Potential effects of offshore wind farms on the marine environment
In this report the current knowledge on potential impacts of wind farms on the marine ecosystem is summarized, with a focus on updated knowledge since the previous report from 2008. Potential physical changes are described in Chapter 2, and the potential impacts on the marine ecosystem are divided in potential effect of the introduction of new structures (Ch 3), Potential effects of electromagnetism from the cables (Ch 4) and potential effects of sound produced during construction and operation of the wind turbines (Ch 5). Conclusions can be found in Chapter 6 and IMR’s current advice in Chapter 7.
Potential physical changes include changes in ocean currents and upwelling- and down-welling patterns, as well as increased ambient noise-levels and electromagnetism from the cables that transport electricity to and from the wind farm. Noise-levels will be louder in the construction phase, in particular during the construction of bottom-mounted turbines. During the construction phase, noise-levels will be lower, but the ambient noise level in the area will be increased continuously when the turbines are in operation.
There are clear indications that bottom-mounted wind farms attract several species of underwater animals and that many of these species reproduce and forage successfully within the wind farm areas. The structures can function as an artificial reef and provide increased food availability and shelter for fish after bottom fauna has established on the substrate. The distribution of planktonic organisms could also be affected by the structures due to changes in water movements such as currents. It is yet unknown whether floating wind turbines will have similar effects as bottom-mounted turbines, and what the combined effects of several wind farms will be.
Subsea cables from wind turbines produce static and time-varying electromagnetic fields. Many animals use electromagnetism for orientation and movement behavior. Therefore, it is likely that migration and establishment settlement of such species will be affected by changes in the electromagnetic field around cables that transfer the energy from the turbines. Impacts are expected to be greater for benthic species, but effects on transient planktonic organisms cannot be excluded. However, little is currently known on the scale and magnitude of such effects.
The sound associated with piledriving during the construction phase can damage hearing and cause animals to avoid the area, but mitigation measures, such as bubble curtains, can significantly decrease sound exposure. Research on the effects of wind farms on marine organisms and ecosystems has mainly focused on the construction phase. However, even for this phase it is still unclear whether effects that haven been reported on individual animals will translate to population-level effects.
During the operational phase, the turbines produce continuous low-frequency sound. Continuous low-frequency sound can affect important behaviours, such as sediment mixing, foraging, anti-predator behaviour, reproduction and communication. However, the magnitude of these negative impacts compared to positive effects such as the increased availability of food and shelter availability, is unknown. There are also significant knowledge gaps on the potential for habituation of marine organisms to noise exposure over extended periods of time.
We conclude that with the state of current knowledge, it is impossible to predict whether the total impact of offshore wind farms on underwater life will be positive or negative. In addition to the effects that have been mentioned before, invasive species could use the wind farms as opportunities for establishment. In addition, we know little about how changes in light properties and shadow flickering will affect animals within the wind farms. To document the effects a wind farms an approach that combines controlled experiments and large-scale field studies is needed. Controlled experiments can provide insights on the response of marine organisms to different wind farm-generated stressors and establish causality. Field studies before and after construction can provide insights on impacts in situ for a specific area. It will be important to test whether such effects are local or whether they can lead to population-level effects. The baseline study for such comparisons will need to run over a period that is long enough to document inter-annual variation. In addition to studies during the operation of the wind farm, studies are needed after decommissioning, to assess reversibility of the intervention. A standardized research protocol that is widely accepted is needed to rigorously compare impacts between different wind farms in Norwegian waters.
Recommendations:
IMR recommends the development of a standardised protocol for wind farms adjusted to the Norwegian circumstances. The protocol should contain substantial baseline studies of the areas before construction, as well as monitoring of physical and biological changes in the area during the operational phase and after decommissioning. Due to large variations in pelagic stocks, baseline should studies be conducted for a minimum three year’s period. This protocol should also describe how the data will be openly available.
IMR advices against the construction of wind farms in areas that are of special importance for certain species, such as important spawning areas and migration routes for fish stocks and important feeding and casting areas for certain species of marine mammals.
IMR advices against the construction of wind farms in especially sensitive or valuable areas.
IMR recommends that the approval of concessions is accompanied with clear guidelines for decommissioning after the end of the operational phase.
IMR recommends the use of noise mitigation measures such as bubble curtains during construction and that construction is avoided during fish spawning periods and feeding and casting period for marine mammals if construction is planned near areas where these biological activities take place.
IMR recommends the use of materials in the anchoring of floating turbines that produce least sound during movement of the turbines, e.g. to avoid jerking of anchor chains.
IMR recommends the collection of data to test whether and how wind turbines change the physical environment, such as currents and the acoustic soundscape. Collected data should be openly available.
IMR recommends the collection of baseline data in the areas of planned wind farms before construction. Collected data should be openly available.
IMR recommends the collection of data to test whether and how wind turbines change the biological environment, such as species composition and behaviour. Collected data should be openly available.
Sammendrag
I denne rapporten er dagens kunnskap om potensielle effekter av vindkraftanlegg på det marine økosystemet oppsummert, med fokus på oppdatert kunnskap siden forrige rapport fra 2008. Potensielle fysiske endringer er beskrevet i kapittel 2, og potensielle effekter på det marine økosystemet er delt inn i potensiell effekt av innføring av nye strukturer (kap. 3), elektromagnetisme fra kablene (kap. 4) og lyd produsert under konstruksjon og drift av vindturbinene (kap. 5). Konklusjoner finnes i kapittel 6, og Havforskningsinstituttets nåværende råd i kapittel 7.
Potensielle fysiske endringer inkluderer endringer i havstrømmer og mønstre for opp- og nedstrømning, samt økte støynivåer og elektromagnetisme fra kablene som transporterer strøm til og fra vindkraftanlegg. Støynivået vil være høyere i byggefasen, spesielt under bygging av bunnmonterte turbiner. Under driftsfasen vil støynivåene være lavere, men støynivået i området økes kontinuerlig når turbinene er i drift.
Det er tydelige indikasjoner på at bunnmonterte vindkraftanlegg tiltrekker seg flere marine arter, og at mange av disse artene reproduserer seg og skaffer føde i vindkraftanleggets område. Strukturene kan fungere som et kunstig rev og gi økt mattilgjengelighet og ly for fisk etter at bunnfaunaen er etablert. Distribusjonen av plankton kan også bli påvirket av strukturene på grunn av endringer i vannbevegelser som strømmer. Det er ennå ukjent om flytende vindturbiner vil ha lignende effekter som bunnmonterte turbiner, og hva de samlede effektene av flere vindkraftanlegg vil være.
Undersjøiske kabler fra vindturbiner produserer statiske og tidsvarierende elektromagnetiske felt. Mange dyr bruker elektromagnetisme til orientering og bevegelsesatferd. Derfor er det sannsynlig at migrasjon og etablering av slike arter vil bli påvirket av endringer i det elektromagnetiske feltet rundt kabler som overfører energien fra turbinene. Virkninger forventes å være større for bentiske arter, men effekter på forbigående planktoniske organismer kan ikke utelukkes. Imidlertid er omfanget og størrelsen av slike effekter foreløpig lite kjent.
Lyden fra påling i byggefasen kan skade hørselen til dyr eller føre til at de unngår området, men avbøtende tiltak, som boblegardiner, kan redusere lydeksponeringen betydelig. Forskning på effekter av vindkraftanlegg på marine organismer og økosystemer har hovedsakelig fokusert på byggefasen. Men selv for denne fasen er det fortsatt uklart om effekter som har blitt rapportert på individuelle dyr lar seg overføre til populasjonsnivåeffekter.
I løpet av driftsfasen produserer turbinene kontinuerlig lavfrekvent lyd. Kontinuerlig lavfrekvent lyd kan påvirke viktig atferd, for eksempel sedimentblanding, beiting, forsvarsadferd, reproduksjon og kommunikasjon. Imidlertid er omfanget av disse negative effektene, sammenlignet med positive effekter som økt tilgjengelighet av mat og ly, ukjent. Det er også betydelige kunnskapshull på potensialet for at marine organismer tilvennes støyeksponering over lengre tidsperioder.
Vi konkluderer med at med dagens kunnskap er det umulig å forutsi om den samlede effekten av havvindanlegg på livet under vann vil være positiv eller negativ. I tillegg til effektene som er nevnt tidligere, kan invaderende arter bruke vindkraftanleggene som muligheter for etablering. Vi vet også lite om hvordan endringer i lys og skyggeflimmer vil påvirke dyr i vindkraftanleggene. For å dokumentere effektene av et vindkraftanlegg er det behov for en tilnærming som kombinerer kontrollerte eksperimenter og store feltstudier. Kontrollerte eksperimenter kan gi innsikt i responsen fra marine organismer til forskjellige vindkraftgenererte stressfaktorer og etablere kausalitet. Feltstudier før og etter bygging kan gi innsikt i påvirkninger «in situ» for et bestemt område.
Det vil være viktig å teste om slike effekter er lokale eller om de kan føre til effekter på befolkningsnivå. Forundersøkelser for slike sammenligninger må løpe over en periode som er lang nok til å dokumentere variasjon mellom år. I tillegg til studier av vindkraftanlegg under drift, er det behov for studier etter avvikling for å vurdere om inngrepet lar seg reversere. Det er nødvendig med en standardisert forskningsprotokoll som er allment akseptert for å sammenligne virkningene mellom forskjellige vindkraftanlegg i norske farvann.
Råd:
HI anbefaler at det utformes en standardisert protokoll for havvindanlegg tilpasset norske forhold, som innbefatter grundige forundersøkelser av områdene før utbygging, samt overvåking av både fysiske og biologiske endringer under drift og etter avvikling av anleggene. På grunn av store svingninger i tettheter til pelagiske arter skal forundersøkelsene foregår i minst tre år. Protokollen bør også beskrive hvordan data bør gjøres åpent tilgjengelig.
HI fraråder vindkraftutbygging i områder som er spesielt viktig for enkelte arter, som gyteområder og migrasjonsruter for aktuelle fiskebestander og beite- og kaste områder for enkelte sjøpattedyrarter.
HI fraråder vindkraftutbygging i spesielt sårbare eller verdifulle områder.
HI anbefaler at det ved utsteding av havvindkonsesjoner legges fram tydelige krav om fjerning av strukturene etter avsluttet drift.
HI anbefaler bruk av støydempende tiltak som boblegardin under utbygging, og at man unngår utbyggingsarbeid i gyteperioder for fisk, samt beite og kasteperioder for sjøpattedyr, i og nær områder dette gjelder.
HI anbefaler bruk av materialer i fortøyning av flytende vindturbiner som lager minst mulig støy ved bevegelser av turbinene, f.e unngå rykking og napping i ankerkjettingen.
HI anbefaler innsamling av data for å undersøke i hvilken grad, vindkraftanlegg endrer fysiske forhold som f.eks. strømmønster og lydbilde. Innsamlete data bør være åpent tilgjengelig.
HI anbefaler at det gjennomføres grundige forundersøkelser for å lage en oppdatert oversikt over artene i influensområdene før utbygging. Innsamlete data bør være åpent tilgjengelig.
HI anbefaler innsamling av data for å undersøke i hvilken grad, vindkraftanlegg endrer biologiske forhold som f.eks. artssammenstilling og adferd. Innsamlete data bør være åpent tilgjengelig.
1 - Introduksjon
1.1 - Bakgrunn
Miljøvennlig produksjon av energi er blitt langt viktigere de senere år, og vindturbiner er en av de kraftproduserende metoder som etter hvert har fått stor, internasjonal utbredelse. Foreløpig plasseres de fleste vindturbiner på land, men i løpet av de senere år har det blitt mer vanlig å plassere vindturbiner også i havområder. Havbaserte vindkraftanlegg er allerede i drift langs kysten av flere europeiske land, og i Norge har regjeringen foreslått å åpne spesifikke havområder for vindkraftproduksjon (NVE 2010, 2012, https://www.nve.no/energiforsyning/kraftproduksjon/ vindkraft/vindkraft-til-havs/). Selv om slike vindkraftanlegg representerer en mer miljøvennlig produksjonsform for energi enn f.eks. kraftverk basert på fossile energikilder, er det samtidig viktig å være klar over at også vindkraftanlegg vil påvirke det marine miljø. Vindkraftanleggene vil legge beslag på sjøareal og påvirke strømforhold, samt produsere lyd og elektromagnetisk stråling som vil kunne påvirke marine økosystemer og ressursutnyttelsen av disse. For at havbaserte vindkraftanlegg skal kunne produsere best mulig miljøvennlig energi er det derfor viktig å utrede og redusere eventuelle utilsiktede konsekvenser denne type anlegg vil kunne påføre det marine miljø.
Det har tidligere vært gjennomført undersøkelser om miljøpåvirkning av havvindanlegg i bl.a Belgia (Degraer et al. 2018), Danmark (Brandt et al. 2011, Danish Energy Agency 2013), Nederland (Lindeboom et al. 2011, Coolen et al. 2018), Storbritannia (Bailey et al. 2014), Sverige (Tougaard & Michaelsen 2018) og Tyskland (Krone et al. 2017, Rose et al. 2019). Her kommer det fram at begroing på turbinfundamentene har en tydelig positiv effekt på forekomst av alger og bunndyr, og at mange fiskearter blir tiltrukket til anleggene. Hvorvidt dette har kun lokale effekter eller effekt på populasjonsnivå er imidlertid usikkert. Som negative effekter blir det trukket fram at nye habitater som vindturbinfundamentene representerer muligens kan bli brukt som springbrett for spredning av fremmede arter (Adams et al. 2014, Kerckhof et al. 2015). I tillegg kan lyd (støy) fra turbiner, elektromagnetisk stråling fra kablene, og lysrefleksjoner fra bladene påvirke artene i og rundt havvindanlegg (e.g. Wahlberg og Westerberg 2005, Hutchison et al. 2019). Kunnskapen ervervet gjennom disse studiene er imidlertid ikke nødvendigvis direkte overførbar til områder langs norskekysten, da effektene av slike anlegg vil være stedsspesifikke og avhenge av lokale miljøforhold (Kaldellis et al. 2016). Det finnes, for eksempel, indikasjoner på at fisk blir tiltrukket sterkere i dypere vann (Methratta & Dardick 2019). I norske farvann er det planlagt en satsing på flytende turbiner framfor bunnfaste turbiner. Det er foreløpig ingen kunnskap om miljøeffekter av flytende vindturbiner og om disse er sammenlignbare med effekter av bunnfaste turbiner. For havvindkonsesjoner bør det derfor gjennomføres prosjektspesifikke evalueringer av mulige effekter på miljø og etableres overvåkingsprogram med relevante før- og etterundersøkelser for å skaffe et godt grunnlag om anleggenes påvirkning på marine organismer, både i anleggs- og driftsfasen. Kartlegging av bunntilknyttede organismesamfunn bør gjennomføres i de aktuelle områdene dersom slik informasjon mangler, og anleggene bør i minst mulig grad lokaliseres i områder med forekomster av viktige og sårbare ressurser og naturtyper, inkludert konsentrerte gyteområder og vandringsruter til økonomisk og økologisk viktige fiskebestander (Sivle et al. 2020).
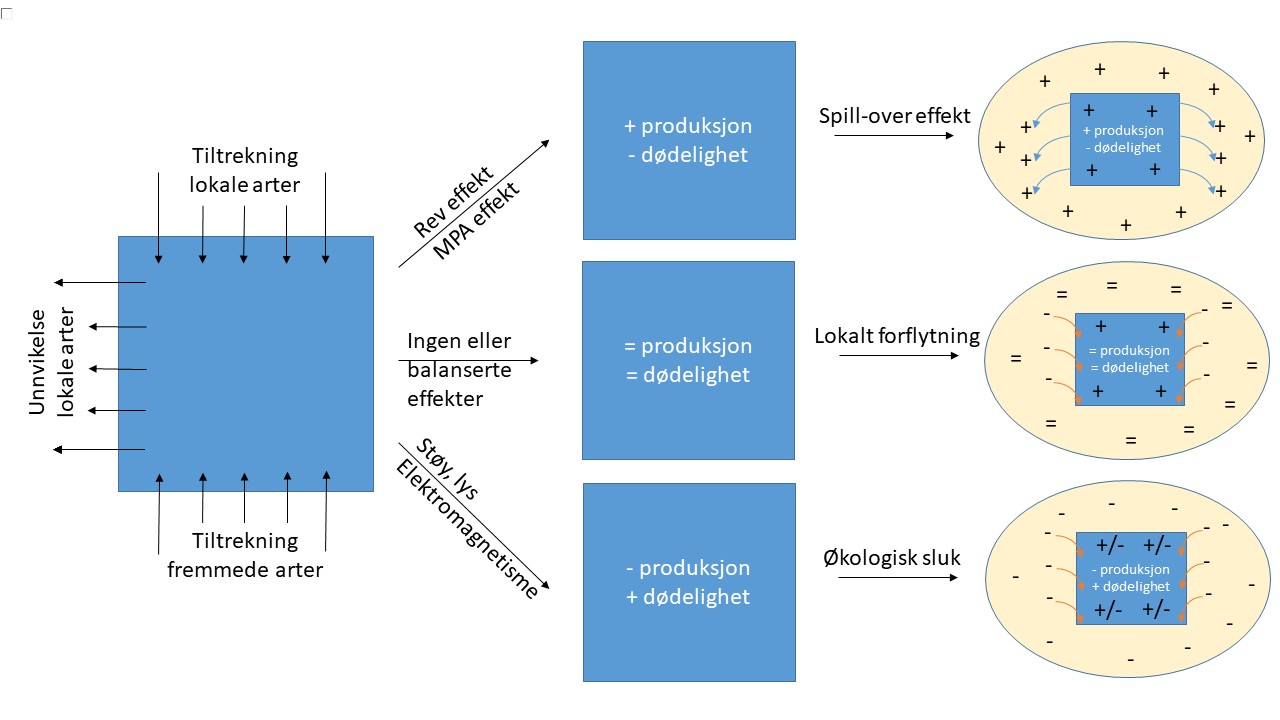
Selv om flere studier på effekter av havvindanlegg har vist tydelige lokale effekter på flere arter, er det fortsatt ikke kjent om effekten påvirker arter på populasjonsnivå (Thomsen et al. 2016; Copping og Hemery 2020). Dersom en art blir tiltrukket av havvindanleggene, vil den relative bestandsstørrelsen innenfor anleggets påvirkningssone øke, og derfor vil populasjonseffekter kunne bli større enn forventet ut ifra anleggets arealbeslag. Innenfor et anlegg kan “kunstig rev effekten” føre til økt tilgang på skjulesteder og mat rundt vindturbiner, mens mindre tilgjengelighet for fiskeri kan føre til redusert dødelighet for fisk. På den andre siden kan endring i habitatet, lybilde og elektromagnetisme også ha negative effekter. Hvis positive effekter fører til at populasjonen rundt et anlegg blir tilført dyr fra anleggssonen kalles det for en “spill-over effekt”. Hvis negative effekter fører til mindre reproduksjon og eller økt dødelighet innenfor et anlegg som dyrene blir tiltrukket til blir området kalt for et “økologisk sluk”. For å få oversikt over de reelle effektene av havvind er det viktig å se effektene i sammenheng og trekke konklusjoner på populasjonsnivå. Uten kunnskap på populasjonsnivå er det ikke mulig å konkludere om effektene er positive, nøytrale eller negative (Figur 1). I denne rapporten skal vi oppsummere kunnskapen siden den forrige rapporten (Steen et al. 2008) om effekter som kan føre til endringer på populasjonsnivå for bunndyr, zooplankton, fisk og sjøpattedyr. Effekter på sjøfugl ligger utenfor Havforskningsinstituttets ekspertise og er derfor ikke nevnt.
Figur 1: Skjematisk skisse av tre mulige effekter av havvindanlegg på populasjonsnivå. Merk at bare de tre mest ekstreme er avbildet og det kan forekomme kombinasjoner eller mellomformer av effektene.
Referanser
Adams TP, Miller RG, Aleynik D, Burrows MT. 2014. Offshore marine renewable energy devices as stepping stones across biogeographical boundaries. Journal of Applied Ecology 51: 330–338.
Bailey H, Brooks KL, Thompson PM. 2014. Assessing environmental impacts of offshore wind farms: lessons learned and recommendations for the future. 2014, 10:8.
Brandt MJ, Diederichs A, Betke K, Nehls G. 2011. Responses of harbor porpoises to pile driving at the Horns Rev II offshore wind farm in the Danish North Sea. Marine Ecology Progress Series 421: 205-216.
Coolen JWP, van der Weide B, Cuperus J, Blomberg M, Van Moorsel GWNM, Faasse MA, Bos OG, Degraer S, Lindeboom HJ. 2018. Benthic biodiversity on old platforms, young wind farms, and rocky reefs. ICES Journal of Marine Science, fsy092, https://doi.org/10.1093/icesjms/fsy092.
Copping AE, Hemery LG. (2020) OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development Around the World. Report for Ocean Energy Systems (OES).
Danish Energy Agency. 2013. Danish Offshore Wind. Key Environmental Issues – a Follow-up. The Environmental Group: The Danish Energy Agency, The Danish Nature Agency, DONG Energy and Vattenfall. 101pp.
Degraer S, Brabant R, Rumes B, Vigin L (eds). 2018. Environmental Impacts of Offshore Wind Farms in the Belgian Parts of the North Sea: Assessing and Managing Effect Spheres of Influence. Brussels: Royal Belgian Institute of Natural Sciences, OD Natural Environment, Marine Ecology and Management. 136pp.
Hutchison ZL, Sigray P, He H, Gill AB, King J, Gibson C. (2019) Electromagnetic Field (EMF) Impacts on Elasmobranch (shark, rays and skates) and American Lobster Movement and Migration from Direct Current Cables. Sterling (VA): U.S. Department of the Interior, Bureau of Ocean Energy Management. OCS Study BOEM 2018-003. 254pp.
Kaldellis JK, Apostelou D, Kapsali M, Kondili E. 2016. Environmental and social footprint of offshore wind energy. Comparison with onshore counterpart. Renewable Energy 92: 543-556.
Kerckhof F, Degraer S, Norro A, Rumes B. 2015. Succession and seasonal dynamics of the epifauna community on offshore wind farm foundations and their role as stepping stones for non-indigenous species. Hydrobiologia 756: 37–50.
Krone R, Dederer G, Kanstinger P, Krämer P, Schneider C, Schmalenbach I. 2017. Mobile demersal megafauna at common offshore wind turbine foundations in the German Bight (North Sea) two years after deployment - increased production rate of Cancer pagurus. Marine Environmental Research 123: 53-61.
Lindeboom HJ, Kouwenhoven HJ, Bergman MJN, Bouma S, Brasseur SMJM, Daan R, Dirken S, van Hal R, Hille Ris Lambers R, ter Hofstede R, Leopold MF, Scheidat M. 2011. Short-term ecological effects of an offshore wind farm in the Dutch coastal zone; a compilation. Environmental Research Letters 6. https://iopscience.iop.org/article/10.1088/1748-9326/6/3/035101.
Methratta E T, Dardick W R. 2019. Meta-Analysis of Finfish Abundance at Offshore Wind Farms. Reviews in Fisheries Science & Aquaculture 27:242–260. DOI: 10.1080/23308249.2019.1584601.
Rose A, Brandt MJ, Vilela R, Diederichs A, Schubert A, Kosarev V, Nehls G, Volkenandt M, Wahl V, Michalik A, Wendeln H, Freund A, Ketzer C, Limmer B, Laczny M, Piper W. 2019. Effects of noise-mitigated offshore pile driving on harbor porpoise abundance in the German Bight 2014-2016 (Gescha 2). Assessment of Noise Effects. 193pp.
Steen H, Nilssen KT, Agnalt A-L, Alvsvåg J, Asplin L, Jelmert A, Dahl E, Dalen J. (2008). Marinøkologiske ringvirkninger av vindkraftanlegger til havs. Fisken og Havet 9-2008: 20 pp.
Thomsen F, Gill A, B, Kosecka M, Andersson M, André M, Degraer S, Folegot T, Gabriel J, Judd A, Neumann T, Norro A, Risch D, Sigray P, Wood D, and Wilson B. (2016) MaRVEN – Environmental Impacts of Noise Vibrations and Electromagnetic Emissions from Marine Renewable Energy (RTD-KI-NA-27738-EN-N), European Commission, Brussels.
Tougaard J, Michaelsen M. 2018. Effects of larger turbines for the offshore wind farm at Krieger’s Flak, Sweden. Assessment of impact on marine mammals. Aarhus University, DCE – Danish Centre for Environment and Energy, Scientific Report No. 286. 112pp.
Wahlberg M. Westerberg H. 2005. Hearing in fish and their reactions to sounds from offshore wind farms. Marine Ecology Progress Series 288: 295-309.
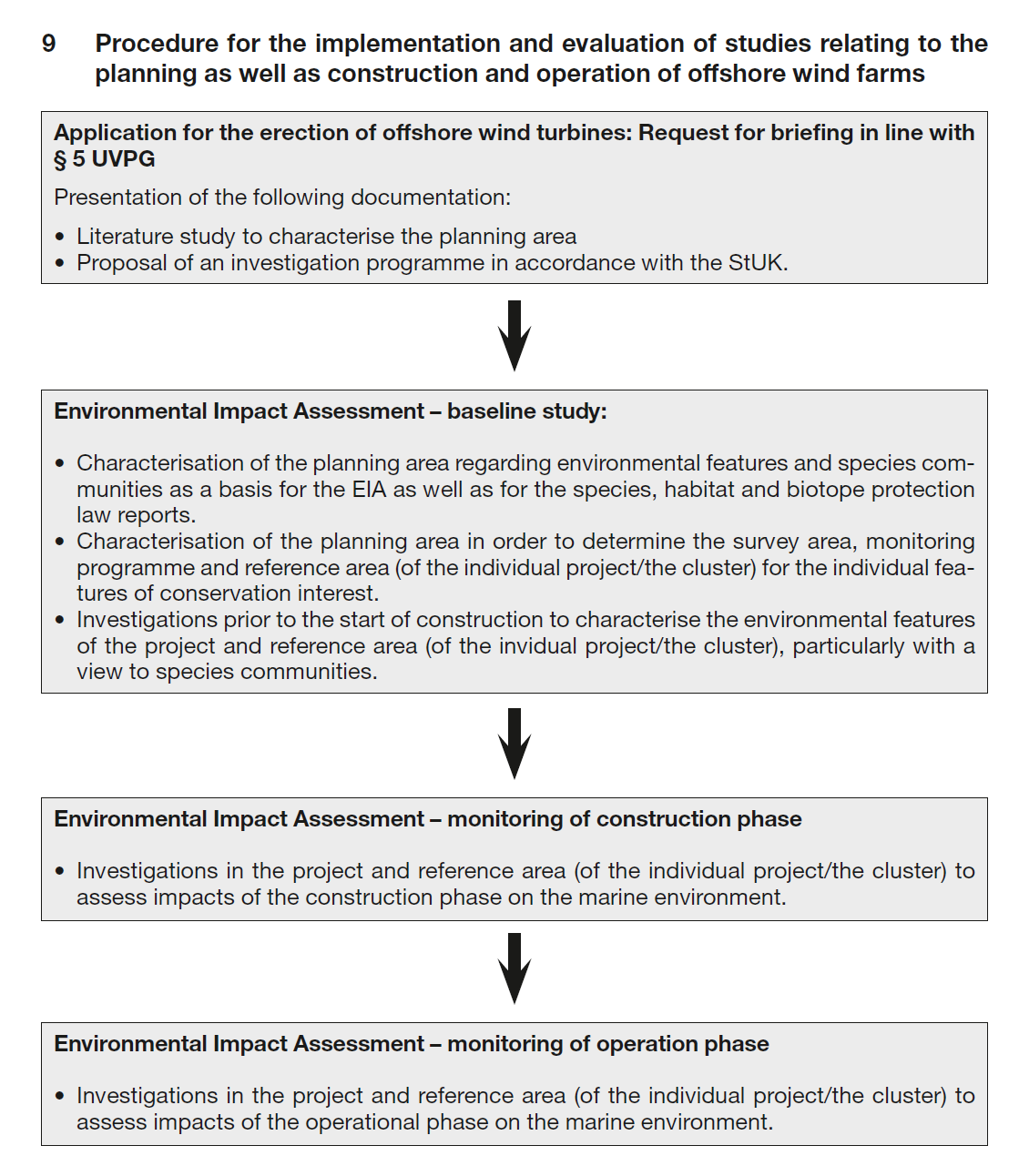
Figur 2: Standard Investigation of the Impacts of Offshore Wind Turbines on the Marine Environment (StUK4) Bundesamt für Seeschifffahrt und Hydrographie (BSH). Federal Maritime and Hydrographic Agency Hamburg und Rostock 2013.
1.2 - Protokoller for miljøovervåkning
Det finnes protokoller i forskjellig land for gjennomføring av miljøundersøkelser ved utbygging av vindkraftanlegg til havs, men i Norge finnes det per i dag ingen sånn protokoll. Her trekker vi frem protokollen fra Tyskland (StUK4; Figur 2) og diskuterer om og hvordan protokollen kan benyttes til miljøundersøkelser for havbaserte vindkraftprosjekt i Norge.
I Tyskland kreves det gjennomføring av sammenhengende “baseline” studier over to hele sesonger før konstruksjon av vindkraftanlegg, med krav om et tredje år i tillegg hvis perioden mellom baseline studien og konstruksjon av anleggene er over to år. Etter dette blir det anbefalt overvåkning under selve konstruksjonsfasen og over en periode fra 3 til 5 år etter at anlegget er satt i drift. Det er krav om gjennomføring av sammenlignbare miljøundersøkelser/overvåking i prosjektområdet og minst et upåvirket referanseområde i disse periodene. Det er spesifikke og detaljerte krav til metoder og evaluering av data for bunnlevende organismer, fisk, fugl, flaggermus, og marine pattedyr.
Den tyske protokollen er svært grundig for alle organismegrupper. De har et utvidet vedlegg med tekniske spesifikasjoner som beskriver hvor ofte og på hvilken måter man skal måle forekomst av bunndyr, fisk og fugl før, under og etter konstruksjon. Mange av disse metodene kunne bli brukt til en utforming av en norsk protokoll for overvåking av effekter av havvindanlegg. For overvåking av fisk er det et stort fokus på trålbaserte undersøkelser i den tyske protokollen, mens det i Norge, der bunnforholdene og dybde ofte vil være mer variable enn på kontinentet, vil være mer naturlig å satse på en kombinasjon av fiskeri og akustikk. For flytende havvindanlegg på dypt vann vil det dessuten være teknisk vanskelig å gjennomføre fiskeriundersøkelser basert på tråling. Det trengs en nyutvikling av metoder for overvåking innenfor flytende anlegg, der autonome farkoster og stasjonære observasjonsplattformer for langtidsovervåking med fordel kan tas i bruk. Dataene bør være åpent digitalt tilgjengelige.
Referanser
Bundesamt für Seeschifffahrt und Hydrographie (BSH) Federal Maritime and Hydrographic Agency Hamburg und Rostock (2013) Standard. Investigation of the Impacts of Offshore Wind Turbines on the Marine Environment (StUK4) BSH-Nr. 7003.
2 - Fysiske endringer introdusert av havvindanlegg
2.1 - Fundamenter
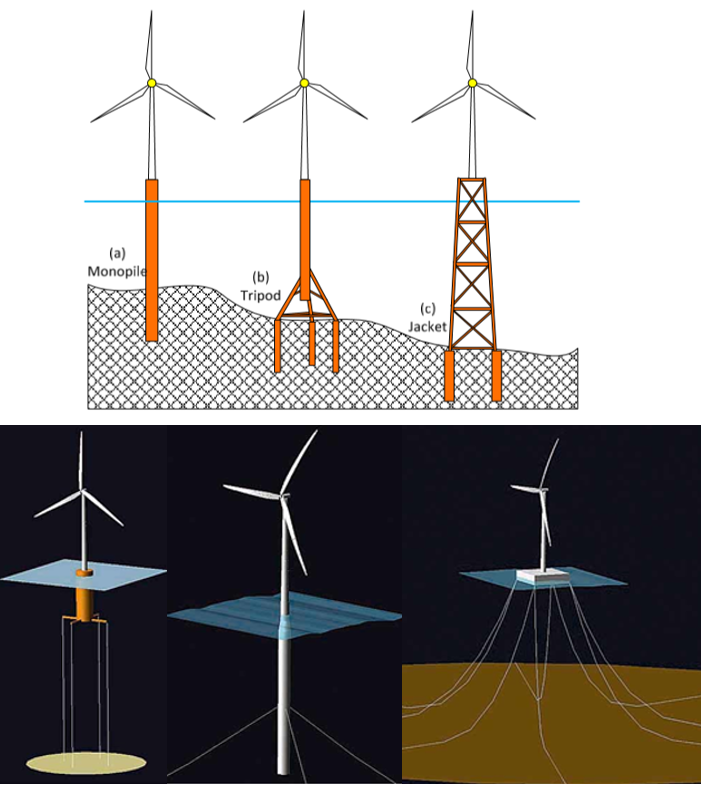
Figur 3: Øverst: Ulike typer bunn monterte vindturbiner. Til nå er de aller fleste vindturbiner montert på havbunnen ved relativt grunne dyp på opptil 50 meter. Monopåle der en påle/stålrør blir banket ned i havbunnen er mest vanlig, men kan bare brukes grunnere enn 30 m. Alternativ som kan brukes mellom 30 - 50 m er tripod med tre påler og eller en “jacket” med fire påler. (Figur fra Plodpradit, 2019). Nederst: Ulike typer ankersystem for flytende vindturbiner. Venstre: Tension leg platform, vertikale, lite elastiske, forankringer som hindrer vertikal bevegelse. Midten: Sparbøye, vertikal flytende bøye med anker til bunnen. Hywind (Skottland, Tampen, og Test/Demo ved Karmøy) er av denne typen. Høyre: flytende lekter med slakke ankerkabler til bunnen. (Figur fra Jonkman et al. 2011).
De fleste marine vindkraftanlegg som har vært i drift til nå er bunnfaste installasjoner, begrenset til relativt grunne sjøområder ned til ca. 50-60 meters dyp. Det finnes flere forskjellige fundamenteringsprinsipper for bunnfaste vindturbiner, hvorav monopeling og gravitasjonsfundamenter er de mest brukte. Som erosjonsbeskyttelse er det også vanlig å anlegge voller av stein rundt fundamentene som vil kunne øke anleggets påvirkningssone både når det gjelder strømforhold og bunneffekter (Glarou et al. 2020). Flytende vindturbiner kan lokaliseres på større dybder enn bunnfaste, og vil øke havarealet tilgjengelig for vindkraftproduksjon betraktelig. Flytende vindkraftanlegg forankres til bunnen i form av moringer og både flytende og nedsenkbare plattformer med en eller flere turbiner er under utvikling.
Miljøpåvirkningene av havbaserte vindkraftanlegg vil sannsynligvis variere mellom bunnfaste og flytende installasjoner både når det gjelder fysisk påvirkning av sjøbunnen og generering av støy (ICES 2018). Per i dag finnes det begrenset kunnskap om hvordan flytende vindkraftanlegg kommer til å se ut nøyaktig, da kun et slikt anlegg (HyWind Scotland som ble etablert utenfor kysten av Skottland i 2017) foreløpig er i drift i tillegg til testsenteret utenfor Karmøy (https://sustainableenergy.no/kompetanse-og-fasiliteter) der en enkelt Hywind turbin har vært forankret siden 2009. Noen ulike typer fundament for både flytende og bunnmonterte vindturbiner er vist i figur 3.
Referanser
Glarou M, Zrust M, Svendsen JC. 2020. Using Artificial-Reef Knowledge to Enhance the Ecological Function of Offshore Wind Turbine Foundations: Implications for Fish Abundance and Diversity. Journal of Marine Science and Engineering. 8, 332; doi:10.3390/jmse8050332.
ICES. 2018. Report of the Working Group on Marine Benthal and Renewable Energy Developments (WGMBRED), 6-9 March 2018, Galway, Ireland. ICES CM 2018/HAPISG:02. 66pp.
Jonkman JM, Matha D. 2011. Dynamics of offshore floating wind turbines-analysis of three concepts. Wind Energy, 14(4), 557–569. https://doi.org/10.1002/we.442.
Plodpradit P, Dinh VN, Kim K-D. 2019. Tripod-Supported Offshore Wind Turbines: Modal and Coupled Analysis and a Parametric Study Using X-SEA and FAST. Journal of Marine Science and Engineering, 7, 181. https://doi.org/10.3390/jmse7060181.
2.2 - Kabler
En undervannskabel skaper elektromagnetiske felt med en elektrisk og en magnetisk komponent. Det elektriske feltet kan vanligvis isoleres inne i kabelen mens de magnetiske feltene vil gå ut i kabelens omgivelser. Selv om det elektriske feltet isoleres i kabelen, kan det magnetiske feltet indusere et elektrisk felt utenfor kabelen, i vannet og/eller i organismer (Skiles 1985). Størrelsen på magnetfeltet kan være omtrent 3000 μT på kabelen, men minker raskt med avstand og er i samme størrelsesorden som jordens magnetfelt på 6 m avstand (Taormina et al. 2018). Likevel kan slike magnetfelt sannsynligvis oppfattes av en del marine dyr opptil flere titalls meter fra kablene, og i enkelte tilfeller også betydelig lengre (Walker 2001, Nyqvist et al. 2020). Styrken på magnetfeltet avhenger av turbin og anleggsstørrelsen. For vindfarmer med stor avstand til land vil det være aktuelt med offshore transformatorstasjoner som omformer vekselstrømmen fra turbinene til likestrøm. I framtiden kan havvindanlegg koples til et nettverk av likestrømskabler som krysser Nordsjøen for kraftutveksling. Kabellegging påvirker det fysiske miljøet ved bunn.
Kablenes elektromagnetiske felt oppfattes videre av dyrene sammen med naturlige magnetiske gradienter og tilfeldige variasjoner. Retningen og intensiteten på jordens magnetfelt varierer med geografisk plassering. Vinkelen mellom horisontalplanet og retningen på totalfeltet er 0° ved ekvator og 90° ved magnetpolene, mens intensiteten er rundt 60 000 nT ved polene, 40 000–50 000 nT ved midtbreddegrader og 30 000 nT ved ekvator. Dette resulterer i en gjennomsnittlig endring på 2–5 nT/km og 0,01°/km mellom ekvator og polene, men dette er kun gjennomsnittlige endringer. Krystallstein og ikke-dipolkomponenter i jordens kjerne produserer også lokale anomalier og forårsaker gradienter på 10–100 nT/km (Kullnick 2000; Walker et al. 2003). I tillegg varierer det naturlige magnetfeltet med solstråling. I norske farvann handler det om noen få til hundrevis av mindre solgenererte magnetiske forstyrrelser (135–158 nT) årlig, med varighet på 0,5–2 timer, og et par mer omfattende solstormer (> 1000 nT), med varighet over flere dager, 5–10 ganger per 11-års-solar-syklus (Nyqvist et al. 2020).
Naturlige elektriske felt blir indusert når saltvann beveger seg i det magnetiske feltet, og varierer med feltstyrke og strømhastighet (Kalmijn 1999). På tilsvarende måte blir også elektriske felt indusert når dyr, som også leder elektrisitet, svømmer i jordens magnetfelt (Kalmijn 1999). En annen kilde til naturlige elektriske felt er levende organismer som skaper elektriske felt som følge av deres fysiologiske livsprosesser – hvilket for eksempel bruskfisk bruker for å jakte (Crampton 2019; Haine et al. 2001).
Referanser
Crampton WG. 2019. Electroreception, electrogenesis and electric signal evolution. Journal of Fish Biology 95, 92–134.
Haine OS, Ridd PV, Rowe RJ. 2001. Range of electrosensory detection of prey by Carcharhinus melanopterus and Himantura granulata. Marine and Freshwater Research 52, 291-296.
Kalmijn A. 1999. Detection and biological significance of electric and magnetic fields in microorganisms and fish, pp. 4-5.
Kullnick U. 2000. Influences of electric and magnetic fields on aquatic ecosystems. ICNIRP: Effects of Electromagnetic Fields on the Living Environment, 113-132.
Nyqvist D, Durif C, Johnsen MG, de Jong K, Forland TN, and Sivle LD. 2020. Electric and magnetic senses in marine animals, and potential behavioral effects of electromagnetic surveys, Marine Environmental Research 155:104888, doi: 10.1016/j.marenvres.2020.104888.
Skiles DD. 1985. Magnetite Biomineralization and Magnetoreception in Organisms, Springer, pp. 43-102.
Taormina B, Bald J, Want A, Thouzeau G, Lejart M, Desroy N, Carlier A. 2018. A review of potential impacts of submarine power cables on the marine environment: Knowledge gaps, recommendations and future directions. Renewable and Sustainable Energy Reviews 96, 380-391.
Walker MM, Diebel CE, Kirschvink JL. 2003. Sensory processing in aquatic environments, Springer, pp. 53-74.
Walker T. 2001. Basslink project review of impacts of high voltage direct current sea cables and electrodes on chondrichthyan fauna and other marine life. Marine and Freshwater Resources Institute Report, 20, 68 pp.
2.3 - Sirkulasjon og turbulens
Et havvindanlegg eller en rekke av vindturbiner vil generelt redusere vindhastigheten betraktelig i den høyden som vindturbinene befinner seg i og senke energinivået i et område rundt og nedstrøms av havvindanlegget. Rotorbevegelsene vil bidra til turbulensdannelse ned mot overflaten, men den viktigste effekten på havoverflaten og sirkulasjonen nedover skjer nedstrøms av havvindanlegget. Flere studier er utført med målsetting om å få mer kunnskap om direkte effekter av havvindanleggene og potensialet for uthenting av energi, men kun et fåtall tar for seg de oseanografiske effektene.
Når vinden passerer at havvindanlegg, vil overflatevinden svekkes. Inntil den igjen får tilført energi fra vindfeltet høyere oppe, vil vinden da være en svakere drivkraft for overflatebølger og for direkte vinddrevet blanding nær overflaten enn før havvindanlegget. I tillegg til disse effektene påpekte Broström (2008) at aktive vindturbiner vil føre til endringer i vannmassenes oppstrømmings- og nedstrømningsdynamikk som følge av vindens endrede egenskaper. Studiet tar for seg et tenkt vindturbinanlegg med vilkårlig utstrekning på et åpent havområde hvor man har et definert aktivt overflatelag (10-20 m dypt), altså typisk situasjon i Nordsjøen og Skagerrak i sommerhalvåret med en oppvarmet overflate.
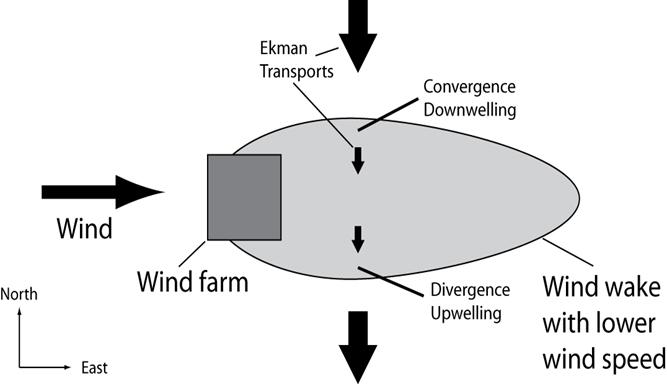
Vinden har den egenskapen på havoverflaten at den setter opp en sirkulasjon til høyre (nordlige halvkule) for vindretningen, og innen overflatelaget vil den totale vinddrevne transporten være rettet 90° mot høyre for vindretningen, den såkalte Ekman-transporten. Ettersom vindkraftanlegget reduserer vindhastigheten nedstrøms av anlegget, får man en konvergens- og en divergenssone i tilknytning til vindkraftanlegget (se Figur 4). Konvergens vil i praksis føre til nedstrømning, mens divergens vil medføre oppvelling av vannmasser, og det er styrken av disse vertikaltransportene som kan ha innflytelse på biomasseforflytninger lokalt rundt og nedstrøms av et vindkraftanlegg.
Figur 4: Skjematisk tegning av innvirkning fra et vindanlegg på sirkulasjonen i havet og tilknyttede konvergens- og divergenssoner (G. Broström, pers. komm.).
En viktig faktor i beregningene er den horisontale utstrekningen av anlegget. Broström (2008) ser primært på endringene i dybden av overflatelaget, som da er et annet mål for oppstrømnings- og nedstrømningshastighet. Han finner store utslag dersom utstrekningen av anlegget overstiger et visst kriterium (Rossby-radien). Kriteriet sier i praksis noe om typisk lengdeskala for de fenomener som dominerer det dynamiske bildet for et havområde, og i Nordsjøen er denne skalaen på rundt 5-10 km (avhenger av breddegrad, dybden av overflatelaget og oppdriftsegenskapene). Merk også at teoretisk sett øker vertikaltransportene i tredjepotens i forhold til dimensjonen av anlegget forutsatt at kriteriet om minimumsutstrekning er oppfylt (ca. 5 km i Nordsjøen). Med andre ord vil da en dobling av dimensjonen av anlegget (firedobling av arealet) medføre at oppstrømningshastigheten åttedobles.
Av andre relevante arbeider som tar for seg oseanografiske effekter av vindkraftanlegg til havs, så fokuserte Carpenter et al. (2016) sin studie på hydrodynamiske endringer på grunt vann i Tyskebukta i sørlige Nordsjøen. De beregnet at når tidevanns-strømmene passerte anleggene, så ville en turbulent bakevje kunne ha betydelig innvirkning på stratifiseringen. Det ble derfor konkludert med at hvis anleggene dekket store arealer så ville de berørte områdene kunne gå fra naturlig stratifiserte til mer gjennomblandete vannmasser, noe som igjen kunne ha en mulig innvirkning på primærproduksjonen. En homogenisering av vannmassene var også en av konklusjonene til Cazenave et al. (2016) som har beskrevet en fullskala modell-studie fra Irskesjøen. De kom i tillegg frem til at vindkraftanlegg på grunt vann utenfor kysten også kan medføre økt vannstand, eller mer konkret 7% er det estimert opp til 7% økning i det astronomiske tidevannet, f.eks. da gjennom økning av vannstanden fra tidevannet.
Clark et al. (2014) ga ut en omfattende litteratur-studie som oppsummerer forskningen som er gjennomført om miljøpåvirkninger fra vindkraftanlegg samt å avdekke forskningshull. De pekte på at det som kan påvirkes av fysiske forhold i sjøen er turbulens (nedstrøms), bølge-energien, strømforhold og vertikalbevegelse. Deres tolkning av ulike forskningsresultater peker på at de fysiske endringene vil være innenfor det som kan betraktes som naturlige svingninger, men noen studier predik erer at store utbygninger av vindkraft til havs kan forårsake irreversible endringer i sediment-deposisjon langs kysten, konveksjons-mønstre (dvs. vertikaltransport og turbulens) og/eller oksygennivåer nær bunn. Og som nevnt tidligere har det blitt påpekt en mulig sammenheng mellom store vindkraftanlegg, forringelse av den naturlige stratifiseringen og dermed endringer i regimene som omhandler næringsstoffer og planteplankton. Til slutt bemerkes det i Clark et al. (2014) at ingen av studiene så langt hverken belyser effekter langt unna vindkraftanleggene eller de kumulative effektene ved utbygging flere steder.
Som en oppfølger til Clark et al. (2014) har Bakhoday-Paskyabi et al. (2018) målt strøm og turbulens oppstrøms og nedstrøms to vindparker i Nordsjøen (FINO1 og Alpha Ventus). De kom frem til at der var liten forskjell i middelstrømmen på begge sider av vindparkene, men at det oppstod noe lavere grad av turbulens på lesiden. Denne svekkingen av turbulens var merkbar helt ned til sjøbunnen, som var rundt 30m i de testede tilfellene.
Referanser
Bakhoday-Paskyabi M, Fer I, Reuder J. 2018. Current and turbulence measurements at the FINO1 offshore wind energy site: analysis using 5-beam ADCPs. Ocean Dynamics 68, 109-130. doi: 10.1007/s10236-017-1109-5.
Broström G. 2008. On the influence of large wind farms on the upper ocean circulation, J. Mar. Syst, doi: 10.1016/j.jmarsys.2008.05.001.
Carpenter JR, Merckelbach L, Callies U, Clark S, Gaslikova L, Baschek B. 2016. Potential Impacts of Offshore Wind Farms on North Sea Stratification. PLoS ONE 11, e0160830. doi: 10.1371/journal.pone.0160830.
Cazenave PW, Torres R, Allen JI 2016. Unstructured grid modelling of offshore wind farm impacts on seasonally stratified shelf seas. Progress in Oceanogography 145, 25-41, doi: 10.1016/j.pocean.2016.04.004.
Clark S, Schroeder F, Baschek B. 2014. The influence of large offshore wind farms on the North Sea and Baltic Sea – a comprehensive literature review. Helmholtz-Zentrum Geesthacht report 2014-6.
2.4 - Støy
Vindkraftanlegg forårsaker lyder som kan høres av de fleste fisk og sjøpattedyr. Lydene kan deles opp i konstruksjonsstøy som er lyder som lages i forbindelse med oppføringen av vindturbinene, og i produksjonsstøy som er lyder vindturbinene lager når de er i drift. Ordet «støy» har flere betydninger, men vil bare bli brukt i betydningen «uønsket lyd» i denne rapporten.
2.4.1 - Konstruksjonsstøy
Tiden det tar å sette opp et vindturbinanlegg er begrenset, men det kan ta flere måneder. I denne tiden kan det bli mye høye lyder. Lydkilder i denne perioden kan være pæling, boring, gravearbeid og økt båttrafikk (Thomsen et al, 2006). Lydene i denne fasen avhenger av antall og type (flytende eller bunnmonterte) vindturbiner.
For bunnmonterte vindturbiner blir det ofte brukt pæling der en hydraulisk hammer dunker ned påler i havbunnen. For vindturbiner er som oftest pålene tykke stålrør med diameter på flere meter (1.5 - 4 m avhengig av fundamenttype) og hammeren slår gjentatte slag, typisk 1-2 slag per sekund (Thomsen et al, 2006). Antall slag varierer, men kan være ca. 5000 per påle, og jobben kan vare i flere timer avhengig av pålens diameter, hammerens størrelse, bunnegenskapene og hvor dypt den skal ned i bunnen. De kan bli banket opptil 50 m ned i havbunnen (Betke 2014). Når hammeren treffer pålen, blir pålen presset sammen og det oppstår en kompresjonsbølge i stålet. Et materiale som blir komprimert i en retning vil utvide seg i en annen retning. I området der det er kompresjon vil det oppstå en midlertidig deformasjon i form av en “utbuling” på pålen som vil flytte seg nedover pålen med hastighet mye høyere enn lydhastigheten i vann. Dette lager en “Mach-bølge effekt” som forårsaker det meste av lyden som blir spredt i havet (Reinhall og Dahl 2011). I tillegg vil det oppstå andre bølgetyper i havbunnen og grensesjiktet mellom hav og bunn. Lavfrekvente seismiske overflatebølger som “ground roll waves” brer seg ikke utover i vannet, men de brer seg langs havbunnen der de kan føre til høye nivå av partikkelbevegelse nær havbunnen (Hazelwood og Macey 2016).
Siden impulsive lyder har så kort varighet er det vanlig å bruke spisstrykk (0-peak verdi) for å beskrive maksimaltrykket som er et mål på den høyeste positive eller laveste negative verdien i en definert periode, eller lydeksponeringsnivå (sound exposure level, SEL) for å beskrive energien i en lydpuls/hammerslag. Den samlede (kumulative) energien fra mange slag kan også beskrives ved å legge sammen energien til enkeltslag over en periode. Spisstrykket kan være omtrent 220 dB re 1 μPa 10 meter fra en påle med diameter på 0.75 cm og 200 dB re 1 uPa 300 meter fra en påle med diameter på 5 meter (Dahl et al. 2015). 1600 m fra pæling av en påle med diamenter på ca 2.5 m ble det målt spisstrykk på 190 dB re 1 μPa og SEL på 165 dB re 1μPa2 s (Betke 2014). For pæling er spisstrykket typisk 20 - 25 dB høyere enn SEL.
Ved avstander på 1600 - 16 000 m fra pælingen er mesteparten av lyden i frekvensområdet rundt 50 -400 Hz, men lydnivået er høyere enn bakgrunnsstøyen fra under 20 Hz til ca 10 kHz (Betke 2014). Varigheten på lydpulsene øker med avstand. 600 m fra kilden kan varigheten være ca 0.1 sekund, men 17 km fra kilden kan varigheten være ca 0.6 sekund (Betke 2014). Med 1 sekund mellom slagene vil lyden kunne oppfattes som kontinuerlig støy ved store avstander.
Det finnes flere tiltak som kan brukes for å begrense lydnivået fra pæling (Stokes et al. 2010):
Boblegardin som er et slør av bobler rundt pålen, kan gi reduksjon i SEL på 10 - 15 dB ( Verfuß 2014)
Ballonggardin ligner på boblegardin, men har fastmonterte ballonger istedenfor bobler ( Verfuß 2014)
Et luftfylt rør rund pålen (kalt cofferdam på engelsk; Verfuß 2014)
Pakke inn pålen med et dempende materiale (Elmer et al. 2007)
Et lag med dempende materiale oppå pålen (Elmer et al. 2007).
Vibrasjons pæling erstatter hammer-slag med en slags vibrerende hammer som rister pålen ned i bakken. Denne metoden lager lavere lyd, men her blir lyden kontinuerlig. Den totale lydenergien blir dermed ikke nødvendigvis lavere for denne metoden ( Verfuß 2014 ).
2.4.2 - Produksjonsstøy
Produksjonslyder er mye lavere enn lydene under konstruksjonsfasen, men dette er permanente lyder som vil være der så lenge vindturbinene er i drift. Lydnivået varierer og vil i stor grad påvirkes av vindstyrke (Pangerc et al. 2016).
Lyden som brer seg i luft fra rotasjonsbladene blir nesten fullstendig reflektert fra havoverflaten og får liten betydning under vann (Tougaard et al. 2009, Betke 2014). Under vann kommer støy fra vindturbiner hovedsakelig fra girkassen og fra generatoren som brer seg nedover tårnet og ut i vannet. Mekaniske deler som roterer resulterer ofte i ekstra høy lyd på smale frekvensbånd (toner), der frekvensen er direkte knyttet til antall mekaniske svingninger/rotasjoner/vibrasjoner per sekund. Resonanser i vindturbinstrukturen kan oppstå ved høyere harmoniske av rotasjonsfrekvensen (Marmo et al. 2013).
Vinden får rotasjonsbladene til å rotere og hovedakselen roterer med samme rotasjonshastighet. Hovedakselen er koblet til en girkasse eller til et drev og dette er igjen koblet til en generator som omdanner mekanisk energi til elektrisk energi. Generatoren består av en stasjonær del (stator) og en bevegelig del (rotor), der den ene delen er magnetisk og den andre er en elektrisk leder. Det norske strømnettverket bruker en frekvens på 50 Hz. For å oppnå dette må rotoren til generatoren ha en rotasjonshastighet på 1500 runder pr. minutt. Vindturbinen roterer omtrent 15 runder per minutt, men ved å bruke gir kan en øke rotasjonshastigheten. Denne typen generator kalles høyhastighets generator. En annen måte å få ønsket frekvens på er å ha flere pol-par, det vil si flere magneter på rotor eller stator for å endre retningen til strømmen mange ganger underveis på en rotasjon. På denne måten kan en klare seg uten gir. Slike generatorer kalles direktedrevne generatorer. Innenfor disse generatortypene finnes det ulike undergrupper og en kan forvente at ulike generatortyper vil produsere ulik støy som kan bre seg i havet (Polinder et al. 2006). Bl.a. for å redusere vekten i toppen av tårnet på vindturbinen, er det nå en klar trend bort fra gir og mot direktedrevne generatorer, spesielt til havs og ettersom turbinene blir større.
Det er flere eksempler på produksjonsstøy fra vindturbiner i litteraturen som er målt i ulike avstander fra en eller flere vindturbiner. Generelt er inntrykket at nær vindturbinene, 1-50 meter avstand, er det forhøyet bakgrunnsstøy over et bredt frekvensspekter fra 10 - 1000 Hz i tillegg til at det er enkelte frekvenser som skiller seg ut med høyere topper (toner; Figur 5). Frekvensen til tonene avhenger av vindturbintype og vindhastighet (Tougard et al. 2009, Pangerc et al. 2016, Cheesman 2016, Wahlberg og Westerberg 2005). Ved større avstander (>100 m) ser man gjerne bare tonene som skiller seg ut (Betke 2014, Thomsen et al, 2006).
I tillegg til gir-system og generator kan måten vindturbinen er fundamentert på kan påvirke hvor mye lyd som blir overført til havet. Ulike fundament kan være tripod som har tre påler, eller “four legged jacket” som har fire påler (Figur 3). I anlegget Alpha Venturs hadde de to ulike typer vindturbiner som var montert på to ulike typer fundament. Da de gjorde støymålinger, kunne de bare detektere støy fra den ene vindturbintypen. Siden både turbin med gir og generator, og fundamentet var ulikt for de to metodene fant de ikke ut hva som var årsaken til at den ene typen var mer stille (Betke 2014).
Basert på teoretiske beregninger viser Marmo et al. (2013) at måten vindturbinen er fundamentert på påvirker lydforplantningen. Produksjonsstøy fra vindturbiner på 6 MW ble modellert for tre ulike fundamenttyper; monopåle, «jacket» og «gravity based». Gravity based er ikke dunket ned i bunnen, men har en tung, tykk sokkel plassert på havbunnen. For frekvenser under 500 Hz hadde monopåle 10 dB høyre lyd enn gravity based, og 50 dB høyere lyd enn jacket. For frekvenser høyere enn 500 Hz hadde jacket høyest lyd av de tre typene, men for disse høye frekvensene var lyden nær eller lavere enn bakgrunnsstøyen. Støyen fra de tre ulike fundamanttypene hadde toner ved omtrent de samme frekvensene fordi turbinene hadde samme gir, generator og vindstyrke, men lydnivået på tonene var veldig ulikt mellom de tre typene på grunn av ulik geometri og materiale og ulik kontakt med havbunnen.
2.4.3 - Lydtrykk vs partikkelbevegelse
Hawkins og Popper (2017) hevder at vurderinger av miljøpåvirkninger fra vindturbiner basert på lydtrykk blir feil fordi de fleste arter er følsomme for partikkelbevegelse. Det går an å beregne partikkelbevegelsen basert på lydtrykksmålinger, men for mange tilfeller blir det for komplisert. Målinger og beregninger fra pæling ved 2.5 m dyp viste at det var et lineært forhold mellom lydtrykk og partikkelbevegelse, men målingene viste også at partikkelhastigheten var mye høyere enn forventet basert på teori. Frekvensfordelingen til lydtrykket og partikkelbevegelsen var også forskjellig (Ceraulo et al. 2016). I tillegg tar modeller av lydtrykk ikke hensyn til “ground roll waves” som kan lage høye nivå av partikkelbevegelsen langs bunnen ved relativt store avstander fra pælingen (Hazelwood og Macey 2016). På den andre siden kan det være utfordrende å gjøre målinger av partikkelbevegelsen fordi det krever spesialutstyr og riktig kompetanse (Nedelec et al. 2016, Gray et al. 2016). For produksjonsstøy er også partikkelbevegelsen av interesse. Sigray og Andersson (2011) utviklet et partikkelakselerometer for dette formålet. Målingene viste at også partikkelakselerasjonen har smale maksimum/ toner på samme måte som lydtrykket.
2.4.4 - Flytende vindturbiner:
For flytende vindturbiner er det ikke så mye informasjon tilgjengelig om lydnivået. Flytende vindturbiner er dyrere og teknisk mer komplisert enn bunnmonterte. Denne teknologien gjør at dypere havområder kan brukes. Det at havet er dypere og turbinene ikke er fundamentert i bunnen vil nok gi et annet lydbilde enn for bunnmonterte vindturbiner, selv om tonene fra mekanisk rotasjon trolig vil bli lignende. Flytende havvindturbiner er under utvikling i flere land. Testing foregår bl.a. ved Karmøy der en enkelt turbin Hywind Demo ble satt i gang i 2009, men det er så vidt vi kjenner til bare et flytende havvindanlegg som er i kontinuerlig drift. Hywind Scotland (Equinor) har vært i operasjon fra 2017 og ligger 25 km utenfor Aberdeen i Skottland med 6 vindturbiner. Hywind Tampen er planlagt fra 2022.
Støy fra en prototype på en flytende vindturbin som ble kalt Hywind Demo ble målt i et testområde vest for Stavanger ved Karmøy i august 2011 (Martin et al. 2011). Høy bakgrunnsstøy i området på grunn av skipstrafikk gjorde det vanskelig å detektere lyden fra vindturbinen. Lydmålingene ble gjort ved 150 m avstand, og en kontroll 10 km unna. I perioder med lavere bakgrunnsstøy kunne smalbånd frekvenstopper bli sett. Toner som er typisk for gir-støy. Ingen av tonene hadde høyere nivå enn 115 dB re 1 uPa^2/Hz på 150 m avstand.
En annen type støy ble også observert fra denne vindturbintypen. Det var transiente impulser, bredbånd 100 -20 000 Hz. Altså korte smell som viste seg å være relatert til forankringssystemet i form av rykking og napping i ankerkjettingen ved bevegelse av vindturbinen (Martin et al. 2011). Det kunne være opptil 23 slike lyder pr dag fra en eneste turbin. Forankringssystemet til prototypen bestod av en kombinasjon av stål wire 465 m og kjetting 330 m. Ferdige modeller kan ha en annen type wire.
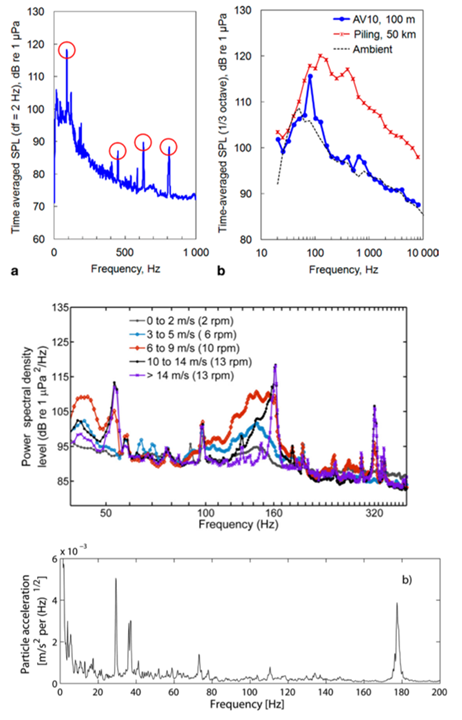
Figur 5: Øverst: lydmålinger fra 100 m avstand fra en vindturbin som funksjon av frekvens fra det bunnmonterte vindturbinanlegget Alpha Ventus i Nordsjøen (Tyskland). Venstre figur viser frekvensspekteret med 2 Hz frekvenssteg der en ser toner ved 90 Hz, og mellom 400 - 800 Hz. Høyre figur viser samme resultat med ⅓ oktav frekvenssteg (som betyr at lyden er midlet over bredere frekvensspekter) og sammenligner nivået fra produksjonsstøy ved 100 m avstand, og pæling ved 50 km avstand med bakgrunnsstøyen. Dette viser at mye av lyden for de lave frekvensene kommer fra bakgrunnsstøy, og at det er de smalbåndede lydene som kommer fra vindturbinen. Selv på 50 km avstand har pæling høyere lydnivå enn produksjonstøyen fra vindturbinen. Når en bruker ⅓ oktavbånd på lyder med smalbånd støy blir det vanskelig å se ved hvilke frekvenser en har støy. (Figur fra Betke, 2014). Midten: Sammenligning av lyd for ulike vindforhold. Målingene er midlet i tid over minst 54 timer. Noen toner oppstår med samme frekvens, men ulik amplitude når vindstyrken endrer seg. Andre toner skifter frekvens med vindstyrken. (Figur fra Pangerc et al, 2016). Nederst: Partikkelakselerasjonen ved 1 m avstand fra en vindturbin ved Utgruden i det Baltiske hav i Sverige med et bunn-ankret akselerometer. Det var en 1.5 MW turbin ved 12 m/s vind. Signalet under 2 Hz er trolig fra bølger på havet. Frekvens-toppene flyttet seg med vindhastighet. Disse kommer mest sannsynligvis fra girkassen. (Figur fra Sigray og Andersson, 2011). Reproduced from Sigray, P., & Andersson, M. H. (2011). Particle motion measured at an operational wind turbine in relation to hearing sensitivity in fish. The Journal of the Acoustical Society of America, 130(1), 200–207. https://doi.org/10.1121/1.3596464 with the permission of the Acoustical Society of America.
Referanser
Betke K. 2014. Underwater construction and operational noise at alpha ventus. In: Federal Maritime and Hydrographic Agency, Federal Ministry for the Environment, Nature Conservation and Nuclear Safety (eds) Ecological Research at the Offshore Windfarm alpha ventus. Springer Spektrum, Wiesbaden.
Ceraulo M, Bruintjes R, Benson T, Rossington K, Farina A, Buscaino G. 2016. Relationships of sound pressure and particle velocity during pile driving in a flooded dock. Fourth International Conference on the Effects of Noise on Aquatic Life. https://doi.org/10.1121/2.0000295.
Cheesman S. 2016. Measurements of Operational Wind Turbine Noise in UK Waters. In The Effects of Noise on Aquatic Life II (pp. 153–160). Springer New York. https://doi.org/10.1007/978-1-4939-2981-8_18.
Dahl PH, de Jong CAF, Popper AN. 2015. The Underwater Sound Field from Impact Pile Driving and Its Potential Effects on Marine Life. Acoustics Today, 11, 18–25
Elmer KH, Gerasch WJ, Neumann T, Gabriel J, Betke K, Schultzvon Glahn M 2007. Measurement and Reduction of Offshore Wind Turbine Construction Noise. DEWI Magazine No. 30. Wilhelmshaven, 33–38.
Gray M, Rogers PH, Zeddies DG. 2016. Acoustic particle motion measurement for bioacousticians: principles and pitfalls. Presented at the Fourth International Conference on the Effects of Noise on Aquatic Life. https://doi.org/10.1121/2.0000290
Hawkins AD, Popper AN. 2017. A sound approach to assessing the impact of underwater noise on marine fishes and invertebrates. ICES Journal of Marine Science 74:635–651. DOI: 10.1093/icesjms/fsw205.
Hazelwood R, Macey P. 2016. Modeling Water Motion near Seismic Waves Propagating across a Graded Seabed, as Generated by Man-Made Impacts. Journal of Marine Science and Engineering, 4(3), 47. https://doi.org/10.3390/jmse4030047.
Marmo B, Roberts I, Buckingham MP, King S, Booth C. 2013. Modelling of Noise Effects of Operational Offshore Wind Turbines including noise transmission through various foundation types. Edinburgh: Scottish Government.
Martin B, MacDonnell J, Vallarta J, Lumsden E, Burns R. 2011. HYWIND Acoustic Measurement Report: Ambient Levels and HYWIND Signature. Technical report for Statoil by JASCO Applied Sciences. https://www.equinor.com/en/how-and-why/impact-assessments/hywind-tampen.html.
Nedelec SL, Campbell J, Radford AN, Simpson SD, Merchant ND. 2016. Particle motion: the missing link in underwater acoustic ecology. Methods in Ecology and Evolution, 7, 836–842. https://doi.org/10.1111/2041-210x.12544.
Pangerc T, Theobald PD, Wang LS, Robinson SP, Lepper PA. 2016. Measurement and characterisation of radiated underwater sound from a 3.6 MW monopile wind turbine. The Journal of the Acoustical Society of America, 140(4), 2913–2922. https://doi.org/10.1121/1.4964824.
Polinder H, van der Pijl FFA, de Vilder G-J, Tavner PJ. 2006. 'Comparison of direct-drive and geared generator concepts for wind turbines.', IEEE transactions on energy conversion, 21, 725-733
Reinhall PG, Dahl PH. 2011. Underwater Mach wave radiation from impact pile driving: Theory and observation. The Journal of the Acoustical Society of America, 130, 1209–1216. https://doi.org/10.1121/1.3614540.
Sigray P, Andersson, MH. 2011. Particle motion measured at an operational wind turbine in relation to hearing sensitivity in fish. The Journal of the Acoustical Society of America, 130, 200–207. https://doi.org/10.1121/1.3596464.
Stokes A, Cockrell K, Wilson J, Davis D, Warwick D. 2010. Mitigation of Underwater Pile Driving Noise During Offshore Construction. Report no. M09PC00019 to Department of the Interior, Minerals Management Service. Groton/CT, January 2010, 104 pp.
Thomsen F, Lüdemann K, Kafemann R, Piper W. 2006. Effects of offshore wind farm noise on marine mammals and fish, biola, Hamburg, Germany on behalf of COWRIE Ltd.
Tougaard J, Henriksen OD, Miller LA. 2009. Underwater noise from three types of offshore wind turbines: Estimation of impact zones for harbor porpoises and harbor seals. The Journal of the Acoustical Society of America, 125, 3766–3773. https://doi.org/10.1121/1.3117444.
Verfuß T. 2014. Noise mitigation systems and low-noise installation technologies. In: Federal Maritime and Hydrographic Agency, Federal Ministry for the Environment, Nature Conservation and Nuclear Safety (eds) Ecological Research at the Offshore Windfarm alpha ventus. Springer Spektrum, Wiesbaden. https://doi.org/10.1007/978-3-658-02462-8_16.
Wahlberg M, Westerberg H. 2005. Hearing in fish and their reactions to sounds from offshore wind farms. Marine Ecology Progress Series, 288, 295–309. https://doi.org/10.3354/meps288295.
2.5 - Lys
Vindkraftanlegg vil påvirke lyset gjennom skyggekast og refleksblink fra turbintårn og rotorblad spesielt. I tillegg vil det ofte plasseres kunstige lys på turbintårnene slik at anleggene også på nattestid skal være synlige både for luftfart og skipstrafikk. Denne type lysforurensning kan ha effekter på nattaktive organismer avhengig av lysstyrke, lysspekter og blinkfrekvens (Gaston et al. 2012). For å redusere effektene av denne type lysforurensning er det f.eks. i Tyskland foreslått reguleringer der lysene på vindturbiner kun skal være aktive i perioder med passerende flytrafikk (https://www.bundesregierung.de/breg-en/news/neue-regeln-bei-windraedern-1710758).
Frekvensen av skyggekast/refleksblink fra vindturbiner vil avhenge av vindstyrke. Refleksblinkenes styrke ved havoverflaten vil avhenge av navhøyde, rotorbladdiameter og rotorbladenes refleksjonsegenskaper, samt intensitet og vinkel av solinnstråling, som igjen vil variere med årstid og skydekke. Bladene til vindturbiner har imidlertid fått en anti-reflekterende lag som gjør at de reflektere lys minimalt. Skygge kast fra rotorblad vil avhenge av skyer og solhøyde. Mye av lyset som reflekteres eller passerer gjennom bladene, vil igjen reflekteres fra havoverflaten, og den del av lyset som trenger ned i vannmassene svekkes raskt med avstand og dyp. Eventuelle effekter av refleksjoner eller skyggekast på marine organismer, vil derfor i overveiende grad være lokale. Mange fiskeslag reagerer kraftig på visuelle stimuli, bl.a. med fluktreaksjoner, og det er derfor ikke usannsynlig at skyggekast og refleksblink fra vindturbiner vil kunne ha skremmeeffekt på fisk. Hvordan fisk påvirkes av denne type stimuli vil også avhenge av fiskens tilvenningsevne. På solrike dager vil også turbin konstruksjonene skape skygger i vannet, og ved å plassere seg i skyggen, vil fisk kunne overvåke sitt territorium for inntrengere og fange byttedyr som flyter eller svømmer forbi i den opplyste vannmassen (Helfman 1981). Effekter av lyspåvirkning fra vindkraftanlegg på fisk og andre marine organismer er det per i dag ingen kunnskap om og det blir derfor ikke behandlet i denne rapporten.
Referanser
Helfman G. 1981. The Advantage to Fishes of Hovering in Shade. Copeia, 1981, 392-400. doi:10.2307/1444228.
Gaston KJ, Davies TW, Bennie J, Hopkins J. 2012. REVIEW: Reducing the ecological consequences of night-time light pollution: options and developments. Journal of Applied Ecology, 49, 1256–1266. https://doi.org/10.1111/j.1365-2664.2012.02212.x.
Det finns flere kunnskapshull om hvordan vindkraftanlegg kommer til å endre de fysiske omgivelsene i havet. Herunder oppsummerer vi de viktigste:
Per i dag finnes det begrenset kunnskap om hvordan fundamentene av flytende vindkraftanlegg kommer til å se ut nøyaktig, da kun et slikt anlegg (HyWind Scotland) foreløpig er i drift.
Vi vet lite om støy fra flytende havvind fordi målinger bare er gjort ved prototypen til Hywind og var påvirket av mye bakgrunnsstøy.
I hvor lang avstand blir «den naturlige» bakgrunnsstøyen påvirket av et vindkraftanlegg. Altså i hvor stor avstand vil den opprinnelige bakgrunnsstøyen bli høyere på grunn av vindkraftanlegget.
Bølger som brer seg i grensesjiktet mellom hav og bunn i forbindelse med pæling (ground roll waves) burde undersøkes nærmere (med geofoner/partikkelakselerometer) og det bør sjekkes hvordan dette påvirker bunnlevende organismer.
Turbulens/havstrøm/strømningsfelter: Det er ikke godt kjent hvordan strømmer og vertikalblanding kan endre seg bak vindkraftanleggene og hva de kumulative effektene er ved utbygging flere steder.
Effekter av lyspåvirkning fra vindkraftanlegg på fisk og andre marine organismer er det ingen kunnskap om.
3 - Effekter av fundamenter på artsfordeling: Lokale bevegelser og endringer i populasjonsnivå.
3.1 - Bakgrunn
Havvindanlegg oppføres hovedsakelig i kystnære gruntvannområder, som ofte huser høyproduktive, artsrike organismesamfunn som vil være sårbare overfor miljøforstyrrelser. I konstruksjonsfasen vil de direkte fysiske påvirkningene av bunnen (f.eks. i form av graving, pæling, sprengning, etc.) kunne ta livet av en del organismer, men det påvirkede arealet vil sannsynligvis være lite sammenlignet med ikke-påvirket areal i området. Erosjonsbeskyttende steinvoller som ofte legges rundt bunnfaste turbinfundamenter vil imidlertid kunne øke det påvirkede arealet og fungere som kunstige rev (Langhamer 2012; Glarou et al. 2020). Mobile organismer vil kunne rømme bort fra området under konstruksjonsfasen, og reetablere seg når denne perioden er over. Installasjoner på bunnen vil over tid bli tilgrodd med marine organismer. Konstruksjonen/pilarene vil her fungere som hardbunn som strekker seg opp til overflaten, men med et begrenset areal i ulike dybdeintervall sammenlignet med grunner/skvalpeskjær. Hastigheten for tilgroing er avhengig av hvilke materiale pilarene/plattformene er laget i, og om det er behandlet med eventuelle antibegroingsmiddel.
Den fysiske påvirkningen på sjøbunnen vil også være ulik for bunnfaste og flytende installasjoner (ICES 2018). Ved montering på bløt bunn vil konstruksjonen og eventuelle erosjonsbeskyttende steinvoller bli kolonisert med typiske solitære hardbunnorganismer som normalt trives i de aktuelle dypene. På denne måten vil anleggsområdene kunne bli tilført arter som ikke naturlig finnes i nærområdet, og dermed kunne bidra til spredning av fremmede arter ved at naturlige spredningsbarrierer brytes ned (Adams et al. 2014, Kerckhof et al. 2015).
Referanser
Adams TP, Miller RG, Aleynik D, Burrows MT. 2014. Offshore marine renewable energy devices as stepping stones across biogeographical boundaries. Journal of Applied Ecology, 51, 330–338.
Glarou M, Zrust M, Svendsen JC. 2020. Using Artificial-Reef Knowledge to Enhance the Ecological Function of Offshore Wind Turbine Foundations: Implications for Fish Abundance and Diversity. Journal of Marine Science and Engineering, 8, 332. doi:10.3390/jmse8050332.
ICES. 2018. Report of the Working Group on Marine Benthal and Renewable Energy Developments (WGMBRED), 6-9 March 2018, Galway, Ireland. ICES CM 2018/HAPISG:02. 66 pp.
Kerckhof F, Degraer S, Norro A, Rumes B. 2015. Succession and seasonal dynamics of the epifauna community on offshore wind farm foundations and their role as stepping stones for non-indigenous species. Hydrobiologia, 756, 37–50.
Langhamer O. 2012. Artificial reef effect in relation to offshore renewable energy conversion: state of the art. The Scientific World Journal, 2012, 1–8. doi: 10.1100/2012/386713.
3.2 - Effekter av fundamenter på bunndyrsamfunn
Havvindanlegg representerer tilførsel av nye faste flater som bunndyr kan etablere seg på, helt på linje med brofester og kaianlegg. Når en ny ren flate er tilgjengelig i havet blir den hurtig koloniseres av bunndyr som i hovedsak er filterspisere, dette føre til økt deponering av deres avfallsprodukter på bunnen rett under den nye flaten de sitter på. Organisk materiale fra vannmassene som tas opp av dyrene overføres da til biomasse på strukturene som er satt ut i havet. Videre vil avføring og døde organismer som faller ned på bunnen stedvis kunne føre til oksygensvinn tett på konstruksjonen. De nyetablerte samfunnene på strukturene vil også filtrere ut larver fra passerende vann og derfor ha effekt på rekrutteringen nedstrøms. Dette er effekter man i liten grad har kunnet kvantifisere nøye og skaden vil være den samme som for alle faste flater som introduseres i havet. Her vil selvfølgelig tettheten av slike havvindanlegg og næringstilgangen på bunnen rundt anlegget ha betydning for hvor store endringene dette vil medføre for bunnsamfunnene.
Mulig endringer forårsaket av vindkraftanlegg som er rapportert i ulike publikasjoner er:
-Effekt på energistrømmer og føde kjede dynamikken til bunndyrsamfunn (Raoux et al. 2017, 2018; Pezy et al. 2018). I en før og etter analyse av bunnfauna-næringskjeden finner Raoux et al. 2017 at: (1) Aktiviteten i økosystemet øker, andel generalister (omnivorer) og omsetningen øker. (2) Høyere nivåer i næringskjeden som fisk, marine pattedyr og sjøfugl responderer positivt på den økte biomassen på og ved anleggene. (3) Nøkkelarter endres til mer strukturerende og dominerende organismegrupper. Kort oppsummert: Rev-/hardbunns-effekt øker bunndyr-biomassen med 55%. Større forekomst av skjell gir en næringskjede som er dominert av detritus-eter. Den økte bunndyr og fisk biomasse tiltrekker seg predatorer.
-Anleggene kan fungere som “stepping stones” for innførsel av fremmede arter eller øke utbredelsen av arter (Coolen 2017) og ifølge Ricciardi and Rasmussen (1998) øke sannsynligheten for invasjoner.
-Vibrasjoner fra vindturbinene kan endre sedimentets sammensetning til en større kornstørrelse noe som så kan ha effekt på biogeokjemiske prosesser som langtids deponering av karbon i sedimentet (Pratt et al, 2014; Solan et al, 2016). Gravende kråkeboller som lever i karbon rike og fine sedimenter kan forsvinne, noe som da vil føre til lavere bioturbasjon (Wieking and Kröncke, 2003; Kröncke et al. 2004).
-Kunnskap om langtids endringer og storskala effekter av kunstige strukturer mangler, og det er ikke mulig å forutsi storskala effekten av slike (Danheim et al. 2019). Samtidig har vi lang erfaring fra kaier, brofundament, oljeinstallasjoner og mange andre strukturer som legges ut på havbunn. Det er helt klart at en havvindanleggene vil tilføre en ny bunntype som vil favorisere organismer som trenger fast underlag for å kunne etablere seg. Dette vil påvirke miljøet lokalt rundt anlegget, men hvordan effekten av disse endringer vil bli på større skala er ukjent.
Referanser
Coolen JWP. (2017) North Sea Reefs. Benthic biodiversity of artificial rocky reefs in the southern North Sea. Dissertation, Wageningen University and Research. 203 pp.
Dannheim J, Bergström L, Birchenough SNR, et al. 2019. Benthic effects of offshore renewables: identification of knowledge gaps and urgently needed research. ICES Journal of Marine Science, 77, 1092–1108.
Kröncke I, Stoeck T, Wieking G, Palojärvi A. 2004. Relationship between structural and functional aspects of microbial and macrofaunal communities in different areas of the North Sea. Marine Ecology Progress Series, 282, 13–31.
Pezy J-P, Raoux A, Dauvin J-C. 2018. An ecosystem approach for studying the impact of offshore wind farms: a French case study. ICES Journal of Marine Science, 77, 1238–1246.
Pratt DR, Lohrer AM, Pilditch CA, Thrush SF. 2014. Changes in ecosystem function across sedimentary gradients in estuaries. Ecosystems, 17, 182–194.
Raoux A, Dambacher JM, Pezy J-P, Mazé C, Dauvin J-C, Niquil N. 2018. Assessing cumulative socio-ecological impacts of offshore wind farm development in the Bay of Seine (English Channel). Marine Policy, 89, 11–20.
Raoux A, Tecchio S, Pezy J-P, Lassalle G, Degraer S, Wilhelmsson D, Cachera M. et al. 2017. Benthic and fish aggregation inside an offshore wind farm: which effects on the trophic web functioning? Ecological Indicators, 72, 33–46.
Ricciardi A, Rasmussen JB. 1998. Predicting the identity and impact of future biological invaders: a priority for aquatic resource management. Canadian Journal of Fisheries and Aquatic Sciences, 55, 1759–1765.
Solan M, Hauton C, Godbold JA, Wood CL, Leighton TG, White P. 2016. Anthropogenic sources of underwater sound can modify how sediment-dwelling invertebrates mediate ecosystem properties. Scientific Reports, 6, 20540.
Wieking G, Kröncke I. 2003. Macrofauna communities of the Dogger Bank (central North Sea) in the late 1990s: spatial distribution, species composition and trophic structure. Helgoland M arine Research, 57, 34–46.
3.3 - Effekter av fundamenter på plankton
Plankton omfatter arter som lever i de frie vannmassene gjennom hele livssyklusen (holoplankton) eller deler av livssyklusen (meroplankton). Meroplankton er som oftest larver av benthiske organismer, men inkluderer også enkelte grupper av maneter. Effekter av vindkraftanlegg på plankton er lite studert, og det finnes få empiriske resultater som påviser effekter på det planktoniske økosystemet. Eksisterende resultater baserer seg på både feltobservasjoner og modellstudier, og viser mulige effekter både som følge av endringer i stratifisering og primærproduksjon og som følge av endringer i bunnsubstrat.
Det er usikkert hvordan primærproduksjon påvirkes av havvindanlegg. Dersom konstruksjoner fører til økt vertikal miksing, vil næringsstoffer transporteres til overflaten, og gi økt primærproduksjon med oppblomstring av planteplankton som igjen har positiv innvirkning på zooplankton (Floeter et al. 2017). Imidlertid kan økt turbiditet i nærheten av konstruksjoner gi dårligere lysforhold for primærprodusenter (van der Molen et al. 2014; Vanhellemont og Ruddick 2014), med påfølgende negativ innvirkning på zooplankton. En økning i næringsstoffer og primærproduksjon i nærheten av anlegg kan gi effekter som minner om eutrofiering, med strukturelle endringer i hele planktonsamfunnet. Det er tidligere rapportert et skifte mot «små» dyreplankton, en økning i mikrozooplankton og flere detritusspisende organismer i nærheten av havvindkonstruksjoner (Wang et al. 2018, 2019).
Havvindanlegg vil gi økt areal av hardbunn. Dette vil føre til en lokal økning i mengden av bunnlevende organismer som igjen vil øke andelen av meroplankton (larver fra bunnlevende organismer) i planktonsamfunnet (Floeter et al. 2017), med påfølgende konkurranse med holoplankton (f.eks hoppekreps). En økning i bunnlevende filterspisere (f.eks blåskjell), vil også konkurrere med planktoniske organismer, og kanalisere energien mot bentisk produksjon, på bekostning av pelagiske organismer (Slavik et al. 2019).
Flere studier antyder at installasjoner i havområder fører til økt forekomst av oppblomstringer av maneter (Purcell et al. 2007; Duarte et al. 2013 og referanser i disse). Maneter, tilhørende gruppen Scyphozoa, har et bunnlevende livsstadium (polypp) som er opphav til medusestadiet gjennom strobilering (ukjønnet formering). En økning i hardbunnsubstrat vil derfor være fordelaktig for en rekke manetarter som f.eks. glassmanet (Aurelia aurita) og brennmanet (Cyanea capillata, C. lamarchii). Studier fra Østersjøen og Middelhavet har vist en økning i mengden maneter som følge av vindkraftanlegg til havs (Janßen et al. 2013; Vodopivec et al. 2017).
Referanser
Duarte CM, et al. 2012. Is global ocean sprawl a cause of jellyfish blooms? Frontiers in Ecology and the Environment, 11, 91–7.
Floeter J, et al. 2017. Pelagic effects of offshore wind farm foundations in the stratified North Sea. Progress in Oceanography 156, 154-173.
Janßen H, Augustin CB, Hinrichsen HH, Kube S 2013. Impact of secondary hard substrate on the distribution and abundance of Aurelia aurita in the western Baltic Sea. Marine Pollution Bulletin, 75, 224-234.
Vodopivec M, et al. 2017. Offshore marine constructions as propagators of moon jellyfish dispersal. Environmental Research Letters, 12, 084003.
Purcell JE. 2012. Jellyfish and ctenophore blooms coincide with human proliferations and environmental perturbations Annu. Reviews in MarineScience, 4, 209–35.
Slavik K, et al. 2019. The large-scale impact of offshore wind farm structures on pelagic primary productivity in the southern North Sea. Hydrobiologia, 845, 35–53. doi: 10.1007/s10750-018-3653-5.
van der Molen J, Smith HCM, Lepper P, Limpenny S, Rees J. 2014. Predicting the large-scale consequences of offshore wind turbine array development on a North Sea ecosystem. Continental Shelf Research, 85, 60–72. doi: 10.1016/j.
Vanhellemont Q, Ruddick K. 2014. Turbid wakes associated with offshore wind turbines observed with Landsat 8. Remote Sensing of the Environment, 145, 105–115.
Wang T, Yu W, Zou X, Zhang D, Li B, Wang J, Zhang H. 2018. Zooplankton community responses and the relation to environmental factors from established offshore wind farms within the Rudong coastal area of China.Journal of Coastal Research, 34, 843 – 855. doi: 10.2112/JCOASTRES-D-17-00058.1.
Wang J, Zou X, Yu W, Zhang D, Wang T. 2019. Effects of established offshore wind farms on energy flow of coastal ecosystems: A case study of the Rudong offshore wind farms in China, Ocean & Coastal Management, 171, 111-118.
3.4 - Effekter av fundamenter på fisk
Fysiske effekter av vindturbiner på fisk kan være både positive og negative. Negative effekter kan være at strukturene i seg selv virker avskrekkende på fisk, som f.eks. kan skremmes bort fra tradisjonelle vandringsruter og gytefelt. Positive effekter, i form av tiltrekning, vil henge sammen med tilkomst av ny struktur, som etter tid vil kunne utvikle seg til et slags kunstig rev eller såkalt FAD (Fish Aggregating Device). En nylig meta-analyse viser en økning i forekomst av fisk innenfor havvindanlegg sammenlignet med kontrollområder (Methratta & Dardick 2019). Det kan bety at fisk trekkes fra omkringliggende områder til installasjonen som vil øke biomassen av fisk i nær tilknytning til de enkelte installasjonene, mens det minsker i området rundt, eller at selve bestanden blir større. Dersom turbinene plasseres i et heterogent område med bløtbunn eller sandbunn vil turbinpilarene kunne bidra til økt struktur og hardbunnareal, med påfølgende mulighet for etablering av hardbunnarter som alger, skjell, koraller, anemoner, sekkedyr osv. og sådan danne et kunstig rev. Etablering av hardbunnarter øker strukturen ytterligere og skaper næringsrike skjul for mindre fisk og yngel. Turbinpilarene vil også kunne gi fisk og andre svømmende organismer le for kraftige havstrømmer.
Det er få studier som sammenligner områder før og etter konstruksjon av vindkraftanlegg, og de studiene som finnes viser forskjellige former for påvirkning. Et 4-årig studie som studerte endringer under utbygging av vindkraftanlegg i Belgia viser at særlig juvenile torsk og skjeggtorsk opphold seg mye innenfor vindkraftanlegg (Reubens et al. 2014). I dette tilfellet fant de ingen tegn til at fiskene var i bedre kondisjon innenfor sammenlignet med utenfor vindkraftanlegget, eller at de fikk mer mat innenfor vindkraftanlegget, selv om skjeggtorsken var større i vindkraftanlegget enn utenfor. Også to telemetri studier innenfor eksisterende vindkraftanlegg viser at torskene forsvinner fra vindkraftanlegg før de blir voksne (Winter et al. 2010; van Hal et al. 2012, 2017). Vandendriesche et al. (2014) studerte endringer i artssammenstilling i områder før og etter bygging av flere vindkraftanlegg. Dette studiet viste store forskjeller mellom to vindkraftanlegg, basert på hvilke arter som forekommer i området. De fant ingen tydelige “edge-effects”, dvs positive ringvirkninger på området rundt vindkraftanlegget, men de fant relativt større rødspette innenfor en av vindkraftanleggene som kan tyde på en skjermende effekt fra fiskeri (refuge effekt).
Kunstig rev-effekten har blitt studert mye på andre strukturer til havs. Disse studier har vist at forekomst av etablerte arter er høyere på vertikale strukturer enn på strukturer som flyter (Glasby and Connell 2001; Langhamer et al. 2009). Hvilke arter som koloniserer blir videre påvirket av en rekke andre faktorer som: strukturens alder (stå-tid), tekstur, dyp, og kompleksitet i utforming (e.g. Connell and Glasby 1999; Connell 2001; Glasby and Connell 2001; Svane and Petersen 2001; Perkol-Finkel et al. 2006). En sammenligningsstudie av stålpilar og betongpilar, har vist at ulike arter slo seg ned på de to materialene, mens pilarmaterialet ikke påvirket artssammensetningen av fisk som oppholdt seg nær pilarene (Andersson et al. 2009). Samme studien viste at der var flere fisk, og da særlig kutling (Gobidae), rundt pilarene sammenlignet med den omkringliggende bløte bunnen, men ikke i forhold til nærliggende hardbunn. Dette studiet ble utført i en grunn bukt (3-10 m dyp) på Sveriges vestkyst. Tilsvarende økte forekomster av fisk (også her mest kutling) i tilknytning til vindturbinfundamenter er blitt observert i Kalmarsundet på den svenske Østersjøkysten (6-8 m dyp, 5 - 10 km fra land; Wilhelmsson et al. 2006) og i vindkraftanlegget ved Horns Rev utenfor vestkysten av Danmark (6-14 m dyp, 14-20 km fra land; Leonhard og Pedersen 2004). Vindturbinene ved Horns Rev er imidlertid fundamentert på jevn sandbunn, slik at den relative effekten av det kunstige hardbunnsubstratet (vindturbinfundamentet) sannsynligvis vil være større enn hvis plassert på eksisterende hardbunn. Ved Vindturbinene på Horns Rev ble det ved enkelte tilfeller observert store mengder brisling og tobis, noe som tyder på at området ble utnyttet også av migrerende fisk, men kutling, leppefisk, ulke, torsk og skjeggtorsk var de mest vanlige artene. Ifølge Leonhard og Pedersen (2005) var artssammensetningen ved vindkraftanleggene lik den som finnes ved båtvrak i Nordsjøen (han refererer til Leewis et al 2000). Hvorvidt vi vil se tiltrekking og kolonisering av samme type arter på de aktuelle vindturbinlokaliteter i Norge er trolig avhengig av avstand fra land, dyp, breddegrad og havstrømmer. For eksempel har Methratta og Dardick (2019) funnet at fisken som lever på hardbunn, blir mer tiltrukket av havvindanlegg enn bløtbunn fisk og pelagiske fisk, og at fiskespisende fisk blir tiltrukket mer enn andre arter. Tiltrekkingseffekten var generelt sterkere i vindkraftanlegg på dypere vann og lenger fra kysten. Dette er viktig i et norsk perspektiv, hvor vindkraftanleggene planlegges lengre til havs enn eksisterende anlegg i Europa.
Studier av fisketetthet og artssammensetning rundt nedlagte plattformer på Ekofisk feltet i Nordsjøen (70-230 m dyp, ca 300 km fra land), viste at det på dagtid samlet seg stimer med pelagisk fisk (makrell) rundt plattformene, mens bunnfisk (dominerende arter: torsk, sei og lange) holdt seg under og i nær tilknytning til plattformene (Soldal et al. 2002; Løkkeborg et al. 2002). Fisken virket å bli tiltrukket til anlegget, da fangstene var 3-4 ganger høyere nær (100 m avstand) sammenlignet med lengre fra (165 m avstand) plattformen (Løkkeborg et al 2002). En meget omfattende langtidsstudie (1995-2013) av fiskeansamlinger rundt 23 olje- og gassrigger utenfor Californiakysten (14-363 m dyp, fra kysten til 20 km fra kysten) viste at tettheten av juvenil fisk (<20 cm) var høyere rundt installasjonene enn den var i omkringliggende naturlige habitat (Love et al. 2019), og forfatterne konkluderer med at plattformene både aggregerer voksen fisk og produserer yngel, og er dermed med å styrke fiskepopulasjonene regionalt (Love 2020). Basert på de tilsvarende studiene har det blitt konkludert at oljeplattformene utenfor California er blant verdens mest produktive marine fiskehabitat (Claisse et al. 2014). En svakhet med disse studiene fra California, og de fleste andre studiene som er diskutert her, er at der ikke er blitt gjennomført skikkelige førstudier, før installasjonene ble bygget. En vet derfor ikke om produksjonene i hele området har økt etter plattformene ble installert eller om fisken har preferanse for habitatet rund plattformen framfor de omkringliggende naturlige habitatene. Det er derfor viktig at det blir lagt inn god tid til gjennomføring av førstudier i de områder til planlagte vindkraftanlegg før utbyggingen begynner.
Referanser
Andersson MH, Berggren M, Wilhelmsson D, Öhman MC. 2009. Epibenthic colonization of concrete and steel pilings in a cold-temperate embayment: a field experiment. Helgoland Marine Research, 63, 249–260. doi: 10.1007/s10152-009-0156-9.
Claisse TJ, Pondella DJ, Love M, Zahn LA, Williams CM, Williams JP, Bull AS. 2014. Oil platforms of California are among the most productive marine fish habitats globaly. PNAS, 43, 15462-15467.
Connell SD. 2001. Urban structures as marine habitats: an experimental comparison of the composition and abundance of subtidal epibiota among pilings, pontoons and rocky reefs. Marine Environmental Research, 52, 115–125.
Connell SD, Glasby TM. 1999. Do urban structures influence local abundance anddiversity of subtidal epibiota? A case study from Sydney Harbour, Australia.Marine Environmental Research, 47, 373–387.
Glasby TM, Connell SD. 2001. Orientation and position of substrata have large effects on epibiotic assemblages. Marine Ecology—Progress Series, 214, 12–135.
Langhamer O, Wilhelmsson D, Engström J. 2009. Artificial reef effect and fouling impacts on offshore wave power foundations and buoys – a pilot study. Estuarine, Coastal and Shelf Science, 82, 426–432.
Leewis RG, van Moorsel H, Waardenburg H. 2000. Shipwrecks on the Dutch Continental Shelf as Artificial Reefs - In A. C. Jensen et al. (eds.): Artificial Reefs in European Seas, pp 419-434.
Leonhard SB, Pedersen J. 2005. Hard bottom substrate monitoring Horns rev offshore wind farm. Annual status report 2004. Bio/consult as. Doc. No. 2438-03-005 rev.
Love MS, Claisse JT, Roeper A. 2019. An analysis of the fish assemblage around 23 oil and gas platforms off California with comparison with natural habitats. Bulletin of Marine Science, 95, 477-514.
Love MS. 2020. A 45-year career in marine science—better than a sharp stick in the eye. ICES Journal of Marine Science, doi:10.1093/icesjms/fsaa002.
Løkkeborg S, Humborstad O-B, Jørgensen T, Soldal AV. 2002. Spatio-temporal variations in gillnet catch rates in the vicinity of North Sea oil platforms. ICES Journal of Marine Science, 59, S294–S299.
Methratta ET, Dardick WR. 2019. Meta-Analysis of Finfish Abundance at Offshore Wind Farms. Reviews in Fisheries Science & Aquaculture 27:242–260. doi: 10.1080/23308249.2019.1584601.
Perkol-Finkel S, Shashar N, Benayahu Y. 2006. Can artificial reefs mimic natural reef communities? The role of structural features and age. Marine Environmental Research, 61, 121–135.
Reubens J T, Degraer S, Vincx M. 2014. The ecology of benthopelagic fishes at offshore wind farms: a synthesis of 4 years of research. Hydrobiologia, 727, 121–136. doi: 10.1007/s10750-013-1793-1.
Soldal AV, Svellingen I, Jørgensen T, Løkkeborg S. 2002. Rigs-to-reefs in the North Sea: hydroacoustic quantification of fish associated with a ‘‘semi-cold’’ platform. ICES Journal of Marine Science, 59, S281–S287.
Svane I, Petersen JK. 2001. On the problems of epibioses, fouling and artificial reefs, a review. Marine Ecology, 22, 169–188
Vandendriessche S, Derweduwen J, Hostens K. 2014. Equivocal effects of offshore wind farms in Belgium on soft substrate epibenthos and fish assemblages. Hydrobiologia, 756, 19–35. DOI: 10.1007/s10750-014-1997-z.
van Hal R, Griffioen B, van Keeken OA. 2017. Changes in fish communities on a small spatial scale, an effect increased habitat complexity by an offshore wind farm. Marine Environmental Research, 126, 26–36. doi:10.1016/j.marenvres.2017.01.009.
van Hal R, Couperus B, Fassler S, Gastauer S, Griffioen B, Hintzen N, Teal L, van Keeken O, Winter E. 2012. IMARES Report C059/12. Prepared for Noordzeewind. 161 pp.
Wilhelmsson D, Malm T, Öhman MC 2006. Influence of offshore wind power on demersal fish. ICES Journal of Marine Science, 63. doi: 10.1016/j.icesjms.2006.02.007.
Winter HV, Aarts G, van Keeken OA. 2010. Residence time and behaviour ofsole and cod in the offshore windfarm Egmond aan Zee (OWEZ). IMARES. Report number OWEZ_R_265_T1_20100916.
3.5 - Effekter av fundamenter på sjøpattedyr
I enkelte områder kan det se ut som om sjøpattedyr tiltrekkes av vindkraftanlegg (Russell et al. 2014). Det kan tyde på at installasjonene fungerer som kunstige rev med økt biologisk produksjon. Det har også vært foreslått at vindkraftanlegg kan fungere som beskyttelse for sel og nise i områder med stor båttrafikk, og at tilstedeværelse (eller fravær) av sjøpattedyr kan ha sammenheng med økologiske variasjoner som påvirker utbredelse og tetthet av fiskebestander og som er uavhengig av havvindanlegg (Russell et al. 2014). Farvannene langs Norges kyst besøkes av flere arter av sjøpattedyr, men bare noen få av disse er til stede på kontinuerlig basis. Det dreier seg i første hånd om de to selartene steinkobbe og havert, samt hvalarten nise som er mest hyppig i kystnære farvann og fjorder. Andre arter som i perioder forekommer i aktuelle områder er vågehval, spekkhogger, grindhval og knølhval. I tillegg til sel har vi her valgt å fokusere på nise, dels fordi den er den mest stasjonære hvalarten i aktuelle områder, men også fordi den er desidert mest sensitiv for undervanns-støy. Av hvalartene er det derfor mest hensiktsmessig å bruke nise som indikatorart, som bør holdes under oppsyn under en utbygging av havvind-anlegg.
3.5.1 - Eksisterende kunnskap
For å kunne evaluere hvordan sjøpattedyr påvirkes av vindkraftanlegg langs norskekysten, kreves det grunnleggende data om disse dyrenes habitatbruk, altså hvor de har viktige beite og oppholdsområder. Slik kunnskap finnes det lite av langs norskekysten. Den mest aktuelle metoden for å skaffe slike data på selartene steinkobbe og havert er å utstyre dem med GPS/GSM-sendere som samler inn store mengder data på hvor selene oppholder seg i sjøen og hvor de har viktige beiteområder. Dette er nødvendig kunnskap for å minske interaksjoner ved etablering av vindkraftanlegg. Slike data er blitt innhentet i stort omfang rundt de britiske øyene (Figur 6; Russell et al. 2017). I Norge er det kun i Porsangerfjorden, ytre områder i Vesterålen og ytre Oslofjorden at det finnes slike data for steinkobbe. For havert finnes det svært lite kunnskap om habitatbruk, kun litt basert på merkinger gjort i Storbritannia (Figur 6). Det er også viktig å ta hensyn til kystselenes kasteområder på land, hvor de føder ungene sine, slik at vindkraftanlegg ikke legges tett inntil disse. Kasteområdene for havert og steinkobbe er godt kartlagt langs norskekysten (Figur 7).
For nise finnes det lite data om habitatbruk i norske farvann, både for områder brukt til reproduksjon og viktige beiteområder. I andre land (Sverige, Danmark, Tyskland, Nederland og Storbritannia) har det vært vanlig å undersøke påvirkning på niser, som følge av etablering av vindkraftanlegg. Dette gjøres ved tellinger av niser langs faste transsekter på forhånd der vindkraftanlegget planlegges og i det omkringliggende området. Slike tellinger gjennomføres også når anleggsfasen pågår og når anlegget er ferdig og driftes. Dette gir gode data for om nisene trekker bort fra området under konstruksjonsfasen og om de vender tilbake når vindkraftanlegget er etablert og i drift. I tillegg brukes det loggere for å registrere lyd fra niser både innenfor og utenfor vindmølleområde for å innhente samme type kunnskap. Slike tellinger bør også gjennomføres i Norge i områder hvor havvindalegg planlegges.
For vindkraftanlegg i drift er det ikke entydige resultater om påvirkning av sel og nise. I noen områder tyder tilstedeværelse og antall sjøpattedyr på at vindmølleområdene ikke påvirker sjøpattedyr (Russell et al. 2016). I andre områder kan det se ut som om sjøpattedyr tiltrekkes vindmøllene (Russell et al. 2014). Det kan tyde på at installasjonene fungerer som kunstige rev med økt biologisk produksjon.
Figur 6: Sporing av 270 havert (øverst til venstre: 1991 to 2016) og 330 steinkobbe (øverst til høyre 2001 to 2016) merket i VK som ble brukt for å generer habitatbrukskartene under. Figur fra Russell et al. 2017.
Figur 7: Faste kaste-lokaliteter til havert (venstre) og hårfellings-lokaliteter til steinkobbe (høyre) langs Norskekysten. For havert er kun lokaliteter markert, mens for steinkobbe angis estimert størrelse representert av størrelse på symbolene. Dette er et minimums-estimat basert på tellinger under hårfellingen. Figur fra Russel et al. 2017.
3.5.2 - Nødvendig ny kunnskap i norske farvann
For sjøpattedyrene er det viktig å innhente kunnskap om habitatbruk før nye vindkraftanlegg anlegges. Dette krever økt innsats i form av merking av sel med sendere i nærliggende områder for å dokumentere om anleggsområdet har betydning for selene, særlig om det er viktig som beiteområde. Det krever også økt innsats på nise ved telling og akustisk registrering av dens ekkolokalisering for å innhente samme type dokumentasjon. Tellinger av nise til nå har vært sjeldne og med mangelfull dekning. Det er heller ikke mye kunnskap om nisenes habitatbruk og migrasjoner langs norskekysten.
Det er utviklet et teoretisk rammeverk for å estimere langsiktig påvirkning på steinkobbepopulasjoner som følge av forstyrrelser fra pælingsfasen for vindkraftanlegg i Skottland (Thompson et al. 2013). Geografisk fordeling av steinkobber modelleres sammen med lyd fra pæling i gitte områder. Kunnskap om selpopulasjonens produksjonsrate og vekst inngår i modellen, noe som gjør det mulig å estimere påvirkning av selpopulasjonen i løpet av 25 år. Eventuell langvarig påvirkning av anleggs- og produksjonsfasen for vindkraftanlegg på populasjonsnivå for kystsel i Norge vil være vanskelig å påvise i ettertid uten at det innhentes ny kunnskap. Bestandsendringer kan påvises i ettertid gjennom etablerte tellinger og bestandsestimeringer, men det vil være vanskelig å evaluere om endringer skyldes påvirkning fra vindkraftanlegg, uten kunnskap om selenes habitatbruk.
Referanser
Russell DJF, Brasseur SMJM, Thompson D, Hastie GD, Janik VM, Aarts G, McClintock BT, Matthiopoulos J, Moss SEW, McConnell B. 2014. Marine mammals trace anthropogenic structures at sea. Current Biology, 24, R638. doi: 10.1016/j.cub.2014.06.033.
Russell DJF, Hastie GD, Thompson D, Janik VM, Hammond PS, Scott-Hayward LAS, Matthiopoulos J, Jones EL, McConnell BJ 2016. Avoidance of wind farms by harbour seals is limited to pile driving activities. Journal of Applied Ecology, 53, 1642-1652. DOI: 10.1111/1365-2664.12678.
Russell DJF, Jones EL, Morris CD. 2017. Updated Seal Usage Maps: The Estimated at-sea Distribution of Grey and Harbour Seals. Scottish Marine and Freshwater Science Report Vol 8 No 25. Published by Marine Scotland Science, ISSN: 2043-7722 doi: 10.7489/2027-1. 30 pp.
Steen H, Nilssen KT, Agnalt A-L, Alvsvåg J, Asplin L, Jelmert A, Dahl E, Dalen J. 2008. Marinøkologiske ringvirkninger av vindkraftanlegger til havs. Fisken og Havet 9-2008. 20 pp.
Thompson PM, Hastie GD, Nedwell J, Barham R, Brookes KL, Cordes LS, Bailey H, McLean N. 2013. Framework for assessing impacts of pile-driving noise from offshore wind farm construction on a harbour seal population. Environmental Impact Assessment Review, 43, 73–85.
3.6 - Oppsummering kunnskapshull effekter av fundamenter
Det finns flere kunnskapshull om hvordan fundamenter av vindkraftanlegg kommer til å påvirke havmiljøet. Herunder oppsummerer vi de viktigste:
Kunnskap om langtids endringer og storskala effekter av kunstige strukturer mangler.
Det er usikkert hvordan primærproduksjon påvirkes av havvindanlegg.
De nyetablerte samfunnene på strukturene vil filtrere ut larver fra det forbipasserende vann og derfor ha effekt på rekrutteringen nedstrøms. Dette er effekter man i liten grad har kunnet kvantifisere nøye.
Det er usikkert om lokale endringer i artssammensetning og mengde fisk som følge av rev-effekt fører til populasjonseffekter.
Det er lite kunnskap om hvordan pelagiske fisk blir påvirket, selv om spesielt flytende strikturer kan tiltrekke seg pelagisk fisk.
For å kvantifisere effekter må man sammenligne områder før og etter utbygging. Det har ikke blitt utført lange nok før-studier for spesielt fisk og sjøpattedyr.
For sjøpattedyr er det spesielt viktig å innhente kunnskap om habitatbruk for nise og sel i området før nye vindkraftanlegg anlegges.
4 - Effekter av elektromagnetisk felt på bevegelsesmønstre og adferd
4.1 - Bakgrunn
Noen marine dyr har utviklet særlige evner til å oppdage svake elektriske felt. Særlig kjent er bruskfisk (haier og skater), men også havmus, niøye, stjernetitterfisk, stør, maller og kvastfinnefisker er elektro følsomme (Alves-Gomes 2001, Collin og Whitehead 2004, Walker 2001). I tillegg har laks, ål og delfin vist responser på svake elektriske felt, og det er indisier på at rødspette potensielt orienterer seg i forhold til elektriske felt som genereres av havstrømmer (Metcalfe et al. 1993, Rommel Jr. og McCleave 1973, Czech-Damal et al. 2011).
Elektriske signaler har et potensielt bredere bruksområde for marine dyr enn magnetiske signaler, spesielt blant bruskfisk. Mens magnetiske signaler hovedsakelig brukes til orientering, brukes elektriske signaler også til å oppdage bytte og unngå predasjon, og i sosiale interaksjoner (Collin og Whitehead 2004). Elektriske felt har derfor potensial til å forstyrre et bredere adferdsspekter enn magnetiske felt. Terskelverdier for elektriske felt i forhold til deteksjon, adferdsendring eller fysiologiske effekter hos marine dyr varierer mye, og bruskfisk er klart mest følsomme (Nyqvist et al. 2020).
4.2 - Effekter av elektromagnetisk felt på bunndyrsamfunn
Hos evertebrater har moderate effekter av elektromagnetiske forstyrrelser blitt observert i enkelte eksperiment. Taskekrabbe (Cancer pagurus), utsatt for maksimale feltstyrker fra undervannskabler (2 800 - 40 000 µT) i 24 timer fikk redusert aktivitet og viste preferanse for magnetisk eksponerte områder i et bur (Scott et al. 2018). Hos juvenil hummer (Homarus gammarus) fant man imidlertid ikke noen effekter på aktivitet og foretrukket område under mer realistiske feltstyrker (maksimal intensitet på 200 μT; Taormina et al. 2019). Undervannskabler påvirket heller ikke merkbart bevegelsen hos en annen krabbeart (Cancer magister) i en feltstudie der krabbene krysset kabler (Love et al 2017).
Spesielt for dyr som oppholder seg i kablenes nærsoner i lengre tidsrom kan de elektromagnetiske feltene tenkes å gi fysiologiske effekter. Formicki et al. (2019) refererer effekter på spermatozobevegelse, befruktningsgrad og egginkubasjonsperiode for en rekke fiskearter, og Juutilainen (2005) rapporterer om utviklingseffekter i fisk og kråkebolle embryoer fra eksponering for magnetiske feltstyrker i området 0,1-10 mT. Dette vil likevel vanligvis bare ramme en svært liten fraksjon av individene i en populasjon, da denne type effekter vil være lokale og begrenset til kablenes nærsoner.
4.3 - Effekter av elektromagnetisk felt på fisk og sjøpattedyr
Elektromagnetiske felt fra undervannskabler kan i teorien påvirke marine dyrs migrasjon, lokale bevegelser (attraksjon/unnvikelse), predator/byttes interaksjoner og dyrenes fysiologi/utvikling (Taormina et al. 2018; Alberta et al. 2020, Nyqvist et al. 2020). Det er utført en del studier på adferd hos marine dyr i relasjon til elektromagnetisme fra undervannskabler eller tilsvarende feltstyrker og resultatene viser marginale effekter på migrasjon og lokal adferd (Taormina et al. 2018, 2019).
For dyr som migrerer over lange avstander kan magnetfeltene fra undervannskablene forstyrre svømmeretning og migrasjon (Taormina et al. 2018). Endringer i svømmeretning og redusert svømmehastighet er observert for ål (Anguila anguila) som passerer elektromagnetiske felt fra undervannskabler, men de elektromagnetiske feltene representerte ingen barriere for ålens migrasjon (Westerberg og Lagenfelt 2008, Öhman et al. 2007). Det har også blitt foreslått at dyr bruker lokale anomalier i det naturlige magnetfeltet, f.eks. daler eller topper i magnetfeltet, til lokal orientering (Klimley 1993, Vanselow et al. 2018). Det er derfor tenkbart at magnetfelt fra undervannskabler kan maskere disse anomaliene og dermed forstyrre dyrenes orientering og bevegelse.
Det finnes noen studier som har påvist effekter av undervannskabler på adferd hos bruskfisk. Her er det trolig at bruskfisken føler det elektriske feltet som induseres av magnetfeltet, enten i vannet eller i fisken selv. Liten skate (Leucoraja erinacea) økte svømmehastigheten sin og beveget seg over et større område når den ble utsatt for elektromagnetiske felt fra en undervannskabel (Hutchison et al. 2018). I en annen studie ble småflekket rødhai (Scyliorhinus canicula) tiltrukket av magnetfelt likt det som produseres av undervannskabler (Gill og Taylor 2001). Bruskfisk kan bruke elektromagnetiske felt for både predasjon flukt, kommunikasjon og orientering (Collin og Whitehead 2004). Det er altså mulig at kabler kan påvirke flere typer adferd hos disse organismene. Man bør huske at bruskfisk kan tilvennes kunstige elektromagnetiske felt, noe som vil kunne modifisere effektene (Kimber et al. 2011, 2014).
Informasjonen om eventuelle påvirkninger av elektromagnetisme på sjøpattedyr er svært begrenset. Det er velkjent at mange sjøpattedyrarter bruker geomagnetisme til navigasjon (Walker et al. 2003), og noen studier har funnet sammenheng mellom mass-strandinger av hval og lokale geomagnetiske aberrasjoner (Kirschvink et al. 1986).
4.4 - Oppsummering kunnskapshull effekter av elektromagnetisme
Det finns flere kunnskapshull om hvordan elaktromagnetisme fra vindkraftanlegg kommer til å påvirke havmiljøet. Herunder oppsummerer vi de viktigste:
På hvor lang avstand fra kabelen finner man effekter?
Kan zooplankton bli påvirket når de blir ført langs kabler med havstrømmer?
Kan kabler forstyrre migrasjonsruter?
Referanser
Alberta L, Deschamps F, Joliveta A, Olivier F, Chauvaud L, Chauvaud S. 2020. A current synthesis on the effects of electric and magnetic fields emitted by submarine power cables on invertebrates. Marine Environmental Research. 159, 104958.
Alves‐Gomes J. 2001. The evolution of electroreception and bioelectrogenesis in teleost fish: a phylogenetic perspective. Journal of Fish Biology, 58, 1489-1511.
Collin SP, Whitehead D. 2004. The functional roles of passive electroreception in non-electric fishes. Animal Biology, 54, 1-25.
Czech-Damal NU, Liebschner A, Miersch L, Klauer G, Hanke FD, Marshall C, Dehnhardt G, Hanke W. 2011. Electroreception in the Guiana dolphin ( Sotalia guianensis ). Proceedings of the Royal Society B: Biological Sciences, 279, 663-668.
Formicki K, Korzelecka‐Orkisz A, Tański A. 2018. Magnetoreception in fish. Journal of Fish Biology, 95, 73– 91. https://doi.org/10.1111/jfb.13998
Hutchison Z, Sigray P, He H, Gill A, King J, Gibson C. 2018. Electromagnetic Field (EMF) Impacts on Elasmobranch (shark, rays, and skates) and American Lobster Movement and Migration from Direct Current Cables. Sterling (VA): US Department of the Interior, Bureau of Ocean Energy Management. OCS Study BOEM 3.
Juutilainen J. 2005. Developmental effects of electromagnetic fields. Bioelectromagnetics 26, S7.
Kimber JA, Sims DW, Bellamy PH, Gill AB. 2011. The ability of a benthic elasmobranch to discriminate between biological and artificial electric fields. Marine Biology, 158, 1-8.
Kimber JA, Sims DW, Bellamy PH, Gill AB. 2014. Elasmobranch cognitive ability: using electroreceptive foraging behaviour to demonstrate learning, habituation and memory in a benthic shark. Animal cognition, 17, 55-65.
Kirschvink JL, Dizon AE, Westphal JA. 1986. Evidence from Strandings for Geomagnetic Sensitivity in Cetaceans. Journal of Experimental Biology, 120, 1–24. https://jeb.biologists.org/content/120/1/1
Klimley A. 1993. Highly directional swimming by scalloped hammerhead sharks, Sphyrna lewini , and subsurface irradiance, temperature, bathymetry, and geomagnetic field. Marine Biology, 117, 1-22.
Love MS, Nishimoto MM, Clark S, McCrea M, Bull AS. 2017. Assessing potential impacts of energized submarine power cables on crab harvests. Continental Shelf Research, 151, 23-29.
Metcalfe J, Holford B, Arnold G. 1993. Orientation of plaice (Pleuronectes platessa) in the open sea: evidence for the use of external directional clues. Marine Biology, 117, 559-566.
Nyqvist D, Durif C, Johnsen MG, de Jong K, Forland TN, Sivle LD. 2020. Electric and magnetic senses in marine animals, and potential behavioral effects of electromagnetic surveys, Marine Environmental Research, doi: 10.1016/j.marenvres.2020.104888.
Rommel Jr S, McCleave JD. 1973. Sensitivity of American eels (Anguilla rostrata) and Atlantic salmon ( Salmo salar ) to weak electric and magnetic fields. Journal of the Fisheries Board of Canada, 30, 657-663.
Taormina B, Bald J, Want A, Thouzeau G, Lejart M, Desroy N, Carlier A. 2018. A review of potential impacts of submarine power cables on the marine environment: Knowledge gaps, recommendations and future directions. Renewable and Sustainable Energy Reviews, 96, 380-391.
Vanselow KH, Jacobsen S, Hall C, Garthe S. 2018. Solar storms may trigger sperm whale strandings: explanation approaches for multiple strandings in the North Sea in 2016. International Journal of Astrobiology, 17, 336-344.
Walker T. 2001. Basslink project review of impacts of high voltage direct current sea cables and electrodes on chondrichthyan fauna and other marine life. Marine and Freshwater Resources Institute Report, 20, 68.
Walker MM, Diebel CE, Kirschvink JL. 2003. Detection and use of the Earth’s Magnetic Field by Aquatic Vertebrates. In S. P. Collins & N. J. Marshall (Eds.), Sensory Processing in Aquatic Environments, Springer, pp. 53–74.
Westerberg H, Lagenfelt I. 2008. Sub‐sea power cables and the migration behaviour of the European eel. Fisheries Management and Ecology, 15, 369-375.
Öhman MC, Sigray P, Westerberg H. 2007. Offshore windmills and the effects of electromagnetic fields on fish. AMBIO: A journal of the Human Environment, 36, 630-633.
5 - Effekter av støy på artsfordeling og atferd
5.1 - Bakgrunn
Vindkraftanlegg øker støy i områder både under utbygging og under driftsfasen (se avsnitt 3.5). Flere rapporter har nylig beskrevet effekten og kunnskapshull om effekter av støy i utbyggingsfase (Thomsen et al. 2016; Copping og Hemery 2020), og vi skal kort oppsummere kunnskapen nedenfor. Det blir ofte argumentert med at fordi støy produsert i utbyggingsfase er mye høyere en støy produsert i driftsfasen, er risiko for påvirkning høyere i driftsfasen. Men, mens utbyggingsfasen er avgrenset over tid, kan kontinuerlig lyd under driftsfasen øke støynivået i havområder i flere år. Dette kan spesielt påvirke de dyrene som lever i eller nær vindkraftanleggene (Tougaard et al. 2009; Marmo et al. 2013). Mange dyr, fra krepsdyr til hval, kan bli forstyrret av lydendringer (f.eks. Hawkins og Popper 2017; Popper og Hawkins 2019; Kvadsheim et al. 2020). Støy kan dermed påvirke for eksempel predator-byttedyr interaksjoner (Simpson et al. 2016), reproduksjon (de Jong et al. 2020) og hvordan evertebrater påvirker økosystemet (Solan et al. 2016). Det har vært vanskelig å finne en måte å kvantifisere artenes eksponering for kontinuerlig lyd i havet, men nylig har det blitt foreslått å bruke en kombinasjon av andelen av en populasjon som utsettes for økt lydnivå og varigheten av støyeksponeringen (Merchant et al. 2017). Den metoden gir en målestokk for hvor utbredt støypåvirkningen er, men den er fortsatt avhengig av at man setter en grenseverdi for antatt akseptabelt støynivå. Fordelen med å bruke denne metoden er at man kan planlegge vindkraftanlegg på et sted hvor man unngår overlapp med viktige populasjoner (Figur 8).
Figur 8: En mulig tilnærming for å kalkulere eksponeringsrisiko for lyd i et område til en bestemt art (Figur fra Merchant et al. 2017). Merk at kurven i E er sterkt avhengig av terskelverdien som er satt for lydnivået man kaller for eksponering, fordi den blir beregnet ut ifra hvilken del av populasjonen som blir utsatt (x-akse) og i hvilken del av perioden den blir utsatt (y-akse) for et støynivå som ligger over terskelverdien.
Referanser
Copping AE, Hemery LG. 2020. OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development Around the World. Report for Ocean Energy Systems (OES).
de Jong K, Forland TN, Amorim MCP, Rieucau G, Slabbekoorn H, Sivle LD. 2020. Predicting the effects of anthropogenic noise on fish reproduction. Reviews in Fish Biology and Fisheries. doi: 10.1007/s11160-020-09598-9.
Hawkins AD, Popper AN. 2017. A sound approach to assessing the impact of underwater noise on marine fishes and invertebrates. ICES Journal of Marine Science 74:635–651. doi: 10.1093/icesjms/fsw205.
Kvadsheim PH, Forland TN, de Jong K, Nyqvist D, Grimsbo E, Sivle LD. 2020. Effekter av støyforurensning på havmiljø - kunnskapsstatus og forvalningsrådgiving. FFI-Rapport 20/01015.
Merchant ND, Faulkner RC, Martinez R. 2017. Marine Noise Budgets in Practice. Conservation Letters, 11, e12420. doi: 10.1111/conl.12420.
Marmo B, Roberts I, Buckingham MP, King S, Booth C 2013. Modelling of Operational Offshore Wind Turbines including noise transmission through various foundation types, Scottish Government Edingburgh.
Popper AN, Hawkins AD. 2019. An overview of fish bioacoustics and the impacts of anthropogenic sounds on fishes. Journal of Fish Biology, 94, 692–713. doi: 10.1111/jfb.13948.
Simpson SD, Radford AN, Nedelec SL, Ferrari MC, Chivers DP, McCormick MI, Meekan MG. 2016. Anthropogenic noise increases fish mortality by predation. Nature Communications, 7, 10544. doi: 10.1038/ncomms10544.
Solan M, Hauton C, Godbold JA, Wood CL, Leighton TG, White P. 2016. Anthropogenic sources of underwater sound can modify how sediment-dwelling invertebrates mediate ecosystem properties. Scientific reports, 6, 20540.
Thomsen F, Gill A, B, Kosecka M, Andersson M, André M, Degraer S, Folegot T, Gabriel J, Judd A, Neumann T, Norro A, Risch D, Sigray P, Wood D, Wilson B. 2016. MaRVEN – Environmental Impacts of Noise Vibrations and Electromagnetic Emissions from Marine Renewable Energy (RTD-KI-NA-27738-EN-N), European Commission, Brussels.
Tougaard J, Henriksen OD, Miller LA. 2009. Underwater noise from three types of offshore wind turbines:Estimation of impact zones for harbor porpoises and harbor seals. Journal of the Acoustical Society of America, 125, 3766-3773.
5.2 - Effekter av støy på bunndyrsamfunn
Mange evertebrater kan oppfatte vibrasjoner som henger sammen med lavfrekvent lyd, og reagere med endringer i adferd, fysikk eller fysiologi (Roberts og Elliot 2017; Popper og Hawkins 2018). Det har vært mest fokus i litteraturen på skadeeffekter av impulsiv støy og vibrasjoner på evertebrater (Edmonds et al. 2016; Roberts & Breithaupt 2016), men kontinuerlig støy kan teoretisk også føre til adferdsendringer. Det er, for eksempel, vist at både muslinger (Mytilus edulis) og sekkedyr (Ciona intestinalis) slår seg ned hurtigere når de hører båtstøy (McDonald et al. 2014; Jolivet et al. 2016), dette kan føre til mer begroing på mer støyete strukturer (Stanley et al. 2014). Det er derfor en mulighet at økt støynivå er en av grunnene at anleggene raskt blir kolonisert av bunnlevende evertebrater. McDonald et al. 2014 fant også en økt overlevelse i laboratorietester med støy sammenlignet med kontrolltester. På den andre siden har laboratoriestudier vist at støy kan påvirke evertebrater negativt og føre til atferdsendringer som kan påvirke økosystemprosesser, som, for eksempel, sediment miksing (Solan et al. 2016). Det finnes derfor foreløpig for lite data for å konkludere om støy fra vindturbiner vil ha mest positive eller mest negative effekter på evertebrater.
Referanser
Edmonds NJ, Firmin CJ, Goldsmith D, Faulkner RC, Wood DT. 2016. A review of crustacean sensitivity to high amplitude underwater noise: data needs for effective risk assessment in relation to UK commercial species. Marine Pollution Bulletin, 108, 5–11.
Jolivet A, Tremblay R, Olivier F, Gervaise C, Sonier R, Genard B, Chauvaud L. 2016. Validation of trophic and anthropic underwater noise as settlement trigger in blue mussels. Scientific Reports, 6. doi: 10.1038/srep33829.
McDonald JI, Wilkens SL, Stanley JA, Jeffs AG. 2014. Vessel generator noise as a settlement cue for marine biofouling species. Biofouling, 30, 741–749. doi: 10.1080/08927014.2014.919630.
Roberts L, Breithaupt T. 2016. Sensitivity of Crustaceans to Substrate-Borne Vibration. In: The Effects of Noise on Aquatic Life II. The Effects of Noise on Aquatic Life II. Springer, 925–931.
Roberts L, Elliott M. 2017. Good or bad vibrations? Impacts of anthropogenic vibration on the marine epibenthos. Science of the Total Environment, 595, 255–268. doi: 10.1016/j.scitotenv.2017.03.117.
Solan M, Hauton C, Godbold JA, Wood CL, Leighton TG, White P. 2016. Anthropogenic sources of underwater sound can modify how sediment-dwelling invertebrates mediate ecosystem properties. Scientific reports, 6, 20540.
Stanley JA, Wilkens SL, Jeffs AG. 2014. Fouling in your own nest: vessel noise increases biofouling. Biofouling, 30, 837–844. doi: 10.1080/08927014.2014.938062.
5.3 - Effekter av støy på zooplankton
Det er fortsatt diskusjon om over hvilke avstander plankton kan skades av lyd fra seismikk (McCauley et al. 2017; Fields et al. 2019), men det er tydelig at høy impulsiv lyd kan skade zooplankton nær lydkilden. Det er derfor mulig at zooplankton innenfor anleggsområdet kan bli skadet av lyd fra sprenging og pæling i utbyggingsfasen. Det er imidlertid mindre sannsynlig at den lavere kontinuerlige støyen i driftsfasen vil skade plankton fysisk. Det er usikkert om plankton bruker lyd i adferd, selv om vi vet at de lager lyd (Kuhn 2018). Fiskelarver derimot, bruker vannbevegelse til å oppdage byttedyr. Fordi lydbølger også beveger partikler i vannet, kan disse signalene bli forstyrret av støy. Det finnes så vidt vi vet, ingen studier som har testet effekter av støy fra vindturbiner på fiskelarver, men studier som tester effekter av skipsstøy kan brukes til å anta hva som skjer. Dette er støy som ligger i samme frekvensområde og på omtrent samme nivå som den som produseres av vindturbiner. Eksperimenter med eksponering av fiskelarver til skipsstøy har generelt ikke vist økt dødelighet (Kvadsheim et al. 2020). Unntaket er en studie (Nedelec et al 2016) som viste økt bruk av plommesekk og en tidvis redusert vekst tidlig i utviklingen hos torskelarver, men som larvene hentet inn over tid. Kroppsformen derimot ble annerledes for larvene som var eksponert for skipstøy. Lignende resultater ble observert for egg til to arter av revfisk, hvor embryoene hadde en økt hjertefrekvens og mindre plommesekk ved klekking når eggene ble utsatt for skipstøy. Om dette skal ha en effekt på populasjonsnivå, må en stor del av eggene og larvene bli utsatt av støy i den tiden rundt klekking og når de begynner å spise selv. Det kan muligens skje hvis vindkraftanlegg blir bygd ut innenfor viktige gyteområder eller områder hvor en stor del av larvene driver gjennom. Forskjellige arter av fiskelarver anvender lyd for å finne foretrukne habitater til å etablere seg. (e.g. Radford et al. 2011; Parmentier et al. 2015). Maskering av naturlig lyd fra en kontinuerlig øking i støynivå i havet kan derfor teoretisk påvirke orienteringsevne i fiskelarver, men det er ennå ikke påvist at detter skjer i realitet, og hvor viktig lyd er sammenlignet med annet sensorisk informasjon (Siebeck et al. 2015; Huijberts et al 2015).
Referanser
Fakan EP, McCormick MI. 2019. Boat noise affects the early life history of two damselfishes. Marine Pollution Bulletin, 141, 493–500. doi: 10.1016/j.marpolbul.2019.02.054.
Fields DM, Handegard NO, Dalen J, Eichner C, Malde K, Karlsen Ø, Skiftesvik AB, Durif CMF, Browman HI. 2019. Airgun blasts used in marine seismic surveys have limited effects on mortality, and no sublethal effects on behaviour or gene expression, in the copepod Calanus finmarchicus. ICES Journal of Marine Science, 76, 2033–2044. doi: 10.1093/icesjms/fsz126.
Kuhn, S. 2018. Can you hear the small talk? Master Thesis. University of Groningen.
Kvadsheim PH, Forland TN, de Jong K, Nyqvist D, Grimsbo E, Sivle LD. 2020. Effekter av støyforurensning på havmiljø - kunnskapsstatus og forvalningsrådgiving. FFI-Rapport 20/01015.
McCauley RD, Day RD, Swadling KM, Fitzgibbon QP, Watson RA, Semmens JM. 2017. Widely used marine seismic survey air gun operations negatively impact zooplankton. Nature Ecology & Evolution, 1. doi: 10.1038/s41559-017-0195.
Nedelec SL, Radford AN, Pearl L, Nedelec B, McCormick MI, Meekan MG, Simpson SD. 2017. Motorboat noise impacts parental behaviour and offspring survival in a reef fish. Proceedings of the Royal Society B, 284, 20170143. DOI: 10.1098/rspb.2017.0143.
Radford CA, Stanley JA, Simpson SD, et al. 2011. Juvenile coral reef fish use sound to locate habitats. Coral Reefs, 30, 295-305.
Siebeck UE, O'Connor J, Braun C, Leis JM. 2015. Do human activities influence survival and orientation abilities of larval fishes in the ocean? Integrative Zoology, 10, 65–82.
5.4 - Effekter av støy på voksen fisk
Støy fra pæling i utbyggingsfase kan skade fisk direkte, og føre til skader i svømmeblære, lever og hørselsorganet (Popper et al. 2014). Dette skjer bare svært nært selve lydkilden og kan begrenses med boblegardin (se avsnitt 2.4). Ved større avstander kan imidlertid atferden til fisk bli påvirket (Herbert-Read et al, 2017). De fleste studier på effekter av støy fra havvindanlegg på fisk har fokusert på effekter av støy i utbyggingsfasen (Thomsen et al. 2016). For seismikk fraråder Havforskningsinstituttet seismikk i gyteområder og konsentrerte gytevandringsområder for å unngå at unnvikelse på grunn av støy kan føre til effekter på populasjonsnivå (Sivle et al. 2020). Det kan være aktuelt å unngå konstruksjonsarbeid i gyteperioder, slik at støy fra anleggsarbeidet ikke fører til unnvikelse av gyteområder eller viktige migrasjonsruter. I Norge er gyteområdene og gyteperioder relativt godt kartlagt for økonomisk viktige arter, men det er mindre kunnskap om hvor og når ikke-kommersielle arter, herunder også enkelte rødlistede arter og nøkkelarter for økosystemet, gyter (Sivle et al. 2020). Derfor er grundige før-studier av områdene som er foreslått til utbygging viktig for å unngå forstyrrelse av gyteområder for viktige arter som er ikke kartlagt.
Støy i driftsfasen har ikke blitt antatt å gi nevneverdige effekter fordi lyd er antageligvis ikke er på mye høyere nivå enn skipsstøy (e.g. Thomsen et al. 2016: Copping og Hemery 2020). For marine pattedyr har lydnivået blitt sett på som ufarlig, fordi risikoen for at de oppholder seg lengre enn 24 timer innenfor rekkevidden av der hvor lydnivået kunne føre til hørselsskader er lav (Tougaard et al. 2009; Weissenberger 2019). Fisk kan imidlertid oppholde seg i vindturbinenes nærsoner i lengre perioder og dermed eksponeres for mer langvarige forhøyede støynivåer (se avsnitt 3.4). I tillegg viser nyere studier at skipsstøy kan påvirke fisk både fysiologisk og atferdsmessig (se de Jong et al. 2020 og referanser derin). Skipstøy kan påvirke reproduksjon, beiteadferd og unnvikelse av predatorer hos fisk (Nedelec et al. 2016; Simpson et al. 2016; Magnhagen et al. 2017), og kontinuerlig lavfrekvent støy kan muligens også ha en større påvirkning på reproduksjon hos fisk enn impulsiv støy (de Jong et al. 2020). Teoretisk er det også mulig at maskering av naturlig lyd kan påvirke orienteringsevner til fisk hvis de bruker lyd til orientering (Siebeck et al. 2015). I utbyggingsfasen kan fisk tidvis unngå området, men når anlegget er ferdig vil den kontinuerlige støyen vedvare i hele anleggets levetid. I tillegg kan forankring av flytende turbiner produsere kraftige smell (se avsnitt 2.4.4). Dersom fisk blir tiltrukket av strukturene i vindkraftanlegg (se avsnitt 3.4), og der opplever nedsatt overlevelse eller reproduksjon, kan det føre til at anlegget påvirker fiskepopulasjoner negativt (Figur 1). Det er uvisst om fiskene vil venne seg til støy over tid eller om reproduksjonen og beiteadferd kan bli påvirket over flere måneder eller år (de Jong et al. 2020; Kvadsheim et al. 2020).
Referanser
de Jong K, Forland TN, Amorim MCP, Rieucau G, Slabbekoorn H, Sivle LD. 2020. Predicting the effects of anthropogenic noise on fish reproduction. Reviews in Fish Biology and Fisheries. doi: 10.1007/s11160-020-09598-9.
Herbert-Read JE, Kremer L, Bruintjes R, Radford AN, Ioannou CC. 2017. Anthropogenic noise pollution from pile-driving disrupts the structure and dynamics of fish shoals. Proceedings of the Royal Society B: Biological Sciences, 284, 20171627. https://doi.org/10.1098/rspb.2017.1627
Kvadsheim PH, Forland TN, de Jong K, Nyqvist D, Grimsbo E, Sivle LD. 2020. Effekter av støyforurensning på havmiljø - kunnskapsstatus og forvalningsrådgiving. FFI-Rapport 20/01015.
Magnhagen C, Johansson K, Sigray P. 2017. Effects of motorboat noise on foraging behaviour in Eurasian perch and roach: a field experiment. Marine Ecology Progress Series, 564, 115–125. doi: 10.3354/meps11997.
Nedelec SL, Radford AN, Pearl L, Nedelec B, McCormick MI, Meekan MG, Simpson SD. 2017. Motorboat noise impacts parental behaviour and offspring survival in a reef fish. Proceedings of the Royal Society B, 284, 20170143. doi: 10.1098/rspb.2017.0143.
Popper AN, Hawkins AD, Fay RR, Mann DA, Bartol S, Carlson TJ, Coombs S, Ellison WT, Gentry RL, Halvorsen MB, Løkkeborg S, Rogers PH, Southall BL, Zeddies DG, Tavolga WN. 2014. ASA S3/SC1.4 TR-2014 Sound Exposure Guidelines for Fishes and Sea Turtles: A Technical Report prepared by ANSI-Accredited Standards Committee S3/SC1 and registered with ANSI. Springer International Publishing. doi: 10.1007/978-3-319-06659-2.
Siebeck UE, O'Connor J, Braun C, Leis JM. 2015. Do human activities influence survival and orientation abilities of larval fishes in the ocean? Integrative Zoology, 10, 65–82.
Simpson SD, Radford AN, Nedelec SL, Ferrari MC, Chivers DP, McCormick MI, Meekan MG. 2016. Anthropogenic noise increases fish mortality by predation. Nature Communications, 7, 10544. doi: 10.1038/ncomms10544.
Sivle LD, Forland TN, de Jong K, Nyqvist D, Grimsbø E. 2020. Havforskningsinstituttets rådgivning for menneskeskapt støy i havet: seismikk, elektromagnetiske undersøkelser og undersjøiske sprengninger. Rapport fra havforskningen 2020-1. ISSN: 1893-4536.
Thomsen F, Gill A, B, Kosecka M, Andersson M, André M, Degraer S, Folegot T, Gabriel J, Judd A, Neumann T, Norro A, Risch D, Sigray P, Wood D, and Wilson B. 2016. MaRVEN – Environmental Impacts of Noise Vibrations and Electromagnetic Emissions from Marine Renewable Energy (RTD-KI-NA-27738-EN-N), European Commission, Brussels.
Tougaard J, Henriksen OD, Miller LA. 2009. Underwater noise from three types of offshore wind turbines:Estimation of impact zones for harbor porpoises and harbor seals. Journal of the Acoustical Society of America, 125, 3766-3773.
De mest aktuelle artene av sjøpattedyr, som kan komme i konflikt med vindkraftanlegg til havs, er nise og kystselartene steinkobbe og havert. Lyd fra vindkraftanlegg kan påvirke enten gjennom å gi hørselskader, eller ved å påvirke adferd (f.eks. habitatsunnvikelse). Hørselskader er mest sannsynlig i anleggsfasen, hvor det gjerne brukes pæling for å plassere fundamentene i grunnen. Det er anleggsfasen av vindkraftanlegg som er mest forstyrrende for disse artene. Anleggsfasen er også preget av økt skipstrafikk, som kan medføre støy av båter og rigger som kan forstyrre sjøpattedyr, men antagelig er det mest forstyrrende når fundamentene pæles ned i bunnen.
Hørselsskade kan være enten midlertidig (temporær; TTS) eller varig (permanent; PTS), og hvilke lydnivåer som antas å forårsake slike skader er oppgitt for ulike grupper av sjøpattedyr (Southall et al. 2019). Lydnivåene (lavfrekvent lyd) når fundamenter pæles kan være opp til 250 dB rel. 1 µPa (Hastie et al. 2015), mens antatt hørselsskade hos sel oppstår for slike impulsive lyder oppstår ved 212 og 218 dB rel. 1 µPa for henholdsvis midlertidig og permanent skade (Southall et al. 2007). Niser er regnet som særlig sensitive, og har lavere terskel, og kan få midlertidig og varig skade ved nivåer helt ned i henholdsvis 196 og 202 dB rel. 1 µPa. Risikoen for at hørselsskade skal inntreffe er sterkt avhengig av avstand fra lydkilden, da lyden svekkes raskt med avstand. Risikoen for skade er derfor begrenset til nærområdet (innen noen hundre meter) rundt selve pælingen. Det er imidlertid utviklet støydempende tiltak når fundamentene pæles hvor særlig «boblegardiner» rundt pæleriggen kan redusere lyden betydelig (Tougaard & Mikaelsen 2018). De senere årene er det også utviklet flytende vindkraftanlegg hvor støyproblemet ved forankring til bunn er betydelig mindre.
Adferdseffekter kan potensielt inntreffe over et mye større område enn hørselsskade, og i teorien i hele området hvor lyden er hørbar. Det er påvist unnvikelse av habitat hos både sel og nise i områder hvor det foregår pæling (Brandt et al. 2011, Tougaard et al. 2009, Dähne et al. 2013, Russel et al. 2016), derfor vil dette kunne være et sannsynlig scenario i anleggsfasen. I tillegg er det blitt utført en rekke fokuserte studier på sensitivitet og adferds-forandringer hos steinkobbe, havert og andre sjøpattedyr i tilknytting til vindkraftanlegg i Nordsjøen (f.eks. Madsen et al. 2006, Thompson et al. 2013, Hastie et al. 2015, Tougard & Mikaelsen 2018), som indikerer at forstyrrelser og unnvikelse av habitat kan forekomme på ulike avstander, men at effekter på populasjonsnivå er usikkert.
Det finnes relativt lite kunnskap om hvordan sel og nise påvirkes av støy fra selve produksjonsfasen av et vindkraftanlegg. Lyd fra vindturbiner i drift kan spres ned i fundamentet og videre ut i havet, men disse støynivåer antas å være svært begrenset, og ikke utgjøre en alvorlig trussel mot sjøpattedyr (Madsen et al, 2006). Tilstedeværelse og antall sjøpattedyr i områder med aktive vindturbiner kan tyde på at sjøpattedyr ikke påvirkes negativt i anleggets driftsfase (Russell et al. 2016).
Referanser
Brandt M, Diederichs A, Betke K, Nehls G 2011. Responses of harbour porpoises to pile-driving at the Horns Rev II offshore wind farm in the Danish North Sea. Marine Ecology Progress Series, 421, 205–16.
Dähne M, Gilles A, Lucke K, Peschko V, Adler S, Krügel K, Sundermeyer J, Siebert U. 2013. Effects of pile-driving on harbour porpoises (Phocoena phocoena) at the first offshore wind farm in Germany. Environmental Research Letters 8, 025002. doi:10.1088/1748-9326/8/2/025002.
Hastie GD, Russell DJF, McConnell B, Moss S, Thompson D, Janik VM. 2015. Sound exposure in harbor seals during the installation of an offshore wind farm: predictions of auditory damage. Journal of applied Ecology, 52, 631-640.
Madsen PT, Wahlberg M, Tougaard J, Lucke K, Tyack P. 2006. Wind turbine underwater noise and marine mammals: Implications of current knowledge and data needs. Marine Ecology Progress Series, 309, 279–295. doi: 10.3354/meps309279.
Russell DJF, Hastie GD, Thompson D, Janik VM, Hammond PS, Scott-Hayward LAS, Matthiopoulos J, Jones EL, McConnell BJ. 2016. Avoidance of wind farms by harbour seals is limited to pile driving activities. Journal of Applied Ecology, 53, 1642-1652. doi: 10.1111/1365-2664.12678.
Thompson PM, Hastie GD, Nedwell J, Barham R, Brookes KL, Cordes LS, Bailey H, McLean N. 2013. Framework for assessing impacts of pile-driving noise from offshore wind farm construction on a harbour seal population. Environmental Impact Assessment Review, 43, 73–85.
Tougaard J, Carstensen J, Teilmann J, Skov H, Rasmussen P. 2009. Pile driving zone of responsiveness extends beyond 20 km for harbor porpoises (Phocoena phocoena). Journal of the Acoustic Society of America, 126, 11–4.
Tougaard J, Mikaelsen M. 2018. Effects of larger turbins for the offshore wind farm at Krieger’s Flak, Sweden. Assessment of impact on marine mammals. Scientific Report from DCE – Danish Centre for Environment and Energy No. 286 2018, 116 pp.
5.6 - Oppsummering kunnskapshull effekter av støy
Det finns flere kunnskapshull om hvordan støy fra vindkraftanlegg kommer til å påvirke havmiljøet. Herunder oppsummerer vi de viktigste:
Kan effekter fra impulsiv støy i utbyggingsfasen føre til populasjonseffekter?
De fleste studier har fokusert på anleggsfasen og pæleaktivitet, men man vet lite om hvordan lyd fra selve driftsfasen påvirker dyrene i og rundt anlegget: Hvordan påvirker langvarig kontinuerlig støy habitatbruk og adferd i et område?
Tilvenner fiskene som blir tiltrukket seg støynivået, eller blir reproduksjon og/ eller beiteadferd påvirket over lengre tid?
Hvor mye og hvilke arter av fiskeegg og yngel oppholder seg i vindkraftanlegg og i hvilken grad blir de påvirket av støy?
I hvilken grad er positive og negative effekter av kontinuerlig støy på evertebrater i balanse?
Blir potensielle negative effekter av støy for fisk utbalansert av positive effekter av økt mattilgang og ly?
6 - Konklusjoner
Det finns tydelige indikasjoner på at bunnfaste vindkraftanlegg tiltrekker flere arter og at mange arter beiter og reproduserer i anleggene. Kunstige rev-effekter av strukturene kan føre til økt mat og ly for fisk etter at bunndyr har etablert seg. Distribusjonen til arter og livsstadier som har liten egenbevegelse, kan bli påvirket av endringer i havstrømmer pga turbinene. Det er ennå ikke kjent i hvilken grad flytende vindturbiner har de samme effektene, eller hvordan flere anlegg samlet påvirker omgivelsene.
De elektromagnetiske feltene generert av undervannskabler kan oppfattes av mange marine dyr, med størst effekt innen noen titalls meter fra kabelen. Slike elektromagnetiske forstyrrelser kan teoretisk påvirke marine dyrs migrasjon, lokale bevegelser (attraksjon/unnvikelse), predator/byttes interaksjoner og dyrenes fysiologi/utvikling. Bruskfisk og dyr som bruker magnetiske felt for å orientere seg vil være spesielt sårbare overfor denne type forstyrrelser. Fysiologiske- eller utviklingseffekter kan ikke utelukkes for individer som oppholder seg svært nær kablene der feltstyrkene er høyest. Studiene som er gjennomført per i dag viser relativt marginale effekter av elektromagnetiske forstyrrelser på marine organismer, men behovet for mer kunnskap er stort.
I utbyggingsfasen kan lyd fra konstruksjonsarbeid som pæling skade hørsel og føre til en skremmeeffekt for både fisk og sjøpattedyr, men tiltak som boblegardiner kan redusere lydnivået betydelig. Selv om forskning på effekter av lyd fra vindkraftanlegg på sjødyr har primært fokusert på utbyggingsfasen, er det fortsatt ukjent om utbyggingsstøy kan føre til effekter på populasjonsnivå.
Kontinuerlig lavfrekvent støy fra turbinene vil først og fremst påvirke dyrene inne i og nært vindkraftanleggene. Siden bunndyr etablerer seg i området og fisk blir tiltrukket til anleggene, blir disse gruppene utsatt for produksjonsstøy over lengre tid. Kontinuerlig lavfrekvent støy kan påvirke atferd som, for eksempel, bunnmiksing, beiting, reproduksjon, anti-predator adferd og kommunikasjon, men hvor stor disse effektene er sammenlignet med positive effekter av økt mattilgang og ly i vindkraftanlegg er ukjent. Det er også kunnskapshull omkring marine dyrs evne til å tilpasse seg støypåvirkninger over tid.
Totalt sett er det med dagens kunnskap ikke mulig å forutsi om den totale effekten av vindkraftanlegg blir positive, negative eller nøytrale i norske havområder. Ved siden av de effektene som er allerede nevnt, kan fremmede arter bruke muligheten et nytt habitat gir for å etablere seg. Vi vet også lite om hvordan et vindkraftanlegg endrer lysforholdene i et område og hvordan dette påvirker lyssensitive organismer. For å dokumentere effekter av vindkraftanlegg trengs det en kombinasjon av kontrollerte eksperimenter som ser på ulike effekter separat og større feltstudier som sammenligner et område i og rundt et anlegg før og etter utbygging. Det er spesielt viktig å teste om effektene påvirker artsfordeling lokalt eller om de fører til endringer på populasjonsnivå (Figur 1). Derfor bør en forundersøkelse ha en varighet slik at år til år variasjoner fanges opp. I tillegg til undersøkelsene i driftsfasen, bør det også gjennomføres undersøkelser etter at anleggene er avviklet for å kontrollere inngrepets reversibilitet. En standardisert protokoll for undersøkelser som gjennomføres i forbindelse med utplassering av vindkraftanlegg i sjø kan gjøre det enklere å sammenlikne effekter av slike anlegg i norske havområder.
7 - Råd
Havforskningsinstituttets råd om seismikk og annen menneskeskapt støy blir oppdatert hvert år (Sivle et al. 2020). Fra i år skal rapporten også inneholde Havforskningsinstituttets råd om utbygging av vindkraftanlegg. Kunnskapen om miljøeffekter av vindkraftanlegg til havs er fortsatt mangelfull, og derfor er det sannsynlig at rådene kommer til å endres de neste årene, etter hvert som mer kunnskap kommer til.
For å få mer kunnskap om miljøeffekter fra havvindanlegg er det viktig å utforme en protokoll for å lage et sammenligningsgrunnlag for de forskjellige områdene hvor havvindkonstruksjoner blir planlagt. En slik protokoll må tilpasses norske forhold, f.eks. bør forundersøkelsene vare lenge nok til å avdekke eventuelle bestandssvingninger for mulig påvirkede bestander. Alle artene som bruker området til reproduksjon eller annen biologisk viktig adferd bør kartlegges nøye før og etter utbygging. Artene som etablerer seg etter utbygging bør registreres, med spesielt fokus på forekomster av fremmede arter.
Grunnet lite kunnskap om hvordan vindparker kan komme til å endre fysiske forhold som f.eks. havstrømmer og støy, bør første prioritet være å måle endringer i vannbevegelser, elektromagnetisme og støy i og rundt vindkraftanlegg.
Da det er knyttet betydelig usikkerhet til omfanget av miljøeffekter fra havvindanlegg i norske farvann fraråder HI foreløpig utbygginger i områder som er spesielt viktige for enkelte arter. Dette omfatter gyteområder for fisk og kaste- og beiteområder for enkelte sjøpattedyrarter. Utbygging bør også unngås i områder som er viktig for det marine økosystemet som for eksempel områdene rundt Lofoten, eller sårbare områder som korallrev.
I konstruksjonsperioden bør det brukes boblegardin, eller liknede, for å redusere kraftig støy, og alt konstruksjonsarbeid som medfører støy bør unngås nær gyteområder for fisk eller nær kasteområder for sel i gyte- eller kasteperioder.
Råd:
HI anbefaler at det utformes en standardisert protokoll for havvindanlegg tilpasset norske forhold, som innbefatter grundige forundersøkelser av områdene før utbygging, samt overvåking av både fysiske og biologiske endringer under drift og etter avvikling av anleggene. På grunn av store svingninger i tettheter til pelagiske arter skal forundersøkelsene foregår i minst tre år. Protokollen bør også beskrive hvordan data bør gjøres åpent tilgjengelig.
HI fraråder vindkraftutbygging i områder som er spesielt viktig for enkelte arter, som gyteområder og migrasjonsruter for aktuelle fiskebestander og beite- og kaste områder for enkelte sjøpattedyrarter.
HI fraråder vindkraftutbygging i spesielt sårbare eller verdifulle områder.
HI anbefaler at det ved utsteding av havvindkonsesjoner legges fram tydelige krav om fjerning av strukturene etter avsluttet drift.
HI anbefaler bruk av støydempende tiltak som boblegardin under utbygging, og at man unngår utbyggingsarbeid i gyteperioder for fisk, samt beite og kasteperioder for sjøpattedyr, i og nær områder dette gjelder.
HI anbefaler bruk av materialer i fortøyning av flytende vindturbiner som lager minst mulig støy ved bevegelser av turbinene, f.e unngå rykking og napping i ankerkjettingen.
HI anbefaler innsamling av data for å undersøke i hvilken grad, vindkraftanlegg endrer fysiske forhold som f.eks. strømmønster og lydbilde. Innsamlete data bør være åpent tilgjengelig.
HI anbefaler at det gjennomføres grundige forundersøkelser for å lage en oppdatert oversikt over artene i influensområdene før utbygging. Innsamlete data bør være åpent tilgjengelig.
HI anbefaler innsamling av data for å undersøke i hvilken grad, vindkraftanlegg endrer biologiske forhold som f.eks. artssammenstilling og adferd. Innsamlete data bør være åpent tilgjengelig.