The Porsangerfjord went from being a fish-rich fjord in the first half of the 20th century to a much more fish-poor fjord in the second half. The fish-poor condition has persisted to this day. The invading king crab has compensated for the value loss, but only to a small extent the cultural loss, caused by reduced fish stocks. Based on discussions between fjord users with local knowledge and researchers, as well as scientific literature, this report presents some hypotheses, reasoning and speculation about the reasons for the change. From February to the end of April, there was previously a winter fishery for cod in connection with at least five spawning grounds on the west side of the fjord. When spawning was completed at the end of April, most of the fishery moved over to the east side of the fjord. The fishermen's opinion was that the cod, both local and migrating, largely followed the capelin to the innermost parts of the fjord. Large saithe joined the feeding bonanza a little later but avoided the eastern innermost parts of the fjord. Today there is a small fishery (approx. 150 tons per year) in connection with one of the spawning fjords on the west side but no following fishery on the east side or in the innermost parts, neither for cod nor saithe. In some places in the inner areas on the west side, you could fish for cod that had foraged on capelin all year round, which indicates that the fjord has a local capelin stock. Recent studies suggest that this capelin stock is small today, perhaps because they are outcompeted by the massive occurrence of shrimp in the inner, cold and capelin-friendly areas. The local herring stock with elements of large herring disappeared in the early 1990s. There is still a lot of herring in the fjord today, but it originates mainly from the strain Norwegian Spring Spawning Herring (NVG) and is largely spawned on the coast of Nordland and Troms. Some of this herring drifts into the fjord in the summer in its first year of life, and swims out again as small herring one to two years later. There is a probable overlap between the juvenile herring's wintering areas and the fjord cod's freshly spawned eggs. Since few predate on the herring, it can lead to large consumption of cod eggs. Cod fry that survive the summer are exposed to predation from sculpins and larger cod and saithe when they in the autumn settle in the shallow areas. High fishing pressure on potential sea urchin predators was probably the reason why the sea urchin population exploded and grazed down the kelp forest in the fjord from the 1970-ties. The seabirds probably have more difficulty finding food because fewer eggs are spawned on the bottom, and because large predator fish chase herring, sand eel and capelin to the surface to a lesser extent. Seals have always been present in the fjord and have been viewed by locals as a priced resource. After 1970, the number of seals in the inner fjord areas increased, but it is uncertain whether the presence of seals frightens cod from entering the inner parts of the fjord to utilize the food resources there. The level of ambition for the reconstruction of the cod stock must be that it is re-established in the abandoned spawning fjords, that both the quantity and the individual size increase, and that the stock utilizes the entire fjord as grazing area.
Essay om fiskerier i Porsangerfjorden før og nå
— et resultat av møter mellom fjordbrukere og forskere
Rapportserie:
Rapport fra havforskningen 2020-41
ISSN: 1893-4536
Publisert: 25.11.2020
Prosjektnr: 15474
Forskningsgruppe(r):
Økosystemprosesser
Tema:
Hav, kyst og fjord
Program:
Kystøkosystemer
Godkjent av:
Forskningsdirektør(er):
Geir Huse
Programleder(e):
Jan Atle Knutsen
English summary
Sammendrag
Porsangerfjorden gikk fra å være en fiskerik fjord i første halvdelen av 1900-tallet, til en mye mer fiskefattig fjord i den andre halvdelen. Den fiskefattige tilstanden har vedvart fram til i dag. Den invaderende kongekrabben har i stor grad kompensert det verdimessige, men bare i liten grad det kulturelle tapet. Basert på diskusjoner mellom fjordbrukere med lokalkunnskap og forskere, samt vitenskapelig litteratur, presenteres det i denne rapporten noen hypoteser og resonnementer rundt årsakene til endringen. Fra februar og ut april foregikk det tidligere vinterfiske etter torsk i tilknytning til minst fem gyteplasser på vestsiden av Porsangerfjorden. Da gytingen var ferdig i slutten av april flyttet mesteparten av fisket seg over til østsiden og innover i de indre delene av fjorden. Fiskernes oppfatning er at torsken, både lokal utgytt og innsigstorsk, i stor grad fulgte lodda innover i fjorden. Siden kom stabb- og storseien til og beitet på sil og sild i tillegg til lodde. Dette fiskeriet var best tidlig på sommeren, men varte til godt utpå høsten. Noen steder i de indre områdene på vestsiden kunne man fiske torsk som hadde spist lodde hele året, noe som tyder på at fjorden har en lokal loddestamme. I dag foregår det et lite fiskeri etter torsk (ca. 150 tonn per år) i forbindelse med én av gytefjordene på vestsiden, men der er intet påfølgende fiskeri på østsiden eller innover i fjorden, verken av torsk eller sei. Det pekes på flere mulige faktorer, som enten hver for seg eller i samvirke, kan være årsaken til disse endringene. Nyere undersøkelser tyder på at loddestammen i fjorden er liten i dag, kanskje fordi den blir utkonkurrert av den massive forekomsten av reker i de indre, kalde og loddevennlige områdene. Siste rest av den lokale sildestammen med innslag av mye stor sild forsvant på begynnelsen av 1990-tallet. Det er likevel mye sild i fjorden i dag, men den stammer i hovedsak fra stammen Norsk Vårgytende Sild (NVG), som i stor grad gyter på kysten av Nordland og Troms. En del av denne silda driver inn i fjorden om sommeren i sitt første leveår, og svømmer ut igjen som småsild ett til to år senere. Det er en sannsynlig overlapp mellom ungsildas overvintringsområder og fjordtorskens nygytte egg. Siden få beiter på silda kan den bli tallrik og føre til stor predasjon på torskeeggene. Torskeyngel som klarer seg gjennom sommeren er utsatt for predasjon fra ulker og større torsk og sei, når de om høsten slår seg ned på bunnen i grunne områder. Hardt fiske på mulige kråkebollepredatorer var sannsynligvis årsaken til at kråkebollebestanden eksploderte og beitet ned tareskogen i fjorden på 1970-tallet, noe som igjen kan ha redusert oppvekstområder for spesielt yngel og ungfisk. Sjøfuglene har sannsynligvis fått det vanskeligere fordi det gytes færre egg på bunnen, og fordi store predatorfisker i mindre grad jager sild, sil og lodde til overflata. Sel har alltid vært til stede i fjorden, og har blitt jaktet på av lokalbefolkningen som en ettertraktet ressurs. Etter 1970 økte antall sel i de indre fjordområdene, men det er usikkert om tilstedeværelse av sel skremmer torsk fra å gå inn i de indre delene av fjorden for å utnytte åte-ressursene der. Ambisjonsnivået for gjenoppbygging av torskestammen må være at den på ny etablerer seg i de forlatte gytefjordene, at mengden og individstørrelsen øker, og at bestanden tar i bruk hele fjorden som beiteområde.
1 - Innledning
Fiskebestander varierer i mengde fra år til år. Det vet både forskere, fiskere og andre som høster av havets ressurser. Ofte er svingningene naturlige, men noen ganger er de menneskeskapte. Overfiske kan som kjent føre til reduserte bestander, men lavere kvoter får vanligvis biomassen raskt opp igjen. Om bestandene tas for langt ned kan imidlertid økosystemet gå inn i en ny tilstand, som ikke enkelt lar seg reversere ved å redusere fiskepresset. Slike regimeskifter (Beisner et al., 2003) kan være stabile over tiår, fordi tilbakekoblingsmekanismer hindrer bestandene i å flippe tilbake til originaltilstanden. For eksempel; om torsk fiskes ned til veldig lave nivåer kan sildefisk og andre planktonspisende byttedyr øke i mengde, slik at de beiter på torskens egg og larver og hindrer, eller i alle fall bremser, tilbakekomsten av sin ivrigste predator (Daan et al., 1985; Koster og Möllmann, 2000; Segers et al., 2007; Fauchald, 2010). Økosystemet endrer seg i så tilfelle fra å være torskedominert til å bli sildedominert.
Fisket etter gytemoden sild som hadde overvintret i Norskehavet (NVG-sild) kollapset fra fangstnivåer på opp mot 2 millioner tonn i perioden fra 1950-1968 til noen få tonn i 1969 (Bogstad et al., 1995). En del av flåten gikk etter kollapsen inn i de nordnorske fjordene og fortsatte fisket, til også dette ble stoppet etter et par år. Lokale porsangerværinger forteller at fjorden var opplyst som en liten landsby når de tallrike snurperne gjorde sine kast i høstmørket. Mer effektive fiskefartøy reduserte fra 1950-tallet ifølge Norderhaug et al. (2020) kystnære bestander av arter som torsk, hyse og steinbit til så lave nivåer, at de sannsynligvis forårsaket eksplosjonen i kråkebollebestanden med påfølgende nedbeiting av tareskog fra 1970-tallet og utover. Også fiskere i Porsangerfjorden opplevde fra 1950-tallet og utover at fiskebestandene ble kraftig reduserte, og også de mente nedgangen skyldtes økt deltakelsen fra en stadig mer effektiv flåte (Persen og Andersen, 2011). Mens det i Porsangers bygdebok estimeres en fangstmengde på 550-680 tonn fisk årlig i perioden 1936-1939 (Petterson, 1994), har det vært anslått at fangstene har vært oppe i mer enn 1000 tonn årlig i de gode årene etter 2. verdenskrig. I dag foregår det et mindre fiskeri på rundt 150 tonn, hovedsakelig etter kysttorsk, i forbindelse med torskegytingen i Smørfjord.
Ble økologiske mekanismer som tidligere hadde resultert i mye stor torsk, hyse, sei og rødspette forrykket da sildesnurperne fisket på planktonspiserne, og torsken, hysa, seien og rødspetta ble fisket hardt på med snurrevad og andre redskaper fra en stadig mer effektiv flåte?
Planktonspisere som sild, lodde, sil og reker er ikke bare en viktig matressurs for torsk, sei og hyse. Også sjøfugl nyter godt av eggene som blir lagt på bunnen og eksemplarene som blir jagd til overflata av de store predatorfiskene. Det var ved å følge med på fuglelivet folk fikk med seg hvor og når fisken kom inn. Så vanlig var det at stor fisk fulgte med åta og fuglelivet at de gangene det ikke skjedde kalte man fenomenet tørre sildeknuter, eller goike salletbállot, på samisk.
Selv om det er lite torsk, flyndre og sei i forhold til tidligere, foregår det i dag et verdifullt fiskeri i fjorden etter den invaderende kongekrabben. Det er imidlertid sterke begrensninger i lokalbefolkningens muligheter til å høste av krabberessursen. I folkemøter og i andre fora gir mange uttrykk for at de først og fremst ønsker torsk, sei, flyndre og stor sild tilbake i de områdene der disse artene tidligere var tallrike. Modellberegninger tyder på at det er lite interessekonflikt i matfatet mellom torsk og kongekrabbe (Pedersen et al., 2018), noe som er en indikasjon på tilstedeværelsen av at den ene bestanden ikke nødvendigvis må gå på bekostning av den andre. Oppfatningen av hvor rikt det var, hvilke arter som dominerte når og hvor, og hva som har ført til nedgangen vil variere, alt etter hvem man spør. Denne rapporten er basert på møter mellom forskning og tradisjonskunnskap, og er et forsøk på å skape en felles framstilling av bestandene det tidligere ble høstet av i rikt monn over hele fjorden, og drivkreftene som kan ha ført til kollaps og i tillegg har hindret gjenoppbygging av bestandene. Det betyr imidlertid ikke at alle forskere eller alle de som bor langs og bruker fjorden, er enige i framstillingen. Men samarbeidet har utvilsom gitt noen nye perspektiver på når fisken og fiskeriene flyttet seg fra vest til østsiden av fjorden, den lokale loddestammens betydning som mat og magnet som trekker til seg fjordtorsken, småsildas mulighet til å beite på torskeegg, sjøfuglenes avhengighet av et velfungerende økosystem, kråkebollens oppblomstring og nedbeiting av tareskogen, og sistnevntes mulige bidrag til at den lokale sildestammen forsvant på begynnelsen av 1990-tallet.
Kunnskapsgrunnlaget for denne rapporten er basert på samtaler mellom førsteforfatteren og personer som var aktive på fjorden etter 2. verdenskrig, og som har ønsket å dele sin lokalkunnskap og sine innsikter. Vanligvis hadde vi sjøkartet mellom oss, slik at piler, streker og stikkord kunne knytte hendelser til spesifikke geografiske steder. Tilsvarende har førsteforfatteren fått hjelp av forskerkolleger med god innsikt i kystøkologi og loddebiologi til å tolke de store sammenhengende, og til å sammenstille Porsangerfjordens fiskeriutvikling til en mer sammenhengende historie enn hva som ellers ville vært mulig. Alle som har bidratt med nøkkelinnsikter er medforfattere på rapporten. Navn og adresse til medvirkende fjordbrukere er i tillegg vist i vedlegg.
2 - Fjordtorsken
Torsken er mer sofistikert enn man kanskje skulle tro. Under parringen kommuniserer den med sin mulige partner ved å tromme med en muskel mot svømmeblæra. Om kjemien stemmer svømmer paret buk mot buk mens det gytes en porsjon egg og melke, som i løpet av sekunder starter tusenvis av nye liv. Deretter starter jakten på en ny partner, og for en erfaren flergangsgyter gjentar prosessen seg omtrent 15-20 ganger over en periode på opptil 48 dager (Kjesbu, 1989; Kjesbu et al., 1996).
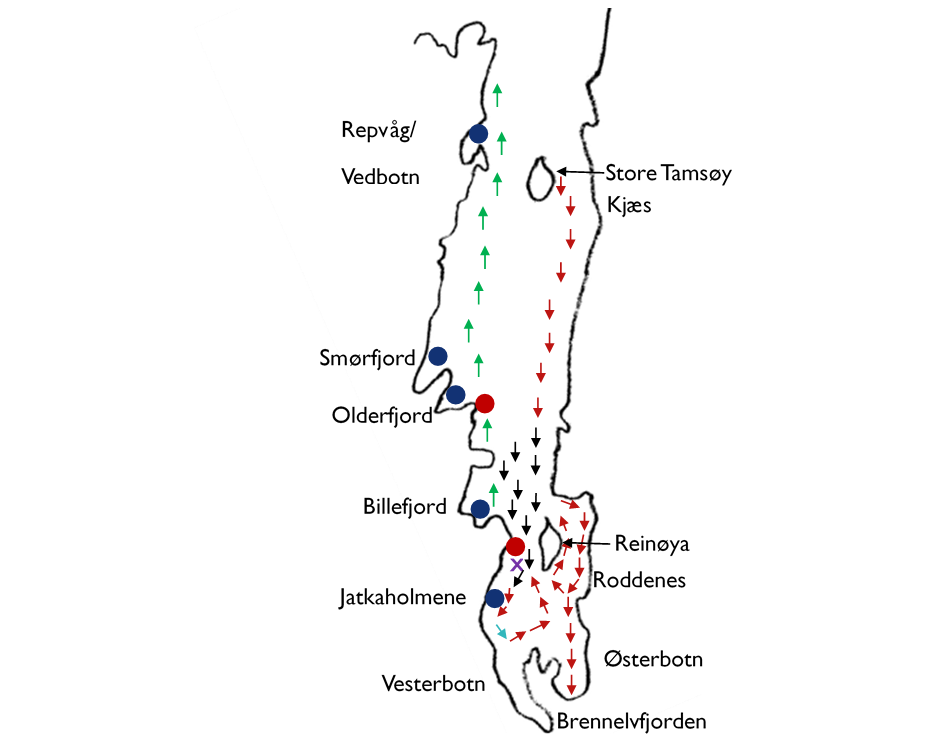 Figur 2. Noe av den informasjonen som ble nedtegnet på kartet under samtalene med fiskere og andre fjordbrukere. Steder det om våren ble fisket torsk som beitet på lodde er indikert med røde og svarte piler. Svarte piler viser områder det ble fisket mye storsei i tillegg til torsken. Seien ser ut til å ha styrt unna de kaldeste områdene øst og sør for Reinøya. Turkis pil markerer område hvor det ble fisket torsk på isen om vinteren, som hadde spist lodde. Grønne piler indikerer sannsynlig utvandringsrute for ungsild. Gyteplasser for lodde og torsk er markert med henholdsvis røde og blå sirkler. Lilla kryss viser en av plassene det ble fisket mye stor hyse fram til de store eksemplarene forsvant fra området rundt 1970.
Figur 2. Noe av den informasjonen som ble nedtegnet på kartet under samtalene med fiskere og andre fjordbrukere. Steder det om våren ble fisket torsk som beitet på lodde er indikert med røde og svarte piler. Svarte piler viser områder det ble fisket mye storsei i tillegg til torsken. Seien ser ut til å ha styrt unna de kaldeste områdene øst og sør for Reinøya. Turkis pil markerer område hvor det ble fisket torsk på isen om vinteren, som hadde spist lodde. Grønne piler indikerer sannsynlig utvandringsrute for ungsild. Gyteplasser for lodde og torsk er markert med henholdsvis røde og blå sirkler. Lilla kryss viser en av plassene det ble fisket mye stor hyse fram til de store eksemplarene forsvant fra området rundt 1970.
For den kjønnsmodne torsken handler ferden fra oppvekst- og beiteområder i og utenfor fjorden og tilbake til gytefjorden om å slutte livssyklusen. Det er en reise torsken starter som et befruktet egg og som varer til den selv gir opphav til nye befruktede egg. Vandringen og den påfølgende gytingen har derfor høyeste prioritet. I løpet av de omtrent to månedene gyteseansen varer er spising kraftig nedprioritert. Torsken kan spise litt innimellom om mat er lett tilgjengelig, men typisk etter gyting er den avmagret og sulten. I et normalår vil torsken i Porsangerfjorden ha gytt i Vedbotn, Smørfjorden, Olderfjorden, Billefjorden, og i sjøområdet mellom Gåradak og Jatkaholmene – for å ta de viktigste (Figur 2). De siste tiårene har det bare vært gyting av betydning i Smørfjorden. De viktigste fiskeriene finner sted når torsken samler seg i tette konsentrasjoner i gytefjordene, og i teorien kan man på disse stedene fiske ut tilnærmet hvert eneste kjønnsmodent fjordtorskindivid i bestanden. Alle gyteområdene ligger på vestsiden av fjorden der den fysiske utformingen er slik at det dannes strømvirvler, som bremser uttransporten av egg og larver med kyststrømmen. Etter hvert utvikler yngelen nok egenbevegelse til selv å bestemme hvor den vil være, og på ettersommeren og høsten forlater yngelen de frie vannmassene og slår seg ned på bunnen i de grunneste områdene. Slik starter en fjordtorsk livet.
Ifølge fiskerne starter torskens gyting i slutten av februar og varer til rundt 20. april. Torsken er etter endt gyting sulten og motivert til å fylle energilagre, og vokse fram mot neste gytesyklus. Hva kan få den til å svømme over til østsiden av fjorden, snarere enn å forbli der den er eller svømme utover i fjorden? Kanskje lodda.
3 - Lodda
Lodde er torskens foretrukne byttedyr (ICES, 2018) fordi den er energirik og lett å fange. Porsangerfjorden har sin egen bestand, men før den diskuteres må vi en tur innom Barentshavlodda. Barentshavbestanden av lodde regnes som den største loddebestanden i verden, med en stående biomasse som enkelte år når opp til 6–8 millioner tonn, og en årlig produksjon som overgår den stående biomassen. Bestanden er spredt over hele Barentshavet, både den arktiske og atlantiske delen. Om høsten finner man lodde i både arktisk og atlantisk vann ved temperaturer mellom -1 og 2 °C, og overvintring finner sted i nord, nært iskanten. I februar forlater den modnende del av bestanden overvintringsområdet og går inn til norskekysten, og i enkelte år også Kolakysten, for å gyte. Mesteparten av Barentshav-lodda dør etter gyting (Christiansen et al., 2008). Under våroppblomstringen litt senere på året kommer også den umodne delen av bestanden etter inn til kysten for å beite på de rike planktonforekomstene, og får da sannsynligvis selskap av de få individene som har overlevd gytingen. I kjølvannet av at iskanten trekker nordover utover våren og sommeren følger en planteplankton- og deretter dyreplanktonoppblomstring. Lodda trekker nordover mens den beiter på disse forekomstene, og den når sin nordligste utbredelse i september/oktober (Gjøsæter, 1998).
Eggene som blir gytt langs norskekysten klekker i løpet av noen uker og de nyklekte larvene driver mot nord mens de vokser og utvikler seg.
Porsangerfjorden framstår på mange måter som Barentshavet i miniatyr, bare speilvendt. I Barentshavet finner vi det varmeste vannet langs norskekysten i sør og en strømretning fra sør mot nord inn i de arktiske vannmassene. I Porsangerfjorden finner vi det varmeste vannet ute ved kysten i nord, og en strømretning mot sør langs vestsiden av fjorden og inn i arktiske vannmasser i bunnen av fjorden (Figur 3).
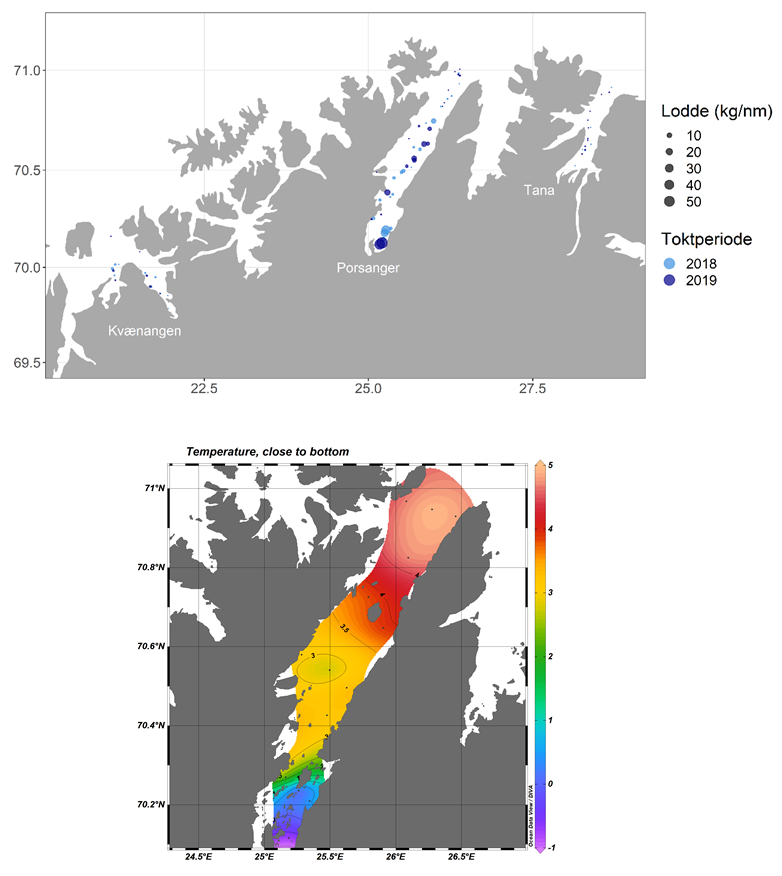
Om den speilvendte sammenligningen med Barentshavet holder inn i biologien, vil den gytemodne Porsangerlodda forlate overvintringsområdene i bunnen av fjorden og gyte langs land lenger ut i fjorden. Lokale fiskere har identifisert områdene utenfor Sandvik og Prestøya (se Figur 2) som gyteområder for lodde. Om våren kunne hunder vasse ut på fjære sjø rundt Trollholmene, og komme på land med levende lodde. Loddas gytefelter er plassert lenger ut enn torskens, og de nyklekte larvene vil derfor sannsynligvis ikke fanges i strømvirvler på samme måte, men heller transporteres innover mot de kalde indre delene av fjorden. Noen uker etter gytelodda går så unglodda ut og møter våren lenger ute i fjorden. Hedly Josefsen anslår området nord til Brenna/Kjæs som aktuell nordlig utbredelse for lodde, fordi de fisket mye torsk med lodde i magen fra dette området og innover om våren og tidlig på sommeren. Denne observasjonen stemmer godt overens med de tetteste registreringene av de sannsynligvis lokale loddeforekomstene i fjorden (Figur 4). Høsten 2019 ble det filmet betydelige mengder loddeyngel i det indre østlige bassenget i fjorden, noe som ytterligere underbygger og forsterker fiskernes og fjordbrukernes oppfatning av en stedegen loddebestand (https://www.dropbox.com/s/6xs18jco1tt987e/Lodde%20og%20reker%20i%20Porsangerfjorden.mp4?dl=0 Foto: Dag Olav Mollan)
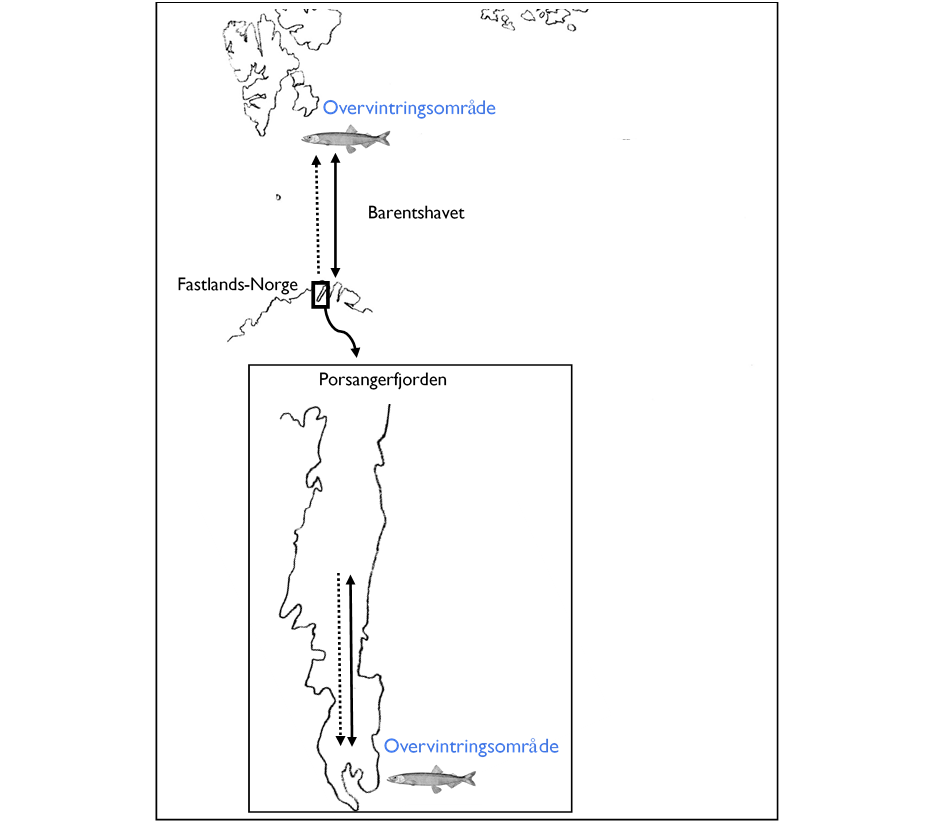 Figur 3. I Barentshavet finner vi de arktiske vannmassene i nord mens vi i Porsangerfjorden finner dem i sør og øst. Det vil gi en geografisk speilvendt dynamikk for årstidsvandringer fra overvintringsområdet til gyte- og beiteområder (heltrukken linje), samt for larvedrift (prikkete linje) mellom de to bestandene, men bevegelsene vil likevel samsvare når det gjelder vandring mellom kaldt og varmt vann i de samme periodene.
Figur 3. I Barentshavet finner vi de arktiske vannmassene i nord mens vi i Porsangerfjorden finner dem i sør og øst. Det vil gi en geografisk speilvendt dynamikk for årstidsvandringer fra overvintringsområdet til gyte- og beiteområder (heltrukken linje), samt for larvedrift (prikkete linje) mellom de to bestandene, men bevegelsene vil likevel samsvare når det gjelder vandring mellom kaldt og varmt vann i de samme periodene.
4 - Sulten torsk møter lodde
Fra slutten av april foregikk det i Porsangerfjorden årlig fram til midt på 1980-tallet et betydelig fiske etter torsk som beitet på lodde. Dette fisket foregikk i hovedsak på østsiden av fjorden inn til Reinøya, og på begge sidene innenfor denne øya (Figur 2). Lodda foretrekker kaldt vann og på den østlige siden er vannet kaldere enn på den vestlige siden lenger ute, mens det er kaldt i begge bassengene innenfor Reinøya (Figur 4). Torsken ble fisket på mens den fulgte etter lodda helt inn til bunnen av fjorden, og Thor Follesø kan for eksempel berette at han fikk smellfeit torsk helt innerst i Østerbotn i mai/juni, og at laksenota en gang gikk full av torsk også på den innerste plassen i Vesterbotn. Tore Eliassen, som fisket ut fra Gåradak, sier fisket på torsk som beitet på lodde startet i slutten av april, altså like etter at gytingen var ferdig. Fiskeriene strakte seg inn i juni, men mai var den beste måneden. Selv fisket han i de strømrike sundene på begge sidene av Reinøya, og mellom holmene innover på vestsiden. Og han beretter om et rikt fiske etter torsk som beitet på lodde. Det gjør også Terje Martinsen, som på 1960- og 70-tallet ofte fisket loddetorsk om våren på østsiden av Reinøya og i hele Roddenesbassenget. Han forteller at stortorsken gjerne jagde loddestimene helt grunt, og at de mer enn én gang holdt på å drive inn i fjæresteinene, fordi de var så opptatt med fisket at de glemte å følge med på landet. I tillegg til den typiske fjordtorsken kom det også inn betydelige mengder torsk som ble betegnet som innsigstorsk, for å beite i loddestimene. Jan Daleng er oppvokst i samme område som Martinsen, og også han fisket mye på 1950 – og 60-tallet. I tillegg til loddetorsken husker han at fisket på en mer lokal torskesort fortsatte utover sommeren. Denne torsken var mer rødbrune i fargen, men kunne som loddetorsken veie flere kilo. De var ofte bitevillige, og kunne gå på redskapen rett under båten. Torskefisket var også godt utover høsten når nettene begynte å bli mørke igjen. Dagny Larsen fra Stabbursnes og Igeldas på andre siden av fjorden fisket en gang båten så full at hun måtte få hjelp til å komme til land, og en sommerdag på slutten av 1970-tallet fisket sønnen hennes en torsk på 23,6 kilo i det samme området (Figur 9). Det var kanskje en av de siste store fjordtorskene.
Det er ikke urimelig å tenke seg at den loddebeitende torsken i øst i stor grad bestod i torsk som hadde gytt i de vestlige gytefjordene (se Figur 2) tidligere på vinteren, siden det ene fiskeriet sluttet omtrent samtidig som det andre startet. Men er det rimelig å anta at det var lodda som fristet torsken til å gå over til den kalde østlige siden, heller enn å beite utover i de varmere delene av fjorden, eller helt ute av fjordsystemet? Fra Tanafjorden forteller en fisker følgende: Midt på 70-tallet var det lodde inne på fjorden om høsten, i oktober-november. Det var smålodde, og det fulgte etter kolossalt med annen fisk og det ble gode fangster for dem som rodde. Det har jeg ikke opplevd verken før eller siden (Persen og Andersen, 2011).
Registreringer om høsten i 2018 og 2019 tyder på at Porsangerfjorden har en lokal loddebestand, og at bestanden er mye større enn forekomstene i Kvænangen og Tanafjorden (Figur 4). Man kan anta at lodde som registreres i fjordene i oktober stort sett vil være av lokalt opphav fordi Barentshavlodda på denne tiden har forlatt kysten og er på vei mot overvintringsområdet nord i Barentshavet.
Lodda er som sagt torskens foretrukne byttedyr fordi den er energirik og lett å fange. Det er i utgangspunktet en livshistoriestrategi som borger for selvutslettelse. Men lodda liker seg i kaldere vann enn torsken, og kan på denne måten unngå å bli spist. I de innerste delene av Porsangerfjorden, og nord i Barentshavet, er det områder med temperatur rundt 0 °C året rundt. Om torsken er sulten nok kan den følge etter inn i det kalde området av Barentshavet (Gjøsæter, pers.medd.), men fordøyelsen og omsetningen av byttet til vekst og nye gyteprodukter vil nødvendigvis gå tregere enn om torsken hadde spist et tilsvarende måltid i varmere vann. Analyser av øresteinene til torsk fanget i de kalde, indre, bassengene av Porsangerfjorden viste at de vokste seinere enn torsk fanget i de varmere delene lenger ute (Larsen, 2010). Fiskernes og fjordbrukernes observasjoner tyder på at storseien i stor grad unngår de indre, østlige, kalde og lodderike bassengene (Figur 2), noe som ytterligere underbygger en antakelse om at tilgang på vann rundt 0 °C deler av året eller i deler av livssyklusen kan være viktig for loddas overlevelse.
5 - Loddebestanden kollapser med ujevne mellomrom
Bestanden av lodde i Barentshavet er registrert helt opp i nesten 8 millioner tonn, men den har også opplevd noen spektakulære kollapser (Figur 5). Den store økologiske og økonomiske betydningen til lodda, har ført til en massiv forskningsinnsats for å forstå hva som bestemmer de store fluktuasjonene. En gjennomgang av forskningen på lodde foretatt av Gjøsæter et al. (2015)konkluderte at predasjon på de tidligste livsstadiene har mest å si for årsklassestyrken og senere rekruttering til fiskeriene. I alle fall når det gjelder å forklare de tre store kollapsene vist i Figur 5. I 2014 gikk bestanden inn i en fjerde kollaps, men denne gangen skyldes det mest sannsynlig stort beitetrykk fra torsk kombinert med svake årsklasser (Havforskningsrapporten 2017). Predasjon på de tidlige livsstadiene finner sted mens de nyklekte loddelarvene driver fra utklekningsområdene på kysten av Troms og Finnmark, og inn i de arktiske områdene nord i Barentshavet.
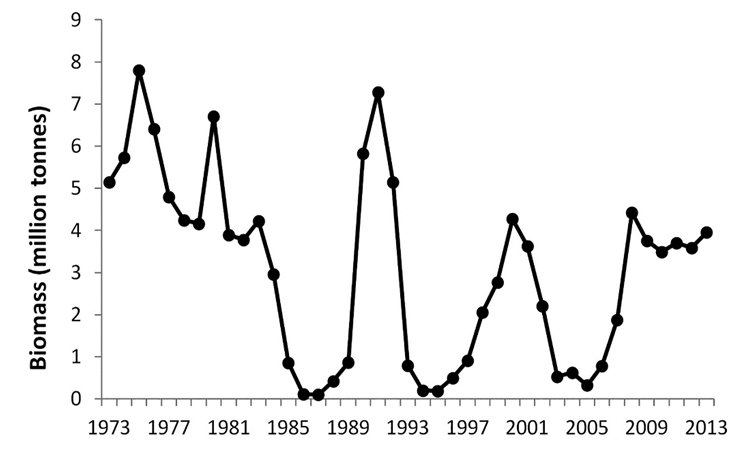
Den viktigste regulatoren av de tidligste livsstadiene til lodde og dermed årsklassestyrken antas å være ung NVG-sild. Det er stor årlig variasjon i hvor mange silderekrutter som driver inn i Barentshavet, og som tilbringer 3-4 år der før de slutter seg til den voksne delen av bestanden i Norskehavet. I 1983 kom den første store årsklassen med sild inn i Barentshavet siden 1959, og denne årsklassen forlot Barentshavet i 1986. Tilsvarende kom det inn sterke årsklasser i 1991 og 1992 og fra 2001 til 2004 (Gjøsæter et al., 2015). Alle disse hendelsene ble fulgt av kollapser i loddebestanden, og basert på undersøkelser av sildemager, som dokumenterer deres evne til å spise loddelarver, blir det konkludert at ungsild er den mest sannsynlige årsaken til loddekollapsene. Forfatterne understreker imidlertid at mye sild i Barentshavet ikke konsekvent medfører kollaps i loddebestanden, fordi det også må være en god overlapp både i tid og rom mellom de to artenes tilstedeværelse, en forutsetning som ikke alltid er til stede.
Det finnes betydelige mengder ung NVG-sild i Porsangerfjorden (Pedersen et al., 2018), som driver inn og svømmer ut igjen etter samme mønster som til Barentshavet, og det er derfor rimelig å anta at de følger de samme mengdefluktuasjonene, det vil si tre store pulser med innstrømmende sildeyngel i hhv. 1983, 1991–1992 og 2001–2004. Et relevant spørsmål er derfor om silda hadde en tilsvarende effekt på loddebestanden i Porsangerfjorden, som i Barentshavet, det vil si at tilstedeværelse i store mengder sild førte til kollaps i loddebestanden. De rike fiskeriene på torsk som beiter lodde nevnt ovenfor varte til rundt midten av 1980-tallet, mens torsken under det kortvarige oppsvinget på begynnelsen av 1990-tallet hovedsakelig spiste sild.
6 - Kanskje reiste ikke Porsangerfjordlodda seg igjen etter kollapsen i 1985/86?
Mens loddebestanden i Barentshavet tar seg opp igjen mellom kollapsene kan vi stille spørsmål ved om den har gjort det i Porsangerfjorden. Etter 1985/86 har det ikke vært noe fiske av betydning på torsk som følger etter lodde innover i fjorden på forsommeren. Det kan skyldes at det er lite torsk, og at den finner maten den trenger i de varmere deler av fjorden, eller at loddebestanden er kraftig redusert i forhold til tidligere.
Fisker Hedly Josefsen fra Billefjord kan ikke huske å ha fått loddetorsk etter kollapsen/selinvasjonen i 1986/1987. Han forteller at torsken under det kortvarige oppsvinget på 1990-tallet hovedsakelig spiste stor sild. Tore Eliassen forteller at fisket på denne torsken hovedsakelig foregikk lenger ut i fjorden og at torsken ikke fulgte det tradisjonelle loddesporet fra Reinøya og innover i fjorden. Om det er mindre lodde enn tidligere, hva kan grunnen til det i så fall være? Som det framgår av Figur 6 tyder registreringene på at rekebestanden i de kalde indre bassengene, spesielt det østlige, er mye større enn loddebestanden, noe som også ble vist av Pedersen et al. (2018).
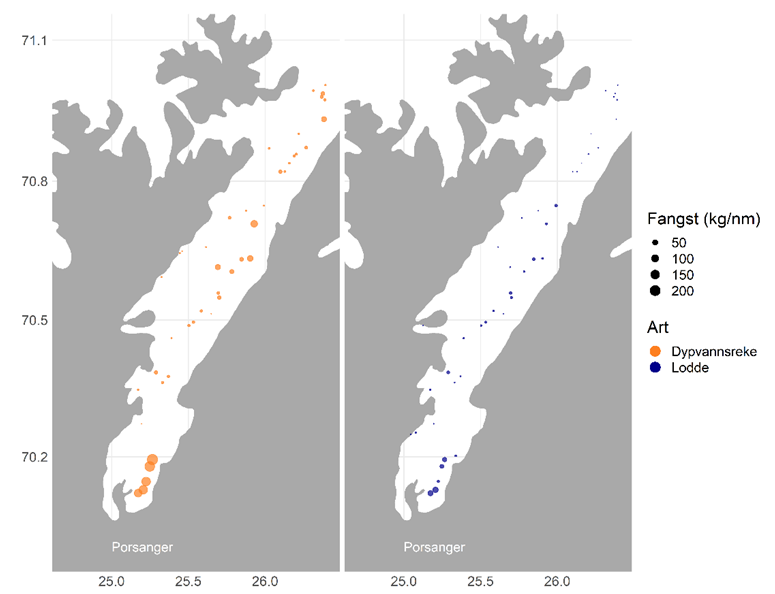
Rekene vandrer ikke på samme måten som lodda, og trekker derfor nok ikke med seg gytetorsk på vei ut av sidefjordene, på samme måte som lodda. Reker inneholder mye mindre energi per vektenhet enn lodde (Temming og Herrmann, 2003), og kan derfor være mindre regningssvarende for torsken å forfølge inn i kaldt vann.
Kan det tenkes at beiting fra ungsild fra den sterke 1983-årsklassen kollapset loddebestanden i Porsangerfjorden slik den gjorde det med Barentshavbestanden, og at reker utnyttet åpningen til å bygge seg opp til nivåer som hindrer gjenvekst i loddebestanden? De store rekeforekomstene kan i dag tenkes å utkonkurrere lodda i matfatet, ettersom reker og lodde i stor grad spiser samme type mat, som for eksempel krill, raudåte og amfipoder (Shumway, 1985; Gjøsæter, 1998). Det kan også tenkes at rekene beiter direkte på de nyklekte loddelarvene når de driver inn i de kalde, østlige bassengene, men hvorvidt reker kan spise nyklekte loddelarver eller fiskelarver generelt, er ukjent, og bør undersøkes.
Det er interessant at loddebestanden i de indre delene av Porsangerfjorden mer enn doblet seg fra 2018 til 2019 (Figur 7). Men estimatene kommer med stor varians og endringen er derfor ikke signifikant, slik at det ikke er mulig å fastslå hvorvidt økningen skyldes naturlig variasjon, lavere rekebestand som følge av teinefiske, den begrensete dekningen av toktet og observasjonsvarians i toktdata, eller en kombinasjon av de tre faktorene.
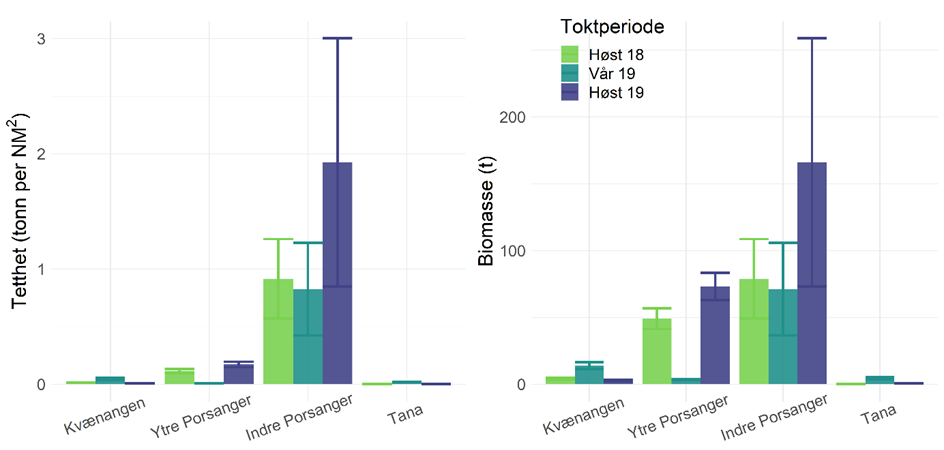
Foreløpig anses uttaket av reker i teinefisket i de indre områdene som beskjedent i forhold til totalbestanden, men om teinefiskeriene utvikler seg videre vil det utvilsom bli interessant å følge endringer i styrkeforholdet mellom sild, lodde og reker framover.
7 - Ungsild kan overbeite dyreplankton og torskeegg
Sild, lodde, sil, reker og andre dyreplanktonspisende arter er vanligvis byttedyr for torsken. Men noen ganger kan rollene snus. Da torsken forsvant fra de canadiske bankene, økte biomassen av de pelagiske byttedyrene lodde, tobis (Ammodytes sp.) og reker. Den økte filtrerings- og partikkelbeitingskapasiteten disse artene representerer når biomassen øker, bidro muligens til å beite ned egg og larver hos torsk, slik at sistnevnte ikke produserte nok rekrutter til å få gjenvekst i bestanden, tross fiskeforbud (Steneck, 2012; Collie et al., 2013). Fra Nordsjøen og Østersjøen er det vist at filterspisere som ungsild og brisling kan konsumere betydelige mengder egg og larver fra torsk og rødspette (Daan et al., 1985; Koster og Möllmann, 2000; Segers et al., 2007). Det er også foreslått at økte sildeforekomster, som følge av reduserte torskebestander i Nordsjøen, kan beite på torskeegg og -larver, og i tillegg torskelarvenes foretrukne byttedyr raudåte, i en slik grad at det hindrer gjenoppbygging av torskebestanden (Fauchald, 2010). Ironisk nok kan altså torskens foretrukne byttedyr holde sin verste fiende i sjakk ved å spise opp avkommet dens.
Om de store ungsildforekomstene i Porsangerfjorden skal kunne utgjøre en trussel mot torskeegg og larver, må det være overlapp i tid og rom. Fiskere i Porsangerfjorden anslår at overvintringen av sild i stor grad foregår fra Billefjorden og innover. Dette stemmer godt overens med småsildundersøkelsene til Hognestad i november 1960 (Figur 8), samt undersøkelsene til Pedersen et al. (2018) på 2000-tallet.
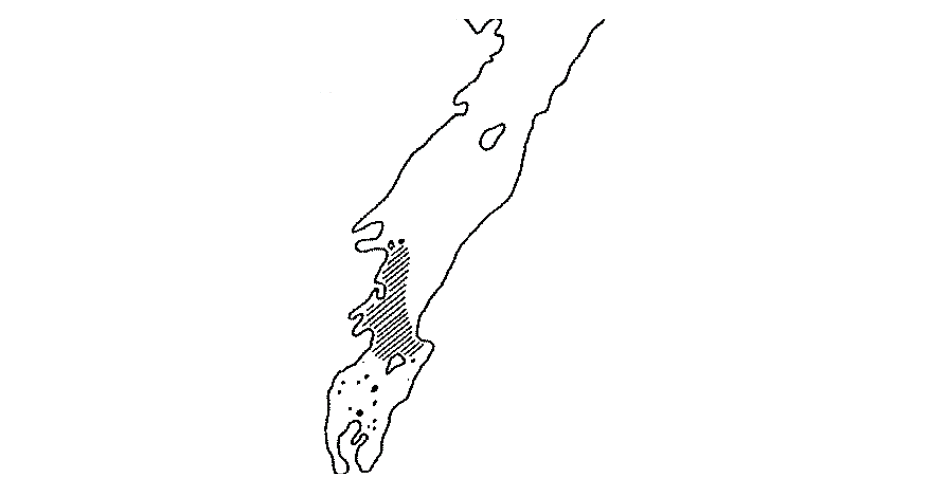
I nordvestenden av det skraverte området i Figur 8 finner vi Olderfjorden, som tidligere var en av de viktigste gytefjorden for torsk i Porsangerfjorden, og i denne fjorden fant Torstensen og Røttingen (2002) betydelige mengder ungsild (1+) på 8-10 cm i tillegg til at de også fant betydelige mengder nord for Smørfjorden, som ligger litt lenger mot nord, og er den andre store gytefjorden for torsk. Man kan følgelig mistenke at alle de viktige gytefeltene Smørfjorden, Olderfjorden, Billefjorden og feltet mellom Gåradak og Jatkaholmene blir påvirket av silda. Når 1-2 år gammel overvintrende ungsild beiter seg utover fjorden om våren for etter hvert å slutte seg til moderbestanden i Norskehavet, vil det sannsynligvis også være overlapp mellom sildebeitingen og torskeegg og -larver som gytes lenger ut i fjorden, men betydningen av denne beitingen vil være vanskelig å kvantifisere.
På begynnelsen av 1900-tallet ble det gjort følgende observasjon fra iskanten rundt holmene på innsiden av Reinøya, et område som tilsvarer nedre del det skraverte området i Figur 8: En kobbebåt var ute ved iskanten omkring den 25. mars. Ved iskanten var det en mengde alker og måser som tok sild og lodde. I båten hadde disse juksa-redskap fra høsten av. De prøvde juksa, og det beit godt av stortorsk. Etter hvert som nordausten brøt isen i stykker innover, fulgte fiskerne iskanten (Petterson, 1994). Dette er midt i gytetiden og midt i kjerneområdet for gytingen til den lokale torsken, og viser hvordan torsk tidligere beitet på den overvintrende ungsilda.
Men på 1970-tallet opplevde de lokale fiskerne en kraftig nedgang i fisket etter stabb- og storsei (Lund, 1988) og den begynnende svikten i torskefiskeriene ble også tydelig i denne perioden (Persen og Andersen, 2011). Disse opplysningene samsvarer også med de lokale fjordbrukernes fortelling om et betydelig fiske etter sei og torsk i det aktuelle overvintringsområdet for ungsild fram til 1980-tallet, slik det framgår av Figur 2.
Det er rimelig å anta at fraværet av stabb- og storsei og torsk i det aktuelle området fra 1980-tallet, kan ha økt sildas overlevelse, og derved dens kapasitet til å beite på torskeegg og larver. Hvor mye torskegg og -larver konsumerte for eksempel den sterke 1983-årsklassen (Gjøsæter et al., 2015) med sild, som sannsynligvis bare i liten grad ble beitet på av større fisk, fram til den forlot fjorden i 1985/86?
Det var mange svake årsklasser med kysttorsk på 1980-tallet, men den som kom i 1987 var sterk. Berg og Pedersen (2001) spekulerte i om den sterke 1987-årsklassen skyldtes at grønlandssel under invasjonen de to foregående årene hadde spist opp eller skremt bort aktuelle torskeyngelpredatorer (hovedsakelig større torsk). En annen bidragende faktor kan imidlertid være at den sterke sildeårsklassen fra 1983 i 1986 var tre år gammel, og da hadde forlatt fjordene og Barentshavet for etter hvert å slå seg sammen med den voksne delen av sildebestanden i Norskehavet. Utreisen til en sterk sildeårsklasse året i forveien kan ha redusert sildas predasjon på fjordtorskens egg og larver i 1987.
Store mengder ungsild som bare i liten grad blir beitet på av predatorer kan også ha andre konsekvenser i fjordøkosystemet, enn at de beiter ned torskeegg og -larver. Blant annet kan det ha ført til overbeiting av dyreplankton, og dermed store mengder uspist planteplankton, som kan ha sedimentert ut av vannsøylen og ført til en forråtnelsesprosess på bunnen. Laksefiskere som fisket i de indre vestlige områdene av fjorden merket store endringer på midten av 1980-tallet, med nedsliming av bruk. «Det var som å dra garnene opp av møkkakjelleren», kan Hans O. Hansen fra Gåradak fortelle. Tore Eliassen husker det samme fenomenet fra midten av 1990-tallet. Også andre fiskere kan fortelle om liknende fenomener: Nedslamming av bunnen på midten av 1980-tallet fikk også konsekvenser for områder der det tidligere hadde vært ruglbunn: Flere av kjente av flyndrefelt, hvor det før var områder hvor det kunne være rugl og man måtte være kjent for å kunne sette flyndregarn, der var kjente flyndrefelt hvor man kunne få masse flyndre – i dag eksisterer det ikke lenger. Det er bare slam på botn, og rugl finner du knapt nok noen steder (Persen og Andersen, 2011). Begge disse hendelsene samsvarer i tid med sterke årganger av ungsild i Barentshavet og påfølgende loddekollaps (Gjøsæter et al., 2015). Overbeite av dyreplankton og økning i mengden av planteplankton er selvsagt ikke et unikt Porsangerfjordfenomen. Da torskefiskeriene kollapset utenfor kysten av Canada på 1980-tallet, økte mengden av torskens dyreplanktonspisende byttedyr, med en påfølgende økning i planteplanktonindeksen (Frank et al., 2005; Shackell et al., 2010; Frank et al., 2011).
8 - Kråkeboller spiser egg som gytes på bunnen
Det var tidligere også mye stor sild i Porsangerfjorden, og sildefisket fra slutten av 1800-tallet foregikk i hovedsak fra september og fram mot jul. Var fjorden frosset lagde man råker i isen og satte sildegarnene i disse. I desember 1877 ble det fisket ca. 100 tønner i Brennelvfjorden, som ligger vest helt innerst i Porsangerfjorden. I Porsangerfjorden under ett ble det fisket hele 3000 tønner sild i 1880, fordelt på not og garnfiskere (Hanssen, 1986), noe som skulle tilsvare ca. 300 tonn matsild. Det ble også fisket noe sild om somrene, men da hovedsakelig småsild til bruk som agn. Det er kjent fra historien at sildeforekomstene langs kysten kan variere voldsomt, men lokalbefolkningen i Porsanger har fram til 1990-tallet kunnet sette sine sildegarn i høstmørket i trygg forvissning om fangst av matsild. Det er utvilsomt at sildesnurpingen på 1950- og 1960-tallet gjorde store innhogg i bestanden. Likevel overlevde sannsynligvis i alle fall deler av bestanden de voldsomme fiskeriene, siden torsken på begynnelsen av 1990-tallet ble observert å beite på stor sild. Fra da av og utover har imidlertid den store silda vært så å si helt fraværende i fjorden.
Kråkebollebestanden økte kraftig på 1970- og 1980-tallet, og de indre områdene ble spesielt hardt rammet. Man vet ikke med sikkerhet hva som skjedde. Kanskje førte det harde fisket på sildebestanden til økt overlevelse hos kråkebollelarver, fordi sild ellers kan filtrere disse ut av vannmassene, og derved til en eksplosjon i kråkebollebestanden (Hagen, 1983). Lokale fiskere observerte begynnende nedbeiting av tareskogen allerede rundt 1965 og flere fiskere synes det er høres fornuftig ut å koble kråkebolleoppblomstringen til det harde fisket på sild de var vitne til: Nå er det mange som mener, og det virker fornuftig, at på 50- og 60-tallet så hadde vi store ansamlinger av sildesnurpere her, i mange av fjordene her. Og vi hadde masse sild inni her, overalt. Bortover her så kunne det faktisk se ut som en stor by når de svære sildesnurperne kom og satte her. Så sier forskerne, og det tror jeg på, det virker logisk – og det er nok mest sannsynlig at da startet hele elendigheten. Da tok man opp all silda. Som egentlig var plassert der for å meske seg på rogn av kråkeboller. For kråkebollerogna, den er sånn frittsvevende i vannmassene med det samme, og da føres den bortover og legger seg på bunnen noen plasser og så gror dem til å bli til kråkeboller. Og det er jo klart at silda ble jo tatt, så var det jo ingen som meska seg med kråkebollerogn (Persen og Andersen, 2011). Hardt kystnært fiske fra 1950-tallet og utover, med påfølgende reduksjon i bestandene av mulige kråkebollepredatorer som torsk, hyse og steinbit er andre sannsynlige årsaker til oppblomstring av kråkeboller og kollaps i tareskogen fra begynnelsen av 1970-tallet (Norderhaug et al., 2020). Tore Eliassen sier den store hysa forsvant fra de indre områdene rundt 1970 (se Figur 2). Steinbit var tidligere vanlig i fjorden, i alle fall i enkelte områder. Jan Daleng husker for eksempel at de som gutter fisket gråsteinbit for moro skyld i de strøm- og kamskjellrike sundene litt nord for Reinøya. «Det var bare å slippe pilken ned, så bar det på en steinbit», forteller Daleng. Enkelte linefiskere betegnet østsiden av fjorden og utover, fra omtrent på høyde med Store Tamsøy, som steinbitland. Det er imidlertid varierende oppfatning av om gråsteinbiten noen gang har vært spesielt tallrik over hele fjorden, men fra midten av 1980-tallet er det enighet om at det generelt har vært svært lite å se (Persen og Andersen, 2011).
Uansett årsak: Strandlinjen og de grunne flatene innover i Porsangerfjorden var tidligere tett tilvokst med tare, men en kråkebollebestand som på 2000-tallet hadde vokst til mer enn 2,6 milliarder individer (Gundersen et al., 2010) forvandlet frodige undersjøiske skoger til et goldt månelandskap mellom 1970 og 1990.
Om fjordsilda gyter direkte på tareblader, kan bortfall av denne ressursen ha bidratt til lavere gytesuksess. Kråkeboller kan imidlertid også beite direkte på sildeegg og loddeegg på bunnen. Det er vist at kråkeboller kan spise opptil 6,5 % av kroppsvekten sin i form av sildeegg (Haegele, 1993). Kan kråkebollens beiting på egg ha vært så omfattende og vedvarende at det har hatt bestandsregulerende effekt på sild og kanskje også lodde? Eller kan forekomstene av kråkeboller på sand og steinbunn ha vært så tette at silda har skydd enkelte gyteområder? En dykker som svømte på grunnene mellom holmene på 1980-tallet ble slått over hvordan tidligere tett tilvokste områder var forvandlet til en steinørken, befolket av tette forekomster med kråkeboller. Laksefiskerne fikk i en trettiårsperiode (ca. 1980-2010) store mengder kråkeboller i redskapen. Tareskog er også et viktig oppvekstområde for fiskeyngel (Norderhaug og Christie, 2009). Sannsynligvis på grunn av kongekrabbens beiting på kråkeboller, er tarevegetasjonen i dag på vei tilbake over store deler av fjorden.
9 - Stor torsk og sei kan være nøkkelen
Lodde er som sagt torskens foretrukne byttedyr, men det finnes sannsynligvis mer ungsild enn lodde i Porsangerfjorden. Sild er også et utmerket byttedyr for torsken, men den er sannsynligvis vanskeligere å fange. Under kartleggingstoktet med reketrål i fjorden i 2018 og 2019, ble det for eksempel fanget slank torsk uten mageinnhold bare noen få hundre meter fra der det ble tatt betydelige mengder ungsild. Men med størrelsen på torsken øker både farten og ferdighetene. Porsangerfjorden var også viden kjent for sitt fiske etter stor og fin sei om somrene. De første eksemplarene kunne man få på torskegarnene allerede i slutten av april, men fra et stykke ut i juni tok fisket seg opp og levra blei feit nok til å kunne brukes til seimølja. Det var ofte sånn at vøer med storsei jagde sil opp i fjæresteinene, men den tok også for seg av silda. Sammen med seien var det også torsk og et yrende fugleliv i overflata. Storseien er en rask og effektiv jeger og kanskje skapte den så mye kaos i sildestimene at det ble enklere for torsken å fange sild? En analyse av fiskefeltene utenfor Nova Scotia viste at biomassen av predatorfisk var omtrent den samme, men at individstørrelsen ble redusert med omtrent 60 % som følge av hardt fiskepress på de store individene. Dette førte igjen til en eksplosjon i byttedyrforekomstene, som de småfalne predatorfiskene ikke klarte å fange. Forfatterne konkluderte at med den nåværende småfalne kroppsstørrelsen vil predatorene bare utnytte en brøkdel av den tilgjengelige byttedyrmengden (Shackell et al., 2010). At stortorsken tidligere tok for seg av silda som overvintret i Porsangerfjorden kommer godt fram i ovenfor siterte (Kap. 7) beretning fra begynnelsen av 1900-tallet: Ved iskanten var det en mengde alker og måser som tok sild og lodde. I båten hadde disse juksa-redskap fra høsten av. De prøvde juksa, og det beit godt av stortorsk. Etter hvert som nordausten brøt isen i stykker innover, fulgte fiskerne iskanten. Nedenfor skal vi se at den beskrevne dynamikken mellom åte og torsk også kan ha betydning for sjøfuglbestandene i fjorden.

Tore Eliassen, Hans O. Hansen og flere andre forteller at det beit på mye stor fin torsk når de fisket på storseien som boltret seg i åte. At torsken fulgte seien inn i stimene av åte var så regelbundet at enkelte, ifølge Hans O. Hansen, mente torsken ikke fant veien på egen hånd, men måtte finne fram ved å følge etter seien.
10 - Lodde og en lokal sildebestand kan være viktig også for sjøfuglene
Også fugler tar godt for seg av lodde. Lodde er viktig for alker og lundefugler, som har spesialisert seg på stimende pelagisk fisk. Tilgang på den energirike lodda kan være spesielt viktig tidlig i hekkesesongen (Barrett et al., 2006). Lodda legger eggene sine på bunnen, og både praktærfugl og vanlig ærfugl er dokumentert å beite på loddeegg ned til 50 m dyp (Gjøsæter, 1998). Det kan gi et viktig energibidrag til disse fuglene og føre til flere fugleegg av høyere kvalitet når tilgangen er god, og ditto til færre egg av dårligere kvalitet når lodda uteblir. Det ble registrert sultedød hos fugler i kjølvannet av loddekollapsen på 1980-tallet. Selv om bestanden av ærfugl på nasjonalt nivå ser ut til å ha vært relativt stabil og noen steder økende til rundt 2000-tallet (Barrett et al., 2006), var det lokalt flere steder dokumentert kraftig tilbakegang fra tidlig på 1980-tallet (Lorentsen, 2007).
Porsangerfjorden utpeker seg som det kanskje viktigste hekkeområdet for ærfugl i Finnmark, og bestanden ble ansett som god i alle fall fram til 1990 (Strann, 1992). Mange som er bosatt i de indre delene av Porsangerfjorden mener imidlertid bestanden er blitt mye mindre etter 1990-tallet og Norsk rødliste for arter 2015 karakteriserer nå ærfuglbestanden nasjonalt som nær truet (Henriksen og Hilmo, 2015). Og mye tyder på at ærfuglbestanden er enda kraftigere redusert om man sammenligner med enda tidligere tider (Figur 10).

På Store Tamsøy ble det for omtrent hundre år siden samlet opp imot 250 kg dun i løpet av sesongen, og om man legger 15 gram dun per rede til grunn, tilsvarer det i overkant av 30 000 voksne fugler. Rundt 1990 ble det talt færre enn 1500 voksne ærfugler på øya (Strann, 1992), eller bare 5 % av estimatet basert på dunsanking.
Høye ærfuglbestander i et dunvær skyldes til dels aktiv forvaltning i form av tilrettelegging for redebygging og uttak av predatorer. Men på tiden Store Tamsøy hadde sin høye produksjon av dun ble det fisket opp mot flere hundre tonn stor sild i året i fjorden (Hanssen, 1986), mens den store silda og eggene som gytes på bunnen i dag er forsvunnet.
I tillegg til lodde vil altså lokal sild gyte i fjorden, og bidra med egg på bunnen og yngel i vannmassene. Yngelen fra begge artene kan beites på av blant annet terner når den driver fra gyteplassene mot de indre delene av fjorden. Om sei og torsk kommer inn i de indre områdene og jakter på lodde, sild og tobis, vil måkefugler fange yngel i overflaten, og dermed ha alternative byttedyr til ærfuglkyllinger, og kanskje føre til høyere overlevelse hos sistnevnte. Hvordan store predatorfisker jaget sild og lodde til overflaten slik at sjøfugler kunne forsyne seg, ble godt illustrert i følgende observasjon i Porsangerfjorden fra begynnelsen av 1900-tallet: En kobbebåt var ute ved iskanten omkring den 25.mars. Ved iskanten var det en mengde alker og måser som tok sild og lodde. I båten hadde disse juksa-redskap fra høsten av. De prøvde juksa, og det beit godt av stortorsk.(Petterson, 1994).
Tidligere daglig leder ved Mearrasiida, Thomas Hansen, sier at de eldre med lokalkunnskap, rundt mange av holmene på sensommeren ser betydelig færre overlevende kyllinger enn det de gjorde tidligere. I disse områdene er det ikke lenger stor fisk som kan jage på åta. Og om ærfugltallene sank drastisk fra dunvær-tida og fram til 1990, har bestanden falt ytterligere fram til i dag: Fugleforsker Sveinn Are Hanssen fra Nina anslår at det var maksimalt 400 ærfugler (200 par) som hekket på øya i 2019 (Hanssen, pers.medd.).
Rundt 2010 gikk kongekrabben også inn i de indre kalde områdene av Porsangerfjorden (Oug et al., 2013), og med det var den invaderende arten for alvor etablert i hele fjorden. Siden både kongekrabber og ærfugl beiter på blant annet blåskjell og kråkeboller (Sivertsen, 2006; Pedersen et al., 2018), kan invasjonen ha redusert matfatet til ærfugl ytterligere.
Oppsummert kan sild og lodde være gunstig for ærfugler via to ruter; 1. foreldrefuglene spiser egg på bunnen om våren, og 2. stormåser og andre ærfuglpredatorer vil ha tilgang på alternative byttedyr til ærfuglunger. Sistnevnte forutsetter at fisk og andre predatorer jager lodde og sild til overflata, slik at de blir tilgjengelig for fuglene. Det er i tillegg en mulig konkurranse i matfatet mellom kongekrabber og ærfugler i hele fjordsystemet.
I dag er dypvannsreke den dominerende arten i de indre bassengene, og den vil ikke være tilgjengelig som mat til sjøfugler på samme måte som lodda. Vi kan ikke med sikkerhet slå fast at dypvannsreke har fortrengt lodde i de indre kalde bassengene, men de formidable forekomstene har ført til at det har utviklet seg et teinefiske etter reker der. I perioden 2017 - 2019 ble det fanget opp mot 10 tonn reker årlig i de indre delene av Porsangerfjorden. Teinefangst-næringen er fortsatt i en tidlig fase og det er derfor knyttet usikkerhet til utviklingen framover. Som sagt finnes det heller ikke stor, og derfor sannsynligvis ikke lokalt gytende sild i fjorden lenger.
11 - Sel, ulker og mangel på gråsteinbit som mulige årsaker til lite fisk i fjorden
Tidligere var selkjøtt et kjærkomment tilskudd på menyen til porsangerværingene, ikke minst om våren når lagrene av annen mat begynte å bli skrinne. Om man fikk øye på en sel som tok seg en hvil på en nesten nedflødd stein, «sprang man komaggan av seg» for å få has på den. Men selen ble raskt sky, og etter noen dagers jakt kunne bare de beste og mest erfarne jegerne regne med å komme hjem med fangst. Hartvig Johansen fra Jonsnes på vestsiden av fjorden er en erfaren seljeger, men hans far var regnet som en mester i faget. Med sin Krag Jørgensen rifle felte Johan Hartvig dyr fra avstander mange bare kunne drømme om (Figur 11). Hartvig forteller at interessen for selkjøtt dabbet av utover 1960- og 70-tallet, og med det ble det lite jakt også. Terje Martinsen fra østsiden sier at de så mye sel på iskanten også på 1960- og 70-tallet, og at kjøpmann Vik hadde utviklet en spesiell jaktteknikk for å komme på skuddhold før selen kastet seg i vannet. Teknikken bestod i å montere en hvit skjerm frampå en spark, og bevege seg nedhuket bak sparken til han kom på hold. Jan Daleng forteller at Nils Eriksen, som jaktet i samme område som Vik, benyttet seg av en annen teknikk; han lå på magen på en kjelke med et kobbeskinn over seg, og dro seg fram til skuddhold av dyrene som lå ved siden av pustehullene. Også om sommeren ble det observert sel i området, men Daleng sier kobbene ble jaktet på og var mye mer spredt og lettskremt enn i dag. På en sandbanke utenfor Børselv-munningen kan man i dag ofte telle mellom 20 og 50 dyr. Daleng forteller at han aldri observerte denne type ansamlinger i sin ungdom. Den stedegne selbestanden med fødeplasser for unger på holmer innerst i fjorden, økte etter hvert til mellom 200 og 400 dyr (Ramasco, 2015), men ligger i dag trolig nært den laveste delen av dette intervallet. Dette betyr at det tidligere har vært sel i de indre delene av fjorden, om enn i mindre mengder enn i dag, samtidig med rike forekomster av torsk og sei. Faktisk ble det fisket loddebeitende torsk rundt Reinøya i alle fall til rundt midten av 1980-tallet, det vil si en periode det også må ha vært en betydelig selbestand i området. Det er ikke utenkelig at selen først og fremst jaktet på lodda, og at torsken vet den har lite å frykte så lenge det er tette loddestimer i området.
Fjordtorsken har også forlatt de innerste gytefjordene på vestsiden, og det er lite sannsynlig av tilstedeværelse av sel er en viktig årsak til dette. Vinteren 2018/19 var det for eksempel torskegyting i Olderfjorden etter tiår med fravær. Det spekuleres lokalt i om torsken kan ha blitt presset dit fra en gytefjord lenger nord (Vedbotn) på grunn av etablering av et lakseanlegg der. Lakseanlegget ble lagt brakt i 2019/20, og da var det heller ikke gyting i Olderfjorden lenger.
Slik lakseyngel preges på og finner tilbake til elven de klekker i, selv etter flere års matsøk i åpne havområder, slik preges og vandrer også torsk og mange andre arter mellom fødested og beiteområder (Sinclair og Iles, 1988). Kanskje er det slik at vi må få i gang igjen gyting av torsk som naturlig søker seg til Olderfjorden, Billefjorden, og ved Jatkaholmene, fordi det er disse fiskene som i stor grad trekker over til østsiden og inn i de indre delene av fjorden etter gyting? Et mulig tiltak kan i så fall være å stimulere gyting så vel som tilbakeholdelse av yngel i disse områdene. Og kan rekefiskeriene som er under utvikling bidra til mer lodde?
En viktig grunn til at sel ligger langt framme i bevisstheten når folk diskuterer fjordøkologi og fiskekollaps, er invasjonene av grønlandssel langs kysten og i fjordene fra midten av 1980-tallet. Det er den kystnære selarten steinkobbe som er viktigst i fjordene, men lenger nord og øst i Barentshavet finnes det en mye større bestand av grønlandssel, som livnærer seg der og utnytter iskanten til ungeproduksjon. Rundt 1985 kollapset imidlertid matfatet i Barentshavet, og horder av sultne seler bega seg på matjakt inn til norskekysten og inn i de nordnorske fjordene. Invasjonshæren bestod av anslagsvis flere hundre tusen individer (Nilssen et al., 1998) og lokalbefolkningen i Porsanger observerte flak på det de anslår må ha vært flere tusen seler på matjakt innover fjorden.
Etter grønlandsselinvasjonen var det svart hav i Porsangerfjorden. Men så skjedde det noe merkelig; på begynnelsen av 1990-tallet kom fjordtorskbestanden tilbake med stor kraft, og fisket ble usedvanlig godt igjen, ikke bare i Porsangerfjorden, men i hele Nord-Norge (Figur 11).
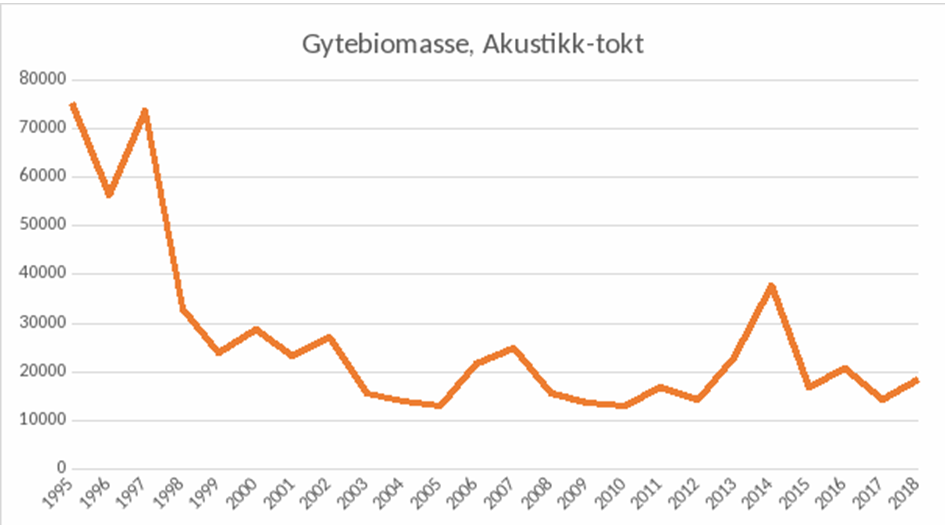
Gleden vedvarte imidlertid ikke. Etter et noen få år kollapset fisket på ny, og denne gangen uten å reise seg igjen i de påfølgende tiårene. Gytebestanden av kysttorsk nord for 62. breddegrad beregnes i dag til rundt 20 000 tonn (Aglen et al., 2020).
Det berettes imidlertid om såkalte kobbeår i fjorden også i 1902-04 og 1916-18 (Petterson, 1994), og selv om fisket var dårlig og fiskerne måtte leite på de store dypene etter fisk mens invasjonen pågikk, berettes det ikke om noen langvarig depresjon i etterkant av invasjonene. Kanskje var det et hardere presset økosystem som ble invadert på 1980-tallet, enn 60-80 år tidligere?
Som nevnt ovenfor kan paradoksalt nok grønlandsselinvasjonen være en viktig årsak til det midlertidige oppsvinget i torskefiskeriene, fordi torsken rakk å gyte før selene kom inn og skremte ut viktige yngelpredatorer, som stor sei og torsk (Berg og Pedersen, 2001). Sannsynligvis spiste de også en god del ulker. Hypotesen om at selen kan ha skremt ut yngelpredatorer stemmer også godt overens med observasjonene til fisker Bernt Wilhelmsen, som forteller at mens de under invasjonen ikke fikk fisk i fjorden, var fangstene av torsk i utkanten av fjorden gode (Lund, 1988).
Predasjon på yngel kan derfor være en viktig årsak til variasjon i mengden stor torsk det kan fiskes på noen år senere. Feltundersøkelser og laboratoriestudier i Porsangerfjorden har vist at ulker er den viktigste predatoren på nylig bunnslått yngel av torsk og sei, og at ulker og ungtorsk på 300-500 g kan jobbe i tospann og øke hverandres predasjonssuksess betraktelig (Pedersen et al., 2020; Strand et al., 2020). De samme undersøkelsene viste at gråsteinbit når den var til stede jagde bort ulkene og bidro til at flere torske- og seiyngler overlevde. Gråsteinbit var tidligere mer tallrik enn i dag, og en eventuell positiv effekt på yngeloverlevelse trenger ikke nødvendigvis utelukkende finne sted ved at steinbiten jager ulke i de grunne områdene der yngelen bunnslår om høsten. Det er nemlig ikke slik at alle ulkene ligger på grunna gjennom hele året, og venter på at yngel av torsk og sei skal slå seg ned der på ettersommeren. Rekefiskere og Havforskningsinstituttets forskere har i Porsangerfjorden opplevd å få betydelige mengder vanlig ulke i teinene, når disse i de indre områdene settes på dyp ned mot 100 meter om somrene. Det er også i disse områdene Jan Daleng kan fortelle det tidligere var et godt fiske etter stor torsk om somrene. Om store torsker beiter på ulker som er på næringsvandring om vinter, vår og sommer, eller steinbit nekter dem tilgang til større beitearealer i denne perioden, kan det påvirke ulkebestanden, og dermed hvor mange ulker som kan gå inn i de grunne områdene om høsten for å jakte på sei- og torskeyngel. Vi kan derfor spørre oss om fravær av de virkelig store torskene (Figur 9) så vel som gråsteinbit, kan bidra til en større bestand ulker på grunna om høsten, enn det som ellers ville vært tilfelle.

Kunnskapen om hvordan store mengder grønlandssel fikk fisk til å søke ut av fjorden og ned på store dyp under invasjonene i 1902-04, 1916-18, og midt på 1980-tallet, samt opplevelsen til fiskerne om at fisken forsvinner når selhodene dukker opp rundt båten, er overbevisende vitnesbyrd om at sel kan skremme fisk. Men den trenger ikke alltid gjøre det. Selv om prøvetakinger i de indre områdene i dag vil vise at det er veldig lite torsk der, samtidig som det er en betydelig selbestand i området, var det sel der i 1992 også, og da fant et forskningstokt om høsten faktisk mest torsk i disse områdene (Larsen, 2010). I tillegg ble det fra etter 2. verdenskrig og fram til midten av 1980-tallet fisket loddetorsk i det indre området, samtidig som det var en betydelig selbestand der. Det tyder i sum på at sel og torsk i alle fall under noen omstendigheter kan være i området samtidig, men at det vil være en grense for hvor mange sel som kan være i området før torsken skyr det. Uavhengig av hvorvidt selen skremmer større fisk fra å gå inn i et område, kan den imidlertid beite på ungfisk av torsk og sei. På den måten kan sel bidra til seinere gjenvekst i en torskebestand, som i utgangspunktet er lav, kanskje med omtrent samme påvirkningsgrad som skarvebestanden. Den muligheten diskuteres mer grundig i Porsangerfjord 2.0-rapporten (Strand, 2019).
Vi kan spekulere i følgende: Torsk har i stor grad forlatt de tre innerste gyteområdene på vestsiden, og det er fra disse områdene torsken har kortest vei til de kalde, tidligere lodderike områdene på østsiden av, og sør for Reinøya. Kanskje må vi hjelpe torsken å reetablere seg i de indre vestlige gyteområdene ved å 1. sette opp gytemærer og strukturer som kan bidra til økt yngeloverlevelse, som igjen kan bidra med mer yngel med lokal tilhørighet, 2. stimulere til loddedominans, slik teinefisket etter reker kanskje vil bidra til, slik at torsken i større grad blir fristet til å beite i de indre områdene, og 3. holde et visst jakttrykk på sel, slik at bestanden ikke blir for dominerende.
Om vi i tillegg legger til rette for at torskebestanden får anledning til å bygge seg opp til et høyere nivå og til en bestand som består av flere store individer enn i dag (se Figur 9), er det sannsynlig at den i større grad vil beite på og gjøre det vanskeligere for sterke sildeårsklasser å beite på torskeegg og -larver. De store torskene vil kanskje også beite på små ulker, når disse er på næringssøk på dypere vann om sommeren. Det vil i så fall sannsynligvis føre til færre ulker, som kan jakte på nylig bunnslått torskeyngel, i grunne områder om høsten.
12 - Referanser
Aglen A,Nedreaas K,Knutsen JA,Huse G. 2020. Kysttorsk nord for 62-grader nord — Vurdering av status og forslag til forvaltningstiltak og ny gjenoppbyggingsplan. In: Fisken og havet, https://www.hi.no/hi/nettrapporter/fisken-og-havet-2020-2 .
Barrett RT,Lorentsen S-H,Anker-Nilssen T. 2006. The status of breeding seabirds in mainland Norway. Atlantic Seabirds 8:97-126.
Beisner, BE., Haydon, DT, Cuddington, K. 2003. Alternative stable states in ecology. Frontiers in Ecology and the Environment 1: 376-382.
Berg E,Pedersen T. 2001. Variability in recruitment, growth and sexual maturity of coastal cod (Gadus morhua L.) in a fjord system in northern Norway. Fisheries research 52:179-189.
Bogstad, B. Røttingen, I. Sandberg, P. Solheim, BTV. Steinshamn, SI, Tjelmeland, S. 1995. Beskatningsstrategi for Norsk vårgytende sild. Fiskeridirektoratet; Rapporter og meldinger, 1995 NR 3.
Christiansen JS,Præbel K,Siikavuopio SI,Carscadden JE. 2008. Facultative semelparity in capelin Mallotus villosus (Osmeridae)-an experimental test of a life history phenomenon in a sub-arctic fish. Journal of Experimental Marine Biology and Ecology 360:47-55.
Collie J,Minto C,Worm B,R. B. 2013. PREDATION ON PRERECRUITS CAN DELAY REBUILDING OF DEPLETED COD STOCKS. BULLETIN OF MARINE SCIENCE 89(0):15.
Daan N,Rijnsdorp AD,Vanoverbeeke GR. 1985. PREDATION BY NORTH-SEA HERRING CLUPEA-HARENGUS ON EGGS OF PLAICE PLEURONECTES-PLATESSA AND COD GADUS-MORHUA. Transactions of the American Fisheries Society 114:499-506 DOI: 10.1577/1548-8659(1985)114<499:pbnshc>2.0.co;2.
Fauchald P. 2010. Predator-prey reversal: a possible mechanism for ecosystem hysteresis in the North Sea? Ecology 91:2191-2197.
Frank KT,Petrie B,Choi JS,Leggett WC. 2005. Trophic cascades in a formerly cod-dominated ecosystem. Science 308:1621-1623 DOI: 10.1126/science.1113075.
Frank KT,Petrie B,Fisher JA,Leggett WC. 2011. Transient dynamics of an altered large marine ecosystem. Nature 477:86.
Gjøsæter H. 1998. The population biology and exploitation of capelin (Mallotus villosus) in the Barents Sea. Sarsia 83:453-496.
Gjøsæter H. pers.medd. Forsker. Havforskningsinstituttet, harald@hino, Mob: 414 79 177.
Gjøsæter H,Hallfredsson EH,Mikkelsen N,Bogstad B,Pedersen T. 2015. Predation on early life stages is decisive for year-class strength in the Barents Sea capelin (Mallotus villosus) stock. ICES Journal of Marine Science 73:182-195.
Gundersen H,Christie H,Rinde E. 2010. Perspektivstudie av kråkeboller-fra problem til ressurs.-Analyse av ressursgrunnlaget for høsting av kråkeboller og veurdering av økologiske perspektiver knyttet til høstingen.
Haegele C. 1993. Epibenthic Invertebrate Predation of Pacific Herring, Clupea pallasi, Spawn in British Columbia. Canadian Field-Naturalist 1070:83-91.
Hagen NT. 1983. Destructive grazing of kelp beds by sea urchins in Vestfjorden, northern Norway. Sarsia 68:177-190.
Hanssen ER. 1986. Porsanger bygdebok, bind 1. Fra eldre steinalder til 1910.
Hanssen SA . pers.medd. Seniorforsker ved Nina; e.mail: sveinn.a.hanssen@nina.no; www.nina.no . In.
Henriksen S,Hilmo O. 2015. Norsk rødliste for arter 2015. Artsdatabanken, Norge 6.
Hognestad PT. 1961. Rapport over småsildundersøkelser i Nord-Norge med F/F "Asterias" høsten 1960. Forekomst av egg og yngel av fisk i vest- og nord-norske kyst- og bankfarvann våren 1960 ; Rapport over småsildundersøkelser i Nord-Norge med F/F "Asterias" høsten 1960:7-11.
ICES. 2018. Report of the Arctic Fisheries Working Group (AFWG), 18–24 April 2018, Ispra, Italy. In: ICES CM 2018/ACOM:06. p 857.
ICES. 2020. Cod (Gadus morhua) in subareas 1 and 2 (Norwegian coastal waters cod). In Report of the ICES Advisory Committee, 2020. ICES Advice 2020, cod.27.1-2coast. https://doi.org/10.17895/ices.advice.5893 .
Kjesbu O. 1989. The spawning activity of cod, Gadus morhua L. Journal of Fish Biology 34:195-206.
Kjesbu OS,Solemdal P,Bratland P,Fonn M. 1996. Variation in annual egg production in individual captive Atlantic cod (Gadus morhua). Canadian Journal of Fisheries and Aquatic Sciences 53:610-620.
Koster FW,Möllmann C. 2000. Trophodynamic control by clupeid predators on recruitment success in Baltic cod? Ices Journal of Marine Science 57:310-323 DOI: 10.1006/jmsc.1999.0528.
Larsen TH. 2010. Population dynamics of cod, Gadus morhua, in Porsangerfjord, Northern Norway-Analysis of back-calculated length at age from otoliths in 1992 and 2009. In: Universitetet i Tromsø.
Lorentsen SH. 2007. Det nasjonale overvåkingsprogrammet for sjøfugl. Resultater til og med hekkesesongen 2007. NINA rapport.
Lund S. 1988. Stille på hamna i Smørfjord. Artikkel i avisa Klassekampen: http://sveinlund.info/fisk/stille.htm . In.
Nilssen KT,Haug T,Øritsland T,Lindblom L,Kjellqwist SA. 1998. Invasions of harp seals Phoca groenlandica Erxleben to coastal waters of nor way in 1995: Ecological and demographic implications. Sarsia 83:337-345.
Norderhaug KM,Christie HC. 2009. Sea urchin grazing and kelp re-vegetation in the NE Atlantic. Marine Biology Research 5:515-528 DOI: 10.1080/17451000902932985.
Norderhaug KM,Nedreaas K,Huserbråten M,Moland E. 2020. Depletion of coastal predatory fish sub-stocks coincided with the largest sea urchin grazing event observed in the NE Atlantic. Ambio:1-11.
Oug E,Fuhrmann MM,Oug EPM. 2013. Bunndyrsamfunn i foreslått marint verneområde i indre Porsangerfjorden. Artssammensetning og biomasse før invasjon av kongekrabben 2011. In: Norsk institutt for vannforskning.
Pedersen T,Fuhrmann MM,Lindstrøm U,Nilssen EM,Ivarjord T,Ramasco V,Jørgensen LL,Sundet JH,Sivertsen K,Källgren E. 2018. Effects of the invasive red king crab on food web structure and ecosystem properties in an Atlantic fjord. Marine Ecology Progress Series 596:13-31.
Pedersen T,Strand HK,Christie H,Moy F. 2020. Predation mortality from ambush and cruising predators on newly-settled 0-group gadoids. Journal of Experimental Marine Biology and Ecology 529:151396.
Persen S,Andersen S. 2011. "Den gang var det jo rikelig med fisk" : lokal kunnskap fra Porsanger og andre fjorder : artikler med utgangspunkt i Fávllis-nettverket. Universitetet i Tromsø; Sjøsamisk kompetansesenter.
Petterson A. 1994. Småfolk og drivkrefter. Porsanger bygdebok, bind 2. Fra 1900 til 1960-årene.: Porsanger kommune.
Ramasco V. 2015. Spatial and temporal patterns of foraging of harbour seals (Phoca vitulina) in Porsangerfjord: from behavioural interpretation to resource selection.
Schuppe B. 2019. How does shrimp fishery affect the ecosystem in a previous closed fjord for bottom trawling? . In: Lehrstuhl für Physische Geographie und Geoökologie des Geographischen Instituts der Rheinisch-Westfälischen Technischen Hochschule Aachen RWTH Aachen University, Germany. p 141.
Segers F,Dickey-Collas M,Rijnsdorp AD. 2007. Prey selection by North Sea herring (Clupea harengus), with special reference to fish eggs. Ices Journal of Marine Science 64:60-68 DOI: 10.1093/icesjms/fsl002.
Shackell NL,Frank KT,Fisher JA,Petrie B,Leggett WC. 2010. Decline in top predator body size and changing climate alter trophic structure in an oceanic ecosystem. Proceedings of the Royal Society of London B: Biological Sciences 277:1353-1360.
Shumway SE. 1985. Synopsis of biological data on the pink shrimp, Pandalus borealis Krøyer, 1838.
Sinclair M,Iles TD. 1988. Population richness of marine fish species. Aquatic Living Resources 1:71-83 DOI: doi:10.1051/alr:1988009.
Sivertsen K. 2006. Overgrazing of kelp beds along the coast of Norway. Journal of Applied Phycology 18:599-610.
Steneck RS. 2012. Apex predators and trophic cascades in large marine ecosystems: Learning from serendipity. Proceedings of the National Academy of Sciences of the United States of America 109:7953-7954 DOI: 10.1073/pnas.1205591109.
Strand H,Pedersen T,Christie H,Moy F. 2020. Synergism between cruising cod and ambush sculpin predators on 0-group gadoids is modified by daylight cycle and presence of aggressive wolffish. Journal of Experimental Marine Biology and Ecology 526:151356.
Strand HK. 2019. Porsangerfjorden 2.0 — En mulighetsstudie. https://www.hi.no/hi/nettrapporter/porsangerfjorden-2.0-final.docx . In: Rapport fra Havforskningen: Havforskningsinstituttet.
Strann K. 1992. Sjøfuglundersøkelser Porsanger1988-90 Med hovedvekt på hekkende ærfugl. In: NINA Oppdragsmelding 104: 1-13.: Nina.
Temming A,Herrmann J-P. 2003. Gastric evacuation in cod: prey-specific evacuation rates for use in North Sea, Baltic Sea and Barents Sea multi-species models. Fisheries Research 63:21-41.
Torstensen E,Røttingen J. 2002. Silde- og brislingundersøkelser i fjordene 2002 : 2. november-16. desember 2002. In. Bergen: Havforskningsinstituttet.
13 - Vedlegg
| Navn | Når sitert | Adresse |
| Jan Daleng | 2020 | Mølleveien 15B, 3440 Røyken |
| Tore Eliassen | 2019 | Kolvik, 9710 Indre Billefjord |
| Thor Follesø | 2019 | 9700 Lakselv |
| Hans Oliver Hansen | 2019 | Gåradak, 9710 Indre Billefjord |
| Thomas Hansen | 2019 | Gåradak, 9710 Indre Billefjord |
| Hartvig Johansen | 2019 | Jonsnes, 9710 Indre Billefjord |
| Hedly Josefsen | 2019 | Indre Billefjord, 9710 Indre Billefjord |
| Dagny Larsen | 2019 | 9710 Indre Billefjord |
| Terje Martinsen | 2020 | 9716 Børselv |