—
Arbeidspakke 2: Egg- og yngelundersøkelser på gyte- og oppvekstområder på Smøla og i Aure i forbindelse med etablering av oppdrettsanlegg nær lokale gyteområder for kysttorsk.
Work package 2: Egg and juvenile surveys on the spawning and nursery areas at Smøla and in Aure in relation to establishment of a new salmon farm in the vicinity of local spawning grounds for coastal Atlantic cod.
The ICOD project was established to investigate potential interactions between salmon farms and nearby spawning areas for coastal Atlantic cod. This report presents results from ICOD which cover the egg and juvenile recruitment investigations carried out in connection with the launch of a new salmon farm at Nørholmen, Smøla. Also, results from a biochemical survey of fatty acids in cod caught on selected spawning areas at Smøla through the spawning season are presented. In addition to the spawning grounds at Smøla, the investigations include surveys from three reference spawning grounds in Aure municipality, that has no salmon farms in the close vicinity.
Variation in amounts of eggs and juveniles was significant, both between years and between the spawning and juvenile nursery areas. However, no signifficant correlation was found between the amounts of eggs collected during the spring surveys and the number of yearlings caught the following autumn. Simulation of egg drift using a hydrodynamic circulation model showed that the investigated spawning areas had relatively low degree of retention and therefore considerable advection of eggs. A prevalent current direction from south west inwards the Edøy fjord made advection of eggs from spawning areas upstream possible. No direct and clear interactions were observed between amounts of eggs and juveniles and periods when the Nørholmen farm was in full operation. However, these results are not immediately transferable to other locations, e.g. in more protected fjord systems, because the waters of the investigated areas in Smøla and Aure are highly dynamic and therefore will result in considerably advection of fish eggs and larvae. Furthermore, no genetic differentiation was observed, indicating lack of isolation in local populations due to the highly dynamic waters of this region.
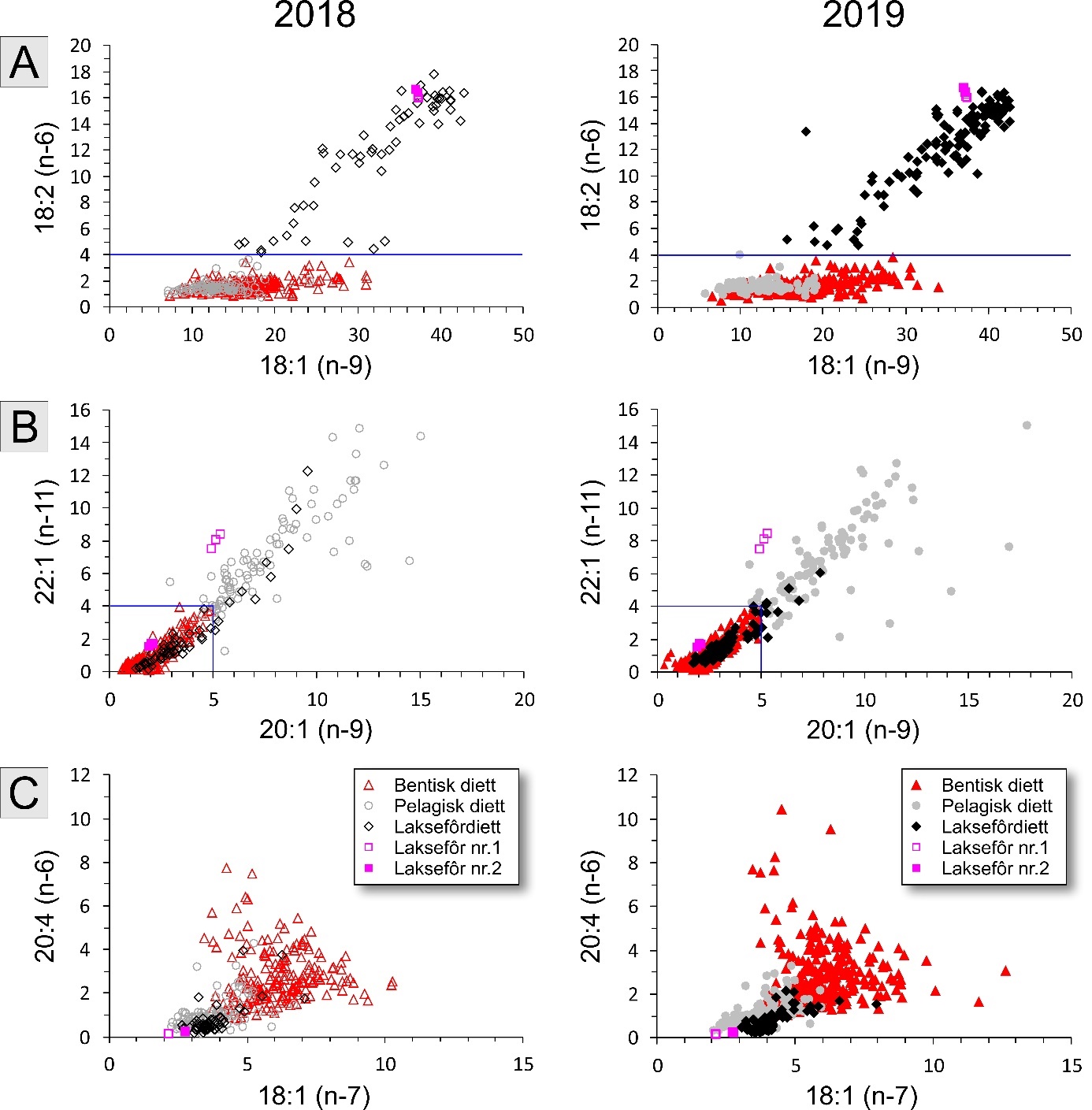
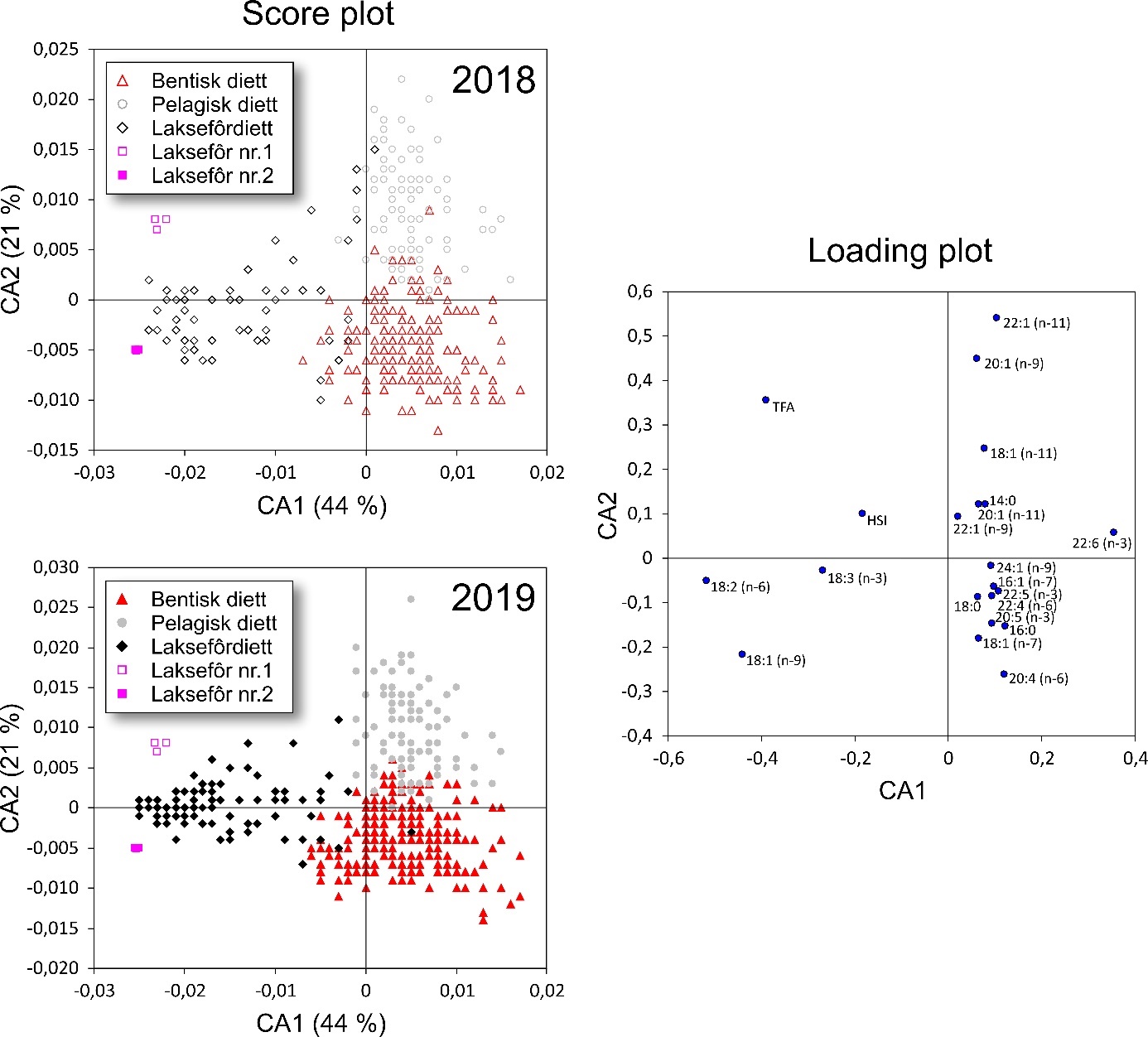
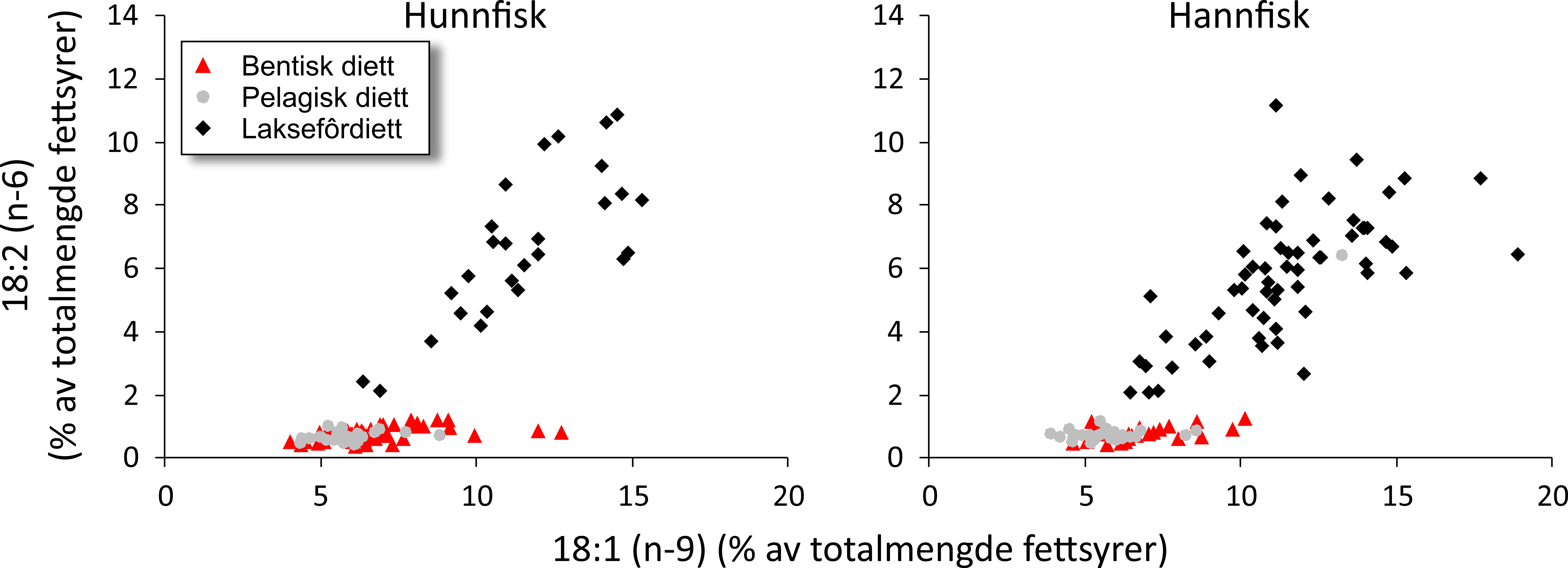
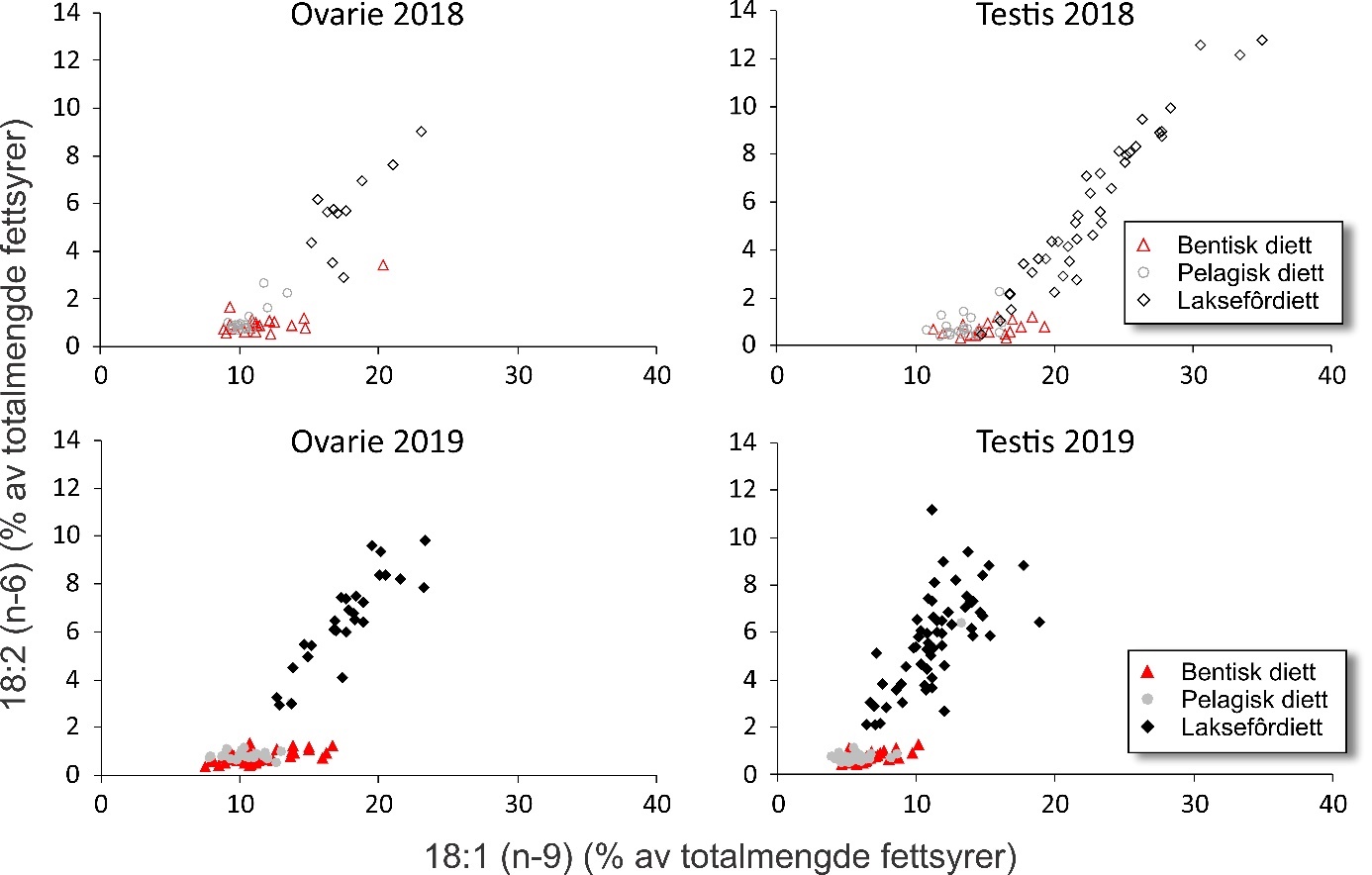
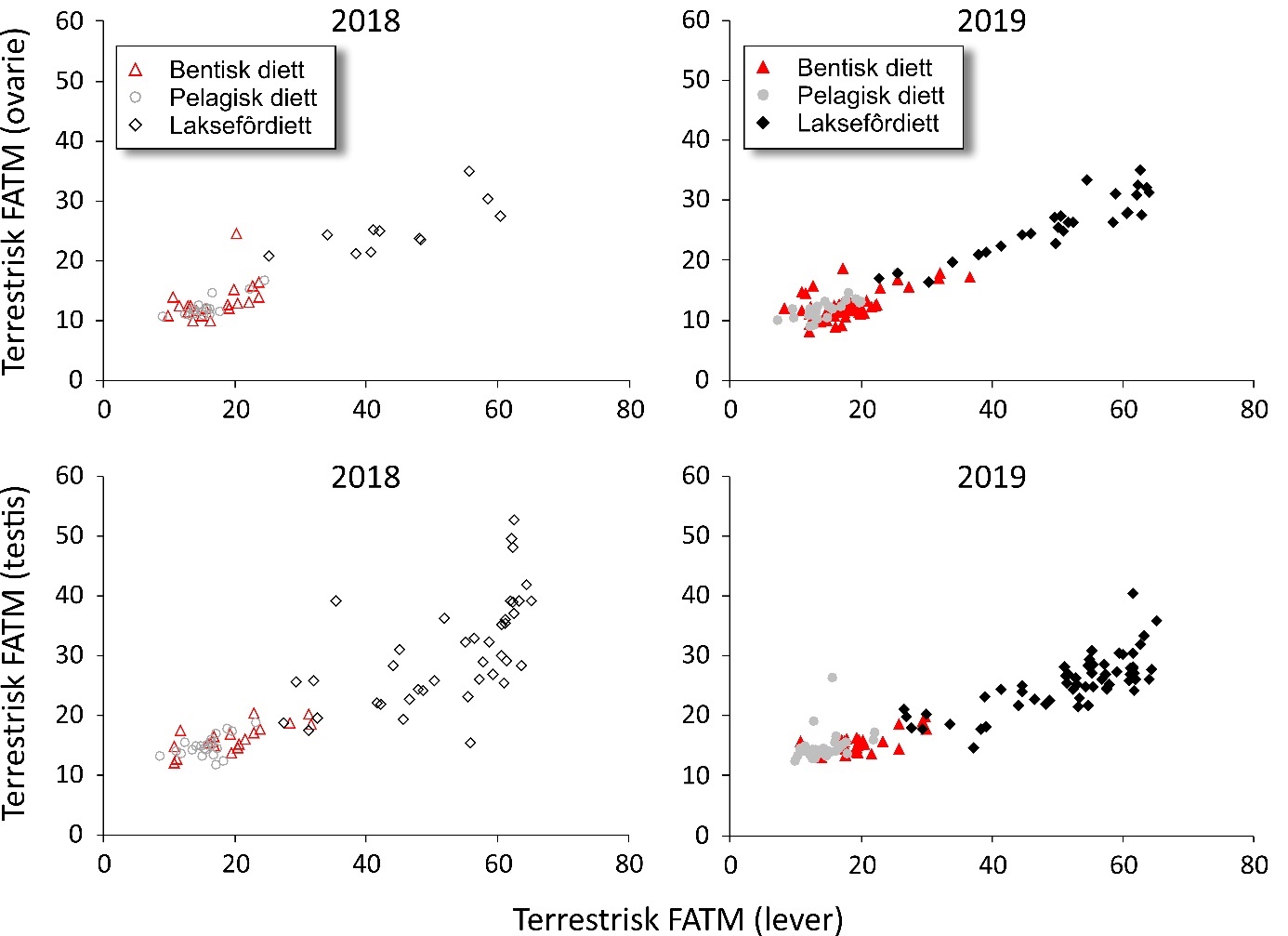
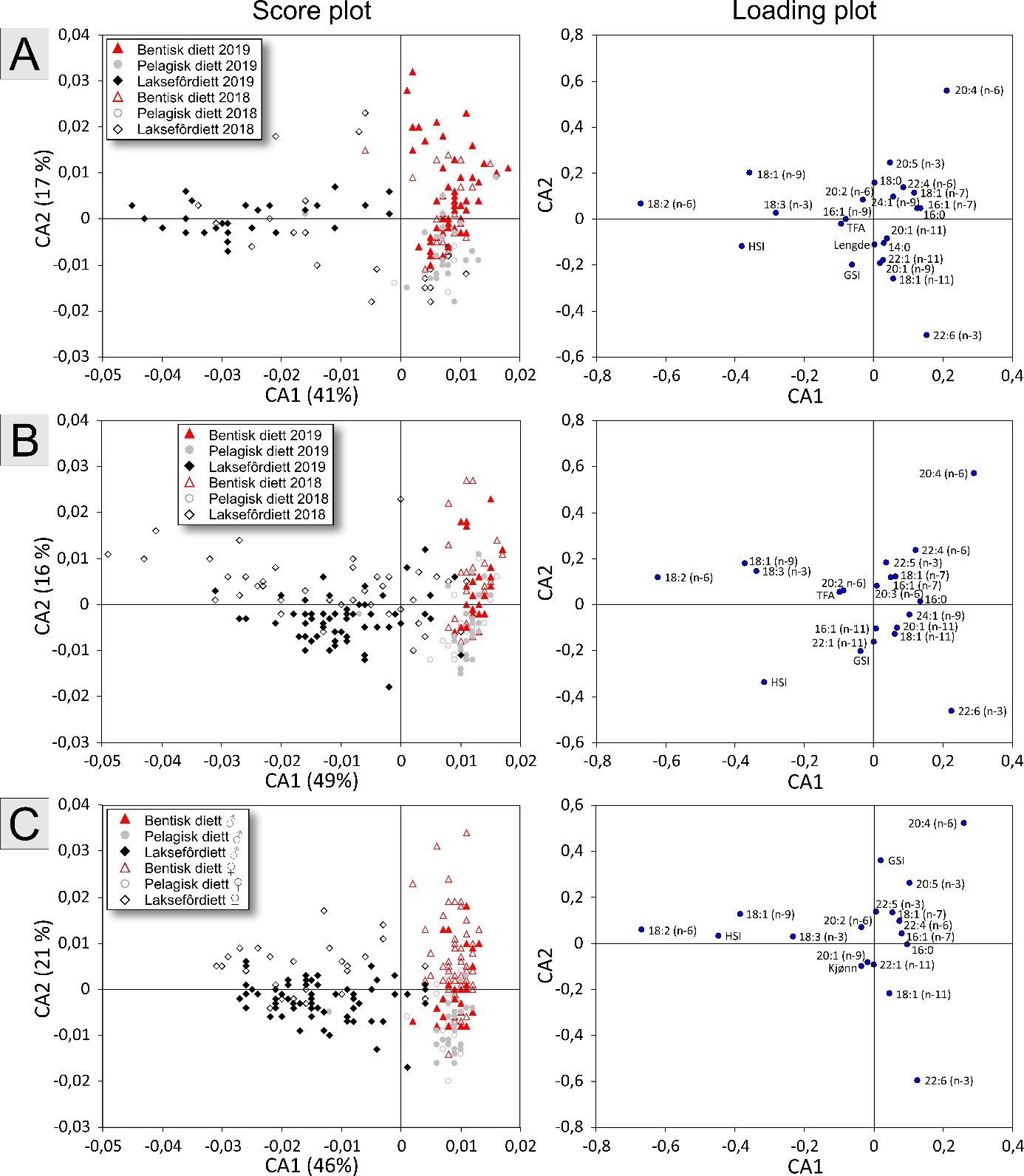
Fatty acid trophic markers from the biochemical investigations showed that a considerable fraction of the cod in the spawning areas had been eating spill-feed under salmon farms, and that the remaining cod could be divided into two components with a different diet background: feeding in the pelagic or benthic food web. It was not possible to determine if cod with pelagic or benthic diets represented migrating or more stationary and local stock components, or if the cod feeding under the fish farms had its origin in cod with one or both these diet types. The observed changes in fatty acid profiles of the liver and gonads due to a high fraction of terrestrial feed ingredients in salmon feed, indicate the possibility for biological effects regarding development and survival in early life stages of marine fish. However, it is unclear if such effects with impacts on recruitment exist, and further investigations on this topic are needed.
Sammendrag
ICOD-prosjektet ble etablert for å undersøke mulige interaksjoner mellom oppdrettsanlegg og nærliggende gyteområder for kysttorsk. Rapporten presenterer resultater fra ICOD som omfatter egg- og yngelundersøkelser foretatt i forbindelse med etablering av et oppdrettsanlegg for laks på lokaliteten Nørholmen på Smøla. Også resultater fra biokjemiske undersøkelser av fettsyrer i torsk fanget gjennom gytesesongen på utvalgte gyteområder ved Smøla presenteres. I tillegg til gyteområdene på Smøla omfattet egg- og yngelundersøkelsene tre referanseområder i Aure kommune uten umiddelbar nærhet til oppdrettsanlegg.
Det ble funnet betydelig variasjon i mengde egg og yngel, både mellom år og mellom de ulike gyte- og oppvekstområdene. Det ble ikke funnet noen signifikant sammenheng mellom eggmengde om våren og årsyngel samme høst. Simuleringer av eggdrift ved hjelp av en hydrodynamisk modell viste at de undersøkte gyteområdene hadde relativt liten grad av retensjon og derfor betydelig spredning av egg. Fremherskende strømretning inn Edøyfjorden fra sydvest gjorde det sannsynlig at egg og larver transporteres fra gyteområder oppstrøms inn i de undersøkte områdene. Det ble ikke funnet direkte og tydelige interaksjoner mellom egg- og yngelmengder og perioder med fisk i oppdrettsanlegget på Nørholmen. Resultatene er ikke umiddelbart overførbare til andre lokaliteter, som for eksempel til et mer beskyttet fjordsystem, fordi vannmassene i undersøkelsesområdene på Smøla og i Aure er svært dynamiske og derfor vil gi god spredning av fiskeegg og larver. Det ble heller ikke funnet genetisk differensiering, noe som kan ha sammenheng med manglende isolering i lokale bestander grunnet de dynamiske vannmassene.
De biokjemiske undersøkelsene viste ved hjelp av fettsyremarkører at en betydelig andel av torsken på gyteområdene hadde beitet på spillfôr under oppdrettsanlegg, og at den resterende torsken kunne deles inn i to komponenter med annen og ulik diettbakgrunn: pelagisk og bentisk diett. Det var ikke mulig å avklare om bentisk og pelagisk diett representerer henholdsvis en stasjonær og en vandrende type kysttorsk, eller om torsk som beiter under oppdrettsanlegg har sitt utspring i torsk med en eller begge disse diettypene. Endringen i fettsyreprofil av lever og gonader hos torsken som følge av at laksefôr i stor grad tilvirkes av fôrråstoffer fra landbruk, kan tenkes å gi biologiske effekter med hensyn til utvikling og overlevelse i tidlige livsstadier hos marin fisk, men om slike effekter og eventuell påvirkning på rekruttering eksisterer, er ukjent og trenger derfor ytterligere forskningsinnsats.
1 - Innledning
Gjentatte rapporter fra fiskere om at torsk forsvinner fra fangstområder på kysten og i fjordene når det etableres oppdrettsanlegg i nærheten, har så langt ikke vært mulig å verifisere vitenskapelig (Karlsen & van der Meeren 2013). Det er eksperimentelt vist at torsk kan reagere negativt på luktstoffer fra laks og torsk i oppdrett (Sæther m.fl. 2007), men i felt var det ikke mulig å verifisere at lukt var en faktor som kunne endre vandrings- og gyteatferd hos torsk i en fjord med lakseoppdrett (Bjørn m.fl. 2009). Hvorvidt ulike bestander eller populasjoner av torsk vil reagere forskjellig på oppdrettsanlegg, er også ukjent. Det er imidlertid registrert at torsk kan tiltrekkes og oppholde seg under oppdrettsanlegg (Dempster m.fl. 2009; 2010; 2011). Oppdrettsfôr for laks, som på grunn av fôrkilder fra landbruk har annerledes fettsyresammensetning enn marine fôrkilder, kan endre sammensetningen av fettstoffer i vill torsk som beiter under anleggene (Barrett m.fl. 2018). Dette åpner for mulig påvirkning av reproduksjonen siden langkjedede omega-3 fettsyrer fra fôr spiller en viktig rolle i modningen av fisken og muligens også eggenes næringsinnhold og kvalitet (Salze m.fl. 2005; Røjbek m.fl. 2014; Norberg m.fl. 2017).
Feltstudier av problemstillingen om vill torsk skremmes eller påvirkes negativt av oppdrettsanlegg krever omfattende undersøkelser da naturlig variasjon i mengde gytefisk på et gyteområde forventes å være betydelig i tid og rom. Ideelt bør denne variasjonen kartlegges over tid før et oppdrettsanlegg etableres, men dette kan være vanskelig da anlegg vanligvis legges ut relativt kort tid etter at en tillatelse er gitt. Imidlertid har det stor forvaltningsmessig betydning å få mer kunnskap om torskens atferd i forbindelse med etablering av akvakulturanlegg nær gyteområder.
I forbindelse med etablering av en ny lokalitet for oppdrett av laks, «Nørholmen» nordøst for Edøya i Smøla kommune, ble prosjektet «Interaksjoner mellom lakseoppdrett og gyteområder for torsk» (Interactions of salmon farming on Atlantic cod spawning grounds: ICOD-prosjektet) etablert våren 2015 som et samarbeid mellom Havforskningsinstituttet, Mowi AS og Smøla fiskerlag. I tillegg ble ICOD-prosjektet fra 2016 støttet av Fiskeri- og havbruksnæringens forskningsfond (FHF, prosjekt nr. 901230). ICOD bestod av fem arbeidspakker (WP) hvor denne rapporten fremlegger resultater hovedsakelig fra WP2 som ble støttet av FHF. Dette omfatter egg- og yngelundersøkelser, samt biokjemiske undersøkelser av torsk på utvalgte gyteområder. Også data fra WP4 som omfatter modellering av strøm, er inkludert i rapporten da dette er viktig for å vurdere drift av egg fra gyteområdene.
Undersøkelsene i ICOD ble foretatt både ved lokaliteter der oppdrettsanlegget skulle plasseres (Nørholmen på Smøla) og på to referanseområder i Aure kommune, Åkvika og Araneset, som ikke har oppdrettsanlegg i umiddelbar nærhet. Studien var tenkt gjennomført med et BACI-design (Before, After, Control, Impact) der nærliggende gytefelt og oppvekstområder overvåkes både før og etter etablering av oppdrettsanlegget. Imidlertid ble anlegget på Nørholmen etablert allerede våren 2016, og det ble satt fisk i anlegget i mai 2016. Derfor ble ytterligere ett referanseområde i Aure uten nærhet til oppdrettsanlegg, Dromnessundet, inkludert fra 2016 for å styrke datagrunnlaget. Gytelokalitetene på Smøla omfattet Lauvøysvaet og Glasøysvaet nord for Edøya, hvor førstnevnte er i nærheten av det nye oppdrettsanlegget ved Nørholmen. Glasøysvaet har i tillegg et annet oppdrettsanlegg i umiddelbar nærhet (Brettingen). I tillegg ble det høsten 2016 etablert et visningsanlegg ved Hestøya i nærheten av den sydvestlige delen av Lauvøysvaet.
2 - Prosjektmål
Hensikten med undersøkelsene i WP2 av ICOD-prosjektet var å skaffe til veie data om eggproduksjon og rekruttering av umoden torsk i gyte- og oppvekstområder som kan tenkes bli påvirket av etableringen av et nytt oppdrettsanlegg i nærområdet. Hovedmålet var derfor å etablere kvantitative data på variasjon og endringer i gyting og rekruttering hos torsk, både på gyte- og oppvekstområder som kan bli påvirket av at det etableres oppdrettsanlegg i nærheten, og i referanseområder uten oppdrettsanlegg i sitt nærområde. Det ble satt opp tre delmål for undersøkelsene: 1) Undersøke om eggmengde på gyteområder nær det nye oppdrettsanlegget på Nørholmen ble påvirket av dette, 2) Undersøke om mengde og størrelsessammensetning av torskeyngel i nærliggende oppvekstområder til det nye oppdrettsanlegget ble påvirket av dette, og 3) Undersøke om etableringen av det nye anlegget på Nørholmen ville påvirke den genetiske strukturen i torskepopulasjoner i de nærliggende gyte- og oppvekstområdene.
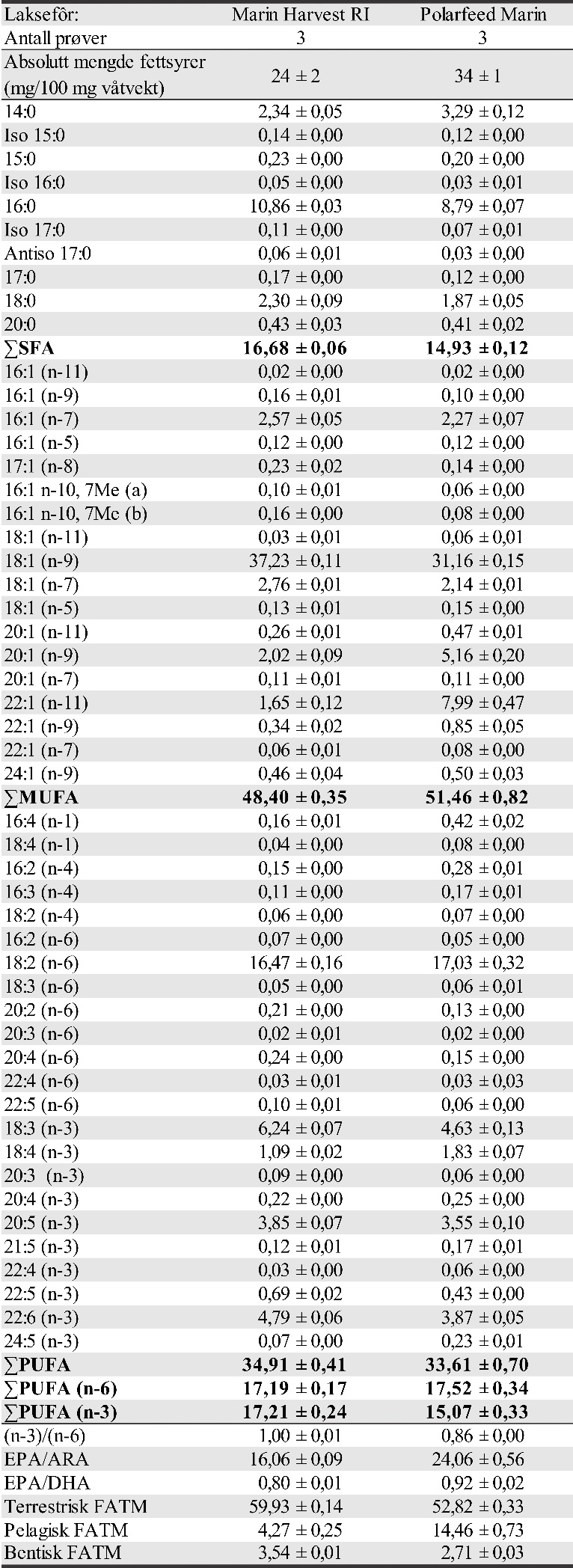
I tillegg ble innsamling av torsk gjennom gytesesongen på gyteområder på Smøla støttet av FHF for analyse av bestandstilknytning, og om fôrmidler fra landbruk som finnes i laksefôr, kan spores i torsken fra disse gyteområdene. Dette arbeidet ble gjennomført gytesesongen 2018 og 2019 i nært samarbeid med Nordmøre Fiskarlag (tidligere Smøla fiskerlag) som stod for fangst av torsk og sikring av prøver.
Eggundersøkelser med vertikale håvtrekk ble igangsatt ved prosjektstart i slutten av mars 2015 og ble gjennomført årlig tre ganger i mars fra 2016 til 2019. Variasjon i forekomst av egg vurderes også ut fra hydrodynamisk modellering av eggdrift (WP4 i ICOD-prosjektet). Videre ble årlige yngelundersøkelser gjennomført som rusefiske på grunt vann sist i oktober og begynnelsen av november fra 2015 til 2019. Genetiske metoder er benyttet for undersøkelser av både egg og yngel.
I henhold til prosjektleveransene er det skrevet årlige fagrapporter om egg- og yngelundersøkelsene (van der Meeren 2016; 2017; 2018; 2019). Den foreliggende rapporten er en faglig sluttrapport for WP2 i ICOD-prosjektet som viser og analyserer data fra egg- og yngeltoktene gjennom hele prosjektperioden, med vurderinger av resultatene ut fra problemstillingen om lokalisering av oppdrettsanlegg vil påvirke eggproduksjon og rekruttering hos torsk på nærliggende gyte- og oppvekstområder. I disse vurderingene trekkes også modellering av hydrodynamiske forhold inn (WP4 i ICOD). Videre presenterer og vurderer rapporten resultater vedrørende omfanget av gytemoden torsk på gyteområder på Smøla som har spist laksefôr under oppdrettsanlegg.
3 - Studieområder
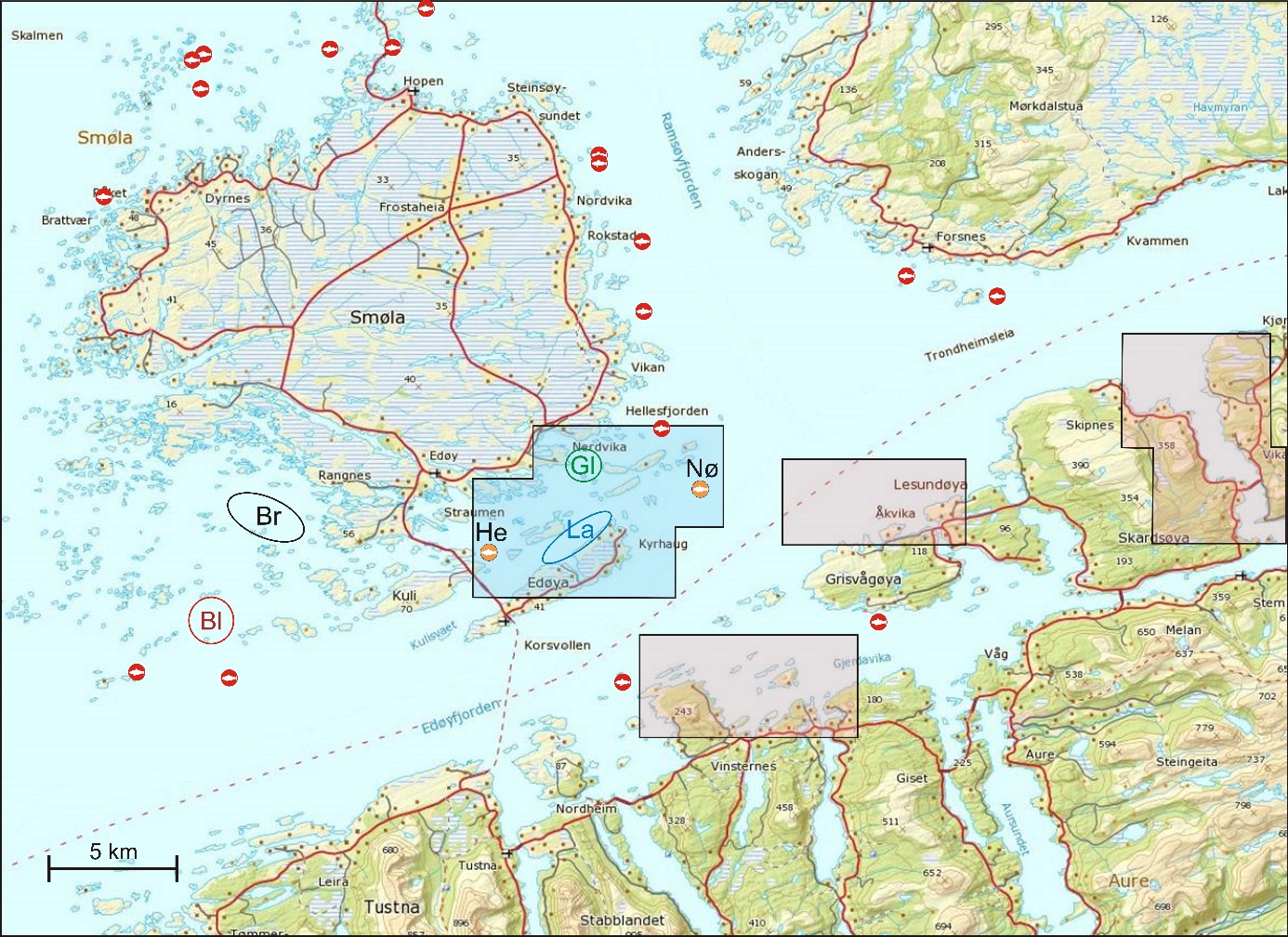
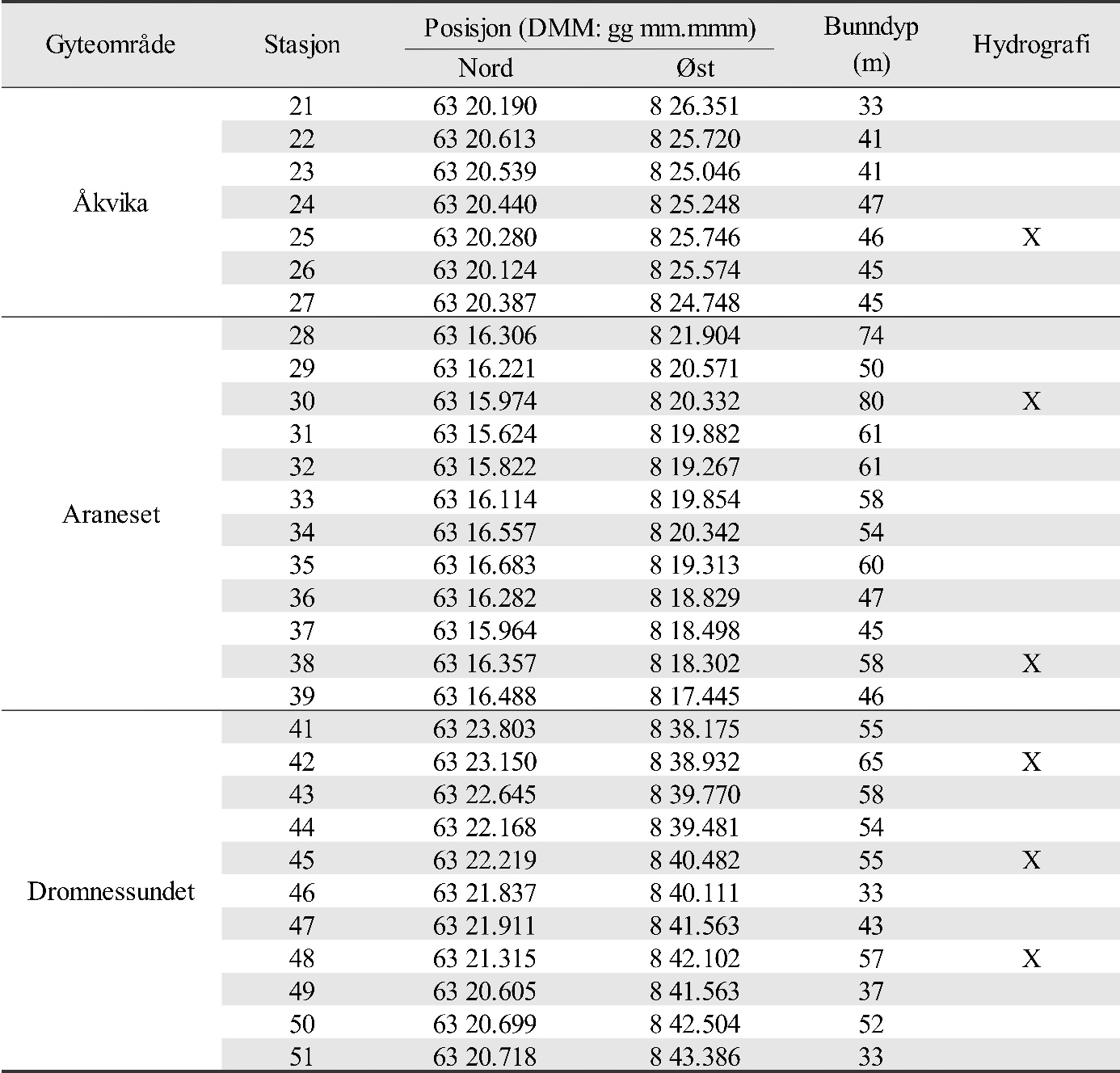
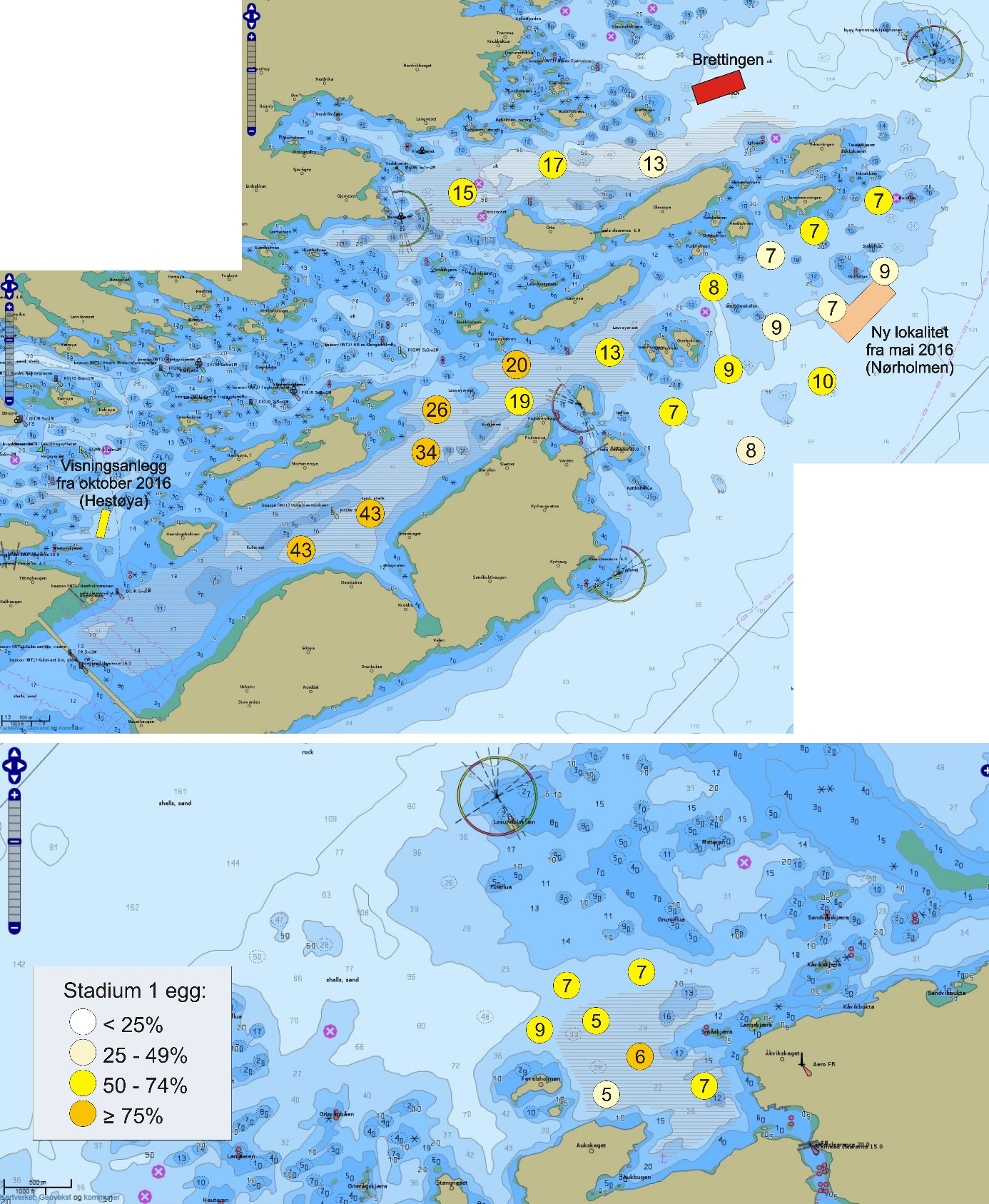
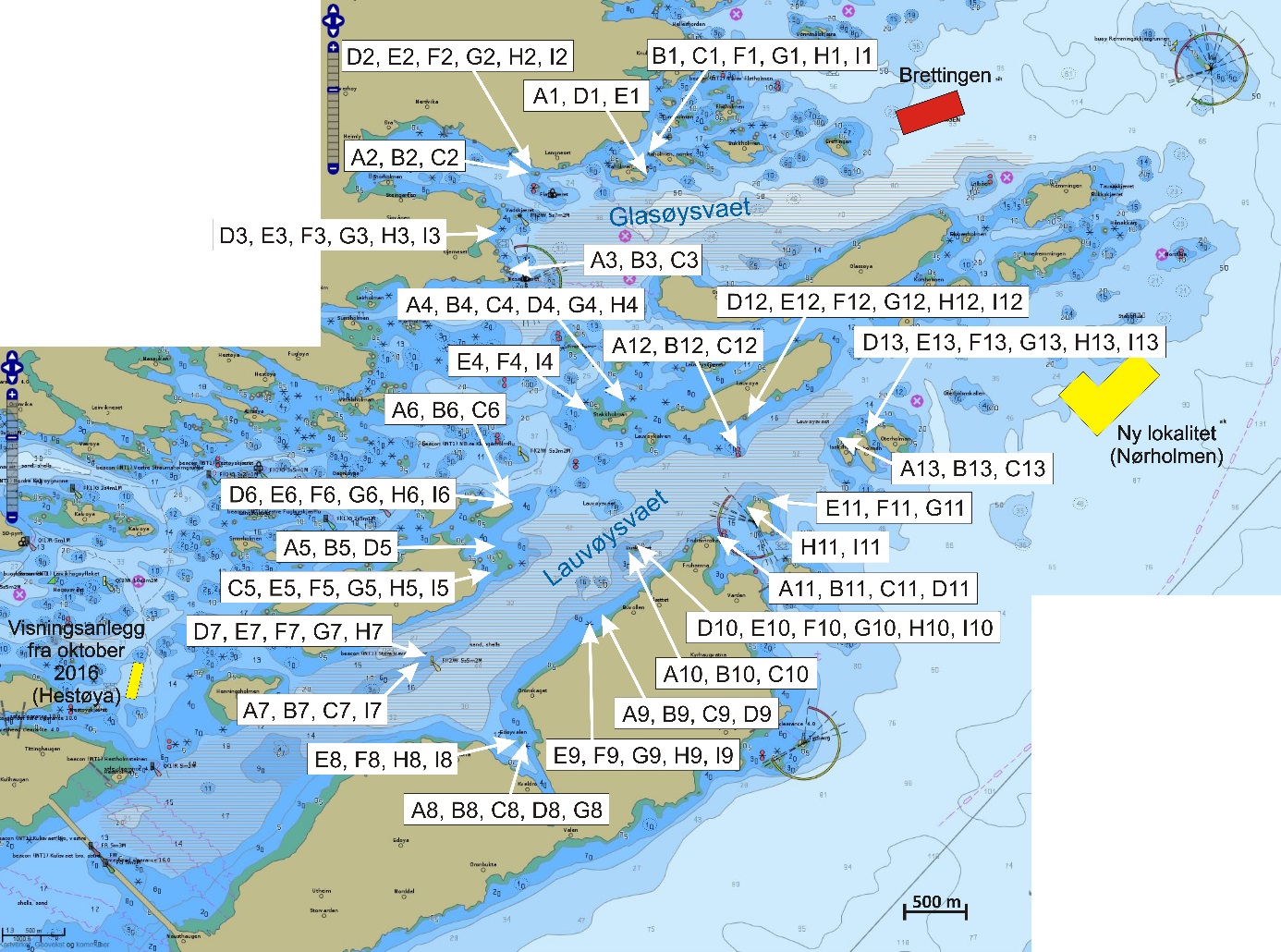
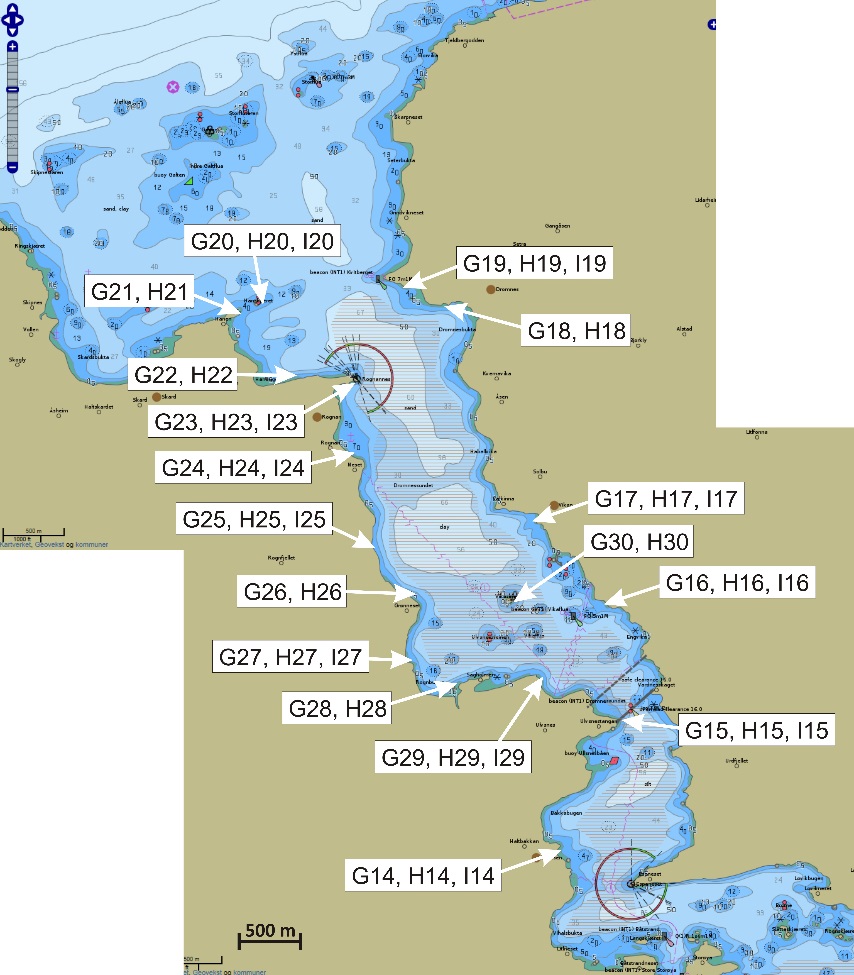
Figur 1 angir kartutsnittene for undersøkelsesområdene som er brukt i denne rapporten. Lyseblått utsnitt er hovedområdet med Glasøysvaet og Lauvøysvaet, mens rosa utsnitt er referanseområdene Dromnessundet (lengst til høyre), Åkvika (i midten) og Araneset (nederst). Videre vises aktive oppdrettsanlegg med røde symboler, og de to anleggene som ble etablert ved Hestøya (He) og Nørholmen (Nø) på Smøla i 2016 er vist med oransje symboler. Valg av referanseområder ble gjort ut fra et kompromiss ut fra at det ikke skulle være oppdrettsanlegg i umiddelbar nærhet til mulige vandringsruter for torsken inn til disse gyteområdene, samtidig som at lokalitetene skulle være mulige å undersøke for egg under ulike værforhold.
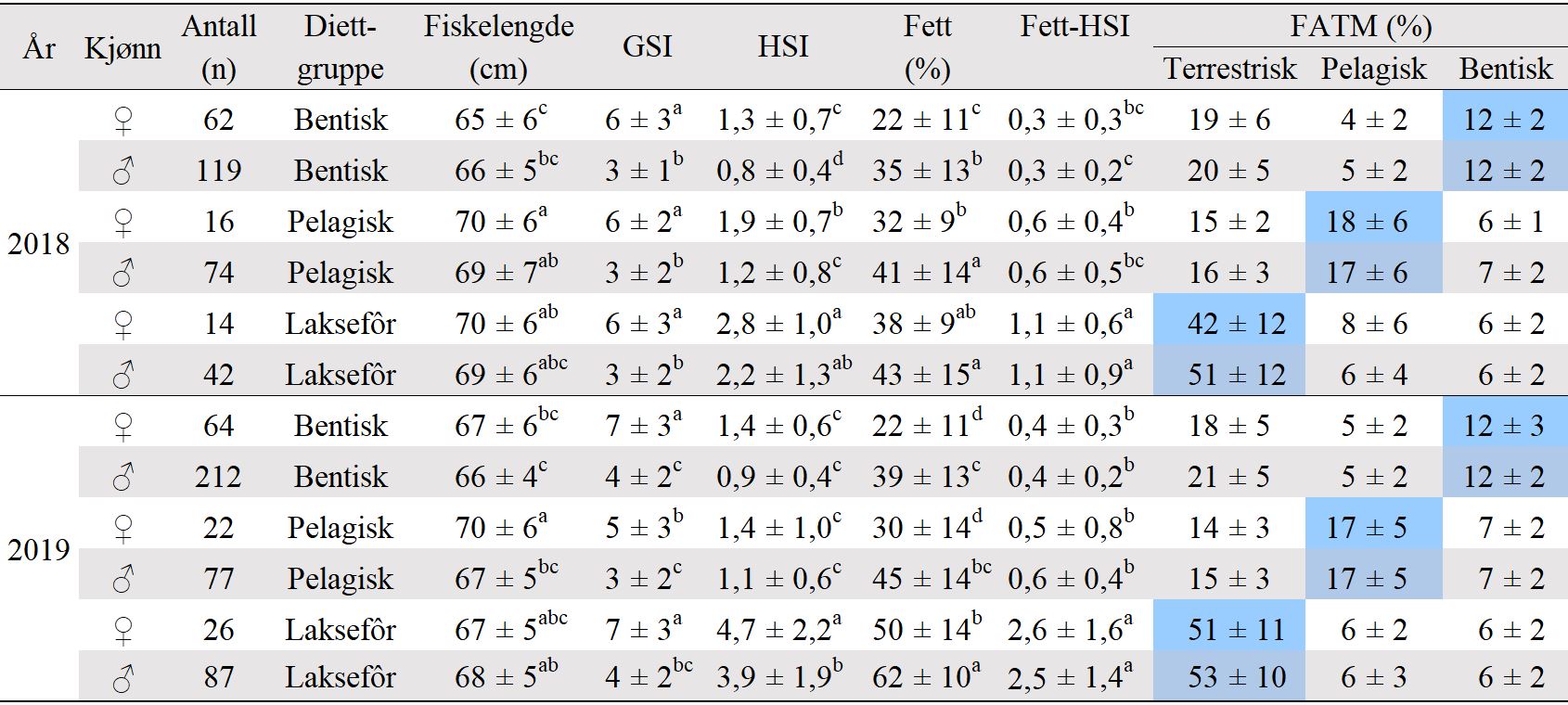
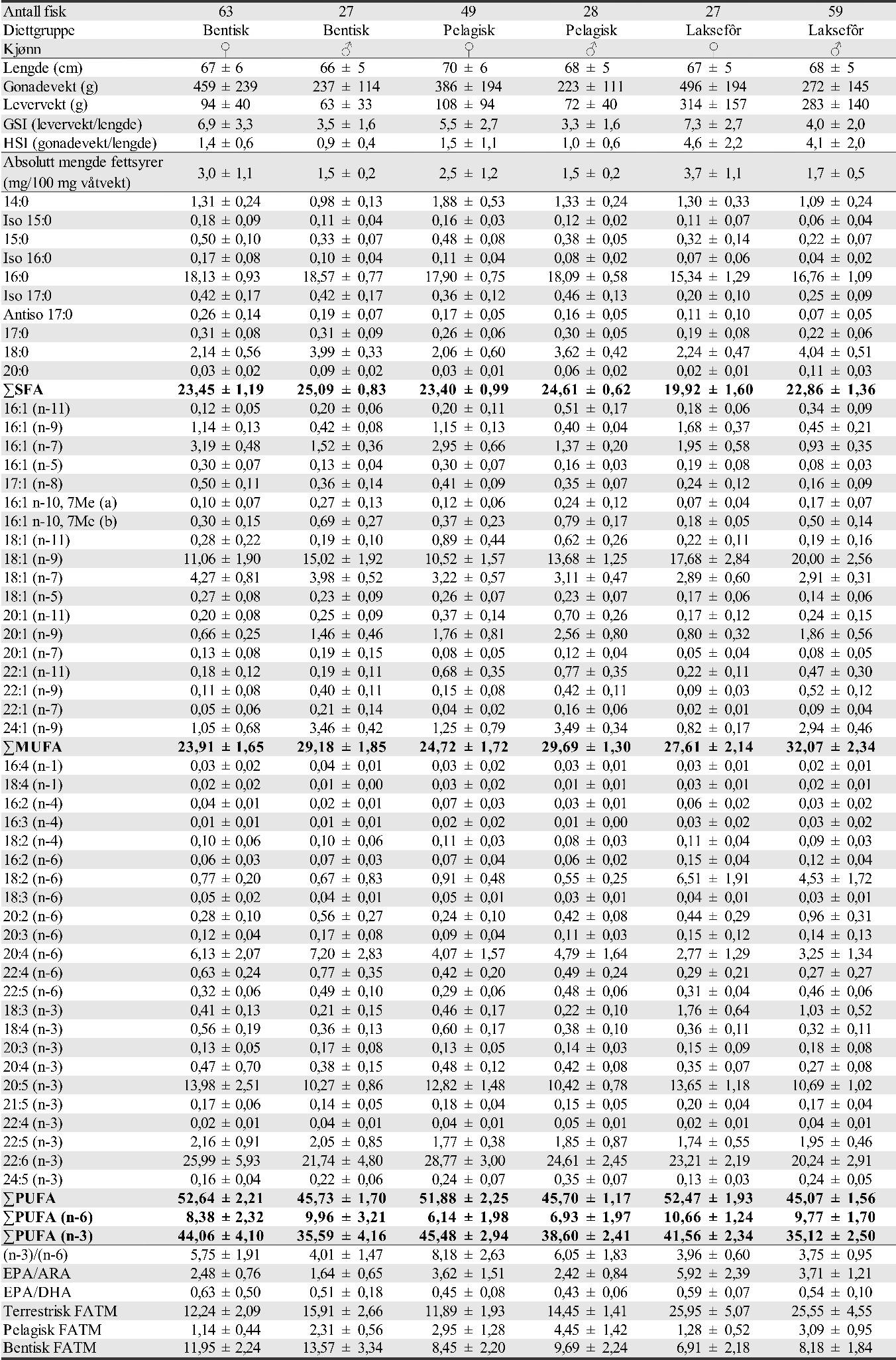
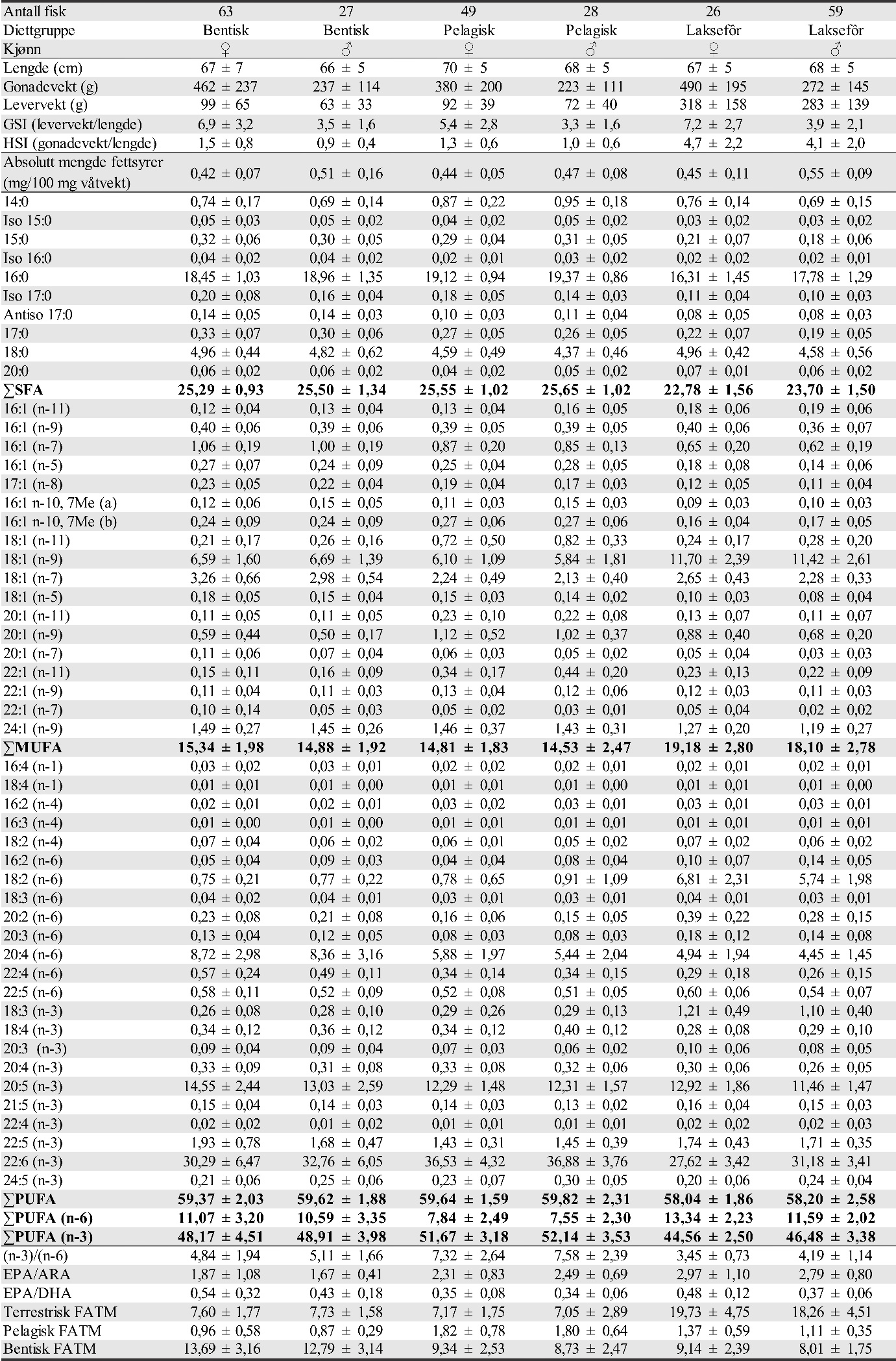
Også fiskeområder for gytemoden torsk som ble undersøkt med hensyn til fett og fettsyrer for å identifisere beiting på oppdrettsfôr, er angitt i kartet. Dette omfatter Blåsværsvaet (Bl), Breisvaet (Br), Lauvøysvaet (La) og Glasøysvaet (Gl), og sirklene i kartet angir område hvor garnsetting foregikk under fangsten.
Figur 1. Undersøkelsesområder på Smøla (lyseblått) og i Aure (rosa). Fargede sirkler og ellipser angir områder at torsk ble fanget inn gjennom gytesesongen for biokjemiske undersøkelser. Se teksten over for mer detaljert forklaring.
4 - Eggundersøkelser
4.1 - Metoder
4.1.1 - Innsamling av egg
Undersøkelsene ble gjennomført i samarbeid med Mowi AS (tidligere Marine Harvest AS) som stilte med egnet båt og båtfører. Base for undersøkelsene var Mowi sine fasiliteter på Vikan på Smøla, der opparbeiding av eggprøvene også skjedde umiddelbart etter prøvetaking. Eggundersøkelsene omfattet Lauvøysvaet og Glasøysvaet på Smøla, samt Araneset, Åkvika og Dromnessundet i Aure.
Undersøkelsen ble gjennomført etter metoder beskrevet av Espeland m.fl. (2013). Det ble benyttet en WP2-håv med 500 µm maskevidde og 56 cm diameter åpning. Håven ble senket ned til åpningen var 30 m under overflaten og trukket opp med en fart av ca. 0,5 m/sek. Dette er mindre dybde enn 50 m som beskrevet i Espeland m.fl. (2013), men 30 m ble valgt ut fra begrensninger i bunndyp på de aktuelle lokalitetene og stasjonene. Etter opptrekk ble håven forsiktig skylt med sjøvann, og prøven ble silt gjennom 2500 µm silkopp for å fjerne maneter og annet stort plankton. Deretter ble prøven silt gjennom en 750 µm duk for å fjerne det minste planktonet. Prøven med egg og resterende plankton som ble igjen på 750 µm-silen, ble oppbevart på 0,5 liters plastflasker som ble satt i skyggen i lufttemperatur (4-7 °C).
Innen 6-7 timer etter prøvetaking ble eggene i prøven manuelt skilt fra planktonet og fotografert. Dette skjedde i hallen på kaiområdet på Vikan, som har åpen port og er uten oppvarming. Opparbeiding av prøvene skjedde da ved 3-7 °C. Det ble benyttet en Olympus SZ61 stereolupe med fototubus og Moticam 10 (10 Megapixler) kamera koblet til bærbar PC med USB 2-kabel.
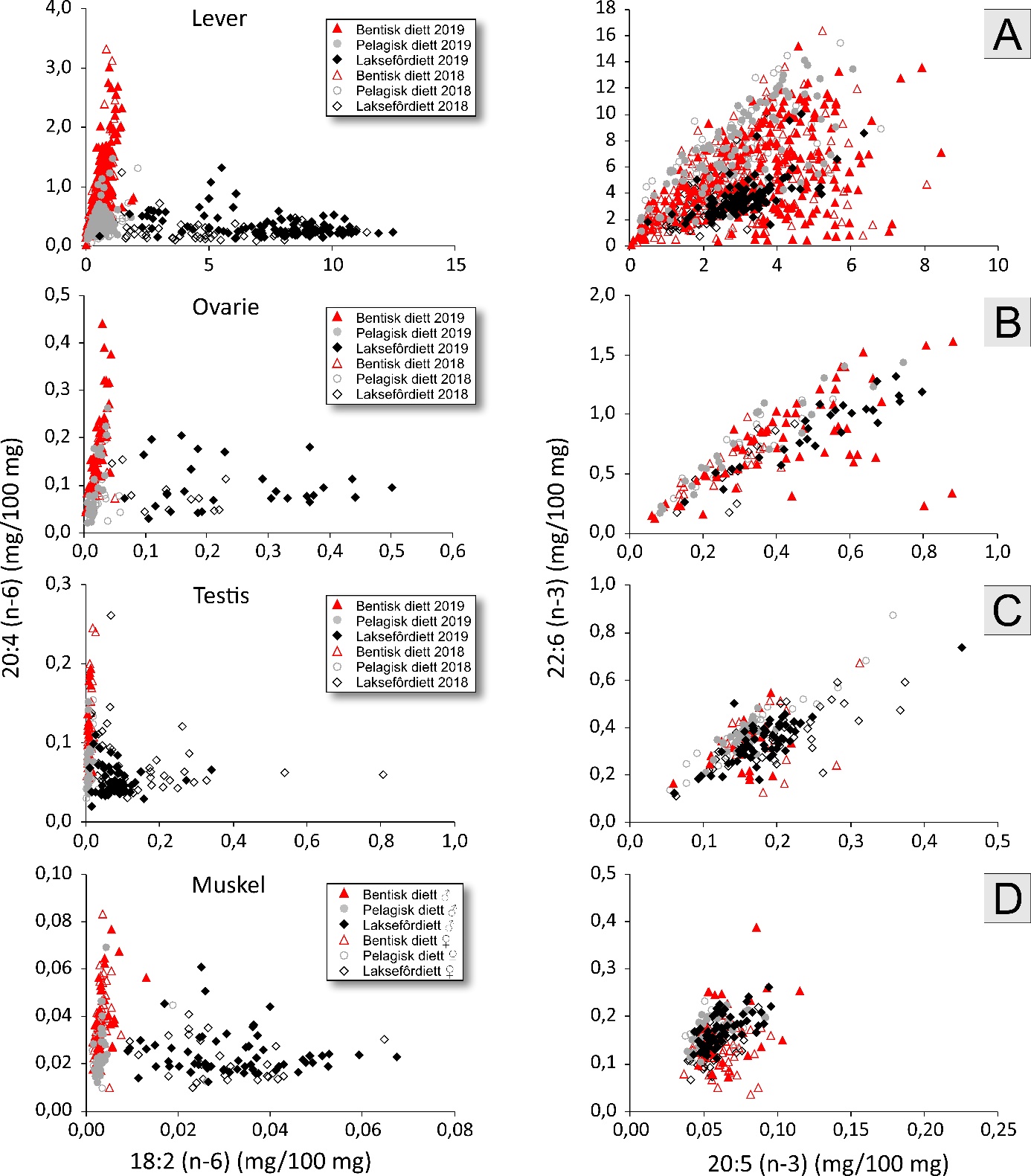
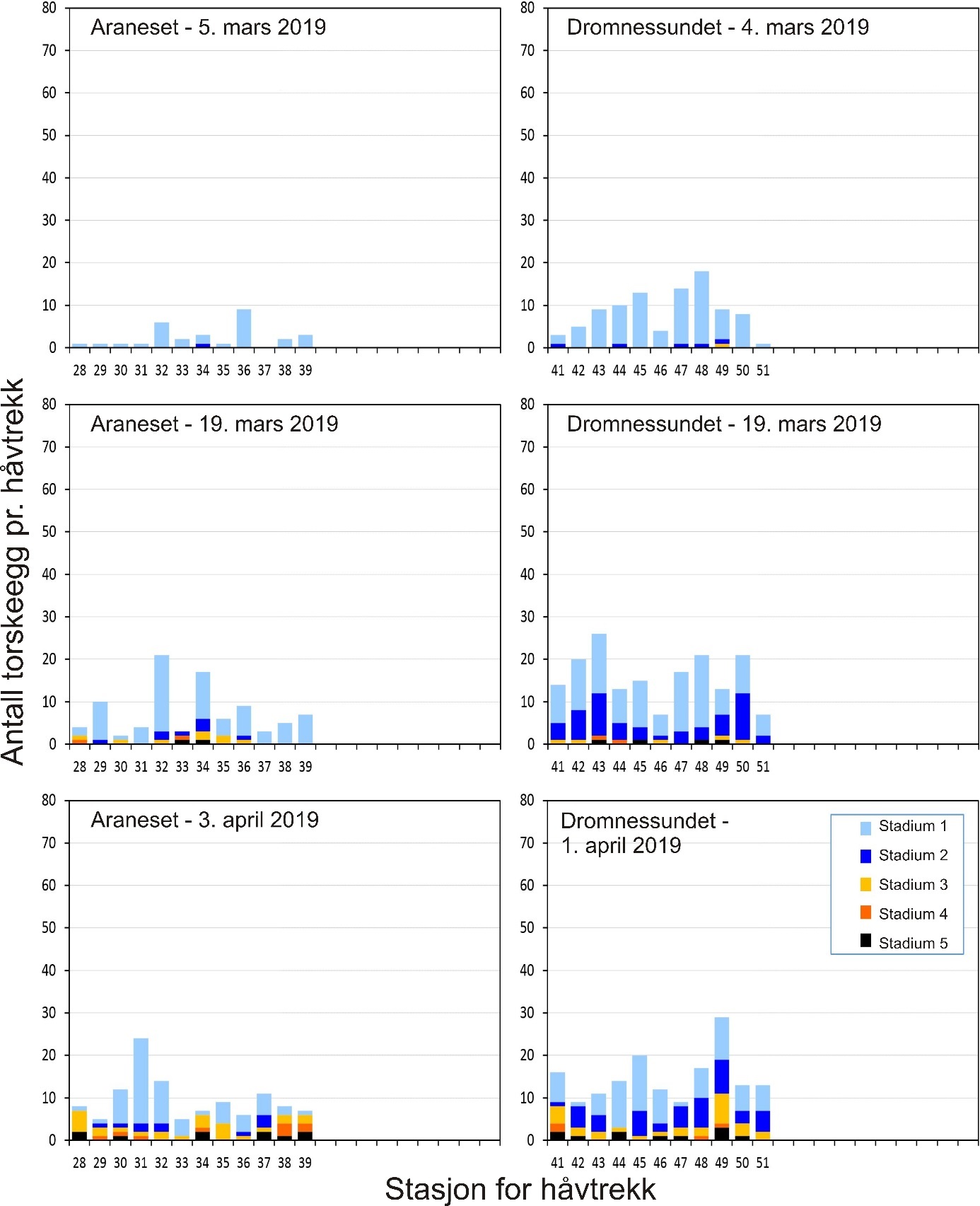
Figur 2. Egg fra stasjon 17 på Lauvøysvaet 31. mars 2018 (mørkefelt til venstre og lysfelt til høyre).
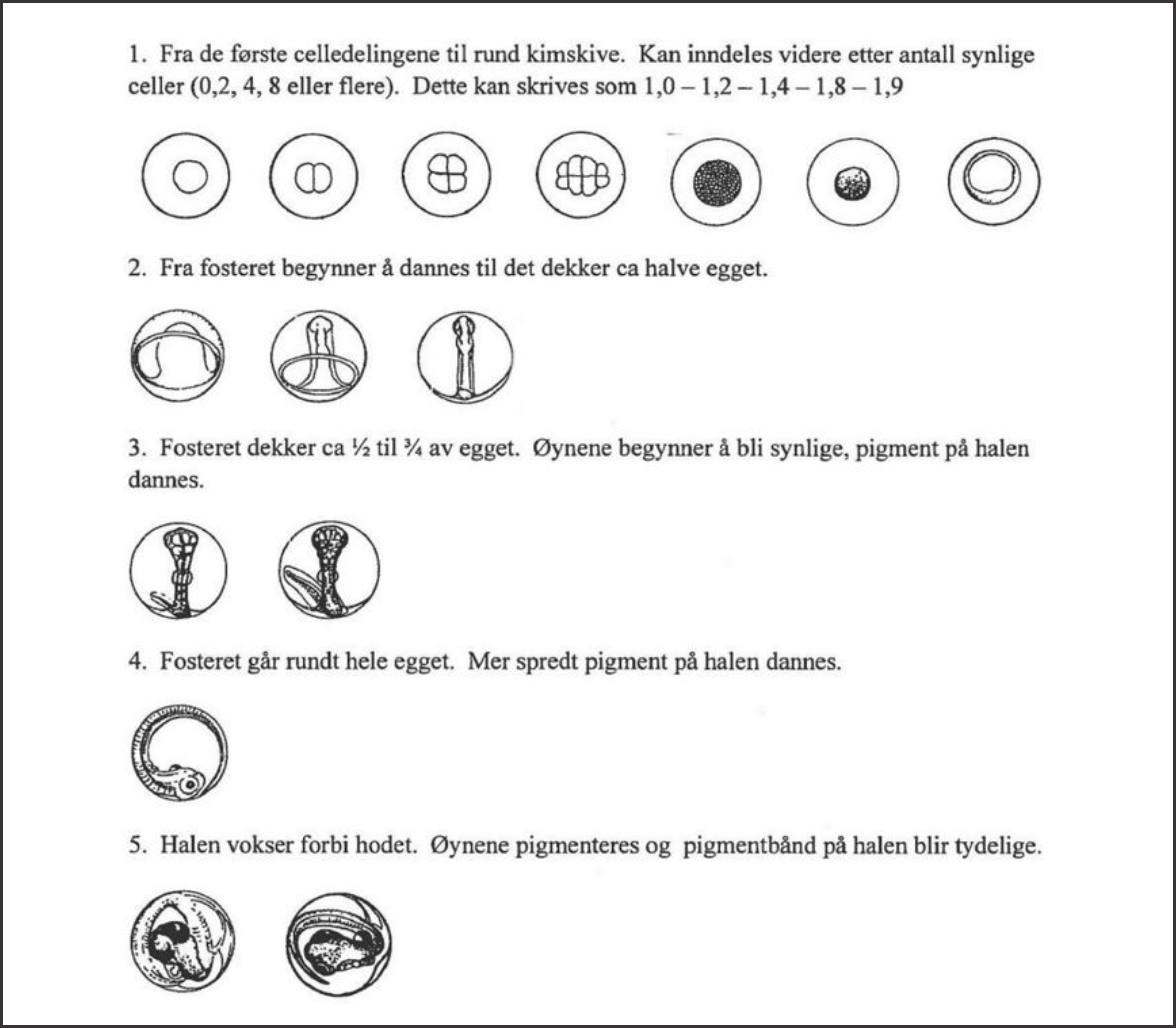
Denne lupen har LED kaldtlys som ikke vil føre til økt temperatur, og derved ikke påvirke eggenes overlevelse under fotograferingen. Bilder av en eggprøve ble tatt både i lysfelt (Oblique) og mørkefelt (DF) under 0,8 (8X) forstørrelse (figur 2). Eggstørrelse og utviklingsstadier ble senere bestemt fra fotografiene ved hjelp av Motic Images Plus 2.0 programvare. Stadier ble bestemt etter Thompson & Riley (1981), med modifikasjon av at stadium 1 ble delt inn i flere understadier som tilsvarte celleantall (1,0 – 1,2 – 1,4 – 1,8 og 1,9 for henholdsvis 1, 2, 4, 8 og flere celler, se figur 3). Stadier tidligere enn 1,9 ble sjelden observert, noe som kan forklares ut fra tidsrommet mellom da prøven ble tatt og til den ble fotografert. Bestemmelse av stadium gir derfor et litt forsinket bilde av faktisk eggutvikling på prøvetakingstidspunktet, særlig for egg like etter gyting som utvikler seg hurtig. Stadium 1 representerer derfor nærhet til gyteområdet, da disse eggene er relativt nylig gytt. En del egg var døde ved opparbeiding og kunne derfor ikke sikkert bestemmes til stadium. Mest sannsynlig var disse i stadium 1,9. Eggstørrelser på 1,2-1,6 mm diameter angir mulige torskeegg, og det er egg i dette størrelsesintervallet som presenteres i figurene. Alle egg ble fiksert på absolutt alkohol til senere DNA-analyser for å bestemme art. I tillegg til håvtrekk ble det innhentet hydrografiske data (saltholdighet, temperatur og oksygen) fra overflaten til bunn ved hjelp av en SAIV (SD 204) CTD-sonde med en Rinko III Oksygen-optode (modell ARO CAV-SA).
Figur 3. Bestemmelse av utviklingsstadium for torskeegg. Fra Espeland m.fl. (2013), modifisert etter Thompson & Riley (1981).
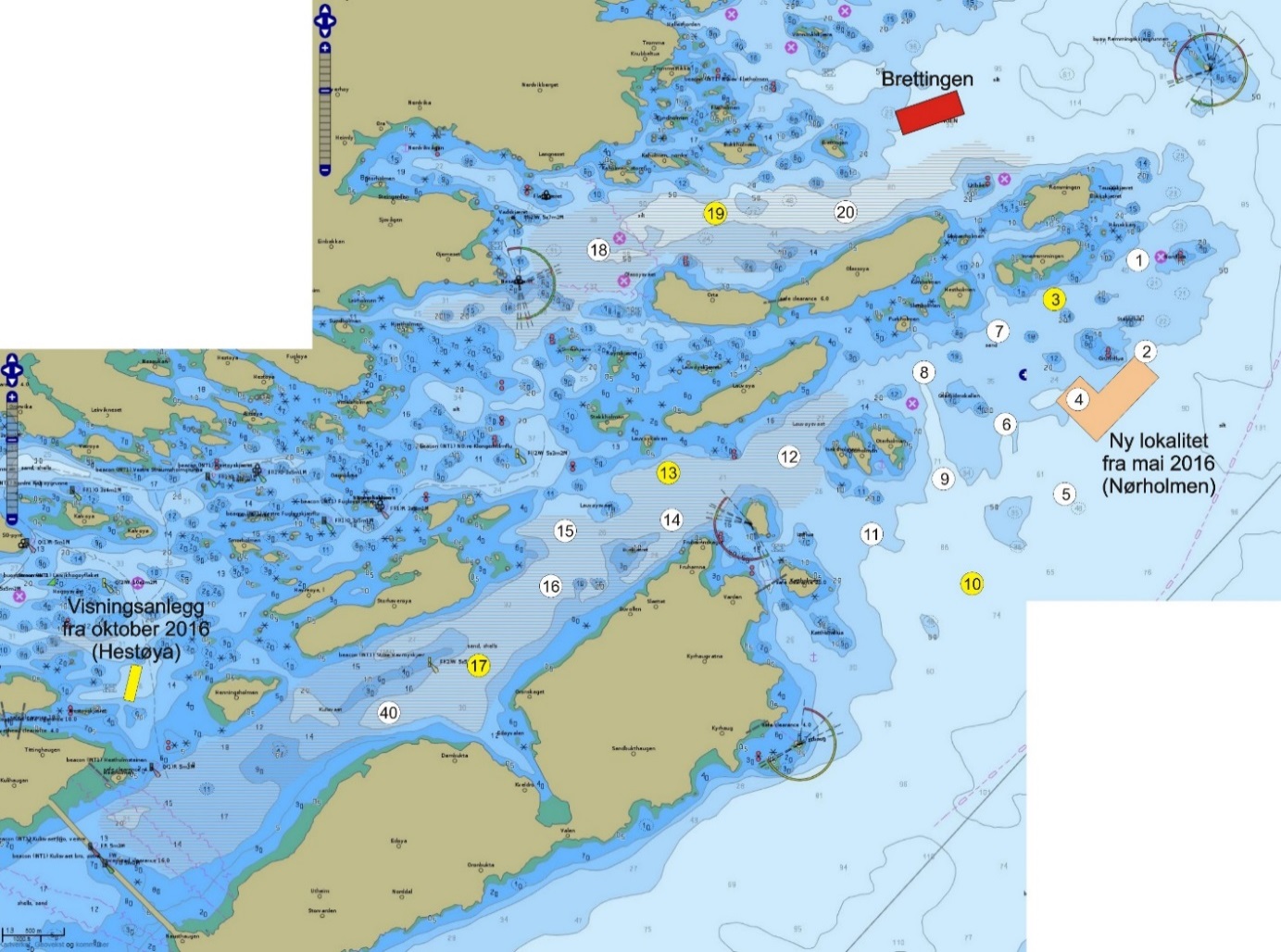
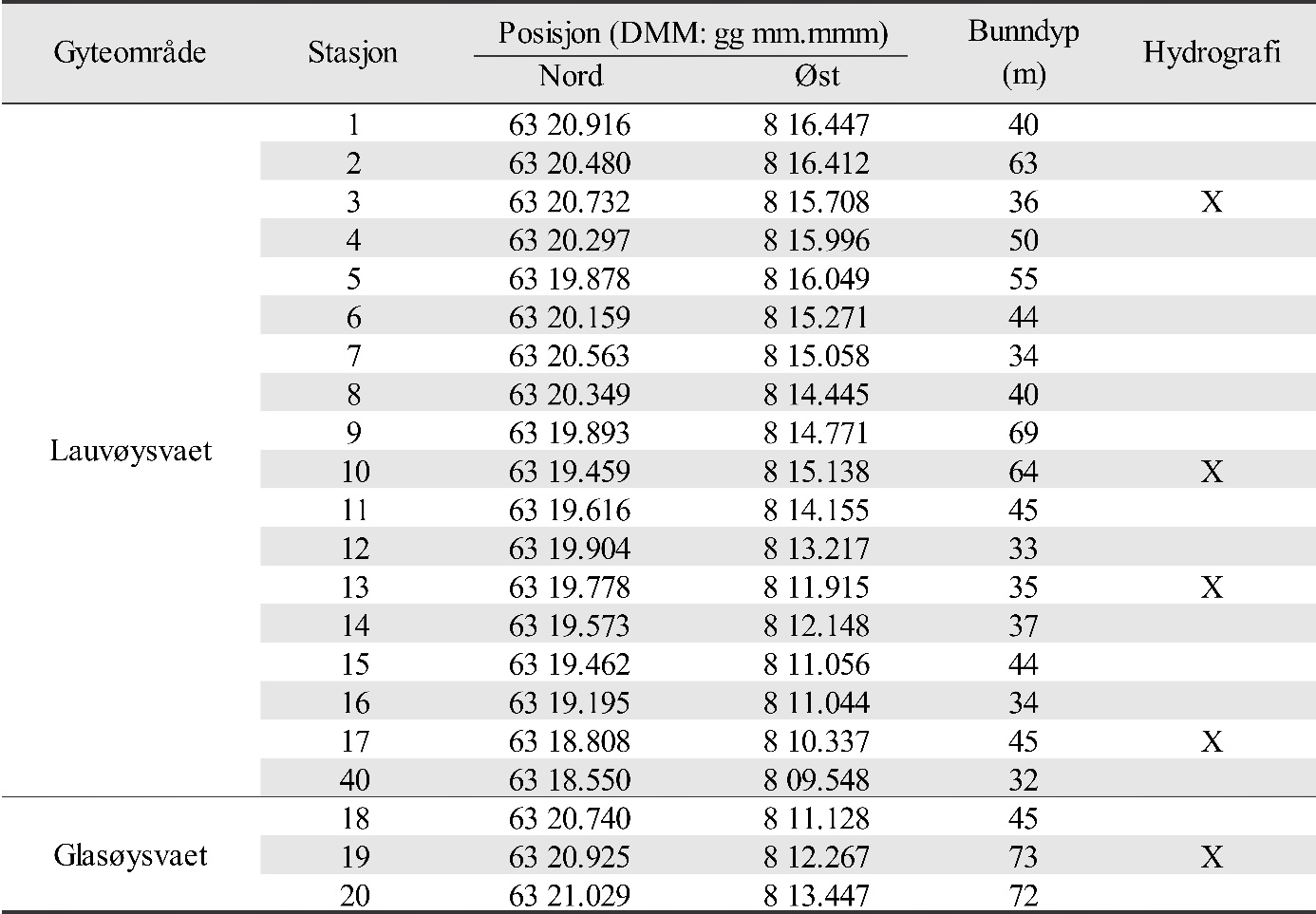
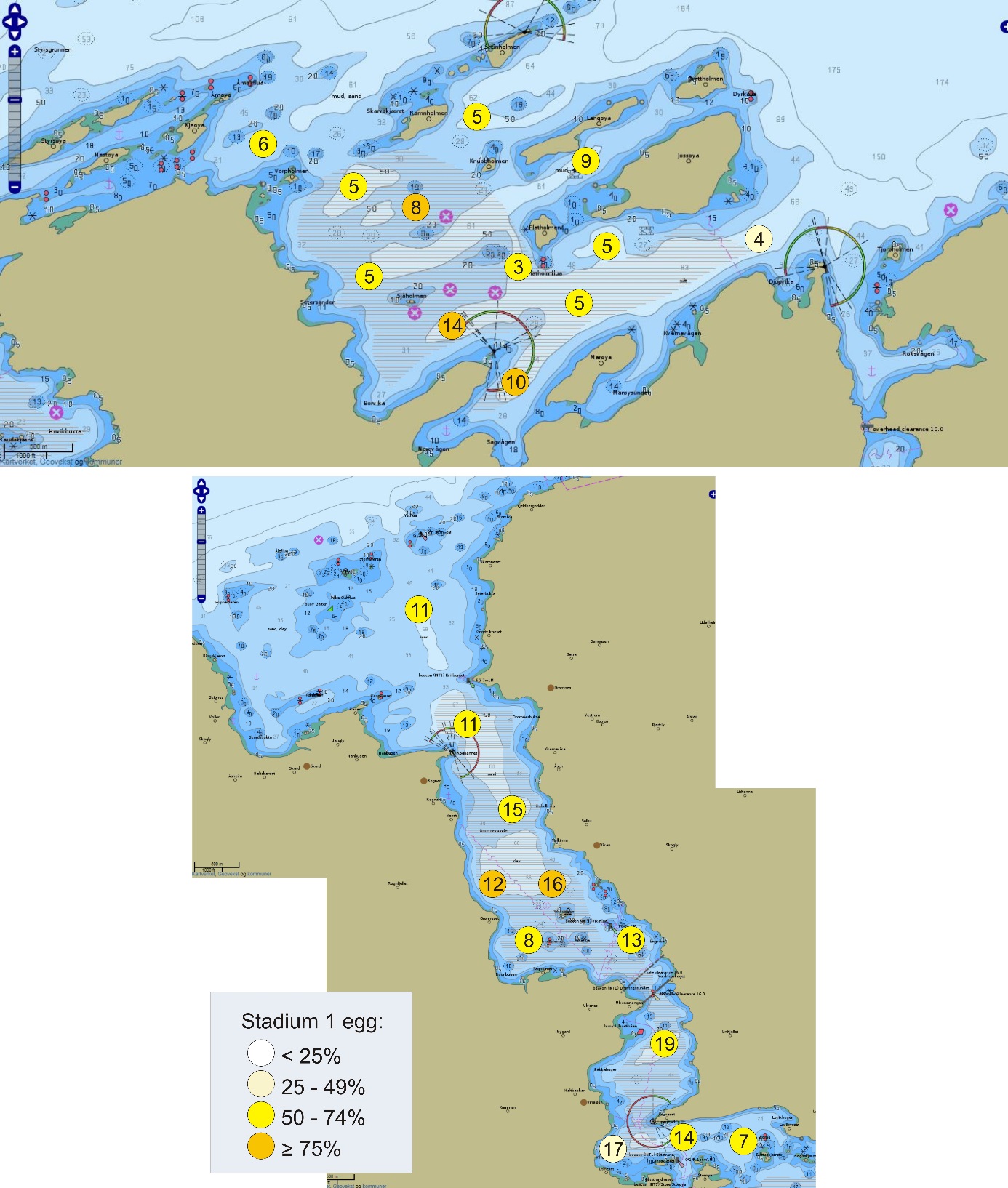
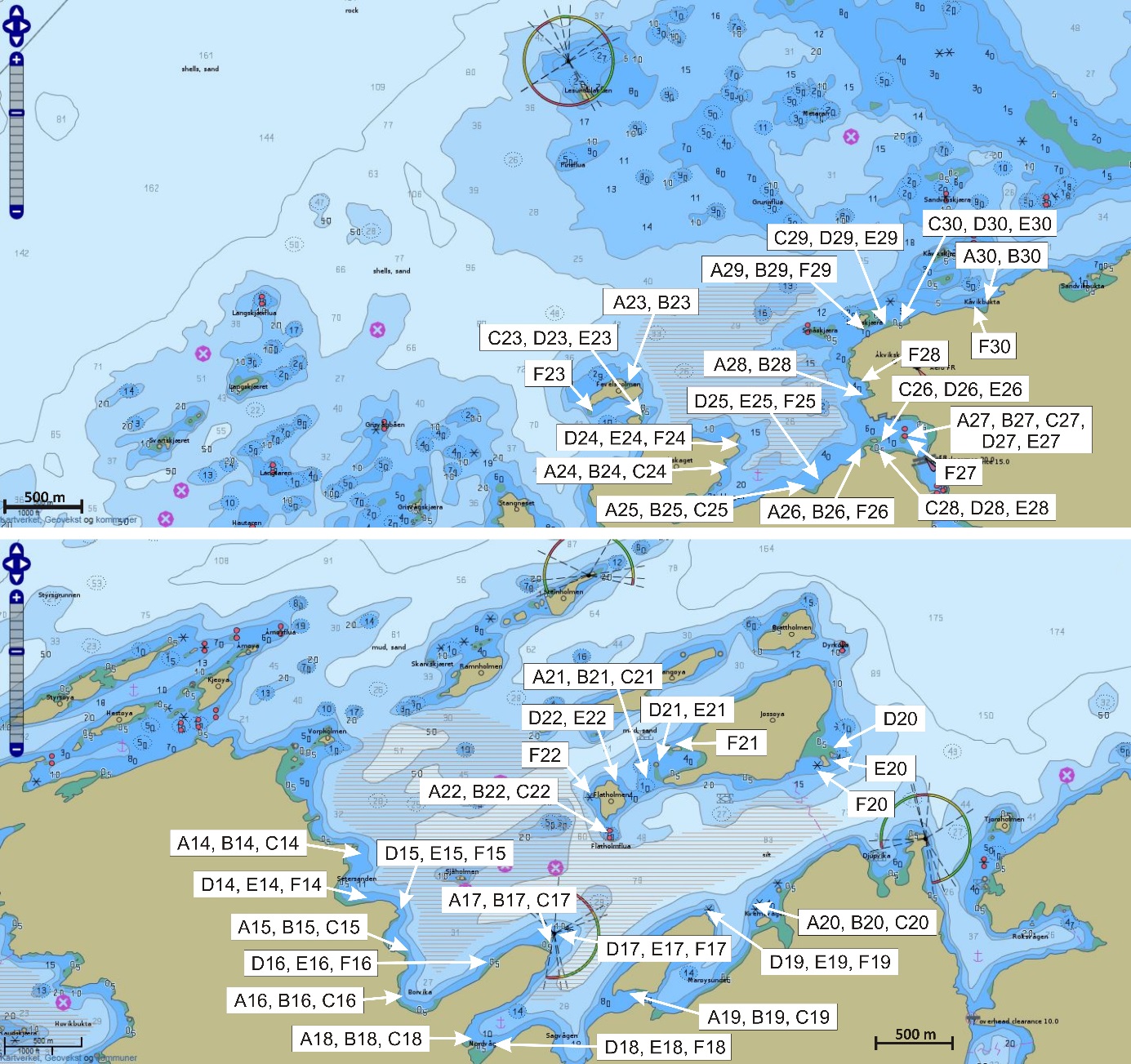
De undersøkte områdene og stasjonsnettet var de samme i perioden 2015-2019 (van der Meeren 2016; 2017; 2018; 2019), med unntak av at stasjon 40 (figur 4) og det nye referanseområdet i Dromnessundet (figur 6) ble lagt til i 2016. Stasjonsnettet ble valgt med utgangspunkt i avmerkede gyteområder som Fiskeridirektoratet i sitt kartverk har basert på informasjon fra fiskere (figur 4, 5 og 6). Da prosjektet startet i mars 2015 og stasjonsnettet ble bestemt, strakte gyteområdet på Lauvøysvaet seg helt ut til stasjon 1 (figur 4). Dette ble korrigert i kartverket i april 2015 da den østligste delen av Lauvøysvaet ble trukket tilbake til området ved stasjon 12, slik det er angitt på kartet (figur 4). Stasjonene ble valgt med tanke på å dekke det meste av disse gyteområdene, og samtidig ha nok dybde ned til 30 m (figur 4, 5, og 6, samt tabell 1.1 og 1.2 i Vedlegg 1). Det undersøkte hovedområdet ved Edøya er delt i to med flere grunne kanaler mellom dem. Den nordlige delen av dette området (Glasøysvaet, stasjon 18 til 20) har allerede et oppdrettsanlegg plassert ut mot Ramsøyfjorden i nordøst (12466 Brettingen, Mowi AS). Den sørlige delen (Lauvøysvaet, stasjon 1 til 17 og stasjon 40) er det området som har størst utstrekning og nærhet til nye oppdrettslokaliteter som ble satt i drift i løpet av 2016 (figur 4).
Figur 4. Hovedundersøkelsesområdet. Kartet viser Fiskeridirektoratets angivelse av gyteområder (brun skravering) pr. 1. mars 2016. Stasjonsnett for håvtrekk er vist som sirkler med stasjonsnummer inni. Gule sirkler angir hydrografiprøve i tillegg til håvtrekk. Gyteområde Lauvøysvaet omfatter stasjon 1 til 17 og stasjon 40, mens gyteområde Glasøysvaet omfatter stasjon 18 til 20. Nye oppdrettslokaliteter i 2016 er angitt sammen med eksisterende anlegg på Brettingen. Kilde: https://www.fiskeridir.no
4.1.2 - Statistiske metoder
Resultatene fra egginnsamlingen ble analysert med statistikkpakken R studio versjon 4.0.2 (R Core Team 2020, https://www.r-project.org/index.html). For å se om det var konsistente forskjeller mellom stasjoner innen gyteområder over år, ble det utført en standard ANOVA test etterfulgt av en Tukey post-hoc test for hvert område. I denne testen ble stasjon brukt som en kategorisk forklaringsvariabel for antall egg.
Forskjeller i antall egg mellom gyteområdene ble så testet ved hjelp av en lineær «mixed effect model» hvor område ble brukt som en kategorisk uavhengig forklaringsvariabel, og stasjoner nøstet innen år ble brukt som en «tilfeldig» effekt.
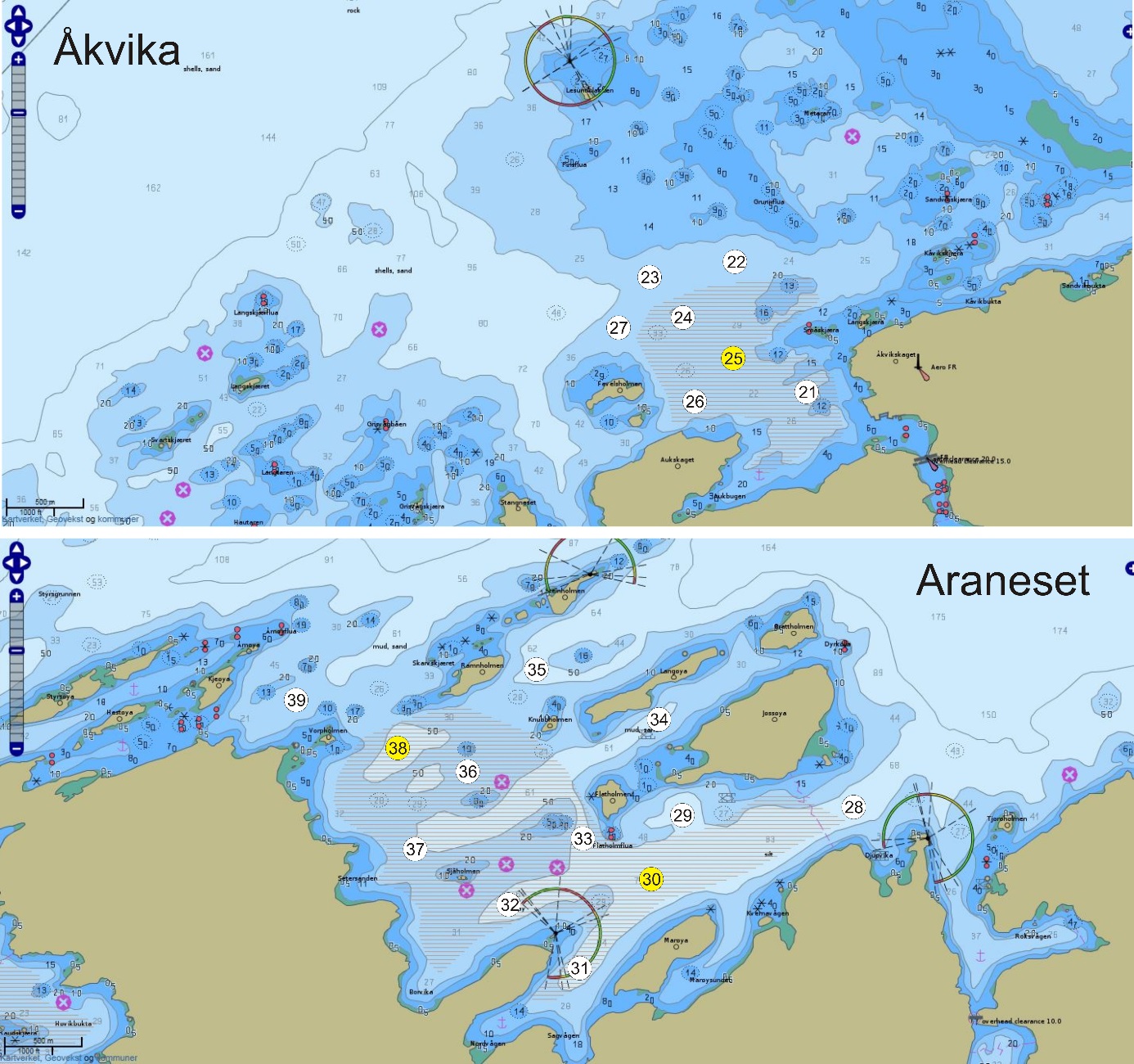
Figur 5. Referanseområdene Åkvika og Araneset. Kartet viser Fiskeridirektoratets angivelse av gyteområder pr. 1. mars 2016 (brun skravering) og stasjonsnett for håvtrekk med hensyn til dette (sirkler med stasjonsnummer). Gule sirkler angir hydrografiprøve i tillegg til håvtrekk. Kilde: https://www.fiskeridir.no
Dette ble etterfulgt av en Tukey post-hoc test for å sammenligne alle gyteområder mot hverandre. De tre eggtoktene tidlig, i midten og på slutten av mars ble rekodet til en kontinuerlig variabel med verdiene 1, 2 og 3. Denne testen ble etterfulgt av en Tukey Post hoc test for å sammenligne alle områder mot hverandre.
For å teste variasjoner mellom år innen gyteområder ble det benyttet en generell modell der hvert område ble testet for seg med år som kategorisk uavhengig variabel, innsamlingsperiode (tidlig, i midten og på slutten av mars) som kontinuerlig uavhengig variabel med verdiene 1, 2 og 3, og stasjon som en «tilfeldig» effekt. Disse modellene ble så etterfulgt av en Tukey post-hoc test.
Figur 6. Referanseområdet Dromnessundet. Kartet viser Fiskeridirektoratets angivelse av gyteområde pr. 1. mars 2016 (brun skravering) og stasjonsnett for håvtrekk med hensyn til dette (sirkler med tall). Gul sirkel angir hydrografiprøve i tillegg til håvtrekk. Kilde: https://www.fiskeridir.no
For å undersøke forskjeller i eggstadier mellom gyteområdene ble det benyttet en logistisk regresjon med antall eldre egg som responsvariabel. Som for de andre analysene ble eggtoktene omkodet til en kontinuerlig variabel med verdiene 1, 2 og 3 for tidlig, midt i og sent i mars. Innenfor hver periode innen et år ble alle stasjonene fra ett gyteområde slått sammen før analysen ble gjennomført. Eggstadium ble kodet til 0 hvis egget var i stadium 1, og ellers gitt verdien 1 for stadiene 2 til 5. Gyteområde (kategorisk variabel), periode og deres interaksjon ble brukt som forklaringsvariabler. År ble kodet som en «tilfeldig» effekt.
Undersøkelse av om eggstørrelse var forskjellig mellom områder og avhengig av tid på året (kalender dag/Juliansk dag), ble utført med en statistisk modell hvor den kategoriske variabelen gyteområde og kalenderdag ble brukt som forklaringsvariabler for eggstørrelse. Stasjon ble nøstet innen år og brukt som en «tilfeldig» effekt i denne modellen.
Alle statistiske tester er gjennomført med et signifikansnivå på 0,05, og p-verdier lavere enn dette gir statistisk signifikante forskjeller i de testene som er gjennomført.
4.1.3 - DNA-analyser av egg
Eggene ble plassert enkeltvis i 200 µl brønner med en 75 μl løsning bestående av 5 % Chelex 100 Resin og 15 μl Proteinase på et 96 brønners brett. Brettet ble så satt ved 56°C i en time, etterfulgt av 10 minutter ved 96°C. Etter en kort sentrifugering ble supernatanten (vesken som inneholder DNA) overført til et nytt 96-brønners brett. COI-genet (som sitter på mitokondrie-DNA) ble så amplifisert med PCR i 12 µl volum som inneholdt kombinasjoner av primer-par (LepF1_t1, VF1_t1, VF1d_t1, VF1i_t1, LepR1_t1, VR1d_t1, VR1_t1 og VR1i_t1). Etter PCR ble prøvene renset ved hjelp av ExoSap-IT. Løsningen ble satt ved 37°C i 15 minutter og deretter 15 minutter ved 80°C. Sekvensering av gener ble gjennomført ved hjelp av 1 μl M13F primer ved sekvenserings-laben til Universitetet i Bergen (https://www.seqlab.uib.no). Analyse av gen-sekvensene ble gjennomført med programvaren Geneious v8.0.5. Gen-sekvensene ble sendt gjennom søkeprogrammet BLASTn i databasen GenBank (https://www.ncbi.nlm.nih.gov/genbank) med standard parametere og Barcode of Life Data System databasen (BOLD; https://www.boldsystems.org) og sammenlignet med referanse-sekvenser for å bestemme tilhørighet til art. Vi forkastet alle resultat hvor det var mindre enn 99 % identitet.
4.1.4 - Modellering av eggdrift
En hydrodynamisk modell (ROMS – Regional Ocean Modeling System) ble benyttet til å simulere havstrømmene på Smøla med omliggende gyteområder for årene 2013-2019. Informasjon om vind, elveavrenning, tidevann og storskala havstrømmer er inkludert i beregningene som inndata til den høyoppløselige modellen med 160 m horisontal oppløsning (Myksvoll m.fl. 2020). Kystmodellen NorKyst800, med 800 m horisontal oppløsning, gir informasjon om kyststrømmen inn til den høyoppløselige modellen (Asplin m.fl. 2020). Modellen produserer havstrøm, saltholdighet og temperatur som blir lagret hver time.
Resultatene fra strømmodellen blir brukt som inndata til en larvedriftsmodell. Partikkelspredningsmodellen LADIM (Lagrangian Advection and DIffusion Model, https://github.com/bjornaa/ladim) bruker en egg- og larvemodul spesifikk for torsk (https://github.com/pnsaevik/ladim_plugins) med konstant gyteperiode fra 11.mars til 9.april hvert år. I simuleringene blir eggene blir sluppet daglig ut på kjente gyteområder i et avgrenset rutenett der hver rute tilsvarer den høyoppløslige strømmodellen (160 x 160 m). Størrelsen på området eggene ble sluppet ut i undere simuleringene omfattet 145 ruter for Glasøysvaet og 418 ruter for hovedområdet på Lauvøysvaet. Videre omfattet referanseområdene Åkvika 96 ruter, Araneset 193 ruter og Dromnessundet 185 ruter. I tillegg ble de omkringliggende gyteområdene Breisvaet og Blåsværsvaet (til sammen 381 ruter) og Hustadvika (738 ruter) inkludert for å vurdere tilførsel fra oppstrøms gyteområder.
4.2 - Resultater
4.2.1 - Eggundersøkelser
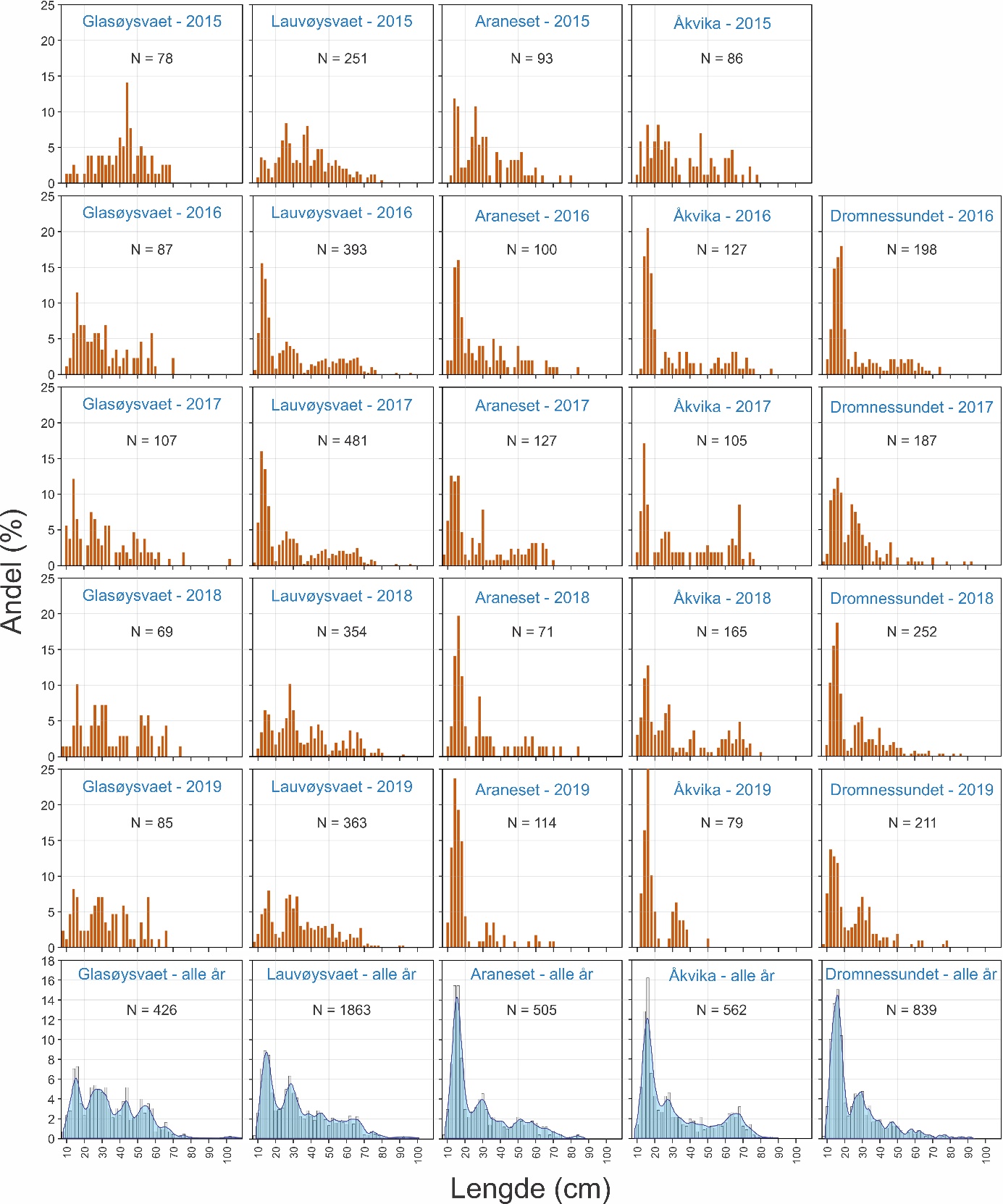
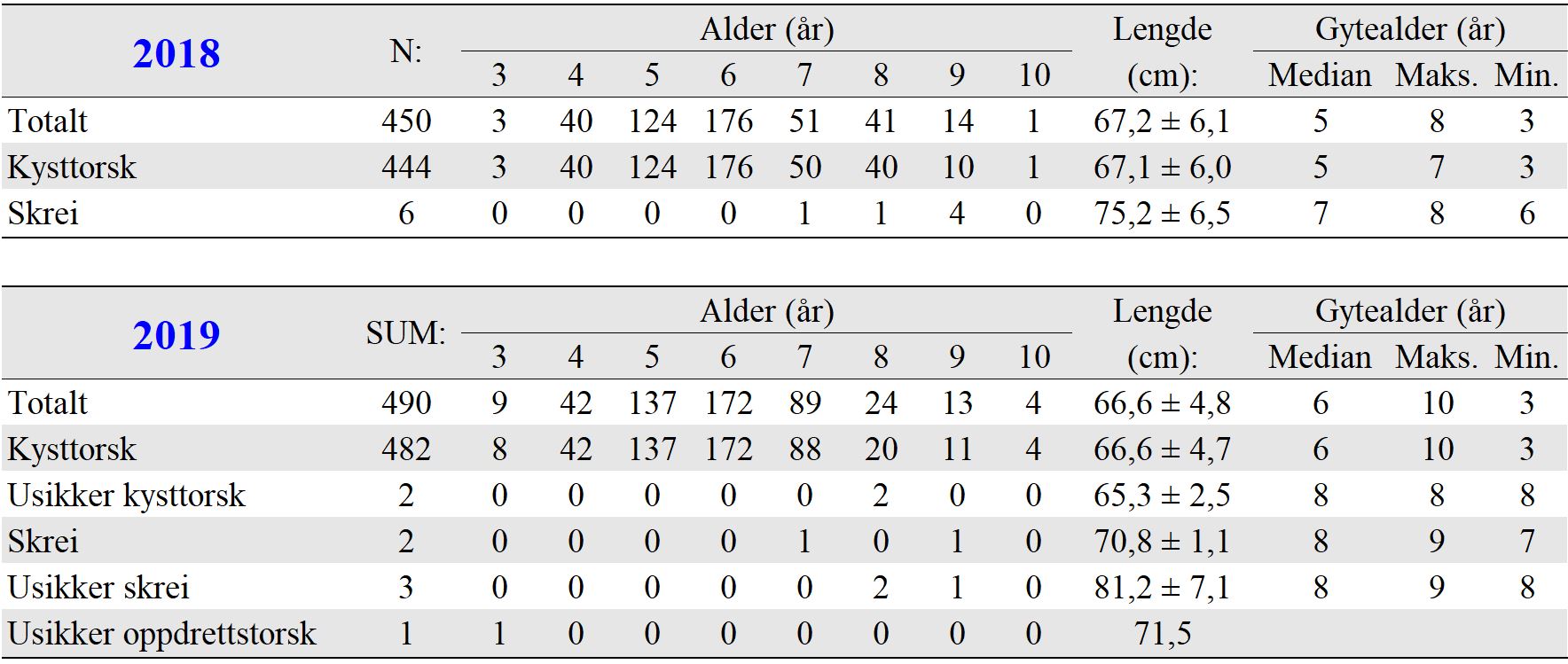
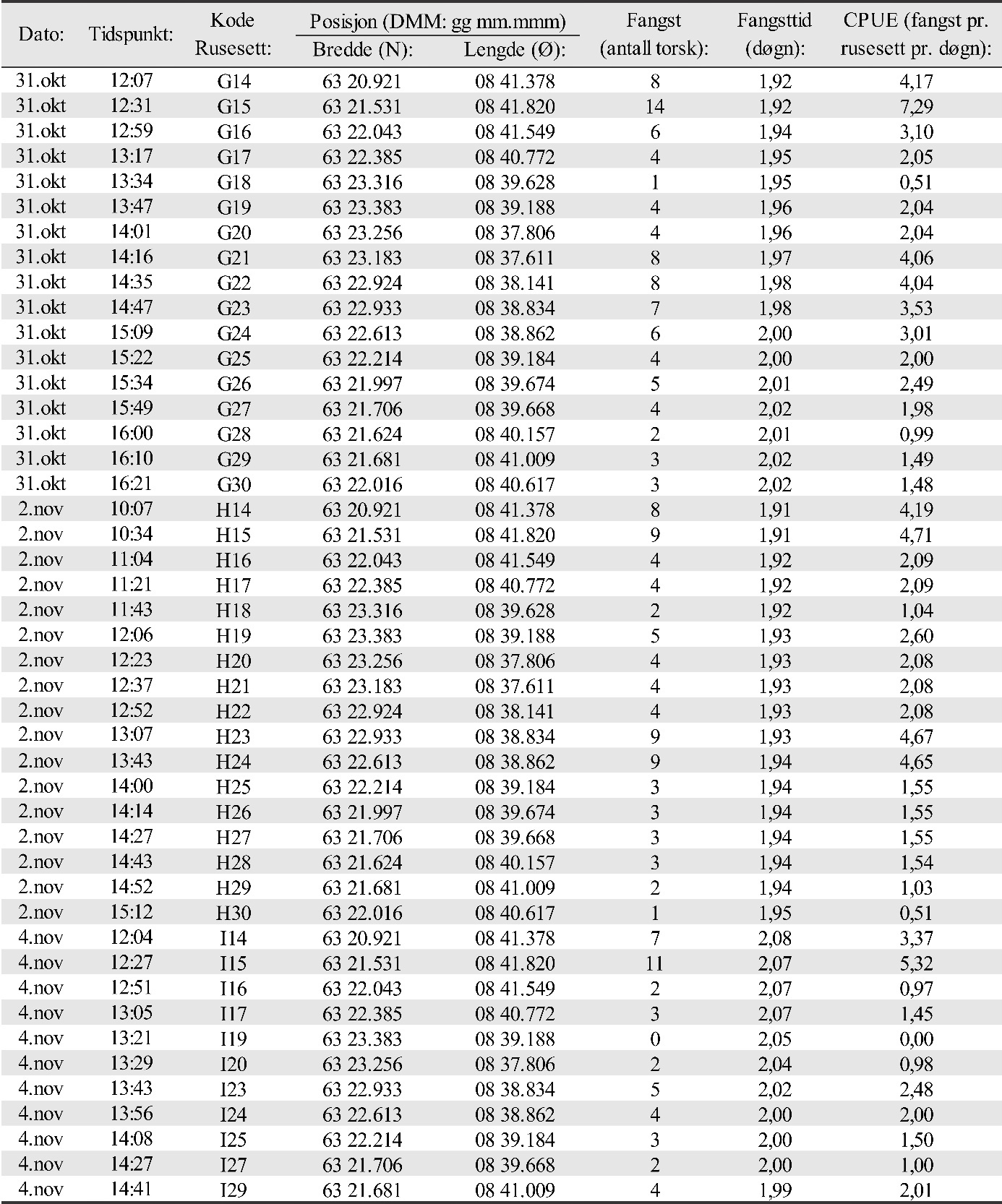
Gyteområder ble valgt ut fra kartverket til Fiskeridirektoratet som basert på innmeldinger fra fiskere. Eggundersøkelsene startet umiddelbart etter prosjektstart i mars 2015, og det første eggtoktet ble derfor gjennomført 25. mars for å verifisere at de valgte gyteområdene faktisk var gyteområder. Resultatene viste at gyting foregikk på alle de valgte områdene (van der Meeren, 2016). Det ble til sammen i løpet av prosjektperioden gjennomført 29 eggtokt der totalt 14300 egg ble samlet inn. Egg pr. håvtrekk ble benyttet som et standardisert mål på mengde av egg i de øverste 30 m av vannsøylen. Detaljer om mengde, fordeling og stadier av eggene er tidligere publisert i egne rapporten for 2015 til 2018 (van der Meeren, 2016; 2017; 2018; 2019), og tilsvarende data for 2019 er gitt i Vedlegg 1 og 2.
Det ble funnet egg fra en rekke forskjellige arter. Arter kan identifiseres ut fra eggets utseende (morfologiske karakterer), blant annet størrelse, strukturer på eggeskallet, plommemassens utseende samt forekomst, farge og størrelse av oljedråper. I godt utviklede egg som nærmer seg klekking er det også mulig å identifisere art ut fra pigmentering på embryoet inne i egget. I tillegg til torsk og hyse ( Melanogrammus aeglefinus ) som enkelt kunne identifiseres på denne måten i stadium 5, var det mulig å identifisere en rekke andre arter. Blant disse var rødspette (Pleuronectes platessa) og gapeflyndre (Hippoglossoides platessoides) som begge har store egg med gule pigmentceller i seine embyonalstadier. Gapeflyndre skilles lett fra rødspette ved at gapeflyndre har stort perivitellint rom (stor avstand mellom eggeskall og plommesekk). Det ble også observert en del små egg som tilsvarte størrelsen av sandflyndre (Limandalimanda). Videre ble brosme (Brosme brosme) observert da denne har en tydelig oransje oljedråpe. Det ble også funnet egg fra hvitting (Merlangius merlangus) og brisling (Sprattus sprattus), sistnevnte hovedsakelig i innerste del av Dromnessundet. Også egg av laksesild (Maurolicus muelleri) ble funnet ved flere anledninger. I tillegg ble det funnet noen små egg med en eller flere oljedråper som ikke lot seg identifisere. Disse kan være fra ulike arter av tangbrosme. Det ble også observert egg med oljedråpe som tilsvarte størrelse som hos makrell (både diameter på egg og oljedråpe).
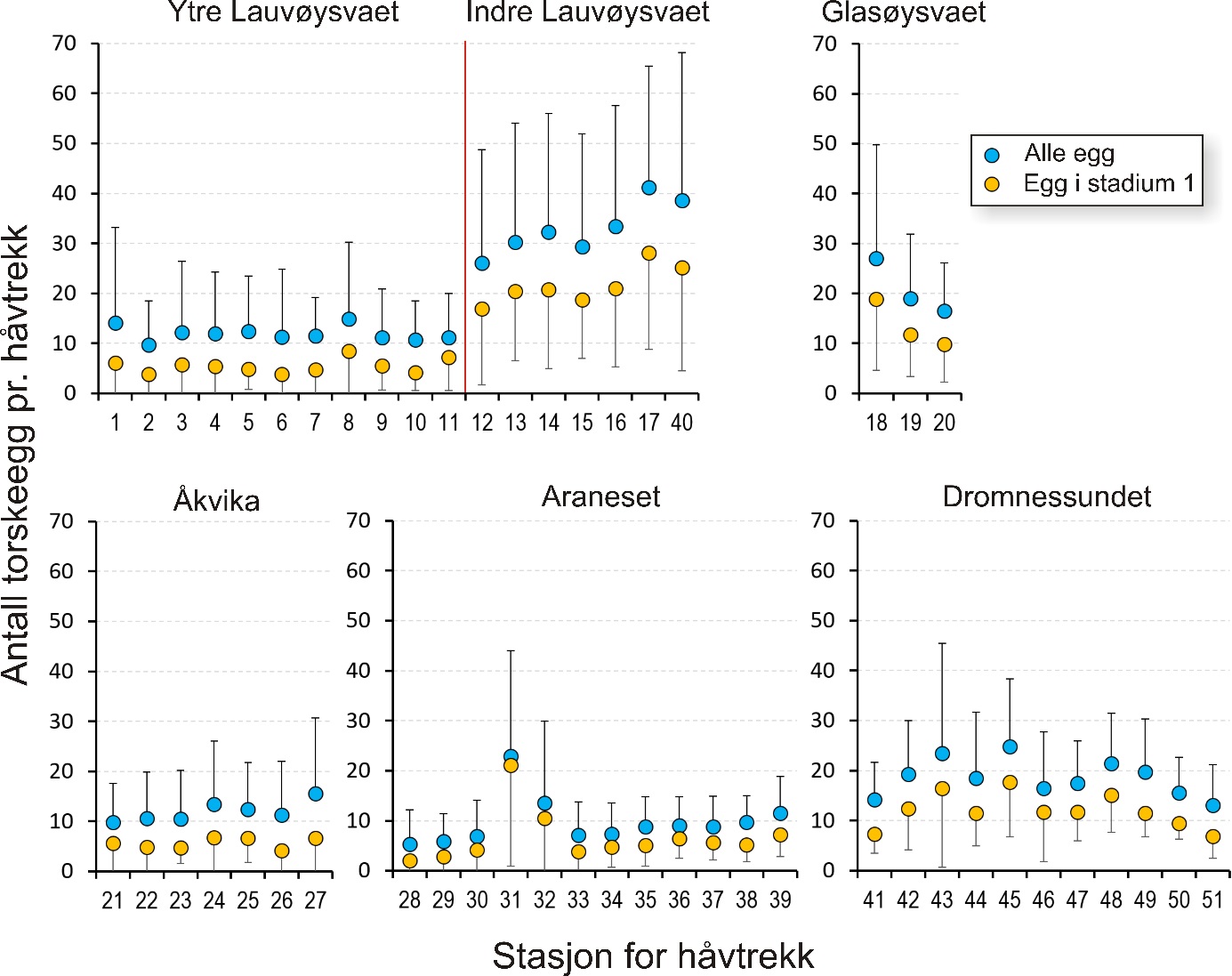
Eggmengde innen gyteområder
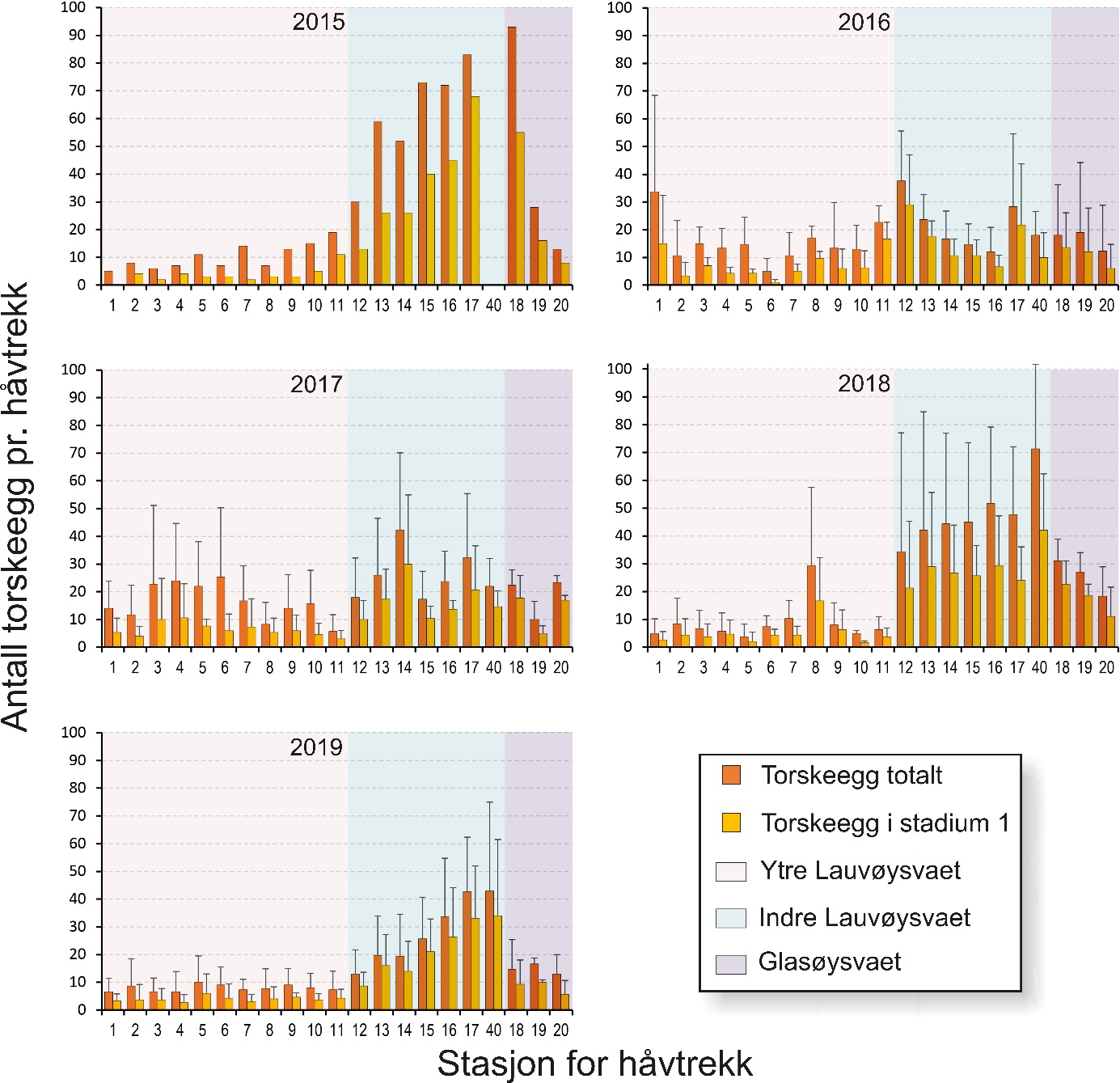
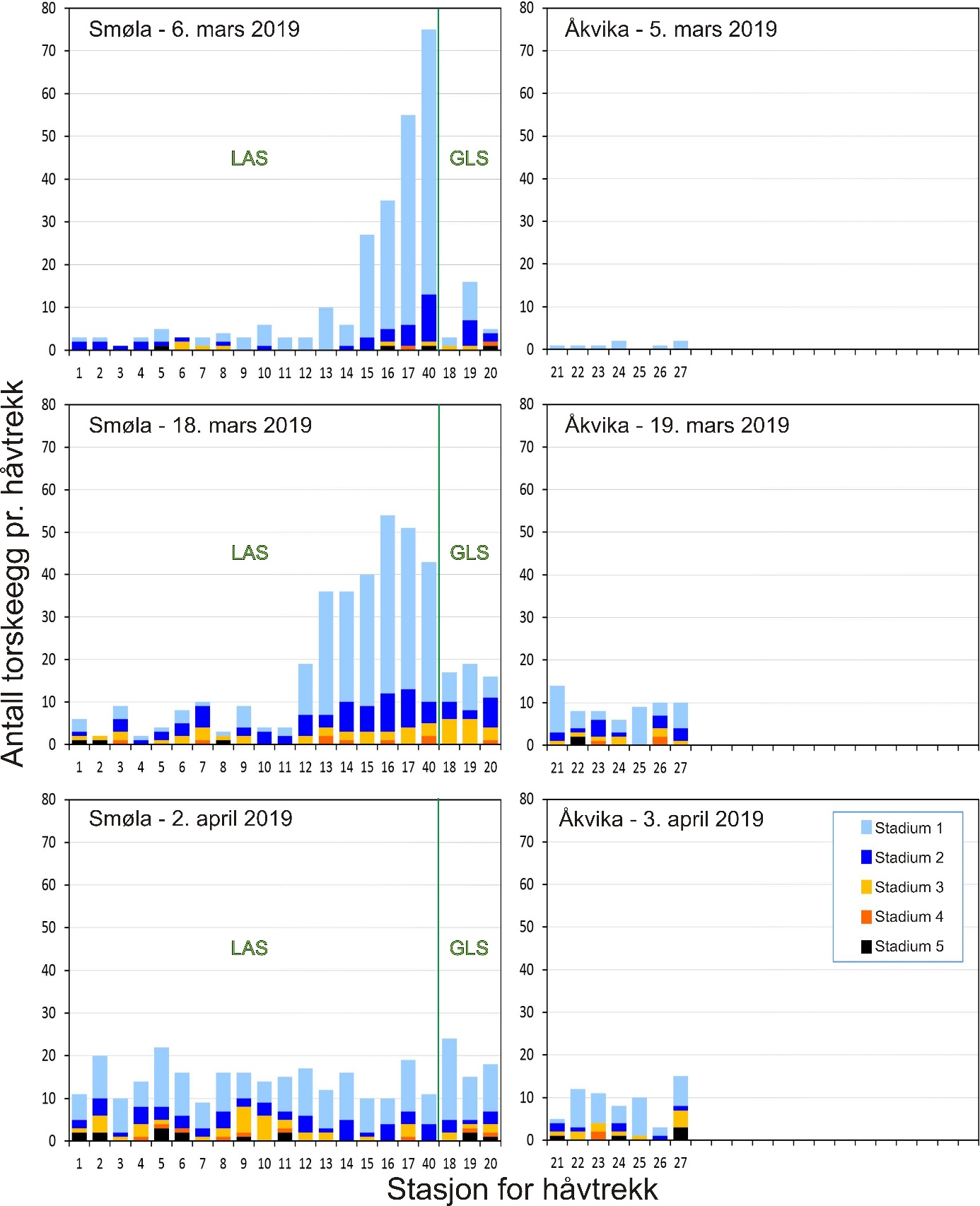
Gjennomsnittlig antall egg i torskestørrelse pr. håvtrekk av alle toktene for hele perioden 2015-2019 er vist i figur 7 for hver stasjon i alle de undersøkte gyteområdene. For gyteområdene på Smøla var det en tendens til flere egg innerst på Glasøysvaet (stasjon 18) som også hadde høyest andel av egg i stadium 1 (70 %). Videre var det et klart mønster med færre egg i den ytre og mer eksponerte delen av Lauvøysvaet (stasjon 1-11) sammenlignet med de mer beskyttede områdene av Lauvøysvaet på innsiden av Edøya (stasjon 12-17 og 40). Imidlertid var det kun stasjon 17 som var signifikant forskjellig fra den ytre delen av Lauvøysvaet, noe som kan forklares med det store antallet stasjoner på Lauvøysvaet som brukes til å justere signifikansverdien i post-hoc testene. En oppdeling av Lauvøysvaet i to områder syntes likevel fornuftig for videre statistisk behandling av resultatene, der ytre Lauvøysvaet omfatter stasjon 1-11 og indre Lauvøysvaet omfatter stasjon 12-17 og stasjon 40. Stasjonene på ytre Lauvøysvaet var da nærmest det nye oppdrettsanlegget nordøst for Nørholmen (figur 4). På stasjonene i indre delen av Lauvøysvaet var andelen egg i stadium 1 på mellom 63 og 68 %, mens den på ytre Lauvøysvaet var mellom 34 og 57 % på stasjonene 1 til 10 og 65 % på stasjon 11.
Innen referanseområdene ble det ikke funnet stasjoner på Åkvika som skilte seg fra andre stasjoner her (figur 7). Her lå andelen av egg i stadium 1 mellom 37 og 57 %. På gyteområdet ved Araneset var det imidlertid signifikant flere egg på stasjon 31 enn alle andre stasjoner her, med unntak av stasjon 32 og 39. Stasjon 31 befinner seg innerst på dette gyteområdet, i bukten sørøst for Araneset lykt. Denne stasjonen utmerket seg også med en svært høy andel av egg i stadium 1 (92 %), og til dels mange egg tidlig i stadium 1, mens stasjon 32 like nordvest for Araneset lykt hadde 78 % av eggene i stadium 1. I Dromnessundet ble det ikke funnet signifikante forskjeller mellom stasjonene med hensyn til mengde egg, men de fleste eggene ble funnet sentralt i sundet på stasjon 43, 45 og 48. På alle disse stasjonene var også andelen av egg i stadium 1 på vel 70 %.
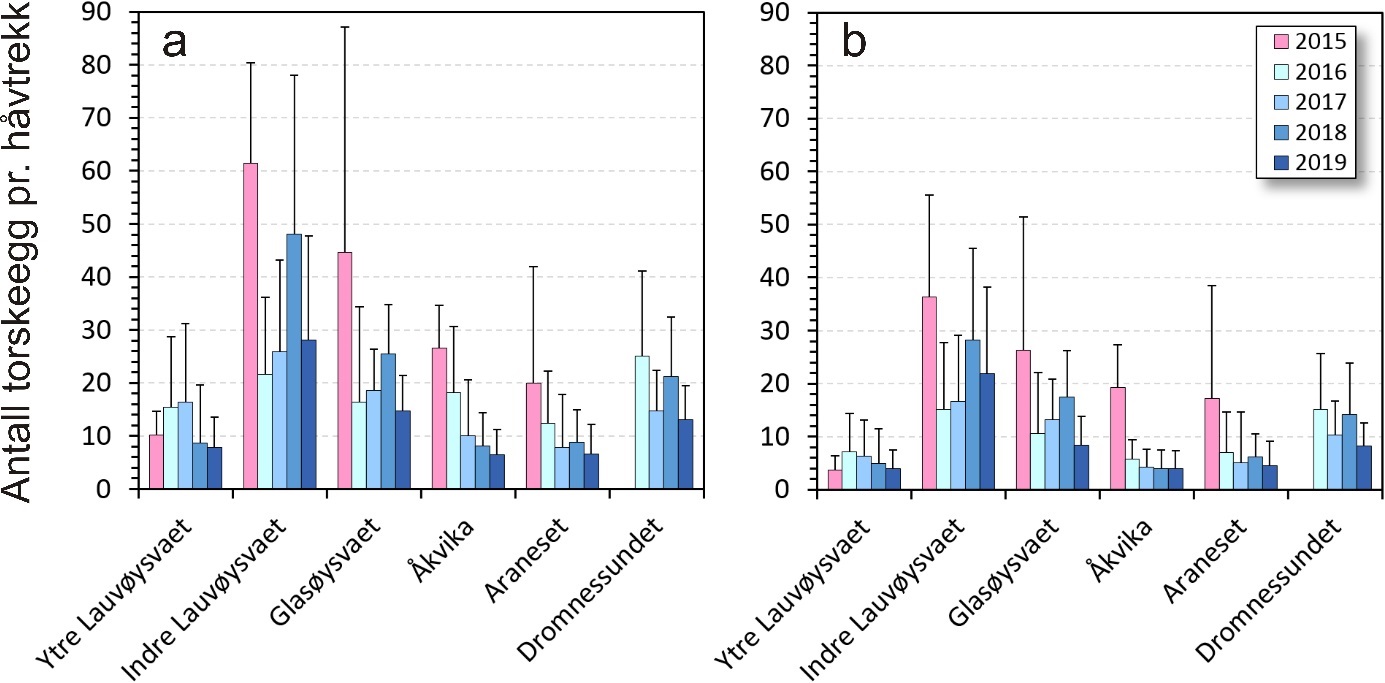
Eggmengde mellom gyteområder
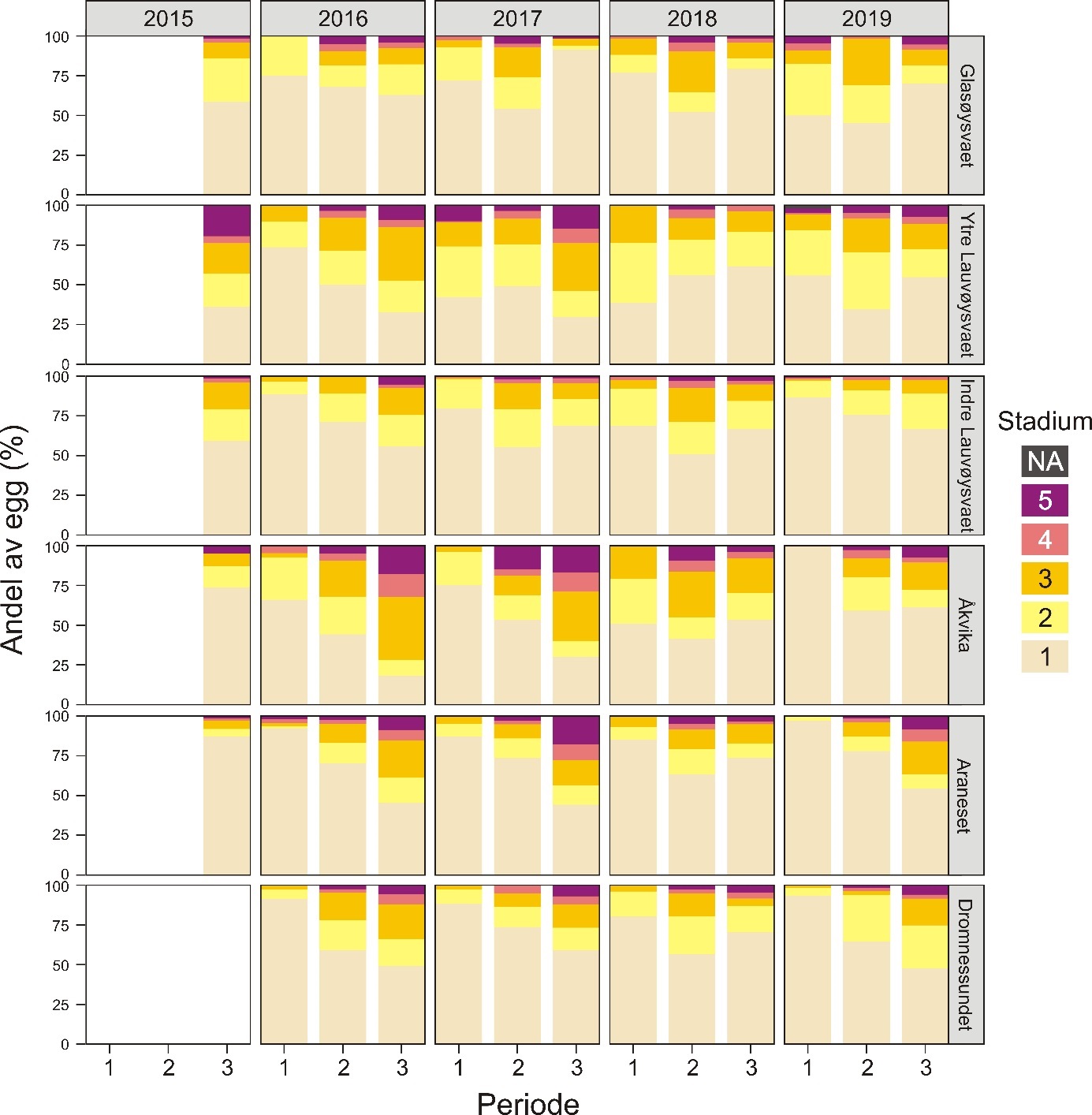
Gjennomsnittlig eggmengde på hvert av gyteområdene i de ulike årene toktene ble gjennomført er vist i figur 8. Generelt var det færre egg i de to referanseområdene Åkvika og Araneset og på ytre Lauvøysvaet enn i de andre områdene (p < 0,001). Det tredje referanseområdet i Dromnessundet hadde flere egg enn de to andre referanseområdene (p < 0,001). Ute på Smøla lå Glasøysvaet på samme nivå som Dromnessundet (p = 0,99). Indre Lauvøysvaet var statistisk sett forskjellig fra alle de andre gyteområdene og hadde de høyeste eggmengdene (p < 0,001).
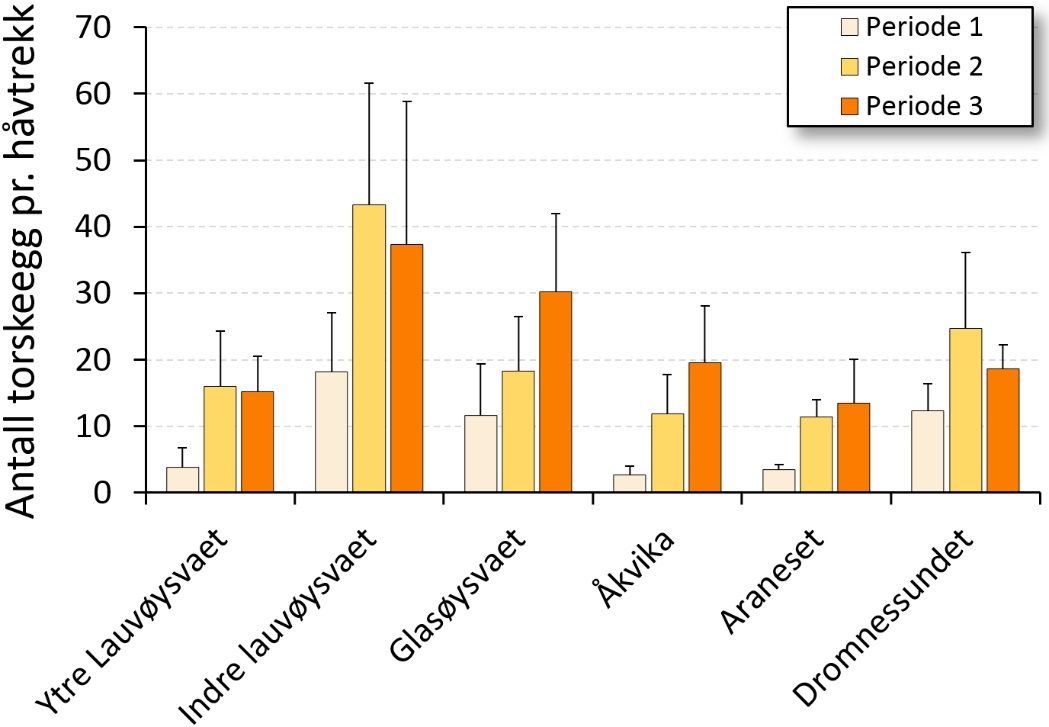
Det ble også funnet signifikant flere egg på de seine eggtoktene (p), og figur 9 viser eggmengde på toktene fra tidlig til seint i mars (periode 1, 2 og 3). I denne sammenheng er det verd å merke seg at det grunnet sein prosjektstart i 2015 kun lot seg gjøre å gjennomføre ett tokt seint i mars, og at dette til dels forklarer de høye eggverdiene i 2015 i figur 8.
Variasjoner innen områder mellom år
Det er til dels betydelige årlige variasjoner i eggmengde innen de undersøkte gyteområdene (figur 8 og 9). Med unntak av ytre Lauvøysvaet skiller de høye eggverdiene i 2015 seg ut for alle gyteområdene som ble undersøkt dette året. Imidlertid ble det som tidligere nevnt, kun gjennomført eggtokt seint i mars (periode 3) i 2015. Da var det også forventet å finne flest egg ut fra gytesyklusens forløp.
Variasjon i årlig eggmengde for de undersøkte gyteområdene på Smøla er gitt i figur 10. Her går det klart frem at indre Lauvøysvaet har flere egg, og særlig egg i stadium 1, enn ytre Lauvøysvaet. Videre er det betydelig variasjon mellom år, der 2015 og 2018 (og til dels 2019) er «gode» år for indre Lauvøysvaet, mens i 2016 og 2017 var det her lite egg.
Figur 7. Gjennomsnittlig mengde egg med diameter tilsvarende torsk på eggstasjoner i undersøkelsesområdene på Smøla (øverst) og referanseområdene (nederst) for alle toktene i perioden 2015 til 2019. Strekene angir standardavvik.
En statistisk test som tar hensyn til periode (figur 9), viste ingen forskjeller mellom år på Glasøysvaet. På indre Lauvøysvaet var det derimot mer egg i 2015 og 2018 enn i 2016 (p < 0,05), mens 2017 og 2019 var i en mellomstilling. Dette var i kontrast til ytre Lauvøysvaet der det var færre egg i 2015 enn i 2016 og 2017 (p < 0,05). Her var det også mer egg i 2017 enn i 2018 og 2019 (p < 0,05), og 2016 hadde i tillegg mer egg enn i 2019 (p < 0,05).
I referanseområdene viste den statistiske testen at på Åkvika var det mer egg i 2015 og 2016 enn i de tre årene fra 2017 til 2019 (p < 0,05). På gyteområdet ved Araneset var det mer egg i 2015 og 2016 enn i 2019 (p < 0,05), og her var 2017 og 2018 i en mellomstilling. I Dromnessundet ble det ikke gjennomført eggtokt i 2015, og her var det mer egg i 2016 og 2018 enn i 2017 og 2019 (p < 0,05).
Det nye anlegget ved Nørholmen ble montert under gytesesongen 2016, og det var i drift i tilknytning til gytesesongen 2017 og 2018. Det ble ikke funnet noen systematisk sammenheng mellom eggmengde på noen av de undersøkte gyteområdene og år det var drift på anlegget ved Nørholmen.
Figur 8. Mengde egg med diameter tilsvarende torsk fra 2015 til 2019 på eggstasjoner i undersøkelsesområdene på Smøla (Lauvøysvaet og Glasøysvaet) og i referanseområdene (Åkvika, Araneset og Dromnessundet). a) Egg i alle stadier, og b) egg i stadium 1. Eggmengde er gitt som gjennomsnitt med standardavvik for alle eggstasjoner og tokt på de ulike gyteområdene. Dromnessundet var ikke inkludert i 2015. Merk at det kun ble gjennomført ett tokt i slutten av mars i 2015 mens alle andre år er middel av tre tokt (tidlig, midt i og seint i mars).
Figur 9. Sesongvariasjon i eggmengde, gitt som mengde egg med diameter tilsvarende torsk på de ulike gyteområdene tidlig i mars (Periode 1), midt i mars (Periode 2) og seint i mars (Periode 3). Eggmengde er gitt som gjennomsnitt og standardavvik for alle eggstasjoner på de ulike gyteområdene, for alle tokt innen hver av de tre periodene.
Figur 10. Mengde egg med diameter tilsvarende torsk på eggstasjoner i undersøkelsesområdene på Smøla fra 2015 til 2019. Eggmengde er gitt som gjennomsnitt med standardavvik for tre eggtokt gjennom gytesesongen, med unntak av 2015 da kun ett tokt ble gjennomført ved prosjektstart seint i mars. Nummerering av eggstasjoner tilsvarer posisjonene i figur 4 og tabell 1.1 i Vedlegg 1. Stasjon 40 var ikke inkludert i undersøkelsene i 2015.
Eggstadier
Stadiefordeling av egg på hver av gyteområdene er vist i figur 11 for de tre toktene som ble gjennomført i en gytesesong. Med unntak av Glasøysvaet og ytre Lauvøysvaet ble det funnet signifikant økende andel av eldre egg mot slutten av mars. Videre så 2018 ut til å skille seg ut ved at det gjennomgående på alle gyteområdene var noe færre eldre egg på det siste toktet i slutten av mars. Ytre Lauvøysvaet og Åkvika hadde signifikant høyere andel av eldre egg enn alle de andre gyteområdene (p < 0,01) , mens Araneset hadde signifikant færre eldre egg sammenlignet med de andre gyteområdene (p < 0,01) , med unntak av Glasøysvaet og Dromnessundet.
Eggstørrelse
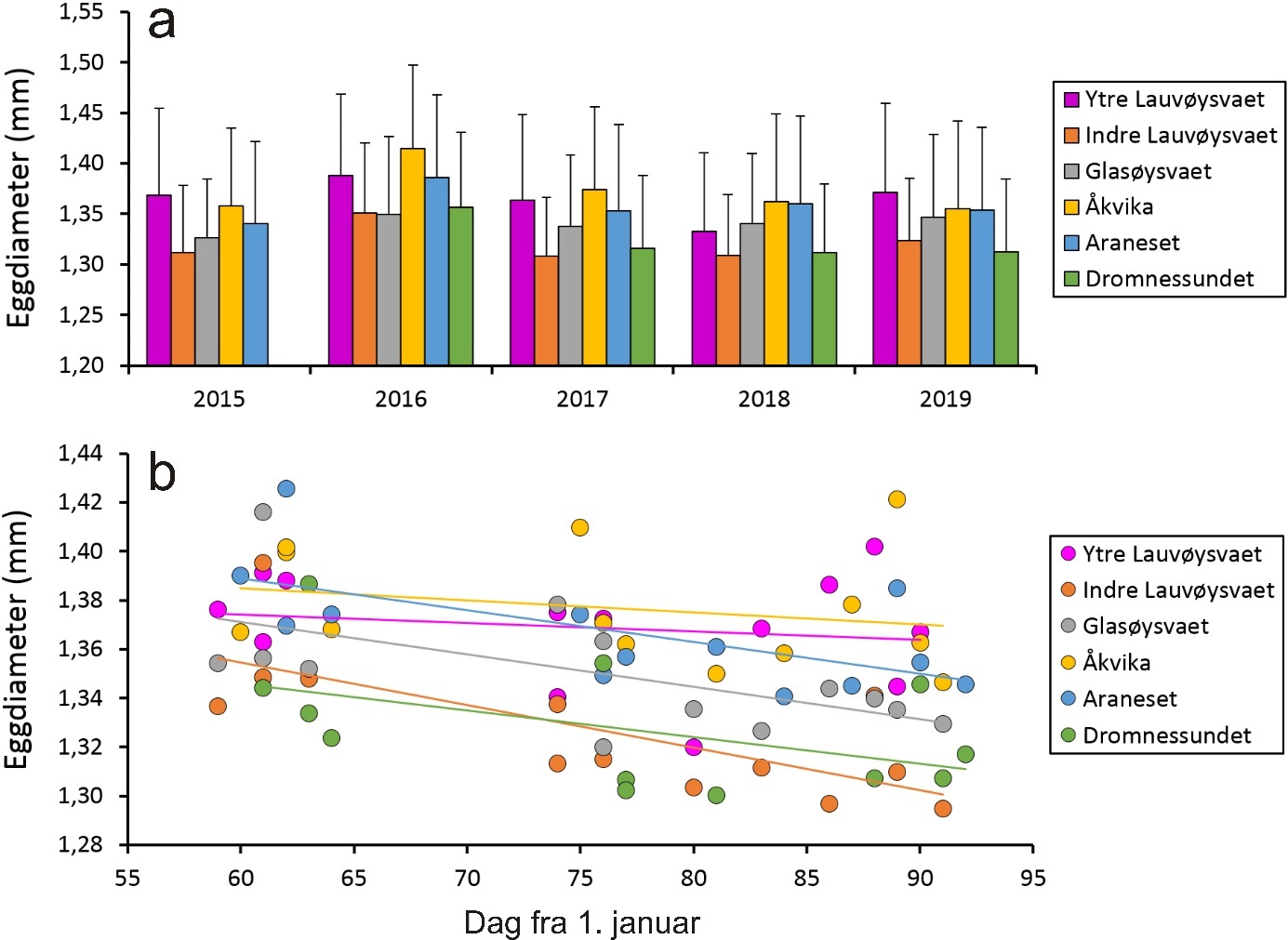
Eggdiameter varierte mellom de ulike gyteområdene, og variasjonen synes å være konsistent for noen av disse (figur 12). Det ble i denne sammenheng funnet signifikant større egg på ytre Lauvøysvaet, Åkvika og Araneset enn i de andre gyteområdene (p < 0,0001) . Glasøysvaet var i en mellomstilling med større egg enn det som ble funnet på indre Lauvøysvaet og Dromnessundet (p < 0,001) .
Videre ble det funnet at eggdiameter ble signifikant mindre med økende kalenderdag, det vil si dag fra 1. januar (p < 0,0001) . Reduksjonen i observert eggdiameter over to uker utgjorde i gjennomsnitt 0,014 mm. Forskjellen i gjennomsnittlig eggdiameter for de ulike gyteområdene innen et år varierte mellom 0,053 og 0,066 mm.
Figur 11. Fordeling av utviklingsstadium (1-5) for egg i torskestørrelse fra alle toktene der NA er ubestemmelig stadium. Periode 1 refererer til tidlig i mars (1.-6. mars), periode 2 til midt i mars (15.-23 mars) og periode 3 fra månedsskiftet mars-april (25.mars-4.april).
Figur 12. Eggstørrelse gitt som a) gjennomsnittlig diameter med standardavvik av alle håvtrekk for de ulike gyteområdene og årene de er undersøkt, og b) eggdiameter for gyteområdene som funksjon av dag i året med trendlinjer.
4.2.2 - DNA-analyser av egg
Identifisering av egg til art fra de ulike gyteområdene ble gjennomført etter en dokumentert prosedyre (Mateos-Rivera m.fl. 2020). Artsidentifisering av egg er et omfattende arbeid, og det ble tatt ut stikkprøver i 2015, 2017 og 2019 for å få et overblikk over artssammensetningen. I første og siste året ble egg på stasjoner fra alle gyteområdene undersøkt, og det var særlig forekomst av torsk og hyse som er interessant da disse overlapper betydelig i eggstørrelse. Andre arter enn torsk og hyse identifisert ved DNA-analysene var brisling, øyepål (Trisopterus esmarkii), sei (Pollachius virens), lange (Molva molva), rødspette, sandflyndre, skrubbe (Platichtys flesus) og gapeflyndre.
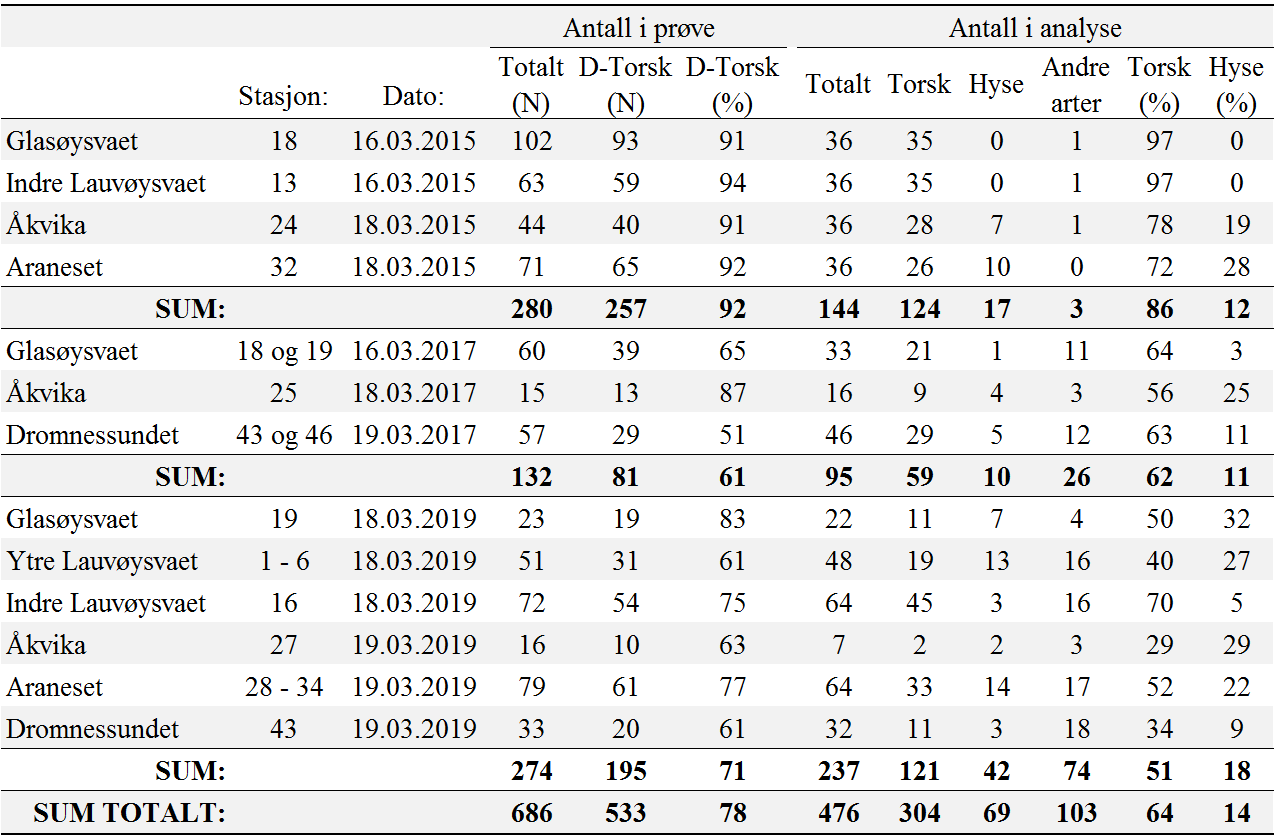
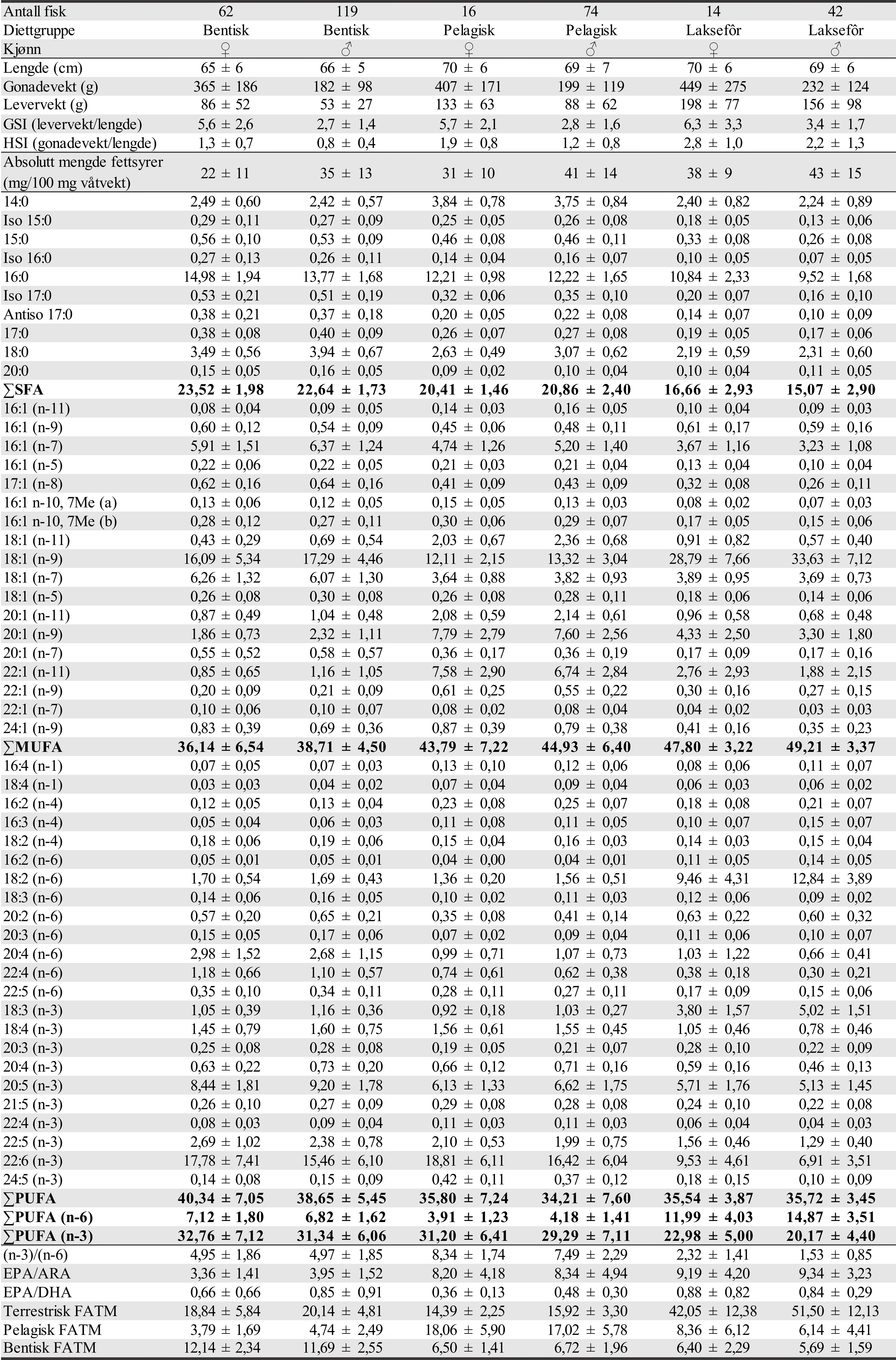
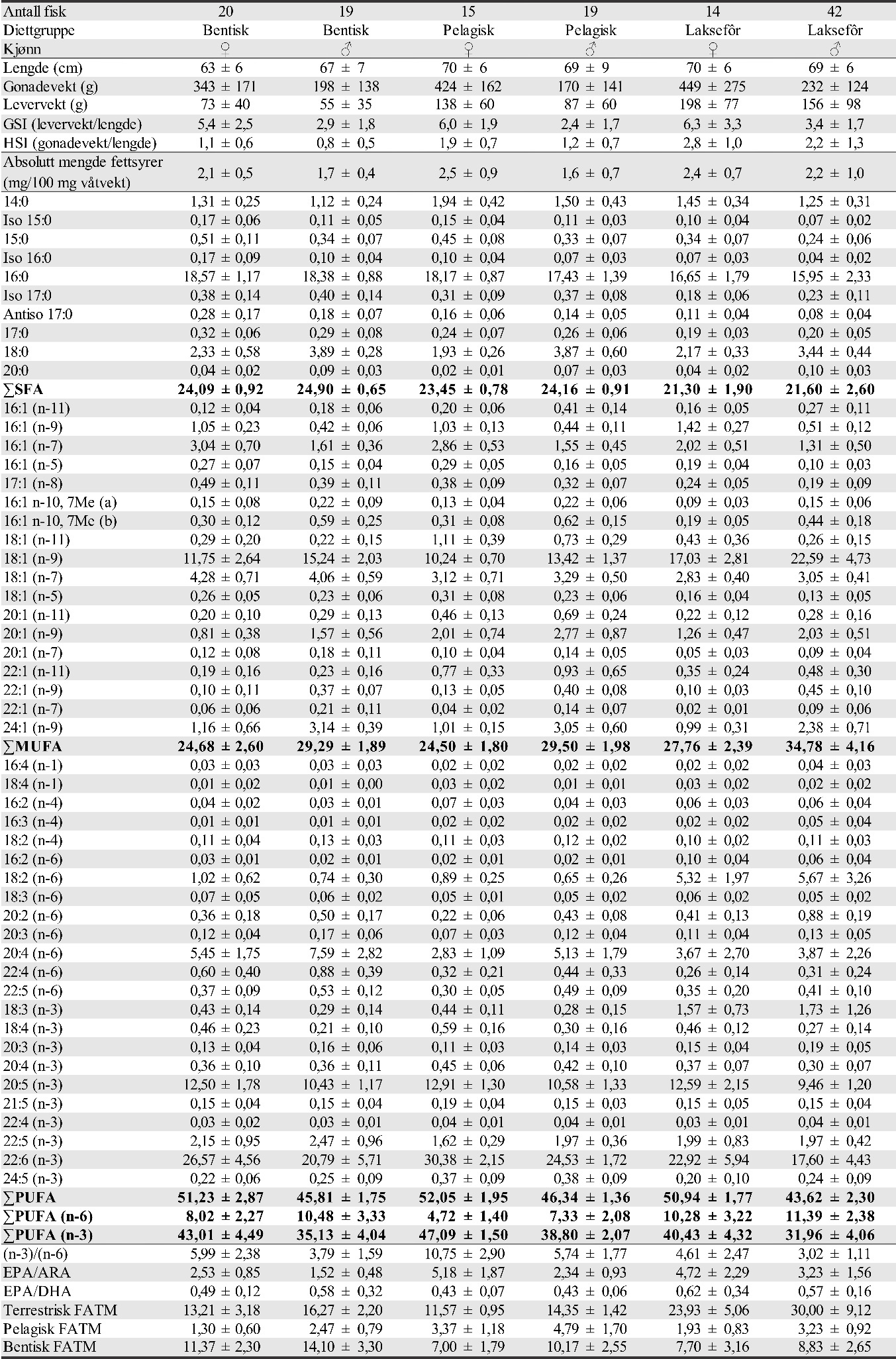
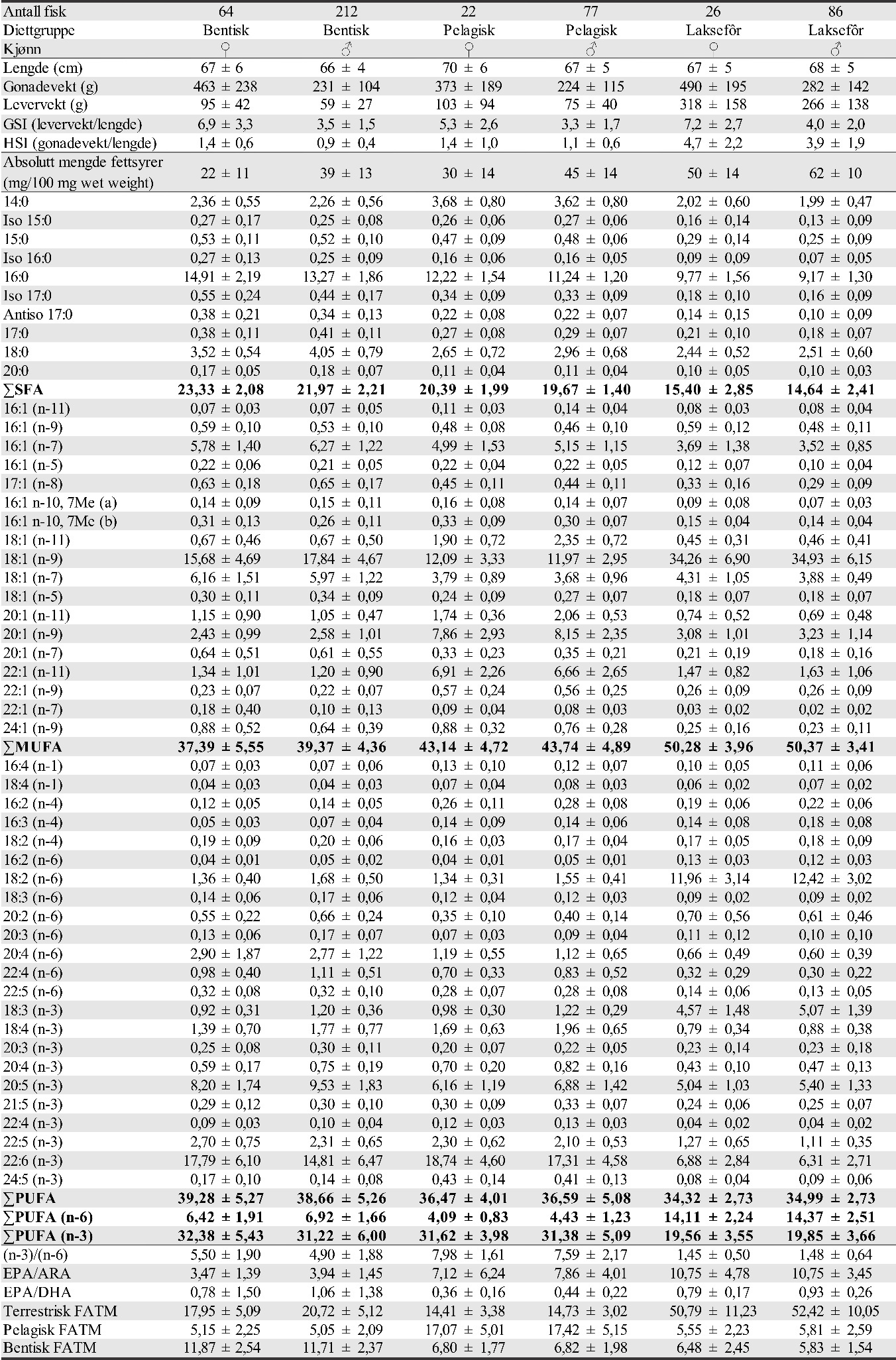
I 2015 ble det funnet høy andel (97 %) av torsk på Glasøysvaet og indre Lauvøysvaet, mens hyse utgjorde 10 og 28 % på henholdsvis Åkvika og Araneset (tabell 1). I 2017 og 2019 var det en noe lavere andel av torsk på Glasøysvaet og Lauvøysvaet (40-70 %), mens andelen av andre arter var høyere. Hyse varierte mellom 3 og 32 % i eggprøvene disse årene. Generelt for de ulike gyteområdene dominerte torskeegg på indre Lauvøysvaet og Glasøysvaet med henholdsvis 77 og 80 % av eggene, mens på ytre Lauvøysvaet, Åkvika, Araneset og Dromnessundet var andelen torskeegg mellom 40 og 66 %. Tilsvarende for hyseegg var 3 % på indre Lauvøysvaet, 27 % på ytre Lauvøysvaet, 18 % på Glasøysvaet, 22 % på Åkvika, 24 % på Araneset, og 10 % i Dromnessundet. På ytre Lauvøysvaet og i Dromnessundet og var også innslaget av andre arter betydelig (henholdsvis 33 og 38 %). For alle DNA-prøvene sett under ett var andelen torskeegg 64 %, hyseegg 14 % og andre arter 22 % (tabell 1).
Tabell 1. Oversikt over DNA-analyser av egg. D-Torsk er egg med diameter 1,2-1,6 mm.
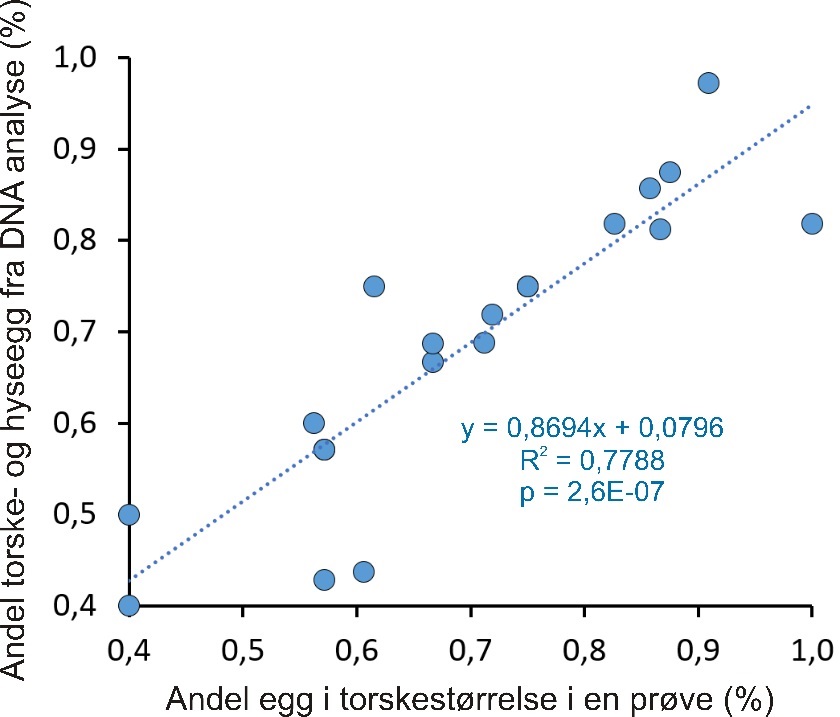
Siden resultatene i denne rapporten presenterer egg med størrelse fra 1,2 til 1,6 mm som «mulige torskeegg», men hvor hyseegg også overlapper i stor grad, er det av interesse å se hvordan egg i denne størrelsesgruppen samsvarer med resultatene fra DNA-analysene. Fordi muligheten for at andelen av torsk og hyse i en prøve kan ha større feil hvis kun en del av eggprøven er analysert, ble denne sammenligningen gjort for prøver der 80 % eller mer av eggene i prøven ble analysert med hensyn til identifikasjon av arter ved hjelp av DNA. Resultatet viser en sterk sammenheng mellom egg observert i dette størrelsesområdet og forekomst av hyse- og torskeegg fra DNA-analysene (figur 13).
Når det gjelder eggstørrelse og identifisering av egg til arter, ble det funnet noen avvik i Dromnessundet. For eksempel ble det 19. mars 2017 funnet at kun 6 egg på stasjon 43 var innenfor størrelsesområdet til torsk og hyse, mens DNA-analysen det identifiserte hele 11 torskeegg og ingen hyseegg i denne prøven. Torskeegg må derfor ha vært utenfor størrelsesområdet 1,2-1,6 mm, og trolig ned til 1,18 mm da flere egg i denne størrelsen ble funnet i prøven. Videre ble det 19. mars 2019 funnet at 20 egg var innenfor dette størrelsesområdet på stasjon 43 i Dromnessundet. DNA-analysen viste kun 11 torskeegg og 4 hyseegg, og følgelig overlappet egg fra andre arter som ikke lot seg identifisere fra eggbildene, med egg i størrelsesområdet 1,2-1,6 mm. Mest sannsynlig gjelder dette øyepål som det var 5 egg av i denne prøven, og som er oppgitt å ha egg med diameter opp til 1,19 mm (Munk og Nielsen, 2005).
Figur 13. Sammenheng mellom andel egg i en prøve i torskestørrelse og andel hyse- og torskeegg i prøvene fra DNA-analysene, for alle prøver der mer enn 80 % av eggene ble analysert og bestemt til art. En p-verdi på mindre enn 0,05 angir statistisk signifikans.
4.2.3 - Resultater modellering av eggdrift
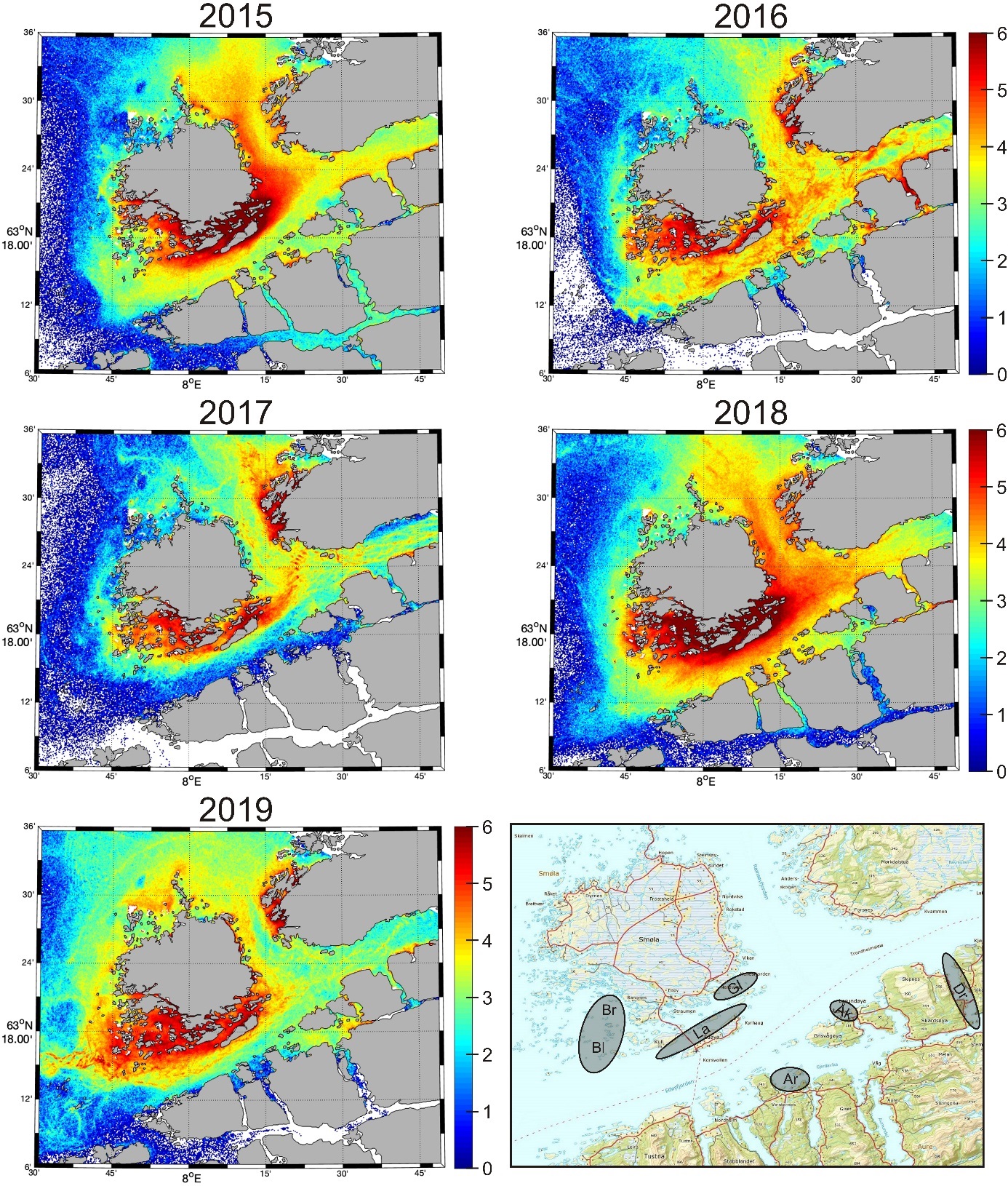
Transport fra hovedgytefeltet på Lauvøysvaet for alle årene fra 2015 til 2019 er vist i figur 14. Det er stor mellomårlig variasjon i transport og spredning av gyteprodukter fra Lauvøysvaet. I enkelte år (2015, 2018) blir en stor del av partiklene værende igjen i nærheten av gyteområdene på sørsiden av Smøla. De partiklene som ikke blir værende igjen i området, blir transportert videre med kyststrømmen hovedsakelig på nordsiden av Frøya.
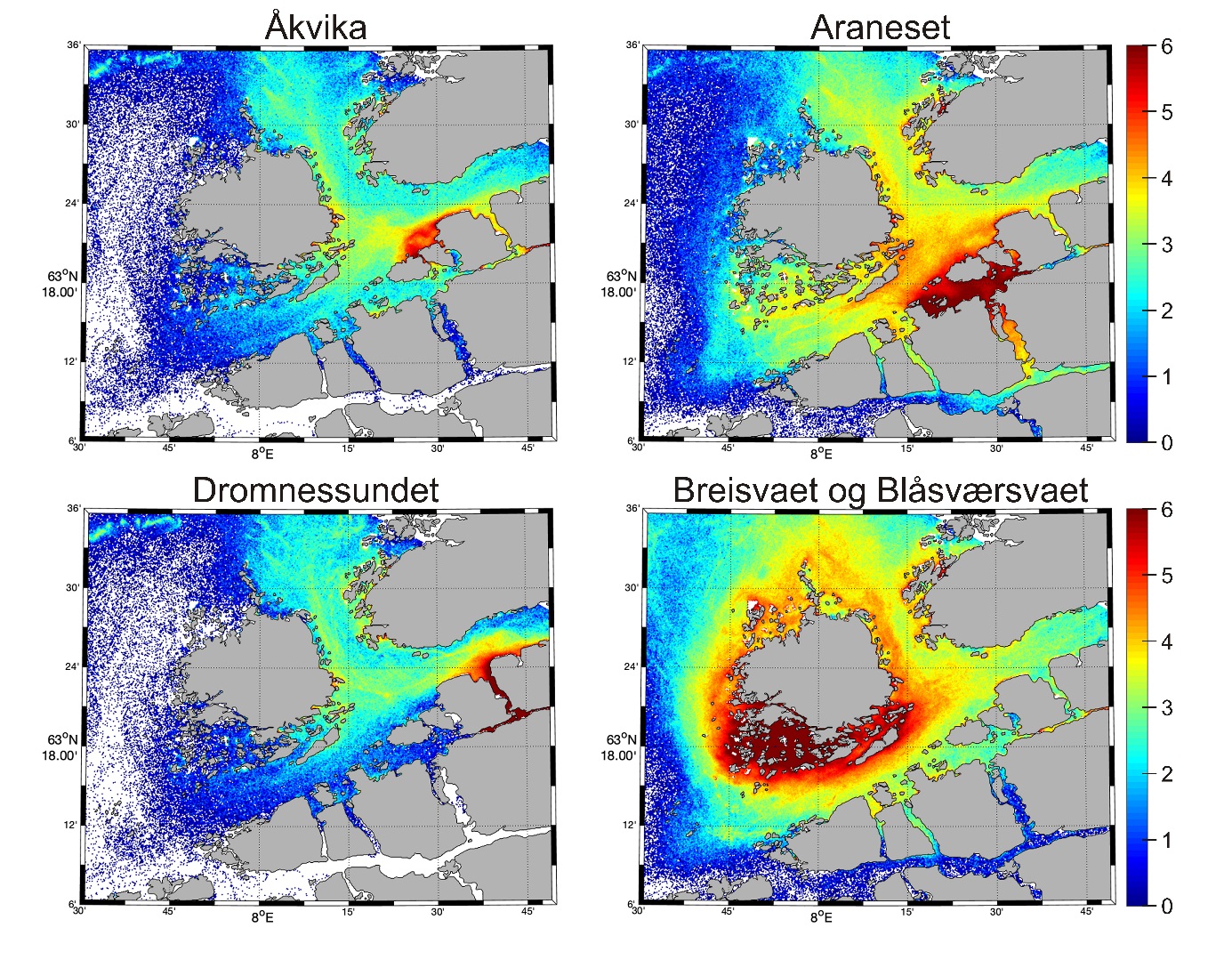
Figur 15 viser spredning av gyteprodukter fra referanseområdene a) Åkvika, b) Araneset og c) Dromnessundet i 2018, i tillegg til de nærliggende gyteplassene d) Breisvaet og Blåsværsvaet. Dromnessundet ligger veldig skjermet til og har derfor liten utveksling med andre områdene. Gytefeltet i Åkvika ligger ganske åpent og partiklene spres derfor raskt utover et større område og i lave konsentrasjoner.
Partikler sluppet ut fra Araneset blir stort sett værende i kystsonen med litt utveksling på tvers av Edøyfjorden over til Lauvøysvaet. Breisvaet og Blåsværsvaet ligger mye mer eksponert, og partiklene spres derfor rundt hele Smøla, samtidig som det er ganske høy oppholdstid tett på kysten (figur 15).
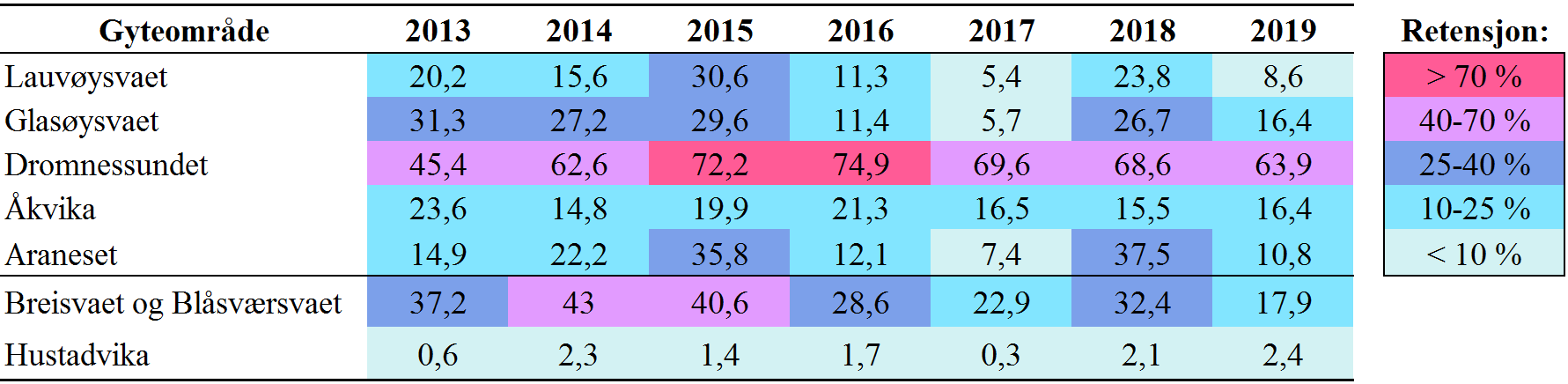
Retensjon er en måte å kvantifisere oppholdstid på, her er retensjon definert som prosentandel av partiklene som oppholder seg innenfor 5 km fra utslippspunktet 5 dager etter utslippet. Definisjonen av retensjon kan variere mye, og det er valgt en relativt liten avstand og en kort periode fordi Smøla ligger i et veldig dynamisk område sterkt påvirket av kyststrømmen. Men hvis man skulle brukt samme definisjon som inne i fjordene, så ville hele dette området kommet ut med lav retensjon (Myksvoll m.fl. 2014). Resultatene er vist i Tabell 2 for alle gyteområdene i årene 2013-2019.
Figur 14. Døgnmidlet konsentrasjon av gyteprodukter i april i simuleringen av eggdrift i april fra Lauvøysvaet for årene 2015-2019. Skalaen angir partikkeltetthet og er logaritmisk: ln (antall partikler/m2). Kartet nederst til høyre viser områdene hvor partiklene er sluppet ut, både for Lauvøysvaet (La) og for simuleringene foretatt i figur 15 og tabell 2. Dette omfatter Breisvaet (Br), Blåsværsvaet (Bl), Glasøysvaet (Gl), Åkvika (Åk), Araneset (Ar) og Dromnessundet (Dr).
Figur 15. Døgnmidlet konsentrasjon av gyteprodukter i simuleringen av eggdrift i april 2018 fra referanseområdene Åkvika, Araneset og Dromnessundet, samt Breisvaet og Blåsværsvaet som er nærliggende oppstrøms gyteområder til gyteområdene Lauvøysvaet og Glasøysvaet på Smøla. Skala til høyre angir partikkeltetthet i simuleringen og er logaritmisk: ln (antall partikler/m2). Kart i figur 1 viser områdene partiklene er sluppet ut.
Tabell 2. Mellomårlig variasjon i retensjon fra simulert utslipp av partikler i gyteområdene, beregnet som andel partikler (%) som befinner seg innenfor 5 km fra utslippspunktet etter 5 dager. Farge viser styrken på retensjonen, og grenseverdiene er forklart i panelet til høyre. Hustadvika omfatter et 18,9 km2 stort område like nordvest for Farstad.
Dromnessundet er det gyteområdet som skiller seg ut med stabilt høy retensjon nesten alle år, kun med unntak av 2013. Lauvøysvaet og Glasøysvaet variere mye og stort sett i takt, med høyest retensjon (30-31 %) i 2015 og lavest (5-6 %) i 2017. Åkvika varierer minst mellom år og ligger på moderat til lav retensjon mellom 15-24 %. Araneset har høy mellomårlig variasjon og har høyest retensjon (38 %) i 2018. Når man vurdere alle gyteområdene samlet så er det lavest retensjon i 2017 og 2019, mens 2015 og 2018 har høyest retensjon.
4.3 - Diskusjon eggundersøkelser
Analysene av DNA på eggene ble først og fremst utført for å identifisere arter. En del arter var lett å skille fra torskeegg ut fra utseende og eggstørrelse, mens andelen av hyseegg var ikke mulig å bedømme uten DNA-analyse da hyseegg overlapper fullstendig med torsk i størrelse og kun er mulig å skille fra torsk i seine eggstadier ut fra utseende. «Egg i torskestørrelse» (1,2 – 1,6 mm) vil derfor omfatte både torsk og hyse. De genetiske analysene viste at det var flest torskeegg (64 %) mens hyse (14 %) utgjorde gjennomsnittlig litt mindre enn en femtedel av alle torske- og hyseegg i prøvene. Den visuelle bedømmelsen av mengden torske- og hyseegg ut fra eggstørrelse viste også god sammenheng med mengden egg av disse to artene bestemt fra DNA. I studien var derfor «egg i torskestørrelse» et brukbart mål på mengde torskeegg med de angitte forholdstall for torsk og hyse som grunnlag. I ett tilfelle ble det funnet at torskeegg i Dromnessundet var mindre enn 1,2 mm, og i ett annet tilfelle her ble det funnet at egg av øyepål må ha vært større enn 1,2 mm. For øyepål er dette større eggdiameter enn det som er angitt i litteraturen, mens torskeegg er oppgitt å kunne være ned mot 1,16 mm (Munk & Nielsen 2005). At slike avvik kun forekom i to av de analyserte prøvene underbygger at den nedre grensen på 1,2 mm er fornuftig. Utvidelse av grensen nedover vil innebære mulighet for at flere egg fra andre arter vil inngå i «egg i torskestørrelse».
Variasjon i eggmengde gjennom undersøkelsesperioden og alder på eggene viste at eggtoktene ble gjennomført i første halvdel av gytesyklusen, og at gytetoppen var som ventet i siste halvdel og mot slutten av mars. Med hensyn til variasjon i eggmengde innenfor hvert gyteområde viste Glasøysvaet en tendens til å ha flest egg og høyest andel av egg i stadium 1 innerst på stasjon 18. Dette antyder at gyting foregår i den indre delen av Glasøysvaet. Egg kan imidlertid vare opp til 3 dager i stadium 1 ved 5 til 6°C, og det vil derfor være noe usikkerhet knyttet til eksakt stedfesting av gyteplass ut fra egg i stadium 1 generelt. Området som omfatter indre Lauvøysvaet, hadde gjennomgående flere egg og til dels høyere andel av egg i stadium 1 enn ytre Lauvøysvaet. Dette vises blant annet tydelig i data fra 2019 (Vedlegg 2, figur 2.1 og 2.3) og kan tolkes som at gyting forgår på indre Lauvøysvaet. Samtidig er det sannsynlig at indre Lauvøysvaet har høyere retensjon av egg, mens ytre Lauvøysvaet er mye mer eksponert for den fremherskende strømretningen fra sydvest som modellsimuleringen av vannbevegelse indikerer. Dette betyr ikke at gyting ikke kan foregå på ytre Lauvøysvaet, men at egg gytt her ute vil ha mye større sannsynlighet for å transporteres vekk fra gyteområdet relativt raskt.
Egg tidlig i stadium 1 (opp til og med 8-celler) vil være gytt samme dag som de ble samlet inn. Her var i gjennomsnitt kun 0,8 % av eggene på ytre Lauvøysvaet i denne kategorien, mens tilsvarende andel på indre Lauvøysvaet var 4,7 %. Glasøysvaet hadde også en lav andel av egg tidlig i stadium 1 (0,4 %). Dette viser at Glasøysvaet og ytre Lauvøysvaet trolig har mer dynamiske vannmasser med aktiv transport av egg både til og fra disse områdene sammenlignet med indre Lauvøysvaet. Egg transportert til disse områdene vil da i større grad være eldre og ikke nygytte egg. I tillegg er det sannsynlig at gyting med rimelig stor sikkerhet foregår på indre Lauvøysvaet.
I referanseområdet Åkvika var mengden egg i stadium 1 på nivå med ytre Lauvøysvaet, men andelen av egg tidlig i stadium 1 (5,2 %) var høy og sammenlignbar med indre Lauvøysvaet. Dette viser at gyting skjer på Åkvika i større grad enn ytre Lauvøysvaet, men at den lave andelen av egg i hele stadium 1 tyder på betydelig transport av egg inn og ut av dette området. I referanseområdet Araneset kan det virke som at gyting foregår hovedsakelig på et lite område ved stasjon 31. Likevel var andelen av egg tidlig i stadium 1 ved Araneset kun 1,7 %, noe som indikerer en viss dynamikk i vannmassene også i dette området. I referanseområdet Dromnessundet var det ingen stasjoner som hadde vesentlig flere egg enn de andre stasjonene, men resultatene tyder på at gytingen foregår flere steder i den sentrale delen av sundet. Modellsimuleringen av eggtransport viste også at Dromnessundet hadde høy grad av retensjon, og følgelig var høy andel av egg i stadium 1 i den sentrale delen av sundet ventet siden det foregikk vedvarende gyting her. Andelen egg tidlig i stadium 1 (3,8 %) støtter dette.
Indre Lauvøysvaet var området med mest egg, med Glasøysvaet og Dromnessundet i en mellomstilling sammenlignet med Åkvika, Araneset og Ytre Lauvøysvaet. Det ble funnet betydelige variasjoner mellom år, og særlig på indre Lauvøysvaet. Det var imidlertid ikke systematiske endringer her som kan knyttes opp til perioden med fisk i det nye anlegget på Nørholmen (2017-2018). I referanseområdene ble det også funnet mellomårlige variasjoner, og tidlig i undersøkelsene (2015-2016) var det gjennomgående mer egg her enn de siste årene av undersøkelsen. Slike mellomårlige variasjoner kan skyldes både at strømforhold som transporterer egg og gytebiomassen er forskjellig fra år til år.
Kystområdet ved Smøla er sterkt påvirket av kyststrømmen som deler seg i to grener og passerer på både på nord- og sørsiden av øyen. Potensialet for spredning av pelagiske fiskeegg som hos torsk og hyse, er derfor betydelig. Samtidig kan lokale forhold gjøre at oppholdstiden øker, spesielt økt virveldannelse i trange sund og mellom små øyer som for eksempel på indre Lauvøysvaet innenfor Edøya. De konsistente forskjellene mellom gyteområdene vil være en funksjon av hydrodynamisk transport, retensjon og biomasse av gytefisk som bidrar til gytingen. Med unntak av Dromnessundet så viser modellsimuleringene at ingen av de undersøkte gyteområdene har spesielt høy retensjon. Den sterke dynamikken i strømforholdene i Edøyfjorden kan derfor bidra til fortynning av eggmengdene på Glasøysvaet, ytre Lauvøysvaet, Åkvika og Araneset. Høy retensjon i Dromnessundet betyr imidlertid ikke at dette området er isolert, og i 2016 viser simuleringene at er det transport fra både Lauvøysvaet og Glasøysvaet inn til Dromnessundet. Et annet aspekt er at eggdiameter er gjennomgående større på ytre Lauvøysvaet, Åkvika og Araneset. Eggdiameter forventes å minke gjennom gytesesongen for det enkelte individ, og derved for hele gytepopulasjonen utover i gytesesongen. I tillegg vil eldre fisk ha egg med større eggdiameter enn yngre fisk. Forekomst av eldre fisk på gyteområdene og transport av egg fra motstrøms gyteområder med eldre fisk er de faktorene som best kan forklare de observerte forskjellene i eggdiameter mellom gyteområder. I undersøkelsesområdene er den mellomårlige variasjonen i retensjon betydelig, hovedsakelig på grunn av forskjeller i vinden og kyststrømmen mellom år (Myksvoll m.fl. 2020).
Det er også viktig å merke seg at gyteområdene lengre vest på Smøla, på Breisvaet og Blåsværsvaet, ifølge simuleringene har lokalt høy retensjon (tabell 1) i tillegg til vesentlig transport av egg til Lauvøysvaet. Breisvaet og Blåsværsvaet har ikke vært et fokusområde for eggundersøkelsene i ICOD-prosjektet, selv om det er rimelig å anta at gyting her vil bidra betydelig til rekrutteringen i studieområdet. Samtidig kan resultatene fra simuleringene også tyde på at egg som kommer lengre sørfra med kyststrømmen, ender opp på Smøla. Dette innebærer at farvannene rundt Smøla trolig er viktige oppvekstområder for yngel fra en større kystregion, inkludert yngel fra de lokale gyteområdene når forholdene ligger til rette for dette. Den hydrodynamiske situasjonen i studieområdet med muligheter for transport av egg over større områder i løpet av kort tid, gjør at eventuelle sammenhenger mellom lokalisering av oppdrettsanlegget på Nørholmen og eggproduksjon på de nærliggende gytefeltene ikke er mulig å påvise.
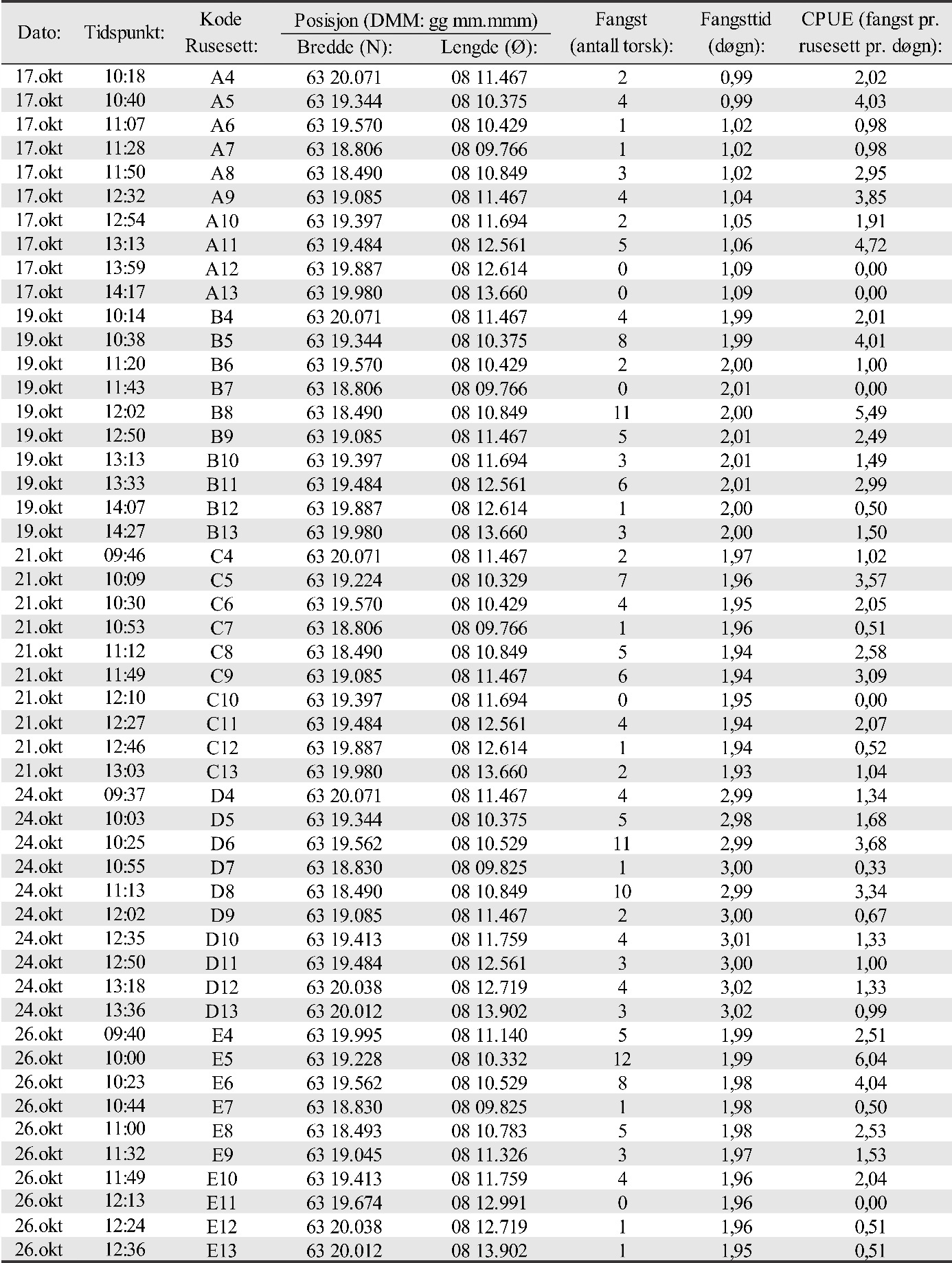
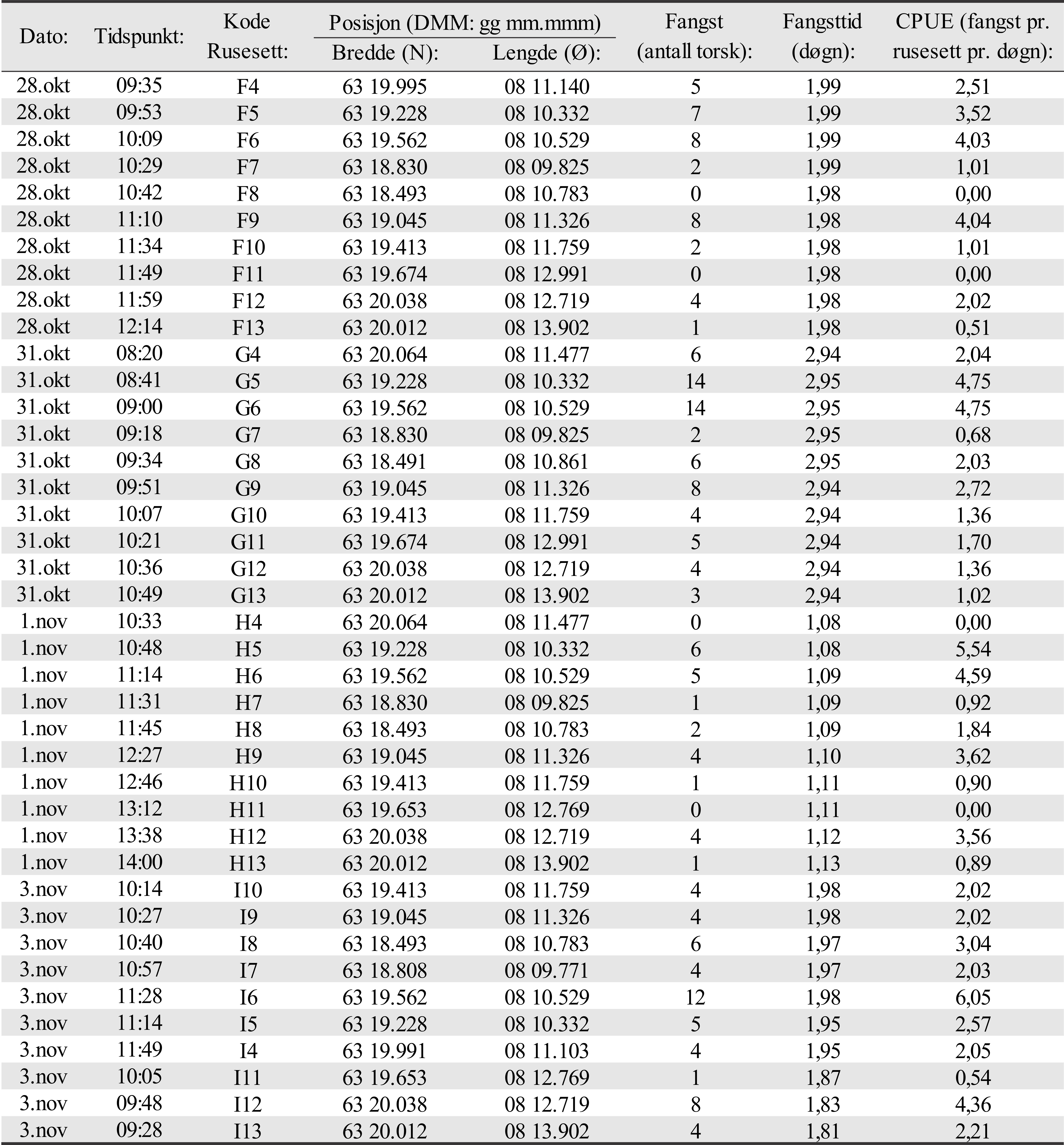
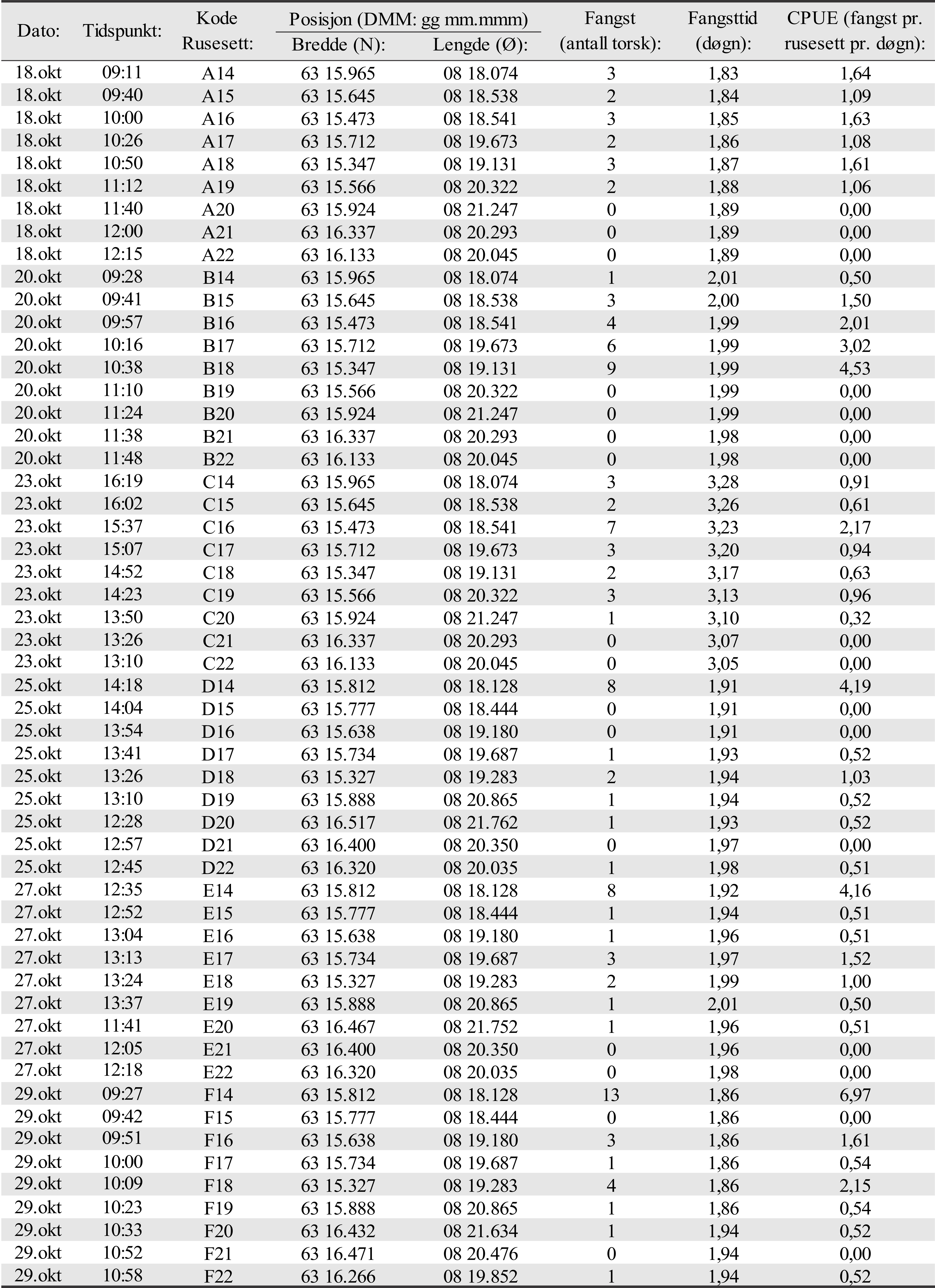
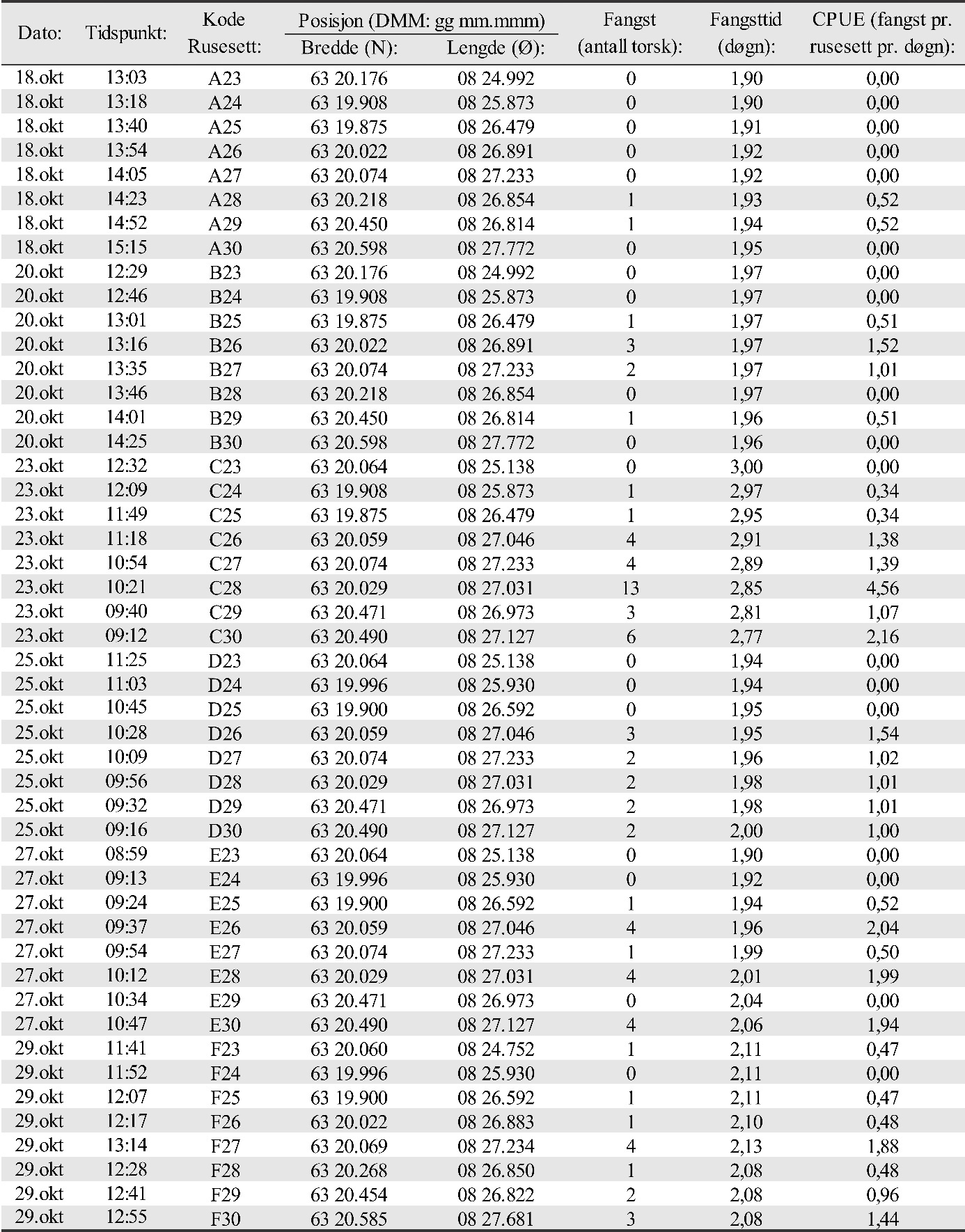
5 - Yngelundersøkelser
I perioden fra midten av oktober til første uken i november ble det fra 2015 til 2019 gjennomført undersøkelser av oppvekstområder for torsk i tilknytning til alle gyteområdene beskrevet foran i kapitlet med eggundersøkelser. Det ble foretatt et fiske med danske åleruser på grunt vann for å fange torsk i alle størrelser fra ca. 10 cm lengde og oppover, noe som innebærer fangst av alle årsklasser helt ned til 0-gruppen (årsyngel). I tillegg ble det i 2019 tatt opp videoer av bunnen for å undersøke bunntyper på fangstposisjonene. Undersøkelsene ble gjennomført i samarbeid med Mowi AS som leide inn egnede båter og mannskap med utgangspunkt i basen på Vikan på Smøla.
5.1 - Metoder
5.1.1 - Innsamling av yngel
Rusefisket ble gjennomført i gyteområdene som ble kartlagt i mars 2015. Dromnessundet ble inkludert i 2016. Erfaringsmessig vil det finnes nærliggende oppvekstområder for torskeyngel og umoden torsk på grunt vann i tilknytning til gyteområder. Dette ble også verifisert i 2015 og 2016 for Dromnessundet (van der Meeren 2016; 2017). Gode oppvekstområder er lokaliteter med tare eller ålegras på grunt vann. I slike områder vil yngel og umoden torsk finne beskyttelse og god tilgang på mat. Også enkelte større individer av torsk vil kunne finnes her. Det kan også finnes torskeyngel på stein, grus og sandbunn til dels bevokst med rødalger, for eksempel av typene smalving (Membranoptera alata) eller smalblekke (Phyllophora crispa). Denne bunntypen er vanlig på grunt vann i Dromnessundet. Posisjoner for ålerusene ble valgt på grunnlag av at bunnen var mørk i størst mulig grad, og omfattet blant annet ulike typer tareskog og ålegraseng.
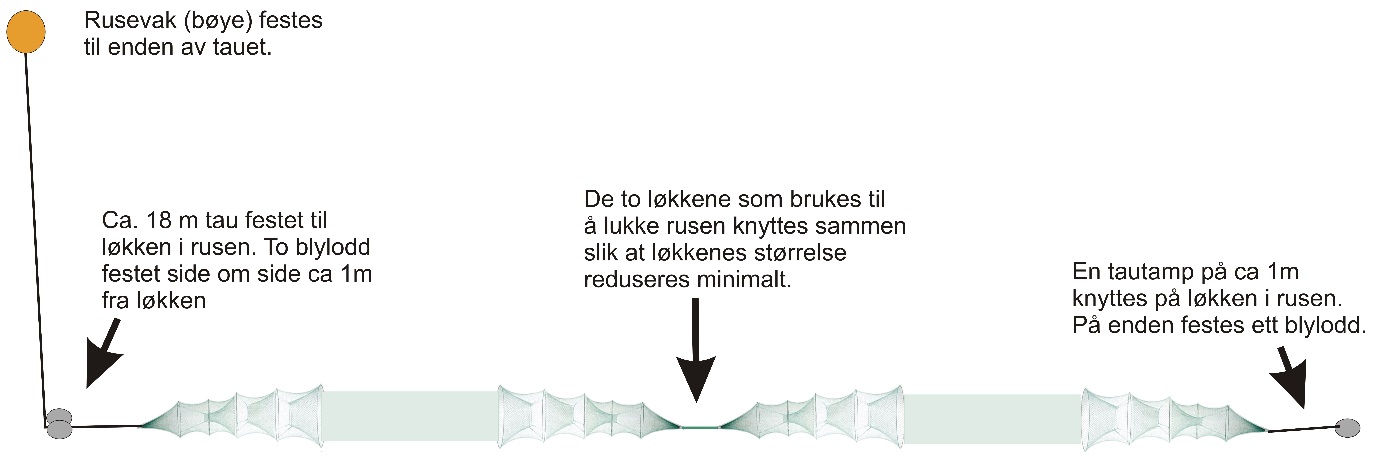
Figur 16. Montering av et ruseoppsett, som i rapporten betegnes som en ruse (van der Meeren 2015).
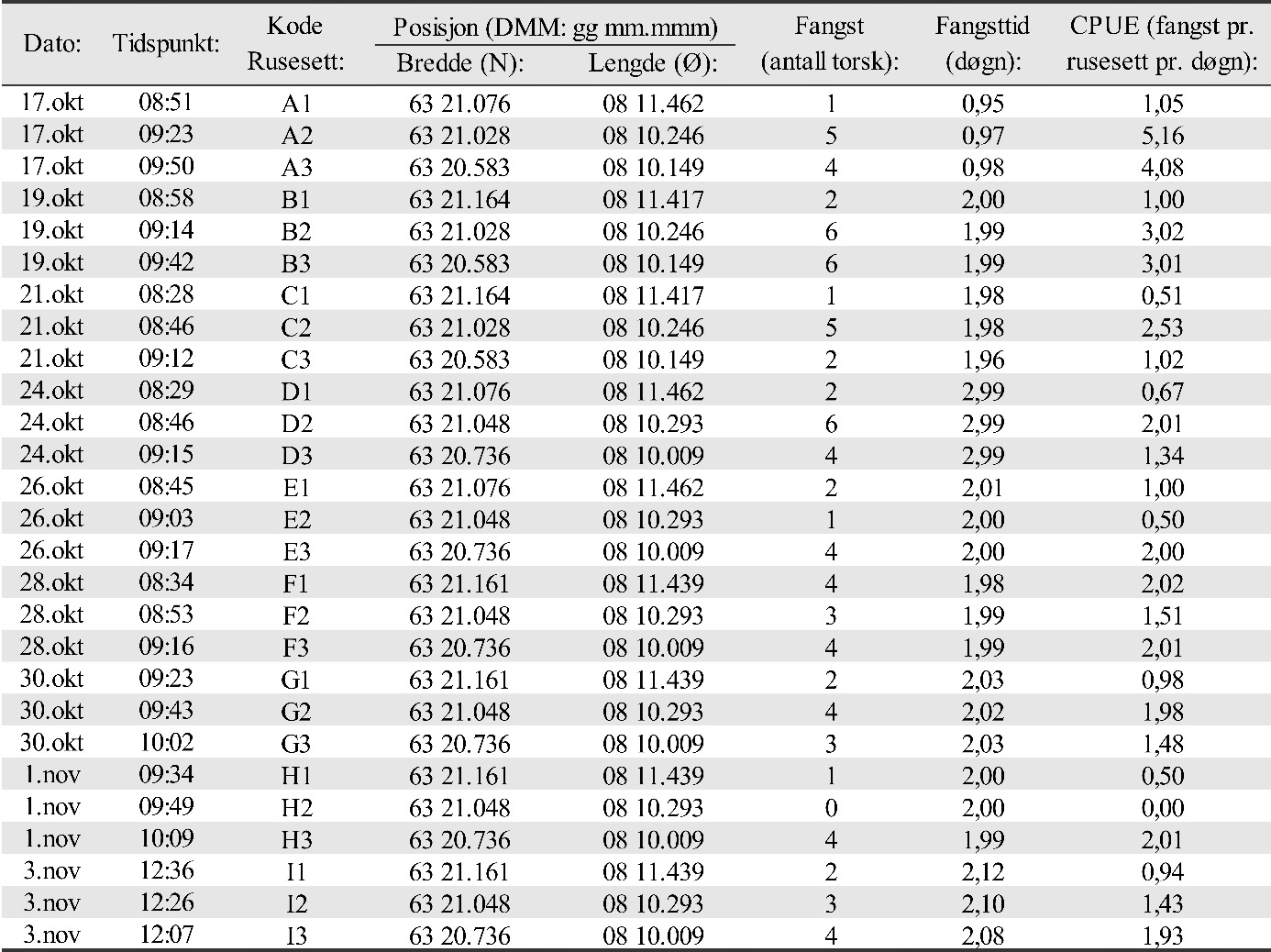
Det ble benyttet danske åleruser av type 79/5 dobbel, med grønn knuteløs not og 6 m ledegarn festet til et bur i hver ende. Hvert bur hadde 5 ringer i 4 mm rustfritt stål med 79 masker på første ring som hadde en diameter på 52,5 cm. Videre hadde burene tre rom med en kalv til hvert av disse. Maskeviddene var 11 mm i bakrommet, 14 mm i mellomrommet, 17 mm i framrommet og 18 mm i ledegarnet. To åleruser ble knyttet sammen i lenke og er i denne rapporten betegnet som en «ruse» (figur 16). Et utsett av en slik ruse for et avgrenset tidsrom med påfølgende opphal og røkting benevnes et «rusesett». Fisket foregikk med i alt 30 slike ruser. Det ble benyttet ett stk. 2 kg blylodd på den ene enden av rusen, og to stk. 2 kg blylodd med 18 m teinetau til rusevak (flytebøye) i den andre enden. Like under hvert rusevak ble det festet en laminert papirlapp med adressen til Havforskningsinstituttet, prosjekt-informasjon, kontaktinformasjon med telefon-nummer, og referanse til tillatelser fra Fiskeridirektoratet til å fiske torsk under minstemål (ref. nr. 14/14305, 16/2748 og 18/9815). Før fisket startet ble det gitt beskjed med kopi av tillatelser på e-post til lensmannskontoret på Smøla og i Aure kommune, samt Statens naturoppsyn. Tillatelser til å ta prøver ved finneklipping var også innhentet på forhånd fra Mattilsynet (ref. 18/143861 og søknad FOTS: id. nr. 16488).
Mowi AS stilte med en Targa 27 innleid fra Norsk Sjøservice AS som følgebåt, mens selve fisket foregikk fra en åpen Øien 530 eller 620 med Honda 30 eller 50 Hk utenbordsmotor (figur 17). Mowi AS stilte med fører til begge båtene. Det ble montert en medbrakt Garmin echomap 52DV kartplotter med et Garmin GT21-TM-ekkolodd på Øien-båten. Dette var nødvendig for sikker navigering, god registrering av posisjon samt å lokalisere gode områder for å sette ut ruser på grunt vann (1-10 m dyp) hvor fisket foregikk. De samme ruseposisjonene fra tidligere års undersøkelser ble benyttet, og rekkefølge og antall rusesett på en ruseposisjon ble justert etter værforholdene. Foruten posisjon og tidsrom hvert enkelt rusesett stod i sjøen, ble biologiske data samlet inn fra all torsk som ble fanget i rusene (Figur 18). I tillegg ble antall av andre fiskearter fanget i rusene registrert. For torsk ble det registrert både lengde og vekt. Vekten ble målt med en nøyaktighet på 10 gram ved hjelp av en håndholdt Rapala 0-8 kg fiskevekt. Torsken ble lagt levende i en bøtte som ble tarert før veiing. Veiingen var imidlertid sensitiv for vind og bølger (spesielt for små individer) og bevegelser hos fisken (for større individer). For de minste fiskene (< 60 g) lot vekten seg vanskelig bestemme sikkert.
Figur 17. Båtene som ble benyttet til yngelundersøkelsene: Targa 27 følgebåt til venstre og arbeidsbåt Øien 580 med 50 Hk Honda utenbordsmotor til høyre (van der Meeren 2019).
I tillegg ble det tatt en vevsprøve (ved finneklipp) fra fremre tupp av bakre gattfinne av rundt 40 torsk fra hvert undersøkelsesområdene. Denne prøven ble konservert i absolutt etanol for senere DNA-analyser. For at fisket ikke skulle påvirke bestandene av torsk lokalt i undersøkelsesområdene, ble innfanget torsk sluppet levende ut i sjøen umiddelbart etter innsamling av biologiske data.
Det ble fisket i de samme områdene hvert år (van der Meeren 2016; 2017; 2018; 2019). Stasjonsnettet for rusene ble valgt med tanke på å dekke det meste av de grunne farvannene rundt gyteområdene som ble verifisert i 2015 og 2016. Det ble benyttet de samme posisjonene for rusene alle årene undersøkelsene foregikk, og der hver ruse hadde minst to posisjoner avhengig av vindretning og bølger. Antall rusesett på en posisjon ble derfor justert etter vindretning og vindstyrke og er vist for 2019 i Vedlegg 3 (figur 3.1, 3.2, 3.3 og tabell 3.1, 3.2a, 3.2b, 3.3, 3.4, 3.5). Grunnet ulike værforhold var det ikke mulig å få rusesettene helt identisk alle årene. Noen posisjoner ble derfor benyttet flere eller færre ganger de ulike årene undersøkelsen varte. Rusene ble typisk satt på 2 til 8 m dyp, og sjekket hovedsakelig annet hvert døgn så langt det lot seg gjøre ut fra været. På det meste ble ruser stående uten røkting i 4 døgn grunnet dårlig vær. I noen tilfeller ble ruser også røktet etter kun ett døgn. Antall stasjoner ble begrenset av antall tilgjengelige ruser (30 stk. i alt) og av det som var mulig å røkte av ruser pr. dag, inkludert biologisk prøvetaking og opptelling av fangst.
Figur 18. Fiske med ruser og biologisk prøvetaking.
Rusesett for Lauvøysvaet i 2019 er vist på kartet i Vedlegg 3 (figur 3.1). Eksisterende oppdrettsanlegg er også angitt på kartet. Glasøysvaet har et oppdrettsanlegg plassert ut mot Ramsøyfjorden i nordøst (12466 Brettingen, Mowi AS). Lauvøysvaet er det området som har størst utstrekning og nærhet til de nye oppdrettslokalitetene (33937 Nørholmen, Mowi AS og 34677 Hestøya, Nekton Havbruk AS) som ble begge ble satt i drift i 2016 .
Det ble satt opp 6 stasjoner ved Glasøysvaet med totalt 27 rusesett her (ruse nr. 1-3), og 22 stasjoner i tilknytning til Lauvøysvaet med totalt 90 rusesett (ruse nr. 4-13) (se figur 3.1 i Vedlegg 3 for oversikt). Fisket foregikk her i 3 uker. Det ble fisket i 2 uker på 21 stasjoner i referanseområdet ved Araneset (figur 3.2 i Vedlegg 3), med i alt 54 rusesett (ruse nr. 14-22). Videre ble 19 stasjoner samtidig med Araneset benyttet i to uker i referanseområdet på Åkvika (Vedlegg 3, figur 3.2), med i alt 48 rusesett (ruse nr. 23-30). I referanseområdet i Dromnessundet ble det den siste uken av fisket satt opp 17 stasjoner med i alt 45 rusesett (ruse nr. 14-30 som de to første ukene ble brukt på Araneset og Åkvika. Flest rusesett ble benyttet på Lauvøysvaet fordi dette området har størst areal og i tillegg er hovedområdet for undersøkelsene. Oversikt over posisjoner for rusesettene for 2019 finnes i vedlegg 3, tabell 3.1, 3.2a, 3.2b, 3.3, 3.4 og 3.5.
Bunntypen på ruseposisjonen ble bestemt ved hjelp av en liten sleperigg beskrevet i figur 19. Sleperiggen ble senket til bunnen og så slept langs rusen for å få et overblikk over bunntype og vegetasjon.
Figur 19. Sleperigg med videokamera og ulike bunntyper på noen av ruseposisjonene.
5.1.2 - Statistiske metoder
Antallet torsk i fangstene (fangst pr. rusesett pr. døgn: CPUE) ble delt opp og analysert for 3 størrelseskategorier av torsk i) antatt 0-gruppe bestående av fisk < 20 cm, ii) flerårig umoden torsk i størrelsesområdet 20-50 cm og iii) moden torsk > 50 cm. Analysene ble gjort på samme måte for disse størrelseskategoriene. Først ble det undersøkt om det var en generell forskjell mellom områdene med hensyn til fangst. Her ble det benyttet en «linear mixed effect model» (LME-modell) hvor område ble brukt som en kategorisk uavhengig forklaringsvariabel, og rusestasjoner nøstet innen år ble brukt som en «tilfeldig» effekt. Dette ble etterfulgt av en Tukey post-hoc test for å sammenligne alle gyteområder mot hverandre. Deretter ble det testet om det var forskjeller i fangster mellom år innen hvert område. Dette ble også gjort ved hjelp av en LME-modell med år som en kategorisk uavhengig forklaringsvariabel og rusestasjon som en tilfeldig effekt etterfulgt av en Tukey post-hoc test for å sammenligne alle år mot hverandre. Til slutt ble det undersøkt om avstand til nærmeste oppdrettsanlegg fra hver enkelt ruseposisjon påvirket fangstene. I denne LME-modellen ble avstand til nærmeste oppdrettsanlegg brukt som en kontinuerlig forklaringsvariabel og rusestasjoner nøstet innen år ble modellert som en «tilfeldig» effekt.
Andel torsk i fangstene som ble definert som 0-gruppe, ble analysert med hensyn til forskjeller både mellom år og mellom områder. Enveis ANOVA fulgt av Bonferroni-Holm post-hoc test ble benyttet etter arc-sinus transformasjon av data. Forskjeller i lengde, vekt og kondisjon mellom områder innen et år ble analysert med enveis ANOVA fulgt av Bonferroni-Holm post-hoc test. Lengde og kondisjon av fisk mellom områdene ble også sammenlignet ved hjelp av en LME-modell med område som en kategorisk forklaringsvariabel og rusestasjon nøstet innen år som en tilfeldig effekt. Dette ble etterfulgt av en Tukey post-hoc test som sammenlignet alle områder mot hverandre.
Alle statistiske tester er gjennomført med et signifikansnivå på 0,05, og p-verdier lavere enn dette betyr statistisk signifikante forskjeller i de testene som er gjennomført.
5.1.3 - DNA-analyser av yngel og umoden torsk
DNA-analysene ble gjennomført i første omgang på torsk samlet ett av årene yngelundersøkelsene foregikk. Resultatene fra disse analysene var bestemmende for om ytterligere prøver fra andre år skulle analyseres. Torsk, inkludert yngel, ble analyser for genetiske forskjeller ved hjelp av SNP-genotyping i henhold til iPLEX-protokollen (https://agenabio.com/) der MassARRAY ™ analysatoren fra Agena ble benyttet. Genotyper ble tilordnet ved å bruke MassARRAY SpectroTYPER RT v3.4 programvare fulgt av manuell inspeksjon av genotyper med programmet MassARRAY TyperAnalyzer v4.0. Metoden er basert på PCR amplifisering av DNA-områder med påfølgende seperasjon av de ulike SNPene i et massespektrometer der det benyttes vakuum og høy spenning mellom to plater.
5.2 - Resultater
5.2.1 - Torsk
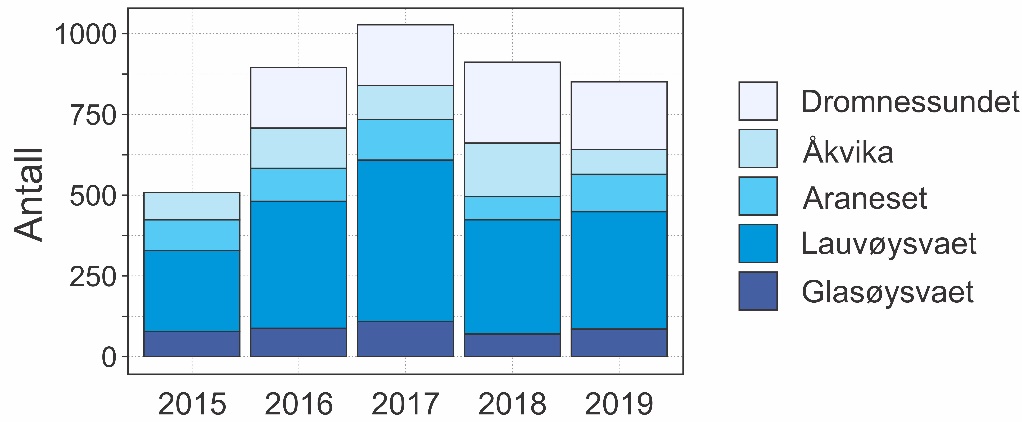
I rusefisket ble det fanget mellom 508 og 1027 torsk årlig fra 2015 til 2019, i alt 4194 torsk (Figur 20). Flest torsk ble fanget på Lauvøysvaet på Smøla, som også hadde lengst fiskeperiode. Færrest torsk ble fanget i 2015 da fiskeperioden var noe kortere og Dromnessundet ikke var med i undersøkelsene. Gjennomsnittlig antall torsk fanget pr. rusesett i et år støtter opp om at tilgjengelighet av torsk 2015 ikke var ulik tilgjengeligheten i perioden 2016-2019. Det ble ved flere anledninger observert at større torsk i rusene hadde spist både leppefisk og mindre torsk.
Det ble observert skader på torsk i rusene fra angrep av skarv, og dette ble vurdert å være en av årsakene til dødelighet, særlig hos yngre torsk. Noen av disse individene ble frosset ned, og alder på ung torsk med lengde mellom 14 og 30 cm ble senere bestemt for 14 individer ut fra øresteiner (otolitter). Her ble 9 torsk mellom 14 og 18 cm alle funnet å være 0-gruppe (årsyngel), mens 4 torsk mellom 22,5 og 30 cm var 1-gruppe (ettåringer). Den siste torsken på 28,5 cm var 2-gruppe (toåring). For 0-gruppen ble det i flere tilfeller også funnet en reduksjon i lengde på opptil 1 cm for det samme individet målt fersk og etter opptining. Ut fra dette ble det i videre beregninger satt en øvre grense på 20 cm for 0-gruppe torsk.
Størrelse av torsk i fangstene
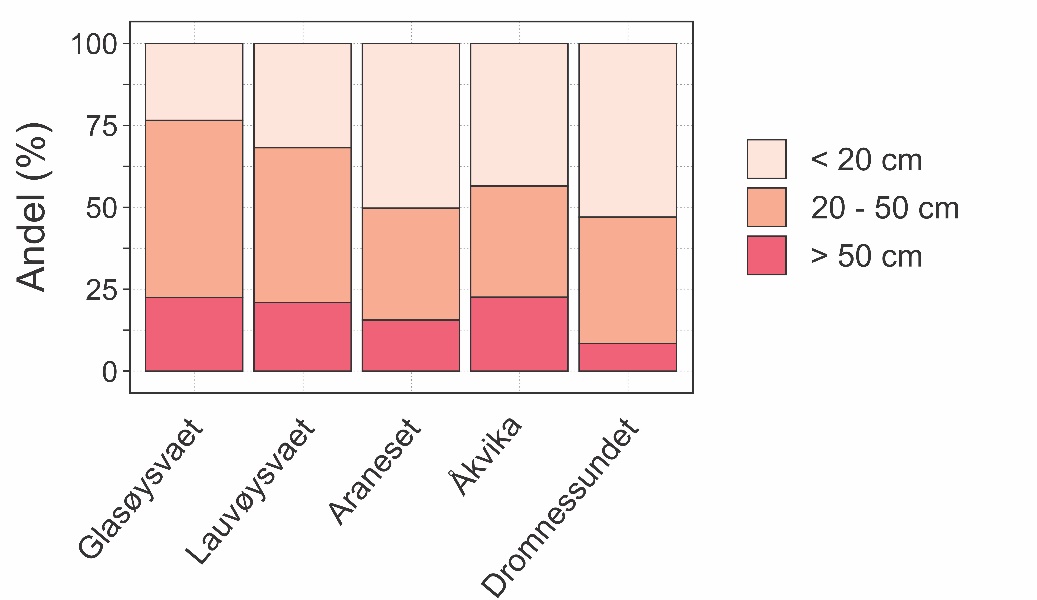
Andelen torsk totalt i tre størrelsesgrupper (< 20 cm: 0-gruppe, 20-50 cm: flerårig umoden torsk og > 50 cm: moden torsk) er vist i figur 21 for hvert av områdene det ble fisket i. Forholdvis flere stor torsk (> 50 cm) ble fanget i Åkvika, Glasøysvaet og Lauvøysvaet, mens den største andelen av 0-gruppe torsk (< 20 cm) ble observert i Dromnessundet og Araneset. Andelen av hovedsakelig umoden torsk (20-50 cm) viste en tendens til å være større på Glasøysvaet og Lauvøysvaet ute på Smøla enn i de tre referanseområdene.
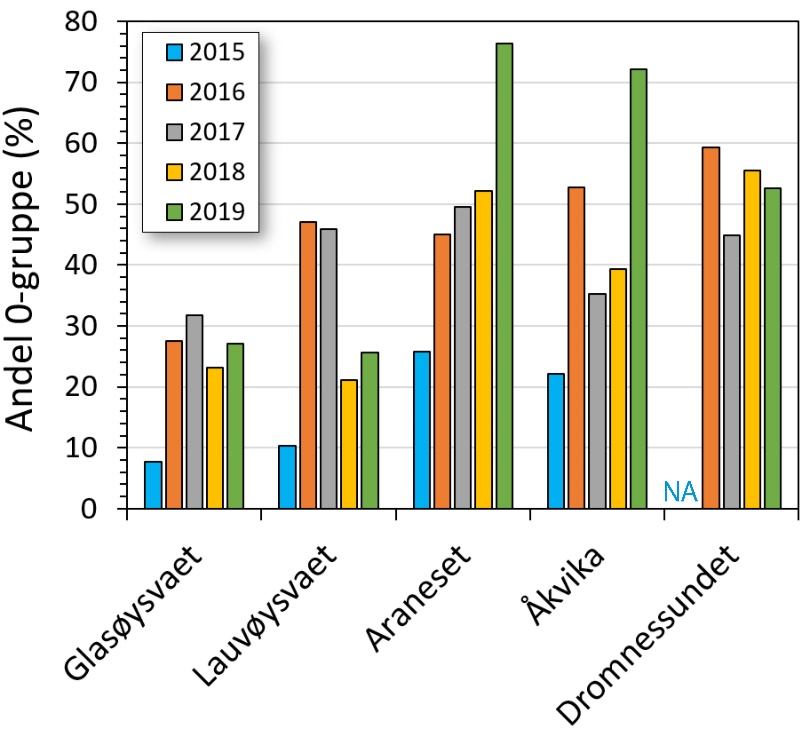
Andelen torsk i fangstene som var 0-gruppe, er vist i figur 22 for alle år og områder, og 2015 hadde generelt lavest andel av 0-gruppe, mens Dromnessundet viste en relativt stabil andel på rundt 50 % 0-gruppe i fangstene. Alle år sett under ett viste signifikante forskjeller i andel 0-gruppe mellom områder, der Dromnessundet hadde høyere andel 0-gruppe enn på Glasøysvaet (p = 0,028). Alle områdene sett under ett viste signifikant lavere andel av 0-gruppe i 2015 enn i 2016 og 2017 (p = 0,021).
Figur 20. Oversikt over antall torsk fanget i rusefisket hvert år for de ulike områdene.
Figur 21. Andel i tre ulike størrelsesgrupper av torsk fanget i rusefisket totalt for de ulike områdene.
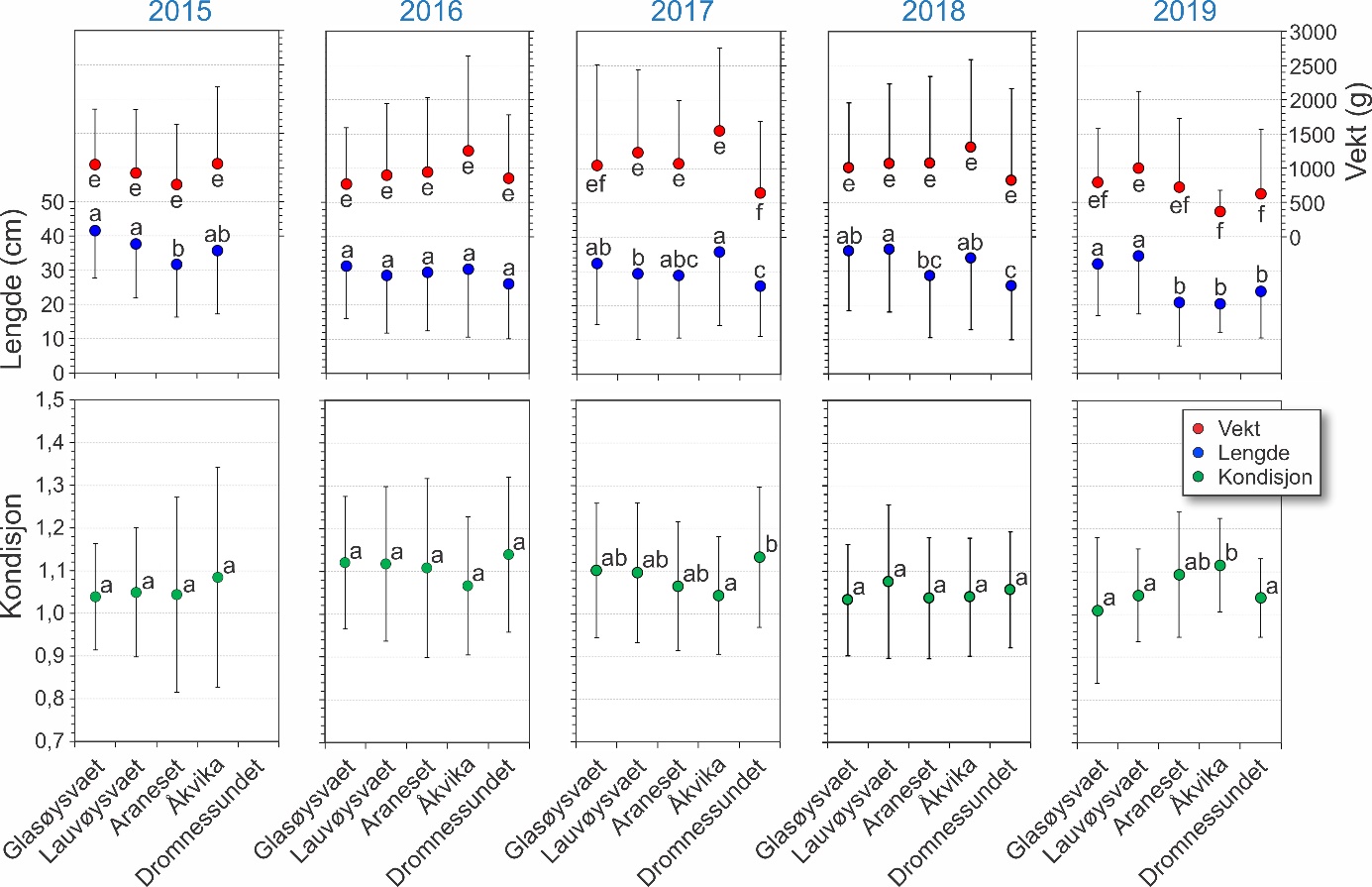
Gjennomsnittlig lengde, vekt og kondisjon med standardavvik hos torsk fanget fra de ulike undersøkelsesområdene for perioden 2015-2019 er vist i figur 23. Kondisjon ble regnet ut som Fultons kondisjonsfaktor [K = Vekt (g) x 100 / Lengde (cm)3] for all torsk over 18 cm siden vekt var vanskelig å måle på torsk under denne størrelsen (ca. 100 gram) i en åpen båt utsatt for bevegelser fra bølger og vind. Lengden hos torsken var fra 8,5 til 111 cm. Noen år var forskjeller i lengde mellom områdene signifikante, mens dette for eksempel ikke ble observert i 2016 (figur 23). Det ble videre funnet at lengen hos torsken fanget ved Araneset og i Dromnessundet var signifikant mindre enn i de tre andre områdene, sett alle årene under ett (p < 0,0001). Tilsvarende ble det funnet signifikante forskjeller i kondisjon mellom områder innen årene 2017 og 2019 (figur 23), mens for alle årene vurdert under ett var det ikke systematiske forskjeller i kondisjon mellom områdene (p > 0,05).
Figur 22. Andel torsk < 20 cm (0-gruppe) i fangstene for de ulike områdene og årene. NA angir år og område det ikke ble fisket.
Frekvensfordelingene viser tydelig to årsklasser, med en topp på 15-17 cm og en topp mellom 27-29 cm i alle områder (figur 24). Ut fra analyse av øresteiner ble dette antatt å være henholdsvis 0-gruppe (0,5 år gammel) og 1-gruppe (1,5 år gammel). Eldre årsklasser var mer utflytende og lot seg ikke skille klart fra hverandre. Gruppering av torsk i de samme lengdegrupper som brukt i figur 21, viste at andelen av små og stor fisk varierte noe med distanse fra nærmeste oppdrettsanlegg innen områder, men denne variasjonen var ikke konsistent mellom områder. Alt i alt var det derfor ingen tydelig trend i størrelse av torsk i rusefangstene og distanse fra nærmeste oppdrettsanlegg.
I hvert av de undersøkte områdene var det mellom 2 og 4 ruseposisjoner som bidro mest med antall små torsk (0-gruppe). Bunnhabitatet på disse stasjonene kan beskrives som sandholdig, blandet i noen tilfeller med grus og stein, og i de fleste tilfeller med forekomst av ålegress eller sukkertare.
Figur 23. Gjennomsnitt og standardavvik for lengde, vekt og kondisjon for torsk fanget i områdene som ble undersøkt. Kun torsk >18 cm er inkludert i beregningen av kondisjon grunnet usikker bestemmelse av vekt for individer ≤18 cm. Ulike bokstaver innen hver av parameterene (lengde, vekt og kondisjon) angir signifikante statistiske forskjeller mellom lokalitetene innen et år.
Mengde torsk i fangstene
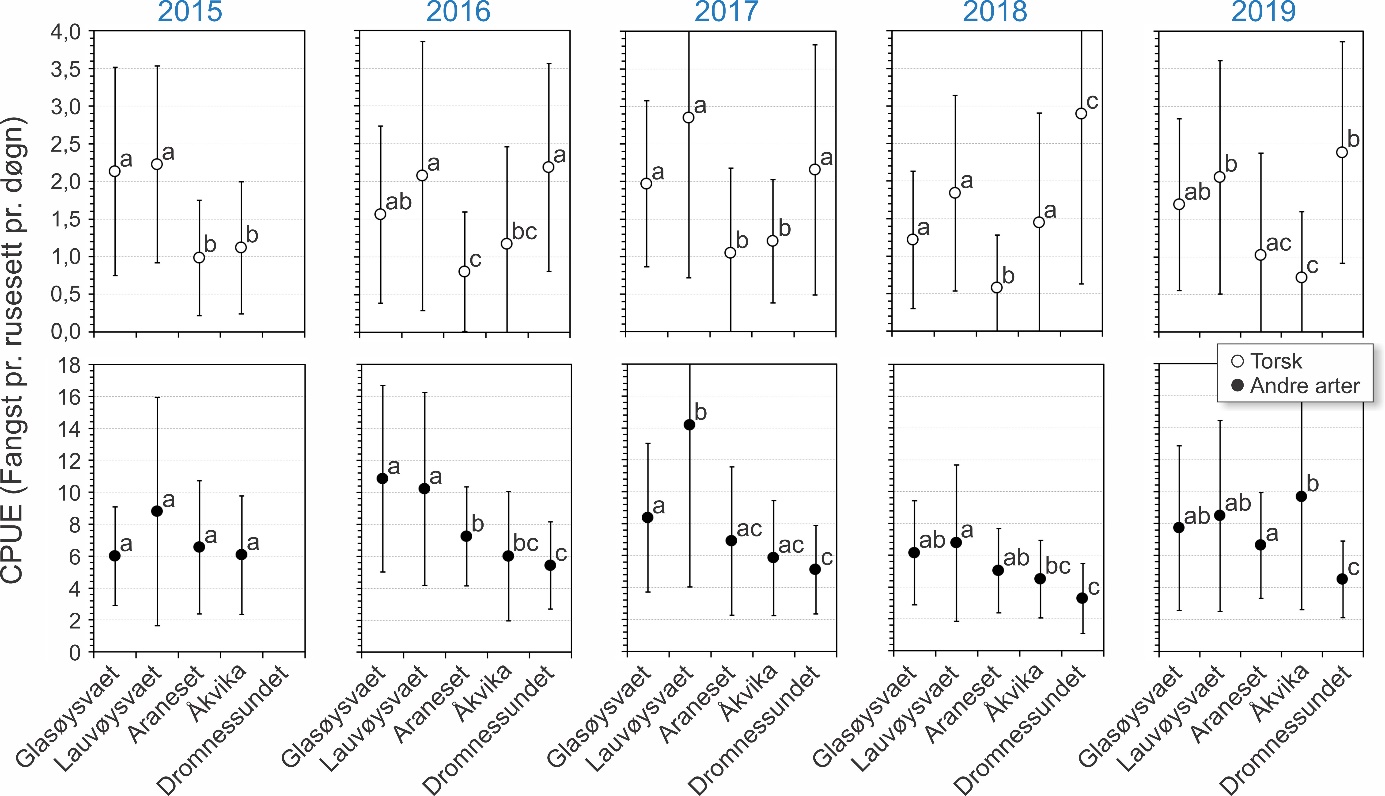
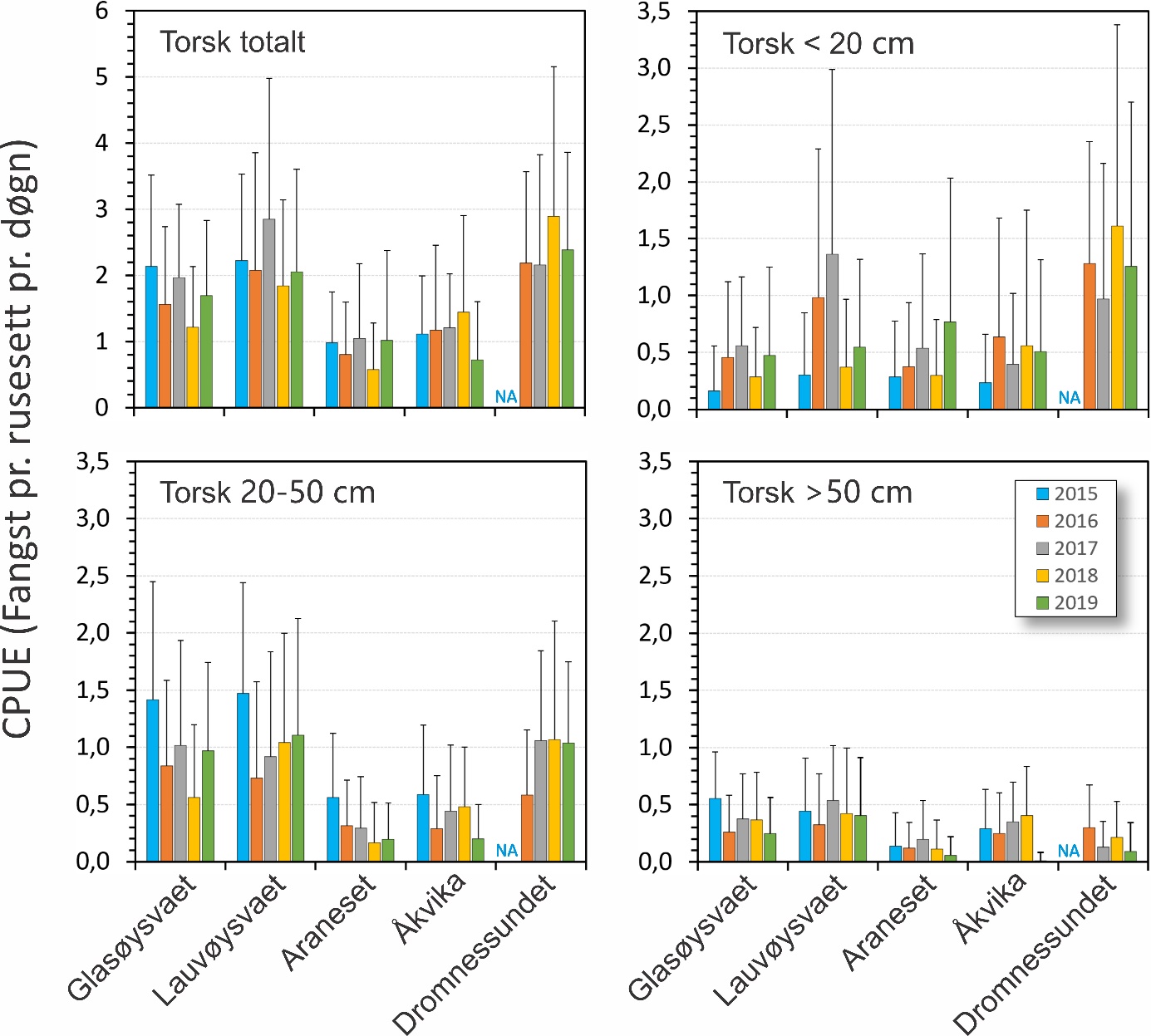
Fangst som funksjon av innsats (CPUE) vil være et indirekte mål på mengde torsk i fangstområdene, og ble beregnet som antall torsk fanget pr. rusesett pr. døgn. Dette er vist for hvert område og hvert år i figur 25. For alle størrelsesgrupper av torsk sett under ett varierte fangstene i gjennomsnitt mellom 0,58 og 2,89 torsk pr. rusesett pr. døgn de ulike områdene og årene. Det var signifikant flere torsk i undersøkelsesområdene Glasøysvaet og Lauvøysvaet på Smøla med 1,21-2,85 torsk pr. rusesett pr. døgn, sammenlignet med referanseområdene Araneset og Åkvika som hadde 0,58-1,45 torsk pr. rusesett pr. døgn (p < 0,0001). Derimot var mengden torsk i Dromnessundet med 2,18-2,89 torsk pr. rusesett pr. døgn på nivå med Lauvøysvaet på Smøla (p = 0,5) men signifikant høyere enn på Glasøysvaet (p < 0,01).
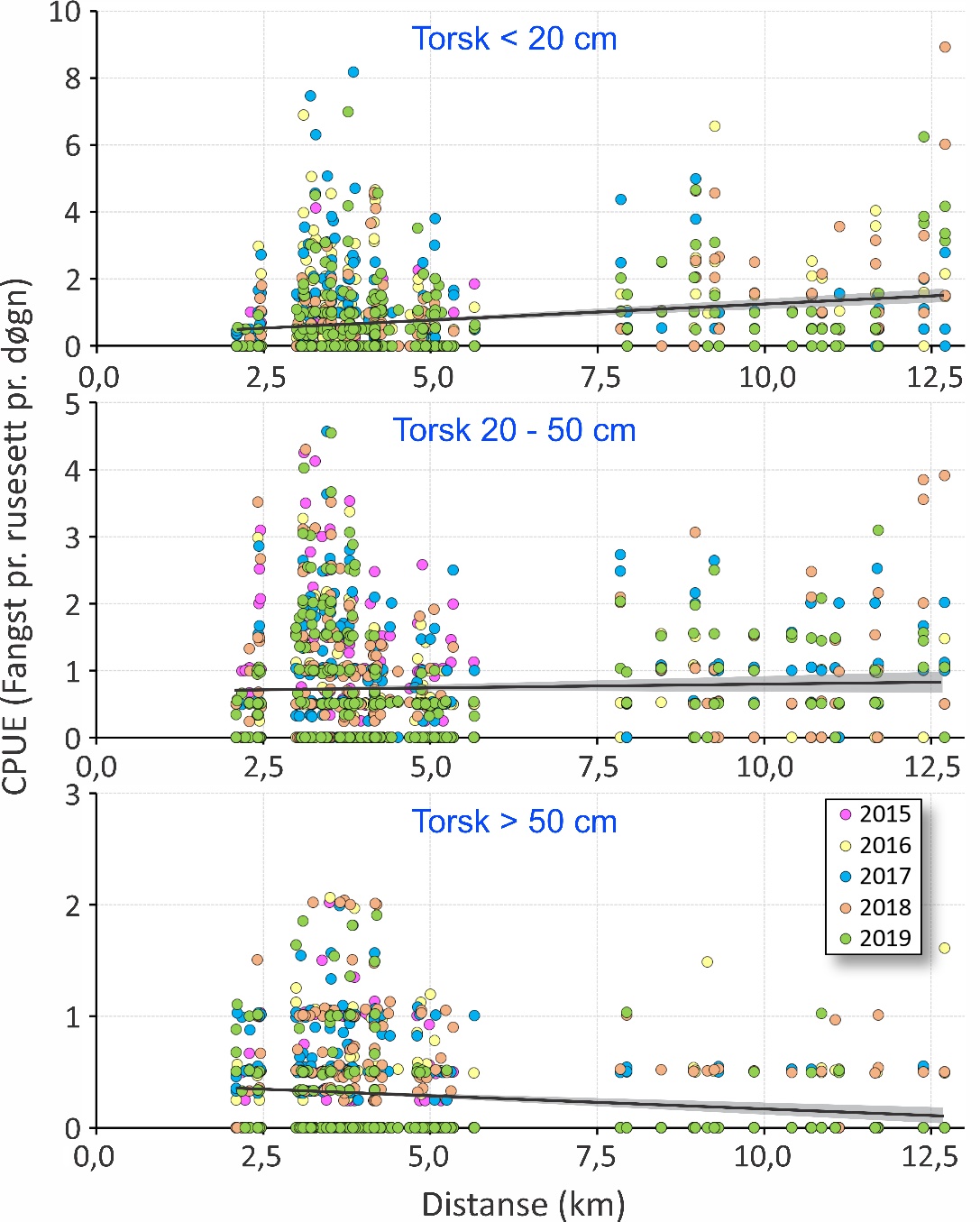
De ulike områdene sammenlignet med hverandre (figur 26) viste at det ble fanget mer årsyngel av torsk (< 20 cm) i Dromnessundet enn i alle andre områder (p < 0,0001). Det ble også fanget mer av årsyngelen på Lauøysvaet enn i de andre områdene utenom Dromnessundet (p < 0,05). Det var ingen signifikante forskjeller for denne størrelsesgruppen av torsk mellom Glasøysvaet, Araneset og Åkvika. Ved sammenligning innenfor hvert av områdene var fangstene av årsyngel på Lauvøysvaet i 2016 og 2017 like, men større enn i de andre årene (p < 0,05). Ved Araneset var fangstene i 2019 større enn i 2015 og 2018 (p < 0,05). Ingen annen mellomårlig variasjon ble funnet innen de andre områdene. Det ble funnet en klart signifikant men svak positiv effekt av avstand til anlegg på fangst av småtorsk (figur 27), der fangstene var større jo lenger unna nærmeste lakseoppdrettsanlegg rusestasjonen var (p < 0,0001). Denne effekten av distanse forklarte likevel mindre enn 2 % av den observerte variasjonen i rusefangstene av årsyngelen.
Figur 24. Frekvensfordelinger av lengde (2 cm lengdeklasser) hos torsk i de ulike områdene, hvert år og alle år samlet.
Figur 25. Fangst pr. enhet innsats (CPUE) for torsk og andre arter fanget i rusene i hovedområdet (Glasøysvaet og Lauvøysvaet) og i referanseområdene (Araneset, Åkvika og Dromnessundet). Data er gitt som gjennomsnitt av fangst pr. rusesett pr. døgn med standardavvik. Ulike bokstaver angir signifikante statistiske forskjeller mellom lokalitetene innen et år.
Når flerårig umoden torsk (20-50 cm) ble sammenlignet mellom områder (figur 26) var det mer av denne størrelsesgruppen torsk på Lauvøysvaet enn i alle områder bortsett fra i Dromnessundet (p < 0,001). Åkvika og Araneset var ikke forskjellig fra hverandre (p = 0,59), men hadde mindre fangster enn i de andre områdene (p < 0,001). Ut over dette ble ingen andre forskjeller funnet. Ved sammenligning innenfor hvert av områdene var fangstene av flerårig umoden torsk større på Glasøysvaet i 2015 enn i 2016 og 2018 (p < 0,01). På Lauvøsyavet var det større fangster av denne torskegruppen i 2015 enn alle de andre årene (p < 0,05). Fangstene var også signifikant lavere her i 2016 enn i 2019 (p < 0,001). Ved Araneset var fangstene av flerårig umoden torsk større i 2015 enn de andre årene (p < 0,01), mens på Åkvika var fangstene i 2015 større enn i 2019 (p < 0,01). I Dromnessundet var fangstene lavere i 2016 enn i de andre årene (p < 0,01). Ellers ble ingen andre forskjeller funnet mellom ulike år innen hvert av områdene. Det ble heller ikke funnet noen effekt av avstand til nærmeste oppdrettsanlegg på rusefangstene av flerårig umoden torsk (p = 0,25) (figur 27).
Sammenligning mellom områder med hensyn til stor torsk (> 50 cm) viste at det var mer av denne størrelsesgruppen på Lauvøysvaet enn i alle andre områder bortsett fra Glasøysvaet (p < 0.01) (figur 26). Ved Araneset ble det fanget lignende antall stor torsk som i Dromnessundet (p = 0,41), men færre enn i de andre områdene (p < 0,001). Det ble også fanget mer stor torsk på Glasøysvaet enn i Dromnessundet (p < 0.01). Årsvariasjon innen områder for den store torsken viste at på Lauvøsyavet ble det fanget mer i 2017 enn i 2016 (p < 0.05). Ved Araneset ble det fanget mer stor torsk i 2017 enn i 2019 (p < 0.05), mens på Åkvika ble det fanget mindre stor torsk i 2019 enn i alle andre år (p < 0.01). I Dromnessundet var fangsten av stor torsk større i 2016 enn i 2017 og 2019 (p < 0.01). Ingen andre forskjeller ble funnet mellom ulike år innen områdene. Når det gjelder avstand til nærmeste oppdrettsanlegg (figur 27) ble det observert en svak men signifikant negativ effekt med hensyn til rusefangstene av stor torsk (p < 0,0001).
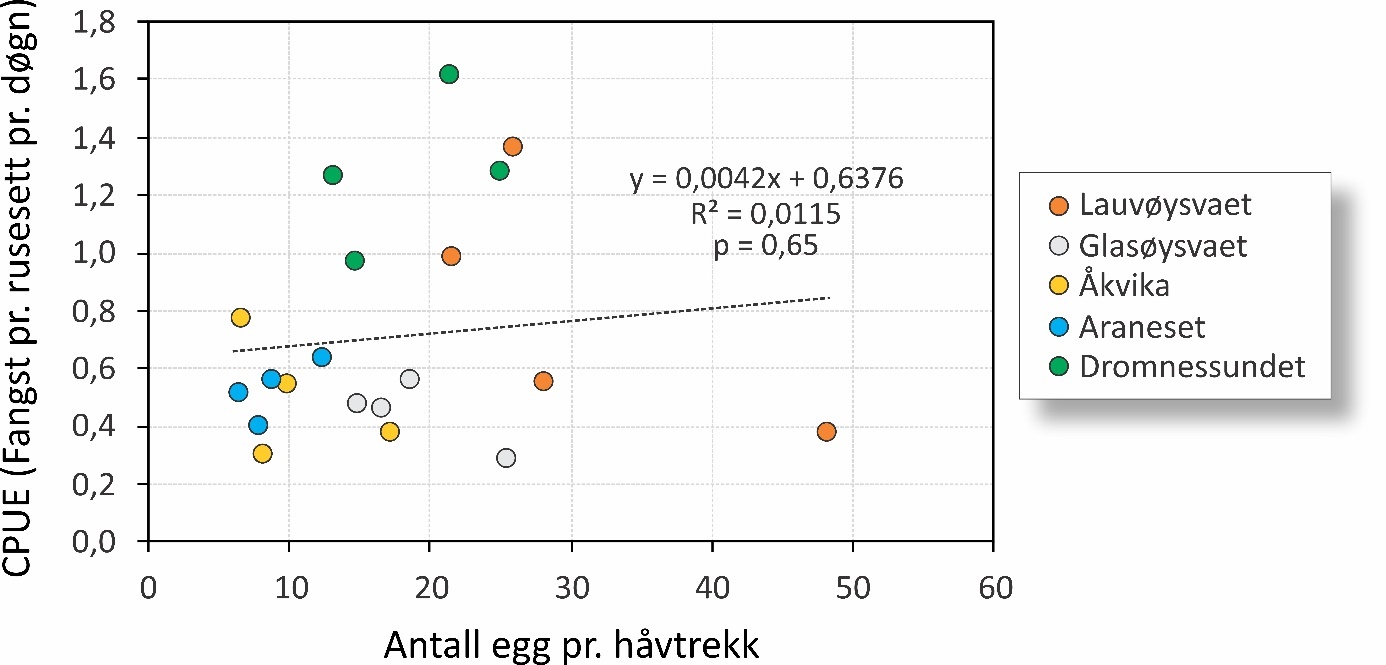
Det ble ikke funnet noen signifikant sammenheng mellom eggmengder målt på eggtoktene i mars og påfølgende fangst av 0-gruppe samme høst (figur 28). Data fra 2015 ble utelatt da det dette året kun ble foretatt ett eggtokt i slutten av mars, som tilsvarer toppen av gyteforløpet. Det ble heller ikke funnet noen statistisk sammenheng innen hvert av områdene de fire årene fra 2016 til 2019.
Figur 26. Fangst pr. enhet innsats (CPUE) for torsk totalt og ulike størrelsesgrupper av torsk, der lengde opptil 20 cm regnes som årsyngel (0-gruppe). Figuren viser fangst i rusene i hovedområdet (Glasøysvaet og Lauvøysvaet) og i referanseområdene (Åkvika, Araneset og Dromnessundet). Data er gitt som gjennomsnitt pr. rusesett med standardavvik.
Figur 27. Fangst pr. enhet innsats (CPUE) for ulike størrelsesgrupper av torsk med hensyn til avstand til et oppdrettsanlegg.
Figur 28. Fangst pr. enhet innsats (CPUE) plottet mot mengde egg i torskestørrelse pr. håvtrekk (gjennomsnitt av tre eggtokt pr. år). Data vises for årene 2016 til 2019. Data for 2015 er utelatt fordi det kun ble gjennomført ett tokt ved gytetoppen dette året. Stiplet linje viser statistisk sammenheng (korrelasjon) med tilhørende formel, men som ikke er signifikant da p > 0,05.
5.2.2 - Andre arter
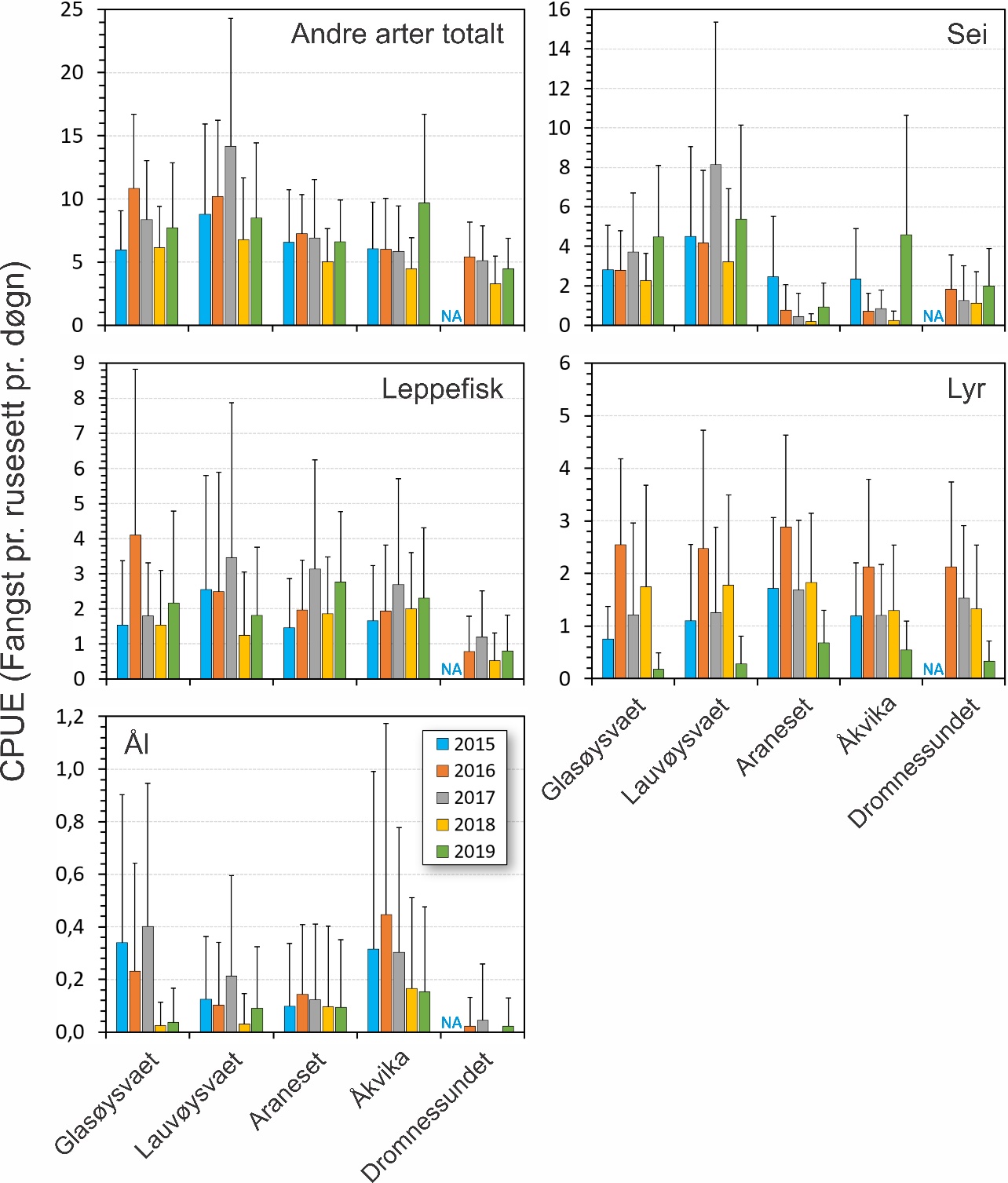
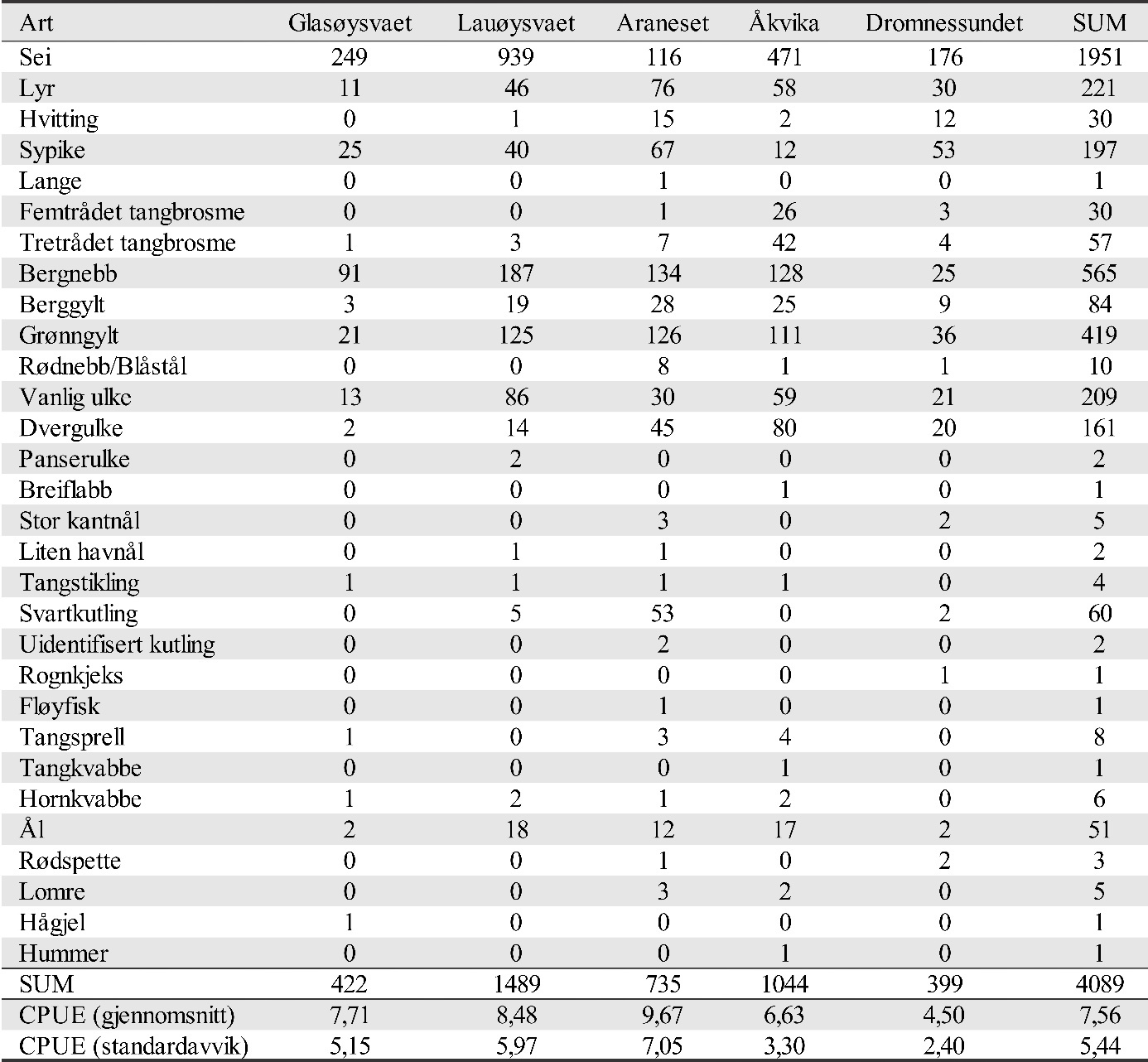
Bifangst i form av andre fiskearter er vist i figur 29. Siden andre fiskearter ikke er det primære målet i studien, er det ikke gjennomført statistiske analyser for hvordan disse variere mellom områder og år. Sei, leppefisk og lyr utgjorde mesteparten av bifangsten. Figur 29 viser at det trolig er forskjeller i forekomst for enkelte av artene mellom år. Spesielt gjelder dette lyr og sei. Et annet trekk er at sei ser ut til å være mer vanlig i de ytre områdene som omfatter lokalitetene på Smøla. Leppefisk og ål ser også ut til å ha lavere forekomst i Dromnessundet enn i de andre områdene.
5.2.3 - DNA-analyser av yngel og umoden torsk
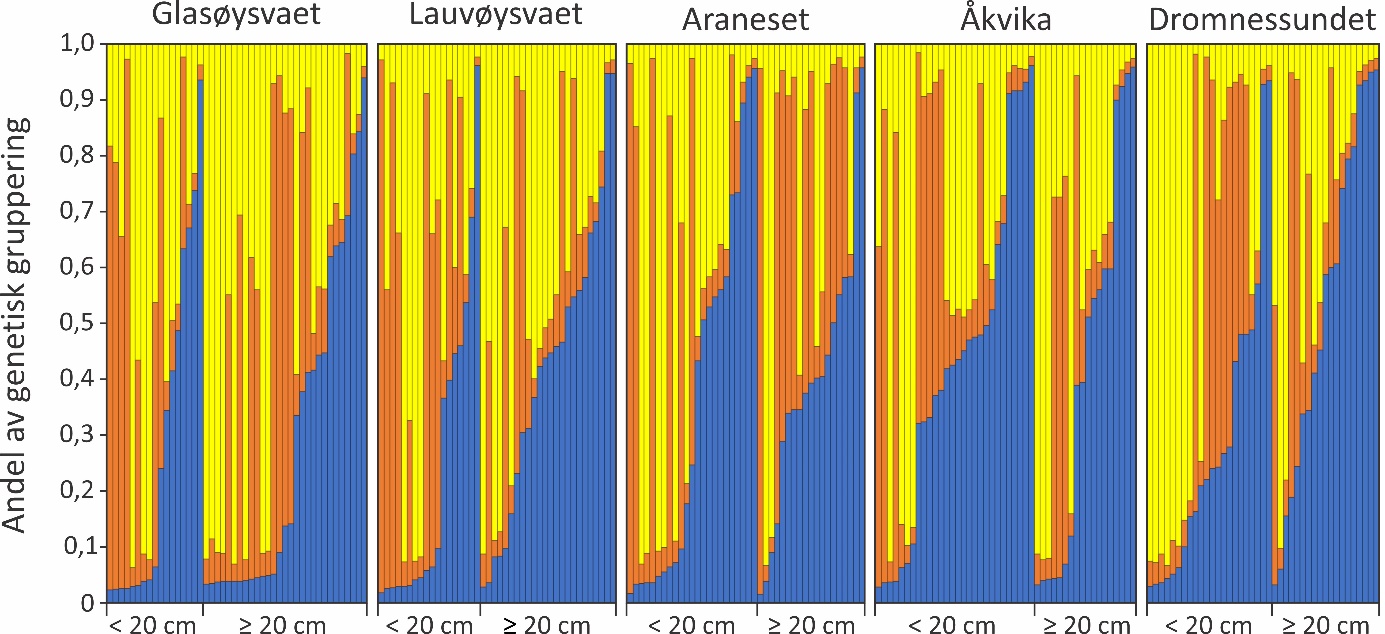
DNA-analysene ble gjennomført på torsk samlet inn ved yngelundersøkelsene i 2017. Ut fra den genetiske informasjonen ble torsken fra yngelundersøkelsen tilordnet tre genetiske typer (grupper eller «clustere» som det også kalles). I figur 30 er denne tilordningen fremstilt grafisk der andelen av de tre gruppene er vist for hvert individ. Det vises tydelig at en av de tre genetiske gruppene dominerer i ulike individer, men det er ikke forskjell i forekomst av genetisk gruppering mellom gyteområdene som er undersøkt eller mellom årsyngel (< 20 cm) og eldre torsk (≥ 20 cm). Ut fra mangel på genetiske forskjeller ble derfor ikke ytterligere analyser foretatt.
Figur 29. Fangst pr. enhet innsats (CPUE) for andre arter totalt, leppefisk, ål, sei og lyr. Figuren viser fangst i rusene i hovedområdet (Glasøysvaet og Lauvøysvaet) og i referanseområdene (Åkvika, Araneset og Dromnessundet). Data er gitt som gjennomsnitt pr. rusesett med standardavvik.
Figur 30. Torsk fra yngelundersøkelsene 2017 gruppert genetisk i tre ulike «clustere» angitt med hver sin farge (blå, rød og gul) for hver av de fem gyteområdene som ble undersøkt. Hver søyle angir en torsk, i alt 217 individer, og innenfor hvert gyteområde er årsyngel (< 20 cm) og eldre torsk (≥ 20 cm) vist hver for seg.
5.3 - Diskusjon yngelundersøkelser
Bruk av åleruser ser ut til å være en god metode for å vurdere rekruttering og størrelsesfordeling av torsk i de undersøkte områdene. Rusene fanger et vidt spekter av størrelser, fra det minste individet på 8,5 cm til det største individet på 111 cm (11,2 kg). Undersøkelsene viste at en betydelig andel av torsken fanget i rusene var årsyngel (0-gruppe), og at denne yngelen er overraskende stor til å være ca. 7 måneder gammel. Størrelsesfordelingene antyder at torsk opp mot 20 cm kan være 0-gruppe, men dette er foreløpig ikke slått fast gjennom undersøkelser av øresteiner som kun påviste 0-gruppe hos torsk med lengder opp til 18 cm.
Med unntak av Dromnessundet var det en betydelig variasjon i andel 0-gruppe i fangstene mellom år innen gyteområdene. Også mellom gyteområdene var det stor variasjon i andel av 0-gruppe, også innen samme år. Mindre variasjon i Dromnessundet kan skyldes at dette området ser ut til å ha større retensjon og derfor i mindre grad er utsatt for transport av egg og larver ut av sundet sammenlignet med de andre og mer eksponerte gyteområdene der mer vannutskifting forventes ut fra en høyere grad av hydrodynamikk. Dromnessundet ligger mer skjermet for de mer utsatte vindforholdene i Edøy- og Ramsøyfjorden, som igjen er en viktig drivkraft for strømforholdene på de andre gyteområdene. I tillegg vil det også være større dynamikk i strømforholdene på de andre gyteområdene på grunn av kyststrømmens påvirkning innover Edøyfjorden. Disse forholdene kan ha hatt betydning for å kunne påvise en sammenheng mellom etablering av oppdrettsanlegg og rekruttering av torskeyngel på nærliggende gyte- og oppvekstområder, om en slik sammenheng skulle finnes. I det foreliggende datamaterialet var det ikke mulig å påvise en sammenheng mellom tidsrommet at oppdrettsanlegget på Nørholmen var etablert og i aktivitet og andel av 0-gruppeyngel på de nærliggende gyteområdene. Variasjonen i andel 0-gruppe skyldes derfor trolig andre forhold av fysisk og biologisk karakter. For eksempel hadde de to gyteområdene Glasøy- og Lauvøysvaet størst andel av umoden torsk mellom 20 og 50 cm. Dette kan bidra til å redusere andelen av 0-gruppe, fordi årsyngelen kan ha vært utsatt for en større grad av predasjon fra større artsfrender (kannibalisme) i disse gyteområdene ute på Smøla. Kannibalisme på årsyngel av torsk, sei og lyr samt beiting på bifangst av leppefisk ble noen få ganger observert hos flerårig torsk fanget i rusene, men dette kan være et resultat av predasjon som har foregått inne i rusene. Ut fra de få observasjonene og kort tid rusene hadde i sjøen mellom hvert hal, ansees det ikke at dette kan ha underestimert andelen 0-gruppe i særlig grad på de to gyteområdene ute på Smøla. Men det viser at en slik kannibalisme kan forekomme på grunt vann i oppvekstområdene til torskeyngelen.
Forskjeller i størrelse og kondisjon hos torsken viste også noe variasjon mellom år og gyteområder. Torsken fanget i Dromnessundet og ved Araneset var i gjennomsnitt mindre enn i Åkvika og i de to gyteområdene ute på Smøla, og forskjeller i andel 0-gruppe kan være med å forklare dette. Det var heller ingen trend i størrelse av torsken og nærhet til oppdrettsanlegg. Torsk som beiter under oppdrettsanlegg, vil kunne få høyere kondisjon grunnet tilgang på energirikt fôr. Forskjeller i kondisjon mellom gyteområder ble kun påvist noen år, men dette kunne ikke knyttes til drift av nærliggende oppdrettsanlegg.
Siden ruser er passive redskaper, er fangst avhengig av torskens aktivitet og mengde torsk på lokaliteten hvor det fiskes. Mengde torsk i fangstene pr. døgn blir derfor et indirekte mål på mengde torsk i et område av en viss utstrekning rundt rusene, det vil si et indirekte pål på bestandstetthet. Resultatene viste at Dromnessundet og områdene på Smøla hadde større tetthet av torsk enn i de to områdene Araneset og Åkvika. Dette skyldtes at Glasøysvaet og Lauvøysvaet på Smøla hadde mer av både flerårig umoden og eldre moden torsk enn Araneset og Åkvika, mens Dromnessundet hadde mer 0-gruppe og umoden flerårig torsk. Årsaken til disse forskjellene er ikke kjent, men både fiske, bunnforhold, mattilgang, predasjon og rekruttering av yngel kan spille en rolle. Når det gjelder avstand til et oppdrettsanlegg fra en ruseposisjon, så ble det funnet mengde 0-gruppe torsk i fangstene med økende avstand fra et oppdrettsanlegg, mens det motsatte var tilfelle for moden torsk. Disse sammenhengene var svake, men signifikante statistisk sett. For flerårig umoden torsk ble det ikke funnet noen slike sammenhenger. Forskjeller i fangst av torsk ulike år på Lauvøysvaet kunne heller ikke knyttes til drift av oppdrettsanlegget på Nørholmen.
Det ble ikke funnet noen signifikant sammenheng mellom eggmengder og rekruttering til 0-gruppen, hverken for alle områdene samlet eller innen områdene rusefisket ble gjennomført. En forutsetning for en slik sammenheng er at egg og larver fra gyting i et område holdes tilbake (retensjon), slik at lokal gyting også vil bidra til rekruttering lokalt. Manglende retensjon i områdene støttes av simuleringene av strøm gjennomført ved hjelp av de hydrodynamiske modellene (se tabell 2 i kap. 4.2.3). Manglende retensjon kan også forklare hvorfor det ut fra DNA-analysene ikke ble funnet genetiske forskjeller mellom torsken fra de ulike områdene. Den høye dynamikken i vannmassene i hele det undersøkte området som ble studert i ICOD-prosjektet, vil være en effektiv barriere mot geografisk isolasjon som er nødvendig for utvikling av genetiske forskjeller.
Dromnessundet viser imidlertid størst grad av retensjon (tabell 2), men retensjonsindeksen er kun beregnet for en periode på 5 dager. Torskeegg vil bruke mellom 2 og 3 uker på å klekke og larvene ytterligere 2 måneder på å søke til bunnen. Retensjonsindeksen i tabell 2 indikerer derfor at det vil være en betydelig transport av egg og larver inn og ut av Dromnessundet. Imidlertid utmerker Dromnessundet seg ved å ha god rekruttering som tidligere nevnt. Dette var uventet fordi bunnsubstratet på grunt vann her består mye av stein, grus og sand, nesten uten beskyttende vekster som sukkertareskog eller større ålegrasenger. Sammenlignet med de andre områdene hadde Dromnessundet i tillegg den laveste forekomsten av stor torsk, og dette kan også ha bidratt til bedre rekruttering gjennom redusert kannibalisme.
Andre fiskearter som utgjorde bifangsten i rusefisket, viste en betydelig variasjon, både mellom områder og mellom år innen områder. Lavere forekomst av ål og leppefisk i Dromnessundet kan ha sammenheng med at sukkertare og ålegras ikke er vanlig her. Stor torsk beiter blant annet på leppefisk og kan derfor forekomme i mindre mengder på grunt vann i Dromnessundet fordi det er lite leppefisk her. Yngel av sei var mest vanlig i områdene ute på Smøla, og ser ut til å være assosiert med forekomst av tareskog. Lyr viste tydelige sterke og svake årsklasser samtidig i alle de undersøkte områdene, noe som indikerer et regionalt signal i rekrutteringen hos denne arten. Det ble ikke funnet noen nedgang i leppefisk fra 2015 til 2019 i noen av områdene, til tross for et omfattende fiske på leppefisk til bruk som rensefisk i lakseoppdrett. Lengden til leppefisken ble ikke målt, og mulige endringer i størrelse over tid grunnet minstemål i fisket kunne derfor ikke undersøkes.
6 - Biokjemiske undersøkelser av gytefisk på Smøla
6.1 - Innledning
Det er velkjent at vill fisk tiltrekkes oppdrettsanlegg for å beite på fôrspill (Callier m.fl. 2018). Langs norskekysten er det godt dokumentert at særlig sei og torsk samler seg i betydelige mengder og beiter på laksepellet under oppdrettsanlegg (Bagdonas m.fl. 2012; Dempster m.fl. 2009; Skog m.fl. 2003; Uglem m.fl. 2009; Uglem m.fl. 2014). Det finnes ingen eksakte tall på mengde fôrspill, men det estimeres at omkring 3-5 % går gjennom merdene uten å bli spist av laksen (Uglem m.fl. 2020). Med et årlig fôr-forbruk på over 1,6 millioner tonn med pellets i norsk laksenæring (Aas m.fl. 2019), så betyr det at mer enn 48 tusen tonn med laksefôr kommer ut i naturen. Dette utgjør med andre ord en betydelig mengde organisk stoff og energi som punktvis tilføres det marine miljøet langs kysten.
Analyse av spesifikke fettsyrer (trofiske markører: FATM) er en veletablert metode for å studere trofisk interaksjon og «hvem spiser hvem» i økosystemene (Dalsgaard m.fl. 2003). FATM-metodene baserer seg på at de fleste av de marine fettsyrene syntetiseres på de laveste trofiske nivåene (bakterier, planteplankton og dyreplankton) og deretter akkumulerer opp gjennom næringskjeden med kun mindre biokjemiske modifikasjoner.
Sammensetningen av laksefôret har endret seg betydelig de siste tiårene, og bruken av marine ingredienser har falt fra 90 % i 1990 til 25 % i 2016. Tilsvarende har mengden av plantebaserte ingredienser økt fra nesten 0 til 71 % i samme periode (Aas m.fl. 2019; Ytrestøyl m.fl. 2015). Denne endringen i sammensetning av laksefôret har særlig hatt betydning for fettsyreprofilen i fôret. Mengden av terrestriske fettsyrer som oljesyre (18:1n-9), linolsyre (18:2(n-6)) og α-linolensyre (18:3(n-3)), har økt svært mye sammenlignet med hva som vanligvis finnes i marine dyr. Dette har gjort at FATM-analyser er et meget nyttig verktøy til å påvise om fisk har spist laksefôr eller en naturlig marin diet, både for rømt laks (Skilbrei m.fl. 2015) og vill fisk (Fernandez-Jover m.fl. 2011).
I forbindelse med tidligere planer for storstilt torskeoppdrett er det gjort mange diettstudier på fôring av torsk, og det er vist at torsk tåler en diet basert på planteproteiner og oljer uten at det påvirker vekst og helse til fisken (Albrektsen m.fl. 2006; Colburn m.fl. 2012; Hansen m.fl. 2006; Hansen m.fl. 2007a; Hansen m.fl. 2007b; Hansen & Hemre 2013; Karalazos m.fl. 2007; Olsen m.fl. 2007). Torsk som spiser en fettrik diett, får en kraftig økning i leverstørrelse på grunn av at torsken lagrer fett i leveren og ikke i musklene slik som hos laksefisk. I tillegg vil fettsyreprofilen i leveren endres hurtig og avspeile fettsyreprofilen i dietten (Albrektsen m.fl. 2006; Hansen m.fl. 2008; Jobling & Leknes 2010; Jobling m.fl. 2008; Lie m.fl. 1986; Standal m.fl. 2008).
Selv om torsk ikke tar skade av å spise plantebasert diet i oppdrett, så er det uklart om slike fôrkilder av ikke-marint opphav kan ha negative konsekvenser for villfisk sin reproduksjon (Gonzalez-Silvera m.fl. 2020). Det er viktig å ha en optimal stamfiskdiet for å få god eggkvalitet og høy overlevelse hos fiskelarver (Tocher 2010). Studier har vist at høy eggproduksjon (fekunditet) og god larveoverlevelse er positivt korrelert med langkjedede flerumettede fettsyrer (PUFA) i dietten (Norberg m.fl. 2017; Pickova m.fl. 1997; Rojbek m.fl. 2014; Salze m.fl. 2005; Sawanboonchun m.fl. 2008). Forsøkene tyder på at det er viktig å ha høye nivåer av arakidonsyre (20:4(n-6)), men også at forholdstallet mellom 20:4(n-6) og andre PUFA som eikosapentaensyre (20:5(n-3)), har stor betydning. Alle disse studiene er gjennomført med variasjoner av marine fettsyrer, men ingen av studiene har undersøkt reproduksjonssuksessen hos torsk som har spist en diet med så avvikende lipid sammensetninger som finnes i laksefôr.
Andre FATM som er nyttige for å beskrive dietten hos kysttorsk er langkjedede enumettetede fettsyrer (MUFA): gadoleinsyre (20:1(n-11)), gondoinsyre (20:1(n-9)), erucinsyre (22:1(n-9)) og ketolinsyre (22:1(n-11)). Disse syntetiseres av copepoder som raudåte (Calanus sp.) og akkumuleres i høye nivåer hos pelagiske fisk som tobis, lodde, sild og makrell. Disse fettsyrene kan dermed brukes som FATM for beiting i den pelagiske næringskjeden (Petursdottir m.fl. 2008). Tilsvarende så akkumulerer den bentiske næringskjeden relativt høyere nivåer av (n-7) MUFA og (n-6) PUFA: 18:1(n-7), 20:1(n-7), 22:1(n-7), 20:2(n-6), 20:3(n-6), 20:4(n-6), 22:4(n-6) og 22:5(n-6) (Kelly & Scheibling 2012).
Regionen rundt Smøla har stor tilstedeværelse av oppdrettsanlegg, og målsetningen med undersøkelsene av biokjemisk innhold hos torsk har vært å studere i hvilken grad torsk som benytter gyteområdene på Smøla har vært og spist laksefôr under oppdrettsanlegg. I motsetning til tidligere studier som har undersøkt biokjemisk sammensetning hos villtorsk, omfatter våre undersøkelser torsk som er fanget over tid på gytefeltene og ikke i umiddelbar nærhet av oppdrettsanleggene. Dette vil dermed gi bedre oversikt over hvor stor andel av gytemoden torsk som beiter under oppdrettsanleggene.
Ut fra lipidanalyser av torskelever har vi brukt FATM-metoden til å klassifisere individer av gyteklar torsk sin fortrukne diettpreferanse forut for gytingen: laksefôr, pelagisk fisk eller bentiske dyr. Fettsyreprofilene i lever, gonader (rogn og melke) og muskel ble sammenlignet, og resultatene diskuteres ut fra mulighetene av at beiting på laksefôr kan ha en effekt på torsken sin reproduksjon.
6.2 - Metoder
6.2.1 - Innsamling av gytetorsk og prøvetaking
Prøver av torsk fra fiske på gyteområdene Breisvaet, Blåsværsvaet, Lauvøysvaet og Glasøysvaet på Smøla (se kartet i kapittel 3) ble samlet av en lokal fisker som drev med garnfiske. Torsken ble samlet inn fra februar til april over to gytesesonger (2018 og 2019). Det ble fanget inn 450 torsk i 2018 og 490 torsk i 2019. Innsamlingen foregikk ved at hode, lever og gonader (rogn og melke) ble pakket i plastposer hver for seg og samlet i en litt større plastpose, sammen med informasjon om lengde, kjønn, posisjon og dato for fangsten. Alle prøver ble frosset ved -20°C og senere sendt til Havforskningsinstituttets forskningsstasjon i Austevoll for opparbeiding. Det ble tatt biokjemiske prøver av 329 og 488 torsk fra henholdsvis 2018 og 2019. I 2018 hevdet fiskeren at det var skrei til stede i fangstene, og derfor ble en bit av muskelen i nakken lagret på 100 % etanol for analyse med hensyn til scnDNA markøren pantophysin (Pan I) for å kunne skille eventuell skrei fra kysttorsk (Dahle m.fl. 2018). Det ble også tatt otolitter (øresteiner) fra torskehodene for å bestemme alder, alder ved første gyting og bestandstilhørighet (skrei eller kysttorsk). Videre ble lever, rogn og melke veid, og det ble tatt små vevsprøver av disse vevene for fettsyreanalyser. Prøvene ble frosset i flytende nitrogen og siden oppbevart på -80°C frem til analyse. I tillegg ble det i 2019 også på samme måte tatt muskelprøver fra nakken for fettsyreanalyser.
Også laksefôr fra to anlegg i nærheten av gyteområdene ble analysert med hensyn til sammensetning av fettsyrer: Polarfeed Marin og Marin Harvest RI.
6.2.2 - Analyser av fettsyrer
Mengde og sammensetning av fettsyrer ble bestemt ved direkte metanolyse og gasskromatografi som angitt i Meier m.fl. (2006). Kort beskrevet så ble en liten prøve med vev (50-100 mg) veid i et 16 ml glassrør. En kjent mengde med nonadecanoic syre (19:0) ble tilsatt som intern standard, og alle fettsyrer ble omdannet til fettsyremetylesters (FAME) med metanolysereagens (2,5 M HCl i vannfri metanol). Metylesterene ble ekstrahert med heksan og deretter analysert med gasskromatografi. Metoden kan skille mellom 51 forskjellige fettsyrer som identifiseres ut fra sammenligning av retensjonstid med standard blandinger av FAME, retensjonsindeks-kort og massespektra bibliotek (GC-MS) (Wasta & Mjøs 2013). Når det i rapporten refereres til fett og fettinnhold, er dette beregnet fra den samlede mengden fettsyrer som er bestemt i analysene.
6.2.3 - Statistiske analyser og klassifisering av fettsyredata
Ut fra gruppering av fettsyreprofiler i leverprøver ble det satt opp tre grupper med hensyn til fôrpreferanse. Klassifiseringen ble gjort med utgangspunkt i fettsyremarkører angitt i tabell 3 der Terrestrisk FATM er karakteristisk for laksefôr, Pelagisk FATM er karakteristisk for pelagisk fisk og Bentisk FATM er karakteristisk for bunnlevende fôrorganismer. Denne klassifiseringen markerer ikke klart adskilte diettgrupper eller næringskjeder, tvert imot er det naturlige overlapp i området omkring grensene mellom gruppene, og mange fisk vil spise i alle tre gruppene. Men disse diettgruppene kan brukes til å estimere torskens foretrukne spiseadferd.
Tabell 3. Trofiske fettsyremarkører (FATM) for tre ulike diettgrupper (næringskjeder).
Torsk som har spesialisert seg på å spise laksefôr, får unaturlig høye nivåer av de terrestriske FATM, og særlig 18:2 (n-6) egner seg godt for å identifisere disse individene. I de marine miljøene utgjør denne fettsyren sjeldent mer enn 2-4 % av den totale mengden fettsyrer, og derfor var det første kriteriet for å tilhøre laksefôrgruppen at 18:2 (n-6) i torskens lever utgjorde mer enn 4 % av den totale fettsyremengden. Forskjeller mellom gytefelt med hensyn til andelen torsk som spiser laksefôr ble testet med en logistisk regresjon med binomial responsvariabel og gytefelt som en kategorisk forklaringsvariabel, og der data er slått sammen over år siden det var data fra Blåsværsvaet kun i 2019.
Det neste kriteriet deler de resterende fisk inn i torsk som foretrekker å spise i den pelagiske næringskjeden, og «andre» som tilsvarer den bentiske næringskjeden. Fettsyremarkører fra MUFA med opprinnelse i raudåte (Calanus-FATM) er et velbrukt kriterium for om en organisme spiser i den pelagiske næringskjeden, det vil si beiter på fet fisk som lodde, tobis, sild og makrell, eller direkte på dyreplankton som copepoder av typen Calanus, amphipoder og krill. Dette er helt klart et dynamisk kriterium, og det viser ikke «enten/eller» men kan brukes til å indikere graden av pelagisk diet fremfor benthisk diet. I dette studiet er det satt en klassifiseringsgrense ut fra to dominerende Calanus-FATM. Torsk som spiser i den pelagiske næringskjeden, har enten et nivå av 20:1 (n-9) på mer enn 5 % eller 22:1 (n-11) på mer enn 4 % av den totale mengden fettsyrer.
Torsk som ikke klassifiseres til de første to gruppene, plasseres i den bentiske næringskjeden. Kriteriet for dette er at bentiske FATM for disse torskene ligger generelt over 10 % av den totale mengden fettsyrer.
Ut fra disse tre kriteriene har vi analysert datamaterialet og testet hvordan laksefôrgruppen skiller seg ut fra de to andre «naturlige» gruppene på fiskelengde, kondisjon (leverindeks og innhold av fett i leveren) og størrelse av gonadene. Statistiske forskjeller er undersøkt med enveis ANOVA og Tukey-Kramer multiple comparison post-hoc tester. Korrelasjonen mellom nivået av terrestrisk FATM i lever og gonade ble analysert for å undersøke i hvilken grad fettsyrer fra dietten overføres til fosfolipider og akkumuleres i gonaden. På samme måte ble fettsyreprofilen av muskelprøver analysert i 2019. FATM-grupperingen av torsk er vist i multivariat form ved hjelp av correspondence analysis (CA) (Meier m.fl. 2016). Signifikansnivå for de statistiske testene er satt til 0,05.
6.2.4 - Genetisk analyse og lesing av otolitter
DNA ble isolert fra 384 torsk ved å bruke HotSHOT-metoden. En meget liten vevsbit fra en torsk ble plassert i en brønn på en 96-brønners plate. Hver brønn ble tilsatt alkalisk løsning (NaOH), og det hele ble varmet ved 95°C i 30 minutter. Deretter ble alle løsningene nøytralisert med Tris-HCl, og 2 µl fra hver brønn ble overført til et nytt brett der hver brønn ble tilsatt en PCR-løsning som inneholder primere for både PanI-A og PanI-B. Fragmentene ble amplifisert i en PCR-maskin og til slutt identifisert i en sekvenseringsmaskin.
Otolittene ble opparbeidet og lest etter prosedyrer beskrevet i kvalitetshåndboken til Havforskningsinstituttet. Otolitten ble knekt i to så nær kjernen som mulig og montert i kitt med bruddflaten opp. Alder ble lest fra soneinndeling under et stereomikroskop med 16 gangers forstørrelse, og bestemt ved å telle antall mørke soner (sommersoner) ved gjennomfallende lys og trekke fra den siste sommersonen. Gytesoner ble identifisert ved at de er tynnere enn de forutgående sonene, samt at vintersonene før og etter gytesonen er mer gjennomskinnelige.
6.3 - Resultater
6.3.1 - Bestandstilhørighet og alderssammensetning
Det meste av innfanget torsk gjennom gytesesongene i 2018 og 2019 på gytefeltene på Smøla var 5 og 6 år gammel fisk (tabell 4). Ut fra lesing av otolitter ble det identifisert noen få skrei blant den innfangede torsken (tabell 4), mens 98,7 og 98,4 % var kysttorsk for henholdsvis 2018 og 2019. Generelt var individene som ble identifisert til skrei, større og eldre enn kysttorsken begge årene. Det samme gjaldt alder ved førstegangs gyting som hadde en medianverdi på 5 år for kysttorsken i 2018 og 6 år i 2019, mens tilsvarende for skrei var 7 og 8 år (tabell 4). Det ble observert et stort spenn i alder ved førstegangs gyting for kysttorsken, tilsvarende 3-10 år for begge årene sett under ett. En betydelig andel hadde ikke gytesone i otolitten, tilsvarende 57,7 og 39,4 % av kysttorsken for henholdsvis 2018 og 2019. Imidlertid var det ubetydelig forskjell i gjennomsnittlig gonadestørrelse mellom torsk med og uten gytegytesone, tilsvarende 6,9 g i 2018 og 28 g i 2019. Dette indikerer at de aller fleste av torskene uten gytesone var førstegangsgytere under innsamlingen av materialet. Alder ved førstegangs gyting kan derfor være noe overestimert i tabell 4.
I 2018 hevdet fiskeren som gjennomførte fangsten, at det var skrei til stede på gytefeltene. Fiskeren merket av 7 av 450 torsk som skrei, og otolittdata viste at 4 av disse også ble karakterisert som skrei ut fra otolittavlesningene. Det ble ikke angitt skrei av fiskeren i 2019. Imidlertid ble det ikke funnet noen skrei fra de genetiske analysene. «Skrei» i tabell 4 må derfor anses å være en kysttorsk som har en utforming av otolittene som kan forveksles med skreien.
Tabell 4. Fiskelengde, aldersfordeling og alder ved første gyting for de ulike bestandskomponentene identifisert fra otolitter (øresteiner) for innfanget torsk på gytefeltene på Smøla gjennom gytesesongen i 2018 og 2019. Fiskelengde er oppgitt med standardavvik. Gytealder er alder ved første gangs gyting.
6.3.2 - Klassifisering av fôrpreferanse
Data fra fettsyreanalysene er gitt i Vedlegg 4 der tabell 4.1 og 4.3 viser resultater fra analyse av lever i 2018 og 2019, tabell 4.2 og 4.4 viser resultater fra analyse av gonader i 2018 og 2019, tabell 4.5 viser resultatene fra analyse av muskel i 2019, og tabell 4.6 viser resultatene fra analysen av de to laksefôrene som ble benyttet på anlegg nær gytefeltene Glasøysvaet og Lauvøysvaet.
En betydelig andel av torsken skilte seg ut ved at de hadde høye nivåer av terrestriske FATM. Omtrent 20 % (17% i 2018 og 23 % i 2019) av torsken fanget på gyteplassene på Smøla ble klassifisert i gruppen som spiser laksefôr ut fra deres fettsyresammensetning i leveren (figur 31A og tabell 5). Det var svært høy korrelasjon (R2 = 0,8, p < 0,0001) mellom de to terrestriske FATM 18:1 (n-9) og 18:2 (n-6) hos torsken som ble klassifisert i gruppen som spiser laksefôr, mens denne korrelasjonen var betydelig lavere blant de resterende torskene (R2 = 0,18, p < 0,0001). I laksefôrgruppen vil det være individer som også spiser i de andre to næringskjedene, noe som gir nivåer av 18:2 (n-6) i leveren på mellom 4 og 10 %, samtidig som andre individer tilsynelatende har spesialisert seg hovedsakelig på å spise laksefôr, noe som gir nivåer av 18:2 (n-6) høyere enn 10 %. I 2018 og 2019 hadde henholdsvis 75 % og 79 % av torsken som ble allokert til laksefôrgruppen mer enn 10 % med 18:2 (n-6) i leverfettet. Nivåene av 18:2 (n-6) i de to laksefôrene som ble analysert, utgjorde til sammenligning 16 % av totalmengden fettsyrer i fôret (figur 31A). Det ble ikke funnet noen statistiske forskjeller mellom gytefeltene i andel av torsk som spiste laksefôr (p > 0,2389).
Tabell 5. Kriterier i prioritert rekkefølge for å plassere en torsk til en diettgruppe. Nivå av trofisk fettsyremarkør (FATM) er angitt som % av total mengde fettsyrer (TFA). Antall torsk i de ulike diettgruppene med tilhørende andel av totalt antall torsk undersøkt de to årene, er gitt til høyre i tabellen.
Figur 31B viser innhold av de to viktigste Calanus-FATM 20:1 (n-9) og 22:1 (n-11) i torskelever, og som brukes til å klassifisere fisk som har høy preferanse for å spise i den pelagiske næringskjeden. Det var svært høy korrelasjon mellom disse to FATM for alle torskene uavhengig av diettgruppering (R2 = 0,9, p < 0,0001), det var ingen klar gruppering ut fra korrelasjonen, men en kontinuerlig lineær korrelasjon. Klassifiseringen med Calanus-FATM av torsk med pelagisk og bentisk diett er derfor overlappende og ikke tydelig adskilt. Med hensyn til de kriterier vi har valgt (tabell 4), så klassifiseres 28 % av torsken i 2018 og 22 % i 2019 til den pelagiske diettgruppen, mens de resterende 55 % (både 2018 og 2019) klassifiseres i den bentiske diettgruppen.
Multivariat correspondence analyse (CA) i figur 32 er en annen måte å vise klassifisering i ulike diettgrupper. I CA brukes alle fettsyrer sammen med fiskedata som lengde, leverindeks (HSI) og gonadeindeks (GSI) for å gruppere prøvene. Dette gjøres ved beskrive dataene med såkalte latente variabler som forklarer dominerende trender i det multivariate rom, og ved å plotte dette langs de to første aksene slik at multivariate trender visualiseres. Prøver som ligger tett på hverandre i scoreplottene er like, mens de som ligger lang fra hverandre har veldig forskjellige fettsyreprofiler.
Score-plottene for 2018 og 2019 viser at prøvene fra begge år ble gruppert på samme måte, og gruppen som spiste laksefôr sammenfalt med de to fôrprøvene. Dette er visualisert i figur 32 der torsk som beiter på laksefôr, er separert fra de to andre diettgruppene langs akse 1 (CA1), mens torsk som spiste i pelagisk eller bentisk næringskjede er separert lang akse 2 (CA2). Loading-plottet viser hvilke variabler som er dominerende i hver av diettgruppene. Her finner vi at terrestriske FATM som 18:1 (n-9), 18:2 (n-6) og 18:3 (n-3) grupperer seg langt til venstre langs CA1, og dette sammenfaller med gruppen som spiser laksefôr. Lengst til høyre finnes DHA, 22:6 (n-3), som indikerer at denne fettsyren finnes i forholdsvis større mengder i torsk som beiter i den bentiske eller pelagiske næringskjeden sammenlignet med laksefôrgruppen.
Figur 31. Trofiske fettsyremarkører (FATM) fra torskelever samlet inn på gytefelt på Smøla i 2018 og 2019. A. Terrestriske FATM; 18:1 (n-9) og 18:2 (n-6), B. Pelagiske FATM; 20:1 (n-9) og 22:1 (n-11), C. Bentiske FATM; 18:1 (n-7) og 20:4 (n-6). Prøvene er klassifisert som bentisk diett (rød trekant), pelagisk diett (grå sirkel) og diett bestående av laksefôr (sorte ruter). Rosa firkanter viser FATM fra to ulike typer laksefôr som er benyttet på oppdrettsanlegg i nærområdet. Blå linjer angir grenser for klassifisering i diettgrupper i tabell 4.
Figur 32. Multivariant correspondance analysis (CA) av 51 fettsyrer fra leverprøver av alle torsk (både 2018 og 2019) sammen med total mengde fettsyrer (TFA), leverindeks (HSI) og gonadeindeks (GSI). Score-plottene viser grupperingen av torsken, og data fra 2018 og 2019 er vist hver for seg for bedre å kunne sammenligne diettgruppene, selv om begge plottene stammer fra sammen analyse. I loading-plottet vises kun de variablene som ligger over gjennomsnittet for loading-verdier og derfor bidrar mest til grupperingen i CA. De to første aksene i CA forklarer 65 % av den totale variansen i datasettet (akse 1 = 44 %, akse 2 = 21 %).
Fettsyrene som separerer prøvene langs CA2, har typiske pelagiske FATM som 14:0, 20:1 (n-9) og 22:1 (n-11) oppover aksen, og bentiske FATM som 20:4 (n-6) og 18:1 (n-7) nedover aksen. Fra fiskedataene ligger både leverindeksen (HSI) og totalmengde fett (TFA) oppe til høyre i diagrammet. Dette viser at 1) laksefôrgruppen har større lever med høyere fettinnhold enn de to andre gruppene, og 2) torsken på pelagisk diett har større lever og høyere fettinnhold i leveren enn torsk på bentisk diett. Som nevnt er klassifiseringen ikke absolutt, men overlappende. Flere torsk har spist i alle tre diettgruppene, mens mange individer tydeligvis så ut til å ha spesialisert seg innenfor en av de tre diettgruppene.
6.3.3 - Fôrpreferanse og påvirkning av torskens kondisjon og energistatus
Gruppen som spiste laksefôr, hadde større lever med høyere fettinnhold, og dermed betydelig høyere energimengde sammenlignet med de to gruppene som spiste en marin diett. I tabell 6 er dette angitt med parameteren Fett-HSI som er total mengde fett i leveren delt på fiskens lengde. I 2018 hadde gruppene med henholdsvis bentisk og pelagisk diett kun 30 og 55 % så mye fett i leveren relativt til fiskelengde sammenlignet med laksefôrgruppen. I 2019 hadde laksefôrgruppen enda bedre energistatus, og nå var energistatus for gruppene med bentisk og pelagisk diett henholdsvis kun 14 og 20 % av nivåene som ble funnet i laksefôrgruppen.
Tabell 6. Oversikt over antall torsk og biologiske data som kjønn, fiskelengde, gonadeindeks (GSI: gonadevekt (g)/fiskelengde), leverindeks (HSI: levervekt (g)/fiskelengde), fettnivå i leveren (% av våtvekt), energistatus gitt som Fett-HSI (levervekt (g) x fett %/fiskelengde) og mengde trofiske fettsyremarkører (FATM som % av mengde fettsyrer totalt) for de tre diettgruppene. Blå farge angir mengde FATM i torsk i en diettgruppe. Statistiske forskjeller innen en kolonne er angitt med ulike små bokstaver (hevet skrift).
Frekvensfordelingen av fettinnhold hos hunn- og hanntorsk fra 2018 og 2019, delt opp på de tre ulike diettgruppene er vist i figur 33. Blant torsken på bentisk diett var det en betydelig andel (10-40 %) som hadde lave fettnivåer (< 20 %) i leveren. Laksefôrgruppene disse to årene hadde generelt mye høyere fettnivåer i leveren, og i 2018 hadde 40-45 % av denne torsken over 40 % fett i leveren, mens i 2019 var laksefôrgruppen i meget god kondisjon der 40-90 % av disse torskene hadde et fettinnhold i leveren på over 50 %. Det er en positiv lineær sammenheng mellom leverstørrelse og fettinnhold opp til en HSI på ca. 3 %, og deretter flater denne trenden ut (figur 34).
6.3.4 - Kjønnsforskjeller og påvirkning av gonadestatus
Der var en skjev fordeling med hensyn til kjønn hos torsken som ble fanget på gyteplassene i dette garnfisket, og hunnfiskene utgjorde i 2018 og 2019 henholdsvis kun 28 og 23 % av den innfangede torsken. Dette kan enten forklares med at det var flere hannfisk som oppholdt seg på gytefeltene, eller at disse hadde en annen atferd som for eksempel at de var mer aktive eller hadde en annen fordeling i vannmassene enn hunnene.
Det var små (2018) eller ingen (2019) forskjeller i fordelingen av kjønn mellom de tre diettgruppene. Derimot var det kjønnsforskjeller med hensyn til energistatus og gonadestørrelse, og disse var like for alle de tre diettgruppene:
Hunnfisken hadde betydelig større gonader sammenlignet med hannfisken.
Hunnfisken hadde større lever enn hannfisken.
Dette viser at hunnfisken har investert mer energi til gonadevekst enn hannfisken, og at hunnfisken har prioritert levervekst (energilagring) høyere enn hannfisken. Gonadeveksten skjer ved at det produseres et lipoprotein (vitellogenin) i leveren, og dette sendes via blodet til gonadene hvor det tas opp i de modnende oocytter. Vitellogenin er sammensatt av protein, fosfolipider og lagringsfett (triacylglycerol) (Silversand & Haux 1995). Hunnfisken hadde lavere relativ fettmengde i leveren (% av vekt) enn hannfisken, men siden de også hadde større lever, så var totalmengden med fett relativ til fiskelengde ikke forskjellig mellom kjønnene.
Det er velkjent at energimengden i leveren (leverstørrelse og fettprosent) er positivt korrelert med eggproduksjon i torsk (Marshall m.fl. 1999; Skjæraasen m.fl. 2006), og det er blitt foreslått at den store økningen i energiopptak og kondisjonsfaktor (leverstørrelse og fettinnhold) som observeres hos gruppen som spiser laksefôr, kan ha en positiv effekt på reproduksjonspotensialet hos torsken og gi en økning i gyteprodukter (Dempster m.fl. 2011).
Figur 35 viser at den store økningen i leverstørrelse (HSI) hos torsken som spiste laksefôr, ikke resulterte i økt gonadestørrelse (GSI) hos hunnfisken (R2 = 0,015, p = 0,45). Derimot var det en lineær korrelasjon mellom HSI og GSI hos hunntorsken fra bentisk (R2 = 0,3, p < 0,0001) og pelagisk (R2 = 0,25, p < 0,002) diettgruppe. For hannfisken var det en positiv korrelasjon mellom HSI og GSI for alle grupper, bentisk (R2 = 0,28, p < 0,0001), pelagisk (R2 = 0,24, p < 0,0001) og laksefôrdiett (R2 = 0,07, p < 0,003). Men det er verd å merke seg at vekst i gonader med økende leverstørrelse hos hannfisken med laksefôrdiett var svært lav sammenlignet med de to andre fôrgruppene.
Figur 33. Innhold av fett i leveren (% av våtvekt) til torsk fanget på gyteplassene ved Smøla i 2018 og 2019. Ulike farger angir fettinnhold, mens andelen av torskene med en gitt fettmengde kan leses av langs y-aksen. Torsken er kategorisert i tre grupper i henhold til profiler av fettsyremarkører (FATM) i leveren: bentisk diett, pelagisk diett og laksefôrdiett.
Figur 34. Sammenheng mellom leverindeks (levervekt/fiskelengde) og fettinnhold i leveren hos torsk fanget på gyteplassene ved Smøla i 2018 og 2019. Torsken er kategorisert i tre grupper i henhold til profiler av fettsyremarkører (FATM) i leveren: bentisk diett, pelagisk diett og laksefôrdiett.
6.3.5 - Terrestriske fettsyremarkører (FATM) i gonader og muskel
De terrestriske fettsyremarkørene som akkumuleres i lagringsfettet i leveren hos torsken som spiser laksefôr, ble overført til fosfolipider og transportert til både muskel og gonader (figur 36 og 37). Det er sterk korrelasjon mellom summen av de tre terrestriske fettsyremarkørene 18:1 (n-9), 18:2 (n-6) og 18:3 (n-3) i lever og gonader (R2 = 0,8, p < 0,0001), all torsk sett under ett (figur 38). Tilsvarende sterk korrelasjon ble også funnet mellom disse fettsyremarkørene i lever og muskel (R2 = 0,9, p < 0,0001). Torsken som ble vurdert å spise laksefôr ut fra FATM i lever, hadde de høyeste nivåene av disse tre fettsyremarkørene i både lever og gonader (figur 38).
Multivariat correspondence analyse (CA) for rogn (ovarie), melke (testis) og muskel i figur 39 viser et tilsvarende bilde som for leverprøvene i figur 32. Langs den første aksen (CA1) separerer laksefôrgruppen seg fra de to gruppene som beiter pelagisk og bentisk med hensyn til de terrestriske FATM. Langs den andre aksen (CA2) separeres pelagisk og bentisk diettgruppe. Loading-plottene i figur 39 viser at den viktigste forskjellen i fettsyreprofil som separerer langs CA2, er at fosfolipidene hos gruppen med bentisk diett har relativ høyere nivåer av ARA, 20:4 (n-6), mens den pelagiske diettgruppen har relativ høyere nivåer av DHA, 22:6 (n-3).
Figur 35. Sammenheng mellom leverindeks (levervekt/fiskelengde) og gonadeindeks (gonadevekt/fiskelengde) hos torsk fanget på gyteplassene ved Smøla i 2018 og 2019. Torsken er kategorisert i tre grupper i henhold til profiler av fettsyremarkører (FATM) i leveren: bentisk diett, pelagisk diett og laksefôrdiett.
Figur 36. Sammenheng mellom de terrestriske fettsyremarkørene 18:1 (n-9) og 18:2 (n-6) i muskelprøver fra hunnfisk og hannfisk i 2019. Torskene er klassifisert til bentisk diett, pelagisk diett og laksefôrdiett ut fra deres innhold av FATM i leveren.
Figur 37. Sammenheng mellom de terrestriske fettsyremarkørene 18:1 (n-9) og 18:2 (n-6) i gonadene hos hunnfisk (ovarie) og hannfisk (testis) i 2018 og 2019. Torskene er klassifisert til bentisk diett, pelagisk diett og laksefôrdiett ut fra deres innhold av FATM i leveren.
6.3.6 - Absolutte mengder av essensielle fettsyrer
Torsken som spiser laksefôr, får svært høye nivåer av terrestriske fettsyrer som 18:1 (n-9), 18:2 (n-6) og 18:3 (n-3). Når fettsyreprofilene uttrykkes som % av total mengde fettsyrer, har disse torskene relative lavere nivåer av de viktige og essensielle lang-kjedede fettsyrene 20:4 (n-6), 22:5 (n-3) og 22:6 (n-3) sammenlignet med de to andre diettgruppene. For å undersøke i hvilken grad dette skyldes «lukke-effekten» (normalisering til 100 % medfører at når noe går opp, må noe annet gå ned), må data uttrykkes i absolutte verdier. I figur 40 vises de absolutte nivåene (% av vevsvekten) av 18:2 (n-6) mot 20:4 (n-6), og 20:5 (n-3) mot 22:6 (n-3) fra lever, ovarie, testis og muskel for torsken fra både 2018 og 2019. I alle disse vevene er 18:2 (n-6) mye høyere i gruppen som har spist laksefôr sammenlignet med de to andre diettgruppene, hvor de absolutte nivåene av denne fettsyren kun utgjorde 5-17 % (beregnet ut fra gjennomsnittsverdier) av nivåene i torsk som spiste laksefôr. Figur 40A viser at 18:2 (n-6) alene i det mest ekstreme tilfellet utgjorde 12,5 % av levervekten for en torsk i laksefôrgruppen, mens 18:2 (n-6) var mindre enn 2 % av levervekten hos torsk i gruppene bentisk og pelagisk diett.
For de tre andre fettsyrene var det stor variasjon innenfor hver diettgruppe, men den bentiske diettgruppen hadde likevel høyere nivåer av 20:4 (n-6) i alle vev sammenlignet med både laksefôrgruppen (134-337 %) og gruppen med pelagisk diett (144-215 %). Videre hadde torsken i den pelagiske diettgruppen høyere nivåer av 22:6 (n-3) i leveren enn laksefôrgruppen (160-224 %), men ikke i gonadene eller muskel. For 20:5 (n-3) var der ingen klare forskjeller mellom de tre diettgruppene.
Figur 38. Sammenheng mellom summen av de terrestriske fettsyremarkørene 18:1 (n-9), 18:2 (n-6) og 18:3 (n-3) i lever og gonader for torsken fra 2018 og 2019. Torskene er klassifisert til bentisk diett, pelagisk diett og laksefôrdiett ut fra deres innhold av FATM i leveren.
6.4 - Diskusjon biokjemiske undersøkelser
Det ble ikke funnet skrei på gytefeltene på Smøla til tross for at avlesninger av otolitter indikerte at skrei kunne forekomme og den lokale fiskeren mente at det var skrei ut fra farge og ytre kjennetegn (morfologi). Bruk av genetiske metoder regnes som den sikreste måten å skille skrei fra kysttorsk. Kysttorsk med otolitter og morfologi som ligner dem man finner hos skrei, er derfor interessant ut fra en teori om at det kan finnes en type kysttorsk som har et levesett som ligner mer på skrei? Det er påvist morfologiske forskjeller mellom kysttorsk og skrei der skreien er slankere, noe som kan være en tilpasning til lengre vandringer slik skreien foretar fra Barentshavet til gyteområdene på Norskekysten (Svåsand m.fl. 1996). Det kan også tenkes at en vandrende kysttorsk som i utstrakt grad benytter større områder på den ytre kysten og bankene med tilliggende havområder som beiteområder, vil kunne fremvise otolitter som ligner mer på skreiens otolitter. Dette er imidlertid en hypotese som må underbygges av forskning. Et annet moment er at 5 av de 6 kysttorskene fra 2018 som ble lest som skrei ut fra otolittene, hadde beitet i den pelagiske næringskjeden. En pelagisk signatur i fettsyreprofilen i leveren er også typisk for skrei (upubliserte data fra skrei fanget i Lofoten).
En betydelig andel (39-58%) av torsken som ble undersøkt på gyteområdene på Smøla, så ut til å være førstegangsgytere. Denne høye andelen kan tyde på at kysttorsken i denne regionen er utsatt for et betydelig fiskepress.
Figur 39. Multivariant correspondance analysis (CA) av 51 fettsyrer fra A) ovarieprøver, B) prøver av testis og C) muskelprøver av torsken. I tillegg er total mengde fettsyrer (TFA), leverindeks (HSI), gonadeindeks (GSI), kjønn og fiskelengde inkludert i analysene. Ovarie og testis omfatter prøver fra torsk i både 2018 og 2019, mens muskelprøvene kun er fra 2019. CA for muskelprøvene er gjort samlet på både hunn- og hannfisk. I loading-plottene vises kun de variablene som ligger over gjennomsnittet for loading-verdier og derfor bidrar mest til grupperingen i CA. Summen av prosentverdier for de to CA-aksene angir andel av den totale variasjonen i datasettet som forklares samlet av CA1 og CA2. Dette utgjør 58, 65 og 67 % for analysen av henholdsvis ovarier, testis og muskel.
Figur 40. Absolutte mengder (mg/100 mg vevsvekt) av 18:2 (n-6) plottet mot 20:4 (n-6), og 20:5 (n-3) plottet mot 22:6 (n-3) i A) lever fra 2018 og 2019, B) ovarie fra 2018 og 2019, C) testis fra 2018 og 2019, og D) muskel fra hunn- og hannfisk fra 2019. Torskene er klassifisert til bentisk diett, pelagisk diett og laksefôrdiett ut fra deres innhold av FATM i leveren.
Det er veldokumentert at vill fisk tiltrekkes til oppdrettsanlegg for å spise fôrspill (Callier m.fl. 2018). Langs norskekysten er det godt dokumentert at særlig sei og torsk samler seg i store mengder og beiter på overskudd av laksefôr under oppdrettsanlegg (Bagdonas m.fl. 2012; Dempster m.fl. 2010; Dempster m.fl. 2009; Skog m.fl. 2003; Uglem m.fl. 2009; Uglem m.fl. 2014). Derimot er det usikkert i hvilken grad denne akkumuleringen av villfisk rundt oppdrettsanlegg har negative effekter på populasjoner av vill fisk langs kysten, eller om det faktisk kan ha en positiv effekt på torskefisk sin reproduksjon gjennom å øke energinivå og kondisjon hos fisken (Barrett m.fl. 2018; Dempster m.fl. 2011).
Undersøkelsene av biokjemisk innhold i torsk fanget på gytefeltene på Smøla tilfører ny kunnskap på flere områder:
Resultatene viser at ca. 20 % av torsken som gyter i de undersøkte områder på Smøla, beiter på laksefôr i større eller mindre grad. Siden torsken er fisket på gytefeltene og ikke i nærheten av fiskeoppdrettsanleggene, så gir dette et mer realistisk bilde på hvor stor en del av gytepopulasjonen som beiter på laksefôr.
Fettsyreanalyser bekrefter tidligere studier om at torsken som beiter på laksefôr, får en tydelig modifisert fettsyreprofil i leveren ved at fisken akkumulerer betydelige mengder med terrestriske fettsyrer i lagrinsfettet i leveren. Resultatene viser også for første gang at disse fettsyrene i høy grad bygges inn i fosfolipider og overføres til muskel og gonader (både ovarier og testis). Dette er i kontrast til tidligere funn som viser at fettsyreprofilen i lever kan være svært variabel innen fiskearter, mens fettsyreprofilen i fosfolipidene som utgjør mesteparten av fettet i gonadene hos torsken, er relativt konstant (Silversand & Haux 1995).
Fettsyreanalysene viser også at torsk som ikke beiter på fôrspill fra lakseoppdrett, kan inndeles i to diettgrupper der den ene hovedsakelig beiter i den pelagiske næringskjeden og den andre beiter i den bentiske næringskjeden.
Torsken som beiter på laksefôr, får 2-7 ganger høyere energinivå der både størrelse av lever og lipidinnhold øker sammenlignet med torsk som beiter bentisk eller pelagisk.
Økningen i energiinnhold hos gruppen som spiser laksefôr, medfører derimot ikke noen økt investering i gonadevekst.
Fôringsstudier har vist at torsk tåler en diet basert på planteproteiner og oljer uten at det påvirker vekst og helsen til fisken (Albrektsen m.fl. 2006; Colburn m.fl. 2012; Hansen & Hemre 2013; Hansen m.fl. 2007a; Hansen m.fl. 2007b; Hansen m.fl. 2006; Karalazos m.fl. 2007; Olsen m.fl. 2007). Det er likevel uklart om disse ikke-marine fôrkildene kan ha negative konsekvenser for vill fisk sin reproduksjon og rekruttering (Gonzalez-Silvera m.fl. 2020). God egg- og spermkvalitet med påfølgende høy overlevelse av larvene er avhengig av optimal stamfiskdiett (Tocher 2010). Eksperimentelle studier på stamfiskdiett hos torsk (Norberg m.fl. 2017; Røjbek m.fl. 2014; Salze m.fl. 2005; Sawanboonchun m.fl. 2008), samt feltstudier på vill torsk (Pickova m.fl. 1997) har vist at høy fekunditet og god overlevelse av torskelarver er positiv korrelert med langkjedet PUFA. Disse forsøkene tyder på at det er spesielt viktig å ha høye nivåer av 20:4 (n-6), men også at forholdstallet mellom 20:4 (n-6) og andre PUFA som for eksempel 20:5 (n-3) har stor betydning.
Alle tidlige studier er gjort med variasjoner av marine fettsyrer, og ingen har undersøkt suksess i reproduksjonen hos torsk som har spist en diett med så høye nivåer og avvikende sammensetning av fettet som det som finnes i laksefôr. Der er derfor behov for nye studier for å undersøke hvordan den sterkt modifiserte fettsyresammensetningen som fremkommer ved å spise laksefôr, påvirker torskens reproduksjonssuksess og larveoverlevelse. Slike studier vil være viktige både for å kunne vurdere eventuelle skadelige reproduksjonseffekter av at villfisk beiter under lakseoppdrettsanlegg.
Et annet moment er den økende interessen for torskeoppdrett, der oppdrettstorsken må konkurrere om de samme fôrressurser som benyttes for lakseoppdrett. Allerede i dag benyttes terrestriske fôrråstoffer til torskeoppdrett, og modifisering av biokjemisk innhold i villfisk som beiter under slike anlegg kan derfor forekomme. Videre er kjønnsmodning hos torsk i oppdrett fremdeles ikke fullt ut kontrollert, og gyting i merd kan også skje (Jørstad m.fl. 2008; Uglem m.fl. 2012). Gyting i merd kan potensielt endre den genetiske sammensetningen hos lokale stammer av torsk, og mulige effekter av terrestriske fôrmidler på utvikling og overlevelse av egg og larver fra gyting i merd er derfor også viktig å få kartlagt for å vurdere om dette kan utgjøre et betydelig miljøproblem.
7 - Konklusjoner
Resultatene fra undersøkelsene med hensyn til de fire delmålene i arbeidspakke 2 (WP2) av ICOD-prosjektet kan kort oppsummeres som følger:
Det ble ikke påvist at eggmengdene på de nærliggende gyteområdene «Lauvøysvaet» og «Glasøysvaet» hadde noen sammenheng med tidspunkt at de to nye oppdrettsanleggene (Nørholmen og Hestøya) ble satt i drift i 2016.
Det ble heller ikke påvist at rekruttering av årsyngel av torsk hadde noen sammenheng med at disse to oppdrettsanleggene var i drift.
Det ble ikke funnet genetiske ulikheter knyttet til geografisk lokasjon hos torsken i noen av de undersøkte gyte- og oppvekstområdene. Genetisk sett ble all undersøkt torsk funnet å være kysttorsk.
Torsk samlet inn gjennom gytesesongen fra utvalgte gyteområder på Smøla kunne deles inn i tre grupper med utgangspunkt i diett, der om lag 20 % at torsken hadde beitet på laksefôr.
Ved å trekke inn resultatene fra arbeidspakke 4 (WP4) i ICOD-prosjektet er det sannsynlig at de svært aktive hydrodynamiske forholdene i regionen undersøkelsene ble gjennomført, med fremherskende strømretning innover Edøyfjorden, vil kunne maskere eventuelle lokale effekter knyttet til lokalisering av oppdrettsanlegg. For eksempel viste simuleringer av eggdrift ved hjelp av hydrodynamiske modeller at egg gytt i områdene vest for Smøla eller enda lengre unna motstrøms, i ulikt omfang kan ende opp på Lauvøysvaet og Glasøysvaet forskjellige år. Videre vil egg gytt på Lauvøysvaet og Glasøysvaet kunne ende opp på andre gytefelt.
Tilbakeholdelse av egg i samme område der de ble gytt (høy retensjon) er en av forutsetningene for at genetiske forskjeller kan etableres. Simuleringene i ICOD-WP4 konkluderte med at det kun var Dromnessundet av de undersøkte gyteområdene som hadde middels grad av retensjon. I tillegg har den landsomfattende kartleggingen av gyteområder (Espeland m.fl. 2013) vist at Blåsværsvaet og områdene herfra og inn mot Vest-Smøla også har middels grad av retensjon og i tillegg høye eggtettheter (https://portal.fiskeridir.no). Blåsværsvaet, som ligger oppstrøms til Lauvøysvaet og Glasøysvaet, vil derfor kunne bidra med egg til de sistnevnte gytefeltene. Alt i alt vurderes det derfor at mangel på høy grad av retensjon i de undersøkte områdene kan forklare fravær av genetisk spesialisering.
Det samme vil være tilfelle med hensyn til rekruttering av årsyngel av torsk. Egg vil drive fritt i de øvre vannlagene 2-3 uker før klekking, mens larvene vil leve fritt i vannmassene opp mot to måneder etter klekking før yngelen søker mot bunn på grunt vann. Mangel på høy retensjon grunnet aktive hydrodynamiske forhold i farvannene rundt Smøla vil derfor med høy sannsynlighet redusere andelen av bunnslått torskeyngel i et oppvekstområde som også er gytt lokalt. Det ble funnet til dels høye eggtettheter i gytefeltene på Smøla, og disse gytefeltene kan derfor være viktige i en regional sammenheng.
Undersøkelsene viste at en betydelig andel av torsken på gytefeltene beiter under oppdrettsanlegg, og at den resterende torsken kan deles inn i to komponenter med annen og ulik diettbakgrunn: pelagisk og bentisk diett. Torsk som beiter under oppdrettsanlegg er kjent fra tidligere (Dempster m.fl. 2010), men det er første gang at andel av slik torsk på gytefelter er kartlagt. Det var ikke mulig å avklare om bentisk og pelagisk diett representerer henholdsvis en stasjonær og en vandrende type kysttorsk, eller om torsk som beiter under oppdrettsanlegg har sitt utspring i torsk med en eller begge disse diettypene. Endringen i fettsyreprofil av lever og gonader hos torsken som følge av at laksefôr i stor grad tilvirkes av fôrråstoffer fra landbruk, kan tenkes å gi biologiske effekter med hensyn til utvikling og overlevelse i tidlige livsstadier, men eventuelle effekter på rekruttering av dette er ikke utforsket. Det anbefales derfor at dette forskningsfeltet gis høy prioritet, siden oppdrett av laks er planlagt økt de neste tiårene, og oppdrett av torsk som også bruker fôrråstoffer fra landbruk, ser ut til å være på rask fremvekst.
Det synes klart at en del torsk tiltrekkes oppdrettsanlegg, og at denne torsken bidrar til gytingen på gytefeltene. Men det kan heller ikke utelukkes at torsk skremmes vekk av oppdrettsanlegg, fordi gytingen ble evaluert ut fra eggmengder påvirket av en svært aktiv hydrodynamikk i denne regionen. Heller ikke er det mulig å vurdere om en eventuell skremmeeffekt kan være knyttet til en bestemt type torsk, som for eksempel kan sees i sammenheng med diettype. Mer forskning er nødvendig for å avklare slike forhold.
Grunnet de hydrodynamiske forholdene er resultatene fra ICOD-WP2 i liten grad egnet til å gi sikre konklusjoner, og i den sammenheng er resultatene ikke overførbare til andre og mer innelukkede fjordsystemer med høyere grad av retensjon.
8 - Referanser
Albrektsen S, Mundheim H & Aksnes A (2006). Growth, feed efficiency, digestibility and nutrient distribution in Atlantic cod (Gadus morhua) fed two different fish meal qualities at three dietary levels of vegetable protein sources. Aquaculture 261: 626-640. https://doi.org/10.1016/j.aquaculture.2006.08.031
Asplin L, Albretsen J, Johnsen IA & Sandvik AD (2020). The hydrodynamic foundation for salmon lice dispersion modelling along the Norwegian coast. Ocean Dynamics. 70:1151-1167. https://doi.org/10.1007/s10236-020-01378-0
Bagdonas K, Humborstad OB & Løkkeborg S (2012). Capture of wild saithe (Pollachius virens) and cod (Gadus morhua) in the vicinity of salmon farms: Three pot types compared. Fisheries Research 134: 1-5. https://doi.org/10.1016/j.fishres.2012.06.020
Barrett LT, Swearer SE, Harboe T, Karlsen Ø, Meier S & Dempster T (2018). Limited evidence for differential reproductive fitness of wild Atlantic cod in areas of high and low salmon farming density. Aquaculture Environment Interactions 10: 369-383. https://doi.org/10.3354/aei00275
Bjørn PA, Uglem I, Kerwath S, Sæther BS & Nilsen R (2009). Spatiotemporal distribution of Atlantic cod (Gadus morhua L.) with intact and blocked olfactory sense during the spawning season in a Norwegian fjord with intensive salmon farming. Aquaculture 286: 36-44. https://doi.org/10.1016/j.aquaculture.2008.08.038
Callier MD, Byron CJ, Bengtson DA, Cranford PJ, Cross SF, Focken U, Jansen HM, Kamermans P, Kiessling A, Landry T, O'Beirn F, Petersson E, Rheault RB, Strand Ø, Sundell K, Svåsand T, Wikfors GH & McKindsey CW (2018). Attraction and repulsion of mobile wild organisms to finfish and shellfish aquaculture: a review. Reviews in Aquaculture 10: 924-949. https://doi.org/10.1111/raq.12208
Colburn HR, Walker AB, Breton TS, Stilwell JM, Sidor IF & Gannam AL (2012). Partial replacement of fishmeal with soybean meal and soy protein concentrate in diets of Atlantic cod. North American Journal of Aquaculture 74: 330-337. https://doi.org/10.1080/15222055.2012.676008
Dahle G, Johansen T, Westgaard J-I, Aglen A & Glover KA (2018). Genetic management of mixed-stock fisheries “real-time”: The case of the largest remaining cod fishery operating in the Atlantic in 2007–2017. Fisheries Research 205: 77-85. https://doi.org/10.1016/j.fishres.2018.04.006
Dalsgaard J, St John M, Kattner G, Müller-Navarra D & Hagen W (2003). Fatty acid trophic markers in the pelagic marine environment. Advances in Marine Biology 46: 225-340. https://doi.org/10.1016/s0065-2881(03)46005-7
Dempster T, Uglem I, Sanchez-Jerez P, Fernandez-Jover D, Bayle-Sempere J, Nilsen R & Bjørn PA (2009). Coastal salmon farms attract large and persistent aggregations of wild fish: an ecosystem effect. Marine Ecology Progress Series 385: 1–14. https://doi.org/10.3354/meps08050
Dempster T, Sanchez-Jerez P, Uglem I & Bjørn PA (2010). Species-specific patterns of aggregation of wild fish around fish farms. Estuarine. Coastal and Shelf Science 86: 271-275. https://doi.org/10.1016/j.ecss.2009.11.007
Dempster T, Sanchez-Jerez P, Fernandez-Jover D, Bayle-Sempere J, Nilsen R & Bjørn PA (2011). Proxy measures of fitness suggest coastal fish farms can act as population sources and not ecological traps for wild gadoid fish. PlosOne 6: 1-9. https://doi.org/10.1371/journal.pone.0015646
Fernandez-Jover D, Martinez-Rubio L, Sanchez-Jerez P, Bayle-Sempere JT, Jimenez JAL, Lopez FJM, Bjørn P-A, Uglem I & Dempster T (2011). Waste feed from coastal fish farms: A trophic subsidy with compositional side-effects for wild gadoids. Estuarine Coastal and Shelf Science 91: 559-568. https://doi.org/10.1016/j.ecss.2010.12.009
Gonzalez-Silvera D, Izquierdo-Gomez D, Sanchez-Jerez P, Elbal MT, Lopez-Jimenez JA, Martinez-Lopez FJ (2020). Influence of aquaculture waste on fatty acid profiles and gonad maturation of wild fish aggregations at fish farms. Marine Environmental Research 156: 104902. https://doi.org/10.1016/j.marenvres.2020.104902
Hansen AC, Rosenlund G, Karlsen R, Olsvik PA & Hemre GI (2006). The inclusion of plant protein in cod diets, its effects on macronutrient digestibility, gut and liver histology and heat shock protein transcription. Aquaculture Research 37: 773-784. https://doi.org/10.1111/j.1365-2109.2006.01490.x
Hansen AC, Karlsen O, Rosenlund G, Rimbach M & Hemre GI (2007a). Dietary plant protein utilization in Atlantic cod, Gadus morhua L. Aquaculture Nutrition 13: 200-215. https://doi.org/10.1111/j.1365-2095.2007.00486.x
Hansen AC, Rosenlund G, Karlsen O, Koppe W & Hemre GI (2007b). Total replacement of fish meal with plant proteins in diets for Atlantic cod (Gadus morhua L.) I - Effects on growth and protein retention. Aquaculture 272: 599-611. https://doi.org/10.1016/j.aquaculture.2007.08.034
Hansen AC & Hemre GI (2013). Effects of replacing fish meal and oil with plant resources in on-growing diets for Atlantic cod Gadus morhua L. Aquaculture Nutrition 19: 641-650. https://doi.org/10.1111/anu.12078
Hansen JO, Berge GM, Hillestad M, Krogdahl Å, Galloway TF, Holm H, Holm J & Ruyter B (2008). Apparent digestion and apparent retention of lipid and fatty acids in Atlantic cod (Gadus morhua) fed increasing dietary lipid levels. Aquaculture 284: 159-166. https://doi.org/10.1016/j.aquaculture.2008.07.043
Jobling M & Leknes O (2010). Cod liver oil: feed oil influences on fatty acid composition. Aquaculture International 18: 223-230. https://doi.org/10.1007/s10499-008-9238-y
Jobling M, Leknes O, Sæther BS & Bendiksen EA (2008). Lipid and fatty acid dynamics in Atlantic cod, Gadus morhua , tissues: Influence of dietary lipid concentrations and feed oil sources. Aquaculture 281: 87-94. https://doi.org/10.1016/j.aquaculture.2008.05.027
Jørstad KE, van der Meeren T, Paulsen OI, Thomsen T, Thorsen A & Svåsand T (2008). "Escapes" of eggs from farmed cod spawning in net pens: Recruitment to wild stocks. Reviews in Fisheries Science 16: 285-295. https://doi.org/10.1080/10641260701678017
Karalazos V, Treasurer J, Cutts CJ, Alderson R, Galloway TF, Albrektsen S, Arnason J, MacDonald N, Pike I & Bell JG (2007). Effects of fish meal replacement with full-fat soy meal on growth and tissue fatty acid composition in Atlantic cod (Gadus morhua). Journal of Agricultural and Food Chemistry 55: 5788-5795. https://doi.org/10.1021/jf0629383
Kelly JR & Scheibling RE (2012). Fatty acids as dietary tracers in benthic food webs. Marine Ecology Progress Series 446: 1-22. https://doi.org/10.3354/meps09559
Lie Ø, Lied E & Lambertsen G (1986). Liver retention of fat and of fatty acids in cod (Gadus morhua) fed different oils. Aquaculture 59: 187-196. https://doi.org/10.1016/0044-8486(86)90003-7
Marshall CT, Yaragina NA, Lambert Y & Kjesbu OS (1999). Total lipid energy as a proxy for total egg production by fish stocks. Nature 402: 288-290. https://doi.org/10.1038/46272
Mateos-Rivera A, Mozfar B, Skern-Mauritzen R, Dahle G, Thorsen A, Sundby S, Wehde H & Krafft BA (2020). Comparison of visual and molecular taxonomic methods to identify ichthyoplankton in the North Sea. Limnol. Oceanogr. Methods 18: 599-605. https://doi.org/10.1002/lom3.10387
Meier S, Falk-Petersen S, Gade-Sorensen LA, Greenacre M, Haug T & Lindstrom U (2016). Fatty acids in common minke whale (Balaenoptera acutorostrata) blubber reflect the feeding area and food selection, but also high endogenous metabolism. Marine Biology Research 12: 221-238. https://doi.org/10.1080/17451000.2015.1118513
Meier S, Mjøs SA, Joensen H & Grahl-Nielsen O (2006). Validation of a one-step extraction/methylation method for determination of fatty acids and cholesterol in marine tissues. Journal of Chromatography A 1104: 291-298. https://doi.org/10.1016/j.chroma.2005.11.045
Munk P & Nielsen JG (2005). Eggs and larvae of North Sea fishes. Biofolia, Rosenoerns Allé 9, Fredriksberg C, Danmark. 215 pp.
Myksvoll MS, Jung K-M, Albretsen J & Sundby S (2014). Modelling dispersal of eggs and quantifying connectivity among Norwegian coastal cod subpopulations. ICES Journal of Marine Science. 71: 957-969. https://doi.org/10.1093/icesjms/fst022
Myksvoll MS, Sandvik AD, Johnsen IA, Skarðhamar J & Albretsen J (2020). The impact of variable physical conditions and future increased aquaculture production on lice infestation pressure and its sustainability in Norway. Aquaculture Environment Interactions 12:193-204. https://www.int-res.com/articles/aei2020/12/q012p193.pdf
Norberg B, Kleppe L, Andersson E, Thorsen A, Rosenlund G & Hamre K (2017). Effects of dietary arachidonic acid on the reproductive physiology of female Atlantic cod (Gadus morhua L.). Gen. Comp. Endocrinol. 250: 21-35. https://doi.org/10.1016/j.ygcen.2017.05.020
Olsen RE, Hansen AC, Rosenlund G, Hemre G-I, Mayhew TM, Knudsen DL, Eroldoğan OT, Myklebust R & Karlsen Ø (2007). Total replacement of fish meal with plant proteins in diets for Atlantic cod (Gadus morhua L.) II - Health aspects. Aquaculture 272: 612-624. https://doi.org/10.1016/j.aquaculture.2007.05.010
Petursdottir H, Gislason A, Falk-Petersen S, Hop H & Svavarsson J (2008). Trophic interactions of the pelagic ecosystem over the Reykjanes Ridge as evaluated by fatty acid and stable isotope analyses. Deep-Sea Research Part II: Topical Studies in Oceanography 55: 83-93. https://doi.org/10.1016/j.dsr2.2007.09.003
Pickova J, Dutta PC, Larsson PO & Kiessling A (1997). Early embryonic cleavage pattern, hatching success, and egg-lipid fatty acid composition: comparison between two cod (Gadus morhua) stocks. Canadian Journal of Fisheries and Aquatic Sciences 54: 2410-2416. https://doi.org/10.1139/f97-148
Røjbek MC, Støttrup JG, Jacobsen C, Tomkiewicz J, Nielsen A & Trippel EA (2014). Effects of dietary fatty acids on the production and quality of eggs and larvae of Atlantic cod (Gadus morhua L.). Aquaculture Nutrition 20: 654-666. https://doi.org/10.1111/anu.12124
Standal IB, Praël A, McEvoy L, Axelson DE & Aursand M (2008). Discrimination of cod liver oil according to Wild/Farmed and geographical origins by GC and C-13 NMR. Journal of The American Oil Chemists Society 85: 105-112. https://doi.org/10.1007/s11746-007-1174-x
Salze G, Tocher DR, Roy WJ & Robertson DA (2005). Egg quality determinants in cod (Gadus morhua L.): egg performance and lipids in eggs from farmed and wild broodstock. Aquaculture Research 36: 1488-1499. https://doi.org/10.1111/j.1365-2109.2005.01367.x
Sawanboonchun J, Roy WJ, Robertson DA & Bell JG (2008). The impact of dietary supplementation with astaxanthin on egg quality in Atlantic cod broodstock (Gadus morhua , L.). Aquaculture 283: 97-101. https://doi.org/10.1016/j.aquaculture.2008.06.024
Silversand C & Haux C (1995). Fatty acid composition of vitellogenin from four teleost species. Journal of Comparative Physiology B-Biochemical Systemic and Environmental Physiology 164: 593-599. https://doi.org/10.1007/BF00389799
Skilbrei OT, Normann E, Meier S & Olsen RE (2015). Use of fatty acid profiles to monitor the escape history of farmed Atlantic salmon. Aquaculture Environment Interactions 7: 1-13. https://doi.org/10.3354/aei00132
Skjæraasen JE, Nilsen T & Kjesbu OS (2006). Timing and determination of potential fecundity in Atlantic cod (Gadus morhua). Canadian Journal of Fisheries and Aquatic Sciences 63: 310-320. https://doi.org/10.1139/f05-218
Skog T-E, Hylland K, Torstensen BE & Berntssen MHG (2003). Salmon farming affects the fatty acid composition and taste of wild saithe Pollachius virens L. Aquaculture Research 34: 999-1007. https://doi.org/10.1046/j.1365-2109.2003.00901.x
Svåsand T, Jørstad KE, Otterå H & Kjesbu OS (1996). Differences in growth performance between Arcto-Norwegian and Norwegian coastal cod reared under identical conditions. Journal of Fish Biology 49: 108-119. https://doi.org/10.1111/j.1095-8649.1996.tb00008.x
Thompson, BM & Riley, JD (1981). Egg and larval development studies in the North Sea cod (Gadus morhua L.). Rapp. P.-v. Reun. Cons. Int. Explor. Mer 178: 553-559.
Uglem I, Dempster T, Bjørn PA, Sanchez-Jerez P & Økland F (2009). High connectivity of salmon farms revealed by aggregation, residence and repeated movements of wild fish among farms. Marine Ecology Progress Series 384: 251-260. https://doi.org/10.3354/meps08001
Uglem I, Knutsen O, Kjesbu OS, Hansen ØJ, Mork J, Bjørn PA, Varne R, Nilsen R, Ellingsen I & Dempster T (2012). Extent and ecological importance of escape through spawning in sea-cages for Atlantic cod. Aquaculture Environment Interactions 3: 35-51. https://doi.org/10.3354/aei00049
Uglem I, Karlsen Ø, Sanchez-Jerez P & Sæther BS (2014). Impacts of wild fishes attracted to open-cage salmonid farms in Norway. Aquaculture Environment Interactions 6: 91-103. https://doi.org/10.3354/aei00112
Uglem I, Toledo-Guedes K, Sanchez-Jerez P, Ulvan EM, Evensen T & Sæther BS (2020). Does waste feed from salmon farming affect the quality of saithe (Pollachius virens L.) attracted to fish farms? Aquaculture Research 51: 1720-1730. https://doi.org/10.1111/are.14519
Wasta Z & Mjøs SA (2013). A database of chromatographic properties and mass spectra of fatty acid methyl esters from omega-3 products. Journal of Chromatography A 1299: 94-102. https://doi.org/10.1016/j.chroma.2013.05.056
Ytrestøyl T, Aas TS & Åsgård T (2015). Utilisation of feed resources in production of Atlantic salmon (Salmo salar) in Norway. Aquaculture 448: 365-374. https://doi.org/10.1016/j.aquaculture.2015.06.023
Aas TS, Ytrestøyl T, Åsgård T (2019). Utilization of feed resources in the production of Atlantic salmon (Salmo salar) in Norway: An update for 2016. Aquaculture Reports 15: 100216 https://doi.org/10.1016/j.aqrep.2019.100216
9 - Vedlegg 1, Posisjoner for eggstasjoner
Tabell 1.1. Posisjon og bunndyp for valgte eggstasjoner i hovedundersøkelsesområdet på Smøla. Stasjoner med hydrografiprøve er også angitt.
Tabell 1.2. Posisjon og bunndyp for valgte eggstasjoner på referanseområdene i Aure. Stasjoner med hydrografiprøve er også angitt.
10 - Vedlegg 2, Eggdata 2019
Figur 2.1. Eggdata fra undersøkelsene av de to gyteområdene ved Edøya (LAS: Lauvøysvaet og GLS: Glasøysvaet) og av referanseområdet i Åkvika. Data vises for de tre undersøkelsene i 2019. Stasjonene 1 til 11 (ytre Lauvøysvaet) representerer nærhet til det nye oppdrettsanlegget ved Nørholmen som ble satt i drift mai 2016. Figurene angir mulige torskeegg (1,2-1,6 mm i diameter) og utviklingsstadium av eggene, der Stadium 1 er de yngste eggene.
Figur 2.2. Eggdata fra undersøkelsene i referanseområdene Araneset og Dromnessundet. Data vises for de tre eggtoktene i 2019. Figurene angir mulige torskeegg (1,2-1,6 mm i diameter) og utviklingsstadium av eggene, der Stadium 1 er de yngste eggene.
Figur 2.3. Eggdata fra undersøkelsene hovedområdet (Lauvøysvaet og Glasøysvaet) og i referanseområdet Åkvika i 2019. Tall i sirklene viser gjennomsnittlig antall egg pr. håvtrekk på en stasjon for de tre undersøkelsene i 2019, mens farge i sirkelen angir hvor stor andel av eggene som var egg i Stadium 1. Brun skravering angir gyteområde pr. 1. mars 2016. Kilde: https://www.fiskeridir.no
Figur 2.4. Eggdata fra undersøkelsene i referanseområdene Araneset og Dromnessundet i 2019. Tall i sirklene viser gjennomsnittlig antall egg pr. håvtrekk på en stasjon for de tre undersøkelsene i 2019, mens farge i sirkelen angir hvor stor andel av eggene som var egg i Stadium 1. Brun skravering angir gyteområde pr. 1. mars 2016. Kilde: https://www.fiskeridir.no
11 - Vedlegg 3, Rusefiske 2019
Tabell 3.1. Oversikt over rusesett, posisjoner og fangst av torsk ved Glasøysvaet, 2019.Tabell 3.2a. Oversikt over rusesett, posisjoner og fangst av torsk ved Lauvøysvaet, 2019.Tabell 3.2b. Oversikt over rusesett, posisjoner og fangst av torsk ved Lauvøysvaet, 2019.Tabell 3.3. Oversikt over rusesett, posisjoner og fangst av torsk ved Araneset, 2019.Tabell 3.4. Oversikt over rusesett, posisjoner og fangst av torsk ved Åkvika, 2019.Tabell 3.5. Oversikt over rusesett, posisjoner og fangst av torsk ved Dromnessundet, 2019.Tabell 3.6. Fangst av andre arter i rusefisket i 2019.Figur 3.1. Hovedundersøkelsesområdet (Lauvøysvaet og Glasøysvaet) nord for Edøy. Kartet viser Fiskeridirektoratets angivelse av gyteområde (brun skravering) i mars 2016 og stasjonsnett for åleruser i 2019 (hvite piler). Alfanumeriske koder angir hvert enkelt rusesett (tall er rusenummer, og bokstaver fra A til I angir rekkefølgen fra 1 til 6 ganger at en ruse er satt på en posisjon under fisket i 2019). Oppdrettsanlegg er vist i gult (nye anlegg) eller rødt (gamle anlegg).
Figur 3.2. Referanseområdene Araneset (øverst) og Åkvika (nederst). Kartet viser Fiskeridirektoratets angivelse av gyteområde (brun skravering) og stasjonsnett for åleruser i 2019 (hvite piler). Alfanumeriske koder angir hvert enkelt rusesett der tall er rusenummer og bokstaver fra A til F angir rekkefølge fra 1 til 5 ganger at en ruse er satt på en posisjon).
Figur 3.3. Referanseområdet ved Dromnessundet. Kartet viser Fiskeridirektoratets angivelse av gyteområde (brun skravering) og stasjonsnett for åleruser i 2019 (hvite piler). Alfanumeriske koder angir hvert enkelt rusesett (tall er rusenummer, og bokstav fra A til E angir rekkefølgen fra 2 til 3 ganger at en ruse er satt på en posisjon).
12 - Vedlegg 4, Tabeller fettsyreanalyser
Tabell 4.1. Fiskedata og fettsyreprofiler i leveren fra torsk (hunn- og hannfisk) fanget på gytefeltene på Smøla i 2018. Prøvene er klassifisert ut fra deres FATM i leveren i tre grupper: bentisk diett, pelagisk diett og laksefôrdiett.
Tabell 4.2. Fiskedata og fettsyreprofiler i gonadene fra torsk (hunn- og hannfisk) fanget på gytefeltene på Smøla i 2018. Prøvene er klassifisert ut fra deres FATM i leveren i tre grupper: bentisk diett, pelagisk diett og laksefôrdiett.Tabell 4.3. Fiskedata og fettsyreprofiler i leveren fra torsk (hunn- og hannfisk) fanget på gytefeltene på Smøla i 2019. Prøvene er klassifisert ut fra deres FATM i leveren i tre grupper: bentisk diett, pelagisk diett og laksefôrdiett.Tabell 4.4. Fiskedata og fettsyreprofiler i gonadene fra torsk (hunn- og hannfisk) fanget på gytefeltene på Smøla i 2019. Prøvene er klassifisert ut fra deres FATM i leveren i tre grupper: bentisk diett, pelagisk diett og laksefôrdiett.Tabell 4.5. Fiskedata og fettsyreprofiler i muskel fra torsk (hunn- og hannfisk) fanget på gytefeltene på Smøla i 2019. Prøvene er klassifisert ut fra deres FATM i leveren i tre grupper: bentisk diett, pelagisk diett og laksefôrdiett.Tabell 4.6. Fettsyreprofiler i laksefôr brukt ved to nærliggende oppdrettsanlegg til gyteområdene Glasøysvaet og Lauvøysvaet på Smøla.