunnshabitater og muliggjør kvantitativ miljø-DNA-prøvetaking for beregning av «Bakteriell Meta-strekkoding Biotisk Indeks» (b-MBI). Resultatene tyder på at mikrobielle samfunn kan respondere på tilførselsgradienter langt utover områder der synlige bunneffekter er åpenbare, og fremhever metodens potensial som et tidlig varslingsverktøy for overvåking av oppdrettspåvirkning. Imidlertid er en videre utvikling av en regionalt validert indikator-database nødvendig, og fremtidige b-MBI-versjoner bør inkludere relativ forekomst og bedre normalisering. b-MBI er basert på en referansedatabase fra New Zealand (Cawthron Institute), og regional validering mot norske mikrobielle samfunn er nødvendig før metoden kan anses som validert for norske forhold. På grunn av strømretningen kan områder nedstrøms få mer sedimentasjon selv om de ligger lenger unna enn områder nærmere anlegget, men utenfor hovedstrømretningen. Disse funnene understøtter et skifte fra avstandsbaserte buffersoner til steds-spesifikke vurderinger basert på hydrodynamisk modellering og predikert sedimentavsetning. Modellering av dominerende strømretning og deposisjonsområder bør derfor være sentralt for å definere det faktiske påvirkningsområdet rundt akvakulturanlegg.
1.9 - Metoder for å dokumentere endringer i helsetilstand til dypvannsarter på grunn av akvakulturutslipp
Visuelle kriterier for helsetilstand kan brukes som en ikke-inngripende metode for å oppdage og overvåke subletale effekter av utslipp fra akvakultur på viftesvamp og risengrynkorall. Kriteriet vevmisfarging av kålrabisvamp og sjøbusk er nødvendig for å vurdere endringer i helsetilstand visuelt, men i det nåværende datasettet kunne dette kun vurderes fra innsamlede individer. Det anbefales å utvikle metoder for å vurdere fargeendringer under vann, og å teste kriteriene og den visuelle helsetilstandsindeksen på flere datasett fra ulike lokaliteter. Kombinasjonen av visuell helsetilstand hos organismer med standardiserte tetthetsestimater gjør det mulig å vurdere habitattilstand og artsfordeling, og gir en bredere økologisk kontekst for å tolke individnivå-responser på stress. Lysosomal membranstabilitet (LMS) er også en effektiv, men inngripende metode for å dokumentere subletale effekter hos viftesvamp, kålrabisvamp og bergskjell. Siden andelen destabiliserte lysosomale celler er korrelert med de visuelle kriteriene for svampene, vil den visuelle vurderingen være å foretrekke som en ikke-inngripende metode. Lysosomal membranstabilitet kan også benyttes til vurdering av korallhelse. Under prosjektet viste LMS-metoden sterkt potensial som et raskt og kostnadseffektivt verktøy for å vurdere fysiologisk stress hos sårbare bunnorganismer i felt. Pågående forbedringer inkluderer mulig integrasjon av AI-basert bildeanalyse for å standardisere celletolkning og redusere observatør-bias ved manuell mikroskopi. Selv om fettsyre (FA)-analyser fortsatt er et kraftig verktøy for å undersøke diett, energireserver og økologiske responser, er terrestriske FA-markører alene utilstrekkelige for å påvise oppdrettsrelaterte innslag i disse dypvannsartene. Kombinert med bredere analyser kan de imidlertid gi et mer komplett bilde av bunnorganismenes respons på organisk tilførsel og sedimentasjon. Basert på denne studien fremstår modellert spredning av organiske partikler , visuell helsevurdering , LMS og b-MBI som metoder som kan bidra til å støtte forvaltningsmessige avgjørelser, særlig i vurdering av påvirkningsgradienter og identifikasjon av sensitive arter og habitater. Molekylære responser (transkriptomikk og mikrobiom) bidrar til forståelsen av påvirkningsprosesser, men krever store datasett, spesialisert analytisk og bioinformatisk kompetanse, standardiserte terskelverdier og kalibrering mot økologiske endepunkter før de kan brukes direkte i regelmessig overvåking. Den sammensatte påvirkningsindeksen (visuell helsetilstand, b-MBI og fysiologiske målinger for kalibrering) viser imidlertid lovende potensiale for å integrere flere responssignaler fra flere metoder i én samlet vurdering, men betydelig videre arbeid er nødvendig før den kan anses som en operativ metode.
1.10 - Tiltak for å redusere effekten av akvakulturutslipp på sårbare bunndyrarter
For arter med moderat til svært høy følsomhet for utslipp fra akvakultur er den eneste måten å redusere påvirkning på å unngå overlapp med utslipp av organisk materiale. Høytoppløselig modellering av avtrykket av partikulært organisk materialer kan identifisere faktisk overlapp med disse artene, i stedet for å bruke en enkel sirkulær avstand, og legge til rette for bedre plassering av anlegg. Ny teknologi som samler en del av partikulært utslipp kan også redusere sedimentasjonspresset på følsomme arter, og prioritering av slike systemer på aktuelle lokaliteter vil bidra til å redusere påvirkningen fra oppdrett. Dødelige og subdødelige effekter av legemidlene hydrogenperoksid og azametifos på larver av øyekorall kan reduseres ved å unngå bruk under øyekorallens gyting om våren på anlegg hvor øyekoraller som ligger innenfor 1 km fra merd. Voksne øyekoraller viser også fysiologisk respons på emamectin, og å unngå bruk av dette medikamentet på anlegg med øyekorall innenfor påvirkningsområdet vil redusere effekten på arten.
2 - Bakgrunn
Dypvannshabitater som kaldtvannskorallrev, svampsamfunn, korall- og sjøfjærhager domineres av filterfødere og artene som danner disse habitatene er vurdert sårbare for menneskelig påvirkning, har langsom vekst og gjenopprettingskapasitet. Derfor regnes kaldtvannskorallrev, svampsamfunn, korall- og sjøfjærhager som truede naturtyper og i tilbakegang etter flere nasjonale og internasjonale kriterier for bevaring av marint biologisk mangfold; OSPAR, ICES og Artsdatabanken. Som enkeltarter regnes bambuskoraller (Isidella lofotensis), sjøtre (Paragorgia arborea) og øyekorall (Desmophyllum pertusum) som nær truet, men andre koraller og svamper, inkludert sjøbusk (Paramuricea placomus), risengrynkorall (Primnoa resedaeformis), blomkålkorall (Drifa glomerata), viftsvamp (Phakellia ventilabrum) og kålrabivamp (Geodia barretti), regnes som livskraftige. Koraller og svamphabitater finnes langs kysten og kontinentalsokkelen, der de utsettes for økte mengder oppvirvlet bunnsediment som følge av menneskelig aktivitet, inkludert bunntråling og oljeutvinning. Langs kysten er de viktigste kildene til økt partikulært organisk materiale utslipp fra havbruk, undersjøisk deponering av gruveavfall og bunntråling.
Sjeldenhet er vurdert til sårbarhet i rødliste- og bevaringskriterier, både nasjonalt og internasjonalt. Disse sårbare bunnhabitatene og artene kan være vanlige regionalt der forholdene samsvarer med deres spesifikke krav, men de er samtidig begrenset til disse områdene og sjeldne på nasjonalt eller internasjonalt nivå. For eksempel, koraller og svamper er avhengige av moderate til sterke strømmer for transport av næringsstoffer og kan bli tallrik i slike områder langs kysten. Kystnære oppdrettsanlegg er også avhengige av strømmer for å fjerne organiske partikulært utslipp og sikre god oksygentilførsel til fisken. Derfor overlapper sårbare bunnhabitater og -arter med flere hundre eksisterende og planlagte oppdrettslokaliteter. Dette overlappet har gitt motivasjon for forskning på de mulige effektene og konsekvenser av utslipp (næringssalter, legemidler, fremmedstoffer og partikulært organisk materiale) fra oppdrettsanlegg på artene som danner disse sårbare bentiske marine habitatene. Oppdrettsanlegg for matfisk påvirker bunnhabitatet hovedsakelig gjennom utslipp av fôrrester, kjemikalier og fekalier. Nær anleggene oppstår sterk organisk anrikning av bunnen, men effektene avtar med økende avstand og naturlige forhold gjenfinnes vanligvis innen 200–1000 m (Hargrave mfl., 2008; Keeley mfl., 2012; 2019). Til tross for eksisterende kunnskap er det fortsatt betydelige kunnskapshull om hvordan utslipp fra oppdrettsanlegg påvirker arters helse, deres tålegrenser og hvordan effektene best kan måles og overvåkes. Denne mangelen på kunnskap gjør at myndighetene i mange tilfeller kun kan fatte beslutninger om nye eller utvidede lokaliteter basert på et føre-var-prinsipp, med en antakelse om at koraller og svamphabitater er særlig sårbare for forstyrrelser.
Svamparter som ofte observeres individuelt eller som del av en aggregasjon i svampehager i kystområder med fiskeoppdrett inkluderer: kålrabisvamp, viftesvamp og Mycale lingua, ved rev og fjordvegger (Klitgaard og Tendal, 2004; Kutti mfl., 2013). Andre svamper av slektene Craniella, Weberella og Polymastia er funnet rundt oppdrettsanlegg på blandede substrater (Dunlop mfl., 2021; Laroche mfl., 2022). Svamper filtrerer aktivt store mengder vann (500–1000 L vann per dag) for næringsstoffer og bakterier (Kutti mfl., 2013) og vil derfor også filtrere organisk material fra oppdrettsanlegg. Flere studier antyder at svamper er følsomme for sedimentasjon. Dette kan redusere vekst, overlevelse og reproduksjon ved å svekke filtrering, oksygentilførsel og næringsopptak (Tjensvoll mfl., 2013). Et kortidsforsøk med kålrabisvamper i relativ høye (50 mg·L⁻¹) og lave (5 mg·L⁻¹) konsentrasjoner av suspendert partikulært oppdrettsfiskefôr indikerer at forhøyede nivåer av oppdrettsfôr negativt påvirker svampers helsetilstand. Denne konklusjonen er basert på økt respirasjon og energiforbruk under høye utslippsnivåer, i tillegg til analyser av fettinnholdet og fettsyreprofiler (Kutti mfl., 2016a). Polymastia, Weberella bursa og Craniella, plassert ved, et oppdrettsanlegg i Nord-Norge i 7 måneder, hadde redusert overlevelse, men ikke redusert oksygenforbruk hos de som overlevde (Laroche mfl., 2021, 2022). Dette samsvare med observasjonene at Polymastia var vanlige mer enn 2 km fra anlegget, men mindre vanlige nær anlegget, noe som viser at utslipp påvirker helse og overlevelse utover det målte markører avdekket (Dunlop mfl., 2021). Andre resultater viste ingen signifikant effekt av oppdrettsutslipp på mikrobiomet til svamp (W. bursa), noe som tyder på at terskelen for å utløse en slik respons kan være relativt høy (Laroche mfl., 2021). Konsekvensene av oppdrettsutslipp for svamper er studert ved endringer i tetthet og sammensetning av svampesamfunn langs påvirkningsgradienter at tre anlegg i Vest-Finnmark (Dunlop mfl., 2021). Viftesvamp (Phakellia spp.) var vanlige mer enn 1 km fra anlegget, men sjeldne innen 250 m, mens kålrabisvamp (Geodia spp.) viste uendret tetthet og en høyere motstandskraft mot utslippspåvirkning. Tilstedeværelsen av glassvampene, inkludert viftesvamp (Phakellia spp.), viste også en negativ sammenheng med sedimentasjon fra oppdrettsanlegg langs kysten av British Columbia (Sutherland mfl., 2018).
Sårbare korall-naturtyper (korallrev og korallskog) og korallarter som hornkoraller og bløtkoraller (blomkålskoraller og sjøfjær), som er på rødlisten, er funnet både på bløtbunn, blandet bunn og fjordvegg. Spredte forekomster av øyekorallrev er registrert langs norskekysten og i fjordområdene. Øyekorall kan overleve en viss eksponering av partikulært materiale, men dette utløser en stressrespons som viser seg gjennom økt stoffskifte og slimproduksjon. Dette antas å være en mekanisme for å fjerne partikulært materiale. Økt energiforbruk til slike forsvarsmekanismer fører til redusert vekst (Kutti mfl., 2015). Utslipp fra oppdrett kan redusere metabolismen hos øyekorall og mulig redusere vekst og energireserver med henholdsvis 70 og 50 % innenfor influensområdet til oppdrettsanlegg (250–1 km), noe som er dokumentert i en ettårig felttransplantasjonsstudie på Vestlandet (Kutti mfl., 2022). Effektene av både høye og lave konsentrasjoner av impregneringsstoffene tralopyril og sinkpyrition (ZnPt) på korallfragmenter ble undersøkt under eksperimentelle forhold. Øyekorall som ble eksponert for høye konsentrasjoner av disse stoffene (100 μg·L⁻¹ sinkpyrition + 10 μg·L⁻¹ tralopyril) viste redusert respirasjonsrate, og koraller eksponert for ZnPt viste tegn til oppløsning av skjelettet (Hunvik, 2025).
Det er gjort begrenset forskning på effektene av partikulært materiale fra havbruk på hornkorallartene sjøbusk, sjøtre og risengrynkorall, men disse artene er observert i nærheten av oppdrettsanlegg. Filterfødende muslinger, som bergskjell (Acesta excavata), er også vurdert sårbare for partikulært organiske materialer, noe som reduserer ernæringseffektiviteten. Arten forekommer på bratte fjordvegger og ofte i tilknytning til korallrev og korallhager. Oppvirvlet sediment fra gruveavfall reduserte den fysiologiske helsen til bergskjell under en 40-dagers eksperimentell eksponering. Dette ble konkludert ut fra en økning i andelen destabilisert lysosomale celler, lavere O:N-forhold og økt ammoniumutskillelse ved forhøyede sedimentnivåer (Scanes mfl., 2024).
FAO definerer sårbare marine økosystemer som områder der «sårbarheten til bestander, samfunn og habitater må vurderes i forhold til spesifikke trusler» (Food og Organization, 2009). Til tross for eksisterende kunnskap om hvordan kystnære koraller og svamper påvirkes av utslipp fra havbruk, er deres sårbarhet ennå ikke fullt ut vurdert.
2.1 - Formål
Hovedmålet med prosjektet VDWS Transition var å dokumentere effekter og toleransegrenser for utslipp fra oppdrettsanlegg på relevante arter av korall og svamp samt å utvikle nye metoder for vurdering av helsetilstanden til utvalgte korall- og svamparter ved eksisterende havbrukslokaliteter.
Delmål
Å samle inn fysiske prøver av utvalgte korall- og svamparter fra flere forskjellige regioner, anlegg og utslippsgradienter og dokumentere individenes helsetilstand ved målinger av cellulære, fysiologiske og biologiske responsparametere.
Å gjennomføre laboratoriestudier for å definere relevante arters relative følsomhet/toleranse for forskjellige typer utslipp.
Å indentifisere enkle og reproduserbare metoder (basert på fysisk prøvetaking og visuelle undersøkelser) som kan brukes til å vurdere relevante arters helsetilstand, og eventuelt dødelighetsrisiko, i felt.
Å rapportere prosjektets funn om effekter, toleransegrenser og nye overvåkningsmetoder til forvaltningen og næringen og bidra til en forbedret forutsigbarhet for næringsutvikling i kystsonen.
Prosjektutvidelse primo 2024
Å gjennomføre ekstra tokt for å ta prøver av flere lokaliteter og samle flere arter for laboratorieprøver og vevsanalyser.
Å modifisere prøvetaking med ROV for å optimalisere prøvetaking på dypvannsarter.
2.2 - Prosjektets omfang
2.2.1 - Prosjektorganisering
Prosjektet startet i januar 2023 og sluttrapport ble levert i februar 2026. Det første toktet ble gjennomført i september 2023 i Møre og Romsdal og i Nordland for å samle faunaprøver ved hjelp av ROV-systemer langs en anrikningsgradient samt ved referansestasjoner én kilometer fra sju oppdrettsanlegg, antatt upåvirket av utslipp. Forsinkelser i anskaffelse av HI sin kyst-ROV førte til at en ROV ble leid fra NTNU. Tekniske problemer på første tokt gjorde at to ekstra tokt ble gjennomført i 2024 i Møre og Romsdal og i Nordland. Anrikningsgradienten ved hvert oppdrettsanlegg ble beskrevet gjennom modellert sedimentasjon, verifisert med sedimentfeller og en mikrobiell indeks (Bakteriell Meta-strekkoding Biotisk Indeks, b-MBI). Bilder og fysiske prøver av koraller, svamper og muslingen bergskjell ble brukt til å teste ulike metoder for å måle effekter av utslipp. Disse inkluderte visuelle indekser, molekylære multi-omics-analyser, lipid- og fettsyreanalyser og lysosomal membranstabilitet. Et siste tokt i august 2025 i Nordland muliggjorde videre testing av den visuelle indeksen og LMS-metodene. Toktene ble ledet av Havforskningsinstituttet på forskningsfartøyet «Prinsesse Ingrid Alexandra», og den visuelle og fysiske prøvetakingen ble gjennomført av Nigel Keeley, Katherine Dunlop, Ana S. Gomes, Birte Schuppe, Tina Kutti og Mona Fuhrmann. Utviklingen av SIBs-prøvetakeren for innsamling av utslipp på harde substrater i dypt vann ble utført av Nigel Keeley, og DNA-ekstraksjon, sekvensering og bioinformatikk ble utført av Xavier Pochon og Olivier Laroche ved Cawthron Institute i New Zealand. Modellering av spredning og sedimentering av partikulært organisk materiale ved lokalitetene ble utført av Pål Næverlid Sævik ved Havforskningsinstituttet. Artsidentifisering av viftesvamp med molekylær identifikasjon ble utført av Møreforsking (Trond Roger Oskars, Jenny Elisabet Ullgren og Eirin Svanoe-Hafstad). Utviklingen av den visuelle helsetilstandsindeksen ble ledet av Katherine Dunlop med Mona Fuhrmann. Analysen av gametogenese og fekunditet hos risengrynkorall og sjøtre ved bruk av histologiske metoder ble ledet av Johanna Järnegren (NINA). Analysen av fysiske prøver for lysosomal membranstabilitet ble utført av Tina Kutti, Katherine Dunlop og Birte Schuppe, og dataanalysen ble utført av Ana S. Gomes. Målinger av oksygenforbruk og ammoniumsekresjon ble utført om bord av Birte Schuppe, Tina Kutti, Mona Fuhrmann og Ana S. Gomes, og dataene ble analysert av Ana S. Gomes. Sonnich Meier ledet analysen av prøver for lipid- og fettsyreanalyser ved hjelp fra Siri Aaserud Olsen og Ana S. Gomes mens molekylære multi-omics-analyser (sekvensering av 16S rRNA, transkriptomanalyse og DNA-metyleringsanalyser) var et samarbeid mellom Olivier Laroche og Ana S. Gomes. Arbeid om telomerlengde på sjøtre-prøver ble ledet av Pierre Blévin med kolleger; Astrid Harendza, Sabine Cochrane og Jocelyn Palerud (Akvaplan-niva), og laboratorieanalysene ble utført av Andreaz Dupoué (Ifremer) og Alexandre Ottaviani (Université Côte d’Azur). Ekstra finansiering ble skaffet av Akvaplan-niva og Havforskningsinstituttet fra Fram Centre Incentive Funding for å analysere prøver samlet inn under prosjektets tokt for telomerlengde. Arts-spesifikk sammensatt påvirkningsindeks (Composite Impact Index) ble utviklet av Ana S. Gomes.
Tre ulike eksponeringsforsøk ble gjennomført. Ved Nord universitet i Bodø ble effekten av emamektin benzoat på voksen øyekorall undersøkt. Forsøket ble gjennomført av Birte Schuppe under veiledning av Pål Asgeir Olsen og Henning Reiss. Bildedata ble samlet inn ved hjelp av et timelapse-kamerasystem fra NORCE sette opp av Christian Andreas Hansen og Alan Le Tressoler, og bildene ble analysert av Thierry Baussant. Ved NTNU i Trondheim ble effekten av hydrogenperoksid og azametifos på larver av øyekorall testet. Forsøket ble gjennomført av Johanna Järnegren fra NINA. Et tredje eksponeringsforsøk ble gjennomført ved Havforskningsinstituttets anlegg i Austevoll, der fire korallarter (risengrynkorall, sjøtre, sjøbusk og øyekorall) ble utsatt for fire nivåer av sedimentasjon av organisk partikulært materiale (laksefekalier). Nivåene tilsvarte den organiske utslippssedimentasjonen observert innen 300 m fra oppdrettsanleggene i studien. Eksperimentet ble utformet og ledet av Tina Kutti, gjennomført av Helen Rastrick, Cathinka Krogness og Siri Aaserud Olsen, og finansiert av Havforskningsinstituttets program for miljøpåvirkninger av akvakultur. Bildedata ble samlet inn ved hjelp av et timelapse-kamera fra NORCE, og bildene ble analysert av Thierry Baussant.
Ansvarlig institusjon er HI, i samarbeid med kollegaer fra NINA, NORCE, Nord universitet, Møreforsking og Cawthron Instituttet (Tabell 1). Prosjektet ble ledet av Nigel Keeley i 2023 og 2024, og av Katherine Dunlop fra mars 2025. Prosjektets referansegruppe ble etablet for å gi innspill til prosjektets gjennomføring, bidra til å utvikle forvaltningsrelevant kunnskap, samt gi tilgang til informasjon om spesifikke lokaliteter.
Navn
Institusjon
Gruppe
Katherine Mary Dunlop
Havforskningsinstituttet
Prosjektgruppe
Ana S. Gomes
Havforskningsinstituttet
Prosjektgruppe
Nigel Keeley
Havforskningsinstituttet
Prosjektgruppe
Tina Kutti
Havforskningsinstituttet
Prosjektgruppe
Sonnich Meier
Havforskningsinstituttet
Prosjektgruppe
Pål Næverlid Sævik
Havforskningsinstituttet
Prosjektgruppe
Mona Maria Fuhrmann
Havforskningsinstituttet
Prosjektgruppe
Siri Aaserud Olsen
Havforskningsinstituttet
Prosjektgruppe
Olivier Laroche
Cawthron Institute
Prosjektgruppe
Xavier Pochon
Cawthron Institute
Prosjektgruppe
Henning Reiss
Nord Universitet
Prosjektgruppe
Pål Asgeir Olsvik
Nord Universitet
Prosjektgruppe
Birte Katarina Schuppe
Nord Universitet
Prosjektgruppe
Johanna Järnegren
NINA
Prosjektgruppe
Thierry Baussant
NORCE
Prosjektgruppe
Trond Roger Oskars
Møreforsking
Prosjektgruppe
Jenny Elisabet Ullgren
Møreforsking
Prosjektgruppe
Eirin Svanoe-Hafstad
Møreforsking
Prosjektgruppe
Randi Nygaard Grøntvedt
Sjømat Norge
Referansegruppe
Arne Kvalvik
MOWI
Referansegruppe
Silje Ramsvatn
Cermaq
Referansegruppe
Trond Baarset
SalMar
Referansegruppe
Sigfrid Lundekvam
Lerøy Seafood Group
Referansegruppe
Erik Bårdseng
Lerøy Seafood Group
Referansegruppe
Andreaz Dupoué
Ifremer
Samarbeidspartnere
Jocelyn Palerud
Akvaplan niva
Samarbeidspartnere
Alexandre Ottaviani
Université Côte d’Azur
Samarbeidspartnere
Sabine Cochrane
Akvaplan niva
Samarbeidspartnere
Astrid Harendza
Akvaplan niva
Samarbeidspartnere
Pierre Blévin
Akvaplan niva
Samarbeidspartnere
Tabell 1: Medlemmer av prosjektgruppen (de som utfører prosjektet), referansegruppen (rådgivende) og samarbeidspartnere (bidrar med kompetanse, data og analyser).
3 - Metodikk og Resultater
I denne studien ble det benyttet to hovedtilnærminger for datainnsamling: (1) feltbaserte prøver og (2) laboratorieeksperimenter.
Valg av metoder ble styrt av behovet for å vurdere både romlige mønstre i påvirkning fra oppdrettsanleggsutslipp og biologiske responser hos bunnlevende arter langs miljøgradienter som inkluderer anrikningsgradient og dybde. Tidligere studier har vist at kombinasjoner av visuelle helsevurderinger, fysiologiske målinger og molekylære analyser gir økt sensitivitet for å påvise subletale effekter av organisk belastning og andre stressfaktorer i marine økosystemer. På bakgrunn av dette ble det valgt en flerlaget metodisk tilnærming som integrerer feltbaserte observasjoner, biokjemiske og fysiologiske analyser, samt molekylære metoder.
Samtidig stilte både studieartene og de fysiske rammene (dypvann, ROV-basert prøvetaking, begrenset prøvemateriale) krav til betydelig metodisk tilpasning. Flere av metodene som ble benyttet er basert på etablerte protokoller, men måtte testes og optimaliseres som en del av prosjektet for å kunne anvendes på de undersøkte artene og prøvetypene på dypt vann. Dette gjaldt blant annet utvikling og videre optimalisering, som for eksempel av ROV-monterte SIBS-samlere for sedimentprøver, samt lagring og samtidig ekstraksjon av DNA og RNA fra vevsprøver, arbeid som resulterte i en egen metodisk publikasjon (Gomes mfl., 2025). Den kombinerte bruken av etablerte og videreutviklede metoder muliggjorde en robust og helhetlig vurdering av biologiske responser på akvakulturrelaterte påvirkninger.
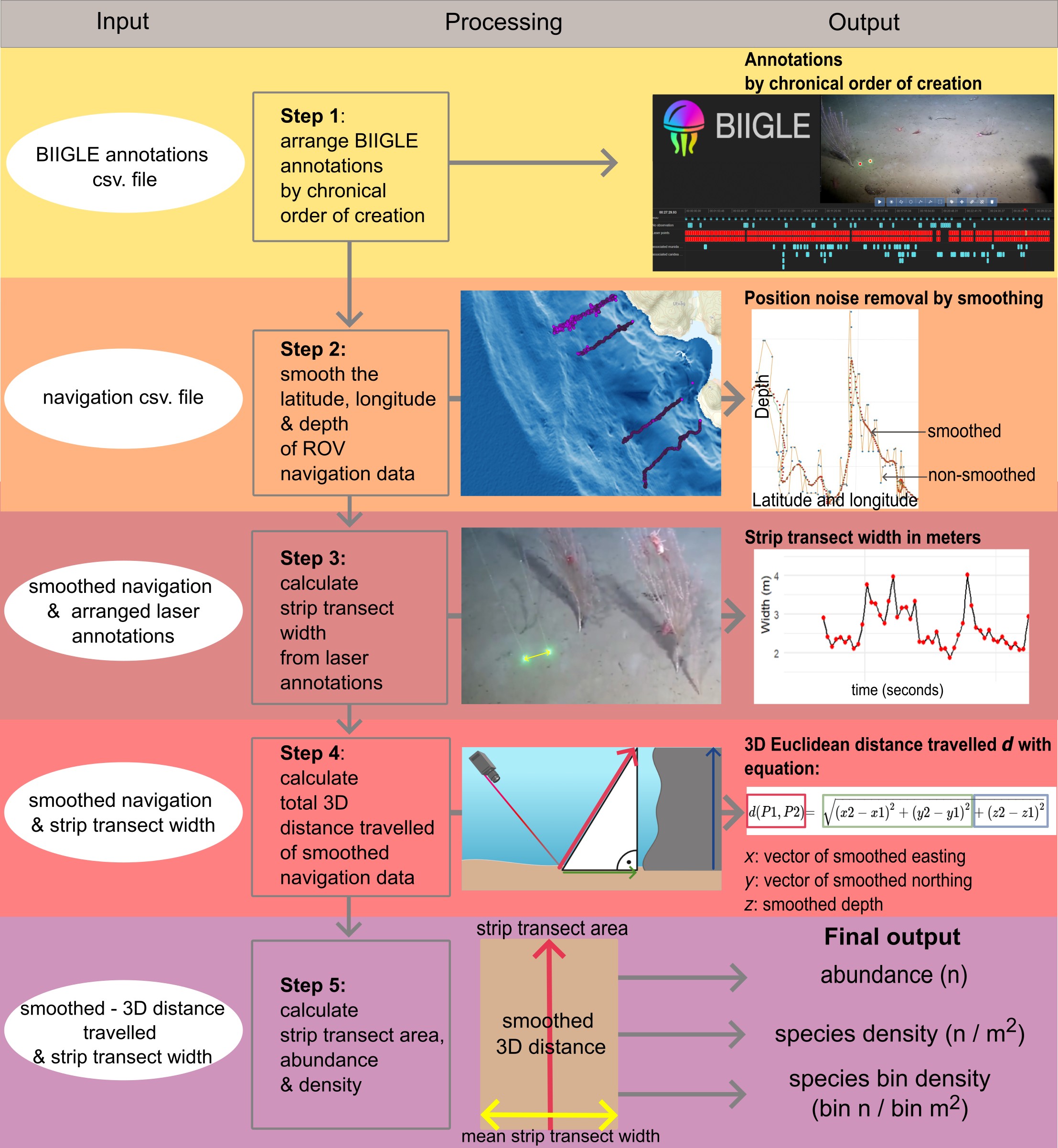
Omfanget og nivået av anrikningsgradienten av partikulært organiske materialer fra anleggene ble kvantifisert med modellering av sedimentasjon kombinert med direkte prøvetaking av sedimenter fra hardbunn og bratte vegger ved hjelp av ROV-monterte Substrate Independent Benthic Sampler (SIBS)-enheter. SIBS-prøvene ble brukt til eDNA-ekstraksjon og 16S-sekvensering for å karakterisere mikrobiell sammensetning i sedimentene. SIBS-sampleren ble tilpasset og videreutviklet for bruk i dypvann (< 500 m) som en integrert del av prosjektet. Dette inkluderte både mekaniske modifikasjoner av prøvetakeren og utvikling av en dypvannskontrollboks som muliggjorde sikker utløsning av prøvetakeren fra overflaten via ROV.
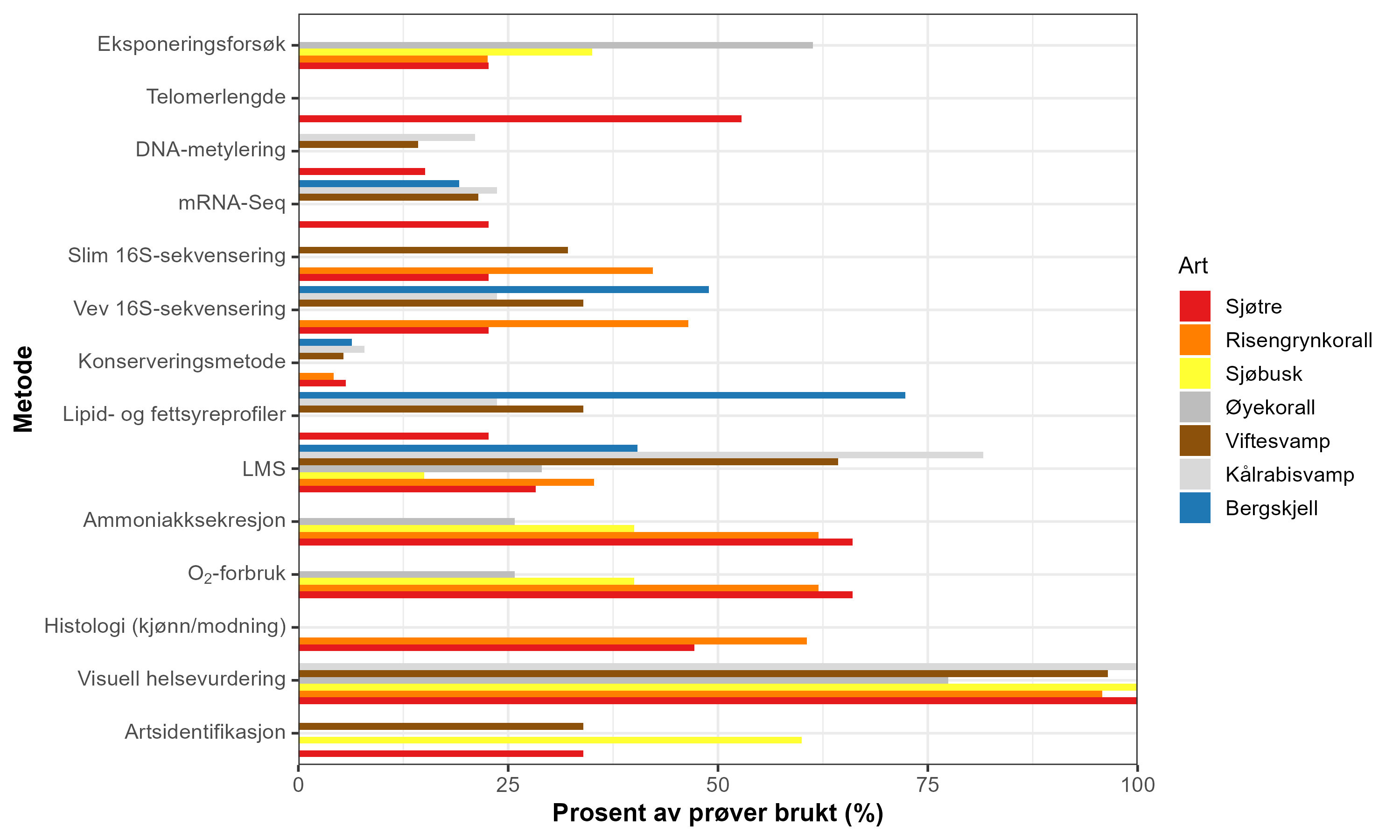
Feltarbeidet ble gjennomført i løpet av fire tokt: ett i 2023, to i 2024 og ett i 2025. Toktene dekket sju lokaliteter i tilknytning til oppdrettsanlegg og referanseområder, og omfattet både visuelle observasjoner og innsamling av biologisk materiale fra utvalgte arter ved hjelp av ROV. For å vurdere helsetilstand og stressresponser hos bunnarter nær oppdrettsanlegg, benyttet vi en kombinasjon av ikke-invasive og invasive metoder som dekker visuelle, fysiologiske, biokjemiske og molekylære måleparametere (Tabell 2). Ikke-invasive tilnærminger inkluderte visuelle helsevurderinger basert på ROV-bilder og laboratoriefotografi, som gir raske og pålitelige indikatorer på vevsskade, kolonitilstand og biotiske interaksjoner uten å påføre skade på organismen. Visuelle vurderinger basert på kriterier som andel vevstap, skadet vev, slimproduksjon og sedimentdekning er brukt tidligere for å dokumentere effekter av termisk stress og oljeeksponering hos gorgoniske koraller (Girard mfl., 2018, 2019; Gómez-Gras mfl., 2022). Negative effekter av termisk stress på kålrabisvamp er også påvist gjennom visuelle stressindikatorer i eksperimentelle studier (Strand mfl., 2017). Invasive metoder omfattet vevsprøver for måling av oksygenforbruk og ammoniumsekresjon, som reflekterer metabolsk aktivitet og energibalanse (Kutti mfl., 2022); lysosomal membranstabilitet, en tidlig og sensitiv indikator på cellulært stress (Strand mfl., 2017; Scanes mfl., 2018); samt lipid- og fettsyreprofilering, som gir informasjon om kosthold og mulig eksponering for organisk materiale fra oppdrett (Redmond mfl., 2010; Meier mfl., 2023). De spesialiserte lipidsammensetningene hos enkelte arter krevde metodiske tilpasninger, som beskrevet i detalj i seksjon 5.9 Lipid- og fettsyreanalyser. Det ble også gjennomført histologiske analyser for å vurdere gametogenese og reproduksjonstilstand, basert på en standard protokoll som ble tilpasset med artsspesifikke modifikasjoner for å sikre anvendelighet, for å undersøke om miljøpåvirkning kunne ha påvirket artenes reproduktive kapasitet.
På molekylært nivå inkluderte vi artsidentifikasjon, 16S rRNA-sekvensering, RNA-seq og DNA-metyleringsanalyser for å oppdage arts-spesifikke responser og undersøke de underliggende mekanismene for stress på cellulært og epigenetisk nivå. Nøyaktig artsidentifikasjon er avgjørende for å evaluere arts-spesifikke tålegrenser, sårbarhet og responser hos sensitive naturtyper (Hofmann og Gaines, 2008). Mikrobiom-analyser (16S rRNA-sekvensering) har blitt brukt for å karakterisere vertstilknyttede mikrobielle samfunn og deres potensielle endringer som følge av miljøforhold. For eksempel viste studier på korallene Acropora hemprichii og Pocillopora verrucosa at mikrobiomsammensetningen responderer arts-spesifikt på antropogen påvirkning (Ziegler mfl., 2019). Tilsvarende ble 16S-metabarkoding anvendt på svampene Craniella og Weberella, bløtkorallen Duva florida og sjøanemonen Hormathia digitata for å undersøke effekter av lakseoppdrett på vertens mikrobiom og funksjonelle profil (Laroche mfl., 2022). RNA-seq (mRNA-sekvensering) har muliggjort påvisning av differensielt genuttrykk hos bentiske evertebrater eksponert for oppdrettspåvirkning. Hos svampen Weberella bursa ble det identifisert endringer i genuttrykk relatert til apoptose, ribosomal aktivitet og stressrespons langs en avstandsgradient fra oppdrettsanlegg (Laroche mfl., 2021). DNA-metyleringsanalyser har blitt brukt for å identifisere potensielle epigenetiske endringer knyttet til miljøstress. Studier på koraller, inkludert Montastraea cavernosa, har vist at endringer i symbiontsammensetning og varmeeksponering kan være assosiert med genom-omfattende endringer i DNA-metyleringsmønstre (Rodriguez-Casariego mfl., 2022), og slike mekanismer er generelt diskutert som sentrale for marin akklimatisering og adaptiv respons (Eirin-Lopez og Putnam, 2019). I tillegg samarbeidet vi med Akvaplan-Niva om analyser av telomerlengde hos sjøtre som en indikator på kumulativt stress og cellulær aldring. Studier på revbyggende koraller som Pocillopora spp. og Porites spp. har vist at telomerlengde påvirkes av miljøforhold og kan reflektere både kort- og langtidseksponering for stress (Rouan mfl., 2023). Disse analysene krevde også betydelig utvikling og optimalisering av DNA-ekstraksjon og analyseteknikker for å kunne anvendes på denne arten. Samlet sett gir denne flerlagede tilnærmingen komplementære data på visuelle, fysiologiske, biokjemiske og molekylære nivåer, noe som muliggjør en robust vurdering av organismenes helsetilstand og respons på akvakulturrelaterte stressfaktorer. En mer detaljert beskrivelse av metodene og begrunnelsen for deres bruk er gitt under hver respektive metodeseksjon. Resultatene ble koblet til avstand fra oppdrettsanlegg, dybde og estimert (modellert) akkumulert sedimentering av organisk materiale.
Metode
Sjøtre
Risengrynkorall
Sjøbusk
Øyekorall
Viftesvamp
Kålrabisvamp
Bergskjell
SIBS
Visuell helsevurdering
✔️
✔️
✔️
✔️
✔️
✔️
-
-
O₂-forbruk
✔️
✔️
-
⚠️
-
-
-
-
Ammoniakksekresjon
✔️
✔️
-
⚠️
-
-
-
-
Lysosomal membranstabilitet
⚠️
⚠️
-
-
✔️
✔️
✔️
-
Lipid- og fettsyreprofiler
✔️
-
-
-
✔️
✔️
✔️
-
Artsidentifikasjon
-
-
-
-
✔️
-
-
-
16S sekvensering
✔️
✔️
-
-
✔️
✔️
✔️
✔️
RNA-seq
✔️
-
-
-
✔️
✔️
✔️
-
DNA-metylering
❌
-
-
-
❌
⚠️
-
-
Telomerlengde
✔️
-
-
-
-
-
-
-
Tabell 2: Oversikt over analyser per art og sedimentprøver (SIBS) fra feltet. Symbolforklaring: ✔️ = analysert/utført, ⚠️ = delvis analysert/tolk med forsiktighet, ❌ = analysen mislyktes, - = ikke aktuelt eller ikke utført
For dataanalysene ble avstand til oppdrettsanlegg beregnet som den korteste avstanden fra hvert observasjonspunkt til nærmeste anleggs polygongrense. Beregningene ble utført i R-versjon 4.4.3 ved bruk av sf-pakken. Geografiske data om norske oppdrettsanlegg ble hentet fra Fiskeridirektoratet sitt oppdrettsregister, inkludert både punktlokasjoner og polygonrepresentasjoner av godkjente produksjonsområder. Kun marine lokaliteter ble beholdt, og landbaserte lokaliteter ble ekskludert før analysene. Alle romlige lag ble transformert til et felles projisert koordinatsystem for å sikre at avstander ble beregnet i meter. For hver artobservasjon ble det nærmeste oppdrettsanlegget identifisert ved et nærmeste-nabo-romlig søk basert på punktlokasjoner, og avstanden til oppdrettsanlegget ble deretter beregnet som den korteste euklidske avstanden mellom hvert observasjonspunkt og grensen til det nærmeste oppdrettsanleggets polygon.
For å undersøke sammenhenger mellom biologiske responser og miljøfaktorer, benyttet vi et utvalg statistiske og modelleringsmetoder som er tilpasset datatypen og økologisk kontekst. Generaliserte additive modeller (GAM) ble brukt for kontinuerlige responsvariabler for å fange fleksible, ikke-lineære trender langs miljøgradienter. I alle GAM-er ble forklaringsvariabler inkludert som smooth terms med en basisdimensjon på k = 3, noe som gir tilstrekkelig fleksibilitet til å fange brede trender samtidig som overtilpasning til småskala støy unngås. Prediktorvariabler (avstand fra oppdrettsanlegg, dybde og estimert akkumulert sedimentering) ble valgt basert på deres økologiske relevans. For å unngå problemer med multikollinearitet ble både korrelasjonsmatriser og Variance Inflation Factor (VIF) vurdert; alle prediktorer hadde lave VIF-verdier (< 2), og korrelasjonene var moderate, noe som indikerer at de trygt kunne inkluderes i modellene. Analysene ble gjennomført i R ved bruk av pakken mgcv (Wood, 2017). Generaliserte lineære modeller (GLM) ble brukt for proporsjons- eller telledata, som lipidinnhold og mikrobiell tilstedeværelse/fravær. Lineære modeller (LM) ble benyttet for å teste sammenhenger mellom mikrobielle taxa og miljøgradienter, da disse modellene gir en enkel og lett tolkbar ramme for sparsomme tilstedeværelse/fravær- eller relativ abundans-data. Multivariate metoder, inkludert hovedkomponentanalyse (PCA) og PERMANOVA, ble brukt for å utforske transkriptomiske data, noe som muliggjør påvisning av mønstre og funksjonelle responser i store, multidimensjonale datasett.
Laboratorieeksperimenter fokuserte på å bestemme toleranser og fysiologiske responser hos voksne individer og larver av målarter for stressfaktorer knyttet til akvakultur under kontrollerte forhold. Voksne øyekoraller ble eksponert for det vanlige brukte legemidlet emamektin benzoate for å vurdere potensielle subletale effekter på fysiologi og atferd. Larver av øyekorall ble testet med hydrogenperoksid og azametifos, to vanlige brukte avlusningsmidler, for å undersøke effektene på tidlige livsstadier. Valg av arter og kjemikalier ble styrt av kunnskapshull i litteraturen og sannsynligheten for eksponering i naturlige habitater nær oppdrettsanlegg. Disse eksperimentene gjorde det mulig å undersøke fysiologiske og atferdsmessige effekter i et kontrollert miljø, og gir komplementære data til feltobservasjoner, samt mulighet til å identifisere spesifikke toleransegrenser for både voksne og larvestadier av sårbare arter. Et eksponeringsforsøk ble gjennomført på voksne individer av fire korallarter (risengrynkorall, sjøtre, sjøbusk og øyekorall) for å undersøke effekter av ulike nivåer av partikulære organiske utslipp. Resultatene vises som endringer i visuelle kriterier (polyppaktivitet og vevsnedbrytning).
3.1 - Studiesteder
Oppdrettsanlegg i regionene Møre og Romsdal og Nordland ble valgt fordi disse regionene har flere eksisterende oppdrettsanlegg som ligger nær sårbare bunnhabitater som korallrev, korallskog og svampsamfunn. Anleggene i studiet ble valgt med en gjennomgang av alle eksisterende kartleggingsrapporter fra konsulentselskaper i 2023, samt materiale levert av Statsforvalteren og referansegruppen. Egnetheten til hvert mulig oppdrettsanlegg som studieområde ble vurdert basert på avstanden mellom målarter og anlegget, med mål om å velge lokaliteter der alle målarter dekkes gjennom hele anrikningsgradienten – fra merdkanten og ut til over 500 m. Tabell 3 gir en oversikt over avstanden mellom målarten og anlegget mellom 0 og mer enn 500 m for flere kandidatanlegg i Nordland og Møre og Romsdal. Tilstedeværelsen av lignende habitattyper langs hele anrikningsgradienten ble også vurdert som et kriterium for utvelgelse, og detaljert batymetri levert av Kartverket ble brukt i denne vurderingen. Produksjonshistorikken til anleggene ble også tatt i betraktning.
Møre og Romsdal (anlegg identifikasjonsnummer)
Nordland (anlegg identifikasjonsnummer)
Art
Avstand
12871
12872
12870
32197
12244
22335
12988
12239
37717
30156
11320
10518
19098
32397
27856
Øyekorall
< 50 m
✔
✔
50-200 m
✔
✔
✔
✔
✔
200-500 m
✔
✔
✔
✔
✔
✔
✔
✔
> 500 m
✔
✔
✔
✔
✔
✔
Sjøtre
< 50 m
✔
✔
✔
✔
50-200 m
✔
✔
✔
✔
✔
✔
✔
200-500 m
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
> 500 m
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
Risengrynkorall
< 50 m
✔
✔
✔
✔
✔
✔
✔
50-200 m
✔
✔
✔
✔
✔
✔
✔
✔
200-500 m
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
> 500 m
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
Sjøbusk
< 50 m
✔
50-200 m
✔
✔
✔
✔
✔
200-500 m
✔
✔
✔
✔
✔
✔
> 500 m
✔
✔
✔
✔
✔
✔
✔
Kålrabisvamp
< 50 m
✔
✔
✔
50-200 m
✔
✔
✔
✔
200-500 m
✔
✔
✔
✔
✔
✔
> 500 m
✔
✔
✔
✔
✔
✔
✔
Viftesvamp
< 50 m
✔
✔
✔
✔
✔
✔
50-200 m
✔
✔
✔
✔
✔
✔
✔
✔
200-500 m
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
> 500 m
✔
✔
✔
✔
✔
✔
✔
✔
✔
✔
Tabell 3: Utvalgte oppdrettsanlegg i Møre og Romsdal og Nordland, med forekomst av målarter (✔) innen definerte avstandsklasser fra anleggene. Tabellen viser anleggenes identifikasjonsnummer.
Oppdrettsanleggene som ble valgt i Møre og Romsdal-regionen var Setevika Nord, Storvika og Kråknes i Julsundet, og Hogsneset Nord i Bremnesfjorden. Julsundet er et 15 km langt sund som forbinder Norskehavet med Romsdalsfjorden og det har sterke bunnstrømmer og bratte fjordsider som strekker seg ned til dyp på opptil 500 m. Norges geologiske undersøkelse (NGU) har kartlagt sannsynligheten for korallforekomster i Julsundet basert på multistråleekkoloddata (detaljert bunntopografi og bunnreflektivitet) samt observasjoner fra MAREANO i 2012. De identifiserte 100 punkter i «mulig»-klassen som ligger i de bratte fjordsidene. Setevika Nord (33017) ble etablert i 2012 og produsere atlantisk laks og har en kapasitet på 3120 tonn (Tabell 4). Storvika (22335) ble etablert i 2004 og har en kapasitet på 5460 tonn. Ved Setevika Nord og Storvika i Julsundet ble det funnet både korallrev, korallskog, blomkålkorall og svampskog mellom < 50 til > 500 m fra anleggene i kartleggingsundersøkelser utført av konsulentselskaper. Kråknes (12239) ble etablert i 1997 og produserer atlantisk laks med en kapasitet på opptil 3900 tonn (Tabell 4). Øyekorall, sjøtre, risengrynkorall, sjøbusk og viftsvamp ble også kartlagt rundt Kråknes, men ble kun observert etter 200 m avstand fra anlegget (Tabell 3). Legemidlene azametifos, deltametrin, hydrogenperoksid og emamektin har blitt brukt til lusebehandling, i tillegg til rensefisk, i perioden 2015–2020 i Setevika Nord, Kråknes og Storvika. Etter 2020 har mer mekanisk behandling blitt brukt til lusebehandling ved lokaliteter i Julsundet, kombinert med azametifos, emamektin, deltametrin og diflubenzuron.
Området
Anlegg
Etableringsår
MTB
Fôrbruk (kg; 1.1.2015–21.20.2025)
Lusbehandling 2020–2025
Møre og Romsdal
33017 Setevika Nord
2012
3120
18633111
Emamektin og mekanisk behandling
12239 Kråknes
1997
3900
28226637
Emamektin, Diflubensuron, termisk og mekanisk behandling
22335 Storvika
2004
5460
38325264
Deltamethrin og Emamektin
30377 Hogsneset N
2009
3120
17250369
Hydrogenperoksid
Nordland
11320 Oksøy
1999
5400
36234620
Azametifos, Hydrogenperoksid og Emamektin
30156 Anevik
2009
5400
37963504
Ememektin og mekanisk behandling
10518 Horsvågen
1999
2340
12741193
Emamektin og termisk behandling
Tabell 4: Oversikt over oppdrettsanlegg i Møre og Romsdal og Nordland, inkludert etableringsår, maksimal tillatt biomasse (MTB), fôrbruk (2015–2025) og lusbehandlinger (2015–2020 og 2020–2025).
Hogsneset og Hogsneset Nord er i Bremnerfjorden som ligger mellom øyene Averøya (Averøy kommune) og Frei (Kristiansund kommune) i Møre og Romsdal. I likhet med Julsundet er det sterke bunnstrømmer og bratte fjordsider der, men det er grunnere med dybder opptil 240 m. Hogsneset Nord (30377) ble etablert i 2009 og produserer atlantisk laks og har en kapasitet på 3120 tonn (Tabell 4). Legemidlene azametifos, deltametrin, hydrogenperoksid og emamektin har blitt brukt til lusebehandling, i tillegg til rensefisk, i perioden 2015–2020 i Hogsneset. Etter 2020 har medikamentell behandling ikke blitt brukt i Hogsnes, og hydrogenperoksid ble brukt i Hogsneset Nord i 2022. Korallskog (sjøtre, risengrynkorall, sjøbusk) og viftesvamp ble observert fra 50 til > 500 m fra anleggene i kartleggingsundersøkelsene.
I Nordland ble de tre anleggene i Økssundet valgt for studier i prosjektet: Anevik, Oksøy og Horsvågen. Økssundet ligger i kommunene Hamarøy og Steigen i Nordland og er karakterisert med bratte fjordsider, et kompleks terreng og et maksimum dyp på 630 m. Oksøy (11320) og Anevik (30156) ligger på østsiden av Lundøya i Økssundet og ble etablert i 1999 og 2009. Begge produserer atlantisk laks og har en kapasitet på 5400 tonn (Tabell 4). Horsvågen (10518) ble etablert i 1999 og har en kapasitet på 2340 tonn. Mellom 2015 og 2020 ble azametifos og emamektin brukt til lusebehandling i Oksøy og Anevik, i tillegg til bruk av rensefisk i Anevik. I Horsvågen ble emamektin brukt til lusebehandling i samme periode (2015–2020). Etter 2020 har azametifos, emamektin og hydrogenperoksid blitt brukt i Oksøy og Anevik, mens det i Horsvågen har blitt benyttet emamektin samt mekanisk og termisk behandling mot lus. For alle de tre lokalitetene i Økssundet ble det gjennomført visuelle undersøkelser av koraller og svamper innenfor en radius på 1 km rundt anleggene, i henhold til krav fra Statsforvalteren i Nordland. Disse ROV-undersøkelsene påviste korallrev, korallskog og svampskog i avstander mellom < 50 og > 500 m fra lokalitetene (Tabell 3).
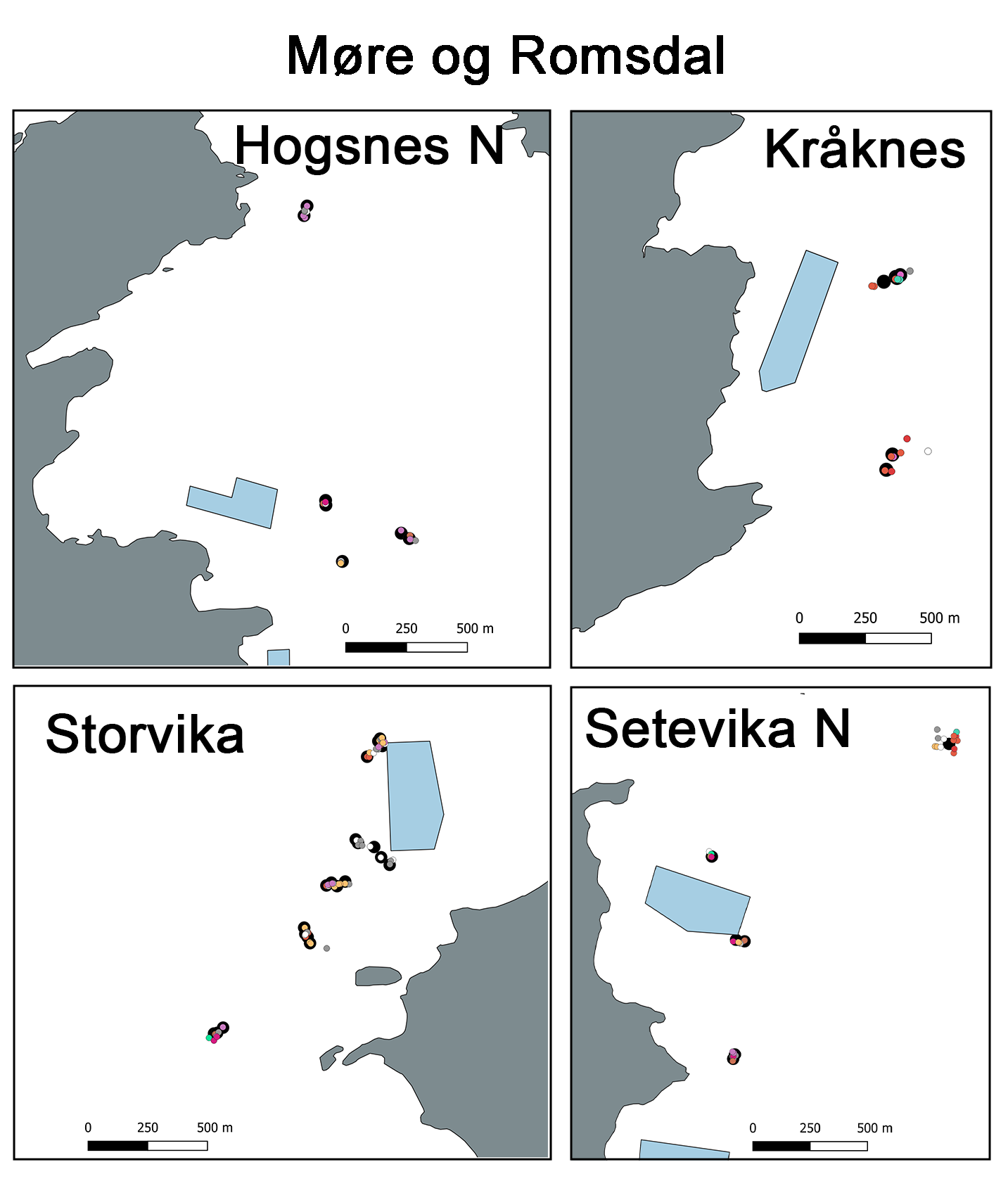
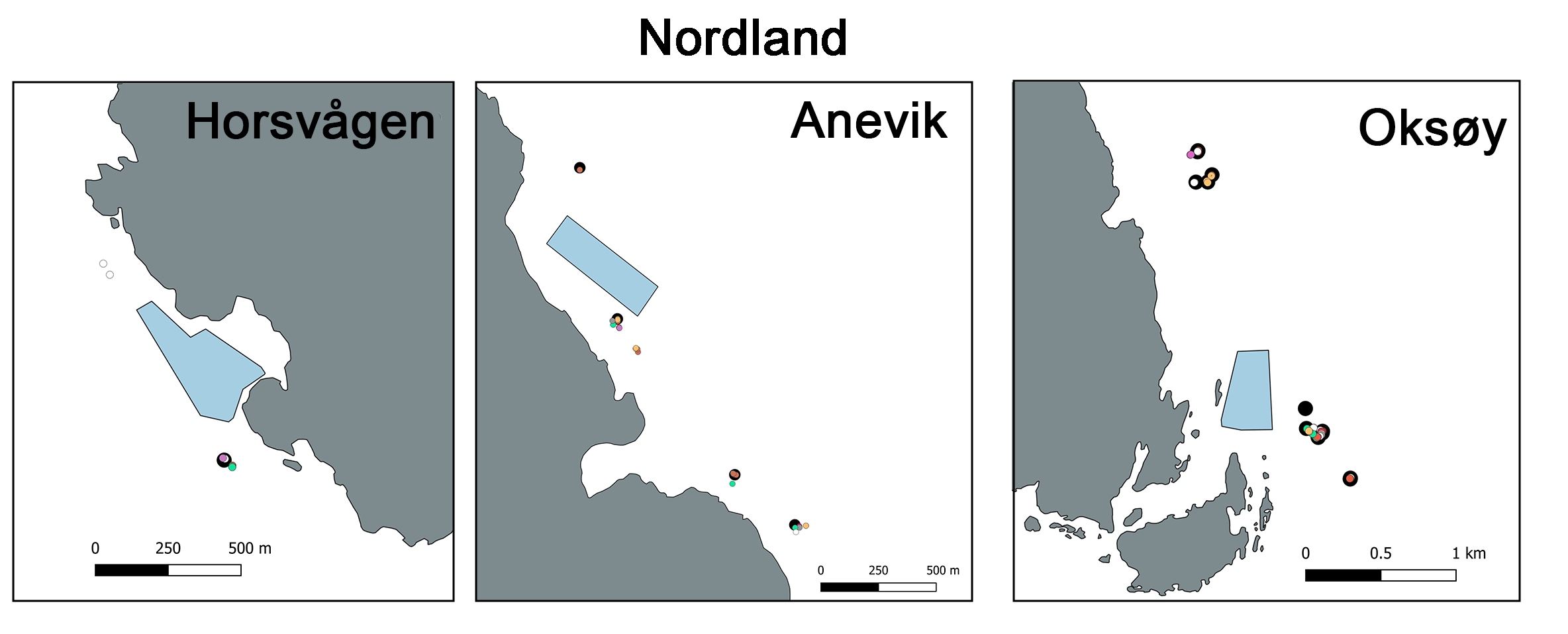
Figur 1: Kart over studieoppdrettsanlegg i Nordland, Julsundet og Bremnesfjorden og ROV transketer
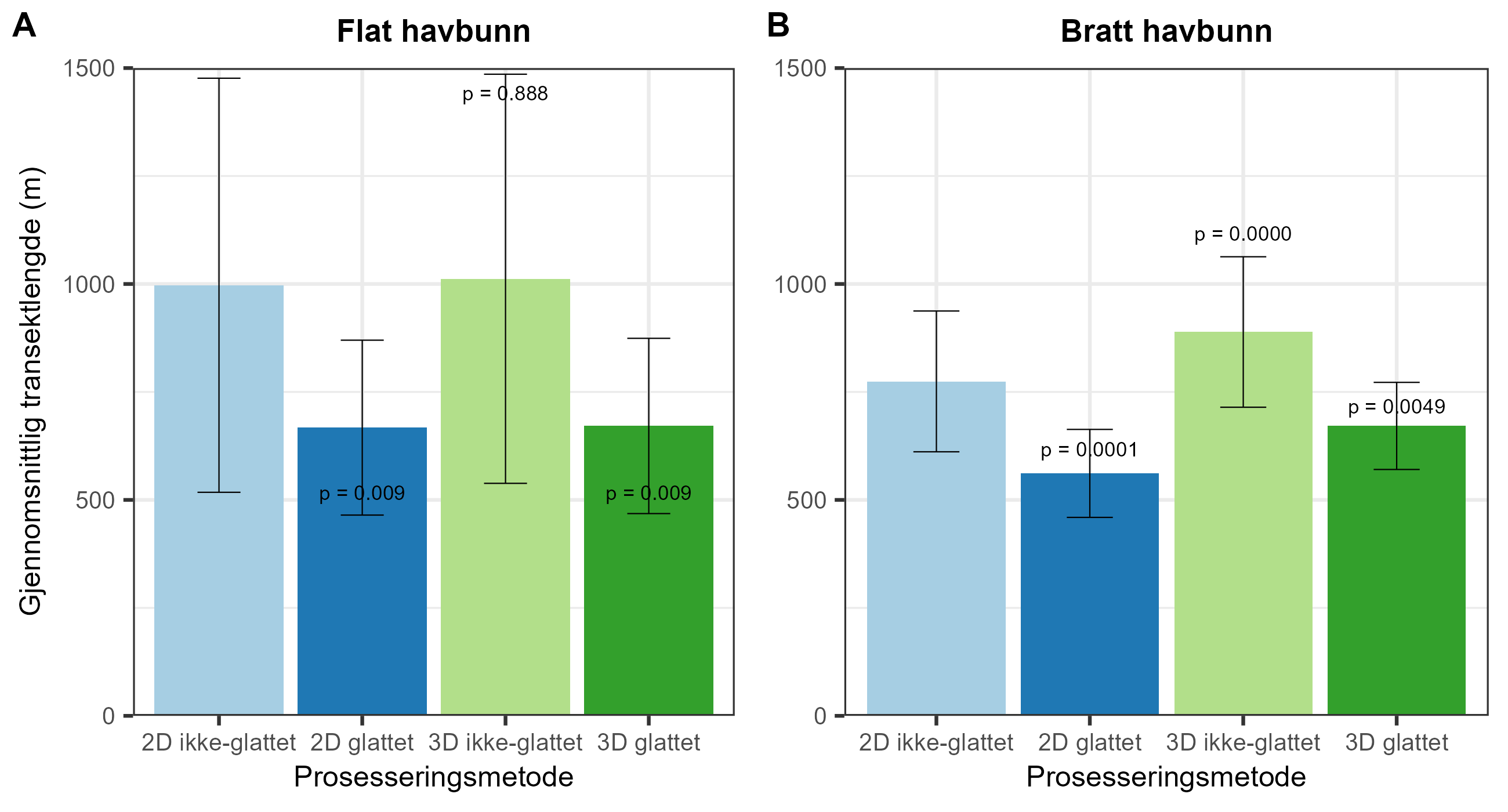
Studiedesignet hadde som mål å oppnå sammenlignbare innsamlingspunkter i sonen påvirket av utslipp, fra så nært oppdrettsmerden som mulig og ut til en avstand som overstiger 1 km, som fungerte som referansestasjon. ROV-dykkeposisjonene langs denne anrikelsesgradienten ble plassert på sammenlignbart terreng, dybder og innenfor hovedretningen til strømmen, så langt det lot seg gjøre logistisk. Totalt ble det gjennomført 46 ROV-dykk fordelt på fire forskningstokt i 2023, 2024 og 2025 rundt de sju anleggene som ble undersøk. Det ble gjennomført 25 dykk i Julsundet og Bremnesfjorden i Møre og Romsdal-området, mens 21 dykk ble gjennomført i Økssundet, Nordland. Faunainnsamlingsdykkene rundt oppdrettsanleggene i Julsundet var på dyp mellom 107 og 290 m, og i Bremnesfjorden mellom 96 og 196 m, mens dykkene rundt anleggene i Økssundet var dypere og dekket et bredere spekter av dybder (116 til 462 m). En oversikt over ROV-dykkeposisjonene finnes på kartet i figur 1, og fullstendige detaljer er gitt i Vedlegg A1. 11 dykk ble gjennomført innenfor 250 m fra oppdrettsanleggene i Møre og Romsdal, 8 ROV-dykk mellom 250 og 750 m fra anleggene og 6 ROV-dykk på avstander større enn 750 m fra anleggene i Møre og Romsdal. I Nordland ble 8 ROV-dykk gjennomført innenfor 250 m fra anleggene, 3 mellom 250 og 750 m fra anleggene og til slutt ble det 10 ROV-dykk mer enn 750 m fra anleggene i Nordland. En oversikt finnes i Tabell 5 og i Vedlegg A1.
Området
Avstand
ROV-dykk
Bergskjell
Viftesvamp
Kålrabisvamp
Øyekorall
Sjøbusk
Sjøtre
Risengrynkorall
Total
Møre og Romsdal
< 250 m
11
8
14
10
3
5
11
13
64
Møre og Romsdal
250 - 750
8
14
13
11
0
3
15
16
72
Møre og Romsdal
> 750
6
3
6
7
9
6
5
10
46
Total
25
25
33
28
12
14
31
39
182
Nordland
< 250 m
8
11
5
4
4
0
8
13
45
Nordland
250 - 750
3
2
7
2
4
3
5
7
30
Nordland
> 750
10
9
11
4
11
3
9
12
59
Total
21
22
23
10
19
6
22
32
134
Tabell 5: Oversikt over ROV-dykk og antall individer av utvalgte bunndyr og koraller som ble prøvetatt ved ulike avstander fra oppdrettsanlegg i Møre og Romsdal og Nordland.
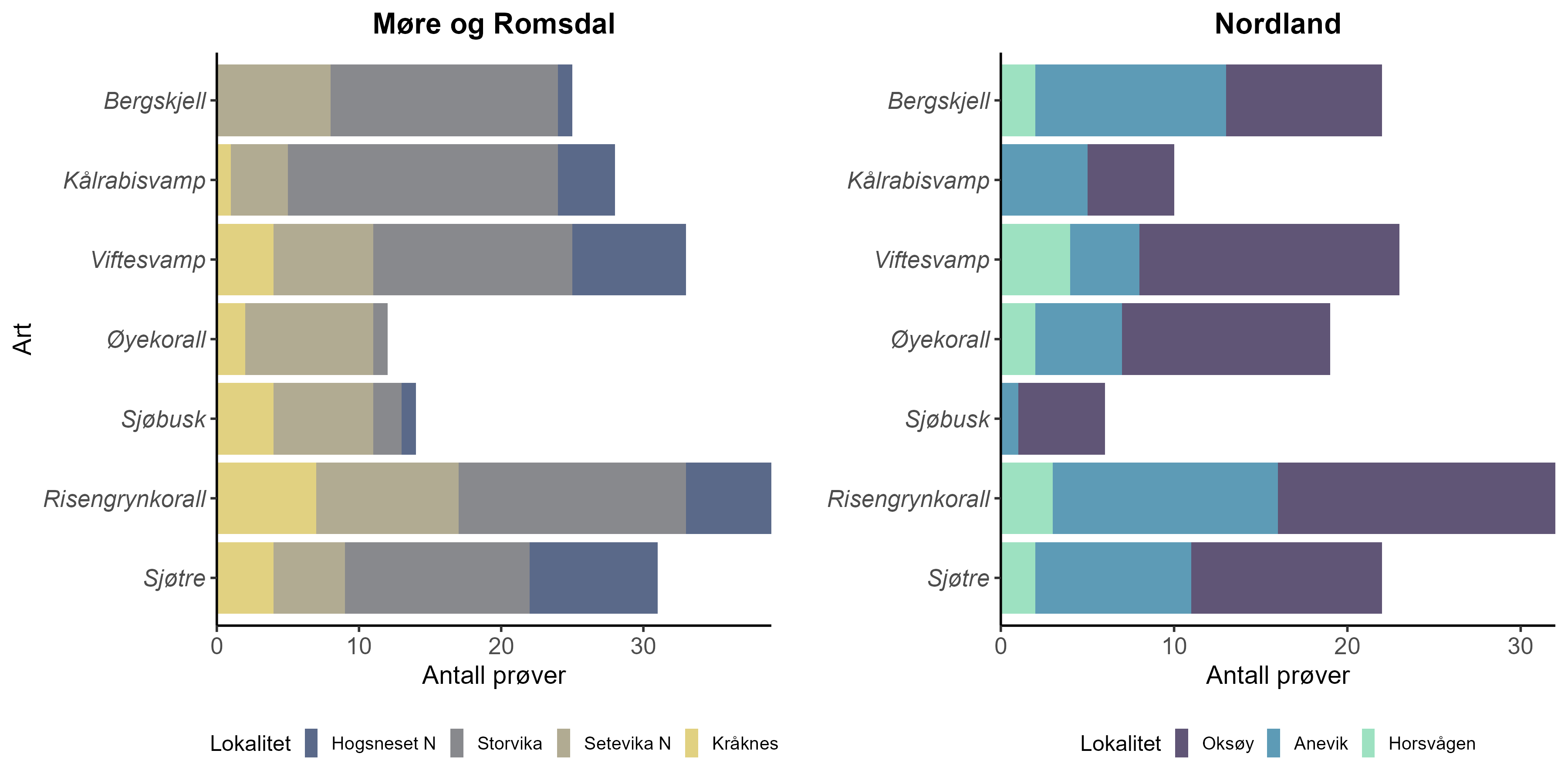
3.2 - Spredning og sedimentering av partikulært organisk materiale
3.2.1 - Modellering
Vi har modellert spredning av organisk materiale fra modellanleggene ved hjelp av metoden som er beskrevet i Carvajalino-Fernández mfl. (2020), men uten resuspensjon. Vår modell er basert på ROMS (Regional Ocean Modelling System) (Shchepetkin og McWilliams, 2005) med inngangsdata som i Norkyst (Albretsen mfl., 2011), men med horisontal oppløsning på 160 m. Modellen beregner strømhastigheten rundt anlegget i ulike dyp, time for time, basert på tidevann, vind, topografi, elveavrenning og andre faktorer. Det organiske materialet blir representert ved partikler som slippes fra merdposisjonene på 25 m dyp, med synkehastigheter som i Bannister mfl. (2016). Det slippes 10000 partikler per dag, for hvert anlegg. Partiklene blir vektet i henhold til fôringsintensiteten på utslippstidspunktet. Vi antar at 1 kg utfôret mengde gir 240 g organisk materiale (POM tørrvekt), i henhold til Cubillo mfl. (2016). Modellperioden er 1.9.2021 til 1.11.2024. Akkumulert sedimentering av organisk materiale i denne perioden er vist i Figur 2. Modellering av sedimentasjonsrate i ytterkanten av nedslagsområdet er en krevende øvelse, og resultatet er sensitivt for en rekke usikre faktorer. I vår modell anslår vi at forenklet batymetri er den største usikkerhetsfaktoren. Havmodeller som ROMS kan ikke kjøre stabilt på batymetri med bratte skrenter, og batymetrien blir derfor glattet med et Laplace-filter først. Dermed forsvinner lokale topografiske terrengformer som skrenter, lokale søkk, små undersjøiske klipper og andre faktorer som kan påvirke den lokale fordelingen av sedimentert organisk materiale. Det er i prinsippet mulig å unngå problemet ved å øke den horisontale oppløsningen i modellen, noe som reduserer behovet for glatting. Dette vil imidlertid kreve vesentlig mer regnekraft, og muligens kreve at modellen inkluderer en høyoppløst beskrivelse av fysikken (bedre turbulensparametrisering, ikke-hydrostatiske ligninger etc.). Bratte skrenter vil også kunne gi behov for å inkludere en validert resuspensjonsmodell for partikkelspredningen, selv på dypere vann. Merdene sin påvirkning på sirkulasjonsmønsteret vil også kunne ha betydning i en høyoppløst modell. En annen usikkerhetsfaktor i vår modell er utfôringsraten. Vi har brukt data fra Fiskeridirektoratet, som registrerer utfôret mengde per måned. Denne informasjonen finnes imidlertid bare på anleggsnivå, ikke på merdnivå. Det har vist seg utfordrende å skaffe til veie mer detaljerte fôringsdata for alle modellanlegg i hele modellperioden. Vi har derfor valgt å spre utfôret mengde likt på alle merder i anlegget, også der vi har bedre informasjon i deler av perioden. På denne måten blir modelleringen konsistent for alle anlegg og lettere å sammenligne. En tredje usikkerhetsfaktor er synkehastigheten. Bannister mfl. (2016) gir synkehastighetsprofil for både små og store partikler, men det er knyttet usikkerhet til om metoden som er benyttet gir sikker informasjon om de minste partiklene, som dominerer i ytterkanten av nedslagsområdet. Det er behov for å gjenta forsøkene med tanke på de minste partiklene for å avgjøre dette.
Figur 2: Nedslagsområde for sedimentering for hvert av de 7 modell-anleggene. Fargen viser sedimentert mengde i løpet av perioden 1.9.2021 til 1.11.2024, fra 0,1 kg·m⁻² (gul) til 10 kg·m⁻² (mørk blå). Cellene i modellgitteret er 80 m × 80 m. Røde ringer viser merder. Svarte kryss viser prøvetakningsposisjoner.
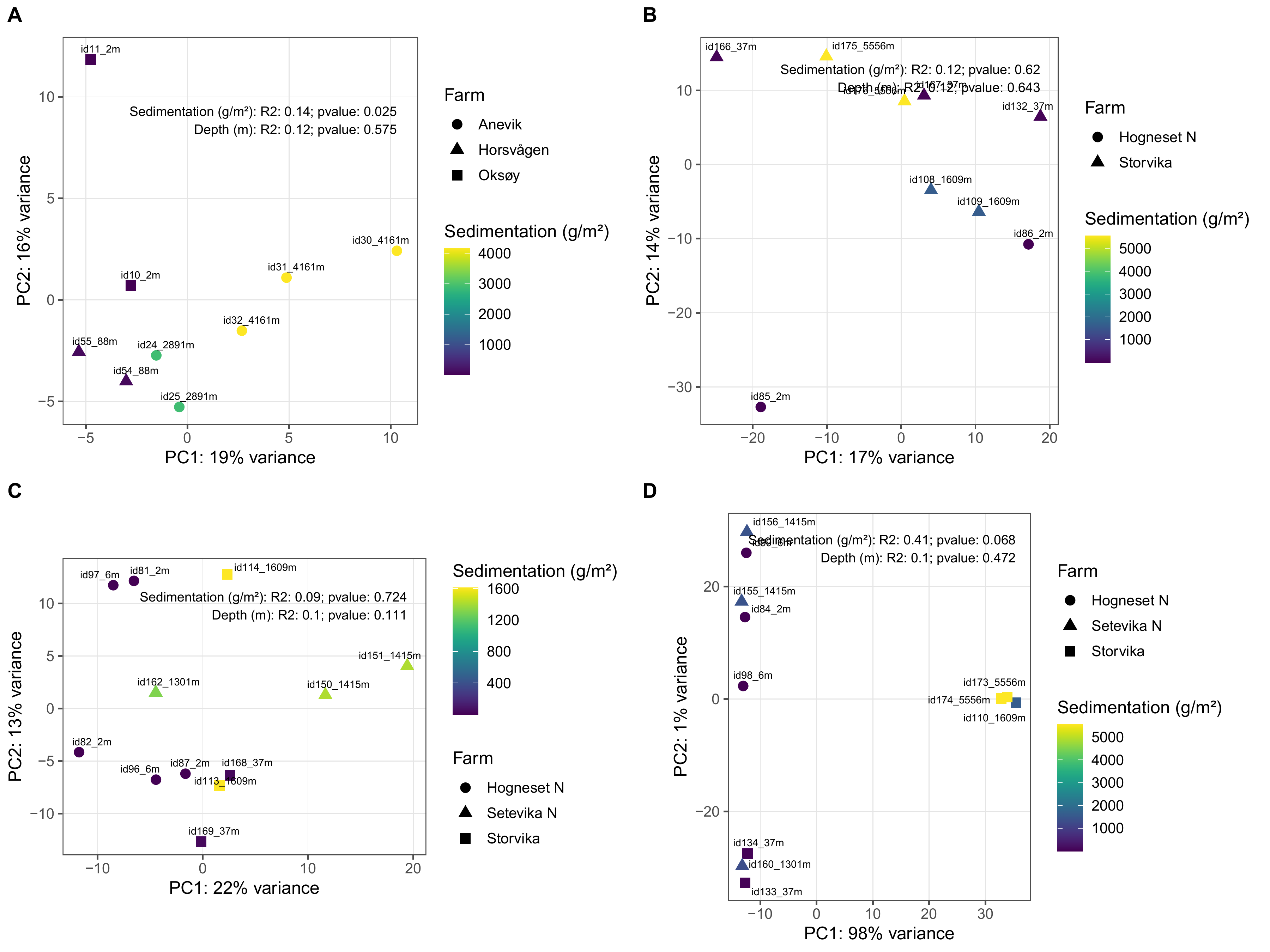
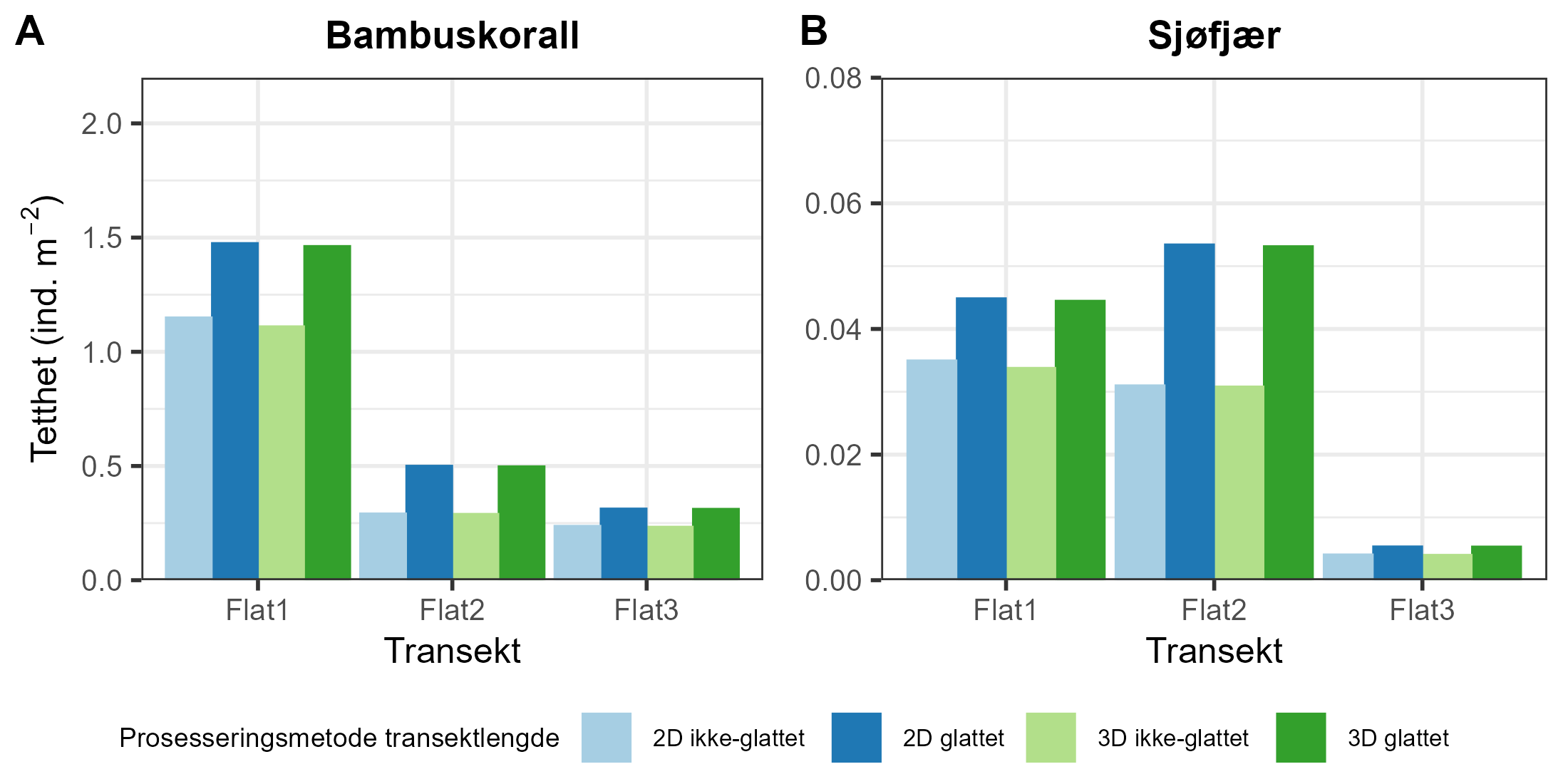
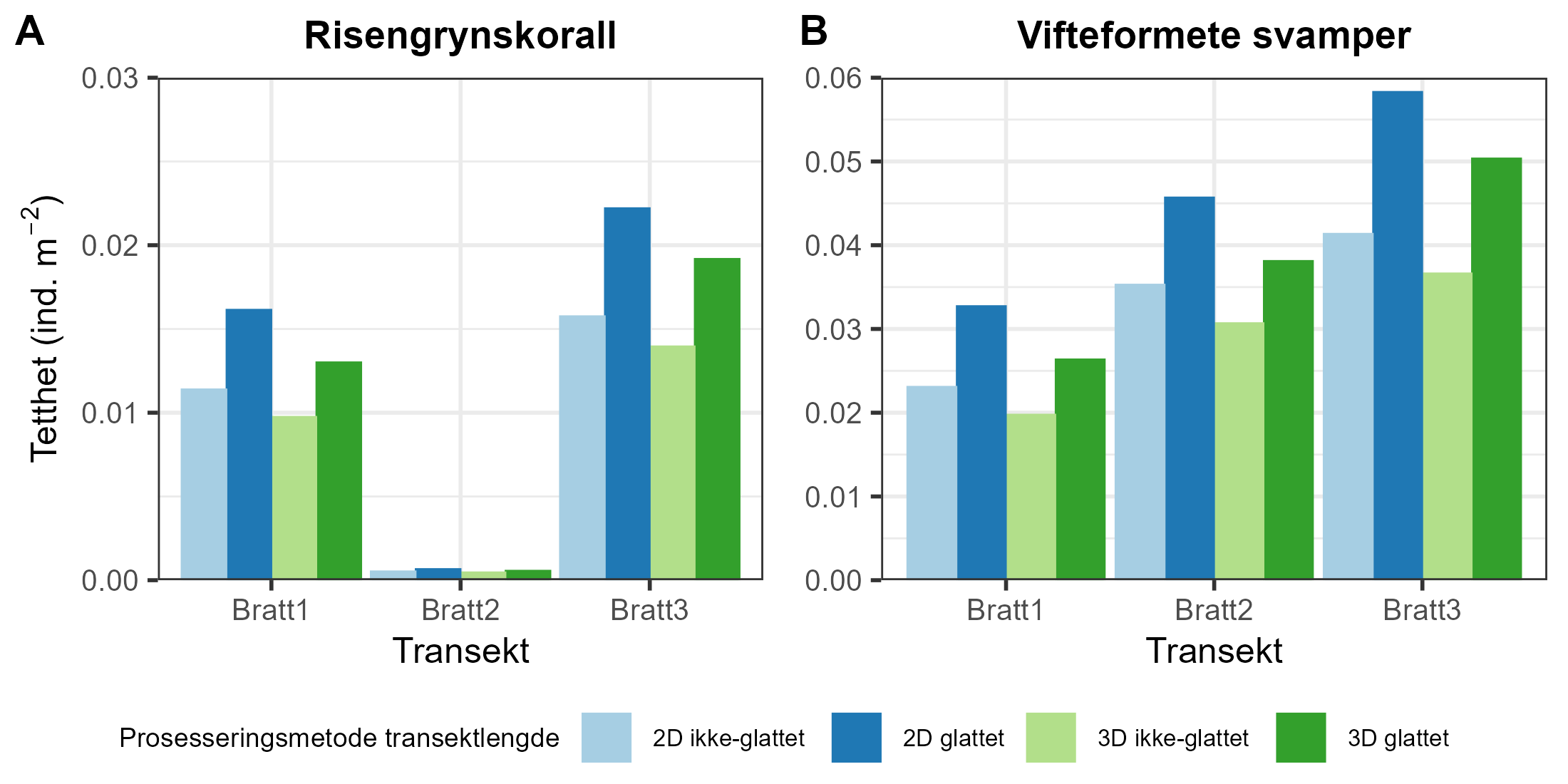
Modellerte akkumulerte sedimentasjonsverdier var betydelig høyere nær merdene ved anleggene i både Nordland (F = 175, p < 0,0001) og Møre og Romsdal (F = 0,7, p < 0,0001) (Figur 3).
Akkumulert sedimentasjon av organisk materiale over to år ved alle anleggene var høyest innenfor 200 m fra anlegget. Ved Anevik i Nordland var verdiene innenfor 150 m over 2 kg·m⁻² og nådde opptil 4 kg·m⁻² for to ROV-dykkposisjoner mindre enn 100 m fra anlegget. Mellom 200 m og 1 km fra Anevik var den akkumulerte sedimentasjonen rundt 200 g·m⁻² og under 100 g·m⁻² mer enn 1 km fra anlegget. Ved Oksøy ble de nærmeste prøvene tatt 300 m fra anlegget, og sedimentasjonen var mellom 163 og 110 g·m⁻² i området mellom 300 og 400 m fra anlegget. Sedimentasjonen fra partikulært organisk utslipp avtok til 33 g·m⁻² 600 m fra anlegget og deretter til 2 g·m⁻² eller mindre mer enn 1 km fra anlegget. Det ble gjennomført to ROV-dykk rundt Horsvågen som samlet inn fauna mellom 125 og 155 m på hver side av anlegget, hvor mellom 367 og 904 g·m⁻² partikulært utslipp hadde akkumulert over en toårsperiode. Lokaliteter ble inkludert som en tilfeldig effekt i den generaliserte additive modellen (GAM) og for anleggene i Nordland gav den tilfeldige effekten av lokaliteter ingen signifikant effekt (F = 251, p = 0,998), noe som indikerer at sedimentasjonsmønstrene i forhold til avstand fra anlegget var mer like mellom de tre anleggene i denne regionen.
Lokaliteter som en tilfeldig effekt i den generaliserte additive modellen (GAM) hadde en signifikant effekt (F = 1,2, p = 0,0407) på sedimentasjonsnivåene for anleggene i Møre og Romsdal, noe som indikerer høy variasjon i sedimentasjonsmønstre mellom disse anleggene.
Utslippsfotavtrykket fra Storvika strakte seg over 1 km fra anlegget, og fauna ble samlet inn gjennom seks ROV-dykk innenfor 150 m fra anlegget, der akkumulert sedimentasjon over en toårsperiode var mellom 1 og 5 kg·m⁻². Mellom omtrent 200 og 500 m fra anlegget var akkumulert sedimentasjon mellom 278 og 124 g·m⁻², og 36 g·m⁻² ved ROV-dykkposisjoner 1 km fra anlegget. Akkumulert sedimentasjon ved Setevika N var også 1,3–1,4 kg·m⁻² innenfor 150 m fra anlegget, 400 g·m⁻² ved ROV-posisjoner rundt 400 m fra anlegget, og mellom 1 og 2,5 g·m⁻² 1–1,5 km fra anlegget.
Akkumulert sedimentasjon var spesielt lav ved ROV-dykkposisjonene ved Hogsneset N og Kråknes, fordi de lå utenfor hovedretningen for sedimentasjon. Ved Hogsneset N var akkumulert sedimentasjon 6 g·m⁻² i området 200–300 m fra anlegget og under 2 g·m⁻² mer enn 500 m fra anlegget. Ved Kråknes var akkumulert sedimentasjon mellom 7,6 og 4,9 g·m⁻² i området mellom 150 og 600 m fra anlegget.
Figur 3: Akkumulert sedimentasjon (g·m⁻²) over to år før ROV-dykking for fauna-prøvetaking, mot avstand til nærmeste anlegg (m) ved for åtte lokaliteter i Møre og Romsdal og Nordland. Linjen som beskriver sammenhengen, er basert på en lokal regresjon (LOESS).
3.2.2 - SIBS-enhet og prøvetakingsprosedyre
En SIBS (Substrate Independent Benthic Sampler) (Keeley mfl., 2025) ble modifisert for bruk i dyphavet (< 500 m) på en ROV for å kunne samle sedimentflak fra havbunnen på valgte punkter av interesse (f.eks. ved siden av spesifikke fauna eller grupper av fauna). Dette involverte nyutforming av enkelte komponenter og utvikling av en dypvannskontrollboks som konverterer det overflate-initierte COMS-signalet til å utløse frigjøringsmekanismen i den aktuelle prøvetakeren. Tre SIBS-samplere ble montert på et spesialbygd stativ der de kunne nås av en syvfunksjonsmanipulator, visuelt støttet av ROV-kamera og lys.
For å ta en prøve ble en prøvetaker fjernet fra stativet og forsiktig plassert på ønsket sted, hvor den ble utløst ved kontakt med bunnen. Et område ble ansett som egnet for prøvetaking når ROV-en befant seg på målhabitatet, omtrent i ønsket avstand fra oppdrettsanlegget, og observatøren hadde identifisert minst to relevante faunaprøver innenfor en radius på cirka 10 m.
Under de første dykkene på toktet i 2023 var denne prosedyren bare delvis vellykket. Videre utvikling av prøvetakeren ble imidlertid gjennomført, og metoden viste seg å være særlig vellykket under toktet i 2024.
På prøvetakingsstedet ble 500 mL med overfladiske flak og nærbunnsvann trukket inn i en steril sprøyte. Deretter ble prøvetakeren plassert tilbake i stativet. Ved retur til overflaten ble sprøytene forseglet, fjernet fra prøvetakeren og oppbevart kjølig frem til videre prosessering etter metodene beskrevet i Keeley mfl. (2025). Fordi prøvene naturlig varierer i sedimentkonsentrasjon, avhengig av bunntype, ble prøver uten synlig sediment filtrert gjennom ett filter (47 mm Whatman GFF-filtre, Sigma-Aldrich) for å øke sjansen for å kunne ekstrahere tilstrekkelig DNA, mens prøver med høyere turbiditet (mye sediment) ble delt opp i 2–3 filtre for å lage tekniske replikater. De resulterende prøvene (fanget på 47 mm Whatman GFF-filtre) ble lagret ved −20 °C under toktet, og deretter ved −80 °C frem til videre prosessering. Prøver med færre enn 10 bioindikator-ASV-er, som representerer antall ASV-er i hver prøve inkludert i bioindikator-databasen, basert på tilstedeværelse/fravær fremfor antall reads, og som kan tilhøre samme eller flere økologiske grupper, får ikke tildelt en b-MBI. Disse prøvene får i stedet verdien NA, og blir dermed ekskludert fra videre analyser.
DNA-ekstraksjon og 16S-sekvensering
Totalt DNA ble ekstrahert fra halve filteret med sediment ved bruk av DNeasy PowerSoil-settet (Qiagen, California, USA), i henhold til produsentens instruksjoner. Prøvene ble deretter konservert i 1 % PVA-løsning med 200 nM Tris-HCl, pH 8. Før sending til Cawthron Institute (Nelson, New Zealand) ble 5 µL PVA-løsning og 20 µL eluert DNA (4 ng·µL⁻¹) tilsatt per brønn i 96-brønns plate, platen tørket over natten under laminær strømning ved romtemperatur og deretter forseglet. Det påfølgende molekylære arbeidet ved instituttet benyttet enveis, kontaminasjonskontrollerte arbeidsflyter. Polymerasekjedereaksjon (PCR) oppsett og tilsetting av mal ble utført i separate sterile laboratorier utstyrt med laminær luftstrømsbenk skap (HEPA-filtrering) og romomfattende ultrafiolett sterilisering, aktivert i > 15 min før og etter bruk. Aerosolresistente pipettespisse ble brukt gjennomgående, og negative (uten mal) PCR-kontroller ble inkludert med hver kjøring. Pre-ekstrahert eDNA fra alle prøver ble amplifisert for bakteriesamfunn ved bruk av 16S rRNA V3–V4-regionen med primerne 341F (5′-CCT ACG GGN GGC WGC AG-3′) og 805R (5′-GAC TAC HVG GGT ATC TAA TCC-3′); primerne bar Illumina overhang-adaptere i henhold til Illumina 16S bibliotekforberedelsesprotokoll. PCR-reaksjoner ble kjørt på en Eppendorf Mastercycler (Eppendorf, Hamburg, Tyskland) med MyFi™ PCR Master Mix (Bioline Meridian Bioscience, Tennessee, USA) i et totalt volum på 50 µL, med 2 µL av hver primer (10 µM lager) og 2 µL mal. Syklusen for 16S bestod av 95 °C i 5 min, etterfulgt av 35 sykluser med 94 °C i 30 sekunder, 54 °C i 30 sekunder og 72 °C i 45 sekunder, og en sluttforlengelse på 72 °C i 7 min. Amplifikonene ble renset og normalisert ved bruk av SequalPrep™ Normalization Plates (Thermo Fisher Scientific, USA), dobbeltindeksert med Nextera XT-adaptere, sammenslått og sekvensert (parvis endesekvensering) hos Sequench Ltd (Nelson, New Zealand) på en Illumina NextSeq™ plattform med V2-kjemi.
Bioinformatikk og dataforbehandling
Sekvenserte data ble demultipleksert og primere fjernet ved bruk av CUTADAPT versjon 2.6 (Martin, 2011). Deretter ble dataene kvalitetsfiltrert, denoiserte, slått sammen som parvise sekvenser og renset for kimærer ved hjelp av DADA2 R-pakken versjon 1.14 (Callahan mfl., 2016). Før kvalitetsfiltreringen ble fremoversekvenser og reverssekvenser trunkert ved henholdsvis 226 og 220 bp på 5′-enden for å fjerne lavkvalitetsregioner. Både kvalitetsfiltrering og denoising ble utført med standardparametrene i DADA2. Forward- og reverse-sekvenser ble slått sammen ved bruk av standardinnstillingene, med et perfekt minimumsoverlapp på 10 bp. Kimæresekvenser ble fjernet med den innebygde «consensus»-metoden i DADA2, og amplikonsekvensvarianter (ASVs) ble taksonomisk tilordnet ved bruk av SILVA-databasen versjon 138 (Quast mfl., 2012) og RDP Naïve Bayesian Classifier-algoritmen (Wang mfl., 2007). Potensiell kontaminasjon i negative kontroller ble undersøkt og fjernet ved bruk av microDecon R-pakken versjon 1.0.2 (McKnight mfl., 2019), med standardinnstillinger. Sekvenseringsdybden per prøve ble visualisert med rarefaksjonskurver ved hjelp av vegan R-pakken versjon 2.5.7 (Oksanen mfl., 2020), og prøver med ≤ 1800 sekvenser ble ekskludert fra videre analyser.
En b-MBI – en indikator for organisk anrikelsesgradient basert på andelen av nøkkelindikator-taxa (sekvenser) som samsvarer med en database utviklet ved Cawthron Institute (Nelson, New Zealand) – ble beregnet for hver prøve som beskrevet i Pochon mfl. (2019, 2021). b-MBI er en kontinuerlig variabel på en skala fra 1 til 7, der 1 tilsvarer praktisk talt «uberørte» bunnforhold uten organisk enrikelse, 5 markerer toppnivå av makrofaunatetthet under svært høyt anrikede forhold, og 7 representerer ekstremt organisk anrikede og anaerobe forhold som ikke kan opprettholde liv av bentiske makrofauna. Prøver med færre enn 10 bioindikator-ASV-er, som representerer antall ASV-er i hver prøve inkludert i bioindikator-databasen, basert på tilstedeværelse/fravær fremfor antall reads, og som kan tilhøre samme eller flere økologiske grupper, får ikke tildelt en b-MBI-verdi. Disse prøvene får i stedet verdien NA, og blir dermed ekskludert fra videre analyser.
b-MBI-analyser
For SIBS-prøver med tekniske replikater ble variasjonen i b-MBI vurdert. Replikater med et standardavvik større enn 0,5 ble redusert til sekvensen med høyest antall ikke-kimære reads, mens replikater med lavere variasjon ble tatt som gjennomsnitt. Generaliserte additive modeller (GAM) ved hjelp av pakken mgcv (Wood, 2017) i R versjon (4.4.3) ble brukt for å undersøke effekten av avstand til anlegg, vanndybde og akkumulert sedimentasjon på b-MBI for SIBS-prøver. Modellene inkluderte smooth terms funksjoner for hver prediktor per fylke, med Gamma-fordeling og log-link. Modellens diagnostikk ble vurdert, og forklart devians samt p-verdier ble annotert på figurene.
Modellen forklarte 70,2 % av deviansen (justert R² = 0,688) (Figur 4). Det var signifikante forskjeller i baseline b-MBI mellom fylkene, der Nordland hadde høyere estimert b-MBI enn Møre og Romsdal (Møre og Romsdal: 0,85 ± 0,05, p < 0,001; Nordland: 2,20 ± 0,66, p = 0,002).
Avstand til anlegg hadde ingen signifikant effekt på b-MBI i noen av fylkene (Møre og Romsdal: F = 0,49, p = 0,488; Nordland: F = 0,02, p = 0,886). Dybde hadde derimot en signifikant effekt i begge fylker (Møre og Romsdal: F = 5,09, p = 0,031; Nordland: F = 6,68, p = 0,015). Akkumulert sedimentasjon hadde en sterk og signifikant effekt på b-MBI i Møre og Romsdal (F = 10,51, p = 0,003), mens denne effekten ikke var signifikant i Nordland (F = 1,14, p = 0,296).
Figur 4: b-MBI-verdier for SIBS-prøver i forhold til avstand til oppdrettsanlegg (A), dybde (B) og akkumulert sedimentasjon (C). Generaliserte additive modeller (GAM) ble tilpasset separat for hvert fylke med Gamma-fordeling og log-link. Punktene viser observerte b-MBI-verdier, linjene viser modellprediksjoner, og skyggelagte områder angir 95 % konfidensintervall. Forklart devians og p-verdier for smooth terms er annotert på figurene.
Disse resultatene indikerer at lokale miljøfaktorer, spesielt dybde og sedimentasjon, spiller en viktigere rolle enn avstand til anlegg for å påvirke b-MBI, med noe regional variasjon mellom de to fylkene.
3.3 - Studiearter - antall prøver og prøvetaking
Målartene som ble samlet inn, ble valgt på bakgrunn av at disse artene anses som sårbare for menneskelig påvirkning fra økt sedimentering eller kjemisk utslipp, i henhold til nasjonale og internasjonale retningslinjer. Det ble også tatt hensyn til eksisterende kunnskap om artenes sensitivitet overfor utslipp fra akvakultur, samt om artene var observert i tilstrekkelig antall under kartlegging av oppdrettsområder til å kunne studere endringer i deres fysiologiske og visuelle helsetilstand.
De forvaltningsrelevante bunnartene som ble valgt som fokus i studien var de tre hornkorallartene sjøtre (Paragorgia arborea), sjøbusk (Paramuricea placomus) og risengrynkorall (Primnoa resedaeformis), samt den revbyggende steinkorallen øyekorall (Desmophyllum pertusum). Svampeartene som var i fokus for studien var viftesvamp av slekten Phakellia sp. og kålrabisvamp (Geodia barretti). Studien inkluderte også muslingen bergskjell (Acesta excavata) (Figur 5).
De utvalgte artene regnes som nøkkelarter i forvaltningsrelevante marine naturenheter som hardbunnskorallskog (risengrynkorall, sjøtre, sjøbusk), svampsamfunn (kålrabisvamp og viftesvamp) og korallrev (øyekorall) (Bekkby mfl., 2021). De fire utvalgte korallartene er også oppført som viktige arter i de rødlistede naturtypene kysttilknyttet korallrev, noe beskyttet dyp hornkorallskog på fastbunn i atlanterhavsvann og eksponert dyp hornkorallskog på fastbunn i kystvann (Artsdatabanken, 2025).
Figur 5: Bilder av de sju målartene fra prøvetaking i VDWS-Transition-prosjektet.
Prøvetaking
Under toktet i 2023 ble prøvene samlet inn ved hjelp av ROV-en Minerva, en arbeidsklasse-ROV utviklet av Sperre AS for NTNU AUR-Lab. ROV-en opererer i «free flying»-modus, koblet direkte til fartøyet med en 900 m lang kabel, og er utstyrt med sonar, stereokamera, HD- og 4K-videokamera, samt en sju-funksjons manipulatorarm (7F Orion Extended) som ble brukt til prøvetaking. I 2024 og 2025 ble prøver samlet inn ved hjelp av Havforskningsinstituttets ROV Freyja (Argus Remote Systems, modell Mariner 141). Visuelle observasjoner ble gjort ved hjelp av et Sony FCB-EV7520A-kamera (uten direktestrøm) montert i omtrent 60° vinkel, som ble bekreftet ved start av hver stasjon og holdt konstant under transektene. To lasere med 10 cm avstand ble brukt for skalering av objekter. For begge ROV-er ble posisjonering utført ved bruk av HIPAP-systemer (HiPAP 502 for Minerva og µpap 200 med Cnode mini S-transponder for Freyja). Høyden over havbunnen ble overvåket ved hjelp av altimeter, logget i SFO, og opprettholdt på omtrent 1 m under prøvetaking.
Målarter ble lokalisert basert på tidligere kartleggingsrapporter og høyoppløselige batymetrikart rundt forsøksområdene. Når eksemplarer av målartene ble funnet, ble posisjonen registrert ved hjelp av HI Seabed Field Observer SFO-systemet, og videomateriale ble tatt før prøvetaking for å vurdere visuelle helsekriterier. Prøver av målartene ble samlet inn ved hjelp av ROV-ens gripeverktøy (Figur 6). Korallfragmenter ble klippet fra hovedkolonien ved hjelp av ROV-manipulatorarmen for risengrynkorall, sjøtre, sjøbusk og øyekorall. For små korallkolonier ble hele individet samlet inn. For svamper (viftesvamp og kålrabisvamp) ble hele individet samlet inn med manipulatorarmen, mens individer av bergskjell ble samlet inn ved hjelp av en spesialdesignet skje som minimaliserte skade på skall og vev. Posisjon, tidspunkt for prøvetaking og hvilken del av ROV-kurven prøven ble plassert i, ble registrert ved bruk av IMR SFOs sanntids videomerkingssystem.
Når ROV-kurven var full, ble kjøretøyet hevet om bord på forskningsfartøyet. Prøvene ble umiddelbart overført til bøtter eller tanker fylt med avkjølt bunnvann, som tidligere samme dag hadde blitt hentet omtrent 1 km fra forsøksområdet ved hjelp av en CTD-rosett. Temperaturen ble opprettholdt ved hjelp av kjøleenheter. Prøvene ble deretter bearbeidet så raskt som mulig i laboratoriet om bord for de ulike analytiske metodene.
Figur 6: Bilder av prøvetaking med ROV-manipulatorarmen: A) klipping av en sjøtre-korallfragment, B) innsamling av en viftesvamp, og C) innsamling av et bergskjell med den spesialtilpassede skuffen.
Prøvetakingsinnsats
I løpet av de 46 ROV-dykkene i Møre og Romsdal og Nordland ble det samlet inn totalt 309 fysiske prøver ved hjelp av ROV (Figur 6). Avstanden fra prøvetakingsposisjonene til oppdrettsanleggene i Møre og Romsdal kunne deles inn i tre kategorier: mindre enn 250 m, 250–750 m, og mer enn 750 m fra anlegget. I Økssundet var det mer utfordrende å undersøke områder nært merdene på grunn av komplekse fortøyningssystemer forankret i bratte fjordvegger og den nærmeste prøven som ble tatt fra oppdrettsanlegget, var omtrent 65 m fra anlegget.
Figur 7: Prøver samlet inn per art ved hvert oppdrettsanlegg, fordelt etter fylke. Y-aksen viser artene, X-aksen viser totalt antall prøver, og fargene representerer de ulike oppdrettsanleggene.
Totalt ble det samlet inn 182 prøver gjennom 25 dykk i Møre og Romsdal. Det ble samlet inn 39 risengrynkorall, 33 viftesvamp, 31 sjøtre, 28 kålrabisvamp og 25 bergskjell. Færre prøver samlet inn av øyekorall (12) og sjøbusk (14), ettersom disse artene ble observert sjeldnere (Figur 7). Innenfor 250 m fra oppdrettsanleggene i Møre og Romsdal ble det samlet inn 64 faunaprøver på denne nærmeste avstanden til merdene. 72 faunaprøver ble samlet inn mellom 250 og 750 m fra anleggene i Møre og Romsdal, etterfulgt av ytterligere 46 prøver som ble tatt mer enn 750 m fra anleggene (Vedlegg A1, Vedlegg A2).
I Økssundet, Nordland, ble det samlet inn 134 prøver fordelt på 21 ROV-dykk. Det ble samlet inn 32 risengrynkorall, 23 viftesvamp, 22 sjøtre og 22 bergskjell. I tillegg ble det samlet inn 19 øyekorall, 10 kålrabisvamp og 6 sjøbusk. I Nordland ble 45 faunaprøver samlet inn innenfor 250 m fra oppdrettsanleggene. Mellom 250 og 750 m fra anleggene i Nordland ble det samlet inn 30 faunaprøver, og til slutt ble 59 prøver samlet inn mer enn 750 m fra anleggene i Nordland (Tabell 5). Fullstendige detaljer over antall prøver fordelt på art og avstandskategori finnes i Vedlegg A1 og Vedlegg A2.
Prosentandelen av prøver per art som ble brukt i de ulike analytiske metodene er vist i Figur 8. Metodene inkluderer visuell helsevurdering, målinger av respirasjon og ammoniakkutskillelse, LMS, lipid- og fettsyreprofiler, molekylære analyser (mRNA-sekvensering, 16S-sekvensering av vev og slim, og DNA-metylering), samt tilleggstester som artsidentifikasjon og eksponeringsforsøk. Prosentandelene er beregnet basert på det totale antallet prøver samlet inn for hver art under prosjektets tokt i perioden 2023–2025. Enkelte metoder, spesielt molekylære og biokjemiske analyser, ble brukt på færre prøver på grunn av tid og kostnader, samt begrenset suksess eller gjennomførbarhet for enkelte arter.
Figur 8: Prosentandel av prøver per art som ble brukt i de ulike analytiske metodene. Prosentandelene er beregnet basert på det totale antallet prøver samlet inn under prosjektets tokt i perioden 2023–2025.
3.4 - Artsidentifikasjon
Korrekt artsidentifikasjon er essensielt i økologiske og fysiologiske studier, særlig for bentiske kaldtvannsorganismer som ofte er morfologisk like eller kryptiske. Riktig identifikasjon sikrer at målte responser, som metabolisme, molekylære markører eller stressindikatorer, faktisk er artsspesifikke, og gjør det mulig å tolke miljøeffekter pålitelig. Videre understøtter det reproduserbarhet, sammenlignbarhet mellom studier og kunnskapsbaserte beslutninger om forvaltning og bevaring i sårbare marine økosystemer.
I lys av dette var det avgjørende å verifisere artsidentiteten til prøvematerialet vårt. Selv om individene morfologisk ble identifisert som viftesvamp (Phakellia ventilabrum), er denne gruppen kjent for betydelig morfologisk variasjon og potensielt kryptiske arter. Det var derfor ikke mulig å være helt sikker basert utelukkende på morfologiske kjennetegn. For å sikre korrekt artsbestemmelse og styrke den vitenskapelige robustheten i studien, ble arten derfor bekreftet ved molekylær identifikasjon.
Metode
Kort fortalt ble vevsprøver bevart ved hurtig frysing i flytende nitrogen og lagret ved -80 °C. Vevsprøvene ble ekstrahert for både DNA og RNA ved av protokollen beskrevet av Gomes mfl. (2025). DNA-ekstraktene ble benyttet til artsidentifikasjon (sendt til Møreforsking), samt for kapitlene 4.10.1 og 4.10.3, mens RNA-ekstraktene ble brukt til kapittel 4.10.2. Partielle sekvenser av de mitokondrielle genene cytochrome c oxidase subunit I (COI; primere: LCO1490 (Fremover) GGTCAACAAATCATAAAGATATTGG, HCO2198 (omvendt) TAAACTTCAGGGTGACCAAAAATCA) (Folmer mfl., 1994) og genmarkøren 28S rRNA C-region (primere: C2 (F) GAAAAGAACTTTGRARAGAGAGT; D2 (R) TCCGTGTTTCAAGACGGG) (Chombard mfl., 1998) ble amplifisert og sekvensert for svampen Phakellia sp. Amplifiseringen for COI fulgte protokoller modifisert fra Diaz mfl. (2021), mens amplifiseringen for 28S rRNA fulgte protokollene beskrevet av Chombard mfl. (1998). Polymerasekjedereaksjoner (PCR) ble utført i et totalt volum på 50 µL, bestående av 17,5 µL vann, 5 µL CoraLLoad-buffer, 5 µL dNTP, 10 µL Q-solution, 7 µL MgCl₂, 2 µL av hver primer, 0,5 µL Qiagen Taq og 1 µL DNA. Rensing og Sanger-sekvensering ble utført av Eurofins Europe.
Alle sekvenser ble kontrollert for kontaminasjon og identitet ved hjelp av BLAST-søk i GenBank. For sekvenser som grupperte innenfor de antatte taksonomiske gruppene, ble de tre nærmeste samsvarende sekvensene samt sekvenser fra morfologisk lignende arter i Nordøst-Atlanteren vurdert. Enkelgen-sekvenser ble justert ved hjelp av Muscle (Edgar, 2004a, 2004b), implementert i MEGA 12 (Kumar mfl., 2024). Et maksimum likelihood-tre ble konstruert for hver genmarkør i MEGA 12 ved bruk av standard bootstrap (langsom) med 1000 bootstrap-replikasjoner, basert på Tamura-Nei-modellen og inkludert alle kodonposisjoner samt ikke-kodende områder. I tillegg ble en Bayesiansk analyse utført på de samme datasett i MrBayes 3.2.5 (Huelsenbeck og Ronquist, 2001; Ronquist og Huelsenbeck, 2003), med tre parallelle kjøringer på fem millioner generasjoner og prøvetaking hver 100 generasjon.
Resultater
Levedyktige sekvenser ble oppnådd for et utvalg av Phakellia -prøvene samlet inn under prosjektet. Av totalt 19 prøver ble 2 prøver sekvensert for det mitokondrielle COI-markøret, og 5 prøver ga sekvenser for det nukleære 28S rRNA-markøret. BLAST-søk og fylogenetiske analyser plasserte konsekvent alle vellykkede sekvenser innenfor Phakellia ventilabrum -kladen. Dendrogrammene basert på både COI og 28S rRNA (Vedlegg A3) viser sterk støtte for plasseringen av disse prøvene, med Bayesian posterior sannsynligheter > 0,95 og ML-bootstrap-verdier mer enn 75 for de fleste grenene. Referansesekvenser fra GenBank, inkludert Phakellia robusta (Spania, MT506127; Mallorca, PQ778214) og Phakellia hirondellei (Mallorca, PQ766449/PQ778218), ble inkludert for å rotfeste (root) de fylogenetiske trærne og bekrefte identiteten til prøvene. Alle sekvenserte prøver grupperte med P. ventilabrum fra Skottland og Norge, noe som bekrefter at de analyserte prøvene tilhører denne arten.
I dette prosjektet ble artsidentifikasjon og utvikling av genetiske markører for DNA-databaser gjennomført for Phakellia. Flere prøver feilet i amplifisering eller ga sekvenser av lav kvalitet, noe som fremhever utfordringene knyttet til DNA-ekstraksjon fra svampvev. Framgangsmåten ble også benyttet for flere av korallartene dekket av prosjektet. Det oppsto imidlertid flere utfordringer knyttet til isolering av DNA og påfølgende analyser, og resultatene som omfatter koraller vil derfor bli videreført i andre prosjekter og publikasjoner i tilknytning til VDWS-prosjektet. Videre arbeid er nødvendig for å utvikle en standardisert og reproduserbar protokoll for molekylære analyser av disse organismene.
3.5 - Visuelle indikatorer
Vurdering av helsetilstanden hos målarter var basert på bilder tatt in situ med ROV samt fysiske prøver analysert i laboratoriet. Hovedmålet for rutinemessig overvåking av disse artene er å bruke ikke-invasive metoder, som bilder og video, som kan gi pålitelige indikasjoner på helsetilstanden. Rapporten tydeliggjør derfor hvilke kriterier som kan vurderes utelukkende med ROV, og dermed egner seg for langtidsovervåking in situ, og hvilke som også krever innsamling av fysiske prøver. Resultatene viser sammenhenger mellom den samlede helsetilstandsindeksen, sett opp mot anrikningsnivå målt etter avstand til oppdrettsanlegg, modellerte sedimentasjonsverdier og dybde.
HD-video- og bildeopptak ble samlet inn med ROV-en «Freyja» (tekniske detaljer finnes i tilleggsrapporten «Protokoll for visuelle vurdering på helsestilstand til kystnære koraller, svamper og bergskjell i nærheten av oppdrettsanlegg»). Det ble samlet inn tre typer bilder for å visuelt vurdere tilstanden til individer og kolonier: 1. Undervannsbilde for vurdering av visuelle helsekriterier tilhel individer. 2. Nærbilde for detaljert vurdering av vev og tentakelstatus og 3. Bilder for vurdering av visuelle helsekriterier i laboratoriet. Tilleggsundervannsbilder kunne samles inn for å vurdere dekning og tilhørende bunnsamfunn.
De visuelle helsekriteriene for hver målart gruppert i hornkorall, øyekorall, bergskjell og svamp, er forklart i «Protokoll for visuelle vurdering på helsestilstand til kystnære koraller, svamper og bergskjell i nærheten av oppdrettsanlegg» (Dunlop mfl., under utarbeidelse). De visuelle helsekriteriene ble valgt basert på tidligere rapporterte visuelle helsekriterier brukt eller anbefalt for å vurdere helsetilstand hos kaldtvannskoraller og svamp (Kutti mfl., 2016b; Vad mfl., 2017; Weinnig mfl., 2020; Norge, 2024) eller basert på tidligere forskning som indikerer at disse visuelle kriteriene kan kobles til negative helseeffekter av utslipp fra oppdrett eller andre påvirkninger. Kriteriene er enten binære (0 eller 1) eller ordinale (0–2). Binære kategorier ble brukt for kriterier som enten er til stede (1) eller ikke (0), som for eksempel biobegroing, mens ordinale kategorier (0–2) ble brukt for kriterier som kunne vurderes på ulike nivåer av dekning, for eksempel graden av vevsmisfarging. Den samlede helsetilstandindeksen ble beregnet for hver målart, normalisert til 0–2, og gjennomsnittet av de relevante scorene ble brukt. En høyere score fra helsekriterier indikerer en dårligere helsetilstand. Oversikt over de visuelle helsekriteriene er tilgjengelig i tilleggsrapporten «Protokoll for visuelle vurdering på helsestilstand til kystnære koraller, svamper og bergskjell i nærheten av oppdrettsanlegg» (Dunlop mfl., under utarbeidelse).
For å undersøke hvordan avstand til oppdrettsanlegg, akkumulert sedimentasjon og andre miljøvariabler påvirket den visuelle helsetilstanden for ulike bunndyrarter, ble det kjørt GAM med Gaussisk fordeling ved hjelp av pakken mgcv i R (4.4.3). GAM hadde forklaringsvariabler inkludert som smooth terms med en basisdimensjon på k = 3.
De følgende visuelle helsekriteriene ble brukt for sjøtre; sedimentering, misfarging, biobegroing og tentakelaktivitet og alle helsekriterier kunne observeres in-situ med ROV-en. Imidlertid var det vanskelig å finne sjøtre med visuelle tegn på negativ påvirkning med disse kriteriene og derfor fikk mange en visuell helsetilstands på 0 på alle avstander fra anlegget. For sjøtre var det kun dybde (F = 5,991, p = 0,006) som hadde en signifikant effekt på visuell helsetilstand, og koloniene hadde dårligere helsetilstand med økt dybde, ned til 300 m (Figur 11). Avstand til oppdrettsanlegget (F = 0,422, p = 0,519) (Figur 9) og sedimentasjon (F = 0,036, p = 0,851) (Figur 10) hadde ikke noen signifikant effekt på visuell helsetilstandsindeks. Dybde, avstand til oppdrettsanlegget og sedimentasjon forklarte 24,5 % av variasjonen, basert på 46 kolonier. Dette anses som relativt lav til moderat forklaringsgraden, men forventet for denne typen biologiske feltdata. Størstedelen av den forklarte variasjonen var knyttet til dybde, men andre miljø- eller biologiske kriterier kan være viktigere, eller at kriteriene brukt ikke er tilstrekkelige indikatorer for helsetilstand.
Figur 9: Visuell helsetilstand som funksjon av avstand til oppdrettsanlegg for hornkorall (sjøtre, sjøbusk og risengrynkorall). Heltrukne linjer viser GAM-prediksjoner, med skraverte områder som angir 95 % konfidensintervaller. Punktene viser de observerte dataene. p-verdier er vist i panelene.
Figur 10: Visuell helsetilstand som funksjon av sedimentasjon for hornkorall (sjøtre, sjøbusk og risengrynkorall). Heltrukne linjer viser GAM-prediksjoner, med skraverte områder som angir 95 % konfidensintervaller. Punktene viser de observerte dataene. p-verdier er vist i panelene.
Den visuelle helsetilstandindeksen hos sjøbusk ble vurdert basert på følgende kriterier; nakent skjelett, misfarging, biobegroing og tentakelaktivitet. Det ble ikke observert akkumulering av sediment på sjøbuskkolonier. Nakent skjelett, misfarging og biobegroing kunne observeres på ROV-bildene, men ikke misfarging hos denne arten, hvor laboratoriebilder måtte brukes. Sjøbuskkolonier nærmere anlegg hadde en tendens til signifikant dårligere visuelle helsetilstandsindeks (F = 4,478, p = 0,051) (Figur 9), men akkumulert sedimentasjon eller dybde hadde ingen signifikant effekt på den visuelle helsetilstanden (F = 1,187, p = 0,282) (Figur 10) og (F = 2,217, p = 0,157) (Figur 11). Modellen forklarte 50,2 % av variasjonen i datasettet visuell helsetilstand basert på 20 individer. Til tross for den relativt høye forklaringsgraden, bør resultantene tolkes med forsiktighet på grunn av det begrensede antallet. De fem sjøbuskene som ble samlet inn innen 250 m fra oppdrettsanlegget, hadde helseindeksen over 0,75, hovedsakelig på grunn av tapt og misfarget vev, mens de fleste individene som ble samlet inn mer enn 1 km fra anlegget, hadde en helsetilstand under 0,5. Ingen individer ble samlet inn i avstandssonen mellom 500 m og 1 km fra oppdrettsanlegget.
De følgende visuelle helsekriteriene ble brukt for risengrynkorall; nakent skjelett, misfarging, biobegroing og tentakelaktivitet. Nakent skjelett, misfarging og biobegroing kunne observeres på ROV-bildene, men ikke misfarging hos risengrynkorall, hvor laboratoriebilder måtte brukes. Kolonier av risengrynkorall nærmere anlegg (F = 10,674, p < 0,0001) (Figur 9) og ved høyere akkumulert sedimentasjon (F = 4,239, p = 0,012) hadde en signifikant dårligere visuell helsetilstand (Figur 10). Dybde hadde ingen signifikant effekt på den visuelle helsetilstanden (F = 2,149, p = 0,148) (Figur 11). Modellen forklarte 50,4 % av variasjonen basert på 63 koraller, noe som regnes som en relativt høy forklaringsgrad. De fleste individene som ble samlet inn innen 500 m fra oppdrettsanlegget, der sedimentasjonsnivået var høyt, hadde en helsetilstand over 0,75. Kolonier som var mer enn 500 m fra anlegget, viste god visuell helsetilstand, og de fleste hadde helsetilstand under 0,5. Det ble observert en generell trend med dårligere helsetilstand langs anrikingsgradienten, indikert av tapt vev og misfarging forårsaket av nekrose, samt mangel på tentakkelaktivitet. Det var imidlertid fortsatt en del variasjon, spesielt innen 500 m fra anlegget, hvor noen kolonier nær anlegget hadde bedre visuell helsetilstand.
Figur 11: Visuell helsetilstand som funksjon av dybde for hornkorall (sjøtre, sjøbusk og risengrynkorall). Heltrukne linjer viser GAM-prediksjoner, med skraverte områder som angir 95 % konfidensintervaller. Punktene viser de observerte dataene. p-verdier er vist i panelene.
Den visuelle helsetilstandsindeksen hos kålrabisvamp ble vurdert basert på følgende kriterier; sedimentering, vevsmisfarging, biobegroing og arrdannelse og revet vev. Alle helsekriteriene kunne observeres in-situ med ROV-en, med unntak av vevsmisfarging, hvor laboratoriebilder måtte brukes. Kålrabisvamp hadde i generelt en bedre visuelltilstand lengre fra oppdrettsanlegget, særlig mellom 100 til 600 m fra oppdrettsanlegg, men det var ikke en signifikant sammenheng (F = 1,387, p = 0,247) (Figur 12). Tilstanden ble dårligere med økende sedimentasjon mellom 0 og 2000 g·m⁻², men sedimentasjon hadde ingen signifikant effekt på helsetilstand (F = 2,377, p = 0,086) (Figur 13). Flere individer innenfor fra 100 m fra anlegg og dermed utsatt for høye akkumulert sedimentasjon, hadde imidlertid relativt god helsetilstand. Dybde hadde ingen signifikant effekt på visuelle helsetilstand (F = 0,529, p = 0,473) (Figur 14). Modellen forklarte 40 % av variasjonen basert på 32 individer.
Figur 12: Visuell helsetilstand som funksjon av avstand til oppdrettsanlegg for svamp (kålrabisvamp og viftesvamp). Heltrukne linjer viser GAM-prediksjoner, med skraverte områder som angir 95 % konfidensintervaller. Punktene viser de observerte dataene. p-verdier er vist i panelene.
Den visuelle helsetilstandsindeksen hos viftesvamp ble vurdert basert på følgende kriterier; sedimentering, vevsmisfarging, biobegroing, arrdannelse og revet vev og regenerering av vev. Alle helsekriteriene kunne observeres in-situ med ROV-en, med unntak av vevsmisfarging, hvor laboratoriebilder måtte brukes. For de 50 viftesvampene som ble undersøkt, ble den visuelle helsetilstand signifikant dårligere for individer nærmere oppdrettsanlegget (F = 3,666, p = 0,059) (Figur 12) og ved høyere akkumulert sedimentasjon (F = 5,429, p = 0,008) (Figur 13). Dybde hadde ingen signifikant effekt på helsetilstanden (F = 3,267, p = 0,103) (Figur 14). Modellen forklarte 61,7 % av variasjonen i datasettet. Den høye forklaringsgraden tyder på at sedimentasjon er en viktig faktor for variasjon i helsetilstand, mens effekten av avstand til anlegg og dybde er mindre tydelig. Alle individene innenfor 200 m fra oppdrettsanlegget hadde en tilstand over 0,6 og de fleste individene mer enn 750 m fra anlegget, hadde en tilstand under 0,4. Alle individene innen 200 m fra anlegget viste tegn på sedimentasjon, vevsregenerasjon og skader, og hos noen også misfarging. Alle individene som ble utsatt for høy akkumulert sedimentasjon over 1000 g·m⁻² hadde tilstand over 0,6.
Figur 13: Visuell helsetilstand som funksjon av sedimentasjon for svamp (kålrabisvamp og viftesvamp). Heltrukne linjer viser GAM-prediksjoner, med skraverte områder som angir 95 % konfidensintervaller. Punktene viser de observerte dataene. p-verdier er vist i panelene.
Figur 14: Visuell helsetilstand som funksjon av dybde for svamp (kålrabisvamp og viftesvamp). Heltrukne linjer viser GAM-prediksjoner, med skraverte områder som angir 95 % konfidensintervaller. Punktene viser de observerte dataene. p-verdier er vist i panelene.
Bergskjell viste begrensede visuelle tegn på negativ påvirkning. Biobegroing på skallet er en naturlig forekomst og kunne ikke knyttes til negative påvirkninger, og tentakkelaktiviteten ble lett forstyrret av ROV-en. Sedimentasjonsnivået var dermed det eneste mulige kriteriet, men dette var ikke tilstrekkelig til å bygge en tilstandsindeks, og data er ikke presentert for bergskjell.
Avstand til oppdrettsanlegget, dybde eller sedimentasjon hadde ingen signifikant effekt på de visuelle helsetilstandene for øyekorall (n = 24) og modellen forklarte 16,8 %. Ettersom var prøvestørrelsen for øyekorall lav, og modellen forklarte begrenset variasjon, er det ikke mulig å trekke konklusjoner om sammenhengen mellom den visuelle helsetilstanden til øyekorallene og anrikelse av utslipp.
Resultatene viste generelt at visuelle kriterier på dårlig helsetilstand var korrelert med anrikelse av utslipp for korallene sjøbusk og risengrynkorall, samt svampen viftesvamp, hvor de to sistnevnte har en sterk korrelasjon med både avstand til anlegg og sedimentasjon. Økt sedimentasjon var til dels knyttet til dårligere visuell helsetilstand hos kålrabisvamp, selv om enkelte individer opprettholdt god visuell helsetilstand ved høye sedimentasjonsnivåer. Kriterier for visuell helsetilstand utviklet for sjøtre viste sterkere sammenheng med dybde.
Målet med den visuelle metoden er å være en ikke-invasiv metode for å vurdere helsen til koraller og svamper in-situ ved hjelp av bilder og video fra fjernstyrte (ROV) eller autonome undervannsfarkoster. De fleste visuelle helsekriteriene kunne vurderes ved bruk av ROV alene, men misfarging av vevet var ikke mulig å vurdere ut fra ROV-bildene som ble samlet inn i dette prosjektet. Visuelle helsetilstandsindekser ble derfor beregnet på nytt kun for kriterier som kunne vurderes med ROV, for å avgjøre om metoden kunne brukes uten innsamling av prøver for de utvalgte artene. Dette innebærer at kriteriet misfarging, som må vurderes ved hjelp av laboratoriebilder, ble utelatt fra indeksene beregnet for risengrynkorall, sjøbusk, kålrabisvamp og viftesvamp. Den visuelle helsetilstand til risengrynkorall var fortsatt dårligere nærmere anleggene (F = 7,297, p = 0,0009) og ved høyere akkumulert sedimentasjon (F = 5,302, p = 0,006). Den visuelle helsetilstanden til viftesvamp viste også fortsatt samme mønster, med dårligere helsetilstand nærmere anleggene (F = 3,958, p = 0,0526) og ved høyere akkumulert sedimentasjon (F = 8,053, p = 0,007). Resultatene indikerer at ROV-basert vurdering ved bruk av kun visuelle kriterier som kan vurderes in-situ uten prøvetaking, er effektivt for risengrynkorall og viftesvamp.
For kålrabisvamp og sjøbusk var imidlertid vurdering av vevsmisfarging et nødvendig kriterium for å kunne påvise endringer i visuell helsetilstand langs en anrikningsgradient fra oppdrettsanlegg. Uten vevsmisfarging som kriterium viste helsetilstandsindeksen for sjøbusk ingen signifikant sammenheng med avstand til oppdrettsanlegget (F = 0,476, p = 0,500). Heller ikke for kålrabisvamp ble det funnet noen signifikant sammenheng, verken med avstand til oppdrettsanlegget (F = 0,001, p = 0,978) eller med sedimentasjon (F = 1,396, p = 0,248). Resultatene indikerer at vevsmisfarging er et viktig kriterium for å vurdere den visuelle helsetilstandsindeksen for sjøbusk og kålrabisvamp langs en anrikningsgradient fra oppdrettsanlegg, og at dette bør vurderes enten med laboratorieprøver eller i felt med en ROV-kamera med effektiv fargebalanse og et kamera med god fargeresolusjon.
3.6 - Histologiske analyser av kjønn og modning
Negative effekter kan påvirke koraller og andre arter enten direkte (for eksempel gjennom vevstap eller fysiologiske forstyrrelser) eller indirekte ved å skifte energifordelingen fra vedlikehold til cellulær stressrespons eller vevsreparasjon. Endringer i energifordelingen kan føre til redusert investering i mindre kritiske funksjoner, som reproduksjon, og dermed potensielt redusere eggstørrelse og fekunditet.
Vi har analysert gametogenese og fekunditet hos risengrynkorall og sjøtre ved bruk av histologiske metoder. Individer av begge arter ble samlet inn i 2023 (september) og 2024 (juni og oktober) fra syv lokaliteter fordelt på to områder langs norskekysten (se kapitlene 4.1 og 4.3 for detaljer). Prøvene ble samlet inn fra ulike dybder (139–462 m) og på forskjellige avstander fra oppdrettsanlegg (25–1444 m). Hver prøve besto av én eller flere små grener som ble plassert i 10 % formalin (VWR, Pennsylvania, USA). Før prøvene ble sendt til analyse, ble formalinen erstattet med 75 % etanol (VWR, Pennsylvania, USA).
De histologiske analysene ble utført ved Florida State University Coastal and Marine Laboratory i USA av Sandra Brooke, Zoie Hill og Laura Anthony. Korallene ble prosessert ved bruk av standardiserte histologiske teknikker (se for eksempel Brooke og Järnegren, (2013)), med noen artsspesifikke modifikasjoner.
Metode for risengrynkorall
Omtrent to centimeter av en gren av risengrynkorall ble kuttet fra hvert individ og de harde kalsiumskjellene som beskytter vevet ble fjernet ved bruk av 10 % saltsyre (HCL). Prøvene ble dehydrert i en serie etanolkonsentrasjoner (30 %, 70 %, 95 %, tre x 100 %), 30 min i hver løsning. Etter dehydrering ble den indre hornaktige skjelettaksen fjernet ved bruk av pinsett og skalpell. Hver prøve ble renset i Toluen i 10 min før infiltrering med Epredia Histoplast Parafinvoks i 24 timer. Prøver og voks ble plassert i små brett for avkjøling, for å siden kuttes i 9 µm tynne snitt ved bruk av et Shandon Finesse 325 mikrotom. Snittene ble plassert på et objektglass (fem snitt per prøve) for å tørke. Vevet ble farget ved bruk av Hetoxylin fra Ricca Chemical Company og Eosin fra Scytek Laboratories, og objektglasset til dekket med dekkglass. Et Olympus BX41 mikroskop ble brukt for observasjon og bilder ble tatt av et AmScope MU1403 digitalkamera. Arealene av 100 oocyter ble målt hos alle hunnene ved bruk av bildeanalyseprogrammet ImageJ. Feret diameteren (maksimal bredde) ble beregnet for hvert egg. Disse diameterne ble bukt for å beregne gjennomsnittlig eggdiameter for hver prøve for å kunne avgjøre om reproduksjonen er kontinuerlig (stor variasjon i eggstørrelser) eller sesongmessig (liten distribusjon innenfor begrensede størrelsesklasser).
Modne egg hos risengrynkorall er store og fulle av lipider. Dette gjør histologisk prosessering utfordrende da parafinvoksen ikke klarer å penetrere eggene, noe som gjør det vanskelig å kutte vevet og et mindre antall egg blir med i hvert snitt. For å kunne bestemme øyeblikkelig fekunditet (antall modne egg), ble fem polypper fra hver hunn brukt hvor hver polypp ble målt og eggene fjernet ved bruk av skalpell og pinsett. Modne egg (< 350 µm) ble fotografert, målt og arealet beregnet. Feret diameteren ble også beregnet for hvert egg.
I tillegg til å måle eggene, ble de tildelt et utviklingsstadium i samsvar med nedenstående klassifisering (modifisert fra Waller og Tyler (2005)).
Stadium I: Oogonia: minste oocytene som knoppskyter fra den mesenteriale lamellen.
Stadium II: Previtallogene oocyter: små oocyter med tynne vegger og et basofilt cytoplasma.
Stadium III: Vitallogene oocyter: store oocyter som utvikler eggeplomme i cytoplasma.
Stadium IV: Sene vitallogene oocyter: granulerte oocyter som har ett tykt kortikalt lag rundt ooplasmen.
Metode for sjøtre
Vev ble skåret ut fra den delen av grenene som inneholder polypper. Spiklene som beskytter vevet ble fjernet ved bruk av 10 % saltsyre (HCl). Vevsprøvene ble så dehydrert i en serie etanolkonsentrasjoner i 8 timer hver (30 %, 70 %, 95 %, 3 x 100 %). Etterpå ble prøvene renset i Histoclear i to til åtte timer før de ble infiltrert med parafinvoks i 24 timer og lagt i små brett for avkjøling. Voksblokkene ble kuttet i 8 µm tynne snitt ved bruk av en Shandon Finesse 325 mikrotom og deretter behandlet på samme måte som risengrynkorall. Fekunditet kunne ikke bestemmes for sjøtre da de reproduktive polyppene var for små for disseksjon.
Resultater for risengrynkorall
Kjønnsfordeling
Totalt inneholdt prøvene av risengrynkorall 19 hunner og 21 hanner (48/52 %), ett omtrentlig jevnt forhold, og tre individer uten noen gameter.
Eggstørrelse
Gjennomsnitt og standardavvik (SD) av eggenes diameter fra prøver i de ulike kategoriene sesong, dybde og avstand fra anlegg vises i tabell 6. Dette er data fra histologiske snitt og inkluderer ikke størrelse og fekunditetsinformasjonen fra dissekerte store egg som presenteres separat.
Sesong
Gjennomsnitt
SD
Antall ♀
sep. 23
114,85
30,66
12
jun. 24
116,70
29,52
4
okt. 24
105,77
35,94
3
Dybde (m)
< 200
114,48
29,63
12
200-300
102,68
27,03
4
300-400
152,61
120,29
1
> 400
112,62
43,85
2
Avstand (m)
< 100
111,26
20,15
3
100-300
109,18
24,86
4
300-500
105,63
36,91
4
500-700
112,31
48,09
3
700-900
137,94
89,57
1
> 1000
123,61
31,91
4
Tabell 6: Gjennomsnittlig diameter (µm) og standard avvik (SD) av eggene til risengrynkorall angitt for sesong, dybde og avstand til oppdrettsanlegg. Dette er data fra histologiske snitt og inkluderer ikke informasjon om de store eggene fra dissekerte store egg.
Statistiske analyser (one-way ANOVA) viser ingen signifikante forskjeller for sesong (F = 0,124, p = 0,884), dybde (F = 0,730, p = 0,550) eller avstand til oppdrettsanlegg (F = 0,243, p = 0,936). Eggene til risengrynkorall hadde en gjennomsnittlig diameter på 114 µm (SD = 29,6 µm). Hoveddelen av eggene var i stadium 3 (53,6 %) fulgt av stadium 2 (24,5 %), stadium 4 (18,0 %) og stadium 1 (3,9 %). Eggenes diameter varierer med utviklingsstadium (Figur 15), med stadium 1 gjennomsnittlig diameter = 29,1 µm (SD: 11,5 µm); stadium 2 = 55,9 µm (SD: 14,2 µm); stadium 3 = 128,2 µm (SD: 49,3 µm) og stadium 4 = 601,3 µm (SD: 135,2).
Figur 15: Gjennomsnitt og standardavvik (SD) av eggenes diameter (µm) i hvert utviklingsstadium. Boksene viser 25/75 persentiler og linjene 10/90 persentiler.
Størrelsen på eggene viste ingen signifikante forskjeller relatert til innsamlingsmåned, men prosentandelen av eggene i forskjellige stadium og antall i hvert stadium relatert til innsamlingsmåned indikerer at der er flere modne egg i september enn i juni og oktober (Figur 16). Dette indikerer sesongmessig gyting, til tross for at alle stadier er til stede i alle innsamlingsmåneder.
Figur 16: Prosent (A) og antall oocytter (B) i de forskjellige utviklingsstadiene (1-4) i prøver av risengrynkorall fra juni, september og oktober.
Fekunditet
Modne (stadium 4) egg ble fjernet fra fem polypper per prøve, telt opp og diameteren på eggene målt. Den totale gjennomsnittlige diameteren var 603,45 µm (SD: 92,51), noe som er nesten det samme som Feret-diameteren hos stadium 4 eggene i den histologiske prosesseringen (601,3 µm (SD: 135,2)). Fekunditet ble kun bestemt for prøvene fra september 2023, hvilket gjør at der ikke er andre perioder å sammenligne med. Fekunditet viste ingen sammenheng med dybde og avstand fra annlegg (Figur 17).
Figur 17: Fekunditet (modne oocytter per polypp) hos risengrynkorall relatert til (A) dybde og (B) avstand fra oppdrettsanlegg.
Resultater for sjøtre
Kjønnsfordeling
Totalt inneholdt prøvene av sjøtre ti hunner og to hanner. Disse var fordelt på september 2023: 4 hunner og 1 hann (80/20 %), juni 2024: 5 hunner og 1 hann (83:17 %) og oktober 2024: 1 hunn og 0 hanner (100:0 %). Andelen hunner var betydelig høyere enn hanner Overlag var kjønnsfordelingen sterkt skjev mot hunner og denne trenden var konsekvent over tid.
Eggstørrelse
Sjøtrærenes egg hadde totalt sett en gjennomsnittlig diameter på 87 µm (SD = 33,9 µm) og hoveddelen av eggene var i utviklingsstadium 2 og 3, i likhet med risengrynkorall. Dog er ikke kategoriseringen av utviklingsstadium komplett ennå for denne arten. Gjennomsnitt og standardavvik (SD) av eggenes diameter fra prøver i de ulike kategoriene sesong, dybde og avstand fra anlegg vises i (Tabell 7).
Sesong
Gjennomsnitt
SD
Antall ♀
sep. 23
104,48
49,37
4
jun. 24
76,22
16,97
5
okt. 24
75,32
13,84
1
Dybde (m)
< 200
85,07
39,85
7
200-300
92,95
19,26
3
300-400
0
0
0
> 400
0
0
0
Avstand (m)
< 100
86,62
28,37
1
100-300
61,21
23,41
3
300-500
100,14
44,39
4
500-700
0
0
0
700-900
101,77
16,59
2
> 1000
0
0
0
Tabell 7: Gjennomsnittlig eggstørrelse og standardavvik (SD) relatert til sesong, dybde og avstand til oppdrettsanlegg.
Statistiske analyser (one-way ANOVA) viser ingen signifikante forskjeller for sesong (F = 0,801, p = 0,486), dybde (F = 0,025, p = 0,994) eller avstand fra oppdrettsanlegg (F = 0,343, p = 0,864). Siden kategorisering av utviklingsstadium ennå ikke er komplett for denne arten, var disseksjon av de små reproduksjonspolyppene ikke mulig for å evaluere umiddelbar fekunditet. Informasjonen som er samlet inn til nå indikerer at reproduksjon hos sjøtre ikke er sesongbetont, og at det ikke er noen signifikante effekter av dybde eller avstand fra anlegg på reproduksjonen hos denne arten.
3.7 - Lysosomal membranstabilitet
Lysosomal membranstabilitet (LMS) er et tidlig og følsomt mål på cellulært stress, og brukes som indikator på påvirkning fra miljøfaktorer som forhøyet temperatur, økt sedimentbelastning eller kjemisk forurensning. Lysosomene spiller en sentral rolle i cellens nedbrytning og resirkulering av komponenter, og tap av membranstabilitet kan føre til lekkasje av enzymer ut i cytoplasmaet og tap av cellulær funksjon (f.eks. (Mamaca mfl., 2005; Edge mfl., 2014, 2016; Mukherjee mfl., 2015; Strand mfl., 2017; Kutti mfl., 2022; Scanes mfl., 2024)).
For å vurdere lysosomal integritet i svamper (viftesvamp og kålrabisvamp) og bergskjell ble nøytralrød-retensjonsmetoden (Neutral Red assay) benyttet, basert på en protokoll utviklet av Edge mfl. (2014, 2015, 2016) etter Lowe mfl. (1992) og Ringwood mfl. (1998, 2005).
Metode
Svamper ble holdt i kaldt sjøvann og mørke inntil prosessering. Vevet ble kuttet i små biter (ca. 1 cm) på objektglass og deretter overført til 24-brønners cellekulturplater med 1 mL kaldt Ca²⁺- og Mg²⁺-fritt sjøvann (CMFS) per brønn. Platene ble dekket med aluminiumsfolie og inkubert i kjøleskap i 30 min. Etter inkubasjon ble cellene forsiktig frigjort ved hjelp av en Pasteurpipette med avknekt spiss, ved gjentatt opptrekk og utslipp av prøven. Celleoppløsningen ble deretter filtrert gjennom et nylonfilter (40 µm) og filtratet ble sentrifugert ved 2000× g i 5 min ved 10–15 °C. Supernatanten ble kastet, og pellet ble vasket med 1 mL kald CMFS og sentrifugert på nytt under samme betingelser før den ble resuspendert i 50 µL CMFS.
For bergskjell ble 50 µL hemolymfe samlet med sprøyte og nål, deretter overført til et 1,5 mL rør og oppbevart kaldt inntil farging. Ingen ytterligere filtrering eller vevsprosessering ble utført for denne arten.
De ferdigpreparerte prøvene ble tilsatt 50 µL 0,08 mg·L⁻¹ nøytralrød-arbeidsløsning (Sigma-Aldrich, Missouri, USA; løsningen ble forberedt fersk hver dag) og 50 µL kald CMFS. Etter forsiktig blanding ble prøvene inkubert ved 10–15 °C i mørke i 60 min. Rett før vurdering ble 20 µL av cellesuspensjonen plassert på et objektglass.
Cellene ble observert i mikroskop ved bruk av 40×-objektiv, og minst 50 celler per prøve ble vurdert der det var mulig. Celler ble kategorisert som stabile , der fargestoffet forble innenfor lysosomene, eller ustabile , der fargestoffet hadde lekket ut i cytoplasmaet (Vedlegg A4). Prosentvis destabilisering ble beregnet som andelen ustabile celler av totalt antall observerte celler.
For å analysere sammenhenger mellom miljøvariabler og andelen destabiliserte celler i svampvev ble det brukt GAM med betaregresjon og logit-link, implementert i mgcv-pakken i R. Andeler ble beregnet som prosentandel destabiliserte celler dividert på 100 og transformert til det åpne intervallet (0–1) for å unngå eksakte 0- og 1-verdier. Separate modeller ble kjørt for hver art. Fullmodellen inkluderte glatte ledd for avstand til nærmeste oppdrettsanlegg, dybde og sedimentasjonsrate. For data fra siste tokt (2025005014) ble det i tillegg kjørt forenklede modeller uten sedimentasjon, da modellerte sedimentasjonsdata for 2025 ikke var tilgjengelige. Modelltilpasning og betydningen av de glatte leddene ble vurdert basert på forklart devians, justert R² og tilnærmede χ²-tester.
Resultater
For bergskjell forklarte den fulle modellen med avstand til anlegg, dybde og akkumulert sedimentasjon 43,1 % av deviansen (justert R² = 0,20). Dybde hadde en signifikant effekt på andelen destabiliserte celler (χ² = 4,32, p = 0,038), mens avstand til anlegg viste en svak, marginalt signifikant trend (χ² = 3,45, p = 0,063). Akkumulert sedimentasjon hadde ingen signifikant effekt (χ² = 4,06, p = 0,122) (Figur 18). Den forenklede modellen uten sedimentasjon forklarte derimot 70,2 % av deviansen (justert R² = 0,71), med sterke og signifikante effekter av både avstand til anlegg (χ² = 22,83, p < 0,001) og dybde (χ² = 28,35, p < 0,001). Den økte forklaringsgraden i den forenklede modellen skyldes sannsynligvis både et større antall observasjoner (n = 18 vs. n = 11) og fraværet av en ikke-signifikant kovariat, snarere enn manglende biologisk relevans av sedimentasjon.
Figur 18: Lysosomal membranendestabilisering hos bergskjell. Predikerte sammenhenger mellom andelen destabiliserte celler og miljøvariabler basert på generaliserte additive modeller (GAM). Venstre kolonne viser modellene som inkluderer sedimentasjon (full modell), mens høyre kolonne viser modellene uten sedimentasjon (forenklet modell). Paneler A–B viser effekten av avstand til nærmeste oppdrettsanlegg, paneler C–D viser effekten av dybde, og panel E viser effekten av sedimentasjonsrate. Grå prikker representerer observerte verdier, heltrukne linjer viser predikerte midler, og skyggelagte områder viser 95 % konfidensintervall.
For kålrabisvamp forklarte den fulle modellen med avstand til anlegg, dybde og akkumulert sedimentasjon 45,4 % av deviansen (justert R² = 0,33). Alle forklaringsvariablene hadde signifikante effekter på andelen destabiliserte celler, med sterkest effekt av avstand til anlegg (χ² = 12,99, p < 0,001), etterfulgt av dybde (χ² = 8,38, p = 0,013) og akkumulert sedimentasjon (χ² = 6,96, p = 0,008) (Figur 19). Når sedimentasjon ble utelatt og et større datasett ble benyttet, var verken avstand til anlegg (χ² = 2,67, p = 0,196) eller dybde (χ² = 2,34, p = 0,126) signifikante prediktorer, og modellen forklarte kun 24,7 % av deviansen (justert R² = 0,17). Dette indikerer at akkumulert sedimentasjon fanger opp en viktig miljøgradient som påvirker cellestabiliteten hos kålrabisvamp, og at utelatelse av denne variabelen reduserer modellens forklaringskraft, til tross for økt prøvestørrelse.
Figur 19: Lysosomal membranendestabilisering hos kålrabisvamp. Predikerte sammenhenger mellom andelen destabiliserte celler og miljøvariabler basert på generaliserte additive modeller (GAM). Venstre kolonne viser modellene som inkluderer sedimentasjon (full modell), mens høyre kolonne viser modellene uten sedimentasjon (forenklet modell). Paneler A–B viser effekten av avstand til nærmeste oppdrettsanlegg, paneler C–D viser effekten av dybde, og panel E viser effekten av sedimentasjonsrate. Grå prikker representerer observerte verdier, heltrukne linjer viser predikerte midler, og skyggelagte områder viser 95 % konfidensintervall.
For viftesvamp forklarte den fulle modellen med avstand til anlegg, dybde og akkumulert sedimentasjon 35,2 % av deviansen (justert R² = 0,25). Avstand til anlegg viste en marginalt ikke-signifikant sammenheng med andelen destabiliserte celler (χ² = 3,85, p = 0,054), mens verken dybde (χ² = 0,47, p = 0,494) eller akkumulert sedimentasjon (χ² = 0,42, p = 0,519) hadde signifikante effekter (Figur 20). I den forenklede modellen uten sedimentasjon forklarte modellen 43,0 % av deviansen (justert R² = 0,37), og her var avstand til anlegg en signifikant prediktor for andelen destabiliserte celler (χ² = 6,62, p = 0,010), mens dybde fortsatt ikke hadde noen signifikant effekt (χ² = 0,74, p = 0,389). Dette indikerer at viftesvamp i begrenset grad påvirkes av avstand til oppdrettsanlegg, mens øvrige miljøvariabler hadde liten betydning innenfor det undersøkte området.
Figur 20: Lysosomal membranendestabilisering hos viftesvamp. Predikerte sammenhenger mellom andelen destabiliserte celler og miljøvariabler basert på generaliserte additive modeller (GAM). Venstre kolonne viser modellene som inkluderer sedimentasjon (full modell), mens høyre kolonne viser modellene uten sedimentasjon (forenklet modell). Paneler A–B viser effekten av avstand til nærmeste oppdrettsanlegg, paneler C–D viser effekten av dybde, og panel E viser effekten av sedimentasjonsrate. Grå prikker representerer observerte verdier, heltrukne linjer viser predikerte midler, og skyggelagte områder viser 95 % konfidensintervall.
Bergskjell de tydeligste responsene på de ulike miljøgradientene, med sterke effekter av både dybde og avstand fra oppdrettsanlegg, mens kålrabisvamp var mest korrelert til sedimentasjonsnivå. Viftesvamp viste svakere og mer variable mønstre, noe som indikerer artsforskjeller i sensitivitet overfor miljøfaktorer knyttet til oppdrettsaktivitet.
Under siste tokt (nr. 2025005014) ble metoden videreutviklet og tilpasset for de to korallartene risengrynkorall og sjøtre. For disse ble vevet mekanisk homogenisert (henholdsvis ved knusing med morter og ved oppdeling på objektglass), filtrert gjennom henholdsvis 100 µm og 70 µm filter, og behandlet etter samme prinsipp som svampeprøvene. Antall tilgjengelige prøver var imidlertid for lavt til kvantitativ analyse, men metoden ble testet vellykket i felt. Mikroskopbilder som viser representative celler og fargemønstre fra disse forsøkene er inkludert i (Figur 21).
Figur 21: Mikroskopbilder som viser lysosomalceller fra sjøtre: A) stabile og B) destabiliserte celler, og fra risengrynkorall: C) stabile og D) destabiliserte celler.
Et sett med individuelle kålrabisvamper og viftesvamper ble analysert for både lysosomal membranendestabilisering og visuell helsetilstandsindeks. Sammenhengen mellom andelen destabiliserte celler og visuell helsetilstandsindeks ble undersøkt ved bruk av generaliserte additive modeller (GAM). For kålrabisvamp og viftesvamp var det en generell tendens til flere destabiliserte celler ved høyere visuelle helsetilstandsindekser, men sammenhengen var ikke signifikant; kålrabisvamp (F = 2,144, p = 0,154) og viftesvamp (F = 2,338, p = 0,081).
Figur 22: Lysosomal membranendestabilisering (%) og visuell helseindeks hos kålrabisvamp og viftesvamp. Predikerte sammenhenger mellom andelen destabiliserte celler og visuell helseindeks basert på generaliserte additive modeller (GAM). Grå prikker representerer observerte verdier, heltrukne linjer viser predikerte midler, og skyggelagte områder viser 95 % konfidensintervall.
3.8 - Oksygenforbruk og ammoniumsekresjon
Oksygenforbruk og ammoniumsekresjon er viktige indikatorer på korallers metabolske aktivitet og energibalanse. Oksygenforbruk målt ved respirometri gir direkte informasjon om korallens respiratoriske aktivitet og kan brukes til å vurdere hvor mye energi organismen bruker under ulike miljøforhold. Ammoniumsekresjon reflekterer nitrogenmetabolismen og kan gi innsikt i korallens ernæringsstatus og samspill med symbionter og mikrobielle samfunn.
Oksygenforbruk
Etter innsamling og en 24 timers akklimatiseringsperiode i de nye tankene, ble flere korallarter inkubert i mørke i sirkulære kamre for å måle oksygenforbruksrater og rater for ammoniakk-utskillelse. Ett tomt kammer ble brukt som kontroll i hver måleserie. Kontrollkamrene, uten koraller, ble brukt til å bestemme bakgrunnsnivåer av mikrobiell respirasjon. Inkubasjonskamrene (860 mL) besto av en akrylglass-sylinder (høyde: 20 cm, indre diameter: 7,4 cm, ytre diameter: 8 cm), lukket i topp med 3D-printede nylonlokk utstyrt med O-ring-tetning. Vannpumper (JOVTOP-dc 12 V; 3 L·min⁻¹ koblet til kamrene med to fleksible PVC-slanger ble brukt for å sikre homogenisering av sjøvannet inne i kamrene.
Oksygenkonsentrasjonen i hvert kammer ble registrert hvert 2. sekund ved bruk av fiberoptiske oksygensensorer (DP-PSt3, PreSens) koblet til en flerkanals oksygenmåler (OXY-4 SMA, PreSens), til den hadde sunket til ≤ 10 % av startverdien. Temperaturen ble målt hvert 2. sekund i kontrollkamrene ved hjelp av en temperatursensor (Pt100, PreSens). Oksygenforbruksraten ble beregnet ved hjelp av R-pakken respR (Harianto mfl., 2019). Alle verdier ble korrigert for bakgrunnsverdier målt i kontrollkamrene. Oksygenforbruksratene er uttrykt som μmol O2/gAFDW/t og ble beregnet som:
Etter inkubasjon ble korallene prøvetatt i vialer og fryst ved −20 °C for senere analyser av tørrvekt og aske-fri tørrvekt (AFDW), som ble brukt til å normalisere respirasjonsdataene.
For å analysere hvordan oksygenforbruk og ammoniumsekresjon varierte med miljøvariabler, ble det brukt GAM med Gamma-fordeling og log-link. Denne modellspesifikasjonen er egnet for kontinuerlige og positive responsvariabler. Modellene inkluderte glatte ledd for avstand til oppdrettsanlegg, sedimentasjon og dybde.
Figur 23: Predikert oksygenforbruk for sjøtre og risengrynkorall som funksjon av miljøvariabler. Heltrukne linjer viser GAM-prediksjoner, med skraverte områder som angir 95 % konfidensintervaller. Punktene viser de observerte dataene. Modellene inkluderte smooth terms for avstand til oppdrettsanlegg, sedimentasjon og dybde, og ble tilpasset med Gamma-fordeling og log-link. Panelene A, C og E viser sjøtre for henholdsvis avstand til oppdrettsanlegg, sedimentasjon og dybde, mens panelene B, D og F viser risengrynkorall for de samme variablene i samme rekkefølge. Forklart variasjon og p-verdier for hver glatt funksjon er vist i panelene.
For sjøtre viste modellen tegn til sammenhenger mellom oksygenforbruk og miljøvariabler (Figur 23). Sedimentasjon hadde en signifikant effekt på oksygenforbruket (F = 6,009, p = 0,021), mens avstand til anlegg (F = 3,584, p = 0,055) og dybde (F = 3,897, p = 0,059) viste svake, men ikke-signifikante tendenser. Modellen forklarte 31,4 % av variasjonen, og den justerte R²-verdien var 0,169, noe som indikerer moderat forklaringskraft. Resultatene antyder at oksygenforbruket hos sjøtre kan påvirkes av miljøfaktorer relatert til oppdrettsaktivitet og sedimentasjonsnivå.
For risengrynkorall ble det ikke funnet noen signifikante sammenhenger mellom oksygenforbruk og de undersøkte miljøvariablene (Figur 23). Verken avstand til anlegg (F = 0,495, p = 0,486), dybde (F = 1,428, p = 0,369) eller sedimentasjon (F = 0,483, p = 0,492) hadde målbar effekt. Modellen forklarte 7,5 % av deviansen, og den justerte R²-verdien var –0.015, noe som viser at modellen ikke hadde forklaringskraft utover tilfeldig variasjon. Dette tyder på at oksygenforbruket hos risengrynkorall ikke var systematisk påvirket av de inkluderte miljøfaktorene i dette datasettet (n = 40).
For øyekorall viste modellen tilsynelatende høy forklart variasjon (85,7 %) og justert R² = 0,441, men datagrunnlaget var svært begrenset (n = 7) (Figur 24). Ingen av de undersøkte variablene var signifikante (alle p > 0,18), og resultatene bør derfor tolkes med stor forsiktighet. Det lave antallet observasjoner gjør at modellen ikke kan brukes til å trekke sikre konklusjoner om sammenhenger mellom oksygenforbruk og miljøfaktorer hos øyekorall.
Til sammenligning viser resultatene at sjøtre responderer noe mer på miljøvariabler enn risengrynkorall, mens øyekorallanalysen var for databegrenset til å vurdere mønstre. Generelt indikerer analysene at variasjon i oksygenforbruk i stor grad kan være påvirket av lokale forhold eller biologiske forskjeller som ikke fanges opp av de målte miljøfaktorene.
Ammoniumsekresjon
For å måle ammoniumsekresjon, ble det tatt to 20 mL-prøver med sjøvann fra hvert kammer ved starten av hver inkubasjon, samt én 20 mL-prøve ved slutten av inkubasjonsperioden. Prøvene ble filtrert gjennom 0,22 μm-filter og oppbevart i scintillasjonsglass ved −20 °C til analyse. Ammoniumkonsentrasjonen ble målt ved fluorometrisk metode beskrevet av Kérouel og Aminot (1997) og Holmes mfl. (1999) ved bruk av Skalar autoanalysator (Skalar, Nederland) ved HI i Bergen. Ammoniumsekresjonsraten ble uttrykt som μmol NH₄⁺/g AFDW/t. Her er NH₄⁺ forskjellen i ammoniumkonsentrasjon mellom slutten og starten av inkubasjonen (μmol NH₄⁺) og AFDW er aske-fri tørrvekt av den inkuberte prøven (g). Kontrollkamrene ble brukt til bakgrunnskorreksjon av sekresjonsraten.
Figur 24: Predikert oksygenforbruk og ammoniumsekresjon for øyekorall som funksjon av miljøvariabler. Heltrukne linjer viser GAM-prediksjoner, med skraverte områder som angir 95 % konfidensintervaller. Punktene viser de observerte dataene. Modellene inkluderte smooth terms for avstand til oppdrettsanlegg, sedimentasjon og dybde, og ble tilpasset med Gamma-fordeling og log-link. Forklart variasjon og p-verdier for hver glatt funksjon er vist i panelene.
For sjøtre viste modellen for ammoniumsekresjon generelt lave forklaringsverdier (Figur 25). Ingen av de undersøkte miljøvariablene hadde signifikant effekt: avstand til anlegg (F = 1,075, p = 0,309), dybde (F = 0,004, p = 0,949) og sedimentasjon (F = 0,729, p = 0,419). Modellen forklarte 16,5 % av deviansen, og den justerte R²-verdien var 0,074, noe som tyder på at variasjonen i ammoniumsekresjon hos sjøtre i liten grad kan tilskrives de målte miljøfaktorene.
For risengrynkorall ble det heller ikke funnet noen signifikante sammenhenger mellom ammoniumsekresjon og miljøvariabler (Figur 25). Avstand til anlegg (F = 1,982, p = 0,168), dybde (F = 0,128, p = 0,893) og sedimentasjon (F = 0,425, p = 0,468) hadde ingen målbare effekter. Modellen forklarte 17,2 % av deviansen, og den justerte R²-verdien var 0,089. Dette indikerer svak forklaringskraft og antyder at ammoniumsekresjonen hos risengrynkorall i hovedsak bestemmes av andre faktorer enn de som inngikk i denne analysen (n = 40).
Figur 25: Predikert ammoniumsekresjon for sjøtre og risengrynkorall som funksjon av miljøvariabler. Heltrukne linjer viser GAM-prediksjoner, med skraverte områder som angir 95 % konfidensintervaller. Punktene viser de observerte dataene. Modellene inkluderte smooth terms for avstand til oppdrettsanlegg, sedimentasjon og dybde, og ble tilpasset med Gamma-fordeling og log-link. Panelene A, C og E viser sjøtre for henholdsvis avstand til oppdrettsanlegg, sedimentasjon og dybde, mens panelene B, D og F viser risengrynkorall for de samme variablene i samme rekkefølge. Forklart variasjon og p-verdier for hver glatt funksjon er vist i panelene.
For øyekorall viste modellen ingen signifikante sammenhenger mellom ammoniumsekresjon og miljøvariabler (alle p > 0,5), og verken avstand til anlegg (F = 0,004, p = 0,955), dybde (F = 0,486, p = 0,536) eller sedimentasjon (F = 0,174, p = 0,705) hadde målbar effekt (Figur 24). Selv om modellen formelt forklarte 36 % av deviansen, var antall observasjoner svært lavt (n = 7), og resultatene må derfor tolkes med stor forsiktighet. Datagrunnlaget er for begrenset til å trekke pålitelige konklusjoner om hvordan miljøfaktorer påvirker ammoniumsekresjon hos øyekorall.
Sammenlignet viser modellene at ammoniumsekresjonen hos alle tre artene varierer betydelig mellom individer og lokaliteter, og at de undersøkte miljøvariablene forklarer bare en liten del av denne variasjonen. Dette antyder at andre biologiske eller miljømessige faktorer kan ha større betydning for variasjonen i ammoniumsekresjon hos koraller.
Figur 26: Sammenheng mellom oksygenforbruk og ammoniumsekresjon for sjøtre (A) og risengrynkorall (B). Punktene viser individuelle korallprøver, mens linjene viser predikerte sammenhenger fra generaliserte additive modeller (GAM) med 95 % konfidensintervaller (skyggefelter). Responsvariabelen er ammoniumsekresjon (µmol N·g⁻¹·AFDW t⁻¹), modellert som en funksjon av oksygenforbruk (µmol O₂·g⁻¹·AFDW·h⁻¹).
For sjøtre viste modellen en svak sammenheng mellom oksygenforbruk og ammoniumsekresjon (Figur 26). Den smooth term for oksygenforbruk var ikke signifikant (F = 1,211, p = 0,28), og modellen forklarte kun 3,78 % av deviansen, med en justert R² på –0,0017. Dette indikerer at variasjonen i ammoniumsekresjon hos sjøtre bare i liten grad kan forklares av oksygenforbruk under de målte forholdene.
For risengrynkorall ble det derimot funnet en signifikant sammenheng mellom oksygenforbruk og ammoniumsekresjon (Figur 26). Den smooth term for oksygenforbruk var signifikant (F = 4,26, p = 0,0354), og modellen forklarte 19,4 % av deviansen, med en justert R² på 0,167. Dette indikerer en moderat forklaringskraft og tyder på at høyere oksygenforbruk generelt er assosiert med økt ammoniumsekresjon hos denne arten.
For øyekorall viste modellen en sterk og signifikant sammenheng mellom oksygenforbruk og ammoniumsekresjon (Figur 26). Den smooth term var sterkt signifikant (F = 18,01, p = 0,0078), og modellen forklarte 83,5 % av deviansen, med en justert R² på 0,822. Selv om antallet observasjoner var svært lavt (n = 7), tyder resultatene på en sterk positiv sammenheng mellom oksygenforbruk og ammoniumsekresjon hos øyekorall.
Samlet viser modellene at ammoniumsekresjon hos alle tre artene påvirkes av metabolsk aktivitet (oksygenforbruk), men styrken på denne sammenhengen varierer. Hos øyekorall forklarer oksygenforbruk mesteparten av variasjonen, mens hos sjøtre og risengrynkorall antyder resultatene at også andre faktorer bidrar til individuelle og lokalitetsbaserte forskjeller i ammoniumsekresjon.
Derimot viste analysene av forholdet mellom oksidativt stoffskifte (O₂-forbruk) og ammoniumsekresjon en signifikant positiv sammenheng for begge arter, med moderat forklaringskraft. Dette tyder på at høyere metabolsk aktivitet er assosiert med økt nitrogenutskillelse, i tråd med forventninger basert på fysiologiske prosesser. Resultatene understøtter dermed at korallenes ammoniumsekresjon i stor grad reflekterer variasjoner i energimetabolisme snarere enn direkte miljøpåvirkning.
3.9 - Lipid- og fettsyreanalyser
Lipider, inkludert fettsyrer (FA), utgjør en viktig energikilde for marint liv. Fettsyresignaturer, kjent som «fettsyretrofiske markører» (FATM), kan brukes til å studere kosthold og energikilder hos marine organismer (Madgett mfl., 2019), og de terrestriske FATM-ene er spesielt gode biomarkører for å påvise om organismer ernærer seg på organisk materiale fra akvakultur. Sammensetningen av laksefôr har endret seg betydelig de siste tiårene, der andelen marine ingredienser har sunket fra 90 % i 1990 til 25 % i 2016, mens plantebaserte ingredienser har økt fra omtrent 0 % til 71 % (Ytrestøyl mfl., 2015; Aas mfl., 2019). I norsk lakseoppdrett har fiskeolje i stor grad blitt erstattet med rapsolje (Aas mfl., 2019), noe som har endret fettsyreprofilen i fôret markant. Rapsolje har høye nivåer av terrestriske fettsyrer som oljesyre (18:1n-9), linolsyre (18:2n-6) og α-linolensyre (18:3n-3), som utgjør henholdsvis ca. 44 %, 15 % og 8 % av de totale fettsyrene, sammenlignet med 12 %, 2,5 % og 1,5 % i fiskeolje (Bell mfl., 2003). Som følge av dette øker konsentrasjonen av terrestriske fettsyrer i sedimentene rundt oppdrettsanlegg (White mfl., 2017; Woodcock mfl., 2019).
FA-analyse har derfor blitt en effektiv biomarkør for fôring på organisk materiale fra oppdrettsanlegg hos både marin fisk (Fernandez-Jover mfl., 2011; Meier mfl., 2023) og flere virvelløse arter, inkludert blåskjell (Mytilus edulis) (Redmond mfl., 2010; Handå mfl., 2012; Woodcock mfl., 2017), slangestjerner (Ophiocomina nigra), kråkeboller (Echinus esculentus), taskekrabber (Cancer pagurus) (Woodcock mfl., 2018), børstemark (Capitella sp.) (Woodcock mfl., 2019), trollkrabbe (Cancer irroratus) (Sardenne mfl., 2020), europeisk hummer (Homarus gammarus) (Baltadakis mfl., 2020) og dypvannsreke (Olsen mfl., 2009; 2012; Søvik mfl., 2025).
Videre er det uttrykt bekymring for at unormalt høye nivåer av terrestriske fettsyrer hos bunnlevende virvelløse dyr (White mfl., 2017; White mfl., 2018; Sardenne mfl., 2020; Drolet mfl., 2022) og villfisk (Gonzalez-Silvera mfl., 2020; Meier mfl., 2023) kan påvirke lipidmetabolismen. Dette kan føre til endret sammensetning av membranlipider og energilagre, samt redusert tilgjengelighet av essensielle marine langkjedede flerumettede fettsyrer (som EPA og DHA), som er avgjørende for normal gonadeutvikling, eggkvalitet og embryonal utvikling. Endringer i fettsyresammensetningen kan dermed påvirke reproduksjonssuksess, larveoverlevelse og rekruttering, og potensielt få konsekvenser for bestandsdynamikk i områder påvirket av akvakultur.
Metode
Lipidmengde (% av tørrvekt) og fettsyresammensetning (% av total fettsyre) ble analysert i fire forskjellige bunnarter; kålrabisvamp, viftesvamp, sjøtre og bergskjell.
Totalt lipid ble ekstrahert med kloroform: metanol (2:1 v/v) og lipidmengden ble bestemt gravimetrisk ved Folch metode (Meier mfl., 2006). Alle prøvene ble metylert, og de respektive fettsyremetylesterne (FAME) ble analysert på en HP-7890A gasskromatograf (Agilent, USA) med en flammeioniseringsdetektor (GC-FID) i henhold til en metode beskrevet i (Meier mfl., 2006) med fettsyren 19:0 tilsatt som en intern standard. 2,5 M tørr HCl i metanol ble brukt som metyleringsreagens. FAME-ene ble ekstrahert ved bruk av 2×2 mL heksan. Det kombinerte ekstraktet ble fortynnet eller konsentrert for å oppnå en passende konsentrasjon for gasskromatografi før 1 µL ble injisert (splittfri) på GC.
De fire forskjellige artene som ble analysert har meget spesialiserte lipidsammensetninger, noe som gav utfordringer i det analystiske arbeide (Tabell 8).
Dyr gruppe
Taxa
Lipidkarakteristikker
Metode modifikasjon
Svamp
Geodia barretti
Inneholder store mengder med bakterie-FA i form av forgrenet mettet FA (SFA). I tillegg består lipidene også av store mengder meget langkjedete “none-methyl interupted FAs (NMI-FA)
FAME er analysert på 60 m DP5 GC-kolonne for at sikre god separasjon av forgrenet isomere
Svamp
Phakellia ventilabrum
Inneholder meget mono-umette FA (MUFA) i (n-7)-familien og meget langkjedete NMI-FA
FAME er analysert ved standard metoden
Koral
Paragorgia arborea
Inneholder store mengder med voks-ester (fettsyre og fettalkoholer (FAOH) koblet sammen av en ester-binding)
Etter metanolysen er FAME og FAOH separert på SPE kolonner og deretter analyser hver for seg med standard metoden)
Muslinger
Acesta excavata
Inneholder store mengder NMI-FA og ether-lipider
FAME er analysert med standard metoden
Tabell 8: Metode modifikasjoner for fettsyremetylesterne (FAME) og fettalkoholene (FAOH) analyse.
Siden korallprøvene inneholder store mengder voksestere, ble FAME og fettalkoholene (FAOH) fraksjonert på en fastfasekolonne (500 mg aminopropyl-SPE, Supelco). FAME- og FAOH-fraksjonene ble analysert separat på GC-FID for å unngå koekluering. Korallprøvene ble først metylert, og de resulterende heksanekstraktene fra den direkte metanolysen ble tilsatt nonadekanol (19:0 alkohol) som intern standard for FAOH-ene. Heksanekstraktet ble lastet på SPE-kolonnen, og FAME-fraksjonen ble eluert med 3 mL heksan + 2 mL heksan:etylacetat (9:1 v/v), og fettalkoholene ble eluert med 4 mL kloroform. De to fraksjonene ble inndampet til tørrhet og løst opp i heksan før GC-FID-analyse.
FAME/FAOH-ekstraktene fra sjøtre og FAME ekstraktene fra viftesvamp og bergskjell ble analysert ved hjelp av en CP-Wax 52 CB-kolonne (25 m x 0,25 mm x 0,25 μm filmtykkelse) (Varian-Chrompack, Middelburg, Nederland). Ovnstemperaturen ble programmert til: 90 °C i 2 min, deretter fra 90 °C til 150 °C ved 30 °C/min og deretter til 225 °C ved 2,5 °C/min og holdt der i 35 minutter, for en total analysetid på 75 min. Helium (99,9999 %) ble brukt som mobil fase ved 1 mL·min i 45 min, etterfulgt av en økning til 3 mL/min som ble holdt i 30 min. Injektor- og detektortemperaturene var henholdsvis 270 °C og 300 °C. FAME-ekstraktet fra Kålrabisvamp ble analysert på Agilent 122-5062 DB-5 (60 m x 0,25 mm x 0,25 μm) med helium som bærergass med en konstant hastighet på 1 mL·min. Ovnstemperaturen ble holdt på 60 °C og økt til 130 °C med en hastighet på 60 °C/min, deretter økt til 325 °C med en hastighet på 1 °C/min og holdt ved denne temperaturen i 7 min. Injeksjons- og deteksjonsforholdene var som beskrevet ovenfor. Kromatogrammene ble integrert ved hjelp av EZChrom Elite-programvaren (Agilent Technologies). Identifisering av FAME og andre lipider ble utført ved å sammenligne retensjonstider med en FAME-standard (GLC-463 fra Nu-Chek Prep. Elysian, MN, USA) utført under lignende kromatografiske forhold som GC-FID (Wasta og Mjøs, 2013). Gasskromatografi-massespektrometri-analyse av alle prøver ble utført ved bruk av elektronstøtionisering (Agilent Technologies, 6890N/5975B GC-MS-system, Santa Clara, CA, USA).
Visualisering av lipidklassene til utvalgte lipidekstrakter (ett ekstrakt av hver art plukket i nærheten av oppdrettsanlegget og på referansestasjon) ble utført ved hjelp av tynnsjiktskromatografi (HPTLC) i henhold til metodene beskrevet i Olsen og Henderson (1989). HPTLC-platene ble utviklet med to mobile faser; først ble en polar fase (metylacetat: isopropanol: kloroform: metanol: 0,25 % KCl (25:25:25:10:9)) brukt til en høyde på 5 cm fra origo. Platen ble deretter tørket i en eksikkator i 30 min og utviklet i det andre (nøytrale) elueringsmiddelet (heksan:dietyleter:eddiksyre (80:30:2 v/v til toppen av platen). De separerte lipidklassene ble detektert ved å spraye platen med 3 % kobberacetat i 8 % fosforsyre, etterfulgt av oppvarming ved 160 °C i 20 min.
Totalt lipidinnhold (mg lipid per 100 mg tørrvekt) ble analysert ved hjelp av generaliserte lineære modeller (GLM) med Gamma-fordeling og log-link. Avstand til oppdrettsanlegg ble delt inn i tre grupper: < 250 m, 250–750 m og > 750 m. Sedimentasjon ble delt inn i tre grupper basert på akkumulert sedimentasjonsmengde: < 50 g·m⁻², 50–1000 g·m⁻² og >10 g·m⁻². For viftesvamp, sjøtre og bergskjell ble både avstand og sedimentasjonsgruppe inkludert som forklaringsvariabler, mens for kålrabisvamp var disse to faktorene sammenfallende, og kun avstandsgruppe ble brukt. For bergskjell ble vevstype (gjeller eller hel kropp) inkludert som en ekstra faktor. Post-hoc sammenligninger ble gjort med Tukey-korrigerte estimerte marginale middelverdier.
Resultater
Totalt lipidinnhold og terrestriske fettsyremarkører
Totalt lipidinnhold hos bergskjell var sterkt påvirket av vevstype, der hele kroppen hadde signifikant høyere lipidinnhold enn gjeller (p < 0,001). Avstand til oppdrettsanlegg viste en moderat effekt i den globale modellen, med høyere lipidinnhold ved mellomliggende avstander (250–750 m; p = 0,032), men parvise sammenligninger mellom avstandskategorier var ikke signifikante etter Tukey-justering (p > 0,07). Sedimentasjon hadde derimot en tydelig effekt, der lokaliteter med høy sedimentasjon (> 1000) hadde signifikant høyere lipidinnhold enn lokaliteter med moderat sedimentasjon (50–1000; p = 0,015). Dette indikerer at sedimentasjon er en viktig miljøgradient for lipidakkumulering hos bergskjell, mens avstandseffektene fremstår mer gradvise. For sjøtre ble det ikke påvist noen signifikant effekt av avstand til oppdrettsanlegg på totalt lipidinnhold (p = 0,94). Sedimentasjon viste en svak, men ikke-signifikant trend mot høyere lipidinnhold ved høy sedimentbelastning (> 1000) sammenlignet med lave nivåer (< 50; p = 0,082). Samlet sett tyder dette på at lipidinnholdet hos sjøtre i liten grad påvirkes av avstand til anlegg, og kun svakt av sedimentasjon. For viftesvamp var totalt lipidinnhold ikke signifikant påvirket av avstand til oppdrettsanlegg (p = 0,41). Sedimentasjon hadde en signifikant negativ effekt i den globale modellen for moderat sedimentasjon (50–1000) sammenlignet med lav sedimentasjon (< 50; p = ,032), men ingen parvise sammenligninger var signifikante etter Tukey-justering (p > 0,07). Dette indikerer en mulig, men svak sammenheng mellom sedimentasjon og lipidinnhold hos viftesvamp. For kålrabisvamp ble det ikke påvist noen signifikant effekt av avstand til oppdrettsanlegg på totalt lipidinnhold (p = 0,46), noe som tyder på at lipidinnholdet er relativt stabilt på tvers av avstandskategorier innenfor det undersøkte området.
For å undersøke variasjon i summen av terrestriske fettsyrer (18:1n-9, 18:2n-6, 18:3n-3), fettsyrer som ofte forekommer i laksefôr, ble generaliserte additive modeller (GAM) med Gamma-fordeling og log-link tilpasset ved hjelp av pakken mgcv i R. Modellene inkluderte smooth terms for avstand til nærmeste oppdrettsanlegg, dybde og akkumulert sedimentasjonsrate, der alle smooth terms ble spesifisert med k = 3 for å begrense modellens kompleksitet og unngå overtilpasning. For bergskjell ble vevstype også inkludert som en fast faktor.
Figur 27: Predikert relativ andel terrestriske fettsyremarkører i kålrabisvamp og viftesvamp som funksjon av miljøvariabler. Heltrukne linjer viser GAM-prediksjoner, og skraverte områder angir 95 % konfidensintervaller. Punktene viser observerte data. Modellene inkluderte smooth terms for avstand til oppdrettsanlegg, sedimentasjon og dybde, og ble tilpasset med Gamma-fordeling og log-link. Panelene A, C og E viser kålrabisvamp for henholdsvis avstand til oppdrettsanlegg, sedimentasjon og dybde, mens panelene B, D og F viser viftesvamp for de samme variablene i samme rekkefølge. Forklart variasjon og p-verdier for hver glatt funksjon er vist i panelene.
For de terrestriske fettsyrene ble det ikke funnet signifikante sammenhenger med avstand eller sedimentasjon hos kålrabisvamp (alle p > 0,1) (Figur 27). Hos viftesvamp var høyere sedimentasjon (p = 0,017) og større dybde (p = 0,049) assosiert med økte nivåer av terrestriske fettsyrer (Figur 27). Hos sjøtre påvirket både avstand til oppdrett (p = 0,035) og sedimentasjonsrate (p = 0,011) fettsyreinnholdet, med forhøyede nivåer nær oppdrettsanlegg og ved høy sedimentasjon (Figur 28). Hos bergskjell var nivået av terrestriske fettsyrer høyere i hele kroppen enn i gjeller (p < 0,001) og økte med sedimentasjonsrate (p = 0,023), mens avstand ikke hadde signifikant effekt (Figur 28). Selv om terrestriske fettsyrer ble påvist i alle arter, var de observerte andelene relativt lave sammenlignet med nivåer som forventes i laksefôr, der disse fettsyrene vanligvis utgjør omkring 30 % av total lipidmengde.
Figur 28: Predikert relativ andel terrestriske fettsyremarkører i sjøtre og bergskjell (gjeller og hele kroppen) som funksjon av miljøvariabler. Heltrukne linjer viser GAM-prediksjoner, og skraverte områder angir 95 % konfidensintervaller. Punktene viser observerte data. Modellene inkluderte smooth terms for avstand til oppdrettsanlegg, sedimentasjon og dybde, og ble tilpasset med Gamma-fordeling og log-link. Panelene A, C og E viser sjøtre for henholdsvis avstand til oppdrettsanlegg, sedimentasjon og dybde, mens panelene B, D og F viser bergskjell for de samme variablene i samme rekkefølge. Forklart variasjon og p-verdier for hver glatt funksjon er vist i panelene.
Totalt lipidinnhold viste begrenset variasjon med avstand fra oppdrett, men økte noe med høyere sedimentasjon, særlig hos bergskjell. Terrestriske fettsyrer karakteristiske for laksefôr ble påvist i alle arter, men i lav konsentrasjon, noe som indikerer begrenset opptak av oppdrettsrelatert materiale. Samlet tyder resultatene på at organisk tilførsel fra oppdrett kan påvirke lipidsammensetning lokalt, men den biokjemiske påvirkningen på disse bunnartene er generelt svak.
Fettsyresammensetning per art
For å teste romlige og miljømessige effekter på FA-sammensetning, ble generaliserte lineære modeller (GLM) med gammafordeling og log-lenke benyttet. For kålrabisvamp inkluderte modellen fettsyretype og avstand fra oppdrettsanlegg. For viftesvamp og sjøtre ble både avstand og akkumulert sedimentasjonsrate inkludert i modellen. Post-hoc sammenligninger (estimerte marginale midler) ble brukt for å identifisere signifikante forskjeller mellom avstands- eller sedimentgrupper (p < 0.05).
Fettsyreprofilen hos kålrabisvamp var dominert av forgrenede og enumettede fettsyrer (f.eks. i-15:0, ai-15:0, 16:1n-7 og Br 19:0 (11Me)) (Figur 29). Mettede fettsyrer (ΣSFA) utgjorde omtrent 45 % av de totale fettsyrene, hvorav bakterielt avledede SFA sto for ca. 38 %. Enumettede fettsyrer (ΣMUFA) bidro med 22–23 %, med bakterielle MUFA som utgjorde omtrent halvparten av denne fraksjonen (11–12 %). Flerumettede fettsyrer (ΣPUFA) utgjorde ca. 32 % av totalen, dominert av ikke-metabolske (NMI) fettsyrer (ca. 27–28 %), mens essensielle n-3 og n-6 PUFA var lave (< 4 %) (Vedlegg A5).
GLM-analysen viste at enkelte fettsyrer, som Br 17:0 (9Me), 22:0, i-23:0 og 26:3Δ5,9,19 NMI, forekom i høyere andeler nær oppdrettsanlegg (< 250 m) sammenlignet med > 750 m, mens den generelle fettsyresammensetningen var relativt upåvirket av avstand (Figur 29). Samlet sett viser resultatene at kålrabisvamp har en stabil, bakteriedominert fettsyresammensetning, noe som reflekterer artens høye mikrobielle tetthet (HMA) og sterke avhengighet av symbiotiske bakterier som energikilde.
Figur 29: Predikert fettsyreprofil (% av totale fettsyrer) for kålrabisvamp ved ulike avstander fra oppdrettsanlegg (< 250 m og > 750 m). Søylene viser modell-predikerte middelverdier (± 95 % konfidensintervall) fra en generalisert lineær modell (GLM) med Gamma-fordeling og log-link. Stjerner (*) markerer fettsyrer med signifikante forskjeller mellom avstandsgrupper (p < 0,05; Tukey-post hoc-test). Kun fettsyrer som bidrar med mer enn 1 % av de totale fettsyrene i minst én av de to avstandsgruppene, er vist i figuren.
Fettsyreprofilen hos viftesvamp bestod av både bakterielle og svampespesifikke non-methylene interrupted (NMI) fettsyrer, samt langkjedede enumettede fettsyrer (Figur 30). Totalt lipidinnhold var 3,02 ± 1,33 % nær oppdrettsanlegg (< 250 m) og 3,91 ± 0,93 % ved referansestasjoner (> 750 m). Mettede fettsyrer (∑SFA) utgjorde 33,7 ± 11,7 % nær anlegg og 25,8 ± 0,6 % ved referansepunktene, med 18:0 som økte markant i svært organiske sedimenter. Enumettede fettsyrer (∑MUFA) utgjorde 30,8 ± 4,6 % og 35,1 ± 2,6 %, inkludert langkjedede MUFA som 22:1n-7 og 24:1n-7 (Vedlegg A5). NMI-fettsyrer, inkludert 25:2D5,9 og 26:2D5,9, utgjorde 7–8 % av total fettsyremengde. Flerumettede fettsyrer (∑PUFA) var dominert av n-3-fettsyrer (20,8–24,0 %), mens n-6 bidro med 4–5 %.
Både sedimentets organiske innhold og avstand til oppdrettsanlegg påvirket fettsyresammensetningen. Svært organiske sedimenter (> 1000 g·m⁻²) var assosiert med høyere andeler av 18:0 og 18:1n-9 og lavere andeler av ai21:0, mens avstandseffektene var mer moderate, men signifikante for ai21:0, 24:1n-9, 18:2n-6 og 18:4n-3. Samlet tyder fettsyresammensetningen på at viftesvamp har lav mikrobiell tetthet (LMA), og at lipidprofilen i noen grad formes av miljøfaktorer, særlig sedimentets organiske innhold.
Figur 30: Predikert fettsyreprofil (% av totale fettsyrer) for viftesvamp som funksjon av (A) avstand til nærmeste oppdrettsanlegg og (B) akkumulert sedimentasjonsrate (g·m⁻²). Søylene viser modell-predikerte middelverdier (± 95 % konfidensintervall) fra en generalisert lineær modell (GLM) med Gamma-fordeling og log-link. Panel A viser forskjeller mellom avstandsgrupper (< 250 m og > 750 m), mens panel B viser forskjeller mellom sedimentasjonsgrupper (< 50, 50–1000, > 1000 g·m⁻²). Ulike bokstaver (B) eller stjerner (A) angir signifikante forskjeller mellom grupper (p < 0,05; Tukey post hoc-test). Kun fettsyrer som bidrar med mer enn 1 % av de totale fettsyrene i minst én av de to gruppene, er vist i figuren.
Fettsyreprofilen til sjøtre var dominert av langkjedede enumettede fettsyrer, særlig 20:1n-9 og 22:1n-11, og flerumettede fettsyrer som 20:4n-6, 20:5n-3 og 24:6n-3 (Figur 31). Verken avstand til oppdrettsanlegg eller sedimentets organiske innhold hadde signifikant effekt på den generelle fettsyresammensetningen, selv om enkelte små variasjoner forekom. Sjøtre hadde også et betydelig innhold av eterlipider (4–6 % av totale lipider), noe som er typisk for nesledyr, og FAOH-profilen viste at de samme langkjedede enumettede fettsyrene dominerte blant alkoholer (Figur 32), særlig 20:1n-9 og 22:1n-11, som utgjorde over 60 % av totale FAOH.
Blant PUFA var n-6-fettsyrer, spesielt kombinasjonen 20:4n-6 + 24:5n-6, bemerkelsesverdig høy, noe som kan reflektere spesifikke fysiologiske eller økologiske tilpasninger hos arten. Samlet viser fettsyre- og FAOH-profilen at sjøtre har en karakteristisk lipidprofil med langkjedede MUFA og n-6 PUFA, supplert med betydelige mengder eterlipider og alkohol-lipider (Vedlegg A5), uten tydelige effekter fra oppdrettsanlegg eller det organiske innholdet av sedimentasjonen.
Figur 31: Predikert fettsyreprofil (% av totale fettsyrer) for sjøtre som funksjon av (A) avstand til nærmeste oppdrettsanlegg og (B) akkumulert sedimentasjonsrate (g·m⁻²). Søylene viser modell-predikerte middelverdier (± 95 % konfidensintervall) fra en generalisert lineær modell (GLM) med Gamma-fordeling og log-link. Panel A viser forskjeller mellom avstandsgrupper (< 250 m og > 750 m), mens panel B viser forskjeller mellom sedimentasjonsgrupper (< 50, 50–1000 og > 1000 g·m⁻²). Stjerner angir signifikante forskjeller mellom grupper (p < 0,05; Tukey post hoc-test). Kun fettsyrer som bidrar med mer enn 1 % av de totale fettsyrene i minst én av de to gruppene, er vist i figuren.
Figur 32: Predikert profil av fettspritalkoholer (FAOH, % av totale alkoholer) i sjøtre som funksjon av (A) avstand til nærmeste oppdrettsanlegg og (B) akkumulert sedimentasjonsrate (g·m⁻²). Søylene viser modell-predikerte middelverdier (± 95 % konfidensintervall) fra en GLM med Gamma-fordeling og log-link. Ingen signifikante forskjeller ble funnet mellom grupper (p > 0,05). Kun fettsyrer som bidrar med mer enn 1 % av de totale fettsyrene i minst én av de to gruppene, er vist i figuren.
Fettsyreprofilen til bergskjell var tydelig påvirket av vevstype (hele kroppen uten gjeller eller kun gjeller), mens miljøfaktorer påvirket kun enkelte fettsyrer. Blant MUFA ble 20:1n‑7 påvist i lavere nivåer på lokaliteter nær oppdrettsanlegg (< 250 m) sammenlignet med mellomliggende lokaliteter (250–750 m), mens 18:3n‑3 var lavere på mellomliggende lokaliteter sammenlignet med referanselokaliteter (> 750 m) (Figur 33). Sedimentasjon påvirket også fettsyresammensetningen; nivåene av 20:1n‑7, 20:2n‑6 og 20:4n‑3 var lavere ved lav sedimentasjon (< 50 eller 50–1000 g·m⁻²) sammenlignet med høy sedimentasjon (> 1000 g·m⁻²).
Langkjedede MUFA som 20:1n‑9 og 22:1n‑11, markører for Calanus, var rikelige og konsistente på tvers av lokaliteter, noe som tyder på at bergskjell akkumulerer lipider fra zooplankton. Generelt var n‑3 PUFA dominerende, spesielt 20:5n‑3 og 22:6n‑3, mens n‑6 PUFA (20:4n‑6 og 20:2n‑6) ble påvirket av både avstand til anlegg og sedimentinnhold.
Metta fettsyrer (∑SFA) var høyest nær anlegg (21,2 ± 1,7 %), og MUFA dominerte den samlede profilen (∑MUFA ca. 41–42 %), med langkjedede MUFA fra Calanus som bidro betydelig. Totalt PUFA‑innhold var 32,9–34,9 %, der n‑3 PUFA representerte hoveddelen (29–30 %). Mindre bidrag fra NMI‑ og hydroksylerte fettsyrer ble observert, men de holdt seg stabile på tvers av lokaliteter (Vedlegg A5).
Oppsummert viser bergskjell en vevsspesifikk lipidprofil dominert av n‑3 PUFA og langkjedede MUFA, inkludert zooplankton‑avledede 20:1n‑9 og 22:1n‑11. Miljøfaktorer modulerer nivåene av enkelte fettsyrer, særlig 20:1n‑7, 20:2n‑6 og 20:4n‑3, uten at den generelle fettsyresammensetningen endres dramatisk.
Figur 33: Predikert fettsyreprofil (% av totale fettsyrer) i bergskjell (hel kropp) som funksjon av (A) avstand til nærmeste oppdrettsanlegg og (B) akkumulert sedimentasjonsrate (g·m⁻²). Søylene viser modell-predikerte middelverdier (± 95 % konfidensintervall) fra en generalisert lineær modell (GLM) med Gamma-fordeling og log-link. Bokstaver over søylene indikerer grupper som er statistisk forskjellige (Tukey post hoc-test, p < 0,05). Kun fettsyrer som bidrar med mer enn 1 % av de totale fettsyrene i minst én av de to gruppene, er vist i figuren.
Analyse av fettsyresammensetningen i fosfolipidfraksjonen hos bergskjell i forhold til lysosommembranstabilitet (LMD) viste ingen signifikante sammenhenger (Vedlegg A5). Beta-regresjonsmodeller for individuelle n-3 PUFAer (20:5n-3 og 22:6n-3), total PUFA, total n-3 PUFA og total n-6 PUFA viste gjennomgående ikke-signifikante effekter (p = 0,57–0,97). Modellene forklarte svært liten andel av variasjonen i LMD, med forklart devians mellom 0,7 og 4,4 % og negative justerte R²-verdier. Dette indikerer at variasjon i disse fosfolipidfettsyrene ikke bidrar til å forklare variasjonen i lysosommembranstabilitet hos bergskjell i dette datasettet.
3.10 - Molekylære multi-omics-analyser
Tradisjonelle metoder basert på morfologisk identifisering og miljøparametere kan være utilstrekkelige for å fange opp subletale, artsavhengige fysiologiske responser på miljøforstyrrelser. Multi-omiske tilnærminger, som integrerer profilering av mikrobielle samfunn, transkriptomikk, epigenomikk og cellulære biomarkører, kan derfor gi et mer helhetlig og sensitivt rammeverk for å karakterisere hvordan organismer responderer på miljøstressorer på flere biologiske nivåer, fra genuttrykk til holobiont-sammensetning. Genomiske og molekylære verktøy er viktige for standardisert konsekvensvurdering på tvers av marine taksa, særlig for fastsittende organismer som ikke kan unngå stressorer atferdsmessig, men må respondere gjennom fysiologisk og molekylær plastisitet (Oppen og Coleman, 2022; Vellani mfl., 2025). Utvikling av slike molekylære indikatorer for kaldtvannskoraller, svamper og tilhørende samfunn er derfor et viktig steg mot etablering av robuste og økologisk relevante overvåkingsverktøy tilpasset hardbunnshabitater.
Vevs- og slimprøver (fra sjøtre, risengrynkorall og viftesvamp) ble bevart ved hurtig frysing i flytende nitrogen og lagret ved -80 °C. Vevs- og slimprøvene ble ekstrahert for både DNA og RNA i henhold til protokollen beskrevet av Gomes mfl. (2025). For slimprøver ble 100 µl brukt til ekstraksjonen.
3.10.1 - Sekvensering 16s rRNA
Mikrobiomet hos koraller og svamper er følsomt for miljøforstyrrelser, og endringer i mikrobielle samfunnsstrukturer er dokumentert under termisk stress, sedimentasjon og næringsbelastning. Hos kaldtvannskorallene øyekorall og sikksakkorall (Madrepora oculata) responderte holobiontene på forhøyet temperatur med omstrukturering av mikrobiomet før det var mulig å påvise fysiologisk svekkelse, inkludert artsavhengig tap av deres karakteristiske bakterielle signaturer ved ekstreme temperaturer (Chapron mfl., 2021). Dette fremhever mikrobiomet som en tidlig varslingsindikator for stress. Tilsvarende har varmestress hos svamper vist å øke den relative forekomsten av forbigående og opportunistiske bakterier, der graden av dysbiose varierer mellom vertsarter og opprinnelig habitat (De Castro-Fernández mfl., 2023).
I sammenheng med akvakulturpåvirkning har 16S rRNA-metabarkoding blitt brukt til å karakterisere funksjonell mikrobiell omsetning hos suspensjonsspisere eksponert for utslipp fra lakseoppdrett, inkludert en økning av svovel- og nitrogenomsetningsbakterier samt taksa med evne til nedbrytning av xenobiotika i W. bursa nær oppdrettsanlegg (Laroche mfl., 2021). Tilsvarende er det rapportert redusert mikrobiomdiversitet og forekomst av fiskepatogener i Patella caerulea som lever på akvakulturbur (Palladino mfl., 2021). Sammensetningen av mikrobiomet hos oktokoraller har videre vist seg å reflektere lokale menneskeskapte påvirkninger, der vertsartens identitet og genetiske differensiering påvirker graden av mikrobiomets respons på miljøforstyrrelser (Reigel og Hellberg, 2023). Samlet sett understøtter disse studiene bruk av 16S rRNA-sekvensering som et effektivt verktøy for å påvise subletale, artsavhengige responser på miljøstressorer hos bentiske virvelløse dyr.
For å forstå hvordan bentiske mikrobielle samfunn responderer på miljøforhold og potensielle påvirkninger fra fiskeoppdrett, analyserte vi 16S rRNA-gensekvenser for å karakterisere den taksonomiske sammensetningen og mangfoldet i vertstilknyttede og SIBS-avledede mikrobielle samfunn, vurdere differensiell forekomst av bakterier langs miljøgradienter (avstand fra anlegg, sedimentasjon og dybde), identifisere felles og unike taxa mellom verter og sedimenter, og evaluere forekomsten av potensielle patogener.
Dataanalyse
Mikrobielle samfunn i sjøtre (vev og slim), risengrynkorall (vev og slim), viftesvamp (vev og slim), kålrabisvamp (vev), bergskjell (gjellevev) og SIBS-prøver ble analysert med 16S rRNA-gen ampliconsekvenser prosessert i phyloseq. For analyser basert på avstand til oppdrett ble prøver gruppert etter art, prøvetype og avstandsgruppe (< 250 m, 250–750 m, > 750 m), og for sedimentasjonsanalyser etter art, prøvetype og sedimentasjonskategori (< 50, 50–1000, > 1000 g·m⁻²). Sammenhenger mellom mikrobielle taxa (familie- og slektsnivå) og miljøgradienter (avstand til oppdrett, akkumulert sedimentasjon, dybde) ble testet med lineære modeller (LM) i microViz, inkludert kun taxa til stede i ≥ 5 % av prøvene, med FDR ≤ 0,05 som signifikansgrense. Tilstedeværelse/fravær-matriser for familier og ASV-er ble brukt til å kvantifisere felles og unike taxa mellom arter og SIBS-prøver, visualisert med Venn-diagrammer laget i eulerr, mens effekstørrelser for signifikante taxa ble vist i barplott. Potensielle patogen-ASVer ble identifisert fra vev-, slim- og SIBS-prøver ved å søke mot MBPD-databasen (multiple bacterial pathogen detection pipeline and database; https://github.com/LorMeBioAI/MBPD) ved bruk av vsearch versjon 2.22.1 (Rognes mfl. , 2016) og med parametere som beskrevet i Yang mfl. (2023). Kun ASVer med 100 % likhet med sekvenser i databasen ble betraktet som potensielle patogener og beholdt for videre analyser. Denne terskelen ble valgt basert på Edgar (2018) for å minimere falske positive. Sammenhengen mellom de foreslåtte patogenene og dybde, sedimentasjon og avstandskategori ble vurdert med generaliserte lineære modeller (GLM) med binomial logit-lenke, med separate modeller for vevs- og slim-/SIBS-prøver. Alle analyser ble utført i R versjon (4.4.3) ved bruk av pakkene phyloseq, microViz, dplyr, tidyr og ggplot2.
Resultater
Taksonomisk sammensetning
På tvers av alle arter var de mikrobielle samfunnene dominert av Proteobacteria, Bacteroidetes og flere andre funksjonelt relevante, tallrike linjer gruppert under «others». Det ble observert merkbare artsspesifikke mønstre: vev fra sjøtre hadde høy relativ forekomst av Spirochaetota, mens vev fra risengrynkorall var beriket med Pseudomonadota. Bergskjell viste berikelse av Campylobacterota, og kålrabisvamp hadde høyere relativ forekomst av Chloroflexi når prøvene ble gruppert etter avstand til anlegg (Figur 34).
Figur 34: Relativ forekomst av bakterielle taxa i henholdsvis slim og vev fra sjøtre, risengrynkorall, viftesvamp, kålrabisvamp og bergskjell, samt i SIBS-prøver (sedimentprøver). Stolpediagrammene er facetert etter art og prøvetype. Farger viser bakterielle rekker og familier, med fylumoverskrifter i fet skrift. Avstandsgrupper: < 250 m, 250–750 m, > 750 m.
Kun mindre variasjon i relativ forekomst ble observert mellom avstandskategorier for de fleste arter og SIBS-prøver. Derimot strukturerte akkumulert sedimentasjon mikrobielle samfunn på tvers av nesten alle vertsarter og SIBS-prøver (Figur 35). De observerte forskjellene i taksonomisk sammensetning mellom avstands- og sedimentasjonsbaserte grupperinger reflekterer sannsynligvis både økologiske og metodiske faktorer. Avstand til anlegg representerer en romlig gradient som ikke nødvendigvis fanger opp lokal miljøvariasjon fullt ut, mens akkumulert sedimentasjon ble modellert for å predikere deponering av organisk materiale og utøver derfor sterkere selektivt press på mikrobielle samfunn. Følgelig varierer enkelte taxa mellom sedimentasjonsbaserte analyser og avstandsbaserte sammenligninger.
Figur 35: Relativ forekomst av bakterielle taxa i henholdsvis slim og vev fra sjøtre, risengrynkorall, viftesvamp, kålrabisvamp, bergskjell, samt i SIBS-prøver (sedimentprøver). Stolpediagrammene er facetert etter art og prøvetype. Farger viser bakterielle rekker og familier, med fylumoverskrifter i fet skrift. Sedimentasjonsgrupper: < 50 g·m⁻², 5–1000 g·m⁻², > 1000 g·m⁻².
Differensiell forekomst langs miljøgradienter
Antallet signifikante taxa på tvers av arter og prøvetyper er oppsummert i Vedlegg A6.
Effektene av avstand til oppdrettsanlegg var generelt svake (Figur 36). Hos kålrabisvamp økte Gaiellales (1 slekter/familier, effekt ca. 14,4). I vev fra risengrynkorall økte Arcobacteraceae (1 slekter/familier, effekt ca. 3,94), mens Flavicella gikk ned i sjøtre-slim (1 slekter, effekt ca. ‑3,54). Viftesvamp og bergskjell viste ingen signifikante taxa knyttet til avstand, og mikrobielle samfunn i SIBS-prøver ble upåvirket.
Figur 36: De viktigste differensielt forekommende taxa per art og prøvetype i forhold til avstand fra anlegget. Stolpene viser effekten av avstand på mikrobiell forekomst. Positive verdier indikerer økt relativ forekomst, negative verdier redusert relativ forekomst. Kun taxa med statistisk signifikant justert p-verdi (padj ≤ 0,05) er vist. Panelene er delt etter art og prøvetype.
Dybde påvirket mange taxa med moderate effekter (Figur 37). Slim fra viftesvamp hadde 35 slekter og 22 familier med positive helninger, mens vev viste 29 slekter og 20 familier som økte med dybde. Kålrabisvamp hadde 1 slekter/familier som responderte. Bergskjell hadde én taxon med negativ helning. SIBS-prøver hadde 172 taxa som responderte, for det meste med moderate positive effekter.
Figur 37: De viktigste differensielt forekommende taxa per art og prøvetype i forhold til dybde. Stolpene viser effekten av dybde på mikrobiell forekomst. Positive verdier indikerer økt relativ forekomst (grønne), negative verdier redusert relativ forekomst (oransje). Kun taxa med statistisk signifikant justert p-verdi (padj ≤ 0,05) er vist. Panelene er delt etter art og prøvetype.
Sedimentasjon var en tydelig driver for mikrobielle samfunn (Figur 38). Slim fra viftesvamp viste 114 slekter og 66 familier som responderte, mens vev viste 37 slekter og 29 familier. Risengrynkorall (vev og slim) hadde 24–25 taxa som responderte. Bergskjell viste kun berikelse av SUP05 og Helicobacteraceae, mens kålrabisvamp hadde økning i Gaiellales. Sjøtre-slim hadde én taxon som ble negativt påvirket, og SIBS-prøver viste 172 taxa med for det meste moderate positive responser.
Figur 38: De viktigste differensielt forekommende taxa per art og prøvetype i forhold til sedimentasjon. Stolpene viser effekten av akkumulerte sedimenter på mikrobiell forekomst. Positive verdier indikerer økt relativ forekomst, negative verdier redusert relativ forekomst. Kun taxa med statistisk signifikant justert p-verdi (padj ≤ 0,05) er vist. Panelene er delt etter art og prøvetype.
Samlet sett var sedimentasjon og dybde de sterkeste driverne for mikrobielle variasjoner på tvers av arter, mens avstand til anlegg hadde begrenset effekt. Viftesvamp og SIBS-prøver viste konsekvent høyest antall responsive taxa.
Felles og unike taxa
På familienivå var overlappet mellom vertsarter og SIBS moderat, men på ASV-nivå var overlappen generelt lav, noe som indikerer et kjerne-mikrobiom på familienivå, men høy habitatspesifikk ASV-diversitet (Vedlegg A6). SIBS-prøver gir en miljømessig mikrobiell baselinje, og viser at mens enkelte mikrobielle familier deles med vertsmikrobiomer, er den fine strukturen (ASV-er) i stor grad verts- eller habitatspesifikk (Vedlegg A6).
Denne separasjonen antyder at effekter av oppdrett på mikrobielt sedimentsamfunn kan delvis buffres i vertsmikrobiomer, men endringer i delte familier kan indirekte påvirke vert-tilknyttede mikrober, spesielt i slimlag eksponert mot omkringliggende sediment.
Patogenprevalens
Generelt var patogenprevalensen lav (median < 6 % av OTUs) (Vedlegg A6). Litt høyere forekomst ble observert i slimprøver fra begge koraller (sjøtre og risengrynkorall) og hos viftesvamp (både vev og slim). Kålrabisvamp hadde nesten ingen antatte patogener, med null oppdaget ved høyeste avstand fra anlegg og i laveste sedimentasjonskategori (Vedlegg A6). GLM-prediksjoner reflekterte arts-spesifikk variasjon, men ingen konsistent effekt av de testede miljøfaktorene (avstand, sedimentasjon eller dybde) (Figur 40).
Figur 39: Andel patogener (%) i prøver fra ulike arter og prøvetyper i forhold til avstand til anlegg. Bokser viser median og kvartiler; whiskers viser minimum og maksimum innenfor 1,5 × interkvartilområdet. Panelene er delt etter art og prøvetype.
Figur 40: Andel patogener (%) i prøver fra ulike arter og prøvetyper i forhold til akkumulert sedimentasjon (g·m⁻²). Bokser viser median og kvartiler; whiskers viser minimum og maksimum innenfor 1,5 × interkvartilområdet. Panelene er delt etter art og prøvetype.
3.10.2 - Transkriptomanalyse
RNA-sekvensering (RNA-seq) er en veletablert metode for å undersøke hvordan organismer responderer på miljøstress på nivået av genaktivitet, inkludert aktivering av stressbeskyttende mekanismer som varmesjokkproteiner og immunforsvar. Hos revbyggende koraller har transkriptomiske studier vist at stressresponser er sterkt arts- og populasjonsspesifikke, der kystnære og offshore-populasjoner av samme art kan vise markant ulike genuttrykksprofiler under termisk stress, noe som understreker betydningen av lokal tilpasning (Ruggeri mfl., 2023). Hos oktokoraller har kombinerte transkriptomiske og proteomiske analyser gitt mekanistisk innsikt i hvorfor enkelte taksa fremstår som mer robuste mot varme og havforsuring enn andre (Vargas mfl., 2022). I en akvakulturkontekst viste genuttrykksanalyser av svamper som ble translokert langs en avstandsgradient fra et oppdrettsanlegg redusert cellulær aktivitet og oppregulering av gener relatert til cellestress og apoptose hos individer nær anlegget, noe som representerer potensielle ikke-letale molekylære indikatorer på påvirkning (Laroche mfl., 2021). Mer generelt har fremskritt innen sekvenseringsteknologi muliggjort et skifte fra målrettede genmarkører til heltranskriptom-tilnærminger, noe som betydelig har forbedret oppløsningen i karakterisering av interaksjoner mellom organismer og miljø.
For å undersøke hvordan våre studerte arter og SIBS-prøver responderer på miljøforhold og potensielle påvirkninger fra fiskeoppdrett på nivå av genuttrykk, analyserte vi transkriptomsekvenser (mRNA) for å karakterisere funksjonelle profiler og differensielle transkripter mellom arter og miljøgradienter (avstand fra anlegg og sedimentasjon). Denne tilnærmingen gjør det mulig å utforske de molekylære mekanismene bak organismenes respons og å identifisere subtile funksjonelle endringer som kan ha oppstått.
Transkriptomikk – bibliotekpreparering og sekvensering
RNA-prøvene ble sendt til Novogene GmbH (Tyskland) for bibliotekpreparering og sekvensering. Et volum på 15 µL per prøve, med en konsentrasjon på minst ≥ 20 ng·µL⁻¹, ble sendt på tørris. Etter ankomst hos Novogene ble prøvene underlagt kvalitetskontroll (QC) før videre behandling, for å sikre at RNA-kvaliteten tilfredsstilte kravene til bibliotekpreparering og sekvensering. Budbringer-RNA (mRNA) ble renset fra total RNA ved hjelp av poly-T oligo-bundne magnetiske kuler. Etter fragmentering ble første tråd cDNA syntetisert med tilfeldige heksamer-primere, etterfulgt av syntese av den andre tråden. Biblioteket ble deretter ferdigstilt gjennom endereparasjon, A-tailing, adapterligering, størrelsesseleksjon, amplifikasjon og rensing.
Etter kvantifisering ble de totalt 42 bibliotekene poolet og sekvensert på en Illumina NovaSeq X Plus-plattform (paired-end 150 bp, PE150).
Bioinformatikk
Rå sekvensdata ble prosessert med Trimmomatic versjon 0.39 (Bolger mfl., 2014) for å fjerne Illumina-sekvenseringsadaptere og nukleotider med lav Phred-score (≤ 2), og deretter kvalitetsfiltrert og feilrettet med Rcorrector-programmet (Song og Florea, 2015).
For hver art ble et de-novo transkriptom konstruert ved hjelp av Oyster River Protocol (ORP) versjon 2.3.3 (MacManes, 2018), et benchmarket bioinformatikkprogram spesialisert for de-novo transkriptommontering og analyse. Fire ulike assemblyer ble opprettet gjennom ORP-protokollen: to med SPAdes (kmer 55 og 75) (Bankevich mfl., 2012), én med Trans-ABySS (kmer 32) (Robertson mfl., 2010), og én med Trinity (Grabherr mfl., 2011). Disse assemblyene ble deretter slått sammen ved bruk av ORTHOFUSE (MacManes, 2018), og kvaliteten ble evaluert både med TRANSRATE versjon 1.0.3 (Smith-Unna mfl., 2016) og Benchmarking sets of Universal Single-Copy Orthologs (BUSCO) versjon 5.1.2 (Seppey mfl., 2019) mot referansedatabasen mollusca_odb10.
Contigs ble annotert med ENTAP versjon 0.10.8 (Hart mfl., 2020) ved bruk av databasene EggNOG (Huerta-Cepas mfl., 2018), UniRef90 (Suzek mfl., 2014), RefSeq (O’Leary mfl., 2015) og UniProt (inkludert Swiss-Prot og TrEMBL) (Consortium, 2024). Sekvenser fra hver prøve ble mappet til det endelige de-novo transkriptomet, og kvantifisert med SALMON versjon 1.4 (Patro mfl., 2017).
Dataanalyse
Transkriptomiske data ble importert til RStudio (RCoreTeam, 2025) med pakken tximport (Soneson mfl., 2016). Sjeldne gener (definert som gener uten minst 10 reads i minst én prøve) ble fjernet fra videre analyser. Dataene ble først utforsket ved hjelp av hovedkomponentanalyse (PCA) på variansstabiliserte data med funksjonen vst i DESeq2 (Love mfl., 2014), samt Euclidisk ulikhet (avstand) beregnet med pakken vegan (Oksanen mfl., 2020), og visualisert med ggplot2 (Wickham, 2016). Effektene av sedimentasjon og dybde ble testet på ulikhetsmatrisen ved bruk av PERMANOVA (adonis2 i vegan, 999 permutasjoner, blokker = anlegg). Differensielt uttrykte transkripter og gener assosiert med sedimentasjon ble identifisert med DESeq2 og visualisert med stolpediagrammer. Signifikant berikede pathways (GO-termer; p-verdi ≤ 0.05) fra sedimentasjon ble identifisert ved hjelp av gene set enrichment analysis (GSEA) (Subramanian mfl., 2005), basert på gen-nivå data og funksjonen fgseaMultilevel i pakken fgsea (Korotkevich mfl., 2021) og visualisert med ggplot2.
Resultater
Differensielt uttrykte transkripter (DETs)
Antall differensielt uttrykte transkripter (DETs) per art som respons på sedimentasjon vises i (Figur 41). Sjøtre hadde det høyeste antallet DETs, etterfulgt av kålrabisvamp, viftesvamp og bergskjell. Både opp- og nedregulerte gener ble observert, men mønstrene varierte sterkt mellom artene, noe som indikerer arts-spesifikke responser.
Figur 41: Antall differensielt uttrykte transkripter (DET) i respons til sedimentasjon for de fire artene. Søylene viser antall opp- og nedregulerte transkripter for AE (bergskjell), GB (kålrabisvamp), PA (sjøtre) og PV (viftesvamp).
Transkriptomrespons på sedimentasjon
Sedimentasjon hadde størst effekt på viftesvamp transkriptom (R² = 0,41, p-verdi = 0,07), etterfulgt av bergskjell (R² = 0,14, p-verdi = 0,03), mens det samlede transkriptomet til både kålrabisvamp (R² = 0,12, p-verdi = 0,62) og sjøtre (R² = 0,09, p-verdi = 0,72) tilsynelatende ikke ble påvirket (Figur 42). Ingen signifikant effekt av dybde ble observert hos noen av artene.
Figur 42: Principal Component Analysis (PCA) av miljø- og biologiprøver fra ulike lokaliteter og sedimentasjonsgradienter. Panelene A) bergskjell, B) kålrabisvamp, C) sjøtre og D) viftesvamp viser de to første hovedkomponentene (PC1 og PC2), som forklarer den største variasjonen i hvert datasett. Punktene er farget etter sedimentasjonsrate (g·m⁻²) og formet etter lokalitet/farm. R²-verdier og p-verdier fra PERMANOVA-tester for effektene av sedimentasjon og dybde er angitt i hvert panel når tilgjengelig. Panelene sammenligner ulike lokaliteter og dybdegrupper, og illustrerer hvordan miljøgradienter og farmplassering påvirker prøvenes gruppering.
mRNA-pathways relatert til sedimentasjon
Responsen til funksjonelle veier på sedimentering varierte sterkt mellom de fire artene, og viste i stor grad arts-spesifikke molekylære tilpasninger (Figur 43). Kun regulering av intracellulær transport, var delt mellom viftesvamp og sjøtre (Figur 44).
Figur 43: Signifikante biologiske signalveier assosiert med sedimentasjon hos ulike arter. Forkortelser: AE = bergskjell, GB = kålrabisvamp, PA = sjøtre, PV = viftesvamp. Kun de 15 mest signifikante signalveiene er vist for hver art. Størrelsen på prikkene angir antall transkripter som er knyttet til hver signalvei – større prikker indikerer et høyere antall involverte transkripter.
Hos bergskjell ble flere metabolske og enzymatiske prosesser betydelig påvirket av sedimentasjon. Nedregulerte veier inkluderte dipeptidaseaktivitet, fosfolipaseaktivitet, negativ regulering av proteolyse og etermetabolisme, noe som tyder på redusert proteolytisk omsetning og lipidmetabolisme. Denne nedreguleringen kan reflektere energisparing eller lavere katabolsk aktivitet under sedimentstress. Oppregulerte veier var hovedsakelig relatert til polyaminmetabolisme og cellulær stressrespons, inkludert putrescin-biosyntese fra ornithin, ornithin-dekarboksylaseaktivitet og putrescinmetabolisme. Ytterligere oppregulering av ikke-kanonisk NF-κB-signalisering, peptidase-regulering og epitelcelle-differensiering støtter en immun- og reparasjonsrelatert respons. Berikelse av monosakkarid- og heksemetsabolisme antyder også en metabolsk omorganisering mot karbohydratbruk. Samlet tyder dette på at bergskjell tilpasser metabolismen for å opprettholde cellulær integritet og energibalanse fremfor å bredt undertrykke genaktivitet.
Hos kålrabisvamp var sedimentasjon assosiert med sterk oppregulering av proteinsyntese, translaterings- og membrantargeterte veier. Disse inkluderte cytosolisk ribosom, ribosomkomponenter, SRP-avhengig cotranslationell proteinsortering til membraner, og proteinlokalisering til endoplasmatisk retikulum. Dette mønsteret reflekterer aktivering av translateringsmaskineri og sekretorisk kapasitet, som muligens støtter cellulær fornyelse og stresskompensasjon. Berikelse av epitelial væsketransport og mRNA-katabolske prosesser indikerer koordinert kontroll av osmoregulering og mRNA-omsetning, noe som tyder på at kålrabisvamp responderer ved å styrke biosyntese og vedlikehold fremfor å undertrykke aktivitet.
Figur 44: UpSet-plot som viser overlapp av berikede biologiske prosesser mellom arter basert på transkriptomiske responser på predikert akkumulert sedimentasjon. Hver søyle representerer antall unike eller delte genontologi (GO)-termer for hver art (AE = bergskjell, GB = kålrabisvamp, PA = sjøtre, PV = viftesvamp). Kun én GO-term, «Regulering av intracellulær transport», var delt mellom sjøtre (PA) og viftesvamp (PV).
Hos sjøtre dominerte oppregulering av symbiont- og virusrelaterte veier, inkludert symbiontmediert påvirkning av vertsgeneuttrykk, undertrykkelse av vertstranslasjon, viral RNA-replikasjon og RNA-avhengig RNA-polymerase-aktivitet. Dette antyder at sedimentasjon kan forstyrre vert–symbiont- eller vert–virus-interaksjoner, muligens gjennom endringer i mikrobiomet eller viral aktivering. Berikelse av vertens immunmodulering og autophagy-induksjon av virus peker på stressinduserte interaksjoner mellom vertens immunforsvar og assosierte mikroorganismer.
Hos viftesvamp var sedimentasjon hovedsakelig assosiert med nedregulering av metabolske og signalveier, inkludert hedgehog-proteinbinding, CAMKK–AMPK-signalering, cAMP-avhengig kinaseaktivitet og lipidrelaterte prosesser som sphingolipid- og ceramidbiosyntese. Disse endringene indikerer redusert energisignalering og lipidmetabolisme under stress. Oppregulering av axonemal central pair projection kan reflektere forbedret cilievedlikehold eller partikkelrensingskapasitet.
3.10.3 - DNA-metyleringsanalyser
Epigenetiske mekanismer, særlig DNA-metylering, kan regulere hvordan gener slås av eller på som respons på miljøstress, og studeres i økende grad som en mekanisme for hvordan organismer akklimatiserer seg til endrede forhold. Hos koraller er det vist at mønstre i DNA-metylering endres systematisk i respons til sesongmessige temperaturvariasjoner, noe som indikerer at organismer registrerer og responderer på miljøsignaler på molekylært nivå (Rodríguez-Casariego mfl., 2020). Det finnes også økende dokumentasjon på at tidligere eksponering for stress kan påvirke hvordan en organisme responderer på senere stressorer, et fenomen kjent som miljøminne, og at DNA-metylering kan være en av de underliggende mekanismene (Hackerott mfl., 2021). Disse studiene tyder på at analyse av DNA-metylering kan være et nyttig verktøy for å påvise kumulative effekter av miljøeksponering hos langlivede, fastsittende organismer som kaldtvannskoraller og svamper, og dermed være relevant for vurdering av kroniske påvirkninger fra akvakulturaktiviteter.
For å undersøke potensielle epigenetiske responser ble DNA-metyleringsanalyser planlagt for tre arter: kålrabisvamp, viftesvamp og sjøtre. Totalt åtte prøver per art ble valgt ut, med fire prøver fra lokaliteter nær oppdrettsanlegg (< 250 m) og fire fra referanseområder (> 750 m). DNA ble først ekstrahert ved bruk av AllPrep DNA/RNA Mini Kit (Qiagen, Hilden, Germany). Alle prøver ble deretter pakket på tørris og sendt til Novogene for kvalitetskontroll og videre prosessering.
Kvalitetskontrollen viste at DNA fra kålrabisvamp hadde tilstrekkelig kvalitet for sekvensering, mens prøver fra sjøtre viste moderat degradering og prøver fra viftesvamp tegn til RNA-kontaminasjon og ekstra fragmenter. Et rensetrinn for RNA ble forsøkt, men førte til ytterligere degradering.
Et nytt forsøk på DNA-ekstraksjon ved bruk av QIAamp Fast DNA Kit (Qiagen), som inkluderer et RNA-rensetrinn, ble deretter gjennomført for alle arter. Dessverre viste disse prøvene alvorlig degradering og besto ikke kvalitetskontrollen. Kun prøvene fra kålrabisvamp fra den første ekstraksjonen ble derfor brukt til bibliotekpreparering og helgenom bisulfittsekvensering (WGBS).
Etter mottak av sekvenseringsdata viste kartleggingen imidlertid svært lav «mapping rate» (< 20 %) mot det tilgjengelige referansegenomet. Dette tyder på at referansegenomet ikke er tilstrekkelig representativt for arten, og at dataene derfor ikke egner seg for videre analyse i denne rapporten.
Selv om analysene ikke kunne fullføres, gir arbeidet verdifull innsikt i utfordringene knyttet til DNA-isolering fra svamper og koraller, samt behovet for høyoppløselig helgenomsekvensering av god kvalitet. En foreløpig analyse ble utført, men resultatene er usikre og bør tolkes med forsiktighet inntil et mer komplett og artsrelevant referansegenom blir tilgjengelig. Dette vil være nødvendig for å muliggjøre pålitelige epigenetiske analyser i fremtidige studier.
3.10.4 - Telomeranalyser
Telomerlengde er en veletablert biomarkør for cellulær aldring og fysiologisk tilstand hos virveldyr, og reflekterer kumulativ eksponering for oksidativt stress, immunaktivering og miljøstressorer (Salmón og Burraco, 2022). Anvendelsen på marine virvelløse dyr er imidlertid svært ny. Hos revbyggende koraller er det vist at telomer-DNA-lengde varierer systematisk med termisk regime, med distinkte regulatoriske mønstre mellom korall-slekter som gjenspeiler forskjeller i livshistorie: Telomerlengdene hos de kortlivede og stressfølsomme Pocillopora spp. var primært styrt av sesongmessige temperaturvariasjoner, mens de hos de langlivede og stresstolerante Porites spp. i større grad ble påvirket av historiske termiske anomalier (Rouan mfl., 2023). For langlivede bentiske arter som kaldtvannskoraller, svamper og muslinger representerer telomerlengde derfor en potensielt informativ, integrerende biomarkør for kronisk miljøpåvirkning, inkludert påvirkning fra akvakulturaktiviteter.
Metode
Genomisk DNA med høy molekylvekt fra sjøtre ble ekstrahert ved bruk av Qiagen Midi Kit for Cell and Blood Culture. Mellom 110–130 mg våtvekt vev ble benyttet per prøve. DNA ble eluert i QF-buffer i henhold til produsentens protokoll. DNA-konsentrasjon og renhet ble målt ved hjelp av NanoDrop, og DNA-kvalitet ble verifisert ved agarosegelelektroforese. Kun prøver med høy molekylvekt og akseptable absorbansforhold (A260/280 og A260/230) ble brukt videre i analysene.
Individuell telomerlengde ble estimert ved bruk av telomere restriction fragment (TRF)-metoden med TeloTAGGG assay kit (Sigma Aldrich, Roche), i henhold til produsentens instruksjoner. Kort fortalt ble 1 µg DNA fordøyd med restriksjonsenzymene HinfI og RsaI ved 37 °C i 3 timer, etterfulgt av elektroforese på 0,8 % agarosegel ved 50 V i 3 timer. Etter elektroforese ble DNA denaturert, nøytralisert og overført til en positivt ladet nylonmembran ved hjelp av Southern blot over natten. DNA ble deretter fiksert på membranen ved 120 °C i 20 min. Hybridisering ble utført med en digoksigenin-merket telomerprobe, etterfulgt av inkubasjon med et antidigoksigenin-spesifikt antistoff. Signalene ble visualisert med Chemidoc (Bio-Rad).
Telomerlengde ble beregnet ved hjelp av ImageJ, basert på tetthetsmålinger av telomersmøret. Bakgrunn ble korrigert per bane, og gjennomsnittlig telomerlengde ble beregnet innenfor et intervall på 2,7–21,2 kb, med ekstrapolering opp til 150 kb for å inkludere hele smøret. Gjennomsnittlig telomerlengde ble estimert for 28 prøver (område: 13–34 kb). Intra-assay variasjon ble beregnet ved bruk av tre replikatprøver per gel (CV: 5,0 %), og inter-assay repeterbarhet ble vurdert ved å inkludere tre prøver på begge geler (CV: 8,0 %).
Resultater
Statistiske analyser viste at telomerlengde hos sjøtre ikke var signifikant påvirket av avstand til oppdrettsanlegg (F = 2,81, p = 0,107) eller av akkumulert sedimentasjon (F = 0,91, p = 0,307), mens dybde viste en svak, men ikke-signifikant tendens (F = 3,60, p = 0,070) (Figur 45). Den generaliserte additive modellen forklarte 29,1 % av variasjonen i telomerlengde, med en justert R²-verdi på 0,176, noe som indikerer moderat forklaringskraft. Samlet sett tyder resultatene på at telomerlengde hos sjøtre ikke viser sterke responser på de undersøkte miljøgradientene, selv om dybde kan ha en viss innflytelse som bør undersøkes nærmere med et større datagrunnlag.
Figur 45: Predikert telomerlengde hos sjøtre (Paragorgia arborea) som funksjon av miljøvariabler. Heltrukne linjer viser prediksjoner fra generaliserte additive modeller (GAM), mens skraverte områder angir 95 % konfidensintervaller. Punktene viser observerte målinger av telomerlengde. Modellene inkluderte glatte funksjoner for avstand til oppdrettsanlegg, dybde og akkumulert sedimentasjon, og ble tilpasset med Gamma-fordeling og log-linkfunksjon. Panel A viser effekten av avstand til oppdrettsanlegg, panel B viser effekten av dybde, og panel C viser effekten av akkumulert sedimentasjon. Forklart variasjon og p-verdier for hver glatt funksjon er vist i hvert panel.
3.11 - Arts-spesifikke responser på tvers av datatyper og miljødrivere
For å gi en helhetlig vurdering av effektens av utslipp fra akvakulturens på kaldtvanns bunnarter benyttet vi en flertrinns analytisk tilnærming som tar høyde for at miljøstress kan manifestere seg ulikt mellom arter, og at enkeltmetoder kan overse viktige påvirkningsmekanismer eller overvekte kortvarige responser uten langsiktig økologisk betydning.
Analysen integrerte komplementære responskategorier med ulik sensitivitet, tidsintegrasjon og økologisk relevans. Visuell helsetilstand vurdering dokumenterte organismenes tilstand, vevsintegritet og synlige stressindikatorer som nekrose og sedimentdekning. Fysiologiske målinger hos risengrynkorall og sjøtre kvantifiserte oksygenforbruk og ammoniakkutskillelse som mål på metabolsk belastning. På cellulært nivå ble lysosomal membranstabilitet (særlig hos svamper og bergskjell) og telomerlengde (hos sjøtre) brukt som biomarkører for cellulært stress og akkumulert belastning. Biokjemiske analyser omfattet totalt lipidinnhold, fettsyreprofiler og terrestriske fettsyresignaturer som indikatorer på ernæringsstatus og mulig sedimentpåvirkning. Mikrobielle samfunn ble karakterisert ved 16S rRNA-sekvensering, mens RNA-seq ble brukt for å identifisere differensielt genuttrykk og aktiverte stressveier. For koraller ble histologiske analyser av kjønnsfordeling, eggstørrelse og fekunditet gjennomført for å vurdere reproduktive effekter. På økosystemnivå ble bunnsamfunn vurdert gjennom b-MBI basert på sedimentprøver.
Prøvene ble samlet langs gradienter av tre sentrale miljødrivere: avstand fra oppdrettsanlegg (30–1200 m), dybde (40–300 m) og modellert akkumulert partikulært organisk materiale over to år. Detaljerte artsvise responser presenteres i Tabell 9.
Art
Variabel
Avstand (til anlegg)
Dybde
Sedimentasjon
Kommentar
Sjøtre
Visual
0
−
0
Ingen tydelig visuell stressrespons
Histologi – Kjønn
0
0
N/A
Ingen respons
Histologi – Eggstørrelse
0
0
N/A
Stabil
O₂-forbruk
±
(−)
+
Ikke-lineær respons: høyest O₂-forbruk ved mellomavstand (30–600 m); økt metabolisme med sedimentasjon
NH₄⁺-eksresjon
0
0
0
Ingen tydelig respons
Totalt lipider
0
0
(+)
Svak økning ved høy sedimentasjon, mulig energilagring
Terrestriske fettsyrer
±
0
+
Ikke-lineær respons med avstand og ved høy sedimentasjon, lavt absoluttnivå ellers
Fettsyrer (profil)
0
N/A
(±)
Enkelte lipider redusert, andre økt ved høy sedimentasjon
16S – Samfunnsstruktur
Δ
N/A
Δ
Struktur endres
16S – Diff. forekomst
(−)
0
(−)
Kun én slekt (Flavicella) negativt påvirket av avstand
RNA-seq – DETs
N/A
N/A
0
Mange DETs, men svak samlet transkriptomrespons
RNA-seq – Veier
N/A
N/A
(+)
Oppregulering av stress-, immun- og symbiont/virus-relaterte veier
Telomerlengde
0
(+)
0
Svak trend mot lengre telomerer med dybde
Risengrynkorall
Visual
+
0
+
Økt visuell stress nær anlegg og ved sedimentasjon
Histologi – Kjønn
0
0
N/A
Stabil kjønnsfordeling
Histologi – Eggstørrelse
0
0
N/A
Stabil
Histologi – Fekunditet
0
0
N/A
Stabil
O₂-forbruk
0
0
0
Ingen tydelig respons
NH₄⁺-eksresjon
0
0
0
Ingen tydelig respons
16S – Samfunnsstruktur
Δ
N/A
Δ
Struktur endres
16S – Diff. forekomst
(+)
0
±
Moderat respons hos noen taxa på sedimentasjon; noen øker, andre reduseres
Kålrabisvamp
Visual
0
0
(−)
Svak visuell respons
Lysosomal membranendestabilisering
+
±
−
Sterk stressrespons; dybderespons modell-avhengig
Totalt lipider
0
0
0
Stabil
Terrestriske fettsyrer
0
0
0
Ingen respons
Fettsyrer (profil)
+
N/A
N/A
Høyere nivåer av Br 17:0 (9Me), 22:0, i-23:0 og 26:3 NMI nær anlegg
16S – Samfunnsstruktur
Δ
N/A
Δ
Struktur endres
16S – Diff. forekomst
(+)
(+)
(+)
Gaiellales øker med sedimentasjon
RNA-seq – DETs
N/A
N/A
+
Mange DETs assosiert med sedimentasjon
RNA-seq – Veier
N/A
N/A
+
Sterk oppregulering av translasjon, ribosom- og ER-relaterte veier
Viftesvamp
Visual
(+)
0
+
Økt stressrespons nær anlegg og ved sedimentasjon
Lysosomal membranendestabilisering
+
0
0
Moderat stressrespons nær anlegg
Totalt lipider
0
0
−
Reduserte lipidreserver ved høy sedimentasjon
Terrestriske fettsyrer
0
+
+
Øker med dybde og sedimentasjon
Fettsyrer (profil)
(+)
N/A
+
Miljøpåvirket sammensetning; sedimentasjon har sterkest effekt
16S – Samfunnsstruktur
Δ
N/A
Δ
Sterk strukturering av sedimentasjon
16S – Diff. forekomst
0
+
±
Mange taxa responderer på dybde og sedimentasjon; noen øker, noen reduseres
RNA-seq – DETs
N/A
N/A
+
Moderat–sterk transkriptomrespons på sedimentasjon
RNA-seq – Veier
N/A
N/A
−
Nedregulering av energi- og lipidmetabolisme; stress-tilpasning
Bergskjell
Visual
N/A
N/A
N/A
Begrenset visuell vurdering
Lysosomal membranstabilitet
+
±
0
Dybderespons med motsatt retning i ulike modeller
Totalt lipider
0
0
+
Økt lipidnivå ved høy sedimentasjon
Terrestriske fettsyrer
0
0
+
Sedimentasjon påvirker nivåer
Fettsyrer (profil)
+
N/A
+
20:1n-7 lavere nær anlegg; 3 FA øker med sedimentasjon
16S – Samfunnsstruktur
Δ
N/A
Δ
Struktur endres
16S – Diff. forekomst
0
(-)
(+)
SUP05 og Helicobacteraceae øker
RNA-seq – DETs
N/A
N/A
+
Færre DETs, men signifikant samlet transkriptomrespons
RNA-seq – Veier
N/A
N/A
±
Nedregulert metabolisme kombinert med oppregulert stress- og immunrespons
Tabell 9: Oversikt over biologiske responser hos ulike arter langs miljøgradienter nær oppdrettsanlegg. Symbolene indikerer retning og type respons, der 0 betyr ingen tydelig effekt, + representerer en positiv effekt eller økning, − angir en negativ effekt eller reduksjon, ± viser en ikke-lineær respons eller blandet effekt, Δ indikerer endret sammensetning, (−) eller (+) viser en svak negativ eller positiv trend, og N/A betyr ikke aktuelt eller data ikke tilgjengelig. Kommentarene i siste kolonne gir tolkning av effekter og mønstre observert i dataene.
For å sammenstille visuell helse og bunnsamfunnsdata i én integrert metrikk utviklet vi to varianter av en arts-spesifikk sammensatt påvirkningsindeks (Composite Impact Index) som «proof-of-concept»-tilnærminger. Dette representerer prosjektets første forsøk på å integrere flere signaler i samlede indikatorer, og indeksene må betraktes som arbeid under utvikling som krever betydelig videreutvikling og validering før eventuell operativ anvendelse. Begge tilnærminger baserer seg på visuell helseindeks beregnet utelukkende fra ROV-opptak, og representerer dermed ikke-invasive metoder.
Tilnærming 1: Like vekter (Figur 46). Den første indeksen kombinerer to sentrale overvåkingsmetrikker med lik vekting: visuell helsetilstandsindeks (som reflekterer direkte organismetilstand) og b-MBI (som representerer strukturen i bunnsamfunnet i nærheten av fokusartene). Integrasjonen bygger på en erkjennelse av at organismestress ofte opptrer parallelt med bredere degradering av bunnsamfunn, og at samtidig overvåking av begge nivåer gir en mer helhetlig vurdering av økosystemhelse. Alle metrikker ble standardisert innen art ved bruk av z-transformasjon for å muliggjøre sammenligninger på tvers av arter.
Figur 46: Sammensatt påvirkningsindeks for fire bunnlevende indikatorarter langs miljøgradienter fra oppdrettsanlegg. Indeksen integrerer visuell helsetlstandsindeks og bakteriell metabarcoding biotisk indeks (b-MBI) med like vekter (0,5/0,5) for alle arter. Generaliserte additive modeller (GAM) viser responser mot avstand til oppdrettsanlegg, dybde, og akkumulert sedimentasjon for sjøtre (rød, A-C), risengrynkorall (oransje, D-F), viftesvamp (brun, G-I), og kålrabisvamp (grå, J-L). Grå punkter representerer individuelle observasjoner, fargede linjer viser predikerte trender med 95% konfidensintervaller (skyggelagte områder). Forklart variasjon og p-verdier er angitt for hver smooth term. n = antall observasjoner per art.
Tilnærming 2: Fysiologisk kalibrert vekting (Figur 47). Den andre indeksen benytter arts-spesifikk vekting basert på korrelasjoner med historiske fysiologiske målinger. For svamper (viftesvamp, kålrabisvamp) ble vektene kalibrert mot lysosomal membranstabilitet (LMS), slik at visuell score tillegges høyere vekt når den korrelerer sterkt med cellulært stress. For koraller (sjøtre, risengrynkorall) ble vektene basert på gjennomsnittlige korrelasjoner med oksygenforbruk og ammoniakkutskillelse. Vekten til b-MBI i den kalibrerte tilnærmingen er beregnet som det resterende (1 − visuell vekten) og er dermed ikke uavhengig kalibrert mot fysiologiske målinger. Denne tilnærmingen sikrer at metrikker som er validert mot fysiologisk stress tillegges passende vekt for hver art. For å sikre at kun individer med tilgjengelige kalibreringsdata ble inkludert, ble datasettet begrenset til svamper med LMS-målinger og koraller med oksygenforbruk-data. Dette reduserte prøvestørrelsen sammenlignet med tilnærming 1, men ga en mer robust kalibrering.
Figur 47: Sammensatt påvirkningsindeks for fire bunnlevende indikatorarter med arts-spesifikk vekting basert på fysiologiske kalibreringer. For svamper (viftesvamp, kålrabisvamp) ble vekter beregnet fra korrelasjoner mellom visuell helsetlstandsindek og lysosomal membranstabilitet (LMS). For koraller (sjøtre, risengrynkorall) ble vekter basert på gjennomsnittlige korrelasjoner mellom visuell helsetlstandsindek og oksygenforbruk samt ammoniakkutskillelse. eneraliserte additive modeller (GAM) viser responser mot avstand til oppdrettsanlegg, dybde, og akkumulert sedimentasjon for sjøtre (rød, A-C), risengrynkorall (oransje, D-F), viftesvamp (brun, G-I), og kålrabisvamp (grå, J-L).
En sentral begrensning for begge tilnærminger var ufullstendig dekning av b-MBI-data. Dette ble håndtert ved å imputere manglende verdier ved hjelp av arts-spesifikke lineære modeller basert på miljøprediktorer (avstand, dybde og sedimentasjon). Selv om dette muliggjorde beregning av indeksverdier for alle lokaliteter, introduserer det økt usikkerhet fordi enkelte verdier er predikerte snarere enn observerte. De sammensatte indeksene ble modellert mot miljødriverne ved bruk av generaliserte additive modeller (GAM) for å håndtere ikke-lineære sammenhenger og kvantifisere effektstørrelser og tilhørende usikkerhet.
Sammenligning av tilnærminger. Begge indeksene viste varierende ytelse mellom arter, men med noen viktige forskjeller. Den like-vektede indeksen (Figur 46) ga større konsistens på tvers av arter og enklere tolkning, men skilte ikke mellom metrikkenes relative pålitelighet. Viftesvamp viste høy sensitivitet i begge tilnærminger (like vekter: R² = 0,51, 52,2 % forklart devians, n = 50; kalibrert: R² = 0,47, 53,9 %, n = 32), med sedimentasjon som dominerende driver (begge p < 0,001) og signifikant avstandseffekt (like vekter p = 0,005; kalibrert p = 0,012). Kålrabisvamp viste også høy sensitivitet i begge tilnærminger (like vekter: R² = 0,53, 58,7 %, n = 32; kalibrert: R² = 0,77, 81,2 %, n = 25), med signifikante effekter av både sedimentasjon (p < 0,001) og avstand i den kalibrerte tilnærmingen (p = 0,014).
For korallene var forskjellene mer markante. Risengrynkorall viste moderat sensitivitet med like vekter (R² = 0,24, 29,8 %, n = 63) med signifikant avstandseffekt (p = 0,003), men svakere respons med kalibrert vekting (R² = 0,13, 22,0 %, n = 40) uten signifikante effekter. Sjøtre viste bedre ytelse med kalibrert vekting (R² = 0,50, 57,0 %, n = 32) med signifikante effekter av både dybde (p < 0,001) og sedimentasjon (p = 0,006), sammenlignet med like vekter (R² = 0,27, 34,8 %, n = 47) hvor kun avstand (p = 0,026) og dybde (p = 0,004) var signifikante.
Den kalibrerte tilnærmingen ga høyere forklart variasjon for kålrabisvamp og sjøtre, men lavere for risengrynkorall. Korrelasjoner mellom visuell helsetilstandsindeks og metabolske parametere var svake for begge korallarter (sjøtre: r = -0,09 med O₂, r = 0,35 med NH₄; risengrynkorall: r = 0,01 med O₂, r = -0,22 med NH, men den noe høyere NH₄-korrelasjonen hos sjøtre bidro til bedre modellytelse ved kalibrering. Dette indikerer at visuell påvirkning og metabolsk stress kan være delvis utkoblet hos koraller, noe som understreker behovet for multimetriske tilnærminger.
Samlet sett fremsto viftesvamp som den mest sensitive arten på tvers av nær sagt alle parametere i begge tilnærminger. Korrelasjonen mellom indeksene og de enkelte målte responsene (Tabell 9: reduserte lipidreserver, nedregulert energimetabolisme, endret mikrobiom, lysosomal destabilisering) gir høy grad av sikkerhet for at arten utsettes for fysiologisk betydelig påvirkning fra anleggene.
Kålrabisvamp viste også høy sensitivitet i begge tilnærminger, særlig med den kalibrerte indeksen som forklarte 81,2 % av variasjonen. Lysosomal membrandestabilisering og oppregulering av stressrelaterte transkriptomveier (Tabell 9) avdekket påvirkning som ikke ville blitt fanget opp gjennom visuell vurdering alene, og illustrerer verdien av «tidlig-varsel»-indikatorer.
Risengrynkorall viste metodeavhengig sensitivitet, med bedre ytelse under like vekter enn kalibrert vekting. Diskrepansen mellom høy visuell sensitivitet og fraværet av metabolsk respons (Tabell 9: O₂ og NH₄ uendret) synliggjør utfordringer knyttet til ulike tidsskalaer for integrasjon og antyder behov for alternative fysiologiske kalibreringer.
Sjøtre viste moderat sensitivitet, med markant bedre ytelse under kalibrert vekting hvor både dybde og sedimentasjon var signifikante drivere. De komplekse responsene i Tabell 9 (ikke-lineær O₂-respons, ingen visuell stressrespons, men mange differensielt uttrykte gener) indikerer subtile påvirkningsmekanismer. Det begrensede utvalget i den kalibrerte tilnærmingen (n = 32 vs. 47) tilsier likevel forsiktighet i tolkningen.
Basert på oppsummeringen av alle feltbaserte responser (Tabell 9) og ytelsen til de sammensatte indeksene (Figur 46 og Figur 47), kan artenes relative sensitivitet rangeres (Tabell 10). Denne rangeringen reflekterer organismenes responser under reelle miljøforhold ved oppdrettsanlegg, og ekskluderer laboratoriebaserte eksponeringseksperimenter.
Art
Sensitivitet
Hovedstressor
Notater
Sjøtre
Moderat
Sedimentasjon og dybde
Komplekse responser: ikke-lineær metabolsk respons, ingen visuell stressrespons, men mange differensielt uttrykte gener; reproduksjon stabil
Risengrynkorall
Moderat (usikker)
Avstand til anlegg
Motstridende signaler: visuell tilstand påvirket, men fraværende metabolsk respons; endringer i mikrobielt samfunn; reproduksjon robust; svak ytelse i kalibrert indeks; begrenset metodisk dekning
Viftesvamp
Svært høy
Sedimentasjon og avstand
Konsistent sterk respons på tvers av alle metoder: molekylær og biokjemisk stressrespons, tydelig visuell påvirkning, reduserte lipidreserver, endret mikrobiom
Generelt robuste; enkelte molekylære responser på sedimentasjon; infaunal livsstrategi gir beskyttelse
Tabell 10: Samlet vurdering av artsvis sensitivitet og hovedstressorer basert på integrert analyse av alle responskategorier fra feltarbeid (Tabell 9) og sammensatte påvirkningsindekser. Vurderingen ekskluderer laboratoriebaserte eksponeringseksperimenter og reflekterer kun responser observert i prøver fra feltarbeid.
3.12 - Eksponeringsforsøk i laboratorium
Eksponeringsforsøk ble gjennomført for å bestemme toleransen hos voksne individer og larver av øyekorall, samt hos voksne individer av risengrynkorall, sjøtre og sjøbusk, overfor relevante akvakulturrelaterte stressfaktorer.
Effektene av de mye brukte legemidlene hydrogenperoksid og azametifos ble testet på larver av øyekorall under laboratorieforhold ved NINA/NTNU-laboratoriet i Trondheim. I tillegg ble effekten av det ofte brukte lusemiddelet emamektinbenzoat undersøkt på voksne øyekoraller under laboratorieforhold ved Nord universitet i Bodø.
For å vurdere toleransen hos voksne individer av risengrynkorall, sjøtre, sjøbusk og øyekorall overfor partikulært organisk materiale (fiskslam) fra akvakultur, ble det gjennomført et eksponeringsforsøk ved Havforskningsinstituttets forskningsstasjon i Austevoll.
Fokuset i eksperimentene, valg av målarter og type stressfaktorer, ble basert på identifiserte kunnskapshull og hvilke påvirkningsfaktorer som mest sannsynlig kunne påvirke de undersøkte artene.
3.12.1 - Øyekorallarve - eksponeringsforsøk med lakselusmidler
Hydrogenperoksid (H₂O₂)
For å behandle laks mot lakselus ved bruk av H₂O₂ anbefales en konsentrasjon på 1500–2100 mg per liter (avhengig på temperatur) i 20 min (Felleskatalogen Pasientutgave: Nemona Nouryon). En tidligere studie (Escobar-Lux mfl., 2019) har vist att konsentrasjoner over 340 mg per liter var 100 % dødelig for kopepoder (både hunner og kopepoditt stadie V). Konsentrasjonene som ble valg for denne studien var 340, 170, 85, 17, 8,5 og 0 mg per liter, noe som tilsvarer 20 %, 10 %, 5 %, 1 %, 0,5 % og 0 % av anbefalt konsentrasjon på 1700 mg per liter.
Voksne kolonier av øyekorall fra toktene i 2023 (tre kolonier) og 2024 (åtte kolonier) fra Møre og Romsdal ble holdt i laboratoriet ved NINA/NTNU. I 2023 var alle koloniene som slapp egg hunner og i 2024 ble det kun observert sperm fra kolonien. Ingen larver fra innsamlede koraller kunne derfor brukes i forsøkene. Larvene som ble brukt i forsøkene kom fra lokale populasjoner i Trondheimsfjorden. De var 22 dager gamle og ble tilfeldig fordelt blant de seks ulike konsentrasjonene og fire replikasjonene. Det ble brukt 20 larver per replikasjon, totalt 480 larver som ble eksponert i en time. Deretter ble de undersøkt under mikroskop og delt inn i kategorier basert på adferd. Siden ble de overført til rent sjøvann for restitusjon i 24 timer før de ble undersøk igjen. Adferd ble kategorisert som normal (svømmer), subletal (svømmer langsomt, urørlig) eller letal (død, abnormal). Temperaturen var konstant på 7–8 °C. Den totale dødeligheten er definert som summen av dødelighet fra en times eksponeringen pluss den etter 24 timer med restitusjon.
Figur 48: Påvirkning av hydrogenperoksid på 22 dager gamle øyekorallarver etter en times eksponering og 24 timers restitusjon.
Det var ikke noen dødelighet i kontrollgruppen etter en times eksponering mens etter 24 timers restitusjon var det noen abnormale larver (Figur 48). Det var 100 % dødelighet ved 85 g·L⁻¹ mens ingen dødelighet eller problemer med bevegelse ble observert ved den laveste testede konsentrasjonen (8,5 mg·L⁻¹ eller 0,05 % av anbefalt konsentrasjon). Konsentrasjonen av hydrogenperoksid som er dødelig for 50 % av populasjonen (LC50-verdien) ble beregnet til 27,20 mg·L⁻¹ (Figur 49). Dette er i samme område som hos voksne kopepoder (Calanus spp.) hvor en LC50 verdi på 30,6 mg·L⁻¹ er påvist (Escobar-Lux mfl., 2020) og betydelig lavere enn pelagiske hummerlarver (stadie I) med LC50 på 177 mg·L⁻¹ (Escobar-Lux mfl., 2020). LC10 verdien var 12,32 mg·L⁻¹ og LC90 57,84 mg·L⁻¹.
Figur 49: Total dødelighet av hydrogenperoksid på 22 dager gamle larver av øyekorall etter en times eksponering og 24 timers restitusjon.
Azametiphos
Ved behandling av laks mot lakselus ved bruk av azametiphos (Salmosan vet.) brukes en behandlingsdose på 100 µg·L⁻¹ i 30–60 min (Salmosan vet. «Benchmark Animal Health» - Felleskatalogen). Konsentrasjonene som ble brukt i denne studie var 0, 1, 10, 50, 100 og 500 µg·L⁻¹, basert på en tidligere studie på hummerlarver (Parsons mfl., 2020). Larver fra lokale populasjoner i Trondheimsfjorden, som var 33 dager gamle, ble tilfeldig fordelt blant de seks ulike konsentrasjonene og fire replikasjonene. Det ble brukt 20 larver per replikasjon, totalt 480 larver som ble eksponert i en time. Deretter ble de undersøkt under mikroskop og delt inn i kategorier basert på adferd. Videre ble de overført til rent sjøvann for restitusjon i 24 timer, før før de igjen ble undersøkt. Adferd ble kategorisert som normal (svømmer), subletal (svømmer langsomt, urørlig) eller letal (død, abnormal). Temperaturen var konstant på 7–8 °C. I denne studien har vi valgt å beregne konsentrasjonen hvor larvene ble synlig påvirket av kjemikalien (EC - Effect Concentration) i stedet for dødelighet (LC – Lethal Concentration) som ble gjort med hydrogenperoksid. Da azametiphos påvirker nervesystemet på dyrene mens hydrogenperoksid har en mekanisk påvirkning (Parsons mfl., 2025) blir effektene ikke like synlige i direkte dødelighet uten på svømmehastighet og paralysering, noe som til slutt resulterer i død. Dette er ansett som en mer relevant variabel for å vurdere nevrotoksiske kjemikalier (Van Geest mfl., 2014). EC er definert som summen av subletale og letale effekter. Det var ikke noen dødelighet i kontrollgruppen etter én times eksponering, men etter 24 timers restitusjon var det noen abnormale larver. Konsentrasjonen av azametiphos som påvirker 50 % av populasjonen (EC50-verdien) ble beregnet til 1,89 µg·L⁻¹ (Figur 50). Dette er lavere enn hos pelagiske hummerlarver (stadium I og II) med EC50 på henholdsvis 15,5 µg·L⁻¹ og 9,2 µg·L⁻¹ (Parsons mfl., 2020). EC10 verdien var 1,13 µg·L⁻¹ og EC90 3,17 µg·L⁻¹.
Figur 50: Effektkonsentrasjon (EC) av azametiphos på 33 dager gamle larver av øyekorall etter en times eksponering og 24 timers restitusjon.
Hydrogenperoksid
mg·L⁻¹
Azametiphos
µg·L⁻¹
LC10
12,32
EC10
1,13
LC50
27,20
EC50
1,89
LC90
57,84
EC90
3,17
Tabell 11: Beregnede LC- og EC-verdier (mg·L⁻¹ og µg·L⁻¹) for hydrogenperoksid og azametiphos basert på dødelighets- og effektdata. LC = lethal concentration, EC = effect concentration.
3.12.2 - Øyekorall - eksponeringsforsøk med emamektin
Per i dag er det lite kunnskap om effekter av avlusingsmidler på sårbare virvelløse bunndyr og det er uklart hvordan avlusingsmidler påvirker bunndyrs adferd og fysiologi, og om disse midlene akkumuleres i dyret. To studier i Norge har nylig påvist rest-konsentrasjoner av avlusingsmiddelet emamektin benzoate (det aktive stoffet brukt i avlusingsmiddelet Slice vet) i sjøfjær (Virgularia mirabilis og Pennatula phosphorea), både i dyr fra feltundersøkelser og etter eksponering i laboratorie-forsøk. Det ble også påvist variabel produksjon av slim, endring av fysiologi og av polyppadferd, slik som tentakkelutstrekning, i felt, men ikke i laboratorium (Taormina mfl., 2024a, 2024b). Ettersom emamektin benzoate, på grunn av sine kjemiske egenskaper, kan vedvare i havbunnsedimenter i lang tid, har dette avlusingsmiddelet blitt valgt som test-kjemikalie i flere laboratorieforsøk (Langford mfl., 2014; Benskin mfl., 2016; Benson mfl., 2017; Hamoutene mfl., 2018; Strachan og Kennedy, 2021). Selv om virkningsmekanismene til emamektin benzoate i koraller er ukjente, er det kjent at derivatet avermectin kan gi neurotoksisk virkning i virvelløse dyr (Casida og Durkin, 2015; El-Saber Batiha mfl., 2020). De siste årene har mange akvakulturlokasjoner blitt flyttet til dype fjorder med hardbunn og sterk vanngjennomstrømming. Dermed blir filtrerende og stedfaste hardbunnsarter som øyekorall mer utsatte for eksponering av organisk materiale og kjemikalier brukt av oppdrettsnæringen.
Metode
Det ble utført et 16-dagers laboratorieforsøk med emamektin benzoate og øyekorall i juni/juli 2024. Små fragmenter (tre koraller per tank) av øyekorall ble eksponert for to konsentrasjoner av emamektin benzoate i et mini-resirkuleringsanlegg (RAS): en lav dose på 0,4 mg·L⁻¹ og en høy dose på 1,6 mg·L⁻¹ (tre tanker for hver konsentrasjon). I den fem dager lange eksponeringsperioden ble emamektin benzoate tilsatt tankene daglig. En kontrollgruppe med koraller uten slik tilsetting, ble brukt som referanse (tre fragmenter per tank, tre tanker). Flere metoder ble brukt for å vurdere effekten av emamektin benzoate på øyekorall sin fysiologi og adferd: i) prøvetaking av korallvev til kjemisk analyse av emamektin benzoate, ii) respirasjonsrate, iii) ammoniumekskresjon og iv) bildeopptak fra kamera ble brukt til å studere visuelle korall-endepunkter. Disse metodene ble gjort før eksponering (for referansemålinger), under eksponering og etter eksponering.
i) Prøver av korallfragmenter ble tatt på ulike dager etter første eksponering. De 18 eksponerte korallprøvene pluss tre korallprøver fra kontrollgruppen ble analysert for rester av emamektin benzoate ved laboratoriet til HI i Bergen. Frossent korallvev ble manuelt homogenisert (blanding av knust korallskjelett og bløtvev). Acetonitril ble brukt til å ekstrahere emamektin fra prøvene, og ekstraktet ble analysert for emamektin benzoate (B1a; hovedkomponenten i emamektin benzoate) med Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS) som tidligere beskrevet (Taormina mfl., 2024a). Vannprøver fra både lav og høy dose ble tatt etter andre eksponering og frem til slutten av eksperimentet for å validere nivået av emamektin benzoate i eksponeringsvannet. Prøvene ble sendt til laboratoriet til HI i Bergen for analyse av emamektin B1a ved bruk av LC-MS/MS som tidligere beskrevet (Taormina mfl., 2024a).
ii) Respirasjonsmålinger ble gjort med alle korallfragmentene ved bruk av 2 L inkubasjonskammer i opptil ca. 60 timer. Oksygennivå og temperatur ble målt hver sjette time med en fiberoptisk oksygenmåler (Fibox 4, PreSens, Tyskland). Kontrollkamrene som kun inneholdt sjøvann ble brukt for å bestemme mikrobiell bakgrunnsrespirasjon, som ble trukket fra respirasjon målt i kamrene med koraller. Respirasjon ble standardisert til biomasse (ash free dry weight AFDW). Respirasjonsrate er angitt i μmol O₂/gAFDW/t og beregnet som lineær regressionskoeffisient av korallens oksygenmålinger over inkubasjonsstid (timer) multiplisert med vannvolum og delt på biomasse. Metningskurven for oksygen viste at to målepunkter fra pre-eksponeringsperidoen åpenbart var avvikende og disse ble fjernet før beregning av regressionskoeffisient.
iii) Målinger av ammonium i vannet ble gjort i sammenheng med respirasjonsmålinger for å kvantifisere ekskresjon av ammonium. På starten og på slutten av hver respirasjonsmåling ble vannprøver tatt fra hver tank og fra korallenes inkubasjonskammer. I tillegg ble vannprøver tatt fra kontrollkamrene, som kun inneholdt vann fra den respektive gruppen, for bestemmelse av vannets bakgrunnsproduksjon av ammonium. Vannprøvene ble analysert for ammonium ved laboratoriet til NIVA i Oslo med et fluorescens-analyseinstrument (Skalar San++ Autoanalysator 5000). Ammoniumekskresjon (NH₄⁺) er angitt i μmol NH₄⁺/gAFDW/t og beregnet som differensen mellom ammonium ved slutt og start delt på inkubasjonstid (timer) multiplisert med vannvolum og delt på biomasse.
iv) Visuelle kamerabaserte observasjoner av koraller ble gjennomført under forsøket. Et GoPro-kompaktkamera ble plassert foran glasstankene til de to emamektin-behandlede gruppene og kontrollgruppen. Ett kamera ble brukt for hver gruppe. Kameraet ble justert for å ta bilde av to av de tre korallfragmentene i hver tank, og deretter flyttet til de replikate tankene, slik at det ble observert totalt seks korallfragmenter per gruppe. For hver korall ble det tatt bilder i løpet av 4 timer, med 30 minutters intervaller. Dette resulterte i omtrent 8 bilder for hver periode, det vil si før eksponering (dag 1), under eksponering (dag 7 og 9), og etter eksponering (dag 12). På grunn av et teknisk problem med GoPro-kameraet ble kun tatt bilde av 1 korall i de lavdosebehandlede tankene før eksponering. Det ble heller ikke gjort observasjoner av hver korall etter eksponering (se Tabell 12). For hvert bilde ble det gjort visuelle observasjoner og telling av polyppaktivitet (antall polypper med utstrakte tentakler, antall polypper med inntrukne tentakler og antall polypper med halvt utstrakte tentakler), slimutløsning (slimtråder) og nedbrytning av korallens coenosarc-vev.
Antall bilder
Gruppe
Behandling
Antall polypper
Tidspunkt 1
Tidspunkt 2-4
Totalt
KONTROLL
C1R1P4
6
8
16
24
C1R1P5
6
8
14
22
C1R2P20
3
8
20
28
C1R2P21
6
1
11
12
C1R3P22
2
8
13
21
C1R3P24
3
8
20
28
Totalt C
26
41
94
135
LAV DOSE
T1R1P16
5
4
69 (24)
97
T1R1P17
8
N/A
30
30
T1R2P2
5
N/A
27 (24)
51
T1R2P3
10
N/A
21
21
T1R2P25
4
N/A
33 (16)
49
T1R2P27
4
N/A
12
12
Totalt T1
36
4
168
212
HØY DOSE
T2R1P13
6
8
16
24
T2R1P14
5
7
22 (8)
29
T2R2P8
6
8
12
20
T2R2P9
9
8
11
19
T2R3P10
4
8
11
19
T2R3P12
5
8
12
20
Totalt T2
35
47
84
131
Tabell 12: Antall bilder tatt av hver korall (P) gjennom forsøksperioden i hver replikat-tank (R1, R2, R3) for hver emamektin-behandlet gruppe (T1 = 0,4 mg·L⁻¹; T2 = 1,6 mg·L⁻¹)samt kontrollgruppenn (C1). Tidspunkt 1: før eksponering; Tidspunkt 2--4: under eksponering og etter eksponering (tall i parentes). NA = ingen data tilgjengelig på grunn av tekniske problemer med kameraet.
Statistisk analyse ble utført med lineære blandede modeller (LMM) med korallfragmenter brukt som tilfeldige effekter (beregnet med restricted maximum likelihood (REML) og under bruk av Satterthwaite’s metodikk) for å ta hensyn til gjentatte målinger av samme korallfragment og for å statistisk teste om forskjellige doser (lav og høy behandling) i interaksjon med tidsperioder (før, under og etter eksponering) har en signifikant effekt på respirasjonsrate og ammoniumsutslipp. Kontrollgruppen ble brukt som referanse (intercept) i LMM. LMM ble kjørt i RStudio (versjon 2025.09.2) med R (versjon 4.5.1) og pakken lmerTest (versjon 3.1-3).
Forskjeller i de visuelle kamerabaserte observasjonene ble testet for signifikanse ved bruk av ikke-parametriske analyse under null hypotesen «ingen forskjell mellom gruppene» (Kruskal–Wallis-test, p > 0,05 med Bonferroni korrigering, IBM SPSS ver.30.0.00).
Resultater
Emamektin B1a ble påvist i alle 37 analyserte vannprøver fra lav og høy behandling, med generelt høyere verdier under eksponeringsperioden og med lavere verdier etter eksponering. Gjennomsnittlig konsentrasjon av emamektin B1a i vannet i lavdosegruppen var 0,129 mg·L⁻¹ (± 0,13 mg·L⁻¹), mens gjennomsnittlig emamektin B1a konsentrasjon i høydosegruppen var 0,596 mg·L⁻¹ (± 0,577 mg·L⁻¹). Konsentrasjonene i vannet var imidlertid avhengige av tidspunktet for prøvetaking, dvs. hvor lenge etter tilsetning av emamektin prøvene ble tatt (resultater ikke vist).
Emamektin B1a ble påvist i alle de 18 eksponerte korallfragmentene, men med stor variasjon i forhold til tid og dosering. Gjennomsnittlig emamektin B1a konsentrasjon i koraller fra lavdosegruppen var 0,0040 µg·mg⁻¹ (± 0,0033 µg·mg⁻¹). Gjennomsnittlig emamektin B1a konsentrasjon i koraller fra høydosegruppen var 0.0031 µg·mg⁻¹ (± 0,0023 µg·mg⁻¹). Konsentrasjonen av emamektin B1a i de tre korallfragmentene fra kontrollgruppen som ble analysert, var under kvantifiseringsgrensen (< LOQ). Korrelasjonsanalyse (Pearson test med bruk av corr.test i R pakken ‘psych’ versjon 2.4.6.26) ble utført for å teste sammenheng mellom korallkonsentrasjonene og 1) tidspunkt (dag) og 2) biomasse (AFDW). Resultat viste ingen signifikant sammenheng mellom biokonsentrasjon av emamektin B1a og prøvetidspunkt og biomasse. Årsaken til de store forskjellene i biokonsentrasjon mellom fragmentene bør studeres nærmere. Gitt den store variasjonen i opptak, kan ikke resultatene fra dette forsøket brukes til å konkludere om bioakkumulering over tid.
Under eksponering og i post-eksponeringsperioden ble korallpolyppmortalitet observert hos to korallfragmenter fra lavdosegruppen, fem korallfragmenter fra høydosegruppen og ett korallfragment fra kontrollgruppen.
I pre-eksponeringsperioden viste alle tre grupper relativt lave respirasjonsverdier. En forskjell i respirasjonsrate mellom kontrollgruppen og begge de eksponerte gruppene ble funnet i eksponeringsperioden samtidig med at alle tre gruppene viste en økning i respirasjon i forhold til pre-eksponering (Figur 51). Sammenlignet med pre-eksponeringsperioden, ble det under eksponeringen funnet en signifikant økning i kontrollgruppens respirasjonsrate samt i behandlingsgruppen med høy dosering (indikasjon på signifikant interaksjon mellom høy dosering og eksponeringsperioden) (LMM intercept;TimeExposure: estimate = 5,29; df = 13,82; t = 3,32; p = 0,0051; Dosehigh dose:TimeExposure: estimate = 7,28; df = 15,04; t = 2,5; p = 0,0244). I post-eksponeringsperioden ble det funnet en signifikant forskjell i respirasjon mellom eksponerte fragmenter og kontrollgruppen (indikasjon på signifikant interaksjon mellom høy og lav dosering og post eksponeringsperioden) (LMM Doselow dose:TimePost-exposure: estimate = 12,07; df = 14,45; t = 3,63; p = 0,0026; Dosehigh dose:TimePost-exposure: estimate = 9,16; df = 14,91; t = 2,29; p = 0,0370). En mulig tolkning på hvorfor respirasjonen også økte i kontrollgruppen under eksponeringsperioden for deretter å holde seg stabil på samme nivå i post-eksponeringsperioden, kan være generelt forhøyet stressnivå (pga. ulike kumulative faktorer inkludert håndtering).
Figur 51: Oksygenforbruk til øyekorall A) før eksponering, B) under eksponering og C) etter eksponering i kontroll, lavdose- og høydosegruppene. Oksygenforbruk er angitt i μmol O₂·gAFDW⁻¹·t⁻¹ og fremstilt i boksplott som viser median, og 25 og 75 % kvartil.
I pre-eksponeringsperioden viste alle gruppene relativ lav ekskresjon av ammonium, mens i eksponeringsperioden ble det påvist en tydelig forskjell i ekskresjon av ammonium mellom kontrollgruppen og de eksponerte gruppene (Figur 52). I forhold til pre-eksponeringsperioden, ble det under eksponeringen funnet en signifikant økning i kontrollgruppens produksjon av ammonium samt i fragmentene i lavdosegruppen (indikasjon på signifikant interaksjon mellom lav dosering og eksponeringsperioden) (LMM intercept;TimeExposure: estimate = 0,61; df = 18,55; t = 2,16; p = 0,044; Doselow dose:TimeExposure estimate = 1,41; df = 19,42; t = 3,04; p = 0,0066). I post-eksponeringsperioden ble det påvist redusert ekskresjon av ammonium i begge eksponeringsgruppene sammenlignet med eksponeringsperioden.
Figur 52: Ammoniumekskresjon hos øyekorall A) før eksponering, B) under eksponering og C) etter eksponering i kontroll, lavdose- og høydosegruppen. Ammoniumekskresjoner angitt i μmol NH₄⁺·gAFDW⁻¹·t⁻¹ og fremstillt i boksplott som viser median, og 25 og 75 % kvartil.
Redusert antall korallfragmenter i begge behandlingsgruppene som ble brukt til måling av respirasjon og prøvetaking av ammonium i eksponerings og post-eksponeringsperioden skyldes hovedsakelig polyppdødelighet. Selv om LMM metoden er spesielt egnet for å håndtere ubalanserte longitudinale data, bør en være forsiktig med å trekke for vide konklusjoner fra forsøk med så lav antall. Oppfølgende forsøk for å validere funnene anbefales.
Polyppaktiviteten fra tentakkelutstrekning var i utgangspunktet lav hos kontrollkorallene, men viste en svak økning i løpet av forsøksperioden. For de emamektin-behandlede korallene var polyppaktiviteten også lav før eksponering, uten signifikante forskjeller mellom lav- og høydosegruppene sammenlignet med kontrollgruppen (Kruskal–Wallis-test, n = 13, p > 0,05). Etter eksponering avtok aktiviteten ytterligere og var signifikant lavere i høydosegruppen sammenlignet med kontrollgruppen (Kruskal–Wallis-test, n = 18, p = 0,034). Polyppene hos korallene i alle grupper, inkludert kontrollene, forble imidlertid hovedsakelig inntrukket under og etter eksponering (Figur 53). Basert på polyppaktivitet alene er det derfor vanskelig å trekke klare konklusjoner om effektene av emamektinbehandling, ettersom alle koraller, også kontrollene, viste gjennomgående lav aktivitet gjennom hele forsøket.
Figur 53: Prosentandel (%) av korallobservasjoner med polypper med fullt utstrakt (blå), inntrukne (grå) eller halvt utstrakte (oransje) tentakler for hvert korallfragment (x-akse) i kontroll-, lav- og høydosegruppene av emamektin. Panel A viser målinger før eksponering, og panel B viser målinger under og etter eksponering kombinert. Figuren viser også oksygenforbruk (μmol O₂·gAFDW⁻¹·t⁻¹) (sekundær y-akse), representert med svarte trekanter for målinger før eksponering, svarte romber for målinger under eksponering, og gule sirkler for målinger etter eksponering. For lavdosegruppen før eksponering foreligger det ingen observasjoner for noen av replikatene på grunn av teknisk feil med kameraet.
Når det gjelder prosentandelen polypper med slimproduksjon viser Tabell 13 den relative andelen av polypper med slimproduksjon før (Tidspunkt 1), under (Tidspunkt 2 og 3) og etter eksponeringsperioden (Tidspunkt 4). Det var ingen (eller lav) synlig utskilling av slim i kontrollkorallene. I motsetning til dette produserte alle polyppene fra koraller eksponert for lav og høy dose av emamektin slim både under og etter eksponering. Det var ingen forskjell i prosentandel polypper med utskilling av slim mellom lavt og høyt eksponerte koraller, men det ble observert en større mengde utskilt slim fra noen av de høydoseeksponert korallene (se Figur 54). Når det gjelder nedbrytning av korallens coenosarc (det ytre slimlaget som dekker skjelettet), ble det ikke observert noen synlige effekter hos koraller behandlet med emamektin verken ved lav eller høy dose.
Slim utskilt fra antall polypper
Gruppe
Behandling
Tidspunkt 1
Tidspunkt 2
Tidspunkt 3
Tidspunkt 4
KONTROLL
C1R1P4
0 %
17 %
17 %
-
C1R1P5
0 %
0 %
0 %
-
C1R2P20
0 %
0 %
0 %
0 %
C1R2P21
0 %
0 %
0 %
-
C1R3P22
0 %
0 %
0 %
-
C1R3P24
0 %
0 %
0 %
0 %
LAV DOSE
T1R1P16
0 %
40 %
80 %
100 %
T1R1P17
-
25 %
0 %
-
T1R2P2
-
20 %
20 %
80 %
T1R2P3
-
30 %
-
-
T1R2P25
-
0 %
25 %
50 %
T1R2P27
-
-
50 %
-
HØY DOSE
T2R1P13
0 %
0 %
16 %
-
T2R1P14
0 %
20 %
40 %
60 %
T2R2P8
0 %
0 %
17 %
-
T2R2P9
0 %
22 %
44 %
-
T2R3P10
0 %
50 %
50 %
-
T2R3P12
0 %
60 %
80 %
-
Tabell 13: Prosentandel polypper med slimutskilling for hver korallreplikat i hver behandling (T1, T2) og kontroll (C1), basert på visuelle observasjoner før (Tidspunkt 1), under (Tidspunkt 2--3) og etter eksponering (Tidspunkt 4).
Figur 54: To bilder A og B av samme korall i høydose behandling tatt med en times mellomrom under eksponeringsperioden. Bildene viser tydelig slimproduksjon i form av tråder. De originalfargete bildene (255 farger) ble omgjort til negative bilder for å synliggjøre slimproduksjon.
3.12.3 - Øyekorall, sjøtre, sjøbusk og risengrynkorall - eksponeringsforsøk med fiskslam
Et tredje eksponeringsforsøk ble gjennomført for å bestemme og sammenligne toleransen hos voksne risengrynkoraller, sjøtrær, sjøbusker og øyekoraller overfor partikulært organisk materiale fra akvakultur. Forsøket ble utført ved Havforskningsinstituttets eksperimentelle akvarieanlegg i Austevoll i mars 2025.
Metode
Korallkolonier av de nevnte artene ble samlet inn fra en bratt vegg i Korsfjorden og et rev i Hardangerfjorden ved hjelp av ROV-en «Freyja» om bord på RV «Prinsesse Ingrid Alexandra» mellom 26. februar og 1. mars 2025. Koloniene ble oppbevart om bord i nedkjølte oppbevaringstanker med dypvann fra innsamlingsstedene før de ble direkte overført til laboratoriefasilitetene i Austevoll. To separate eksponeringsforsøk ble gjennomført: det første med risengrynkorall og sjøtre mellom 3. og 19. mars, og et andre forsøk med sjøbusk og øyekorall fra 24. mars til 9. april. For hvert forsøk ble 45 korallfragmenter på omtrent 10 cm kuttet fra kolonigrenene og plassert i holdere laget av ikke-giftige materialer. Korallfragmenter av risengrynkorall og sjøtre ble overført direkte til forsøkskarene og akklimatisert i 1 dag før eksponeringen startet, mens sjøbusk og øyekorall ble akklimatisert i oppbevaringstanker i 3 uker. Eksponeringsforsøket ble gjennomført i 15 gjennomstrømningskar som ble tilført ufiltrert dyphavsvann pumpet fra 160 m dyp med en strømningshastighet på 50 L·t⁻¹.
Karene ble eksponert for fem ulike konsentrasjoner (0, 0,5, 1, 5 og 10 g·m⁻²·dag⁻¹) av tørket lakseavføring fra Bioretur, med partikkelstørrelser mellom 100 og 400 µm. Dosene ble valgt basert på modellerte og målte sedimentasjonsrater (g·m⁻²·dag⁻¹) rundt oppdrettsanlegg (Kutti mfl., 2007; Kutti mfl., 2022), og tilsvarer nivåene av sedimentasjon som er modellert direkte under merdene og innenfor 300 m i hovedstrømretningen fra oppdrettsanleggene som inngår i denne studien (Tabell 14).
Forsøkdose (g·m⁻²·dag⁻¹)
Akkumulert forsøkdose over 2 år (g·m⁻²)
Omtrentlige avstandsintervall til anlegg (m)
0
0
> 1000
0,5
360
225 – 300
1
720
175 – 250
5
3600
merdkant - 50
10
7200
under merden
Tabell 14: Sammenligning mellom forsøksdose (g·m⁻²·dag⁻¹), akkumulert sedimentasjon over to år (g·m⁻²) og omtrentlig avstand fra studieoppdrettsanleggene (basert på sju studieanlegg i prosjektet).
Totalt ble 15 kar brukt i hvert eksperiment. I det første eksperimentet ble tre risengrynkoraller og tre sjøtrær plassert i hvert kar, mens i det andre eksperimentet ble tre øyekoraller og tre sjøbusker fordelt i hvert kar. Tre replikatkar ble tildelt hver av de fem dosekonsentrasjonene. Alle koraller som ble ansett for å ha dødd under eksperimentet, vurdert ut fra et betydelig tap av vev, ble fjernet.
Eksperimentet varte i 20 dager. Hver morgen kl. 09:00 ble korallene eksponert ved å drysse tørket lakseavføring jevnt over vannoverflaten i hvert kar, unntatt i kontrollbehandlingen. Vanntilførselen ble slått av før eksponering og gjenopptatt etter 10 minutter for å la partiklene synke til bunnen. Temperatur, oksygenkonsentrasjon og pH ble overvåket daglig og holdt seg stabile gjennom hele forsøket. Temperaturen var mellom 8,5 og 8,8 °C, oksygenkonsentrasjonen mellom 95 og 105 mg·L⁻¹, og pH mellom 8,1 og 9,1.
Et tidsforløpskamerasystem ble brukt for å vurdere polyppaktivitet fra tentakkelutstrekning og den generelle tilstanden til korallene under begge eksperimentene. Kameraer ble installert for fire behandlingsgrupper (ett kar per gruppe): kontroll, 0,5, 5 og 10 g·m⁻²·dag⁻¹. Et GoPro 12-kamera, synkronisert med LED-belysning, ble montert over midten av hvert kar, med kameralinsen nedsenket i vannet (Figur 55A). Systemet tok bilder av alle korallene én gang per time gjennom hele forsøksperioden (Figur 55B).
Figur 55: Eksperimentelt oppsett ved Havforskningsinstituttets forskningsstasjon i Austevoll. (A) Gjennomstrømningskar med tidsforløpskamerasystem montert over karet for kontinuerlig fotografering av korallfragmentene under eksponering. (B) Eksempel på bilde tatt av kamerasystemet under forsøksperioden.
Resultater
Tidsseriebilder av koraller eksponert for 0, 0,5, 5 og 10 g·m⁻²·dag⁻¹ ble tatt gjennom hele forsøksperioden i begge eksperimentene (Figur 56 og 57). Midtveis i eksperiment 1, mellom dag 10 og 12, hadde noen av risnegyrnkorallene mistet en betydelig mengde kalkskjell og vev og ble ansett som døde og fjernet. Én risengrynkorall døde i kontrollen og i behandlingsgruppen 5 g·m⁻²·dag⁻¹, og tre i 10 g·m⁻²·dag⁻¹. Mot slutten av eksperimentet 1, mellom dag 1 og 19, døde ytterligere 6 risengrynkorall i behandlingsgruppen 5 g·m⁻²·dag⁻¹ og 5 i 10 g·m⁻²·dag⁻¹. Alle risengrynkorall overlevde i behandlingene 0,5 og 1 g·m⁻²·dag⁻¹. Alle sjøtrær overlevde forsøksperioden og klarte å fjerne noen partikler, men under de høyere eksponeringene (5 og 10 g·m⁻²·dag⁻¹) viste sjøtrærne omfattende dekning av sedimenter og produksjon av slim og vevsnedbrytning, som observert i Figur 56.
Alle øyekoraller og sjøbusk som ble eksponert, overlevde hele 20 dager av forsøk 2. Imidlertid var greinene til sjøbusk kraftig dekket av sedimenterte partikler ved slutten av eksperimentet under de høyere eksponeringsbehandlingene (5 og 10 g·m⁻²·dag⁻¹), men polyppene var fortsatt aktive og strakte ut tentaklene sine. Øyekoraller viste derimot bedre evne til å fjerne sedimenterte partikler enn sjøbusk, og det ble observert minimal sedimentering på øyekorallene ved slutten av eksperimentet (Figur 57).
Polyppaktivitet ble registrert med omtrent én times intervaller for hver art og behandling. Enkelte dataserier ble ekskludert på grunn av ugunstig korallplassering, delvis tildekking av karkonstruksjoner, sedimentakkumulering som forstyrret segmenteringen, eller manglende GoPro-tidsstempler. En oversikt over gjennomsnittlig polyppaktivitet over tid, inkludert når data ble samlet inn, er vist i Vedlegg A7, Figur 76.
Figur 56: Tidsforløpsbilder av risengrynkorall og sjøtre i karene eksponert for 0 (kontroll), 0,5, 5 og 10 g·m⁻²·dag⁻¹ tatt ved starten og slutten av forsøk 1.
Figur 57: Tidsforløpsbilder av øyekorall og sjøbusk i karene eksponert for 0 (kontroll), 0,5, 5 og 10 g·m⁻²·dag⁻¹ tatt ved starten og slutten av forsøk 2.
Tidsseriene for polyppaktivitet fra tentakkelutstrekning ble analysert med en cosinor-modell for å vurdere døgnrytme. Modellen dekomponerer tidsserien i en rytmisk komponent med en fast 24-timersperiode. Modellene ble tilpasset ved hjelp av en beta-fordelt generalisert lineær blandet modell (GLMM; glmmTMB i R), egnet for proporsjonsdata mellom 0 og 1. Når flere kolonier var tilgjengelige innen samme behandling, ble koloni inkludert som tilfeldig intercept for å ta hensyn til grunnleggende forskjeller mellom kolonier. Fra modellene ble tre biologisk relevante parametere estimert: MESOR (middelverdien rytmen oscillerer rundt), amplitude (rytmeutslag, uttrykt som halvparten av topp–bunn-variasjonen og tilbakeført til prosentskala), og akrofase (estimert tidspunkt for maksimal aktivitet). Rytmisitet ble testet ved hjelp av likelihood ratio test (LRT), hvor full cosinor-modell ble sammenlignet med en nullmodell som kun inkluderte tilfeldig intercept. Marginal R² ble beregnet for å kvantifisere andelen varians forklart av den faste rytmiske komponenten.
Døgnrytmen varierte betydelig mellom artene og ble påvirket ulikt av behandlingskonsentrasjonene (Figur 58).
Figur 58: Tidsforløpsbilder ble brukt for å registrere polyppaktivitet under fire behandlinger (Kontroll, 0,5, 5 og 10 g·m⁻²·dag⁻¹). Observasjoner av polyppaktivitet per time ble gjennomsnittet over alle dager og replikater for hver art. Cosinor-modeller med 24-timers periode ble tilpasset disse dataene ved hjelp av generaliserte lineære blandede modeller med beta-fordeling (glmmTMB) for å estimere døgnrytmer. Tilpassede kurver ble generert fra modellkoeffisientene og plottet som stiplede linjer. De endelige 2×2-panelfigurene viser fire arter per panel, med observerte gjennomsnitt (punkter og bånd for ± SE) og modellprediksjoner (stiplede linjer) for hver behandling.
Risengrynkorall viste en svak, men signifikant 24-timers rytme i kontroll (Tabell 15). Ingen av behandlingskonsentrasjonene (0,5, 5 og 10 g·m⁻²·dag⁻¹) viste statistisk påvisbar døgnrytme, og både amplitude og marginal R² var lave. Døgnrytme identifisert med MESOR avtok med økende behandlingskonsentrasjon (50,3 % i kontroll mot 40,0–44,8 % i behandlingene) og ble påvirket ved alle konsentrasjoner. Resultatene indikerer en doseavhengig reduksjon i total polyppaktivitet i tillegg til tap av rytmisitet.
Sjøtre viste den tydeligste døgnrytmen. Kontrollindivider hadde en klar og sterkt signifikant 24-timers rytme (Tabell 15). Ved 0,5 g·m⁻²·dag⁻¹ var rytmen fortsatt til stede, men betydelig dempet, og akrofase var forskjøvet til ca. kl. 08:00, nesten 10 timer tidligere enn i kontroll. Data for høyere konsentrasjoner var ikke tilgjengelige for denne arten. Kontroll og 0,5 g·m⁻²·dag⁻¹ dekket dessuten delvis ikke-overlappende perioder (2.–11. mars og 10.–21. mars). Selv om miljøforholdene ble holdt konstante, kan tidsmessige effekter ikke helt utelukkes.
Sjøbusk ved 0,5 g·m⁻²·dag⁻¹ viste en signifikant døgnrytme (Tabell 15). Kontrolldata ble imidlertid ekskludert fra den endelige modellen på grunn av singularitet, da de to replikatkoloniene viste sterkt divergerende aktivitetsmønstre, noe som ga ustabile estimater for tilfeldig effekt. Dette begrenser direkte sammenligning mellom tilgjengelige grupper.
Øyekorall viste ingen statistisk påvisbar døgnrytme under noen av de testede betingelsene. Alle p-verdier var ikke-signifikante, amplitudene var neglisjerbare og marginal R² var nær null (Tabell 15). MESOR økte imidlertid med økende konsentrasjon (36,1 % i kontroll mot 43,2 % ved 5 g·m⁻²·dag⁻¹ og 63,8 % ved 10 g·m⁻²·dag⁻¹), noe som kan indikere en mulig konsentrasjonsavhengig stimulering av total polyppaktivitet.
Samlet sett viser eksponeringsforsøkene at fiskefekalier påvirker både døgnrytme på en artsavhengig måte. Risengrynkorall hadde den høyeste relative følsomheten for fiskefekalier sammenlignet med de andre fire korallartene og døgnrytmen ble forsytyrret ved laveste eksponeringsnivå. Sjøtre hadde den tydeligste døgnrytmen under kontrollforhold, men rytmen ble allerede dempet og faseforskjøvet ved laveste eksponeringsnivå. Øyekorall viste ingen statistisk påvisbar døgnrytme under noen av de testede betingelsene. Totalt aktivitetsnivå (MESOR) økte imidlertid med konsentrasjon av partikler, noe som kan indikere en økt aktivitet av tentaklene for å fjerne partikler.
Art
Behandling
Mesor
Amplitude
Amplitude (%)
Akrofase (timer)
R²
p-verdi
N obs.
N. replikater
Risengrynkorall
0,5
44,84
0,09
2,21
4,25
0,014
0,256
674
3
Risengrynkorall
10
39,98
0,1
2,43
4,06
0,009
0,211
541
2
Risengrynkorall
5
43,97
0,11
2,77
11,81
0,019
0,319
375
3
Risengrynkorall
Kontroll
50,26
0,31
7,65
2,19
0,167
< 0,001
402
2
Sjøbusk
0,5
53,92
0,17
4,29
22,61
0,093
< 0,001
1471
3
Sjøtre
0,5
45,77
0,21
5,09
8,05
0,057
0,006
535
2
Sjøtre
Kontroll
50,73
0,47
11,61
18,28
0,348
< 0,001
401
2
Øyekorall
10
63,8
0,06
1,33
7,62
0,006
0,159
791
3
Øyekorall
5
43,16
0,03
0,84
1,77
0,001
0,667
1282
3
Øyekorall
Kontroll
36,14
0,05
1,09
0,25
0,007
0,279
916
2
Tabell 15: Resultater fra cosinor-analyse av døgnrytme i tentakkelskår for hver art og behandling. Tabellen viser mesor (rytmejustert gjennomsnitt), amplitude (absolutt og i prosent av mesor), akrofase (tidspunkt for maksimal aktivitet i timer), forklaringsgrad (R²), p-verdi for rytmisitet, totalt antall observasjoner (n) og antall replikattanker.
3.13 - Kvantifisering av artstettheter fra video
Bunndyrarters tetthet (antall individer per areal, oftest standardisert til m⁻²) er en kvantitativ måte å sammenligne endringer knyttet til menneskelig påvirkning på arters utbredelse mellom tidspunkter og geografiske steder. Frem til nå har det ikke eksistert en standardisert metodikk som beskriver hvordan artstettheten og spesielt hvordan havbunnsarealet fra video som er tatt på bratt eller variert terreng skal beregnes (i motsetning til standardisert prosedyre som tradisjonelt anvendes på bløtbunn for miljøovervåkning rundt akvakulturanlegg). Dette har i de siste årene blitt påpekt av norske miljømyndigheter og konsulentselskaper at de trenger både en definisjon på sårbare naturtyper (spesielt på hardbunn) og en konsekvensvurdering av akvakulturutslipp som er basert på nøyaktige tetthetsestimater. Konsulentselskaper rapporterer per i dag artssammensetning kun semi-kvantitativ, dvs. antall individer per transekt. I et samarbeid mellom NORD universitet, Havforskningsinstituttet og Akvaplan Niva AS ble det i 2025 utviklet en ny prosedyre BVides (Biigle Video density estimation) for å beregne tetthet av bunnarter fra videotransekter.
Figur 59: Fremgangsmåte til BVides med skjematisk visualisering av de fem stegene for tetthetsberegning. For hvert steg er data input, prosessering og output illustrert.
Det åpent tilgjengelige bilde- og videoannoteringsverktøyet BIIGLE 2.0 (BioImage Indexing, Graphical Labeling and Exploration) (Langenkämper mfl., 2017) brukes for å indentifisere og annotere bunndyr. Deretter brukes BVides for å beregne havbunnsarealet, modellert som et rektangel av transektlengde multiplisert med transektbredde. BVides inkluderer her to prosesseringsmetoder som per i dag ikke ble tatt hensyn til for å presisere transektlengden: glatting (engelsk: smoothing) dvs. fjerning av støy fra ROV posisjon (GPS koordinater og dybde) og inkludering av tre-dimensjonal transektlengde (heretter 3D), dvs. inkludere dybdeforskjell langs transektet. BVides estimerer transektbredden utfra laseren montert til ROV. Til slutt kan BVides koble sammen bunndyrannoteringer fra BIIGLE med havbunnsarealet fra video for å kvantifisere tettheten over hele transektet. Som alternativ kan brukeren velge å kvantifisere tettheten i mindre transektavsnitter, f.eks. tettheten innafør 50 m.
BVides ble testet med bruk av videotransekter tatt opp fra to miljøundersøkelser i Økssundet, tre på flat bløtbunn og tre på bratt hardbunn. De ulike prosesseringsmetodene for å beregne transektlengden ble vurdert mot standardmetodikken brukt per i dag (dvs. ikke-glattete GPS data, uten inklusjon av dybde, kalt “2D ikke-glattet” heretter). For hver videotransekt ble fire transektlengder beregnet ved bruk og kombinasjon av de ulike prosesseringsmetodene. Disse observasjoner er derfor ikke uavhengige, men representerer gjentatte beregninger på samme videotransekt under ulike prosesseringsmetoder. Det ble derfor tilpasset to lineære blandede modeller (LMM), én for flat og én for bratt havbunn, estimert med restricted maximum likelihood (REML), for å teste effekten av prosesseringsmetode på transektlengde. Transektlengde ble log10-transformert før analyse og brukt som responsvariabel. Beregnet varians av transektlengden viste betydelige forskjeller mellom prosesseringsmetodene. For å håndtere slik heterogen varians ble det inkludert en varIdent-variansstruktur i modellen, slik at hver prosesseringsmetode kunne ha sin egen varians. Prosesseringsmetode (2D ikke-glattet, 2D glattet, 3D ikke-glattet og 3D glattet) ble inkludert som fast effekt, med standardmetodikken (2D ikke-glattet) definert som referansekategori (intercept). Her refererer 2D til beregning av transektlengde basert utelukkende på horisontale GPS koordinater (x–y), uten inkludering av dybdeforskjeller langs transektet. Individuelle videotransekter ble inkludert som tilfeldig effekt for å ta høyde for gjentatte målinger innen samme transekt.
For alle tre transektene på flat bløtbunn ble de samme effektene observert: glatting førte til kortere transektlengder, mens inkludering av 3D‑dybdeforskjell ga en svært liten økning i transektlengden. Bruk av glatting (på 2D og 3D) resulterte i en statistisk signifikant reduksjon av transektlengden; gjennomsnittlig (n = 3) mellom 32 og 33 %. I motsetning resulterte ikke-glattet 3D-inkludering i en veldig liten ikke-signifikant økning; gjennomsnittlig (n = 3) 1 % (Figur 60 og Tabell 16).
Figur 60: Hver søyle representerer gjennomsnittlig transektlengde (i meter), standardavvik og p-verdier (fra LMM) til de fire prosesseringsmetodene til flat og bratt havbunn (n = 3). Mørkere fargetoner indikerer bruk av glatting, lyse fargetoner indikerer ikke-glattet, grønn farge inkludering av 3D-dybdeforskjell og blå farge bruk av 2D.
Også for alle tre transektene på bratt hardbunn ble de samme effektene observert: glatting førte til kortere transektlengder, mens inkludering av 3D‑dybdeforskjell ga i dette tilfelle en signifikant økning i transektlengden. Bruk av glatting (på 2D og 3D) resulterte i ei statistisk signifikant redusering av transektlengden som er gjennomsnittlig (n = 3) mellom 13–27 %. I motsetning til flate transekter, resulterte ikke-glattet 3D inkludering i en signifikant økning som er gjennomsnittlig (n = 3) betraktelig større med 14 %. Når glatting og 3D-inkludering ble brukt samtidig på bratte transekter (kalt 3D glattet), blir den 3D-forårsakete økningen av transektlengden kompensert av glattings-redusering av transektlengden (Figur 60).
Habitat
Metode
Beskrivelse
Endring i lengde (%)
Estimat
t
df
P
Flat bløtbunn
(n = 3)
2D ikke-glattet
Referanse (standardmetodikk)
0
0
–
–
–
2D glattet
Glatting av GPS-posisjoner
–33
–0,156
–3,81
6
0,009
3D ikke-glattet
Dybde inkludert
1
0,008
0,15
6
0,888
3D glattet
Dybde inkludert + glatting av GPS-posisjoner
–32
–0,153
–3,75
6
0,009
Bratt hardbunn
(n = 3)
2D ikke-glattet
Referanse (standardmetodikk)
0
0
–
–
–
2D glattet
Glatting av GPS-posisjoner
–27
–0,138
–10,11
6
< 0,001
3D ikke-glattet
Dybde inkludert
14
0,060
13,69
6
< 0,001
3D glattet
Dybde inkludert + glatting av GPS-posisjoner
–13
–0,059
–4,33
6
0,005
Tabell 16: Oversikt over prosesseringsmetodene, deres statistiske signifikans og prosentuell endring i transektlengden på flat og bratt terreng.
Tetthet av utvalgte sårbare hardbunnsarter (risengrynkorall og den morfologiske artsgruppen vifteformede svamper), bløtbunnsarten bambuskorall (Isidella lofotensis) og artsfamilien sjøfjær (Pennatuloidea) ble kvantifisert med de fire ulike prosesseringsmetodene (Figur 61 og 62, Tabell 17).
Art
n
Metode for transektlengde
Min–maks (ind·m⁻²)
Bambuskorall
3
2D ikke-glattet (standard)
0,23–1,15
2D glattet
0,31–1,47
3D ikke-glattet
0,23–1,11
3D glattet
0,31–1,46
Sjøfjær familie
3
2D ikke-glattet (standard)
0,004–0,035
2D glattet
0,005–0,053
3D ikke-glattet
0,004–0,034
3D glattet
0,005–0,053
Risengrynkorall
3
2D ikke-glattet (standard)
0,0005–0,0157
2D glattet
0,0006–0,0221
3D ikke-glattet
0,0004–0,0139
3D glattet
0,0005–0,0191
Vifteformete svamper
3
2D ikke-glattet (standard)
0,023–0,041
2D glattet
0,033–0,058
3D ikke-glattet
0,02–0,037
3D glattet
0,026–0,05
Tabell 17: Kvantifisert tetthet over hele transektlengden per prosesseringsmetode for hver art på flat og bratt terreng. Tetthet er fremstillt som minimum og maksimum tetthet per prosesseringsmetode, sammenført over alle tre analyserte transekter per art
Figur 61: Total tetthet (ind·m⁻²) til bambuskorallen og sjøfjær beregnet for hver prosesseringsmetode. Hver søyle representerer tettheten beregnet over den totale transektlengden. Mørkere fargetoner indikerer bruk av glatting, lyse fargetoner indikerer ikke-glattet, grønn farge inkludering av 3D-dybdeforskjell og blå farge bruk av 2D.
Figur 62: Total tetthet (ind·m⁻² ) til risengrynkorallen og viftesvamper beregnet for hver prosesseringsmetode. Hver søyle representerer beregnet tettheten over den totale transektlengden. Mørkere fargetoner indikerer bruk av glatting, lyse fargetoner indikerer ikke-glattet, grønn farge inkludering av 3D-dybdeforskjell og blå farge bruk av 2D.
Som forventet hadde beregning av tettheten med ulike prosesseringsmetoder motsatt effekt: bruk av glatting resulterte i en relativ stor økning av tettheten per arealet mens 3D-inkludering resulterte i en redusering i tetthet i forhold til 2D fordi transektlengden økte. På flate havbunnstransekter resulterte bruk av glatting (2D glattet) i en relativt høy prosentvis økning (mellom 28–73 %, resultater ikke vist) i tetthet hos bambuskorall og sjøfjær, sammenlignet med standardmetodikken. Detter er fordi støy i GPS posisjonene til de flate transektene var høyere. På bratte havbunnstransekter, resulterte bruk av ikke-glattet 3D-inkludering i en prosentvis reduksjon i artens tetthet (mellom -11 og -14 %, resultater ikke vist), sammenlignet med standardmetodikken. Bruk av den kombinerte metodikken (3D glattet) resulterte i en prosentvis økning i artens tetthet (mellom 8 % og 22 %, resultater ikke vist), sammenlignet med standardmetodikken.
Det anbefales å bruke glatting på flate og bratte havbunnstransekter og 3D-justering kun på bratte havbunnstransekter. Den nye prosedyren beskrevet med BVides kan være en standardisert måte for konsulentselskaper og forskningsinstitutter til å kvantifisere tetthet av videotransekter kjørt ved miljøundersøkelser. Spesielt når brukeren skal beregne tetthet i transektavsnitter (se eksempel i bakgrunn) kan dette brukes for å analysere hvordan arters tetthet i avsnitter er fordelt langs hele transektet, og hvordan tettheten forandrer seg i henhold til avstand til merd. Bruk av BVides kan være nyttig for å kvantifisere tetthet til arter tilhørende sårbare naturtyper som korall- og svampskog, og i fremtiden kan prosedyren også benyttes til å støtte bestemmelse av arters terskeltetthet.
4 - Diskusjon
For å vurdere effektene av organiske akvakulturutslipp på forvaltningsrelevante bunnsamfunn kombinerte vi modellerte sedimentasjonsdata med arts-spesifikke indikatorer, inkludert visuell helsetilstandsvurdering, lysosomal membranstabilitet, lipid- og fettsyreprofilering, respirasjonsmålinger og multi-omikk-analyser. Substrate Independent Benthic Sampler (SIBS), kombinert med Bacterial Metabarcoding Biotic Index (b-MBI), muliggjorde kvantitativ evaluering av sedimentassosierte mikrobielle samfunn i komplekse hardbunnshabitater. Denne integrerte tilnærmingen hadde som mål å oppdage subletale stressresponser, identifisere arts-spesifikke responser og forstå hvordan organisk tilførsel fra oppdrett påvirker marine økosystemer.
Sedimentasjon og b-MBI
Modellerte sedimentasjonsdata viste at avsetning av partikulært organisk materiale fra de anleggene vi undersøkte var høyest innenfor 100–200 m fra merdene, og avtok raskt med avstand. Lokal variasjon ble observert, spesielt ved Storvika i Møre og Romsdal, sannsynligvis på grunn av bratte fjordvegger som fører til ujevn sedimentakkumulering. SIBS ble med suksess tilpasset for ROV-rigging på dypt vann (< 500 m), og muliggjorde direkte prøvetaking av overflatesedimentflak og tilhørende mikrobielle samfunn fra bratte vegger og hardbunn (Keeley mfl., 2021, 2025). Denne metoden gir et pålitelig, ikke-inngripende alternativ til tradisjonell grabb-prøvetaking på hardbunnshabitat, løser “hard-bunn-problemet” og muliggjør kvantitativ miljø-DNA-prøvetaking for beregning av b-MBI.
b-MBI-analysene viste klare sammenhenger mellom sedimentasjon og mikrobiell samfunnssammensetning i sedimentet. Generaliserte additive modeller (GAMs) indikerte at kumulativ sedimentasjon og dybde, snarere enn bare avstand fra oppdrettsanlegg, var de sterkeste prediktorene for b-MBI-verdier. Lavere b-MBI-verdier ble generelt observert ved større dybder, noe som kan reflektere redusert tilførsel av organisk materiale eller lavere sedimentasjonsrater, for eksempel på grunn av strømforhold. b-MBI-verdier tilsvarte moderat til høy anrikelse nær oppdrettsanleggene og på steder med høy akkumulert sedimentasjon, og lavere verdier når sedimentasjonen avtok utover 1 km, i tråd med observasjoner fra New Zealand-fjordene (Pochon mfl., 2019; Keeley mfl., 2021). Resultatene tyder på at mikrobielle samfunn kan respondere på anrikningsgradienter langt utover områder der det er synlige bunneffekter, og fremhever metodens potensial som et tidlig varslingsverktøy for overvåking av oppdrettspåvirkning.
Likevel gjenstår flere begrensninger. b-MBI-referansedatabasen er basert på mikrober fra New Zealand og norske fjordsamfunn kan avvike fra disse, dermed er videre utvikling av en regionalt validert indikator-database nødvendig. I tillegg kan dagens tilstedeværelse/fravær-modell begrense sensitivitet, og fremtidige b-MBI-versjoner bør inkludere relativ forekomst og bedre normalisering. Til tross for disse forbeholdene gir kombinasjonen av SIBS og b-MBI en standardisert, skalerbar og minimalt invasiv tilnærming for miljøovervåking av oppdrettseffekter i dype og komplekse habitater.
Artsidentifikasjon
Artsnivåidentifikasjon er fortsatt avgjørende for vurdering av sårbarhet for oppdrettsrelatert stress. I Norge er kunnskap om marine virvelløse dyr, taksonomi og økologi vurdert til moderat til lav (score 1–3 av 5, på Artsdatabankens kunnskapsskala, der 1 = svært lav og 5 = omfattende), med svamper og steinkoraller vurdert til 3, og bløtkoraller som Alcyonacea (3) og sjøfjær (Pennatulacea, 2) har svakere kunnskapsgrunnlag (Elven og Søli, 2016). Basert på dette er det rimelig å anta at mange norske bunnarter fortsatt er ubeskrevet. Selv små DNA-forskjeller mellom nært beslektede taxa kan gi divergerende responser på miljøstress, inkludert oppdrettsutslipp. Nøyaktig artsidentifikasjon er derfor avgjørende for å vurdere arts-spesifikk toleranse, sårbarhet og respons.
I dette prosjektet ble artsidentiteten til viftesvamp bekreftet molekylært ved hjelp av COI- og 28S rRNA-markører. Alle sekvenserte prøver grupperte konsekvent med Phakellia ventilabrum, noe som bekrefter at det analyserte materialet tilhører denne arten og styrker den vitenskapelige robustheten i de arts-spesifikke analysene. COI- og 28S rRNA-markørene som ble brukt, kan forbedre referansedatabaser og fremtidig deteksjon av lokale og utbredte arter, og er også sentrale for å definere forvaltningsrelevante habitater.
Visuell helsetilstandsindeks, histologi og tetthetsvurdering
Visuelle kriterier for organismers helse ble utviklet for svamper (viftesvamp og kålrabisvamp) og koraller (risengrynkorall, sjøtre, sjøbusk og øyekorall). Denne metoden viste klare sammenhenger med stress fra oppdrettsutslipp for risengrynkorall og viftesvamp og kan brukes til å påvise effekten av organiske utslipp på disse artene ikke-invasivt i felt. De fleste risengrynkorall innenfor 500 m fra oppdrettsanlegg, der sedimentasjonsnivåene var høye, hadde relativt dårligere helsetilstand enn de som lå mer enn 500 m unna. Viftesvamp nært oppdrettsanlegg (innen 200 m) viste dårligere visuell helse enn svamper i upåvirkede områder. Den visuelle helsetilstandsindeksen for kålrabisvamp viste noe effekt av oppdrettsutslipp, men noen kålrabisvamp-individer som var utsatt for høy sedimentasjon hadde imidlertid god visuell tilstand, noe som antyder en viss motstandskraft. Den visuelle helsetilstanden til sjøbusk var dårligere nærmere oppdrettsanlegget, men vi hadde et lavt antall prøver. Sjøtre viste få visuelle endringer når de var utsatt for sedimentasjon eller nærhet til oppdrett, sannsynligvis på grunn av lavere respons for de kriteriene som ble brukt. Øyekorall viste ingen sammenheng mellom visuell helsevurdering og sedimentasjon eller nærhet til oppdrett, men prøveantallet var lite (24 individer), noe som begrenser statistisk styrke. Imidlertid er det observert fysiologiske effekter hos øyekorall i et ettårig felteksponeringsforsøk, som nedgang i metabolisme, reduserte vekst og energireserver ( Kutti mfl., 2022) . Den økte polyppaktiviteten hos noen øyekorall under høy eksponering til organiske utslipp under forsøket, for å fjerne partikler, kan føre til redusert energi til vekst. Høy sedimentasjon har tidligere blitt knyttet til redusert aktivitet hos gorgoniakoraller, overføring av sykdommer og koralldød (Tseng mfl., 2011; Pollock mfl., 2014). Misfarging eller tap av gorgoniakorallvev har også blitt knyttet til stress fra oljeeksponering (Etnoyer mfl., 2015), temperaturendringer (Canessa mfl., 2024; Gobbato mfl., 2024) og sykdom (Hall-Spencer mfl., 2007). Begroing ble brukt som et visuelt kriterium for negativ helsepåvirkning, fordi tidligere studier har koblet dekning av koraller med epibionter til sykdom eller død hos koraller (Hall-Spencer mfl., 2007). Uttynning av svampevev, slik at strukturen blir synlig, samt nedbrytning av vev er kjente indikatorer på redusert svampehelse (Luter mfl., 2011).
Den visuelle helsetilstandsindeksen viser godt potensial som metode for rutinemessig vurdering av korall- og svampetilstand rundt oppdrettslokaliteter. Resultatene indikerer en sammenheng mellom indeksen og påvirkningsgradienter for forvaltningsrelevante svamp- og korallarter. Selv om denne studien representerer ett av de største datasettene som hittil er brukt for å undersøke effekter av akvakultur på kystnære koraller og svamper, bør neste steg være videre validering av indeksen på et bredere utvalg av lokaliteter og eksponeringsgradienter. Metoden er relativt kostnadseffektiv, kan implementeres bredt og krever et lavt til middels kompetansenivå.
Visuelle observasjoner av endringer i polyppaktivitet ved bruk av et time-lapse-kamerasystem og en semiautomatisk bildeanalysemetode registrerte effekter av ulike konsentrasjoner av fiskefekalier på døgnrytmen i korallenes tentakelaktivitet. Metodens evne til å oppdage endringer var imidlertid sensitiv for utfordringer knyttet til bildedeteksjon, som forstyrrelser fra bakgrunnen, overlappende koraller og sedimentering. Derfor kunne bare et begrenset antall enkeltkoraller følges gjennom hele eksperimentperioden
For å evaluere potensielle subletale og energimessige effekter, ble histologiske analyser av gametogenese (utvikling og modning av reproduktive celler, dvs. egg og sperm) og fertilitet utført for risengrynkorall og sjøtre på ulike dybder og avstander fra oppdrettsanlegg. Ingen signifikante effekter av sesong, dybde eller nærhet ble observert, og artene viste konsistente gametogenetiske stadier på tvers av prøvetakingsperioder men effektene av sedimentasjkon ble ikke analysert. Til tross for et begrenset antall prøver gir disse analysene viktig basisinformasjon for fremtidig vurdering av effekten på reproduksjonskapasitet. De understreker at endringer i energifordeling på grunn av stress, for eksempel redusert investering i reproduksjon, bør vurderes når langsiktige effekter av utslipp fra oppdrett på økosystemet skal vurderes.
Kombinasjonen av visuelle helsevurderinger og standardiserte tetthetsestimater gir et mer helhetlig økologisk rammeverk for å tolke stressresponser på individnivå. Behovet for metodisk standardisering er tydelig, ettersom nyere studier har vist at ulike metoder for beregning av areal i ROV-transekter video kan gi systematisk forskjellige tetthetsestimater (opptil 13 % variasjon), avhengig av hvordan transektbredde, -lengde og posisjonsdata behandles (Vickery mfl., 2026). I dette prosjektet ble en standardisert prosedyre for tetthetsberegning fra videotransekter utviklet. Ved å inkludere glatting av posisjonsdata og beregning av tredimensjonal (3D) transektlengde adresserer metoden sentrale kilder til metodisk variasjon. Testing viste at valg av prosesseringsmetode har betydelig innvirkning på både estimert transektlengde og beregnet tetthet. Metoden legger dermed til rette for mer robuste og sammenlignbare tetthetsvurderinger, særlig for sårbare hardbunnsarter.
Lysosomal membranstabilitet (LMS)
LMS fungerte som en sensitiv biomarkør for subletalt stress hos svamper (viftesvamp og kålrabisvamp) og bergskjell, og reflekterte tidlige cellulære responser på miljøstress, inkludert sedimentasjon, temperatur og kjemisk eksponering (Edge mfl., 2016, 2016; Elven og Søli, 2016). Nøytralrød-retensjonsmetoden, tilpasset feltforhold, ble brukt til å kvantifisere lysosomal stabilitet og viste artsspesifikke mønstre. Kålrabisvamp viste de sterkeste og mest konsistente LMS-responsene, med signifikante effekter av alle tre miljøvariabler (avstand til anlegg, dybde og akkumulert sedimentasjon). I tråd med disse funnene viste laboratorieeksponeringer LMS-sensitivitet for suspendert fiskefôr og annet partikulært materiale, med opptil 50 % lysosomal destabilisering i kålrabisvamp etter 7 dager ved 50 g·L⁻¹ fôr (Kutti mfl., 2016a). Bergskjell viste tydelige effekter av dybde og nærhet til oppdrettsanlegg, men imidlertid var prøvestørrelsen liten, og resultatene bør tolkes med forsiktighet. Viftesvamp viste svakere mønstre, med avstand som eneste marginalt signifikante predikator i den forenklede modellen, mens ingen variabler var signifikante i fullmodellen. Disse artsspesifikke forskjellene indikerer at kålrabisvamp er mest sensitiv for sedimentasjonsrelatert cellulært stress, mens viftesvamp og bergskjell responderer mer variabelt avhengig av hvilke miljøvariabler som inkluderes i modellen.
LMS kan fungere som en tidlig indikator før synlige morfologiske endringer, og fremhever verdien av biomarkøren som tidlig-varslingsindikator. Ingen signifikant sammenheng ble funnet mellom lysosomal membrandestabilisering (LMD) og visuell helsetilstandsindeks hos svampene. Likevel kan LMS fungere som en tidlig indikator før synlige morfologiske endringer oppstår, særlig hos kålrabisvamp, når den inngår i påvirkningsindeksen, og dermed fremheve verdien av biomarkøren som et tidlig varslingsverktøy som kan fange opp stressresponser utover observerbare morfologiske endringer. Under det siste toktet ble LMS-metoden vellykket tilpasset og testet på risengrynkorall og sjøtre, noe som åpner for fremtidig anvendelse av biomarkøren også hos koraller, selv om antall prøver var for lavt til kvantitativ analyse. Under prosjektet viste LMS-metoden et sterkt potensial som et raskt og kostnadseffektivt verktøy for å vurdere fysiologisk stress hos sårbare bunnorganismer i naturen. Pågående forbedringer inkluderer mulig integrasjon av AI-basert bildeanalyse for å standardisere celletolkning og redusere observatør-bias ved manuell mikroskopi.
Respirasjon og ammoniakkutskillelse
Oksygenforbruk og ammoniakkutskillelse ble målt hos koraller (risengrynkorall, sjøtre og øyekorall). Resultatene indikerer begrensede direkte effekter av sedimentasjon eller nærhet til oppdrettsanlegg på metabolske hastigheter. Sjøtre viste en signifikant påvirkning av sedimentasjon på O₂-forbruk, og en ikke-lineær respons på avstand til oppdrettsanlegg med det høyeste oksygenforbruket observert ved mellomliggende avstander. Risengrynkorall og øyekorall viste ingen signifikante sammenhenger mellom oksygenforbruk eller ammoniakkutskillelse og de undersøkte miljøvariablene; øyekorall-analysene var d dessuten sterkt begrenset av lav prøvestørrelse (n = 7) og bør tolkes med stor forsiktighet. For ammoniakkutskillelse ble det ikke funnet signifikante sammenhenger med miljøvariabler hos noen av artene, noe som antyder at nitrogenutskillelse i hovedsak drives av metabolsk aktivitet snarere enn direkte miljøstress.
Signifikante positive korrelasjoner mellom O₂-forbruk og ammoniakkutskillelse ble funnet hos risengrynkorall og øyekorall, mens sjøtre ikke viste en slik sammenheng. Dette tyder på at nitrogenutskillelse i hovedsak drives av metabolsk aktivitet, snarere enn direkte miljøstress. Dette stemmer med litteratur som viser varierende metabolske responser hos filterfødere under sedimentasjon, der noen arter opprettholder respirasjon for å støtte slimproduksjon og partikkelfjerning (Erftemeijer mfl., 2012). Målt respirasjon er i samsvar med andre kaldtvannskoraller (Brafield og Chapman, 1965; Migné og Davoult, 1997; Dorey mfl., 2020) og med tidligere studier under emamektin benzoat eksponering (Taormina mfl., 2024a).
Lipider og fettsyreprofiler
Totalt lipidinnhold var generelt stabilt hos alle de fire studerte artene, og ingen hadde forhøyede nivåer av terrestriske fettsyrer, noe som indikerer minimalt opptak av organisk materiale fra oppdrettsfôr. Terrestriske fettsyremarkører (18:1n-9, 18:2n-6, 18:3n-3) var sporbare i alle arter, men forekom i lave andeler (< 10 % av total FA) sammenlignet med de ca. 30 % som kan finnes i laksefôr, noe som støtter at disse artene i liten grad ernærer seg direkte på oppdrettsrelatert materiale. Transplantasjonsseksperimenter støtter at fastsittende sessile arter primært ernærer seg gjennom symbionter eller plankton/detritus, og ikke laksefôr (Laroche mfl., 2021, 2022). Fraværet av forhøyede terrestriske fettsyrer og stabile marine PUFA-profiler tyder på at reproduksjonssuksess og energimetabolisme i de studerte artene sannsynligvis ikke påvirkes negativt av direkte opptak av oppdrettsfôr, og at eventuelle akvakulturrelaterte effekter i større grad skyldes indirekte miljøfaktorer som sedimentasjon fremfor redusert ernæringskvalitet.
Arts-spesifikke fettsyreprofiler reflekterte tydelige biologiske og økologiske tilpasninger. Kålrabisvamp hadde en bakteriedominert fettsyreprofil med høy andel forgrenede mettede fettsyrer (SFA ca. 45 %, hvorav ca. 38 % bakterielt avledet) og ikke-metabolske NMI-fettsyrer (ca. 27–28 %), og svært lave nivåer av essensielle n-3 og n-6 PUFA (< 4 %), konsistent med artens høy-mikrobiell-abundans (HMA)-strategi og sterke avhengighet av symbiotiske bakterier. Viftesvamp, derimot, viste en profil mer typisk for lav-mikrobiell-abundans (LMA)-svamper, dominert av langkjedede enumettede fettsyrer (MUFA i n-7-familien) og NMI-fettsyrer, med sedimentasjon som sterkeste miljødriver for endringer i fettsyresammensetning. Sjøtre hadde en karakteristisk lipidprofil dominert av langkjedede MUFA (særlig 20:1n-9 og 22:1n-11) og n-6 PUFA, supplert med et betydelig innhold av voksestere (4–6 % av total lipid) og tilhørende fettsyrealkohoer (FAOH), et distinkt lipidologisk trekk som reflekterer artens nesledyrbiologi og som ikke ble signifikant påvirket av miljøgradienter. Bergskjell viste en tydelig vevsspesifikk lipidprofil der vevstype (hel kropp vs. gjeller) hadde sterkere effekt på fettsyresammensetningen enn miljøfaktorer, med langkjedede MUFA typiske for calanus-akkumulering (20:1n-9 og 22:1n-11) og dominans av n-3 PUFA. Sedimentasjon påvirket nivåene av enkeltfettsyrer hos bergskjell, særlig 20:1n-7, 20:2n-6 og 20:4n-3, uten at den generelle fettsyreprofilen ble vesentlig endret. Samlet sett reflekterer endringene i fettsyresammensetning metabolsk tilpasning til miljøgradienter snarere enn direkte opptak av laksefôr fra oppdrettsanlegg.
Selv om FA-analyser fortsatt er et kraftig verktøy for å undersøke diett, energireserver og økologiske responser, er terrestriske FA-markører alene utilstrekkelige for å påvise oppdrettsutslippsrelaterte påvirkning i disse artene. Kombinert med andre analyser kan de imidlertid gi et mer komplett bilde av bunnorganismenes respons på organisk tilførsel og sedimentasjon.
Mikrobiomsammensetning og miljødrivere
Hos alle undersøkte taxa var mikrobielle samfunn dominert av Proteobacteria, Bacteroidetes og andre vanlige marine bakteriegrupper, men tydelige vertsspesifikke mønstre var også synlige: sjøtre-vev var beriket med Spirochaetota, bergskjellets gjeller med Campylobacterota, og kålrabisvamp med Chloroflexi. Overlapp mellom amplicon-sekvensvarianter (ASVer) hos de studerte artene og de som ble påvist i sediment (SIBS-prøver) var lavt, noe som tyder på arts-spesifikke mikrobielle samfunn, selv om høyere taksonomiske nivåer viste moderat overlapp. Predikert akkumulert sedimentasjon og dybde var de viktigste miljødriverne for mikrobielle samfunn, mens avstand fra oppdrettsanlegg hadde begrenset effekt. Dette stemmer med tidligere funn som viser at mikrobielle samfunn hos svamper og koraller er relativt stabile under subletale stressforhold, og i større grad påvirkes av abiotiske faktorer som sedimentasjon eller hydrodynamikk (Baquiran og Conaco, 2018; Laroche mfl., 2021, 2022).
Tydelig forskjell ble observert mellom viftesvamp og kålrabisvamp: viftesvamp viste et høyt antall differensielt forekommende taxa langs sedimentasjonsgradienter (opptil 114 signifikante slekter i slim og 37 i vev), mens kålrabisvamp hadde svært få (kun én slekt og én familie i vev), men med ekstremt høye effektestimater. Dette mønsteret speiler den taksonomiske spesifisiteten beskrevet i lav- og høymikrobiell-abundans (LMA/HMA) svampsystemer, og antyder at sedimentasjon modulerer vert–mikrobiom-interaksjoner forskjellig avhengig av vertens egenskaper. Samlet sett understreker disse resultatene at vert–mikrobiom-partnerskap hos virvelløse dyr er både spesialiserte og funksjonelt konvergente, og gir metabolsk fleksibilitet under miljømessige forstyrrelser.
Transkriptomiske responser
RNA-seq-analyser viste at hver art hadde distinkte sett av transkripter og metabolske veier som responderte på predikert akkumulert sedimentasjon. Sjøtre hadde det høyeste antallet differensielt uttrykte transkripter (DETs), etterfulgt av kålrabisvamp, viftesvamp og bergskjell. Bergskjell viste metabolsk modulering og aktivering av cellulære beskyttelsesveier; kålrabisvamp oppregulerte gener relatert til proteinsyntese; sjøtre viste symbiont- og virusregulerte endringer; og viftesvamp nedregulerte energisignalisering og intracellulær transport. Slike idiosynkratiske transkripsjonsresponser er konsistente med mønstre observert hos W. bursa (Laroche mfl., 2021), der differensielt uttrykte gener nær oppdrettsanlegg primært involverte ribosomal aktivitet og apoptose-regulering, mens de fleste andre metabolske veier forble stabile.
Den begrensede overlappen mellom arter, kun intracellulær transport-regulering som delt respons mellom viftesvamp og sjøtre, fremhever hvor individualiserte stressresponser er hos ulike arter. Denne individualiteten kan reflektere arts-spesifikke strategier for transkripsjonell regulering og ressursallokering under sedimentasjonsstress. I tillegg antyder frakoblingen mellom mikrobielle og transkriptomiske responser, som også notert av Laroche mfl. (2021), at vertens genuttrykk kan reagere raskere på kortsiktige stressfaktorer, mens mikrobiomstrukturen forblir buffret med mindre forstyrrelses-grenseverdier overskrides.
Epigenetikk og telomerlengde
DNA-metyleringsanalyser ble forsøkt gjennomført for kålrabisvamp, viftesvamp og sjøtre, men tekniske begrensninger, lav DNA-kvalitet hos de fleste arter og svært lav kartleggingsrate mot tilgjengelig referansegenom hos kålrabisvamp (< 20 %), gjorde at dataene ikke egnet seg for videre analyse. Ingen epigenetiske slutninger kan trekkes fra dette materialet. Erfaringene understreker behovet for fremtidig investering i høykvalitets, artsrelevante referansegenomer som en grunnleggende forutsetning for epigenetiske studier av disse organismegrupper.
Telomerlengde hos sjøtre viste ingen signifikant sammenheng med avstand til oppdrettsanlegg eller akkumulert sedimentasjon, og fanget ikke opp de samme påvirkningsgradientene som andre biomarkører i denne studien. Metoden er likevel interessant som indikator på kumulativt cellulært stress, men krever større prøvestørrelser og bredere arts-dekning før potensialet kan vurderes fullt ut (Rouan mfl., 2023).
Metodevalg, anvendelse og ressursbehov
Tabell 18 gir en sammenlignende oversikt over metodene som er brukt i prosjektet, med vekt på kostnader, tidsbruk, krav til utstyr og kompetanse, samt egnethet for overvåking og/eller forskning. Metodene spenner fra enkle og kostnadseffektive tilnærminger som kan implementeres raskt i operativ overvåking, til mer ressurs- og kompetansekrevende analyser som primært er egnet for mekanistisk forskning og metodeutvikling.
Det er viktig å merke seg at oppgitte kostnader per prøve/analyse ikke inkluderer feltinnsamling ved bruk av ROV. ROV-operasjoner krever betydelig teknisk infrastruktur, fartøy og spesialisert personell, og kan representere den største kostnadskomponenten i feltbaserte studier. Typiske dagrater ligger i størrelsesorden 30 000–150 000 NOK, mens totale toktkostnader kan variere fra omtrent 120 000 til 300 000 NOK per dag, avhengig av fartøytype, ROV-system og bemanning. Tidsrammen for gjennomføring varierer fra umiddelbar feltbasert vurdering (visuell helse) til flere uker for sekvenseringsbaserte analyser som RNA-seq og DNA-metylering.
Metode
Kostnad (per prøve / analyse) i NOK
Tidsbruk
Primært bruksområde
Kommentarer
Visuell helsevurdering
Lav (kun tidsbruk)
Rask
Overvåkning
Avhenger av bildekvalitet og observatørens erfaring; ingen labkostnader; krever begrenset spesialisert kompetanse
O₂-forbruk (respirasjon)
Lav–moderat (100-200)
Moderat
Overvåkning & forskning
Krever levende dyr; løpende kostnader er i hovedsak knyttet til drift, kalibrering og vedlikehold av utstyr etter initial investering; moderat teknisk kompetanse (erfaring med respirometriutstyr)
Ammoniakksekresjon
Lav–moderat (250-600)
Moderat
Forskning
Vanligvis kombinert med respirasjonsmålinger; reagenser og instrumentbruker; standard laboratoriekompetanse
Lysosomal membranstabilitet (LMS)
Lav–moderat (50-100)
Moderat
Overvåkning & forskning
Krever ferskt vev og mikroskopi; spesialisert laboratoriekompetanse
Lipid- og fettsyreprofiler
Moderat (100 + tidsbruk)
Moderat–lang
Forskning
Krever kjemisk ekstraksjon og kromatografiske analyser; spesialisert analytisk kompetanse
Artsidentifikasjon (morfologi)
Lav (kun tidsbruk)
Rask
Overvåkning
Avhenger av bildekvalitet og observatørens erfaring; ingen labkostnader; krever taksonomisk erfaring
Tetthetsestimering
Lav (kun tidsbruk)
Moderat-rask
Overvåking
Gratis og åpen programvare og videoannotering kan utføres ute i felt. Krever noe kompetanse innen datahåndtering og programmering.
RNA-ekstraksjon, bibliotek, dyp sekvensering; krever høy grad av spesialisert bioinformatisk kompetanse
DNA-metylering (epigenetikk) *
Høy (2200-5500 + tidsbruk)
Lang (2–6 uker)
Forskning
DNA-ekstraksjon, bibliotek, sekvensering; krever svært spesialisert analytisk og bioinformatisk kompetanse
b-MBI (benthic Microbial Biodiversity Index)
Moderat–høy (1100-1300 + tidsbruk)
Rask–moderat (avhengig av sekvenseringsdata)
Overvåkning
Basert på 16S-sekvensering med minimale tilleggskostnader; b-MBI-beregningen er inkludert i prisen og utføres ved bruk av tilgjengelig database i Cawthron; krever spesialisert økologisk og bioinformatisk kompetanse; videre dataanalyser er ikke inkludert i prisen.
Telomerer lengde
Lav–moderat (400-600 + tidsbruk)
Moderat
Forskning
Basert på DNA-ekstraksjon og kvantitativ analyse; egnet som indikator på kronisk stress og cellulær aldring; krever molekylærbiologisk kompetanse og standard laboratorieinfrastruktur
Presisjon varierer med inputdata og modell; best i kombinasjon med sedimentfeller; kreverkrever spesialisert modell- og miljøfaglig kompetanse
Tabell 18: Oversikt over metoder brukt i prosjektet, med estimert kostnad per prøve/analyse (i NOK), tidsbruk, primært bruksområde og sentrale kommentarer. Oppgitte kostnader inkluderer ikke arbeidstid knyttet til prøvehåndtering, analyser eller databehandling. Metoder merket med * innebærer sekvensering utført av ekstern tjenesteleverandør. Kostnads- og tidsestimater er veiledende og kan variere avhengig av utstyr, analysetilnærming og institusjonelle forhold.
Enkle feltbaserte metoder som visuell helsevurdering og modellert sedimentasjon krever begrenset laboratorieinfrastruktur, men forutsetter høy kvalitet på bildemateriale og erfarne observatører eller modellkompetanse. Respirasjonsmålinger og ammoniakkanalyser krever levende organismer og tilgang til respirometriutstyr og standard laboratoriefasiliteter. LMS-analyser forutsetter ferskt vev, mikroskopi og spesialisert laboratoriekompetanse. Under det siste toktet ble LMS-metoden vellykket tilpasset og testet på risengrynkorall og sjøtre, noe som åpner for fremtidig bruk av biomarkøren også på koraller. Antall tilgjengelige prøver var for lavt til kvantitativ analyse, men den metodiske gjennomførbarheten ble bekreftet. Molekylære metoder som 16S-sekvensering, RNA-seq og DNA-metylering krever molekylærbiologisk infrastruktur, tilgang til sekvenseringsplattformer (ofte via ekstern leverandør), samt betydelig bioinformatisk kapasitet for databehandling og tolkning. Disse metodene innebærer høy analytisk kompleksitet og lang behandlingstid (uker), men gir detaljert innsikt i cellulære og molekylære mekanismer. b-MBI representerer en mellomposisjon: metoden er basert på 16S-sekvensering, men beregningen kan gjennomføres raskt når sekvenseringsdata foreligger. Metoden krever bioinformatisk og økologisk kompetanse, men gir direkte anvendbare indekser for påvirkningsgrad.
Visuell helsevurdering, LMS, modellert sedimentasjon og b-MBI fremstår som mest direkte anvendbare for å dokumentere endringer i helsetilstand fra akvakulturutslipp. Disse metodene gir relativt rask respons, moderate kostnader og resultater som kan tolkes innenfor etablerte forvaltningsrammer. Respirasjonsmålinger kan anvendes både i forskning og overvåking, men krever levende materiale og logistisk tilrettelegging som kan begrense rutinemessig bruk. Lipidprofiler, mikrobiomanalyser, RNA-seq, DNA-metylering og telomerlengde er primært forskningsorienterte metoder. De gir verdifull innsikt i mekanismer og tidlige stressresponser, men krever videre standardisering og validering før de kan inngå som faste elementer i kostnadseffektiv overvåking. Prosjektet utviklet også en ny standardisert prosedyre for tetthetskvantifisering av bunnarter fra ROV-videotransekter, som kan være et viktig verktøy for fremtidig konsekvensvurdering av akvakulturutslipp på sårbare hardbunnshabitat.
Basert på denne studien fremstår modellert sedimentasjon, visuell helsevurdering, LMS og b-MBI som metoder som allerede nå kan bidra til å støtte vurdering av påvirkningsgradienter fra akvakulturutslipp og identifikasjon av sensitive arter og habitater. Molekylære responser (transkriptomikk, epigenetikk, mikrobiom) gir derimot sterk mekanistisk støtte til forståelsen av påvirkningsprosesser, men krever større datasett, standardiserte terskelverdier og kalibrering mot økologiske endepunkter før de kan brukes direkte i reguleringssammenheng.
Et foreløpig forsøk på å integrere flere metodiske tilnærminger i én samlet vurdering ble gjennomført gjennom utviklingen av en sammensatt påvirkningsindeks. Denne indeksen viser lovende potensial for å integrere flere responssignaler i én samlet vurdering, men betydelig videre arbeid er nødvendig før den kan anses som operativ i overvåking. De ulike resultatene mellom tilnærmingene for enkelte arter (særlig koraller) understreker at b-MBI og visuell helseindeks fanger opp ulike aspekter av påvirkningsgradienten, og at kombinasjonen av begge metoder gir en mer robust vurdering enn enkeltmetoder alene. Videre utvikling bør: (1) valideres mot uavhengige datasett og inkludere flere arter med større prøvestørrelser, (2) undersøke inkludering av ytterligere responsvariabler fra denne studien, herunder transkriptomiske pathway-scorer, lipiddeplesjonsmetrikker og mikrobiomindikatorer, (3) videreutvikle vektingssystemer basert på større LMS-kalibreringsdatasett, (4) teste ytelse på ulike tidsmessige skalaer og under restitusjonsscenarier, samt (5) identifisere optimale kombinasjoner av feltobserverbare og laboratorieintensive metoder med hensyn til kostnadseffektiv overvåking. Den nåværende versjonen bør derfor betraktes som en demonstrasjon av gjennomførbarhet og potensialet i multisignal-integrasjon snarere enn et ferdig utviklet overvåkingsverktøy for å dokumentere endringer i helsetilstand av forvaltningsrelevante bunnarters fra akvakulturutslipp .
Laboratorieeksponeringer
Laboratorieforsøkene bidro til å etablere toleransegrenser for øyekorallarver overfor to avlusningsmidler. Øyekorall-larver eksponert for hydrogenperoksid hadde LC50 på 27,2 mg·L⁻¹, sammenlignbart med hoppekreps (30,7 mg·L⁻¹), noe som antyder potensielle effekter innen 1 km fra oppdrettsanlegg (Refseth mfl., 2016). Eksponering for azamethiphos viste høy larvesensitivitet med en EC50 på 1,9 µg·L⁻¹ noe som er betydelig lavere enn pelagiske hummerlarver (9,2 µg·L⁻¹) og indikerer mulige effekter på nervesystem og fôring. Resultatene peker mot at hensyn bør tas i gyteperioden for øyekorall og utslipp av avlusningsmidler minimeres i denne periode. Det planlegges å gjennomføres flere eksperimenter for å verifisere resultatene.
Hos voksne øyekorall ble emamektin detektert i alle eksponerte fragmenter, med variabel biokonsentrasjon og stressrespons, inkludert økt oksygenforbruk, økt slimproduksjon og polyppdød, spesielt ved høye doser. Disse funnene står i kontrast til tidligere studier på sjøfjær, som ikke viste noen signifikante metabolske eller atferdsmessige effekter (Taormina mfl., 2024a). Økt oksygenforbruk står også i motsetning til tidligere observasjoner av redusert respirasjon etter eksponering for tralopyril og sinkpyrition i øyekorall (Hunvik, 2025), samt redusert oksygenforbruk nær anlegg med modellert høy partikulær organisk belastning (Kutti mfl., 2022). Tilsvarende førte eksponering for suspendert sediment til redusert polyppaktivitet, men bare lett økt respirasjon (Larsson mfl., 2013). Kombinasjonen av høyt oksygenforbruk og lav polyppaktivitet motsier en av de foreslåtte mekanismene for regulering av oksygenopptak hos øyekorall; ekspansjon av tentaklene for å øke vevsoverflatens kontakt med vann, noe som fører til økt oksygendiffusjon fra vannet til vevet (Dodds mfl., 2007). Eksponering for lavt til moderat stress hos organismer fører vanligvis til økt metabolsk aktivitet og ATP omsetning. Dette kan komme til uttrykk som økt oksygenforbruk og ammoniumekskresjon, noe som gjør organismen i stand til å kompensere for de ekstra energikostnadene knyttet til økt fysiologisk aktivitet, cellulært vedlikehold og reparasjon av skader (Sokolova mfl., 2012). Dette kan bidra til å forklare den økte respirasjonsraten som ble målt i begge eksponeringsgruppene, hvor alle korallene hadde opptak av emamektin i vevet. Siden virkningsmekanismen til emamektin benzoate på cellulært nivå fortsatt er ukjent, er det også ukjent hvilke cellulære responser - f.eks. ulike detoksifikasjonsmekanismer – som aktiveres i korallens celler. Dette studiet gir ikke noe endelig svar på hvordan eksponering for emamektin påvirker cellulære stressresponser og detoksifiseringsmekanismer hos koraller.
Tidsserier av polyppaktivitet av tentakkelutstrekningen i dypvannskoraller er godt dokumentert fra feltobservasjoner og er rapportert å følge døgnrytmer relatert til mattilgjengelighet, som påvirkes av faktorer som strømretning, tidevann og daglige vertikale migrasjoner av zooplankton (Buhl-Mortensen mfl., 2015; Johanson mfl., 2017; Girard mfl., 2022; Prado mfl., 2023; Salvati mfl., 2023). Korallene ser ut til å tilpasse seg disse døgnrytmesyklusene for å utnytte mat effektivt når den er tilgjengelig (tentakler utstrakt) og spare energi og redusere metabolisme når mattilgangen er begrenset (tentakler inntrukket).
Samlet sett viser eksponeringsforsøkene at fiskefekalier påvirker både total tentakkelutstrekning og døgnrytme på en artsavhengig måte. Risengrynkorall hadde den høyeste relative følsomheten for fiskefekalier sammenlignet med de andre tre korallartene. Høy dødelighet hos risengrynkorall ble observert under de høyere eksponeringene (5 og 10 g m⁻² dag⁻¹), med henholdsvis 78 % og 91 % dødelighet. Dette tilsvarer sedimentasjonsnivåer som man kan finne rett under oppdrettsanleggene som ble studert i dette prosjektet. Alle sjøtre og sjøbusk overlevde under alle eksperimentelle behandlinger, men viste betydelig sedimentering på vevet under behandlingene med 5 og 10 g m⁻² dag⁻¹. Til tross for at flere sjøtre overlevde, viste de tydelige tegn på vevsnedbrytning. Det ble heller ikke observert dødelighet hos øyekorall under noen av de eksperimentelle behandlingene, og dette var arten med minst sedimentering ved slutten av eksperimentene.
Øyekorall viste ingen statistisk påvisbar døgnrytme under noen av de testede betingelsene. Totalt aktivitetsnivå (MESOR) økte imidlertid med konsentrasjon av partikler, noe som kan indikere en økt aktivitet av tenkaler for å fjerne partikler. Denne arten klarer raskt (2–3 timer) å rense seg for små partikler som sedimenterer fra gjentatte pulser, og er også kjent for å tåle relativt høye doser av annet suspendert materiale, som for eksempel fra borevirksomhet innen olje og gass, og er generelt ganske robust mot effekter av suspendert materiale i vann (Larsson og Purser, 2011; Larsson mfl., 2013; Baussant mfl., 2018, 2022).
Sjøtre hadde den tydeligste døgnrytmen under kontrollforhold, men rytmen ble allerede dempet og faseforskjøvet ved laveste eksponeringsnivå. Tilførsel av mengden partikulært organisk materiale registert fra modellering innenfor 300 m fra oppdrettsanleggene som ble studert i dette prosjektet, forstyrret døgnrytmen hos risengrynkorall og dermed deres evne til å spise og rense vevet sitt. Disse funnene understreker at partikulært organisk materiale fra akvakultur kan ha biologiske effekter på koraller i nærheten av oppdrettsanlegg, og bør vurderes sammen med kjemiske stressorer som hydrogenperoksid og azametifos i miljøvurderinger.
Integrert perspektiv og videre utvikling
Integreringen av sedimentasjonsmodellering, b-MBI, visuelle vurderinger, LMS, lipid-analyser og transkriptomprofilering gir en helhetlig forståelse av hvordan organisk tilførsel fra akvakultur påvirker bunnøkosystemer. På tvers av arter og metodiske nivåer fremstod sedimentasjon som den dominerende stressfaktoren, med effektstørrelser som generelt oversteg både avstand og dybde. Konsistensen i responsene, fra genuttrykk og lipidmetabolisme, via cellulære biomarkører, til b-MBI, viser at partikkelavsetning er den viktigste prediktoren for biologisk stress.
Blant de studerte artene var viftesvamp mest sårbar, med synlig vevsdegradering og sterke transkriptomiske responser på sedimentasjon. Bergskjell viste større motstandskraft, med intakt struktur og relativt stabile metabolske profiler. Disse artsforskjellene reflekterer sannsynligvis variasjon i fôringsmåte, vevsarkitektur og symbiotiske assosiasjoner.
Sterk metodisk konvergens ble observert for viftesvamp og kålrabisvamp, der visuelle, cellulære, biokjemiske, mikrobielle og molekylære metoder alle avdekket dose-respons-forhold, noe som gir høy sikkerhet i vurdering av påvirkning. I kontrast viste sjøtre visuell stress som ikke fullt ut ble fanget av den sammensatte indeksen, mens kålrabisvamp viste molekylære stressresponser som oversteg synlige skader. Denne divergensen fremhever både begrensningene ved enkeltmetoder og verdien av fler-nivå-vurderinger for tidlig påvisning av subletale effekter. Sikkerheten er høyest der uavhengige metoder konvergerer, mens større usikkerhet knytter seg til molekylære endepunkter uten etablerte økologiske terskler eller standardisert kalibrering mellom arter. Selv om disse molekylære verktøyene gir verdifull mekanistisk innsikt, kreves videre validering før rutinemessig regulatorisk bruk.
Resultatene viser også at avstand fra oppdrettsanlegg alene ofte er en svak prediktor for biologisk påvirkning. Selv om avstand var statistisk signifikant for noen arter, forklarte sedimentasjon en betydelig større andel av variasjonen på tvers av metoder. Strømretning medfører dessuten store forskjeller i eksponering, slik at områder nedstrøms for dominerende strømmer kan oppleve høyere sedimentbelastning selv på større avstand enn områder som ligger nærmere men utenfor hoveddispersjonsbanen. Disse funnene understøtter et skifte fra rigide avstandsbaserte buffersoner til steds-spesifikke vurderinger basert på hydrodynamisk modellering og predikert sedimentavsetning. Modellering av dominerende strømretning og deposisjonsområder bør derfor være sentralt for å definere det faktiske påvirkningsområdet rundt akvakulturanlegg.
Sammenlagt indikerer resultatene at de undersøkte lokalitetene opplevde målbar sedimentasjonsdrevet stress, med størrelse avhengig av artssensitivitet og lokal eksponering. Mens noen arter viste motstandskraft ved observerte nivåer, hadde sensitive arter klare biologiske responser innenfor det modellerte deposisjonsområdet. Fravær av abrupt økologisk kollaps betyr ikke fravær av effekt; heller tyder bevisene på en gradient av subletale og tidlig-varslingsresponser knyttet til sedimenteksponering.
Fremtidig metodikkutvikling bør utvide biomarkørporteføljen til å inkludere indikatorer på kronisk stress (f.eks. telomerlengde), ikke-invasive proxyer som slimproduksjon, og høyoppløselige bildeverktøy som hyperspektral fotogrammetri for kvantifisering av vevstilstand og sedimentdekning. Metabolomikk og epigenetikk kan avsløre adaptive biokjemiske mekanismer, mens atferdsmålinger som polyppaktivitet kan fungere som feltbaserte, ikke-invasive stressindikatorer. Videre integrering av disse tilnærmingene, sammen med forbedret kalibrering av LMS og b-MBI, vil styrke kapasiteten for tidlig påvisning og forbedre vurdering av påvirkning i bunnhabitater.
5 - Referanser
Albretsen, J., Sperrevik, A. K., Staalstrøm, A., Sandvik, A. D., Vikebø, F., og Asplin, L. 2011. NorKyst-800 report no. 1: User manual and technical descriptions. Fisken og havet, 2.
Baltadakis, A., Casserly, J., Falconer, L., Sprague, M., og Telfer, T. C. 2020. European lobsters utilise Atlantic salmon wastes in coastal integrated multi-trophic aquaculture systems. Aquaculture Environment Interactions, 12: 485–494. https://www.int-res.com/abstracts/aei/v12/aei00378.
Bankevich, A., Nurk, S., Antipov, D., Gurevich, A. A., Dvorkin, M., Kulikov, A. S., Lesin, V. M., mfl. 2012. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. Journal of Computational Biology, 19: 455–477. https://doi.org/10.1089/cmb.2012.0021.
Bannister, R. J., Johnsen, I. A., Hansen, P. K., Kutti, T., og Asplin, L. 2016. Near- and far-field dispersal modelling of organic waste from Atlantic salmon aquaculture in fjord systems. ICES Journal of Marine Science, 73: 2408–2419. https://doi.org/10.1093/icesjms/fsw027.
Baussant, T., Nilsen, M., Ravagnan, E., Westerlund, S., og Ramanand, S. 2018. Effects of suspended drill cuttings on the coral Lophelia pertusa using pulsed and continuous exposure scenarios. Journal of Toxicology and Environmental Health, Part A, 81: 361–382. https://doi.org/10.1080/15287394.2018.1444375.
Baussant, T., Arnberg, M., Lyng, E., Ramanand, S., Bamber, S., Berry, M., Myrnes Hansen, I., mfl. 2022. Identification of tolerance levels on the cold-water coral Desmophyllum pertusum (Lophelia pertusa) from realistic exposure conditions to suspended bentonite, barite and drill cutting particles. PLOS ONE, 17: e0263061. https://doi.org/10.1371/journal.pone.0263061.
Bekkby, T., Rinde, E., Oug, E., Buhl-Mortensen, P., Thormar, J., Dolan, M., Mjelde, M., mfl. 2021. Forslag til forvaltningsrelevante marine naturenheter.
Bell, J. G., McGhee, F., Campbell, P. J., og Sargent, J. R. 2003. Rapeseed oil as an alternative to marine fish oil in diets of post-smolt Atlantic salmon (Salmo salar): changes in flesh fatty acid composition and effectiveness of subsequent fish oil «wash out». Aquaculture, 218: 515–528. https://www.sciencedirect.com/science/article/pii/S0044848602004623.
Benskin, J. P., Ikonomou, M. G., Surridge, B. D., Dubetz, C., og Klaassen, E. 2016. Biodegradation potential of aquaculture chemotherapeutants in marine sediments. Aquaculture Research, 47: 482–497. https://onlinelibrary.wiley.com/doi/abs/10.1111/are.12509.
Benson, V., Aldous, E., og Clementson, A. 2017. Review of environmental quality standard for emamectin benzoate. Report.
Bolger, A. M., Lohse, M., og Usadel, B. 2014. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics, 30: 2114–2120. https://doi.org/10.1093/bioinformatics/btu170.
Brafield, A. E., og Chapman, G. 1965. The oxygen consumption of Pennatula rubra ellis and some other anthozoans. Zeitschrift für vergleichende Physiologie, 50: 363–370. https://doi.org/10.1007/BF00339424.
Brooke, S., og Järnegren, J. 2013. Reproductive periodicity of the scleractinian coral Lophelia pertusa from the Trondheim Fjord, Norway. Marine Biology, 160: 139–153. https://doi.org/10.1007/s00227-012-2071-x.
Buhl-Mortensen, P., Tenningen, E., og Tysseland, A. B. S. 2015. Effects of water flow and drilling waste exposure on polyp behaviour in Lophelia pertusa. Marine Biology Research, 11: 725–737. https://doi.org/10.1080/17451000.2014.993651.
Callahan, B. J., McMurdie, P. J., Rosen, M. J., Han, A. W., Johnson, A. J. A., og Holmes, S. P. 2016. DADA2: High-resolution sample inference from Illumina amplicon data. Nature Methods, 13: 581–583. https://doi.org/10.1038/nmeth.3869.
Canessa, M., Bertolotto, R., Betti, F., Bo, M., Dagnino, A., Enrichetti, F., Toma, M., mfl. 2024. Variation in the Health Status of the Mediterranean Gorgonian Forests: The Synergistic Effect of Marine Heat Waves and Fishing Activity. Biology, 13: 642. http://dx.doi.org/10.3390/biology13080642.
Carvajalino-Fernández, M. A., Sævik, P. N., Johnsen, I. A., Albretsen, J., og Keeley, N. B. 2020. Simulating particle organic matter dispersal beneath Atlantic salmon fish farms using different resuspension approaches. Marine Pollution Bulletin, 161: 111685. https://www.sciencedirect.com/science/article/pii/S0025326X20308031.
Chapron, L., Galand, P. E., Pruski, A. M., Peru, E., Vétion, G., Robin, S., og Lartaud, F. 2021. Resilience of cold-water coral holobionts to thermal stress. Proceedings of the Royal Society B: Biological Sciences, 288: 20212117. https://doi.org/10.1098/rspb.2021.2117.
Chombard, C., Boury-Esnault, N., og Tillier, S. 1998. Reassessment of Homology of Morphological Characters in Tetractinellid Sponges Based on Molecular Data. Systematic Biology, 47: 351–366. https://doi.org/10.1080/106351598260761.
Consortium, T. U. 2024. UniProt: the Universal Protein Knowledgebase in 2025. Nucleic Acids Research, 53: D609–D617. https://doi.org/10.1093/nar/gkae1010.
Cubillo, A. M., Ferreira, J. G., Robinson, S. M. C., Pearce, C. M., Corner, R. A., og Johansen, J. 2016. Role of deposit feeders in integrated multi-trophic aquaculture — A model analysis. Aquaculture, 453: 54–66. https://www.sciencedirect.com/science/article/pii/S0044848615302593.
De Castro-Fernández, P., Ballesté, E., Angulo-Preckler, C., Biggs, J., Avila, C., og García-Aljaro, C. 2023. How does heat stress affect sponge microbiomes? Structure and resilience of microbial communities of marine sponges from different habitats. Frontiers in Marine Science, Volume 9 - 2022. https://www.frontiersin.org/journals/marine-science/articles/10.3389/fmars.2022.1072696.
Díaz, J. A., Ramírez-Amaro, S., og Ordines, F. 2021. Sponges of Western Mediterranean seamounts: new genera, new species and new records. PeerJ, 9: e11879. https://doi.org/10.7717/peerj.11879.
Dodds, L. A., Roberts, J. M., Taylor, A. C., og Marubini, F. 2007. Metabolic tolerance of the cold-water coral Lophelia pertusa (Scleractinia) to temperature and dissolved oxygen change. Journal of Experimental Marine Biology and Ecology, 349: 205–214. https://www.sciencedirect.com/science/article/pii/S0022098107002559.
Dunlop, K., Harendza, A., Bannister, R., og Keeley, N. 2021. Spatial response of hard- and mixed-bottom benthic epifauna to organic enrichment from salmon aquaculture in northern Norway. Aquaculture Environment Interactions, 13: 455–475. https://www.int-res.com/abstracts/aei/v13/p455-475/.
Dunlop, K. M., Gomes, A. S., og Fuhrmann, M. M. under utarbeidelse. Protokoll for visuelle vurderinger av helsetilstand hos kystnære koraller, svamper og bergskjell i nærheten av oppdrettsanlegg. Report.
Edgar, R. C. 2004a. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research, 32: 1792–1797. https://doi.org/10.1093/nar/gkh340.
Edgar, R. C. 2004b. MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC Bioinformatics, 5: 113. https://doi.org/10.1186/1471-2105-5-113.
Edge, K. J., Dafforn, K. A., Simpson, S. L., Roach, A. C., og Johnston, E. L. 2014. A biomarker of contaminant exposure is effective in large scale assessment of ten estuaries. Chemosphere, 100: 16–26. https://www.sciencedirect.com/science/article/pii/S0045653514000356.
Edge, K. J., Dafforn, K. A., Simpson, S. L., Ringwood, A. H., og Johnston, E. L. 2015. Resuspended contaminated sediments cause sublethal stress to oysters: A biomarker differentiates total suspended solids and contaminant effects. Environmental Toxicology and Chemistry, 34: 1345–1353. https://doi.org/10.1002/etc.2929.
Edge, K. J., Johnston, E. L., Dafforn, K. A., Simpson, S. L., Kutti, T., og Bannister, R. J. 2016. Sub-lethal effects of water-based drilling muds on the deep-water sponge Geodia barretti. Environmental Pollution, 212: 525–534. https://www.sciencedirect.com/science/article/pii/S0269749116301737.
El-Saber Batiha, G., Alqahtani, A., Ilesanmi, O. B., Saati, A. A., El-Mleeh, A., Hetta, H. F., og Magdy Beshbishy, A. 2020. Avermectin Derivatives, Pharmacokinetics, Therapeutic and Toxic Dosages, Mechanism of Action, and Their Biological Effects. Pharmaceuticals, 13: 196. https://www.mdpi.com/1424-8247/13/8/196.
Elven, H., og Søli, G. 2016. Kunnskapsstatus for artsmangfoldet i Norge 2015. Utredning for Artsdatabanken, 1: 2016.
Erftemeijer, P. L. A., Riegl, B., Hoeksema, B. W., og Todd, P. A. 2012. Environmental impacts of dredging and other sediment disturbances on corals: A review. Marine Pollution Bulletin, 64: 1737–1765. https://www.sciencedirect.com/science/article/pii/S0025326X12001981.
Escobar-Lux, R. H., Fields, D. M., Browman, H. I., Shema, S. D., Bjelland, R. M., Agnalt, A.-L., Skiftesvik, A. B., mfl. 2019. The effects of hydrogen peroxide on mortality, escape response, and oxygen consumption of Calanus spp. FACETS, 4: 626–637. https://www.facetsjournal.com/doi/abs/10.1139/facets-2019-0011.
Escobar-Lux, R. H., Parsons, A. E., Samuelsen, O. B., og Agnalt, A.-L. 2020. Short-term exposure to hydrogen peroxide induces mortality and alters exploratory behaviour of European lobster (Homarus gammarus). Ecotoxicology and Environmental Safety, 204: 111111. https://www.sciencedirect.com/science/article/pii/S0147651320309507.
Etnoyer, P. J., Wickes, L. N., Silva, M., Dubick, J. D., Balthis, L., Salgado, E., og MacDonald, I. R. 2015. Decline in condition of gorgonian octocorals on mesophotic reefs in the northern Gulf of Mexico: before and after the Deepwater Horizon oil spill. Coral Reefs, 35: 77–90. http://dx.doi.org/10.1007/s00338-015-1363-2.
Fernandez-Jover, D., Martinez-Rubio, L., Sanchez-Jerez, P., Bayle-Sempere, J. T., Lopez Jimenez, J. A., Martínez Lopez, F. J., Bjørn, P.-A., mfl. 2011. Waste feed from coastal fish farms: A trophic subsidy with compositional side-effects for wild gadoids. Estuarine, Coastal and Shelf Science, 91: 559–568. https://www.sciencedirect.com/science/article/pii/S0272771410004099.
Folmer, O., Black, M., Hoeh, W., Lutz, R., og Vrijenhoek, R. 1994. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol, 3: 294–9.
Food, og Organization, A. 2009. International guidelines for the management of deep-sea fisheries in the high seas. Book. FAO.
Girard, F., Cruz, R., Glickman, O., Harpster, T., og Fisher, C. R. 2019. In situ growth of deep-sea octocorals after the Deepwater Horizon oil spill. Elementa: Science of the Anthropocene, 7. https://doi.org/10.1525/elementa.349.
Girard, F., Litvin, S. Y., Sherman, A., McGill, P., Gannon, A., Lovera, C., DeVogelaere, A., mfl. 2022. Phenology in the deep sea: seasonal and tidal feeding rhythms in a keystone octocoral. Proceedings of the Royal Society B: Biological Sciences, 289. https://doi.org/10.1098/rspb.2022.1033.
Gobbato, J., Work, T. M., Facchinelli, M. P., Siena, F. M., Montalbetti, E., Seveso, D., Louis, Y. D., mfl. 2024. Pathology of tissue loss in three key gorgonian species in the Mediterranean Sea. Journal of Invertebrate Pathology, 207: 108197. http://dx.doi.org/10.1016/j.jip.2024.108197.
Gomes, A. S., Guerreiro, E. M., Pochon, X., Keeley, N., og Laroche, O. 2025. A Comparative Protocol for Preserving Deep-Water Marine Invertebrate Tissues: DNA/RNA Shield vs. Liquid Nitrogen for Dual Extraction of High-Quality Nucleic Acids. Bio-protocol, 15: e5521. https://doi.org/10.21769/BioProtoc.5521.
Gómez-Gras, D., Bensoussan, N., Ledoux, J. B., López-Sendino, P., Cerrano, C., Ferretti, E., Kipson, S., mfl. 2022. Exploring the response of a key Mediterranean gorgonian to heat stress across biological and spatial scales. Scientific Reports, 12: 21064. https://doi.org/10.1038/s41598-022-25565-9.
Gonzalez-Silvera, D., Izquierdo-Gomez, D., Sanchez-Jerez, P., Elbal, M. T., López-Jiménez, J. A., og Martínez-López, F. J. 2020. Influence of aquaculture waste on fatty acid profiles and gonad maturation of wild fish aggregations at fish farms. Marine Environmental Research, 156: 104902. https://www.sciencedirect.com/science/article/pii/S0141113619306506.
Grabherr, M. G., Haas, B. J., Yassour, M., Levin, J. Z., Thompson, D. A., Amit, I., Adiconis, X., mfl. 2011. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nature Biotechnology, 29: 644–652. https://doi.org/10.1038/nbt.1883.
Hall-Spencer, J., Pike, J., og Munn, C. 2007. Diseases affect cold-water corals too: Eunicella verrucosa (Cnidaria: Gorgonacea) necrosis in SW England. Diseases of Aquatic Organisms, 76: 87–97. http://dx.doi.org/10.3354/dao076087.
Hamoutene, D., Salvo, F., Egli, S. N., Modir-Rousta, A., Knight, R., Perry, G., Bottaro, C. S., mfl. 2018. Measurement of Aquaculture Chemotherapeutants in Flocculent Matter Collected at a Hard-Bottom Dominated Finfish Site on the South Coast of Newfoundland (Canada) After 2 Years of Fallow. Frontiers in Marine Science, Volume 5 - 2018. https://www.frontiersin.org/journals/marine-science/articles/10.3389/fmars.2018.00228.
Handå, A., Ranheim, A., Olsen, A. J., Altin, D., Reitan, K. I., Olsen, Y., og Reinertsen, H. 2012. Incorporation of salmon fish feed and feces components in mussels (Mytilus edulis): Implications for integrated multi-trophic aquaculture in cool-temperate North Atlantic waters. Aquaculture, 370-371: 40–53. https://www.sciencedirect.com/science/article/pii/S0044848612005674.
Hart, A. J., Ginzburg, S., Xu, M., Fisher, C. R., Rahmatpour, N., Mitton, J. B., Paul, R., mfl. 2020. EnTAP: Bringing faster and smarter functional annotation to non-model eukaryotic transcriptomes. Molecular Ecology Resources, 20: 591–604. https://onlinelibrary.wiley.com/doi/abs/10.1111/1755-0998.13106.
Hofmann, G. E., og Gaines, S. D. 2008. New Tools to Meet New Challenges: Emerging Technologies for Managing Marine Ecosystems for Resilience. BioScience, 58: 43–52. https://doi.org/10.1641/B580109.
Holmes, R. M., Aminot, A., Kérouel, R., Hooker, B. A., og Peterson, B. J. 1999. A simple and precise method for measuring ammonium in marine and freshwater ecosystems. Canadian Journal of Fisheries and Aquatic Sciences, 56: 1801–1808. https://cdnsciencepub.com/doi/abs/10.1139/f99-128.
Huerta-Cepas, J., Szklarczyk, D., Heller, D., Hernández-Plaza, A., Forslund, S. K., Cook, H., Mende, D. R., mfl. 2018. eggNOG 5.0: a hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Research, 47: D309–D314. https://doi.org/10.1093/nar/gky1085.
Hunvik, G. 2025. Impacts of new aquaculture antifouling agents tralopyril and zinc pyrithione on the deep-water coral Lophelia pertusa (Linnaeus, 1758). https://hdl.handle.net/11250/3210757.
Keeley, N., Valdemarsen, T., Woodcock, S., Holmer, M., Husa, V., og Bannister, R. 2019. Resilience of dynamic coastal benthic ecosystems in response to large-scale finfish farming. Aquaculture Environment Interactions, 11: 161–179. https://www.int-res.com/abstracts/aei/v11/aei00301.
Keeley, N., Laroche, O., Birch, M., og Pochon, X. 2021. A Substrate-Independent Benthic Sampler (SIBS) for Hard and Mixed-Bottom Marine Habitats: A Proof-of-Concept Study. Frontiers in Marine Science, 8. https://www.frontiersin.org/articles/10.3389/fmars.2021.627687.
Keeley, N., Dunlop, K., Laroche, O., Hansen, P. K., Angell, I. L., og Rudi, K. 2025. An approach for quantifying the influence of fish farm waste on hard-bottom habitats. Marine Pollution Bulletin, 217: 118039. https://www.sciencedirect.com/science/article/pii/S0025326X25005144.
Keeley, N. B., Forrest, B. M., Crawford, C., og Macleod, C. K. 2012. Exploiting salmon farm benthic enrichment gradients to evaluate the regional performance of biotic indices and environmental indicators. Ecological Indicators, 23: 453–466. https://www.sciencedirect.com/science/article/pii/S1470160X12001975.
Kumar, S., Stecher, G., Suleski, M., Sanderford, M., Sharma, S., og Tamura, K. 2024. MEGA12: Molecular Evolutionary Genetic Analysis Version 12 for Adaptive and Green Computing. Molecular Biology and Evolution, 41. https://doi.org/10.1093/molbev/msae263.
Kutti, T., Bannister, R. J., og Fosså, J. H. 2013. Community structure and ecological function of deep-water sponge grounds in the Traenadypet MPA—Northern Norwegian continental shelf. Continental Shelf Research, 69: 21–30. https://www.sciencedirect.com/science/article/pii/S0278434313003051.
Kutti, T., Nordbø, K., Bannister, R. J., og Husa, V. 2015. Oppdrettsanlegg kan true korallrev i fjordene. Report. Havforskningsinstituttet.
Kutti, T., Ólafsdóttir, S. H., og Burgos, J. M. 2016b. Health state of cold-water coral and sponge populations in Norwegian and Icelandic waters. Institute of Marine Research; Marine & Freshwater Research Institute.
Kutti, T., Legrand, E., Husa, V., Olsen, S. A., Gjelsvik, Ø., Carvajalino-Fernandez, M., og Johnsen, I. A. 2022. Fish farm effluents cause metabolic depression, reducing energy stores and growth in the reef-forming coral Lophelia pertusa. Aquaculture Environment Interactions, 14: 279–293. https://www.int-res.com/abstracts/aei/v14/p279-293/.
Langford, K. H., Øxnevad, S., Schøyen, M., og Thomas, K. V. 2014. Do Antiparasitic Medicines Used in Aquaculture Pose a Risk to the Norwegian Aquatic Environment? Environmental Science & Technology, 48: 7774–7780. https://doi.org/10.1021/es5005329.
Laroche, O., Meier, S., Mjøs, S. A., og Keeley, N. 2022. Suspension-Feeding Benthic Species’ Physiological and Microbiome Response to Salmon Farming and Associated Environmental Changes. Frontiers in Marine Science, 9. https://www.frontiersin.org/articles/10.3389/fmars.2022.841806.
Larsson, A. I., og Purser, A. 2011. Sedimentation on the cold-water coral Lophelia pertusa: Cleaning efficiency from natural sediments and drill cuttings. Marine Pollution Bulletin, 62: 1159–1168. http://dx.doi.org/10.1016/j.marpolbul.2011.03.041.
Larsson, A. I., Oevelen, D. van, Purser, A., og Thomsen, L. 2013. Tolerance to long-term exposure of suspended benthic sediments and drill cuttings in the cold-water coral Lophelia pertusa. Marine Pollution Bulletin, 70: 176–188. https://www.sciencedirect.com/science/article/pii/S0025326X13001136.
Love, M. I., Huber, W., og Anders, S. 2014. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biology, 15: 550. https://doi.org/10.1186/s13059-014-0550-8.
Lowe, D. M., Moore, M. N., og Evans, B. M. 1992. Contaminant impact on interactions of molecular probes with lysosomes in living hepatocytes from dab Limanda limanda. Marine Ecology Progress Series, 91: 135–140. http://www.jstor.org/stable/24832651.
Luter, H. M., Whalan, S., og Webster, N. S. 2011. The marine sponge Ianthella basta can recover from stress-induced tissue regression. In Ancient Animals, New Challenges, pp. 227–235. http://dx.doi.org/10.1007/978-94-007-4688-6_19.
Madgett, A. S., Yates, K., Webster, L., McKenzie, C., og Moffat, C. F. 2019. Understanding marine food web dynamics using fatty acid signatures and stable isotope ratios: Improving contaminant impacts assessments across trophic levels. Estuarine, Coastal and Shelf Science, 227: 106327. https://www.sciencedirect.com/science/article/pii/S0272771419300411.
Mamaca, E., Bechmann, R. K., Torgrimsen, S., Aas, E., Bjørnstad, A., Baussant, T., og Floch, S. L. 2005. The neutral red lysosomal retention assay and Comet assay on haemolymph cells from mussels (Mytilus edulis) and fish (Symphodus melops) exposed to styrene. Aquatic Toxicology, 75: 191–201. https://www.sciencedirect.com/science/article/pii/S0166445X05002596.
McKnight, D. T., Huerlimann, R., Bower, D. S., Schwarzkopf, L., Alford, R. A., og Zenger, K. R. 2019. microDecon: A highly accurate read-subtraction tool for the post-sequencing removal of contamination in metabarcoding studies. Environmental DNA, 1: 14–25. https://onlinelibrary.wiley.com/doi/abs/10.1002/edn3.11.
Meier, S., Mjøs, S. A., Joensen, H., og Grahl-Nielsen, O. 2006. Validation of a one-step extraction/methylation method for determination of fatty acids and cholesterol in marine tissues. Journal of Chromatography A, 1104: 291–298. https://www.sciencedirect.com/science/article/pii/S0021967305022065.
Meier, S., Meeren, T. van der, Skjæraasen, J. E., Bannister, R. J., Rasinger, J. D., og Karlsen, Ø. 2023. Terrestrial fatty acids from feed oil in feed for farmed salmonids are transferred to the liver, gonads, and muscle of wild Atlantic cod (Gadus morhua). ICES Journal of Marine Science, 80: 1143–1154. https://doi.org/10.1093/icesjms/fsad051.
Migné, A., og Davoult, D. 1997. Oxygen consumption in two benthic cnidarians: Alcyonium digitatum (Linnaeus, 1758) and Urticina felina (Linnaeus, 1767). In Proc. 6th int. Conf. on Coelenterate Biology, pp. 321–328.
Mukherjee, S., Ray, M., Dutta, M. K., Acharya, A., Mukhopadhyay, S. K., og Ray, S. 2015. Morphological alteration, lysosomal membrane fragility and apoptosis of the cells of Indian freshwater sponge exposed to washing soda (sodium carbonate). Ecotoxicology and Environmental Safety, 122: 331–342. https://www.sciencedirect.com/science/article/pii/S0147651315300579.
Norge, O. 2024. Species and Habitats of Environmental Concern: Mapping, Risk Assessment, Mitigation and Monitoring. - In Relation to Offshore Activities. Report, 2019-007, Rev. 1. Offshore Norge.
O’Leary, N. A., Wright, M. W., Brister, J. R., Ciufo, S., Haddad, D., McVeigh, R., Rajput, B., mfl. 2015. Reference sequence (RefSeq) database at NCBI: current status, taxonomic expansion, and functional annotation. Nucleic Acids Research, 44: D733–D745. https://doi.org/10.1093/nar/gkv1189.
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., Minchin, P. R., mfl. 2020. vegan: Community ecology package (2.5-7). Ordination methods, diversity analysis and other functions for community and vegetation ecologists.
Olsen, R. E., og Henderson, R. J. 1989. The rapid analysis of neutral and polar marine lipids using double-development HPTLC and scanning densitometry. Journal of Experimental Marine Biology and Ecology, 129: 189–197. https://www.sciencedirect.com/science/article/pii/0022098189900567.
Olsen, S. A., Ervik, A., og Grahl-Nielsen, O. 2009. Deep-water shrimp (Pandalus borealis, Krøyer 1838) as indicator organism for fish-farm waste. Journal of Experimental Marine Biology and Ecology, 381: 82–89. https://www.sciencedirect.com/science/article/pii/S0022098109003943.
Olsen, S. A., Ervik, A., og Grahl-Nielsen, O. 2012. Tracing fish farm waste in the northern shrimp Pandalus borealis (Krøyer, 1838) using lipid biomarkers. Aquaculture Environment Interactions, 2: 133–144. https://www.int-res.com/abstracts/aei/v2/aei00036.
Oppen, M. J. H. van, og Coleman, M. A. 2022. Advancing the protection of marine life through genomics. PLOS Biology, 20: e3001801. https://doi.org/10.1371/journal.pbio.3001801.
Palladino, G., Rampelli, S., Scicchitano, D., Musella, M., Quero, G. M., Prada, F., Mancuso, A., mfl. 2021. Impact of Marine Aquaculture on the Microbiome Associated with Nearby Holobionts: The Case of Patella caerulea Living in Proximity of Sea Bream Aquaculture Cages. Microorganisms, 9: 455. https://www.mdpi.com/2076-2607/9/2/455.
Parsons, A. E., Escobar-Lux, R. H., Sævik, P. N., Samuelsen, O. B., og Agnalt, A.-L. 2020. The impact of anti-sea lice pesticides, azamethiphos and deltamethrin, on European lobster (Homarus gammarus) larvae in the Norwegian marine environment. Environmental Pollution, 264: 114725. https://www.sciencedirect.com/science/article/pii/S0269749120302451.
Parsons, A. E., Escobar-Lux, R. H., Hannisdal, R., Agnalt, A.-L., og Samuelsen, O. B. 2025. Anti-Sea Lice Veterinary Medicinal Products on Salmon Farms: A Review and Analysis of Their Usage Patterns, Environmental Fate and Hazard Potential. Reviews in Aquaculture, 17: e13006. https://onlinelibrary.wiley.com/doi/abs/10.1111/raq.13006.
Patro, R., Duggal, G., Love, M. I., Irizarry, R. A., og Kingsford, C. 2017. Salmon provides fast and bias-aware quantification of transcript expression. Nature Methods, 14: 417–419. https://doi.org/10.1038/nmeth.4197.
Pochon, X., Wood, S., Atalah, J., Laroche, O., Zaiko, A., og Keeley, N. 2019. A Validated Protocol for Benthic Monitoring of New Zealand’s Salmon Farms Using Environmental DNA. Report. Cawthron Institute.
Pochon, X., Atalah, J., Fletcher, L., Laroche, O., Elvines, D., Zaiko, A., Wood, S., mfl. 2021. A Molecular Index Refined for Benthic Monitoring of Salmon Farms in the Marlborough Sounds. Report. Cawthron Institute.
Pollock, F. J., Lamb, J. B., Field, S. N., Heron, S. F., Schaffelke, B., Shedrawi, G., Bourne, D. G., mfl. 2014. Sediment and Turbidity Associated with Offshore Dredging Increase Coral Disease Prevalence on Nearby Reefs. PLoS ONE, 9: e102498. http://dx.doi.org/10.1371/journal.pone.0102498.
Prado, E., Abad-Uribarren, A., Ramo, R., Sierra, S., González-Pola, C., Cristobo, J., Ríos, P., mfl. 2023. Describing Polyps Behavior of a Deep-Sea Gorgonian, Placogorgia sp., Using a Deep-Learning Approach. Remote Sensing, 15: 2777. https://www.mdpi.com/2072-4292/15/11/2777.
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., Peplies, J., mfl. 2012. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Research, 41: D590–D596. https://doi.org/10.1093/nar/gks1219.
RCoreTeam. 2025. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. https://www.R-project.org/.
Redmond, K. J., Magnesen, T., Hansen, P. K., Strand, Ø., og Meier, S. 2010. Stable isotopes and fatty acids as tracers of the assimilation of salmon fish feed in blue mussels (Mytilus edulis). Aquaculture, 298: 202–210. https://www.sciencedirect.com/science/article/pii/S0044848609008291.
Refseth, G., Sæther, K., Drivdal, M., Nøst, O., Augustine, S., Camus, L., Tassara, L., mfl. 2016. Miljørisiko ved bruk av hydrogenperoksid Økotoksikologisk vurdering og grenseverdi for effekt. Report.
Reigel, A. M., og Hellberg, M. E. 2023. Microbiome environmental shifts differ between two co-occurring octocoral hosts. Marine Ecology Progress Series, 720: 59–83. https://www.int-res.com/abstracts/meps/v720/meps14401.
Ringwood, A. H., Conners, D. E., og Hoguet, J. 1998. Effects of natural and anthropogenic stressors on lysosomal destabilization in oysters Crassostrea virginica. Marine Ecology Progress Series, 166: 163–171. https://www.int-res.com/abstracts/meps/v166/meps166163.
Ringwood, A. H., Conners, D. E., Hoguet, J., og Ringwood, L. A. 2005. Lysosomal destabilization assays for estuarine organisms. In Techniques in Aquatic Toxicology, Volume 2, 1st Edition, pp. 287–300. Red. G. K. Ostrander. CRC Press.
Robertson, G., Schein, J., Chiu, R., Corbett, R., Field, M., Jackman, S. D., Mungall, K., mfl. 2010. De novo assembly and analysis of RNA-seq data. Nature Methods, 7: 909–912. https://doi.org/10.1038/nmeth.1517.
Rodriguez-Casariego, J. A., Cunning, R., Baker, A. C., og Eirin-Lopez, J. M. 2022. Symbiont shuffling induces differential DNA methylation responses to thermal stress in the coral Montastraea cavernosa. Molecular Ecology, 31: 588–602. https://onlinelibrary.wiley.com/doi/abs/10.1111/mec.16246.
Rodríguez-Casariego, J. A., Mercado-Molina, A. E., Garcia-Souto, D., Ortiz-Rivera, I. M., Lopes, C., Baums, I. B., Sabat, A. M., mfl. 2020. Genome-Wide DNA Methylation Analysis Reveals a Conserved Epigenetic Response to Seasonal Environmental Variation in the Staghorn Coral Acropora cervicornis. Frontiers in Marine Science, Volume 7 - 2020. https://www.frontiersin.org/journals/marine-science/articles/10.3389/fmars.2020.560424.
Ronquist, F., og Huelsenbeck, J. P. 2003. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics, 19: 1572–1574. https://doi.org/10.1093/bioinformatics/btg180.
Rouan, A., Pousse, M., Djerbi, N., Porro, B., Bourdin, G., Carradec, Q., Hume, B. C. C., mfl. 2023. Telomere DNA length regulation is influenced by seasonal temperature differences in short-lived but not in long-lived reef-building corals. Nature Communications, 14: 3038. https://doi.org/10.1038/s41467-023-38499-1.
Salvati, E. V. A., Provenzani, C., D’Ambrosi, A., Finoia, M. G., Romano, E., og Canese, S. 2023. In situ observation of circadian rhythm of polyps’ opening and closing of the coral Dendrophyllia ramea (Linnaeus, 1758). Mediterranean Marine Science, 24: 574–585. https://ejournals.epublishing.ekt.gr/index.php/hcmr-med-mar-sc/article/view/34250.
Scanes, E., Kutti, T., Fang, J. K. H., Johnston, E. L., Ross, P. M., og Bannister, R. J. 2018. Mine Waste and Acute Warming Induce Energetic Stress in the Deep-Sea Sponge Geodia atlantica and Coral Primnoa resedeaformis; Results From a Mesocosm Study. Frontiers in Marine Science, Volume 5 - 2018. https://www.frontiersin.org/journals/marine-science/articles/10.3389/fmars.2018.00129.
Scanes, E., Kutti, T., Fang, J. K. H., Johnston, E. L., Ross, P. M., og Bannister, R. J. 2024. The long-lived deep-sea bivalve Acesta excavata is sensitive to the dual stressors of sediment and warming. Marine Pollution Bulletin, 202: 116323. https://www.sciencedirect.com/science/article/pii/S0025326X2400300X.
Seppey, M., Manni, M., og Zdobnov, E. M. 2019. BUSCO: Assessing Genome Assembly and Annotation Completeness. In Gene Prediction: Methods and Protocols, pp. 227–245. Red. M. Kollmar. Springer New York, New York, NY. https://doi.org/10.1007/978-1-4939-9173-0_14.
Shchepetkin, A. F., og McWilliams, J. C. 2005. The regional oceanic modeling system (ROMS): a split-explicit, free-surface, topography-following-coordinate oceanic model. Ocean Modelling, 9: 347–404. https://www.sciencedirect.com/science/article/pii/S1463500304000484.
Smith-Unna, R., Boursnell, C., Patro, R., Hibberd, J. M., og Kelly, S. 2016. TransRate: reference-free quality assessment of de novo transcriptome assemblies. Genome Research, 26: 1134–1144. http://genome.cshlp.org/content/26/8/1134.abstract.
Sokolova, I. M., Frederich, M., Bagwe, R., Lannig, G., og Sukhotin, A. A. 2012. Energy homeostasis as an integrative tool for assessing limits of environmental stress tolerance in aquatic invertebrates. Marine Environmental Research, 79: 1–15. https://www.sciencedirect.com/science/article/pii/S0141113612000712.
Soneson, C., Love, M., og Robinson, M. 2016. Differential analyses for RNA-seq: transcript-level estimates improve gene-level inferences. F1000Research, 4. https://f1000research.com/articles/4-1521/v2.
Song, L., og Florea, L. 2015. Rcorrector: efficient and accurate error correction for Illumina RNA-seq reads. GigaScience, 4: 48. https://doi.org/10.1186/s13742-015-0089-y.
Strand, R., Whalan, S., Webster, N. S., Kutti, T., Fang, J. K. H., Luter, H. M., og Bannister, R. J. 2017. The response of a boreal deep-sea sponge holobiont to acute thermal stress. Scientific Reports, 7: 1660. https://doi.org/10.1038/s41598-017-01091-x.
Subramanian, A., Tamayo, P., Mootha, V. K., Mukherjee, S., Ebert, B. L., Gillette, M. A., Paulovich, A., mfl. 2005. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proceedings of the National Academy of Sciences, 102: 15545–15550. https://www.pnas.org/doi/abs/10.1073/pnas.0506580102.
Sutherland, T. F., Sterling, A. M., og Ou, M. 2018. Influence of salmonid aquaculture activities on a rock-cliff epifaunal community in Jervis Inlet, British Columbia. Marine Pollution Bulletin, 127: 297–309. https://www.sciencedirect.com/science/article/pii/S0025326X17310329.
Suzek, B. E., Wang, Y., Huang, H., McGarvey, P. B., Wu, C. H., og Consortium, the U. 2014. UniRef clusters: a comprehensive and scalable alternative for improving sequence similarity searches. Bioinformatics, 31: 926–932. https://doi.org/10.1093/bioinformatics/btu739.
Søvik, G., Olsen, S. A., Hansen, A., Stiansen, S., Nilssen, E. M., Bjørn, P. A., og Meier, S. 2025. Fatty acid composition of northern shrimp Pandalus borealis in relation to salmon aquaculture locations in northern Norway. Aquaculture Environment Interactions, 17: 161–174. https://www.int-res.com/abstracts/aei/v17/aei00500.
Taormina, B., Escobar-Lux, R. H., Legrand, E., Parsons, A. E., Kutti, T., Husa, V., Hannisdal, R., mfl. 2024a. Effects of the sea lice chemotherapeutant, emamectin benzoate, on metabolism and behaviour of the sea-pen Pennatula phosphorea. Marine Pollution Bulletin, 198: 115903. https://www.sciencedirect.com/science/article/pii/S0025326X23013383.
Taormina, B., Kutti, T., Olsen, S. A., Sævik, P. N., Hannisdal, R., Husa, V., og Legrand, E. 2024b. Effects of aquaculture effluents on the slender sea pen Virgularia mirabilis. Scientific Reports, 14: 9385. https://doi.org/10.1038/s41598-024-59613-3.
Tjensvoll, I., Kutti, T., Fosså, J. H., og Bannister, R. J. 2013. Rapid respiratory responses of the deep-water sponge Geodia barretti exposed to suspended sediments. Aquatic Biology, 19: 65–73. https://www.int-res.com/abstracts/ab/v19/ab00522.
Tseng, L.-C., Dahms, H.-U., Hsu, N.-J., og Hwang, J.-S. 2011. Effects of sedimentation on the gorgonian Subergorgia suberosa (Pallas, 1766). Marine Biology, 158: 1301–1310. http://dx.doi.org/10.1007/s00227-011-1649-z.
Vad, J., Orejas, C., Moreno-Navas, J., Findlay, H. S., og Roberts, J. M. 2017. Assessing the living and dead proportions of cold-water coral colonies: implications for deep-water Marine Protected Area monitoring in a changing ocean. PeerJ, 5: e3705. https://doi.org/10.7717/peerj.3705.
Van Geest, J. L., Burridge, L. E., og Kidd, K. A. 2014. Toxicity of two pyrethroid-based anti-sea lice pesticides, AlphaMax® and Excis®, to a marine amphipod in aqueous and sediment exposures. Aquaculture, 434: 233–240. https://www.sciencedirect.com/science/article/pii/S0044848614004177.
Vargas, S., Zimmer, T., Conci, N., Lehmann, M., og Wörheide, G. 2022. Transcriptional response of the calcification and stress response toolkits in an octocoral under heat and pH stress. Molecular Ecology, 31: 798–810. https://onlinelibrary.wiley.com/doi/abs/10.1111/mec.16266.
Vellani, V., Oliva, M., Pretti, C., og Renzi, M. 2025. Stress-Related Molecular Biomarkers to Monitor the Effects of Global Changes on Calcifying Reef-Forming Organisms: A Review in the Mediterranean. Journal of Marine Science and Engineering, 13: 4. https://www.mdpi.com/2077-1312/13/1/4.
Vickery, G., Zimmermann, F., Thompson, F., og Hvingel, C. 2026. Area estimation methods in underwater video surveys: Biases, errors and their impacts on density estimates. Ecological Informatics: 103648. https://www.sciencedirect.com/science/article/pii/S1574954126000543.
Waller, R. G., og Tyler, P. A. 2005. The reproductive biology of two deep-water, reef-building scleractinians from the NE Atlantic Ocean. Coral Reefs, 24: 514–522. https://doi.org/10.1007/s00338-005-0501-7.
Wang, Q., Garrity, G. M., Tiedje, J. M., og Cole, J. R. 2007. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Applied and Environmental Microbiology, 73: 5261–5267. https://journals.asm.org/doi/abs/10.1128/aem.00062-07.
Weinnig, A. M., Gómez, C. E., Hallaj, A., og Cordes, E. E. 2020. Cold-water coral (Lophelia pertusa) response to multiple stressors: High temperature affects recovery from short-term pollution exposure. Scientific Reports, 10: 1768. https://doi.org/10.1038/s41598-020-58556-9.
White, C. A., Bannister, R. J., Dworjanyn, S. A., Husa, V., Nichols, P. D., Kutti, T., og Dempster, T. 2017. Consumption of aquaculture waste affects the fatty acid metabolism of a benthic invertebrate. Science of The Total Environment, 586: 1170–1181. https://www.sciencedirect.com/science/article/pii/S0048969717303571.
White, C. A., Bannister, R. J., Dworjanyn, S. A., Husa, V., Nichols, P. D., og Dempster, T. 2018. Aquaculture-derived trophic subsidy boosts populations of an ecosystem engineer. Aquaculture Environment Interactions, 10: 279–289. https://www.int-res.com/abstracts/aei/v10/aei00270.
Wickham, H. 2016. ggplot2: Elegant Graphics for Data Analysis. Book. Springer-Verlag, New York, NY. https://ggplot2.tidyverse.org.
Woodcock, S. H., Troedsson, C., Strohmeier, T., Balseiro, P., Skaar, K. S., og Strand, Ø. 2017. Combining biochemical methods to trace organic effluent from fish farms. Aquaculture Environment Interactions, 9: 429–443. https://www.int-res.com/abstracts/aei/v9/aei00242.
Woodcock, S. H., Strohmeier, T., Strand, Ø., Olsen, S. A., og Bannister, R. J. 2018. Mobile epibenthic fauna consume organic waste from coastal fin-fish aquaculture. Marine Environmental Research, 137: 16–23. https://www.sciencedirect.com/science/article/pii/S0141113617307432.
Woodcock, S. H., Meier, S., Keeley, N. B., og Bannister, R. J. 2019. Fate and longevity of terrestrial fatty acids from caged fin-fish aquaculture in dynamic coastal marine systems. Ecological Indicators, 103: 43–54. https://www.sciencedirect.com/science/article/pii/S1470160X19302389.
Yang, X., Jiang, G., Zhang, Y., Wang, N., Zhang, Y., Wang, X., Zhao, F.-J., mfl. 2023. MBPD: A multiple bacterial pathogen detection pipeline for One Health practices. iMeta, 2: e82. https://onlinelibrary.wiley.com/doi/abs/10.1002/imt2.82.
Ziegler, M., Grupstra, C. G. B., Barreto, M. M., Eaton, M., BaOmar, J., Zubier, K., Al-Sofyani, A., mfl. 2019. Coral bacterial community structure responds to environmental change in a host-specific manner. Nature Communications, 10: 3092. https://doi.org/10.1038/s41467-019-10969-5.
Tabell 19: Oversikt over ROV-dykk og antall individer av utvalgte bunndyr som ble prøvetatt ved ulike avstander fra oppdrettsanlegg i Møre og Romsdal og Nordland. Tabellen inkluderer lokalitetsnavn, dykk-ID, koordinater for midtpunkter, avstand fra anlegget, dato, dybdeintervall og antall observerte nøkkelarter.
6.2 - Vedlegg A2
Figur 63: Kart over lokaliteter for fauna-prøvetaking (fargede) og Substrate Independent Benthic Sampler (SIBS)-prøvetaking (svarte) for beregning av Bakteriell Meta-strekkoding Biotisk Indeks (b-MBI) ved studieoppdrettsanlegg i Møre og Romsdal.
Figur 64: Kart over lokaliteter for fauna-prøvetaking (fargede) og Substrate Independent Benthic Sampler (SIBS)-prøvetaking (svarte) for beregning av Bakteriell Meta-strekkoding Biotisk Indeks (b-MBI) ved studieoppdrettsanlegg i Nordland.
6.3 - Vedlegg A3
Figur 65: Dendrogram basert på bayesisk fylogeni av Phakellia basert på mitokondrielt COI. Treet er rotet med Phakellia hirondellei. Tallene over grenene viser bayesiske posterior-sannsynligheter (MrBayes 3.2.7), der verdier over 0,95 anses som sterkt støttet, mens tallene under grenene viser bootstrap-verdier fra en maksimum likelihood-fylogeni, der verdier over 75 anses som sterkt støttet (MEGA 11). Skalaen viser grenlengder fra den bayesiske analysen.
Figur 66: Dendrogram basert på bayesisk fylogeni av Phakellia basert på genomisk 28S rRNA. Treet er rotet med Phakellia hirondellei. Tallene over grenene viser bayesiske posterior-sannsynligheter (MrBayes 3.2.7), der verdier over 0,95 anses som sterkt støttet, mens tallene under grenene viser bootstrap-verdier fra en maksimum likelihood-fylogeni, der verdier over 75 anses som sterkt støttet (MEGA 11). Skalaen viser grenlengder fra den bayesiske analysen.
6.4 - Vedlegg A4
Figur 67: Mikroskopbilde av A) stabile og B) hovne lysosomalceller fra kålrabisvamp, viftesvamp, bergskjell.
6.5 - Vedlegg A5
Variable
Nær oppdrett (< 250 m, n=7)
Referanse (> 750 m, n=5)
Lipid %
9,49 ± 2,47
7,27 ± 1,84
14:0
1,83 ± 0,28
2,21 ± 0,51
16:0
7,90 ± 1,10
8,83 ± 1,92
18:0
1,69 ± 0,14
1,79 ± 0,22
∑SFA
12,97 ± 1,43
14,47 ± 2,93
16:1n-7
1,23 ± 0,17
1,33 ± 0,10
18:1n-9
5,01 ± 0,85
5,20 ± 0,76
18:1n-7
1,39 ± 0,23
1,15 ± 0,23
20:1n-9
13,91 ± 1,68
12,05 ± 2,61
20:1n-7
2,07 ± 0,45
1,67 ± 0,50
22:1n-11
21,46 ± 2,03
19,40 ± 3,90
∑MUFA
48,84 ± 4,07
44,49 ± 7,63
20:4n-6
13,23 ± 2,49
16,34 ± 3,55
22:4n-6
1,28 ± 0,16
1,39 ± 0,22
24:5n-6
4,81 ± 0,78
5,96 ± 1,52
20:4n-3
1,28 ± 0,12
1,09 ± 0,24
20:5n-3
5,03 ± 0,52
4,68 ± 0,30
22:6n-3
1,96 ± 0,37
2,03 ± 0,34
24:5n-3
1,00 ± 0,07
0,89 ± 0,12
24:6n-3
4,31 ± 0,28
3,93 ± 0,27
∑PUFA
38,19 ± 2,84
41,04 ± 4,72
∑PUFA (n-6)
20,87 ± 2,99
25,00 ± 5,02
∑PUFA (n-3)
16,77 ± 0,81
15,49 ± 1,00
Ether lipid 16:0
2,88 ± 0,77
3,69 ± 1,37
Ether lipid 16:1
0,98 ± 0,30
1,51 ± 0,97
Ether lipid 18:0
0,58 ± 0,15
0,70 ± 0,20
∑ Ether lipid
4,45 ± 1,21
5,90 ± 2,52
FAOH-profil (% av totale FAOH)
14:0 ALK
1,67 ± 0,50
2,03 ± 0,29
16:0 ALK
15,35 ± 2,82
16,84 ± 0,69
18:0 ALK
1,05 ± 0,18
1,18 ± 0,12
16:1n-7 ALK
3,35 ± 0,44
3,36 ± 0,56
18:1n-9 ALK
1,74 ± 0,20
1,72 ± 0,11
18:1n-7 ALK
1,05 ± 0,19
1,00 ± 0,07
20:1n-9 ALK
17,31 ± 0,93
16,78 ± 0,81
20:1n-7 ALK
1,98 ± 0,23
1,92 ± 0,20
22:1n-11 ALK
44,88 ± 1,99
44,70 ± 1,53
22:1n-9 ALK
5,64 ± 1,38
4,84 ± 0,71
22:1n-7 ALK
2,72 ± 0,41
1,99 ± 0,86
Tabell 20: Lipidinnhold og fettsyresammensetning i prøver av sjøtre fra ulike avstander fra oppdrett, vist som gjennomsnitt ± standardavvik (n angitt for hver gruppe).
Variabel
Nær oppdrett (< 250 m, n=11)
Referanse (> 750 m, n=7)
Lipid %
3,02 ± 1,33
3,91 ± 0,93
14:0
2,89 ± 1,85
1,69 ± 0,69
i-15:0
1,84 ± 0,66
1,68 ± 0,22
16:0
4,94 ± 2,42
4,32 ± 1,37
i-17:0
0,77 ± 0,29
1,00 ± 0,09
18:0
4,15 ± 4,85
1,70 ± 0,81
20:0
1,39 ± 0,93
1,35 ± 0,07
ai21:0
2,45 ± 1,25
1,74 ± 0,81
22:0
0,51 ± 0,10
0,55 ± 0,12
Phytanic acid
5,33 ± 1,28
4,30 ± 1,67
∑SFA
33,73 ± 11,73
25,76 ± 0,60
16:1n-7
2,99 ± 0,63
3,14 ± 0,93
18:1n-9
2,16 ± 1,47
2,32 ± 1,59
18:1n-7
1,62 ± 0,68
1,30 ± 0,27
20:1n-9
1,38 ± 0,41
1,40 ± 0,44
20:1n-7
1,55 ± 0,40
1,67 ± 0,26
22:1n-11
2,60 ± 1,57
2,74 ± 0,79
22:1n-9
0,19 ± 0,20
0,17 ± 0,08
22:1n-7
4,39 ± 3,05
7,01 ± 0,94
24:1n-9
0,69 ± 0,43
1,24 ± 0,30
24:1n-7
5,50 ± 2,93
6,27 ± 0,66
26:1n-7
0,23 ± 0,17
0,18 ± 0,07
∑MUFA
30,80 ± 4,64
35,09 ± 2,59
18:2n-6
0,61 ± 0,48
0,78 ± 0,60
20:4n-6
3,57 ± 0,90
2,95 ± 0,40
18:4n-3
0,40 ± 0,23
0,88 ± 0,44
20:5n-3
16,17 ± 4,05
18,22 ± 2,93
22:6n-3
2,43 ± 0,95
3,24 ± 0,88
25:2D5,9 NMI
3,68 ± 2,02
4,13 ± 0,67
26:2D5,9 NMI
1,90 ± 1,27
2,91 ± 0,31
28:2D5,9 NMI
1,00 ± 0,22
0,84 ± 0,26
29:2D5,9 NMI
0,56 ± 0,26
0,36 ± 0,17
∑PUFA
34,71 ± 5,99
38,05 ± 2,42
∑PUFA (n-6)
5,08 ± 1,03
4,26 ± 0,52
∑PUFA (n-3)
20,82 ± 4,10
23,97 ± 2,01
∑PUFA (NMI)
7,18 ± 2,83
8,29 ± 1,19
Tabell 21: Lipidinnhold og fettsyresammensetning i prøver av viftesvamp fra ulike avstander fra oppdrett, vist som gjennomsnitt ± standardavvik (n angitt for hver gruppe).
Variabel
Nær oppdrett (< 250 m, n=4)
Referanse (>750 m, n=5)
Lipid % (mg/100 mg dry weight)
2,26 ± 0,36
2,45 ± 0,37
14:0
1,12 ± 0,04
1,10 ± 0,09
i-15:0
3,14 ± 0,12
3,28 ± 0,26
ai-15:0
2,31 ± 0,32
2,31 ± 0,17
16:0
2,91 ± 0,10
3,05 ± 0,36
Br 17:0 (8Me)
1,50 ± 0,33
1,53 ± 0,15
Br 17:0 (9Me)
1,40 ± 0,23
1,13 ± 0,38
Br 17:0 (10Me)
2,68 ± 0,10
2,84 ± 0,22
i-17:0
1,01 ± 0,20
1,16 ± 0,12
ai-17:0
1,64 ± 0,09
1,63 ± 0,02
18:0
2,33 ± 0,09
2,46 ± 0,08
Br 19:0 (10Me)
4,10 ± 0,35
3,92 ± 0,49
Br 19:0 (11Me)
6,11 ± 0,46
6,60 ± 0,46
22:00:00
1,11 ± 0,16
0,83 ± 0,20
i-23:0
1,09 ± 0,25
0,87 ± 0,21
∑SFA
44,83 ± 0,79
45,08 ± 1,93
∑ Bacteria-SFA
37,57 ± 0,71
37,94 ± 1,77
∑ Other-SFA
8,38 ± 0,25
8,22 ± 0,37
16:1 (n-7)
4,84 ± 0,17
4,91 ± 0,31
16:1 n-5
1,27 ± 0,13
1,28 ± 0,07
Br 17:1n-x_2
3,70 ± 0,27
4,05 ± 0,31
18:1 n-7
2,52 ± 0,18
2,63 ± 0,11
Br 18:1 n-x, xMe_2
1,23 ± 0,10
1,29 ± 0,09
Br 18:1 n-x, xMe_3
1,19 ± 0,07
1,25 ± 0,12
25:1 n-7
1,68 ± 0,39
1,54 ± 0,14
∑MUFA
22,54 ± 0,57
23,23 ± 0,88
∑ Bacteria MUFA
11,27 ± 0,68
12,04 ± 0,46
∑ Other MUFA
6,63 ± 0,34
6,49 ± 0,47
22:6 (n-3)
1,68 ± 0,17
1,90 ± 0,51
18:2d5,9 NMI
2,47 ± 0,14
2,30 ± 0,28
26:2D5,9 NMI
5,58 ± 0,25
4,98 ± 0,53
26:2D9,19 NMI
3,06 ± 0,54
2,74 ± 0,18
26:3D5,9,19 NMI
1,02 ± 0,09
0,85 ± 0,09
27:2Dxx NMI_1
1,70 ± 0,04
1,75 ± 0,11
27:2Dxx NMI_2
1,02 ± 0,35
1,04 ± 0,22
27:2Dxx NMI_3
1,97 ± 0,25
2,09 ± 0,33
27:2Dxx NMI_4
2,85 ± 0,34
2,59 ± 0,28
27:2Dxx NMI_6
1,23 ± 0,10
1,27 ± 0,14
27:2Dxx NMI_7
1,29 ± 0,13
1,23 ± 0,18
28:2Dxx NMI_4
1,34 ± 0,19
1,34 ± 0,32
∑PUFA
32,63 ± 1,21
31,70 ± 2,51
∑PUFA (n-6)
0,52 ± 0,04
0,60 ± 0,23
∑PUFA (n-3)
3,11 ± 0,38
3,49 ± 0,70
∑PUFA (NMI)
28,60 ± 0,97
27,17 ± 2,17
Tabell 22: Lipidinnhold og fettsyresammensetning i prøver av kålrabisvamp fra ulike avstander fra oppdrett, vist som gjennomsnitt ± standardavvik (n angitt for hver gruppe).
Variable
Nær oppdrett (<250 m, n=4)
Middels avstand (250–750 m, n=4)
Referanse (>750 m, n=3)
Lipid (%)
18,39 ± 6,75
15,24 ± 3,47
14,30 ± 7,15
14:0
4,13 ± 0,34
3,85 ± 0,42
2,91 ± 0,38
i-15:0
0,08 ± 0,01
0,07 ± 0,01
0,08 ± 0,06
ai-15:0
0,10 ± 0,04
0,10 ± 0,03
0,08 ± 0,06
15:0
0,27 ± 0,06
0,28 ± 0,06
0,22 ± 0,13
i-16:0
0,10 ± 0,02
0,10 ± 0,03
0,10 ± 0,06
16:0
11,79 ± 0,89
11,23 ± 0,76
10,23 ± 2,19
i-17:0
0,78 ± 0,19
0,81 ± 0,13
0,99 ± 0,48
ai-17:0
0,37 ± 0,06
0,37 ± 0,05
0,40 ± 0,21
17:0
0,54 ± 0,12
0,56 ± 0,08
0,64 ± 0,26
i-18:0
0,39 ± 0,12
0,40 ± 0,12
0,50 ± 0,26
18:0
2,37 ± 0,45
2,33 ± 0,12
2,49 ± 0,70
20:0
0,21 ± 0,08
0,21 ± 0,06
0,28 ± 0,14
22:0
0,06 ± 0,04
0,04 ± 0,03
0,11 ± 0,10
4,8,12-Me 13:0
0,81 ± 0,12
0,80 ± 0,06
0,61 ± 0,14
∑SFA
21,19 ± 1,71
20,34 ± 0,87
19,03 ± 4,54
16:1n-7
4,27 ± 0,40
4,42 ± 0,47
4,45 ± 0,53
16:1n-5
0,27 ± 0,01
0,27 ± 0,01
0,24 ± 0,02
16:1n-10,7Me (b)
0,16 ± 0,02
0,15 ± 0,03
0,16 ± 0,03
18:1n-11
0,37 ± 0,08
0,31 ± 0,06
0,42 ± 0,04
18:1n-9
2,75 ± 0,69
2,99 ± 0,93
2,43 ± 0,57
18:1n-7
2,39 ± 0,59
2,34 ± 0,50
2,77 ± 0,33
18:1n-5
0,46 ± 0,05
0,48 ± 0,02
0,45 ± 0,02
20:1n-11
1,28 ± 0,24
1,28 ± 0,11
1,12 ± 0,14
20:1n-9
15,85 ± 1,56
15,04 ± 1,13
16,17 ± 4,21
20:1n-7
1,35 ± 0,55
1,24 ± 0,57
1,48 ± 0,29
22:1n-11
11,29 ± 2,22
11,67 ± 1,39
10,14 ± 3,56
22:1n-9
0,59 ± 0,08
0,60 ± 0,08
0,53 ± 0,19
22:1n-7
0,35 ± 0,03
0,32 ± 0,03
0,50 ± 0,42
24:1n-9
0,30 ± 0,04
0,30 ± 0,05
0,29 ± 0,11
∑MUFA
41,67 ± 2,68
41,41 ± 1,59
41,17 ± 8,19
18:2n-6
0,65 ± 0,18
0,68 ± 0,18
0,53 ± 0,18
20:2n-6
1,59 ± 0,47
1,49 ± 0,50
1,74 ± 0,37
20:4n-6
1,21 ± 0,28
1,31 ± 0,10
1,75 ± 0,88
22:4n-6
0,17 ± 0,05
0,18 ± 0,04
0,20 ± 0,05
22:5n-6
0,26 ± 0,03
0,28 ± 0,02
0,34 ± 0,05
18:3n-3
1,09 ± 0,19
1,16 ± 0,15
0,78 ± 0,05
18:4n-3
1,85 ± 0,63
2,00 ± 0,57
0,96 ± 0,28
20:3n-3
0,56 ± 0,16
0,53 ± 0,18
0,55 ± 0,20
20:4n-3
1,36 ± 0,36
1,26 ± 0,34
1,17 ± 0,44
20:5n-3
7,82 ± 0,81
7,87 ± 0,41
7,42 ± 0,05
22:5n-3
1,79 ± 0,12
1,89 ± 0,08
2,40 ± 0,53
22:6n-3
13,48 ± 1,01
14,31 ± 0,56
15,49 ± 3,15
24:5n-3
0,87 ± 0,10
0,88 ± 0,06
1,28 ± 0,78
24:6n-3
0,22 ± 0,04
0,23 ± 0,01
0,27 ± 0,09
∑PUFA
32,93 ± 1,70
34,07 ± 0,96
34,88 ± 2,28
∑PUFA (n-6)
3,88 ± 0,42
3,93 ± 0,33
4,56 ± 0,80
∑PUFA (n-3)
29,05 ± 1,34
30,14 ± 0,71
30,32 ± 1,54
20:2 D5,11 NMI
0,60 ± 0,03
0,58 ± 0,10
0,65 ± 0,08
20:2 D5,13 NMI
0,11 ± 0,03
0,11 ± 0,03
0,14 ± 0,05
20:3 D5,11,14 NMI
0,38 ± 0,12
0,39 ± 0,03
0,49 ± 0,26
22:2 D7,13 NMI
1,30 ± 0,23
1,25 ± 0,09
1,87 ± 0,65
22:2 D7,15 NMI
0,19 ± 0,05
0,20 ± 0,03
0,25 ± 0,09
22.3 D7,13,16 NMI
0,58 ± 0,28
0,60 ± 0,26
0,64 ± 0,18
∑PUFA (NMI)
3,15 ± 0,54
3,13 ± 0,15
4,04 ± 1,21
16:0-2OH
0,16 ± 0,04
0,15 ± 0,02
0,16 ± 0,10
17:0-2OH
0,04 ± 0,01
0,03 ± 0,02
0,06 ± 0,03
17:0-2OH
0,05 ± 0,02
0,05 ± 0,02
0,06 ± 0,05
∑ Hydroksylerte FA
0,24 ± 0,04
0,24 ± 0,03
0,28 ± 0,11
∑MAE+DMA
4,05 ± 0,44
4,15 ± 0,39
4,48 ± 2,72
∑Kolesterol
2,23 ± 0,22
2,22 ± 0,14
3,52 ± 1,56
∑ Desmosterol
0,35 ± 0,09
0,35 ± 0,06
0,84 ± 0,59
Tabell 23: Lipidinnhold og fettsyresammensetning i prøver av bergskjell fra ulike avstander fra oppdrett, vist som gjennomsnitt ± standardavvik (n angitt for hver gruppe).
Figur 68: Sammenheng mellom andel destabiliserte lysosomale membranceller og utvalgte fosfolipidfettsyrer hos bergskjell. Prediksjoner fra beta-regresjonsmodeller (logit-link) med 95 % konfidensintervall vises for: (A) 20:5n-3 (EPA, PL), (B) 22:6n-3 (DHA, PL), (C) totale PUFA (PL), (D) totale n-3 PUFA (PL) og (E) totale n-6 PUFA (PL). Punkter viser observerte data, og linjer med skygge viser modellprediksjoner med konfidensintervall.
6.6 - Vedlegg A6
Oppsummering av differensielt uttrykte mikrobielle taxa i prøvene
Miljøgradient
Art
Prøvetype
Rang
Antall signifikante taxa
Høyeste positive effekt
Høyeste negative effekt
Median effekt
Avstand
Sjøtre
slim
Genus
1
-3,544
-3,544
-3,544
Risengrynkorall
vev
Family
1
3,944
3,944
3,944
Risengrynkorall
vev
Genus
1
3,944
3,944
3,944
Kålrabisvamp
vev
Family
1
14,382
14,382
14,382
Kålrabisvamp
vev
Genus
1
14,382
14,382
14,382
Sedimentasjon
Sjøtre
slim
Genus
1
-3,477
-3,477
-3,477
Risengrynkorall
slim
Family
25
3,731
-1,741
2,417
Risengrynkorall
slim
Genus
25
3,731
-1,741
2,417
Risengrynkorall
vev
Family
24
4,481
-4,715
1,579
Risengrynkorall
vev
Genus
24
4,481
-4,715
1,579
Viftesvamp
slim
Family
66
2,459
-2,883
1,415
Viftesvamp
slim
Genus
114
2,726
-3,040
1,387
Viftesvamp
vev
Family
29
3,455
-2,691
0,866
Viftesvamp
vev
Genus
37
3,853
-3,011
1,014
Kålrabisvamp
vev
Family
1
14,539
14,539
14,539
Kålrabisvamp
vev
Genus
1
14,539
14,539
14,539
Bergskjell
vev
Genus
2
4,324
2,959
3,642
SIBS
Family
156
1,783
-1,446
0,835
SIBS
Genus
156
1,783
-1,446
0,835
Dybde
Viftesvamp
slim
Family
22
2,796
1,158
1,926
Viftesvamp
slim
Genus
35
3,695
-1,840
2,026
Viftesvamp
vev
Family
20
4,178
-0,395
2,198
Viftesvamp
vev
Genus
29
4,589
-0,395
2,592
Kålrabisvamp
vev
Family
1
9,741
9,741
9,741
Kålrabisvamp
vev
Genus
1
9,741
9,741
9,741
Bergskjell
vev
Family
1
-5,354
-5,354
-5,354
Bergskjell
vev
Genus
1
-5,354
-5,354
-5,354
SIBS
Family
172
2,076
-1,640
0,714
SIBS
Genus
172
2,076
-1,640
0,714
Tabell 24: Oppsummering av differensielt uttrykte mikrobielle taxa (slekt og familie) på tvers av arter, prøvetyper og miljøgradienter. Miljøgradienten angir faktoren som differensialanalyse ble utført langs: avstand til anlegg, akkumulert sedimentasjon eller dybde. Tabellen viser antall signifikante taxa, største positive og negative effektestimat og median effektestimat.
Felles og unike taxa mellom vertsarter og SIBS
Figur 69: Overlapp av bakterielle familier og ASV-er mellom vev, slim fra sjøtre og SIBS-prøver. Øverste panel: overlapp på familienivå. Nederste panel: overlapp på ASV-nivå. Delte og unike funksjoner er angitt for hver prøvetype.
Figur 70: Delte bakterielle familier og ASV-er mellom vev, slim fra risengrynkorall og SIBS-prøver. Øverste panel viser overlapp på familienivå, nederste panel viser overlapp på ASV-nivå. Tallene angir unike og delte funksjoner.
Figur 71: Delte bakterielle familier og ASV-er mellom vev, slim fra viftesvamp og SIBS-prøver. Overlapp på familienivå vises i øverste panel, overlapp på ASV-nivå i nederste panel. Tallene viser unike og delte funksjoner for hver prøvetype.
Figur 72: Delte bakterielle familier og ASV-er mellom vev fra kålrabisvamp og SIBS-prøver. Øverste panel viser overlapp på familienivå, og nederste panel viser overlapp på ASV-nivå. Tallene viser unike og delte funksjoner mellom prøvetypene.
Figur 73: Delte bakterielle familier og ASV-er mellom vev fra bergskjell og SIBS-prøver. Øverste panel viser overlapp på familienivå, mens nederste panel viser overlapp på ASV-nivå. Størrelsen på settene indikerer antall unike eller delte funksjoner for hver prøvetype.
Taksonomisk sammensetning av patogene taxa
Figur 74: Relativ forekomst (%) av bakterielle patogener i vev og slim for de ulike artene (sjøtre, risengrynkorall, viftesvamp, kålrabisvamp, bergskjell) og SIBS-prøver i forhold til avstand til anlegg (< 250 m, 250–750 m, > 750 m). Fargene viser bakterielle rekker og familier, med rekker markert i legendens overskrift. Panelene er delt etter art og prøvetype.
Figur 75: Relativ forekomst (%) av bakterielle patogener i vev og slim for de ulike artene (sjøtre, risengrynkorall, viftesvamp, kålrabisvamp, bergskjell) og SIBS-prøver i forhold til akkumulert sedimentasjon (< 50, 50–1000, > 1000 g·m⁻²). Fargene viser bakterielle rekker og familier, med rekker markert i legendens overskrift. Panelene er delt etter art og prøvetype.
6.7 - Vedlegg A7
Figur 76: Gjennomsnittlig polyppaktivitet over tid, gjennomsnittet beregnet på tvers av replikater innen hver behandling, med skyggede bånd som representerer ± SD. Behandlingene er fargekodet: Kontroll (teal), 0,5 g·m⁻²·dag⁻¹ (blå), 5 g·m⁻²·dag⁻¹ (oransje) og 10 g·m⁻²·dag⁻¹ (rosa).
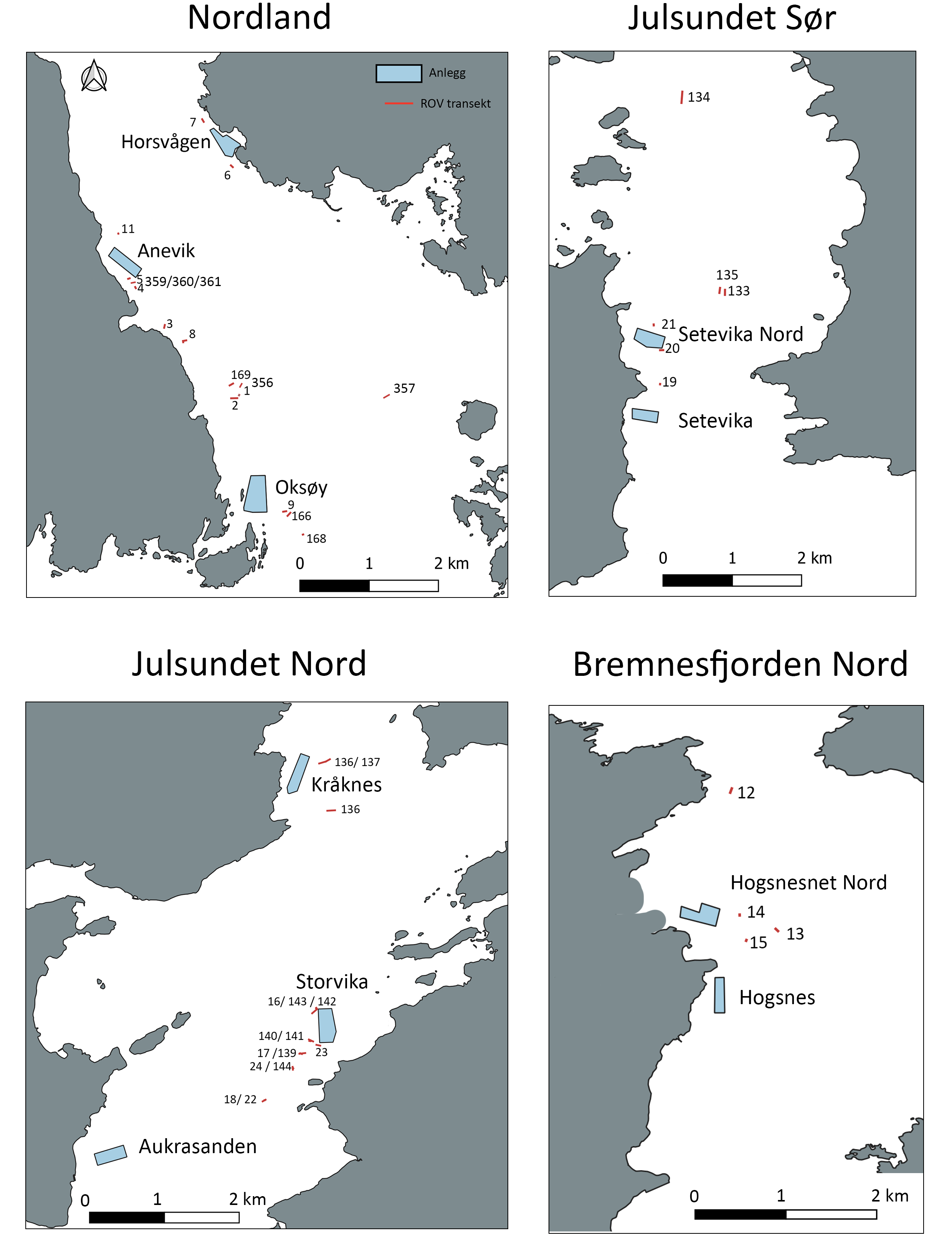
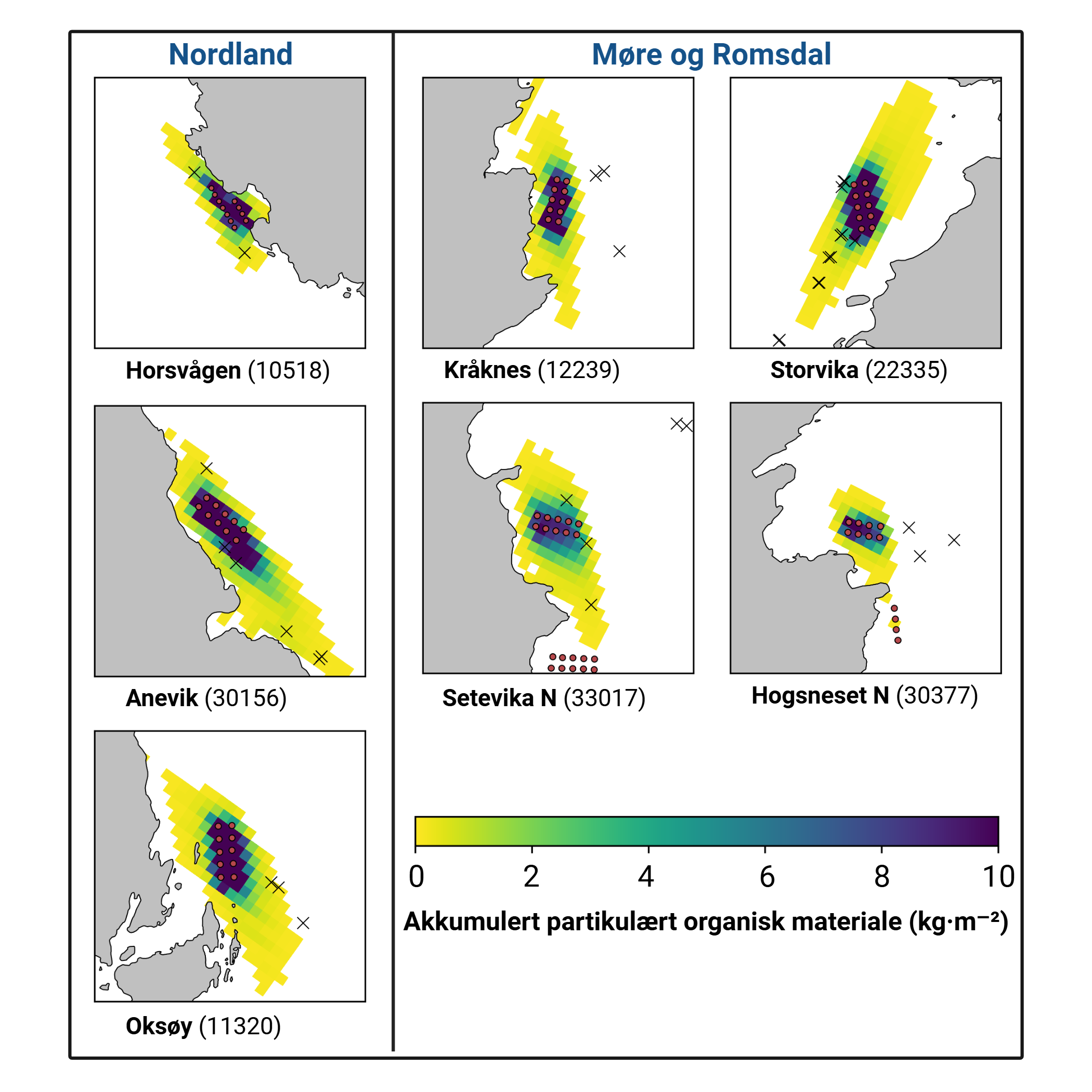
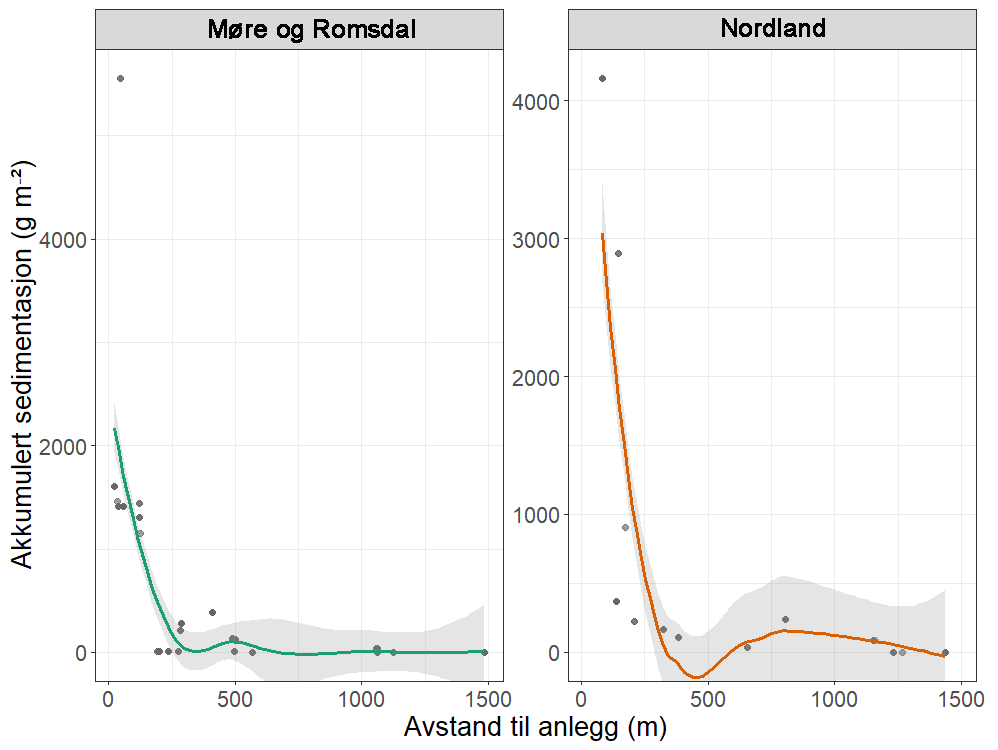
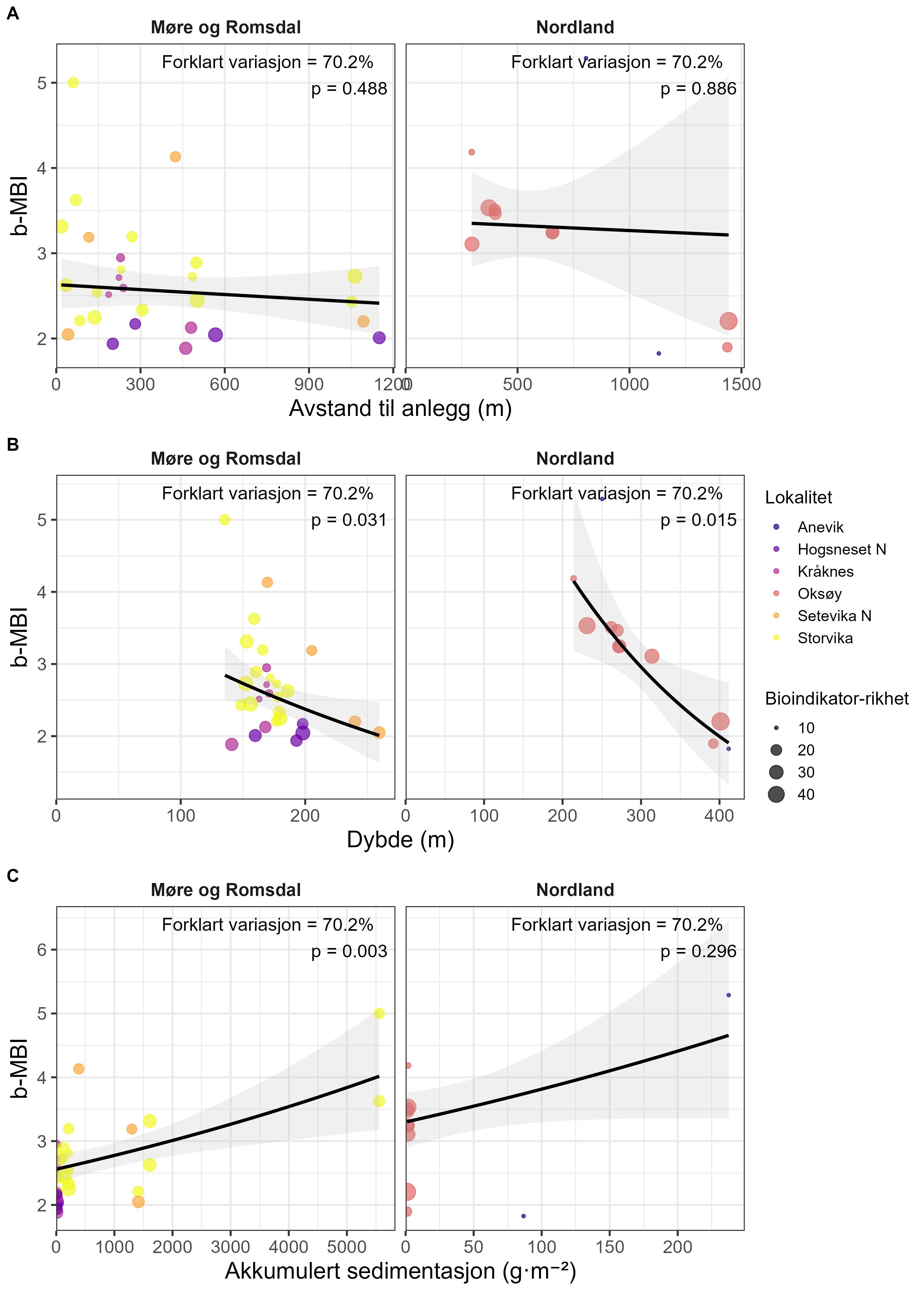
 Figur 5: Bilder av de sju målartene fra prøvetaking i VDWS-Transition-prosjektet.
Figur 5: Bilder av de sju målartene fra prøvetaking i VDWS-Transition-prosjektet.